Three Neuropeptide Y Receptor Genes in the Spiny Dogfish, Squalus acanthias, Support en Bloc Duplications in Early Vertebrate Evolution Erik Salaneck,* David H. Ardell, Earl T. Larson,* and Dan Larhammar* *Department of Neuroscience, Unit of Pharmacology, and Department of Molecular Evolution, Evolutionary Biology Center, Uppsala University, Uppsala, Sweden It has been debated whether the increase in gene number during early vertebrate evolution was due to multiple independent gene duplications or synchronous duplications of many genes. We describe here the cloning of three neuropeptide Y (NPY) receptor genes belonging to the Y1 subfamily in the spiny dogfish, Squalus acanthias, a cartilaginous fish. The three genes are orthologs of the mammalian subtypes Y1, Y4, and Y6, which are located in paralogous gene regions on different chromosomes in mammals. Thus, these genes arose by duplications of a chromosome region before the radiation of gnathostomes (jawed vertebrates). Estimates of duplication times from linearized trees together with evidence from other gene families supports two rounds of chromosome duplications or tetraploidizations early in vertebrate evolution. The anatomical distribution of mRNA was determined by reverse- transcriptase PCR and was found to differ from mammals, suggesting differential functional diversification of the new gene copies during the radiation of the vertebrate classes. Introduction Vertebrate genomes contain considerably more genes than most invertebrate genomes. Ohno (1970) and others have proposed that this difference arose through early vertebrate tetraploidizations. Gene mapping data support gene duplications involving large chromosomal regions or entire chromosomes (Lundin 1993; Popovici et al. 2001a, 2001b). Such sets of paralogous chromosomal regions are called paralogons (Coulier et al. 2000). The growing number and sizes of paralogons that have been identified in vertebrate genomes suggest that there was an expansion during early vertebrate evolution via two tetraploidizations (Holland et al. 1994; Kasahara et al. 1996; Sidow 1996). This hypothesis is called the one-to-four rule (Ohno 1999), or the 2R hypothesis, for two rounds of genome doubling (Hughes 1999). Despite the evidence, the 2R hypothesis remains controversial (Hughes 1999; Hughes, da Silva, and Friedman 2001; Martin 2001; Pennisi 2001). Alternative hypotheses include independent gene duplications fol- lowed by parallel aggregation into similar clusters (Hughes 2000) or independent chromosome or block duplications rather than simultaneously in tetraploidizations (Skrabanec and Wolfe 1998; Smith, Knight, and Hurst 1999). The arguments are discussed in recent reviews (Wolfe 2001; Larhammar, Lundin, and Hallbo ¨o ¨k 2002). Another issue is when the duplications took place. Several authors have suggested that they occurred before the origin of gnathostomes but after amphioxus branched off (Holland et al. 1994; Sidow 1996; Spring 1997). This is supported by the presence of a single Hox cluster in amphioxus as opposed to four in tetrapods (Garcia- Ferna `ndez and Holland 1994). This is also true for the large chromosomal region carrying the major histocom- patibility complex (Abi-Rached et al. 2002). After the divergence of amphioxus and before the appearance of gnathostomes, the agnathan (jawless fishes) hagfishes and lampreys appeared, either independently or as a mono- phyletic group (Delarbre et al. 2002). It has been proposed that the first tetraploidization took place before the agnathans diverged and that the second tetraploidization happened subsequently in the lineage leading to gnatho- stomes (Holland et al. 1994). This interpretation is com- plicated by the discovery of three to four Hox clusters in lampreys, but these clusters may have occurred indepen- dently (Force, Amores, and Postlethwait 2002; Irvine et al. 2002; Sharman and Holland 1998). The time point for the second proposed tetraploidiza- tion is also uncertain, as only two Hox clusters have yet been identified in any cartilaginous fish (Kim et al. 2000). A recent review placed the second tetraploidization after divergence of sharks (Pennisi 2001). Regardless, cartilag- inous fishes and agnathans are important groups for determining time points and mechanisms for the increase in vertebrate gene number. We have previously investigated the evolution of the neuropeptide Y (NPY) family of neuroendocrine peptides in vertebrates and found that the first gene duplication leading to NPY and peptide YY (PYY) probably took place before the origin of lampreys, as both the river lamprey and gnathostomes have NPY and PYY (Cerda ´- Reverter and Larhammar 2000; Larhammar 1996; So ¨der- berg et al. 1994). More recently, we have described an NPY receptor in the river lamprey Lampetra fluviatilis that appears to be a pro-ortholog of the tetrapod receptor Y4 and the teleost receptor Yb (Salaneck et al. 2001). Both NPY peptide and receptor genes belong to paralogons that have likely arisen by duplications in early vertebrate evolution, as the genes for NPY and PYY are linked to Hox clusters (Larhammar 1996), and the receptor genes belong to the paralogon consisting of human chromosomes Hsa4, Hsa5, Hsa8, and Hsa10 (Wraith et al. 2000). The NPY receptors belong to the superfamily of G protein–coupled receptors that include three Y1 subfamily subtypes named Y1, Y4, and Y6 and are located on human chromosomes 4, 10, and 5, respectively (Wraith et al. 2000). These three mammalian receptors are Key words: neuropeptide Y, G protein–coupled receptor, 2R hypothesis, linearized tree, gnathostome, chondrichthyes, Squalus acanthias. E-mail: [email protected]. 1271 Mol. Biol. Evol. 20(8):1271–1280. 2003 DOI: 10.1093/molbev/msg133 Molecular Biology and Evolution, Vol. 20, No. 8, Ó Society for Molecular Biology and Evolution 2003; all rights reserved. by guest on July 5, 2014 http://mbe.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Three Neuropeptide Y Receptor Genes in the Spiny Dogfish, Squalus acanthias,Support en Bloc Duplications in Early Vertebrate Evolution
Erik Salaneck,* David H. Ardell,� Earl T. Larson,* and Dan Larhammar**Department of Neuroscience, Unit of Pharmacology, and �Department of Molecular Evolution, Evolutionary Biology Center,Uppsala University, Uppsala, Sweden
It has been debated whether the increase in gene number during early vertebrate evolution was due to multipleindependent gene duplications or synchronous duplications of many genes. We describe here the cloning of threeneuropeptide Y (NPY) receptor genes belonging to the Y1 subfamily in the spiny dogfish, Squalus acanthias,a cartilaginous fish. The three genes are orthologs of the mammalian subtypes Y1, Y4, and Y6, which are located inparalogous gene regions on different chromosomes in mammals. Thus, these genes arose by duplications ofa chromosome region before the radiation of gnathostomes (jawed vertebrates). Estimates of duplication times fromlinearized trees together with evidence from other gene families supports two rounds of chromosome duplications ortetraploidizations early in vertebrate evolution. The anatomical distribution of mRNA was determined by reverse-transcriptase PCR and was found to differ from mammals, suggesting differential functional diversification of the newgene copies during the radiation of the vertebrate classes.
Introduction
Vertebrate genomes contain considerably more genesthan most invertebrate genomes. Ohno (1970) and othershave proposed that this difference arose through earlyvertebrate tetraploidizations. Gene mapping data supportgene duplications involving large chromosomal regions orentire chromosomes (Lundin 1993; Popovici et al. 2001a,2001b). Such sets of paralogous chromosomal regions arecalled paralogons (Coulier et al. 2000). The growingnumber and sizes of paralogons that have been identifiedin vertebrate genomes suggest that there was an expansionduring early vertebrate evolution via two tetraploidizations(Holland et al. 1994; Kasahara et al. 1996; Sidow 1996).This hypothesis is called the one-to-four rule (Ohno 1999),or the 2R hypothesis, for two rounds of genome doubling(Hughes 1999).
Despite the evidence, the 2R hypothesis remainscontroversial (Hughes 1999; Hughes, da Silva, andFriedman 2001; Martin 2001; Pennisi 2001). Alternativehypotheses include independent gene duplications fol-lowed by parallel aggregation into similar clusters (Hughes2000) or independent chromosome or block duplicationsrather than simultaneously in tetraploidizations (Skrabanecand Wolfe 1998; Smith, Knight, and Hurst 1999). Thearguments are discussed in recent reviews (Wolfe 2001;Larhammar, Lundin, and Hallbook 2002).
