Trace Elements in Plankton, Benthic Organisms, and Forage Fish of Lake Moreno, Northern Patagonia, Argentina Maria A. Arribére & Linda M. Campbell & Andrea P. Rizzo & Marina Arcagni & Jorge Revenga & Sergio Ribeiro Guevara Received: 24 July 2009 / Accepted: 13 January 2010 / Published online: 17 February 2010 # Springer Science+Business Media B.V. 2010 Abstract The Northern Patagonian Andean range shared by Chile and Argentina has numerous glacial oligotrophic lakes protected in a series of National Parks. Recent baseline surveys indicated that concen- trations in muscle and liver tissues from various fish species from across Nahuel Huapi and Los Alerces National Parks in Argentina were comparable or higher than similar fish species from other parts of the world. As a result, Lake Moreno, in Nahuel Huapi National Park, was chosen to investigate multiple element sinks, trends, and transfer in a representative Patagonia aquatic food web. The metals and metal- loids Ag, As, Ba, Br, Cs, Co, Cr, Fe, Hg, K, Na, Rb, Se, and Zn were analyzed in three size plankton fractions, submerged macrophytes, biofilm, insect larvae, amphipods, decapods, gastropods (snails), annelids (earthworms), and forage fish. Except for nanoplankton (10–53 μm; small-celled algae, rotifers) and microplankton (53–200 μm; larger algae, ciliates, zooplankton nauplii), which share elemental compo- sitional similarities, each taxon category had its own distinctive compositional pattern, revealed by princi- pal component analysis. Nano- and microplankton tend to be relatively elevated in some metals, including As, Co, Cr, Fe, Hg, Zn, and Rb, followed by biofilm. Shredder-scrapper Trichoptera (caddis- flies) have higher concentration of most of the studied elements than other insect larvae taxa, especially carnivorous Odonata (Anisoptera, dragonflies), which were associated with lower elemental contents. Those trends point to an overall tendency for biodiminishing element concentrations with trophic level in the benthos of Lake Moreno. Keywords Metals . Aquatic food web . Lake Moreno . Functional feeding groups Water Air Soil Pollut (2010) 212:167–182 DOI 10.1007/s11270-010-0330-3 M. A. Arribére (*) : A. P. Rizzo : M. Arcagni : S. Ribeiro Guevara Laboratorio de Análisis por Activación Neutrónica, UAIN, Centro Atómico Bariloche, Comisión Nacional de Energía Atómica (CNEA), Bustillo 9500, 8400 Bariloche, Argentina e-mail: [email protected] M. A. Arribére Instituto Balseiro, Universidad Nacional de Cuyo y CNEA, Bustillo 9500, 8400 Bariloche, Argentina L. M. Campbell Department of Biology and School of Environmental Studies, Queen’ s University, Kingston, ON K7L 3N6, Canada A. P. Rizzo CONICET, Buenos Aires, Argentina M. Arcagni : J. Revenga Centro Regional Universitario Bariloche (CRUB), Universidad Nacional del Comahue, Quintral 1250, 8400 Bariloche, Argentina

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Trace Elements in Plankton, Benthic Organisms, and ForageFish of Lake Moreno, Northern Patagonia, Argentina
Maria A. Arribére & Linda M. Campbell &Andrea P. Rizzo & Marina Arcagni &Jorge Revenga & Sergio Ribeiro Guevara
Received: 24 July 2009 /Accepted: 13 January 2010 /Published online: 17 February 2010# Springer Science+Business Media B.V. 2010
Abstract The Northern Patagonian Andean rangeshared by Chile and Argentina has numerous glacialoligotrophic lakes protected in a series of NationalParks. Recent baseline surveys indicated that concen-trations in muscle and liver tissues from various fishspecies from across Nahuel Huapi and Los Alerces
National Parks in Argentina were comparable orhigher than similar fish species from other parts ofthe world. As a result, Lake Moreno, in Nahuel HuapiNational Park, was chosen to investigate multipleelement sinks, trends, and transfer in a representativePatagonia aquatic food web. The metals and metal-loids Ag, As, Ba, Br, Cs, Co, Cr, Fe, Hg, K, Na, Rb,Se, and Zn were analyzed in three size planktonfractions, submerged macrophytes, biofilm, insectlarvae, amphipods, decapods, gastropods (snails),annelids (earthworms), and forage fish. Except fornanoplankton (10–53 μm; small-celled algae, rotifers)and microplankton (53–200 μm; larger algae, ciliates,zooplankton nauplii), which share elemental compo-sitional similarities, each taxon category had its owndistinctive compositional pattern, revealed by princi-pal component analysis. Nano- and microplanktontend to be relatively elevated in some metals,including As, Co, Cr, Fe, Hg, Zn, and Rb, followedby biofilm. Shredder-scrapper Trichoptera (caddis-flies) have higher concentration of most of the studiedelements than other insect larvae taxa, especiallycarnivorous Odonata (Anisoptera, dragonflies), whichwere associated with lower elemental contents. Thosetrends point to an overall tendency for biodiminishingelement concentrations with trophic level in thebenthos of Lake Moreno.
Keywords Metals . Aquatic food web . LakeMoreno .
Functional feeding groups
Water Air Soil Pollut (2010) 212:167–182DOI 10.1007/s11270-010-0330-3
M. A. Arribére (*) :A. P. Rizzo :M. Arcagni :S. Ribeiro GuevaraLaboratorio de Análisis por Activación Neutrónica, UAIN,Centro Atómico Bariloche,Comisión Nacional de Energía Atómica (CNEA),Bustillo 9500,8400 Bariloche, Argentinae-mail: [email protected]
M. A. ArribéreInstituto Balseiro, Universidad Nacional de Cuyo y CNEA,Bustillo 9500,8400 Bariloche, Argentina
L. M. CampbellDepartment of Biology and School of EnvironmentalStudies, Queen’s University,Kingston, ON K7L 3N6, Canada
A. P. RizzoCONICET,Buenos Aires, Argentina
M. Arcagni : J. RevengaCentro Regional Universitario Bariloche (CRUB),Universidad Nacional del Comahue,Quintral 1250,8400 Bariloche, Argentina

1 Introduction
The Northern Patagonian Andean mountain range(39° to 45° S, 71° W; 500 to 3,554 m a.s.l.), shared byChile and Argentina, have numerous glacial lakes,which drain toward the Atlantic and Pacific oceans.Both countries have established national parks alongtheir shared border to protect the “Lake District.” TheNahuel Huapi National Park (NHNP) on the Argen-tine side encompasses 785,000 ha, including drainagebasins of three major river systems.
Because of its isolation and low population density,this area has been historically protected from anthro-pogenic contamination, with the most importantsource of metals being from the frequent Andesvolcanic activity (Stern 2004; Campbell et al. 2007).However, over the past 60 years, the human populationin the region has rapidly increased by a factor of ten.Tourism, both from within Argentina and outside,fishing, and recreational water sports are importantsources of revenue and are fuelling a major populationgrowth with subsequent increase in environmentalcontaminants. Recently, a baseline survey indicated thatmetal (Cs, Fe, Br, Se, Rb, Na, and Zn) concentrations inmuscle and liver tissues from various fish species fromacross Nahuel Huapi National Park and the neighboringLos Alerces National Park were comparable or higherthan similar fish species from other parts of the world(Ribeiro Guevara et al. 2004; Campbell et al. 2005;Arribére et al. 2006; Arribére et al. 2008). The actualtrophodynamics and metal transfer routes within foodwebs leading to fish still require analysis and quanti-fication for better management of the economicallyimportant sport fisheries.
Lake Moreno, a double-basined lake situated byLake Nahuel Huapi and the City of San Carlos deBariloche, provides an ideal case study due to itsaccessibility and similarity to other less accessiblelakes of Nahuel Huapi National Park. The aim of thisstudy is to investigate metal trends and transferthrough the lower food web of Lake Moreno usingmultivariate approaches and how that relates to metalconcentrations in sportfish.
2 Methods
Study Site Lake Moreno (41°5′ S; 71°33′ W, 758 m a.s.l.), in Nahuel Huapi National Park, is an hour-glass-
shaped warm monomictic and ultraoligatrophic sys-tem. It is stratified from late spring to early autumnwith a thermocline at about 30 m depth. The twobasins, Lake Moreno West (MW) and Lake MorenoEast (ME), are connected by a short narrow channel(Fig. 1). Lake Moreno West, in turn, is connected toLake Nahuel Huapi, the largest lake of the Park(560 km2), through a short channel. Table 1 shows themain characteristics of both basins.
Lake Moreno West has a highly irregular coast line,with peninsulas, bays, and flooded areas, surrounded bytemperate rainforest dominated by Nothofagus dombeyiand Austrocedrus chilensis (Queimaliños et al. 1999).The coast line of Lake Moreno East is more regularand present rocky beaches, and the drier rainforest–steppe ecotone vegetation coverage in the surroundingsof the lake is less dense. This region is increasinglybecoming periurban, with family housing and hoteldevelopment along the shores, in addition to a rainbowtrout fish farm in ME.
Field Sampling Lakes Moreno West and East weresampled four consecutive seasons (spring, summer,autumn, and winter), from 2005 to 2006. The samplingsite chosen from Moreno West was on the westernmostside, with dense rainforest in the surroundings, wherethe lowlands are flooded in spring time during thesnow melting period (GP, in Fig. 1). The siteschosen on Moreno East were the North and Southmargins (S and N; Fig. 1), in rocky beaches. Thetraps on the North side were damaged duringsampling; this site was abandoned, remaining onlythe South margin site. All plastic and glass vesselsand bottles used in the field or in the laboratory werewashed for 15 days in 50% HNO3 and thoroughlyrinsed in ASTM grade 1 water before use. Filteringdevices and filters were washed in 50% HNO3
whenever possible. The filters (made out of the netmaterial NITEX®), which could not stand acidcleaning, were washed with EXTRAN® detergentand thoroughly rinsed, using a bath with ultrasound.
