Topical application of Gallic acid suppresses the 7,12-DMBA/Croton oil induced two-step skin carcinogenesis by modulating anti-oxidants and MMP-2/MMP-9 in Swiss albino mice Vimala Subramanian, Balaji Venkatesan, Anusha Tumala, Elangovan Vellaichamy ⇑ Department of Biochemistry, University of Madras, Guindy Campus, Chennai 600025, India article info Article history: Received 17 September 2013 Accepted 13 January 2014 Available online 18 January 2014 Keywords: Gallic acid Skin cancer Anticancer activity DMBA/Croton oil GST MMP abstract Gallic acid (GA – 3,4,5-trihydroxybenzoic acid), a dietary anti-oxidant has been shown to inhibit cancer cell growth in in vitro. Herein, we investigated the in vivo chemo preventive activity of GA on 7,12-Dim- ethylbenz[a]anthracene (DMBA)/Croton oil induced two-step skin carcinogenesis in Swiss albino mice. Skin tumor incidence and tumor volume were recorded during the 16 weeks of experimental period. In addition, LDH-isozyme shift, skin collagen content, activities of matrix metalloproteinases (MMP-2/ MMP-9) enzymes and enzymatic and non-enzymatic antioxidant were studied in the skin and serum of experimental mice. Tumor incidence was significantly increased in the DMBA/Croton oil induced mice (100%; p < 0.001) when compared to GA co-treated mice (60%; p < 0.01) and 5-FU treated mice (50%; p < 0.01). Skin collagen content, MMPs activities, LDH-isoenzymes and MMP-2/-9 expressions were increased in DMBA/Croton oil induced skin while decreased levels of enzymatic (GST, SOD, CAT & GPx) and non-enzymatic anti-oxidant (GSH) were noticed. On the other hand, GA co-treatment exhibited a sig- nificant protection by reverting back the altered levels of LDH-isoenzymes, antioxidants, collagen and MMP-2/MMP-9 activities. The results of this study indicate that topical application of GA inhibits DMBA/Croton oil induced two-stage skin carcinogenic process by modulating the antioxidants and MMPs (-2 & -9) in the mouse skin. Ó 2014 Elsevier Ltd. All rights reserved. 1. Introduction Skin is the largest organ of the human body, and is a major tar- get for toxic insult by a chemical mutagens and carcinogens in a day-to-day life (Simna et al., 2012). Throughout the world, the number of people affected by skin cancer is increasing due to in- creased exposure to environmental pollutants and UV rays (Narendhirakannan and Angeline Christie Hannah, 2013). Recent studies have also predicted that the number of people affected by this malady will be doubled by the year 2050 (Hayat et al., 2007). Although many synthetic drugs are commercially available to treat skin cancer; However, these drugs causes serious side ef- fects to patients such as nephrotoxicity, neurotoxicity, infertility, thromboembolic complications, hair loss, nausea, and myocardial infarction (Shabaruddin et al., 2013). Hence, there is a dire need to look for more efficacious agents with lesser side effects for the prevention and eradication of skin tumors. Number of recent evidence has shown that natural dietary anti- oxidants are capable of inhibiting cancer cell growth. Moreover, these natural antioxidants are reported to have the ability to en- hance the activities of detoxification enzyme components such as glutathione-S-transferase (GST). The activation of GST not only neutralizes the chemical carcinogens, but also preventing them from forming a DNA adducts in the genome (Liu et al., 1992). Fur- thermore, it has been suggested that these natural antioxidant compounds can be used in multiple ways, either as cancer preven- tive agents or even as cancer therapy drugs (Thornthwaite et al., 2013). Gallic acid (GA), a polyhydroxyphenolic compound, is abun- dantly present in leafy vegetables, fruits, and nuts such as gallnut, grapes, sumach, oak bark, green tea, apple peels (Atkinson et al., 2004; Chakraborty et al., 2009; Kim et al., 2006; Ng et al., 2004; Liao et al., 2012) and also present in onion and potato varieties (Albishi et al., 2013). GA exhibits variety of biological activities including antioxidant (Isuzugawa et al., 2001; Yogendra Kumar et al., 2013), anti-tumor (Agarwal et al., 2006; Liao et al., 2012; Lee et al., 2013; Locatelli et al., 2013), anti-inflammatory (Albishi et al., 2013) and anti-bacterial (Kang et al., 2008; Lee and Je, 2013). GA has been shown to inhibit gastric cancer cell growth in vitro by modulating the levels of MMP-2 and -9 and cytoskeletal reorganization (Ho et al., 2010). Furthermore, GA has also been http://dx.doi.org/10.1016/j.fct.2014.01.017 0278-6915/Ó 2014 Elsevier Ltd. All rights reserved. ⇑ Corresponding author. Tel.: +91 44 22202734. E-mail address: [email protected] (E. Vellaichamy). Food and Chemical Toxicology 66 (2014) 44–55 Contents lists available at ScienceDirect Food and Chemical Toxicology journal homepage: www.elsevier.com/locate/foodchemtox

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Food and Chemical Toxicology 66 (2014) 44–55
Contents lists available at ScienceDirect
Food and Chemical Toxicology
journal homepage: www.elsevier .com/locate/ foodchemtox
Topical application of Gallic acid suppresses the 7,12-DMBA/Croton oilinduced two-step skin carcinogenesis by modulating anti-oxidantsand MMP-2/MMP-9 in Swiss albino mice
http://dx.doi.org/10.1016/j.fct.2014.01.0170278-6915/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author. Tel.: +91 44 22202734.E-mail address: [email protected] (E. Vellaichamy).
Vimala Subramanian, Balaji Venkatesan, Anusha Tumala, Elangovan Vellaichamy ⇑Department of Biochemistry, University of Madras, Guindy Campus, Chennai 600025, India
a r t i c l e i n f o
Article history:Received 17 September 2013Accepted 13 January 2014Available online 18 January 2014
Keywords:Gallic acidSkin cancerAnticancer activityDMBA/Croton oilGSTMMP
a b s t r a c t
Gallic acid (GA – 3,4,5-trihydroxybenzoic acid), a dietary anti-oxidant has been shown to inhibit cancercell growth in in vitro. Herein, we investigated the in vivo chemo preventive activity of GA on 7,12-Dim-ethylbenz[a]anthracene (DMBA)/Croton oil induced two-step skin carcinogenesis in Swiss albino mice.Skin tumor incidence and tumor volume were recorded during the 16 weeks of experimental period.In addition, LDH-isozyme shift, skin collagen content, activities of matrix metalloproteinases (MMP-2/MMP-9) enzymes and enzymatic and non-enzymatic antioxidant were studied in the skin and serumof experimental mice. Tumor incidence was significantly increased in the DMBA/Croton oil induced mice(100%; p < 0.001) when compared to GA co-treated mice (60%; p < 0.01) and 5-FU treated mice (50%;p < 0.01). Skin collagen content, MMPs activities, LDH-isoenzymes and MMP-2/-9 expressions wereincreased in DMBA/Croton oil induced skin while decreased levels of enzymatic (GST, SOD, CAT & GPx)and non-enzymatic anti-oxidant (GSH) were noticed. On the other hand, GA co-treatment exhibited a sig-nificant protection by reverting back the altered levels of LDH-isoenzymes, antioxidants, collagen andMMP-2/MMP-9 activities. The results of this study indicate that topical application of GA inhibitsDMBA/Croton oil induced two-stage skin carcinogenic process by modulating the antioxidants and MMPs(-2 & -9) in the mouse skin.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction these natural antioxidants are reported to have the ability to en-
Skin is the largest organ of the human body, and is a major tar-get for toxic insult by a chemical mutagens and carcinogens in aday-to-day life (Simna et al., 2012). Throughout the world, thenumber of people affected by skin cancer is increasing due to in-creased exposure to environmental pollutants and UV rays(Narendhirakannan and Angeline Christie Hannah, 2013). Recentstudies have also predicted that the number of people affectedby this malady will be doubled by the year 2050 (Hayat et al.,2007). Although many synthetic drugs are commercially availableto treat skin cancer; However, these drugs causes serious side ef-fects to patients such as nephrotoxicity, neurotoxicity, infertility,thromboembolic complications, hair loss, nausea, and myocardialinfarction (Shabaruddin et al., 2013). Hence, there is a dire needto look for more efficacious agents with lesser side effects for theprevention and eradication of skin tumors.
