Time Course of Neurotrophic Factor Upregulation and Retinal Protection against Light-Induced Damage after Optic Nerve Section Krisztina Valter, 1 Silvia Bisti, 2 Claudia Gargini, 3 Silvia Di Loreto, 4 Rita Maccarone, 2 Luigi Cervetto, 3 and Jonathan Stone 1 PURPOSE. To assess neurotrophic factor upregulation in the retina after damage to the optic nerve and relate that regulation to changes in photoreceptor stability and function. METHODS. Retinas of adult pigmented (Long-Evans) rats were examined at successive times (1– 60 days) after unilateral optic nerve section. The distribution and expression of ciliary neu- rotrophic factor (CNTF) and basic fibroblast growth factor (FGF-2) and their receptor elements FGFR1 and CNTFR were studied with immunohistochemistry and Western blot analysis. FGF-2 and CNTF mRNA levels were also assessed, with semi- quantitative reverse transcription-PCR. Levels and localization of the intracellular signaling molecule ERK and its activated, phosphorylated form pERK, were examined by immunohisto- chemistry. To assess the correlation between neurotrophic factor levels and their protective effect against light damage, albino (Sprague-Dawley) rats were exposed to bright continu- ous light (1000 lux) for 24 or 48 hours at successive times after nerve section. The TUNEL technique was used to visualize neuronal cell death in the retina. RESULTS. CNTF upregulation was detected 1 week after optic nerve section, peaked at 2 weeks, and fell to control levels at 4 weeks. CNTF appeared first in the inner retina in the gan- glion cells, then in the Mu ¨ller cells in which it became prom- inent at the outer limiting membrane (OLM) and in the outer segment (OS) region of photoreceptors. FGF-2 upregulation became prominent, particularly in photoreceptors, 21 to 28 days after surgery, continued to 2 months, and slowly declined thereafter. Double labeling with antibodies to ligand and the receptor showed colocalization of CNTF to its receptor at the OS region, whereas FGF-2-to-FGFR1 binding was found in the outer nuclear (ONL) and outer plexiform (OPL) layers. Optic nerve section provided a significant protective effect against light-induced damage in the first 2 weeks. There was no pro- tection when animals were exposed to damaging light 1 month after nerve section. CONCLUSIONS. The upregulation of CNTF 7 to 14 days after nerve section correlates with a reduction in the a-wave de- scribed previously. Colocalization of CNTF and CNTFR on the outer segments suggests that CNTF acts at the photoreceptor membrane. The slower upregulation of FGF-2 correlates with a reduction of the b-wave. FGF-2/FGFR1 colocalization in the OPL suggests that this factor acts at the synaptic terminals of photoreceptors, modulating the release of neurotransmitters. The time course of pERK upregulation suggests that the suc- cessive upregulation of CNTF and FGF-2 activates the ERK pathway. Based on the time course of protection against bright continuous light, it seems that CNTF plays a major role in this effect, and FGF-2 has a less important role in the protection against light-induced damage. (Invest Ophthalmol Vis Sci. 2005;46:1748 –1754) DOI:10.1167/iovs.04-0657 D amage to the optic nerve causes retrograde degeneration of retinal ganglion cells, but also induces changes in the cell biology of the retina, including its outer layers. In partic- ular, nerve section increases the resistance of photoreceptors to damaging light, 1 the expression of the trophic factor FGF-2 2,3 in photoreceptor somas and the expression of the cytokine CNTF in Mu ¨ller cells and is followed by a decrease in the amplitude of the a-wave (photoreceptor) and the b-wave (postsynaptic) component of the ERG (Cervetto et al. IOVS 2001;42:Abstract 3396). 3–5 Previously, we 3 presented evi- dence that several of these effects can be accounted for by an upregulation of FGF-2 levels in the retina. The effect of nerve section of the b-wave, for example, is mimicked by intraocular injection of FGF-2, and FGF-2 and its receptor FGFR1 are found in the synaptic terminals of photoreceptors, 6,7 specifically on synaptic vesicles within those terminals. 7 It has been clear for some time, however, that stress regu- lates the expression of several trophic factors and cytokines in the retina 8 and evidence has emerged recently 5 that different components of the ERG are affected at different times after the section of the optic nerve. To test whether different factors have differential effects on retinal function, we tested the working hypothesis that the regulation of CNTF and FGF-2 after nerve damage may differ in time course and that the difference may relate to differences in regulation of the a- and b-wave of the ERG (see Ref. 5). Herein, we present evidence that the time course of upregulation induced by nerve section differs between CNTF and FGF-2 and that the differences cor- relate well with ERG changes reported previously 3,9 in, respec- tively, the a- and b-waves of the ERG. METHODS All experiments were performed in compliance with the Animal Ex- perimentation Ethics Committee guidelines and in accordance with the ARVO Statement on the Use of Animals in Ophthalmic and Vision Research. Experimental animals were adult Long-Evans pigmented rats, born and raised in 12-hour dark–light cyclic light, with the light cycle From the 1 CNS Stability and Degeneration Group, Research School of Biological Sciences, The Australian National University, Can- berra, Australia; 2 Dipartimento di Scienze e Tecnologie Biomediche, Universita ` di L’Aquila, L’Aquila, Italy; 3 Dipartimento di Psichiatria, Neurobiologia, Farmacologia e Biotecnologie, Universita ` di Pisa, Pisa, Italy; and 4 Istituto Tipizzazione Tissutale Consiglio Nazionale delle Ricerche, L’Aquila, Italy. Supported by the Sir Zelman Cowen Universities Fund; the Na- tional Health and Medical Research Council of Australia; the Medical Foundation of the University of Sydney; Retina Australia; Telethon E.746 Italy; and CNR, Agenzia 2000. Submitted for publication June 6, 2004; revised November 9, 2004; accepted November 20, 2004. Disclosure: K. Valter, None; S. Bisti, None; C. Gargini, None; S. Di Loreto, None; R. Maccarone, None; L. Cervetto, None; J. Stone, None The publication costs of this article were defrayed in part by page charge payment. This article must therefore be marked “advertise- ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact. Corresponding author: Jonathan Stone, Research School of Bio- logical Sciences, The Australian National University, Canberra, PO Box 475, Canberra City, ACT 2601, Australia; [email protected]. Investigative Ophthalmology & Visual Science, May 2005, Vol. 46, No. 5 1748 Copyright © Association for Research in Vision and Ophthalmology

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Time Course of Neurotrophic Factor Upregulation andRetinal Protection against Light-Induced Damage afterOptic Nerve Section
Krisztina Valter,1 Silvia Bisti,2 Claudia Gargini,3 Silvia Di Loreto,4 Rita Maccarone,2
Luigi Cervetto,3 and Jonathan Stone1
PURPOSE. To assess neurotrophic factor upregulation in theretina after damage to the optic nerve and relate that regulationto changes in photoreceptor stability and function.
