THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 263, No. 33, Issue of November 25, PP. 17643-17649,1988 Printed in U.S.A. Thyroid HormoneRegulates a and a+ Isoforms ofNa,K-ATPase during Development in Neonatal Rat Brain* (Received for publication, June 24, 1988) Cheryl A. SchmittS and Alicia A. McDonoughs From the Department of Physiology and Biophysics, University of Southern California, Los Angeles, California 90033 The brain contains two molecular forms of Na,K- ATPase designated a found in non-neuronal cells and neuronal soma and a+ found in axolemma. Previously we have shown that the abundance of both forms (de- termined by immunoblots) as well as Na,K-ATPase activity increases 10-fold between 4 days before and 20 days after birth (Schmitt, C. A., and McDonough, A. A. (1986) J. Biol. Chem. 261, 10439-10444). Hy- pothyroidism in neonates blunts these increases. Neo- natal, but not adult brain Na,K-ATPase is thyroid hor- mone (triiodothyronine, T3) responsive. This study defines the period during which brain Na,K-ATPase responds to T3. The start of the critical period was defined by comparing Na,K-ATPase activity and a and a+ abundance in hypothyroid and euthyroid neonates (birth to 30 days of age). For all parameters, euthyroid was significantly higher by 15 days of age. The end of the critical period was defined by dosing hypothyroid neonates with T3 daily (0.1 pg/g body weight) begin- ning at increasing days of age, and sacrificing all at 30 days then assaying enzyme activity and abundance. Those starting T3 treatment on or before day 19 were restored to euthyroid levels of Na,K-ATPase activity and abundance, while those starting T3 treatment on or after day 22 remained at hypothyroid levels of enzyme activity and abundance. We conclude that brain Na,K-ATPase (Y and a+ isoforms are sensitive to T3 by as late as 15 days of age and that the period of thyroid hormone responsiveness is over by 22 days. Na,K-ATPase,the sodium pump,anubiquitousplasma membrane-bound enzyme consisting of catalytic a subunit (a M, 112,573 and/or a+ M, 111,736) (1) and a glycoprotein p subunit (M, 34,639) (2), is a crucial enzyme in the central nervous system. By transducing thechemical energy of ATP into transmembrane gradients by the forced exchange of three sodium out of the cell for two potassium into the cell, the pump generates electrochemical gradients that control the ionic environment essential for neuronal activity in the cen- tral nervous system. The electrochemical gradient supplies energy for sodium-dependent transport systems allowing reuptake of neurotransmitters during nerve impulses (3-5), * This work was supported in part by National Institutes of Health Grant DK 34316. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solelyto indicate this fact. Grant GM08017-03. $ Supported by National Institutes of Health Predoctoral Training 5 Established Investigator of the American Heart Association. To whom correspondence should be addressed Dept. of Physiology and Biophysics, University of Southern California, 1333 San Pablo, Los Angeles, CA 90033. and organic solute movement (6). Evidence is accumulating for multiple forms of Na,K- ATPase a and p subunits. Twomolecular forms of the a catalytic subunit (a and a+) were first identified in brain by Sweadner (7) and have since been identified in eye (8,9) and in canine cardiac muscle (10, 11). Three unique a isozyme cDNAs (a, a+ and aIII) have been isolatedfrom a brain cDNA library, although no peptide form of a111 has been identified (1). Four sizes of /3 subunit mRNA derived from different start and stop sites have been identified in rat brain, and the expression of thedifferent sizes of p mRNAs is regulated in a tissue-specific manner (2). These mRNAs ap- pear to code for a single p peptide (12). Brain Na,K-ATPase activity increases 10-15-fold during development beginning a few days before birth and approach- ing adult values 20-30 days after birth (13-15), coincident with a change in ionic composition of brain tissue (13, 16) and the onset of electrical activity (17). The increase in Na,K- ATPase activity is due to accumulation of functional pumps as measured by sodium-dependent phosphorylation (14). We have previously shown, by immunoblotanalysis,thatthe abundance of both the a and a+ isozymes increases during this period (15). Thyroid hormone (triiodothyronine, T3),l required for nor- mal growth and differentiationof the central nervous system (18), regulates Na,K-ATPase activity in developing brain (13), but does not regulate Na,K-ATPase activityor abundance in adult brain (19, 20). In a previous study we have shown that thyroid hormone regulates Na,K-ATPase activity and abun- dance during development. In 15-day hypothyroid neonates, sodium pump activity and a isozyme abundance were 50-70% of euthyroid values (15). These results are supported by those of Attenvill et al. (21) who showed significant decreases in both a and a+ activities in hypothyroid neonatal rat brain discriminated on the basis of differential ouabain sensitivities. This paper tests the hypothesis that a critical period exists during brain development when TB regulates the abundance of a and a+ isozymes. We delimit this period during devel- opment by measuring Na,K-ATPase activity and a and a+ isozyme abundance in hypothyroid, T3-injected hypothyroid, and euthyroid neonates, to test when TB exerts its regulatory effects. A preliminary report of a portion of this study was published previously (22). EXPERIMENTALPROCEDURES Animals-In the first study, Na,K-ATPase in brains of hypothy- roid and euthyroid neonates were compared. Pregnant Sprague-Daw- ley rats were transported on gestational day 13 and housed on a 12-h light/dark schedule with Purina rat chow and water ad libitum. On 15.5 days gestation hypothyroidism was established by treating the pregnant rats with low iodine test diet (ICN, Radiochemicals), 0.5% The abbreviations used are: T4, thyroxine; TS, triiodothyronine; SDS, sodium dodecyl sulfate. 17643

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

T H E JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 263, No. 33, Issue of November 25, PP. 17643-17649,1988 Printed i n U.S.A.
