Molecular and Cellular Pharmacology Third intracellular loop of glucagon like-peptide-1 receptor is coupled with endogenous mono-ADP-ribosyltransferase — Novel type of receptor regulation? Matjaž Deželak, Aljoša Bavec ⁎ Institute of Biochemistry, Faculty of Medicine, University of Ljubljana, Vrazov trg 2, 1000 Ljubljana, Slovenia abstract article info Article history: Received 14 February 2011 Received in revised form 13 May 2011 Accepted 17 May 2011 Available online 27 May 2011 Keywords: Glucagon like peptide-1 receptor Synthetic receptor-loop peptides ADP-ribosylation Posttranslational modification Diabetes Our previous studies revealed the main role of the third intracellular loop (IC 3 ) of glucagon-like peptide-1 receptor (GLP-1 receptor), in G-protein activation, where the presence or absence of agonist and the receptor phosphorylation seemed to be the only regulatory mechanisms. In order to further study the signaling mechanisms of GLP-1 receptor, we investigated the effect of the third intracellular loop-derived peptide on endogenous mono-ADP-ribosyltransferase mediated mono-ADP-ribosylation of G-proteins β subunit in CHO cells. Results showed an inhibitory effect of IC 3 peptide on mono-ADP-ribosylation of β subunit, obviously via the mechanism of competitive inhibition. Excluding the activity of this inhibitory mechanism via pertussis toxin-sensitive G proteins, the direct functional coupling of IC 3 of GLP-1 receptor and endogenous mono-ADP- ribosyltransferase was confirmed. We suggest that this arginine specific enzymatic posttranslational modification of third intracellular loop of GLP-1 receptor might represent a possible novel mechanism of receptor activity regulation and the pharmacological potential in treatment of diabetes mellitus type 2. © 2011 Elsevier B.V. All rights reserved. 1. Introduction The glucagon-like peptide-1 receptor (GLP-1 receptor) belongs to the G-protein coupled receptors and it is a member of secretin/ vasointestinal peptide receptor family B (Dillon et al., 1993; Thorens, 1992). The physiological ligand of GLP-1 receptor is a 30 or 31 amino acid long peptide hormone glucagon like peptide-1 (GLP-1) with effects both in the central nervous system (During et al., 2003; Navaro et al., 1996) and in the periphery part of the body (Barragan et al., 1994; Kreymann et al., 1987). Supporting its therapeutic potential, GLP-1 stimulates insulin secretion not only in normal subjects but also in those with non-insulin-dependent diabetes mellitus type 2, as indicated by clinical studies (Gutniak et al., 1992). GLP-1 receptor mediated signaling involves activation of at least two signaling pathways: i) adenylyl cyclase/cAMP/protein kinase A pathway which increases the level of cAMP and glucose-stimulated insulin secretion (Göke and Conlon, 1988; Wheeler et al., 1993), and ii) phosphatydilinositol 3-kinase/extracellular signal-related kinase/ protein kinase Cξ pathway, which affects β-cell insulin gene transcription and proliferation (Buteau et al., 1999; Edvell and Lindstrom, 1999). Overexpressed GLP-1 receptor couples to different types of α subunits of G-proteins in CHO cells (Montrose-Rafizadeh et al., 1999) and sf9 cells (Bavec et al., 2003). Point and block deletion mutations and peptide based interaction mapping of the receptor revealed the importance of the third intracellular (IC 3 ) loop in activation of G-proteins and adenylyl cyclase (Bavec et al., 2007; Mathi et al., 1997; Takhar et al., 1996). More specifically, our previous study using three synthetic peptide sequences derived from the three intracellular loops of the GLP-1 receptor showed different role of these loops in G-protein coupling; third loop is the main switch that mediates signaling via GLP-1 receptor to G-proteins, while first and second loops are important in discrimination between different types of G-proteins (Bavec et al. 2003). GLP-1 receptor can be negatively modulated by homologous and heterologous desensitization, mechanisms that involve receptor phosphorylation (Widmann et al., 1996). Another possible type of regulation of GLP-1 receptor downstream effects would be via mono- ADP-ribosylation of G-proteins β (Lupi et al., 2002) and α (Di Girolamo and Corda, 2003) subunits, as seen in CHO cells. This reaction is catalyzed by an arginine-specific, plasma membrane associated, mono-ADP-ribosyltransferase (mono-ART) that specifi- cally modifies arginine at position 129, a crucial residue in the common effector-binding surface of the β subunit (Corda and Di Girolamo, 2003). Mono-ADP-ribosylation of this residue prevents β subunit-dependent modulation of effectors such as type 1 adenylyl cyclase (Lupi et al., 2000), phosphoinositide 3-kinase-γ and phos- pholipase C-β2(Lupi et al., 2002). The modified β subunit is further a substrate for a cytosolic mono-ADP-ribosylhydrolase that releases the bound ADP-ribose from the mono-ADP-ribosylated protein. There- fore, these activities are part of a cycle of intracellular mono-ADP- European Journal of Pharmacology 666 (2011) 35–42 ⁎ Corresponding author. Tel.: +386 1 543 76 59; fax: +386 1 543 76 41. E-mail address: [email protected] (A. Bavec). 0014-2999/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2011.05.033 Contents lists available at ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

European Journal of Pharmacology 666 (2011) 35–42
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
Molecular and Cellular Pharmacology
Third intracellular loop of glucagon like-peptide-1 receptor is coupled withendogenous mono-ADP-ribosyltransferase — Novel type of receptor regulation?
Matjaž Deželak, Aljoša Bavec ⁎Institute of Biochemistry, Faculty of Medicine, University of Ljubljana, Vrazov trg 2, 1000 Ljubljana, Slovenia
⁎ Corresponding author. Tel.: +386 1 543 76 59; fax:E-mail address: [email protected] (A. Bavec).
0014-2999/$ – see front matter © 2011 Elsevier B.V. Aldoi:10.1016/j.ejphar.2011.05.033
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 February 2011Received in revised form 13 May 2011Accepted 17 May 2011Available online 27 May 2011
Keywords:Glucagon like peptide-1 receptorSynthetic receptor-loop peptidesADP-ribosylationPosttranslational modificationDiabetes
Our previous studies revealed the main role of the third intracellular loop (IC3) of glucagon-like peptide-1receptor (GLP-1 receptor), in G-protein activation, where the presence or absence of agonist and the receptorphosphorylation seemed to be the only regulatory mechanisms. In order to further study the signalingmechanisms of GLP-1 receptor, we investigated the effect of the third intracellular loop-derived peptide onendogenous mono-ADP-ribosyltransferase mediated mono-ADP-ribosylation of G-proteins β subunit in CHOcells. Results showed an inhibitory effect of IC3 peptide on mono-ADP-ribosylation of β subunit, obviously viathe mechanism of competitive inhibition. Excluding the activity of this inhibitory mechanism via pertussistoxin-sensitive G proteins, the direct functional coupling of IC3 of GLP-1 receptor and endogenous mono-ADP-ribosyltransferase was confirmed. We suggest that this arginine specific enzymatic posttranslationalmodification of third intracellular loop of GLP-1 receptor might represent a possible novel mechanism ofreceptor activity regulation and the pharmacological potential in treatment of diabetes mellitus type 2.
