THE JOURNAL OP BKILOGICAL CHEMISTRY 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 265, No. 6, Issue of February 25, pp. 3474-3478,199O Printed in U.S.A. Thyroid Hormone and Dietary Carbohydrate Interact to Regulate Rat Liver S14 Gene Transcription and Chromatin Structure* (Received for publication, October 5, 1989) Donald B. Jump, Aneka Bell, and Vivian Santiago From the Department of Physiology, Cell and Molecular Biology Program, Michigan State University, East Lansing, Michigan 48824 Dietary carbohydrate and thyroid hormone (T3) in- teract to regulate rat liver S14 gene expression. The molecular basis for this interaction was examined by analysis of hepatic mRNAslr levels, S14 gene tran- scription, and chromatin structure. While starvation of euthyroid rats inhibited hepatic S 14 gene transcrip- tion r90%, sucrose administration induced mRNAslr and 514 transcription to 82% of euthyroid-fed levels within 4 h. In contrast, administration of sucrose or Ts to starved hypothyroid animals restored S14 gene transcription to only 30% of euthyroid-fedvalues. Both Ts and sucrose were required to restore S14 run-on activity and mRNAs14 to euthyroid-fed levels within 4 h. Thus, TI and sucrose interact synergistically and rapidly to induce 514 gene transcription. Analysis of 514 chromatin structure showed that starvation of hypothyroid rats inhibited the formation of three DNase I-hypersensitive sites flanking the 5’-end of the S14 gene (Hss-1 at -65 to -265 base pairs; Hss-2 at -1.2 kilobases and Hss-3 at -2.67 kilobases). The loss of these sites correlated with the repression of 514 gene transcription in starved hypothyroid rats. Whereas administration of sucrose to starved hypothy- roid rats consistently induced the Hss-1 and Hss-2 sites, T3 consistently induced all three DNase I-hyper- sensitive sites. Yet, neither treatment alone induced 514 gene transcription to euthyroid levels. The com- bination of T3 and sucrose induced no additional change in S14 chromatin structure over that induced by Ts alone. Thus, regulation of S14 chromatin struc- ture alone is not the sole mechanism by which these stimuli regulate 514 gene transcription. We speculate that the synergistic regulation of 514 gene transcrip- tion by Ta and dietary carbohydrate involves a complex interaction between factors which regulate the acces- sibility of putative c®ulatory elements through changes in chromatin structure and the regulation of “transcription factors” which interact with these ele- ments. The mRNA (mRNA& coding for the rat liver S14 protein (Mr = 17,000, p1 4.9) is subject to tissue-specific, develop- mental, hormonal, and nutritional regulation (l-9). The over- all pattern of regulation of this sequence shares many char- acteristics in common with the regulation of hepatic lipogenic enzymes and their mRNAs (9, 10). Although the function of * This work was supported by National Institutes of Health Grant GM36851. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. the S14 protein is unknown, these similarities suggest that the S14 protein plays some role in lipid metabolism (2, 7, 9). We have focused on defining the molecular basis of the multifactorial regulation of S14 gene expression by analysis of S14 gene transcription and chromatin structure. Previous studies suggest that specific loci located upstream from the 5’-end of the S14 gene harbor key &-acting elements affect- ing gene transcription (3, 6, 7, 11). For example, the Tzl induction of S14 gene transcription in hypothyroid rats is preceded by induction of a single DNase I-hypersensitive site located nearly 3 kb upstream from the S14 cap site. Based on studies of steroid-responsive genes (12-15), we suggested the TB-regulated DNase I-hypersensitive site marks the location of a thyroid hormone response element (TRE) controlling S14 gene transcription (6). Changes in nutritional status have major effects on S14 gene expression in the adult rat. Starvation reduces hepatic mRNAsll levels, while feeding animals a high carbohydrate- fat-free diet or administering sucrose induces hepatic mRNAel, (1, 4, 5, 7, 8). In contrast to euthyroid animals, mRNAsld was only marginally induced following sucrose ga- vage of starved hypothyroid rats implicating a role for T3 in the carbohydrate-mediated regulation of this gene. Although Mariash et al. (4) previously reported that TS and carbohy- drate interact synergistically to regulate hepatic levels of mRNAs14, the mechanism for this interaction is unknown. This report examines the mechanism of nutritional regu- lation of Sl4 gene expression. Activation of S14 gene tran- scription was established as the principal mechanism account- ing for the induction of mRNAsll following sucrose adminis- tration to starved euthyroid rats. Although both sucrose and T3 regulate S14 gene transcription in the starved hypothyroid rat, neither stimulus alone was sufficient to induce S14 gene transcription to euthyroid-fed levels. Induction of S14 gene transcription to euthyroid-fed values required both stimuli. Analysis of S14 chromatin structure showed that both T3 and dietary carbohydrate induce major changes in the structure of specific DNase I-hypersensitive sites flanking the S14 gene. Our findings suggest that each stimuli regulates S14 gene transcription, at least in part, through local modification of chromatin structure. MATERIALS AND METHODS Animals-Euthyroid Sprague-Dawley male rats were obtained from Charles River Breeding Laboratories. Rats were made hypothy- roid by maintenance on methimazole (0.025% in drinking water) for 4 weeks. Euthyroid animals were made hyperthyroid by injection of TS (15 rg/lOO g body weight) for 5 days (6). All animals used in this 1 The abbreviations used are: TB, thyroid hormone; kb, kilobase( bp, base pair(s); rGH, growth hormone; TRE, thyroid hormone re- sponse element. 3474 by guest on July 15, 2018 http://www.jbc.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOURNAL OP BKILOGICAL CHEMISTRY 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 6, Issue of February 25, pp. 3474-3478,199O Printed in U.S.A.
