Toxoplasma gondii: Uptake and survival of oocysts in free-living amoebae Jadwiga Winiecka-Krusnell a, * , Isabel Dellacasa-Lindberg a,b , J.P. Dubey c , Antonio Barragan a,b a Department of Parasitology, Mycology and Environmental Microbiology, Swedish Institute for Infectious Disease Control, 171 82 Solna, Sweden b Center for Infectious Medicine, Department of Medicine, Karolinska Institutet, 141 86 Stockholm, Sweden c United States Department of Agriculture, Agricultural Research Service, Animal and Natural Resources Institute, Animal Parasitic Diseases Laboratory, Building 1001, Beltsville, MD 20705-2350, USA article info Article history: Received 11 December 2007 Received in revised form 23 July 2008 Accepted 30 September 2008 Available online 18 October 2008 Keywords: Protozoa Apicomplexa Toxoplasma gondii Acanthamoeba spp. Free-living amoebae Waterborne infections Transmission of Toxoplasma infection abstract Waterborne transmission of the oocyst stage of Toxoplasma gondii can cause outbreaks of clinical toxo- plasmosis in humans and infection of marine mammals. In water-related environments and soil, free-liv- ing amoebae are considered potential carriers of various pathogens, but knowledge on interactions with parasitic protozoa remains elusive. In the present study, we assessed whether the free-living Acantha- moeba castellanii, due to its phagocytic activity, can interact with T. gondii oocysts. We report that amoe- bae can internalize T. gondii oocysts by active uptake. Intracellular oocysts in amoebae rarely underwent phagocytic lysis, retained viability and established infection in mice. Interaction of T. gondii with amoebae did not reduce the infectivity and pathogenicity of oocysts even after prolonged co-cultivation. Our results show that uptake of oocysts by A. castellanii does not restrain the transmission of T. gondii in a murine infection model. Ó 2008 Elsevier Inc. All rights reserved. 1. Introduction Toxoplasmosis is a prevalent zoonotic infection in humans and warm-blooded animals. Domestic and wild felids are definitive hosts and shed oocysts in their feces. Mammals and birds serve as intermediate hosts developing tissue cysts in their organs. Infec- tion occurs after ingestion of tissue cysts or contact with soil or water contaminated with oocysts. Oral infections with tissue cysts and oocysts cannot be distinguished and therefore the impact of either stage on the epidemiology of toxoplasmosis remains unde- termined (Dubey, 2004). Several waterborne outbreaks of toxoplasmosis have been doc- umented (Bahia-Oliveira et al., 2003; Benenson et al., 1982; Bowie et al., 1997; de Moura et al., 2006) and in one instance viable Tox- oplasma gondii oocysts were found in drinking water (de Moura et al., 2006). Infections with T. gondii also occur in marine mam- mals, and can be lethal for sea otters and seals (Dubey et al., 2003; Miller et al., 2002). The transmission route in the marine environment remains unknown. Occasional ingestion of oocysts from seawater or ingestion of infected animals has been consid- ered (Conrad et al., 2005). There is no known cold-blooded inter- mediate host for T. gondii. However, mollusks are able to accumulate oocysts in their bodies due to their filter-feeding activ- ity and are thus a potential source of infection in aquatic environ- ments (Arkush et al., 2003; Lindsay et al., 2001). Acanthamoeba spp. represent free-living amoebae and are com- monly present in water-related environments including fresh-, sea-, and brackish water. Evolved along a line of phagocytic cells, Acanthamoeba spp. can be compared to non-professional macro- phages. They participate in nutrient and energy turnover in nature, essentially preying on bacteria in biofilms (Rodriguez-Zaragoza, 1994). The features of the lysosomal system of acanthamoebae ap- pear to be typical of these found in other phagocytic cells. Ingested particles are surrounded by plasma membrane to form phago- somes that subsequently acquire hydrolytic enzymes by fusion with lysosomes (Korn and Weisman, 1967; Wetzel and Korn, 1969). Some of the engulfed microorganisms, however, can avoid destruction and survive inside amoebae. Recently, Acanthamoeba spp. have been recognized as potential carriers of several patho- genic bacteria, yeasts and viruses (Greub and Raoult, 2004; Steen- bergen et al., 2004). It has been postulated that amoebae not only act as a vehicle enabling spread of bacteria in the environment but also influence bacterial virulence and resistance to drugs and disin- fectants (Cirillo et al., 1994, 1997; Miltner and Bermudez, 2000; Steenbergen et al., 2001; Storey et al., 2004). The knowledge on possible interactions between free-living amoebae and other pro- tozoan organisms is rather unclear. A microsporidia-like natural endosymbiont in Vannella spp. isolated from tap water and from inflamed human cornea has been reported (Hoffmann et al., 1998; Michel et al., 2000). Recent reports describe in vitro observa- 0014-4894/$ - see front matter Ó 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.exppara.2008.09.022 * Corresponding author. Fax: +46 08 310525. E-mail address: [email protected] (J. Winiecka-Krusnell). Experimental Parasitology 121 (2009) 124–131 Contents lists available at ScienceDirect Experimental Parasitology journal homepage: www.elsevier.com/locate/yexpr

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Experimental Parasitology 121 (2009) 124–131
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/ locate /yexpr
Toxoplasma gondii: Uptake and survival of oocysts in free-living amoebae
Jadwiga Winiecka-Krusnell a,*, Isabel Dellacasa-Lindberg a,b, J.P. Dubey c, Antonio Barragan a,b
a Department of Parasitology, Mycology and Environmental Microbiology, Swedish Institute for Infectious Disease Control, 171 82 Solna, Swedenb Center for Infectious Medicine, Department of Medicine, Karolinska Institutet, 141 86 Stockholm, Swedenc United States Department of Agriculture, Agricultural Research Service, Animal and Natural Resources Institute, Animal Parasitic Diseases Laboratory, Building 1001,Beltsville, MD 20705-2350, USA
a r t i c l e i n f o
Article history:Received 11 December 2007Received in revised form 23 July 2008Accepted 30 September 2008Available online 18 October 2008
Keywords:ProtozoaApicomplexaToxoplasma gondiiAcanthamoeba spp.Free-living amoebaeWaterborne infectionsTransmission of Toxoplasma infection
0014-4894/$ - see front matter � 2008 Elsevier Inc. Adoi:10.1016/j.exppara.2008.09.022
* Corresponding author. Fax: +46 08 310525.E-mail address: [email protected]
a b s t r a c t
Waterborne transmission of the oocyst stage of Toxoplasma gondii can cause outbreaks of clinical toxo-plasmosis in humans and infection of marine mammals. In water-related environments and soil, free-liv-ing amoebae are considered potential carriers of various pathogens, but knowledge on interactions withparasitic protozoa remains elusive. In the present study, we assessed whether the free-living Acantha-moeba castellanii, due to its phagocytic activity, can interact with T. gondii oocysts. We report that amoe-bae can internalize T. gondii oocysts by active uptake. Intracellular oocysts in amoebae rarely underwentphagocytic lysis, retained viability and established infection in mice. Interaction of T. gondii with amoebaedid not reduce the infectivity and pathogenicity of oocysts even after prolonged co-cultivation. Ourresults show that uptake of oocysts by A. castellanii does not restrain the transmission of T. gondii in amurine infection model.