+386 1 543 76 41.
l rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
The glucagon-like peptide-1 receptor (GLP-1 receptor) belongs tothe G-protein coupled receptors and it is a member of secretin/vasointestinal peptide receptor family B (Dillon et al., 1993; Thorens,1992). The physiological ligand of GLP-1 receptor is a 30 or 31 aminoacid long peptide hormone glucagon like peptide-1 (GLP-1) witheffects both in the central nervous system (During et al., 2003; Navaroet al., 1996) and in the periphery part of the body (Barragan et al.,1994; Kreymann et al., 1987). Supporting its therapeutic potential,GLP-1 stimulates insulin secretion not only in normal subjects but alsoin those with non-insulin-dependent diabetes mellitus type 2, asindicated by clinical studies (Gutniak et al., 1992).
GLP-1 receptor mediated signaling involves activation of at leasttwo signaling pathways: i) adenylyl cyclase/cAMP/protein kinase Apathway which increases the level of cAMP and glucose-stimulatedinsulin secretion (Göke and Conlon, 1988; Wheeler et al., 1993), andii) phosphatydilinositol 3-kinase/extracellular signal-related kinase/protein kinase Cξ pathway, which affects β-cell insulin genetranscription and proliferation (Buteau et al., 1999; Edvell andLindstrom, 1999). Overexpressed GLP-1 receptor couples to differenttypes of α subunits of G-proteins in CHO cells (Montrose-Rafizadeh etal., 1999) and sf9 cells (Bavec et al., 2003). Point and block deletion
mutations and peptide based interaction mapping of the receptorrevealed the importance of the third intracellular (IC3) loop inactivation of G-proteins and adenylyl cyclase (Bavec et al., 2007;Mathi et al., 1997; Takhar et al., 1996). More specifically, our previousstudy using three synthetic peptide sequences derived from the threeintracellular loops of the GLP-1 receptor showed different role ofthese loops in G-protein coupling; third loop is the main switch thatmediates signaling via GLP-1 receptor to G-proteins, while first andsecond loops are important in discrimination between different typesof G-proteins (Bavec et al. 2003).
GLP-1 receptor can be negatively modulated by homologous andheterologous desensitization, mechanisms that involve receptorphosphorylation (Widmann et al., 1996). Another possible type ofregulation of GLP-1 receptor downstream effects would be via mono-ADP-ribosylation of G-proteins β (Lupi et al., 2002) and α (DiGirolamo and Corda, 2003) subunits, as seen in CHO cells. Thisreaction is catalyzed by an arginine-specific, plasma membraneassociated, mono-ADP-ribosyltransferase (mono-ART) that specifi-cally modifies arginine at position 129, a crucial residue in thecommon effector-binding surface of the β subunit (Corda and DiGirolamo, 2003). Mono-ADP-ribosylation of this residue prevents βsubunit-dependent modulation of effectors such as type 1 adenylylcyclase (Lupi et al., 2000), phosphoinositide 3-kinase-γ and phos-pholipase C-β2 (Lupi et al., 2002). The modified β subunit is further asubstrate for a cytosolic mono-ADP-ribosylhydrolase that releases thebound ADP-ribose from the mono-ADP-ribosylated protein. There-fore, these activities are part of a cycle of intracellular mono-ADP-

36 M. Deželak, A. Bavec / European Journal of Pharmacology 666 (2011) 35–42
ribosylation and de-mono-ADP-ribosylation, regulating the functionof the βγ dimer. The physiological role of this reaction has beensupported further by the demonstration of its hormonal controlexerted by thrombin, serotonin and cholecystokinin (Lupi et al.,2002).
In this study, we investigated the functional coupling of GLP-1receptor and endogenous mono-ART in CHO cells. In this manner wehave synthesized a 23 amino acid peptide corresponding to sequencederived from the third intracellular loop of GLP-1 receptor (329–351),CIVIAKLKANLMCKTDIKCRLAK, andwe followed the endogenousmono-ART-mediated mono-ADP-ribosylation of β-subunit of G-proteins.
2. Materials and methods
2.1. Cell cultures
Chinese hamster ovary (CHO) cells were grown as a monolayerculture in Dulbecco's modified Eagle's medium (DMEM) supplemen-ted with 10% fetal bovine serum, 0.39% NaHCO3, 58 mg/l proline,53 mg/l aspartatic acid, 59 mg/l glutamatic acid, 60 mg/l asparagine,100 units/ml penicillin, 100 μg/ml streptomycin at 37 °C in a 5%CO2 atm.
2.2. Plasma membrane preparation
CHO plasma membranes were prepared as described previously(Bavec, 2004), but again with minor modifications. Briefly, cells(approximately 2×108) were washed in Hanks' Balanced SaltSolution (HBSS) buffer without Ca2+ and Mg2+, detached byhypotonic buffer containing 10 mM TES (pH 7.5) and 1 mM EDTA.Everything was carried out at 4 °C. Detached cells were homogenizedin Teflon/glass Potter homogenizer and centrifuged at 300×g for10 min. Crude membranes were collected from the supernatant by atwenty-minute spin at 42.000×g. The pellet membranes wereresuspended in 10 mM TES (pH 7.5) containing 0.25 M sucrose,followed by purification of plasma membranes with an aqueous two-phase system with dextran and polyethylene glycol. Crude mem-branes of 1 ml were mixed with 2.72 g of 20% dextran 500, 1.156 g of40% polyethylene glycol 4000, 800 μl of 1 M sucrose, 2.124 ml ofdeionized water, 200 μl 0.2 M potassium phosphate buffer (pH 7.4).After continuous mixing in dextran and polyethylene glycol two-phase system for 15 min, the crude membranes were centrifuged at2500×g for 20 min. The upper dextran phase was transferred into theaqueous two-phase system and extracted for 15 min. After the secondextraction, the crude membranes were centrifuged at 2500×g for20 min, the upper phase containing plasmamembranes, was removedand combined with 5 volumes of 10 mM TES (pH 7.5) buffercontaining 0.25 M sucrose, and centrifuged at 48.000×g for 35 min.The pellet was suspended in 25 mM Hepes (pH 7.5), 150 mM NaCl,1 mM EDTA and protease inhibitors 500 μM O-fenantroline, 2 μMpepstatine and 1 mM phenylmethylsulphonyl fluoride were added.The concentration of proteins in the plasma membranes wasdetermined by Biorad Protein Assay at 595 nm.
2.3. GTPγS binding studies
The rate of [35S]GTPγS binding to G-proteins from plasmamembranes was followed as previously described (Bavec et al.,1999) with minor modification. Briefly, the membranes (final proteinconcentration in the assay mixture in plasma was 500 μg/ml,respectively) were incubated with 5 mM MgCl2, 1 mM dithiothreitol,150 mM NaCl, 1 μM GDP and 0.5–1 nM [35S]GTPγS (approximately140 000 cpm per assay) at 25 °C in TE buffer (pH 7.5), for 2 min withor without peptides. The unbound [35S]GTPγS was washed out byrapid filtration of the reaction mixture through Millipore GF/C glass-fiber filters under vacuum three times with 5 ml of TE buffer. After
extraction of the radioactivematerial overnight in 20 ml of Emulsifier-Safe (Packard, USA) scintillation liquid, radioactivity was determinedwith LKB 1214 Rackbeta liquid scintillation counter. Blank valuesweredetermined by the same procedure in samples in which themembranes were replaced with buffer.