All sampled specimens were rinsed with lake waterto remove excess sediment and particulates, taken tothe laboratory immediately after sampling where theywere rinsed with ASTM-grade water, stored at −20°Cuntil freeze drying, and then homogenized usingTeflon and titanium tools.
The food web sampling focused on items impor-tant to the larger sport fish diet: galaxids (Galaxias
168 Water Air Soil Pollut (2010) 212:167–182

maculatus), salmonid young of the year (YOY),decapods (Samastacus sp., Aegla sp.), amphipods(Hyalella sp.), insect larvae (Plecoptera, Ephemerop-tera, Odonata, Trichoptera, and Coleoptera), gastro-pods (Chilina sp.), and Hirudinea.
Galaxids, salmonid juveniles, and macrocrusta-ceans (Samastacus sp. and Aegla sp.) were caughtwith baited traps set out for 2 to 4 days in the littoralzone. Brook trout and rainbow trout juveniles rangedin length from 7 to 18 cm and were pooled by size.
Fig. 1 Map of the study area
Units Moreno East Moreno West
Areaa km2 5.4 4.8
Conductivity μS cm−1 37 37
Maximum deptha m 128 110
Average deptha m 69 60
Maximum lengtha km 4.4 4.7
Maximum widtha km 1.8 2.0
Surface temperature °C 8 (winter)–16.5 (summer) 8 (winter)–17.5 (summer)
pH 6.8 6.8
Chlorophyll ab μg L−1 1 1
Table 1 Chemical, physi-cal, and limnologicalparameters of Lake Moreno
a Quirós (1988)b Souza et al. (2007)
Water Air Soil Pollut (2010) 212:167–182 169

Because of their abundance and small masses,galaxids were pooled into five size classes to ensuresufficient analytical mass: G1, 3 to 4 cm length; G2, 4to 5 cm; G3, 5 to 6 cm; G4, 6 to 7 cm; and G5>7 cm.Each pooled set consisted of ten to 30 individuals;and to check for individual variability, in the springseason, when the largest set of small fish was caught(belonging to groups G1, G2, and G3), three sets of30 fish of each group were prepared as independentsamples to search for sampling variability. Since theshell of adult decapods and gastropods is notcompletely digested by fish, the majority of Samas-tacus, Aegla, and Chilina samples were dissected formuscle and digestive tissues (hepatopancreas), whichwere analyzed separately. Only a few selected adultSamastacus and Aegla samples were analyzed whole.All other invertebrates, insect larvae, Hyalella, Hiru-dinea, and soft-shelled juvenile Aegla sp (<2 cm,collected in summer and fall), were collected by handalong the shoreline. Pooled samples were created withhomogenized whole-body organisms since fish candigest those organisms in its entirety.
Nanoplankton (10–53 µm; consisting of hapto-phyte, cryptophytes, dinoflagellates, and rotifers),microplankton (53–200 µm; consisting of largerphytoplankton, ciliates, and smaller zooplankton),and zooplankton (≥200 μm; consisting of largercopepods and cladocerans) were collected using aseries of conical NITEX® nets in four different sites.Since Moreno East has a rather regular basin with themaximum depth in the middle, a central samplingpoint was chosen (ME, Fig. 1). Three sampling siteswere chosen in Moreno West: a site where the lakereaches its maximum depth (PP), a shallow and rathernarrow bay (LL, 18 m deep), and an open bay on theWest side (MW) 45 m deep. Samples were collectedby vertical hauls from 50 m depth to the lake surfacein deeper sites (ME, PP) and from about 2 to 4 mabove the lake bottom in the shallower sites (LL,MW). All samples were concentrated in the laborato-ry by filtering through 0.2 μm Nucleopore® polycar-bonate acid-washed filters and freeze-dried untilconstant weight.
Submerged macrophytes, moss, and benthic algaewere collected by hand. Biofilm (composed of algae,bacteria, decomposing organic matter, and very finedetritical particulate) was very abundant in spring andsummer and was sampled by gently scraping thesurface of folded NITEX® nets set in the littoral zone,
macrophyte leaves, and rock surfaces with acid-washed plastic or titanium utensils. Biofilm wasscarce in autumn and winter, and it was not possibleto collect enough mass for analysis, with the onlywinter sample obtained after one unusually sunny andwarm week in Moreno East (S).
Elemental Analyses Aliquots of about 100 mg ofdried homogenized and powdered sample were sealedin SUPRASIL AN® quartz ampoules, irradiated for24 h, and analyzed by Instrumental Neutron Activa-tion Analysis (INAA) at the RA-6 nuclear researchreactor at Centro Atómico Bariloche. Ag, As, Ba, Br,Cs, Co, Cr, Fe, Hf, Hg, K, La, Na, Rb, Sb, Se, Sm, U,and Zn were measured for all samples, while Ca wasalso measured in macrocrustacean samples. Gamma-ray spectra were collected using an intrinsic HighPurity Germanium (HPGe) n-type detector, 12.3%relative efficiency, and a 4096-channel analyzer.Spectra were analyzed by using the GAMANALroutine included in the GANAAS package, distributedby International Atomic Energy Agency (IAEA).Corrections for spectral interferences were includedwhen necessary. The concentrations reported arereferred to dry weight basis. Standard referencematerials from the National Research Council ofCanada (NRCC)-DORM-2 Dogfish Muscle, NRCC-TORT-2 Lobster, NRCC-DOLT-2 Dogfish Liver, andfrom the International Atomic Energy Agency, IAEA-336 lichen material, IAEA-392 Algae material, aswell as sample replicates, were analyzed to check onthe quality of analysis. The analyses of the standardreference materials showed good agreement withcertified and informed values, and replicate sampleswere consistent.
Data Analyses Bivariate correlation and principalcomponent analysis (PCA) were used to identifyassociation of food items as sources of an elementor set of elements. Bivariate correlation allowsidentifying elemental contents related to geogenicmaterial, especially in insect larvae and other sampleswhich could retain entrapped geological particulate.Since Ba was determined only in a limited number ofsamples of fish and macroinvertebrates due toanalytical detection limits, it was included in the PCanalysis when only macrophyte, biofilm, plankton,and insect larvae were considered. Cr contents insome samples were below all other measurements in
170 Water Air Soil Pollut (2010) 212:167–182

any sample considered; for these cases, the detectionlimit was included as input data for the PCA. Fish Asand Ba were not included in the PCA because the datawere not available for all samples.
Previously published trace element data for sevenfish species and one native mussel species from thislake were also included in the data analyses to enabledirect comparison of invertebrate and fish data. Thefish species included are Diplomystes viedmensis(velvet catfish), Odontesthes hatcheri (creole silver-side or pejerrey), Oncorhynchus mykiss (rainbowtrout), Percichthys trucha (creole perch), Salmo trutta(brown trout), and Salvelinus fontinalis (brook trout),and the sampling and analysis details can be found inRibeiro Guevara et al. (2006), while sampling andanalysis information for the native mussel Diplodonchilensis were taken from Ribeiro Guevara et al.(2004). Since the literature reports that musselpredators consume only the soft tissue (Lara andMoreno 1995), data for mussels without valves werethe only included.
3 Results and Discussion
The lithophile and the rare earth elements (REE) ofthe lanthanide and actinide series, Hf, U, La, Sb, andSm, are highly correlated with each other in the lowerfood web species (bivariate analysis, r2>0.9). REEand other insoluble elements are sometimes found infish and decapod tissues, but in significantly lowerconcentrations than in other organisms such as insectlarvae. REE are lithophile elements associated withgeological material trapped within the sample andthus having little biological significance within softtissues of animals. As a result, REE were notconsidered in the subsequent statistical analysis.