Number of recent evidence has shown that natural dietary anti-oxidants are capable of inhibiting cancer cell growth. Moreover,
hance the activities of detoxification enzyme components such asglutathione-S-transferase (GST). The activation of GST not onlyneutralizes the chemical carcinogens, but also preventing themfrom forming a DNA adducts in the genome (Liu et al., 1992). Fur-thermore, it has been suggested that these natural antioxidantcompounds can be used in multiple ways, either as cancer preven-tive agents or even as cancer therapy drugs (Thornthwaite et al.,2013).
Gallic acid (GA), a polyhydroxyphenolic compound, is abun-dantly present in leafy vegetables, fruits, and nuts such as gallnut,grapes, sumach, oak bark, green tea, apple peels (Atkinson et al.,2004; Chakraborty et al., 2009; Kim et al., 2006; Ng et al., 2004;Liao et al., 2012) and also present in onion and potato varieties(Albishi et al., 2013). GA exhibits variety of biological activitiesincluding antioxidant (Isuzugawa et al., 2001; Yogendra Kumaret al., 2013), anti-tumor (Agarwal et al., 2006; Liao et al., 2012;Lee et al., 2013; Locatelli et al., 2013), anti-inflammatory (Albishiet al., 2013) and anti-bacterial (Kang et al., 2008; Lee and Je,2013). GA has been shown to inhibit gastric cancer cell growthin vitro by modulating the levels of MMP-2 and -9 and cytoskeletalreorganization (Ho et al., 2010). Furthermore, GA has also been

V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55 45
shown to suppress cell viability, proliferation, invasion and angio-genesis in human glioma cells, in vitro (Lu et al., 2010; Lee et al.,2013). Lo et al. (2011) have also demonstrated that GA inhibitsthe migration and invasion of cultured A375.S2 human melanomacells by inhibiting matrix metalloproteinase-2. GA derivatives suchas tannic acid and ellagic acid have been shown to exhibit antitu-mor activities in mouse skin (Perchellet et al., 1992). GA has beenshown to selectively induce apoptosis only in the pancreatic cancercells without affecting the survival of normal cells in in vitro condi-tion (Liu et al., 2012). Although several line of evidence has shownthat GA inhibits tumor cells growth, migration and invasion in vitro(Chen et al., 2012). However, a detailed in vivo studies have notbeen carried out to examine the role of GA on tumor cell growth,invasion and on MMPs.
In this study, we have utilized a two stage in vivo skin carcino-genesis model, because it is a suitable model system to study boththe initiation and progression phases of the tumorigenesis. The re-sults of this study indicates that topical application of GA inhibitsthe skin tumor initiation and progression phases of the skin carcin-ogenic process by modulating the antioxidant enzyme system andMMPs (-2 & -9) in the mouse skin tissue.
2. Materials and methods
2.1. Chemicals
7,12-Dimethylbenzanthracene (DMBA) (Cat. No. D3254), and Gallic acid (Cat.No. 27645) were purchased from Sigma, St. Louis, MO, USA. Croton oil was procuredfrom Aroma essential oils, Haryana, India. 5-Fluorouracil was purchased from FlukaBio Chemika (Buchs, Switzerland). Primary antibodies for MMP-2 (Cat. No. sc-10736), MMP-9 (Cat. No. sc-10737) and HRP-labeled secondary antibody (Cat. No.sc-2749) were procured from Santa Cruz biotechnology, San Diego, CA, USA. Nitro-blue tetrazolium (NBT), Nucleotide adenosine diphosphate (NAD+), riboflavin andPhenazine methasulphate (PMS) were obtained from SD Fine and SRL ChemicalsLtd., India. All other chemicals used were of analytical grade, purchased from SiscoResearch Laboratories Pvt. Ltd., Mumbai, India.
2.2. Animal
Adult male Swiss albino mice, 8–10 weeks old (25–30 g) were obtained fromTamil Nadu Veterinary and Animal Science University, Chennai, India, and werehoused in polypropylene cages. The animals were acclimatized for 1 week priorto the start of the experiment. Mice were fed with commercially available GoldMohr pelleted diet supplied by Lipton India Pvt. Ltd. (Bangalore, India) and waterad libitum; maintained on standard housing conditions under controlled atmo-sphere with 12:12 h light/dark cycles with an ambient temperature of 25 ± 5 �Cand humidity at 50 ± 10%. This study was conducted according to the ethical normsapproved by Ministry of Social Justices and Empowerment, Government of Indiaand by Animal Ethics Committee Guidelines of our Institution (IAEC NO. 12/02/2012).
2.3. Experimental design
Animals were divided into five groups (10 mice/Group) as follows.
� Group I (Control): Mice were topically applied with acetone alone throughoutthe course of the experiment.� Group II (Induction): Mice received topical application of single dose of DMBA
(100 lg/100 lL in acetone) over the shaved dorsal area of the skin. After2 weeks, Croton oil (1% in acetone) was applied 3 times a week for 16 weeks.� Group III (Co-treatment): Mice received topical application of GA (25 mg/kg body
weight per day) 10 min before the application of DMBA/Croton oil as that ofGroup II mice.� Group IV (Positive control): Mice received topical application of 0.5% of 5-Fluro-
uracil (5-FU) 10 min before the application of DMBA/Croton oil as that of GroupII mice (Wilgus et al., 2004; Olson and Wattenberg, 1969).� Group V (Drug control): Mice received topical application of GA (25 mg/kg body
weight per day) alone three times a week for 16 weeks.
During the 16 weeks of experiments, mice were observed daily for the appear-ance of skin papillomas and tumor volume was recorded. At the end of 16 weeks, allthe mice were sacrificed and skin tissue and serum samples were collected.
2.4. Preparation of skin and tissue homogenate
The skin tissue was separated and washed several times in an ice-cold saline.Skin tissue (100 mg) was carefully chopped into fine pieces using a surgical blade.The minced skin tissue was homogenized in an ice-cold RIPA buffer, pH 7.4 with ahelp of homogenizer (Remi model-RQ-127A/D). Typically, the homogenizer was setat a speed of 1000 rpm, 10 strokes for 30 s for 2–3 times at 4 �C were applied to geta complete homogenate of the skin tissue. The resultant homogenate was centri-fuged at 20,000 rpm for 5 min at 4 �C. The tissue debris was separated as a pelletand the tissue supernatant was pipette out. The skin tissue homogenates and serumwere stored at �20 �C until further use. Protein quantification was carried out usingthe method of Bardford Protein Assay (Simna et al., 2012).
2.5. Electrophoretic separation of Lactate Dehydrogenase (LDH) Isoenzymes
LDH isoenzymes consist of five isoforms, and are encoded by LDHA and LDHBgene. The polypeptide subunits combine to form two pure types of isoenzymeswhich referred as H and M, LDH-1 consists of 4-polypeptide subunits of H type,while LDH-5 consists of 4-polypeptide subunits of M type and LDH-2 (H3M),LDH-3 (H2M2), and LDH-4 (HM3) consists of hybrids of H and M type subunits(Markert, 1963). The LDH isoenzymes were separated as described by Van Der Helmet al. (1962). Briefly the serum protein (10 lg) was loaded on a 1% agarose gel pre-pared with 25 mM Tris (pH 9.5). Tris–glycine buffer, pH 9.5 was used as a runningbuffer and the electrophoresis was carried out for 1 h at 100 V. After the electropho-resis, the gel was washed in 0.1 M Tris–HCl (pH 8.5). Then, gel was incubated in aLDH staining solution containing lactate (3.24 mg/ml), nicotinamide adenine dinu-cleotide (0.3 mg/ml), nitroblue tetrazolium (0.8 mg/ml) and phenazine methosul-fate (0.167 mg/ml) dissolved in 0.01 M Tris–HCl (pH 8.5). LDH isoenzymes wereseparated depending on their charge and molecular weight and appeared as a clearzone. LDH-5 isoenzyme moved toward negative pole because of its net positivecharge while isoenzymes move toward positive pole. The relative activity of eachisoenzyme was determined by densitometry analysis.
