PHYSIOLOGIA PLANTARUM 92: 645-653. 1994 Copyri^hr © Physioiogia Phnrarum 1994 printed in Denmark - all ri^hlK reaervtd Tissue-specific expression of multiple forms of cyprosin (aspartic proteinase) in flowers of Cynara carduncubis Maria C, Cordeiro, M. Salome Pais and Peter E, Brodelius Cordeiro, M.C, Pais, M.S. and Brodelius, P.E. 1994. Tissue-specific expression of multiple forms of cyprosin {aspartic proteinase) in flowers of Cynara carduncuhis. - Physiol. Plant. 92: 645-65.'5. Tliree heterodimeric aspartic proteinases (cyprosin I, 2 and 3) wilh milk-clotting activity have previously been purified from flowers of Cynara cardunculus and partly characterized (U. Heimgartner et al. !990, Phytochemistiy 29: 1405-1410). These proteinases have now been further studied, isoeiectric focusing has revealed a micro- heterogeneity of the apparently pure cyprosins. Three isozymes with close isoeiectric points around 4.0 have been found. Reversed-phase high performance liquid chromato- graphy of e!ectrophoretically purified large stibunits of cyprosin has also shown a microheterogeneity. Peptide mapping of cyprosins 2 and 3 by trypsin or BrCN cleav- age indicate that they are derived from common procyprosin(s). Studies on the organ- specific accumulation of the enzyme were carried out using flower buds and flowers at different stages of development and styles and coro!las from open flowers, leaves and seeds. !mmunostained western hlots revealed the presence of cyprosin in very young flowers in low amounts. The amount of enzyme increased towards later stages of development and it was mostly present in the violet parts of styles and coro!!as. The enzyme couid not be detected in leaves or seeds. Proteolytic and milk-clotting activities correlate well with these findings. The enzyme was localized by immunolabelling in the epidermal cell layer of styles. Mature flowers collected at 8 different locations in Portugal showed some variation in proteolytic activity while the milk-clotting activity was essentially the same for all extracts. Key words - Aspartic proteinase, Cynara cardunculus, cyprosin, immunolocaiization, milk clotting, tissuc-specific accumulation. M, C. Cordeiro (present address: Depto Biologia. Univ. of Evora. Apartado 94. P-7001 Evora Codex. Portugal) and M. S. Pais, Depto Biologia Vegetal, ECL, Bloco C2, Campo Grande, P-17(X) Lisboa, Portugal: P,E, Brodelius (corresponding author), Dept of Plant Biochemistry. Univ. of Lund. P.O. Box 117, S-22I00 Lund, Sweden, , . . .. recent revision of the genus (Wik!und 1992), Only C Introduction ji a .t • J ,, %. cardunculus ^pp, flavescens cv. cardoon is referred to be A plant coagulant obtained from flowers of Cynara car- used in cheese making. However, C, humilis and C. dunculus (cardoon) is widely used in Portugal to produce scolymus (now C. cardunculus spp. flavescens cv. ar- ewe's milk cheese. The Serpa and Serra cheeses are well tichoke) have been found to possess clotting activity. In known and appreciated for their characteristic taste. They Spain, flowers of C, humilis are used in the manufacture are typical products from the southem and northem re- of various cheeses. Examples of cheeses are Serena, Torta gions of Portugal, respectively. del Casar, Pedroches and Grazalema. Besides the genus The cardoon plant belongs to the Asteraceae and grows Cynara other Asteraceae species have been used to clot wild in Portugal and the Madeira and Canary Islands. It is milk; however, generally showing a much lower clotting also found in the southem and western parts of the Medi- activity. That is the case for Centaurea calcitrapa terranean region and in northem Africa. In total eight (Domingos et al. 1992), Sv/ifcum mananMm (Fevereiroet different species are described for the genus Cynara in a al, 1986) and Onopordum turcicum (Tamer 1993), Received 15 December, 1993; revised 20 June, 1994; in final form 24 August, 1994 Ptiysiol. Ptanl. 92. \mi 645

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PHYSIOLOGIA PLANTARUM 92: 645-653. 1994 Copyri^hr © Physioiogia Phnrarum 1994printed in Denmark - all ri^hlK reaervtd
Tissue-specific expression of multiple forms of cyprosin (asparticproteinase) in flowers of Cynara carduncubis
Maria C, Cordeiro, M. Salome Pais and Peter E, Brodelius
Cordeiro, M.C, Pais, M.S. and Brodelius, P.E. 1994. Tissue-specific expression ofmultiple forms of cyprosin {aspartic proteinase) in flowers of Cynara carduncuhis. -Physiol. Plant. 92: 645-65.'5.
Tliree heterodimeric aspartic proteinases (cyprosin I, 2 and 3) wilh milk-clottingactivity have previously been purified from flowers of Cynara cardunculus and partlycharacterized (U. Heimgartner et al. !990, Phytochemistiy 29: 1405-1410). Theseproteinases have now been further studied, isoeiectric focusing has revealed a micro-heterogeneity of the apparently pure cyprosins. Three isozymes with close isoeiectricpoints around 4.0 have been found. Reversed-phase high performance liquid chromato-graphy of e!ectrophoretically purified large stibunits of cyprosin has also shown amicroheterogeneity. Peptide mapping of cyprosins 2 and 3 by trypsin or BrCN cleav-age indicate that they are derived from common procyprosin(s). Studies on the organ-specific accumulation of the enzyme were carried out using flower buds and flowers atdifferent stages of development and styles and coro!las from open flowers, leaves andseeds. !mmunostained western hlots revealed the presence of cyprosin in very youngflowers in low amounts. The amount of enzyme increased towards later stages ofdevelopment and it was mostly present in the violet parts of styles and coro!!as. Theenzyme couid not be detected in leaves or seeds. Proteolytic and milk-clotting activitiescorrelate well with these findings. The enzyme was localized by immunolabelling inthe epidermal cell layer of styles. Mature flowers collected at 8 different locations inPortugal showed some variation in proteolytic activity while the milk-clotting activitywas essentially the same for all extracts.