2.4. Mono-ADP-ribosylation assay
ADP-ribosylation followed the incorporation of [32P] ADP-riboseinto the β subunit of heterotrimeric G-protein and was performedwith 5 μg of CHO plasma membrane preparation as describedpreviously (Bavec, 2004) with minor modification. The 50 μl ofplasma membranes in the ADP-ribosylation buffer (50 mM potassiumphosphate buffer (pH=7.5, 0.5 mM MgCl2, 4 mM dithiothreitol,5×106 cpm or 2.3 μCi [32P]NAD+ (specific activity=1000 Ci/mmol)and 6 μM β-NAD+))were incubated at 37 °C for 1 hwith the indicatedpeptides.
When the experiments were performed in the presence ofpertussis toxin (preactivated in 62.5 mM dithiothreitol at roomtemperature for 1 h), 50 μl of reaction mixture (10 μg of plasmamembranes, 13 μg/ml of pertussis toxin, 50 mM Tris (pH=8.0),25 mM dithiothreitol, 0.5 mM EDTA, 1 mM ATP, 5×106 cpm [32P]NAD+, 10 μM β-NAD+) was incubated at 37 °C for 1 h. Afterincubation, the reaction mixture was centrifuged at 10.000×g for15 min. The resulting pellet was washed in 100 μl of 1 M KCl at10.000×g for 15 min, resuspended in 50 μl of ADP-ribosylation bufferand manipulated as the protocol above describes.
The ADP-ribosylation reactions were stopped by diluting thesamples with 50 μl Laemmli buffer, then boiled, and analyzed by 11%SDS-PAGE followed by electroblot onto nitrocellulose membrane. Theradioactivity incorporated into the membrane proteins was detectedand quantified by Instant imager (Packard) or probed with specificanti-β antibodies (1:1000, Santa Cruz).
2.5. Thin layer chromatography
In order to determine non-specific ADP-ribosylation the 50 μl ofADP-ribosylation buffer (50 mM potassium phosphate buffer(pH=7.5), 0.5 mM MgCl2, 4 mM dithiothreitol, 5×106 cpm or2.3 μCi [32P]NAD+ (specific activity=1000 Ci/mmol) and (β-NAD+)) were incubated at 37 °C for 1 h with or without the peptideIC3 and with or without 0.1 U/ml NAD+-glycohydrolase (EC 3.2.2.5).[32P]NAD+ and [32P]ADP-ribose were then resolved by thin layerchromatography in 0.28 M NH4HCO3 for 1 h. The radioactivity wasdetected by Instant Imager (Packard).
2.6. Immunobloting analysis
After the run on SDS-PAGE (Hoefer SE600, 16 cm long) with acurrent of 30 mA (60 mA for two gels) for stacking (2.5 ml 0,5 Tris–HCl, pH=6.8, 1.5 ml 40% acrylamide mix 37.5:1, 100 μl 10% SDS,100 μl 10% ammonium persulphate, 10 μl TEMED, 5.8 ml distillatedwater) and 45 mA (80 mA for two gels) for running gel (7.5 ml 1,5Tris–HCl, pH=8.8, 8.25 ml 40% acrylamide mix 37.5:1, 300 μl 10%SDS, 300 μl 10% ammonium persulphate, 20 μl TEMED, 14.25 mldistillated water) for 2–3 h (EPS 601, Amersham Biosciences) inrunning buffer (3.0 g Tris base, 14.4 g glycine, 1.0 g SDS in 1 ldistillated water), proteins were transferred from the gel ontonitrocellulose membranes with a constant current of 400 mA for 5 h(TE 62 Transphor II) in transfer buffer (1 l methanol plus 3.5 ldistillated water plus 500 ml TB 10x, 60 g Tris base, 288 g glycine).The run and transfer of proteins were cooled at 6 °C (MultiTemp III).The proteins on the blot were visualized with 0.2% Ponceou S in 5%CH3COOH. After staining, the proteins were incubated for 1 h inblocking buffer (2% bovine serum albumin in TBS; 20 mM Tris–HCl,500 mM NaCl; pH=7.5). The blocking buffer was then replaced by a

Fig. 1. Effect of IC3 peptide on mono-ADP-ribosylation of β subunit of G-proteins bymono-ART from CHO cells. (A) Quantitative analysis of mono-ADP-ribosylation of βsubunit. Each bar with the indicated standard deviation represents the mean value offive independent experiments. ANOVA: F(9, 40)=46.21; Pb0,0001 (B) Analysis ofendogenous mono-ADP-ribosylation of β subunit with SDS-PAGE and proteinstransferre from the gel onto nitrocellulose membrane followed by the detection ofradioactivity. (C) Western blot analysis of β subunit (calibrator) with anti-β antibodies.
37M. Deželak, A. Bavec / European Journal of Pharmacology 666 (2011) 35–42
buffer TTBS (0.05% Tween 20 in TBS) and incubated with the primarypolyclonal rabbit IgG anti-β antibodies (T20, 1:1000, Santa Cruz) atroom temperature for 2 h (at this stage blot should be stored in TTBSat 4 °C and is stable for few days). The primary antibodies werewashed three-times in TTBS buffer for 10 min and incubated withsecondary peroxidase-conjugated goat anti-rabbit IgG antibodies(1:1000, Calbiochem) at room temperature for 1 h. After the washin TTBS buffer three-times for 10 min and once in TBS for 10 min, theimmunoreactive proteins caught on the blot were exposed for 15–60 sto Kodak X-Omat film using ECL chemiluminescence detection kit(Amersham Pharmacia Biotech, RPN 2106) (Bavec and Licar, 2009).
2.7. CD spectroscopy
Circular-dichroism measurements were performed on the auto-mated Aviv CD spectrometer model 62A DS at 25 °C. The parametersused were a bandwidth of 1 nm, 1 s averaging time, and step size of a0.5 nm. Peptide was dissolved in 5 mM potassium phosphate(pH=7.4) and measurements at different peptide concentrationswere performed in 10 mm cell (1 μM, 5 μM), 1 mm cell (50 μM) and in0.1 mm demutable cell (500 μM). Spectra at the 50 μM and 500 μMconcentration were scanned between 185 and 250 nm, whereasspectra at the 1 and 5 μM concentration were scanned only between200 and 250 nmdue to strong signal of the 10 mm cell bellow 200 nm.
The average spectra of four measurements were corrected for thebaseline and presented in units of mean residue molar ellipticity.Secondary structure calculations were provided by PROSEC computerprogram supplied with the instrument (Yang et al., 1986).
2.8. Statistical methods
Data are expressed as means±S.D. of at least three independentexperiments. The non-linear least squares fitting of the curves andstatistical analysis (one-way ANOVA) were carried out by PRISM5computer program (GraphPad Software, USA), which was used alsofor graphical presentation of the results. In ANOVA analysis Tukey–Kramer multiple comparison test was used to reveal significantdifferences between groups of interest, PN0.05 was not consideredsignificant.
2.9. Materials
Chinese hamster ovary cells (CHO-K1) were obtained from ECACC,UK. [32P]NAD+, ESL kit and Dextran 500 were supplied fromAmersham Pharmacia Biotech, UK. Hank's Balanced Salt Solution(HBSS) was provided by Gibco, UK and fetal bovine serum was fromBiochrom (Germany). Dulbecco's modified Eagle's medium (DMEM),acrylamide/bis-acrylamide and other electrophoresis reagents werefrom Eurobio, France. Nitrocellulose transfer membranes were fromSchleicher and Schuell, Germany. Ultima Gold scintillation liquid wasfrom Packard, USA. Polyethylene glycol 4000 was from Merck (USA).Rabbit polyclonal anti-β (T-20) and anti-α1 were from Santa CruzBiotechnology (USA). Goat anti-rabbit IgG horseradish peroxidaseconjugates were from Calbiochem, USA. All other chemicals werefrom Sigma Aldrich Co, USA and were of analytical grade.