2.6. Glutathione-S-transferase (GST), enzymatic and non-enzymatic antioxidants
Skin GST enzyme activity was measured by modified method of Habig et al.,1974. The GST activity was expressed as lmoles of 1-chloro-2,4-dinitro benzene(CDNB) conjugated per minutes per mg of protein. Enzymatic antioxidants – super-oxide dismutase (SOD) activity was based on the inhibition of epinephrine–adreno-chrome transition and the activity of enzyme was expressed as units/min/mgprotein (Marklund and Marklund, 1974). Catalase (CAT) activity was quantifiedby the amount of H2O2 decomposed/min/mg protein (Sinha, 1972). Glutathioneperoxidase (GPx) activity was estimated based on its substrate glutathione as de-scribed by Rotruck et al. (1973), and non-enzymatic antioxidant reduced glutathi-one (GSH) which involves the reaction of glutathione with dithio bis-nitrobenzoic acid (DTNB) to give a compound which has an absorption maximum at412 nm (Moron et al., 1979) were estimated as described previously.
2.7. Total serum activity of Lactate Dehydrogenase (LDH) and lipid peroxidation (LPO)
Serum LDH was estimated as described by King (1965), which is a ubiquitousglycolytic enzyme that catalyzes reversible oxidation of lactate to pyruvate withconcomitant reduction of NAD. When the enzyme is supplied with lactate andNAD+, the LDH catalyzed reaction starts to produce pyruvate. LDH activity was ex-pressed as lg/dl of serum. Lipid peroxidation (LPO) was analyzed by previously de-scribed method (Okhawa et al., 1979). LPO activity was expressed as (TBARS) mmol/lg protein.
2.8. Substrate gel analysis of SOD and Catalase CAT activity
To further validate the enzymatic activity assay of SOD and CAT, substrate nativegel electrophoresis was carried out. The SOD activity was detected in a native PAGEas described by the method of Beauchamp and Fridovich (1971). The CAT enzymeactivity was detected in a Native PAGE as described by the method of Woodburyet al. (1971). Protein sample (30 lg/lane) was mixed with 50% glycerol and loadingdye and run on the 8% Native PAGE gel constantly at 100 V, �1 h in 4 �C. For SODactivity analysis, the gel was soaked in a Riboflavin-NBT solution (2–5 ml) andincubated at RT for 15 min in dark (Riboflavin is light sensitive). After incubation,the Riboflavin-NBT solution was removed and 2–5 ml of 0.1% TEMED was addedand incubated at RT for15 min in dark. Remove the staining solution and keep thegel in light which induces super oxide synthesis. For CAT activity, the gel was exten-sively rinsed with double distilled water followed by incubation with 0.003% H2O2
for 10 min. The gel was stained with 2% potassium ferric cyanide and 2% ferric chlo-ride solution. The clear zone in the gel indicates the CAT activity.

Fig. 1. Effect of GA on tumor incidence and tumor volume in the control and experimental group of animals. Where, plate 1a – control, plate 1b – DMBA induced, plate 1c –DMBA + GA treated mice, and plate 1d – DMBA + 5-FU treated mice.
46 V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55
2.9. Quantification of collagen and gelatin zymography of matrix metalloproteinases
Skin collagen content was quantified using a modified method of Bergman andLoxley. The MMP-2 & -9 activities were studied by substrate-gelatin zymography(Vellaichamy et al., 2005). The skin tissue homogenate was separated on a 7.5% na-tive-PAGE containing 1 g/L gelatin under non-reducing conditions without priorboiling. After the electrophoresis, the gel was washed with 2.5% Triton X-100 for1 h at RT and the protein was allowed to denature with subsequent immersion ina mixture containing 50 mM Tris (pH 7.5) and 20 mM CaCl2 for 16 h at 37 �C. Thegel was then stained with 0.25% Coomassie Brilliant Blue R250/40% aqueous/10%acetic acid and destained in 25% aqueous/8% acetic acid. The appearance of clearband indicates the gelatin lysis by enzymatic action. The density of white bandwas calculated as relative expression of enzyme activity.
2.10. Western blot analysis of MMP-2 and MMP-9
The SDS–PAGE resolved proteins were electrophoretically transferred to a poly-vinylidene fluoride (PVDF) membrane. For immuno detection, the PVDF membranewas blocked with 5% non fat dry milk (sigma) in phosphate-buffered saline for1.30 h at room temperature, and then probed with MMP-2 (rabbit polyclonal anti-body at a dilution of 1:1000); b-actin (mouse monoclonal antibody at a dilution of1:2000) overnight at 4 �C. The blots were then extensively washed in PBST and thenincubated with anti-rabbit and anti-mouse HRP labeled secondary antibody (SantaCruz) respectively, at a dilution of 1:2500 for 2 h at room temperature. After exten-sive washes in PBST, the bands were visualized by treating the membranes in asolution of 0.02% DAB containing 0.01% H2O2. The membranes were photographedand quantitated with image analysis software, ImageJ. Densitometry data are ex-pressed as ‘‘fold change’’ as compared with control (Gopi et al., 2013).
2.11. Histological evaluation
The skin tissues were fixed in 10% formaldehyde, processed and embedded inparaffin wax, sectioned to the thickness of 2–3 lm in a rotary microtome andstained with hematoxylin and eosin. Sections were evaluated under Nikon EclipseE-200 Photomicrographic System (Ortega et al., 2003).
2.12. Sirius red staining of skin collagen
To determine the collagen content, 2–3 lm thickened skin tissue sections werestained with 0.1% picrosirius red (10 mg of sirius red in 10 ml of aqueous picric acidsolution) as described previously (Aparna and Charu, 2010). These sections wereanalyzed morphometrically by light microscope. Collagen deposition appeared asred color.
2.13. Statistical analysis
All data obtained are represented as mean ± SEM for 10 animals in each group.The results were computed statistically (SPSS/10 Software Package; SPSS Inc.) usingone-way ANOVA. In all tests, the level of statistical significance was set at p < 0.05.
3. Results
3.1. Skin papillomas assessments
Table 1 shows the tumor incidence, tumor delay, papilloma/mouse, and tumor volume in control and experimental groups ofmice. DMBA/Croton oil treated (Group II) mice showed 100% tumorincidence, while GA and 5-FU co-treated mice exhibited a markedreduction in the tumor incidence (60% and 50%; respectively). Skinpapillomas started appearing in the 5th week of promoter applica-tion, whereas, the papilloma appearance was delayed by 4 weeks,respectively in the GA and 5-FU treated mice group. Further, a sig-nificant reduction (p < 0.01) in number of papilloma per mousewas noticed in the GA and 5-FU treated mice as compared withDMBA/Croton oil treated mice group (10 ± 0.5 vs. 2.09 ± 0.13 and10 ± 0.5 vs. 2.53 ± 0.03 papillomas/mouse, respectively). The tumorvolume was significantly reduced in GA and 5-FU co-treated micegroup as compared with DMBA/Croton oil alone induced mice

Table 1Effect of GA on tumor incidence, tumor delay, tumor number and tumor volume of control and experimental group of animals.
Groups Tumor incidence (%) Tumor appearance/delay in weeks No of tumors/mouse Tumor volume (mm3) Mortality
Control 0 – 0 0 0GA alone 0NS – 0NS 0NS 0DMBA Induced 100a 5 10.05 ± 0.2a 126 ± 3.06a 3DMBA + GA 60b 9 2.09 ± 0.13b 18 ± 0.9b 0DMBA + 5-FU 50b 9 2.53 ± 0.03b 28 ± 0.9b 1
Values are expressed as mean ± SD (n = 10 mice). NS (Non significant)-control vs. drug control.a p < 0.001, control vs. DMBA induced.b p < 0.01, DMBA induced vs. DMBA + GA treated, DMBA induced vs. DMBA + 5-FU treated mice.
