THÈSE DE DOCTORAT Soutenue à Aix-Marseille Université le 09 avril 2021 par Evgeniia Rusina Focal seizure induction using deep brain stimulation via temporally interfering electric fields Discipline Biologie santé Composition du jury Spécialité Neurosciences Brian LITT University of Pennsylvania Lyle MULLER Western University Rapporteur Rapporteur École doctorale Sciences de la vie et de la santé / ED 62 Mary DONAHUE Linkoping University Yuri ZILBERTER Aix-Marseille Université Examinatrice/ présidente du jury Directeur du thèse Laboratoire/Partenaires de recherche PhysioNet Adam WILLIAMSON Aix-Marseille Université Co-directeur du thèse

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THÈSE DE DOCTORATSoutenue à Aix-Marseille Université le 09 avril 2021 par
Evgeniia Rusina
Focal seizure induction using deep brain stimulation via temporally interfering
electric fields
Discipline
Biologie santé
Composition du jury
Spécialité
Neurosciences
Brian LITT University of Pennsylvania
Lyle MULLERWestern University
Rapporteur
Rapporteur
École doctorale
Sciences de la vie et de la santé / ED 62
Mary DONAHUELinkoping University
Yuri ZILBERTERAix-Marseille Université
Examinatrice/présidente du jury
Directeur du thèse
Laboratoire/Partenaires de recherche
PhysioNet
Adam WILLIAMSON Aix-Marseille Université
Co-directeur du thèse

Affidavit
I, undersigned, Evgeniia Rusina, hereby declare that the work presented in this manuscript is my own work, carried out under the scientific direction of Dr Yuri Zilberter and Dr Adam Williamson, in accordance with the principles of honesty, integrity and responsibility inherent to the research mission. The research work and the writing of this manuscript have been carried out in compliance with both the French national charter for Research Integrity and the Aix-Marseille University charter on the fight against plagiarism.
This work has not been submitted previously either in this country or in another country in the same or in a similar version to any other examination body.
Place: Marseille…………… …….. .Date: 01/10/2020 ............................................................

Affidavit
Je soussigné, Evgeniia RUSINA, déclare par la présente que le travail présenté dans ce manuscrit est mon propre travail, réalisé sous la direction scientifique de Yuri ZILBERTER et Adam WILLIAMSON, dans le respect des principes d’honnêteté, d'intégrité et de responsabilité inhérents à la mission de recherche. Les travaux de recherche et la rédaction de ce manuscrit ont été réalisées dans le respect à la fois de la charte nationale de déontologie des métiers de la recherche et de la charte d’Aix-Marseille Université relative à la lutte contre le plagiat.
Ce travail n'a pas été précédemment soumis en France ou à l'étranger dans une version identique ou similaire à un organisme examinateur.
Fait à Marseille le 01/10/2020 ................................ .................................................................

Acknowledgements
I acknowledge the financial support from A*MIDEX, a chair of excellence in the University of Aix-Marseille and from European Research Council.
I am grateful to my both supervisors, for their patience and understanding. I would like to thank Dr Yuri Zilberter for his kindness and devotion to be my advisor. Thanks to Dr Adam Williamson, whose ideas and scientific inspiration made this all possible.
I recognize the contribution of my Neuroengineering team and I would like to sincerely thank all of them, without these people I would not have made it up to this point. Thank you, Emma Acerbo, for your infinite patience and all your help, you have become my right hand and my best friend in Marseille. Thanks to Boris Botzanowski and Florian Missey for your witty humor and bright young minds, you two certainly made my PhD one hell of a fun. Thanks to Sawssan Safieddine, who has become not only my colleague but also my travel partner; I will remember all our trips with warmth. I especially want to thank Dr Andrea Slezia and Dr Martin Baca, who helped me in so many matters I lost count months ago.
I am lucky to have been a part of the Physionet Team at Institut de Neurosciences des Systèmes. I would like to thank: Dr Christophe Bernard for his help on the epilepsy research, on writing and publishing of my papers, and on general PhD survival; Antoine Ghestem and Anton Ivanov for teaching me and providing lots of support; Dr Pascale Quilichini and Dr Monique Esclapez for being examples of strong women in science; Conchetta Taverna and Sonia Timourian for their excellent job and warm smiles. And of course thanks to Dr Viktor Jirsa, whose leadership has made INS what it is.I also want to thank Institut de Neuroscience de la Timone for collaboration. Many thanks to Dr Nicolas Wanaverbecq, Pascal Weber, Anne Duhoux and all the people of INT who helped to achieve my goals.
To conclude, I would like to acknowledge the help of all the people who have not been mentioned. And of course I would like to thank my loving family and my boyfriend Pierre, who supported me at all times, no matter how hard I struggled and how many times I wanted to give up. Thank you all for making this happen.

