Behavioural Brain Research 275 (2014) 252–258 Contents lists available at ScienceDirect Behavioural Brain Research jou rn al hom epage: www.elsevier.com/locate/bbr Research report Time course of motor and cognitive functions after chronic cerebral ischemia in rats Thenmoly Damodaran a,∗ , Zurina Hassan a,b , Visweswaran Navaratnam a , Mustapha Muzaimi b,c , Gandi Ng f , Christian P. Müller e , Ping Liao f,g , Hans C. Dringenberg d a Centre for Drug Research, Universiti Sains Malaysia, 11800 Penang, Malaysia b Centre for Neuroscience Services and Research, Universiti Sains Malaysia, Health Campus, 16150 Kubang Kerian, Kelantan, Malaysia c Department of Neurosciences, School of Medical Sciences, Universiti Sains Malaysia, Health Campus, 16150 Kubang Kerian, Kelantan, Malaysia d Department of Psychology and Centre for Neuroscience Studies, Queen’s University, Kingston, ON, Canada K7L 3N6 e Department of Psychiatry and Psychotherapy, Friedrich-Alexander-University Erlangen-Nuremberg, Schwabachanlage 6, 91054 Erlangen, Germany f Calcium Signaling Laboratory, National Neuroscience Institute, Singapore g DUKE-NUS Graduate Medical School, Singapore h i g h l i g h t s • PBOCCA in rats leads to learning and memory dysfunctions. • PBOCCA in rats did not impair motor performance. • PBOCCA rats exhibit long-lasting cognitive impairment. a r t i c l e i n f o Article history: Received 18 February 2014 Received in revised form 5 September 2014 Accepted 8 September 2014 Available online 17 September 2014 Keywords: PBOCCA Locomotor activity Passive avoidance task Morris water maze Learning Memory, BDNF a b s t r a c t Cerebral ischemia is one of the leading causes of death and long-term disability in aging populations, due to the frequent occurrence of irreversible brain damage and subsequent loss of neuronal function which lead to cognitive impairment and some motor dysfunction. In the present study, the real time course of motor and cognitive functions were evaluated following the chronic cerebral ischemia induced by permanent, bilateral occlusion of the common carotid arteries (PBOCCA). Male Sprague Dawley rats (200–300 g) were subjected to PBOCCA or sham-operated surgery and tested 1, 2, 3 and 4 weeks following the ischemic insult. The results showed that PBOCCA significantly reduced step-through latency in a passive avoidance task at all time points when compared to the sham-operated group. PBOCCA rats also showed significant increase in escape latencies during training in the Morris water maze, as well as a reduction of the percentage of times spend in target quadrant of the maze at all time points following the occlusion. Importantly, there were no significant changes in locomotor activity between PBOCCA and sham-operated groups. The BDNF expression in the hippocampus was 29.3 ± 3.1% and 40.1 ± 2.6% on day 14 and 28 post PBOCCA, respectively compared to sham-operated group. Present data suggest that the PBOCCA procedure effectively induces behavioral, cognitive symptoms associated with cerebral ischemia and, consequently, provides a valuable model to study ischemia and related neurodegenerative disorder such as Alzheimer’s disease and vascular dementia. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Cerebral ischemia is defined as condition characterized by insuf- ficient blood flow to the brain, which will alter normal cellular function. Moreover, cerebral ischemia is one of the leading causes ∗ Corresponding author. Tel.: +60 4 6532726; fax: +60 4 6568669. E-mail address: [email protected] (T. Damodaran). of death and long-term disability in aging populations, due to the frequency occurrence of irreversible brain damage and subsequent loss of neuronal function [12]. Chronic cerebral ischemia (CCI) can induce the accumulation of nitric oxide (NO) and reactive oxygen species (ROS), leading to neurons injury in selective, vulnerable regions of the brain, especially the hippocampus and cerebral cor- tex, and subsequent cognitive impairments and motor dysfunction [11,18,25]. Furthermore, chronic cerebral ischemia has been con- sidered as the major cause of vascular dementia and Alzheimer’s http://dx.doi.org/10.1016/j.bbr.2014.09.014 0166-4328/© 2014 Elsevier B.V. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R
Ti
TMa
b
c
d
e
f
g
h
•••
a
ARRAA
KPLPMLM
1
fif
h0
Behavioural Brain Research 275 (2014) 252–258
Contents lists available at ScienceDirect
Behavioural Brain Research
jou rn al hom epage: www.elsev ier .com/ locate /bbr
esearch report
ime course of motor and cognitive functions after chronic cerebralschemia in rats
henmoly Damodarana,∗, Zurina Hassana,b, Visweswaran Navaratnama,ustapha Muzaimib,c, Gandi Ngf, Christian P. Müllere, Ping Liao f,g, Hans C. Dringenbergd
Centre for Drug Research, Universiti Sains Malaysia, 11800 Penang, MalaysiaCentre for Neuroscience Services and Research, Universiti Sains Malaysia, Health Campus, 16150 Kubang Kerian, Kelantan, MalaysiaDepartment of Neurosciences, School of Medical Sciences, Universiti Sains Malaysia, Health Campus, 16150 Kubang Kerian, Kelantan, MalaysiaDepartment of Psychology and Centre for Neuroscience Studies, Queen’s University, Kingston, ON, Canada K7L 3N6Department of Psychiatry and Psychotherapy, Friedrich-Alexander-University Erlangen-Nuremberg, Schwabachanlage 6, 91054 Erlangen, GermanyCalcium Signaling Laboratory, National Neuroscience Institute, SingaporeDUKE-NUS Graduate Medical School, Singapore
i g h l i g h t s
PBOCCA in rats leads to learning and memory dysfunctions.PBOCCA in rats did not impair motor performance.PBOCCA rats exhibit long-lasting cognitive impairment.