3. Results
3.1. Effect of IC3 peptide on mono-ADP-ribosylation of β subunit ofG-proteins by mono-ART from CHO cells
Endogenous mono-ART is well expressed in plasma membranefrom CHO cells (Lupi et al. 2000). The best characterized substrate bydate of this arginine-specific mono-ART is β subunit of heterotrimericG-proteins in plasma membranes. At first, activity of aforementionedendogenous mono-ARTwas followed bymono-ADP-ribosylation of β-
subunit in the presence or absence of IC3 peptide. The mono-ADP-ribosylation protein bandswith an apparentmolecularmass≈36 kDacorresponded to β subunit of G-proteins and its mono-ADP-ribosyla-tion was carefully quantified (Fig. 1A and B). They were additionallyconfirmed by ECL Western blot analysis using anti-β antibodies(Fig. 1C).
As expected, the effect of IC3 was dose-dependent but surprisingly,there was obvious deviation from expected relation. IC3 shouldstimulate mono-ADP-ribosylation of β subunit in single-phase dose-dependent manner by mimicking the activated receptor and thusincreasing the level of free βγ dimer. No stimulatory effect wasobserved at all tested IC3 concentrations; at low concentration there isno statistical significant effect relative to basal and at higher IC3
concentration rate of mono-ADP-ribosylation even begin to decreaseexceedingly at 10 μM and higher. At 50 μM mono-ADP-ribosylationof β subunit was even lower than the basal (61% of basal) and itdecreased further at 100 and 200 μM (48% and 30% of basal,respectively) (Fig. 1A).
3.2. Effect of IC3 peptide on mono-ADP-ribosylation of β subunit of G-proteins by mono-ART from CHO cells in presence or absence of pertussistoxin
To further investigate mechanism of IC3 peptide action on mono-ADP-ribosylation of β subunit, pertussis toxin was used together withdifferent concentration of IC3 peptide to show if enzymatic inhibitionis mediated through G0/i proteins. If α subunit of G0/i proteins isinvolved in inhibition of mono-ART there would not be any inhibitionafter treatment with pertussis toxin. But, as it can be noticed fromFig. 2A and B, pertussis toxin has no significant effect on mono-ADP-ribosylation of β subunit. This result suggests direct interaction of IC3
with mono-ART. The expression of the β subunit during theexperiment was unchanged (Fig. 2C).
3.3. Mono-ADP-ribosylation of IC3 peptide by endogenous mono-ARTfrom plasma membrane of CHO cells
Next we tested most the probable explanation of the apparentinhibition of mono-ADP-ribosylation of β subunit by IC3 peptide.

Fig. 2. Effect of IC3 peptide onmono-ADP-ribosylation of β subunit of G-proteins by mono-ART from CHO cells in presence or absence of PTX. (A) Quantitative analysis of mono-ADP-ribosylation of β subunit. Each bar with the indicated standard deviation represents the mean value of three independent experiments. (B) Analysis of endogenous mono-ADP-ribosylation of β subunit with SDS-PAGE and proteins transferre from the gel onto nitrocellulose membrane followed by the detection of radioactivity. (C) Western blot analysis of βsubunit (calibrator) with anti-β antibodies.
38 M. Deželak, A. Bavec / European Journal of Pharmacology 666 (2011) 35–42
Arginine-specific mono-ART participates inmono-ADP-ribosylation ofβ subunit and since IC3 peptide contains one arginine (Arg348 in nativereceptor) as well, the competitive inhibition was hypothesized. In thismannerwe analyze if mono-ADP-ribosylation of IC3 peptide occur andthe results are shown in Fig. 3A and B (empty bars). Band of apparentmolecular mass of ~3.0 kDa (which is sum of Mw(IC3)=2577 Da andMw(ADP-ribose)=576) corresponded to ADP-ribosylated IC3 peptideand single-phase dose-dependent relation was prominent. Addition-ally, meta-iodobenzylguanidine (MIBG), a competitive inhibitor ofarginine-dependent mono-ADP-ribosylation, prevented mono-ADP-ribosylation of IC3 peptide (Fig. 3A and B, patterned bars). Thin layerchromatography revealed nor non-specific binding of [32P]β-NAD+
Fig. 3. Mono-ADP-ribosylation of IC3 peptide with or without MIBG and IC3 (R348A)peptide by endogenous mono-ART from plasma membrane of CHO cells.(A) Quantitative analysis of mono-ADP-ribosylation of IC3 (empty bars), IC3 (R348A)(filled bars) and IC3+MIBG (patterned bars). Each bar with the indicated standarddeviation represents the mean value of three independent experiments. (B) Analysis ofendogenous mono-ADP-ribosylation of IC3 (empty), IC3 (R348A) (filled) and IC3+MIBG (patterned) with SDS-PAGE and proteins transferre from the gel ontonitrocellulose membranes followed by the detection of radioactivity. (C) Westernblot analysis of β subunit (calibrator) with anti-β antibodies.
(Fig. 4, Lane 2) neither non-specific binding of [32P]ADP-ribose ornon-enzymatic modification by [32P]ADP-ribose of IC3 (Fig. 4, Lane 4).These results strongly suggest the competitive inhibition of mono-ADP-ribosylation of β subunit by IC3 peptide.
3.4. Mono-ADP-ribosylation of IC3(R348A) peptide and its effect onmono-ADP-ribosylation of β subunit of G proteins by endogenous mono-ART from plasma membrane of CHO cells
If competitive inhibition is present, arginine to alanine mutant ofIC3 peptide should exert drastic decline in rate of its mono-ADP-ribosylation. Moreover, effect of IC3(R348A) on mono-ADP-ribosyla-tion of β subunit should not be significant; no inhibition should beobserved because mono-ADP-ribosylation site is absent. As it is clearfrom Fig. 5, increasing concentrations of IC3(R348A) peptide has nodistinctive effect on mono-ADP-ribosylation of β subunit in compar-ison with basal and statistical analysis of data did not reveal anysignificant differences between groups of interest.
Assessment of IC3(R348A) peptide mono-ADP-ribosylation givesmore or less expected results (Fig. 3A and B, filled bars). Althoughdecrease is not complete, it is still very significant; the rate of mono-ADP-ribosylation of IC3(R348A) is ~50% or less of value for IC3 peptide.Nevertheless, these results indicate involvement of at least one othertype of binding; probably an enzymatic modification of other aminoacid residues.
Fig. 4.Determination of non-enzymatic mono-ADP-ribosylation of IC3 peptide with thinlayer chromatography. Lane 1: [32P]β-NAD+. Lane 2: [32P]β-NAD++IC3 peptide. Lane3: [32P]β-NAD++NAD-ase. Lane 4: [32P]β-NAD++NAD-ase+IC3 peptide. One ofthree independent experiments is shown in the figure.