� 2008 Elsevier Inc. All rights reserved.
1. Introduction
Toxoplasmosis is a prevalent zoonotic infection in humans andwarm-blooded animals. Domestic and wild felids are definitivehosts and shed oocysts in their feces. Mammals and birds serveas intermediate hosts developing tissue cysts in their organs. Infec-tion occurs after ingestion of tissue cysts or contact with soil orwater contaminated with oocysts. Oral infections with tissue cystsand oocysts cannot be distinguished and therefore the impact ofeither stage on the epidemiology of toxoplasmosis remains unde-termined (Dubey, 2004).
Several waterborne outbreaks of toxoplasmosis have been doc-umented (Bahia-Oliveira et al., 2003; Benenson et al., 1982; Bowieet al., 1997; de Moura et al., 2006) and in one instance viable Tox-oplasma gondii oocysts were found in drinking water (de Mouraet al., 2006). Infections with T. gondii also occur in marine mam-mals, and can be lethal for sea otters and seals (Dubey et al.,2003; Miller et al., 2002). The transmission route in the marineenvironment remains unknown. Occasional ingestion of oocystsfrom seawater or ingestion of infected animals has been consid-ered (Conrad et al., 2005). There is no known cold-blooded inter-mediate host for T. gondii. However, mollusks are able toaccumulate oocysts in their bodies due to their filter-feeding activ-
ll rights reserved.
(J. Winiecka-Krusnell).
ity and are thus a potential source of infection in aquatic environ-ments (Arkush et al., 2003; Lindsay et al., 2001).
Acanthamoeba spp. represent free-living amoebae and are com-monly present in water-related environments including fresh-,sea-, and brackish water. Evolved along a line of phagocytic cells,Acanthamoeba spp. can be compared to non-professional macro-phages. They participate in nutrient and energy turnover in nature,essentially preying on bacteria in biofilms (Rodriguez-Zaragoza,1994). The features of the lysosomal system of acanthamoebae ap-pear to be typical of these found in other phagocytic cells. Ingestedparticles are surrounded by plasma membrane to form phago-somes that subsequently acquire hydrolytic enzymes by fusionwith lysosomes (Korn and Weisman, 1967; Wetzel and Korn,1969). Some of the engulfed microorganisms, however, can avoiddestruction and survive inside amoebae. Recently, Acanthamoebaspp. have been recognized as potential carriers of several patho-genic bacteria, yeasts and viruses (Greub and Raoult, 2004; Steen-bergen et al., 2004). It has been postulated that amoebae not onlyact as a vehicle enabling spread of bacteria in the environment butalso influence bacterial virulence and resistance to drugs and disin-fectants (Cirillo et al., 1994, 1997; Miltner and Bermudez, 2000;Steenbergen et al., 2001; Storey et al., 2004). The knowledge onpossible interactions between free-living amoebae and other pro-tozoan organisms is rather unclear. A microsporidia-like naturalendosymbiont in Vannella spp. isolated from tap water and frominflamed human cornea has been reported (Hoffmann et al.,1998; Michel et al., 2000). Recent reports describe in vitro observa-

J. Winiecka-Krusnell et al. / Experimental Parasitology 121 (2009) 124–131 125
tions on phagocytosis of oocysts of Cryptosporidium parvum byAcanthamoeba spp. (Gomez-Couso et al., 2006; Stott et al., 2003).
We have tested the hypothesis that free-living amoebae caninteract with oocysts of T. gondii and participate in the transmis-sion of the infection. We have assessed the uptake of T. gondii oo-cysts by trophozoites of Acanthamoeba castellanii and the ability ofintra-amoebic oocysts to establish infection in laboratory animals.
2. Materials and methods
2.1. Parasites and amoebae
Oocysts were obtained from the feces of a cat infected with theVEG strain of T. gondii as described (Dubey, 1995; Dubey et al.,1996). VEG is a genetic Type III strain and was isolated from bloodof an AIDS patient (Dubey et al., 1996). Oocysts were concentratedand separated from cat feces by sugar flotation. After sporulation in2% sulfuric acid at 22 �C for one week oocysts were stored in 2%sulfuric acid at 4 �C until use (Dubey, 1995). Purification of oocystsfrom stool was performed using saturated NaCl solution. Stool sus-pension was washed with phosphate buffer salt (PBS), pH 7 untilclear supernatant was obtained (5 � 5 min, 1000g). Sediment wassuspended in 10 ml of saturated NaCl, well-mixed and centrifugedfor 3 min at 1000g. Following the centrifugation, 5 ml of dH2O con-taining 0.2% Tween 20 was applied on the surface of the tube.Water layer was carefully agitated with the pipette and transferredto collection tube. NaCl layer was mixed and centrifuged again3 min at 1000g. Purification was repeated 3 times. Collected waterlayers were centrifuged for 10 min at 1000g and sediment waswashed twice with water/Tween 20.
Acanthamoeba castellanii trophozoites were cultivated axeni-cally in peptone–yeast extract–glucose medium (PYG) medium(ATCC 712) at 30 �C.
Leishmania tropica promastigotes were maintained in RPMImedium supplemented with 10% FCS, 2 mM L-glutamine and50 lg/ml of gentamicin (all reagents from Sigma, St. Louis, MO)at 23 �C.
2.2. Uptake of T. gondii oocysts by amoebae
To determine the uptake of T. gondii oocysts by A. castellanii tro-phozoites, a suspension containing 92% of sporulated oocysts wasadded to amoebae monolayers at the ratio 5:1. Cells were incu-bated at 30 �C in PYG medium containing 50 lg/ml of gentamicin.Autofluorescent oocysts were visualized in UV light (Lindquistet al., 2003). The number of extracellular oocysts and amoebaecontaining intracellular oocysts was assessed at indicated timeintervals in phase contrast and UV light (340–380 nm) using a Lei-ca DRMB epifluorescence microscope. Cultures were incubated onice for 20 min to detach cells from the growth surface and 500 ll ofcell suspension was removed and centrifuged to assess the numberof free oocysts. The number of amoebae containing oocysts wasevaluated by counting 103 amoebae trophozoites. Latex-coatedpolystyrene beads, 10 lm in diameter, (Sigma, Cat. No. 55463,Macro beads), were added to amoebae at the same ratio as oocystsand incubated for indicated time intervals. To assess phagocyticactivity of A. castellanii, L. tropica promastigotes were added to tro-phozoites at the ratio 10:1 and incubated for the same time inter-vals as amoebae-oocysts co-cultures.