METHODS. Retinas of adult pigmented (Long-Evans) rats wereexamined at successive times (1–60 days) after unilateral opticnerve section. The distribution and expression of ciliary neu-rotrophic factor (CNTF) and basic fibroblast growth factor(FGF-2) and their receptor elements FGFR1 and CNTFR� werestudied with immunohistochemistry and Western blot analysis.FGF-2 and CNTF mRNA levels were also assessed, with semi-quantitative reverse transcription-PCR. Levels and localizationof the intracellular signaling molecule ERK and its activated,phosphorylated form pERK, were examined by immunohisto-chemistry. To assess the correlation between neurotrophicfactor levels and their protective effect against light damage,albino (Sprague-Dawley) rats were exposed to bright continu-ous light (1000 lux) for 24 or 48 hours at successive times afternerve section. The TUNEL technique was used to visualizeneuronal cell death in the retina.
RESULTS. CNTF upregulation was detected 1 week after opticnerve section, peaked at 2 weeks, and fell to control levels at4 weeks. CNTF appeared first in the inner retina in the gan-glion cells, then in the Muller cells in which it became prom-inent at the outer limiting membrane (OLM) and in the outersegment (OS) region of photoreceptors. FGF-2 upregulationbecame prominent, particularly in photoreceptors, 21 to 28days after surgery, continued to 2 months, and slowly declinedthereafter. Double labeling with antibodies to ligand and thereceptor showed colocalization of CNTF to its receptor at theOS region, whereas FGF-2-to-FGFR1 binding was found in theouter nuclear (ONL) and outer plexiform (OPL) layers. Opticnerve section provided a significant protective effect againstlight-induced damage in the first 2 weeks. There was no pro-
tection when animals were exposed to damaging light 1 monthafter nerve section.
CONCLUSIONS. The upregulation of CNTF 7 to 14 days afternerve section correlates with a reduction in the a-wave de-scribed previously. Colocalization of CNTF and CNTFR� on theouter segments suggests that CNTF acts at the photoreceptormembrane. The slower upregulation of FGF-2 correlates with areduction of the b-wave. FGF-2/FGFR1 colocalization in theOPL suggests that this factor acts at the synaptic terminals ofphotoreceptors, modulating the release of neurotransmitters.The time course of pERK upregulation suggests that the suc-cessive upregulation of CNTF and FGF-2 activates the ERKpathway. Based on the time course of protection against brightcontinuous light, it seems that CNTF plays a major role in thiseffect, and FGF-2 has a less important role in the protectionagainst light-induced damage. (Invest Ophthalmol Vis Sci.2005;46:1748–1754) DOI:10.1167/iovs.04-0657
Damage to the optic nerve causes retrograde degenerationof retinal ganglion cells, but also induces changes in the
cell biology of the retina, including its outer layers. In partic-ular, nerve section increases the resistance of photoreceptorsto damaging light,1 the expression of the trophic factorFGF-22,3 in photoreceptor somas and the expression of thecytokine CNTF in Muller cells and is followed by a decrease inthe amplitude of the a-wave (photoreceptor) and the b-wave(postsynaptic) component of the ERG (Cervetto et al. IOVS2001;42:Abstract 3396).3–5 Previously, we3 presented evi-dence that several of these effects can be accounted for by anupregulation of FGF-2 levels in the retina. The effect of nervesection of the b-wave, for example, is mimicked by intraocularinjection of FGF-2, and FGF-2 and its receptor FGFR1 are foundin the synaptic terminals of photoreceptors,6,7 specifically onsynaptic vesicles within those terminals.7
It has been clear for some time, however, that stress regu-lates the expression of several trophic factors and cytokines inthe retina8 and evidence has emerged recently5 that differentcomponents of the ERG are affected at different times after thesection of the optic nerve. To test whether different factorshave differential effects on retinal function, we tested theworking hypothesis that the regulation of CNTF and FGF-2after nerve damage may differ in time course and that thedifference may relate to differences in regulation of the a- andb-wave of the ERG (see Ref. 5). Herein, we present evidencethat the time course of upregulation induced by nerve sectiondiffers between CNTF and FGF-2 and that the differences cor-relate well with ERG changes reported previously3,9 in, respec-tively, the a- and b-waves of the ERG.
METHODS
All experiments were performed in compliance with the Animal Ex-perimentation Ethics Committee guidelines and in accordance with theARVO Statement on the Use of Animals in Ophthalmic and VisionResearch. Experimental animals were adult Long-Evans pigmented rats,born and raised in 12-hour dark–light cyclic light, with the light cycle
From the 1CNS Stability and Degeneration Group, ResearchSchool of Biological Sciences, The Australian National University, Can-berra, Australia; 2Dipartimento di Scienze e Tecnologie Biomediche,Universita di L’Aquila, L’Aquila, Italy; 3Dipartimento di Psichiatria,Neurobiologia, Farmacologia e Biotecnologie, Universita di Pisa, Pisa,Italy; and 4Istituto Tipizzazione Tissutale Consiglio Nazionale delleRicerche, L’Aquila, Italy.
Supported by the Sir Zelman Cowen Universities Fund; the Na-tional Health and Medical Research Council of Australia; the MedicalFoundation of the University of Sydney; Retina Australia; TelethonE.746 Italy; and CNR, Agenzia 2000.
Submitted for publication June 6, 2004; revised November 9,2004; accepted November 20, 2004.
Disclosure: K. Valter, None; S. Bisti, None; C. Gargini, None; S.Di Loreto, None; R. Maccarone, None; L. Cervetto, None; J. Stone,None
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be marked “advertise-ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Corresponding author: Jonathan Stone, Research School of Bio-logical Sciences, The Australian National University, Canberra, PO Box475, Canberra City, ACT 2601, Australia; [email protected].
Investigative Ophthalmology & Visual Science, May 2005, Vol. 46, No. 51748 Copyright © Association for Research in Vision and Ophthalmology

set at 200 lux. In experiments involving light-induced damage, animalswere adult Sprague–Dawley albino rats, born and raised in 12-hourdark–light cyclic light set at 5 to 10 lux.
