Tissue Engineering by Self-Assembly of Cells Printed into Topologically Defined Structures KAROLY JAKAB, Ph.D., 1 CYRILLE NOROTTE, 1 BROOK DAMON, Ph.D., 1 FRANCOISE MARGA, Ph.D., 1 ADRIAN NEAGU, Ph.D., 1,2 CYNTHIA L. BESCH-WILLIFORD, Ph.D., 3 ANATOLY KACHURIN, Ph.D., 4 KENNETH H. CHURCH, Ph.D., 5 HYOUNGSHIN PARK, Ph.D., 6 VLADIMIR MIRONOV, M.D., Ph.D., 7 ROGER MARKWALD, Ph.D., 7 GORDANA VUNJAK-NOVAKOVIC, Ph.D., 8 and GABOR FORGACS, Ph.D. 1,9,10 ABSTRACT Understanding the principles of biological self-assembly is indispensable for developing efficient strategies to build living tissues and organs. We exploit the self-organizing capacity of cells and tissues to construct functional living structures of prescribed shape. In our technology, multicellular spheroids (bio-ink par- ticles) are placed into biocompatible environment (bio-paper) by the use of a three-dimensional delivery device (bio-printer). Our approach mimics early morphogenesis and is based on the realization that the genetic control of developmental patterning through self-assembly involves physical mechanisms. Three- dimensional tissue structures are formed through the postprinting fusion of the bio-ink particles, in analogy with early structure-forming processes in the embryo that utilize the apparent liquid-like behavior of tissues composed of motile and adhesive cells. We modeled the process of self-assembly by fusion of bio- ink particles, and employed this novel technology to print extended cellular structures of various shapes. Functionality was tested on cardiac constructs built from embryonic cardiac and endothelial cells. The postprinting self-assembly of bio-ink particles resulted in synchronously beating solid tissue blocks, showing signs of early vascularization, with the endothelial cells organized into vessel-like conduits. INTRODUCTION G ENES, THROUGH DIFFERENTIATION, SET UP the inherent physical and chemical properties of cells, extracellular matrix, and tissues. These in turn generate forces that drive structure formation and cause subsequent alterations in gene activity. It is this delicate interplay of genetic, molecular, and physical factors that underlies the evolving modern understanding of morphogenesis as a self-assembly pro- cess. 1–3 To paraphrase Whitesides and Grzybowski, 4 self- assembly is the autonomous organization of components, from an initial state into final pattern or structure without 1 Department of Physics, University of Missouri, Columbia, Missouri. 2 Department of Biophysics, Victor Babes University of Medicine and Pharmacy, Timisoara, Romania. 3 Department of Veterinary Pathobiology, University of Missouri, Columbia, Missouri. 4 vaxDesign, Inc., Orlando, Florida. 5 nScrypt, Inc., Orlando, Florida. 6 Department of Chemical Engineering, Massachusetts Institute of Technology, Cambridge, Massachusetts. 7 Department of Cell Biology and Anatomy, Medical University of South Carolina, Charleston, South Carolina. 8 Department of Biomedical Engineering, Columbia University, New York, New York. 9 Department of Biology, University of Missouri, Columbia, Missouri. 10 Department of Biomedical Engineering, University of Missouri, Columbia, Missouri. TISSUE ENGINEERING: Part A Volume 14, Number 3, 2008 # Mary Ann Liebert, Inc. DOI: 10.1089/tea.2007.0173 413

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Tissue Engineering by Self-Assembly of Cells Printed
into Topologically Defined Structures
KAROLY JAKAB, Ph.D.,1 CYRILLE NOROTTE,1 BROOK DAMON, Ph.D.,1
FRANCOISE MARGA, Ph.D.,1 ADRIAN NEAGU, Ph.D.,1,2 CYNTHIA L. BESCH-WILLIFORD, Ph.D.,3
ANATOLY KACHURIN, Ph.D.,4 KENNETH H. CHURCH, Ph.D.,5 HYOUNGSHIN PARK, Ph.D.,6
VLADIMIR MIRONOV, M.D., Ph.D.,7 ROGER MARKWALD, Ph.D.,7
GORDANA VUNJAK-NOVAKOVIC, Ph.D.,8 and GABOR FORGACS, Ph.D.1,9,10
ABSTRACT
Understanding the principles of biological self-assembly is indispensable for developing efficient strategiesto build living tissues and organs. We exploit the self-organizing capacity of cells and tissues to constructfunctional living structures of prescribed shape. In our technology, multicellular spheroids (bio-ink par-ticles) are placed into biocompatible environment (bio-paper) by the use of a three-dimensional deliverydevice (bio-printer). Our approach mimics early morphogenesis and is based on the realization that thegenetic control of developmental patterning through self-assembly involves physical mechanisms. Three-dimensional tissue structures are formed through the postprinting fusion of the bio-ink particles, inanalogy with early structure-forming processes in the embryo that utilize the apparent liquid-like behaviorof tissues composed of motile and adhesive cells. We modeled the process of self-assembly by fusion of bio-ink particles, and employed this novel technology to print extended cellular structures of various shapes.Functionality was tested on cardiac constructs built from embryonic cardiac and endothelial cells. Thepostprinting self-assembly of bio-ink particles resulted in synchronously beating solid tissue blocks,showing signs of early vascularization, with the endothelial cells organized into vessel-like conduits.
INTRODUCTION
GENES, THROUGH DIFFERENTIATION, SET UP the inherent
physical and chemical properties of cells, extracellular
matrix, and tissues. These in turn generate forces that drive
structure formation and cause subsequent alterations in gene
activity. It is this delicate interplay of genetic, molecular,
and physical factors that underlies the evolving modern
understanding of morphogenesis as a self-assembly pro-
cess.1–3 To paraphrase Whitesides and Grzybowski,4 self-
assembly is the autonomous organization of components,
from an initial state into final pattern or structure without
1Department of Physics, University of Missouri, Columbia, Missouri.2Department of Biophysics, Victor Babes University of Medicine and Pharmacy, Timisoara, Romania.3Department of Veterinary Pathobiology, University of Missouri, Columbia, Missouri.4vaxDesign, Inc., Orlando, Florida.5nScrypt, Inc., Orlando, Florida.6Department of Chemical Engineering, Massachusetts Institute of Technology, Cambridge, Massachusetts.7Department of Cell Biology and Anatomy, Medical University of South Carolina, Charleston, South Carolina.8Department of Biomedical Engineering, Columbia University, New York, New York.9Department of Biology, University of Missouri, Columbia, Missouri.10Department of Biomedical Engineering, University of Missouri, Columbia, Missouri.