Fig. 2. Histopathological analysis of skin sections of control and experimental groups of animals (hematoxylin and eosin staining-10�). Sections from control, DMBA, andDMBA + Croton oil treated skin tissues were stained with H&E for morphological assessment as described in the materials and methods section. Where, plate 2a – skin sectionfrom control mice, plate 2b – skin section from DMBA induced for 2 weeks mice, plate 2c – DMBA/Croton oil induced for 12 weeks mice skin section, plate 2d – DMBA/Crotonoil induced for 16 weeks mice skin section, plate 2e and plate 2f – skin sections from GA and 5-FU co-treated animals, respectively. Green arrow indicates the intact skinlayers with normal structure of epidermis and secretary glands in the dermis. Yellow arrow indicates the disturbed and ruptured secretary glands in the dermis region. Blackarrow indicates the hyperplasic, increased proliferative cells and melanocytes in the epidermal area. Blue arrow indicates the decreases in the cell count and melanocytes inthe recovering epidermis to normal structure. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55 47

48 V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55
group (126 ± 3.06 mm3 vs. 18 ± 0.9 mm3; and 126 ± 3.06 mm3 vs.28 ± 0.9 mm3, p < 0.01 respectively). GA alone applied mice skindid not exhibit any adverse effects.
Fig. 1 shows the representative photographs of the skin papillo-mas in control and experimental group of mice. Plate 1a shows thecontrol mouse skin. DMBA/Croton oil treated mice skin exhibitsskin papillomas (plate 1b). A significant reduction in the numberof papilloma and tumor volume was observed in the GA and 5-FU treated mice (plates 1c and Id, respectively).
3.2. Histopathological/morphological observation
Fig. 2(a–f) shows the H&E staining of control and experimentalgroup of mice skin sections. The vehicle alone treated control miceskin sections exhibited well defined subcutaneous tissue and intact
Fig. 3. Effect of GA on LDH levels and LDH-isoenzyme profile in serum of control and expserum of control and experimental group of mice. LDH-isoenzyme pattern of experimenDMBA + GA treated mice, lane 4 – DMBA + 5-FU treated mice, and lane 5 – GA alone treatbands. Values are expressed as mean ± SD (n = 10 mice). NS (Non significant)-control vsDMBA induced; b p < 0.01, control vs. DMBA induced, DMBA induced vs. DMBA + GA tDMBA + GA treated, DMBA induced vs. DMBA + 5-FU treated mice.
epithelial layer (Fig. 2a). DMBA alone painted (2 weeks) skin tissuesection showed initial stages of hyperplasia, hyperkeratosis, anddysplasia (Fig. 2b). A moderate to severe hyperplasia, hyperkerato-sis and increased melanocytes are seen in DMBA/Croton oil(12 weeks) treated mouse skin section (Fig. 2c). Severe hyperplasiawith well-differentiated squamous cell carcinoma is evident in16 weeks of DMBA/Croton oil treatment (Fig. 2d). GA and 5-FUtreated mice sections showed a significant decrease in hyperplasia,and skin tissue recovering to normal structure (Fig. 2e and f,respectively).
3.3. Effect of GA on serum marker enzyme
Serum LDH activity and LDH-isoenzymes profile of control andexperimental group of mice are represented in Fig. 3A and B. It is
erimental groups of mice. A. shows the lactate dehydrogenase (LDH) activity in thetal animals was shown in the B. Lane 1 – control, lane 2 – DMBA induced, lane 3 –ed mice. C. shows the representative densitometric analysis of clear LDH-isoenzyme. drug control, DMBA induced vs. DMBA + 5-FU treated mice; a p < 0.001, control vs.reated, DMBA induced vs. DMBA + 5-FU treated; and c p < 0.05, DMBA induced vs.

Table 2Effect of GA on phase II detoxification enzyme in skin tissue of control andexperimental animals.
Groups GST (lmol of CDNB conjugated/min mgprotein)
GSH (lg/mgprotein)
Control 6.46 ± 0.57 3.03 ± 0.20GA alone 6.40 ± 0.31NS 2.94 ± 0.09NS
DMBAinduced
2.80 ± 0.23a 1.52 ± 0.12a
DMBA + GA 6.03 ± 0.51b 2.44 ± 0.10b
DMBA + 5-FU
5.90 ± 0.29b 2.57 ± 0.11b
Values are expressed as mean ± SD (n = 10 mice). NS (Non significant)-control vs.drug control.
a p < 0.001, control vs. DMBA induced.b p < 0.01, DMBA induced vs. DMBA + GA treated, DMBA induced vs. DMBA + 5-
FU treated.
V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55 49
evident that, the total LDH activity significantly increased(p < 0.001) in DMBA/Croton oil induced (Group II) animals whencompared with control mice (Group I). In addition, there was a sig-nificant increase in the levels of LDH-isoenzymes [LDH-5 (3-fold),LDH-4 (14-fold), LDH-3 (9-fold), LDH-2 (2-fold) and a slight in-crease in LDH-1] were noticed in the serum of DMBA/Croton oil in-duced (Group II) mice. GA co-treatment (Group III) significantlydecreased the elevated serum levels of LDH total activity andLDH-isoenzymes to near normal level as compared with DMBA/Croton oil induced (Group II) mice. No significant change wasfound in the serum of GA alone treated (Group V) mice as com-pared with that of control mice (Group I).
3.4. Effect of GA on phase II xenobiotic enzyme and reducedglutathione
Table 2 shows the phase II enzyme activity in skin tissue of con-trol and experimental group of mice. The levels of GST and GSHwere significantly decreased (p < 0.001, respectively) in DMBA/Croton oil induced (Group II) mice skin when compared with con-trol (Group I). GA co-treated (Group III) mice showed a significantincrease (p < 0.01) in GST activity and GSH when compared withDMBA/Croton oil induced mice (Group II). Similar treatment wasnoticed in 5-FU (positive drug) treated mice (Group IV). No signif-icant changes were observed in GA alone treated (Group V) mice.
3.5. Effect of GA on enzymatic anti-oxidants and lipid peroxidation(LPO)
Effect of GA on enzymatic anti-oxidants levels in skin of controland experimental groups of mice are depicted in Table 3. The levelsof enzymatic anti-oxidants in DMBA/Croton oil induced models(Group II) were decreased (p < 0.01, respectively) when comparedto control (Group I). GA co-treatment (Group III) normalized thedecreased levels of enzymatic anti-oxidants when compared to
Table 3Effect of GA on status of enzymatic, non-enzymatic antioxidants in skin tissue of control
Groups SOD (units/min/mg protein) CAT (lmoles of H2O2 u
Control 8.41 ± 0.38 40.17 ± 3.13GA alone 7.98 ± 0.31NS 39.6 ± 2.42NS
DMBA induced 4.92 ± 0.22a 25.57 ± 2.76a
DMBA + GA 7.19 ± 0.59a 38.94 ± 3.09a
DMBA + 5-FU 7.91 ± 0.52a 39.74 ± 4.53a
Values are expressed as mean ± SD (n = 10 mice). NS (Non significant)-control vs. drug ca p < 0.01, control vs. DMBA induced, DMBA induced vs. DMBA + GA treated, DMBA in
DMBA/Croton oil induced (Group II) mice. GA effect was compara-ble with the positive drug 5-FU co-treatment.
Fig. 4 reveals the substrate gel activities of SOD, CAT and the le-vel of LPO in control and experimental group of mice skin tissue.The lipid peroxide level was significantly increased (p < 0.001) inDMBA/Croton oil induced group of mice (Group II) while this levelwas reverted back to near normal level upon treatment with GA(Group III) and 5-FU (Group IV). The significant decrease was ob-served in the activities of SOD (2-fold) and CAT (4-fold) inDMBA/Croton oil treated mice when compared to control mice.Upon co-treatment with GA and 5-FU the decreased activities ofSOD and CAT were significantly reverted back to near normal levelas compared with Group II experimental animals. No significantchanges were seen in drug control mice (Group V).
3.6. Effects of GA on MMP-2/-9 activities and expression
Fig. 5A and B reveals the gelatin zymographic activity assay ofMMP-9, MMP-2 and densitometric assay of MMPs in skin tissueof control and experimental group of mice. DMBA/Croton oil trea-ted mice skin exhibited a 4-fold increase in MMP-2 and a 3-fold in-crease in MMP-9 activities when compared to control mice skin.GA co-treatment significantly reduced the activities of bothMMP-2 and MMP-9 and GA effect was comparable to 5-FU co-treatment. Drug control (Group V) mice did not show anyabnormalities.