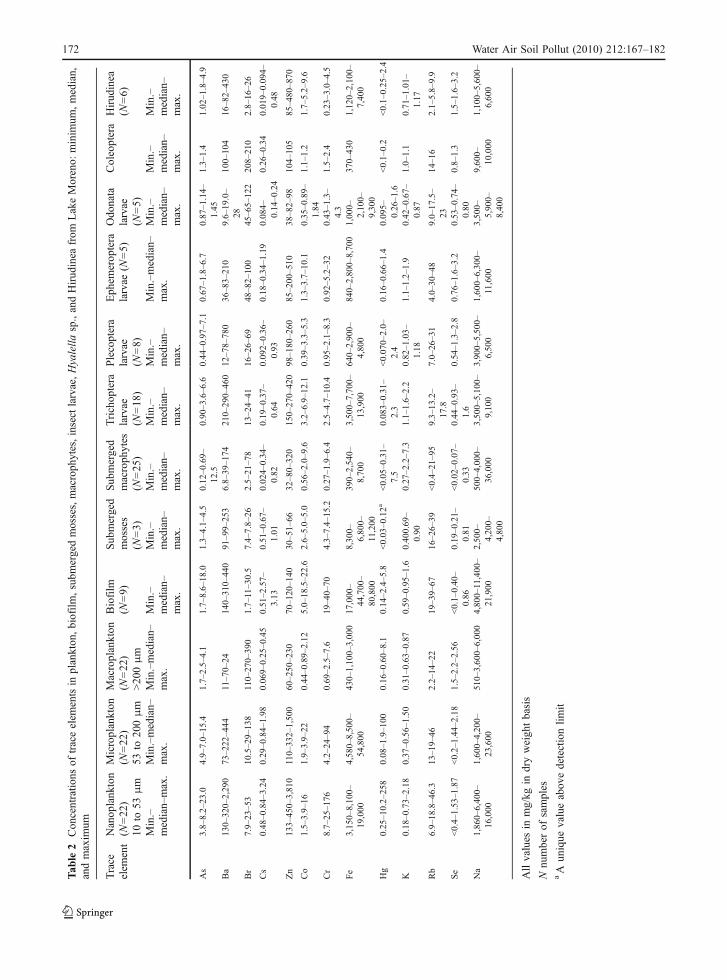
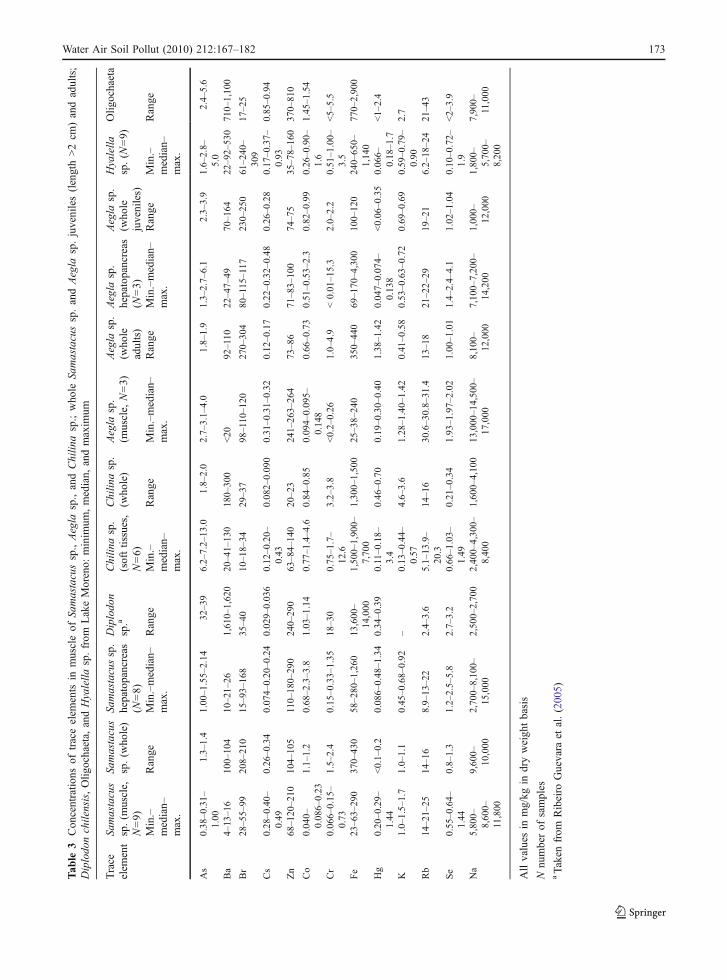
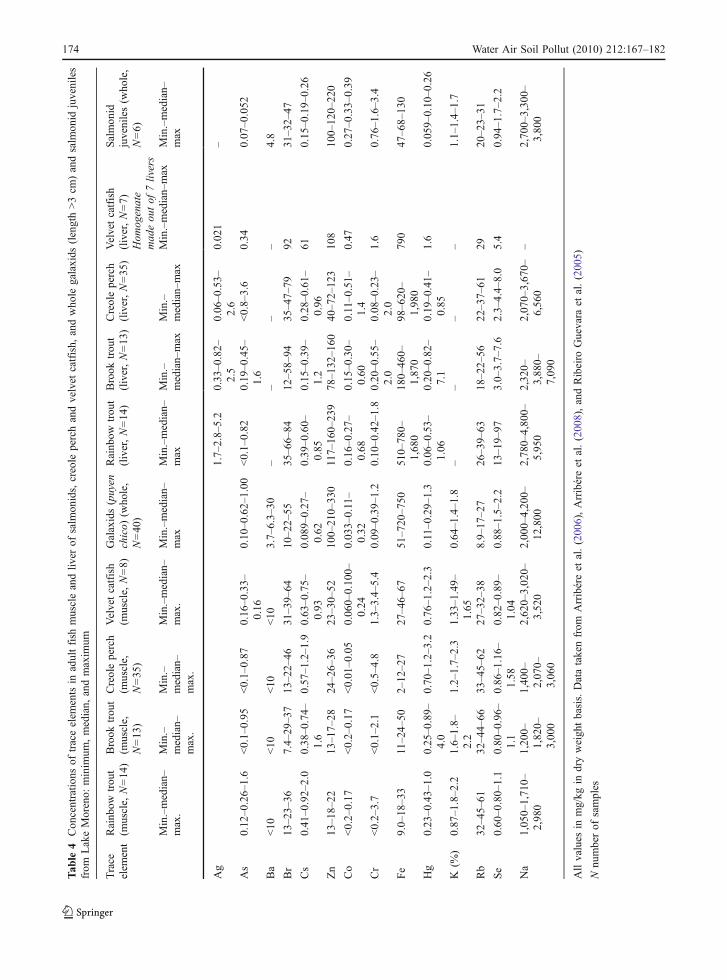
The minimum, median, maximum, average, andstandard deviation of As, Ba, Br, Cs, Co, Cr, Fe, Hg,K, Na, Rb, Se, and Zn concentrations of each type oforganisms are shown in Tables 2, 3, and 4.
Table 5 compares the elemental contents measuredin this work with some data found in the literature forsimilar species. Although comparisons are not alwayspossible due to the heterogeneity of statisticalmeasurements (ranks and means), data suggestenriched Zn, Fe, and Hg concentrations in planktonfrom this work (with the exception of Zn in macro-
plankton) compared to other published data. Inbenthic organisms, Ephemeroptera and Hyalella sp.have higher Hg burdens, but contents in whole fishare similar or lower.
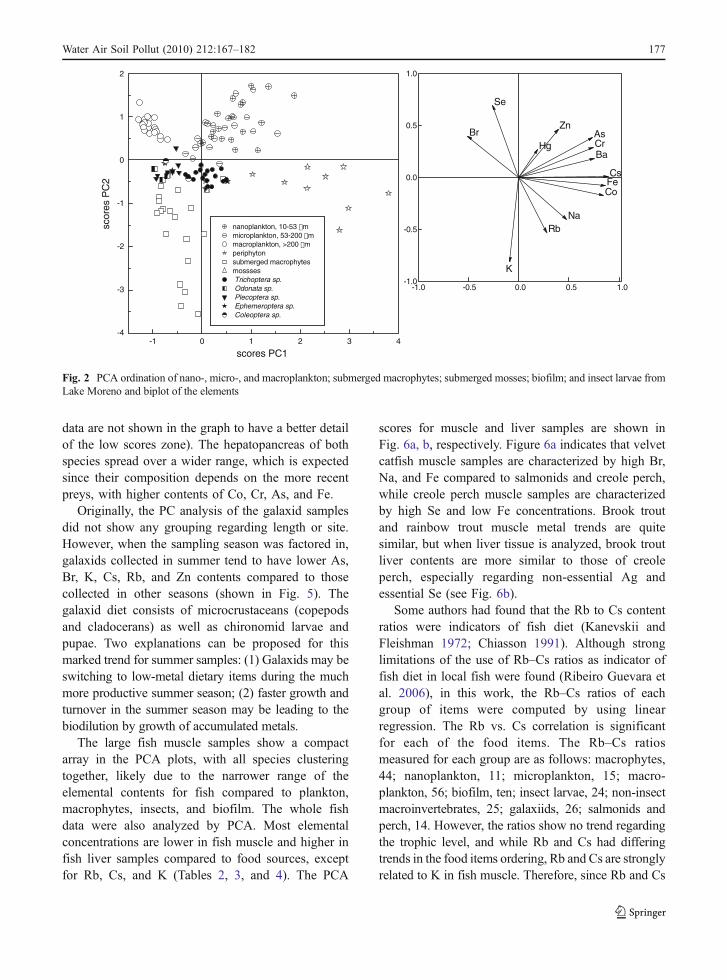
The primary producers fell across a broad range forall the other transitional and semi-metalloid elements.The first principal component of the PCA for theprimary producers and zooplankton showed distinc-tive separation of four groups (Fig. 2). The firstgroup, nano- (10–53 μm) and microplankton (53–200 μm), was clustered apart from the second groupconsisting of zooplankton (≥200 μm). Macrophytesand mosses formed a tight third cluster, while biofilmformed the fourth. Biofilm and nano- and micro-plankton samples tend to be richer in As, Ba, Co, Cr,Cs, and Fe and poorer in Br and Se. All planktonfractions are poorer in K compared to macrophytes,mosses, and biofilm and tend to be richer in Se andZn. The PCA components 1 and 2 plot for the insectlarvae show that insects are located between planktonsamples and macrophytes (below), having biofilm tothe right. Within this cluster, the five Odonatasamples fall to the left end of the distribution andshares similar PC1 scores with zooplankton. Trichop-tera is closer to the phytoplankton and biofilm PCscores. Plecoptera and Ephemeroptera samples liebetween Odonata and Trichoptera. According to thisplot, Trichoptera metal trends are more closelyrelated to biofilm, which suggests that Trichopteramay be obtaining their metal burdens from grazingon biofilm. Odonata are known to be predatory(Westfall and Tennesseen 1996), and the PC1 scoregradient from Trichoptera to Odonata suggests afeeding gradient from biofilm grazing to feedingupon other organisms. Relating PC1 with elements,an increase in carnivory is accompanied with adecrease in As, Ba, Co, Cr, Cs, and Fe and anincrease in Br. This biodilution effect in macro-invertebrates food chain has been observed by Faraget al. (1998) for As, Hg, and Zn and by Watanabe etal. (2008) among different functional feeding groupsof benthic macroinvertebrates.