Another issue is when the duplications took place.Several authors have suggested that they occurred beforethe origin of gnathostomes but after amphioxus branchedoff (Holland et al. 1994; Sidow 1996; Spring 1997). Thisis supported by the presence of a single Hox cluster inamphioxus as opposed to four in tetrapods (Garcia-Fernandez and Holland 1994). This is also true for thelarge chromosomal region carrying the major histocom-patibility complex (Abi-Rached et al. 2002). After the
divergence of amphioxus and before the appearance ofgnathostomes, the agnathan (jawless fishes) hagfishes andlampreys appeared, either independently or as a mono-phyletic group (Delarbre et al. 2002). It has been proposedthat the first tetraploidization took place before theagnathans diverged and that the second tetraploidizationhappened subsequently in the lineage leading to gnatho-stomes (Holland et al. 1994). This interpretation is com-plicated by the discovery of three to four Hox clusters inlampreys, but these clusters may have occurred indepen-dently (Force, Amores, and Postlethwait 2002; Irvine et al.2002; Sharman and Holland 1998).
The time point for the second proposed tetraploidiza-tion is also uncertain, as only two Hox clusters have yetbeen identified in any cartilaginous fish (Kim et al. 2000).A recent review placed the second tetraploidization afterdivergence of sharks (Pennisi 2001). Regardless, cartilag-inous fishes and agnathans are important groups fordetermining time points and mechanisms for the increasein vertebrate gene number.
We have previously investigated the evolution of theneuropeptide Y (NPY) family of neuroendocrine peptidesin vertebrates and found that the first gene duplicationleading to NPY and peptide YY (PYY) probably tookplace before the origin of lampreys, as both the riverlamprey and gnathostomes have NPY and PYY (Cerda-Reverter and Larhammar 2000; Larhammar 1996; Soder-berg et al. 1994). More recently, we have described anNPY receptor in the river lamprey Lampetra fluviatilis thatappears to be a pro-ortholog of the tetrapod receptor Y4and the teleost receptor Yb (Salaneck et al. 2001). BothNPY peptide and receptor genes belong to paralogons thathave likely arisen by duplications in early vertebrateevolution, as the genes for NPY and PYY are linked toHox clusters (Larhammar 1996), and the receptor genesbelong to the paralogon consisting of human chromosomesHsa4, Hsa5, Hsa8, and Hsa10 (Wraith et al. 2000).
The NPY receptors belong to the superfamily of Gprotein–coupled receptors that include three Y1 subfamilysubtypes named Y1, Y4, and Y6 and are located onhuman chromosomes 4, 10, and 5, respectively (Wraithet al. 2000). These three mammalian receptors are
Key words: neuropeptide Y, G protein–coupled receptor, 2Rhypothesis, linearized tree, gnathostome, chondrichthyes, Squalusacanthias.
E-mail: [email protected].
1271
Mol. Biol. Evol. 20(8):1271–1280. 2003DOI: 10.1093/molbev/msg133Molecular Biology and Evolution, Vol. 20, No. 8,� Society for Molecular Biology and Evolution 2003; all rights reserved.
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from

approximately 50% identical to each other as well as tothree teleost fish receptors Ya, Yb, and Yc (Lundell et al.1997; Ringvall, Berglund, and Larhammar 1997; Starbacket al. 1999). Relationships between the mammalian andteleostean receptors have been unclear. Ya may be anortholog of mammalian Y4, whereas Yb and Yc areclosely related to each other, and their teleost pro-ortholog,Yb/c (represented in our trees by cod Yb/c), could be thefourth member of the quartet comprising this paralogon.The mammalian Yb/c ortholog was probably lost. Thereceptors named Y2 and Y5 are more distantly related tothe Y1 subfamily, and Y3 has only been defined phar-macologically and probably does not exist as a separategene (Larhammar et al. 2001).
As the phylogenetic resolution of agnathan sequencesrelative to the vertebrate gene duplication events is oftenambiguous in gene family trees (Ono-Koyanagi et al.2000; Germot et al. 2001; Neidert et al. 2001; Salanecket al. 2001), it is important to obtain additional sequenceinformation from agnathans and cartilaginous fishes. Thelatter group is poorly represented in molecular evolution-ary studies. We describe here the cloning and phylogeneticanalyses of three NPY receptor subtypes belonging to theY1 subfamily in the spiny dogfish, Squalus acanthias.These were identified as orthologs of mammalian receptorsY1, Y4, and Y6, thereby showing that block duplicationsof this paralogon preceded the origin of gnathostomes.We also describe unexpected anatomical distribution ofreceptor mRNA.
Materials and MethodsPCR Cloning
Alignments were made using amino acid sequencesfrom all available Y1 subfamily receptors using MegalignLASERGENE software. Highly conserved domains wereidentified and evaluated for construction of degeneratePCR primers. Genomic DNA was extracted from muscletissue from one frozen Squalus acanthias from the NorthSea (Hambergs Fisk, Uppsala, Sweden). Primers codingfor all possible combinations of the amino acid sequencesat positions indicated in figure 1 were designed (seeSupplementary Material for primer sequences). PCRreactions were run and analyzed according to Salanecket al. (2001).
Generation of Full-Length Clones by Inverse PCR
Squalus acanthias genomic DNA was digested witheither of four restriction enzymes: EcoRI, HindIII, BamHI,or XhoI (New England Biolabs) overnight at 378C. Eachdigest was then ligated at 148C overnight with T4 DNAligase (New England Biolabs) and precipitated. Two setsof PCR primers were designed for each receptor fragment;one inner primer pair and one outer primer. The primerswere constructed in reverse or ‘‘outward’’ direction toamplify nucleotide sequences upstream and downstream ofthe cloned fragment sequences. A primary PCR reactionwas run with the inner primer pairs and 1 ll of theligations as a template with the following cycling con-ditions: 958C for 30 s, 558C for 30 s, and 728C for 2 minfor 40 cycles. One ll of the primary PCR reactions wasused as template in a second PCR using the second primercombination for each fragment with the followingconditions: 958C for 30 s, 508C for 30 s, and 728C for 2min for 40 cycles. The second PCR reactions wereanalyzed on agarose gels, and the bands were excised andpurified. The purified fragments were cloned into pCR IIvector and transformed into TOP 10 cells (Invitrogen).Plasmid inserts were sequenced with vector-specificprimers according to the sequencing protocol above. Toconfirm the sequences, specific PCR primers weredesigned to generate full-length products for SacY4 andSacY6 and for the entire ORF of the first SacY1 exon.PCR with Taq polymerase (Gibco BRL) and Platinum Pfx(Gibco BRL) proofreading DNA polymerase generatedproducts identical to the sequences obtained by inversePCR.
Sequence Alignments
We made separate alignments of the Y1 subfamily(Y1, Y4, and Y6, hence ‘‘146’’) and all availablevertebrate NPY receptors (adding Y2 and Y5, hence‘‘12456’’) to assess alignment uncertainty and to reducethe effect of this uncertainty on analyses requiring only thesmaller data set.
The alignments were made with ClustalW (Thomp-son, Higgins, and Gibson 1994) and optimized withMultiClustal (Yuan et al. 1999). The MultiClustal align-ments and parameters were similar to those optimized by
FIG. 1.—The three Squalus Y receptor sequences (aligned in DNASTAR Megalign software). Putative transmembrane regions are boxed andnumbered. Positions and names of the degenerate primers used for PCR cloning are shown with arrows. Cystein residues conserved in all Y receptorsequences are also marked with boxes. Possible glycosylation sites are marked with shaded boxes. The ‘‘ERH’’ motif found in most Y receptorsequences is also indicated by a shaded box.
1272 Salaneck et al.
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from
hand with ClustalW alignment quality guides. Theparameter values and models used in the creation of fouralignments are shown in table 1. Because gaps werecontained almost exclusively in loops and at the termini,all gap-free alignments consisted almost entirely of trans-membrane regions and were highly consistent with oneanother.
Using Tree-Puzzle version 5.0 (Strimmer and vonHaeseler 1996), we found that the Danio rerio Yasequence (DreYa) was significantly different in its aminoacid composition from maximum likelihood estimates forboth 146 and 12456 data sets (v219 . 15:9, P, 0.05).
The MultiClustal-optimized alignments of the datasets without DreYa (‘‘146�DreYa’’ and ‘‘12456�DreYa’’)were different both qualitatively and quantitatively fromthose of the data sets with DreYa (table 1). For anal-yses without DreYa, we therefore used these separatealignments.
We initially removed all gap-containing sites. Thisforced us to exclude two truncated or incomplete se-quences (human Y6 and sturgeon Y1) from all data sets.We later found it necessary to retain gaps in the analysis ofthe largest data set, 12456�DreYa (see below). In analysisof gap-containing alignments, we ensured that all softwarewe used was consistent in treating gaps as missing data.