Fig. 6A and B shows the Western blot expression analysis ofMMP-9 and MMP-2 in control and experimental group of mice.DMBA/Croton oil treatment significantly increases the expressionof MMP-9 (3.6-fold) and MMP-2 (2.7-fold) when compared to con-trol (Group I). Upon co-treatment with GA and 5-FU, a significantdecrease was observed in the MMP-2/-9 expression as comparedwith DMBA/Croton oil induced (Group II) mice. No significantchanges were noticed in GA alone treated (Group V) mice.
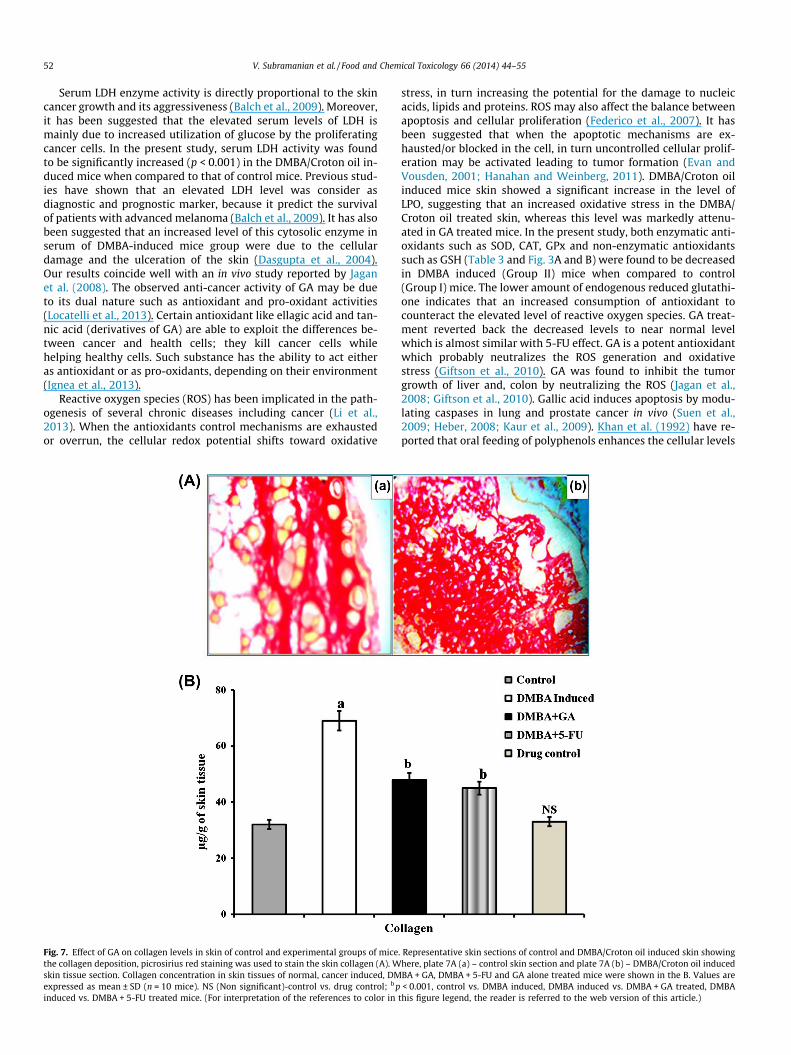
Fig. 7A illustrates the collagen deposition by picro sirius redstaining. Fig. 7B shows the collagen content in skin tissue of controland experimental mice. It is evident that the DMBA/Croton oil trea-ted skin section showed increased deposition of collagen (in-creased picro sirius red staining), as compared with normalcontrol skin section. The collagen concentration was elevated(p < 0.001) in DMBA induced mice skin while co-treatment withGA and 5-FU, normalized the altered level to near normal level(p < 0.01). No significant changes were noticed in GA alone treated(Group V) mice.
4. Discussion
The results of the present study demonstrate that topical appli-cation of GA, a natural antioxidant, markedly suppresses the initi-ation and progression phases of DMBA/Croton oil induced twostage skin carcinogenesis by modulating the phase II detoxificationenzyme, antioxidants, and MMP-2/-9 activities.
and experimental animals.
tilized/min/mg protein) GPx (lmoles of GSH utilized/min/mg protein)
43.70 ± 2.0542.02 ± 2.83NS
28.52 ± 1.31a
42.66 ± 2.10a
41.04 ± 4.23a
ontrol;duced vs. DMBA + 5-FU treated mice.

50 V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55
The sequential steps of skin cancer development include: tumorinitiation – stage 1 (conversion/neoplasia), promotion – stage 2(propagation/dysplasia or hyperplasia), and progression – stage 3(metastatic). The papilloma appearance on the skin is consideredas the primary indication of carcinogenic process. Our resultsshows that there was 100% tumor incidence in DMBA/Croton oilapplied mice while the tumor incidence was markedly reducedupon co-treatment with GA (60%). Histopathological observation
Fig. 4. Effect of GA on SOD, CAT and LPO activity in skin of control and experimental gro(B) activities in control and experimental group of mice. Each lane was loaded with 50 lfigure A lane 1 – control, lane 2 – DMBA induced, lane 3 – DMBA + GA treated mice, lane 4densitometric analysis of SOD and CAT bands respectively. E elucidates the LPO levels inmean ± SD (n = 10 mice). NS (Non significant)-control vs. drug control; a p < 0.001, conDMBA + GA treated, DMBA induced vs. DMBA + 5-FU treated.
of skin sections obtained from experimental mice also confirmsthat there was a stage wise increase in cell proliferation, hyperpla-sia and dysplasia of skin epidermis only in the DMBA/Croton oiltreated mice group (Fig. 2b–d). GA treated mice skin showed an in-tact epidermis, suggesting the anti-proliferative effect of GA. Ourresults coincide well with the previous report of Ortega et al.(2003).
ups of mice. Representative native polyacrylamide gels stained for SOD (A) and CATg of protein and electrophoresed through an 8% polyacrylamide gel at 4 �C. In this– DMBA + 5-FU treated mice, and lane 5 – GA alone treated mice. C and D show the
the skin of control and various experimental groups of mice. Values are expressed astrol vs. DMBA induced; b p < 0.01, control vs. DMBA induced, DMBA induced vs.

Fig. 5. Effect of GA on MMP activity in skin of control and experimental groups of mice. Representative gelatin zymographic analysis of normal, DMBA applied and treatedmice skin was shown in the A. Gels were stained with 0.25% coommassie brilliant blue and destained until the clear lytic bands were visible. Lane 1 – control, lane 2 – DMBAinduced, lane 3 – DMBA + GA treated mice, lane 3 – DMBA + 5-FU treated mice, and lane 5 – GA alone treated mice. B depicts the densitometric analysis of MMP-2 and MMP-9bands. Data are expressed as mean ± SEM (n = 10 mice). Values are expressed as mean ± SD (n = 10 mice). NS (Non significant)-control vs. drug control; a p < 0.001, control vs.DMBA induced; b p < 0.01, DMBA induced vs. DMBA + GA treated, DMBA induced vs. DMBA + 5-FU treated mice. (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
Fig. 6. Effect of GA on MMP-2 and MMP-9 expression in skin tissue of control and experimental groups of mice. Representative immunoblot analysis for MMP-9, MMP-2expression (A). PVDF membrane was developed with DAB and band intensity was analyzed with imageJ software (B). Lane 1 – control, lane 2 – DMBA induced, lane 3 –DMBA + GA treated mice, lane 3 – DMBA + 5-FU treated mice, and lane 5 – GA alone treated mice. Data are represented as mean ± SEM (n = 10 mice). Values are expressed asmean ± SD (n = 10 mice). NS (Non significant)-control vs. drug control; a p < 0.001, control vs. DMBA induced; b p < 0.01, DMBA induced vs. DMBA + GA treated, DMBA inducedvs. DMBA + 5-FU treated mice.