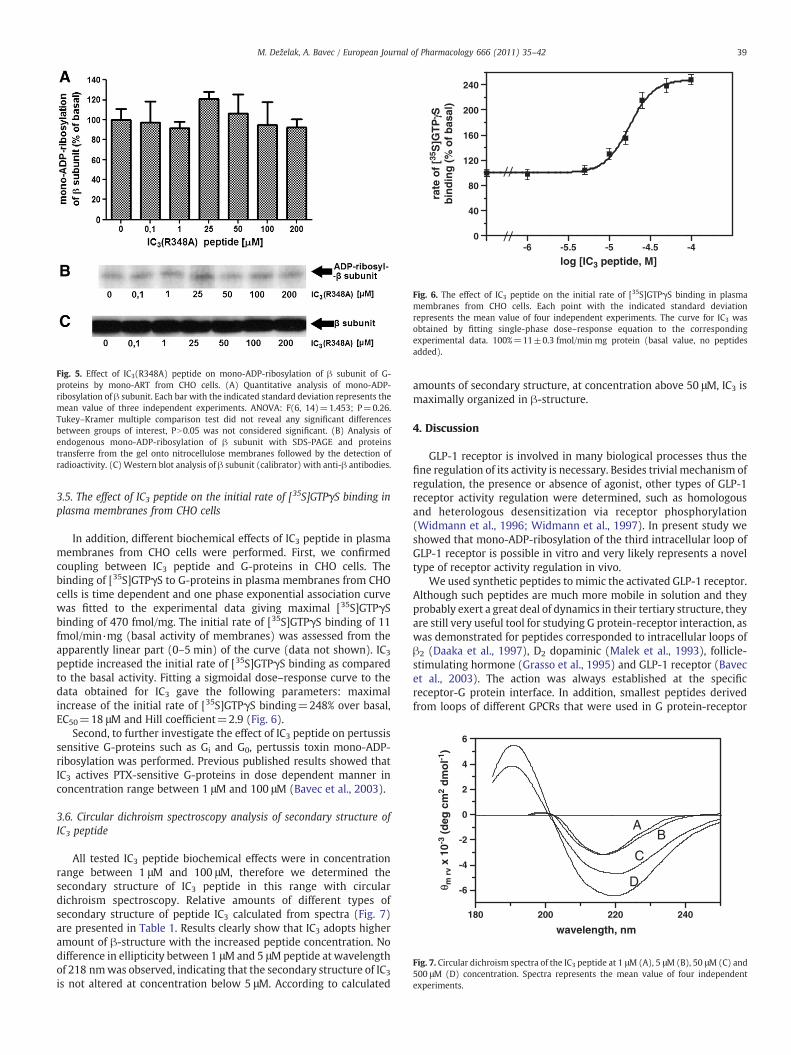
Fig. 5. Effect of IC3(R348A) peptide on mono-ADP-ribosylation of β subunit of G-proteins by mono-ART from CHO cells. (A) Quantitative analysis of mono-ADP-ribosylation of β subunit. Each bar with the indicated standard deviation represents themean value of three independent experiments. ANOVA: F(6, 14)=1.453; P=0.26.Tukey–Kramer multiple comparison test did not reveal any significant differencesbetween groups of interest, PN0.05 was not considered significant. (B) Analysis ofendogenous mono-ADP-ribosylation of β subunit with SDS-PAGE and proteinstransferre from the gel onto nitrocellulose membranes followed by the detection ofradioactivity. (C) Western blot analysis of β subunit (calibrator) with anti-β antibodies.
0
40
80
120
160
200
240
-6 -5.5 -5 -4.5 -4
log [IC3 peptide, M]
rate
of
[35S
]GT
PγS
bin
din
g (
% o
f b
asal
)
Fig. 6. The effect of IC3 peptide on the initial rate of [35S]GTPγS binding in plasmamembranes from CHO cells. Each point with the indicated standard deviationrepresents the mean value of four independent experiments. The curve for IC3 wasobtained by fitting single-phase dose–response equation to the correspondingexperimental data. 100%=11±0.3 fmol/min mg protein (basal value, no peptidesadded).
180 200 220 240
-6
-4
-2
0
2
4
6
wavelength, nm
B
C
D
A
θ m r
v x
10-3
(d
eg c
m2
dm
ol-1
)
Fig. 7. Circular dichroism spectra of the IC3 peptide at 1 μM (A), 5 μM (B), 50 μM (C) and500 μM (D) concentration. Spectra represents the mean value of four independentexperiments.
39M. Deželak, A. Bavec / European Journal of Pharmacology 666 (2011) 35–42
3.5. The effect of IC3 peptide on the initial rate of [35S]GTPγS binding inplasma membranes from CHO cells
In addition, different biochemical effects of IC3 peptide in plasmamembranes from CHO cells were performed. First, we confirmedcoupling between IC3 peptide and G-proteins in CHO cells. Thebinding of [35S]GTPγS to G-proteins in plasma membranes from CHOcells is time dependent and one phase exponential association curvewas fitted to the experimental data giving maximal [35S]GTPγSbinding of 470 fmol/mg. The initial rate of [35S]GTPγS binding of 11fmol/min·mg (basal activity of membranes) was assessed from theapparently linear part (0–5 min) of the curve (data not shown). IC3
peptide increased the initial rate of [35S]GTPγS binding as comparedto the basal activity. Fitting a sigmoidal dose–response curve to thedata obtained for IC3 gave the following parameters: maximalincrease of the initial rate of [35S]GTPγS binding=248% over basal,EC50=18 μM and Hill coefficient=2.9 (Fig. 6).
Second, to further investigate the effect of IC3 peptide on pertussissensitive G-proteins such as Gi and G0, pertussis toxin mono-ADP-ribosylation was performed. Previous published results showed thatIC3 actives PTX-sensitive G-proteins in dose dependent manner inconcentration range between 1 μM and 100 μM (Bavec et al., 2003).
3.6. Circular dichroism spectroscopy analysis of secondary structure ofIC3 peptide
All tested IC3 peptide biochemical effects were in concentrationrange between 1 μM and 100 μM, therefore we determined thesecondary structure of IC3 peptide in this range with circulardichroism spectroscopy. Relative amounts of different types ofsecondary structure of peptide IC3 calculated from spectra (Fig. 7)are presented in Table 1. Results clearly show that IC3 adopts higheramount of β-structure with the increased peptide concentration. Nodifference in ellipticity between 1 μMand 5 μMpeptide at wavelengthof 218 nmwas observed, indicating that the secondary structure of IC3
is not altered at concentration below 5 μM. According to calculated
amounts of secondary structure, at concentration above 50 μM, IC3 ismaximally organized in β-structure.
4. Discussion
GLP-1 receptor is involved in many biological processes thus thefine regulation of its activity is necessary. Besides trivial mechanism ofregulation, the presence or absence of agonist, other types of GLP-1receptor activity regulation were determined, such as homologousand heterologous desensitization via receptor phosphorylation(Widmann et al., 1996; Widmann et al., 1997). In present study weshowed that mono-ADP-ribosylation of the third intracellular loop ofGLP-1 receptor is possible in vitro and very likely represents a noveltype of receptor activity regulation in vivo.
We used synthetic peptides to mimic the activated GLP-1 receptor.Although such peptides are much more mobile in solution and theyprobably exert a great deal of dynamics in their tertiary structure, theyare still very useful tool for studying G protein-receptor interaction, aswas demonstrated for peptides corresponded to intracellular loops ofβ2 (Daaka et al., 1997), D2 dopaminic (Malek et al., 1993), follicle-stimulating hormone (Grasso et al., 1995) and GLP-1 receptor (Bavecet al., 2003). The action was always established at the specificreceptor-G protein interface. In addition, smallest peptides derivedfrom loops of different GPCRs that were used in G protein-receptor

Table 1Relative proportion of different types of secondary structure of peptide derived fromthe third intracellular loop of the glucagon-like peptide-1 receptor peptide (IC3) at1 μM, 5 μM, 50 μM, and 500 μM concentration.