TISSUE ENGINEERING: Part AVolume 14, Number 3, 2008# Mary Ann Liebert, Inc.DOI: 10.1089/tea.2007.0173
413

external intervention. This definition clearly applies to
processes taking place in living organisms, in particular
during early morphogenesis. We have demonstrated that
biological self-assembly can be employed to construct liv-
ing structures of prescribed geometry.5 Our approach re-
lies on morphogenetic driving forces known to operate in
early embryonic development, specifically on the liquid-like
properties of tissues composed of motile and adhesive
cells.6–9 In particular, aggregates of such cells spontaneously
round up,10 and upon contact, fuse, similarly to coalescing
liquid drops.5,11 We used our earlier findings to develop a
novel tissue engineering technology with sound scientific
underpinning rooted in developmental biology that has
a number of advantages when compared with classical
approaches.
The basic idea underlying classical tissue engineering12 is
to seed living cells into biocompatible and eventually bio-
degradable environment (the scaffold), and then culture the
system in a bioreactor so that the initial cell population can
expand into a tissue.13 With an appropriate scaffold that
mimics the biological extracellular matrix, it is expected that
the developing tissue will adopt both the form and function
of the desired organ. The organ would then be implanted into
the recipient.14 This approach has lead to some spectacular
results.15,16 Despite these successes, the classical tissue en-
gineering approach faces serious hurdles. Selection of the
ideal biomaterial scaffold for a given cell type is problem-
atic17 and has been accomplished to date mostly by trial and
error. Even if the right biomaterial is available, achiev-
ing high enough cell density to construct a viable tissue is
extremely time consuming. Preshaping the scaffold may
present further difficulties. Recent efforts have concentrated
on scaffoldless tissue engineering18–21 and produced prom-
ising results even if they are limited to specific applications
and are not scalable. A major unresolved problem in tissue
engineering that hampers its commercial viability is that no
technology exists at present to produce off-the-shelf prod-
ucts. To address some of these issues, it has been suggested
that the process of building three-dimensional (3D) biolog-
ical structures could significantly be accelerated by the
technology of bioprinting: the automated, computer-aided
deposition of cells and cell aggregates.22 Commercially
available inkjet printers have been successfully redesigned
or new ones built to specifically deliver biological material
into scaffold fabricated according to a computer-aided de-
sign template.23,24 Pressure-operated mechanical extruders
have been developed to handle biomolecular assemblies or
live cells.25–28 The novel technology described in this
work is compatible with methods of rapid prototyping, is
scalable, exploits intrinsic properties of cells and tissues,
and makes it possible to reliably and reproducibly build
organoids of defined topology and functionality in vitro.
The 3D structures are arrived at via a three-phase process:
(i) preprocessing, or bio-ink preparation; (ii) processing, that
is, the actual automated delivery/printing of the bio-ink
particles into the bio-paper by the bio-printer; and (iii)
postprocessing, that is, the maturation/incubation of the
printed construct in the bioreactor. Final structure formation
takes place during postprocessing via the fusion of the bio-
ink particles (hence ‘‘ink’’).
MATERIALS AND METHODS
Cushion tissue fusion
Leghorn chicken eggs (Ozark Hatcheries, Neosho, MO)
were incubated at 408C in 80% humidity for 4–5 days.
Excised cushion tissue explants were washed in phosphate-
buffered saline (PBS) and cut into similar-size fragments.
Fragments were incubated in gyratory shaker (at 120 rpm,
5% CO2, 378C). Within 24–36 h (depending on size) this
procedure reproducibly yielded round aggregates. For
in vivo fusion studies, the eggs were cut open, embryos
were placed in Earle’s balanced salt solution (EBSS; Invi-
trogen, Carlsbad, CA), and the heart was dissected. Fusing
atrio-ventricular (AV) cushion tissue buds of approximately
400 mm were pinched from the opened myocardial tube at
regular time intervals between Hamburger-Hamilton (HH)
stages 26 and 28 (5–6 days), washed in PBS, placed in a
Petri dish with fresh EBSS, and photographed with a charge-
coupled device camera (Diagnostic Instruments, Sterling
Heights, MI) attached to a dissecting microscope (Olympus,
Center Valley, PA).
Preparation of bio-ink particles
and cardiac constructs
AV tissue fragments from 9-day-old Leghorn chicken
embryos were dissociated into single cells, then plated in
tissue culture dishes in Dulbecco’s modified Eagle’s me-
dium (DMEM) (Invitrogen) supplemented with 10% fetal
bovine serum (FBS) (US Biotechnologies, Parkerford, PA),
and allowed to grow to confluence. Human vascular endo-
thelial cells (CRL1730) were purchased from ATCC
(Manassas, VA) and grown in F12K medium (Invitrogen)
supplemented with 10% FBS, 0.1mg/mL heparin (Sigma,
St. Louis, MO), and 0.03mg/mL endothelial cell growth
supplement (Upstate, Lake Placid, NY). To prepare spheri-
cal cell aggregates, cell cultures were washed twice with
Hank’s balanced salt solution containing 2mM calcium
chloride, and treated with 0.1% Trypsin for 10min. Cardiac
and endothelial cells were counted using a hemacytometer,
mixed in 4:1 ratio, and then centrifuged at 2500 rpm. Cul-
tures of endothelial cells between seven and nine passage
doublings were used. The resulting pellet was aspirated into
capillary micropipettes of 500 mm diameter and incubated at
378C with 5% CO2 for 10min. The micropipette was
transferred into an in-house-built special apparatus, which
extruded the cellular ‘‘sausage’’ and cut it into equal-size
cylinders (height-to-diameter ratio¼ 1). Cellular cylinders
414 JAKAB ET AL.

rapidly rounded into spheres. Spheroids were aspirated into
the capillary micropipettes to form the bio-ink cartridge.
Similar procedure was followed with the Chinese hamster
ovary (CHO) cells.