r t i c l e i n f o
rticle history:eceived 18 February 2014eceived in revised form 5 September 2014ccepted 8 September 2014vailable online 17 September 2014
eywords:BOCCAocomotor activityassive avoidance taskorris water maze
earning
a b s t r a c t
Cerebral ischemia is one of the leading causes of death and long-term disability in aging populations,due to the frequent occurrence of irreversible brain damage and subsequent loss of neuronal functionwhich lead to cognitive impairment and some motor dysfunction. In the present study, the real timecourse of motor and cognitive functions were evaluated following the chronic cerebral ischemia inducedby permanent, bilateral occlusion of the common carotid arteries (PBOCCA). Male Sprague Dawley rats(200–300 g) were subjected to PBOCCA or sham-operated surgery and tested 1, 2, 3 and 4 weeks followingthe ischemic insult. The results showed that PBOCCA significantly reduced step-through latency in apassive avoidance task at all time points when compared to the sham-operated group. PBOCCA rats alsoshowed significant increase in escape latencies during training in the Morris water maze, as well as areduction of the percentage of times spend in target quadrant of the maze at all time points followingthe occlusion. Importantly, there were no significant changes in locomotor activity between PBOCCA and
emory, BDNF sham-operated groups. The BDNF expression in the hippocampus was 29.3 ± 3.1% and 40.1 ± 2.6% on day14 and 28 post PBOCCA, respectively compared to sham-operated group. Present data suggest that thePBOCCA procedure effectively induces behavioral, cognitive symptoms associated with cerebral ischemiaand, consequently, provides a valuable model to study ischemia and related neurodegenerative disordersuch as Alzheimer’s disease and vascular dementia.
© 2014 Elsevier B.V. All rights reserved.
. Introduction
Cerebral ischemia is defined as condition characterized by insuf-cient blood flow to the brain, which will alter normal cellular
unction. Moreover, cerebral ischemia is one of the leading causes
∗ Corresponding author. Tel.: +60 4 6532726; fax: +60 4 6568669.E-mail address: [email protected] (T. Damodaran).
ttp://dx.doi.org/10.1016/j.bbr.2014.09.014166-4328/© 2014 Elsevier B.V. All rights reserved.
of death and long-term disability in aging populations, due to thefrequency occurrence of irreversible brain damage and subsequentloss of neuronal function [12]. Chronic cerebral ischemia (CCI) caninduce the accumulation of nitric oxide (NO) and reactive oxygenspecies (ROS), leading to neurons injury in selective, vulnerable
regions of the brain, especially the hippocampus and cerebral cor-tex, and subsequent cognitive impairments and motor dysfunction[11,18,25]. Furthermore, chronic cerebral ischemia has been con-sidered as the major cause of vascular dementia and Alzheimer’s
l Brai
di
idaTttdttCias
ic[itactdctemoie
ndfsgmll
oiwPmpaccs
2
2
oShaFd
T. Damodaran et al. / Behavioura
isease (AD), characterized by neurodegeneration and cognitivempairment with advancing age [27].
Several animal models have been developed to create ischemicnjuries with various degrees of severity in the rat brain, allowingetailed examinations of the cellular and metabolic mechanismsnd behavioral consequences of disrupted cerebral circulation.echniques such as ligation of specific, large arteries that supplyhe brain create conditions of cerebral hypoperfusion and triggerhe pathophysiological changes in the brain, leading to neuronalamage of the brain [1,8]. Further, some occlusion models, such ashe two-vessel occlusion of the common carotid arteries, give riseo ischemic cell changes in brain areas including the hippocampalA1 subfield, striatum, and neocortex. Interestingly, the damage
nduced by two-vessel occlusion of the common carotid arteriesppears to be similar in size and location as the injury typicallyeen with four-vessel occlusion models [13].
It is noteworthy that, in addition to mimicking acute ischemicnsults, some occlusion models also capture conditions of morehronic hypoperfusion of the cerebrum. According to Tsuchiya et al.21], permanent bilateral occlusion of the common carotid arter-es is a model of chronic cerebral hypoperfusion, which could leado cognitive impairments such as those seen in Alzheimer’s diseasend vascular dementia. The progressive neuronal degeneration andholinergic dysfunction following permanent bilateral occlusion ofhe common carotid arteries can result in long-lasting cognitioneficits in rat [14]. Furthermore, Scherr at al. [17] suggested thathronic blood flow restrictions may cause or exacerbate the cogni-ive decline seen in various forms of human dementia, a notion thatmphasizes the importance of developing and evaluating novel ani-al models of chronic hypoperfusion. In fact, a thorough evaluation
f the functional outcomes over the course of several weeks afterschemia is a key component in improving the clinical relevance ofxperimental dementia studies.
Brain-derived neurotrophic factor (BDNF) is a member of theerve growth factor family, potently promote cell proliferation, cellifferentiation, and neuronal protection and regulate the synapticunction in the central nervous system by stimulating intracellularignaling pathway [15]. BDNF also play important role in neuro-enesis, long-term potentiation (LTP), learning and memory, andood changes [24]. Moreover, clinical studies have suggested that
ow level of BDNF cause reduction in hippocampal volume whicheads to age related spatial memory impairments [6].