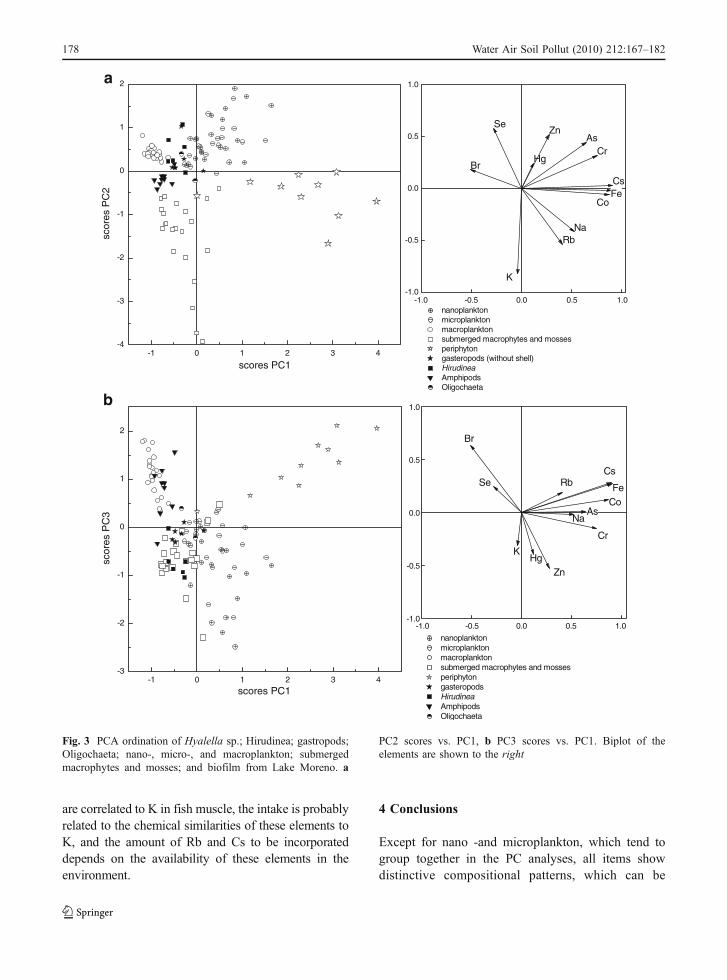
The PC1 vs. PC2 scores for the non-insectinvertebrates, Hyalella, Hirudinea, Oligochaeta, andgastropods (Fig. 3a), in addition to primary producers,indicate that Hyalella samples are clustered close tothe macrophytes and zooplankton, while Chilina sp.,Oligochaeata, and Hirudinea cluster between the PCscores for zooplankton and micro- and nanoplankton.
Water Air Soil Pollut (2010) 212:167–182 171
Tab
le2
Con
centratio
nsof
traceelem
entsin
plankton
,biofilm
,subm
ergedmosses,macroph
ytes,insectlarvae,Hyalella
sp.,andHirud
inea
from
LakeMoreno:
minim
um,median,
andmaxim
um
Trace
elem
entNanop
lank
ton
(N=22
)Microplankton
(N=22
)Macroplankton
(N=22
)Biofilm
(N=9)
Sub
merged
mosses
(N=3)
Sub
merged
macroph
ytes
(N=25
)
Trichop
tera
larvae
(N=18
)
Plecoptera
larvae
(N=8)
Eph
emerop
tera
larvae
(N=5)
Odo
nata
larvae
(N=5)
Coleoptera
Hirud
inea
(N=6)
10to
53μm
53to
200μm
>20
0μm
Min.–
median–
max.
Min.–median–
max.
Min.–median–
max.
Min.–
median–
max.
Min.–
median–
max.
Min.–
median–
max.
Min.–
median–
max.
Min.–
median–
max.
Min.–median–
max.
Min.–
median–
max.
Min.–
median–
max.
Min.–
median–
max.
As
3.8–
8.2–23.0
4.9–7.0–
15.4
1.7–2.5–
4.1
1.7–
8.6–18.0
1.3–
4.1–
4.5
0.12–0
.69–
12.5
0.90–3
.6–6.6
0.44
–0.97–
7.1
0.67–1
.8–6.7
0.87–1
.14–
1.45
1.3–
1.4
1.02
–1.8–4.9
Ba
130–
320–2,290
73–222
–444
11–7
0–24
140–310–
440
91–99–
253
6.8–39
–174
210–290–460
12–7
8–780
36–83–
210
9.6–19.0–
28100–104
16–82–
430
Br
7.9–
23–5
310.5–2
9–138
110–270–
390
1.7–
11–3
0.5
7.4–
7.8–
262.5–21
–78
13–24–
4116–2
6–69
48–82–
100
45–65–
122
208–210
2.8–
16–2
6
Cs
0.48
–0.84–
3.24
0.29–0
.84–
1.98
0.069–
0.25–0
.45
0.51
–2.57–
3.13
0.51
–0.67–
1.01
0.024–
0.34–
0.82
0.19–0
.37–
0.64
0.092–
0.36
–0.93
0.18–0
.34–
1.19
0.084–
0.14
–0.24
0.26
–0.34
0.019–
0.094–
0.48
Zn
133–
450–3,810
110–
332–
1,500
60–2
50–2
3070–1
20–140
30–51–
6632–8
0–320
150–270–420
98–1
80–260
85–200
–510
38–82–
98104–105
85–480
–870
Co
1.5–
3.9–16
1.9–3.9–
220.44–0
.89–
2.12
5.0–
18.5–2
2.6
2.6–
5.0–
5.0
0.56–2
.0–9
.63.2–6.9–
12.1
0.39
–3.3–5
.31.3–3.7–
10.1
0.35–0
.89–
1.84
1.1–
1.2
1.7–
5.2–
9.6
Cr
8.7–
25–1
764.2–24–9
40.69–2
.5–7
.619–4
0–70
4.3–
7.4–
15.2
0.27–1
.9–6
.42.5–4.7–
10.4
0.95
–2.1–8
.30.92–5
.2–32
0.43–1
.3–
4.3
1.5–
2.4
0.23
–3.0–4.5
Fe
3,150–
8,100–
19,000
4,580–8,500–
54,800
430–1,100–
3,000
17,000
–44,700
–80,800
8,300–
6,800–
11,200
390–2,540–
8,700
3,500–7,700–
13,900
640–
2,900–
4,800
840–2,800–
8,700
1,000–
2,100–
9,300
370–430
1,120–
2,100–
7,400
Hg
0.25
–10.2–
258
0.08–1
.9–100
0.16–0
.60–
8.1
0.14
–2.4–5
.8<0.03
–0.12a
<0.05
–0.31–
7.5
0.083–0.31
–2.3
<0.070–
2.0–
2.4
0.16–0
.66–
1.4
0.095–
0.26
–1.6
<0.1–
0.2
<0.1–
0.25
–2.4
K0.18
–0.73–
2.18
0.37–0
.56–
1.50
0.31–0
.63–
0.87
0.59
–0.95–
1.6
0.400.69
–0.90
0.27–2
.2–7
.31.1–1.6–
2.2
0.82
–1.03–
1.18
1.1–1.2–
1.9
0.42–0
.67–
0.87
1.0–
1.1
0.71
–1.01–
1.17
Rb
6.9–
18.8–4
6.3
13–19–
462.2–14
–22
19–3
9–67
16–26–
39<0.4–
21–95
9.3–13.2–
17.8
7.0–
26–3
14.0–30–4
89.0–17.5–
2314–1
62.1–
5.8–
9.9
Se
<0.4–
1.53
–1.87
<0.2–1.44
–2.18
1.5–2.2–
2.56
<0.1–
0.40
–0.86
0.19
–0.21–
0.81
<0.02
–0.07–
0.33
0.44–0
.93–
1.6
0.54
–1.3–2
.80.76–1
.6–3.2
0.53–0
.74–
0.80
0.8–
1.3
1.5–
1.6–
3.2
Na
1,860–
6,400–
16,000
1,600–4,200–
23,600
510–3,600–
6,000
4,800–
11,400
–21,900
2,500–
4,200–
4,800
500–4,000–
36,000
3,500–5,100–
9,100
3,900–
5,500–
6,500
1,600–6,300–
11,600
3,500–
5,900–
8,400
9,600–
10,000
1,100–
5,600–
6,600
Allvalues
inmg/kg
indryweigh
tbasis
Nnu
mberof
samples
aA
unique
valueabov
edetectionlim
it
172 Water Air Soil Pollut (2010) 212:167–182
Tab
le3
Con
centratio
nsof
traceelem
ents
inmuscleof
Samastacussp.,Aegla
sp.,andChilin
asp.;who
leSa
mastacussp.andAegla
sp.juveniles(length>2cm
)andadults;
Diplodo
nchilensis,Olig
ochaeta,
andHyalella
sp.from
LakeMoreno:
minim
um,median,
andmaxim
um
Trace
elem
entSa
mastacus
sp.(m
uscle,
N=9)
Samastacus
sp.(w
hole)
Samastacussp.
hepatopancreas
(N=8)
Diplodo
nsp.a
Chilin
asp.
(softtissues,
N=6)
Chilin
asp.
(who
le)
Aegla
sp.
(muscle,
N=3)
Aegla
sp.
(who
leadults)
Aegla
sp.
hepatopancreas
(N=3)
Aegla
sp.
(who
lejuveniles)
Hyalella
sp.(N
=9)
Olig
ochaeta
Min.–
median–
max.
Range
Min.–median–
max.
Range
Min.–
median–
max.
Range
Min.–median–
max.
Range
Min.–median–
max.
Range
Min.–
median–
max.