Key words - Aspartic proteinase, Cynara cardunculus, cyprosin, immunolocaiization,milk clotting, tissuc-specific accumulation.
M, C. Cordeiro (present address: Depto Biologia. Univ. of Evora. Apartado 94. P-7001Evora Codex. Portugal) and M. S. Pais, Depto Biologia Vegetal, ECL, Bloco C2,Campo Grande, P-17(X) Lisboa, Portugal: P,E, Brodelius (corresponding author),Dept of Plant Biochemistry. Univ. of Lund. P.O. Box 117, S-22I00 Lund, Sweden,
, . . . . recent revision of the genus (Wik!und 1992), Only CIntroduction j i a .t • J ,, %.
cardunculus ^pp, flavescens cv. cardoon is referred to beA plant coagulant obtained from flowers of Cynara car- used in cheese making. However, C, humilis and C.dunculus (cardoon) is widely used in Portugal to produce scolymus (now C. cardunculus spp. flavescens cv. ar-ewe's milk cheese. The Serpa and Serra cheeses are well tichoke) have been found to possess clotting activity. Inknown and appreciated for their characteristic taste. They Spain, flowers of C, humilis are used in the manufactureare typical products from the southem and northem re- of various cheeses. Examples of cheeses are Serena, Tortagions of Portugal, respectively. del Casar, Pedroches and Grazalema. Besides the genus
The cardoon plant belongs to the Asteraceae and grows Cynara other Asteraceae species have been used to clotwild in Portugal and the Madeira and Canary Islands. It is milk; however, generally showing a much lower clottingalso found in the southem and western parts of the Medi- activity. That is the case for Centaurea calcitrapaterranean region and in northem Africa. In total eight (Domingos et al. 1992), Sv/ifcum mananMm (Fevereiroetdifferent species are described for the genus Cynara in a al, 1986) and Onopordum turcicum (Tamer 1993),
Received 15 December, 1993; revised 20 June, 1994; in final form 24 August, 1994
Ptiysiol. Ptanl. 92. \mi 645
Fig. 1. .4. Cynara cardunculus capitulum; B, flower hud"- atvarious stages of deveiopment and C, mature flowers.
The clotting activity present in the flowers of C. car-duncidus spp. flavescens cv. cardoon (Fig. 1) is due lothree aspartic proteinases, cypro.sins 1, 2 and 3 (previ-ously called cynarases), which have been purified andpartly characterized (Heimgartner et al. 1990). They areheterodimeric enzymes containing high mannose typegiycosyiations vi'ith native molecular masses of 49 JcDa.They consi.st of one large subunit (32.5-35.5 kDa) andone small subunit (13.5-16.5 kDa) and as other asparticproteinases they preferentially cleave peptide bonds be-tween hydrophobic amino acid residues. They are allstongly inhibited by pepstatin A. Their proteolytic andmilk clotting activities were compared to those of chymo-sin (Cordeiro et al. 1992), which is the most widely usedenzyme in the cheese manufacturing industry, Cyprosin 3showed the highest specific activity among the cyprosins.The clotting activity of cypro.sin 3 is comparable to thatof chymosin, whiie slight differences in tlie specificitytowards milk caseins are observed for these two enzymes(Cordeiro et al. 1992).
A cDNA clone encoding a cyprosin has been isolatedand characterized (Cordeiro et al. 1994). The nucleotidesequence contains a 1419 bp open reading frame coding
for 473 amino acids including a putative full length ma-ture protein (439 amino acids) and a partial prosequence(34 amino acids). The deduced amino acid sequence ofcyprosin shows a relatively high homology to other plantaspartic proteinases: barley (78%) and rice (63%). (Com-pared to other aspartic proteinases the platit enzymescontain an insert of around 100 amino acids (plant specif-ic insert) io the C-terminal part of the protein molecule.Omitting this insert, the homology to mammalian asparticproteinases is relatively high; human cathepsin D (55%)and bovine chymosin b (43%).
Farmers only use mature flowers of C. cardunculus toproduce cheese. Recent studies have indicated that thecj'prosins are organ-specific enzymes and that they aremainly present in the violet part of the flowers (Heim-gartner et al, 1990, Cordeiro et al. 1992). In thi,s study thepresence of the cyprosins in flower buds and flowers atvariotis stages of development, as well as in other tissuesof the plant has been investigated. Also their tisstie-specific accumulation has been studied. The cyprosioshave also been further biochemically characterized.
Abbreviations - lEF, i.soelectric focusing; PBS, phosphate-huf-fered saline; TFA, trifluoroacetic acid.
Materials and methods
Plant materia!
Flower buds and flowers of Cynara carduncubis L. indifferent stages of development were collected in thefield in Portugal from mid-April to mid-July. They wereisolated from the pappus and divided into different parts,i,e, style.-;, corollas and seeds and .subsequently frozen inliquid nitrogen and stored at —20°C. Ver)' young stageswere obtained by cutting the capitula and isolating singlebuds. Leaves of differenl size.s (7 and 30 cm long) werecollected in the field, frozen in liquid nitrogen and storedat -20°C.
Mature flowers were also coliected at 8 differenl loca-tions in Portugal. These flowers were allowed to air dryin the same way as farmers dr\' collected flowers.
Crude protein extract
Crude extracts of the different tissues were obtainedby homogenizing frozen fresh tissue (100 mg) or driedflowers (40 mg) in a mortar under liquid nitrogen. Thepowder obtained was extracted with 50 mM Tris-HCIbuffer, pH 8,3 (2 ml). After centrifugation at 18 000 g for25 min at 4°C, the supematant was collected and ana-lyzed.
Purification of cyprosins
Cyprosins were purified to apparent hornogeneity as de-scribed by Heimgartner et al, (1990),
646 Hiysiol. Ptant. 92, 1994

Protein concentration
Protein concentrations were determined according toBradford (1976).
Proteoiytic activity
Proteolytic activity was determined at pH 5.1 using ca-sein labelled with fluorescein isothiocyanate according tothe fluorometric procedure previously described (Heim-gartner et al. 1990).
