Towards an ecologically-sustainable forestry in the Atlantic Forest Carlos R. Fonseca a,b, * , Gislene Ganade a,b , Ronei Baldissera c , Carlos G. Becker a , Carlos R. Boelter a , Antonio D. Brescovit d , Lucas M. Campos a , Tomás Fleck a , Vanda S. Fonseca a , Sandra M. Hartz c , Fernando Joner a , Marcia I. Käffer a,e , Ana M. Leal-Zanchet a , Marcelo P. Marcelli f , Alex S. Mesquita a , Cláudio A. Mondin g , Claudia P. Paz a , Maria V. Petry a , Fabio N. Piovensan a , Jair Putzke h , Anamaria Stranz a , Micheline Vergara a , Emerson M. Vieira a,i a Centro 2, Universidade do Vale do Rio dos Sinos, São Leopoldo, RS 93022-970, Brazil b Departamento de Botânica, Ecologia e Zoologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte, Natal, RN 59072-970, Brazil c Instituto de Biociências, Centro de Ecologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS 91501-970, Brazil d Seção de Artrópodes Peçonhentos, Instituto Butantan, São Paulo, SP 05503-900, Brazil e Fundação Zoobotânica do Rio Grande do Sul, Porto Alegre, RS 90690-000, Brazil f Instituto de Botânica, São Paulo, SP 04301-902, Brazil g Departamento de Biologia, Faculdade de Biociências, Pontifícia Universidade Católica, Porto Alegre, RS 90619-500, Brazil h Departamento de Biologia, Universidade de Santa Cruz do Sul, Santa Cruz do Sul, RS 96815-900, Brazil i Departamento de Ecologia, Instituto de Biologia, Universidade de Brasília, Brasília, DF 70919-970, Brazil article info Article history: Received 15 September 2008 Received in revised form 9 February 2009 Accepted 14 February 2009 Available online 20 March 2009 Keywords: Araucaria Biodiversity Conservation Eucalyptus Forestry Management Monoculture Multi-taxa Pinus abstract Since pristine Atlantic Forest remnants are vanishing, and biological reserves are in short, conservation of biodiversity will largely depend on proper management of the anthropogenic matrix. Here, we test (1) the effectiveness of ecologically-managed tree monocultures in maintaining Araucaria Forest biodiver- sity, (2) how this effectiveness change among taxa, and (3) we discuss management principles that can be used by the forestry industry in order to contribute positively to biodiversity conservation. The study was conducted in the São Francisco de Paula National Forest, southern Brazil, an environmental mosaic composed mostly of patches of Araucaria Forest and ecologically-managed monocultures of Arau- caria, Pinus and Eucalyptus. Using standardized sampling methods in these four main habitats, we have recorded the richness and species composition of small mammals, birds, leaf-litter frogs, butterflies, gall- ing insects, spiders, opiliones, flatworms, woody plants, epiphytic angiosperms, epiphytic ferns, lichens, and fruit-body producing fungi. Overall, we recorded 506 species in Araucaria Forest, 181 (36%) of which were exclusive of this habitat while 325 (64%) could be found in at least one monoculture. Distribution patterns of species richness and number of records across taxonomic groups showed that a large biodi- versity can be found inside ecologically-managed plantations of Araucaria, Pinus, and Eucalyptus. For all studied taxa, except for epiphytic angiosperms and fruit-body producing fungi, more than half of the Araucaria Forest species could be found living on monocultures. We discuss how the actual management practices of the forest industry can be improved to contribute positively to the conservation of the Atlan- tic Forest biodiversity. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction Araucaria Forest, a subtropical rain forest located on the south most part of the Atlantic Forest, remained relatively untouched till the beginning of the XX century (Galindo-Leal and Câmara, 2003). During the First World War, with the impossibility for Europe to commercialize pine wood from Riga (Letonia), the exploitation of the Brazilian pine Araucaria angustifolia (Bertol.) Kuntze (1898) be- gan. From 1915 to 1960, Brazil exported more than 18 million m 3 of timber, mostly from the Atlantic Forest (Koch and Corrêa, 2002; Fonseca, 1985). A single British company, Lumber and Colo- nisation Comp. Ltda., extracted from Araucaria Forest more than 15 million A. angustifolia trees (Medeiros et al., 2004). Nowadays, con- sidering the 25,379,316 ha previously occupied by Araucaria For- est, only 12.6% (3,202,134 ha) remains intact (Ribeiro et al., 2009; but see Guerra et al., 2002). In parallel to this historical contraction of the Araucaria Forest in particular, and the Atlantic Forest in general (Tabarelli et al., 2005), tree monocultures have been expanding quickly in the last 0006-3207/$ - see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocon.2009.02.017 * Corresponding author. Address: Departamento de Botânica, Ecologia e Zoologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte, Natal, RN 59072-970, Brazil. Tel.: +55 84 32153374. E-mail address: [email protected] (C.R. Fonseca). Biological Conservation 142 (2009) 1209–1219 Contents lists available at ScienceDirect Biological Conservation journal homepage: www.elsevier.com/locate/biocon

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biological Conservation 142 (2009) 1209–1219
Contents lists available at ScienceDirect
Biological Conservation
journal homepage: www.elsevier .com/locate /b iocon
Towards an ecologically-sustainable forestry in the Atlantic Forest
Carlos R. Fonseca a,b,*, Gislene Ganade a,b, Ronei Baldissera c, Carlos G. Becker a, Carlos R. Boelter a,Antonio D. Brescovit d, Lucas M. Campos a, Tomás Fleck a, Vanda S. Fonseca a, Sandra M. Hartz c,Fernando Joner a, Marcia I. Käffer a,e, Ana M. Leal-Zanchet a, Marcelo P. Marcelli f, Alex S. Mesquita a,Cláudio A. Mondin g, Claudia P. Paz a, Maria V. Petry a, Fabio N. Piovensan a, Jair Putzke h, Anamaria Stranz a,Micheline Vergara a, Emerson M. Vieira a,i
a Centro 2, Universidade do Vale do Rio dos Sinos, São Leopoldo, RS 93022-970, Brazilb Departamento de Botânica, Ecologia e Zoologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte, Natal, RN 59072-970, Brazilc Instituto de Biociências, Centro de Ecologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS 91501-970, Brazild Seção de Artrópodes Peçonhentos, Instituto Butantan, São Paulo, SP 05503-900, Brazile Fundação Zoobotânica do Rio Grande do Sul, Porto Alegre, RS 90690-000, Brazilf Instituto de Botânica, São Paulo, SP 04301-902, Brazilg Departamento de Biologia, Faculdade de Biociências, Pontifícia Universidade Católica, Porto Alegre, RS 90619-500, Brazilh Departamento de Biologia, Universidade de Santa Cruz do Sul, Santa Cruz do Sul, RS 96815-900, Brazili Departamento de Ecologia, Instituto de Biologia, Universidade de Brasília, Brasília, DF 70919-970, Brazil
a r t i c l e i n f o a b s t r a c t
Article history:Received 15 September 2008Received in revised form 9 February 2009Accepted 14 February 2009Available online 20 March 2009
Keywords:AraucariaBiodiversityConservationEucalyptusForestryManagementMonocultureMulti-taxaPinus
0006-3207/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.biocon.2009.02.017
* Corresponding author. Address: Departamento deCentro de Biociências, Universidade Federal do Rio59072-970, Brazil. Tel.: +55 84 32153374.
E-mail address: [email protected] (C.R. Fonseca)
Since pristine Atlantic Forest remnants are vanishing, and biological reserves are in short, conservation ofbiodiversity will largely depend on proper management of the anthropogenic matrix. Here, we test (1)the effectiveness of ecologically-managed tree monocultures in maintaining Araucaria Forest biodiver-sity, (2) how this effectiveness change among taxa, and (3) we discuss management principles thatcan be used by the forestry industry in order to contribute positively to biodiversity conservation. Thestudy was conducted in the São Francisco de Paula National Forest, southern Brazil, an environmentalmosaic composed mostly of patches of Araucaria Forest and ecologically-managed monocultures of Arau-caria, Pinus and Eucalyptus. Using standardized sampling methods in these four main habitats, we haverecorded the richness and species composition of small mammals, birds, leaf-litter frogs, butterflies, gall-ing insects, spiders, opiliones, flatworms, woody plants, epiphytic angiosperms, epiphytic ferns, lichens,and fruit-body producing fungi. Overall, we recorded 506 species in Araucaria Forest, 181 (36%) of whichwere exclusive of this habitat while 325 (64%) could be found in at least one monoculture. Distributionpatterns of species richness and number of records across taxonomic groups showed that a large biodi-versity can be found inside ecologically-managed plantations of Araucaria, Pinus, and Eucalyptus. For allstudied taxa, except for epiphytic angiosperms and fruit-body producing fungi, more than half of theAraucaria Forest species could be found living on monocultures. We discuss how the actual managementpractices of the forest industry can be improved to contribute positively to the conservation of the Atlan-tic Forest biodiversity.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Araucaria Forest, a subtropical rain forest located on the southmost part of the Atlantic Forest, remained relatively untouched tillthe beginning of the XX century (Galindo-Leal and Câmara, 2003).During the First World War, with the impossibility for Europe tocommercialize pine wood from Riga (Letonia), the exploitation of
ll rights reserved.