Preparation of bio-paper
Rat tail collagen type 1 (Sigma) was dissolved in 1M
acetic acid, the pH of the solution was adjusted to 7.2
with 2M sodium hydroxide and HEPES buffer, and then
the resulting solution was diluted to 1mg/mL with
DMEM–F12K mixture. Vascular endothelial growth factor
(VEGF165, Sigma) was added to the gel before polymeri-
zation (20 ng/mL).
Visualization
To visualize details of fusion of bio-ink particles made of
CHO cells or mixture of cardiac and endothelial cells, these
were stained with red and green membrane-intercalating
dyes (PKH26 and PKH2, Sigma) according to the proto-
col supplied by the manufacturer. Cells were replated af-
ter labeling to recover from the staining procedure and
then used to make aggregates of different fluorescent col-
ors. Transparency of the collagen gels permitted the use of
bright-field imaging to monitor the assembling structures.
Histology
Cell–gel constructs were fixed in paraformaldehyde
(PFA). PFA was subsequently washed out. Samples were
dehydrated and prepared for immunohistochemistry (sec-
tioning into 5-mm-thick layers, paraffin embedding, etc.).
Immunohistology was performed to assess cellular distri-
bution in the constructs using hematoxylin and eosin (H&E);
evaluate the expression of cardiac proteins, using troponin I
antibody (primary: rabbit anti-troponin antibodies; second-
ary: anti-rabbit immunoglobulin G conjugated with FITC
[fluorescein isothiocyanate; Chemicon International, Inc.,
Temecula, CA]); and visualize the pattern of endothelial
cells, using CD31 antibody (primary: mouse anti-human
CD31 monoclonal antibody; secondary: EnVisionþ, a horse-
radish peroxidase–labeled polymer conjugated with anti-
mouse antibodies [DAKO, Glostrup, Denmark]).
Contractile behavior
Functionality of the engineered cardiac constructs was
evaluated by monitoring contractile activity at 10�magni-
fication using a Nikon Diaphot microscope (Nikon Instru-
ments, Inc., Melville, NY) as in our previous studies.29
To measure the amplitude of contractions, video-recorded
beating sequences were digitized at the rate of 30 frames/
second. The en-face area of each construct was measured as
function of time using image analysis (Scion Image soft-
ware). Amplitude of contraction was expressed as frac-
tional area change. Decrease in the total en-face area was
used as a measure of the contraction amplitude to avoid
determining the axis of maximal shortening in a construct
of an irregular rectangular shape.
RESULTS
Early morphogenesis based on tissue liquidity
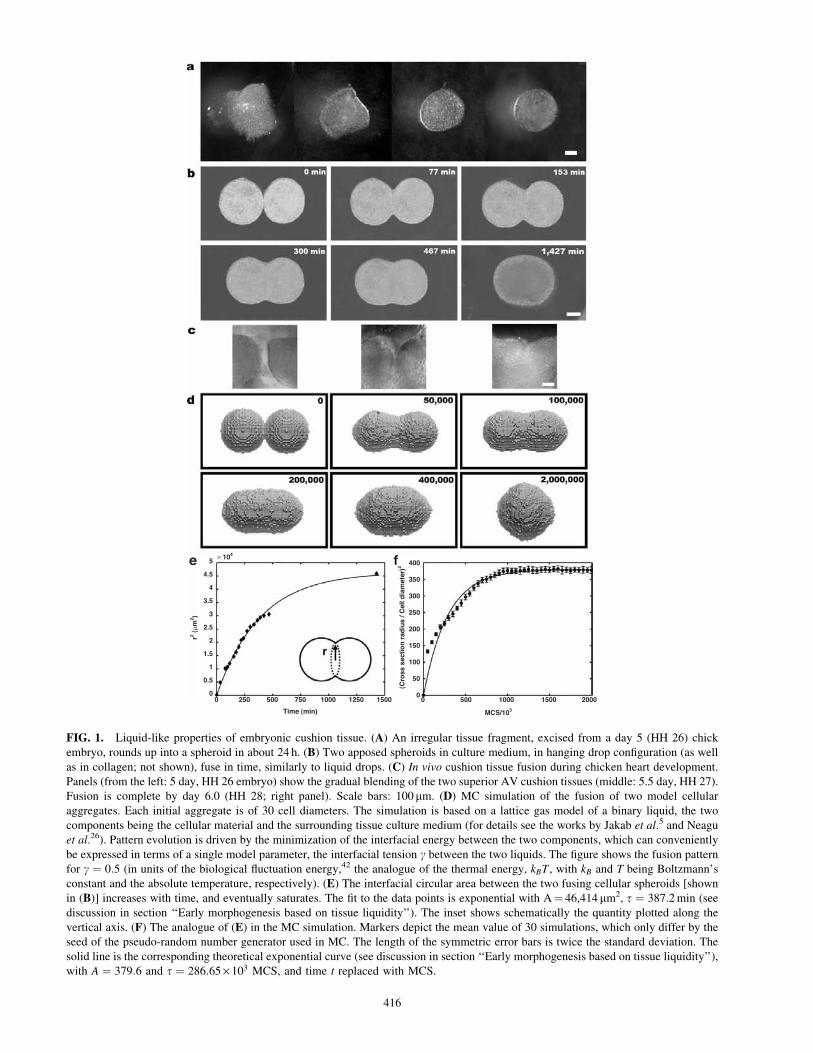
To demonstrate that the outlined tissue engineering
program indeed is executed similarly to morphogenetic
processes, we specifically considered the in vitro, midline
fusion of embryonic chicken cardiac cushions, the process
leading to septation of the heart into the four chambers and
valve formation.30 We dissected early chicken embryonic
heart to extract AV cushions at the stage when their in vivo
fusion would commence (HH 26, day 5). When irregularly
shaped tissue explants were isolated and incubated in cul-
ture medium, they naturally and progressively rounded into
almost perfect spherical shape (Fig. 1A). This observation
suggests that these tissue spheroids have properties in
common with liquid droplets in that they try to minimize
their interfacial area with the surroundings. Indeed, tissues
composed of adhesive and motile cells have measurable
surface tensions and viscosities.1,10,31–34
When two similar cushion tissue spheroids were placed
contiguously, they fused into a single spheroid (Fig. 1B).
