1042 Mycologia, 96(5), 2004, pp. 1042–1058. q 2004 by The Mycological Society of America, Lawrence, KS 66044-8897 Toward a better understanding of the infrageneric relationships in Cortinarius (Agaricales, Basidiomycota) Ursula Peintner 1 University Innsbruck, Institute of Microbiology, Technikerstr. 25, 6020 Innsbruck, Austria Jean-Marc Moncalvo Centre for Biodiversity and Conservation Biology, Royal Ontario Museum, 100 Queen’s Park, Toronto, Ontario, M5S 2C6 Rytas Vilgalys Department of Biology, Duke University, Durham, North Carolina 27708 Abstract: Research on the molecular systematics of Cortinarius, a species-rich mushroom genus with nearly global distribution, is just beginning. The pre- sent study explores infrageneric relationships using rDNA ITS and LSU sequence data. One large dataset of 132 rDNA ITS sequences and one combined da- taset with 54 rDNA ITS and LSU sequences were gen- erated. Hebeloma was used as outgroup. Bayesian analyses and maximum-likelihood (ML) analyses were carried out. Bayesian phylogenetic inference performed equally well or better than ML, especially in large datasets. The phylogenetic analysis of the combined dataset with species representing all cur- rently recognized subgenera recovered seven well- supported clades (Bayesian posterior probabilities BPP . 90%). These major clades are: /Myxacium s.l., /subg. Cortinarius, the /phlegmacioid clade (includ- ing the subclades /Phlegmacium and /Delibuti), the /calochroid clade (/Calochroi, /Ochroleuci and /Allutus), the /telamonioid clade (/Telamonia, /Or- ellani, /Anomali), /Dermocybe s.l. and /Myxotela- monia. Our results show that Cortinarius consists of many lineages, but the relationships among these clades could not be elucidated. On one hand, the low divergence in rDNA sequences can be held re- sponsible for this; on the other hand, taxon sampling is problematic in Cortinarius phylogeny. Because of the incredibly high diversity (;2000 Cortinarius spe- cies), our sampling included ,5% of the known spe- cies. By choosing type species of subgenera and sec- tions, our sampling is strongly biased toward North- ern Hemisphere taxa. More extensive taxon sam- pling, especially of species from the Southern Accepted for publication March 1, 2004. 1 Corresponding author. E-mail: [email protected] Hemisphere, is essential to resolve the phylogeny of this important genus of ectomycorrhizal fungi. Key words: Bayesian analysis, ITS phylogeny, max- imum-likelihood analysis, taxon sampling INTRODUCTION The genus Cortinarius Fr. is one of the largest genera of gilled basidiomycete fungi. The CABI Bioscience and CBS Database of Fungal Names (http:// www.indexfungorum.org/Names/NAMES.ASP) lists 3685 Cortinarius epithets for about 2000 species (Kirk et al 2001). These characters delimit Cortinar- ius taxa: (i) a rusty brown spore print and warty- rough spore ornamentation, (ii) spores lacking a germ pore or loosening perisporium, (iii) and a cor- tinate inner veil. Cortinarius spp. form mycorrhizal associations with ectotrophic trees (Singer 1986). Macroscopically, members of this genus are highly variable: their habit varies from mycenoid to tricho- lomatoid; pilei can be dry, squamose, silky, fibrillose or viscid; the outer veil can vary from very fugacious to persistently belt-like or entirely mucilaginous; and fruit bodies can be colorful or uniformly dull brown. The subdivision of Cortinarius into subgeneric units causes problems because of the overall consid- erable morphological variation in this genus: differ- ent manifestations of each character have been ob- served between, and sometimes even within, species. This difficulty in defining character states, as well as the different weights assigned to such morphological characters by different taxonomists, has been the largest obstacle to the establishment of a broadly ac- ceptable Cortinarius classification. The many contro- versies (Ku ¨hner and Romagnesi 1953, Orton 1958, Ku ¨hner 1980, Moser 1983, Moser in Singer 1986, Melot 1990, Moe ¨nne-Loccoz et al 1990) have resulted in a taxonomic chaos and indicate that morphology alone is insufficient for recognizing natural units in this group of fungi. After the discovery of striking pigments and sec- ondary compounds in Dermocybe and Leprocybe (Ko- panski et al 1982, Steglich and Oertel 1984, Arnold et al 1987, Gill and Steglich 1987, Arnold 1993, Gill 1995a, b), Cortinarius taxonomists had great expec- tations for chemotaxonomy. These hopes have been fulfilled only partly. Molecular phylogenetics is now

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1042
Mycologia, 96(5), 2004, pp. 1042–1058.q 2004 by The Mycological Society of America, Lawrence, KS 66044-8897
Toward a better understanding of the infrageneric relationships in Cortinarius(Agaricales, Basidiomycota)
Ursula Peintner1
University Innsbruck, Institute of Microbiology,Technikerstr. 25, 6020 Innsbruck, Austria
Jean-Marc MoncalvoCentre for Biodiversity and Conservation Biology,Royal Ontario Museum, 100 Queen’s Park, Toronto,Ontario, M5S 2C6
Rytas VilgalysDepartment of Biology, Duke University, Durham,North Carolina 27708
Abstract: Research on the molecular systematics ofCortinarius, a species-rich mushroom genus withnearly global distribution, is just beginning. The pre-sent study explores infrageneric relationships usingrDNA ITS and LSU sequence data. One large datasetof 132 rDNA ITS sequences and one combined da-taset with 54 rDNA ITS and LSU sequences were gen-erated. Hebeloma was used as outgroup. Bayesiananalyses and maximum-likelihood (ML) analyseswere carried out. Bayesian phylogenetic inferenceperformed equally well or better than ML, especiallyin large datasets. The phylogenetic analysis of thecombined dataset with species representing all cur-rently recognized subgenera recovered seven well-supported clades (Bayesian posterior probabilitiesBPP . 90%). These major clades are: /Myxacium s.l.,/subg. Cortinarius, the /phlegmacioid clade (includ-ing the subclades /Phlegmacium and /Delibuti), the/calochroid clade (/Calochroi, /Ochroleuci and/Allutus), the /telamonioid clade (/Telamonia, /Or-ellani, /Anomali), /Dermocybe s.l. and /Myxotela-monia. Our results show that Cortinarius consists ofmany lineages, but the relationships among theseclades could not be elucidated. On one hand, thelow divergence in rDNA sequences can be held re-sponsible for this; on the other hand, taxon samplingis problematic in Cortinarius phylogeny. Because ofthe incredibly high diversity (;2000 Cortinarius spe-cies), our sampling included ,5% of the known spe-cies. By choosing type species of subgenera and sec-tions, our sampling is strongly biased toward North-ern Hemisphere taxa. More extensive taxon sam-pling, especially of species from the Southern
Accepted for publication March 1, 2004.1 Corresponding author. E-mail: [email protected]
Hemisphere, is essential to resolve the phylogeny ofthis important genus of ectomycorrhizal fungi.
Key words: Bayesian analysis, ITS phylogeny, max-imum-likelihood analysis, taxon sampling
INTRODUCTION
The genus Cortinarius Fr. is one of the largest generaof gilled basidiomycete fungi. The CABI Bioscienceand CBS Database of Fungal Names (http://www.indexfungorum.org/Names/NAMES.ASP) lists3685 Cortinarius epithets for about 2000 species(Kirk et al 2001). These characters delimit Cortinar-ius taxa: (i) a rusty brown spore print and warty-rough spore ornamentation, (ii) spores lacking agerm pore or loosening perisporium, (iii) and a cor-tinate inner veil. Cortinarius spp. form mycorrhizalassociations with ectotrophic trees (Singer 1986).Macroscopically, members of this genus are highlyvariable: their habit varies from mycenoid to tricho-lomatoid; pilei can be dry, squamose, silky, fibrilloseor viscid; the outer veil can vary from very fugaciousto persistently belt-like or entirely mucilaginous; andfruit bodies can be colorful or uniformly dull brown.
The subdivision of Cortinarius into subgenericunits causes problems because of the overall consid-erable morphological variation in this genus: differ-ent manifestations of each character have been ob-served between, and sometimes even within, species.This difficulty in defining character states, as well asthe different weights assigned to such morphologicalcharacters by different taxonomists, has been thelargest obstacle to the establishment of a broadly ac-ceptable Cortinarius classification. The many contro-versies (Kuhner and Romagnesi 1953, Orton 1958,Kuhner 1980, Moser 1983, Moser in Singer 1986,Melot 1990, Moenne-Loccoz et al 1990) have resultedin a taxonomic chaos and indicate that morphologyalone is insufficient for recognizing natural units inthis group of fungi.
After the discovery of striking pigments and sec-ondary compounds in Dermocybe and Leprocybe (Ko-panski et al 1982, Steglich and Oertel 1984, Arnoldet al 1987, Gill and Steglich 1987, Arnold 1993, Gill1995a, b), Cortinarius taxonomists had great expec-tations for chemotaxonomy. These hopes have beenfulfilled only partly. Molecular phylogenetics is now

1043PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
regarded as more promising for studying the evolu-tion in this complex genus. Earlier molecular inves-tigations of Cortinarius, based mainly on sequencesof the rDNA internal transcribed spacer (ITS) or therDNA large subunit (LSU), have implied that Corti-narius is monophyletic (Liu et al 1997, Høiland andHolst-Jensen 2000, Moncalvo et al 2000, Peintner etal 2001). But the monophyly of all currently definedlarger subgenera (.10 species), namely Dermocybe,Leprocybe, Myxacium, Phlegmacium, Sericeocybe and Te-lamonia has been rejected (Liu et al 1997, Chamberset al 1999, Høiland and Holst-Jensen 2000, Seidl2000, Peintner et al 2002c, Garnica et al 2003b). Ourown earlier phylogenetic studies demonstrated thatthe Cortinarius clade also included the three mor-phologically similar genera, Rozites, Cuphocybe andRapacea, as well as the sequestrate genera Thaxtero-gaster, Protoglossum and Quadrispora (Peintner et al2001, 2002).
The aim of the present study is to gain a betterunderstanding of the phylogenetic relationships inCortinarius. The effect of taxon sampling versus char-acter sampling is examined and discussed briefly. Ourreconstruction of phylogenetic relationships withinCortinarius is based on a large (132 taxa) ITS dataset.An additional rDNA dataset with 54 combined ITSand LSU sequences of representative taxa also wasused for this purpose. Both datasets were analyzed us-ing two analytical approaches: maximum likelihood(ML) and Bayesian inference of phylogeny.
MATERIALS AND METHODS
Taxonomic sampling.—Representatives of all subgenera andsections of Cortinarius as proposed by Moser (Singer 1986)were sampled, including a broad selection of diverse Cor-tinarius taxa (including Dermocybe, Rozites, Cuphocybe andThaxterogaster) from the Southern Hemisphere. Type spec-imens or specimens collected near the type locality wereused whenever possible. Species interpretations follow Mos-er (1960), Moser and Horak (1975) and Moser (1983). Twosampling strategies were followed: (i) 132 ITS sequenceswere used to produce a large, taxon-rich phylogeny, withrepresentative species selected based on traditional taxon-omy; and (ii) LSU sequences were sampled for represen-tative species from each terminal clade recovered by the ITSphylogeny.