Clotting activity
Clotting activity was routinely determined at 37°C usitigcow's low fat milk preserved with 1.5 mAf NaN3. Crudeextract (100 jil) was added to milk (5 ml) and clottingtime was determined by direct obsen'ation of flocculationatid gel formation. Controls were prepared by addingextraction buffer to preser\'ed milk. Clotting activity wasalso measured with a Formagraph as previously described(Cordeiro et al. 1992). These two tnethods to measureclotting activity are not directly comparable.
Eiectrophoresis
Sodium dodecyl sulfale-polyacrylamide gel electrophore-sis (SDS-PAGE) was performed on 13%-gels accordingto Laemmli (1970) and gels were stained with silvernitrate according to Blum et al. (1987).
Two-dimensiona! ge! eiectrophoresis
Isoeiectric focusing (IEF) of purified cyprosins was per-formed on 10% polyacr)'lamide tube gels in 8 Af ureacontaining 0.5% Ampholyte pH 4-6.5, 0.59c AmpholytepH 3..5-I0 and 1.75% tergitol NP-40. pH gradientsformed during lEF were determined by mea.suring thesolution pH of 1-cm tube gel segtnent.s eiuted in 0.1 MKCl (1.0 ml) overnight at 4°C. Standard proteins (car-bonic anhydrase, pi 6,1, bovine serum albumin, pi 4.9and ovalbumio, pi 4.7) were also run to verify the pHgradients.
Tube gel.s were equilibrated in SDS-PAGE buffer for15 min and placed onto 15% acrylamide slab gels withoutstacking gel. Electrophoresis was earned out according toLaemmli (1970).
Western blot
Western blotting was performed on a semi-dry eleciro-blotter (Bio-Rad, Hercules, CA, USA) according to Bur-nette (1981) with some modificatiotis according toHawkes et al, (1982) and Johtison et al. (i984). Blotswere incubated with polyclonal antibodies (totai serumdiluted 1 ;200) raised in rabbit against the large .subunit ofcyprosin 3 (Heimgartner et ai. 1990) and antigen-anti-body complexes were detected using goat anti-rabbit im-
munogiobulin conjugated with horse radish peroxidase(Bio-Rad, Hercules, CA, USA),
Preparative electroplioresis
Preparative SDS-PAGE was carried out according toLaemmli (1970) in 13% gels (340 x 130x 1,5 mm). Thegels were nm for 1 540 Vh. Visualization of bands wasachieved with 1 M KCl in 10% acetic acid. The bandscorresponding to the large and small subunits of cyprosin2 and 3 were cut out and kept separately in 10% aceticacid.
Peptide mappitig, protein microseqnencing and reversed-phase higli performance liquid chrotnatography
BrCN cleavageSmall gel slices (0.28 g) were cut from each isolated bandfrom preparative electrophoresis and were equilibrated in70% formic acid (2x 10 min). The gel slices were thentreated with BrCN (50 g 1"') in 70% formic acid for 30min at room temperature. The reaction was stopped byperforming several washes with 10% acetic acid. Sub-sequently, the treated gel slices were washed and equili-brated with 0,1 M Tris-HCI buffer, pH 8,0, containing0.4% SDS (4 x20 min) and fmally with the same buffercontaining 2% /?-mercaptoethaooI (1x20 min). Theequilibrated gel pieces were placed in the sample wells ofa SDS-P.AGE slab gel. SDS-PAGE was performed oti15%-gels according to Laemmli (1970). Gels werestained with silver nitrate as above.
Trypsin digestionThe isolated ge! slices from preparative electrophoresiscontaining the large subunit of cyprosin 2 and 3 were cutinto small pieces (2x2 mm) and the protein was electro-eluted for 5 h at 40 m,4 using 25 mM Tris buffer con-taining 192 mM glycine and 0.1% SDS. The collectedprotein was desalted by gel filtration and the proteinsample was divided into portions containing 100 (ig pro-tein in Eppendorf tubes. Subsequently the water wasremoved on a Speedvac and the protein digested withtrypsin (EC 3.4.21.4) according to Stone et al. (1989),The dry protein was redissolved in 50 (tl 8 M ureacontaining 0.4 M NH4HCO3 and 5 ill 45 mM dithiothrei-tol (DTT) was added. The mixture was incubated first at50°C for 15 min, cooled to room temperature and in-cubaied wilh 5 ^I 100 mM iodoacetamide for 15 minbefore adding 140 ill water. Hydrolysis Vi'as carried oulwith 2, 4 or 10 }.tg trj'psin (Fluka, Buchs, Switzerland) at37°C for 24 h and analyzed by SDS-PAGE on 15%-gelsas described above,
MicrosequeticingAfler SDS-PAGE, BrCN and trj'psin treated proteinswere electroblotted otito Problot membranes (AppliedBiosystems, Fo,ster City, CA). The membranes werestained with Coomassie blue R-250 or with copper phtha-
Physiol. Ptanl. 92, 1994 647

locyanine 3,4',4",4"'-tetrasulfonic acid tetrasodium salt(Bickar and Reid 1992). Appropriate bands were excisedand subjected to N-terminal sequencing on an AppliedBiosystems model 477A gas phase sequencer,
Reversed-phase high peiformance liquidchromatographyReversed-phase HPLC of electrophoretically purifiedsubunits of cyprosin 2 and 3 was carried out on a Kroma-sil 100 5|i C8 column (Hichrom, Reading, UK)(250x4,6 mm) using 0,1% trifluoroacetic acid (TFA) inwater (solvent A) and acetonitrile;0.1% TFA (80;20) (sol-vent B). The program used for elution was; 0-2 min 10%B, 2-32 min 10-100% B, 32-34 min 100% B, 34-36 min100-10% B, 36-39 min 10% B. The flow rate was 1.0 mlmin'' and the sample size was 100 j.tl. Detection wascarried out on a dual UV-monitor at 230 and 280 tim.