PhylogeneticsLikelihood Analysis
Using Tree-Puzzle to optimize maximum-likelihood(ML) branch lengths with initial Neighbor-Joining (NJ)tree of the 12456�DreYa data set computed withNEIGHBOR from JTT-F ML pairwise distances computedin Tree-Puzzle, we found that the JTT-F model (Jones,Taylor, and Thornton 1992) of protein evolution hada much larger likelihood with the data (JTT-F: logL ¼ �13149) than other available models available inTree-Puzzle (WAG-F: log L ¼ �13309; VM-F: logL ¼ �13487; and BLOSUM-F: log L ¼ �13492). Thisresult did not change when these other models were usedto calculate the initial pairwise distances.
We conducted likelihood ratio tests for site-rateheterogeneity using initial likelihood trees estimated inPROTML version 2.3b3 (Adachi and Hasegawa 1995),using local rearrangement search and default parameters,from initial Neighbor-Joining trees estimated with NJDISTversion 1.2.5 (Saitou and Nei 1987). Both initial pairwisedistances and likelihood trees were estimated with
PROTML using the JTT-F model. The resultingPROTML-estimated tree topologies were then input toTree-Puzzle for likelihood estimation trees with andwithout a discrete approximation (exact estimation, eightintervals) to a gamma distribution of site-rate hetero-geneity.
Assuming the likelihood ratio test (LRT) statistic forJTT-F versus JTT-F-� is distributed according to a 50:50mixture of the v20 and v21 distributions (denoted �vv21 as inGoldman and Whelan [2000]), then we could reject thenull hypothesis of site-rate homogeneity in all three datasets with very high confidence. The LRT statistics andinitial ML estimates (and their standard errors) of the alphashape parameters for the three data sets are presented intable 2, wherein P(�vv21 � S)¼ [P(v21 � S)/2] (Goldman andWhelan 2000). Similar results were obtained with thegapped 12456 alignment (calculated from an NJ tree fromPAUP*).
Likelihood tree estimation followed a similar processas likelihood ratio testing but started with ML pairwisedistances allowing for site-rate heterogeneity. First, MLpairwise distances were estimated for each data set usingTree-Puzzle and a JTT-F-� model with the initial alphaestimates shown in table 2. These ML pairwise distanceswere then input to NJDIST to make initial NJ trees. Thesewere then fed to PROTML for likelihood estimation oftopology under the JTT-F model and local rearrangementsearch (PROTML cannot model site heterogeneity). Thetopology changed slightly for the 146 tree but not for146�DreYa. The branch lengths and alpha parameter for146 were then reestimated from the data and the PROTMLlocal ML topologies in Tree-Puzzle using the JTT-F-�model. With the final 146 tree, the final alpha estimate wasa146 ¼ 0.73 6 0.07, and the likelihood was comparable tothat of the original estimated tree (log L ¼ �7337).
Bayesian Inference of Phylogeny
We analyzed the full (gap-containing) 12456�DreYaalignment in MrBayes version 3.0B (Huelsenbeck andRonquist 2001) with the JTT-F-� model and the gammashape parameter a coestimated with the trees. The programran for 117,140 steps with a tree sampled every 20generations. The first 857 trees (‘‘burn-in’’) were discardedas a generous estimate of the onset of stationarity, leaving5,000 trees for analysis. The program was initialized froma random tree. As a comparison and control against localmaxima, short runs were initialized from a ‘‘user tree,’’
Table 1Clustal Parameters Optimized with MultiClustal for the Different Data Sets
Number ofPairwise Multiple Sites
Alignment Sequences Matrix GOa GIb Matrix GO GI Total GFc
12456 39 GONNET 8 0.5 GONNET 20 0.1 530 30112456�DreYa 38 BLOSUM 12 0.5 GONNET 4 0.1 532 300146 25 BLOSUM 20 0.5 GONNET 20 0.1 402 323146�DreYa 24 BLOSUM 4 0.5 GONNET 4 0.1 399 324
a Gap-opening penalty.b Gap-insertion penalty.c Gap-free.
Squalus Neuropeptide Y Receptor Genes Support en Bloc Duplications 1273
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from

which was an initial PROTML-optimized topology madeas described above for likelihood ratio testing, and from anadditional random tree.
Distance and Parsimony Analysis
Consensus NJ trees were estimated from 100bootstrap replicates generated from the data sets inSEQBOOT from the PHYLIP package (Felsenstein1993), using ML distances under the JTT-F-� model withinitial alpha parameter estimates as given above or in table2. Pairwise distance matrices were calculated with Tree-Puzzle running under PUZZLEBOOT. NJ trees werecalculated with random addition of sequences usingNEIGHBOR and a consensus tree generated from themunder extended majority-rule with CONSENSE, both fromPHYLIP. Bootstrapped consensus parsimony trees (100replicates) were obtained using PAUP* (Swofford 2000)with mean character values, ‘‘ACCTRAN’’ or ‘‘acceler-ated transformation,’’ and random addition of sequences.
Phylogenetic Tests of the Molecular Clock andLinearized Trees
We estimated absolute and relative times of di-vergence of the Yb,Yc clade and the Y4 clades and of theY1,Y6 clade and the Y4,Yb,Yc clade by generatinga linearized tree (Takezaki, Rzhetsky, and Nei 1995) withthe Y2 and Y5 sequences as outgroups and calibrationdates from estimated Squalus and Lampetra fossil di-vergence times.
The topology of the Bayesian consensus tree of the12456�DreYa data set (gapped) and its correspondingdata set were automatically converted and reordered to theMEGA-style format required by the LINTREE packageusing custom Perl software (FAS2LTRE, NEWICK2-MEGA, and LTRECONFORM) available from David H.Ardell. Two-cluster and branch-length tests were com-puted with TPCV and BRANCH from the LINTREEpackage. We used the amino-gamma distance with thealpha estimate from gap-free data (table 2) becauseLINTREE eliminates gap-containing sites.
Three rounds of sequence elimination were necessaryto produce a nearly ultrametric linearized tree. In the firstround, the two-cluster test indicated heterogeneity betweenthe slow-evolving Squalus Y6 and the more quicklyevolving mammalian Y6 sequences (P , 0.01). Peccaryy6, in this group, was also the only significantly fast (orslow) evolving sequence compared with the branchaverage and so was eliminated using COLLAPSE (avail-able from David H. Ardell). In the second round (reusing
the same alpha estimate), no individual node or branch wassignificantly heterogeneous at the 1% level, and overallrate heterogeneity as quantified by the U statistic for thetwo-cluster test was not significant (U ¼ 15:9; P. 0:5)but the U statistic for the branch-length test was stillsignificant (U ¼ 133:1; P, 10�3), with the Squalus Y4sequence evolving quickest and the Cod Yb sequenceevolving slowest. Cod Yb was most atypical overall andwas removed for a third round. In the third and fourthrounds, heterogeneity between the Squalus and mamma-lian y/Y6 sequences and/or overall rate heterogeneity werestill significant at the 5% level, requiring elimination of themouse y6 and then Squalus Y6 in the third and fourthrounds, respectively. In the fourth round, no clusters orbranches exhibited rate heterogeneity at the 5% level, nordid the data set overall, by either test.
RT-PCR
One female Squalus acanthias was captured in theSkagerack Sea (Larssons Fisk HB, Mollosund, Sweden).Tissue samples were placed in RNAlater (Ambion). TotalRNA preparations were made with ca 50 mg from eachtissue according to RNeasy kit protocol (Qiagen) includingDNase treatment to eliminate genomic DNA contamina-tion. Primers designed from the deduced Squalus se-quences were used (primer sequences available uponrequest) in Titan One-tube RT-PCR reactions (Roche Bio-chemicals) with ca 20 pg total RNA in each reaction ac-cording to protocol. Specific primers for Squalus acanthiasmyelin basic protein (MBP) was used as a positive con-trol (Spivack et al. 1993) (sequences upon request). For anegative control, all utilized RT-PCR primer combinationswere tested in standard PCR conditions with Taq poly-merase (Gibco BRL) on the RNA preparations. Also, asa second negative control, all RT-PCR reactions were runwithout the preliminary reverse transcriptase step. Theproducts were analyzed on standard 1.5% agarose gels andblotted overnight to nylon filters. Filters were hybridizedwith 32P-labeled probes at 658C in ExpressHyb bufferovernight and exposed to film (Amersham) overnight at�708C.