Botânica, Ecologia e Zoologia,Grande do Norte, Natal, RN
.
the Brazilian pine Araucaria angustifolia (Bertol.) Kuntze (1898) be-gan. From 1915 to 1960, Brazil exported more than 18 million m3
of timber, mostly from the Atlantic Forest (Koch and Corrêa,2002; Fonseca, 1985). A single British company, Lumber and Colo-nisation Comp. Ltda., extracted from Araucaria Forest more than 15million A. angustifolia trees (Medeiros et al., 2004). Nowadays, con-sidering the 25,379,316 ha previously occupied by Araucaria For-est, only 12.6% (3,202,134 ha) remains intact (Ribeiro et al., 2009;but see Guerra et al., 2002).
In parallel to this historical contraction of the Araucaria Forestin particular, and the Atlantic Forest in general (Tabarelli et al.,2005), tree monocultures have been expanding quickly in the last

1210 C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219
decades (Baptista and Rudel, 2006; Bracelpa, 2007). In 2000, Brazilbecame the sixth country in area occupied by tree monocultures,mostly of Pinus and Eucalyptus exotic species, reaching a total of5 million ha (Bacha and Barros, 2004). The main responsible forsuch an increase are the companies of cellulose, steal, and woodcrafts, besides state governments and private landowners. In thenear future, monocultures are expected to expand steadily due tothe increasing demand for timber and energy (Bracelpa, 2007).
The expansion of tree monocultures has brought about a tradi-tionally polarized debate between entrepreneurs and brazilianenvironmentalists. In one hand, foresters hold that in order to ob-tain satisfactory revenue tree plantations should be intensivelymanaged, with the use, for instance, of exotic species, reducedrotation periods, high tree densities, and with the use of herbicides,insecticides, and nutrient input. Biodiversity losses are justified inrelation to the economic and social benefits produced by their ini-tiatives. On the other hand, environmentalists argued that in orderto maintain the local biodiversity tree monocultures should belightly managed, otherwise they are nothing but ‘‘green deserts”.
This debate takes place in a biodiversity hotspot (Mittermeieret al., 2004) with obvious limitations on the network of establishedconservation units (Indrusiak and Monteiro, forthcoming). Today,only 0.39% of the Araucaria Forest is effectively protected (Ribeiroet al., 2009). Therefore, the long-term survival of the forest biodi-versity depends on the environmental quality of the private land.The forest industry can contribute to this enterprise by embracingecologically-sustainable management approaches that allow (a)biodiversity to occur in more intensively managed areas, and (b)substantial economic revenue to be generated in lesser intensemanagement regimes. Ecologically-sustainable practices involve,for instance, the use of native species, long rotation periods, andlow tree density allowing the development of a rich and complexunderstorey (Lindenmayer and Franklin, 2002; Hartley, 2002).
Empirical evidence supporting the notion that the forest indus-try may have a larger role in the solution of the forest biodiversitycrisis than previously suspected can be found mostly in the tem-perate zone (see Lindenmayer and Franklin, 2002). In the tropics,some multi-taxa studies support the notion that, under some par-ticular management regimes, tree monocultures can hold a reason-able fraction of the local biodiversity. In Cameroon, a survey ofeight animal taxa revealed that in comparison to a Primary Forest(F) plantation of Terminaria ivorensis (P) hosted a considerablenumber of birds (P: 29 spp, F: 45 spp), butterflies (P: 30, F: 29and 33), Malaise-trapped beetles (P: 27, F: 26 and 31), canopy bee-tles (P: 52 and 80, F: 73), canopy ants (P: 79, F: 55 and 62), termites(P: 53, F: 46), and soil nematodes (P: 69, F: 70) (Lawton et al.,1998). In Amazon, a multi-taxa study demonstrated that youngEucalyptus monocultures, when surrounded by primary forest,can host a large fraction of the forest species (Barlow et al.,2007). In the Atlantic Forest, more than half the species classifiedas forest specialists, considering birds, bats, small mammals, liz-ards, leaf-litter frogs, frugivorous butterflies, trees, and ferns, canbe found inside shade cocoa plantations (Pardini et al., 2009).
The present work investigates how a range of taxonomicalgroups may be able to colonize ecologically-managed tree mono-cultures implemented in the southern Atlantic Forest region. Thespecific aims are: (1) to test the effectiveness of ecologically-man-aged tree monocultures in maintaining Araucaria Forest biodiver-sity, (2) to test how this effectiveness change among taxa, and(3) to discuss management principles that can be used by the forestindustry in order to minimize the impact of their activity on biodi-versity. The study was conducted in a mosaic landscape consti-tuted of Araucaria Forest patches and tree monoculture stands ofPinus, Eucalyptus, and Araucaria. The management regimes of suchmonocultures differ drastically from the ones adopted by econom-ically-driven monocultures of the region. Using a multi-taxa ap-
proach, we compared species richness among habitats and thepercentage of shared species between Araucaria Forest and treemonocultures for thirteen taxonomic groups, including smallmammals, birds, leaf-litter frogs, butterflies, gall-inducing insects,spiders, opiliones, terrestrial flatworms, woody plants, epiphyticangiosperms, epiphytic ferns, lichens, and fruit-body producingfungi.
2. Methods
2.1. Study area
The study was conducted in the São Francisco de Paula NationalForest (29�230–29�270S, 50�230–50�250W), southern Brazil (Fig. 1).The National Forest is a conservation unit of sustainable use, form-ing a heterogeneous mosaic landscape constituted by remnants ofAraucaria Forest together with ecologically-managed tree mono-cultures. However, the National Forest is surrounded by naturalgrasslands used as cattle fields. The reserve encompasses1600.77 ha with elevation ranging between 600 and 950 m abovesee level. The climate is subtropical with mean annual temperatureof 18.5 �C and mean annual rainfall of 2252 mm (Backes, 1999).
The landscape was originally dominated by Araucaria Forestand natural grasslands (Rambo, 2000). Before the National Forestwas established, part of the area was used for selective logging,agriculture and cattle ranching. At the end of the 1940s, a largeportion of the disturbed area was replanted with the native speciesA. angustifolia. At the end of the 1960s, two exotic species of Pinuswere introduced, the loblolly pine Pinus taeda (Elliott, 1824) andthe slash pine Pinus elliotti (Engelm, 1880). Plantations of Eucalyp-tus saligna began around the 1970s. Today, Araucaria Forest is stillthe dominant element of the landscape. Ecologically-managed treeplantations of Araucaria, Pinus, and Eucalyptus are also importantelements (Fig. 1). Other land uses include mixed plantations, plan-tations of Cupressus, besides natural grasslands, roads, lakes, wet-lands, gaps, and buildings. The National Forest is inserted in alandscape matrix dominated by grasslands used as cattle fields.
2.2. Sampling design
In 2002, we selected three site replicates of each one of the fol-lowing four habitats: Araucaria Forest (FO), and ecologically-man-aged plantations of A. angustifolia (PA), Pinus (PP) and Eucalyptus(PE). Site selection was based on availability and accessibility,but an effort was made to spread the sites and treatments acrossthe landscape. In 2002, the stands of A. angustifolia were 43, 55,and 55 years old, P. taeda 30 and 34 years old, P. elliotti 37 yearsold, and E. saligna 8, 14, and 30 years old. These long rotation peri-ods are much longer than those traditionally adopted by econom-ically-driven regional timber companies (typically 7–10 years).Notice that due to field constraints, our design was not able to fullydisentangle stand age from the habitat effect. Initial tree densityfor the plantations of Araucaria (1.5 � 1.5 m) was slight higher thanfor plantations of Pinus (2 � 2 m) and Eucalyptus (2.5 � 2 m;2.5 � 2.5 m), but most stands were submitted to posterior thinningleading to lower tree densities. Studied stands were relativelysmall in size, being in close contact with adjacent habitats(Fig. 1). Most stands hold a close and relatively complex understo-rey, due to the ecologically-sustainable practices adopted by theNational Forest.