Thyroid Hormone Regulates a and a+ Isoforms of Na,K-ATPase during Development in Neonatal Rat Brain*
(Received for publication, June 24, 1988)
Cheryl A. SchmittS and Alicia A. McDonoughs From the Department of Physiology and Biophysics, University of Southern California, Los Angeles, California 90033
The brain contains two molecular forms of Na,K- ATPase designated a found in non-neuronal cells and neuronal soma and a+ found in axolemma. Previously we have shown that the abundance of both forms (de- termined by immunoblots) as well as Na,K-ATPase activity increases 10-fold between 4 days before and 20 days after birth (Schmitt, C. A., and McDonough, A. A. (1986) J. Biol. Chem. 261, 10439-10444). Hy- pothyroidism in neonates blunts these increases. Neo- natal, but not adult brain Na,K-ATPase is thyroid hor- mone (triiodothyronine, T3) responsive. This study defines the period during which brain Na,K-ATPase responds to T3. The start of the critical period was defined by comparing Na,K-ATPase activity and a and a+ abundance in hypothyroid and euthyroid neonates (birth to 30 days of age). For all parameters, euthyroid was significantly higher by 15 days of age. The end of the critical period was defined by dosing hypothyroid neonates with T3 daily (0.1 pg/g body weight) begin- ning at increasing days of age, and sacrificing all at 30 days then assaying enzyme activity and abundance. Those starting T3 treatment on or before day 19 were restored to euthyroid levels of Na,K-ATPase activity and abundance, while those starting T3 treatment on or after day 22 remained at hypothyroid levels of enzyme activity and abundance. We conclude that brain Na,K-ATPase (Y and a+ isoforms are sensitive to T3 by as late as 15 days of age and that the period of thyroid hormone responsiveness is over by 22 days.
Na,K-ATPase, the sodium pump, an ubiquitous plasma membrane-bound enzyme consisting of catalytic a subunit ( a M , 112,573 and/or a+ M , 111,736) (1) and a glycoprotein p subunit ( M , 34,639) (2), is a crucial enzyme in the central nervous system. By transducing the chemical energy of ATP into transmembrane gradients by the forced exchange of three sodium out of the cell for two potassium into the cell, the pump generates electrochemical gradients that control the ionic environment essential for neuronal activity in the cen- tral nervous system. The electrochemical gradient supplies energy for sodium-dependent transport systems allowing reuptake of neurotransmitters during nerve impulses (3-5),
* This work was supported in part by National Institutes of Health Grant DK 34316. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Grant GM08017-03. $ Supported by National Institutes of Health Predoctoral Training
5 Established Investigator of the American Heart Association. To whom correspondence should be addressed Dept. of Physiology
and Biophysics, University of Southern California, 1333 San Pablo, Los Angeles, CA 90033.
and organic solute movement (6). Evidence is accumulating for multiple forms of Na,K-
ATPase a and p subunits. Two molecular forms of the a catalytic subunit ( a and a+) were first identified in brain by Sweadner (7) and have since been identified in eye (8,9) and in canine cardiac muscle (10, 11). Three unique a isozyme cDNAs (a , a+ and aIII) have been isolated from a brain cDNA library, although no peptide form of a111 has been identified (1). Four sizes of /3 subunit mRNA derived from different start and stop sites have been identified in rat brain, and the expression of the different sizes of p mRNAs is regulated in a tissue-specific manner (2). These mRNAs ap- pear to code for a single p peptide (12).
Brain Na,K-ATPase activity increases 10-15-fold during development beginning a few days before birth and approach- ing adult values 20-30 days after birth (13-15), coincident with a change in ionic composition of brain tissue (13, 16) and the onset of electrical activity (17). The increase in Na,K- ATPase activity is due to accumulation of functional pumps as measured by sodium-dependent phosphorylation (14). We have previously shown, by immunoblot analysis, that the abundance of both the a and a+ isozymes increases during this period (15).
Thyroid hormone (triiodothyronine, T3),l required for nor- mal growth and differentiation of the central nervous system (18), regulates Na,K-ATPase activity in developing brain (13), but does not regulate Na,K-ATPase activity or abundance in adult brain (19, 20). In a previous study we have shown that thyroid hormone regulates Na,K-ATPase activity and abun- dance during development. In 15-day hypothyroid neonates, sodium pump activity and a isozyme abundance were 50-70% of euthyroid values (15). These results are supported by those of Attenvill et al. (21) who showed significant decreases in both a and a+ activities in hypothyroid neonatal rat brain discriminated on the basis of differential ouabain sensitivities.
This paper tests the hypothesis that a critical period exists during brain development when TB regulates the abundance of a and a+ isozymes. We delimit this period during devel- opment by measuring Na,K-ATPase activity and a and a+ isozyme abundance in hypothyroid, T3-injected hypothyroid, and euthyroid neonates, to test when TB exerts its regulatory effects. A preliminary report of a portion of this study was published previously (22).
EXPERIMENTAL PROCEDURES
Animals-In the first study, Na,K-ATPase in brains of hypothy- roid and euthyroid neonates were compared. Pregnant Sprague-Daw- ley rats were transported on gestational day 13 and housed on a 12-h light/dark schedule with Purina rat chow and water ad libitum. On 15.5 days gestation hypothyroidism was established by treating the pregnant rats with low iodine test diet (ICN, Radiochemicals), 0.5%
The abbreviations used are: T4, thyroxine; TS, triiodothyronine; SDS, sodium dodecyl sulfate.
17643

17644 Thyroid Regulation of Na,K-ATPase in Developing Brain perchlorate drinking water, and propylthiouracil (25-mg pellet im- planted subcutaneously; Innovative Research of America, Rockville, MD), as described previously (15). After birth the mothers were maintained on this regime in order to maintain hypothyroid neonates. Hypothyroid and euthyroid neonates were killed by carbon dioxide asphyxiation and decapitation. The number of hypothyroid brain samples assayed ranged from one to six and are listed as follows: day 1 (3); day 3 (3); day 6 (3); day 8 (3); day 10 (5); day 12 (2); day 14 (2); day 15 (4); day 29 (3); day 22 (4); day 24 (1); and day 30 (5). The number of euthyroid brain samples assayed were the same as de- scribed for hypothyroid except on day 15 (5) and day 30 (6). Each point plotted is expressed as the mean f the standard error.
In the second study, Na,K-ATPase in brains of T3-supplemented hypothyroid neonates were compared with hypothyroid and euthyroid controls. Hypothyroidism was established as described for the first study. TS was restored to groups of hypothyroid neonates starting at 7, 10, 19,22,25, and 28 days of age by daily subcutaneous injections with 0.1 pg of Ts/g body weight. This dose has been shown to be sufficient to saturate nuclear T3 receptors in the brain (23). The T, was prepared from a 10 mg of Ta/ml 0.1 N NaOH stock, diluted to a final concentration of 0.5 pg of T3/p1 diluent (diluent: 0.005 N NaOH, 1 % bovine serum albumin, pH 7.0 to 7.5 adjusted with HCl), while age-matched hypothyroid and euthyroid controls received diluent only. The mothers of T3-injected neonates were returned to regular food and water ad libitum, while the mothers of diluent-injected hypothyroid neonates remained on the test chow and perchlorate water throughout the study. All neonates were killed at 30 days of age for subsequent Na,K-ATPase studies. The number of brain samples (varying between one and six) that were collected at 30 days for each treatment group beginning daily thyroid hormone injections on sequentially later days of age, are listed as follows: day 7 (2); day 10 (3); day 16 (1); day 19 (3); day 22 (4); day 25 (4); and day 28 (4). Five hypothyroid and six euthyroid control samples were also col- lected on 30 days and assayed (Figs. 6 and 8). Each point plotted is expressed as the mean f the standard error.