The in vitro fusion process resembled the in vivo process
(Fig. 1C) of AV septum formation during heart develop-
ment.30 Monte Carlo (MC) simulations35 based on a model
of binary liquids5,26 were in excellent agreement with ex-
perimental results (Fig. 1D). The finding that the function
r2¼A(1� e�t=t) fitted well the time variation of the cir-
cular interfacial area (of instantaneous radius r) between
the two spherical fragments (Fig. 1E) gave further support
to the notion of tissue liquidity. This expression is consis-
tent with the analytical result r2¼sRt=(Z) for the fusion of
droplets of highly viscous materials (Z: viscosity; s: inter-facial tension between the liquid and the embedding me-
dium; R: initial radius of the fusing drops), valid in the limit
t/t�136 and relates the relaxation time t, characterizing therate of fusion, to the material properties of the tissue:
t¼22=3RZ=s. (Here we used the relation A¼ 22=3R2, which
follows from the conservation of volume during fusion.)
The exponential growth of the interfacial area is similar to
experimental results obtained in real liquids.11 Measure-
ment of cushion tissue–culture medium interfacial tension
with earlier described methods34 resulted in s � 16:0 dyn/
cm (0.016N/m). This value, when combined with the ex-
pression and experimentally obtained information for t,leads to Z � 107 Poise for chicken embryonic cushion tis-
sue, a value consistent with earlier estimates obtained with
an independent method for various embryonic chicken
tissues.33 The exponential function r2 ¼ A(1� e�MCS=t)
described accurately the evolution of fusion also within the
TISSUE ENGINEERING BY PRINTING CELL AGGREGATES 415
× 104
0 250 500 750
Time (min)
1000 1250 1500 000
500
MCS/103
1000 1500 2000
50
100
150
200
250
300
350
400
(Cro
ss s
ecti
on
rad
ius
/ Cel
l dia
met
er)2
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5 f
r2 (µm
2 )
e
FIG. 1. Liquid-like properties of embryonic cushion tissue. (A) An irregular tissue fragment, excised from a day 5 (HH 26) chick
embryo, rounds up into a spheroid in about 24 h. (B) Two apposed spheroids in culture medium, in hanging drop configuration (as well
as in collagen; not shown), fuse in time, similarly to liquid drops. (C) In vivo cushion tissue fusion during chicken heart development.
Panels (from the left: 5 day, HH 26 embryo) show the gradual blending of the two superior AV cushion tissues (middle: 5.5 day, HH 27).
Fusion is complete by day 6.0 (HH 28; right panel). Scale bars: 100mm. (D) MC simulation of the fusion of two model cellular
aggregates. Each initial aggregate is of 30 cell diameters. The simulation is based on a lattice gas model of a binary liquid, the two
components being the cellular material and the surrounding tissue culture medium (for details see the works by Jakab et al.5 and Neagu
et al.26). Pattern evolution is driven by the minimization of the interfacial energy between the two components, which can conveniently
be expressed in terms of a single model parameter, the interfacial tension g between the two liquids. The figure shows the fusion pattern
for g ¼ 0:5 (in units of the biological fluctuation energy,42 the analogue of the thermal energy, kBT , with kB and T being Boltzmann’s
constant and the absolute temperature, respectively). (E) The interfacial circular area between the two fusing cellular spheroids [shown
in (B)] increases with time, and eventually saturates. The fit to the data points is exponential with A¼ 46,414mm2, t ¼ 387:2min (see
discussion in section ‘‘Early morphogenesis based on tissue liquidity’’). The inset shows schematically the quantity plotted along the
vertical axis. (F) The analogue of (E) in the MC simulation. Markers depict the mean value of 30 simulations, which only differ by the
seed of the pseudo-random number generator used in MC. The length of the symmetric error bars is twice the standard deviation. The
solid line is the corresponding theoretical exponential curve (see discussion in section ‘‘Early morphogenesis based on tissue liquidity’’),
with A ¼ 379:6 and t ¼ 286:65· 103 MCS, and time t replaced with MCS.
416

MC simulation as the process approached its completion
but not at its initiation (Fig. 1F). To simulate the fusion pro-
cess, time has been replaced by Monte Carlo Steps (MCSs),
one step being defined as a sequence of computations, which
give each interfacial cell a chance to experience a move
(dictated by the random process implemented in MC sim-
ulations5,35). This is to be expected since in MC pattern
evolution is driven by energy minimization, thus being
more accurate near equilibrium than far from it.
These results imply that the time evolution of the fusion
pattern is indeed similar to that of coalescing liquid drops.
Our findings suggest that fusion of early (primitive) cardiac
cushions during heart development, once under way, may
be driven by the inherent physical properties of the tissue.37
The novel tissue engineering technology
Tissue liquidity, as described above, provides the sci-
entific basis for our tissue engineering technology to
build complex biological structures. The components of this
a
f b
c
d
e
FIG. 2. Components of the novel technology. (A) A patent-
pending device (schematic) cuts the earlier prepared tissue/
cellular slurry into cylindrical aggregates with aspect ratio 1.
(B) The spheroidal bio-ink particles result from the rapid rounding
of the tissue/cellular cylinders upon incubation (the one shown is
of 500 mm and is made of CHO cells fluorescently labeled with
a membrane dye). (C) The printer cartridge is a micropipette
housing the spherical multicellular aggregates. (D) The printer.
The one shown has been designed and built by nScrypt (Orlando,
FL). It is a fully computer-controlled delivery device. Three-
dimensional deposition of bio-ink particles (with accuracy of mm)
is achieved by the movable x–y stage and three z-directional
printing heads. Two of these are used to print the spheroids, which
are extruded from the cartridge/micropipette by the positive dis-
placement of a piston within the micropipette. The third unit is
pressure operated and is used to print the embedding hydrogel
(e.g., collagen gel). Each extruder is equipped with a camera,
providing full visual control of printing. (E) Spheroids are de-
livered one by one into the hydrogel according to a computer
script encoding the printable shape. Note that the hydrogel itself
is also printed, here a 1�1�0.2 cm layer of collagen type 1 gel.
(F) The outcome of the printing process is a set of discrete
spheroids, here 500mm aggregates of CHO cells, fluorescently
labeled with red or green membrane dyes.
a
d
e6
Am
plit
ude (
µm)
4 < A > = 1.78 µm < T > = 1.84 s
2
0
0 5 10 15
Time (s)
20 25 30
b c
FIG. 3. Printing cellular structures of defined topology.