2.3. Survey methodologies
In each one of the 12 study sites, we have established a one-hectare grid (100 � 100 m). From 2002 to 2006, we collected infor-mation on the abundance, richness and species composition of 13

Fig. 1. The mosaic landscape of the São Francisco de Paula National Forest (Rio Grande do Sul, Southern Brazil) includes mostly remnants of Araucaria Forest, plantations ofthe native tree species Araucaria angustifolia, and plantations of exotic Pinus and Eucalyptus species. Other minor land uses includes mixed plantations, natural grasslands,roads, lakes and buildings. Land use after a LandSat TM7 image from 2000.
C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219 1211
taxonomic groups: small mammals, birds, leaf-litter frogs, butter-flies, gall-inducing insects, spiders, opiliones, terrestrial flatworms,woody plants, epiphytic angiosperms, epiphytic ferns, lichens, andfruit-body producing fungi. Sampling methodology and effort var-ied widely across taxonomic group but were the same betweenhabitats and sites. When surveys had to be interrupted for a periodof time which could influence species composition, sampling wasmade by blocks that encompassed one site of each one of the fourhabitats. This procedure controls for possible differences in rich-ness or species composition that may occur due to temporal effectsand not habitat effects. Where necessary, voucher species were
collected to allow posterior identification by expert taxonomists.Below, we give a brief description of the sampling methodologyof each taxon.
Small mammals and leaf-litter amphibians were sampled bi-monthly, from October 2003 to April 2005, by five randomly lo-cated pitfall traps (33 cm diameter, 48 cm height) per site. Eachtrap was associated to four orthogonal drift fences (3 m long,50 cm high, 10 cm below the surface). The pitfalls remained openfor 72 h during each bimonthly census (total sampling effort of1800 traps day, 450 traps day per habitat). After identification theindividuals were released.

1212 C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219
Bird species and abundance were recorded for every site twice aseason, from August 2002 to July 2003, in a single point per siteusing the point count method (10 m radius). The location of thepoints varied between observations, being randomly located alonga 80 m long transect (10 m from the edges) that crossed the middleof the one-hectare plot. Observations were performed from 08:00to 11:00 and from 15:00 to 18:00 with an effort of 15 min in eachsite (total sampling effort: 1440 min, sampling effort per habitat:360 min).
Spiders were sampled six times, between 2003 and 2004, in two25 � 2 m plots randomly located in each site. During the day, spi-ders were actively captured by striking the vegetation with a stick,at a height between 1.0 and 2.5 m above ground, and collecting thefallen material with a canvas sheet held horizontally below thevegetation (total sampling effort: 4800 m2, sampling effort perhabitat: 1200 m2).
Oppiliones were collected in three groups of five pitfall trapsrandomly located in each site. Pitfall traps were transparent plasticcups of 7.5 cm diameter and 10 cm deep that were buried in thesoil until the top edge. In each group, the pitfall traps were dis-posed on a 3 � 3 m plot one in the center and the other four inthe corners. Pitfall traps were open for eight consecutive days dur-ing summer and winter 2003 (total sampling effort: 2880 traps -day, sampling effort per habitat: 720 traps day).
Galling insects were sampled, from October 2003 to March2004, by a single visit per site. In each site, six randomly locatedplots (10 � 10 m) were established and all plants up to 2 m inheight were inspected. For each plot, the number of galls of eachmorphotype was counted (total sampling effort: 7200 m2, sam-pling effort per habitat: 1800 m2).
Butterflies were sampled from January to April 2006, along45 days, with five Van Someren–Rydon traps per site. In each site,the traps were installed in the center and in each corner of a50 � 50 m square plot, being placed one-meter from the groundfloor. The traps were baited with over-riped banana baits and re-mained open 24 h a day (total sampling effort: 2700 traps day,sampling effort per habitat: 675 traps day).
Terrestrial flatworms were sampled on the forest floor twice aseason, from October 2003 to August 2005, in two randomly lo-cated transects (100 � 2 m) per site. For each transect, 10 ran-domly located plots (2 � 2 m) were inspected during 15 min byfour trained individuals (total sampling effort: 7680 m2, samplingeffort per habitat: 1920 m2).
Woody plants were sampled, from April 2003 to March 2004, ina single visit per site. In each site, six randomly located plots(10 � 10 m) were established. For each plot, all woody plants withmore than 15 cm cbh (circumference at breast height) were re-corded. Woody plants with less than 15 cm cbh, but higher thanone-meter tall, were surveyed in 10 � 5 m subplots placed withinthe six 10 � 10 m plots (total sampling effort: 7200 m2 and3600 m2, respectively; sampling effort per habitat: 1800 m2 and900 m2, respectively).
Epiphytic angiosperms and epiphytic ferns were surveyed in asingle visit per site, in 25 trees, from July to August 2006. Treeswere chosen as being the nearest tree (>10 cm dbh) of 25 pointshomogeneously distributed in the one-hectare plot. For each tree,the number of individuals of every epiphytic angiosperm wascounted and the number of fronds of every epiphytic fern was esti-mated using binocular lens and climbing techniques when neces-sary (total sampling effort: 300 trees, sampling effort per habitat:75 trees).
Epiphytic lichens were recorded in 10 randomly selected trees(dbh > 8 cm) per site, from March 2003 to April 2004. Only lichensoccurring in the trunk at a height of 30–150 cm above ground wererecorded (total sampling effort: 120 trees, sampling effort per hab-itat: 30 trees).
Fruit-body producing fungi from the Order Agaricales weresampled monthly, from September 2003 to August 2004, alongtwo randomly located transects (100 � 10 m) per site. The inten-sity of use of each site by each species was measured by the num-ber of fructifications (total sampling effort: 288,000 m2, samplingeffort per habitat: 72,000 m2).
2.4. Data analyses
Species were classified in two main response groups: (1) Arau-caria Forest species – species that were recorded in at least oneAraucaria Forest site. Within this group, we recognize the exclusivespecies that were recorded exclusively on Araucaria Forest sites. (2)Tree monoculture species – species that were recorded in at leastone tree monoculture, but were not recorded in Araucaria Forest.This last group includes mostly species from natural grasslands,but has also a few exotic species and some Araucaria Forest speciesthat were not recorded in Araucaria Forest samples.
This paper focuses on the conservation of Araucaria Forest bio-diversity. All analyses are performed at the habitat level (habitatdiversity) and not at the site level (alfa diversity). This allows usto evaluate the potential role of tree monocultures for hostingAraucaria Forest species. Therefore, we describe the number ofspecies and the number of records at the habitat level, consideringjointly the three site replicates. Also, we analyse the effectivenessof monocultures in hosting Araucaria Forest species as the number(or percentage) of Araucaria Forest species that could also be foundin the monocultures.
To compare how biodiversity varied among habitats, we useddistribution patterns of richness and number of records acrosstaxa. In order to test if the distribution pattern of richness exhib-ited by the monocultures were similar to the distribution patternexhibited by the Araucaria Forest, we used chi-square tests wherethe proportion of species found in each taxon in Araucaria Forestwas taken to generate the expected values. In order to test if thenumber of records of each taxon was equally distributed amongthe four habitats we used chi-squares.
In order to compare the richness of species between habitats,considering that the number of records were also unevenly distrib-uted, we used the rarefaction method (Sanders, 1968; Hurlbert,1971) as implemented in EcoSim 7.0 (Gotelli and Entsminger, 2001).
3. Results
A total of 914 species was recorded in the mosaic landscape ofthe São Francisco de Paula National Forest (N = 119,761 records).Overall, we have recorded a total of 291 species of fruit-body pro-ducing fungi (N = 42,362), 132 spiders (N = 1272), 108 woodyplants (N = 3380), 81 galling insects (N = 13,363), 78 lichens(N = 330), 59 birds (N = 1067), 45 epiphytic angiosperms(N = 6118), 35 flatworms (N = 353), 33 butterflies (N = 506), 18 epi-phytic ferns (N = 49,765), 16 small mammals (N = 784), 11 opili-ones (N = 182), and seven leaf-litter amphibians (N = 279).