ResultsIsolation of Three Squalus acanthias NPY ReceptorDNA Clones
Of approximately 300 plasmid clones containingPCR products, eight had a high degree of identity with Y1subfamily receptors. Clones originally produced withprimers Fa1f and Fa1r showed highest identity with theY1 subtype, clones from Yabc1f and Yabc1r showedhighest identity with Y4 sequences, and clones made withprimers Fa1f and Ls1r showed highest identity with Y6sequences (fig. 1). Inverse PCR performed on circularizedfragments of genomic Squalus acanthias (Sac) DNAallowed the deduction of full-length Y4 and Y6 sequences.The SacY1 sequence could be extended in the 59 directionto reach the initiating ATG codon. The SacY1 gene wasfound to have an intron after codon 232, that is, the sameposition as in all other known Y1 sequences (fig. 1).
Table 2LRT Statistics and Initial Likelihood Parameter Estimatesfor Gap-Free Alignments
Alignment LRT ¼ �2�lnL P(�vv21) � LRT aNJ 6 SE
12456�DreYa 2(8642 � 8333) ¼ 618 ,10�3 0.99 6 0.10146�DreYa 2(7160 � 6864) ¼ 592 ,10�3 0.74 6 0.07146 2(7679 � 7337) ¼ 684 ,10�3 0.73 6 0.07
1274 Salaneck et al.
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from

Approximately 500 bp of intron sequence could bedetermined by the inverse PCR technique, but no codingsequence corresponding to the next exon was reached. Toobtain sequence from exon 2, a separate degenerate PCRcloning was undertaken (primers Y1phf and Y1cp2r) usingthe same PCR procedure as above, and three clones wereobtained. The exon sequences flanking the intron wereobtained with an exon 1 forward primer and an exon 2reverse primer using Sac renal cDNA as template. Noproduct was obtained when performing the PCR reactionwith genomic DNA as a template, despite optimizing thereaction for long PCR products (. 5,000 kb). Thus, theintron appears to be considerably larger than in tetrapods,where it is just over 100 bp. Full-length sequences ofSacY4 and SacY6 as well as the almost complete SacY1were aligned with other Y1 subfamily amino acid se-quences (fig. 1). The overall sequence amino acid identitybetween the Squalus and mammalian orthologs were ca58% to 64% for Y1, 56% to 57% for Y6, and 48% for Y4.Sequence identity between the three Squalus sequenceswas 46% to 55%, just as between other Y1 subfamilysequences.
Monophyly and Rooting of the 146 Clade
Figure 2 shows a cladogram of the complete NPYdata set without Danio rerio Ya (12456�DreYa gapped).Support for monophyly of a Y1,Y6,Y4 clade (‘‘146’’ or‘‘Y1 subfamily’’) was unequivocal.
All methods place the root of the Y1 subfamilybetween the Y4,Yb,Yc clade and the Y1, Y6 clade, asspecified from the Y2 and Y5 sequences as outgroups.However, the placement of the Lampetra Y sequence wassensitive to the inclusion of gap-containing sites and to themethod used. Although NJ and parsimony methods put thelamprey sequence in the Y1,Y6 clade, Mr. Bayescalculated a posterior probability of 0.94 for monophylyof Lampetra Y with the Y4,Yb,Yc clade. Using four-cluster likelihood mapping (Strimmer and von Haeseler1997) as implemented in Tree-Puzzle on the alignmentwith gaps, with Y2,Y5; Y1,Y6; Y4,Yb,Yc; and LampetraY as the four clusters, gave weak support for groupingLampetra Y with the Y1,Y6 clade (8.5%). Among re-solved quartets (33.3%), monophyly with the Y4,Yb,Ycclade was relatively high (65.2%), but most support was ina star-tree (54.9%).
The 146 clade could not be rooted reliably with thegap-free version of this alignment. In the gap-freelikelihood tree made with PROTML, the root fell betweenthe Lampetra Y receptor sequence and the other Y4,Yb,Ycsequences, with no support for the placement as de-termined by RELL relative likelihood values (data notshown). The NJ tree gave a result consistent with thatshown in figure 2, but the support for placing the rootoutside the Lampetra Y receptor sequence was only slightto nonexistent, at 64. The parsimony results, on the otherhand, were less affected by the presence or absence of gapsthan those of the other methods. Also, in the gap-freeanalysis, the monophyly of the 146 clade remained un-equivocally supported by all methods of analysis. In theBayesian analysis of this data set, the posterior mean
log-likelihood of sampled trees was ln L ¼�13798 witha 95% credible interval of ln L 2 (�13812;�13785). Theestimated posterior mean shape parameter for the gammadistribution of site-rates was a¼ 0.89 with a 95% credibleinterval of a 2 (0.75; 1.03).
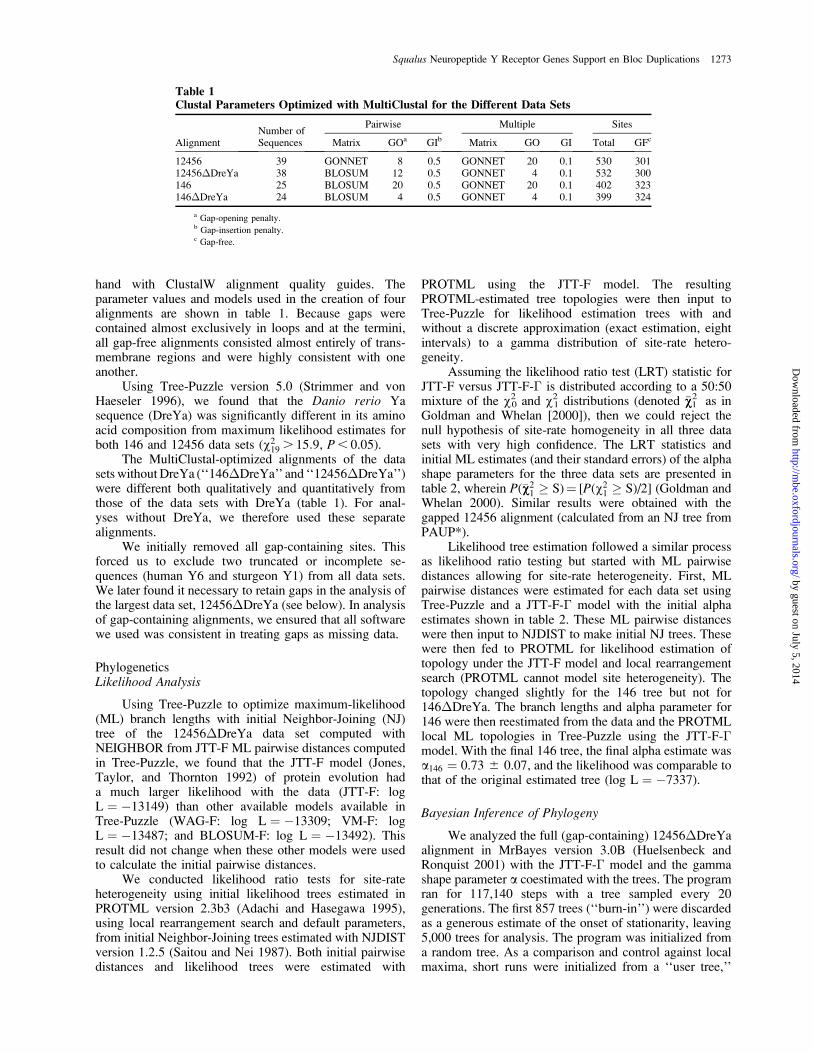
Phylogenetic Placement of Squalus acanthias Sequences
Unrooted likelihood phylograms of the 146 clade areshown in figure 3. With most branches, particularly thoseof the Squalus acanthias and Lampetra fluviatilis se-quences, all methods gave consistent and reliable results.The three SacY1 subfamily homologs segregate clearlyand consistently near the roots of the Y1, Y4 þ Ya, andY6 clades.
Removing the heterogeneous Danio Ya sequenceresulted in a drop of overall support for monophyly of theY4,Yb,Yc clade. Otherwise the phylogenetic placement ofthe Squalus sequences and the shape of the tree in generalwas little affected by the presence or absence of thissequence.
FIG. 2.—Consensus cladogram of all NPY receptor sequences fromthe 12456�DreYa alignment. Clade support values shown are in order ofMrBayes clade credibilities (sample size n ¼ 20,000), then NJ and MPbootstrap support (100 replicates). Shallow branches with uniformly lowsupport (less than 70 with all methods) or no support with any onemethod were collapsed.