The genus Hebeloma was chosen as outgroup to root theCortinarius phylogenies, because earlier studies have shownHebeloma to be a possible sister group (Peintner et al 2001,Moncalvo et al 2002). Sampled taxa, voucher specimen,geographical locations and Genbank accession numbers arelisted in TABLE I. Species names follow the nomenclatureof Garnier (1991). Recently recognized Cortinarius namesare used for former Thaxterogaster, Rozites, Cuphocybe andRapacea epithets (Peintner et al 2002a, b, Kuhnert andPeintner 2003). To distinguish between clade names and
formal taxonomic epithets, clade names are not italicizedand preceded with the symbol ‘‘/’’ (Moncalvo et al 2002).
Molecular techniques.—DNA isolation and sequencing pro-tocols outlined in Peintner et al (2001) were followed. Prim-ers used for PCR amplification and sequencing of the ITSregion were ITS1, ITS1F, ITS2, ITS3 and ITS4 (White et al1990, Gardes and Bruns 1993). The 59 end of the nLSUgene was amplified with either LRØR –LR5 or LRØR –LR7primer pairs. Sequencing primers were LRØR, LR3, LR3R,LR5 and LR16 (Vilgalys and Sun 1994, Hopple and Vilgalys1999, Moncalvo et al 2000). Amplification protocols fol-lowed conditions described by Vilgalys and Hester (1990).
Phylogenetic analyses.—Sequences were aligned manually andregions of ambiguous alignment were excluded from theanalyses. Gaps were treated as missing data. The programModeltest version 3.06 (Posada and Crandall 1998) was usedto choose the best-fit model of DNA substitution for eachdataset, as determined from the Akaike information criterion(AIC). Maximum-likelihood (ML) analyses were carried outwith the computer program PAUP* 4.0b10 (Swofford 2001)and conducted with ‘‘asis’’ addition sequence and TBRbranch swapping. Branch support was assessed by bootstrap-ping with simple taxon addition, NNI, with 100 replicates,five trees held at each step (Salamin et al 2003).
The Bayesian approach to phylogenetics allows simulta-neous estimation of the uncertainty associated with any pa-rameter within a phylogenetic model (e.g., topology) by us-ing posterior probabilities. For these complex estimationproblems, Markov Chain Monte Carlo (MCMC; Largentand Simon 1999) methods were used to estimate the pos-terior distribution of parameters. Bayesian analysis was car-ried out with the computer program MrBayes 2.01 (Huel-senbeck and Ronquist 2001). Random trees were used asthe starting point. Sample frequency and print frequencyoccurred once every 100 generations. To improve mixingof the chain, four simultaneous MCMC chains were run.Branch lengths of the trees were saved. For those trees thatwere sampled after the process had reached stationarity, a50% majority-rule consensus tree was computed withPAUP* to get estimates for posterior probabilities. Bayesianconsensus phylograms were calculated with MrBayes. Bayes-ian analysis was carried out three times to test the indepen-dence of the results from topological priors. Clades withBayesian Posterior Probabilities (BPP) higher than 70%were named, as far as possible, in accordance with the no-menclature of Moser (Singer 1986). Because this representsa preliminary phylogenetic classification of Cortinarius, noformal epithets are applied to clade names.
RESULTS
The alignment of the 132 ITS sequences consisted of840 characters, of which 659 were included, 218 wereconstant and 346 were parsimony informative. TheML analysis of the ITS dataset was performed underthe GTR1I1G model with a user-specified substitu-tion rate matrix and with nucleotide frequencies aslisted in TABLE II. A starting tree for branch swapping
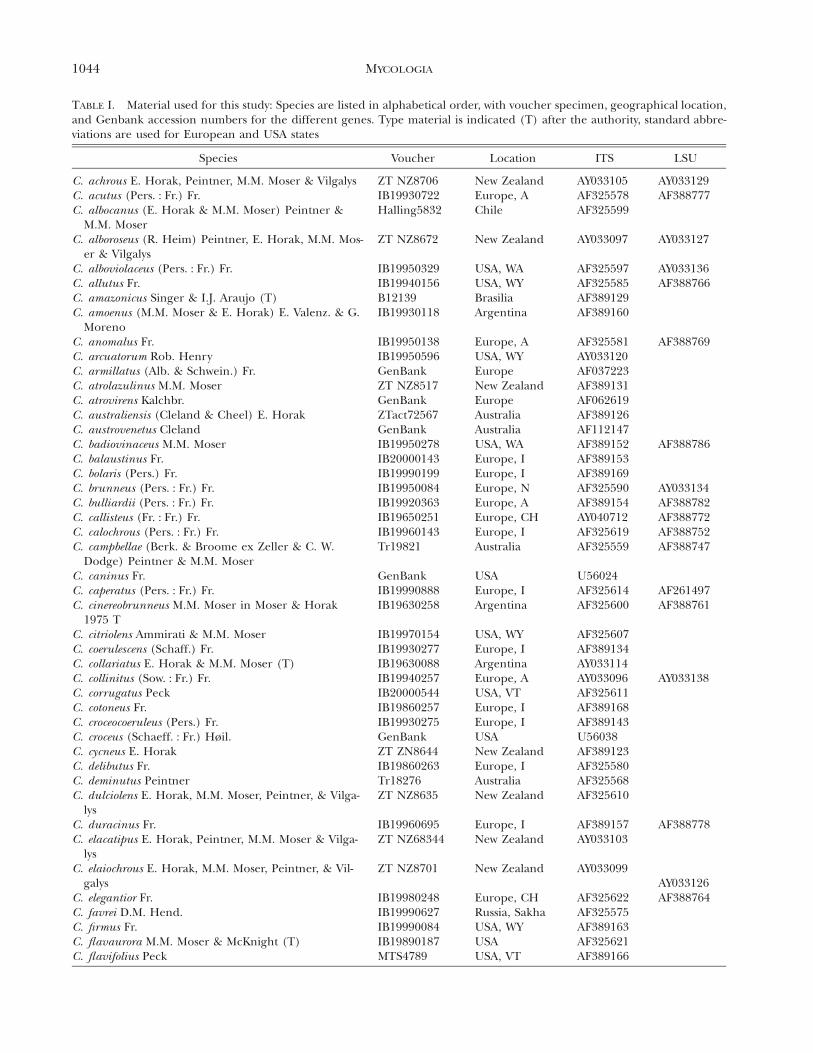
1044 MYCOLOGIA
TABLE I. Material used for this study: Species are listed in alphabetical order, with voucher specimen, geographical location,and Genbank accession numbers for the different genes. Type material is indicated (T) after the authority, standard abbre-viations are used for European and USA states
Species Voucher Location ITS LSU
C. achrous E. Horak, Peintner, M.M. Moser & VilgalysC. acutus (Pers. : Fr.) Fr.
ZT NZ8706IB19930722
New ZealandEurope, A
AY033105AF325578
AY033129AF388777
C. albocanus (E. Horak & M.M. Moser) Peintner &M.M. Moser
C. alboroseus (R. Heim) Peintner, E. Horak, M.M. Mos-er & Vilgalys
Halling5832
ZT NZ8672
Chile
New Zealand
AF325599
AY033097 AY033127
C. alboviolaceus (Pers. : Fr.) Fr.C. allutus Fr.C. amazonicus Singer & I.J. Araujo (T)
IB19950329IB19940156B12139
USA, WAUSA, WYBrasilia
AF325597AF325585AF389129
AY033136AF388766
C. amoenus (M.M. Moser & E. Horak) E. Valenz. & G.Moreno
C. anomalus Fr.C. arcuatorum Rob. HenryC. armillatus (Alb. & Schwein.) Fr.C. atrolazulinus M.M. MoserC. atrovirens Kalchbr.
IB19930118
IB19950138IB19950596GenBankZT NZ8517GenBank
Argentina
Europe, AUSA, WYEuropeNew ZealandEurope
AF389160
AF325581AY033120AF037223AF389131AF062619
AF388769
C. australiensis (Cleland & Cheel) E. HorakC. austrovenetus ClelandC. badiovinaceus M.M. Moser
ZTact72567GenBankIB19950278
AustraliaAustraliaUSA, WA
AF389126AF112147AF389152 AF388786
C. balaustinus Fr.C. bolaris (Pers.) Fr.C. brunneus (Pers. : Fr.) Fr.C. bulliardii (Pers. : Fr.) Fr.C. callisteus (Fr. : Fr.) Fr.C. calochrous (Pers. : Fr.) Fr.
IB20000143IB19990199IB19950084IB19920363IB19650251IB19960143
Europe, IEurope, IEurope, NEurope, AEurope, CHEurope, I
AF389153AF389169AF325590AF389154AY040712AF325619
AY033134AF388782AF388772AF388752
C. campbellae (Berk. & Broome ex Zeller & C. W.Dodge) Peintner & M.M. Moser
C. caninus Fr.C. caperatus (Pers. : Fr.) Fr.
Tr19821
GenBankIB19990888
Australia
USAEurope, I
AF325559
U56024AF325614
AF388747
AF261497C. cinereobrunneus M.M. Moser in Moser & Horak
1975 TC. citriolens Ammirati & M.M. MoserC. coerulescens (Schaff.) Fr.C. collariatus E. Horak & M.M. Moser (T)C. collinitus (Sow. : Fr.) Fr.C. corrugatus Peck
IB19630258
IB19970154IB19930277IB19630088IB19940257IB20000544
Argentina
USA, WYEurope, IArgentinaEurope, AUSA, VT
AF325600
AF325607AF389134AY033114AY033096AF325611
AF388761
AY033138
C. cotoneus Fr.C. croceocoeruleus (Pers.) Fr.C. croceus (Schaeff. : Fr.) Høil.C. cycneus E. HorakC. delibutus Fr.C. deminutus Peintner