Araucaria Forest was the richest habitat (S = 506, N = 82,185),however, species richness was also substantially high in planta-tions of Araucaria (S = 466, N = 22,652), Pinus (S = 397, N = 8042),and Eucalyptus (S = 318, N = 8042, Fig. 2). From the 506 AraucariaForest species, 325 (64.2%) were able to occupy one or more mono-cultures. Plantations of Araucaria had the highest number of Arau-caria Forest species (S = 248, 49%), followed by plantations of Pinus(S = 211, 41.7%) and Eucalyptus (S = 174, 34.4%). About 50% of thespecies in all monocultures were not recorded in Araucaria Forest,218 species (47%) in Plantations of Araucaria, 186 species (47%) inplantations of Pinus and 144 species (45%) in Eucalyptus plantations(Fig. 2).

0
100
200
300
400
500
600
FO PA PP PE
Habitat
Tota
l spe
cies
rich
ness
Fig. 2. Total species richness found in Araucaria Forest (FO), and plantations ofAraucaria (PA), Pinus (PP), and Eucalyptus (PE) considering together 13 taxonomicgroups. Black bars represent species that were recorded in Araucaria Forest whilewhite bars represent species that were not recorded in the Araucaria Forest butwere recorded in at least one of the studied monocultures.
C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219 1213
Araucaria Forest had 181 (36% of 506) exclusive species thatwere recorded in this natural habitat and were not recorded inthe tree monocultures (Fig. 3). The highest numbers of AraucariaForest exclusive species were recorded for fruit-body producingfungi (S = 77), epiphytic angiosperms (S = 29), and galling insects(S = 15). All taxa but mammals had species that were recordedexclusively in Araucaria Forest.
The distribution pattern of species richness across taxa exhib-ited by Araucaria Forest showed its potential to conserve biodiver-sity (Fig. 4a). Araucaria Forest exhibited the highest richness of
0
10
20
30
40
50
60
70
80
Fung
i
Epi
phyt
ic a
ngio
sper
ms
Gal
ling
inse
cts
Spi
ders
Lich
ens
Woo
dy p
lant
s
Flat
wor
ms
Bird
s
Epi
phyt
ic fe
rns
Opi
lione
s
Am
phib
ians
But
terfl
ies
Sm
all m
amm
als
Num
ber o
f exc
lusi
ve s
peci
es
Fig. 3. Number of species recorded exclusively in Araucaria Forest for 13 taxonomicgroups.
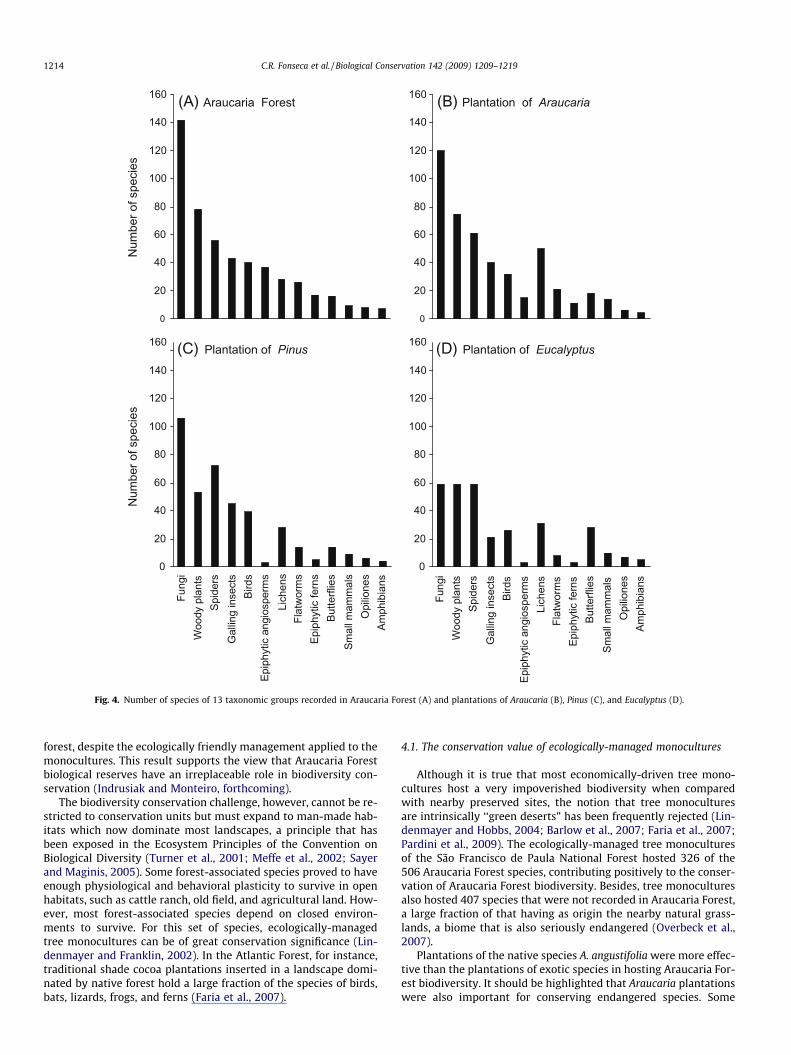
fungi (S = 142), woody plants (S = 78), birds (S = 40), epiphyticangiosperms (S = 37), flatworms (S = 26), epiphytic ferns (S = 17),opiliones (S = 8), and amphibians (S = 7, Fig. 4a). The distributionpattern exhibited by old plantations of Araucaria was very similarto that observed in Araucaria Forest, however, they had excess oflichens (S = 50) and small mammals (S = 14), and shortage of epi-phytic angiosperms (v2 = 44.39, d.f. = 12, P < 0.001, Fig. 4b). Theobserved pattern for plantations of Pinus was different due to anexcess of spiders (S = 72) and galling insects (S = 45) and shortageof fungi, woody plants, epiphytic angiosperms, flatworms, and epi-phytic ferns (v2 = 58.19, d.f. = 12, P < 0.001, Fig. 4c). Plantations ofEucalyptus showed the most distinct pattern when compared toAraucaria Forest, showing substantially fewer species of fungi,woody plants, galling insects, birds, epiphytic angiosperms, epi-phytic ferns, and flatworms, but the richest butterfly community(v2 = 103.48, d.f. = 12, P < 0.001, Fig. 4d).
The large number of records for all taxa exhibited by AraucariaForest showed that an abundant community lives inside this natu-ral habitat (Fig. 5a). The pattern for plantation of Araucaria indi-cated also a high number of records for most taxa (Fig. 5b). Thecommunity in plantations of Pinus (Fig. 5c) and, above all, Eucalyp-tus (Fig. 5d) was considerably impoverished. The number of re-cords per taxon was unevenly distributed across habitat, despitethe standardized sampling effort (v2 test, P < 0.05 for all taxa,Fig. 5). Araucaria Forest had the highest percentage of records forepiphytic ferns (95%), flatworms (65%), fungi (59%), opiliones(57%), epiphytic angiosperms (51%), birds (43%), and galling insects(30%). Plantation of Araucaria had the highest percentage of recordsfor lichens (43%), woody plants (36%), and small mammals (33%),while plantations of Pinus had more spiders (39%) and amphibians(31%). Butterflies were the only taxon overrepresented in planta-tions of Pinus (47%).
The rarefied richness showed that plantation of Araucaria hadlower richness than Araucaria Forest only for amphibians and epi-phytic angiosperms (Table 1). Richness in plantations of Pinus waslower than Araucaria Forest for amphibians, woody plants, epi-phytic angiosperms and butterflies while in plantations of Eucalyp-tus richness was lower than Araucaria Forest for birds, amphibians,galls, woody plants, epiphytic angiosperms and epiphytic ferns(Table 1). Rarefied richness also showed that estimated speciesrichness in Plantations of Eucalyptus tended to be higher than whatwas initially registered for a variety of taxonomic groups (Table 1).
For most studied taxa, more than 50% the species found in Arau-caria Forest could also be found in monocultures (Fig. 6). In con-trast, only 24% of the epiphytic angiosperms and 46% of the fungispecies found in Araucaria Forest were recorded in monocultures.For many taxa, the percentage of shared species was higher forplantations of Araucaria than for plantation of Pinus and Eucalyptus.
We found 18 species at risk in the present survey: four bird spe-cies, eight woody plant species and six epiphytic angiosperms. Tenof these species were found in Araucaria Forest, six in Pinus planta-tion, and six in Eucalyptus plantations (Table 2). Seven of these spe-cies occurred in higher abundance in monocultures, mainlyplantations of Araucaria (Table 2).