Range
As
0.38
–0.31–
1.00
1.3–
1.4
1.00
–1.55–
2.14
32–3
96.2–
7.2–
13.0
1.8–2.0
2.7–
3.1–
4.0
1.8–
1.9
1.3–
2.7–
6.1
2.3–
3.9
1.6–
2.8–
5.0
2.4–
5.6
Ba
4–13
–16
100–
104
10–2
1–26
1,610–1,620
20–4
1–130
180–
300
<20
92–110
22–4
7–49
70–1
6422
–92–530
710–
1,100
Br
28–5
5–99
208–
210
15–9
3–168
35–40
10–1
8–34
29–3
798
–110
–120
270–
304
80–115
–117
230–250
61–2
40–
309
17–2
5
Cs
0.28
–0.40–
0.49
0.26
–0.34
0.074–
0.20
–0.24
0.029–0.036
0.12
–0.20–
0.43
0.082–
0.090
0.31
–0.31–
0.32
0.12
–0.17
0.22
–0.32–
0.48
0.26–0
.28
0.17
–0.37–
0.93
0.85
–0.94
Zn
68–1
20–2
10104–
105
110–
180–
290
240–290
63–8
4–140
20–2
3241–
263–
264
73–8
671
–83–
100
74–7
535
–78–160
370–
810
Co
0.040–
0.086–0.23
1.1–
1.2
0.68
–2.3–3
.81.03–1
.14
0.77
–1.4–4
.60.84
–0.85
0.094–
0.095–
0.148
0.66
–0.73
0.51
–0.53–
2.3
0.82–0
.99
0.26
–0.90–
1.6
1.45
–1.54
Cr
0.066–
0.15
–0.73
1.5–
2.4
0.15
–0.33–
1.35
18–30
0.75
–1.7–
12.6
3.2–
3.8
<0.2–
0.26
1.0–
4.9
<0.01
–15.3
2.0–2.2
0.51
–1.00–
3.5
<5–
5.5
Fe
23–6
3–290
370–
430
58–2
80–1
,260
13,600–
14,000
1,500–
1,900–
7,700
1,300–
1,500
25–3
8–240
350–
440
69–1
70–4
,300
100–120
240–650–
1,140
770–
2,900
Hg
0.20
–0.29–
1.44
<0.1–
0.2
0.086–
0.48
–1.34
0.34–0
.39
0.11–0
.18–
3.4
0.46
–0.70
0.19
–0.30–
0.40
1.38
–1.42
0.047–
0.074–
0.138
<0.06
–0.35
0.066–
0.18
–1.7
<1–
2.4
K1.0–
1.5–
1.7
1.0–
1.1
0.45
–0.68–
0.92
–0.13
–0.44–
0.57
4.6–
3.6
1.28
–1.40–
1.42
0.41
–0.58
0.53
–0.63–
0.72
0.69–0
.69
0.59
–0.79–
0.90
2.7
Rb
14–2
1–25
14–1
68.9–
13–2
22.4–3.6
5.1–
13.9–
20.3
14–1
630.6–3
0.8–
31.4
13–1
821
–22–
2919–2
16.2–
18–2
421
–43
Se
0.55
–0.64–
1.44
0.8–
1.3
1.2–
2.5–
5.8
2.7–3.2
0.66
–1.03–
1.49
0.21
–0.34
1.93
–1.97–
2.02
1.00
–1.01
1.4–
2.4–
4.1
1.02–1
.04
0.10
–0.72–
1.9
<2–
3.9
Na
5,800–
8,600–
11,800
9,600–
10,000
2,700–
8,100–
15,000
2,500–2,700
2,400–
4,300–
8,400
1,600–
4,100
13,000
–14,500–
17,000
8,100–
12,000
7,100–
7,200–
14,200
1,000–
12,000
1,800–
5,700–
8,200
7,900–
11,000
Allvalues
inmg/kg
indryweigh
tbasis
Nnu
mberof
samples
aTaken
from
Ribeiro
Guevara
etal.(200
5)
Water Air Soil Pollut (2010) 212:167–182 173
Tab
le4
Con
centratio
nsof
traceelem
entsin
adultfish
muscleandliv
erof
salm
onids,creole
perchandvelvet
catfish,
andwho
legalaxids
(length>3cm
)andsalm
onid
juveniles
from
LakeMoreno:
minim
um,median,
andmaxim
um
Trace
elem
entRainb
owtrou
t(m
uscle,
N=14
)Brook
trou
t(m
uscle,
N=13
)
Creoleperch
(muscle,
N=35
)
Velvetcatfish
(muscle,
N=8)
Galaxids(puyen
chico)
(who
le,
N=40
)
Rainb
owtrou
t(liver,N=14
)Brook
trou
t(liver,N=13
)Creoleperch
(liver,N=35
)Velvetcatfish
(liver,N=7)
Hom
ogenate
mad
eou
tof
7livers
Salmon
idjuveniles(w
hole,
N=6)
Min.–median–
max.
Min.–
median–
max.
Min.–
median–
max.
Min.–median–
max.
Min.–median–
max
Min.–median–
max
Min.–
median–
max
Min.–
median–
max
Min.–median–
max
Min.–median–
max
Ag
1.7–2.8–
5.2
0.33
–0.82–
2.5
0.06–0
.53–
2.6
0.02
1–
As
0.12
–0.26–
1.6
<0.1–
0.95
<0.1–
0.87
0.16–0
.33–
0.16
0.10
–0.62–
1.00
<0.1–0.82
0.19
–0.45–
1.6
<0.8–3.6
0.34
0.07
–0.052
Ba
<10
<10
<10
<10
3.7–
6.3–30
––
––
4.8
Br
13–23–36
7.4–
29–3
713
–22–46
31–3
9–64
10–22–
5535–6
6–84
12–58–94
35–4
7–79
9231
–32–47
Cs
0.41
–0.92–
2.0
0.38
–0.74–
1.6
0.57
–1.2–1
.90.63–0
.75–
0.93
0.08
9–0.27
–0.62
0.39–0
.60–
0.85
0.15
–0.39–
1.2
0.28–0
.61–
0.96
610.15
–0.19–0.26
Zn
13–18–22
13–1
7–28
24–2
6–36
23–3
0–52
100–
210–
330
117–16
0–23
978
–132–1
6040–7
2–12
310
810
0–12
0–22
0
Co
<0.2–
0.17
<0.2–
0.17
<0.01
–0.05
0.06
0–0.10
0–0.24
0.03
3–0.11–
0.32
0.16–0
.27–
0.68
0.15
–0.30–
0.60
0.11–0.51–
1.4
0.47
0.27
–0.33–
0.39
Cr
<0.2–
3.7
<0.1–
2.1
<0.5–
4.8
1.3–3.4–
5.4
0.09
–0.39–
1.2
0.10–0
.42–1.8
0.20
–0.55–
2.0
0.08–0
.23–
2.0
1.6
0.76
–1.6–3
.4
Fe
9.0–
18–3
311–24–
502–
12–2
727–4
6–67
51–720
–750
510–
780–
1,68
018
0–46
0–1,87
098–6
20–
1,98
079
047
–68–13
0
Hg
0.23
–0.43–
1.0
0.25
–0.89–
4.0
0.70
–1.2–3
.20.76–1
.2–2
.30.11–0
.29–1.3
0.06–0
.53–
1.06
0.20
–0.82–
7.1
0.19–0
.41–
0.85
1.6
0.05
9–0.10–0
.26
K(%
)0.87
–1.8–2
.21.6–
1.8–
2.2
1.2–
1.7–2.3
1.33–1
.49–
1.65
0.64
–1.4–1
.8–
––
–1.1–
1.4–1.7
Rb
32–45–61
32–4
4–66
33–4
5–62
27–3
2–38
8.9–
17–2
726–3
9–63
18–22–56
22–3
7–61
2920
–23–31
Se
0.60
–0.80–
1.1
0.80
–0.96–
1.1
0.86
–1.16–
1.58
0.82–0
.89–
1.04
0.88
–1.5–2
.213–1
9–97
3.0–
3.7–7.6
2.3–4.4–
8.0
5.4
0.94
–1.7–2
.2
Na
1,05
0–1,71
0–2,98
01,20
0–1,82
0–3,00
0
1,40
0–2,07
0–3,06
0
2,62
0–3,02
0–3,52
02,00
0–4,20
0–12
,800
2,78
0–4,80
0–5,95
02,32
0–3,88
0–7,09
0
2,07
0–3,67
0–6,56
0–
2,70
0–3,30
0–3,80
0
Allvalues
inmg/kg
indryweigh
tbasis.Datatakenfrom
Arribéreet
al.(200
6),Arribéreet
al.(200
8),andRibeiro
Guevara
etal.(200
5)
Nnu
mberof
samples
174 Water Air Soil Pollut (2010) 212:167–182
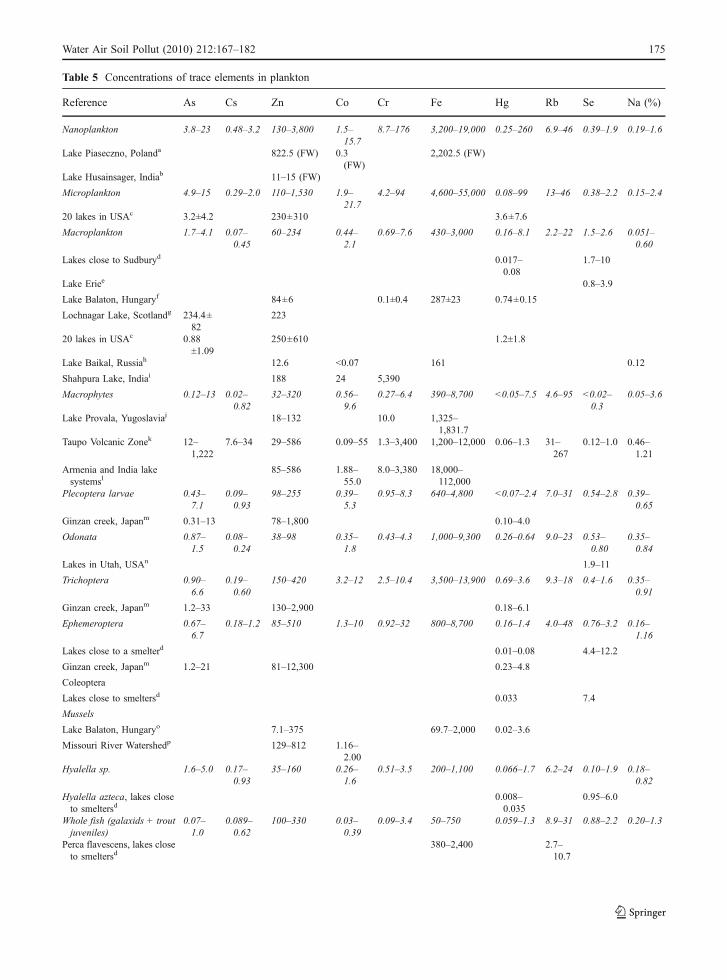
Table 5 Concentrations of trace elements in plankton
Reference As Cs Zn Co Cr Fe Hg Rb Se Na (%)
Nanoplankton 3.8–23 0.48–3.2 130–3,800 1.5–15.7
8.7–176 3,200–19,000 0.25–260 6.9–46 0.39–1.9 0.19–1.6
Lake Piaseczno, Polanda 822.5 (FW) 0.3(FW)
2,202.5 (FW)
Lake Husainsager, Indiab 11–15 (FW)
Microplankton 4.9–15 0.29–2.0 110–1,530 1.9–21.7
4.2–94 4,600–55,000 0.08–99 13–46 0.38–2.2 0.15–2.4
20 lakes in USAc 3.2±4.2 230±310 3.6±7.6
Macroplankton 1.7–4.1 0.07–0.45
60–234 0.44–2.1
0.69–7.6 430–3,000 0.16–8.1 2.2–22 1.5–2.6 0.051–0.60
Lakes close to Sudburyd 0.017–0.08
1.7–10
Lake Eriee 0.8–3.9
Lake Balaton, Hungaryf 84±6 0.1±0.4 287±23 0.74±0.15
Lochnagar Lake, Scotlandg 234.4±82
223