V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55 51
52 V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55
Serum LDH enzyme activity is directly proportional to the skincancer growth and its aggressiveness (Balch et al., 2009). Moreover,it has been suggested that the elevated serum levels of LDH ismainly due to increased utilization of glucose by the proliferatingcancer cells. In the present study, serum LDH activity was foundto be significantly increased (p < 0.001) in the DMBA/Croton oil in-duced mice when compared to that of control mice. Previous stud-ies have shown that an elevated LDH level was consider asdiagnostic and prognostic marker, because it predict the survivalof patients with advanced melanoma (Balch et al., 2009). It has alsobeen suggested that an increased level of this cytosolic enzyme inserum of DMBA-induced mice group were due to the cellulardamage and the ulceration of the skin (Dasgupta et al., 2004).Our results coincide well with an in vivo study reported by Jaganet al. (2008). The observed anti-cancer activity of GA may be dueto its dual nature such as antioxidant and pro-oxidant activities(Locatelli et al., 2013). Certain antioxidant like ellagic acid and tan-nic acid (derivatives of GA) are able to exploit the differences be-tween cancer and health cells; they kill cancer cells whilehelping healthy cells. Such substance has the ability to act eitheras antioxidant or as pro-oxidants, depending on their environment(Ignea et al., 2013).
Reactive oxygen species (ROS) has been implicated in the path-ogenesis of several chronic diseases including cancer (Li et al.,2013). When the antioxidants control mechanisms are exhaustedor overrun, the cellular redox potential shifts toward oxidative
Fig. 7. Effect of GA on collagen levels in skin of control and experimental groups of micethe collagen deposition, picrosirius red staining was used to stain the skin collagen (A). Wskin tissue section. Collagen concentration in skin tissues of normal, cancer induced, DMexpressed as mean ± SD (n = 10 mice). NS (Non significant)-control vs. drug control; b pinduced vs. DMBA + 5-FU treated mice. (For interpretation of the references to color in
stress, in turn increasing the potential for the damage to nucleicacids, lipids and proteins. ROS may also affect the balance betweenapoptosis and cellular proliferation (Federico et al., 2007). It hasbeen suggested that when the apoptotic mechanisms are ex-hausted/or blocked in the cell, in turn uncontrolled cellular prolif-eration may be activated leading to tumor formation (Evan andVousden, 2001; Hanahan and Weinberg, 2011). DMBA/Croton oilinduced mice skin showed a significant increase in the level ofLPO, suggesting that an increased oxidative stress in the DMBA/Croton oil treated skin, whereas this level was markedly attenu-ated in GA treated mice. In the present study, both enzymatic anti-oxidants such as SOD, CAT, GPx and non-enzymatic antioxidantssuch as GSH (Table 3 and Fig. 3A and B) were found to be decreasedin DMBA induced (Group II) mice when compared to control(Group I) mice. The lower amount of endogenous reduced glutathi-one indicates that an increased consumption of antioxidant tocounteract the elevated level of reactive oxygen species. GA treat-ment reverted back the decreased levels to near normal levelwhich is almost similar with 5-FU effect. GA is a potent antioxidantwhich probably neutralizes the ROS generation and oxidativestress (Giftson et al., 2010). GA was found to inhibit the tumorgrowth of liver and, colon by neutralizing the ROS (Jagan et al.,2008; Giftson et al., 2010). Gallic acid induces apoptosis by modu-lating caspases in lung and prostate cancer in vivo (Suen et al.,2009; Heber, 2008; Kaur et al., 2009). Khan et al. (1992) have re-ported that oral feeding of polyphenols enhances the cellular levels
. Representative skin sections of control and DMBA/Croton oil induced skin showinghere, plate 7A (a) – control skin section and plate 7A (b) – DMBA/Croton oil inducedBA + GA, DMBA + 5-FU and GA alone treated mice were shown in the B. Values are< 0.001, control vs. DMBA induced, DMBA induced vs. DMBA + GA treated, DMBA
this figure legend, the reader is referred to the web version of this article.)

Fig. 8. Schematic representation of molecular and cellular events of GA on two-step skin carcinogenesis. Figure shows the molecular and cellular actions of GA on sequentialsteps such as initiation, promotion stages of skin carcinogenesis. GA increases levels of cellular anti-oxidant enzymes thereby delays the initiation process of tumorigenesisand prevents tumor metastasis by inhibiting MMPs activity and collagen accumulation.
V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55 53
of both enzymatic and non-enzymatic antioxidant system andphase II detoxification enzymes system. The phase II enzyme sys-tem comprising glutathione-S-transferase (GST), GSH participatesin the detoxification of xenobiotics which inactivates chemical car-cinogens into a less toxic form and converts DNA damaging entitiesinto metabolites that can be easily excreted from the body (Kaurand Arora, 2013). GST enzyme has an important role as it initiatesthe process of detoxification by neutralizing the active sites of elec-trophiles by binding with the –SH group of glutathione. In the pres-ent study, we found that GST activity and GSH was significantlydecreased in the skin of DMBA induced mice when compared tocontrol mice skin. Upon GA co-treatment, the altered GST andGSH level was reverted back to near normal level (in Table 3), sug-gesting that GA exhibits its chemo preventive activity by enhanc-ing the drug detoxification system (Giftson et al., 2010).
Matrix metalloproteinases (MMPs) are zinc dependent enzymesreported to involved in the initial stage of invasion and metastasisof various tumor cells (Farris et al., 2011; Liao et al., 2012). Overexpression of MMPs has been observed in variety of metastatic tu-mor tissues and also several anticancer drug formulations mainlyfocus to attenuate the expression of MMPs (Rasmussen andMcCann, 1997). Numerous reports have been shown that the inhi-bition of MMP expression and/or inhibition of the activities ofMMP enzymes can be used as early targets for preventing cancermetastasis (Chiodoni et al., 2010; Chien et al., 2012). Therefore,great attention is focused on developing agents or drugs that caninhibit metastasis; however, the anti-metastatic agents are stilllacking. MMP-2 (72-kDa gelatinase A) and MMP-9 (92-kDa gelatin-ase B) are involved in the invasive metastatic potential of tumorcells (Pires et al., 2013). In the present study, 4-fold and 3-fold in-crease in the activities of MMP-9 (p < 0.001) and MMP-2 (p < 0.001)respectively, was observed in the DMBA-induced when comparedto that of control group. The increases in MMP-9 and MMP-2expression were also further confirmed with immunoblot analysis(Fig. 6). Our results demonstrated that anti-metastatic effects ofGA, which are associated with decreased MMPs. It has been sug-
gested that the migration and invasion of human melanoma cellsthrough the MMPs (Chen and Chang, 2012).
Collagen, a main constituent of extracellular matrix (ECM)which has vital role in the attachment of cancer cells to other or-gans during metastatic process (Jeon et al., 2013). Our results alsoshow that there was a significant increase of collagen content(p < 0.001) in DMBA induction when compared to control. This in-creased collagen level was reverted back to near normal level uponGA treatment. Moreover, picrosirius red staining of collagen whichstrongly evidence that, an increased collagen accumulation in theepidermis of DMBA induced skin. It has been suggested that an in-creased accumulation of extracellular matrix may influence tumorprogression (Kligman et al., 1989; Provenzano et al., 2008; Aparnaand Charu, 2010). GA treated skin showed a reduction in collagendeposition which further confirms the anti-migration property ofGA. Nakamuta et al. (2005) showed that gallates from green teasuppress both collagen and collagenase activity in hepatic cells.
Fig. 8 shows the schematic representation of role of GA insequential steps of chemical induced skin carcinogenic process.GA prevents DNA adducts formation by activating GST and enhanc-ing enzymatic anti-oxidants, whereas in progression stage it pre-vents the tumor cells from metastasis by blocking MMP-2 andMMP-9. Taken together, our findings suggest that GA has a potentchemopreventive activity and suppress the DMBA/Croton oil in-duced two step skin carcinogenesis in in vivo model by modulatingthe antioxidant systems and MMPs of mouse skin.
Conflict of Interest
The authors declare that there are no conflicts of interest.
Transparency Document
The Transparency document associated with this article can befound, in the online version.

54 V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55
Acknowledgements
Authors wishes to thank University Grant Commission (UGC),Department of Biotechnology (DBT), and Indian Council of MedicalResearch (ICMR), India for the financial support in the form of re-search fellowship (VM) and research projects (VE).
References
Agarwal, C., Tyagi, A., Agarwal, R., 2006. Gallic acid causes inactivatingphosphorylation of cdc25A/cdc25C-cdc2 via ATM-Chk2 activation, leading tocell cycle arrest, and induces apoptosis in human prostate carcinoma DU145cells. Mol. Cancer Ther. 5, 3294–3302.