Unilateral Optic Nerve Section
Nerve section was performed in rats under surgical anesthesia byintraperitoneal injection of either tribromoethanol (270 mg/100 g,Avertin; Sigma-Aldrich Chemie GmbH, Deisenhofen, Germany) or amixture of ketamine (100 mg/kg) and xylazine (12 mg/kg). The surgi-cal approach to the optic nerve has been described.3 In brief, the skinwas incised at the superior rim of the orbit, and the orbital tissuebehind the eye was blunt dissected to visualize the optic nerve. Thenerve was cut several millimeters behind the eyeball, to avoid damag-ing vessels entering or leaving the eye. The skin was closed withsutures, and an antiseptic/antibiotic powder applied to the wound. Atvarious times after surgery, animals were euthanatized and eyes werecollected for processing.
Light-Induced Damage
Animals were placed in individual transparent Plexiglas cages, withfood available on the floor and water provided from clear plasticbottles, both ad libitum. A light source placed above the cages gener-ated an intensity of 1000 lux at eye level. Animals were dark adaptedovernight and then exposed to the bright light continuously for 48hours. Exposure started at the same time of day for all animals (9 AM).The rats were euthanatized at the end of the exposure, and eyes wereenucleated and immersion fixed in 4% paraformaldehyde for furtherprocessing.
Immunohistochemistry
Immediately after euthanasia, eyes were enucleated and immersionfixed in 4% paraformaldehyde for 1 hour, washed in 0.1 M phosphate-buffered saline (PBS, pH 7.4), and cryoprotected by immersion in 15%sucrose overnight. Eyes were embedded in optimal cutting tempera-ture (OCT) compound (Tissue Tek; Sakura Fintek, Torrance, CA), snapfrozen in liquid nitrogen–isopentane and cryosectioned at 20 �m. Eacheye was oriented so that sections ran from the superior to the inferioredge. Sections were collected on gelatin- and poly-L-lysine-coatedslides. They were then immunolabeled for the following proteins:CNTF (Chemicon, Temecula, CA) 1:200, FGF-2 (Upstate Biotechnol-ogy, Lake Placid, NY) 1:200, CNTFR� (RDI, Flanders, NJ) 1:50, extra-cellular signal-regulated kinase (ERK) 1:50, phosphorylated ERK(pERK) 1:100 (Cell Signaling Technology, Beverly, MA), and FGFR11:100 (Santa Cruz Biotechnology Inc., Santa Cruz, CA). For all labeling,washes were performed three times for 5 minutes each in 0.1 M PBS atroom temperature. To block nonspecific binding, 10% normal horseserum was used. Sections were incubated with antibodies at 37°C for70 minutes. Antibodies against CNTF, FGFR1, ERK, and pERK wererabbit polyclonals, the antibody against CNTFR� was a goat anti-ratpolyclonal, and the antibody against FGF-2 was mouse monoclonal.
Secondary antibodies were goat anti-mouse, goat anti-rabbit, ordonkey anti-goat IgG conjugated to Alexa Fluor 488 (green) or 594(red) diluted 1:200 in PBS (Molecular Probes, Eugene, OR) and incu-bated at 37°C for 1 to 2 hours, followed by three 5-minute washes inPBS, and counterstaining with DNA-specific label, bisbenzamide (Cal-biochem, La Jolla, CA) 1:10,000 for 1 minute at room temperature(RT). Images were taken by laser scanning microscope (Leica, Deer-field, IL) and analyzed with NIH image (ImageJ 1.14c for Linux plat-form; available by ftp at zippy.nimh.nih.gov/ or at http://rsb.info.nih.gov/nih-image; developed by Wayne Rasband, National Institutes ofHealth, Bethesda, MD). Measurements of FGF-2 labeling were madeacross the full thickness of the ONL in the midperipheral retina (ap-proximately half-way between the disc and the superior edge). Mea-surements of CNTF-labeling were made over an area covering thethickness of the INL, to include the most strongly CNTF-labeled ele-ments in the retina, the somas of Muller cells.
TUNEL Labeling
To label apoptotic cells, we used the TUNEL technique, according toearlier published methods.10
Western Blot Analysis
Isolated rat retinas were washed in Western blot buffer (50 mMTris-HCl [pH7.4], 50 mM NaCl, and 5 mM EDTA) and homogenized ina lysis buffer (0.2% Triton X-100, protease inhibitor [Sigma-AldrichChemie GmbH], and 5% DNase diluted 1:100 in wash buffer). Tworetinas were pooled per sample, and homogenates were incubated inthe lysis buffer for 30 minutes at RT. After incubation, samples werecentrifuged at 15,000g for 20 minutes at 4°C. The resultant supernatantwas collected, and protein content was measured (Protein Assay;Bio-Rad, Munich, Germany). Supernatant was diluted in an equal vol-ume of 2� Laemmli buffer (Sigma-Aldrich Chemie GmbH), boiled for5 minutes and stored at �20°C until all samples were collected.
Protein of each sample (100 �g) was electrophoresed on a 15%SDS-polyacrylamide gel. Proteins were transferred to a nitrocellulosemembrane using transfer buffer (25 mM Tris-HCl [pH 8.3], 192 mMglycine, and 20% methanol). The protein blot was then blocked with3% nonfat dried milk and 0.05% Tween 20 in blot buffer (20 mM TrisHCl [pH 8.00], 500 mM NaCl) at RT for 45 minutes. The membranethen was incubated overnight at 4°C with the rabbit anti-rat CNTFantibody (Chemicon), or the anti-FGF-2 clone bFM-2 mouse monoclo-nal IgG (Upstate Biotechnology) diluted 1:1000, or the anti-pERKmouse monoclonal (Santa Cruz Biotechnology Inc.) diluted 1:200 inthe blocking buffer. The reactions were visualized with horse-radishperoxidase–conjugated secondary antibodies (anti-rabbit or anti-mouse IgG) for 2 hours at RT. Bands were visualized and quantifiedusing a chemiluminescence method (ECL Western blot system; Amer-sham Pharmacia Biotech, Freiburg, Germany).