4. Discussion
Araucaria Forest is a unique ecosystem in South America (Ram-bo, 2000). The apparent homogeneity of its canopy contrastsstrongly with the high biodiversity found in its interior. Althoughsome large mammals have become rare along its fragmented dis-tribution, Araucaria Forest develops an unappreciated role in theconservation of a myriad of organisms which cannot survive inman-made habitats. Our results indicate that almost 40% of theAraucaria Forest species were found exclusively inside the primary
0
20
40
60
80
100
120
140
160
0
20
40
60
80
100
120
140
160
0
20
40
60
80
100
120
140
160
Amph
ibia
nsO
pilio
nes
Smal
l mam
mal
s Bu
tterfl
ies
Epip
hytic
fern
s Fl
atw
orm
s Li
chen
s
Bird
s G
allin
g in
sect
s Sp
ider
sW
oody
pla
nts
Fung
i
Epip
hytic
ang
iosp
erm
s
(C) Plantation of Pinus
(B) Plantation of Araucaria (A) Araucaria Forest
Amph
ibia
nsO
pilio
nes
Smal
l mam
mal
s Bu
tterfl
ies
Epip
hytic
fern
s Fl
atw
orm
s Li
chen
s
Bird
s G
allin
g in
sect
s Sp
ider
sW
oody
pla
nts
Fung
i
Epip
hytic
ang
iosp
erm
s
0
20
40
60
80
100
120
140
160 (D) Plantation of Eucalyptus
Num
ber o
f spe
cies
N
umbe
r of s
peci
es
Fig. 4. Number of species of 13 taxonomic groups recorded in Araucaria Forest (A) and plantations of Araucaria (B), Pinus (C), and Eucalyptus (D).
1214 C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219
forest, despite the ecologically friendly management applied to themonocultures. This result supports the view that Araucaria Forestbiological reserves have an irreplaceable role in biodiversity con-servation (Indrusiak and Monteiro, forthcoming).
The biodiversity conservation challenge, however, cannot be re-stricted to conservation units but must expand to man-made hab-itats which now dominate most landscapes, a principle that hasbeen exposed in the Ecosystem Principles of the Convention onBiological Diversity (Turner et al., 2001; Meffe et al., 2002; Sayerand Maginis, 2005). Some forest-associated species proved to haveenough physiological and behavioral plasticity to survive in openhabitats, such as cattle ranch, old field, and agricultural land. How-ever, most forest-associated species depend on closed environ-ments to survive. For this set of species, ecologically-managedtree monocultures can be of great conservation significance (Lin-denmayer and Franklin, 2002). In the Atlantic Forest, for instance,traditional shade cocoa plantations inserted in a landscape domi-nated by native forest hold a large fraction of the species of birds,bats, lizards, frogs, and ferns (Faria et al., 2007).
4.1. The conservation value of ecologically-managed monocultures
Although it is true that most economically-driven tree mono-cultures host a very impoverished biodiversity when comparedwith nearby preserved sites, the notion that tree monoculturesare intrinsically ‘‘green deserts” has been frequently rejected (Lin-denmayer and Hobbs, 2004; Barlow et al., 2007; Faria et al., 2007;Pardini et al., 2009). The ecologically-managed tree monoculturesof the São Francisco de Paula National Forest hosted 326 of the506 Araucaria Forest species, contributing positively to the conser-vation of Araucaria Forest biodiversity. Besides, tree monoculturesalso hosted 407 species that were not recorded in Araucaria Forest,a large fraction of that having as origin the nearby natural grass-lands, a biome that is also seriously endangered (Overbeck et al.,2007).
Plantations of the native species A. angustifolia were more effec-tive than the plantations of exotic species in hosting Araucaria For-est biodiversity. It should be highlighted that Araucaria plantationswere also important for conserving endangered species. Some

Epip
hytic
ang
iosp
erm
s
Woo
dy p
lant
sSp
ider
sG
allin
g in
sect
s Bi
rds
Lich
ens
Flat
wor
ms
Epip
hytic
fern
sBu
tterfl
ies
Smal
l mam
mal
sO
pilio
nes
Amph
ibia
ns
1
10
100
1000
10000
100000
1
10
100
1000
10000
100000
1
10
100
1000
10000
100000
1
10
100
1000
10000
100000
Num
ber o
f rec
ords
N
umbe
r of r
ecor
ds
(A) Araucaria Forest (B) Plantation of Araucaria
(C) Plantation of Pinus (D) Plantation of Eucalyptus
Fung
i
Amph
ibia
nsO
pilio
nes
Smal
l mam
mal
s Bu
tterfl
ies
Epip
hytic
fern
s Fl
atw
orm
s Li
chen
s
Gal
ling
inse
cts
Spid
ers
Woo
dy p
lant
s Fu
ngi
Epip
hytic
ang
iosp
erm
s Bi
rds
Fig. 5. Number of records of 13 taxonomic groups observed in Araucaria Forest (A) and plantations of Araucaria (B), Pinus (C), and Eucalyptus (D). The record unit is individualfor small mammals, birds, leaf-litter frogs, butterflies, galling insects, spiders, opiliones, terrestrial flatworms, woody plants, and epiphytic angiosperms. Epiphytic ferns wererecorded by the number of fronds. Lichens were recorded by the number of trees with their presence. Fungi were recorded by the number of fruit-bodies.
Table 1Richness predicted by rarefaction curves performed for 13 taxonomic groups surveyed in four habitats: Araucaria Forest (FO), Plantations of Araucaria (PA), Plantations of Pinus(PP), and Plantations of Eucalyptus (PE). N represents standard abundance used for comparison between vegetation types. Numbers between brackets represent confidenceintervals.
N FO PA PP PE
Birds 175 31.5 (28–35) 30.7 (28–32) 37.9 (36–39) 26 (26–26)Small mammals 113 7.7 (6–9) 10.3 (8–13) 7.9 (6–9) 10 (10–10)Amphibians 60 7 (7–7) 3.8 (3–4) 3.6 (3–4) 5 (5–5)Spiders 181 49.1 (45–53) 46.7 (41–52) 45.9 (40–52) 59 (59–59)Galling insects 2610 40.4 (38–43) 38.4 (36–40) 45 (45–45) 20.6 (19–21)Woody plants 505 67.1 (62–72) 62.8 (58–68) 52.5 (51–53) 59 (59–59)Lichens 51 25.2 (22–27) 29.9 (25–34) 23.3 (20–26) 30 (30–30)Opiliones 11 3.3 (1–5) 3.4 (2–5) 3.4 (2–5) 7 (7–7)Flatworms 17 8.7 (6–11) 9.9 (7–13) 8.6 (6–11) 8 (8–8)Epiphytic angiosperms 28 12.4 (9–16) 3.8 (2–6) 2.4 (1–3) 3 (3–3)Epiphytic ferns 45 5.3 (4–7) 7.1 (5–9) 4.2 (3–5) 3 (3–3)Butterflies 50 16 (16–16) 14.1 (12–17) 12.5 (10–14) 15.8 (12–19)Fungi 1918 53.8 (47–61) 59.2 (52–67) 106 (106–106) 46.1 (40–51)
C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219 1215
biological arguments can be brought about to support this claim.For instance, its contribution to the conservation of vertebrates isparticularly noteworthy, since the intense production of large
seeds during the Southern hemisphere winter provides food formany animal populations (Iob and Vieira, 2008). Besides, Araucariais a good host tree, providing adequate conditions for the fixation

0
20
40
60
80
100
Sm
all m
amm
als
But
terfl
ies
Woo
dy p
lant
s
Am
phib
ians
Bird
s
Spi
ders
Opi
lione
s
Gal
ling
inse
cts
Flat
wor
ms
Lich
ens
Epi
phyt
ic fe
rns
Fung
i
Epi
phyt
ic a
ngio
sper
ms
0
20
40
60
80
100
Sm
all m
amm
als
Opi
lione
s
Bird
s
But
terfl
ies
Spi
ders
Am
phib
ians
Woo
dy p
lant
s
Gal
ling
inse
cts
Flat
wor
ms
Fung
i
Epi
phyt
ic fe
rns
Lich
ens
Epi
phyt
ic a
ngio
sper
ms
0
20
40
60
80
100
But
terfl
ies
Sm
all m
amm
als
Am
phib
ians
Opi
lione
s
Woo
dy p
lant
s
Spi
ders
Bird
s
Flat
wor
ms
Lich
ens
Gal
ling
inse
cts
Epi
phyt
ic fe
rns
Fung
i
Epi
phyt
ic a
ngio
sper
ms
0
20
40
60
80
100
Sm
all m
amm
als
But
terfl
ies
Woo
dy p
lant
s
Bird
s
Opi
lione
s
Epi
phyt
ic fe
rns
Flat
wor
ms
Am
phib
ians
Spi
ders
Lich
ens
Gal
ling
inse
cts
Fung
i
Epi
phyt
ic a
ngio
sper
ms
(D) Plantation of Eucalyptus(C) Plantation of Pinus
(A) All plantations (B) Plantation of Araucaria
Shar
ed s
peci
es (%
) Sh
ared
spe
cies
(%)
Fig. 6. Percentage of Araucaria Forest species that are shared (black bars) with monocultures (A), and plantations of Araucaria (B), Pinus (C), and Eucalyptus (D) for 13taxonomic groups.