Thyroid Hormone and Dietary Carbohydrate Interact to Regulate Rat Liver S14 Gene Transcription and Chromatin Structure*
(Received for publication, October 5, 1989)
Donald B. Jump, Aneka Bell, and Vivian Santiago From the Department of Physiology, Cell and Molecular Biology Program, Michigan State University, East Lansing, Michigan 48824
Dietary carbohydrate and thyroid hormone (T3) in- teract to regulate rat liver S14 gene expression. The molecular basis for this interaction was examined by analysis of hepatic mRNAslr levels, S14 gene tran- scription, and chromatin structure. While starvation of euthyroid rats inhibited hepatic S 14 gene transcrip- tion r90%, sucrose administration induced mRNAslr and 514 transcription to 82% of euthyroid-fed levels within 4 h. In contrast, administration of sucrose or Ts to starved hypothyroid animals restored S14 gene transcription to only 30% of euthyroid-fedvalues. Both Ts and sucrose were required to restore S14 run-on activity and mRNAs14 to euthyroid-fed levels within 4 h. Thus, TI and sucrose interact synergistically and rapidly to induce 514 gene transcription. Analysis of 514 chromatin structure showed that starvation of hypothyroid rats inhibited the formation of three DNase I-hypersensitive sites flanking the 5’-end of the S14 gene (Hss-1 at -65 to -265 base pairs; Hss-2 at -1.2 kilobases and Hss-3 at -2.67 kilobases). The loss of these sites correlated with the repression of 514 gene transcription in starved hypothyroid rats. Whereas administration of sucrose to starved hypothy- roid rats consistently induced the Hss-1 and Hss-2 sites, T3 consistently induced all three DNase I-hyper- sensitive sites. Yet, neither treatment alone induced 514 gene transcription to euthyroid levels. The com- bination of T3 and sucrose induced no additional change in S14 chromatin structure over that induced by Ts alone. Thus, regulation of S14 chromatin struc- ture alone is not the sole mechanism by which these stimuli regulate 514 gene transcription. We speculate that the synergistic regulation of 514 gene transcrip- tion by Ta and dietary carbohydrate involves a complex interaction between factors which regulate the acces- sibility of putative c®ulatory elements through changes in chromatin structure and the regulation of “transcription factors” which interact with these ele- ments.
The mRNA (mRNA& coding for the rat liver S14 protein (Mr = 17,000, p1 4.9) is subject to tissue-specific, develop- mental, hormonal, and nutritional regulation (l-9). The over- all pattern of regulation of this sequence shares many char- acteristics in common with the regulation of hepatic lipogenic enzymes and their mRNAs (9, 10). Although the function of
* This work was supported by National Institutes of Health Grant GM36851. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
the S14 protein is unknown, these similarities suggest that the S14 protein plays some role in lipid metabolism (2, 7, 9).
We have focused on defining the molecular basis of the multifactorial regulation of S14 gene expression by analysis of S14 gene transcription and chromatin structure. Previous studies suggest that specific loci located upstream from the 5’-end of the S14 gene harbor key &-acting elements affect- ing gene transcription (3, 6, 7, 11). For example, the Tzl induction of S14 gene transcription in hypothyroid rats is preceded by induction of a single DNase I-hypersensitive site located nearly 3 kb upstream from the S14 cap site. Based on studies of steroid-responsive genes (12-15), we suggested the TB-regulated DNase I-hypersensitive site marks the location of a thyroid hormone response element (TRE) controlling S14 gene transcription (6).
Changes in nutritional status have major effects on S14 gene expression in the adult rat. Starvation reduces hepatic mRNAsll levels, while feeding animals a high carbohydrate- fat-free diet or administering sucrose induces hepatic mRNAel, (1, 4, 5, 7, 8). In contrast to euthyroid animals, mRNAsld was only marginally induced following sucrose ga- vage of starved hypothyroid rats implicating a role for T3 in the carbohydrate-mediated regulation of this gene. Although Mariash et al. (4) previously reported that TS and carbohy- drate interact synergistically to regulate hepatic levels of mRNAs14, the mechanism for this interaction is unknown.
This report examines the mechanism of nutritional regu- lation of Sl4 gene expression. Activation of S14 gene tran- scription was established as the principal mechanism account- ing for the induction of mRNAsll following sucrose adminis- tration to starved euthyroid rats. Although both sucrose and T3 regulate S14 gene transcription in the starved hypothyroid rat, neither stimulus alone was sufficient to induce S14 gene transcription to euthyroid-fed levels. Induction of S14 gene transcription to euthyroid-fed values required both stimuli. Analysis of S14 chromatin structure showed that both T3 and dietary carbohydrate induce major changes in the structure of specific DNase I-hypersensitive sites flanking the S14 gene. Our findings suggest that each stimuli regulates S14 gene transcription, at least in part, through local modification of chromatin structure.
MATERIALS AND METHODS
Animals-Euthyroid Sprague-Dawley male rats were obtained from Charles River Breeding Laboratories. Rats were made hypothy- roid by maintenance on methimazole (0.025% in drinking water) for 4 weeks. Euthyroid animals were made hyperthyroid by injection of TS (15 rg/lOO g body weight) for 5 days (6). All animals used in this
1 The abbreviations used are: TB, thyroid hormone; kb, kilobase( bp, base pair(s); rGH, growth hormone; TRE, thyroid hormone re- sponse element.