Preparation of Crude Membrane Fractions-Crude membrane frac- tions were prepared by homogenization and differential centrifuga- tion according to Jorgensen (24) as described previously (15). In the first study, two to five brains (as listed above) from hypothyroid and euthyroid neonates were processed separately. In the second study, one to six brains (as listed above) from 30-day-old T3-supplemented hypothyroid, hypothyroid, and euthyroid neonates were processed separately. Membrane preparations were assayed for Na,K-ATPase activity as described by Lo et al. (25). Each sample was analyzed at least two times in triplicate (+ouabain). 0.1% sodium deoxycholate was added to the membranes immediately prior to the assay to permeabilize vesicles as described previously (15). Protein concentra- tions were determined by the method of Lowry (26). Data were analyzed by the Student’s t test and were accepted as statistically significant when P < 0.01.
Detection and Quantitation of the a Subunits-a and a+ subunits were resolved using 7.5% Laemmli gels (27) as modified by Best et al. (28) and a resolving gel pH of 9.0 as suggested by Sackett et al. (29). Samples were electrophoresed at 20 mA/gel for 2 h after the brom- phenol blue tracking dye had run off the bottom, to enhance resolu- tion of the two isozymes as described previously (15).
Preparation and characterization of antibodies to guinea pig kidney Na,K-ATPase has been described (30). Antibodies to rat kidney Na,K-ATPase were provided by M. Ellisman, University of California at San Diego, and their preparation and characterization has been described (31). The ability of these antisera to bind to the Na,K- ATPase a isozymes has been established (15). The immunoblot method was as described in McDonough and Schmitt (32). Briefly, 25 pg of membrane protein was applied per gel lane and processed by SDS-polyacrylamide gel electrophoresis. Following electrophoresis, the resolved proteins were covalently attached to diazophenyl- thioether paper, incubated with antibodies, and labeled with iodinated protein-A according to the immunoblot method of Renart et al. (33) as described by McDonough and Schmitt (32).
Autoradiograms of immunoblots of a isozymes were scanned with a Hoefer scanning densitometer and data compiled using Chromlab software (Data Translations, Marlborough, MA), as described previ- ously (15). Each sample was analyzed at least two times, and each lane was scanned two times. Areas under curves were calculated by integration and normalized to the mean euthyroid value. In the first study, integrated areas of hypothyroid a and a+ subunits were normalized to age-matched euthyroid controls. In the second study, integrated areas of hypothyroid and Ta-injected hypothyroid a and
a+ subunits were normalized to 30-day euthyroid controls. Standard error of the normalized euthyroid data was calculated by determining the mean value of two to three euthyroid samples run concurrently on each gel, and then dividing each euthyroid data point by this calculated mean. These ratios were calculated for each euthyroid animal in the experiments represented in the first (Fig. 5) and second (Fig. 8) studies and compiled as the euthyroid mean -+ the standard error. For each data point representing hypothyroid and T3-injected hypothyroid values, the normalized mean f the standard error is plotted. Data were compared by the Student’s t test and significance was accepted at P < 0.01.
Materials-Chemicals were reagent grade, spectroquality, or elec- trophoresis purity reagents, except for the sodium lauryl sulfate (Pierce Chemical Co., catalogue No. 28364). Leupeptin, phenylmeth- ylsulfonyl fluoride, triiodothyronine, and the chemicals for immuno- blots were from Sigma. 4-Aminobenzamidine dihydrochloride and the chemicals for preparation of the diazophenylthioether paper were from Aldrich. lZ5I-Protein A was purchased from ICN. T S and T4 concentrations were determined by radioimmunoassay (Amerlax, Amersham Corp).
RESULTS
In the first study, we delimit the beginning of the period for T3 induction of Na,K-ATPase during development by assaying Na,K-ATPase activity and a isozyme abundance in hypothyroid and euthyroid neonates.
T3 Plasma Concentration in Adult and Neonatal Hypothy- roid and Euthyroid Rats-T3 plasma concentrations in the mother rats on the hypothyroid regime described under “Ex- perimental Procedures,” were 25% of euthyroid controls after 8 days and undetectable by 15 days of treatment (data not shown). To ensure that maternal treatment was rendering neonatal rats hypothyroid, we also measured T3 and T4 con- centrations in trunk blood at the time of sacrifice (Fig. 1). T3 concentrations were low in hypothyroid neonates at all ages, diverged from euthyroid after 10 days, and were significantly different by 15 days. Plasma T4 levels in hypothyroid neonates were significantly lower than euthyroid neonates at all ages assayed. T4 levels were undetectable before 10 days in hypo- thyroids. The fact that some neonatal animals have low but significant levels of T3 may reflect incomplete suppression of thyroid function in some mothers, which would supply a potential source of iodine in neonates. Therefore, we cannot completely rule out the possibility that some newborn animals have small amounts of serum thyroid hormone. However, brain weights, body weights (discussed below), and T3 concen- trations suggest newborn neonates are all hypothyroid if not T3-free.
Body and Brain Weight Determinations in Neonatal Hypo- thyroid and Euthyroid Rats-Body and brain weights of hy-
1 [ ’ O0
I I ,= I .
AGE (DAYS)
FIG. 1. Plasma Ts and T4 concentrations in hypothyroid and euthyroid neonates. Plasma T3 and T4 concentrations (nanograms/ p l ) of neonatal rats were determined on trunk bloods of pups from three hypothyroid and three euthyroid litters between 1 and 30 days. A, 0, hypothyroid; A, 0, euthyroid.