Squalus Neuropeptide Y Receptor Genes Support en Bloc Duplications 1275
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from
Absolute and Relative Divergence Time and RateEstimates Within the Y1-Subfamily
As described in the methods section, the Y6 cladewas particularly heterogeneous in evolutionary rate, con-sistent with pharmacological data suggesting partial orcomplete inactivity of this subtype in mammals. TheYb,Yc sequences evolved significantly slower than did Y4sequences (P, 0:001), by the two-cluster test.
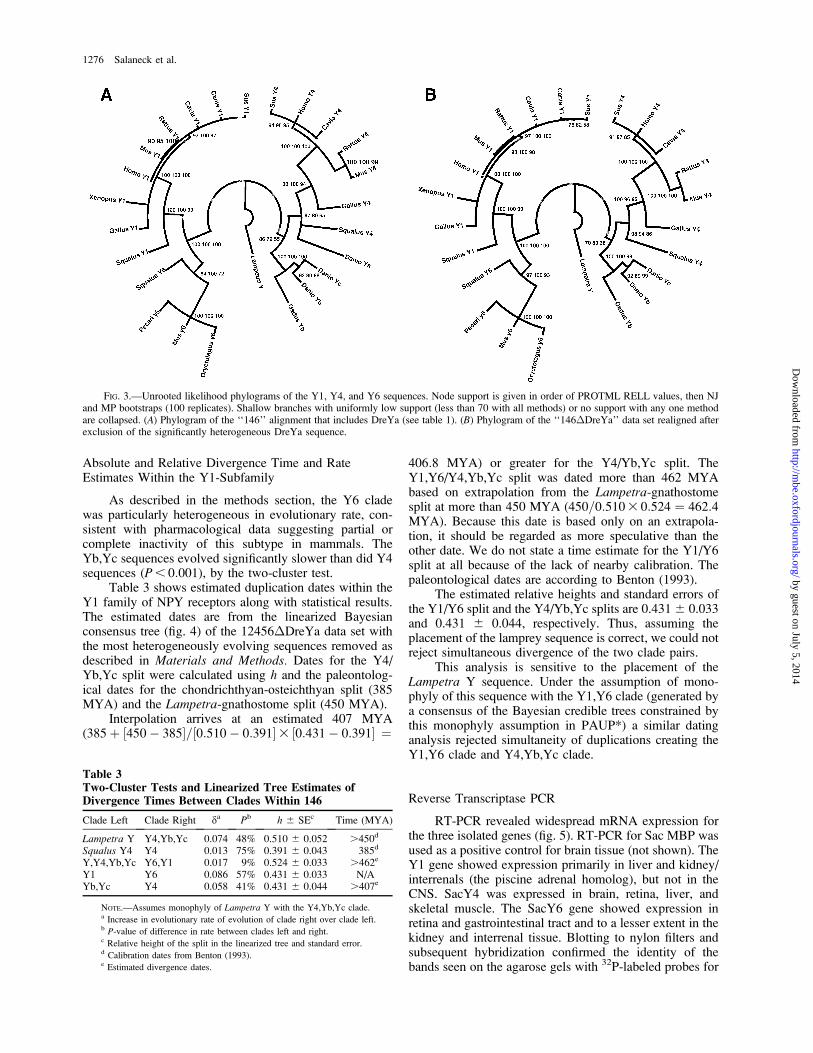
Table 3 shows estimated duplication dates within theY1 family of NPY receptors along with statistical results.The estimated dates are from the linearized Bayesianconsensus tree (fig. 4) of the 12456�DreYa data set withthe most heterogeneously evolving sequences removed asdescribed in Materials and Methods. Dates for the Y4/Yb,Yc split were calculated using h and the paleontolog-ical dates for the chondrichthyan-osteichthyan split (385MYA) and the Lampetra-gnathostome split (450 MYA).
Interpolation arrives at an estimated 407 MYA(385þ ½450� 385�=½0:510� 0:391�3 ½0:431� 0:391� ¼
406:8 MYA) or greater for the Y4/Yb,Yc split. TheY1,Y6/Y4,Yb,Yc split was dated more than 462 MYAbased on extrapolation from the Lampetra-gnathostomesplit at more than 450 MYA (450=0:5103 0:524 ¼ 462:4MYA). Because this date is based only on an extrapola-tion, it should be regarded as more speculative than theother date. We do not state a time estimate for the Y1/Y6split at all because of the lack of nearby calibration. Thepaleontological dates are according to Benton (1993).
The estimated relative heights and standard errors ofthe Y1/Y6 split and the Y4/Yb,Yc splits are 0.4316 0.033and 0.431 6 0.044, respectively. Thus, assuming theplacement of the lamprey sequence is correct, we could notreject simultaneous divergence of the two clade pairs.
This analysis is sensitive to the placement of theLampetra Y sequence. Under the assumption of mono-phyly of this sequence with the Y1,Y6 clade (generated bya consensus of the Bayesian credible trees constrained bythis monophyly assumption in PAUP*) a similar datinganalysis rejected simultaneity of duplications creating theY1,Y6 clade and Y4,Yb,Yc clade.
Reverse Transcriptase PCR
RT-PCR revealed widespread mRNA expression forthe three isolated genes (fig. 5). RT-PCR for Sac MBP wasused as a positive control for brain tissue (not shown). TheY1 gene showed expression primarily in liver and kidney/interrenals (the piscine adrenal homolog), but not in theCNS. SacY4 was expressed in brain, retina, liver, andskeletal muscle. The SacY6 gene showed expression inretina and gastrointestinal tract and to a lesser extent in thekidney and interrenal tissue. Blotting to nylon filters andsubsequent hybridization confirmed the identity of thebands seen on the agarose gels with 32P-labeled probes for
FIG. 3.—Unrooted likelihood phylograms of the Y1, Y4, and Y6 sequences. Node support is given in order of PROTML RELL values, then NJand MP bootstraps (100 replicates). Shallow branches with uniformly low support (less than 70 with all methods) or no support with any one methodare collapsed. (A) Phylogram of the ‘‘146’’ alignment that includes DreYa (see table 1). (B) Phylogram of the ‘‘146�DreYa’’ data set realigned afterexclusion of the significantly heterogeneous DreYa sequence.
Table 3Two-Cluster Tests and Linearized Tree Estimates ofDivergence Times Between Clades Within 146
Clade Left Clade Right da Pb h 6 SEc Time (MYA)
Lampetra Y Y4,Yb,Yc 0.074 48% 0.510 6 0.052 .450d
Squalus Y4 Y4 0.013 75% 0.391 6 0.043 385d
Y,Y4,Yb,Yc Y6,Y1 0.017 9% 0.524 6 0.033 .462e
Y1 Y6 0.086 57% 0.431 6 0.033 N/AYb,Yc Y4 0.058 41% 0.431 6 0.044 .407e
NOTE.—Assumes monophyly of Lampetra Y with the Y4,Yb,Yc clade.a Increase in evolutionary rate of evolution of clade right over clade left.b P-value of difference in rate between clades left and right.c Relative height of the split in the linearized tree and standard error.d Calibration dates from Benton (1993).e Estimated divergence dates.