Table 2List of the species at risk registered in the survey and their abundance in the followingfour habitats: Araucaria Forest (FO), Plantation of Araucaria (PA), Plantation of Pinus(PP), and Plantation of Eucalyptus (PE).
Taxon Status FO PA PP PE
BirdsCampephilus robustus Endangered 0 0 3 0Amazona pretrei Vulnerable 0 4 0 0Procnias nudicollis Endangered 7 4 0 0Triclaria malachitacea Vulnerable 1 0 0 0
Woody plantsWeinmannia paulliniifolia Vulnerable 0 0 0 1Azara uruguayensis Vunerable 1 9 0 0Maytenus aquifolia Vulnerable 0 7 1 0Rhamnus sphaerosperma Vulnerable 3 3 0 6Rollinia emarginata Vulnerable 11 10 0 5Araucaria angustifolia Vulnerable 1 79 8 7Lippia ramboi Vulnerable 2 110 16 46Dicksonia sellowiana Vulnerable 1 0 0 0
Epiphytic angiospermsGriselinea ruscifolia Vulnerable 3 0 0 0Oreopanax fulvum Vulnerable 1 0 0 0Tillandsia mallemontii Endangered 0 0 10 0Tillandsia tenuefolia Vulnerable 0 13 0 0Tillandsia usneoides Vulnerable 51 0 0 0Vriesea reitzii Vulnerable 80 1 10 1
1216 C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219
of epiphytic ferns, epiphytic angiosperms, and lichens. However, itmust be take into account that the higher number of species inplantations of Araucaria could be partially explained by the olderage of such monocultures when compared with the plantationsof Pinus and Eucalyptus.
4.2. How conservation effectiveness varies among taxa
It has been frequently demonstrated that different taxa respondin different ways to habitat alteration (Lawton et al., 1998; Barlowet al., 2007; Faria et al., 2007; Pardini et al., 2009). Therefore, itcould be expected that the effectiveness of tree monocultures tohost Araucaria Forest biodiversity would vary considerably amongtaxa. Indeed, while all mammal species were recorded in themonocultures, only a quarter of the epiphytic angiosperms usedthe ecologically-managed monocultures. The general pattern sug-gests that there is a gradient of effectiveness from taxa exhibitinghigher dispersal ability and lower specific ecological requirementsto those with limited-dispersal ability and niche or habitat special-ization. Most species of the three vertebrate taxonomic groups,which are long-distance travelers and frequently have large homeranges, were recorded in the monocultures. Among the inverte-brates, most butterfly species were also recorded inside the mono-cultures, in part due to their high vagility. Most Araucaria Forest

C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219 1217
woody plants were also found regenerating inside the monocul-tures, indicating that they could in the long run regain their lostground.
On the other side of the gradient we found epiphytic angio-sperms, ferns, and lichens which have more limited dispersal abil-ity and more specialized life-histories. For these taxa, dispersalmust be directional in order to find a new phorophyte to colonize.Besides, the quality of the phorophyte is important (Benzing,1990). This seems to explain, for instance, why only three epiphyticangiosperms and three epiphytic ferns could be found in planta-tions of Eucalyptus, well known for dropping continuously the out-side layer of its trunk bark. Also, Araucaria Forest epiphytes areadapted to high air humidity levels that predominate in the regionand could be suffering with the higher levels of irradiance thatpenetrates the monocultures (Padmawate et al., 2004). Flatwormswere also highly sensitive to the replacement of Araucaria Forestby tree monocultures, probably due to its narrow resistance todrought (Kawaguti, 1932; Carbayo et al., 2001). Again, differencesin plantation age could partially explain differences in colonizationeffectiveness between taxonomic groups. Some of these groupscould be perfectly able to survive in tree monocultures once en-ough time is allowed for them to colonize.
4.3. Principles on ecological management
Ecologically-sustainable forest management in the tropics isstill in its infancy. However, we demonstrated that changes inmanagement practices can bring unsuspected high benefits interms of biodiversity conservation. The management of the SãoFrancisco de Paula National Forest differs substantially from mosteconomically-driven enterprises found in Brazil and other tropicalcountries. We discuss bellow six key principles that once followedby managers and enforced by policy makers may succeed toachieve ecologically friendly landscapes for the Atlantic Forest bio-diversity (Table 3). The relative importance of these principles onincreasing biodiversity or reducing economical outcome remainsunknown and is to be determined by ongoing adaptive manage-ment practices (Meffe et al., 2002).
4.3.1. Biogeographic suitabilityAround the globe, forests are found in places where water avail-
ability is high and soil conditions are adequate (Lomolino et al.,
Table 3Summary of some management principles that influence biodiversity conservation intree monocultures.
Management intensity
Low High
Biogeographic suitabilityBiome Forest Grassland
Landscape planningTotal extension Small LargeLandscape homogeneity Mosaic HomogeneousMatrix quality Natural Non-naturalConectivity High LowCorredors Present Absent
Stand qualityStand size Small LargeStand number Many FewStand shape Complex, linear Round, square
Stand ageRotation period Long Short
Understorey managementThinning Low frequency High frequency
Species choiceBiogeographic origin Native species Exotic species
2006). If these conditions are not met, wood productivity dependson a strong input of external resources, increasing the economicand environmental costs of the enterprise. When tree monocul-tures are established in areas naturally occupied by grasslands,for instance, a large demand for water are generated in order tosustain fast-growing trees, with the potential to exhaust the watersupplies in the long run (Bosch and Hewlett, 1982; Le Maitre et al.,2002). Geographic suitability is also a good predictor of how biodi-versity will cope with tree monocultures. For instance, many grass-land plants adapted to full light (Lambers et al., 2006) can beexpected to suffer considerably under tree monocultures but Atlan-tic Forest plant species can be expected to cope relatively well withthis anthropogenic habitat as shown by the results of this investi-gation. Therefore, the decision to establish tree monocultures inareas previously occupied by the Atlantic Forest can be consideredmore adequate than recent proposals to expand tree monoculturesin South American grasslands (Overbeck et al., 2007).
4.3.2. Landscape planningLandscape ecology provides important guidelines of how to
organize the elements of a landscape in space in order to increasethe connectivity among natural populations, facilitate the persis-tence of metapopulations, and promote gene flow (Turner et al.,2001). In the SFP National Forest, tree monocultures and AraucariaForest patches formed a complex mosaic, allowing the flow of indi-viduals, seeds and other propagules across the landscape. We canbe sure that many taxa recorded inside the tree monocultures wereactually living on them, such as plants, lichens, epiphytes, flat-worms, and fungi. Other taxa, however, such as birds, could be justusing monocultures as corridors. This is certainly an important fac-tor explaining the great number of species found inside monocul-tures. This work indicates that the design of new forestry plans canno longer be done without considering the connectivity betweenthe natural elements of the landscape with the use of corridors,stepping stones, and the quality of the intervening matrix. Addi-tionally, landscape planning can positively contribute to the con-servation of biodiversity without substantial costs for theforestry industry (Lindenmayer and Franklin, 2002).
4.3.3. Stand qualityStand traits are key factors determining the coarseness of the
landscape (Turner et al., 2001). Silvicultures based on a small num-ber of large stands produce a more homogeneous environmentwhen compared with those that are based on a large number ofsmall stands. When stands become larger, the perimeter–area ratiodecreases, increasing the difficulty of seeds, for instance, to reachthe center of the stands. For most invertebrate species with limiteddispersal ability this can lead to lower population sizes (Collingeand Palmer, 2002). The shape of the stands should also be designedto increase the perimeter-area ratio. In the SFP National Forest,stand size was typically small when compared to economically-driven tree monocultures, this probably being an important factordefining the species colonization success in these areas.