Preparatioti of flower tissue for immunocytochemica!!oca!ization of cyprosin
Fiowers at different stages of development were collectedin the field from early May to mid-June. The capitulawere cut into Iwo halves and single flower buds wereisolated from the pappus and immediately inserted infixative. All die steps until polymerization were per-formed at 4°C. Immunogold labellin.g was performed asdescribed by Bergman et al. (1985) but with some modi-fication. Each flower bud was divided into 3 parts de-pending on size, and transversal sections of about 1.5 mmwere made with a sharp razor blade at the beginning ofeach of these regions. These sections were cut in fixative.Fixation was carried out in 2% paraformaldehyde and2.5% glutaraldehyde in 50 mM sodium phosphate buffer,pH 7,2, for 1 h. Next, 3 washes in phosphate btifler wereperformed. Dehydration was carried out in an ascendingseries of ethanol in water; 30, 50, 70, 80, 90 and 100%.The cuts were incubated for 20 min in each solution andfor 60 min in J00% ethanol. Einbedding was performedgradually in LR-White resin (London Resin Co. Ltd.,London, UK) in ethanol; 50%, 30 min; 75%, 60 min;100%, overnight. Then the sections were incubated in100% resin for 2 h before they were encapsulated andoriented as well as possible in 100% resin, and poly-merized at 55°C for 24 h. Semi-thin sections (1—2 \im)were mounted on microscope slides for immunogold la-belling,
Immunogoid !al»e!!ing and siiver en!ianccmetJt for lightmicrosciipj
All steps were performed at room temperature. First, thesections were washed with 10 mM phosphate-bufferedsaline (PBS), pH 7.4, lor 5 min and then treated with 5%>(w/v) bovine serum albumin (BSA), I %• invertase and 5%foetal calf serum in PBS for 20 min. A wash with PBS for5 tnin followed and then a treatment with primary anti-body (a rabbit antLserum raised again.st the large subunit
of cyprosin 3; diluted 1 ;20) in PBS containing 1% BSAand 1% invertase for 60 min followed. Washes (3x15min) with PBS were carried out and then the sectionswere treated with goat anti-rabbit IgG coupled with 5 nmgold particles (diluted l;20) for 30 min (AuroProbe EMGAR G5; Amersham, Berks, UK). Washes with PBS(2x15 min) and water (2x15 min) were performed next.In controls the primary antibody was substituted by 1 %•BSA and/or rabbit preimmurte serum.
Silver enhancement was performed according to theinstructions of the manufacturer described in the lntenSEM Kit (Amersham, Berks, UK). The assay was moni-torted by light microscopy and micrographs were taken atconstant lime inter\'als. After enhancemeol for 16 min,the preparations were washed in water. After drying theywere stained with Azur II-methylene blue (Richardson etal, 1960), and mounted with adhesive (Merckogiass;Merck, Darmstadt, Germany).
Results and disctissron
Cyprositis
Protein extracts of mature flowers of C cardunctdus(Fig. 1) contain relatively high proteolylic atid milk clot-ting activities (Heimgartner et al. 1990, Cordeiro et al.1992), Three proteinases (cyprosins 1, 2 and 3) have beenpurifted from such extracts (Heimgarttier el al. 1990).The three cyprosins are heterodimeric enzymes of a.spar-tic acid type wilh a native molecular mass of ca 49 kDa.They are glycoproteins containing N-linked high mati-nose type glycans. Similar heterodimeric asparlic prolei-nases have been found in resting barley grains (Sarkkinenel al. 1992) while other plant aspartic proteinases aremonomeric (Doi et al. 1980, Polanovvski et al. 1985,Belozersky el al. 1989).
Determination of pi
Apparently pure cyprosin 2 and 3 (according lo SDS-FAGE) were subjected to Iwo-dimensional electrophore-sis to determine their isoeiectric points. The first dimen-sion gel was an IEF gel within the pH range of 3.5 lo 6.5.The second dimension gel was an SDS-PAGB. Both puri-fied enzymes .showed a niicrohelerogeneity wilh proteinspecies of differenl pi. For example, apparently purecyprosin 3 was fractionated into 3 isozymes with pivalues of 3.85, 4,00 and 4.15, respectively, as shown inFig. 2. Weslem blotting showed that the large subunit.s ofall three isozymes hybridized with antibodies raisedagainst the large subunit of cyprosin 3 (Fig. 2B).
A.spartic proteinases are often synthesized as preproen-zymes which are converted to zytrtogeos by the remova!of an N-termitial signal sequence. These zymogens aresubsequently converied to mature enzymes by die re-moval of the prosequence (Tang and Wong 1987). SinceSouthern blot analysis of a nuclear DNA restiction maphas revealed multiple cyprosin genes (at least three; Cor-
Phy.siol. Planl. 92. 1M4

3.5 4.0 -J.S 5.0 5.5I I I I t
kDa
3.5 4.0 4.5I l l
5.0 5.5 G.Ol l i
Fig. 2. T^vo-dimensional eleclrophoresis ol purifted cypro.sin 3.Isoeiectric focu.sing was performed in ihc fins! dimension andSDS-P.AGE in the second as described in Materials andmethods. A. silver-stained gel; B, immunostained western blotof con'esponding gei.
deiro el al. 1994) il is most likely tliat the three isozymesare fomied through the expression of a cyprosin geoefamily. This assumption is further supported by the factlhat we recently have isolated and sequenced a secondcyprosin cDNA (M. Pielrzak, M. S. Pais and P, E. Brode-lius. unpublished results) showing some substitutions inthe N-terminus of the putative tnature proteiti.
Reversed-phase !iigh performance iiquld chromatography
Analytical reversed-phase HPLC of the electrophoret-ically purified large subutiits of cyprosin 2 and 3 (.seecontrols in Fig. 3) also revealed microheterogeneity.These large subunils could aach be resolved into threepeaks (data not shown).
Peplide mapping
The larger subunils ot cypro.sios 2 and 3 were purified bypreparative eiectrophoresis (controls in Fig. 3) and Iheisolated peptides were subjected to cleavage by BrCN ortr\'psin. The treatments resulted in peptide maps as shownin Fig. 3. Some of the peplides generated by the cleavage(arrow.s B4, Tl and T2) appeared to be the same for both
cyprosins indicating some common structural features ofthe two enzymes and that they may be derived from acommon procyprosin. It is possible that the bands la-belled B1 and B5 are generated by cleavage of the pep-tide chains at the same methionine io the two cyprosins.The apparent size difference of the pair of peptidesformed is essentially the same as that of the two non-hydrolyzed large subunits. The occurrence of hetero-dimeric forms of the enzyme containing subunits of dif-ferent sizes could thus be due to proteolytic processing ofthe cyprosin molecule at different sites. We have previ-ously suggested that the cyprositis have peptide regionsin common (Heimgartner et al. 1990).