IB19860257IB19930275GenBankZT ZN8644IB19860263Tr18276
Europe, IEurope, IUSANew ZealandEurope, IAustralia
AF389168AF389143U56038AF389123AF325580AF325568
C. dulciolens E. Horak, M.M. Moser, Peintner, & Vilga-lys
C. duracinus Fr.
ZT NZ8635
IB19960695
New Zealand
Europe, I
AF325610
AF389157 AF388778C. elacatipus E. Horak, Peintner, M.M. Moser & Vilga-
lysC. elaiochrous E. Horak, M.M. Moser, Peintner, & Vil-
galys
ZT NZ68344
ZT NZ8701
New Zealand
New Zealand
AY033103
AY033099AY033126
C. elegantior Fr.C. favrei D.M. Hend.C. firmus Fr.C. flavaurora M.M. Moser & McKnight (T)C. flavifolius Peck
IB19980248IB19990627IB19990084IB19890187MTS4789
Europe, CHRussia, SakhaUSA, WYUSAUSA, VT
AF325622AF325575AF389163AF325621AF389166
AF388764
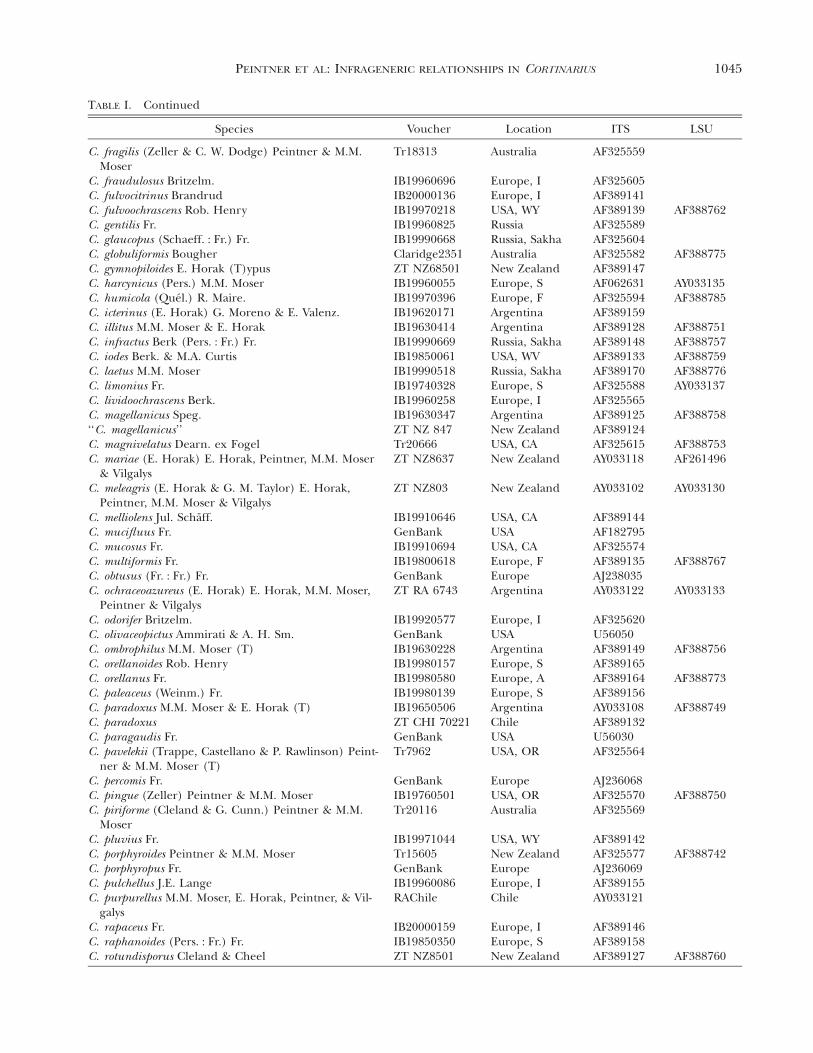
1045PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
TABLE I. Continued
Species Voucher Location ITS LSU
C. fragilis (Zeller & C. W. Dodge) Peintner & M.M.Moser
Tr18313 Australia AF325559
C. fraudulosus Britzelm.C. fulvocitrinus BrandrudC. fulvoochrascens Rob. HenryC. gentilis Fr.C. glaucopus (Schaeff. : Fr.) Fr.C. globuliformis Bougher
IB19960696IB20000136IB19970218IB19960825IB19990668Claridge2351
Europe, IEurope, IUSA, WYRussiaRussia, SakhaAustralia
AF325605AF389141AF389139AF325589AF325604AF325582
AF388762
AF388775C. gymnopiloides E. Horak (T)ypusC. harcynicus (Pers.) M.M. MoserC. humicola (Quel.) R. Maire.C. icterinus (E. Horak) G. Moreno & E. Valenz.C. illitus M.M. Moser & E. HorakC. infractus Berk (Pers. : Fr.) Fr.
ZT NZ68501IB19960055IB19970396IB19620171IB19630414IB19990669
New ZealandEurope, SEurope, FArgentinaArgentinaRussia, Sakha
AF389147AF062631AF325594AF389159AF389128AF389148
AY033135AF388785
AF388751AF388757
C. iodes Berk. & M.A. CurtisC. laetus M.M. MoserC. limonius Fr.C. lividoochrascens Berk.C. magellanicus Speg.‘‘C. magellanicus’’C. magnivelatus Dearn. ex Fogel
IB19850061IB19990518IB19740328IB19960258IB19630347ZT NZ 847Tr20666
USA, WVRussia, SakhaEurope, SEurope, IArgentinaNew ZealandUSA, CA
AF389133AF389170AF325588AF325565AF389125AF389124AF325615
AF388759AF388776AY033137
AF388758
AF388753C. mariae (E. Horak) E. Horak, Peintner, M.M. Moser
& VilgalysC. meleagris (E. Horak & G. M. Taylor) E. Horak,
Peintner, M.M. Moser & Vilgalys
ZT NZ8637
ZT NZ803
New Zealand
New Zealand
AY033118
AY033102
AF261496
AY033130
C. melliolens Jul. Schaff.C. mucifluus Fr.C. mucosus Fr.C. multiformis Fr.C. obtusus (Fr. : Fr.) Fr.
IB19910646GenBankIB19910694IB19800618GenBank
USA, CAUSAUSA, CAEurope, FEurope
AF389144AF182795AF325574AF389135AJ238035
AF388767
C. ochraceoazureus (E. Horak) E. Horak, M.M. Moser,Peintner & Vilgalys
ZT RA 6743 Argentina AY033122 AY033133
C. odorifer Britzelm.C. olivaceopictus Ammirati & A. H. Sm.C. ombrophilus M.M. Moser (T)C. orellanoides Rob. HenryC. orellanus Fr.C. paleaceus (Weinm.) Fr.C. paradoxus M.M. Moser & E. Horak (T)C. paradoxus
IB19920577GenBankIB19630228IB19980157IB19980580IB19980139IB19650506ZT CHI 70221
Europe, IUSAArgentinaEurope, SEurope, AEurope, SArgentinaChile
AF325620U56050AF389149AF389165AF389164AF389156AY033108AF389132
AF388756
AF388773
AF388749
C. paragaudis Fr.C. pavelekii (Trappe, Castellano & P. Rawlinson) Peint-
ner & M.M. Moser (T)C. percomis Fr.C. pingue (Zeller) Peintner & M.M. Moser
GenBankTr7962
GenBankIB19760501
USAUSA, OR
EuropeUSA, OR
U56030AF325564
AJ236068AF325570 AF388750
C. piriforme (Cleland & G. Cunn.) Peintner & M.M.Moser
C. pluvius Fr.C. porphyroides Peintner & M.M. MoserC. porphyropus Fr.C. pulchellus J.E. Lange
Tr20116
IB19971044Tr15605GenBankIB19960086
Australia
USA, WYNew ZealandEuropeEurope, I
AF325569
AF389142AF325577AJ236069AF389155
AF388742
C. purpurellus M.M. Moser, E. Horak, Peintner, & Vil-galys
C. rapaceus Fr.C. raphanoides (Pers. : Fr.) Fr.C. rotundisporus Cleland & Cheel
RAChile
IB20000159IB19850350ZT NZ8501
Chile
Europe, IEurope, SNew Zealand
AY033121
AF389146AF389158AF389127 AF388760

1046 MYCOLOGIA
TABLE I. Continued
Species Voucher Location ITS LSU
C. rubellus CookeC. russeoides M.M. MoserC. saginus Fr.C. salor Fr.
GenBankIB19980112IB19960705IB19940297
EuropeEurope, AEurope, IEurope, A
AJ236064AF389136AF325608AF325579
AF388768
C. sanguineus (Wulfen) Fr.C. scaurus (Fr. : Fr.) Fr.C. scaurusC. semisanguineus (Fr.) Gill.
GenBankIB19980175GenBankIB19990566
USAEurope, SEuropeRussia, Sakha
U56058AF325562AJ236070AF389150
AF388763
AF388770C. subcastanella E. Horak, Peintner, M.M. Moser & Vil-
galysC. subfoetidus A.H. SmithC. sulphurinus Quel.C. talus Fr.C. teraturgus M.M. Moser TC. tophaceus Fr.
ZT NZ 8533
IB19950326IB20000167IB19950301IB19630218IB19740174
New Zealand
USA, WAEurope, IUSA, WAArgentinaEurope, S
AY033110
AF325609AF389140AY033119AF389151AY040714
AY033130
AF388765AF388755
C. trachycystis M.M. MoserC. traganus (Fr. : Fr.) Fr.C. trivialis J.E. LangeC. variicolor (Pers. : Fr.) Fr.C. variipes Rob. HenryC. venetus (Fr.) Fr.
IB19630143GenBankGenBankGenBankIB19890457IB19970371
ArgentinaEuropeEuropeEuropeUSA, WAEurope, I
AF389145AF037224AJ236066AJ238082AF389138AF389167
AF388771
AF388774C. vespertinus Fr.C. vibratilis Fr.C. violaceus (L. : Fr.) Fr.C. violaceusCortinarius sp.Dermocybe austronanceiensis M.M. Moser
IB19840067GenBankIB19980181IB19950556ZT NZ8682IB19630363
Europe, SEuropeEurope, SUSA, WYNew ZealandArgentina
AF389137AJ238032AF325600AF389130AY033117AF389161
D. cardinalis E. HorakD. marylandensis Ammirati & A.H. Sm. nom. inval.
ZT NZ68373GenBank
New ZealandUSA
AF389162U56047
D. spendida E. HorakHebeloma fastibile (Pers. : Fr.) P. Kumm.H. mesophaeum (Pers. : Fr.) Quel.Hymenogaster remyi Dodge & Zeller & Dodge
ZT NZ920IB19940036GenBankMPM1710
New ZealandEurope, AEuropeUSA, E
AF325582AF325643AF126100AF325602
AY033139
H. sublilacinus A.H. Sm.Protoglossum luteum MasseeProtoglossum sp. 1Protoglossum sp. 2
Tr19101Tr18831Beever 996Tr15856
USA, CAAustraliaNew ZealandAustralia
AF325603AF325612AF325613AF325561
AY033128AF388743AF388743
Quadrispora oblongispora (G.W. Beaton, Pegler &T.W.K. Young) Bougher & Castellano
Tr18111 Australia AF325566 AF388746
was constructed by neighbor joining. The heuristicsearch was carried out with TBR branch swapping.The ML search was stopped after 30 d of computingtime. Seven ML trees were obtained with a –Ln like-lihood of 9811.8250.
The three Bayesian analyses of the 132 rDNA ITSsequences were run for 200 000, 2 000 000 and3 000 000 generations, respectively. For Bayesian pos-terior probabilities (BPP), a 50% majority rule con-sensus tree was computed with the overall 41 504trees sampled after the process had reached station-arity. The Bayesian consensus phylogram shown inFIG. 1 was calculated using 28 302 trees from thethird Bayesian analysis. This longest chain reached
stationarity at about 170 000 generations: the first1700 trees were discarded as ‘‘burn in’’, and 28 302were used to calculate the consensus tree usingMrBayes. The consensus phylogram is based on meanbranch lengths and posterior probabilities. Branchlengths are maximum-likelihood estimates and arescaled in terms of expected numbers of nucleotidesubstitutions per site. The posterior distribution ofparameters is listed in TABLE II.