20 lakes in USAc 0.88±1.09
250±610 1.2±1.8
Lake Baikal, Russiah 12.6 <0.07 161 0.12
Shahpura Lake, Indiai 188 24 5,390
Macrophytes 0.12–13 0.02–0.82
32–320 0.56–9.6
0.27–6.4 390–8,700 <0.05–7.5 4.6–95 <0.02–0.3
0.05–3.6
Lake Provala, Yugoslaviaj 18–132 10.0 1,325–1,831.7
Taupo Volcanic Zonek 12–1,222
7.6–34 29–586 0.09–55 1.3–3,400 1,200–12,000 0.06–1.3 31–267
0.12–1.0 0.46–1.21
Armenia and India lakesystemsl
85–586 1.88–55.0
8.0–3,380 18,000–112,000
Plecoptera larvae 0.43–7.1
0.09–0.93
98–255 0.39–5.3
0.95–8.3 640–4,800 <0.07–2.4 7.0–31 0.54–2.8 0.39–0.65
Ginzan creek, Japanm 0.31–13 78–1,800 0.10–4.0
Odonata 0.87–1.5
0.08–0.24
38–98 0.35–1.8
0.43–4.3 1,000–9,300 0.26–0.64 9.0–23 0.53–0.80
0.35–0.84
Lakes in Utah, USAn 1.9–11
Trichoptera 0.90–6.6
0.19–0.60
150–420 3.2–12 2.5–10.4 3,500–13,900 0.69–3.6 9.3–18 0.4–1.6 0.35–0.91
Ginzan creek, Japanm 1.2–33 130–2,900 0.18–6.1
Ephemeroptera 0.67–6.7
0.18–1.2 85–510 1.3–10 0.92–32 800–8,700 0.16–1.4 4.0–48 0.76–3.2 0.16–1.16
Lakes close to a smelterd 0.01–0.08 4.4–12.2
Ginzan creek, Japanm 1.2–21 81–12,300 0.23–4.8
Coleoptera
Lakes close to smeltersd 0.033 7.4
Mussels
Lake Balaton, Hungaryo 7.1–375 69.7–2,000 0.02–3.6
Missouri River Watershedp 129–812 1.16–2.00
Hyalella sp. 1.6–5.0 0.17–0.93
35–160 0.26–1.6
0.51–3.5 200–1,100 0.066–1.7 6.2–24 0.10–1.9 0.18–0.82
Hyalella azteca, lakes closeto smeltersd
0.008–0.035
0.95–6.0
Whole fish (galaxids + troutjuveniles)
0.07–1.0
0.089–0.62
100–330 0.03–0.39
0.09–3.4 50–750 0.059–1.3 8.9–31 0.88–2.2 0.20–1.3
Perca flavescens, lakes closeto smeltersd
380–2,400 2.7–10.7
Water Air Soil Pollut (2010) 212:167–182 175

The gastropod and Hirudinea samples share negativecorrelations with K and positive correlations with Seand Zn. However, when examining PC3 scores(Fig. 3b), which correlates strongly with Br, (p value0.64), gastropods are separated from Hirudinea andOligochaeta, while the other macroinvertebrates retainsimilar associations with primary consumers. Hirudi-nea and Oligochaeta are both annelids, with Oligo-chaeta being detritivorous and Hirudinea acting asscavengers on other organisms. Interestingly, mostelements (except for Br, Co, and Fe) are similar orhigher in Oligochaeta than in Hirudinea, furthersupporting that detritus and biofilm are importantsources of elevated metals to the lower stages of thefood web. Smock (1983) also found that sediment-dependent organisms had the highest concentrationsof most elements.
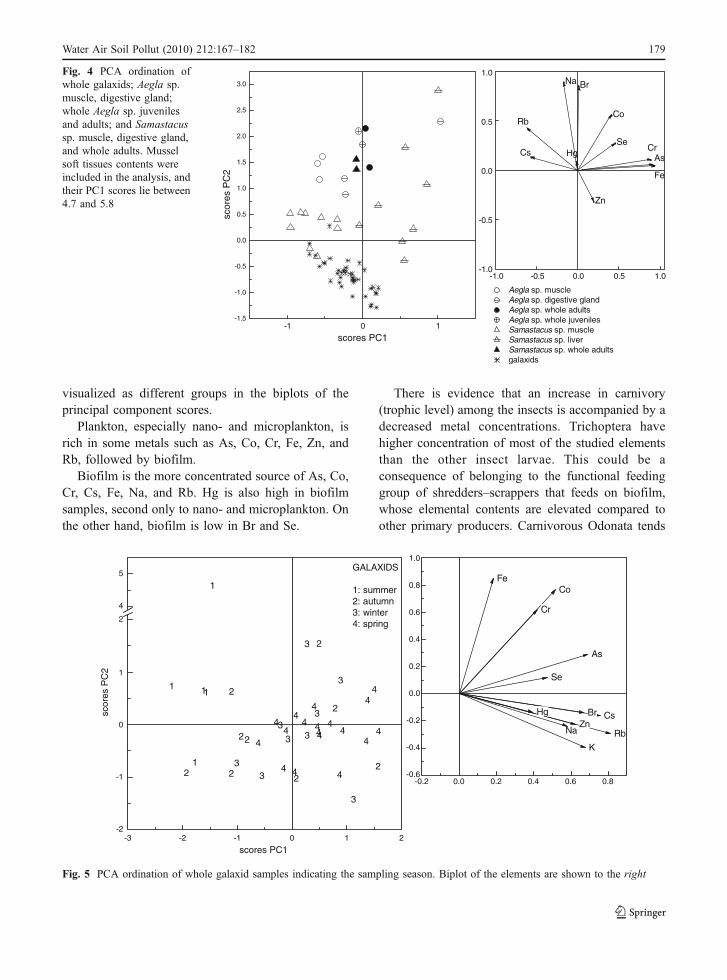
Comparing the PCA scores for similar size preys,whole decapods Samastacus and Aegla, Diplodon,
and whole galaxids (Fig. 4), it is evident that wholegalaxids cluster away from all decapods. The elemen-tal contents of whole adult Samastacus and Aegla aredistributed equally between muscle and hepatopan-creas, while all Aegla values cluster closely, regard-less of tissue type or life stage. Samastacus muscleand hepatopancreas samples spread over a widerange, and as expected, muscle samples of both Aeglaand Samastacus form two compact groups, duemainly to the low variability of contents amongsamples. Small galaxid samples are grouped next toSamastacus muscle samples, which tend to havelower contents of As, Cr, and Fe and larger amountsof Rb and Cs. Aegla muscle samples are separatedfrom Samastacus, with higher contents Br and Na.Mussel data have very high scores for PC1 (4.7 and5.8), due to the high Fe contents, which are ten timeshigher than any of the other crustaceans’ contents (thesamples were included in the analysis, although these
Table 5 (continued)
Reference As Cs Zn Co Cr Fe Hg Rb Se Na (%)
Freshwater fish, Great SaltLake Basins, USAq
0.31–1.2
77–696 0.7–2.9 0.04–0.19 2–5.1
Perch juveniles, Wisconsinlakes, USAr
0.13–0.92
Perch juveniles, N Fork,Alene river, USAs
89 0.21
Data in italics are from this work (overall range for all samples). All values in mg/kg, in dry weight basis, except where indicateda Radwan et al. (1990);b Prahalad and Seenayya (1989)c Chen et al. (2000)d Belzile et al. (2006)e Adams and Johnson (1977)f Balogh (1988)g Yang et al. (2002)h Ciesielski et al. (2006)i Shrivastava et al. (2003)j Stanković et al. (2000)k Robinson et al. (2006)l Vardanyan and Ingole (2006)mWatanabe et al. (2008)n Hillwalker et al. (2006)o Salánki et al. (1982)p Schmitt et al. (2007)qWaddell and Giddings (2004)rWatras et al. (1998)s Farag et al. (1998)
176 Water Air Soil Pollut (2010) 212:167–182
data are not shown in the graph to have a better detailof the low scores zone). The hepatopancreas of bothspecies spread over a wider range, which is expectedsince their composition depends on the more recentpreys, with higher contents of Co, Cr, As, and Fe.
Originally, the PC analysis of the galaxid samplesdid not show any grouping regarding length or site.However, when the sampling season was factored in,galaxids collected in summer tend to have lower As,Br, K, Cs, Rb, and Zn contents compared to thosecollected in other seasons (shown in Fig. 5). Thegalaxid diet consists of microcrustaceans (copepodsand cladocerans) as well as chironomid larvae andpupae. Two explanations can be proposed for thismarked trend for summer samples: (1) Galaxids may beswitching to low-metal dietary items during the muchmore productive summer season; (2) faster growth andturnover in the summer season may be leading to thebiodilution by growth of accumulated metals.