The present study was conducted to provide a real time coursef the neurobehavioral outcome following the chronic cerebralschemia induced by PBOCCA in rats. A battery of behavioral tests
ere administered over the course of a 4-week period following theBOCCA procedure, with tests chosen to assess both cognitive andotoric functions, thus providing a detailed behavioral and tem-
oral profile of the neurobehavioral functions that are impairednd spared as a result of chronic brain ischemia. The effect ofhronic cerebral ischemia on BDNF expression and morphologi-al changes in hippocampal CA1 region also were examined in thistudy.
. Methodology
.1. Animals
Male Sprague Dawley (SD) rats weighing 200–300 g werebtained from the breeding colony of the Animal Research andervice Centre (ARASC), Universiti Sains Malaysia (USM). They were
oused five rats per cage and maintained at a constant temper-ture on a standard 12:12 light/dark cycle with light on at 7 am.ood and water were given ad libitum. The experiments were con-ucted according to the approval of the Animal Ethics Committeen Research 275 (2014) 252–258 253
Universiti Sains Malaysia with the reference number USM/AnimalEthics Approval/2013/(87)(447).
2.2. Surgery
Permanent bilateral common carotid arteries occlusion(PBOCCA) in rats was performed as described previously [14,16].In brief, each rat was anesthetized with ketamine and xylazine(80 mg/kg/10 mg/kg, i.p.). In rats chosen randomly for PBOCCA(n = 48), the common carotid arteries were exposed via a ven-tral midline incision, carefully separate from their sheaths andvagus nerves, and permanently doubly ligate with 5/0 silk sutureapproximately 8–10 mm below the origin of the external carotidartery. The skin incision was closed and the rat was kept in anair-conditioned room at 25 ◦C. Sham-operated rat (n = 36) wassubjected to the same procedure without PBOCCA. The rats wererandomly assigned to 8 groups (n = 7–9/group): Sham-operated(week 1–week 4) and PBOCCA (week 1–week 4).
2.3. Locomotor activity
Locomotor activity in both the PBOCCA group and the sham-operated group was assessed at 1, 2, 3, and 4 weeks after the surgeryby means of an automated open-field apparatus (Pan Lab, Spain).The apparatus consisted of a Perspex cage (height: 40 cm, length:90 cm; width: 90 cm), with the bottom divided into five zones.The detection unit consisted of 45 × 45 cm frame contained total of16 × 16 infrared beams at an interval of 2.5 cm, located on the sidesand there are a total of 32 cells in one frame. The rat’s movementswere stored and analyzed offline. For testing, each rat was placed inthe center of the open field and spontaneous activity (ambulationand rearing) was recorded for 20 min (n = 7–9/group).
2.4. Passive avoidance task
Passive avoidance performance was examined using an appara-tus consisting of two compartments: a lighted box (plexiglass) and adark box (black) of the same size (20 × 20 × 40 cm each) separatedby a guillotine door (8 × 8 cm). The lighted box was illuminatedwith a lamp (60 W positioned above the apparatus). The floor wasconstructed of stainless-steel grids 2 mm in diameter and at 8-mmintervals. Intermittent electric shock (50 Hz, 10 s, 0.5 mA intensity)was delivered to the grid floor of the dark compartment by anisolated stimulator. At 1, 2, 3, and 4 weeks following surgery, anacquisition trial was performed. For this trial, the rat was allowedto habituate in both compartments freely for 3 min. During train-ing, each rat was placed in light compartment; 10 s later the doorbetween the compartments was opened and the latency to enterthe dark compartment with all four paws was timed. Once the ratcrossed with all four paws into the dark compartment, the door wasclosed and the 0.5 mA foot shock was administered for 10 s. The ratwas removed from the apparatus and returned to its cage. If the ratwaited for more than 100 s to cross to the dark compartment, theyare excluded from the experiment. A retention trial was performed24 h after the acquisition trial. It was conducted in a similar man-ner to the acquisition trial except that the shock was not appliedto the grid floor. If the rat did not enter to the dark compartmentwithin 300 s, the retention trial was terminated and the maximalstep through latency of 300 s was recorded. This procedure wascarried out as previously described, with some modification [23].
2.5. Morris water maze
The Morris water maze task was carried out as previouslydescribed, with some modification [23]. One day before the startof training, each rat was given a pretraining session in which they

2 l Brai
wdpv3a2qDiefitimtotwqfpqqr
2
h1Enff(fSatw(wWbfs
2
(bpweSf
2
utad
54 T. Damodaran et al. / Behavioura
ere allowed to swim freely in a pool (160 cm diameter, with aepth of 70 cm of water) for 60 s without an escape platform. Theool was placed in a large test room, and surrounded by variousisual cues. For formal training, the pool was filled to a depth of9 cm with water maintained at 25 ± 1 ◦C and made opaque by theddition of white paint. A platform (10 cm diameter) was situated
cm below the surface of the water. The pool was divided into fouruadrants, with the platform in a fixed position in one quadrant.aily training consisted of four trials in which the rat was placed
n the water from four different starting points and the latency ofscaping onto the platform was recorded. This was conducted forve consecutive days. A maximum of 60 s were allowed for eachrial during which the rat had to find the platform and climb ontot. If the rat failed to find the platform within 60 s, the trial was ter-
inated and a maximum score of 60 s was assigned. The rat washen guided to the hidden platform by hand and allowed to stayn the platform for 10 s before it was removed from the water. Onhe sixth day, each rat was subjected to a probe trial in which thereas no platform present. The time of crossing the former platform
uadrant and the total time of crossing all quadrants were recordedor 1 min. The percentage of crossing the quadrant of the formerlatform represented the ratio between the time of crossing theuadrant of the former platform and the total time of crossing alluadrants. The percentage was calculated as a measure of spatialeference memory.