The combined alignment of ITS and LSU sequenc-es consisted of 2413 characters, 1365 were included,857 were constant and 243 were parsimony infor-mative. The ML analysis was performed as describedabove with the GTR1G1I model. The parameters
1047PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
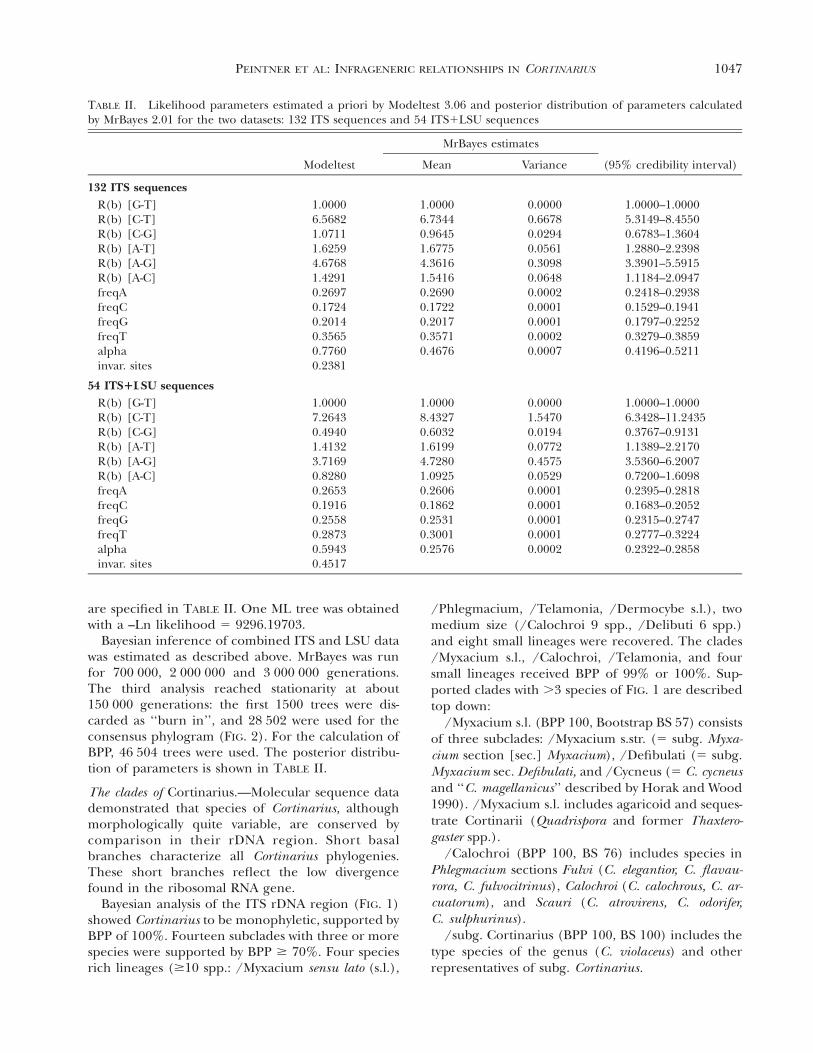
TABLE II. Likelihood parameters estimated a priori by Modeltest 3.06 and posterior distribution of parameters calculatedby MrBayes 2.01 for the two datasets: 132 ITS sequences and 54 ITS1LSU sequences
Modeltest
MrBayes estimates
Mean Variance (95% credibility interval)
132 ITS sequencesR(b) [G-T]R(b) [C-T]R(b) [C-G]R(b) [A-T]R(b) [A-G]R(b) [A-C]freqAfreqCfreqGfreqTalphainvar. sites
1.00006.56821.07111.62594.67681.42910.26970.17240.20140.35650.77600.2381
1.00006.73440.96451.67754.36161.54160.26900.17220.20170.35710.4676
0.00000.66780.02940.05610.30980.06480.00020.00010.00010.00020.0007
1.0000–1.00005.3149–8.45500.6783–1.36041.2880–2.23983.3901–5.59151.1184–2.09470.2418–0.29380.1529–0.19410.1797–0.22520.3279–0.38590.4196–0.5211
54 ITS1LSU sequencesR(b) [G-T]R(b) [C-T]R(b) [C-G]R(b) [A-T]R(b) [A-G]R(b) [A-C]freqAfreqCfreqGfreqT
1.00007.26430.49401.41323.71690.82800.26530.19160.25580.2873
1.00008.43270.60321.61994.72801.09250.26060.18620.25310.3001
0.00001.54700.01940.07720.45750.05290.00010.00010.00010.0001
1.0000–1.00006.3428–11.24350.3767–0.91311.1389–2.21703.5360–6.20070.7200–1.60980.2395–0.28180.1683–0.20520.2315–0.27470.2777–0.3224
alphainvar. sites
0.59430.4517
0.2576 0.0002 0.2322–0.2858
are specified in TABLE II. One ML tree was obtainedwith a –Ln likelihood 5 9296.19703.
Bayesian inference of combined ITS and LSU datawas estimated as described above. MrBayes was runfor 700 000, 2 000 000 and 3 000 000 generations.The third analysis reached stationarity at about150 000 generations: the first 1500 trees were dis-carded as ‘‘burn in’’, and 28 502 were used for theconsensus phylogram (FIG. 2). For the calculation ofBPP, 46 504 trees were used. The posterior distribu-tion of parameters is shown in TABLE II.
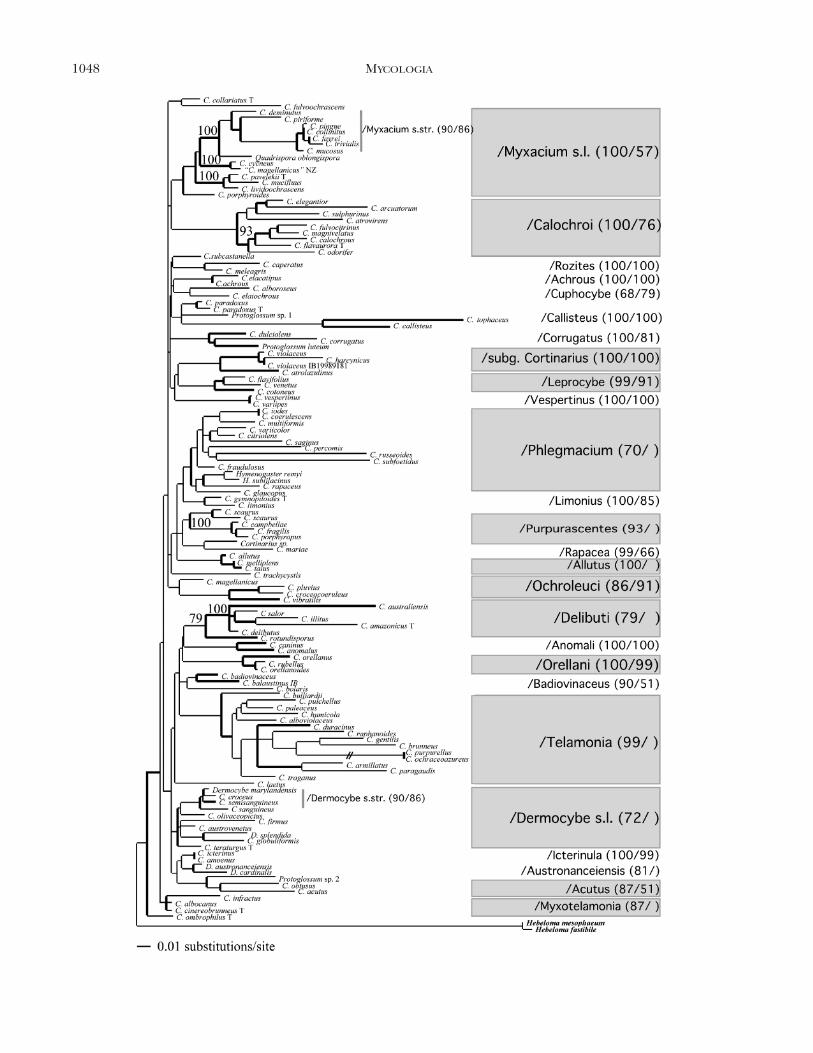
The clades of Cortinarius.—Molecular sequence datademonstrated that species of Cortinarius, althoughmorphologically quite variable, are conserved bycomparison in their rDNA region. Short basalbranches characterize all Cortinarius phylogenies.These short branches reflect the low divergencefound in the ribosomal RNA gene.
Bayesian analysis of the ITS rDNA region (FIG. 1)showed Cortinarius to be monophyletic, supported byBPP of 100%. Fourteen subclades with three or morespecies were supported by BPP $ 70%. Four speciesrich lineages ($10 spp.: /Myxacium sensu lato (s.l.),
/Phlegmacium, /Telamonia, /Dermocybe s.l.), twomedium size (/Calochroi 9 spp., /Delibuti 6 spp.)and eight small lineages were recovered. The clades/Myxacium s.l., /Calochroi, /Telamonia, and foursmall lineages received BPP of 99% or 100%. Sup-ported clades with .3 species of FIG. 1 are describedtop down:
/Myxacium s.l. (BPP 100, Bootstrap BS 57) consistsof three subclades: /Myxacium s.str. (5 subg. Myxa-cium section [sec.] Myxacium), /Defibulati (5 subg.Myxacium sec. Defibulati, and /Cycneus (5 C. cycneusand ‘‘C. magellanicus’’ described by Horak and Wood1990). /Myxacium s.l. includes agaricoid and seques-trate Cortinarii (Quadrispora and former Thaxtero-gaster spp.).
/Calochroi (BPP 100, BS 76) includes species inPhlegmacium sections Fulvi (C. elegantior, C. flavau-rora, C. fulvocitrinus), Calochroi (C. calochrous, C. ar-cuatorum), and Scauri (C. atrovirens, C. odorifer,C. sulphurinus).
/subg. Cortinarius (BPP 100, BS 100) includes thetype species of the genus (C. violaceus) and otherrepresentatives of subg. Cortinarius.
1048 MYCOLOGIA

1049PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
←
FIG. 1. Consensus phylogram of 28 302 trees resulting from the Bayesian analysis of 132 ITS sequences. Bold branchesare supported by Bayesian posterior probabilities (BPP) .70%. BPP and bootstrap (BS) values are given in brackets (BPP/BS) after the clade name. BPP for subclades are shown above branches.
/Leprocybe (BPP 99, BS 91) includes the type spe-cies of subg. Leprocybe (C. cotoneus), and morpholog-ically closely related species (C. venetus, C. flavifolius).
/Phlegmacium (BPP 70, BS , 50) is a lineage of14 species in different sections of subg. Phlegmacium:sec. Phlegmacium (C. multiformis, C. fraudulosus andC. saginus, the typus of subg. Phlegmacium), sec. Cal-ochroi (C. glaucopus), sec. Coerulescentes (C. variicolor,C. coerulescens, C. subfoetidus, C. citriolens), and sec.Scauri (C. percomis, C. russeoides). Two secotioid taxa(Hymenogaster remyi, H. sublilacinus) also belong tothis lineage.
/Purpurascentes (BPP 93, BS , 50) circumscribestaxa in subg. Phlegmacium sec. Scauri. This lineagewas recovered by tree topologies in all analytical ap-proaches but with little and varying support. A recentstudy including seven temperate species in the C.scaurus species complex demonstrated the monophy-ly of /Purpurascentes (Moser and Peintner 2002b).
/Allutus (BPP 100, BS , 50) is a lineage of speciesin Phlegmacium sec. Phlegmacium.
/Ochroleuci (BPP 86, BS 91) circumscribes speciesin subg. Myxacium sec. Ochroleuci.
/Delibuti (BPP 79, BS , 50) is a lineage of species,which have been classified in subg. Myxacium sectionsDelibuti, Cystidiferae and Archeri. When excluding C.rotundisporus, an Australian species of Myxacium, thisclade is strongly supported (BPP 96, BS 51).
/Orellani (BPP 100, BS 99) includes species ofsubg. Leprocybe sec. Orellani.
/Telamonia (BPP 99, BS , 50) represents Corti-narius spp. in subg. Telamonia, and taxa in the sub-genera Sericeocybe (C. alboviolaceus, C. traganus) andLeprocybe (C. gentilis, C. raphanoides, C. humicola), aswell as in the former genus Rozites (C. purpurellus, C.ochraceoazureus).
/Dermocybe s.l. (BPP 72, BS , 50) circumscribesDermocybe spp. from the northern and southernhemisphere. Subgenus Dermocybe is paraphyletic, as/Dermocybe s.l. also includes taxa in subgenera Cys-togenes (C. teraturgus) and Telamonia (C. firmus).Northern hemisphere taxa of Dermocybe form thewell-supported (BPP 90, BS 86) subclade /Dermo-cybe s.str. (C. croceus, C. semisanguineus, C. sangui-neus, D. marylandensis). Tree topologies indicate asister group relationship of /Dermocybe s.l. with twoclades of the Southern Hemisphere Dermocybe spp.,/Icterinula and /Austronanceiensis.