Previous to inoculation in mice, amoebae-oocyst co-cultureswere washed with agitation 5 times for 10 min with warm PBSto remove extracellular oocysts that are normally non-adhesive.Monolayers were then put on ice for 20 min, detached cells weresedimented by centrifugation for 5 min at 1000g and suspendedin 5 ml of PBS. The suspension of 500 ll was centrifuged and the
number of intracellular oocysts was determined by epifluorescencemicroscopy. When indicated, a suspension containing A. castellaniiand oocysts mixed at ratio 5:1 was spread on the surface of non-nutrient agar dishes (1.5% agar in distilled water), covered withthe suspension of heat inactivated Escherichia coli, K 12 and incu-bated at 30 �C in a moist chamber.
2.3. Uptake inhibition assay
Amoebae monolayers were incubated for 2 h with 1, 10 or100 mM solutions of a-methyl-D-mannoside, D(+)mannose,D(+)galactose, D-fructose-6-phosphate, D(+)galactosamine, N-acet-yl-D-galactosamine or cytochalasin D, 10 lg/ml (all reagentsfrom Sigma), dissolved in cultivation medium, prior to additionof oocysts or latex-coated polystyrene beads. After 16 h of co-cultivation the frequency of oocysts or beads contained inamoebae was related to frequencies in non-treated cultures,respectively.
2.4. Lysosomal staining
Labeling of lysosomal vacuoles of A. castellanii was per-formed using LysoTrackerTM probe (Molecular Probes, Eugene,OR). Amoebae monolayers growing in duplicates on chamberslides (NUNC) were incubated with oocysts for 30 min, 1, 5,24, 48, 72, and 144 h washed 5 times with PBS, incubated with25 lM solution of LysoTrackerTM in PYG medium for 2 h,mounted in PBS and assessed by counting 2 � 102 endocyticvacuoles at each time point using epifluorescence microscopy(Leica, DMRB).
2.5. Live microscopy
Acanthamoeba castellanii and oocyst co-cultures containing in-tra-amoebic oocysts were visualized by time-lapse epifluorescencemicroscopy using Leica, DMRB microscope equipped with AxioCamcamera (Carl Zeiss. Oberkochen, Germany) and imaging softwaresOpenlab 4.0.2 (Improvision, Coventry, UK) and Photoshop 6(Adobe, San Jose, CA).
2.6. Infection of mice
Six weeks old male BALB/c mice were infected by oral gavagewith 100 ll of PBS suspension containing indicated numbers ofintra-amoebic oocysts obtained after 48 h, 7 or 14 d of incuba-tion with amoebae, free oocysts or A. castellanii trophozoites.Mice were euthanized 9 or 20 d.p.i. and internal organs werecollected. Blood samples were taken the first and last day ofthe experiment to assess seroconversion of infected animals. Eth-ical approval was granted by the ethical committee at KarolinskaInstitutet.
2.7. Plaque formation assays
Human foreskin fibroblasts (HFF) cells were cultivated on 12-well culture plates in DMEM medium containing 1mM Hepes,10% FCS, 2 mM L-glutamine and 80 lg/ml of gentamicin (all re-agents from Sigma). Murine organs were homogenized in PBS bypressing through 40 lm cell strainers (BD Falcon, Bedford, MA).Cell suspension was washed in PBS and sediment suspended in1 ml of complete DMEM medium and divided into two parts.One aliquot was frozen for further extraction of DNA. The otherwas diluted 1:10 in complete DMEM medium and 100 ll/wellwas added to HFF cells. Plates were incubated at 37 �C and 5%CO2 and ocular plaque counting was performed on days 5, 7,and 10.
126 J. Winiecka-Krusnell et al. / Experimental Parasitology 121 (2009) 124–131
2.8. Indirect immunofluorescence (IF) test
Freshly egressed tachyzoites of CTG strain of T. gondii (Pfef-ferkorn and Pfefferkorn, 1980) were filtered through 3 lm Iso-pore filters (Millipore, Billerica, MA, USA) and washed 3 timeswith PBS. Acanthamoeba castellanii trophozoites cultivated inPYG medium for 48 h, were detached from the growth surfaceby incubation on ice for 20 min, pelleted by centrifugation for5 min at 1000g and washed 3 times with PBS. Toxoplasma gondiiand A. castellanii cell suspensions were then distributed on 8-well teflon-coated slides (Erie Scientific Company, Portsmouth,NH) air dried, fixed in cold acetone for 20 min and kept frozenuntil used. Mouse serum samples were diluted in PBS, addedto amoebae and T. gondii-coated slides and incubated at roomtemperature for 30 min in a humid chamber. After 3 washes inPBS, FITC-conjugated rabbit anti mouse Ig (Sigma) was addedto wells and incubated for 30 min at room temperature. Follow-ing 3 consecutive washes in PBS, slides were mounted in non-fading VectashieldTM medium (Vector Laboratories Inc., Burlin-game, CA) and assessed by epifluorescence microscopy (LeicaDMRB).
2.9. PCR analysis
Extraction of DNA from organs was performed using QIAmpDNA Mini Kit (Qiagen Inc. Valencia, CA, USA) according tomanufacturer’s protocol. Toxoplasma gondii DNA was detectedin extracted material using nested PCR amplification of SAG3genetic marker as described earlier (Grigg et al., 2001). Sam-ples that gave negative results in SAG3 PCR were retestedusing a second PCR test based on the amplification of B1 gene(Burg et al., 1989). DNA extracted from tachyzoites of CTGstrain and DNA from organs of non-infected mice served ascontrols.