.6. Western blot
Hippocampus from the PBOCCA brain tissue was isolated andomogenized in 200 �l of HEPES lysis buffer (20 mM Hepes;37 mM NaCl; 1% Triton X-100; 10% glycerol; 1.5 mM MgCl2; 1 mMGTA) with freshly added protease inhibitor (1:50, Roche Diag-ostics). Homogenized samples were centrifuged at 13,200 rpm
or 30 min at 4 ◦C and the supernatant was collected for Brad-ord protein assay (Cat # 500-0006, Bio-Rad). Extracted protein100 �g) was separated on 12% SDS-polyacrylamide gel and trans-erred onto PVDF membrane for 1 h at 100 V at room temperature.ubsequently, the membrane was blocked with 1% BSA for an hour,nd was probed with a rabbit anti-BDNF antibody (1:3000 dilu-ion, ab108383, abcam) overnight at 4 ◦C. Next, the membraneas washed and probed with HRP-conjugated anti-rabbit antibody
1:5000 dilution, A4914, Sigma–Aldrich) for an hour. The proteinas detected by chemiluminescence substrate AmershamTM ECLTM
estern Blotting Analysis System (RPN2209, GE Healthcare). Thelots were stripped and reprobed with mouse anti-�-actin (1:3000)or loading control. The protein bands were quantified using ImageJoftware.
.7. Histopathological evaluation
Animals were anaesthetized with overdose urethane1.3 mg/kg) intraperitonealy and pre-fixation was achievedy trans-cardiac perfusion with normal saline followed by 4%araformaldehyde (PFA) in 0.1 M phosphate buffer. The brainsere removed, post-fixed in 4% PFA at 4 ◦C. The brain was then
mbedded in paraffin wax and sectioned at multiple levels at 5 �m.ections of hippocampus were stained with hematoxylin–eosinor morphological analysis.
.8. Statistics
Behavioral performance in the water maze task was analyzed
sing a two-way ANOVA, followed by Bonferroni post hoc test. Sta-istical analysis for locomotor activity, passive avoidance latencies,nd probe trial performance were performed by unpaired Stu-ent t-test. The expression of BDNF was analyzed using a one-wayn Research 275 (2014) 252–258
ANOVA followed by Bonferroni post hoc test. Data were expressedas means ± S.E.M. Probability values less than 5% (p < 0.05) wereconsidered significant. Statistical analysis was performed using thesoftware Graph Pad Prism.
3. Results
3.1. Locomotor activity
The locomotor activity during 20 min open field test sessions forboth the PBOCCA and sham-operated group is presented in Fig. 1.Unpaired Student’s t-test did not reveal a significant difference inlocomotor activity between the groups at1 week (p = 0.3035), 2weeks (p = 0.6106), 3 weeks (p = 0.5339), and 4 weeks (p = 0.3961)after the surgery, indicating that PBOCCA did not impair motorperformance.
3.2. Passive avoidance task
Memory functions of PBOCCA rats and sham-operated animalswere assessed by a passive avoidance task. Memory performancewas assessed 24 h after an administration of an initial acquisitiontrial at 1, 2, 3, and 4 weeks following surgery. The latencies to enterthe dark (shock-paired) compartment on the retention trials arepresented in Fig. 2. Data analyzed using unpaired Student’s t-testshowed the step-through latency was decreased significantly inPBOCCA rats relative to sham-operated group at 1 week (p < 0.001),2 weeks (p < 0.05), 3 weeks (p < 0.05), and 4 weeks (p < 0.01) follow-ing the surgery.
3.3. Morris water maze
Two-way ANOVA revealed a statistically significant effectof lesion (PBOCCA vs. sham) on performance in the Morriswater during acquisition training, with PBOCCA animals show-ing longer escape latencies at all time points following theoperation (group effects: 1st week: F1,80 = 44.99; p < 0.0001; 2ndweek: F1,65 = 16.05; p = 0.0002; 3rd week: F1,70 = 193.1; p < 0.0001;4th week: F1,65 = 47.75; p < 0.0001). There were also significanteffects of test day (1st week: F4,80 = 42.13; p < 0.0001; 2nd week:F4,65 = 8.094; p < 0.0001; 3rd week: F4,70 = 30.66; p < 0.0001; 4thweek: F4,65 = 9.14; p < 0.0001), but no significant effect for the onlesion × day interaction (1st week: F4,80 = 0.9863; p = 0.4199; 2ndweek: F4,65 = 1.487; p = 0.2165; 3rd week: F4,70 = 2.221; p = 0.0755;4th week: F4,65 = 0.4654; p = 0.7609) (Fig. 3).
Probe trial performance was assessed on the sixth day by remov-ing the platform and allowing each rat to swim freely for 60 s. Thetime of the rat spent in the target quadrant was recorded. Stu-dent’s t-test (unpaired) indicated that the time spent in the targetquadrant during the probe trial was decreased significantly in thePBOCCA group relative to sham animals at, 1 week (p < 0.01), 2weeks (p < 0.01), 3 weeks (p < 0.001), and 4 weeks following thesurgery (p < 0.05) (Fig. 4).