Secondary structure Relative proportion of different types of secondary structure
1 μM IC3 5 μM IC3 50 μM IC3 500 μM IC3
α-helix 7% 8% 5% 7%β-structure 27% 28% 66% 63%Random coil 32% 31% 29% 30%β-turn 34% 33% 0% 0%
Fig. 8. Comparison of amino acid residues in the same position relative to modifiedarginine in β subunit and IC3 peptide. The comparison was performed in view ofgeneral, qualitative similarity between side chains structure and electric chargedisposition. Double underlined residue: mono-ADP-ribosylated residue. Solid frame:high degree of analogy. Dotted frame: low degree of analogy. No frame: no apparentanalogy.
40 M. Deželak, A. Bavec / European Journal of Pharmacology 666 (2011) 35–42
interactions studies were 9–15 amino acids in length (Grasso et al.1995; Okamoto and Nishimoto, 1992), which is extra confirmation ofour peptides suitability.
Intracellular activation of G proteins via GLP-1 receptor is tightlyregulated by structural determinants primarily located to the N-terminus of the third intracellular loop of receptor (Mathi et al., 1997).Synthetic peptide IC3 efficiently imitates activated GLP-1 receptor(Bavec et al. 2003), causing the GDP to GTP exchange at α subunit inαβγ trimer and consequential dissociation toα subunit and βγ dimer.β subunit in βγ dimer is substrate for mono-ADP ribosylation byendogenous mono-ART. The increasing IC3 concentrations shouldraise the amount of mono-ADP-ribosylated β subunit via raising theamount of the substrate. In such expected relation an ascendingsigmoidal dose–response curve is suitable. But as can be noticed fromFig. 1, the results show a descending sigmoidal dose–response curve.However, the model of an ascending sigmoidal dose–responserelation is still very likely valid, as can be concluded from Fig. 6, butit seems that some other mechanism of suppressing mono-ADP-ribosylation of β subunit exists and it predominates at tested IC3
concentrations range. In comparison with basal mono-ADP-ribosyla-tion rate of β subunit this inhibitory mechanism even lowers it at50 μM IC3 or higher (Fig. 1A and B). It seems the inhibition is weak(EC50 is about 25–50 μM), but it correlates well with binding affinityof IC3 (EC50=18 μM) to G-proteins (Fig. 6). Apparently similar affinityof IC3 as obtained in our case to G-proteins was observed also withother loop peptides (Bavec et al. 2003), such as the third intracellularloop peptide of human α2-adrenergic receptor (Okamoto andNishimoto, 1992), which means that the specific effective concentra-tion for loop peptides is in micromolar range.
We tested if G0/i proteins are involved in inhibition of mono-ART,similarly as it was determined for inhibition of adenylyl cyclase (Okaet al., 1997). If that would be the case, no reduction of mono-ADP-ribosylation of β subunit should be observed, compared to basalbecause subunits of G0/i-proteins would be locked in their trimericform (Abood et al., 1987). Fig. 2 clearly shows that inactivation of G0/i
proteins did not affect mono-ADP-ribosylation of β subunit whichmeans that mono-ART is not inhibited by G0/i proteins. Nevertheless, asmall but not statistically significant reductionwas observed in case ofPTX presence (Fig. 2B), as expected, due to a reduction of free βγdimer because PTX is locking it up in αβγ trimer (Abood et al., 1987).In short, it is evident that the mono-ADP-ribosylation of β subunit isdependent by IC3 concentration, where the inhibiting effect isprevalent.
Then we analyzed amino acids near the mono-ADP-ribosylatedamino acid residue in the β subunit (Arg129) and near the proposedmodifiedmono-ADP-ribosylated amino acid residue in the IC3 peptide(Arg348 in native receptor). Some qualitative similarities regarding tothe structure and electric charge disposition of amino acid residuesside chains were observed (Fig. 8), supporting Arg348 as possible siteof modification. Based on the structure (Jørgensen et al. 2008), theinteraction between mono-ADP-ribosyltransferase and substrate isbased on not only the sequence but also interface secondary structure.If so, IC3 peptide adopts higher amount of β-structure (Fig. 7) whichmight be involved in substrate recognition by the enzyme. Thus we
followed mono-ADP-ribosylation of IC3 peptide with detection ofradioactivity and band of apparent molecular mass of ~3.0 kDa wasobserved which corresponded to ADP-ribosylated IC3 peptide (Fig. 3,empty bars). Non-enzymatic mono-ADP-ribosylation of IC3 peptidewas not observed (Fig. 4). Therefore, a specific enzymatic process isnecessary for IC3 mono-ADP-ribosylation.
Since β subunit is covalently modified at Arg129 by mono-ART, theArg348 of IC3 peptide seems to be the only suitable mono-ADP-ribosylated amino acid residue for specific enzymatic modification. Inthis manner we synthesized amutant form of IC3 peptide, IC3(R348A),having an alanine instead of an arginine. Mono-ADP-ribosylation ofIC3(R348A) and β subunit were followed to clarify the role of Arg348 ininhibition process. Comparison of filled and empty bars in Fig. 3clearly shows a drastic decline of IC3(R348A) mono-ADP-ribosylation.Another confirmation that both β subunit and IC3 peptide are mono-ADP-ribosylated by same mono-ART is inhibition of mono-ADP-ribosylation of IC3 peptide by MIBG (Fig. 3A, patterned bars), a wellknown competitive inhibitor of mono-ART (Kharadia et al. 1992).Similar rate of inhibition was shown for mono-ADP-ribosylation of βsubunit too (data not shown). However, the inhibition of mono-ADP-ribosylation of IC3(R348A) is not absolutely complete which could beinterpreted as an existence of enzymatic modification of other aminoacid residues, e.g. cysteine, and asparagine (Okazaki and Moss, 1999).Cysteine is besides arginine also well defined site for mono-ADP-ribosylation. Action of some cysteine-specific endogenous mono-ARTcould be in place, but it is also possible that the same mono-ART thatmodifies arginines in β subunit and IC3 peptide modifies cysteine inIC3 as well (Han et al., 2001; Koch-Nolte et al., 2001; Maehama et al.,1996; Tsuge et al., 2008). Possible mono-ADP-ribosylation of cysteineresidue is currently under investigation.
If competitive inhibition is present, no decline in mono-ADP-ribosylation of β subunit in presence of IC3(R348A) should beobserved. Obviously, there is neither inhibition of mono-ADP-ribosylation of β subunit – a finding that supports IC3 as competitiveinhibitor – nor stimulation (Fig. 5). Absence of stimulation couldmeanthat ability of IC3(R348A) tomimic the activated receptor was lost, butthat is not the case since our previous study (Bavec et al., 2007)showed that ability of IC3(R348A) peptide to mimic the activatedreceptor is comparable with IC3 peptide for Gs, Gi1, G0 and G11
proteins. Another explanation would be still present competitiveinhibition as a consequence of mono-ADP-ribosylation of other aminoacid residues.