1276 Salaneck et al.
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from
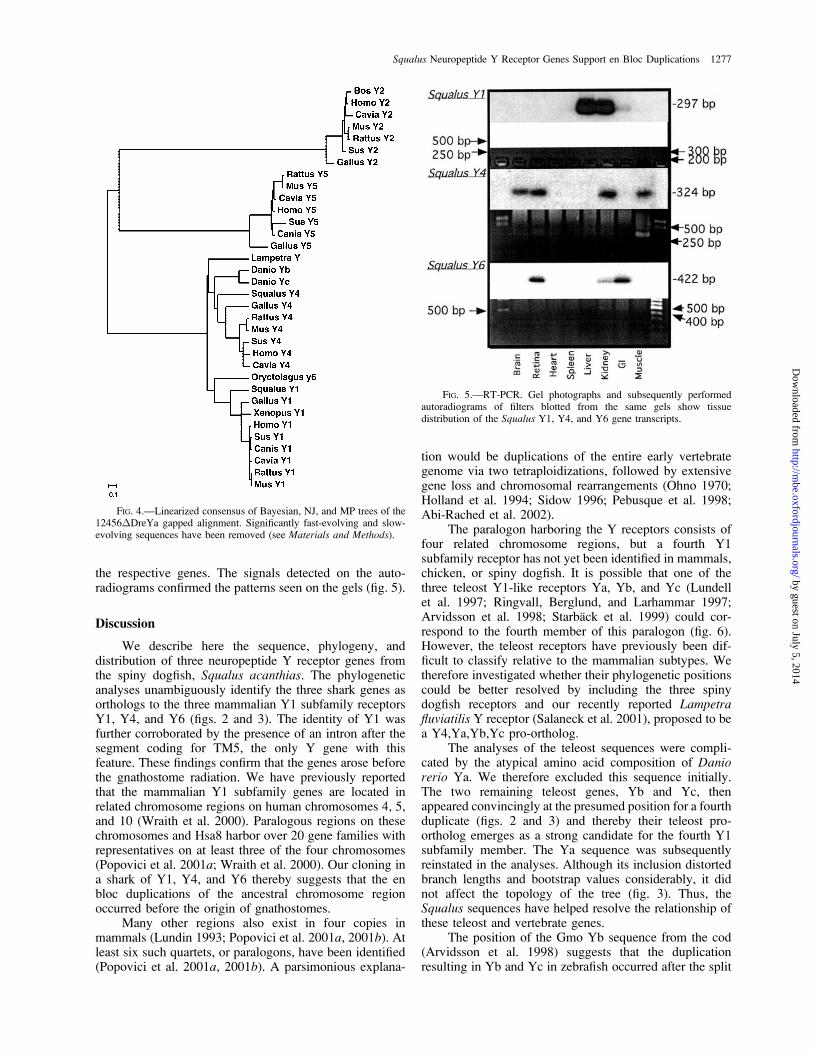
the respective genes. The signals detected on the auto-radiograms confirmed the patterns seen on the gels (fig. 5).
Discussion
We describe here the sequence, phylogeny, anddistribution of three neuropeptide Y receptor genes fromthe spiny dogfish, Squalus acanthias. The phylogeneticanalyses unambiguously identify the three shark genes asorthologs to the three mammalian Y1 subfamily receptorsY1, Y4, and Y6 (figs. 2 and 3). The identity of Y1 wasfurther corroborated by the presence of an intron after thesegment coding for TM5, the only Y gene with thisfeature. These findings confirm that the genes arose beforethe gnathostome radiation. We have previously reportedthat the mammalian Y1 subfamily genes are located inrelated chromosome regions on human chromosomes 4, 5,and 10 (Wraith et al. 2000). Paralogous regions on thesechromosomes and Hsa8 harbor over 20 gene families withrepresentatives on at least three of the four chromosomes(Popovici et al. 2001a; Wraith et al. 2000). Our cloning ina shark of Y1, Y4, and Y6 thereby suggests that the enbloc duplications of the ancestral chromosome regionoccurred before the origin of gnathostomes.
Many other regions also exist in four copies inmammals (Lundin 1993; Popovici et al. 2001a, 2001b). Atleast six such quartets, or paralogons, have been identified(Popovici et al. 2001a, 2001b). A parsimonious explana-
tion would be duplications of the entire early vertebrategenome via two tetraploidizations, followed by extensivegene loss and chromosomal rearrangements (Ohno 1970;Holland et al. 1994; Sidow 1996; Pebusque et al. 1998;Abi-Rached et al. 2002).
The paralogon harboring the Y receptors consists offour related chromosome regions, but a fourth Y1subfamily receptor has not yet been identified in mammals,chicken, or spiny dogfish. It is possible that one of thethree teleost Y1-like receptors Ya, Yb, and Yc (Lundellet al. 1997; Ringvall, Berglund, and Larhammar 1997;Arvidsson et al. 1998; Starback et al. 1999) could cor-respond to the fourth member of this paralogon (fig. 6).However, the teleost receptors have previously been dif-ficult to classify relative to the mammalian subtypes. Wetherefore investigated whether their phylogenetic positionscould be better resolved by including the three spinydogfish receptors and our recently reported Lampetrafluviatilis Y receptor (Salaneck et al. 2001), proposed to bea Y4,Ya,Yb,Yc pro-ortholog.
The analyses of the teleost sequences were compli-cated by the atypical amino acid composition of Daniorerio Ya. We therefore excluded this sequence initially.The two remaining teleost genes, Yb and Yc, thenappeared convincingly at the presumed position for a fourthduplicate (figs. 2 and 3) and thereby their teleost pro-ortholog emerges as a strong candidate for the fourth Y1subfamily member. The Ya sequence was subsequentlyreinstated in the analyses. Although its inclusion distortedbranch lengths and bootstrap values considerably, it didnot affect the topology of the tree (fig. 3). Thus, theSqualus sequences have helped resolve the relationship ofthese teleost and vertebrate genes.
The position of the Gmo Yb sequence from the cod(Arvidsson et al. 1998) suggests that the duplicationresulting in Yb and Yc in zebrafish occurred after the split
FIG. 4.—Linearized consensus of Bayesian, NJ, and MP trees of the12456�DreYa gapped alignment. Significantly fast-evolving and slow-evolving sequences have been removed (see Materials and Methods).
FIG. 5.—RT-PCR. Gel photographs and subsequently performedautoradiograms of filters blotted from the same gels show tissuedistribution of the Squalus Y1, Y4, and Y6 gene transcripts.
Squalus Neuropeptide Y Receptor Genes Support en Bloc Duplications 1277
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from

between these two teleosts and not as a result of the basalteleost tetraploidization (Amores et al. 1998; Taylor et al.2001). However, data from more teleosts are neededbefore the time point for the Yb/Yc duplication can bedetermined with certainty. Whether additional duplicatesof any of these four Y1 subfamily genes exist in teleosts,as a result of their additional tetraploidization (fig. 6),remains to be investigated. According to the proposedduplication scheme, the Yb gene should have arisen beforethe divergence of Chondrichthyes. Further studies areneeded to see if Yb still exists in any species of this classor whether it has been lost, as seems to be the case inmammals. The chromosomal locations of the zebrafish Ya,Yb, and Yc genes (Starback et al. 1999) do not at presentclarify the relationships to their mammalian genes.
The phylogenetic position of the Lampetra fluviatilisreceptor was not clearly resolved by our data. However,the Bayesian analysis provides clear support of itsmembership in the Y4,Ya,Yb,Yc clade at a posteriorprobability of 0.94. If the lamprey sequence is indeed a Y4receptor, the first chromosome duplication (tetraploidiza-tion) would have taken place before the lampreys divergedfrom the gnathostome ancestor and the second chromo-some duplication (tetraploidization) took place in thegnathostome lineage (fig. 6). However, the idea that onetetraploidization took place on each side of the lampreybranch point is questioned by the finding of three to fourHox clusters in the sea lamprey, Petromyzon marinus(Force, Amores, and Postlethwait 2002; Irvine et al. 2002).We should be able to shed more light on this issue bycloning more Y1-like lamprey sequences. In any event, itseems clear that both chromosome duplications took placebefore the radiation of gnathostomes (fig. 6).
To determine whether the four clades of the Y1 sub-family arose via duplication of two pro-orthologs, alinearized tree was generated. Using this tree (fig. 4), wefound that the Y1, Yb þ c, Y4, and Y6 clades arosesimultaneously (i.e., we could not falsify the hypothesis
that the two deep branches became four at a single timepoint). The so-called 2 þ 2 topology we found, alsodenoted (AB)(CD), has not been consistently obtained forother gene families and quadrupled chromosomes(Hughes, da Silva, and Friedman 2001), although variableevolutionary rates may also explain this (Abi-Rached et al.2002). This result depends on the monophyly of thelamprey sequence with the Y4,Yb,Yc clade; we were ableto reject simultaneity on the hypothesis that the lampreysequence groups with the Y1 and Y6 receptors.
The tissue distribution of the three shark receptormRNAs, as determined by RT-PCR, revealed unexpecteddifferences from mammals and chicken. Mammalian Y1 isprimarily expressed in the CNS (Mikkelsen and Larsen1992). We could not detect SacY1 expression in the brain,but instead in peripheral tissues (fig. 5). This was un-expected, considering that receptor binding studies per-formed on CNS sections of the smooth dogfish Mustelacanis seemed to identify Y1-like binding in the hypothal-amus (McVey et al. 1996).
Y4 expression in the shark was also apparentlydifferent than that in mammals, where it is foundpredominantly in the gastrointestinal tract (Feletou et al.1999). We detected SacY4 in retina, muscle, renal/interrenal tissue, and the brain. Mammalian Y4 isa receptor for the gastrointestinal peptide PP (Lundellet al. 1995), which has been observed only in tetrapods(Larhammar 1996). We hypothesize that PP hijacked thepreexisting Y4 receptor at some point during tetrapodevolution, concomitant with divergence in function of theY4 receptor. This is consistent with the different ex-pression pattern of the shark Y4 receptor, the first non-tetrapod Y4 to be cloned.