N-termiual sequencing
The population of cyprosins in flowers of C cardunculusis very complex. They vary in subunit size and in isoeiec-tric points. This complexity complicates detailed studiesof the protein. Attempts to carry otit N-terminal seqtien-cing of electrophoretically (SDS-PAGE) purified sub-units of cyprosins 2 and 3 (controls in Fig. 3) have beenunsuccessful. This appears to be due to a microheteroge-neity of the N-terniinal end since it is clear that theprotein is not N-terminaly blocked. We assume this mi-croheterogeneity to ari,se through the expression of thecyprosin getie family containing genes coding for verj"closely related proteins.
However, an intemal partial N-terminal sequence hasbeen obtained from a peptide obtained by BrCN-cleavageof cyprosio 2 (arrovv B2 in Fig. 3). This internal sequetice(Mel-Leu-Asn-Gln-Gly-Leu-Val-Gln-Glu) was used toidentify a cDNA clone coding for the cyprosin (Cordeiroet al. 1994). This sequence may be part of a highlyconserved region in the cyprosin isozyrnes. Attempts tosequence peptides obtained by trypsin treatment of cy-prosin 2 and 3 (arrow T l , Fig. 3) did not result in anyN-terminal sequence. This is most likely due to the mi-croheterogeneity of the N-temiinal end of this peptide.We assume this N-terminal end to be the same as that ofthe undigested subunit.
Fig. i. SDS-PAGE ofelectrophorelically purifiedlarge siibuiiits of cyprosin 2and 3. Controls are untreatedsiibuoil^. Peptides obtainedafler BrCN and trypsintreatments of the subunitsare labelled B and T,respectively. Ali gels were.stained with silver nitrate.
Standards Control BrCN Trypsin
20-
14.4-
BJ
T l Tl
.T2
Pliysiol. Ptml. 92, 1994 649
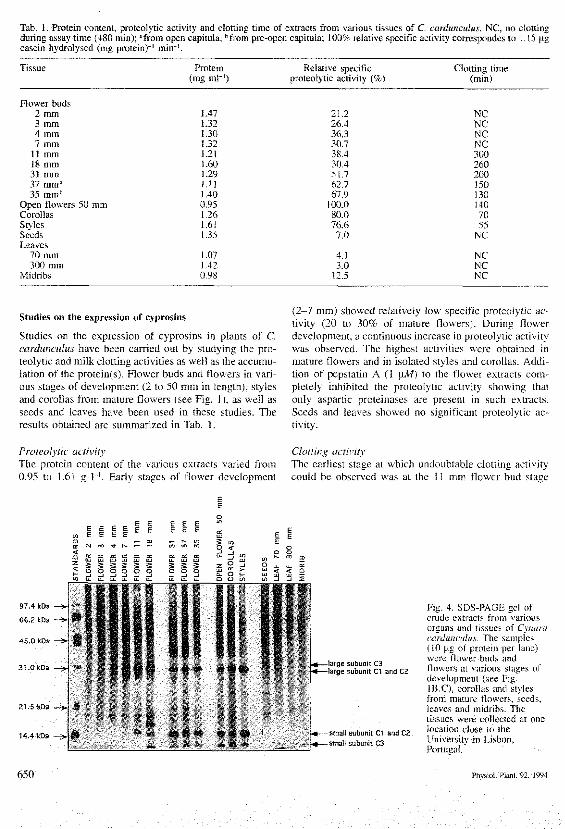
Tab. 1. Protein conteni, proteoiytic activity and clotting time of extracts from various tissues of C cardunculus, NC, no clottingduring assay time (480 min); "from open capitula; "from pre-open capitula; 100% relative specific activity correspondes to 1.15 ngcasein hydrolysed (mg protein)"' min"'.
Tissue
Flower buds2 mm3 mm4 mm7 mm
11 mm18 mm31 mm37 mm"35 mm"
Open flowers 50 mmCorollasStylesSeedsLeaves
70 mm300 mm
Midribs
Protein(mg ml"')
J.471.321.30L32L2]
(
(
.60
.29.11.40
).95.26.61.35
.07
.42).98
Relative specificproteolytic activity (%)
21.226.436.330.738.430.451.762.767.9
100.080.076.67.0
4.13.0
12.5
Clotting time(min)
NCNCNCNC3002602001501301407055
NC
NCNCNC
Sttidies on the expression of cyprosins
Studies on the expression of cyprosins in plants of C,cardunculus have been carried out by studying the pro-teolytic and milk clotting activities as well a.s the accumu-lation of the protein(s). Flower buds and flowers in vari-ous stages of development (2 to 50 mm in length), stylesand corollas from mature flowers (see Fig. 1), as well asseeds and ieaves have been used in these studies. Theresults obtained are summarized in Tab. 1.
(2—7 mm) showed relatively low specific proteolytic ac-tivity (20 to 30% of mature flowers). During flowerdevelopment, a continuous increase in proteolytic activitywas observed. The highest activities were obtained inmature flowers and in isolated styles and corollas. Addi-tion of pepstatin A (! jtiW) to ihe flower extracts corn-pletely inhibited the proteolytic activity showing thatonly a.spartic proteinases are preseni in such extracts.Seeds and leaves showed DO significant proteolytic ac-tivitv.
Proteolytic activityThe protein content of the various extracts varied from0.95 to 1.61 g 1~'. Early stages of flower development
Clotting activityThe earliest stage at which undoubtable clotting activitycouid be observed was at the 11 mm flower bod staee
31.0 kDa
21,5 t<Da
14.4 kDa
-targe subunit C3-targe subunrt Cl and CH
.H*— smalt subunit Cl and C2.- i^i—small subunit C3
Fig. 4. SDS-PAGE gel ofcrude extracts frotn %'ariousorgans and tissues of Cynaracardunculus. The samples(10 [lg of protein per lane)were flower huds andflowers at various stages ofdevelopment (see Fig.IB,C), corollas and stylesfrom mature flower.s, seeds,leaves and midribs. Thetissues were collected al onelocation ciose to IheUniversity in Lisbon,Portugal.