Albishi, T., John, J.A., Al-Khalifa, A.S., Shahidi, F., 2013. Antioxidant, anti-inflammatory and DNA scission inhibitory activities of phenolic compoundsin selected onion and potato varieties. J. Funct. Foods 5, 930–939.
Aparna, V., Charu, S., 2010. Evaluation of collagen in different grades of oralsquamous cell carcinoma by using the Picrosirius red stain – a histochemicalstudy. J. Clin. Diag. Res. 4, 3444–3449.
Atkinson, B.L., Blackman, A.J., Faber, H., 2004. The degradation of the naturalpyrethrins in crop storage. J. Agric. Food Chem. 52, 280–287.
Balch, C.M., Gershenwald, J.E., Soong, S.J., Thompson, J.F., Atkins, M.B., Byrd, D.R.,Buzaid, A.C., Cochran, A.J., Coit, D.G., Ding, S., Eggermont, A.M., Flaherty, K.T.,Gimotty, P.A., Kirkwood, J.M., McMasters, K.M., Mihm Jr., M.C., Morton, D.L.,Ross, M.I., Sober, A.J., Sondak, V.K., 2009. Final version of 2009 AJCC melanomastaging and classification. J. Clin. Oncol. 27, 6199–6206.
Beauchamp, C., Fridovich, I., 1971. Superoxide dismutase: improved assays and anassay applicable to acrylamide gels. Anal. Biochem. 44, 276–287.
Chakraborty, A., Ferk, F., Simic, T., Brantner, A., Dusinska, M., Kundi, M., Hoelzl, C.,Nersesyan, A., Knasmuller, S., 2009. DNA-protective effects of sumach (Rhuscoriaria L.), a common spice. Results of human and animal studies. Mutat. Res.661, 10–17.
Chen, Y.J., Chang, L.S., 2012. Gallic acid downregulates matrix metalloproteinase-2(MMP-2) and MMP-9 in human leukemia cells with expressed Bcr/Abl. Mol.Nutr. Food Res. 56, 1398–1412.
Chien, M.H., Ying, T.H., Hsieh, Y.S., Chang, Y.C., Yeh, C.M., Ko, J.L., Lee, W.S., Chang,J.H., Yang, S.F., 2012. Dioscorea nipponica Makino inhibits migration andinvasion of human oral cancer HSC-3 cells by transcriptional inhibition ofmatrix metalloproteinase-2 through modulation of CREB and AP-1 activity.Food Chem. Toxicol. 50, 558–566.
Chiodoni, C., Colombo, M.P., Sangaletti, S., 2010. Matricellular proteins: fromhomeostasis to inflammation, cancer, and metastasis. Cancer Metast. Rev. 29,295–307.
Dasgupta, T., Rao, A.R., Yadava, P.K., 2004. Chemomodulatory efficacy of basil leaf(Ocimum basilicum) on drug metabolizing and antioxidant enzymes, and oncarcinogen-induced skin and forestomach papillomagenesis. Phytomedicine 11,139–151.
Evan, G.I., Vousden, K.H., 2001. Proliferation, cell cycle and apoptosis in cancer.Nature 411, 342–348.
Farris, S.D., Hu, J.H., Krishnan, R., Emery, I., Chu, T., Du, L., Kremen, M., Dichek, H.L.,Gold, E., Ramsey, S.A., Dichek, D.A., 2011. Mechanisms of urokinaseplasminogen activator (uPA)-mediated atherosclerosis: role of the uPAreceptor and S100A8/A9 proteins. J. Biol. Chem. 286, 22665–22677.
Federico, A., Morgillo, F., Tuccillo, C., Ciardiello, F., Loguercio, C., 2007. Chronicinflammation and oxidative stress in human carcinogenesis. Int. J. Cancer 121,2381–2386.
Giftson, J.S., Jayanthi, S., Nalini, N., 2010. Chemopreventive efficacy of gallic acid, anantioxidant and anticarcinogenic polyphenol, against 1,2-dimethyl hydrazineinduced rat colon carcinogenesis. Invest. New Drugs 28, 251–259.
Gopi, V., Parthasarathi, A., Umadevi, S., Vellaichamy, E., 2013. Angiotensin-II down-regulates cardiac natriuretic peptide receptor-A mediated ant-hypertrophicsignalling in experimental rat hearts. Indian J. Exp. Biol. 51, 48–55.
Habig, W.H., Pabst, M.J., Jakoby, W.B., 1974. Glutathione-S-transferases. The firstenzymatic step in mercapturic acid formation. J. Biol. Chem. 249, 7130–7139.
Hanahan, D., Weinberg, R.A., 2011. Hallmarks of cancer: the next generation. Cell144, 646–674.
Hayat, M.J., Howlader, N., Reichman, M.E., Edwards, B.K., 2007. Cancer statistics,trends, and multiple primary cancer analyses from the Surveillance,Epidemiology, and End Results (SEER) program. Oncologist 12, 20–37.
Heber, D., 2008. Multitargeted therapy of cancer by ellagitannins. Cancer Lett. 269,262–268.
Ho, H.H., Chang, C.S., Ho, W.C., Liao, S.Y., Wu, C.H., Wang, C.J., 2010. Anti-metastasiseffects of gallic acid on gastric cancer cells involves inhibition of NF-kappaBactivity and downregulation of PI3K/AKT/small GTPase signals. Food Chem.Toxicol. 48, 2508–2516.
Ignea, C., Dorobant�u, C.M., Mintoff, C.P., Branza-Nichita, N., Ladomery, M.R., Kefalas,P., Chedea, V.S., 2013. Modulation of the antioxidant/pro-oxidant balance,cytotoxicity and antiviral actions of grape seed extracts. Food Chem. 141, 3967–3976.
Isuzugawa, K., Inoue, M., Ogihara, Y., 2001. Catalase contents in cells determinesensitivity to the apoptosis inducer gallic acid. Biol. Pharm. Bull. 24, 1022–1026.
Jagan, S., Ramakrishnan, G., Anandakumar, P., Kamaraj, S., Devaki, T., 2008.Antiproliferative potential of gallic acid against diethylnitrosamine-inducedrat hepatocellular carcinoma. Mol. Cell. Biochem. 319, 51–59.
Jeon, J.S., Zervantonakis, I.K., Chung, S., Kamm, R.D., Charest, J.L., 2013. In-vitromodel of tumor cell extravasation. PLoS One 8 (2), e56910.
Kang, M.S., Oh, J.S., Kang, I.C., Hong, S.J., Choi, C.H., 2008. Inhibitory effect of methylgallate and gallic acid on oral bacteria. J. Microbiol. 46, 744–750.
Kaur, M., Velmurugan, K.M., Rajamanickam, B., Agarwal, R.S., Agarwal, C., 2009.Gallic acid, an active constituent of grape seed extract, exhibits anti-proliferative, pro-apoptotic and anti-tumorigenic effects against prostatecarcinoma xenograft growth in nude mice. Pharm. Res. 26, 2133–2140.
Kaur, R., Arora, S., 2013. Interactions of betulinic acid with xenobiotic metabolizingand antioxidative enzymes in DMBA-treated Sprague Dawley female rats. FreeRadic. Biol. Med. 65, 131–142.
Khan, S.G., Katiyar, S.K., Agarwal, R., Mukhtar, H., 1992. Enhancement of antioxidantand phase II enzymes by oral feeding of green tea polyphenols in drinking waterto SKH-1 hairless mice. Possible role in cancer chemoprevention. Cancer Res. 52,4050–4052.
Kim, S.H., Jun, C.D., Suk, K., 2006. Gallic acid inhibits histamine release and pro-inflammatory cytokine production in mast cells. Toxicol. Sci. 91, 123–131.
King, J., 1965. Practical Clinical Enzymology. Von Nostrand D, Company Ltd.,London, p. 106.
Kligman, L.H., Gebre, M., Alper, R., Kefalides, N.A., 1989. Collagen metabolism inultraviolet irradiated hairless mouse skin and its correlation to histochemicalobservations. J. Invest. Dermatol. 93, 210–214.
Lee, D.S., Je, J.Y., 2013. Gallic acid-grafted-chitosan inhibits food borne pathogens bya membrane damage mechanism. J. Agric. Food Chem. 61, 6574–6579.