Thyroid Regulation of Na,K-
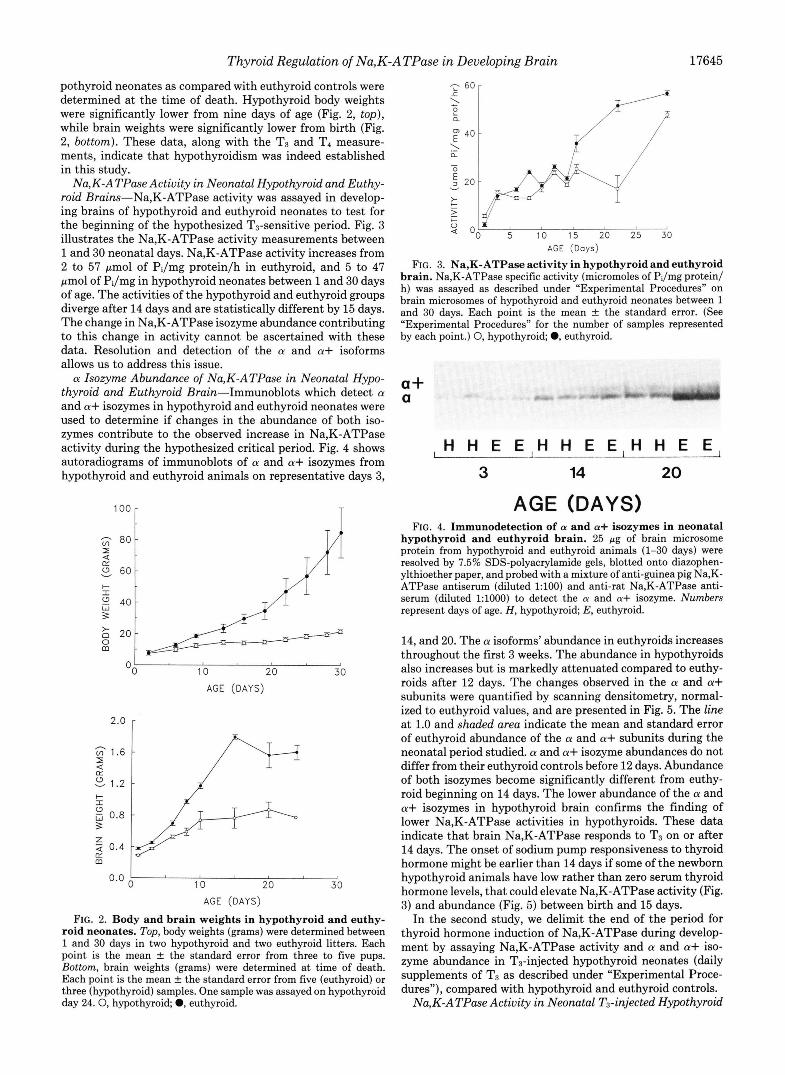
pothyroid neonates as compared with euthyroid controls were determined at the time of death. Hypothyroid body weights were significantly lower from nine days of age (Fig. 2, top), while brain weights were significantly lower from birth (Fig. 2, bottom). These data, along with the T3 and T4 measure- ments, indicate that hypothyroidism was indeed established in this study.
Na,K-ATPase Actiuity in Neonatal Hypothyroid and Euthy- roid Brains-Na,K-ATPase activity was assayed in develop- ing brains of hypothyroid and euthyroid neonates to test for the beginning of the hypothesized T3-sensitive period. Fig. 3 illustrates the Na,K-ATPase activity measurements between 1 and 30 neonatal days. Na,K-ATPase activity increases from 2 to 57 Fmol of PJmg proteinlh in euthyroid, and 5 to 47 pmol of PJmg in hypothyroid neonates between 1 and 30 days of age. The activities of the hypothyroid and euthyroid groups diverge after 14 days and are statistically different by 15 days. The change in Na,K-ATPase isozyme abundance contributing to this change in activity cannot be ascertained with these data. Resolution and detection of the a and a+ isoforms allows us to address this issue. a Isozyme Abundance of Na,K-ATPase in Neonatal Hypo-
thyroid and Euthyroid Brain-Immunoblots which detect a and a+ isozymes in hypothyroid and euthyroid neonates were used to determine if changes in the abundance of both iso- zymes contribute to the observed increase in Na,K-ATPase activity during the hypothesized critical period. Fig. 4 shows autoradiograms of immunoblots of a and a+ isozymes from hypothyroid and euthyroid animals on representative days 3,
v 60
40 3
m 2ol
T II
10 20 30 AGE (DAYS)
2.0 r
m 0.4 ty 0.0
10 20 30 AGE (DAYS)
FIG. 2. Body and brain weights in hypothyroid and euthy- roid neonates. Top, body weights (grams) were determined between 1 and 30 days in two hypothyroid and two euthyroid litters. Each point is the mean f the standard error from three to five pups. Bottom, brain weights (grams) were determined at time of death. Each point is the mean f the standard error from five (euthyroid) or three (hypothyroid) samples. One sample was assayed on hypothyroid day 24 .0 , hypothyroid; 0, euthyroid.
ATPase in Developing Brain 17645
e a *,I
AGE (Days)
FIG. 3. Na,K-ATPase activity in hypothyroid and euthyroid brain. Na,K-ATPase specific activity (micromoles of Pi/mg protein/ h) was assayed as described under “Experimental Procedures” on brain microsomes of hypothyroid and euthyroid neonates between 1 and 30 days. Each point is the mean f the standard error. (See “Experimental Procedures” for the number of samples represented by each point.) 0, hypothyroid; 0, euthyroid.
a+ a
, H H E E , H H E E , H H E E ,
3 14 20
AGE (DAYS) FIG. 4. Immunodetection of a and a+ isozymes in neonatal
hypothyroid and euthyroid brain. 25 pg of brain microsome protein from hypothyroid and euthyroid animals (1-30 days) were resolved by 7.5% SDS-polyacrylamide gels, blotted onto diazophen- ylthioether paper, and probed with a mixture of anti-guinea pig Na,K- ATPase antiserum (diluted 1:lOO) and anti-rat Na,K-ATPase anti- serum (diluted 1:lOOO) to detect the a and a+ isozyme. Numbers represent days of age. H, hypothyroid; E, euthyroid.
14, and 20. The a isoforms’ abundance in euthyroids increases throughout the first 3 weeks. The abundance in hypothyroids also increases but is markedly attenuated compared to euthy- roids after 12 days. The changes observed in the a and a+ subunits were quantified by scanning densitometry, normal- ized to euthyroid values, and are presented in Fig. 5. The line at 1.0 and shaded area indicate the mean and standard error of euthyroid abundance of the a and a+ subunits during the neonatal period studied. a and a+ isozyme abundances do not differ from their euthyroid controls before 12 days. Abundance of both isozymes become significantly different from euthy- roid beginning on 14 days. The lower abundance of the a and a+ isozymes in hypothyroid brain confirms the finding of lower Na,K-ATPase activities in hypothyroids. These data indicate that brain Na,K-ATPase responds to T3 on or after 14 days. The onset of sodium pump responsiveness to thyroid hormone might be earlier than 14 days if some of the newborn hypothyroid animals have low rather than zero serum thyroid hormone levels, that could elevate Na,K-ATPase activity (Fig. 3) and abundance (Fig. 5) between birth and 15 days.