(A) Upper panel: a circular arrangement of bio-ink particles,
composed of spherical aggregates of CHO cells (neighboring
aggregates were labeled alternatively with red and green dyes).
Lower panel: a fused circle, after about 60 h. (B) The time evo-
lution of a vertically printed three-layered tubular structure of
CHO cells, built from circles shown in panel (A). (C) Upper
panel: a horizontally printed tube of 12 layers composed of hu-
man umbilical smooth muscle cells. Lower panel: a horizontally
printed branching CHO tube. (D) Building a pulsatile cardiac
construct. A 6�6 array of cardiac bio-ink particles (without en-
dothelial cells; left) printed into 1mg/mL collagen 1 bio-paper. If
endothelial cells are present (middle panel; labeled with red dye;
cardiac cells labeled with green dye), they first accumulate at the
boundaries between neighboring aggregates and later in the
originally acellular region between four aggregates (although
many remain also inside the aggregates). In a 6�6 array, fusion
is complete in about 70 h (right panel). (E) The synchronous
beating of the fused construct in (D). <A> and <T> are the
amplitude and period of beating, respectively. All bio-ink particles
in this figure are of 500 mm diameter. The colored panels [in
(A) and (D)] were prepared by fluorescently labeling the cells with
red or green membrane dyes.
TISSUE ENGINEERING BY PRINTING CELL AGGREGATES 417

technology are shown in Figure 2. We first prepared
monodisperse spherical bio-ink particles (Fig. 2A, B) with
composition dictated by the properties of the desired struc-
ture. The bio-ink particles were packaged into cartridges
(Fig. 2C; micropipettes of 300–500 mm inner diameter).
Cartridges were inserted into the bio-printer (Fig. 2D) and
delivered into the bio-paper/hydrogel (Fig. 2E) according to
a computer script that encodes the shape of the structure to
be printed (Fig. 2F).
Building organoids of definite topology
Printing results in a set of discrete biological structures,
which subsequently undergo fusion into continuous struc-
tures. As a proof of concept, we have applied this technology
to print bio-ink particles in the form of circles (Fig. 3A),
cylinders (Fig. 3B, C), or sheets (Fig. 3D), which upon fu-
sion yielded constructs of specific shape, in particular hol-
low tubes and solid blocks (Fig. 3B–D). To illustrate the
flexibility of the method, we prepared tubular structures in
two different ways: vertical and horizontal. In the first case
(Fig. 3B), bio-ink particles are delivered in circular pattern,
and circles are printed layer by layer in the vertical direction.
In the second case (Fig. 3C), chains of bio-ink particles are
printed again layer by layer, with the number of chains
varying in each layer. Horizontally printed cylinders can be
arranged to arrive at branching tubular structures upon fu-
sion of the bio-ink particles (Fig. 3C). (Once the right con-
ditions for structure formation had been established [see
‘‘Discussion’’ section], reproducibility was verified by per-
forming the relevant experiments at least four times.)
Functionality of the printed tissue constructs
To verify that postprinting structure formation leads
to functional tissue constructs, we prepared multicellular
spheroids employing fragments of 9-day chicken embryonic
heart tissue, containing various cell types. In particular, the
fragments contained the fused AV cushions, whose mesen-
chyme at this stage begins to differentiate into endothelial
and fibrous (fibroblastic) tissue and, uniquely in the chick,
also into myocardial cells (e.g., the large muscular leaflet of
the triscupid valve38,39). We used the cells of the dissociated
fragments to prepare 500 mm spherical aggregates with or
without endothelial cells (human umbilical endothelial cells,
randomly dispersed in the spheroids in the presence of
VEGF). We then printed a 6�6 array of the spheroids in a
collagen type 1 gel bio-paper (Fig. 3D). After deposition the
aggregate–gel construct was incubated in tissue culture
medium for 5 days. The bio-ink particles completely fused
over a period of 70 h to form a thick graft. The engineered
tissue manifested characteristic properties of authentic
chicken cardiomyocytes as revealed by its synchronous
beating after about 90 h (Fig. 3E) and immunohistochemis-
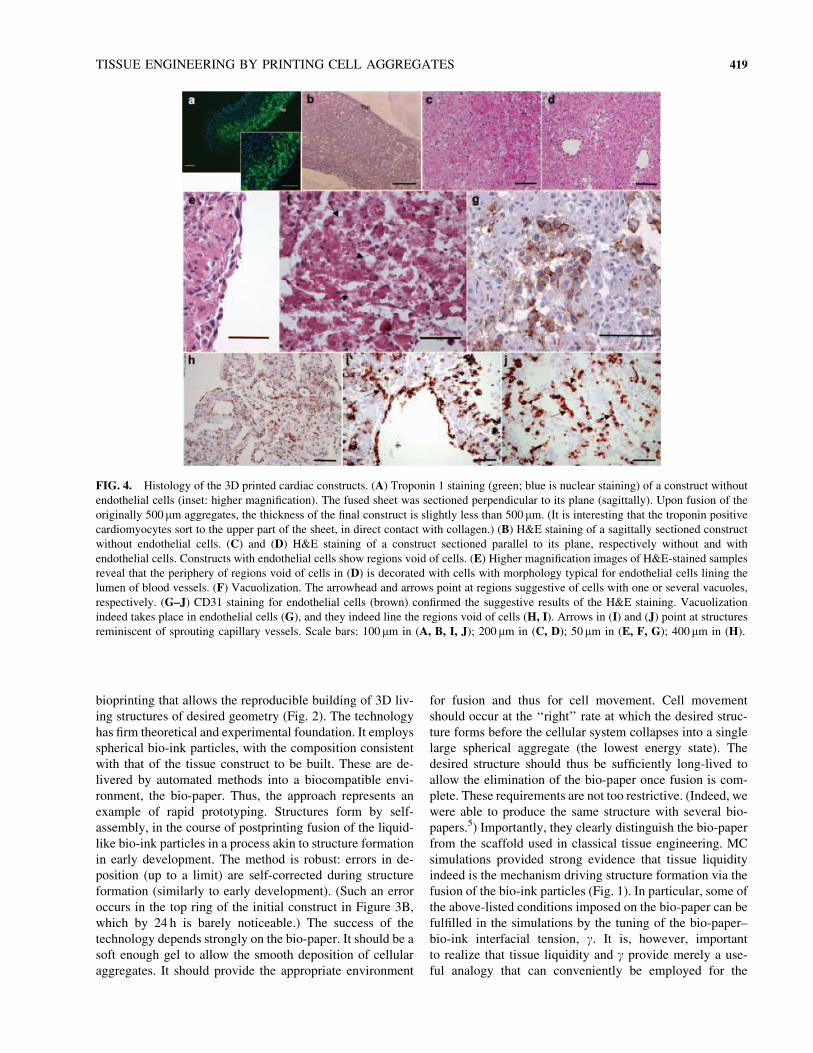
try (Fig. 4).