/Acutus (BPP 87, BS 51) represents species in
subg. Telamonia sec.Obtusi. The mycenoid basidiomehabit, a inflated hyphae in the mediostratum of thelamellar trama, and the presence of cystidia distin-guish species belonging to the Obtusi from other Te-lamonia spp.
/Myxotelamonia (BPP 87, BS , 50) consists of spe-cies in subg. Phlegmacium, sec. Amarescentes (C. in-fractus) and in subg. Telamonia sec. Myxotelamonia(C. cinereobrunneus). A gelatinized epicutis and gray-ish tints to the lamellae characterize both.
When comparing tree topologies obtained by theML and the Bayesian analysis of the ITS dataset, thesame larger clades are recovered. In general, BPP arehigher than bootstrap (BS) values (FIG. 1). As cladesbecome larger, BS values quickly decrease to below50%, while BPP higher than 70% are still obtained(e.g., /Calochroi /Dermocybe s.l., /Myxacium s.l.,/Phlegmacium, /Telamonia).
In the Bayesian estimation of the combined ribo-somal DNA phylogeny (ITS and LSU), 10 clades weresupported by BPP of at least 70% (FIG. 2). Also inthis dataset, BPP generally were higher than BS val-ues. Compared to the ML tree, three new relation-ships were supported (BPP . 85%): (i) the /phleg-macioid group, a lineage of /Delibuti and C. multi-fomis representing /Phlegmacium from the ITS phy-logeny; (ii) the common lineage of /Calochroi,/Ochroleuci and /Allutus, tentatively addressed asthe /calochroid group; (iii) the /telamonioid group,a common lineage of /Telamonia, C. anomalus, C.laetus and C. orellanus. Moreover, little support (BS70, BPP , 50) is obtained for a larger /Purpurascen-tes, which now also includes C. mariae representing/Rapacea from the ITS phylogeny (FIG. 1). /Der-mocybe s.l. is further supported as monophyleticgroup (BPP 92, BS 79). ML tree topologies consis-tently indicate a sister group relationship of /Der-mocybe s.l. and /Myxotelamonia.
When focusing on the phylogeny of classical sub-genera in Cortinarius (Moser in Singer 1986), we fur-ther confirm that most subgenera of Cortinarius arepolyphyletic (TABLE III); a monophyletic exception issubgenus Cortinarius. Species of subg. Dermocybe splitinto /Dermocybe s.l., /Icterinula and /Austronan-ceiensis, and monophyly of these three lineages can-not be excluded. Cortinarius species classified insubg. Leprocybe fall into the clades /Badiovinaceus,/Callisteus, /Leprocybe s.str., /Limonei, /Orellaniand /Telamonia. Myxacium spp. form the lineages

1050 MYCOLOGIA
FIG. 2. Consensus phylogram of 28 502 trees resulting from the Bayesian analysis of 54 ITS and LSU sequences. Boldbranches are supported by BPP . 70%. BPP and BS values are given in brackets (BPP/BS) after the clade name. BPP forlarger clades are shown above branches.
/Myxacium s.l., /Ochroleuci, and /Delibuti. Corti-narii classified in subg. Phlegmacium fall in the line-ages /Calochroi, /Phlegmacium, /Purpurascentes,and /Myxotelamonia. Cortinarius species classified insubg. Sericeocybe can be found in the clades /Tela-monia and /Anomali. Telamonia species are found in/Telamonia, /Dermocybe s.l., /Myxotelamonia and/Acutus.
DISCUSSION
Bayesian inference and ML analysis.—Bayesian infer-ence of phylogeny (Yang and Rannala 1997) offersadvantages over other methods in its ability to usecomplex models of evolution and in its ease of inter-
pretation and computational efficiency. Phylogeneticanalyses of multigene data, estimation of divergencetimes, testing monophyly of clades, inferring the rootof a phylogenetic tree, mapping mutations on phy-logenies, and evaluation of tree topologies have beenaddressed using Bayesian inference (Huelsenbeck etal 2002; Marvaldi et al 2002; Miller et al 2002; Nielsen2002; Thorne and Kishino 2002; Garnica et al 2003a,b). In our datasets, Bayesian phylogenetic inferenceperformed as well or better than ML. Bayesian infer-ence is more practical and much faster than ML, es-pecially when analyzing large datasets. BS values cal-culated using maximum parsimony or ML are gen-erally lower than the corresponding BPP, especiallyin larger clades (Karol et al 2001, Murphy et al 2001,

1051PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
TABLE III. Distribution of classical subgenera and/or genera (Singer 1986) into selected subclades (.2 taxa and BPP .70%) of our Cortinarius phylogenies (in alphabetical order, pp 5 pro parte, all 5 all known species)
SubcladeSG Cor-tinarius
Cysto-genes
Dermo-cybe
Lepro-cybe
Myxa-cium
Serice-ocybe
Phleg-macium
Tela-monia
Hymeno-gaster Rozites
Thaxter-ogaster
Quadri-spora
/Acutus/Allutus/Calochroi/calochroid group/subg. Cortinarius all
pp
pp
pp
pp pp
/Delibuti/Dermocybe s.l./Leprocybe
pp pppp
pppp
/Myxacium s.l./Myxotelamonia/Ochroleuci/Orellani/phlegmacioid group
pp
pp
pp
pp
pp
pp
pp
pp
pppp
all
/Phlegmacium/Purpurascentes/Rozites/Telamonia/telamonioid group
pppp
pppp
pppp
pppp
pp
pppppp
Huelsenbeck et al 2002, Miller et al 2002). The Bayes-ian method was evaluated critically and tested bycomputer simulations in two recent studies. Suzuki etal (2002), using three sets of four nucleotide se-quences, showed that BPP can be excessively liberalwhen concatenated gene sequences are used whereasBS probabilities are slightly conservative. Wilcox et al(2002), using a ML model tree of 23 species ofsnakes, evaluated the reliability of BPP and BS as astatistical confidence of interior branches. Theyshowed that as BPP underestimates less than BS, BPPis a better indicator of statistical confidence.
First consequences: the departure from old genus con-cepts.—If taxonomic recognition is granted only toclearly monophyletic groups, all taxa except the He-beloma outgroup must be regarded as members ofone large genus. Thus, Cortinarius is an extremelyramified lineage of ectomycorrhizal mushrooms witha broad range of morphological variability. The ge-nus circumscription had to be widened based on phy-logenetic and morphological data only recently;Peintner et al (2001, 2002) clearly demonstrated thatrepresentative species of Thaxterogaster, Protoglossumand Quadrispora are reduced forms of Cortinarius.Therefore their generic names were synonymized un-der Cortinarius (Peintner et al 2002a, Kuhnert andPeintner 2003). Dermocybe, Rozites, Cuphocybe and Ra-pacea had been segregated from Cortinarius based onveil characteristics and chemotaxonomy (Heim 1951,Moser 1972, Singer 1986, Horak 1999). But molecu-lar data (Liu et al 1997, Høiland and Holst-Jensen
2000, Seidl 2000, Peintner et al 2001, Peintner et al2002c) suggested that some of these diagnosticallyvaluable characters had been overweighted. The ge-nus Dermocybe was delimited by the presence of an-thraquinonic pigments, once regarded as restrictedto this ‘‘genus’’ (Moser 1972, Singer 1986). But Gilland Steglich (1987) also recorded anthraquinonesfor some Telamonia and Phlegmacium species. Ourcurrent phylogenetic analyses show that Dermocybetaxa from both the Northern and Southern hemi-spheres nest within Cortinarius, supporting Dermocybeas a synonym of Cortinarius. The nesting of Rozites,Cuphocybe and Rapacea spp. also supports synonymyof those genera with Cortinarius (Peintner et al2002c). The nomenclatural consequences proposedby Peintner et al (2002b) were based on both mor-phological and molecular data. Thus, the genus Cor-tinarius encompasses a morphologically highly vari-able group of mushrooms that includes both agaric(Cortinarius, Dermocybe, Rozites, Cuphocybe and Ra-pacea) and sequestrate (Thaxterogaster, Protoglossum,Hymenogaster pro parte and Quadrispora) forms. Allthese taxa, however, do share the typical basidiosporemorphology and ectomycorrhizal strategy.
Toward a phylogeny-based taxonomy of Cortinarius.—Subgenera are exceedingly helpful to subdivide largegenera into smaller taxonomic units; they should bepractical and reflect the hierarchy of evolutionary re-lationships. Phylogenetic research repeatedly has re-jected the monophyly of Dermocybe, Leprocybe, Myxa-cium, Phlegmacium, Sericeocybe and Telamonia (Liu et

1052 MYCOLOGIA
al 1997, Chambers et al 1999, Høiland and Holst-Jensen 2000, Seidl 2000, Peintner et al 2001, Moserand Peintner 2002, Peintner et al 2002c, Garnica etal 2003b). Dermocybe and Telamonia both are para-phyletic and need to be redefined. The morpholog-ical and chemotaxonomical characters used to defineLeprocybe and Sericeocybe (fluorescent substances orsilky to glimmering pilei) appear to have been over-weighted, thus creating artificial subgenera. The sub-genera Myxacium and Phlegmacium, previously delim-ited solely by veil characters, probably deserve sub-division into smaller, monophyletic entities. All thesesmaller and/or partly new subgenera should be de-fined using both morphological and molecular data.Chemotaxonomy alone, while useful in delimitingspecies or sections, has not proven reliable for seg-regating subgenera (e.g., Dermocybe, Leprocybe). De-limitation of groups based on only a few morpholog-ical characters likewise has proven untrustworthy(e.g., Rozites, Phlegmacium, Myxacium).
The existing taxonomy needs to be re-evaluated toreflect an accurate phylogeny of Cortinarius. We baseour Cortinarius phylogeny on the results of the Bayes-ian analysis of the two datasets (FIGS. 1, 2). Becausethe shortness of the basal branches greatly compli-cates generation of a Cortinarius phylogeny, for now,we offer a synthesis of the two Bayesian consensustrees as our best hypothesis.
Several larger monophyletic groups can be distin-guished within Cortinarius (e.g., /Myxacium s.l., te-lamonioid group, /Dermocybe s.l., /Calochroi; FIGS.1, 2), but our results do not support a split of Corti-narius into two distinct lineages as suggested byHøiland and Holst-Jensen (2000). The many lineagesuncovered indicate that this species-rich genus re-quires more subgenera than are defined currently.Each of the well-supported clades discussed belowmay merit the rank of a subgenus, but we were un-able to elucidate evolutionary relationships amongthese Cortinarius subclades. It is possible that the stillcomparatively small sample size is fundamentally re-sponsible for this lack of resolution. This in turn sug-gests the need for combining morphological and mo-lecular studies and including more species on a glob-al scale before proposing any major taxonomicchanges. Below we present the potentially diagnosticcharacters for each well-supported lineage revealedin our analyses.
The three distinct lineages of Myxacium species. Cor-tinarius subg. Myxacium is polyphyletic (Høiland andHolst Jensen 2000, Peintner et al 2001, Seidl 2000).Because C. collinitus is the type species of the sub-genus, subg. Myxacium should be restricted to taxaof /Myxacium s.l. Our results indicate that the Och-roleuci and Delibuti, previously placed in Myxacium,
derived from different lineages of Phegmacium spp.and thus are not closely related to /Myxacium s.l.More extensive taxon sampling is urgently needed tounderstand the evolution of the striking, strongly de-veloped, viscid veils in Cortinarius.