In the second study, we delimit the end of the period for thyroid hormone induction of Na,K-ATPase during develop- ment by assaying Na,K-ATPase activity and a and a+ iso- zyme abundance in T3-injected hypothyroid neonates (daily supplements of T3 as described under “Experimental Proce- dures”), compared with hypothyroid and euthyroid controls.
Na,K-ATPase Activity in Neonatal T3-injected Hypothyroid
17646 Thyroid Regulation of Na,K-ATPase in Developing Brain
0.4 1
0.0 ‘ 0 5 10 15 20 25 30
AGE (Days)
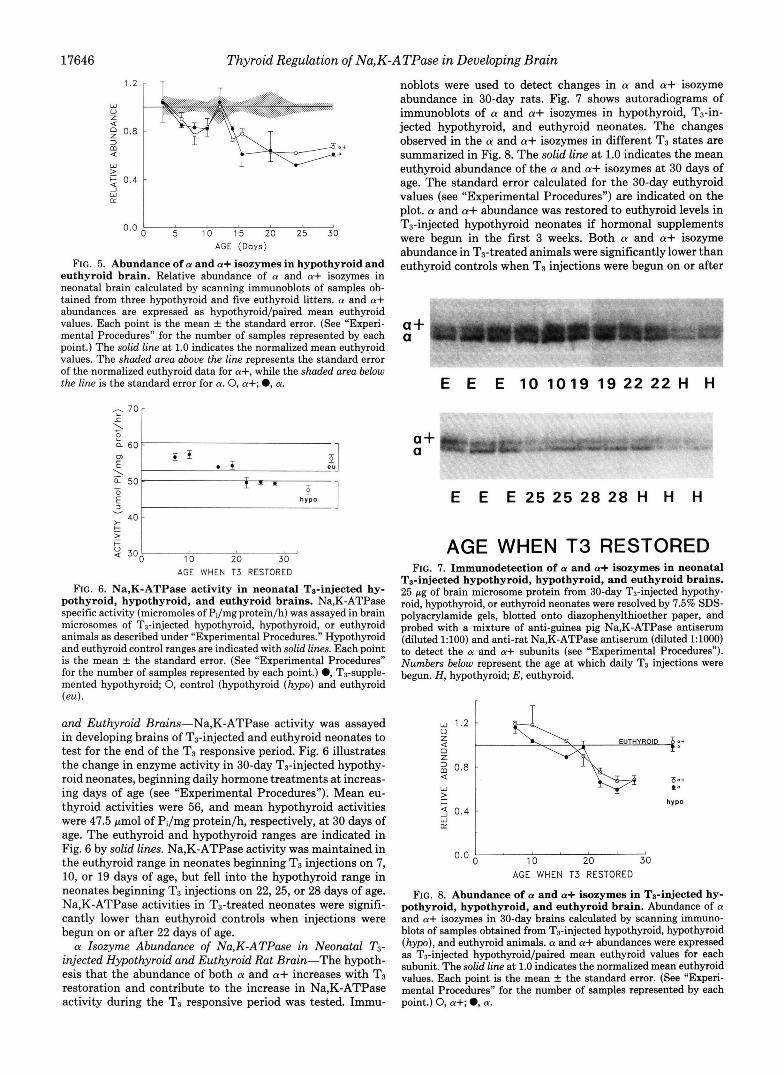
FIG. 5. Abundance of a and a+ isozymes in hypothyroid and euthyroid brain. Relative abundance of a and a+ isozymes in neonatal brain calculated by scanning immunoblots of samples ob- tained from three hypothyroid and five euthyroid litters. a and a+ abundances are expressed as hypothyroid/paired mean euthyroid values. Each point is the mean f the standard error. (See “Experi- mental Procedures” for the number of samples represented by each point.) The solid line at 1.0 indicates the normalized mean euthyroid values. The shaded area above the line represents the standard error of the normalized euthyroid data for a+, while the shaded area below the line is the standard error for a. 0, a+; 0, a.
- 70r
v ; 4 o y l 2
Q 300 c 0
10 20 30 AGE WHEN T3 RESTORED
FIG. 6. Na,K-ATPase activity in neonatal Ts-injected hy- pothyroid, hypothyroid, and euthyroid brains. Na,K-ATPase specific activity (micromoles of PJmg protein/h) was assayed in brain microsomes of T3-injected hypothyroid, hypothyroid, or euthyroid animals as described under “Experimental Procedures.” Hypothyroid and euthyroid control ranges are indicated with solid lines. Each point is the mean f the standard error. (See “Experimental Procedures” for the number of samples represented by each point.) 0, T3-supple- mented hypothyroid; 0, control (hypothyroid (hypo) and euthyroid (eu).