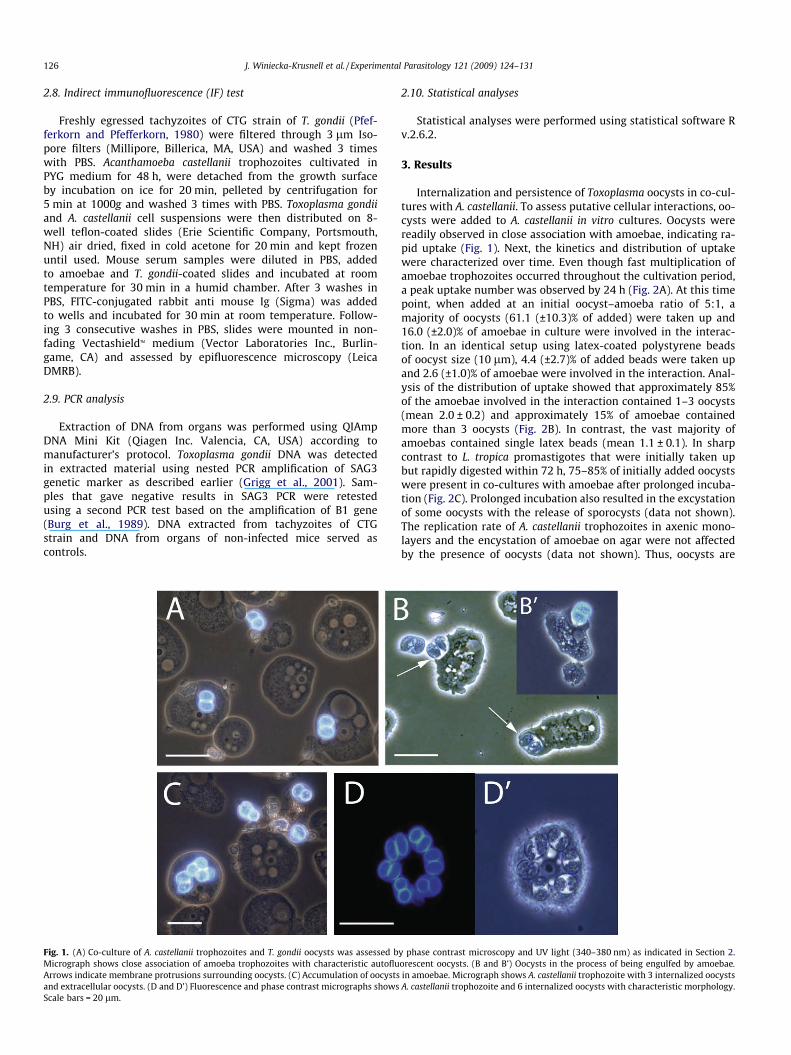
Fig. 1. (A) Co-culture of A. castellanii trophozoites and T. gondii oocysts was assessed bMicrograph shows close association of amoeba trophozoites with characteristic autofluArrows indicate membrane protrusions surrounding oocysts. (C) Accumulation of oocystsand extracellular oocysts. (D and D’) Fluorescence and phase contrast micrographs showsScale bars = 20 lm.
2.10. Statistical analyses
Statistical analyses were performed using statistical software Rv.2.6.2.
3. Results
Internalization and persistence of Toxoplasma oocysts in co-cul-tures with A. castellanii. To assess putative cellular interactions, oo-cysts were added to A. castellanii in vitro cultures. Oocysts werereadily observed in close association with amoebae, indicating ra-pid uptake (Fig. 1). Next, the kinetics and distribution of uptakewere characterized over time. Even though fast multiplication ofamoebae trophozoites occurred throughout the cultivation period,a peak uptake number was observed by 24 h (Fig. 2A). At this timepoint, when added at an initial oocyst–amoeba ratio of 5:1, amajority of oocysts (61.1 (±10.3)% of added) were taken up and16.0 (±2.0)% of amoebae in culture were involved in the interac-tion. In an identical setup using latex-coated polystyrene beadsof oocyst size (10 lm), 4.4 (±2.7)% of added beads were taken upand 2.6 (±1.0)% of amoebae were involved in the interaction. Anal-ysis of the distribution of uptake showed that approximately 85%of the amoebae involved in the interaction contained 1–3 oocysts(mean 2.0 ± 0.2) and approximately 15% of amoebae containedmore than 3 oocysts (Fig. 2B). In contrast, the vast majority ofamoebas contained single latex beads (mean 1.1 ± 0.1). In sharpcontrast to L. tropica promastigotes that were initially taken upbut rapidly digested within 72 h, 75–85% of initially added oocystswere present in co-cultures with amoebae after prolonged incuba-tion (Fig. 2C). Prolonged incubation also resulted in the excystationof some oocysts with the release of sporocysts (data not shown).The replication rate of A. castellanii trophozoites in axenic mono-layers and the encystation of amoebae on agar were not affectedby the presence of oocysts (data not shown). Thus, oocysts are
y phase contrast microscopy and UV light (340–380 nm) as indicated in Section 2.orescent oocysts. (B and B’) Oocysts in the process of being engulfed by amoebae.in amoebae. Micrograph shows A. castellanii trophozoite with 3 internalized oocystsA. castellanii trophozoite and 6 internalized oocysts with characteristic morphology.

Fig. 2. (A) Uptake of T. gondii oocysts by A. castellanii overtime. Assessment of intra-amoebic parasites was performed as indicated in Section 2. Uptake index indicatesfor each time point the frequency of amoeba trophozoites containing intracellularparasites related to the frequency of amoeba trophozoites containing intracellularparasites after 2 h incubation (index 1 = 9.0%). Data were not adjusted for thereplication of amoebic trophozoites over time. Data are means (±SD) from onerepresentative experiment performed in triplicate. (B) Distribution of internalizedoocysts and latex-coated polystyrene beads (Macrobeads) in acanthamoebae after24 h of incubation. Counts indicate number of internalized oocysts or beads. Dataare means (±SD) from one representative experiment performed in triplicate. (C)Persistence of T. gondii oocysts and L. tropica promastigotes in co-cultures with A.castellanii. For each time point, parasites present in co-cultures were assessed asindicated and related to the number of parasites added initially. Data are means(±SD) from one representative experiment performed in triplicate.
Fig. 3. Inhibition of uptake of oocysts and latex beads by A. castellanii trophozoites.Cytochalasin D (10 lg/ml) or monosaccharides (10 mM) were added to cultures andthe number of amoebae, containing oocysts or latex-coated polystyrene beads(Macrobeads) was related to untreated co-cultures as indicated in Section 2. Dataare means (±SD) from three independent experiments.
J. Winiecka-Krusnell et al. / Experimental Parasitology 121 (2009) 124–131 127
internalized in A. castellanii trophozoites and exhibit long-termpersistence in the presence of acanthamoebae in vitro.
3.1. Active uptake of oocysts by amoebae
To further investigate oocyst internalization by A. castellanii, up-take was assessed in the presence of compounds known to targetphagocytic events in amoebae (Allen and Dawidowicz, 1990; Venk-ataraman et al., 1997). Internalization of oocysts was abolished by
treatment with cytochalasin D (Fig. 3). In competition assays usingmonosaccharides, the dose-dependent inhibition of uptake ob-served was most prominent for D(+)mannose and a-methyl-D-mannoside (p < 0.01, Student’s t test) for both oocysts and latex-coated polystyrene beads (Fig. 3 and Supplementary materialFig. S1). In contrast, D(+)galactose, D-fructose-6-phosphate,D(+)galactosamine and N-acetyl-D-galactosamine only had inhibi-tory effect at very high concentrations (Supplementary materialFig. S1), that also affected surface adhesion of amoebae (data notshown).