3.4. BDNF expression after PBOCCA
As BDNF is critical for learning and memory (Bekinschtein et al.,2014 [2]), we decided to examine the expression of BDNF afterPBOCCA. Western blot was performed using the hippocampus col-lected from the rats with PBOCCA or sham operation. Two timepoints, 14 days and 28 days after PBOCCA were studied to illus-trate the temporal change of BDNF. The size of BDNF on the gel
is around 15 kDa. Three samples each from 14 and 28 days postPBOCCA were shown and compared to a sham control (Fig. 5a).BDNF level was decreased in all animals receiving PBOCCA, withthe most reduction occurred in animals 14 days after PBOCCA. We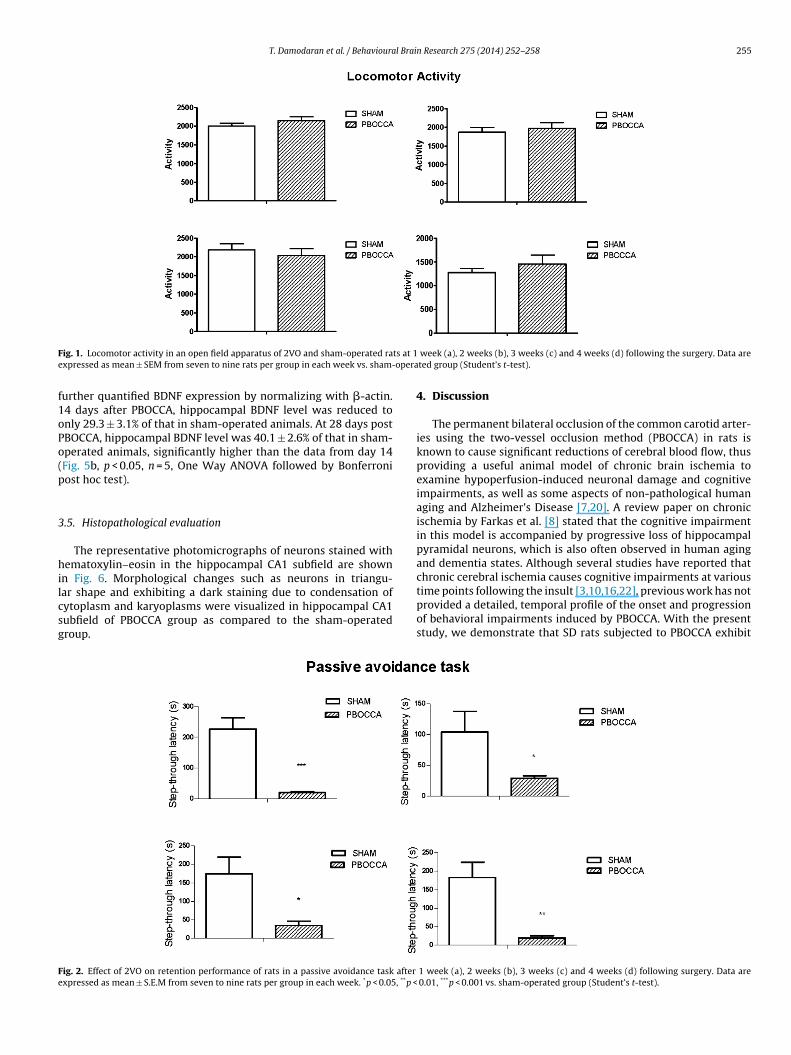
T. Damodaran et al. / Behavioural Brain Research 275 (2014) 252–258 255
F ts at 1e opera
f1oPo(p
3
hilcsg
Fe
ig. 1. Locomotor activity in an open field apparatus of 2VO and sham-operated raxpressed as mean ± SEM from seven to nine rats per group in each week vs. sham-
urther quantified BDNF expression by normalizing with �-actin.4 days after PBOCCA, hippocampal BDNF level was reduced tonly 29.3 ± 3.1% of that in sham-operated animals. At 28 days postBOCCA, hippocampal BDNF level was 40.1 ± 2.6% of that in sham-perated animals, significantly higher than the data from day 14Fig. 5b, p < 0.05, n = 5, One Way ANOVA followed by Bonferroniost hoc test).
.5. Histopathological evaluation
The representative photomicrographs of neurons stained withematoxylin–eosin in the hippocampal CA1 subfield are shown
n Fig. 6. Morphological changes such as neurons in triangu-ar shape and exhibiting a dark staining due to condensation ofytoplasm and karyoplasms were visualized in hippocampal CA1ubfield of PBOCCA group as compared to the sham-operatedroup.
ig. 2. Effect of 2VO on retention performance of rats in a passive avoidance task after
xpressed as mean ± S.E.M from seven to nine rats per group in each week. *p < 0.05, **p <
week (a), 2 weeks (b), 3 weeks (c) and 4 weeks (d) following the surgery. Data areted group (Student’s t-test).
4. Discussion
The permanent bilateral occlusion of the common carotid arter-ies using the two-vessel occlusion method (PBOCCA) in rats isknown to cause significant reductions of cerebral blood flow, thusproviding a useful animal model of chronic brain ischemia toexamine hypoperfusion-induced neuronal damage and cognitiveimpairments, as well as some aspects of non-pathological humanaging and Alzheimer’s Disease [7,20]. A review paper on chronicischemia by Farkas et al. [8] stated that the cognitive impairmentin this model is accompanied by progressive loss of hippocampalpyramidal neurons, which is also often observed in human agingand dementia states. Although several studies have reported thatchronic cerebral ischemia causes cognitive impairments at various
time points following the insult [3,10,16,22], previous work has notprovided a detailed, temporal profile of the onset and progressionof behavioral impairments induced by PBOCCA. With the presentstudy, we demonstrate that SD rats subjected to PBOCCA exhibit1 week (a), 2 weeks (b), 3 weeks (c) and 4 weeks (d) following surgery. Data are 0.01, ***p < 0.001 vs. sham-operated group (Student’s t-test).