4.3.4. Stand ageIn all tree monocultures, land preparation launches a process of
site colonization that will be headed by pioneer species. The colo-nization and establishment of later successional species, however,will be constrained by the time by which the plantation is allowedto grow before the clearcut process takes place. All monoculturesof this study displayed considerable levels of biodiversity and wereallowed to grow longer than 7–10 years which is the classical rota-tion period for commercial tree monocultures. How old the planta-tion should be to achieve maximum diversity is still unknown.However, many species of woody plants, gall-inducing insects,and fungi seem to be able to colonize younger plantations. In

1218 C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219
contrast, for other taxonomic groups such as epiphytic angio-sperms and epiphytic ferns, which depend on large trees to getestablished, longer rotation periods seem to be essential to allowthe establishment of most species. In any case, the results of thiswork strongly support the idea that longer rotation periods bringbenefits for biodiversity conservation.
4.3.5. Understorey managementWhen tree monocultures are established in densities similar to
that found in natural forests, a diverse and productive understoryis allowed to get established, providing resources for herbivores,predators, pollinators, frugivores, parasitoids, and decomposers(Lindenmayer and Franklin, 2002). Indeed, some empirical studiesindicate that understory diversity is the best predictor of the diver-sity of animals in monocultures (López and Moro, 1997; Humphreyet al., 1999). In the São Francisco de Paula National Forest, althoughthe stands were established in high densities, application of thin-ning lead progressively to a lower tree density and to a complexunderstorey which was able to sustain a large fraction of the Arau-caria Forest biodiversity. In contrast, relatively old (<30 years old)Eucalyptus plantations located in the Atlantic Forest domain, butwith a clear understorey, had only eight bird species from the111 species recorded in a nearby biological reserve (Marsdenet al., 2000). Therefore, the maintenance of a complex and diverseunderstorey in monoculture plantations may be a key factor forconserving overall biodiversity.
4.3.6. Species choiceBiological invasion by exotic species is today the second major
cause of species extinction in the world (Meffe and Carroll, 1994;Millennium Ecosystem Assessment, 2005). By spatially escapingtheir natural enemies and finding adequate resources many exoticspecies succeed to reach high population growth rates and expandquickly their geographic range causing strong direct and indirecteffects in natural communities (Elton, 1958; Davis et al., 2000;Sax et al., 2005). Unfortunately, most trees used by the forestryindustry in Brazil are exotic species despite the fact that the coun-try possesses the richest flora of the world (Giullietti et al., 2005).Eucalyptus was introduced from Australia while Pinus came fromEurope and North America. The invader potential of Pinus has beenverified in grasslands, savannas (Brazilian cerrado), coastal dunesand other open habitats in almost all countries in South Americawhere it was introduced for commercial use (Richardson and Hig-gins, 1998; Ziller and Galvão, 2002; Zalba and Villamil, 2002; GISP,2006). Therefore, in principle, the use of exotic species should begradually abandoned.
The use of exotic species is frequently justified in terms of pro-ductivity, but few studies with native species have being made tosupport their supremacy. There is an unattended demand for Bra-zilian research centers and universities to allocate resources to cre-ate new forestry technology based on native species. In southernBrazil, the native tree A. angustifolia is an important alternativeto the exotic cultures. It is a magnificent tree which produces astrait, uniform and cylindrical trunk much appreciated by thewood industry. Twenty years old stands accumulate 254 tonnes/ha of biomass and necromass, corresponding to 108 tonnes/ha ofcarbon (Sanquetta, forthcoming). Nowadays, plantations of A.angustifolia probably extend to more than 20,000 ha, but its usehas the potential to become much more widespread in southernBrazil (Sanquetta, forthcoming).
5. Conclusion
The Brazilian forestry industry adopts management practicesthat has been successful in generate outstanding economic reve-
nues when compared to other countries (Bracelpa, 2007). This isachieved, however, under a relatively high cost in terms of biodi-versity conservation. Our results indicate that monocultures arenot intrinsically poor in terms of biodiversity and that by movingtowards ecologically-sustainable management practices, the for-estry industry can make a great contribution to the conservationof the Atlantic Forest.
Acknowledgements
This project was financially supported by the Conselho Nacionalde Desenvolvimento Científico e Tecnológico (CNPq: 479223/2006–8) and by the Universidade do Vale do Rio dos Sinos (UNISI-NOS). The authors would like to thank CNPq for research fellow-ships (Fonseca, C.R., Ganade, G, Brescovit, A.D., Hartz, S.M., Leal-Zanchet, A.M., Marcelli, M.P., Putzke, J., and Vieira, E.M.), graduatestudenships (Baldissera, R.), and undergraduate studenships (Boel-ter, C.R., Joner, F., Mesquita, A.S., Paz, C.P.); the Coordenação deAperfeiçoamento de Pessoal de Nível Superior (CAPES) for graduatestudentships (Fleck, T., and Fonseca, V.S.); the Fundação de Amparoà Pesquisa do Rio Grande do Sul (FAPERGS) for undergraduate stu-dentship (Vergara, M.); UNISINOS for undergraduate studentships(Becker, C.G., and Miranda, L.). This manuscript was greatly im-proved by two anonymous reviewers. Thanks to Artur Soligo andEdenice Brandão Avila de Souza, from the São Francisco de PaulaNational Forest staff, for their longstanding support. We are alsograteful to a number of volunteers that helped in data collection.
References
Bacha, C.J.C., Barros, A.L.M.B., 2004. Reflorestamento no Brasil: evolução recente eperspectivas para o futuro. Scientia Forestalis 66, 191–203.
Backes, A., 1999. Condicionamento climático e distribuição geográfica de Araucariaangustifolia (Bertol.) Kuntze no Brasil. Pesquisas – Botânica 49, 31–51.
Baptista, S.R., Rudel, T.K., 2006. A re-emerging Atlantic Forest? Urbanization,industrialization and the forest transition in Santa Catarina, southern Brazil.Environmental Conservation 33, 195–202.
Barlow, J., Gardner, T.A., Araujo, I.S., Ávila-Pires, T.C., Bonaldo, A.B., Costa, J.E.,Esposito, M.C., Ferreira, L.V., Hawes, J., Hernandez, M.I.M., Hoogmoed, M.S.,Leite, R.N., Lo-Man-Hung, N.F., Malcolm, J.R., Martins, M.B., Mestre, L.A.M.,Miranda-Santos, R., Nunes-Gutjahr, A.L., Overal, W.L., Parry, L., Peters, S.L.,Ribeiro-Junior, M.A., da Silva, M.N.F., da Silva Motta, C., Peres, C.A., 2007.Quantifying the biodiversity value of tropical primary, secondary, andplantation forests. Proceedings of the National Academy of Science 104,18555–18560.
Benzing, D.H., 1990. Vascular Epiphytes. Cambridge University Press, Cambridge.Bosch, J.M., Hewlett, J.D., 1982. A review of catchment experiments to determine
the effect of vegetation changes on water yield and evapotranspiration. Journalof Hydrology 55, 3–23.
Bracelpa, 2007. Setor de Papel e Celulose. <www.bracelpa.org.br> (accessed30.01.08).
Carbayo, F., Leal-Zanchet, A.M., Vieira, E.M., 2001. Land planarians (Platyhelminthes,Tricladida, Terrícola) as indicators of man-induced disturbance in SouthBrazilian rainforest. Belgian Journal of Zoology 131, 223–224.
Collinge, S.K., Palmer, T.M., 2002. The influence of patch shape and boundarycontrast on insect response to fragmentation in California grasslands.Landscape Ecology 17, 647–656.
Davis, M.A., Grime, J.P., Thompson, K., 2000. Fluctuating resources in plantcommunities: a general theory of invasibility. Journal of Ecology 88, 528–534.
Elton, C.S., 1958. The Ecology of Invasions by Animals and Plants. Chicago UniversityPress, Chicago.
Faria, D., Paciencia, M.B.L., Dixo, M., Laps, R.R., Baungarten, J., 2007. Ferns, frogs,lizards, birds and bats in forest fragments and shade cacao plantations in twocontrasting landscapes in Atlantic forest, Brazil. Biodiversity and Conservation16, 2335–2357.
Fonseca, G.A.B., 1985. The vanishing Brazilian Atlantic Forest. BiologicalConservation 34, 17–34.
Galindo-Leal, C., Câmara, I.G., 2003. Atlantic Forest hotspot status: an overview. In:Galindo-Leal, C., Câmara, I.G. (Eds.), The Atlantic Forest of South America:Biodiversity Status, Threats, and Outlook. Island Press, Washington, DC, pp. 3–11.