Vascularization of the printed tissue constructs
Long-term survival and function of engineered tissues can
be maintained only with appropriate vascularization.40 We
hypothesized that if printing of the cardiac construct, as de-
scribed above, is repeated with aggregates composed of a
mixture of cardiac and endothelial cells (human umbilical
vein endothelial cell), self-organizing properties will result
in the formation of primitive vasculature. Thus, a 6�6 array
(similar to that shown in Fig. 3D) was printed onto the col-
lagen gel bio-paper. VEGF, an endothelial mitogen and
vasculogenic stimulator, was added to the collagen solution
before gelation. For the postprinting incubation, the culture
medium was also supplemented with VEGF. Synchronous
beating was again observed, after approximately 90 h. Con-
structs that formed without endothelial cells (Fig. 4A–C)
were always compact and had cells present throughout
their entire volume. However, tissue constructs that formed
with endothelial cells (Fig. 4D) consistently showed regions
void of cells with internal boundaries notably decorated
with characteristically elongated cells (Fig. 4E), as well as
conduit-like structures (Fig. 4F). The formation of such
structures seems to be facilitated by the intrinsic features of
the method, the acellular regions between four neighboring
spherical aggregates, and the boundaries between pairs of
aggregates, where endothelial cells preferentially accumu-
late (middle panel in Fig. 3D). Further, cells with either nu-
merous intracellular vacuoles or a single large internal
vacuole, characteristic for lumen-forming endothelial cells,41
could be observed (Fig. 4F). Such structures were never
observed in the constructs without endothelial cells. Immu-
nohistochemistry provided evidence that these cells and those
lining the periphery of the hollow acellular regions indeed
were endothelial cells (Fig. 4G–J). Structures suggestive of
forming capillaries could also be discerned (Fig. 4I, J).
DISCUSSION
This work had four objectives. First, to further demon-
strate that embryonic tissues (or, more generally, cell ag-
gregates composed of adhesive and motile cells) have
liquid-like properties. In particular, we showed both exper-
imentally and by modeling that this concept applies not only
to equilibrium cellular arrangements (such as those observed
in sorting10,42) but also to time-dependent phenomena, such
as the fusion of two tissue fragments (Fig. 1). Second, we
wished to provide an example of a true early morphogenetic
phenomenon that can be interpreted in terms of tissue li-
quidity. For this, we considered the in vivo fusion of em-
bryonic chicken cardiac cushions, the hallmark of cardiac
development. We provided evidence that this morphoge-
netic process indeed is similar to its in vitro counterpart,
which in turn obeys the laws of fluid mechanics (Fig. 1).
Third, we described a novel tissue engineering technology,
418 JAKAB ET AL.
bioprinting that allows the reproducible building of 3D liv-
ing structures of desired geometry (Fig. 2). The technology
has firm theoretical and experimental foundation. It employs
spherical bio-ink particles, with the composition consistent
with that of the tissue construct to be built. These are de-
livered by automated methods into a biocompatible envi-
ronment, the bio-paper. Thus, the approach represents an
example of rapid prototyping. Structures form by self-
assembly, in the course of postprinting fusion of the liquid-
like bio-ink particles in a process akin to structure formation
in early development. The method is robust: errors in de-
position (up to a limit) are self-corrected during structure
formation (similarly to early development). (Such an error
occurs in the top ring of the initial construct in Figure 3B,
which by 24 h is barely noticeable.) The success of the
technology depends strongly on the bio-paper. It should be a
soft enough gel to allow the smooth deposition of cellular
aggregates. It should provide the appropriate environment
for fusion and thus for cell movement. Cell movement
should occur at the ‘‘right’’ rate at which the desired struc-
ture forms before the cellular system collapses into a single
large spherical aggregate (the lowest energy state). The
desired structure should thus be sufficiently long-lived to
allow the elimination of the bio-paper once fusion is com-
plete. These requirements are not too restrictive. (Indeed, we
were able to produce the same structure with several bio-
papers.5) Importantly, they clearly distinguish the bio-paper
from the scaffold used in classical tissue engineering. MC
simulations provided strong evidence that tissue liquidity
indeed is the mechanism driving structure formation via the
fusion of the bio-ink particles (Fig. 1). In particular, some of
the above-listed conditions imposed on the bio-paper can be
fulfilled in the simulations by the tuning of the bio-paper–
bio-ink interfacial tension, g. It is, however, important
to realize that tissue liquidity and g provide merely a use-
ful analogy that can conveniently be employed for the
FIG. 4. Histology of the 3D printed cardiac constructs. (A) Troponin 1 staining (green; blue is nuclear staining) of a construct without
endothelial cells (inset: higher magnification). The fused sheet was sectioned perpendicular to its plane (sagittally). Upon fusion of the
originally 500mm aggregates, the thickness of the final construct is slightly less than 500 mm. (It is interesting that the troponin positive
cardiomyocytes sort to the upper part of the sheet, in direct contact with collagen.) (B) H&E staining of a sagittally sectioned construct
without endothelial cells. (C) and (D) H&E staining of a construct sectioned parallel to its plane, respectively without and with
endothelial cells. Constructs with endothelial cells show regions void of cells. (E) Higher magnification images of H&E-stained samples
reveal that the periphery of regions void of cells in (D) is decorated with cells with morphology typical for endothelial cells lining the
lumen of blood vessels. (F) Vacuolization. The arrowhead and arrows point at regions suggestive of cells with one or several vacuoles,
respectively. (G–J) CD31 staining for endothelial cells (brown) confirmed the suggestive results of the H&E staining. Vacuolization
indeed takes place in endothelial cells (G), and they indeed line the regions void of cells (H, I). Arrows in (I) and (J) point at structures
reminiscent of sprouting capillary vessels. Scale bars: 100 mm in (A, B, I, J); 200 mm in (C, D); 50mm in (E, F, G); 400 mm in (H).