We next investigated the fate of internalized oocysts by label-ling acidic vacuoles of amoebae at different time points of co-cul-tivation. Co-localization of oocysts with acidic vacuole staining(LysoTrackerTM) was observed sporadically but absence of co-locali-zation appeared to predominate (Fig. 4). Quantitative analysis ofco-localization over time revealed an increasing number of oocystsco-localizing with LysoTrackerTM staining during the first 24 h and adecrease thereafter. At any time point, the vast majority of inter-nalized oocysts (>93%) did not co-localize with LysoTrackerTM acidicvacuole staining (Fig. 5). Next, the viability of intra-amoebic oo-cysts was assessed by time-lapse microscopy. Internalized oocystsexhibited conserved morphology and organelle motility (Supple-mentary material video S1). Thus, internalization of oocysts byamoebae is an active process that requires actin rearrangementand can be competed out by saccharides. Importantly, phagocyto-sis of internalized oocysts is characterized by low efficiency andconsequently leads to the presence of morphologically intact intra-cellular oocysts.
3.2. Intra-amoebic oocysts establish infection in mice
To test whether intra-amoebic oocysts could establish infection,BALB/c mice were challenged orally with intracellular oocysts ob-tained after 48 h of pre-incubation with A. castellanii and the infec-tion was compared to infection with free oocysts. Infection wasmonitored by antibody response, PCR and plaque forming assay.Regardless of mode of infection, free or intra-amoebic oocysts, allmice became infected with T. gondii and none developed anti-Acan-thamoeba antibodies (Table 1). Differences between groups werenon-significant (p > 0.05, Fisher’s exact test). The parasite burdenin mice infected with 102 oocysts was assessed by plaque formingassays by day 9 or 20 p.i. (Fig. 6). While parasite loads were highestin the spleen during acute infection (Fig. 6A), the brain containedthe highest parasite loads by day 20 p.i. (Fig. 6B). The differencesin parasite loads in mice infected with free oocysts or intra-amoe-

Fig. 4. Acidic vacuole staining of A. castellanii trophozoites with internalized T. gondii oocysts. Co-cultures of amoebae and oocysts were stained with LysoTrackerTM asindicated in Section 2. Arrows indicate internalized oocysts and depicts localization of oocysts (blue) in acidic vacuoles (red arrows) and non-acidic vacuoles (white arrows).Scale bar = 20 lm.
Fig. 5. Co-localization of intracellular oocysts with lysosomal marker. A. castellaniimonolayers with added T. gondii oocysts were stained with LysoTrackerTM andassessed by epifluorescence microscopy at indicated time points as described inSection 2. Diagram shows the frequency (%) of double fluorescent vacuoles [blue(oocyst) + red (LysoTrackerTM)] related to the total number of internalized oocysts.Data were not adjusted for the replication of amoebic trophozoites and are means(±SD) from a representative experiment performed in triplicate.
128 J. Winiecka-Krusnell et al. / Experimental Parasitology 121 (2009) 124–131
bic oocysts 9 days p.i. were non-significant (p > 0.05, Student’s ttest). Interestingly, by day 20 p.i. significantly higher parasite num-bers were demonstrated in brains (p = 0.008) and livers (p = 0.012),but not in spleens (p = 0.596), of mice infected with intra-amoebicoocysts compared to mice infected with free oocysts. We concludethat intra-amoebic oocysts establish primary T. gondii infection asefficiently as free oocysts. Furthermore, establishment of chronicinfection was not hindered, reaching similar or higher parasiteloads in peripheral organs and CNS.
3.3. Infectivity of oocysts after prolonged co-cultivation with amoebae
To determine the effects of prolonged co-incubation on oocystinfectivity, BALB/c mice were orally infected with 102 oocysts
pre-incubated for 7 or 14 d with amoebae. The infectivity of oo-cysts, defined as positive PCR or plaquing assay, remained highwith 5/5 animals infected after 7 d incubation and 4/5 animals in-fected after 14 d incubation (Table 2). Differences in infectivity be-tween intra-amoebic oocysts and free oocysts were non-significantfor both time points (p > 0.05, Fisher’s exact test). Additionally, dif-ferences in infectivity between intra-amoebic oocysts incubatedfor 7 or 14 d were also non-significant (p > 0.05, Fisher’s exact test).The parasite loads in organs of mice infected with material incu-bated for 7 and 14 d was similar to that observed after 48 h incu-bation (Supplementary material Fig. S2 and Fig. 6). Thus,incubation of oocysts with acanthamoebae for extended periods(up to 14 d) did not reduce the infectivity of oocysts.
4. Discussion
In their environmental habitats, Acanthamoeba spp. prey onmicroorganisms, mainly bacteria, present in the biofilms (Rodri-guez-Zaragoza, 1994). Some engulfed microorganisms, however,have developed mechanisms that allow them to resist destructionand remain viable inside amoebae (Barker et al., 1995; Greub andRaoult, 2004). In this study, we have investigated interactions be-tween A. castellanii trophozites and T. gondii oocyts in vitro and as-sessed infectivity of oocysts in vivo. Present results show that A.castellanii can engulf T. gondii oocysts. Further, internalized oocystswere not digested to a significant extent and retained their abilityto establish infection in mice.
Endocytic uptake of microorganisms involves interactions withthe carbohydrate rich surface membrane of acanthamoebae (Ello-way et al., 2004). Mannose-binding protein (MBP) of Acanthamoebaspp. has been previously linked to the phagocytic uptake of yeastsand bacteria, e.g. Legionella pneumophila, (Allen and Dawidowicz,1990; Alsam et al., 2005; Declerck et al, 2007) and to adhesion ofamoebae to the corneal surface (Larkin et al., 1991; Yang et al.,1997). Also, a Gal/GalNac lectin has been identified as a receptorfor attachment and invasion by legionellae in Hartmannella vermi-formis (Venkataraman et al., 1997) but not in A. polyphaga (Harbet al., 1998). The inhibitory effects of saccharides on uptake of T.gondii oocysts by A. castellanii observed here are reminiscent of

Table 1Comparison of mice orally infected with free T. gondii oocysts or with intra-amoebic oocysts obtained after 48 h of incubation with A. castellanii.