Reverse Transcription–PolymeraseChain Reaction
Ten samples for each series were collected and total RNA was ex-tracted (TRIzol reagent; Invitrogen-Life Technologies, Scotland, UK)according to the manufacturer’s directions. Total RNA (1 �g) wasreverse transcribed (RETROscript; Ambion, Cambridgeshire, UK) withrandom decamers used as first-strand primers. The cDNA (2 �L) wasused as the template for amplification in a 20-�L PCR reaction. PCRwas performed in duplicate and in multiplex reactions, using 18S asthebinternal standard (QuantumRNA Classic 18S; Ambion), producinga 489-bp PCR product. The 18S rRNA internal control is used incombination with 18S competimers and a gene-specific PCR primerpair. Competimer technology was used to modulate the amplificationefficiency of a PCR template, and the correct ratio between the 18SrRNA primers and 18S competimers was empirically determined. Thesequences of the FGF-2-specific primer pair were: forward, GGA GTGTGT GCT AAC CGT TAC CTG GCT ATG; reverse, TCA GCT CTT AGCAGA CAT TGG AAG AAA AAG—and produced an amplification prod-uct of 242 bp. The product of the CNTF primers was 1050 bp and thesequences of the primers were: forward, CAC ATT TCT TAT TTG GAC;reverse, ACA ACA ATA CTC TTT TTA TTC. The number of cycles wasdetermined empirically by sampling FGF-2 and CNTF amplicons be-tween 22 and 40 cycles and selecting the approximate midpoint of thelinear amplifications (35 cycles). PCR thermal cycling for FGF-2 mRNAamplification and classic 18S was performed under the following con-ditions: initial denaturation for 10 minutes at 94°C; followed by 35cycles of 40 seconds at 94°C, 40 seconds at 65°C, and 1 minute at 72°C;and a final extension of 5 minutes at 72°C. The multiplex amplificationparameters for CNTF and classic II 18S were: 2 minutes and 30 secondsat 94°C (initial denaturation), 1 minute at 94°C, 1 minute at 50°C, 1minute at 72°C for 35 cycles and a final extension of 5 minutes at 72°C.PCR products were separated on a 2% agarose gel stained withethidium bromide, and the relative densities of the PCR fragmentswere determined and normalized using software for semiquantitativedensitometric analysis (Total Laboratory, Phoretix, Newcastle-upon-Tyne, UK). Results are given as relative units (RU).
IOVS, May 2005, Vol. 46, No. 5 Neurotrophic Factor Upregulation after Optic Nerve Damage 1749
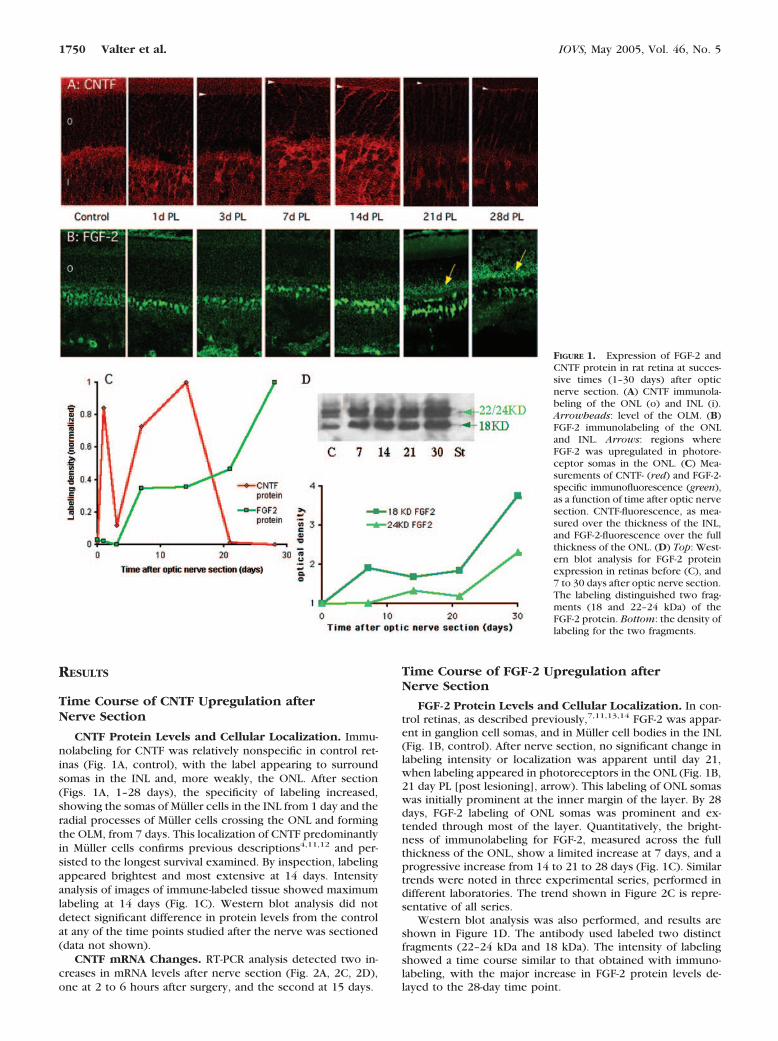
RESULTS
Time Course of CNTF Upregulation afterNerve Section
CNTF Protein Levels and Cellular Localization. Immu-nolabeling for CNTF was relatively nonspecific in control ret-inas (Fig. 1A, control), with the label appearing to surroundsomas in the INL and, more weakly, the ONL. After section(Figs. 1A, 1–28 days), the specificity of labeling increased,showing the somas of Muller cells in the INL from 1 day and theradial processes of Muller cells crossing the ONL and formingthe OLM, from 7 days. This localization of CNTF predominantlyin Muller cells confirms previous descriptions4,11,12 and per-sisted to the longest survival examined. By inspection, labelingappeared brightest and most extensive at 14 days. Intensityanalysis of images of immune-labeled tissue showed maximumlabeling at 14 days (Fig. 1C). Western blot analysis did notdetect significant difference in protein levels from the controlat any of the time points studied after the nerve was sectioned(data not shown).
CNTF mRNA Changes. RT-PCR analysis detected two in-creases in mRNA levels after nerve section (Fig. 2A, 2C, 2D),one at 2 to 6 hours after surgery, and the second at 15 days.
Time Course of FGF-2 Upregulation afterNerve Section
FGF-2 Protein Levels and Cellular Localization. In con-trol retinas, as described previously,7,11,13,14 FGF-2 was appar-ent in ganglion cell somas, and in Muller cell bodies in the INL(Fig. 1B, control). After nerve section, no significant change inlabeling intensity or localization was apparent until day 21,when labeling appeared in photoreceptors in the ONL (Fig. 1B,21 day PL [post lesioning], arrow). This labeling of ONL somaswas initially prominent at the inner margin of the layer. By 28days, FGF-2 labeling of ONL somas was prominent and ex-tended through most of the layer. Quantitatively, the bright-ness of immunolabeling for FGF-2, measured across the fullthickness of the ONL, show a limited increase at 7 days, and aprogressive increase from 14 to 21 to 28 days (Fig. 1C). Similartrends were noted in three experimental series, performed indifferent laboratories. The trend shown in Figure 2C is repre-sentative of all series.