It is notorious that there is a high degree of correlation between astructure and function of biological active molecules. IC3 peptideexerts its competitive inhibition on mono-ADP-ribosylation of βsubunit in micromolar range around 50 μM (Fig. 3, EC50=25–50 μM).We have determined, this is the active amount of peptide in otherbiological processes in vitro, such as [35S]GTPγS binding to G-proteins(Fig. 6, EC50=18 μM). The same IC3 concentration range near 50 μMseems to be effective also for PTX-mediated mono-ADP-ribosylationof α subunit of G0/i-proteins (Bavec et al. 2003). In this manner weperformed a circular dichroism (CD) spectroscopy analysis ofsolutions containing different amount of IC3 peptide (Table 1.).According to the acquired data a change in the relative amount of

41M. Deželak, A. Bavec / European Journal of Pharmacology 666 (2011) 35–42
different types of secondary structures of IC3 happens round 50 μM,i.e. ratio of β-structures double and β-turns vanish. Therefore, moreorganized structure of IC3 from GLP-1 receptor seems to be necessaryfor efficient signal passing downstream to the G-proteins and it ispossible that at concentrations round 50 μM IC3 structure is changedfrom “switch-off” to “switch-on” conformation that stimulates G-proteins by increasing the rate of [35S]GTPγS binding (Fig. 6), affectsthe PTX-sensitive G-proteins (Bavec et al. 2003) and mono-ADP-ribosylation of β subunit (Fig. 1).
In present study we demonstrated functional coupling of the thirdintracellular loop of GLP-1 receptor with mono-ART in CHO cells.Third intracellular loop of GLP-1 receptor works as competitivesubstrate for mono-ART in an enzymatic process, which reducesmono-ADP-ribosylation of β subunit in vitro. This posttranslationalmodification of third loop of GLP-1 receptor could present a noveltype of regulation in cellular processes such as homologous orheterologous desensitization via receptor ADP-ribosylation in vivo.Therefore, mono-ART might be a novel pharmacological target intreatment of diabetes mellitus type 2. GLP-1 stimulates insulinsecretion in subjects with non-insulin-dependent diabetes mellitustype 2 (Gutniak et al., 1992). Molecular mechanism of insulinsecretion by GLP-1 involves activation of GLP-1 receptor-G protein-adenylyl cyclase pathway (Göke and Conlon, 1988; Wheeler et al.,1993). Mono-ADP-ribosylation by mono-ART prevents G protein βγdimer dependent modulation of effectors such as adenylyl cyclase(Lupi et al., 2000). IC3 peptide inhibits mono-ART, in theory; βγ dimeris free to activate adenylyl cyclase type 2 and potentiates insulinrelease in patients with diabetes mellitus type 2. Recently, we havesynthesized and characterized GLP-1 receptor fused with derivativesof GFP in CHO cells (Bavec and Licar, 2009). Using these biologicalfluorescent markers together with different biochemical approaches(Mars et al., 2003; Golicnik et al. 2001; Debeljak and Sytkowski, 2008;Dani et al., 2009) on living cells will give usmore realistic view on verylikely GLP-1 receptor regulation by mono-ART, especially withthoughts on its role in diabetes mellitus type 2.
Conflict of interest statement
All the authors declare that there are no conflicts of interest.
Acknowledgments
We thank Daniela Corda (Napoli, Italy) and Mariela Di Girolamo(Santa Maria Imbaro, Italy) because they were so kind to teach usADP-ribosylation assay. This work was supported by the Ministry ofScience and Technology of the Republic of Slovenia, Grant No. P1-0170.
References
Abood, M.E., Lee, N.M., Loh, H.H., 1987. Modification of opioid agonist binding bypertussis toxin. Brain Res. 417, 70–74.
Barragan, J.M., Rodriguez, R.E., Blazquez, E., 1994. Changes in arterial blood pressureand heart rate induced by glucagon-like peptide-1-(7–36) amide in rats. Am. J.Physiol. 266, 459–466.
Bavec, A., 2004. Novel features of amphiphilic peptide Mas7 in signalling viaheterotrimeric G-proteins. J. Pept. Sci. 10, 691–699.
Bavec, A., Licar, A., 2009. Functional characterization of N-terminally GFP-tagged GLP-1receptor. J. Biomed. Biotechnol. doi:10.1155/2009/498149.
Bavec, A., Juréus, A., Cigić, B., Langel, U., Zorko, M., 1999. Peptitergent PD1 affects theGTPase activity of rat brain cortical membranes. Peptides 20, 177–184.
Bavec, A., Hällbrink, M., Langel, Ü., Zorko, M., 2003. Different role of intracellular loopsof glucagon-like peptide-1 receptor in G-protein coupling. Reg. Peptides 111,137–144.
Bavec, A., Jiang, Y., Langel, U., Zorko, M., 2007. Role of cysteine 341 and arginine 348 ofGLP-1 receptor in G-protein coupling. Mol. Biol. Rep. 34, 53–60.
Buteau, J., Roduit, R., Susini, S., Prentki, M., 1999. Glucagon-like peptide-1 promotesDNA synthesis, activates phosphatidylinositol 3-kinase and increases transcriptionfactor pancreatic and duotenal homeobox gene 1 (PDX-1) DNA binding activity inβ(INS-1)-cells. Diabetologia 42, 856–864.
Corda, D., Di Girolamo, M., 2003. Functional aspects of protein mono-ADP-ribosylation.EMBO J. 22, 1953–1958.
Daaka, Y., Luttrell, L., Lefkowitz, R.J., 1997. Switching of the coupling of the β2-adrenergic receptor to different G proteins by protein kinase A. Nature 390, 88–91.
Dani, N., Stilla, A., Marchegiani, A., Tamburro, A., Till, S., Ladurner, A.G., Corda, D., DiGirolamo, M., 2009. Combining affinity purification by ADP-ribose-binding macrodomains with mass spectrometry to define the mammalian ADP-ribosyl proteome.Proc. Natl. Acad. Sci. U. S. A. 106, 4243–4248.
Debeljak, N., Sytkowski, A.J., 2008. Erythropoietin: new approaches to improvedmolecular designs and therapeutic alternatives. Curr. Pharma. Des. 14, 1302–1310.
Di Girolamo, M., Corda, D., 2003. Mono-ADP-Ribosylation of Heterotrimeric G Proteins.In: Bradshaw, R., Dennis, E. (Eds.), Handbook of Cell Signalling. Academic Press, SanDiego, pp. 613–618.
Dillon, J.S., Tanizawa, Y., Wheeler, M.B., Leng, X.H., Ligon, B.B., Rabin, D.U., Yoo, W.H.,Permutt, M.A., Boyd, A.D., 1993. Cloning and functional expression of the humanglucagon-like peptide-1 (GLP-1) receptor. Endocrinology 133, 1907–1910.
During, M.J., Cao, L., Zuzga, D.S., Francis, J.S., Fitzsimons, H.L., Jiao, X., Bland, R.J.,Klugmann, M., Banks, W.A., Drucker, D.J., Haile, C.N., 2003. Glucagon-like peptide-1receptor is involved in learning and neuroprotection. Nat. Med. 9, 1173–1179.
Edvell, A., Lindstrom, P., 1999. Initiation of increased pancreatic islet growth in youngnormoglycemic mice (Umea +/?). Endocrinology 140, 778–783.
Göke, R., Conlon, J.M., 1988. Receptors for glucagon-like peptide-1(7–36)amide on ratinsulinoma-derived cells. J. Endocrinol. 116, 357–362.