9 d.p.i.a 20 d.p.i.b
102 103 102 103 103 102 102 102
Oocysts in amoebaec Free oocystsd Amoebaee Oocysts in amoebae Free oocysts Amoebaee
n = 4 n = 4 n = 4 n = 2 n = 2 n = 8 n = 6 n = 2T. gondii antibodies, P1/100f 3/4g 4/4 4/4 2/2 0/2 8/8 6/6 0/2A. castellanii antibodies, negative 4/4 4/4 4/4 2/2 2/2 8/8 6/6 2/2Positive PCRh, brain 3/4g 4/4 4/4 2/2 0/2 8/8 6/6 0/2Positive PCR, liver 4/4 4/4 4/4 2/2 0/2 8/8 6/6 0/2Positive PCR, spleen 3/4g 4/4 4/4 2/2 0/2 8/8 6/6 0/2Positive plaquing assayi, brain 4/4 ND 4/4 ND ND 8/8 6/6 NDPositive plaquing assay, liver 4/4 ND 4/4 ND ND 8/8 6/6 NDPositive plaquing assay, spleen 4/4 ND 4/4 ND ND 8/8 6/6 ND
ND, not done.Differences in infectivity between groups infected with free oocysts or intra-amoebic oocysts were non-significant (p > 0.05, Fisher’s exact test).
a Mice euthanized 9 d.p.i. were assessed for T. gondii antibodies, positive PCR and plaque forming assays as indicated in Section 2.b Mice euthanized 20 d.p.i. were assessed as above.c Mice were orally infected with 102 or 103 intra-amoebic oocysts as indicated in Section 2. All mice given 103 oocysts developed severe symptoms and were euthanized 9
d.p.i.d Mice infected with 102 or 103 free oocysts.e Mice infected with 102 or 103 A. castellanii trophozoites.f Antibodies were detected in mice sera using IF test as indicated in Section 2.g One mouse in this group showed lower T. gondii antibody titre: >1/10 <1/50 and its brain and spleen remained PCR negative.h Extraction of DNA from tissues and PCR was performed as indicated in Section 2.i Tissues were processed and plaquing assays were performed as described in Section 2.
J. Winiecka-Krusnell et al. / Experimental Parasitology 121 (2009) 124–131 129
those observed for L. pneumophila and A. castellanii. (Declerck et al,2007). Yet, uptake of latex beads was inhibited in a similar fashion,indicating that the inhibition was not necessarily specific for up-take of oocysts.
Acanthamoeba spp. are capable of internalizing different typesof particles, some of them of no nutritive value like silica, carbon,or plastic. However, amoebae appear to be able to discriminate be-
Fig. 6. Parasite loads in mice infected with 102 free oocysts or with 102 intra-amoebic oocysts obtained after 48 h of incubation with A. castellanii trophozoites.Organs were extracted (A) 9 d.p.i. and (B) 20 d.p.i. and assessment was performedby plaque forming assays as indicated in Section 2. Graphics show means (±SD)from two independent experiments performed in triplicate.
tween digestible and indigestible particles and expel the last (Bow-ers and Olszewski, 1983). Oocysts persevered in co-culture withamoebae for weeks without significant destruction. Remarkably,the vast majority of internalized oocysts did not co-localize withacidic vacuolar staining at any time point. This may reflect a veryearly phase of internalization or normalization of pH in a laterphase. The findings are in contrast to the fate of latex beads andheat-killed or formalin treated legionellae that appear to be chieflyobserved in lysosomal vacuoles but resemble observations withlive Legionella, that inhibits phago-lysosomal maturation (Bozueand Johnson, 1996; Wetzel and Korn, 1969). Given their resistanceto acid environment (Dubey, 2004), it is not unlikely that oocystscan resist acidification of the endocytic vacuoles. Also, at this point,an alteration of lysosmal fusion induced by oocysts cannot beexcluded.
Transient incorporation of oocysts of C. parvum with the aver-age of 2 oocysts per amoeba has been reported recently (Gomez-
Table 2Infectivity of intra-amoebic oocysts compared to free oocysts after prolongedincubation with A. castellanii.
7 days of co-cultivation 14 days of co-cultivation
Oocysts inamoebaea
Freeoocystsb
Oocysts inamoebae
Freeoocysts
n = 5 n = 5 n = 5 n = 5Positive PCRc, spleen 5/5 4/5 4/5 3/5Positive PCR, liver 5/5 2/5 3/5 3/5Positive PCR, brain 5/5 3/5 4/5 3/5Positive plaquing
assayd, spleen5/5 2/5 3/5 3/5
Positive plaquingassay, liver
5/5 2/5 3/5 3/5
Positive plaquingassay, brain
5/5 2/5 3/5 3/5
Differences in infectivity between groups were non-significant (p > 0.05, Fisher’sexact test).
a BALB/c mice were orally infected with 102 intra-amoebic oocysts and eutha-nized 9 d.p.i. as indicated in Section 2.
b Mice were infected with 102 free oocysts and euthanized 9 d.p.i.c Extraction of DNA from tissues and PCR was performed as indicated in Section 2.d Tissues were processed and plaquing assays were performed as described in
Section 2.

130 J. Winiecka-Krusnell et al. / Experimental Parasitology 121 (2009) 124–131
Couso et al., 2006). In our study, the majority of amoebae involvedin the interaction contained 1–3 T. gondii oocysts (average 2.0) andamoebae containing up to 13 oocysts were observed. Gomez-Cousoet al. (2006) reported that amoebae trophozoites retain C. parvumoocysts only temporarily and actively release them with time.Although not experimentally addressed here, it is also likely thata certain turnover may take place for T. gondii oocysts. Further,we never observed intracellular oocysts in encysted amoebae.Steward and Weisman (1972) reported that acanthamoebae expelnon-nutritive particles like latex prior to encystation, but it is notclear if trophozoites containing oocysts were unable to encyst orif oocysts were released before the encystation process started.
Viability of oocysts after exposure to amoebae for up to 2 weekswas indirectly assessed by bioassay in mice. There is no simple via-bility test for T. gondii oocysts and use of techniques applied for cystsof Giardia spp. and Cryptosporidium spp. oocysts is not evident(Dumètre and Dardé, 2003). Bioassay in mice that are sensitive toinfection is still a reference method (Dubey and Beattie, 1988; Du-bey, 2004). Our results show that infectivity of intra-amoebic oo-cysts in mice was not reduced compared to free oocysts.Interactions with amoebae did not interfere with the infectiousproperties of oocysts in terms of parasite burden and disseminationof parasites in the organism. The significance of the higher parasiteload observed during chronic infection in brain and liver of mice in-fected with intra-amoebic oocysts needs to be further evaluated.
Considering the common occurrence of Acanthamoeba spp. inwater, both humans and animals are frequently exposed e.g. bydrinking, eating, and bathing or by soil contact. Although drinkingwater and seawater have been linked to epidemic infections withT. gondii, the demonstration of parasites in water is difficult toachieve. Water may become contaminated with oocysts by directlydeposited cat feces or by oocyst-containing soil washing after rain-fall (Dubey, 2004; Dumètre and Dardé, 2003; Sroka et al., 2006).The number of oocysts in a random sample is probably low (Dumètreand Dardé, 2003) and ingestion of molluscs containing concentratedoocysts has been considered as a mode of transmission of infection insea otters (Conrad et al., 2005). Experimental studies showed that bi-valves could accumulate both oocysts of different coccidian para-sites including T. gondii and cysts of Giardia (Arkush et al., 2003;Graczyk et al., 1998a,b,c, 1999; Lindsay et al., 2001). However, T. gon-dii oocysts in naturally exposed molluscs have not been reported(Conrad et al., 2005). Additionally, infection with T. gondii has beendescribed in several marine mammals that do not consume bivalves(Dubey et al., 2003) and ingestion of oocysts present in prey animalshas been proposed as one of the theoretical modes of transmission ofwaterborne T. gondii infections (Conrad et al., 2005).