Western blot analysis was also performed, and results areshown in Figure 1D. The antibody used labeled two distinctfragments (22–24 kDa and 18 kDa). The intensity of labelingshowed a time course similar to that obtained with immuno-labeling, with the major increase in FGF-2 protein levels de-layed to the 28-day time point.
FIGURE 1. Expression of FGF-2 andCNTF protein in rat retina at succes-sive times (1–30 days) after opticnerve section. (A) CNTF immunola-beling of the ONL (o) and INL (i).Arrowheads: level of the OLM. (B)FGF-2 immunolabeling of the ONLand INL. Arrows: regions whereFGF-2 was upregulated in photore-ceptor somas in the ONL. (C) Mea-surements of CNTF- (red) and FGF-2-specific immunofluorescence (green),as a function of time after optic nervesection. CNTF-fluorescence, as mea-sured over the thickness of the INL,and FGF-2-fluorescence over the fullthickness of the ONL. (D) Top: West-ern blot analysis for FGF-2 proteinexpression in retinas before (C), and7 to 30 days after optic nerve section.The labeling distinguished two frag-ments (18 and 22–24 kDa) of theFGF-2 protein. Bottom: the density oflabeling for the two fragments.
1750 Valter et al. IOVS, May 2005, Vol. 46, No. 5

FGF-2 mRNA Changes. RT-PCR analysis detected an earlyincrease in FGF-2 mRNA levels 2 to 6 hours after optic nervesection. After this time, mRNA levels returned to controllevels, increased to 4 days, and then returned to controllevels (Figs. 2B, 2C, 2D). The persistence of high FGF-2protein levels after the transient upregulation of FGF-2mRNA suggests that the protein is stabilized, by a still un-known mechanism.
Site of Action of Neurotrophic Factors:Receptor Localization
CNTFR�. In other tissues, CNTF has been shown to actthrough a tripartite receptor complex. CNTF binds to the �component of the complex, but that component does not havea cytoplasmic domain and plays no role in the signalingprocess.15,16 The binding of the CNTF to its specific �subunit initiates the formation of a complex, by the recruit-ment of the � components LIF� and gp130 and the forma-tion of a heterodimer of these two components, which theninitiates the signaling process by activating either cytoplas-mic tyrosine kinases,15 or ERK members of the MAP-kinasecascade.17
As previously described,12 we found punctate labeling ofCNTFR� along Muller cell processes across the retina, but themost prominent labeling was on the photoreceptor outer seg-ments (Figs. 3A–C, green). This localization of CNTFR� onphotoreceptors18 or more specifically on its outer segments19
has been confirmed. At 7 days and most clearly at 14 days,CNTF labeling (Figs. 3A–C, red) was upregulated, as evidencedby the prominent yellow signal of the outer segment labeling.The yellow signal suggests colocalization of the ligand andreceptor, as demonstrated previously.12
FGFR1. In control retinas, FGFR1-labeling (Figs. 3D–F, red)was prominent in somas in the ONL, where it was moreintense near the OPL.7 By 21 days after nerve section, theexpression of FGF-2 in the ONL increased, particularly in thecytoplasm of ONL somas. Yellow staining of these somas wasprominent at 1 and 2 months after nerve section, specifically in
the ONL (Figs. 3E, 3F). Again, the yellow staining suggestscolocalization of the ligand and receptor, as demonstratedpreviously.7 Figures 3G and 3H show ONL somas labeled forboth FGF-2 and FGFR1, in control material (Fig. 3G) and 1month after nerve section (Fig. 3H). Again, yellow staining,indicative of ligand/receptor colocalization, is more prominentafter nerve section (bottom panels Figs. 3G, 3H).
Mode of Action of Neurotrophic Factors in theRetina: Intracellular Signaling Using the ERKPathway after Optic Nerve Section
Cytokines act through the Jak-STAT pathway, but an increasingbody of evidence is showing that the MAPK pathway is alsolikely to be involved in mediating their effects.20 Growth fac-tors act by phosphorylation and activation of the ERK family,also known as MAP kinases (MAPKs).20–23 Activation of thesekinases involves their translocation to the nucleus and phos-phorylation; the phosphorylated form then activates transcrip-tion factors. Since the MAPK pathway may be involved in bothcytokine and growth factor signaling, we investigated changesin levels of the phosphorylated (active) form of ERK (pERK)induced by optic nerve section, at key times (7, 14, 21, and 28days) after lesioning, in six experimental series, three exam-ined by immunohistochemistry (Figs. 4A–E), and three byWestern blot analysis (Figs. 4F, 4G).
All series showed a bimodal upregulation of pERK over theperiod examined (0–30-day after lesioning). In control retinas,immunolabeling for pERK was not prominent in any retinallayer (Fig. 4A). One week after nerve section, pERK labelingwas strongly present in Muller cell bodies in the INL, and alongthe full length of their processes from the OLM to the ILM,where astrocytes were also labeled. Horizontally oriented pro-cesses were also labeled in the OPL and IPL. In the IPL theyappeared to mark the sublaminae of the layer (Fig. 4B). Label-ing was reduced 14 days after nerve section (Fig. 4C) and wasupregulated again at 21 days (Fig. 4D). One month after opticnerve section the intensity of labeling was reduced again, beingprominent in only some Muller cell bodies (Fig. 4E).
Western blot analysis showed increases in pERK proteinlevels at 7 and 30 days after nerve section (Figs. 4F, 4G). Thetwo forms of analysis are thus consistent in showing a bimodalupregulation of pERK, with an early peak at 7 days afterlesioning and a sharp reduction at 14 days. The time of thesecond peak was at 21 day in the immunohistochemical anal-ysis (Fig. 4E) but at 30 days or later in the Western blot analysis.