Clade /Myxacium s.l.: Seidl (2000) already hasshown that two sections of subg. Myxacium, sec. My-xacium and sec. Defibulati, form a monophyleticgroup with two distinct lineages. Our phylogeneticanalysis has added species from the southern hemi-sphere. Cortinarius cycneus and ‘‘C. magellanicus’’form a distinct lineage within /Myxacium s.l. This‘‘C. magellanicus’’ from New Zealand is not conspe-cific with the material from the type locality in SouthAmerica. More data from the numerous SouthernHemisphere Myxacium and Paramyxacium speciesare needed to determine whether Northern Hemi-sphere species evolved as a separate lineage due togeographic isolation.
Clade /Ochroleuci: Konrad and Maublanc (1948)established subg. Myxacium sec. Ochroleuci, based onthe basidiome color and the bitter taste of the slimyveil. Bayesian posterior probabilities support sistergroup relationships of this lineage to /Allutus and/Calochroi, suggesting that viscid universal veils arederived secondarily in /Ochroleuci.
Clade /Delibuti comprises Northern and SouthernHemisphere species. Therefore it might be regardedas a comparatively old lineage. Horak and Wood(1990) hypothesized that C. salor and C. rotundispo-rus represent living fossils because they appear tohave been ectomycorrhizal partners in the Nothofa-gus and Eucalyptus-Leptospermum forests in Gondwan-aland millions of years ago. Relationships of C. iodesand C. salor, both classified in sec. Delibuti, remainunresolved. Common morphological characters arethe bluish basidiome colors and (sub)globose basid-iospores. However, as indicated by the numerous tax-onomic shifts, there is considerable uncertainty re-garding the systematic placement of many so-calledDelibuti; C. australiensis has synonyms in the form ge-nus Rozites, and C. rotundisporus also has been placedin Phlegmacium sec. Amarescentes. The combinedanalysis shows C. iodes and C. rotundisporus as sistergroups, indicating a close relationship of /Delibutito /Phlegmacium.
The lineages of Phlegmacium spp. Our results andother recent molecular studies on Phlegmacium spe-cies (Moser and Peintner 2002a; Garnica et al 2003a,b) underscore three urgent reasons for revising tra-ditional concepts of subg. Phlegmacium based onmorphological, molecular and chemotaxonomicaldata: (i) subg. Phlegmacium is polyphyletic, (ii) manysections of Phlegmacium as defined by Moser (in Sing-er 1986) (e.g., Fulvi) are also polyphyletic; (iii) many

1053PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
clades implied by molecular analyses are related toother classically based taxa.
Using chemotaxonomical data, Brandrud (1998,2002) proposed that section Fulvi is polyphyletic de-spite its apparent morphological homogeneity. Phleg-macium species with the most primitive biosyntheticpigment pathways also have high pigment concentra-tions, high pigment diversity and easily oxidized com-pounds. These supposed primitive pigment charac-ters co-occur with such extreme ‘‘phlegmacioid’’ fea-ture as bright coloration, strongly glutinous pileussurfaces, marginate bulbous stipe bases, citriform,coarsely verrucose spores, and absence of brown en-crusted parietal pigments. Derived species arethought to possess more inconspicuous telamonioidcharacters, such as brown coloration, and less bul-bous stipes (Brandrud 2002). We likewise observetwo lineages with viscid universal veils derived fromCortinarius species producing phlegmacioid basidi-omes. It would appear that the evolutionary trend forPhlegmacium spp. and perhaps Cortinarius in generalis a decrease in pigment diversity and a shift awayfrom spectacular toward less conspicuous basidiomes.Another observed trend is from complex agaricoidto reduced sequestrate fruit bodies (Peintner et al2001).
The /calochroid group is a common lineage forPhlegmacium spp. and /Ochroleuci in subg. Myxa-cium. Garnica et al (2003a) also reported speciesfrom different sections of European Phlegmacium in/Calochroi. Morphological characters common tothis group are citriform basidiospores, a well-devel-oped gelatinous layer and epicutis, a simplex pilei-pellis, brightly colored basidiomes and a marginatedbulbous stipe.
In addition, /Phlegmacium circumscribes taxa as-signed to different sections of subg. Phlegmacium.Moreover, transitions between ‘‘classical’’ Phlegma-cium and Myxacium spp. morphotypes can be ob-served also in this lineage; Cortinarius coerulescens hasthe blue colors typical for representatives of the spe-cies complex around C. iodes (subg. Myxacium) butwith a less-developed viscid veil. C. iodes shows a sistergroup relationships with C. rotundisporus in the com-bined analysis, a representative of /Delibuti.
The lineage subg. Cortinarius. Representative spe-cies of subg. Cortinarius, one of the smallest subgen-era with only ;10 species worldwide, form a well-supported lineage. Our results support segregationof small groups based on classical morphological con-cepts; subg. Cortinarius, as delimited from other Cor-tinarius subclades, produces large, fleshy basidiomeswith dry, squamose-tomentose pileus surfaces, lamel-lae bearing cheilo- and pleurocystidia and tissues thatturn red in KOH (Moser in Singer 1986).
Lineages of species with well-developed universal veils.Basal relationships are not resolved for lineages/Rozites, /Cuphocybe, and /Achrous. These cladesrepresent former Rozites or Cuphocybe taxa. Roziteshas been inferred to be polyphyletic (Peintner et al2002c). The amyloid reaction in the trama is themost conspicuous shared morphological character ofspecies of /Rozites. Data including more taxa withamyloid trama reaction support a subg. Rozites cir-cumscribing taxa around the type species, C. capera-tus (Peintner unpublished).
The lineage(s) of Dermocybe. The included Der-mocybe taxa fall into three lineages: /Dermocybe s.l.,/Icterinula and /Austronanceiensis. Our data do notreject monophyly of these three clades. Importantimplications resulting from our phylogenetic analysesare: (i) all Dermocybe spp. analyzed are nested in Cor-tinarius, suggesting for Cortinarius systematics thatthey and probably all other Dermocybe species shouldbear a Cortinarius name; (ii) the lineage /Dermocy-be s.l. can be considered as Cortinarius subg. Dermo-cybe (Fr.) Trog because this clade of Northern Hemi-sphere taxa includes the type of Dermocybe, C. cin-namomeus (L. : Fr.) Wunsche (Liu et al 1997); (iii)Northern Hemisphere taxa of /Dermocybe s.str. (sec.Dermocybe) appear derived from Southern Hemi-sphere taxa; (iv) /Dermocybe s.l. is paraphyletic, in-cluding taxa of subgenera Cystogenes and Telamonia.More species from the Southern Hemisphere shouldbe sampled to test monophyly of Dermocybe and toelucidate the origin and evolution of this interestinggroup of fungi.
Clade /Dermocybe s.l. includes the subclades/Dermocybe s.str., /Splendida (D. splendida, C. glob-uliformis) as well as C. austrovenetus, C. firmus (Te-lamonia), C. olivaceopictus, and C. teraturgus (Cysto-genes). A wide range of pigments can be found intaxa of this clade. Taxa of /Dermocybe s.str. containdermorubin and sometimes also dermocybin (Keller1982). C. olivaceopictus represents a separate lineage(Liu et al 1997). This corresponds with results of pig-ment analyses that showed that C. olivaceofuscus,which is related closely to C. olivaceopictus, to containendocrocin and flavomannin-6,6-dimethylether butno dermorubin (Gruber 1975). Høiland (1983) re-garded the C. olivaceopictus complex basal to North-ern Hemisphere Dermocybe species because its pig-ment spectrum is less complex than found in /Der-mocybe s.str. Pigments of C. firmus have not beeninvestigated. Moser and Horak (1975) placed C. aus-trovenetus in Dermocybe subg. Icterinula, sec. Pauperae.Taxa of stirps Austroveneta typically have olivaceouscolors. The two major pigments are the orange an-thraquinone skyrin and the green-yellow pre-anthra-quinone austrovenetin. Austrovenetin is unstable and

1054 MYCOLOGIA
on exposure to oxygen and sunlight changes chem-ically to protohypericin and the purple extended qui-none hypercin (Gill 1995b). These pigments are re-lated chemically to pigments in /Purpurascentes(sec. Scauri), atrovirin-B, skyrin, and (probably) hy-pericin (Keller et al 1987, Gill 1995a). BecauseD. splendida contains the rare tetrahydroanthraqui-nones, Gill et al (1995b) assume an isolated positionfor this taxon, suggesting a possible relationship toC. umbonatus and D. erythrocephala. Moser and Hor-ak (1975) integrated C. teraturgus in subg. Cystogenesstirps Austrolimonius. Most species in Cystogenes havecheilocystidia, and orange to red-brown colors char-acterize species of stirps Austrolimonius. Chromato-graphy demonstrated the lack of typical Dermocybepigments (Gruber 1975), which is in agreement withthe isolated position of C. teraturgus in /Dermocybes.l.
Clade /Icterinula includes Southern Hemispherespecies in Dermocybe subg. Icterinula, sec. Holoxantha,stirps Amoena. C. icterinus and C. amoenus have pre-dominantly yellow pigments (endocrocin, dermolu-tein) and a slightly gelatinized pileipellis (Moser andHorak 1975, Garnica 2003b). Endocrocin and der-molutein are the principal anthraquinonic pigments(Gruber 1975).
Clade /Austronanceiensis includes SouthernHemisphere species in Dermocybe subg. Icterinula sec.Holoxantha stirps Nothoveneta. C. austronanceiensisdiffers from species in stirps Amoena by more robustbasidiomes and pileipelli that are not gelatinize, butthe main pigments of C. austronanceiensis are alsoendocrocin and dermolutein. Dermocybe cardinaliscombines yellow, red and purple pigments that areproduced by the cardinals, a series of unique naph-thoquinone dimers. Because cardinals are unique forthis species, Gill (1995b) proposed an isolated posi-tion for D. cardinalis. However, our phylogeneticanalyses suggest a close relationship to Dermocybe aus-tronanceiensis, providing further support for mono-phyly of /Icterinula and /Austronanceiensis.
The lineages of Telamonia spp. The /telamonioidgroup is a lineage of /Telamonia /Orellani, /An-omali, and C. laetus. /Telamonia morphologically cir-cumscribes a homogenous group of Cortinarii. Hy-grophanous pilei, the lack of viscid or gelatinous veilsand well-developed cortinas characterize most spe-cies. The fluid transitions among inconspicuous mor-phological characters make the systematics of thisgroup particularly difficult. Compared to the otherCortinarius spp., members of /Telamonia all haveITS sequences with a ;50 bp gap in the ITS1 region.Moreover, there is a low divergence of ITS sequencesin this group that makes it particularly difficult toresolve subclades within /Telamonia. More variable
molecular regions have to be tested to infer for ameaningful phylogeny for this difficult group of‘‘small brown mushrooms’’.