TISSUE ENGINEERING BY PRINTING CELL AGGREGATES 419

characterization of the outlined tissue engineering method.
Ultimately, postprinting structure formation is governed by
molecular mechanisms underlying cell–cell and cell–matrix
adhesion, as well as cell motility. (Indeed, g can be related tothe density and nature of cell–cell31,32 and cell–matrix ad-
hesion molecules.43) Finally, our forth objective was to
present specific examples of structures built using the novel
technology (Fig. 3) and provide evidence that these tissue
structures manifest the functionality of the real tissue they
are supposed to replace. We showed that building small-
diameter tubes (major structures in complex organisms)
or extended 3D grafts can easily be accomplished. Sig-
nificantly, the self-organizing capacity of cells assures the
onset of vascular network formation within the 3D con-
structs (Fig. 4), even without their implantation into a living
organism.44–47 Our work implies that functional tissue
constructs containing multiple cell types could be built by
rapid prototyping (i.e., bio-printing), suggesting that the off-
the-shelf availability of organoid-like structures could be
achieved.
ACKNOWLEDGMENTS
Support for this research was provided by the National
Science Foundation FIBR program (EF-0526854 to VM,
RM, KC, GF), the National Institutes of Health (P41
EB002520 and RO1 HL076485 to GV-N) and the Medical
University of South Carolina Bioprinting Research Center
(to VM).
REFERENCES
1. Forgacs, G., and Newman, S.A. Biological Physics of the
Developing Embryo. Cambridge: Cambridge University Press,
2005.
2. Newman, S.A. Generic physical mechanisms of tissue mor-
phogenesis: a common basis for development and evolution.
J. Evol. Biol. 7, 467, 1994.
3. Wolpert, L. Principles of Development. Oxford: Oxford
University Press, 1998.
4. Whitesides, G.M., and Grzybowski, B. Self-assembly at all
scales. Science 295, 2418, 2002.
5. Jakab, K., Neagu, A., Mironov, V., Markwald, R.R., and
Forgacs, G. Engineering biological structures of prescribed
shape using self-assembling multicellular systems. Proc. Natl.
Acad. Sci. USA 101, 2864, 2004.
6. Godt, D., and Tepass, U. Drosophila oocyte localization is
mediated by differential cadherin-based adhesion. Nature 395,
387, 1998.
7. Gonzalez-Reyes, A., and St. Johnston, D. Patterning of the
follicle cell epithelium along the anterior-posterior axis dur-
ing Drosophila oogenesis. Development 125, 2837, 1998.
8. Hayashi, T., and Carthew, R.W. Surfacemechanics mediate pat-
tern formation in the developing retina. Nature 431, 647, 2004.
9. Steinberg, M.S., and Poole, T.J. Liquid behavior of embry-
onic tissues. In: Bellairs, R., Curtis, A.S.G., and Dunn, G.,
eds. Cell Behaviour. Cambridge: Cambridge University Press,
1982, p. 583.
10. Foty, R.A., Pfleger, C.M., Forgacs, G., and Steinberg, M.S.
Surface tensions of embryonic tissues predict their mutual
envelopment behavior. Development 122, 1611, 1996.
11. Yao, W., Maris, H.J., Pennington, P., and Seidel, G.M.
Coalescence of viscous liquid drops. Phys. Rev. E Stat.
Nonlin. Soft Matter Phys. 71, 016309, 2005.
12. Langer, R., and Vacanti, J.P. Tissue engineering. Science 260,
920, 1993.
13. Khademhosseini, A., Langer, R., Borenstein, J., and Vacanti,
J.P. Microscale technologies for tissue engineering and
biology. Proc. Natl. Acad. Sci. USA 103, 2480, 2006.
14. Griffith, L., and Naughton, G. Tissue engineering—current
challenges and expanding opportunities. Science 295, 1009,
2002.
15. Atala, A., Bauer, S.B., Soker, S., Yoo, J.J., and Retik, A.B.
Tissue-engineered autologous bladders for patients needing
cystoplasty. Lancet 367, 1241, 2006.
16. Oberpenning, F., Meng, J., Yoo, J.J., and Atala, A. De novo
reconstitution of a functional mammalian urinary bladder by
tissue engineering. Nat. Biotechnol. 17, 149, 1999.
17. Dahl, S.L., Rhim, C., Song, Y.C., and Niklason, L.E. Me-
chanical properties and compositions of tissue engineered and
native arteries. Ann. Biomed. Eng. 35, 348, 2007.
18. L’Heureux, N., Dusserre, N., Konig, G., Victor, B., Keire, P.,
Wight, T.N., Chronos, N.A., Kyles, A.E., Gregory, C.R.,
Hoyt, G., Robbins, R.C., and McAllister, T.N. Human tissue-
engineered blood vessels for adult arterial revascularization.
Nat. Med. 12, 361, 2006.
19. L’Heureux, N., Paquet, S., Labbe, R., Germain, L., and Au-
ger, F.A. A completely biological tissue-engineered human
blood vessel. FASEB J. 12, 47, 1998.
20. McGuigan, A.P., and Sefton, M.V. Vascularized organoid
engineered by modular assembly enables blood perfusion.
Proc. Natl. Acad. Sci. USA 103, 11461, 2006.
21. Shimizu, T., Yamato, M., Isoi, Y., Akutsu, T., Setomaru, T.,
Abe, K., Kikuchi, A., Umezu, M., and Okano, T. Fabrication
of pulsatile cardiac tissue grafts using a novel 3-dimensional
cell sheet manipulation technique and temperature-responsive
cell culture surfaces. Circ. Res. 90, e40, 2002.
22. Mironov, V., Boland, T., Trusk, T., Forgacs, G., and Mark-
wald, R.R. Organ printing: computer-aided jet-based 3D tis-
sue engineering. Trends Biotechnol. 21, 157, 2003.
23. Boland, T., Xu, T., Damon, B., andCui, X. Application of inkjet
printing to tissue engineering. Biotechnol. J. 1, 910, 2006.
24. Sun, W., Darling, A., Starly, B., and Nam, J. Computer-aided
tissue engineering: overview, scope and challenges. Bio-
technol. Appl. Biochem. 39, 29, 2004.