3474
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
T3 and Dietary Carbohydrate Regulate Hepatic S14 Gene Transcription
study ranged from 3 to 4 months of age and were killed between 0800 and 1200 h by ether anesthesia and exsanguination.
DNA Probes-All DNA probes, with the exception of pEMBLS14- 0.7P, were described previously (3,6). The pEMBLS14-0.7P plasmid was subcloned from a genomic clone (pEMBLS14-13E). The genomic clone was obtained from a size-selected (EcoRI) rat liver genomic library screened with a S14 5’-exon probe. The sequences in pEMBLS14-0.7P extend from -3350 to -4010 bp relative to the 5’- end of the S14 gene. The cDNA and genomic probes were labeled with [32P]dCTP using either nicked translation or primed synthesis (6).
Measurement of Hepatic mRNAs,,-Total hepatic RNA was ex- tracted from livers as described by Chirgwin et al. (16), and hepatic mRNAsll expression was measured by dot-blot hybridization and quantified (6). Results are expressed as units of mRNAsla (1 unit = 4.8 X 10’ copies of mRNAs,,/pg of total RNA).
Analysis of S14 Gene Activity and Chromatin Structure-Nuclei were isolated and used for gene transcription and chromatin studies (6).
DNA Sequence Analysis of the DNA Flanking the 5’-End of the SI4 Gene-The pS14-6.8 genomic plasmid contains S14 genomic DNA extending from +1.7 kb to -5.2 kb relative to the 5’-end of the S14 gene. Portions of this plasmid were restricted and subcloned into pGEM-1 or pGem-blue vectors (Promega). A series of additional subclones were generated by deletion from either end by unidirec- tional digestion with exonuclease III (17). DNA sequencing was performed by the dideoxy chain termination method of Sanger (18) using [““S]dATP and Tq DNA polymerase (Sequenase; U.S. Biochem- icals). All portions of the sequence were determined independently in both directions from overlapping clones. Sequence data were melded and analyzed using computer programs developed by Deveruex et al. (19).
RESULTS
Nutritional Status Regulates Hepatic S14 Gene Transcrip- tion-when compared to levels of S14 expression in euthy- roid-fed animals, starvation of euthyroid rats for 48 h re- pressed both hepatic mRNAslr and in vitro transcriptional run-on activity by 290% (Fig. 1). Following sucrose admin- istration, a l-h lag period precedes the 35 and 22-fold induc- tion of hepatic mRNAsi4 and S14 run-on activity, respectively, that is observed at 4 h. S14 gene expression was restored to ~80% of euthyroid-fed values within 4 h. During this short term experiment, we found that activation of S14 gene tran- scription accounted for nearly the full induction of mRNAsi.,
FIG. 1. Dietary carbohydrate regulates hepatic 514 gene transcription and mRNAs14 abundance. Male chow-fed euthyroid animals were maintained on rat chow and water ad Zibitum. Euthyroid animals were starved for 48 h, but were provided water ad libitum. Starved rats were administered sucrose (1 ml/100 g body weight; 60%) via gavage and killed at 1 or 4 h afterward. Vehicle-treated (control) animals received water by the same route and were killed 4 h after treatment. All animals were killed between 1000 and 1200 h. Livers were removed for extraction of RNA and isolation of nuclei. Meas- urement of relative abundance of mRNAsll and S14 transcriptional run-on activity was described under “Materials and Methods.” Data are expressed as mean + S. E., and the number of animals at each point ranged from four to six. q l, mRNAsi, expression in units (1 unit = 4.8 X 10’ copies of mRNA&pg of total RNA); I, S14 transcription run-on activity in ppm.