650 Physiol. Plant. 92, 1994
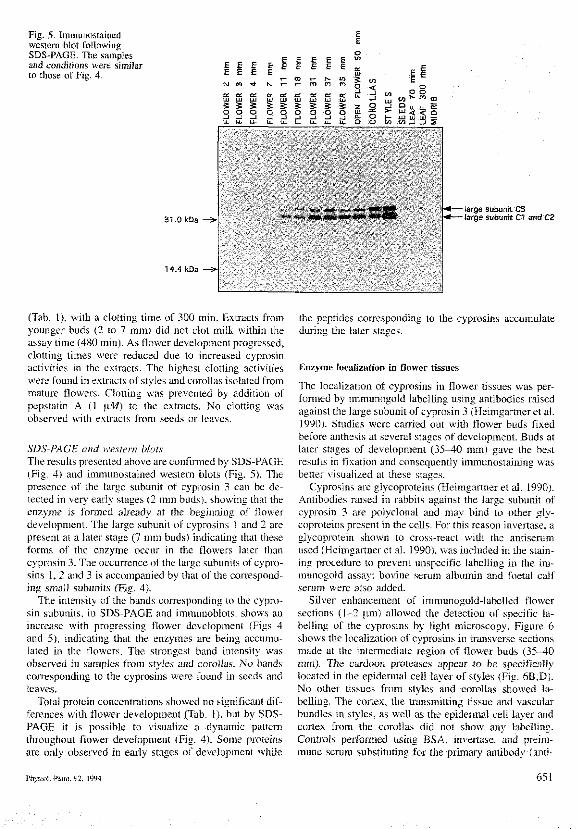
Fig. 5. Immunostainedwestern blot followingSDS-PAGE. Tbe samplesand conditions were simiSarto those of Fig. 4.
6 EE E
UJ Ul
s s3 S
E E
T to»- »- m
S 5 S 3o o o o
3 3
S I ou. D O
Q 3 O u. u. S>" < < nUJ UJ uJ —
31.0 kDa
14.4 kDa •
-targe subunit C3- large subunit Cl and C2
(Tab. 1), with a clotting time of 300 min. Extracts fromyounger buds (2 to 7 mm) did not clot milk within theassay time (480 min). As flovi er development progressed,clotting times were reduced due to increased cyprosinactivities in the extracts. The highest clotting activitieswere found in extracts of styles and corollas isolated frommature flowers. Clotting was prevented by additioti ofpepstatin A (1 \.M) to the extracts. No clotting wasobserved with extracts from seeds or leaves.
SDS-PAGE and wcs'tern blotsThe re.sults presented above are confirmed by SDS-PAGE(Fig. 4) and imtitunostained western blots (Fig. 5). Thepresence of ihe large subunit of cyprosin 3 can be de-lected in very early stages (2 mm buds), showing lhat theenzyme is formed already al the beginning of flowerdevelopment. The large subunit of cyprosins 1 and 2 arepresent at a later stage (7 mm buds) indicating that theseforms of the enzyme occur in the flowers later thancyprosin 3. The occurrence of the large subunits of cypro-sins 1, 2 and 3 is accompanied by that of the correspond-ing small subunits (Fig. 4).
The intensity of the bands corresponding to the cypro-sin subunits, io SDS-PAGE and immutioblots, show.s anincrease with progressing Oower development (Figs 4and 5), indicating that the enzymes are being accumu-lated in the flowers. The strongest band intensity wasobsen'ed in samples from styles and corolla.s. No band.scorresponding to the cyprosins were found in .seeds andleaves.
Total protein concentrations showed no significant dif-ferences witli flower development (Tab. 1), bul by SDS-PAGE it is possible to visualize a dynamic patternthroughoul fiower development (Fig. 4), Some proteinsare otily observed in early stages of development while
the peptides corresponding to the cyprosins accumulateduring the later stages.
Enzyme localizatioti in flower tissues
The localization of cyprosins in flower tissues was per-formed by immunogold labelling using antibodies raisedagainst tbe large subunii of cyprosin 3 (Heimgartner et al.1990). Studies were carried out with flower buds fixedbefore anthesis at several stages of development. Buds atlater stages of development (35-^0 mm) gave the bestresults ill fixation and consequently immunostaining wasbetter visualized at these stages.
Cyprosin.s are glycoproteins (Heimgartner et al. 1990).Antibodies raised in rabbits against the large subunit ofcyprosin 3 are polyclonal and may bind to other gly-coproteios present in the cells. For this reason invertase, aglycoprotein shown to cross-react with the antiserumused (Heimgartner et al. 1990), was included in the stain-iog procedure to prevent unspecific labelling in the im-munogold as,say: bovine serum albumin and foetal calfseriiin were also added.