25. Landers, R., Hubner, U., Schmelzeisen, R., and Mulhaupt, R.
Rapid prototyping of scaffolds derived from thermoreversible
hydrogels and tailored for applications in tissue engineering.
Biomaterials 23, 4437, 2002.
26. Neagu, A., Jakab, K., Jamison, R., and Forgacs, G. Role of
physical mechanisms in biological self-organization. Phys.
Rev. Lett. 95, 178104, 2005.
27. Smith, C.M., Christian, J.J., Warren, W.L., and Williams,
S.K. Characterizing environmental factors that impact the
viability of tissue-engineered constructs fabricated by a
direct-write bioassembly tool. Tissue Eng. 13, 373, 2007.
420 JAKAB ET AL.

28. Smith, C.M., Stone, A.L., Parkhill, R.L., Stewart, R.L., Simp-
kins, M.W., Kachurin, A.M., Warren, W.L., and Williams, S.K.
Three-dimensional bioassembly tool for generating viable tis-
sue-engineered constructs. Tissue Eng. 10, 1566, 2004.
29. Radisic, M., Park, H., Shing, H., Consi, T., Schoen, F.J.,
Langer, R., Freed, L.E., and Vunjak-Novakovic, G. Func-
tional assembly of engineered myocardium by electrical
stimulation of cardiac myocytes cultured on scaffolds. Proc.
Natl. Acad. Sci. USA 101, 18129, 2004.
30. Wessels, A., and Sedmera, D. Developmental anatomy of the
heart: a tale of mice and man. Physiol. Genomics 15, 165,
2003.
31. Forgacs, G., Foty, R.A., Shafrir, Y., and Steinberg, M.S.
Viscoelastic properties of living embryonic tissues: a quan-
titative study. Biophys. J. 74, 2227, 1998.
32. Foty, R.A., and Steinberg, M.S. The differential adhesion
hypothesis: a direct evaluation. Dev. Biol. 278, 255, 2005.
33. Gordon, R., Goel, N.S., Steinberg, M.S., and Wiseman, L.L.
A rheological mechanism sufficient to explain the kinetics of
cell sorting. J. Theor. Biol. 37, 43, 1972.
34. Hegedus, B., Marga, F., Jakab, K., Sharpe-Timms, K.L., and
Forgacs, G. The interplay of cell-cell and cell-matrix inter-
actions in the invasive properties of brain tumors. Biophys.
J. 91, 2708, 2006.
35. Metropolis, N., Rosenbluth, A.W., Rosenbluth, M.N., Teller,
A.H., and Teller, E. Equation of state calculations by fast
computing machines. J. Chem. Phys. 2121, 1087, 1953.
36. Frenkel, J. Viscous flow of crystalline bodies under the action
of surface tension. J. Phys. 9, 351, 1945.
37. Sugi, Y., Yamamura, H., Okagawa, H., and Markwald, R.R.
Bone morphogenetic protein-2 can mediate myocardial reg-
ulation of atrioventricular cushion mesenchymal cell forma-
tion in mice. Dev. Biol. 269, 505, 2004.
38. van den Hoff, M.J., Kruithof, B.P., Moorman, A.F., Mark-
wald, R.R., and Wessels, A. Formation of myocardium after
the initial development of the linear heart tube. Dev. Biol.
240, 61, 2001.
39. van den Hoff, M.J., Moorman, A.F., Ruijter, J.M., Lamers,
W.H., Bennington, R.W., Markwald, R.R., and Wessels, A.
Myocardialization of the cardiac outflow tract. Dev. Biol.
212, 477, 1999.
40. Laschke, M.W., Harder, Y., Amon, M., Martin, I., Farhadi, J.,
Ring, A., Torio-Padron, N., Schramm, R., Rucker, M., Junker,
D., Haufel, J.M., Carvalho, C., Heberer, M., Germann, G.,
Vollmar, B., and Menger, M.D. Angiogenesis in tissue engi-
neering: breathing life into constructed tissue substitutes.
Tissue Eng. 12, 2093, 2006.
41. Kamei, M., Saunders, W.B., Bayless, K.J., Dye, L., Davis,
G.E., and Weinstein, B.M. Endothelial tubes assemble from
intracellular vacuoles in vivo. Nature 442, 453, 2006.
42. Beysens, D.A., Forgacs, G., and Glazier, J.A. Cell sorting is
analogous to phase ordering in fluids. Proc. Natl. Acad. Sci.
USA 97, 9467, 2000.
43. Robinson, E.E., Zazzali, K.M., Corbett, S.A., and Foty, R.A.
Alpha5beta1 integrin mediates strong tissue cohesion. J. Cell
Sci. 116, 377, 2003.
44. Kelm, J.M., Djonov, V., Hoerstrup, S.P., Guenter, C.I., Ittner,
L.M., Greve, F., Hierlemann, A., Sanchez-Bustamante, C.D.,
Perriard, J.C., Ehler, E., and Fussenegger, M. Tissue-
transplant fusion and vascularization of myocardial micro-
tissues and macrotissues implanted into chicken embryos and
rats. Tissue Eng. 12, 2541, 2006.
45. Kelm, J.M., Djonov, V., Ittner, L.M., Fluri, D., Born, W.,
Hoerstrup, S.P., and Fussenegger, M. Design of custom-
shaped vascularized tissues using microtissue spheroids as
minimal building units. Tissue Eng. 12, 2151, 2006.
46. Levenberg, S., Rouwkema, J., Macdonald, M., Garfein, E.S.,
Kohane, D.S., Darland, D.C., Marini, R., van Blitterswijk,
C.A., Mulligan, R.C., D’Amore, P.A., and Langer, R. En-
gineering vascularized skeletal muscle tissue. Nat. Bio-
technol. 23, 879, 2005.
47. Shimizu, T., Sekine, H., Isoi, Y., Yamato, M., Kikuchi, A.,
and Okano, T. Long-term survival and growth of pulsatile
myocardial tissue grafts engineered by the layering of cardi-
omyocyte sheets. Tissue Eng. 12, 499, 2006.
Address reprint requests to:
G. Forgacs, Ph.D.
Departments of Physics, Biology, and Biomedical
Engineering
University of Missouri
Columbia, MO 65211
E-mail: [email protected]
TISSUE ENGINEERING BY PRINTING CELL AGGREGATES 421

Related Documents