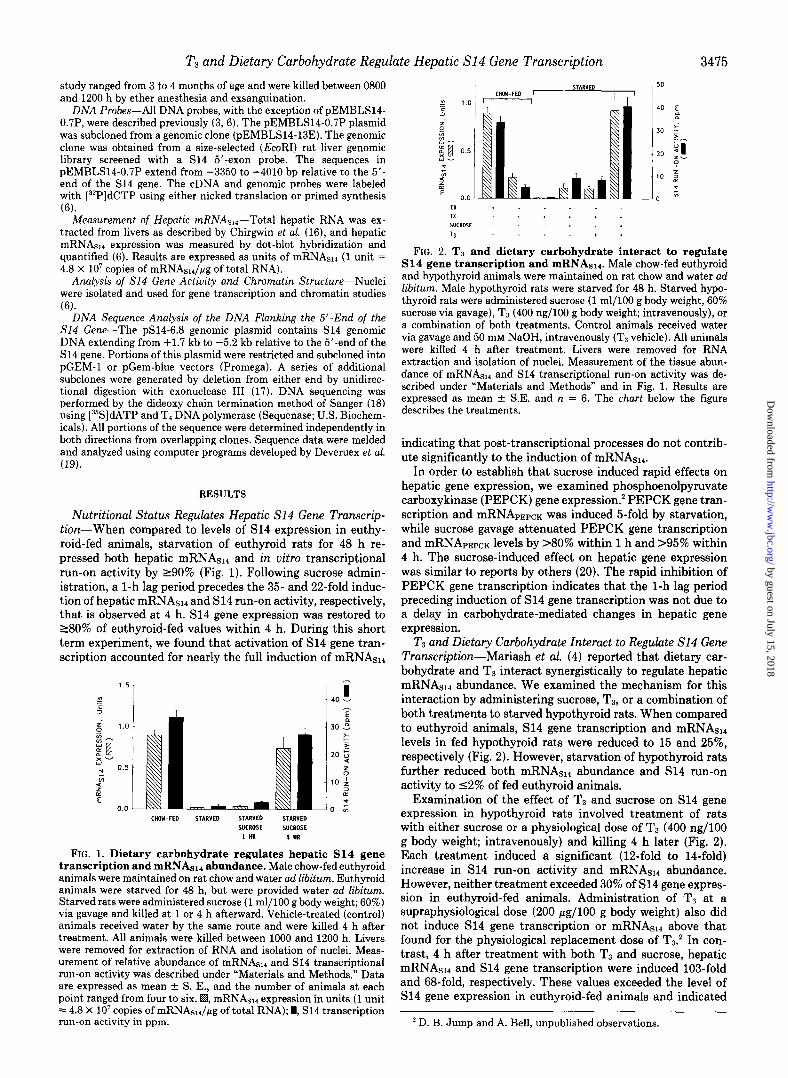
FIG. 2. TB and dietary carbohydrate interact to regulate S14 gene transcription and mRNAslr. Male chow-fed euthyroid and hypothyroid animals were maintained on rat chow and water ad libitum. Male hypothyroid rats were starved for 48 h. Starved hypo- thyroid rats were administered sucrose (1 ml/100 g body weight, 60% sucrose via gavage), T, (400 ng/lOO g body weight; intravenously), or a combination of both treatments. Control animals received water via gavage and 50 mM NaOH, intravenously (T3 vehicle). All animals were killed 4 h after treatment. Livers were removed for RNA extraction and isolation of nuclei. Measurement of the tissue abun- dance of mRNAs14 and S14 transcriptional run-on activity was de- scribed under “Materials and Methods” and in Fig. 1. Results are expressed as mean + S.E. and n = 6. The chart below the figure describes the treatments.
indicating that post-transcriptional processes do not contrib- ute significantly to the induction of mRNAsil.
In order to establish that sucrose induced rapid effects on hepatic gene expression, we examined phosphoenolpyruvate carboxykinase (PEPCK) gene expression.’ PEPCK gene tran- scription and mRNApEpCK was induced 5-fold by starvation, while sucrose gavage attenuated PEPCK gene transcription and mRNApapck levels by >80% within 1 h and >95% within 4 h. The sucrose-induced effect on hepatic gene expression was similar to reports by others (20). The rapid inhibition of PEPCK gene transcription indicates that the l-h lag period preceding induction of Sl4 gene transcription was not due to a delay in carbohydrate-mediated changes in hepatic gene expression.
T3 and Dietary Carbohydrate Interact to Regulate S14 Gene Transcription-Mariash et al. (4) reported that dietary car- bohydrate and T3 interact synergistically to regulate hepatic mRNAsll abundance. We examined the mechanism for this interaction by administering sucrose, TB, or a combination of both treatments to starved hypothyroid rats. When compared to euthyroid animals, S14 gene transcription and mRNAs,, levels in fed hypothyroid rats were reduced to 15 and 25%, respectively (Fig. 2). However, starvation of hypothyroid rats further reduced both mRNAsl, abundance and S14 run-on activity to 12% of fed euthyroid animals.
Examination of the effect of TB and sucrose on S14 gene expression in hypothyroid rats involved treatment of rats with either sucrose or a physiological dose of Ts (400 ng/lOO g body weight; intravenously) and killing 4 h later (Fig. 2). Each treatment induced a significant (12-fold to 14-fold) increase in S14 run-on activity and mRNAs14 abundance. However, neither treatment exceeded 30% of S14 gene expres- sion in euthyroid-fed animals. Administration of T1 at a supraphysiological dose (200 pg/lOO g body weight) also did not induce S14 gene transcription or mRNASi4 above that found for the physiological replacement dose of T3.’ In con- trast, 4 h after treatment with both TB and sucrose, hepatic mRNAsi4 and S14 gene transcription were induced 103-fold and 68-fold, respectively. These values exceeded the level of S14 gene expression in euthyroid-fed animals and indicated
’ D. B. Jump and A. Bell, unpublished observations.
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

3476 Tri and Dietary Carbohydrate Regulate Hepatic S14 Gene Transcription
that T:i and dietary carbohydrate act rapidly and synergisti- cally to regulate hepatic mRNAsl., abundance at the transcrip- tional level.
Roth T:, and Dietary Carbohydrate Regulate S14 Chromatin Structure-In an effort to gain additional insight into the T:< and dietary carbohydrate regulation of Sl4 gene transcription, we examined the chromatin organization of the S14 gene. Previous studies from our laboratory showed that changes in S14 chromatin structure accompanied activation of SI4 gene transcription induced by T:< or during postnatal development (3, 6, 7). A genomic probe (pEMBLS14-0.7P) was used to examine S14 chromatin structure. We mapped the location of the TYi-inducible DNase I-hypersensitive site and determined whether nutritional manipulation induced changes in S14 chromatin structure.