Lee, S.H., Kim, J.K., Kim, D.W., Hwang, H.S., Eum, W.S., Park, J., Han, K.H., Oh, J.S.,Choi, S.Y., 2013. Antitumor activity of methyl gallate by inhibition of focaladhesion formation and Akt phosphorylation in glioma cells. Biochim. Biophys.Acta 1830, 4017–4029.
Liao, C.L., Lai, K.C., Huang, A.C., Yang, J.S., Lin, J.J., Wu, S.H., Gibson Wood, W., Lin, J.G.,Chung, J.G., 2012. Gallic acid inhibits migration and invasion in humanosteosarcoma U-2 OS cells through suppressing the matrix metalloproteinase-2/-9, protein kinase B (PKB) and PKC signaling pathways. Food Chem. Toxicol.50, 1734–1740.
Li, X., Fang, P., Mai, J., Choi, E.T., Wang, H., Yang, X.F., 2013. Targeting mitochondrialreactive oxygen species as novel therapy for inflammatory diseases and cancers.J. Hematol. Oncol. 6, 19.
Liu, J., Lin, R.I., Milner, J.A., 1992. Inhibition of 7,12-dimethylbenz[a]anthracene-induced mammary tumors and DNA adducts by garlic powder. Carcinogenesis13, 1847–1851.
Liu, Z., Li, D., Yu, L., Niu, F., 2012. Gallic acid as a cancer-selective agent inducesapoptosis in pancreatic cancer cells. Chemotherapy 58, 185–194.
Lo, C., Lai, T.Y., Yang, J.S., Yang, J.H., Ma, Y.S., Weng, S.W., Lin, H.Y., Chen, H.Y., Lin,J.G., Chung, J.G., 2011. Gallic acid inhibits the migration and invasion of A375.S2human melanoma cells through the inhibition of matrix metalloproteinase-2and Ras. Melanoma Res. 21, 267–273.
Locatelli, C., Filippin-Monteiro, F.B., Creczynski-Pasa, T.B., 2013. Alkyl esters of gallicacid as anticancer agents: a review. Eur. J. Med. Chem. 60, 233–239.
Lu, Y., Jiang, F., Jiang, H., Wu, K., Zheng, X., Cai, Y., Katakowski, M., Chopp, M., TonyTo, S.S., 2010. Gallic acid suppresses cell viability, proliferation, invasion andangiogenesis in human glioma cells. Eur. J. Pharmacol. 641, 102–107.
Markert, C.L., 1963. Lactate dehydrogenase isozymes: dissociation andrecombination of subunits. Science 140, 1329–1330.
Marklund, S., Marklund, G., 1974. Involvement of superoxide anion radical inautooxidation of pyrogallol and a convenient assay for superoxide dismutase.Eur. J. Biochem. 47, 469–474.
Moron, M.S., Depierre, J.W., Mannervik, B., 1979. Levels of glutathione, glutathionereductase and glutathione S-transferase activities in rat lung and liver. Biochim.Biophys. Acta 582, 67.
Nakamuta, M., Higashi, N., Kohjima, M., Fukushima, M., Ohta, S., Kotoh, K.,Kobayashi, N., Enjoji, M., 2005. Epigallocatechin-3-gallate, a polyphenolcomponent of green tea, suppresses both collagen production and collagenaseactivity in hepatic stellate cells. Int. J. Mol. Med. 16, 677–681.
Narendhirakannan, R.T., Angeline Christie Hannah, M., 2013. Oxidative stress andskin cancer: an overview. Ind. J. Clin. Biochem. 28, 110–115.
Ng, T.B., He, J.S., Niu, S.M., Zhao, L., Pi, Z.F., Shao, W., Liu, F., 2004. A gallic acidderivative and polysaccharides with antioxidative activity from rose (Rosarugosa) flowers. J. Pharm. Pharmacol. 56, 537–545.
Okhawa, H., Onishi, N., Yagi, K., 1979. Assay for lipid peroxides in animal tissues bythiobarbituric acid reaction. Anal. Biochem. 95, 351.
Olson, P.R., Wattenberg, L.W., 1969. Inhibition by 5-fluorouracil of the early stagesof chemical carcinogenesis in mouse skin (34,054). Proc. Soc. Exp. Biol. Med.131, 1135–1137.
Ortega, E., Sadaba, M.C., Ortiz, A.I., Cespon, C., Rocamora, A., Escolano, J.M., Roy,G., Villar, L.M., Gonzalez-Porque, P., 2003. Tumoricidal activity of laurylgallate towards chemically induced skin tumours in mice. Br. J. Cancer 88,940–943.
Perchellet, J.P., Gali, H.U., Perchellet, E.M., Klish, D.S., Armbrust, A.D., 1992.Antitumor-promoting activities of tannic acid, ellagic acid, and several gallicacid derivatives in mouse skin. Basic Life Sci. 59, 783–801.
Pires, I., Gomes, J., Prada, J., Pereira, D., Queiroga, F.L., 2013. MMP-2 and MMP-9expression in canine cutaneous melanocytic tumours: evidence of arelationship with tumoural malignancy. Melanoma –From Early Detection toTreatment/Book 2. Intech, pp. 1–29 (provisional chapter).

V. Subramanian et al. / Food and Chemical Toxicology 66 (2014) 44–55 55
Provenzano, P.P., Inman, D.R., Eliceiri, K.W., Knittel, J.G., Yan, L., Rueden, C.T., White,J.G., Keely, P.J., 2008. Collagen density promotes mammary tumor initiation andprogression. BMC Med. 6, 11.
Rasmussen, H.S., McCann, P.P., 1997. Matrix metalloproteinase inhibition as a novelanticancer strategy: a review with special focus on batimastat and marimastat.Pharmacol. Ther. 75, 69–75.
Rotruck, J.T., Pope, A.L., Ganther, H.E., Swanson, A.B., Hafeman, D.G., Hoekstra, W.G.,1973. Selenuim: biochemical role as a component of glutathione peroxidase.Science 179, 588–590.
Shabaruddin, F.H., Chen, L.C., Elliott, R.A., Payne, K., 2013. A systematic review ofutility values for chemotherapy-related adverse events. Pharmacoeconomics31, 277–288.
Simna, S.P., Umadevi, S., Gopi, V., Elangovan, V., 2012. Toxicological assessment ofherbomineral siddha formulation: Neraadimuthuvallathy Mezhugu in Swissalbino mice. JPR 5, 1684–1689.
Sinha, K.A., 1972. Colorimetric assay of catalase. Anal. Biochem. 47, 389–394.Suen, J.J., Gibson Wood, W., Chung, J.G., 2009. Gallic acid induces apoptosis via
caspase-3 and mitochondrion-dependent pathways in vitro and suppresseslung xenograft tumor growth in vivo. J. Agric. Food Chem. 57, 7596–7604.
Thornthwaite, J.T., Shah, H.R., Shah, P., Peeples, W.C., Respess, H., 2013. Theformulation for cancer prevention & therapy. Adv. Biol Chem. 3, 356–387.
Van Der Helm, H.J., Zondag, H.A., Hertog, H.A., Van Der Kool, M.N., 1962. Lactatedehydrogenase isoenzymes in myocardial infarction. Clin. Chim. Acta 7, 540–554.
Vellaichamy, E., Khurana, M.L., Fink, J., Pandey, K.N., 2005. Involvement of the NF-kappa B/matrix metalloproteinase pathway in cardiac fibrosis of mice lackingguanylyl cyclase/natriuretic peptide receptor A. J. Biol. Chem. (280), 19230–19242.
Wilgus, T.A., Breza, T.S., Tober, K.L., Oberyszyn, T.M., 2004. Treatment with 5-fluorouracil and celecoxib displays synergistic regression of ultraviolet light B-induced skin tumors. J. Invest. Dermatol. 122, 1488–1494.
Woodbury, W., Speacer, A.K., Stahman, M.A., 1971. An improved procedure usingferricyanide for detecting catalase isozymes. Anal. Biochem. 44, 301–305.
Yogendra Kumar, M.S., Tirpude, R.J., Maheshwari, D.T., Bansal, A., Misra, K., 2013.Antioxidant and antimicrobial properties of phenolic rich fraction ofSeabuckthorn (Hippophae rhamnoides L.) leaves in vitro. Food Chem. 141,3443–3450.
Related Documents