Our experimental data indicates that intra-amoebic location ofoocysts does not reduce infectivity in mice. Oocysts are well pre-pared to withstand hostile conditions and, opposite to many bacte-ria, do not essentially depend on protection provided by otherorganisms. Yet, interactions between free-living amoebae and T.gondii oocysts could in theory have an impact on transient accu-mulation and dispersion of oocysts in aquatic environments. How-ever, the role of amoebae in the transmission of waterbornetoxoplasmosis awaits further investigation.
Acknowledgments
The study was supported in part by a grant from Swedish Re-search Council (to A.B.).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.exppara.2008.09.022.
References
Allen, P.G., Dawidowicz, E.A., 1990. Phagocytosis in Acanthamoeba: I. A mannosereceptor is responsible for the binding and phagocytosis of yeast. Journal of CellPhysiology 145, 508–513.
Alsam, S., Sissons, J., Dudley, R., Khan, N.A., 2005. Mechanisms associated withAcanthamoeba castellanii (T4) phagocytosis. Parasitology Research 96, 402–409.
Arkush, K.D., Miller, M.A., Leutenegger, C.M., Gardner, I.A., Packham, A.E., Heckroth,A.R., Tenter, A.M., Barr, B.C., Conrad, P.A., 2003. Molecular and bioassay-baseddetection of Toxoplasma gondii oocysts uptake by mussels (Mytilusgalloprovinciallis). International Journal of Parasitology 33, 1087–1097.
Bahia-Oliveira, L.M., Jones, J.L., Azevedo-Silva, J., Alves, C.C., Orefice, F., Addiss, D.G.,2003. Highly endemic, waterborne toxoplasmosis in north Rio de Janeiro state,Brazil. Emerging Infectious Diseases 9, 55–62.
Barker, J., Scaife, H., Brown, M.R., 1995. Intraphagocytic growth induces anantibiotic-resistant phenotype of Legionella pneumophila. Antimicrobial Agentsand Chemotherapy 39, 2684–2688.
Benenson, M.W., Takafuji, E.T., Lemon, S.M., Greenup, R.L., Sulzer, A.J., 1982. Oocyst-transmitted toxoplasmosis associated with ingestion of contaminated water.New England Journal of Medicine 307, 666–669.
Bowers, B., Olszewski, T.E., 1983. Acanthamoeba discriminates internally betweendigestible and indigestible particles. Journal of Cell Biology 97, 317–322.
Bowie, W.R., King, A.S., Werker, D.H., Isaac-Renton, J.L., Bell, A., Eng, S.B., Marion,S.A., 1997. Outbreak of toxoplasmosis associated with municipal drinkingwater. The BC Toxoplasma Investigation Team. Lancet 350, 173–177.
Bozue, J.A., Johnson, W., 1996. Interaction of Legionella pneumophila withAcanthamoeba castellanii: uptake by coiling phagocytosis and inhibition ofphagpsome–lysosome fusion. Infection and Immunity 64, 668–673.
Burg, J.L., Grover, C.M., Pouletty, P., Boothroyd, J.C., 1989. Direct and sensitivedetection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chainreaction. Journal of Clinical Microbiology 27, 1787–1792.
Cirillo, J.D., Falkow, S., Tompkins, L.S., 1994. Growth of Legionella pneumophila inAcanthamoeba castellanii enhances invasion. Infection and Immunity 62, 3254–3261.
Cirillo, J.D., Falkow, S., Tompkins, L.S., Bermudez, L.E., 1997. Interaction ofMycobacterium avium with environmental amoebae enhances virulence.Infection and Immunity 65, 3759–3767.
Conrad, P.A., Miller, M.A., Kreuder, C., James, E.R., Mazet, J., Dabritz, H., Jessup, D.A.,Gulland, F., Grigg, M.E., 2005. Transmission of Toxoplasma: clues from the studyof sea otters as sentinels of Toxoplasma gondii flow into the marineenvironment. International Journal of Parasitology 35, 1155–1168.
Declerck, P., Behets, J., De Keersmaecker, B., Ollevier, F., 2007. Receptor-mediateduptake of Legionella pneumophila by Acanthamoeba castellanii and Naeglerialovaniensis. Journal of Applied Microbiology 103, 2697–2703.
de Moura, L., Bahia-Oliveira, L.M., Wada, M.Y., Jones, J.L., Tuboi, S.H., Carmo, E.H.,Ramalho, W.M., Camargo, N.J., Trevisan, R., Graca, R.M., da Silva, A.J., Moura, I.,Dubey, J.P., Garret, D.O., 2006. Waterborne toxoplasmosis, Brasil, from field togene. Emerging Infectious Diseases 12, 326–329.
Dubey, J.P., Beattie, C.P., 1988. Toxoplasmosis of Animals and Man. Boca Raton, FL.Dubey, J.P., 1995. Duration of immunity to shedding of Toxoplasma gondii oocysts by
cats. Journal of Parasitology 81, 410–415.Dubey, J.P., Lunney, J.K., Shen, S.K., Kwok, O.C.H., Ashford, D.A., Thulliez, P., 1996.
Infectivity of low numbers of Toxoplasma gondii oocysts to pigs. Journal ofParasitology 82, 438–443.
Dubey, J.P., Zaranke, R., Thomas, N.J., Wong, S.K., Van Bonn, W., Briggs, M., Davis,J.W., Ewing, R., Mense, M., Kwok, O.C., Romand, S., Thulliez, P., 2003. Toxoplasmagondii, Neospora caninum, Sarcocystis neurona and Sarcocystis canis-likeinfections in marine mammals. Veterinary Parasitology 116, 275–296.
Dubey, J.P., 2004. Toxoplasmosis—a waterborne zoonosis. Veterinary Parasitology126, 57–72.
Dumère, A., Dardé, M.L., 2003. How to detect Toxoplasma gondii oocysts inenvironmental samples? FEMS Microbiology Reviews 27, 651–661.
Elloway, E.A.G., Armstrong, R.A., Bird, R.A., Kelly, S.L., Smith, S.N., 2004. Analysis ofAcanthamoeba polyphaga surface carbohydrate exposure by FITC-lectin bindingand fluorescence evaluation. Journal of Applied Microbiology 97, 1319–1325.