Correlation with Light Damage
Exploration of the effects of nerve section on trophic factorexpression in the retina began with the observation1 that nervesection is followed by a period of photoreceptor resistance tolight-induced damage. To confirm this observation and defineits time course we exposed nerve-sectioned rats to brightcontinuous light (BCL; 1000 lux for 48 hours) at successiveintervals after nerve section and assessed the photoreceptordeath induced, using the TUNEL technique. Four animals wereexamined at each survival time (0, 7, 14, and 30 days). Aspreviously,14 BCL induced the DNA fragmentation characteris-tic of dying cells almost exclusively in the photoreceptor layer(Fig. 5A). The rate of photoreceptor death induced by BCL,expressed as the frequency of TUNEL� profiles/mm of retina,decreased with time to a minimum (� 50% of control) at 7 daysafter lesioning (Figs. 5A, 5B) and recovered monotonicallythereafter (Fig. 5B). The minimum at 7 days after lesioningcorresponds to a rise in the expression of CNTF. We observedno minimum in BCL-induced death that corresponded with thepeak upregulation of FGF-2, at �30 days after lesioning.
FIGURE 2. Result of an RT-PCR interrogation for 1092-bp sequence ofCNTF mRNA and 242-bp sequence of FGF-2 mRNA. (A, B) Subscriptsindicate negative control (c�) and positive control (c�) lanes, andmRNA from control rats (C) and from rats examined at 1 to 48 hours,and 4 to 30 days after lesioning. (C, D) Density of labeling in the gelsin (A) and (B) on two different time scales (hours, days).
IOVS, May 2005, Vol. 46, No. 5 Neurotrophic Factor Upregulation after Optic Nerve Damage 1751

DISCUSSION
The present study documents the impact of damage to theoptic nerve on the cell biology of the retina, in particular onthe expression of neuroprotective proteins, specifically thegrowth factor FGF-2 and the cytokine CNTF. The retina reactsto nerve damage by upregulating these proteins, whose bind-ing with their receptors increases, activating intracellular ERKpathways and protecting photoreceptors against light stress.
Trophic Factors and Retinal Function
We have recently described a strong correlation between ERGresponse amplitude and trophic factor expression3,5 in the ratretina. A detailed analysis of the a-wave after optic nervesection5 suggested a reduced dark current in the first weekafter nerve section, temporally related to an upregulation ofCNTF. Although the molecular events leading to a reduced ioncurrent have to be clarified, they may be triggered by thebinding of CNTF ligand to its receptor CNTFR� on the photo-receptor outer segment. The reduced amplitude of the b-waveis related to the a-wave reduction for up to 21 days aftersection, and subsequently seems to be due to postsynapticevents triggered by FGF-2 upregulation.3,5
Value of the Nerve Section Model of Retinal Stress
Continued attention to this model seems warranted for tworeasons. First, the retina’s reaction to damage to the opticnerve resembles in many ways its reaction to light-induceddamage,14,24 mechanical injury,25,26 or hyperoxic injury.27 Inthe nerve section model, however, the photoreceptors are freeof damage,28,29 which is restricted to the retrograde degener-ation of ganglion cells, located in the inner retina. In otherstress models (light-induced damage, hypoxic or hyperoxicdamage, mechanical damage), by contrast, the photoreceptorsare directly affected. In the nerve section model uniquely, thephotoreceptors are protected but undamaged.1–3 Second, thepresent results show that the upregulation of CNTF and FGF-2induced by nerve section follow distinct time courses. Thenerve section model thus presents an opportunity to test pho-toreceptor stability when CNTF is upregulated and FGF-2 is not(e.g., at day 7 after lesioning) and vice versa (day 28).
Protection against Light Damage:Is FGF-2 Important?
The present results confirm the original report of Bush andWilliams1 that nerve section induces an increase in photore-ceptor resistance to damaging light, and the report of Kostyk etal.2 that nerve section induces an upregulation of FGF-2 inphotoreceptors. When we traced the time course of the in-crease in photoreceptor resistance, however, the result wasunexpected: Resistance correlated with the upregulation ofCNTF and not of FGF-2.
Many studies have reported a correlation between upregu-lation of FGF-2 in the retina/photoreceptors and photoreceptorresistance to stress. In most, however, FGF-2 upregulation wasinduced by injury to the retina (needlestick,26 laser,13,30 pre-conditioning light challenge31), which upregulated a range ofpotentially protective factors (reviewed in Ref. 32). Consider-FIGURE 3. Effect of optic nerve section on ligand and receptor bind-
ing of CNTF and FGF-2. (A–C) CNTF and CNTFR� on the outersegments (os). In control retinas, CNTFR� labeling (green) was dom-inant. At 7 days, and more prominently at 14 days, CNTF-labeling (red)appeared and colocalized with CNTFR� labeling (yellow). o, nuclei ofphotoreceptors (blue) in the ONL. (D–F) FGF-2 and FGFR1 labeling ofthe retina. o, ONL; i, INL; g, ganglion cell layer. In control retina (D),FGF-2 was prominent in Muller cell somas in the INL and in ganglioncell somas. FGFR1 labeling (red) was prominent in the ONL and inganglion cell somas whose cytoplasm was labeled for both FGF-2 andFGFR1 (yellow). At 30 days (1 month) and at 60 days (2 months) FGF-2(green) was upregulated in the ONL. The FGF-2 labeling colocalizedwith FGFR1 labeling, in the ONL (yellow). In each of (D–F), the red
signal has been removed over a central portion of the image, to showthe labeling for FGF-2. The scale in (F) refers to (D, E). (G, H) Top in(G): FGF-2 (green) and FGFR1 labeling (red) of the OPL (opl) andsurrounding layers. Bottom: only the yellow pixels in the top panel,where red and green pixels colocalized. Colocalization was evident insomas in both the ONL and INL, but not in the OPL. At 30 days (H, 1mo) after nerve section, colocalization was more prominent in somasof both layers, and was also detected in the OPL (arrows). The scale in(H) refers to (A, B, C, G).
1752 Valter et al. IOVS, May 2005, Vol. 46, No. 5

ing the detail of those studies, those that give evidence of anFGF-2-specific protection of photoreceptors against damaginglight are LaVail et al.33 and Faktorovich et al.25 They usedsubretinal injection of FGF-2, and control experiments (subreti-nal needle insertions) showed that much of the effect resultedfrom the impact of the injection. LaVail et al.33 used intravitrealinjection of FGF-2, which arguably involves less retinal damageand still showed significant FGF-2-related photoreceptor rescuefrom light damage. One other FGF-2-specific study deservesnote: Campochiaro et al.34 showed that dominant-negativecompetition for FGF-2 in transgenic mice caused photorecep-tor degeneration, indicating an important role for FGF-2 inphotoreceptor stability. They did not, however, test whetherdominant negative competition for FGF-2 affects vulnerabilityto light-induced damage.