GISP, 2006. Sudamérica Invadida: el creciente peligro de las especies exóticasinvasoras. Programa Mundial sobre Especies Invasoras. Cape Town, SouthAfrica.
Giullietti, A.M., Harley, R.M., Queiroz, L.P., Wanderley, M.G.L., Van den Berg, C., 2005.Biodiversity and Conservation of Plants in Brazil. Conservation Biology 19, 632–639.

C.R. Fonseca et al. / Biological Conservation 142 (2009) 1209–1219 1219
Gotelli, N.J., Entsminger, G.L., 2001. EcoSim: Null Models Software for Ecology.Version 7.0. Acquired Intelligence Inc. & Kesey-Bear. <http://homepages.together.net/gentsmin/ecosim.htm>.
Guerra, M.P., Silveira, V., Reis, M.S., Schneider, L., 2002. Exploração, manejo econservação de araucária (Araucaria angustifolia). In: Simões, L.L., Lino, C.F.(Eds.), Sustentável Mata Atlântica: exploração de seus recursos florestais. Senac,São Paulo, Brazil, pp. 85–101.
Hartley, M.J., 2002. Rationale and methods for conserving biodiversity in plantationforests. Forest Ecology and Management 155, 81–95.
Humphrey, J.W., Hawes, C., Pearce, A.J., Ferris-Kaan, R., Jukes, M.R., 1999.Relationship between insect diversity and habitat characteristics in plantationforests. Forest Ecology and Management 113, 11–21.
Hurlbert, S.H., 1971. The nonconcept of species diversity: a critique and alternativeparameters. Ecology 52, 577–585.
Indrusiak, C., Monteiro, S.A., forthcoming. Unidades de conservação na área dedistribuição da Araucária. In: Fonseca, C.R., Souza, A.F., Leal-Zanchet, A.M.,Dutra, T., Backes, A., Ganade, G. (Eds.), Floresta com Araucária: Ecologia,Conservação e Desenvolvimento Sustentável. Holos Editora, Ribeirão Preto,Brazil.
Iob, G., Vieira, E.M., 2008. Seed predation of Araucaria angustifolia (Araucariaceae) inthe Brazilian Araucaria Forest: influence of deposition site and comparative roleof small and ‘large’ mammals. Plant Ecology 198, 185–196.
Kawaguti, S., 1932. On the physiology of land planarians. III. The problems ofdesiccation. Memoirs of the Faculty of Science and Agriculture 7, 39–55.
Koch, Z., Corrêa, M.C., 2002. Araucária: A Floresta do Brasil Meridional. OlharBrasileiro Editora, Curitiba, Brazil.
Lambers, H., Chapim III, F.S., Pons, T.L., 2006. Plant Physiological Ecology. Springer,New York.
Lawton, J.H., Bignell, D.E., Bolton, B., Bloemers, G.F., Eggleton, P., Hammond, P.M.,Hodda, M., Holt, R.D., Larsenk, T.B., Mawdsley, N.A., Stork, N.E., Srivastava, D.S.,Watt, A.D., 1998. Biodiversity inventories, indicator taxa and the effects ofhabitat modification in tropical forest. Nature 391, 72–76.
Le Maitre, D.C., van Wilgen, B.W., Gelderblom, C.M., Bailey, C., Chapman, R.A., Nel,J.A., 2002. Invasive alien trees and water resources in South Africa: case studieson costs and benefits of management. Forest Ecology and Management 160,143–159.
Lindenmayer, D.B., Franklin, J.F., 2002. Conserving Forest Biodiversity: AComprehensive Multiscaled Approach. Island Press, Washington.
Lindenmayer, D.B., Hobbs, R.J., 2004. Fauna conservation in Australian plantationforest – a review. Biological Conservation 119, 151–168.
Lomolino, M.V., Riddle, B.R., Brown, J.H., 2006. Biogeography. Sinauer, Sunderland,Massachusetts.
López, G., Moro, M.J., 1997. Birds of Aleppo pine plantations in southeast Spain inrelation to vegetation composition and structure. Journal of Applied Ecology 34,1257–1272.
Marsden, S.J., Whiffin, M., Galetti, M., 2000. Bird diversity and abundance in forestfragments and Eucalyptus plantations around an Atlantic forest reserve, Brazil.Biodiversity and Conservation 10, 737–751.
Medeiros, J.D., Gonçalves, M.A., Prochnow, M., Schäffer, W.B., 2004. Floresta comAraucárias: um símbolo da Mata Atlântica a ser salvo da extinção. Apremavi,Brazil.
Meffe, G.K., Carroll, C.R., 1994. Principles of Conservation Biology. SinauerAssociates, Sunderland, Massachusetts.
Meffe, G.K., Nielsen, L.A., Knight, R.L., Schenborn, D.A., 2002. EcosystemManagement: Adaptive, Community-based Conservation. Island Press,Washington.
Millennium Ecosystem Assessment, 2005. Ecosystems and Human Well-Being:Biodiversity Synthesis. World Resources Institute, Washington, DC.
Mittermeier, R.A., Gil, P.R., Hoffmann, M., Pilgrim, J., Brooks, T., Mittermeier, C.G.,Lamoreaux, J., Fonseca, G.A.B., 2004. Hotspots Revisited: Earth’s BiologicallyRichest and Most Endangered Terrestrial Ecoregions. Cemex & University ofChicago Press, Chicago.
Overbeck, G.E., Müller, S.C., Fidelis, A., Pfadenhauer, J., Pillar, V.D., Blanco, C.C.,Boldrini, I.I., Both, R., Forneck, E.D., 2007. Brazil’s neglected biome: the SouthBrazilian Campos. Perspectives in Plant Ecology, Evolution and Systematics 9,101–116.
Padmawate, R., Quereshi, Q., Rawat, G.S., 2004. Effects of logging on vascularepiphytes diversity in a moist lowland of Eastern Himalaya, India. BiologicalConservation 119, 81–92.
Pardini, R., Faria, D., Accacio, G.M., Laps, R.R., Mariano-Neto, E., Paciencia, M.L.B.,Dixo, M., Baumgarten, J., 2009. The challenge of maintaining Atlantic forestbiodiversity: a multi-taxa conservation assessment of specialist and generalistspecies in an agro-forestry mosaic in southern Bahia. Biological Conservation142, 1178–1190.
Rambo, B., 2000. A Fisionomia do Rio Grande do Sul: Ensaio de Monografia Natural,third ed. Editora Unisinos, São Leopoldo, Brazil.
Ribeiro, M.C., Metzger, J.P., Martensen, A.C., Ponzoni, F., Hirota, M., 2009. BrazilianAtlantic forest: how much is left and how is the remaining forest distributed?Implications for conservation. Biological Conservation 142, 1141–1153.
Richardson, D.M., Higgins, S.I., 1998. Pines as invaders in the southern hemisphere.In: Richardson, D.M. (Ed.), Ecology and Biogeography of Pinus. CambridgeUniversity Press, Cambridge, pp. 450–473.
Sanders, H., 1968. Marine benthic diversity: a comparative study. The AmericanNaturalist 102, 243–282.
Sanquetta, C.R., forthcoming. Fixação de carbono em Floresta com Araucaria. In:Fonseca, C.R., Souza, A.F., Leal-Zanchet, A.M., Dutra, T., Backes, A., Ganade, G.(Eds.) Floresta com Araucária: Ecologia, Conservação e DesenvolvimentoSustentável. Holus Editora, Ribeirão Preto, Brazil.
Sayer, J., Maginis, S., 2005. Forest in landscapes: ecosystem approaches tosustainability. Earthscan, London.
Sax, D.F., Stachowitz, J.J., Gaines, S.D., 2005. Species Invasions: Insights into Ecology,Evolution, and Biogeography. Sinauer, Sunderland, Massachusetts.
Tabarelli, M., Pinto, L.P., Silva, J.M.C., Hirota, M., Bede, L., 2005. Challenges andopportunities for biodiversity conservation in the Brazilian Atlantic Forest.Conservation Biology 19, 695–700.
Turner, M.G., Gardner, R.H., O’Neill, R.V., 2001. Landscape Ecology in Theory andPractice. Patterns and Processes. Springer-Verlag, New York.
Zalba, S.M., Villamil, C.B., 2002. Invasion of woody plants in relictual nativegrasslands. Biological Invasions 4, 55–72.
Ziller, S.R., Galvão, F.A., 2002. Degradação da estepe gramíneo-lenhosa noParaná por contaminação biológica de Pinus ellioti e P. taeda. Forest 32,41–47.
Related Documents