Species of /Orellani share many characters withspecies of /Telamonia. In addition, they containbright blue or blue-green fluorescent substances andorellanine, a lethal mushroom nephrotoxin (Danelet al 2001). The placement of /Anomali in the tela-monioid group also is in agreement with morpholog-ical characters typical for this group (subg. Sericeocybesec. Anomali), e.g., slender basidiomes that lack viscidveils. Many species of sec. Sericeocybe in subg. Sericeo-cybe appear in /Telamonia. Such a taxonomic place-ment already was proposed by Brandrud et al (1995)based on morphological characters. Cortinarius laetusis a Telamonia spp. characterized by a conspicuousyellow universal veil and small, mycenoid aspect.Therefore, it was a surprise at first to discover thatthe morphologically similar species of /Acutus are asegregate of /Telamonia, as already shown byHøiland and Holst-Jensen (2000). Peintner et al(2003) more recently have demonstrated that at leastfive species (including one sequestrate) belong tothis lineage. On closer examination, species of /Acutidiffer from other Telamonia spp. by the typical struc-ture of the lamellar trama consisting of ellipsoid in-flated hyphae and by the presence of numerouscheilocystidia. The independent origin inferred for/Acutus suggests that the mycenoid habit originatedat least twice in Cortinarius: once in the /telamonioidgroup (C. pulchellus, C. laetus) and once in /Acutus.
The lineages of Leprocybe spp. The subg. Leprocybeas currently recognized is polyphyletic: some cladesor species cluster in the /telamonioid group (/Or-ellani, C. bolaris, C. raphanoides, C. gentilis), othersform lineages with unknown relationship (/Callis-teus, /Limonius, /Leprocybe). The fluorescent pig-ments that define this subgenus do not appear to besynapomorphic. Tree topologies indicate a relation-ship of /Callisteus and /Limonius to Phlegmaciumspecies, from which they mainly differ by the reduc-tion of the viscid pileipellis. More extensive samplingwill prove our hypothesis that this subgenus has tobe restricted to /Leprocybe as typified by C. cotoneus.
Can the Cortinarius phylogeny be resolved?—Our re-sults reveal many fundamental problems for Corti-narius systematics. First, it is difficult to identify anddelimit taxa reliably within this species-rich genus, es-pecially taxa from different geographical regions.‘‘C. magellanicus’’ from New Zealand (/Cycneus), forinstance, is not conspecific with (and in fact well sep-arated from) C. magellanicus (/Ochroleuci) from thetype locality in South America (FIG. 1). For reliablephylogenetic data it often is necessary to analyze se-

1055PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
quences from at least two or more collections of onespecies and if possible from different geographicalorigins (Peintner et al 2001, Moser and Peintner2002a, b, Peintner et al 2002c, Garnica et al 2003b).
Resolution and support in a phylogeny can be im-proved by increasing the number of characters (Soltiset al 1998, 1999). Here, character sampling helpedto resolve relationships between (sub)clades of Cor-tinarius. In general, increased character sampling in-creased BPP for larger clades showing low support(/Dermocybe s.l.), or recovered and supported new,larger clades (e.g., the telamonioid group).
On the other hand, increased taxon sampling re-solved relationships in terminal branches of the phy-logeny, e.g., within clades. The lineages /Purpuras-centes and /Phlegmacium were not supported oronly weakly supported in the combined analysis (FIG.2), yet a significantly higher BPP was obtained withincreased taxon sampling (FIG. 1). This indicates thata minimum number of representative species mustbe sampled to recover a clade in multigene phylog-enies. Extensive taxon sampling still is regarded asone of the most reliable ways to increase overall phy-logenetic accuracy (Hillis 1996, 1998, Pollock andBruno 2000, Pollock et al 2002, Zwickl and Hillis2002). Thus, in addition to the low divergence inrDNA sequences, insufficient taxon sampling canconfuse Cortinarius phylogeny. Increased taxon sam-pling has helped to resolve the phylogeny of closelyrelated Cortinarius species (Liu et al 1997, Seidl 2000,Moser and Peintner 2002, Garnica et al 2003, Peint-ner et al 2003). Cortinarius, one of the most species-rich ectomycorrhizal mushrooms genera, has a world-wide distribution, yet most subgenera and sectionsare typified by species from the Northern Hemi-sphere. Selecting the type species for analysis was ap-propriate, but in so doing we strongly biased our re-sults toward the Northern Hemisphere. Consequent-ly, future research should include more species fromthe Southern Hemisphere. A meaningful phylogenyof this morphologically plastic and large genus can-not be based on a mere 5–10% of the known species.It would be appropriate to focus future phylogeneticanalyses first on single clades and to sample taxafrom both hemispheres to better understand the evo-lution of Cortinarius and thus to resolve, step by step,the challenging taxonomic problems of this fascinat-ing genus.
The insights gained by a thorough investigation onCortinarius phylogeny and evolution can be relatedto other fungal groups, especially ectomycorrhizal ba-sidiomycetes. Ectomycorrhizal fungi are concentrat-ed in the bolete clade, thelephoroid clade, canthar-elloid clade, gomphoid-phalloid clade and in the eu-agarics clade (Amanita, Hygrophorus, Tricholoma, In-
ocybe, Cortinarius, Phaeocollybia, Hebeloma andLaccaria) (Hibbett and Thorn 2000). The existenceof numerous, relatively large clades of ectomycorrhi-zal fungi indicates that this ecological habit has beenrelatively stable at least during the recent radiationof euagarics (Moncalvo et al 2000, 2002). The phy-logeny of ectomycorrhizal basidiomycetes has beenaddressed in many studies (e.g., Binder 1999, Jarosch2001, Grubisha et al 2002, bolete clade; Koljalg et al2000, 2002, thelephoroid clade; Dahlman et al 2000,cantharelloid clade; Humpert et al 2001, gomphoid-phalloid clade; Miller et al 2001, russuloid clade;Drehmel et al 1991, Amanita; Aanen et al 2000, He-beloma) most of them using rDNA sequence data. Fewstudies furnish comparatively well-resolved phyloge-nies on different taxonomic levels (Drehmel et al1991, Grubisha et al 2002, Miller 2003), but somesubclades often remain unresolved, and the phylog-enies of large genera are especially difficult to resolve(Binder 1999, Humpert et al 2001, Miller et al 2001).Strategies developed to resolve the phylogeny of Cor-tinarius and the resulting insights in the taxonomicvalue of morphological and ecological characterscould help to address the phylogeny, systematics andevolution of many other groups of fungi.
ACKNOWLEDGMENTS
This paper is based on ideas and suggestions of MeinhardMoser. Unable to thank him personally, we acknowledge hishelp by trying to continue Cortinarius research in his spirit.We thank Lorelei Norvell, Michelle Seidl and an unknownreviewer for their helpful suggestions, which certainlyserved to improve this paper. This work was financed by theNSF grants DEB-9708035 and DEB-0076023 to Jean-MarcMoncalvo and RV and by an Erwin Schrodinger Auslandss-tipendium ( J1821-BIO) grant from the Austrian ScienceFoundation (FWF) to UP.
LITERATURE CITED
Aanen DK, Kuyper TW, Boekhout T, Hoekstra RF. 2000.Phylogenetic relationships in the genus Hebeloma basedon ITS1 and 2 sequences, with special emphasis on theHebeloma crustuliniforme complex. Mycologia 92:269–281.
Arnold N. 1993. Morphologisch-anatomische und chemis-che Untersuchung der Untergattung Telamonia (Cor-tinarius, Agaricales). IHW Verlag, Eching. 214 p.
, Besl H, Bresinsky A, Kemmer H. 1987. Remarks onthe chemotaxonomy of the genus Dermocybe (Agari-cales) and on its distribution in Bavaria (West Germa-ny). Zeitschrift fur Mykologie 53:187–194.
Binder M. 1999. Zur molekularen Systematik der Boletales:Boletinae und Sclerodermatinae subordo nov. Univer-sitat Regensburg, Regensburg.
Brandrud TE. 1998. Cortinarius subgen. Phlegmacium sec.

1056 MYCOLOGIA
Fulvi: chemotaxonomy versus morphological taxono-my. Journales des J.E.C. 1998:4–9.
. 2002. Evolution of subg. Phlegmacium in light of itspigment chemistry a case of ‘‘trivialization’’ and loss ofcharacter diversity? In: International Mycological Con-gress 7 Book of Abstracts, Oslo. p 125.
, Lindstrom H, Marklund H, Melot J, Muskos S. 1995.Cortinarius Flora Photographica Teil 3. Fotoflora. Cor-tinarius HB, Matfors.
Chambers SM, Sawyer NA, Cairney JWG. 1999. Molecularidentification of co-occurring Cortinarius and Dermo-cybe species from southeastern Australian SclerophyllForests. Mycorrhiza 9:85–90.
Dahlman M, Danell E, Spatafora JW. 2000. Molecular sys-tematics of Craterellus: cladistic analysis of nuclear LSUrDNA sequence data. Mycol Res 104:388–394.
Danel VC, Saviuc PF, Garon D. 2001. Main Features of Cor-tinarius spp. poisoning: a literature review. Toxicon 39:1053–1060.
Drehmel D, Moncalvo JM, Vilgalys R. 1999. Molecular phy-logeny of Amanita based on large-subunit ribosomalDNA sequences: implications for taxonomy and char-acter evolution. Mycologia 91:610–618.
Gardes M, Bruns TD. 1993. ITS primers with enhancedspecificity for basidiomycetes application to the iden-tification of mycorrhizae and rusts. Mol Ecol 2:113–118.
Garnica S, Weiß M, Oberwinkler F. 2003b. Morphologicaland molecular phylogenetic studies in South AmericanCortinarius species. Mycol Res 197:1143–1156.
, , . 2003a. Phylogenetic relationshipsof European Phlegmacium species (Cortinarius Agari-cales). Mycologia 95:1155–1170.
Garnier G. 1991. Bibliographie des Cortinaires. Volumes:A–C: 180 p. D–O: 272 p. P–Z: 290 p. Federation my-cologique Dauphine-Savoie. Annecy.
Gill M. 1995a. New pigments of Cortinarius Fr. and Dermo-cybe (Fr.) Wunsche (Agaricales) from Australia andNew Zealand. Sydowia Beihefte 10:73–87.
. 1995b. Pigments of Australasian Dermocybe toad-stools. Aust J Chem 48:1–26.
, Steglich W. 1987. Pigments of Fungi (Macromyce-tes). Prog Chem Org Nat Prod 51:1–317.
Gruber I. 1975. Papierchromatographische Pigmentanalysevon Sudamerikanischen Dermocyben und Cortinarien.In: Moser M, Horak E, eds. Cortinarius Fr.und naheverwandte Gattungen in Sudamerika Nova Hedwigia,Beiheft 52:524–540.
Grubisha LC, Trappe JM, Molina R, Spatafora JW. 2002.Biology of the ectomycorrhizal genus Rhizopogon. VI.Re-examination of infrageneric relationships inferredfrom phylogenetic analyses of ITS sequences. Mycolo-gia 94:607–619.
Heim R. 1951. Cuphocybe, Noveau genre neo-zelandais d’agarics ochrospores. Revue de Mycologie 16:3–10.
Hibbett DS, Thorne RG. 2000. Basidiomycota: Homobasi-diomycetes. In: McLaughlin DS, ed. The Mycota VII:Systematics and Evolution. p 121–160.
Hillis DM. 1996. Inferring complex phylogenies. Nature383:130–131.
. 1998. Taxonomic sampling, phylogenetic accuracy,and investigator bias. Syst Biol 47:3–8.
Høiland K. 1983. Cortinarius Subg. Dermocybe. Opera Bo-tanica 71:5–113.
, Holst-Jensen A. 2000. Cortinarius phylogeny andpossible taxonomic implications of ITS rDNA sequenc-es. Mycologia 92:694–710.