SacY6 expression was detected in retina, GI tract, andkidney/interrenal tissues. Comparisons with mammals areuncertain since, in most mammals, Y6 is either a pseudo-gene or has not been allocated a physiological correlate(Matsumoto et al. 1996; Rose et al. 1997; Starback et al.2000; Wraith et al. 2000). The apparent reduced im-portance of mammalian Y6 is further supported by the factthat all known Y6 pseudogenes have become nonfunc-tional by distinct mutations (Larhammar et al. 2001).
An explanation for these divergent expressionpatterns, considering that the duplications occurred shortlybefore the gnathostome radiation, could be that subfunc-tionalization of the Y1 subfamily genes or their acquisitionof novel functions had probably not yet occurred by thetime of the Chondrichthyan split from all other gnatho-stomes (Osteichthyes, i.e., Sarcopterygians plus Actino-pterygians).
In summary, we have isolated elasmobranch ortho-logs to all three Y1 subfamily receptor genes previouslyfound in tetrapods. Orthology is strongly supported byphylogenetic analyses, overall sequence identity as well asrare genomic changes in the sequences, such as an intronand deletions. The localization of these genes on relatedchromosome segments in mammals and chicken, alongwith their presence in the spiny dogfish, suggest that thegene family expanded by en bloc duplications early invertebrate evolution. Together with data from other genefamilies, this supports the theory of the vertebrate genome
FIG. 6.—A hypothetical gene duplication scheme depicting theexpansion of the Y1 subfamily in chordates. The three Squalus sequencescorrelate well to the three mammalian subtypes Y1, Y4, and y6,indicating that chondrichthyans underwent the same duplication events astetrapods (shaded ovals). The scheme is in agreement with the 2R theorybut may be seen as speculative until further sequences have beenobtained.
1278 Salaneck et al.
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from

having expanded by means of chromosomal or genomicduplications before the rise of gnathostomes.
Supplementary Material
The primer sequences used for the PCR and RT-PCRreactions we performed are available in the online Sup-plementary Material and at http://www.bmc.uu.se/;danl/Sqprimers.htm. Accession numbers for the sequencesreported in this paper are as follows: AY177270,AY177271, AY177272, and AY177273. The ‘‘146,’’‘‘146�DreYa,’’ and ‘‘12456�DreYa’’ alignments havebeen deposited at EMBL-ALIGN with accession numbersALIGN_000531, ALIGN_000532, and ALIGN_0005333.
Acknowledgments
We thank Christina Bergqvist, Ida Solum, Dr. MatsFalk, and Dr. Mikael Thollesson for assistance and advice,the crew of M/S Shetland for acquiring a live shark, andAnna Salaneck, M.D., for dissection assistance and sharkrestraint. The Swedish Natural Science Research Councilsupported this work. D.H.A. was supported by a NationalScience Foundation Fellowship in Biological Informaticsawarded in 1999.
Literature Cited
Abi-Rached, L., A. Gilles, T. Shiina, P. Pontarotti, and H. Inoko.2002. Evidence of en bloc duplication in vertebrate genomes.Nature Genet. 31:100–105.
Adachi, J., and M. Hasegawa, 1995. Molphy version 2.3b3,programs for molecular phylogenetics based on maximumlikelihood. The Institute of Statistical Mathematics, Tokyo,Japan.
Amores, A., A. Force, Y.-L. Yan et al. (13 co-authors). 1998.Zebrafish hox clusters and vertebrate genome evolution.Science. 282:1711–1714.
Arvidsson, A.-K., A. Wraith, A.-C. Jonsson-Rylander, and D.Larhammar. 1998. Cloning of a neuropeptide Y/peptide YYreceptor from the Atlantic cod: the Yb receptor. Regul. Pept.75–76:39–43.
Benton, M. J. 1993. The fossil record 2. Chapman & Hall,London.
Cerda-Reverter, J. M., and D. Larhammar. 2000. Neuropeptide Yfamily of peptides: structure, anatomical expression, function,and molecular evolution. Biochem. Cell Biol. 78:371–392.
Coulier, F., C. Popovici, R. Villet, and D. Birnbaum, 2000.MetaHOX gene clusters. J. Exp. Zool. 288:345–351.
Delarbre, C., C. Gallut, V. Barriel, P. Janvier, and G. Gachelin,2002. Complete mitochondrial DNA of the hagfish, Eptatretusburgeri: the comparative analysis of mitochondrial DNAsequences strongly supports the cyclostome monophyly. Mol.Phylogenet. Evol. 22:184–192.
Feletou, M., J. P. Nicolas, M. Rodriguez, P. Beauverger, J. P.Galizzi, J. A. Boutin, and J. Duhault. 1999. NPY receptorsubtype in the rabbit isolated ileum. Br. J. Pharmacol.127:795–801.
Felsenstein, J. 1993. PHYLIP (phylogeny inference package).Version 3.5c. Distributed by the author, Department ofGenetics, University of Washington, Seattle.
Force, A., A. Amores, and J. H. Postlethwait. 2002. Hox clusterorganization in the jawless vertebrate Petromyzon marinus.J. Exp. Zool. 294:30–46.
Garcia-Fernandez, J., and P. W. H. Holland. 1994. Archetypalorganization of the amphioxus Hox gene cluster. Nature370:563–566.
Germot, A., G. Lecointre, J. L. Plouhinec, C. Le Mentec, F.Girardot, and S. Mazan. 2001. Structural evolution of Otxgenes in craniates. Mol. Biol. Evol. 18:1668–1678.
Goldman, N., and S. Whelan. 2000. Statistical tests of gamma-distributed rate heterogeneity in models of sequence evolutionin phylogenetics. Mol. Biol. Evol. 17:975–978.
Holland, P. W. H., J. Garcia-Fernandez, N. A. Williams, and A.Sidow. 1994. Gene duplications and the origins of vertebratedevelopment. Dev.(Suppl.):125–133.
Huelsenbeck, J. P., and F. Ronquist. 2001. MrBayes: Bayesianinference of phylogenetic trees. Bioinformatics 17:754–755.
Hughes, A. L. 1999. Phylogenies of developmentally importantproteins do not support the hypothesis of two rounds ofgenome duplication early in vertebrate history. J. Mol. Evol.48:565–576.
———. 2000. Polyploidization and vertebrate origins: a reviewof the evidence. Pp. 493–502 in D. Sankoff and J. H. Nadeau,eds. Comparative genomics. Kluwer, Dordrecht, The Nether-lands.
Hughes, A. L., J. da Silva, and R. Friedman. 2001. Ancientgenome duplications did not structure the human Hox-bearingchromosomes. Genome Res. 11:771–780.
Irvine, S. Q., J. L. Carr, W. J. Bailey, K. Kawasaki, N. Shimizu,C. T. Amemiya, and F. H. Ruddle. 2002. Genomic analysis ofHox clusters in the sea lamprey Petromyzon marinus. J. Exp.Zool. 294:47–62.
Jones, D. T., W. R. Taylor, and J. M. Thornton. 1992. The rapidgeneration of mutation data matrices from protein sequences.Comput. Appl. Biosci. 8:275–282.
Kasahara, M., M. Hayashi, K. Tanaka, H. Inoko, K. Sugaya,T. Ikemura, and T. Ishibashi. 1996. Chromosomal locali-zation of the proteasome Z subunit gene reveals an ancientchromosomal duplication involving the major histocom-patibilitiy complex. Proc. Natl. Acad. Sci. USA 93:9096–9101.
Kim, C.-B., C. Amemiya, W. Bailey, K. Kawasaki, J. Mezey, W.Miller, S. Nimoshima, N. Shimizu, G. Wagner, and F. Ruddle.2000. Hox cluster genomics in the horn shark, Heterodontusfrancisci. Proc. Natl. Acad. Sci. USA 97:1655–1660.
Larhammar, D. 1996. Evolution of neuropeptide Y, peptide YY,and pancreatic polypeptide. Regul. Pept. 62:1–11.
Larhammar, D., L. G. Lundin, and F. Hallbook. 2002. The Hox-bearing chromosome regions did arise by block or chromo-some (or even genome) duplications. Genome Res. 12:1910–1920.