Fig. 3 shows the location of three major DNase I hypersen- sitive sites flanking the 5’-end of the S14 gene: Hss-1 (-65 to -265 bp), Hss-2 (-1.2 kb), and Hss-3 (at -2.67 kb). Hss-3’ is a minor site at -1.8 kb. As previously reported, the DNase I sensitivity of the Hss-3 site is the only site significantly affected by a change in thyroidal status and a lo-fold change in S14 gene transcription (6). The Hss-3 site is located 2.7 kb from the S14 gene cap site. This location is closer to the S14
Hypothyroid I
Hyperthyroid
I I / T
ECORI
J--l- ,/’ 1 sl 23452345 I
Probe
lBamHl
Fro. 3. Localization of a Ts-inducible DNase I-hypersensi- tive site. Hepatic nuclei isolated from hyperthyroid and hypothyroid rats were digested with DNase I and extracted for DNA as described under “Materials and Methods.” Isolated DNA (25 pg) was digested with EcoRI and BamHI and electrophoretically separated in 1% agarose-Tris-borate-EDTA gels. The DNA was denatured and trans- ferred to Zetabind. Blots were hybridized with ‘“P-labeled nick- translated insert derived from the pEMBLSl4-0.7P plasmid. The insert contains sequences from -3350 to -4010 bp upstream from the 5’-end of the S14 gene (see probe location on figure). The RamHI site is located at -4314 bp from the 5’-end of the Sl4 gene. Following hybridization and washing, blots were exposed to x-ray film for 3 days. Lane S, DNA standards, X/Hind111 and 4X174/HaeIII. Lanes I, 2, 3, il, and 5 represent 0, 0.1, 0.2, 0.5, and 1.0 unit of DNase I/A260. The location of the DNA standards and the sizes are shown in the figure. The distance of each DNase I-hypersensitive site relative to the HamHI site is: Hss-1 at 4.2 kb. Hss-2 at 3.1 kb. Hss-3’ at 2.5 kb. and Hss-3 at 1.7 kb. These values correspond to -65 to -265 bp, -1 kb, -1.8 kb, and -2.6 kb for the distance of Hss-1, -2, -3’, and -3 from the j’-end of the S14 gene, respectively. Gene map, the thin vertical [ine represents the 6.5.kb BarnHI-EcoRI genomic fragment. The location of each DNase I-hypersensitive site upstream from the S14 gene is denoted bv brackets. The arrow: marks the location of the S14 cap site and the direction of transcription. The sequence homol- ogy between the insert from pEMBLS14-0.7P and the S14 5’.flanking DNA is indicated by the cross-hatched box. This blot is representative of four separate studies.
gene than previously reported (6), and we attribute this dis- crepancy to the inaccuracy of determining the size of large restriction fragments.
Since steroid-inducible DNase I-hypersensitive sites are located at or near hormone response elements (12-15), the DNA sequence in the vicinity of the Hss-3 site was examined for homology to the rat growth hormone TRE (Fig. 44). The region extending from -160 to -200 bp relative to the growth hormone gene (Fig. 4B) has been reported to harbor TRE(s) and to bind salt-extracted TTr receptors (21-24). Comparison of these sequences shows the presence of an element with significant homology (77%) to the growth hormone TRE within the boundaries of the Hss-3 site (Fig. 4B). The region of highest homology corresponds to where Glass et al. (22,23) reported binding of salt-extracted T3 receptors to the growth hormone promot,er. These findings suggest that T,? receptors may bind to the putative TRE within the Hss-3 site.
Does sucrose regulate the structure of the Ta-inducible site or other DNase I-hypersensitive sites flanking the S14 gene? Starvation of euthyroid (7) and hypothyroid rats (Fig. 5) inhibits the formation of the Hss-1, Hss-2, and Hss-3 sites. Two DNase I-hypersensitive sites located at -5.3 kb (Hss-4) and -6.2 kb (Hss-5) were only marginally affected by changes in nutritional status.’ Thus, starvation effects on S14 chro- matin structure appear to be restricted to a region within 3 kb of the Sl4 gene. Loss of the DNase I-hypersensitive sites in this 3-kb region correlated with nearly full inhibition of S14 gene transcription.
Administration of sucrose to starved hypothyroid rats con- sistently induced the DNase I sensitivity of the Hss-1 and Hss-2 sites to levels equal to or greater than that seen in
A. DNA SEQUENCE WITHIN THE HSS-3 DNase I HYPERSENSITIVE SITE.
B.
-2801 CTAGACACTAAGTGACATATGTTCATATAGATAGTGTTTTTAAACTGGAA
-2751 AGACTGTCCCTACCTTGGTTTGTAGCCACCTGGGCTGAGGGCCAGCTGAG
-2701 GTTACTTGGGGCCTGGCAGCCTGGGTACCACTTGCTCATCTTACCCTTGG
-2651 ATCACATTCACATGATGTTCTGGCCAGGCCCTTGACCCCCCAGTTCCTCTGG
-2601 GGCCAATTCAATTTGTCCAGCCTAGGGCATTCTGGGTGAGGTTATTCCAA
-2551 AAGATGTGTCAGCCAACCCTCCGTTTCTCCACAGGGTTGTAGTCATGGAA
-2501 AGACTAGAGAAAGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGT
SEQUENCE COMPARISON BETWEEN THE 514 AND GROWTH HORMONE GENES.