The present evidence seems to be the first to question therelationship between FGF-2 and the resistance of photorecep-tors to damaging light. The present data do not howeverquestion the role of FGF-2 in photoreceptor stability in normallighting conditions, or in the face of other forms of stress, suchas hyperoxia, hypoxia, or genetically induced. We are cur-rently using the nerve section model to assess the roles ofCNTF and FGF-2 in these forms of stress.
Correlation with Light-Induced Damage:A Particular Role for CNTF?
Conversely, the present results suggest a particular role forCNTF in the protection of photoreceptors against damage bylight. Many previous studies have shown a general correlationbetween CNTF upregulation and photoreceptor resistance todamage, and one study,33 using intravitreal injection of CNTF,has shown a CNTF-specific effect. Recent studies of the retinaldistribution of CNTFR� (the element common to the family ofCNTF receptors; reviewed in Refs. 16,35) have shown thatCNTFR� is prominent on the outer segments of photorecep-tors12,19 and that CNTF/CNTFR� colocalization occurs onouter segments and is upregulated in light-stressed retina.12
The available evidence seems to suggest, therefore, that CNTFplays a major role in increasing photoreceptor resistance tolight-induced damage, perhaps by acting at the outer segmentlevel. The present study did not monitor factors such as BDNF
FIGURE 4. Expression of pERK inthe retina after optic nerve section.(A–E) Immunolabeling for pERKacross the rat retina. Labeling waslow in a control retina (A), and be-came prominent in the IPL and OPLand in the somas and radial processesof Muller cells at 7 days and again at21 day after lesioning. o, ONL; i, INL;ipl, IPL. (F) Western blot for pERKprotein. Bands for 42- and 44-kDafragments of pERK were prominentat 7 days and again at 30 days afternerve section. (G) Density of 42- and44-kDa bands in Western blot analy-sis (F) for up to 30 days afterlesioning.
FIGURE 5. Changes in photoreceptor vulnerability to damaging lightafter optic nerve section. (A) Sections of retina from rats exposed tobright light for an extended period (BCL; 1000 lux for 48 hours). Blue:nuclear DNA of cells in the INL (i) and ONL (o), labeled with bisben-zamide. Red: fragmentation of DNA induced by BCL, labeled with theTUNEL technique. Labeling was intense in control animals (0 days’exposure) and again at 30 days. TUNEL labeling was consistentlyreduced (i.e., photoreceptor vulnerability was reduced) at 7 days. DNAfragmentation was specific to the ONL and therefore to photorecep-tors. (B) The data in black show BCL-induced TUNEL labeling of theONL, expressed as TUNEL� profiles/mm and averaged over the fulllength of retinal sections, for successive times after nerve section.Exposure to BCL was for 48 hours. Vulnerability was minimal at 7 daysand significantly (P � 0.0001 on a 2-tailed t-test) lower than in controlsubjects. By 30 days, vulnerability had returned toward control levels(P � 0.2, by two-tailed t-test). The red and green curves show the timecourse of expression of FGF-2 and CNTF over the same period.
IOVS, May 2005, Vol. 46, No. 5 Neurotrophic Factor Upregulation after Optic Nerve Damage 1753

and IL-1�, which have also been shown to protect photorecep-tors from light-induced damage when injected intravitreally.33
The reduction of retinal responsiveness associated previ-ously with CNTF upregulation9,36 suggests, however, that theprotection does not extend to photoreceptor performance,and our own observations in the nerve-sectioned rat5 confirmthis suggestion. In two strains of mouse in which the retinadegenerates, the rd and rds strains, CNTF upregulation hasreported to be associated with an improvement in theERG.37,38 A subsequent study of one of these strains36 re-ported, however, that upregulation of CNTF expression usingan AAV-mediated gene delivery system negated the effects ofgene therapy on retinal function, as assessed by the ERG.Further, the same upregulation of CNTF expression in wild-type mice resulted in a marked reduction of the ERG.
In summary, the present results suggest that CNTF upregu-lation plays a major role in the protection of photoreceptorsand reduction of the ERG which follow damage to the opticnerve. The linkage between increased resistance of photore-ceptors to damage and the loss of their responsiveness to light,first reported by Gargini et al.,3 deserves further investigation.
References
1. Bush RA, Williams TP. The effect of unilateral optic nerve sectionon retinal light damage in rats. Exp Eye Res. 1991;52:139–153.
2. Kostyk SK, D’Amore PA, Herman IM, Wagner JA. Optic nerveinjury alters basic fibroblast growth factor localization in the retinaand optic tract. J Neurosci. 1994;14:1441–1449.
3. Gargini C, Belfiore MS, Bisti S, Cervetto L, Valter K, Stone J. Theimpact of basic fibroblast growth factor on photoreceptor functionand morphology. Invest Ophthalmol Vis Sci. 1999;40:2088–2099.
4. Chun MH, Ju WK, Kim KY, et al. Upregulation of ciliary neurotro-phic factor in reactive Muller cells in the rat retina following opticnerve transection. Brain Res. 2000;868:358–362.
5. Gargini C, Bisti S, Demontis G, Valter K, Stone J, Cervetto L.Electroretinogram changes associated with retinal upregulation oftrophic factors: observations following optic nerve section. Neu-roscience. 2004;126:775–783.
6. Morimoto A, Matsuda S, Uryu K, Fujita H, Okumura N, Sakanaka M.Light- and electron-microscopic localization of basic fibroblast growthfactor in adult rat retina. Okajimas Folia Anat Jpn. 1993;70:7–12.
7. Valter K, van Driel D, Bisti S, Stone J. FGFR1 expression andFGFR1-FGF-2 colocalisation in rat retina: sites of FGF-2 action onrat photoreceptors. Growth Factors. 2002;20:177–188.
8. Steinberg RH. Survival factors in retinal degenerations. Curr OpinNeurobiol. 1994;4:515–524.
9. Casson RJ, Chidlow G, Wood JP, Vidal-Sanz M, Osborne NN. Theeffect of retinal ganglion cell injury on light-induced photorecep-tor degeneration. Invest Ophthalmol Vis Sci. 2004;45:685–693.
10. Maslim J, Valter K, Egensperger R, Hollander H, Stone J. Tissueoxygen during a critical developmental period controls the deathand survival of photoreceptors. Invest Ophthalmol Vis Sci. 1997;38:1667–1677.