Golicnik, M., Fournier, D., Stojan, J., 2001. Interaction of Drosophila acetylcholineesterases with D-tubocurarine: an explanation of the activation by an inhibitor.Biochemistry 40, 1214–1219.
Grasso, P., Deziel, M.R., Reichert, L.E., 1995. Synthetic peptides corresponding toresidues 551 to 555 and 650 to 653 of the rat testicular follicle-stimulatinghormone (FSH) receptor are sufficient for post-receptor modulation of Sortoli cellresponsiveness to FSH stimulation. Regul. Pept. 60, 177–183.
Gutniak, M., Orskov, C., Holst, J.J., Ahrén, B., Efendic, S., 1992. Antidiabetogenic effect ofglucagon-like peptide-1(7–36)amide in normal subjects and patients withmellitus. N. Engl. J. Med. 326, 1316–1322.
Han, S., Arvai, A.S., Clancy, S.B., Tainer, J.A., 2001. Crystal structure and novel recognitionmotif of rho ADP-ribosylating C3 exoenzyme from Clostridium botulinum:structural insights for recognition specificity and catalysis. J. Mol. Biol. 305, 95–107.
Jørgensen, R., Wang, Y., Visschedyk, D., Merrill, A.R., 2008. The nature and character ofthe transition state for the ADP-ribosyltransferase reaction. EMBO Rep. 9,802–809.
Kharadia, S.V., Huiatt, T.W., Huang, H.Y., Peterson, J.E., Graves, D.J., 1992. Effect of anarginine-specific ADP-ribosyltransferase inhibitor on differentiation of embryonicchick skeletal muscle cells in culture. Exp. Cell Res. 201, 33–42.
Koch-Nolte, F., Reche, P., Haag, F., Bazan, F., 2001. ADP-ribosyltransferases: plastic toolsfor inactivating protein and small molecular weight targets. J. Biotechnol. 92,81–87.
Kreymann, B., Williams, G., Ghatei, M.A., Bloom, S.R., 1987. Glucagon-like peptide-1 7–36: a physiological incretin in man. Lancet 2, 1300–1304.
Lupi, R., Corda, D., Di Girolamo, M., 2000. Endogenous ADP-ribosylation of the G proteinbeta subunit prevents the inhibition of type 1 adenylyl cyclase. J. Biol. Chem. 275,9418–9424.
Lupi, R., Dani, N., Dietrich, A., Marchegiani, A., Turacchio, S., Berrie, C.P., Moss, J.,Gierschik, P., Corda, D., Di Girolamo, M., 2002. Endogenous mono-ADP-ribosylationof the free Gβγ prevents stimulation of phosphoinositide 3-kinase-γ andphospholipase C-β2 and is activated by G-protein-coupled receptors. Biochem. J.367, 825–832.
Maehama, T., Hoshino, S., Katada, T., 1996. Increase in ADP-ribosyltransferase activity ofrat T lymphocyte alloantigen RT6.1 by a single amino acid mutation. FEBS Lett. 388,189–191.
Malek, D., Münch, G., Palm, D., 1993. Two sites in the third intracellular loop of the D2receptor are involved in functional G protein-mediated coupling to adenylylatecyclase. FEBS Lett. 325, 215–219.
Mars, T., King, M.P., Miranda, A.F., Walker, W.F., Mis, K., Grubic, Z., 2003. Functionalinnervation of cultured human skeletal muscle proceeds by twomodes with regardto agrin effects. Neuroscience 118, 87–97.
Mathi, S.K., Chan, Y., Li, X., Wheeler, M.B., 1997. Scanning of the glucagon-like peptide-1receptor localizes G protein-activating determinants primarily to the N terminus ofthe third intracellular loop. Mol. Endocrinol. 11, 424–432.
Montrose-Rafizadeh, C., Avdonin, P., Garant, M.J., Rodgers, B.D., Kole, S., Yang, H., Levine,M.A., Schwindinger, W., Bernier, M., 1999. Pancreatic glucagon-like peptide-1receptor couples to multiple G proteins and activate mitogen-activated proteinkinase pathways in chine hamster ovary cells. Endocrinology 140, 1132–1140.
Navarro, M., Rodriquez de Fonseca, F., Alvarez, E., Chowen, J.A., Zueco, J.A., Gomez, R.,Eng, J., Blázquez, E., 1996. Colocalization of glucagon-like peptide-1 (GLP-1)receptors, glucose transporter GLUT-2, and glucokinase mRNAs in rat hypothalamiccells: evidence for a role of GLP-1 receptor agonists as an inhibitory signal for foodand water intake. J. Neurochem. 67, 1982–1991.
Oka, M., Itoh, Y., Shimidzu, T., Ukai, Y., Yoshikuni, Y., Kimura, K., 1997. Involvement ofmetabotropic glutamate receptors in Gi- and Gs-dependent modulation ofadenylate cyclase activity induced by a novel cognition enhancer NS-105 in ratbrain. Brain Res. 754, 121–130.
Okamoto, T., Nishimoto, I., 1992. Detection of G-protein-activator regions in M4subtype muscarinic, cholinergic, and alpha 2-adrenergic receptors based uponcharacteristics in primary structure. J. Biol. Chem. 267, 8342–8346.
Okazaki, I.J., Moss, J., 1999. Characterization of glycosylphosphatidylinositol-anchored,secreted and intracellular vertebrate mono-ADP-ribosyltransferases. Annu. Rev.Nutr. 19, 485–509.

42 M. Deželak, A. Bavec / European Journal of Pharmacology 666 (2011) 35–42
Takhar, S., Gyomorey, S., Su, R.C., Mathi, S.K., Li, X.F., Wheeler, M.B., 1996. The thirdcytoplasmic domain of the GLP-1[7–36]amide receptor is required for coupling tothe adenylyl cyclase system. Endocrinology 137, 2175–2180.
Thorens, B., 1992. Expression cloning of the pancreatic beta cell receptor for the gluco-incretin hormone glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 89, 8641–8645.
Tsuge, H., Nagahama, M., Oda, M., Iwamoto, S., Utsunomiya, H., Marquez, V.E.,Katunuma, N., Nishizawa, M., Sakurai, J., 2008. Structural basis of actin recognitionand arginine ADP-ribosylation by Clostridium perfringens iota-toxin. Proc. Natl.Acad. Sci. USA 105, 7399–7404.
Wheeler, M.B., Lu, M., Dillon, J.S., Leng, X.H., Chen, C., Boyd III, A.E., 1993. Functionalexpression of the rat glucagon-like peptide-1 receptor, evidence for coupling toboth adenylyl cyclase and phospholipase-C. Endocrinology 133, 57–62.
Widmann, C., Dolci, W., Thorens, B., 1996. Heterologous desensitization of theglucagon-like peptide-1 receptor by phorbol esters requires phosphorylation ofthe cytoplasmic tail at four different sites. J. Biol. Chem. 271, 19957–19963.
Widmann, C., Dolci, W., Thorens, B., 1997. Internalization and homologous desensiti-zation of the GLP-1 receptor depend on phosphorylation of the receptor carboxyltail at the same three sites. Mol. Endocrinol. 11, 1094–1102.
Yang, J.T., Wu, C.S., Martinez, H.M., 1986. Calculation of protein conformation fromcircular dichroism. Meth. Enzymol. 130, 208–269.
Related Documents