s14: -2647 AATTCACATGATGTTCTGGCCAGGCCCTTGPLCCCAGTTCC -2607 ::: : .... . . . . : ::::: :::
TGH: -200 CGGCGGTGGAAAGGTAAGATCAGGGACGTGACCGCAGGAGA -160 : F00tl)rl”t ]
FIG. 4. Location of a putative thyroid hormone response element (TRE) upstream from the S14 gene. A, DNA sequence analysis of the Hss-3 region. DNA sequence analysis was performed as described under “Materials and Methods.” A computer-assisted homology search of the 4314 bp of DNA flanking the B/-end of the S14 gent revealed a single sequence resembling the growth hormone TRE. This sequence was found within the Hss-3 DNase I-hypersen- sitive site region. The figure shows only DNA sequence extending from -2801 to -2452 bp relative to the 5’.end of the S14 gene. The sin&e underlined sequences estimate the boundaries of DNase I hy- persensitivity. The double underlined regions correspond to the se- ouences showing high homologv to the rGH-TRE. The seauences in dolcf face are homologous to r??H-TRE. B, sequence comparison of the S14 and rGH-TRE region. A 40.bp region from the rGH gene (-160 to -200 bp) and the rat S14 gene (-2607 to -2647 bp) are aligned. Sequences showing homology are in bold face. The region labeled as Footprint corresponds to the region which confers Tg control to a heterologous promoter and where salt-extracted TR recep- tors bind to rhe growth hormone promoter (22).
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

T:, and Dietary Carbohydrate Regulate Hepatic S14 Gene Transcription 3477
Chow-fed _- Euthyrdid
scription and a slow effect at the post-transcriptional level to Starved Hypothyroid ECORI regulate S14 gene expression.
The sucrose induction of Sl4 gene transcription in the
12342345234523452345 ’
/ I starved euthyroid rat follows a patt.ern described for glucoki-
11 //i,
nase and fatty acid synthase (25-28). Refeeding starved rats rapidly induced hepatic glucokinase and fatty acid synthase gene transcription through an apparent insulin-dependent mechanism. Although our preliminary studies show that in- sulin rapidly induces Sl4 gene transcription in the strepto- zotocin-induced diabetic rat,” the precise role of insulin in the regulation of S14 and perhaps other carbohydrate-responsive genes is not clear. Volpe and Vagelos (32) reported that induction of fatty acid synthase was not a direct effect of insulin, since administration of fructose to diabetic rats in- duced full enzyme activity. Mariash and Oppenheimer (33) and Mariash and Schwartz (34), reported that specific inter-
FIG. 5. T3 and dietary carbohydrate interact to regulate S14 chromatin structure. Hepatic nuclei from euthyroid rats or
mediary metabolites induced mRNAsl.r and malic enzyme in
from starved hypothyroid rats receiving vehicle. sucrose, T:;, or both hepatocyte cultures. Thus, insulin may regulate the cellular
T.; and sucrose were digested with DNase I and processed as described abundance of a specific mediator generated from intermediary in Fig. :3. The details of the physiological manipulation of the starved metabolism rather than a plasma membrane-derived second hypothyroid rats are described in Fig. 2. Blots were exposed to x-ray messenger. The recent report by Kinlaw et al. (35) showing film for 5 davs. The figure is a composite photograph and is repre- sentative of f:ve separate studies.
that administration of glucagon rapidly inhibits S14 gene expression implicates elevated hepatic CAMP as a suppressor
euthyroid-fed animals, while the Hss-3 site was only margin- of S14 gene transcription in the starved rat. From these
ally induced (Fig. 5). The induction of these sites correlated studies, it appears that manipulation of nutritional status will
with the 15.5fold induction of S14 gene transcription. Both set in motion a complex array of regulatory pathways affecting
the pattern of chromatin structure and the transcription rate Sl4 gene transcription. The l-h lag period preceding activa-
in these animals is similar to that found in hypothyroid-fed tion of S14 gene transcription may reflect the accumulation
rats (Figs. 2 and 3). of specific protein(s) or non-protein mediator(s) or the decline
In fed hypothyroid rats, TIi regulates the structure of one in hepatic CAMP levels. Our studies do not allow us to
major DNase I-hypersensitive site and induces a lo-fold in- distinguish between these various possibilities. Thus, addi-
crease in S14 gene transcription (Fig. 3 and Ref. 6). In tional studies will be required to establish the pathway for
contrast, T:, administration to starved hypothyroid rats con- nutritional control of S14 gene transcription.
sistently induced the DNase I sensitivity of the Hss-1, Hss-2, Despite these limitations, our studies show that changes in
and Hss-3 sites to levels comparable to that found in euthy- nutritional status, like changes in thyroidal status, induce
roid-fed rats (Fig. 5). While the formation of these sites rapid changes in chromatin structure at discrete loci upstream
correlated with a 14-fold induction of S14 gene transcription, from the S14 gene. Starvation of hypothyroid rats promotes
this treatment did not restore S14 gene transcription to a loss of three major DNase I-hypersensitive sites flanking
euthyroid-fed levels (Fig. 2). Both TZi and sucrose were re- the S14 gene, and sucrose administration induces two of these sites within 4 h. The induction of these sites correlates with
quired to restore S14 gene transcription to euthyroid-fed levels; however, this treatment induced no further change in
a X-fold induction of Sl4 gene transcription. Since the ki-
S14 chromatin structure over that observed with T, alone. netics of induction of these sites was not examined, we do not
Thus, regulation of chromatin structure alone is not the sole know whether the change in Hss-1 and Hss-2 structure is a
mechanism by which these stimuli regulate S14 gene tran- pre- or co-transcriptional event. However, such a highly lo-
scription. calized change in structure accompanying gene activation suggests the regulation of these structures may be of physio-
DISCUSSION logical importance to the carbohydrate-mediated regulation of Sl4 gene transcription. These carbohydrate-regulated sites
While there are several examples of nutritional regulation may mark the location of carbohydrate-responsive &-acting of hepatic gene transcription, examples of nutritional effects elements. Additional studies will be required to define how on chromatin structure are rare (25-31). In this report, we these elements function in the nutritional control of S14 gene provide evidence for site-specific changes in S14 chromatin transcription and how nuclear factors regulate the structure structure induced by changes in nutritional status. of these sites.