Abstract
In patients with focal drug-resistant epilepsy, electrical stimulation from intracranial electrodes is frequently used for the localization of seizure onset zones and related pathological networks. The ability of electrically stimulated tissue to generate beta and gamma range oscillations, called rapid-discharges, is a frequent indication of an epileptogenic zone. However, a limit of intracranial stimulation is the fixed physical location and number of implanted electrodes, leaving numerous clinically and functionally relevant brain regions unexplored. In this thesis, I present and describe an alternative technique relying exclusively on non-penetrating surface electrodes, namely an orientation-tunable form of temporally-interfering (TI) electric fields to target the CA3 of the mouse hippocampus which focally evokes seizure-like events (SLEs) having the characteristic frequencies of rapid-discharges, but without the necessity of the implanted electrodes. The orientation of the topical electrodes with respect to the orientation of the hippocampus is demonstrated to strongly control the threshold for evoking SLEs. Additionally, I describe the use of Pulse-width-modulation of square waves as an alternative to sine waves for TI stimulation. An orientation-dependent analysis of classic implanted electrodes to evoke SLEs in the hippocampus is subsequently utilized to support the results of the minimally-invasive temporally-interfering fields. The principles of orientation-tunable TI stimulation seen here can be generally applicable in a wide range of other excitable tissues and brain regions, overcoming several limitations of fixed electrodes which penetrate tissue and overcoming several limitations of other noninvasive stimulation methods in epilepsy, such as transcranial magnetic stimulation (TMS)._____________________________________________________________________________
Key words: Epilepsy, Electrical brain stimulation, Seizure localization

Résumé
Chez les patients atteints d’épilepsie focale pharmaco-résistante, la stimulation électrique par électrodes intracrâniennes est fréquemment utilisée lors de la localisation des zones épileptogènes et des réseaux pathologiques associés. Les tissus stimulés électriquement génèrent des oscillations de gamme bêta et gamma, appelées décharges rapides, et sont une indication fréquente d’une zone épileptogène. Cependant, il existe des limites à la stimulation intracrânienne comme l’emplacement fixe des électrodes et leur nombre implantées, laissant de nombreuses régions du cerveau cliniquement et fonctionnellement inexplorées. Dans cette thèse, je présente et décris une technique alternative qui repose exclusivement sur des électrodes de surface non pénétrante, permettant l’application de champs électriques à interférence temporelle (TI) mais aussi avec orientation réglable. Le but est de cibler le CA3 de l’hippocampe chez la souris qui évoque de manière focalisée des événements de type crise (SLE) ayant les fréquences caractéristiques des décharges rapides mais sans la nécessité des électrodes implantées. L’orientation des électrodes topiques par rapport à celle de l’hippocampe contrôle fortement le seuil d’évocation des SLE. En outre, je décris l’utilisation de la modulation de largeur d’impulsion des ondes carrées comme alternative aux ondes sinusoïdales pour la stimulation TI. Une analyse dépendante de l’orientation des électrodes classiques implantées pour évoquer les SLE dans l’hippocampe est ensuite utilisée pour étayer les résultats des champs à interférence temporelle mini-invasive. Les principes de la stimulation TI à orientation réglable que l’on voit ici peuvent être ainsi applicables à un large éventail d’autres tissus et régions cérébrales excitables. Ceci permettrait de surmonter les limitations des électrodes fixes qui pénètrent dans les tissus mais aussi celles d’autres méthodes de stimulation non invasives dans l’épilepsie, comme la stimulation magnétique transcrânienne (TMS)._____________________________________________________________________________
Mots clés: Epilepsie, Stimulation cérébrale électrique, Localisation des crises

Contents
Affidavit 2
Affidavit 3
Acknowledgements 4
Abstract 5
Résumé 6
Contents 7
List of Figures 10
List of Tables 12
1. Epilepsy: Temporal Lobe Epilepsy 13
1.1. Overview 13..................................................................................................................
1.1.1. Definition and epidemiology 13...........................................................................
1.1.2. Classification 14..................................................................................................
1.1.3. Etiology 14...........................................................................................................
1.1.3.1. Structural 14...............................................................................................
1.1.3.2. Genetic 15..................................................................................................
1.1.3.3. Infectious 16...............................................................................................
1.1.3.4. Metabolic 16...............................................................................................
1.1.3.5. Immune 17..................................................................................................
1.1.3.6. Unknown 19................................................................................................
1.1.4. Pathophysiology 19.............................................................................................
1.1.5. Signs and symptoms 20......................................................................................
1.1.6. Diagnosis and treatment 21................................................................................
2. Non-invasive seizure foci localization 23
2.1. Problematic in epilepsy 23...........................................................................................
2.2. Historical overview 23..................................................................................................
2.2.1. EEG-guided localization 23.................................................................................
2.2.1.1. Invasive VS non-invasive 23.......................................................................
2.2.1.2. Stereoelectroencephalography 24..............................................................
2.2.1.3. Electrocorticography 25.............................................................................
2.2.1.4. Hybrid EEG 27............................................................................................
2.2.2. Neuroimaging techniques 30...............................................................................