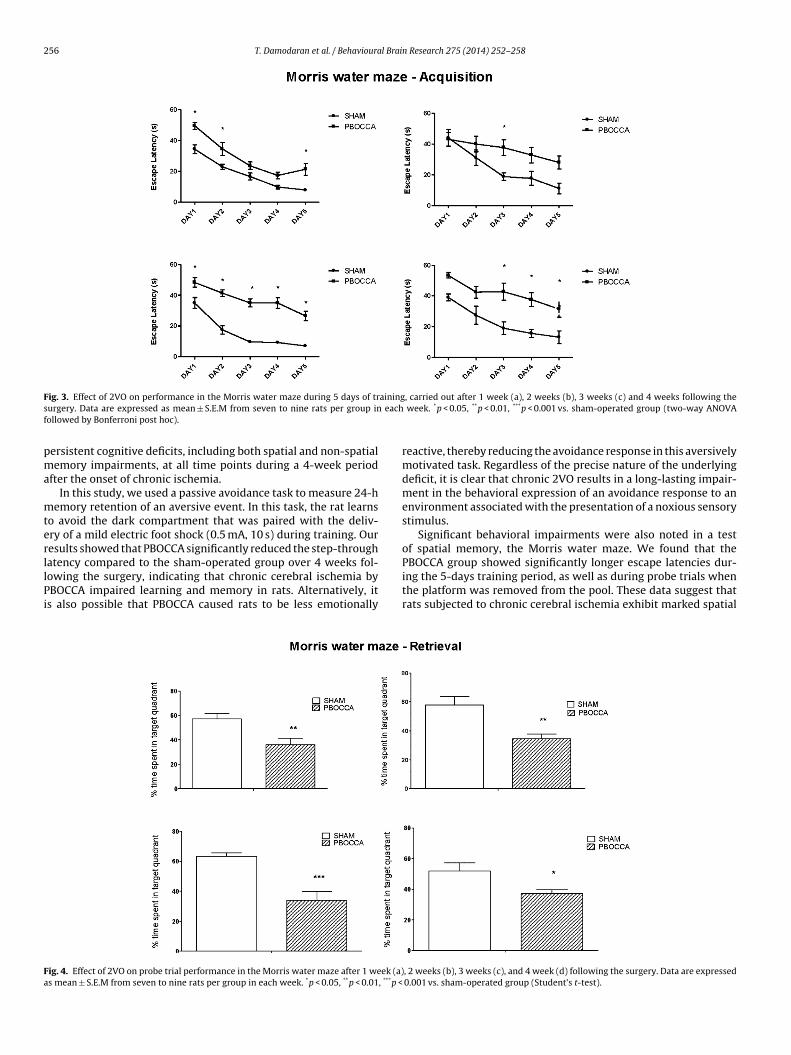
256 T. Damodaran et al. / Behavioural Brain Research 275 (2014) 252–258
F ainings eachf
pma
mterllPi
Fa
ig. 3. Effect of 2VO on performance in the Morris water maze during 5 days of trurgery. Data are expressed as mean ± S.E.M from seven to nine rats per group inollowed by Bonferroni post hoc).
ersistent cognitive deficits, including both spatial and non-spatialemory impairments, at all time points during a 4-week period
fter the onset of chronic ischemia.In this study, we used a passive avoidance task to measure 24-h
emory retention of an aversive event. In this task, the rat learnso avoid the dark compartment that was paired with the deliv-ry of a mild electric foot shock (0.5 mA, 10 s) during training. Ouresults showed that PBOCCA significantly reduced the step-throughatency compared to the sham-operated group over 4 weeks fol-
owing the surgery, indicating that chronic cerebral ischemia byBOCCA impaired learning and memory in rats. Alternatively, its also possible that PBOCCA caused rats to be less emotionallyig. 4. Effect of 2VO on probe trial performance in the Morris water maze after 1 week (a)s mean ± S.E.M from seven to nine rats per group in each week. *p < 0.05, **p < 0.01, ***p <
, carried out after 1 week (a), 2 weeks (b), 3 weeks (c) and 4 weeks following the week. *p < 0.05, **p < 0.01, ***p < 0.001 vs. sham-operated group (two-way ANOVA
reactive, thereby reducing the avoidance response in this aversivelymotivated task. Regardless of the precise nature of the underlyingdeficit, it is clear that chronic 2VO results in a long-lasting impair-ment in the behavioral expression of an avoidance response to anenvironment associated with the presentation of a noxious sensorystimulus.
Significant behavioral impairments were also noted in a testof spatial memory, the Morris water maze. We found that thePBOCCA group showed significantly longer escape latencies dur-
ing the 5-days training period, as well as during probe trials whenthe platform was removed from the pool. These data suggest thatrats subjected to chronic cerebral ischemia exhibit marked spatial, 2 weeks (b), 3 weeks (c), and 4 week (d) following the surgery. Data are expressed 0.001 vs. sham-operated group (Student’s t-test).