Sucrose administration to starved hypothyroid and euthy- In hypothyroid-fed rats, TZi regulates the structure of a roid rats induces S14 gene transcription (Figs. 1 and 2). single DNase I-hypersensitive site (Fig. 3 and Ref. 6). This Within the 4-h interval examined, we find no evidence for the site is located 2.67 & 0.1 kb upstream from the 5’-end sucrose-mediated induction of mRNAsla through activation of the Sl4 gene (Fig. 3) and contains a sequence of a post-transcriptional process. Dozin et al. (29) reported ((‘““‘CAGGCCCTTGACC-‘““) showing high homology to a that feeding rats a high carbohydrate-fat-free diet induced rat pituitary growth hormone TRE (Fig. 4B). T:, induces this hepatic malic enzyme mRNA levels by augmenting RNA site within 5 min of hormone administration, an event that stabilization. We have found that feeding euthyroid rats a precedes the TZl-mediated activation of S14 gene transcription similar diet for 5 days increased hepatic mRNAs,, levels 4- (6). The rapid induction of this site and the presence of a fold while not significantly increasing S14 gene transcription putative TRE supports the view that the Hss-3 site marks the above the control euthyroid levels.” These studies suggest that location of the primary target of T:, on the S14 gene. Whether dietary carbohydrate induces a rapid effect on S14 gene tran- thi.; site functions as a T:: receptor binding site and a distal
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

3478 T3 and Dietary Carbohydrate Regulate Hepatic S14 Gene Transcription
enhancer regulating S14 gene transcription requires func- tional analysis of the S14 gene regulatory elements.
Surprisingly, T3 induced three DNase I-hypersensitive sites in starved hypothyroid rats indicating that T3 effects on S14 chromatin structure may not be restricted to a single locus as previously reported (6). The effect of T3 on the Hss-1 and Hss-2 sites is apparent only in the starved animal, since changes in plasma T3 levels had no effect on these structures in hypothyroid-fed animals (Fig. 3 and Ref. 6). Dietary car- bohydrate regulates these same structures. Thus, chow feeding apparently is sufficient to maintain the DNase I sensitivity of these structures in the hypothyroid-fed rat. It is not clear from our studies whether these structures are regulated by distinct T, and carbohydrate-mediated pathways. Although, we have not found high sequence homology to the growth hormone TRE in the vicinity of the Hss-1 and Hss-2 sites,’ we cannot exclude the possibility that T1 receptors interact with DNA at these sites. Alternatively, TS may influence the structure of the Hss-1 and Hss-2 sites by regulating the abundance of specific trans-acting factors binding to cis- regulatory elements. Such hormone-regulated factors have been reported to bind to the Hss-1 site of the S14 gene (36) and several sites flanking the malic enzyme gene (37). Thus, some actions of Ta on gene transcription may not be restricted to T3 receptors bound to &-linked hormone response ele- ments.
Finally, we have established that the interaction between T3 and dietary carbohydrate to regulate hepatic mRNAslh is directed at the transcriptional level. Although each factor induces changes in chromatin structure which accompany changes in S14 gene transcription, regulation of chromatin structure alone does not appear to be the sole mechanism by which these physiological stimuli affect S14 gene transcrip- tion. For example, T3 induced three major sites in the starved hypothyroid rat without fully activating S14 gene transcrip- tion. The combined treatment of T3 and sucrose induced S14 gene transcription to euthyroid-fed levels, but did not induce significant changes in S14 chromatin structure over that induced by TS alone (Figs. 2 and 5). These studies suggest TS and dietary carbohydrate regulate the activity and/or abun- dance of ancillary factors (“transcription factors”) which function in Sl4 gene transcription, but do not affect S14 chromatin structure. We speculate that the synergistic regu- lation of Sl4 gene transcription by T3 and dietary carbohy- drate requires the regulation of the accessibility of putative &-regulatory elements (chromatin structure effects) and the regulation of factors which interact with these elements (tran- scription factors). The nature of these factors is unknown.
Acknowledgments-We wish to thank Drs. R. Hanson, Case West- ern University, for the pPEPCK-10 cDNA probe used to measure phosphoenolpyruvate carboxykinase gene transcription and mRNApapcx levels. We also wish to thank Gerald Lepar and Ormond MacDougald of the Physiology Department at Michigan State Uni- versity and Steven Clarke of the Upjohn Co. for critical review of this manuscript.
REFERENCES
1. Jump, D. B., Narayan, P., Towle, H., and Oppenheimer, J. H. (1984) J. Biol. Chem. 259,2789-2797
6. 7.
8.
9.
10.
11.
12. 13. 14.
15.
16.
17. 18.
19.
20.
21.
22.
23. 24.
25.
26.
27.
28.
29.
30.
31. 32.
33.
34.
35.
36.