Gomez-Couso, H., Paniagua-Crespo, E., Ares-Mazas, E., 2006. Acanthamoeba as atemporal vehicle of Cryptosporidium. Parasitology Research 100, 1151–1154.
Graczyk, T.K., Farley, C.A., Fayer, R., Lewis, E.J, Trout, J.M., 1998a. Detection ofCryptosporidium oocysts and Giardia cysts in the tissues of eastern oysters(Crassostrea virginica) carrying principal oyster infectious diseases. Journal ofParasitology 84, 1039–1042.
Graczyk, T.K., Fayer, R., Cranfield, M.R., Conn, B.D., 1998b. Recovery of waterborneCryptosporidium parvum oocysts by freshwater benthic clams (Corbiculafluminea). Applied and Environmental Microbiology 64, 427–430.
Graczyk, T.K., Ortega, Y.R., Conn, D.B., 1998c. Recovery of waterborne oocysts ofCyclospora cayetanensis by Asian freshwater clams (Corbicula fluminea).American Journal of Tropical Medicine and Hygiene 59, 928–932.
Graczyk, T.K., Fayer, R., Lewis, E.J., Trout, J.M., Farley, C.A., 1999. Cryptosporidiumoocysts in bent mussels (Ischadium recurvum)in the Chesapeake Bay.Parasitology Research 85, 518–521.
Greub, G., Raoult, D., 2004. Microorganisms resistant to free-living amoebae.Clinical Microbiology Reviews 17, 413–433.
Grigg, M.E., Ganatra, J., Boothroyd, J.C., Margolis, T.P., 2001. Unusual abundance ofatypical strains associated with human ocular toxoplasmosis. Journal ofInfectious Diseases 184, 633–639.

J. Winiecka-Krusnell et al. / Experimental Parasitology 121 (2009) 124–131 131
Harb, O.S., Venkatamaran, C., Haack, B.J., Gao, L.-Y., Abu Kwaik, Y., 1998.Heterogeneity in the attachment and uptake mechanisms of the legionnaire’sdisease bacterium Legionella pneumophila by protozoan hosts. Applied andEnvironmental Microbiology 64, 126–132.
Hoffmann, R., Michel, R., Schmid, E.N., Muller, K.D., 1998. Natural infection withmicrosporidian organisms (KW19) in Vannella spp. (Gymnamoebia) isolatedfrom a domestic tap-water supply. Parasitology Research 84, 164–166.
Korn, E.D., Weisman, R.A., 1967. Phagocytosis of latex beads by Acanthamoeba. II.Electron microscopic study of the initial events. Journal of Cell Biology 34, 219–227.
Larkin, D.F., Berry, M., Easty, D.L., 1991. In vitro corneal pathogenicity ofAcanthamoeba. Eye 5, 560–568.
Lindsay, D.S., Phelps, K.K., Smith, S.A., Flick, G., Sumner, S.S., Dubey, J.P., 2001.Removal of Toxoplasma gondii oocysts from sea water by eastern oysters(Crassostrea virginica). Journal of Eukaryotic Microbiology (Suppl.), 197S–198S.
Lindquist, H.D., Bennett, J.W., Hester, J.D., Ware, M.W., Dubey, J.P., Everson, W.V.,2003. Autofluorescence of Toxoplasma gondii and related coccidian oocysts.Journal of Parasitology 89, 865–867.
Michel, R., Schmid, E.N., Boker, T., Hager, D.G., Muller, K.D., Hoffmann, R., Seitz, H.M.,2000. Vannella sp. harboring Microsporidia-like organisms isolated from thecontact lens and inflamed eye of a female keratitis patient. ParasitologyResearch 86, 514–520.
Miller, M.A., Gardner, I.A., Kreuder, C., Paradies, D.M., Worcester, K.R., Jessup, D.A.,Dodd, E., Harris, M.D., Ames, J.A., Packham, A.E., Conrad, P.A., 2002. Coastalfreshwater runoff is a risk factor for Toxoplasma gondii infection of southern seaotters (Enhydra lutris nereis). International Journal of Parasitology 32, 997–1006.
Miltner, E.C., Bermudez, L.E., 2000. Mycobacterium avium grown in Acanthamoebacastellanii is protected from the effects of antimicrobials. Antimicrobial Agentsand Chemotherapy 44, 1990–1994.
Pfefferkorn, L.C., Pfefferkorn, E.R., 1980. Toxoplasma gondii: genetic recombinationbetween drug resistant mutants. Experimental Parasitology 50, 305–316.
Rodriguez-Zaragoza, S., 1994. Ecology of free-living amoebae. Critical Reviews inMicrobiology 20, 225–241.
Sroka, J., Wojcik-Fatla, A., Dutkiewicz, J., 2006. Occurrence of Toxoplasma gondii inwater from wells located on farms. Annals of Agricultural and EnvironmentalMedicine 13, 169–175.
Steenbergen, J.N., Shuman, H.A., Casadevall, A., 2001. Cryptococcus neoformansinteractions with amoebae suggest an explanation for its virulence andintracellular pathogenic strategy in macrophages. Proceedings of the NationalAcademy of Sciences of the United States of America 98, 15245–15250.
Steenbergen, J.N., Nosanchuk, J.D., Malliaris, S.D., Casadevall, A., 2004. Interaction ofBlastomyces dermatitidis, Sporothrix schenckii, and Histoplasma capsulatum withAcanthamoeba castellanii. Infection and Immunity 72, 3478–3488.
Stewart, J.R., Weisman, R.A., 1972. Exocytosis of latex beads during the encystmentof Acanthamoeba. Journal of Cell Biology 52, 117–130.
Storey, M.V., Winiecka-Krusnell, J., Ashbolt, N.J., Stenstrom, T.A., 2004. The efficacyof heat and chlorine treatment against thermotolerant Acanthamoebae andLegionellae. Scandinavian Journal of Infectious Diseases 36, 656–662.
Stott, R., May, E., Ramirez, E., Warren, A., 2003. Predation of Cryptosporidium oocystsby protozoa and rotifers: implications for water quality and public health.Water Science and Technology 47, 77–83.
Venkataraman, C., Haack, B.J., Bondada, S., Abu Kwaik, Y., 1997. Identification of Gal/GalNAc lectin in the protozoan Hartmannella vermiformis as a potential receptorfor attachment and invasion by the legionnaire’s disease bacterium. Journal ofExperimental Medicine 186, 537–547.
Wetzel, M.G., Korn, E.D., 1969. Phagocytosis of latex beads by Acanthamoebacastellanii (Neff). III. Isolation of the phagocytic vesicles and their membranes.Journal of Cell Biology 43, 90–104.
Yang, Z., Cao, Z., Panjwani, N., 1997. Pathogenesis of Acanthamoeba keratitis:carbohydrate-mediated host–parasite interactions. Infection and Immunity 65,439–445.
Related Documents