Silver enhancemeni of immunogold-labelled flowersections (1-2 (.im) allowed the detection of specific la-belling of the cyprosins by light microscopy. Figure 6shows the localization of cyprosins in transverse sectionsmade al the intermediate region of flower buds ( 3 5 ^ 0mm). Tiie cardoon protea.ses appear to be .specificallylocated in the epidermal cell layer of styles (Fig. 6B,D).No other tissues from .styles atid corollas showed la-belling. The cortex, tbe transmitting tissue and vascularbundles in styles, as well as the epidennal cell layer andcortex from the corollas did nol show any labelling.Controls performed using BSA, invertase, and preim-mune serum substituting for the primary antibody (anti-
Physiol. Ptanl. 92. 1994 651
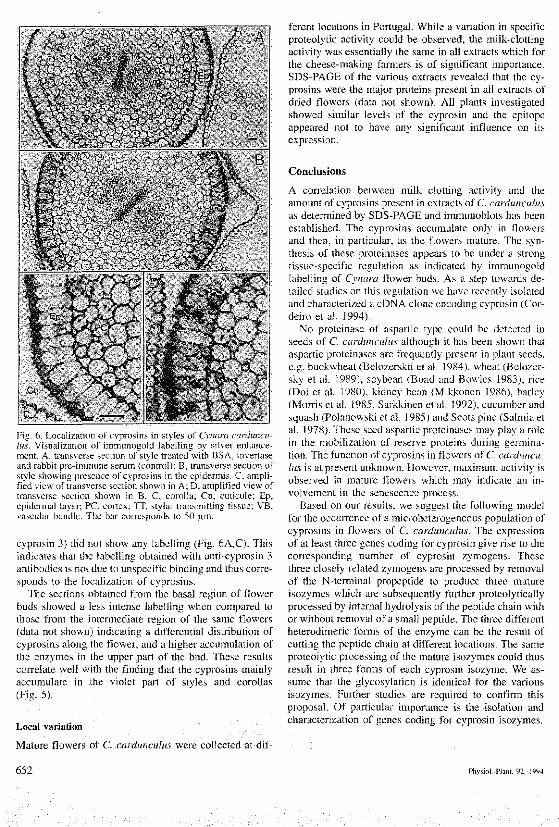
Fig. 6. Localization of cypnisins m styles of Cvnara iarduncu-lus. \'isualizalion of immunogold labelling b) siher enhance-ment. A. trans^'er.se section of .stvle treated with BSA. ]n\erta.seand rabbil pie-immune serum (controll: B, transverse section ofstyle shov^mg presence ol C}pr(}sins 3n the epidermis C, ampli-fied \ien of tran'.^erse seclion shown tn A. D. amplified view oftrans\erse section shown in B. C. corolla; Cu. cuttcule: Ep,epidermal layer; PC, cortex; TT, stjlar transmitting ti.'.suu; VB,va.scular bundle The bar corre.sponds to 50 um
cyprosin 3) did not show any labelling (Fig, 6A,C). Thisindicates that the labelling obtained with anti-cyprosin 3antibodies is not due to unspecific binding and thus corre-sponds to the localization of cyprosins.
The sections obtained from the basal region of flowerbuds showed a less intense labelling when compared tothose from the intermediate region of the same flowers(data not sbown) indicating a differentia! distribution ofcyprosins along the fiower, and a higher accumulation ofthe enzymes in the upper part of the bud. These resultscorrelate well with the fmding that the cyprosins mainlyaccumulate in the violet part of styles and corollas(Fig. 5).
Local variation
Mature flowers of C, cardunculus were collected at dif-
ferent locations in Portugal. While a variation in specificproteolytic activity could be observed, the milk-clottingactivity was essentially the same in all extracts which forthe cheese-making farmers is of significant importance.SDS-PAGE of the various extracts revealed that the cy-prosins were the major proteins present in all extracts ofdried flowers (data not shown). All plants investigatedshowed similar levels of the cyprosin and the epitopeappeared not to have any significant influence on itsexpression.
Conclusions
A correlation between milk clotting activity and theamount of cyprosins present in extracts of C. cardwiculusas determined by SDS-PAGE and immunoblols has beenestablished. The cyprosins accumulate only in flowersand then, in particular, as the flowers mature. The syn-thesis of these proteinases appears to be under a strongtissue-specific regulation as indicated by immunogoldlabelling of Cynara flower buds. As a step towards de-tailed studies on this regulation we have recently isolatedand characterized a cDNA clone encoding cyprosin (Cor-deiro et al. 1994).
No proteinase of aspartic type could be detected inseeds of C, carduncuhis although it has been shown thataspartic proteina.ses are frequently present in plant seeds,e,g. buckwheat (Belozerskii et a!. 1984), wheat (Belozer-sky et al. 1989), soybean (Bond and Bowles 1983), rice(Doi et al. 1980), kidney bean (Mikkonen 1986), barley(Morris et al. 1985, Sarkkinen el al. 1992), cucumber andsquash (Polanowski et al. 1985) and Scots pine (Salmia etal. 1978). These seed aspartic proteinases may play a rolein the mobilization of reserve proteins during germina-tion. The function of cyprosins in flower.s of C. carduncu-lus is at present utiknown. However, maximum activity i.sobserved in mature flowers whicb may indicate an in-volvement in the senescence process.
Based on our results, we suggest the following modelfor the occurrence of a microheterogeneou.s population ofcyprosins in flowers of C. cardunculus. The expressionof at least three genes coding for cyprosin give rise to thecorresponding number of cyprosin zymogens. Tbesethree clo.sely related zymogens are processed by removalof the N-tennina! propeptide to produce three matureisozymes which are subsequently further proleolyticallyprocessed by intemal hydrolysis of the peptide chain withor without removal of a small peptide. The three differentheterodimeric forms of the enzyme can be the result ofcutting the peptide chain at different locations. The sameproteolytic processing of the mature isozymes couid thusresult in three forms of each cyprosin isozyme. We as-sume that the glycosylation is identical for the variousisozymes. Further studies are required to confirm lhi.sproposal. Of particular importance is the isolation andcharacterization of genes coding for cyprosin isozymes.
652 Phvsiot. Plant. 92. 1994

Acknowledgments - The authors would like to thank Prof. Bir-gitta Bergman for valuable .suggestions regarding (he silverenhancement staining. M,C,C was .supported hy a doctoralfellowship from Junta Nacional de Investigacao Cientifica eTecnica (JNICT) Portugal. This work was funded in pan by agram from the Swedish Council for Forestry and AgriculturalRe.search to P.E.B.
ReferencesBelozerskii, M.A., Dunaevskii, Ya.E., Rudenskaya, G.N. &
Stepanov, V. M. 1984. Carhoxvl proteinases from buck-wheat seeds. - Biochemistry (USSR) 49; 4 0 1 ^ 0 7 .
Betozersky, M. A., Sarbakanova, S.T. & Dumaevsky, Ya. A.1989. Aspartic proteinase from wheat seeds; isolation,properties aod action on gliadin. — Pianta 177; 321-326.
Bergman, B., Lindhlad, P., Petlersson, A., Reastrom, E. & Ti-berg, E. 1985. Immuno-gold locaUration of glutamine sy^-thelase in a iiitrogen-fixing cyanobacterium (Anabaena cy-lindrica), - Plania 166; .129-.134.