Hopple JS, Vilgalys R. 1999. Phylogenetic relationships inthe mushroom genus Coprinus and dark-spored alliesbased on sequence data from the nuclear gene codingfor the large ribosomal subunit RNA: divergent do-mains, outgroups, and monophyly. Mol PhylogenetEvol 13:1–19.
Horak E. 1988. New species of Dermocybe (Agaricales) fromNew Zealand. Sydowia 40:81–112.
. 1999. New genera of Agaricales (Basidiomycota) 1.Rapacea gen. nov. Kew Bulletin 54:789–794.
, Wood AE. 1990. Cortinarius Fr. (Agaricales) in Aus-tralasia. 1. Subgen. Myxacium and subgen. Paramyxa-cium. Sydowia 42:88–168.
Huelsenbeck JP, Bollback JP, Levine AM. 2002. Inferring theroot of a phylogenetic tree. Syst Biol 51:32–43.
, Ronquist FR. 2001. MrBayes: Bayesian inference ofphylogeny. Biometrics 17:754–755.
Humpert AJ, Muench EL, Giachini A, Castellano MA, Spa-tafora JW. 2001. Molecular phylogenetics of Ramariaand related genera: evidence from nuclear large sub-unit and mitochondrial small subunit rDNA sequences.Mycologia 93:465–477.
Jarosch M. 2001. Zur molekularen Systematik der Boletales:Coniophorinae Paxillinae und Suillinae. Berlin: J. Cra-mer.
Karol KG, Mccourt RM, Cimino MT, Delwiche CF. 2001.The closest living relatives of land plants. Science 294:2351–2353.
Keller G. 1982. Pigmentuntersuchungen bei europaischenArten aus der Gattung Dermocybe (Fr. Wunsche). Sy-dowia 35:110–126.
, Moser M, Horak E, Steglich W. 1987. Chemotaxo-nomic investigations of species of Dermocybe (Fr. Wun-sche) Agaricales from New Zealand, Papua New Guin-ea and Argentina. Sydowia 40:168–187.
Kirk PM, Cannon PF, David JC, Stalpers J. 2001. Ainsworth& Bisby’s dictionary of the fungi. 9th ed. Oxon, UK:CAB International.
Koljalg U, Dahlberg A, Taylor AFS, Larsson E, HallenbergN, Stenlid J, Larsson KH, Fransson PM, Karen O, Jons-son L. 2000. Diversity and abundance of resupinate the-lephoroid fungi as ectomycorrhizal symbionts in Swed-ish boreal forests. Mol Ecol 9:1985–1996.
, Tammi H, Timoner S, Agerer R, Sen R. 2002. ITSrDNA sequence-based phylogenetic analysis of Tomen-tellopsis species from boreal forests, and the identifica-tion of pink-type ectomycorrhizas. Mycol Progress 1:81–92.
Konrad P, Maublanc A. 1948. Les Agaricales. Classification,Revision des especes- Iconographie, Comestibilite. En-cyclopedie Mycologique XIV. Paul Lechevalier, Paris.
Kopanski L, Klaar M, Steglich W. 1982. Fungus pigments:40. Leprocybin, the fluorescent principle of Cortinarius

1057PEINTNER ET AL: INFRAGENERIC RELATIONSHIPS IN CORTINARIUS
cotoneus and related Leprocybes (Agaricales). LiebigsAnnalen der Chemie: 1280–1296.
Kuhner R. 1980. Les Hymenomycetes agaricoıdes: etude ge-nerale et classification. Numero special du Bulletin dela Societe Linneenne de Lyon, France.
, Romagnesi H. 1953. Flore Analytique des Champi-gnons Superieurs. Masson, Paris.
Kuhnert R, Peintner U. 2003. Revised nomenclature of Cor-tinarius taxa with type specimens deposited in the Inns-bruck (IB) mycological collection. Mycotaxon 81:113–120.
Largent B, Simon DL. 1999. Markov Chain Monte Carloalgorithms for Bayesian analysis of phylogenetic trees.Mol Biol Evol 16:750–759.
Liu YJ, Rogers SO, Ammirati JF. 1997. Phylogenetic rela-tionships in Dermocybe and related Cortinarius taxabased on nuclear ribosomal DNA internal transcribedspacers. Can J Bot 75:519–532.
Marvaldi AE, Sequeira AS, O’Brien CW, Farrell BD. 2002.Molecular and morphological phylogenetics of Weevils(Coleoptera, Curculionoidea): do niche shifts accom-pany diversification? Syst Biol 51:761–785.
Melot J. 1990.Une classification du genre Cortinarius (Pers.S.F.Gray). Documents Mycologiques 20:43–59.
Miller OK. 2003. The Gomphidiaceae revisited: a worldwideperspective. Mycologia 95:176–183.
Miller RE, Buckley TR, Manos PS. 2002. An examination ofthe monophyly of morning glory taxa using bayesianphylogenetic inference. Syst Biol 51:740–753.
Miller SL, McClean TM, Walker JF, Buyck B. 2001. A mo-lecular phylogeny of the Russulales including agari-coid, gasteroid and pleurotoid taxa. Mycologia 93:344–354.
Moenne-Loccoz PR, Reumaux P, Henry R. 1990. Atlas desCortinaires II. Editions Federation mycologique Dau-phine-Savoie. Annecy.
Moncalvo JM, Lutzoni FM, Rehner SA, Johnson J, VilgalysR. 2000. Phylogenetic relationships of agaric fungibased on nuclear large subunit ribosomal DNA se-quences. Syst Biol 49:278–305.
, Vilgalys R, Redhead SA, Johnson JE, James TY,Aime MC, Hofstetter V, Verduin SJW, Larsson E, BaroniTJ, Thorn RG, Jacobsson S, Clemencon H, Miller OK.2002. One hundred and seventeen clades of euagarics.Mol Phylogenet Evol 23:357–400.
Moser M. 1960. Die Gattung Phlegmacium. Verlag JuliusKlinkhardt, Bad Heilbronn. 440 p.
. 1972. Die Gattung Dermocybe (Fr.). Wunsche (DieHautkopfe). Schweizerische Zeitschrift fur Pilzkunde50:153–167.
. 1983. Die Rohrlinge und Blatterpilze. 5th ed. In:Kleine Kryptogamenflora Bd. IIb/2: Stuttgart: GustavFischer Verlag. 533 p.
, Horak E. 1975. Cortinarius Fr. und nahe verwandteGattungen in Sudamerika. Nova Hedwigia Beiheft 52:1–628.
, Peintner U. 2002a. Die phylogenetischen Beziehun-gen der Cortinarius aureopulverulentus Gruppe. Jour-nales des J.E.C. 4:28–38.
, . 2002b. The species complex Cortinarius scau-
rus-C. herpeticus based on morphological and moleculardata. Micologia e Vegetazione Mediterranea 17:3–17.
Murphy WJ, Eizirik E, O’Brien SJ, Madsen O, Scally M,Douady CJ, Teeling E, Ryder OA, Stanhope MJ, DeJong WWW, Springer MS. 2001. Resolution of the earlyplacental mammal radiation using Bayesian phyloge-netics. Science 294:2348–2351.
Nielsen R. 2002. Mapping mutations on phylogenies. SystBiol 51:729–739.
Orton PD. 1958. The genus Cortiniarius II. Inoloma andDermocybe. The Naturalist (Supplement):81–149.
Peintner U, Bougher NL, Castellano MA, Moncalvo JM,Moser M, Trappe JM, Vilgalys R. 2001. Multiple originsof sequestrate fungi related to Cortinarius (Cortinari-aceae). Am J Bot 88:2168–2179.
, Moser M, Vilgalys R. 2002a. Thaxterogaster is a tax-onomic synonym of Cortinarius: new names and newcombinations. Mycotaxon 81:177–184.
, Moser M, Vilgalys R. 2002b. Rozites, Cuphocybe andRapacea are taxonomic synonyms of Cortinarius: newnames and new combinations. Mycotaxon 83:447–452.
, Horak E, Moser M, Vilgalys R. 2002c. Phylogeny ofRozites, Cuphocybe and Rapacea. Mycologia 94:620–629.
, Moser M, Agretious Thomas K, Manimohan P.2003. First records of ectomycorrhizal Cortinarius spe-cies (Agaricales, Basidiomycetes) from tropical Indiaand their phylogenetic position based on rDNA ITSsequences. Mycol Res 107:485–494.
Pollock DD, Bruno WJ. 2000. Assessing an unknown evo-lutionary process: effect of increasing site-specificknowledge through taxon addition. Mol Biol Evol 17:1854–1858.
, Zwickl DJ, McGuire JA, Hillis DM. 2002. Increasedtaxon is advantageous for phylogenetic inference. SystBiol 51:664–671.
Posada D, Crandall KA. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14:817–818.
Salamin N, Chase MW, Hodkinson TR, Savolainen V. 2003.Assessing internal support with large phylogeneticDNA matrices. Mol Phylogenet Evol 27:528–539.
Seidl MT. 2000. Phylogenetic relationships within Cortinar-ius subg. Myxacium, sections Defibulati and Myxacium.Mycologia 92:1091–1102.
Singer R. 1986. The Agaricales in modern taxonomy. 4th ed.Koenigstein, Germany: Koeltz Scientific Books. 981 p.
Steglich W, Oertel B. 1984. Studies on the composition anddistribution of pigments of Cortinarius, subg. Phleg-macium (Agaricales). Sydowia 37:284–295.
Soltis DE, Soltis PS, Chase MW. 1999. Angiosperm phylog-eny inferred from multiple genes as a tool for compar-ative biology. Nature 402:402–404.
, , Mort ME, Chase MW, Savolainen V, HootSB, Morton CM. 1998. Inferring complex phylogeniesusing parsimony: an empirical approach using threelarge DNA data sets for angiosperms. Syst Biol 47:32–42.
Suzuki Y, Glazko GV, Nei M. 2002. Overcredibility of mo-lecular phylogenies obtained by Bayesian phylogenet-ics. Proc Natl Acad Sci USA 99:16138–16143.
Swofford DL. 2001. PAUP*. Phylogenetic analysis using par-

1058 MYCOLOGIA
simony (*and other methods). Version 4.0d10. Sunder-land, Massachusetts: Sinauer Associates.
Thorne JL, Kishino H. 2002. Divergence time and evolu-tionary rate estimation with multilocus data. Syst Biol51:689–703.
Vilgalys R, Hester M. 1990. Rapid genetic identification andmapping of enzymatically amplified ribosomal DNA fromseveral Cryptococcus species. J Bacteriol 172:4238–4246.
, Sun BL. 1994. Ancient and recent patterns of geo-graphic speciation in the oyster mushroom Pleurotusrevealed by phylogenetic analysis of ribosomal DNA se-quences. Proc Natl Acad Sci USA 91:4599–4603, 7832.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and
direct sequencing of fungal ribosomal RNA genes forphylogenetics. In: Michael AI, Gelfand DH, Sninsky JJ,White TJ, eds. PCR protocols: a guide to methods andapplications. New York: Academic Press. p 315–322.
Wilcox TP, Zwickl DJ, Heath TA, Hillis DM. 2002. Phylo-genetic relationships of the dwarf boas and a compar-ison of Bayesian and bootstrap measures of phyloge-netic support. Mol Phylogenet Evol 25:361–371.
Yang Z, Rannala B. 1997. Bayesian phylogenetic inferenceusing DNA sequences: a Markov Chain Monte Carlomethod. Mol Biol Evol 14:717–724.
Zwickl DJ, Hillis DM. 2002. Increased taxon sampling great-ly reduces phylogenetic error. Syst Biol 51:588–598.
Related Documents