37.
Jump, D. B., and Oppenheimer, J. H. (1985) Endocrinology 117, 2259-2266
Jump, D. B., Veit, A., Santiago, V., Lepar, G., and Herberholz, L. (1988) J. Biol. Chem. 263, 7254-7260
Mariash, C. N., Seelig, S., Schwartz, H. L., and Oppenheimer, J. H. (1986) J. Biol. Chem. 261,9583-9586
Carr, F. E., Bingham, C., Oppenheimer, J. H., Kistner, C., and Mariash. C. N. (1984) Proc. Natl. Acad. Sci. U. S. A. 81. 974- 978 ’
Jump, D. B. (1989) J. Biol. Chem. 264,4698-4703 Jump, D. B. (1989) in Gene Regulation by Steroid Hormones IV
(Roy, A., and Clark, J., eds) Springer-Verlag, New York Inc., New York
Clarke, S. D., Armstrong, M. K., and Jump, D. B. (1990) J. Nutr., in press
Oppenheimer, J. H., Schwartz, H. L., Mariash, C. N., Kinlaw, W. B., Wong, N. C. W., and Freake, H. C. (1987) Endocrine Reu. 8.288-307
Mariash. C. N.. Kaiser. F. E.. Schwartz. H. L.. Towle. H. C.. and Oppenheimer, J. H. (1986)‘J. Clin. Zniest. 65, 1126-1134’
Jump, D. B., Wong, N. C. W., and Oppenheimer, J. H. (1987) J. Biol. Chem. 262, 778-784
Burch, J. B. E., and Weintraub, H. (1983) Cell 33,65-76 Zaret. K. S.. and Yamamoto, K. R. (1984) Cell 38, 29-38 Jantzen, H.‘M., Strahle, U.,‘Gloss, B., Stewart, F., Schmid, W.,
Boshart, M., Miksicek, R., and Schutz, G. (1987) Cell 49, 29- 38
Hecht. A.. Berkenstam. A.. Stromstedt. P.-E., Gustafasson. J.-A.. and’sipple, A. (1988) EiBO J. 7, 2663-2473
Chirawin. J. M.. Przvbvla. A. E.. MacDonald. R. J.. and Rutter. WrJ. (1979) l%och&&ry l&,5294-5299 '
Henikoff, S. (1984) Gene (Am&.) 28,351-359 Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.
Acad. Sci. U. S. A. 74, 5463-5467 Devereux, J., Haeberli, P., and Smithies, 0. (1984) Nucleic Acids
Res. 12,387-395 Lamers. W. H.. Hanson. R. W.. and Meisner, H. M. (1982) Proc.
Natl. &ad. &i. U. S. A. 79,5137-5141 Brent, G. A., Larsen, P. R., Harney, J. W., Koenig, R. J., and
Moore, D. D. (1989) J. Biol. Chem. 264,178182 Glass. C. K.. France. R.. Weinbereer. C.. Albert. V. R.. Evans. R.
M.,‘and Rosenfeld, Ml G. (1987j iat&e 329; 738-741 Evans, R. M. (1988) Science 240,889-895 Glass, C. K., Holloway, J. M., Devary, 0. V., and Rosenfeld, M.
G. (1988) Cell 54,313-323 Iynedjian, P. B., Ucla, C., and Mach, B. (1987) J. Biol. Chem.
262,6032-6038 Ivnediian. P. B.. Giinovci. A., and Renold. A. E. (1988) J. Biol.
” Chem. i63,740~744 Paulauskis. J. D.. and Sul. H. S. (1988) J. Biol. Chem. 263,7049-
7054 ’ ’ ’ Paulauskis, J. D., and Sul, H. S. (1989) J. Biol. Chem. 264,574-
577 Dozin, B., Rall, J. E., and Nikodem, V. M. (1986) Proc. Natl.
Acad. Sci. U. S. A. 83,4705-4709 Strobl, W., Gorder, N. L., Fienup, G. A., Lin-Lee, Y.-C., Gotto,
A. M., Jr., and Patsch, W. (1989) J. Biol. Chem. 264, 1190- 1194
Castro, C. E., and Sevall, J. S. (1980) J. N&r. 110, 105-116 Volpe, J. J., and Vagelos, P. R. (1974) Proc. Natl. Acad. Sci.
U. s. A. 71,889-893 Mariash. C. N.. and Gunenheimer. J. H. (1984) Metabolism. 33,
545-552 - - Mariash, C. N., and Schwartz, H. L. (1986) Metabolism 35,452-
456 Kinlaw, W. B., Schwartz, H. L., Towle, H. C., and Oppenheimer,
J. H. (1986) J. Clin. Znuest. 78. 1091-1096 Wong, N. C. W., Perez-Castillo, A. M., Sanders, M. M., Schwartz,
H. L.. and GDDenheimer. J. H. (1989) J. Biol. Chem. 264, 4466-4470 - -
Petty, K. J., Morioka, H., Mitsuhashi, T., and Nikodem, V. M. (1989) J. Biol. Chem. 264, 11483311490
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from

D B Jump, A Bell and V Santiagotranscription and chromatin structure.
Thyroid hormone and dietary carbohydrate interact to regulate rat liver S14 gene
1990, 265:3474-3478.J. Biol. Chem.
http://www.jbc.org/content/265/6/3474Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/6/3474.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 15, 2018http://w
ww
.jbc.org/D
ownloaded from
Related Documents