11. Walsh N, Valter K, Stone J. Cellular and subcellular patterns ofexpression of bFGF and CNTF in the normal and light stressedadult rat retina. Exp Eye Res. 2001;72:495–501.
12. Valter K, Bisti S, Stone J. Location of CNTFRalpha on outersegments: evidence of the site of action of CNTF in rat retina.Brain Res. 2003;985:169–175.
13. Xiao M, Sastry SM, Li ZY, et al. Effects of retinal laser photocoag-ulation on photoreceptor basic fibroblast growth factor and sur-vival. Invest Ophthalmol Vis Sci. 1998;39:618–630.
14. Bowers F, Valter K, Chan S, Walsh N, Maslim J, Stone J. Effects ofoxygen and bFGF on the vulnerability of photoreceptors to lightdamage. Invest Ophthalmol Vis Sci. 2001;42:804–815.
15. Inoue M, Nakayama C, Noguchi H. Activating mechanism of CNTFand related cytokines. Mol Neurobiol. 1996;12:195–209.
16. Hirano T, Nakajima K, Hibi M. Signaling mechanisms throughgp130: a model of the cytokine system. Cytokine Growth FactorRev. 1997;8:241–252.
17. Boulton TG, Stahl N, Yancopoulos GD. Ciliary neurotrophic fac-tor/leukemia inhibitory factor/interleukin 6/oncostatin M family of
cytokines induces tyrosine phosphorylation of a common set ofproteins overlapping those induced by other cytokines andgrowth factors. J Biol Chem. 1994;269:11648–11655.
18. Beltran WA, Zhang Q, Kijas JW, et al. Cloning, mapping, and retinalexpression of the canine ciliary neurotrophic factor receptor alpha(CNTFRalpha). Invest Ophthalmol Vis Sci. 2003;44:3642–3649.
19. Rhee KD, Yang XJ. Expression of cytokine signal transductioncomponents in the postnatal mouse retina. Mol Vis. 2003;9:715–722.
20. Peterson WM, Wang Q, Tzekova R, Wiegand SJ. Ciliary neurotro-phic factor and stress stimuli activate the Jak-STAT pathway inretinal neurons and glia. J Neurosci. 2000;20:4081–4090.
21. Blanquet PR, Jonet L. Signal-regulated proteins and fibroblastgrowth factor receptors: comparative immunolocalization in ratretina. Neurosci Lett. 1996;214:135–138.
22. van Puijenbroek AA, van der Saag PT, Coffer PJ. Cytokine signaltransduction in P19 embryonal carcinoma cells: regulation of Stat3-mediated transactivation occurs independently of p21ras-Erk sig-naling. Exp Cell Res. 1999;251:465–476.
23. Sheu JY, Kulhanek DJ, Eckenstein FP. Differential patterns of ERKand STAT3 phosphorylation after sciatic nerve transection in therat. Exp Neurol. 2000;166:392–402.
24. Gao H, Hollyfield JG. Basic fibroblast growth factor: increased geneexpression in inherited and light-induced photoreceptor degener-ation. Exp Eye Res. 1996;62:181–189.
25. Faktorovich EG, Steinberg RH, Yasumura D, Matthes MT, LaVail MM.Basic fibroblast growth factor and local injury protect photoreceptorsfrom light damage in the rat. J Neurosci. 1992;12:3554–3567.
26. Wen R, Song Y, Cheng T, et al. Injury-induced upregulation ofbFGF and CNTF mRNAs in the rat retina. J Neurosci. 1995;15:7377–7385.
27. Yamada H, Yamada E, Ando A, et al. Fibroblast growth factor-2decreases hyperoxia-induced photoreceptor cell death in mice.Am J Pathol. 2001;159:1113–1120.
28. Schremser JL, Williams TP. Photoreceptor plasticity in the albinorat retina following unilateral optic nerve section. Exp Eye Res.1992;55:393–399.
29. Hollander H, Bisti S, Maffei L, Hebel R. Electroretinographic re-sponses and retrograde changes of retinal morphology after intra-cranial optic nerve section: a quantitative analysis in the cat. ExpBrain Res. 1984;55:483–493.
30. Chu Y, Humphrey MF, Alder VV, Constable IJ. Immunocytochem-ical localization of basic fibroblast growth factor and glial fibrillaryacidic protein after laser photocoagulation in the Royal College ofSurgeons rat. Aust NZ J Ophthalmol. 1998;26:87–96.
31. Liu C, Peng M, Laties AM, Wen R. Preconditioning with bright lightevokes a protective response against light damage in the rat retina.J Neurosci. 1998;18:1337–1344.
32. Stone J, Maslim J, Valter K, et al. Mechanisms of photoreceptordeath and survival in mammalian retina. Prog Retin Eye Res.1999;18:689–735.
33. LaVail MM, Unoki K, Yasumura D, Matthes MT, Yancopoulos GD,Steinberg RH. Multiple growth factors, cytokines, and neurotro-phins rescue photoreceptors from the damaging effects of con-stant light. Proc Natl Acad Sci USA. 1992;89:11249–11253.
34. Campochiaro P, Chang M, Ohsato M, et al. Retinal degeneration intransgenic mice with photoreceptor specific expression of a dom-inant-negative fibroblast growth factor receptor. J Neurosci. 1996;16:1679–1688.
35. Ip NY, McClain J, Barrezueta NX, et al. The alpha component ofthe CNTF receptor is required for signaling and defines potentialCNTF targets in the adult and during development. Neuron. 1993;10:89–102.
36. Schlichtenbrede FC, MacNeil A, Bainbridge JW, et al. Intraoculargene delivery of ciliary neurotrophic factor results in significantloss of retinal function in normal mice and in the Prph2Rd2/Rd2model of retinal degeneration. Gene Ther. 2003;10:523–527.
37. Cayouette M, Gravel C. Adenovirus-mediated gene transfer of cil-iary neurotrophic factor can prevent photoreceptor degenerationin the retinal degeneration (rd) mouses. Hum Gene Ther. 1997;8:423–430.
38. Cayouette M, Behn D, Sendtner M, Lachapelle P, Gravel C. Intraoc-ular gene transfer of ciliary neurotrophic factor prevents death andincreases responsiveness of rod photoreceptors in the retinal de-generation slow mouse. J Neurosci. 1998;18:9282–9293.
1754 Valter et al. IOVS, May 2005, Vol. 46, No. 5
Related Documents