T. Damodaran et al. / Behavioural Brai
Fig. 5. Temporal expression of BDNF after 2VO. (a) Using Western blot, BDNF expres-sion in hippocampus was detected from rats receiving sham operation or PBOCCA.Ts*
rwnmai
Fai
he hippocampus was collected at day 14 and 28 after operation. (b) The expres-ion of BDNF was normalized to �-actin, and compared to sham-operated animals.p < 0.05, n = 5, one way ANOVA followed by Bonferroni post hoc test.
eference learning and memory deficits, results that are consistentith earlier reports [16,22]. It is important to note that there were
o significant differences in locomotor activity, measured in auto-ated open field apparatus, between the two experimental groupst any time following the PBOCCA or sham operation. These resultsndicate that the PBOCCA did not exert any obvious effects on basic
ig. 6. Neuronal damage in hippocampal CA1 subfield of PBOCCA rats. Photomicrograph with magnification 4× and panel c with magnification 400×) and PBOCCA group (paneschemic cell in the hippocampal CAI subfield of PBOCCA rat.
n Research 275 (2014) 252–258 257
motor abilities. These data may also suggest that longer escapelatencies in the PBOCCA group were related exclusively to slowerswim speeds, which were not measured in our experiments, butshould be monitored in future work.
According to Davidson et al. [5], chronic cerebral ischemia byPBOCCA can lead to visual impairments due to retina damage. How-ever, Kim et al. [10] reported that, in contrast to Wistar rats, SDrats had nearly intact optic tracts and showed no loss of papil-lary reflexes. Further, Xi et al. [22] had proven that they did notsee any significant differences in escape latencies between PBOCCAand control groups for training using a visible platform in the Mor-ris water maze. The fact that PBOCCA results in behavioral deficitsin a wide range of learning tasks (e.g., Pappas et al. [16]; Cecchettiet al. [3]; Xi et al., [22]; present results), some of which requirevery little visual processing (e.g., passive avoidance, as used here),also suggest that behavioral deficits are not entirely due to visualimpairments, even though it is possible that retinal damage mayaggravate the behavioral deficits in tasks that require the use ofvisual cues.
BDNF is a key mediator of memory processing by regulatingvarious intracellular signaling pathways. Stable memories requireoptimal levels of endogenous BDNF [2]. After cerebral ischemia,BDNF yields multiple neuroprotective effects including anti-apoptosis, anti-inflammation, anti-neurotoxicity, anti-epilepsy,promotion of neural regeneration, protection against periventric-ular leukomalacia, and contribution to cognitive and memoryacquisition [4]. To elucidate the role of BDNF in cognitive functionalloss, we studied the temporal expression of BDNF after PBOCCA.In consistent with a previous report Sun et al. [19], global hypoxiareduced BDNF expression in hippocampus which was prominent atday 14 after PBOCCA. Such reduction correlates with the cognitivefunctional loss. Interestingly, comparing to day 14, more BDNF wasproduced in hippocampus at day 28, albeit lower than in sham-operated animals. It has been proved that enrichment of BDNFwithin the injury area following cerebral hypoxia/ischemia can
enhance neural regeneration by stimulating neurite outgrowth andpromoting axonal sprouting and synaptogenesis [9,26]. However,we did not observe an improvement in cognitive functions at day28 comparing to at day 14. It is possible that the neuroregenerations showing pyramidal layer of the hippocampal CA1 subfield of sham group (panell b with magnification 4× and panel d with magnification 400×). Arrow shows the

2 l Brai
inai
5
icamtmfiohtc
A
tf(
R
[
[
[[
[
[
[
[
[
[
[
[
[
[
[
[
[after transplantation of neural stem cells modified by brain-derived neu-
58 T. Damodaran et al. / Behavioura
s not enough to yield functional outcome at day 28. More time iseeded or more BDNF-induced synaptogenesis is required to lead to
significant recovery in cognitive functions. One possible approachs to introduce exogenous BDNF into the site of injury.
. Conclusion
This study provides further evidence that chronic cerebralschemia due to PBOCCA in rats results in severe and long-lastingognitive impairments in several learning and memory (passivevoidance, spatial navigation), without affecting spontaneous loco-otor activity measured in an open field apparatus. Furthermore,
he decrease in BDNF expression in hippocampus after PBOCCAay aggravate the cognitive impairment. Our observation also con-
rm the notion that the PBOCCA rat model may capture aspectsf neurodegenerative diseases characterized by chronic, cerebralypoperfusion, such as Alzheimer’s disease and vascular demen-ia, both of which are characterized by significant behavioral andognitive deficits.
cknowledgements
Financial support was received from Ministry of Higher Educa-ion (MOHE) Malaysia and Universiti Sains Malaysia special fundingor the project of Fundamental Neuroscience-Neurobehaviour304/CNEURO/652201/K134).
eferences
[1] Bacigaluppi M, Comi G, Hermann DM. Animal models of ischemic stroke. Parttwo: Modeling cerebral ischemia. Open Neurol J 2010;4:34–8.
[2] Bekinschtein P, Cammarota M, Medina JH. BDNF and memory processing. Neu-ropharmacology 2014;76:677–83.
[3] Cechetti F, Pagnussat AS, Worm PV, Elsner VR, Ben J, da Costa MS, et al. Chronicbrain hypoperfusion causes early glial activation and neuronal death, and sub-sequent long-term memory impairment. Brain Res Bull 2012;87:109–16.
[4] Chen A, Xiong LJ, Tong Y, Mao M. The neuroprotective roles of BDNF in hypoxicischemic brain injury. Biomed Rep 2013;1:167–76.