and Euthyroid Brains-Na,K-ATPase activity was assayed in developing brains of T3-injected and euthyroid neonates to test for the end of the T3 responsive period. Fig. 6 illustrates the change in enzyme activity in 30-day T3-injected hypothy- roid neonates, beginning daily hormone treatments at increas- ing days of age (see “Experimental Procedures”). Mean eu- thyroid activities were 56, and mean hypothyroid activities were 47.5 pmol of PJmg protein/h, respectively, at 30 days of age. The euthyroid and hypothyroid ranges are indicated in Fig. 6 by solid lines. Na,K-ATPase activity was maintained in the euthyroid range in neonates beginning T3 injections on 7, 10, or 19 days of age, but fell into the hypothyroid range in neonates beginning T3 injections on 22,25, or 28 days of age. Na,K-ATPase activities in T3-treated neonates were signifi- cantly lower than euthyroid controls when injections were begun on or after 22 days of age. a Isozyme Abundance of Na,K-ATPase in Neonatal T3-
injected Hypothyroid and Euthyroid Rat Brain-The hypoth- esis that the abundance of both a and a+ increases with T3 restoration and contribute to the increase in Na,K-ATPase activity during the T3 responsive period was tested. Immu-
noblots were used to detect changes in a and a+ isozyme abundance in 30-day rats. Fig. 7 shows autoradiograms of immunoblots of a and a+ isozymes in hypothyroid, Ta-in- jected hypothyroid, and euthyroid neonates. The changes observed in the a and a+ isozymes in different T3 states are summarized in Fig. 8. The solid line at 1.0 indicates the mean euthyroid abundance of the a and a+ isozymes at 30 days of age. The standard error calculated for the 30-day euthyroid values (see “Experimental Procedures”) are indicated on the plot. a and a+ abundance was restored to euthyroid levels in T3-injected hypothyroid neonates if hormonal supplements were begun in the first 3 weeks. Both a and a+ isozyme abundance in T3-treated animals were significantly lower than euthyroid controls when T3 injections were begun on or after
,,. - . . ~ . . . “””“.I.-._”_
a+ a
E E E 10 1019 19 22 22 H H
a+ a
E E E 2 5 2 5 2 8 2 8 H H H
AGE WHEN T3 RESTORED FIG. 7. Immunodetection of a and a+ isozymes in neonatal
Ts-injected hypothyroid, hypothyroid, and euthyroid brains. 25 pg of brain microsome protein from 30-day T3-injected hypothy- roid, hypothyroid, or euthyroid neonates were resolved by 7.5% SDS- polyacrylamide gels, blotted onto diazophenylthioether paper, and probed with a mixture of anti-guinea pig Na,K-ATPase antiserum (diluted 1:lOO) and anti-rat Na,K-ATPase antiserum (diluted 1:lOOO) to detect the a and a+ subunits (see “Experimental Procedures”). Numbers below represent the age at which daily T3 injections were begun. H, hypothyroid; E, euthyroid.
AGE WHEN T3 RESTORED
FIG. 8. Abundance of a and a+ isozymes in Ts-injected hy- pothyroid, hypothyroid, and euthyroid brain. Abundance of a and a+ isozymes in 30-day brains calculated by scanning immuno- blots of samples obtained from Ts-injected hypothyroid, hypothyroid (hypo), and euthyroid animals. a and a+ abundances were expressed as T3-injected hypothyroid/paired mean euthyroid values for each subunit. The solid line at 1.0 indicates the normalized mean euthyroid values. Each point is the mean f the standard error. (See “Experi- mental Procedures” for the number of samples represented by each point.) 0, a+; 0, a.

Thyroid Regulation of Na,K-ATPase in Developing Brain 17647
T
0 E H 7 10 1 6 19 22 25 28
AGE WHEN T3 RESTORED
FIG. 9. Body weight of 30-day T3-injected hypothyroid, hy- pothyroid, and euthyroid neonates. Body weights (grams, wet weight) were determined at 30 days at the time of death in four euthyroid, five hypothyroid, and one to four TB-injected hypothyroid neonates beginning hormone treatment between 7 and 28 days. Num- bers on the abscissa represent the age at which daily T3 injections were begun (hatched bars); E, euthyroid (closed bar); H , hypothyroid (open bar).
22 days of age. These results support the Na,K-ATPase activity data summarized in Fig. 6 and indicate that after 22 days Na,K-ATPase is no longer sensitive to T3 administra- tion.
Body Weight Determinations in Hypothyroid, T3-injected Hypothyroid, and Euthyroid Rats-Body weights were deter- mined on 30 days at the time of death in euthyroid, hypothy- roid, and T3-injected hypothyroid neonates receiving hormone or diluent treatment between 7 and 28 days as described under “Experimental Procedures” (Fig. 9). T3-injected hypothyroid neonates do not attain euthyroid body weights by the time of death. Body weights were significantly higher than hypothy- roid when daily T3 injections were begun before 19 days of age.
DISCUSSION
In this study we consider thyroid hormone regulation of Na,K-ATPase activity and a and a+ isozyme abundance in developing neonatal brains, and show that the thyroid respon- sive period is established as late as 15 days and ends by 22 days.
Thyroid hormone is required for normal growth and devel- opment of the central nervous system (18), and possibly functions as a biological timing mechanism for the normal sequence of brain maturation (34, 35). T3 binds to nuclear chromatin-associated, non-histone proteins in regions be- tween nucleosomes in target tissues (36-38). The binding results in a generalized increase in all classes of mRNA due to an increase in RNA polymerase I and I1 and a subsequent increase in protein synthesis (37). In addition, some mRNAs are increased or decreased directly as a result of the thyroid hormone interaction with its receptor (39, 40). In this study, we have shown that thyroid hormone causes specific altera- tions in Na,K-ATPase enzyme abundance and activity during development when normalized to a constant amount of mem- brane protein, suggesting specific effects, direct or indirect, on Na,K-ATPase synthesis. The increase in abundance might also reflect a T3-regulated decrease in degradation rate of
Na,K-ATPase peptides. However, changes in degradation rate are unlikely given the large increase in Na,K-ATPase abun- dance.
T3 regulates the sodium pump in a number of target tissues, including kidney, liver, heart, and developing brdin. T3 in- creases the synthesis of the a and @ subunits (24, 41, 42) by coordinately increasing the a and @ mRNA levels (43). In adult brain, T3 does not increase Na,K-ATPase activity (19) or abundance (20), and no measurable change in oxygen consumption is evident following T3 treatment (44). However, hypothyroidism does attenuate Na,K-ATPase activity (13, 15) and abundance (15) in neonatal brain. These observations mandate the existence of a specific period of thyroid hormone influence on brain Na,K-ATPase during development.
In this study, Na,K-ATPase activity and abundance is the same in brains of hypothyroid and euthyroid neonates before 15 days of age. Since Na,K-ATPase abundance has been shown to be regulated by ionic substrates as well as hormones (45), the increased sodium influx seen in brain cells during the first 2 weeks of life might result in up-regulation of sodium pump expression. Subsequent differentiation processes be- yond 15 days, including the increase in Na,K-ATPase abun- dance and activity, may be T3-dependent as suggested by this study. The possibility remains that during the responsive period T3 regulates Na,K-ATPase abundance indirectly by up-regulating the fluxes of ionic substrates (46).
In comparing 30-day hypothyroid to euthyroid neonates, the difference in abundance of a isoforms is greater than the difference in Na,K-ATPase activity. The reason for this dif- ference was not established in this study. The difference suggests that other factors contribute to regulation of the enzymatic activity of a given number of sodium pumps (e.g. via covalent modification).