Larhammar, D., A. Wraith, M. M. Berglund, S. K. S. Holmberg,and I. Lundell. 2001. Origins of the multiple NPY-familyreceptors in mammals. Peptides 22:295–307.
Lundell, I., M. M. Berglund, P. Starback, E. Salaneck, D. R.Gehlert, and D. Larhammar. 1997. Cloning and characteriza-tion of a novel neuropeptide Y receptor subtype in thezebrafish. DNA Cell Biol. 16:1357–1363.
Lundell, I., A. G. Blomqvist, M. M. Berglund, D. A. Schober, D.Johnson, M. A. Statnick, R. A. Gadski, D. R. Gehlert, and D.Larhammar. 1995. Cloning of a human receptor of the NPYreceptor family with high affinity for pancreatic polypeptideand peptide YY. J. Biol. Chem. 270:29123–29128.
Lundin, L. G. 1993. Evolution of the vertebrate genome asreflected in paralogous chromosomal regions in man and thehouse mouse. Genomics 16:1–19.
Lynch, M., and J. S. Conery. 2000. The evolutionary fate andconsequences of duplicate genes. Science 290:1151–1155.
Martin, A. 2001. Is tetralogy true? Lack of support for the ‘‘one-to-four’’ rule. Mol. Biol. Evol. 18:89–93.
Squalus Neuropeptide Y Receptor Genes Support en Bloc Duplications 1279
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from

Matsumoto, M., T. Nomura, K. Momoses, Y. Ikeda, Y. Kondou,H. Akiho, J. Togami, Y. Kimura, M. Okada, and T.Yamaguchi. 1996. Inactivation of a novel neuropeptide Y/peptide YY receptor gene in primate species. J. Biol. Chem.271:27217–27220.
McVey, D. C., D. Rittschof, P. J. Mannon, and S. R. Vigna.1996. Localization and characterization of neuropeptide Y/peptide YY receptors in the brain of the smooth dogfish(Mustelis canis). Regul. Pept. 61:167–173.
Mikkelsen, J. D., and P. J. Larsen. 1992. A high concentration ofNPY (Y1)-receptor mRNA-expressing cells in the hypotha-lamic arcuate nucleus. Neurosci. Lett. 148:195–198.
Neidert, A. H., V. Virupannavar, G. W. Hooker, and J. A.Langeland. 2001. Lamprey Dlx genes and early vertebrateevolution. Proc. Natl. Acad. Sci. USA 98:1665–1670.
Ohno, S. 1970. Evolution by gene duplication. Springer-Verlag,Berlin.
Ono-Koyanagi, K., H. Suga, K. Katoh, and T. Miyata. 2000Protein tyrosine phosphatases from amphioxus, hagfish, andray: divergence of tissue-specific isoform genes in the earlyevolution of vertebrates. J. Mol. Evol. 50:302–311.
Pebusque, M. J., F. Coulier, D. Birnbaum, and P. Pontarotti.1998. Ancient large-scale genome duplications: phylogeneticand linkage analyses shed light on chordate genomeevolution. Mol. Biol. Evol. 15:1145–1159.
Pennisi, E. 2001. Genome duplications: the stuff of evolution?Science 294:2458–2460.
Popovici, C., M. Leveugle, D. Birnbaum, and F. Coulier. 2001a.Coparalogy: physical and functional clusterings in the humangenome. Biochem Biophys. Res. Comm. 288:362–370.
———. 2001b. Homeobox gene clusters and the human paralogymap. FEBS Lett. 491:237–242.
Ringvall, M., M. M. Berglund, and D. Larhammar. 1997.Multiplicity of neuropeptide Y receptors: cloning of a thirddistinct subtype in the zebrafish. Biochem. Biophys. Res.Comm. 241:749–755.
Rose, P. M., J. S. Lynch, S. T. Frazier, S. M. Fisher, W. Chung,P. Battaglino, Z. Fathi, R. Leibel, and F. Prabhavathi. 1997.Molecular genetic analysis of a human neuropeptide Yreceptor: the human homolog of the murine ‘‘Y5’’ receptormay be a pseudogene. J. Biol. Chem. 272:3622–3627.
Saitou, N., and M. Nei. 1987. The neighbor joining method:a new method for reconstructing phylogenetic trees. Mol.Biol. Evol. 4:406–425.
Salaneck, E., R. Fredriksson, E. T. Larson, J. M. Conlon, and D.Larhammar. 2001. A neuropeptide Y receptor Y1-subfamilygene from an agnathan, the European river lamprey: a potentialancestral gene. Eur. J. Biochem. 268:6146–6154.
Sharman, A. C., and P. W. H. Holland. 1998. Estimation of Hoxgene cluster number in lampreys. Int. J. Dev. Biol. 42:617–620.
Sidow, A. 1996. Gen(om)e duplications in the evolution of earlyvertebrates. Curr. Opin. Genet. Dev. 6:715–722.
Skrabanec, L., and K. H. Wolfe. 1998. Eukaryote genomeduplication—where’s the evidence? Curr. Opin. Genet. Dev.8:694–700.
Smith, N. G. C., R. Knight, and L. D. Hurst. 1999. Vertebrategenome evolution: a slow shuffle or a big bang? BioEssays21:697–703.
Soderberg, C., V. A. Pieribone, J. Dahlstrand, L. Brodin, and D.Larhammar. 1994. Neuropeptide role of both peptide YY andneuropeptide Y in vertebrates suggested by abundantexpression of their mRNAs in a cyclostome brain. J. Neurosci.Res. 37:633–640.
Spivack, W. D., N. Zhong, S. Salerno, R. A. Saavedra, and R. M.Gould. 1993. Molecular cloning of the myelin basic proteinsin the shark, Squalus acanthias, and the ray, Raja erinacia.J. Neurosci. Res. 35:577–584.
Spring, J. 1997. Vertebrate evolution by interspecific hybrid-isation—are we polyploid? FEBS Lett. 400:2–8.
Starback, P., I. Lundell, R. Fredriksson, M. M. Berglund, Y.-L.Yan, A. Wraith, C. Soderberg, J. H. Postlethwait, and D.Larhammar. 1999. Neuropeptide Y receptor subtype withunique properties cloned in the zebrafish: the zYa receptor.Mol. Brain Res. 70:242–252.
Starback, P., A. Wraith, H. Eriksson, and D. Larhammar.2000. Neuropeptide Y receptor gene y6: multiple deathsor resurrection? Biochem. Biophys. Res. Comm. 277:264–269.
Strimmer, K., and A. von Haeseler. 1996. Quartet puzzling:a quartet maximum-likelihood method for reconstructing treetopologies. Mol. Biol. Evol. 13:964–969.
———. 1997. Likelihood mapping: a simple method to visualizephylogenetic content of a sequence alignment. Proc. Natl.Acad. Sci. USA. 94:6815–6819.
Swofford, D. 2000 PAUP*: phylogenetic analysis underparsimony (*and other methods). Version 4.0b8. SinauerAssociates, Sunderland, Mass.
Takezaki, N., A. Rzhetsky, and M. Nei. 1995. Phylogenetic testof the molecular clock and linearized trees. Mol. Biol. Evol.12:823–833.
Taylor, J. S., Y. Van de Peer, I. Braasch, and A. Meyer. 2001.Comparative genomics provides evidence for an ancientgenome duplication event in fish. Phil. Trans. R. Soc. Lond. BBiol. Sci. 356:1661–1679.
Thompson, J. D., D. G. Higgins, and T. J. Gibson. 1994.ClustalW: improving the sensitivity of progressive multiplesequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. NucleicAcids Res. 22:4673–4680.
Wolfe, K. H. 2001. Yesterday’s polyploids and the mystery ofdiploidization. Nat. Rev. Genet. 2:333–341.
Wraith, A., A. Tornsten, P. Chardon, I. Harbitz, B. P.Chowdhary, L. Andersson, L.-G. Lundin, and D. Larhammar.2000. Evolution of the neuropeptide Y receptor family: geneand chromosome duplications deduced from the cloning ofthe five receptor subtype genes in pig. Genome Res. 10:302–310.
Yuan, J., A. Amend, J. Borkowski, R. DeMarco, W. Bailey, Y.Liu, G. Xie, and R. Blevins. 1999. MultiClustal: a systematicmethod for surveying ClustalW alignment parameters. Bio-informatics 15:862–863.
Axel Meyer, Associate Editor
Accepted March 25, 2003
1280 Salaneck et al.
by guest on July 5, 2014http://m
be.oxfordjournals.org/D
ownloaded from
Related Documents