[5] Davidson CM, Pappas BA, Stevens WD, Fortin T, Bennett SA. Chronic cerebralhypoperfusion: loss of pupillary reflex, visual impairment and retinal neurode-generation. Brain Res 2000;859:96–103.
[6] Erickson KI, Prakash RS, Voss MW, Chaddock L, Heo S, McLaren M, et al.Brain-derived neurotrophic factor is associated with age-related decline inhippocampal volume. J Neurosci 2010;30:5368–75.
[7] Farkas E, Donka G, de Vos RAI, Mihaly A, Bari F, Luiten PGM. Experimental
cerebral hypoperfusion induces white matter injury and microglial activationin the rat brain. Acta Neuropathol (Berl) 2004;108:57–64.[8] Farkas E, Luiten PGM, Bari F. Permanent, bilateral common carotid arteryocclusion in the rat: a model for chronic cerebral hypoperfusion-related neu-rodegenerative diseases. Brain Res Rev 2007;54:162–80.
[
n Research 275 (2014) 252–258
[9] Horch HW, Kruttgen A, Portbury SD, Katz LC. Destabilization of cortical den-drites and spines by BDNF. Neuron 1999;23(2):353–64.
10] Kim SK, Cho KO, Kim SY. White matter damage and hippocampal neurodegen-eration induced by permanent bilateral occlusion of common carotid arteryin the rat: comparison between Wistar and Spraque–Dawley strain. Korean JPhysiol Pharmacol 2008;12:89–94.
11] Li PA, He QP, Nakamura L, Csiszar K. Free radical spin trap �-phenyl-N-tert-butyl-nitron inhibits caspase-3 activation and reduces brain damage followinga severe forebrain ischemic injury. Free Radic Biol Med 2001;31:1191–7.
12] Lipton P. Ischemic cell death in brain neurons. Physiol Rev 1999;79:1431–568.13] McBean DE, Kelly PA. Rodent models of global cerebral ischemia: a com-
parison of two-vessel occlusion and four-vessel occlusion. Gen Pharmacol1998;30(4):431–4.
14] Ni JW, Matsumoto k, Li HB, Murakami Y, Watanobe H. Neuronal damage anddecrease of central acetylcholine level following permanent occlusion of bilat-eral common carotid arteries in rat. Brain Res 1995;673:290–6.
15] Numakawa T, Suzuki S, Kumamaru E, Adachi N, Richards M, Kunugi H.BDNF function and intracellular signaling in neurons. Histol Histopathol2010;25:237–58.
16] Pappas BA, Torre-de la JC, Davidson CM, Keyes MT, Fortin T. Chronic reductionof cerebral blood flow in the adult rat: late-emerging CA1 cell loss and memorydysfunction. Brain Res 1996;708:50–8.
17] Scherr M, Trinka E, Mc Coy M, Krenn Y, Staffen W, Kirschner M, et al. Cere-bral hypoperfusion during carotid artery stenosis can lead to cognitive deficitsthat may be independent of white matter lesion load. Curr Neurovasc Res2012;29(3):193–9.
18] Sinclair AJ, Bayer AJ, Johnston J, Warner C, Maxwell SR. Altered plasma antiox-idant status in subjects with Alzheimer’s disease and vascular dementia. Int JGeriatr Psychiatry 1998;13:840–5.
19] Sun H, Zhang J, Zhang L, Liu H, Zhu H, Yang Y. Environmental enrichmentinfluences BDNF and NR1 levels in the hippocampus and restores cogni-tive impairment in chronic cerebral hypoperfused rats. Curr Neurovasc Res2010;7(4):268–80.
20] Tohda M, Suwanakitch P, Jeenapongsa R, Hayashi H, Watanabe H, MatsumotoK. Expression changes of the mRNA of Alzheimer’s disease related factors inthe permanent ischemic rat brain. Biol Pharm Bull 2004;27(12):2021–3.
21] Tsuchiya M, Sako K, Yura S, Yonemasu Y. Cerebral blood flow and histopatho-logical changes following permanent bilateral carotid artery ligation in Wistarrats. Exp Brain Res 1992;89(1):87–92.
22] Xi Y, Wang M, Zhang W, Bai M, Du Y, Zhang Z, et al. Neuronal damage, cen-tral cholinergic dysfunction and oxidative damage correlate with cognitivedeficits in rats with chronic cerebral hypoperfusion. Neurobiol Learn Mem2014;109:7–19.
23] Xu J, Murakami Y, Matsumoto K, Tohda M, Watanabe H, Zhang S, et al. Protec-tive effect of Oren-gedoku-to (Huang-Lian-Jie-Du-Tang) against impairmentof learning and memory induced by transient cerebral ischemia in mice. JEnthnopharmacol 2000;73:405–13.
24] Yamada K, Mizuno M, Nabeshima T. Role of brain-derived neurotrophic factorin learning and memory. Life Sci 2002;70:735–44.
25] Zarow C, Vinters HV, Ellis WG, Weiner MW, Mungas D, White L, et al. Correlatesof hippocampal neuron number in Alzheimer’s disease and ischemic vasculardementia. Annu Neurol 2005;57:896–903.
26] Zhu JM, Zhao YY, Chen SD, Zhang WH, Lou L, Jin X. Functional recovery
rotrophic factor in rats with cerebral ischemia. J Int Med Res 2011;39(2):488–98.
27] Zlokovic BV. Neurovascular mechanisms of Alzheimer’s neurodegeneration.Trends Neurosci 2005;28:202–8.
Related Documents