Thyroid hormone receptor distribution in developing brain may affect the responsive period for a number of enzymes, including the sodium pump. During euthyroid maturation, T3 binding to brain nuclei receptors is highest in samples from 1- and 2-day neonates, decreasing to one-half this level by 14 days where it remains in adults (44, 47, 48). Our hypothesis, based on the results of this study, is that during the responsive period thyroid receptors are coupled to relevant stretches of DNA associated with Na,K-ATPase genes or genes whose products regulate Na,K-ATPase expression and that the receptors become uncoupled at the end of the T3 responsive period.
Thyroid hormone concentrations may also affect the re- sponsive period. T3 plasma concentrations increase rapidly after 2 days, attaining adult concentration between 11 (44) and 21 (48) days. In our study, the critical period for thyroid hormone influence on brain sodium pump is between 15 and 22 days, corresponding with a time when specific thyroid hormone receptors and T3 plasma concentrations are decreas- ing toward adult levels. In this study two-thirds of hypothy- roid neonates between 1 and 15 days had no measurable serum T3, while one-third showed very low levels of serum T3. Thus, we may have defined the latest possible starting point for the critical period because the presence of low levels of thyroid hormone in some hypothyroid neonates could induce Na,K- ATPase expression before 15 days, thus elevating enzyme abundance and activity.
Since Sweadner (7) reported two molecular forms of the O(
subunit in brain, Shull and coworkers (1) have recently iso- lated and characterized a cDNA corresponding to a third brain a mRNA designated aIII. As of yet no protein identity has been established for aIII. However, it is possible that immuno-cross-reactivity between the a isozymes is occurring

17648 Thyroid Regulation of Na,K-ATPase in Developing Brain
in our study, and some of the results attributed to a or a+ in this study may be associated with changes in aIII. Atterwill et al. (49), using differential ouabain inhibition of Na,K- ATPase activity in rat cerebellum, found kinetic evidence for a and a+ isozymes in both neuronal and glial cells, demon- strating a complex subunit distribution. The existence of isozymes presents the potential for differential regulation of a versus a+. However, in our study a and a+ isozyme abun- dances are inhibited to the same extent in hypothyroid com- pared with control, concomitant with decreases in Na,K- ATPase activity, all during the same T3 responsive period. Whether Na,K-ATPase activity and isozyme abundance are altered differentially by T3 in subsets of the brain requires confirmation.
When hypothyroid neonates were injected with thyroid hormone on sequentially later days of development, Na,K- ATPase activity and isozyme abundance were maintained at euthyroid levels if daily T3 supplements were begun on or before 19 days, but remained at hypothyroid levels when begun on or after 22 days. Our results are supported by the finding of Lindholm (50) that synaptic membrane Na,K- ATPase activity and abundance was reduced in the hypothy- roid state and increased in T4-injected euthyroid and hypo- thyroid neonates on 14 but not 30 neonatal days. In this study, hypothyroid neonates beginning daily T S injections on 7 and 10 days of age showed evidence of being slightly hyper- thyroid, as evidenced by precocious eye opening (1-2 days early) and reduced body weights. In these neonates, a and a+ abundance was greater than that of euthyroid, suggesting that hyperthyroidism increases Na,K-ATPase abundance beyond control values. Thyroid hormone toxicity prevented us from injecting hypothyroid neonates with more than 0.1 pg of T3/ g body weight, for further evaluation of the effects of induced hyperthyroidism.
Effects of malnutrition in association with hypothyroidism on Na,K-ATPase development were not controlled in this study. For example, the effects ascribed to neonatal injection of TI may at least in part be due to the resolution of iodine deficiency in the mothers (since they were returned to control water and chow), which could then affect neonatal enzyme activities by an indirect mechanism. However, Hernandez (51) has demonstrated an increase in Na,K-ATPase activity in malnourished rat brains between 15 and 30 days. Thus, effects of malnutrition in this study might diminish rather than enhance the effects of hypothyroidism on Na,K-ATPase development.
In delimiting the thyroid responsive period, two additional points need consideration. First, the lag period between the binding of T3, the increase in Na,K-ATPase mRNAs, and the increase in abundance observed in this study may alter the beginning of what we have defined as the responsive period. Another study from this lab has shown that in hypothyroid adult rat kidney, a and @ mRNA levels increase significantly by 6 h after a single injection of T3 (43). Because there is a long turnover time for kidney Na,K-ATPase (4.9 days), sub- unit abundance and Na,K-ATPase activity does not increase until 24 h after the TS (43). Thus, there is an 18-h lag between increase in mRNA and increase in enzyme. Assuming similar kinetics in neonatal brain, this alters the described thyroid responsive period by less than 1 day. Secondly, it is possible that the response to T3 requires a number of days to occur. If so, the period between T3 administration and death may be too short for a response to manifest, and we would underes- timate the end of the responsive period. Since we have shown that a single dose of T3 increases Na,K-ATPase abundance and activity by 24 h after treatment, it is unlikely that the 8-
day period between day 22 and the day of death is too short for a T3 effect to become manifest. However, if thyroid status indirectly regulates Na,K-ATPase (e.g. via nutritional status), then this is a viable possibility.
In conclusion, we suggest a general model to explain how changes in thyroid hormone receptors, T3 concentration, and morphological alterations in developing brain might account for changes in Na,K-ATPase activity and abundance during development. Brain T3 receptors and T3 plasma concentra- tions peak rapidly after birth, decreasing to attain adult concentrations between 11 and 21 days. This period corre- sponds to a period when decisive maturational effects are evident in brain, including rapid myelinogenesis and prolif- eration of axons, dendrites, and glial cells. These processes are greatly attenuated in hypothyroid brain. The T3 respon- sive increase in Na,K-ATPase activity between 15 and 22 days, observed in this study, may coincide with the large increase in axons and dendrites. During this period it is possible that thyroid hormone may regulate Na,K-ATPase isozyme synthesis directly or, alternatively, may regulate syn- thesis indirectly by increasing the fluxes of ionic substrates (e.g. by regulating sodium channels). After 22 neonatal days, associated with the decrease in brain thyroid hormone recep- tors, the receptors are no longer coupled to the relevant DNA binding sites, resulting in an uncoupling of thyroid hormone regulation of Na,K-ATPase isoform synthesis.
Acknowledgment-We would like to thank I. S. Edelman for critical reading of the manuscript.
1.
2.
3. 4.
5.
6.
7. 8.
9. 10.
11.
12.
13.
14.