Bickar, D. & Reid, P. D. 1992. A high-affinity proiein stain forwestern blots, tissue prints, and electrophoretic gels. - Anal.Biochem. 203; 109-115.
Blutn, H., Beier, H. & Gross, H.J. 1987. Improved silver stain-ing of plant proteins. RNA and DNA in polyacrylamide gels.- Electrophoresi.s 8; 93-99.
Bond, H.M. & Bowles. D.J. 1983. Characterization of soybeanendopeptidase activity using exogenous and endogenoussubstrates. - Plant Fhysiol. 72; 345-350.
Bradford, M.M. 1976. A rapid and sen.sitive method for thequanutation of microgram cjuanlities of protein utilizing Iheprinciple of protein-dve binding. - Anal. Biocbem. 72; 248-2.54.
Bumette. W. N. 1981. Western blotting; Electrophoretic tran.sferof proteins from sodium dodecyl sulfate polyacrylamide gelsto unmodified nitrocellulose and radiographic detection withantibody and radioiodinated protein A. - Anal. Biochem.112; 195-203.
Cordeiro, M., Jakob, E., Puhan, Z., Pais, M. S. & Brodelius. P. E.1992. Milk clotting and proteolytic activities of purifiedcynarases from Cynara cardunculus; ,\ comparison to chy-mosin. - Milchwissenschaft 47; 683-687.
- , Xue, Z.-T.. Pieirzak, M., Pais, M.S. & Brodelius, RE.1994. Isolation and characterization of a cDNA from flowersof Cynara cardunculus encoding cyprosin (an aspartic pro-teinase) and its use to study the organ-specific expression otcyprosin. - Plant Mol. Eiol. 24; 733-741.
Doi,E., Shibata. D., Matoba, T. & Yonezawa. D. 1980. Charac-terization of pepstatin-sensitive acid protease in resting riceseeds. - Agric. Biol. Cheni. 44; 741-747.
Domingos, A.. Clemente, A. & Pais, .M.S. 1992. Studie.s onproteases with milk clotting activity from Centaurea cal-citrapa, - Sixth Forum for Applied Biotechnology. Bruge,s,Belgiutn, September 24-25, 1992. Meded. Fac. Landbou-wet. Univ. Gent 57 (4 part A-B), 1909-1910.
Fevereiro, P,, Cabral, J., Fonseca, M., Novais, J. & Pais, M.1986. Callus and cell suspension culture of Sylybmn maria-num. Biosynihesis of proteins with clotting activity. - Bio-technol. Lett. 8; 19-24.
Hawkes, R., Niday, E. & Godon, J. 1982. A dot-immuoobindingas.say for monoclonal and other anlibodiesi. - Anal. Bio-chem. !19; 142-147.
Heimgartner, U., Pietrzak, M., Geertsen, R., Brodelius, P., daSilva Figueiredo, A.C. & Pai.s, M.S.S. 1990. Purificationand partial characterization of milk clotting proteases fromflowers of Cynara cardunculus, - Phytochemistry 29; 1405-1410.
Johnson, D. A., Gautsch, J.W., Sportsman, J.R. & Elder, J.H.1984. Improved techmque utilizing non-fat dry milk foranaK'.sis of protein.^ and nucleic acids transferred to nitrocel-lulose. - Gene Anal. Techn. 1; i-%.
Laemmli, U.K. 1970. Cleavage of structural proteins during theassembly of the head of the bacieriophage T4. - Nature 227;680-685'.
Mikkonen, A. 1986. Activities of .some peplidases and protei-na.ses in germinating kidney bean. Pha.'ieolus vulgaris. -Physiol. Planl. 68; 282-286.
Morris', P. C , Miller, R. C. & Bowle.s, D. J. 1985. Endopeptidaseaclivitv in dry harvest-Hpe wheat barley grains. — Plant Sci.39; 12'l-124.'
Polanow,ski, A., Wilusz, T.. Kolaczkowska, M. K., Wieczorek,M. & Wilimowska-Pelc, A. 1985. Puriftcation and charac-terization of aspartic proteinasas from Cucumis sativus andCucurbita maxima .seeds. - In Aspanic Proteinases andTheir Inhibitors (V. Kostka. ed.). pp. 49-52. Walter de Gruy-ter, Berlin. ISBN 3-11-010179-3.
Richardson, K.C., Jarret. L. & Fink, E.H. 1960. Embedding inepoxy resins for ultrathin sectioning in electron microscopy.- Stain Technoi. 35; 3J3-.123.
Salmia. M. A.. Nyman, S. A. & Mikola, J. J. 1978. Characteriza-tion of the proteinases present in germinating seeds of Scotspine, Pinus sylvestris. - Physiol. Plam. 42; 252-256.
Sarkkinen. P., Kalkkinen, N., Tilgmann, C , Siuro, J., Kervinen,J. & Mikola, L. 1992. .Aspartic proteinase from barley grainsh related to mammalian ii.st>.somal catbep.sin D. - Planta186; 317-323.
Slone. K. L., LoPresti, M. B., Crawford, J. U,, DeAngelis, R. &William.s, K. R. 1989. Enzymatic dige.stion of proteins andHPLC peplide i.solation. - In A Practical Guide to Proteinand Peptide Purification for Micro.sequencing (P. T. Mat-sudaira. cd.), pp. 3 1 ^ 7 . Academic Pre.ss, San Diego, CA.ISBN 0-i2-480280-X.
Tamer. I.M. 1993. identification and partial purification of anovel tnilk clotting enzyme from Onopordum turcicum, -Biotechnol. Lett. fS; 42'7-432.
Tang, J. & Wong, R. N. S. 1987. Evolution in the structure andfunction of aspartic proteinases. - J. Cell Biochem. 33;5.3-63.
Wiklund. A. 1992. The genus Cvnara L. Asteraceae cardueae. -Bot. ] . Linn. Soc. 109; 75-123.
Edited by C. H. Bomman
Physiol. Plain. 92, 1994 653

Related Documents