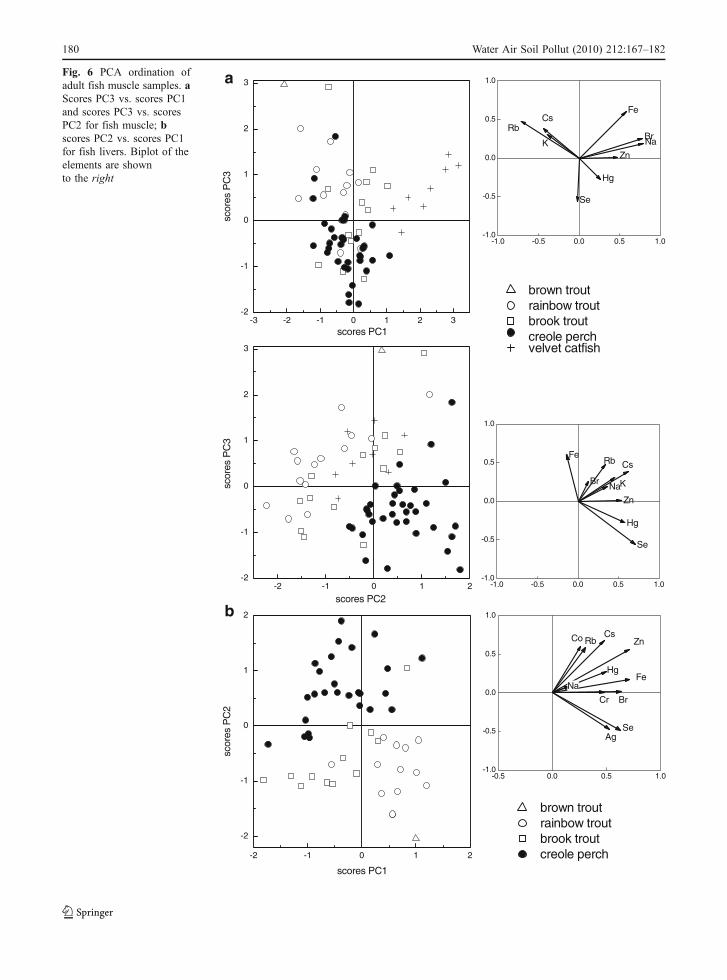
The large fish muscle samples show a compactarray in the PCA plots, with all species clusteringtogether, likely due to the narrower range of theelemental contents for fish compared to plankton,macrophytes, insects, and biofilm. The whole fishdata were also analyzed by PCA. Most elementalconcentrations are lower in fish muscle and higher infish liver samples compared to food sources, exceptfor Rb, Cs, and K (Tables 2, 3, and 4). The PCA
scores for muscle and liver samples are shown inFig. 6a, b, respectively. Figure 6a indicates that velvetcatfish muscle samples are characterized by high Br,Na, and Fe compared to salmonids and creole perch,while creole perch muscle samples are characterizedby high Se and low Fe concentrations. Brook troutand rainbow trout muscle metal trends are quitesimilar, but when liver tissue is analyzed, brook troutliver contents are more similar to those of creoleperch, especially regarding non-essential Ag andessential Se (see Fig. 6b).
Some authors had found that the Rb to Cs contentratios were indicators of fish diet (Kanevskii andFleishman 1972; Chiasson 1991). Although stronglimitations of the use of Rb–Cs ratios as indicator offish diet in local fish were found (Ribeiro Guevara etal. 2006), in this work, the Rb–Cs ratios of eachgroup of items were computed by using linearregression. The Rb vs. Cs correlation is significantfor each of the food items. The Rb–Cs ratiosmeasured for each group are as follows: macrophytes,44; nanoplankton, 11; microplankton, 15; macro-plankton, 56; biofilm, ten; insect larvae, 24; non-insectmacroinvertebrates, 25; galaxiids, 26; salmonids andperch, 14. However, the ratios show no trend regardingthe trophic level, and while Rb and Cs had differingtrends in the food items ordering, Rb and Cs are stronglyrelated to K in fish muscle. Therefore, since Rb and Cs
-1 0 1 2 3 4-4
-3
-2
-1
0
1
2
As
Ba
Br
Co
Cr
CsFe
Hg
NaRb
Se
Zn
-1.0 -0.5 0.0 0.5 1.0-1.0
-0.5
0.0
0.5
1.0
K
nanoplankton, 10-53 µmmicroplankton, 53-200 µmmacroplankton, >200 µmperiphytonsubmerged macrophytes mossses Trichoptera sp. Odonata sp. Plecoptera sp.Ephemeroptera sp. Coleoptera sp.
scor
es P
C2
scores PC1
Fig. 2 PCA ordination of nano-, micro-, and macroplankton; submerged macrophytes; submerged mosses; biofilm; and insect larvae fromLake Moreno and biplot of the elements
Water Air Soil Pollut (2010) 212:167–182 177
are correlated to K in fish muscle, the intake is probablyrelated to the chemical similarities of these elements toK, and the amount of Rb and Cs to be incorporateddepends on the availability of these elements in theenvironment.
4 Conclusions
Except for nano -and microplankton, which tend togroup together in the PC analyses, all items showdistinctive compositional patterns, which can be
-1 0 1 2 3 4-4
-3
-2
-1
0
1
2
As
Br
Cr
Cs
Hg
NaRb
SeZn
nanoplankton microplankton macroplankton submerged macrophytes and mosses periphyton gasteropods (without shell) Hirudinea Amphipods Oligochaeta
scor
es P
C2
scores PC1
-1.0 -0.5 0.0 0.5 1.0-1.0
-0.5
0.0
0.5
1.0
FeCo
K
-1 0 1 2 3 4-3
-2
-1
0
1
2
scor
es P
C3
scores PC1
nanoplankton microplankton macroplankton submerged macrophytes and mosses periphyton gasteropods Hirudinea Amphipods Oligochaeta
-1.0 -0.5 0.0 0.5 1.0-1.0
-0.5
0.0
0.5
1.0
Cr
AsNa
CsRb
ZnHg
Se
Br
CoFe
K
a
b
Fig. 3 PCA ordination of Hyalella sp.; Hirudinea; gastropods;Oligochaeta; nano-, micro-, and macroplankton; submergedmacrophytes and mosses; and biofilm from Lake Moreno. a
PC2 scores vs. PC1, b PC3 scores vs. PC1. Biplot of theelements are shown to the right
178 Water Air Soil Pollut (2010) 212:167–182
visualized as different groups in the biplots of theprincipal component scores.
Plankton, especially nano- and microplankton, isrich in some metals such as As, Co, Cr, Fe, Zn, andRb, followed by biofilm.
Biofilm is the more concentrated source of As, Co,Cr, Cs, Fe, Na, and Rb. Hg is also high in biofilmsamples, second only to nano- and microplankton. Onthe other hand, biofilm is low in Br and Se.
There is evidence that an increase in carnivory(trophic level) among the insects is accompanied by adecreased metal concentrations. Trichoptera havehigher concentration of most of the studied elementsthan the other insect larvae. This could be aconsequence of belonging to the functional feedinggroup of shredders–scrappers that feeds on biofilm,whose elemental contents are elevated compared toother primary producers. Carnivorous Odonata tends
-3 -2 -1 0 1 2-2
-1
0
1
2
4
5
As
Br
Co
Cr
Cs
Fe
Hg
K
Rb
Se
Zn
2
3
1
44
4
4
2
24
3
2
44 4 4
4
4
4
4
32
3
1
21
4 4
4
3
2
1
3
2
3
43
42
1
3
-0.2 0.0 0.2 0.4 0.6 0.8-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
Na
scor
es P
C2
scores PC1
GALAXIDS
1: summer2: autumn3: winter4: spring
Fig. 5 PCA ordination of whole galaxid samples indicating the sampling season. Biplot of the elements are shown to the right
-1.0 -0.5 0.0 0.5 1.0-1.0
-0.5
0.0
0.5
1.0
-1 0 1-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0 Br
Co
Na
Se
Zn
CrHg
Rb
Cs
Fe
As
scor
es P
C2
scores PC1
Aegla sp. muscle Aegla sp. digestive gland Aegla sp. whole adults Aegla sp. whole juveniles Samastacus sp. muscle Samastacus sp. liver Samastacus sp. whole adults galaxids
Fig. 4 PCA ordination ofwhole galaxids; Aegla sp.muscle, digestive gland;whole Aegla sp. juvenilesand adults; and Samastacussp. muscle, digestive gland,and whole adults. Musselsoft tissues contents wereincluded in the analysis, andtheir PC1 scores lie between4.7 and 5.8
Water Air Soil Pollut (2010) 212:167–182 179
-2 -1 0 1 2-2
-1
0
1
2
3
-3 -2 -1 0 1 2 3-2
-1
0
1
2
3
Br
Fe
Na
Se
Zn
Hg
Br
Fe
Na
Se
Zn
Hg
scor
es P
C3
scores PC2
-1.0 -0.5 0.0 0.5 1.0-1.0
-0.5
0.0
0.5
1.0
CsRb
K
scor
es P
C3
scores PC1
brown trout rainbow trout brook trout creole perch velvet catfish
-1.0 -0.5 0.0 0.5 1.0-1.0
-0.5
0.0
0.5
1.0
RbCs
K
-2 -1 0 1 2
-2
-1
0
1
2
-0.5 0.0 0.5 1.0-1.0
-0.5
0.0
0.5
1.0
AgSe
BrCr
FeHg
Na
ZnCsCo Rb
scor
es P
C2
scores PC1
brown trout rainbow trout brook trout creole perch
b
aFig. 6 PCA ordination ofadult fish muscle samples. aScores PC3 vs. scores PC1and scores PC3 vs. scoresPC2 for fish muscle; bscores PC2 vs. scores PC1for fish livers. Biplot of theelements are shownto the right
180 Water Air Soil Pollut (2010) 212:167–182

to have lower elemental contents. Fish muscle, whichare the top predators of the studied species, have thelowest concentrations of most metals.