REFERENCES Shull, G. E., Greeb, J., and Lingrel, J. B. (1986) Biochemistry 26,
Mercer, R. W . , Schneider, J. W., Savitz, A., Emanuel, J., Benz, E. J., and Levenson, R. (1986) Mol. Cell. Biol. 6 , 3884-3890
Iversen, L. L. (1974) Biochem. Pharmacol. 2 3 , 1927-1935 Iversen, L. L., and Kelly, J. S. (1975) Biochem. P h a r m o l . 2 4 ,
Tissari, A. H., Schonhofer, P. S., Bogdanski, D. F., and Brodie,
Riggs, T. R., Pote, K. G., Im, H. S., and Huff, D. W . (1984) J.
Sweadner, K. J. (1979) J. Biol. Chem. 264,6060-6067 Specht, S. C., and Sweadner, K. J. (1984) Proc. Natl. Acud. Sci.
McDonough, A. (1985) Erp. Eye Res. 40,667-674 Matsuda, T., Iwata, H., and Cooper, J. R. (1984) J. Bwl. Chem.
McDonough, A. A., and Schmitt, C. A. (1987) Am. J. Physwl.
Young, R. M., Shull, G. E., and Lingrel, J. B. (1987) J. Bwl.
Valcana, T., and Timiras, P. S. (1969) J. Neurochem. 1 6 , 935-
Bertoni, J. M., and Siegel, G. J. (1978) J. Neurochem. 31 , 1501-
8125-8132
933-938
B. B. (1969) Mol. Pharmacol. 5,593-604
Neurochem. 42,1251-1259
U. S. A. 81, 1234-1238
259,3858-3863
253, C862-C865
Chem. 262,4905-4910
943
1.511 15. Schmitt. C. A., and McDonough, A. A. (1986) J. Biol. Chem. 261.
" ~~
10439110444 ~~~
16. Vernadakis, A., and Woodbury, D. M. (1962) Am. J. Physiol. 203,748-752
17. Abdel-Latif, A. A., Brody, J., and Ramahi, H. (1967) J. Neuro-
18. Balazs, R., Lewis, P. D., and Patel, A. J. (1975) in Growth and Deuelopmnt of the Bruin (Brazier, M. A. B., ed) pp. 83-115, Raven Press, New York
19. Ismail-Beigi, F., and Edelman, I. S. (1971) J. Gen. Physiol. 5 2 , 710-722
20. Lin, M. H., and Akera, T. (1978) J. Biol. Chem. 263 , 723-726 21. Attenvill, C. K., Reid, J., and Athayde, C. M. (1985) Mol. Cell.
Endocrinol. 4 0 , 149-158 22. McDonough, A., and Schmitt, C. (1988) in Iodine and the Bruin
(DeLong, R., ed) Plenum Publishing Corp., New York, in press
them. 14,1133-1141

Thyroid Regulation of Na,K-ATPase in Developing Brain 17649
23. Silva, J. E., and Leonard, J. L. (1985) Endocrinology 116, 1627-
24. Jorgensen, P. L. (1974) Biochim. Biophys. Acta 356,36-52 25. Lo, C.-S., August, T. R., Liberman, U. A., and Edelman, I. S.
(1976) J. Biol. Chem. 251, 7826-7833 26. Lowry, 0. H., Rosehrough, N. J., Farr, A. L., and Randall, R. J.
(1951) J. Biol. Chem. 193,265-275 27. Laemmli, U. K. (1970) Nature 227,680-685 28. Best, D., Warr, P. J., and Gull, K. (1981) A d . Biochem. 114,
29. Sackett, D. L., Bhattacharyya, B., and Wolff, J. (1985) J. Biol.
30. McDonough, A. A., Hiatt, A., and Edelman, I. S. (1982) J. Membr.
31. Ariyasu, R. G., Nichol, J. A., and Ellisman, M. H. (1985) J.
32. McDonough, A., and Schmitt, C. (1985) Am. J. Physiol. 248,
33. Renart, J., Reiser, J., and Stark, G. R. (1979) Proc. Nutl. Acud.
34. Hamburgh, M., Lynn, E., and Weiss, E. P. (1964) Anat. Rec.
35. Hamburgh, M. (1968) Gen. Comp. Endocrinol. 10, 198-213 36. Oppenheimer, J. H. (1985) Ann. Znt. Med. 102,374-384 37. Jump, D. B., and Oppenheimer, J. H. (1983) Mol. Cell. Biochem.
1635
281-284
Chem. 260,43-45
Biol. 69,13-22
Neurosci. 5, 2581-2596
C247-C251
Sci. U. S. A. 76,3116-3120
150,147-159
55,159-176
38.
39.
40.
41.
42. 43.
44.
45.
46.
47.
48.
49.
50. 51.
Oppenheimer, J. H., Dillmann, W. H., Schwartz, H. L., and
Towle, H. C., Dillmann, W. H., Schwartz, H. L., and Oppenhei-
Dillmann, W. H., Barrieux, A., Neeley, W. E., and Contreras, P.
Lo, C.-S., and Edelman, I. S. (1976) J. Biol. Chem. 251, 7834-
Lo, C. S., and Lo, T. N. (1980) J. Biol. Chem. 255, 2131-2136 McDonough, A., Brown, T., Horowitz, B., Chiu, R., Schlotterbeck,
J., Bowen, J., and Schmitt, C. (1988) Am. J. Physiol. 254,
Schwartz, H. L., and Oppenheimer, J. H. (1978) Endocrinology
Bowen, J. W., and McDonough, A. (1987) Am. J. Physiol. 252,
Atterwill, C. K., Atkinson, D. J., Bermudez, I., and Balazs, R.
Valcana, T., and Timiras, P. S. (1978) Mol. Cell. Endocrinol. 11,
Ishiguro, K., Suzuki, Y., and Sato, T. (1980) Acta Endocrinol. 95,
Atterwill, C. K., Cunningham, V. J., and Balazs, R. (1984) J.
Lindholm, D. B. (1984) Deu. Bruin Res. 15,83-88 Hernandez, R. J. (1980) Deu. Neurosci. 3, 277-282
Towle, H. C. (1979) Fed. Proc. 38, 2154-2161
mer, J. H. (1979) Transmembr. Sign. 31,691-701
(1983) J. Biol. Chem. 258, 7738-7745
7840
C323-C329
103,943-948
C179-C189
(1985) Neuroscience 14,361-373
31-41
495-499
Neurochem. 43,8-18
Related Documents