Aegla sp. soft tissues are richer in trace elementsthan Samastacus. Mussels have the highest contentsof iron, above the other items by a factor of 10.
Acknowledgments We acknowledge the assistance of Mr.Ricardo Sánchez in all the field work, Dr. M. Diéguez and Dr.C. Queimaliños for their help with the plankton samplers, Dr.R. Daga for the site characterization, Mr. J. Pérez duringplankton sampling, and the RA-6 reactor staff for the irradiationof the samples. This work was partially funded by projectsPICT2005 33838 and PICT2006 1051 of the ANPCyT(Agencia Nacional de Promoción Científica y Técnológica) ofArgentina.
References
Adams, W. J., & Johnson, H. E. (1977). Survey of the seleniumcontent in the aquatic biota of western Lake Erie. Journalof Great Lakes Research, 3, 10–14.
Arribére, M. A., Ribeiro Guevara, S., Bubach, D. F., &Vigliano, P. H. (2006). Trace elements as fingerprint oflake of provenance and of species of some native andexotic fish of northern patagonian lakes. Biological TraceElement Research, 110, 71–95.
Arribére, M. A., Ribeiro Guevara, S., Bubach, D. F., Arcagni,M., & Vigliano, P. H. (2008). Selenium and mercury innative and introduced fish species of patagonian lakes,Argentina. Biological Trace Element Research, 122, 43–63.
Balogh, K. V. (1988). Comparison of mussels and crustaceanplankton to monitor heavy metal pollution. Water, Air, andSoil Pollution, 37, 281–292.
Belzile, N., Chen, Y. W., Gunn, J. M., Tong, J., Alarie, Y.,Delonchamp, T., et al. (2006). The effect of selenium onmercury assimilation by freshwater organisms. CanadianJournal of Fisheries and Aquatic Science, 63, 1–10.
Campbell, L.M., Arribére, M.A., Klassen, K., Kyser, K.,Ribeiro Guevara, S., Bubach, D.F. (2005). Metal biogeo-chemistry and trophodynamics in Patagonia lakes ofNahuel Huapi National Park, Argentina, Proc. VII Meet-ing of the Society of Environmental Toxicology andChemistry (SETAC) América Latina, Santiago, Chile.
Campbell, L. M., Kyser, K., Klassen, K., Arribére, M. A.,Revenga, J., Daga, R. B., Ribeiro Guevara, S. (2007).Linking volcanoes and fish: metal biomagnification inPatagonia Andean aquatic food webs. Proc. 34th AquaticToxicology Workshop, Halifax, Nova Scotia, Canada.
Chen, C. Y., Stemberger, R. S., Klaue, B., Blum, J. D.,Pickhardt, P. C., & Folt, L. C. (2000). Accumulation ofheavy metals in food web components across a gradient oflakes. Limnology and Oceanography, 45(7), 1525–1536.
Chiasson, A. G. (1991). The ratio of rubidium to caesium inthreespine stickleback (Gasterosteus aculeatus), benthicand limnetic ticklebacks (Gasterosteus), and juvenile
sockeye salmon (Oncorhynchus nerka). Chemistry andEcology, 5, 227–240.
Ciesielski, T., Pastukhov, M. V., Fodor, P., Bertenyi, Z.,Namiesnik, J., & Szefer, P. (2006). Relationships andbioaccumulation of chemical elements in the Baikal seal(Phoca sibirica). Environmental Pollution, 139, 372–384.
Farag, A. M., Woodward, D. F., Goldstein, J. N., Brumbaugh,W., & Meyer, J. S. (1998). Concentrations of metalsassociated with mining waste in sediments, biofilm,benthic macroinvertebrates and fish form from Coeurd’Alene river basin, Idaho. Archives of EnvironmentalContamination and Toxicology, 34(1), 19–27.
Hillwalker, W. E., Jepson, P. C., & Anderson, K. A. (2006).Selenium accumulation patterns in lotic and lentic aquaticsystems. Science of the Total Environment, 366, 367–379.
Kanevskii, Y. P., & Fleishman, D. G. (1972). Investigation offood chains in an ichtyocoenosis of lake Dal’nyi (alsospelt Dalnee) (Kamchatka) according to the concentrationsof rubidium and cesium in hydrobionts. Academy ofScience of the U.S.S.R. Translated from Ékologiya 1971.Soviet Journal of Ecology, 3(5–8), 191–193.
Lara, G. P., & Moreno, C. A. (1995). Efectos de la depredaciónde Aegla abtao (Crustacea, Aeglidae) sobre la distribuciónespacial y abundancia de Diplodon chilensis (Bivalvia,Hyriidae) en el Lago Panguipulli, Chile. Revista Chilenade Historia Natural, 68, 123–129.
Prahalad, A. K., & Seenayya, G. (1989). Physico-chemicalinteractions and bioconcentration of zinc and lead in theindustrially polluted Lake Husainsager, Hyderabad, India.Environmental Pollution, 58A, 139–154.
Queimaliños, C. P., Modenutti, B. E., & Balseiro, E. G. (1999).Symbiotic association of the ciliate Ophryidium naumanniwith Chlorella causing a deep chlorophyll α maximum inan oligotrophic South Andes lake. Journal of PlanktonResearch, 21, 167–178.
Quirós, R. (1988). Relationships between air temperature,depth, nutrients and chlorophyll in 103 Argentinian lakes.Verhandlungen der Internationalen Vereinigung für Lim-nologie, 23, 647–658.
Radwan, S., Kowalik, W., & Kowalczyk, C. (1990). Occur-rence of heavy metals in water, phytoplankton andzooplankton of a mesotrophic lake in Eastern Poland.The Science of the Total Environment, 96, 115–120.
Ribeiro Guevara, S., Bubach, D. F., Vigliano, P. H., Lippolt, G.,& Arribére, M. A. (2004). Heavy metals and other traceelements in native mussel Diplodon chilensis fromNorthern Patagonian lakes, Argentina. Biological TraceElement Research, 102(1–3), 245–263.
Ribeiro Guevara, S., Arribére, M. A., Bubach, D. F., Vigliano,P. H., Rizzo, A. P., Alonso, M., et al. (2005). Silvercontamination on abiotic and biotic compartments of lakeNahuel Huapi National Park lakes, Patagonia, Argentina.Science of the Total Environment, 336(1–3), 119–134.
Ribeiro Guevara, S., Bubach, D. F., Macchi, P. J., Vigliano, P.H., Arribére, M. A., & Colombo, J. C. (2006). Rb–Cs ratioas an indicator of fish diet in lakes of the Patagonia,Argentina. Biological Trace Element Research, 110, 97–119.
Robinson, B., Kim, N., Marchetti, M., Moni, C., Schroeter, L.,van den Dijssel, C., et al. (2006). Arsenic hyperaccumu-lation by aquatic macrophytes in the Taupo Volcanic Zone,
Water Air Soil Pollut (2010) 212:167–182 181

New Zealand. Environmental and Experimental Botany,58(1–3), 206–215.
Salánki, J., Balogh, K. V., & Berta, E. (1982). Heavy metals inanimals of Lake Balaton. Water Research, 16, 1147–1152.
Schmitt, C. J., Brumbaugh, W. G., Besser, J. M., May, T. W.(2007). Concentrations of metals in aquatic invertebratesfrom the Ozark National Scenic Riverways, Missouri.USGSS Open-File Report 2007–1435.
Shrivastava, P., Saxena, A., & Swarup, A. (2003). Heavy metalpollution in a sewage-fed lake of Bhopal, (M. P.) India.Lakes & Reservoirs: Research and Management, 8, 1–4.
Smock, L. A. (1983). The influence of feeding habits on whole-body metal concentrations in aquatic insects. FreshwaterBiology, 13, 301–311.
Souza, M. S., Modenutti, B. E., & Balseiro, E. G. (2007).Antioxidant defenses in planktonic crustaceans exposed todifferent underwater light irradiances in Andean lakes.Water, Air, and Soil Pollution, 183, 49–57.
Stanković, Ž., Pajević, S., Vučković, M., & Stojanović, S.(2000). Concentrations of trace metals in dominant aquaticplants of the Lake Provala (Vojvodina, Yugoslavia).Biologia Plantarum, 43(4), 583–585.
Stern, C. (2004). Active Andean volcanism: its geologic andtectonic setting. Revista Geológica de Chile, 31(2), 161–206.
Vardanyan, L. G., & Ingole, B. S. (2006). Studies on heavymetal accumulation in aquatic macrophytes from Sevan(Armenia) and Carambolim (India) lake systems. Environ-mental International, 32(2), 208–218.
Waddell, K.M., Giddings, E.M. (2004). Trace elements andorganic compounds in sediment and fish tissue from theGreat Salt Lake basins, Utah, Idaho, and Wyoming, 1998–99. U.S. GEOLOGICAL SURVEY Water-ResourcesInvestigations Report 03-4283. Salt Lake City, Utah, USA.
Watanabe, K., Monaghan, M. T., Takemon, Y., & Omura, T.(2008). Biodilution of heavy metals in a stream macro-invertebrate food web: Evidence from stable isotopeanalysis. Science of the Total Environment, 394, 57–67.
Watras, C. J., Back, R. C., Halvorsen, S., Hudson, R. J. M.,Morrison, K. A., & Wente, S. P. (1998). Bioaccumulationof mercury in pelagic freshwater food webs. Science of theTotal Environment, 219, 183–208.
Westfall, M. J., & Tennesseen, K. J. (1996). Odonata. In R. W.Merritt & K. W. Cummins (Eds.), An introduction to theaquatic insects of North America (pp. 164–211). Dubuque:Kendall/Hunt.
Yang, H., Rose, N. L., & Battarbee, R. W. (2002). Distributionof some trace metals in Lochnagar, a Scottish mountainlake ecosystem and its catchment. Science of the TotalEnvironment, 285, 197–208.
182 Water Air Soil Pollut (2010) 212:167–182
Related Documents