2.2.2.1. Computed tomography (CT) 30..................................................................
2.2.2.2. Magnetic resonance imaging (MRI) 30.......................................................
2.2.2.3. Positron emission tomography (PET) 32....................................................
2.2.2.4. Single-photon emission computed tomography (SPECT) 33.....................
2.2.2.5. Magnetoencephalography (MEG) 34..........................................................
2.3. Modern non-invasive brain stimulation approach 35...................................................
2.3.1. Transcranial magnetic stimulation 35..................................................................
2.3.1.1. Technical outline 35....................................................................................
2.3.1.2. Seizure detection 35...................................................................................
2.3.2. Transcranial electrical stimulation 37...................................................................
2.3.2.1. Technical outline 37....................................................................................
2.3.2.2. Seizure detection 37...................................................................................
2.3.3. TI stimulation: a potential diagnostic tool? 37.....................................................
3. Temporal Interference (TI) electrical stimulation 39
3.1. Basic concept 39..........................................................................................................
3.1.1. Neurophysiological basis 39................................................................................
3.1.2. Technical outline 40.............................................................................................
3.2. Research: Orientation-Tunable TI for Non-Invasive DBS in Epilepsy 41......................
3.2.1. Introduction 41....................................................................................................
3.2.2. Methodology 43...................................................................................................
3.2.2.1. Animals 43..................................................................................................
3.2.2.2. TI Electrical stimulation 43..........................................................................
3.2.2.3. Direct stimulation 44...................................................................................
3.2.2.4. Behavioral evaluation 44.............................................................................
3.2.2.5. Virtual simulation 44...................................................................................
3.2.2.6. Statistical analysis 45.................................................................................
3.2.3. Results 45............................................................................................................
3.2.4. Discussion 51......................................................................................................
ot-TI: Supplementary materials 53......................................................................................
4. Conclusion and future directions 55
4.1. Summary 55.................................................................................................................
4.2. Future directions 56......................................................................................................
4.2.1. Quadropole TI stimulation 56..............................................................................
4.2.2. Therapeutic application 56..................................................................................
4.2.3. Diagnostics 56.....................................................................................................

Annex 57
A Future directions: The Kainic Acid models of TLE 57
A.1. Introduction 57.......................................................................................................
A.2. Models of TLE: a brief overview 57........................................................................
A.3. The kainic acid model 59.......................................................................................
A.3.1. Kainic acid receptors 60................................................................................
A.3.2. Mechanism of action 60................................................................................
A.4. Administration routes 62........................................................................................
A.4.1. Systemic administration 62...........................................................................
A.4.2. Intraventricular injection 66...........................................................................
A.4.3. Intrahippocampal and intra-amygdaloid injections 67..................................
A.4.4. Suprahippocampal 70...................................................................................
A.4.5. Intranasal injection 71...................................................................................
A.5. Electrophysiology 73..............................................................................................
A.6. Age, sex and strain specificity 75..........................................................................
A.7. Concluding remarks 77..........................................................................................
Bibliography 78

List of Figures
Figure 1.1. Trends in epilepsy from 1990 to 2017. 13...............................................................
Figure 1.2. Updated epilepsy classification (ILAE, 2017). 14....................................................
Figure 1.3. Examples of epilepsies of structural etiology. 15....................................................
Figure 1.4. Examples of epilepsies of infectious etiology. 16....................................................
Figure 1.5. Papez circuit. 19......................................................................................................
Figure 1.6. Pathophysiological mechanisms of epileptogenesis. 20........................................
Figure 1.7. Characteristic EEG-pattern for TLE. 22...................................................................
Figure 2.1. Epilepsy surgery using sEEG, Sainte-Anne hospital, 1974. 24...............................
Figure 2.2. Invasive seizure foci localization techniques. 25.....................................................
Figure 2.3. Hybrid EEG: a combination between sEEG and ECoG. 27....................................
Figure 2.4. Spatio-temporal source localization approach (FINE). 28.......................................
Figure 2.5. Theta EEG source localization using LORETA. 29..................................................
Figure 2.6. CT brain scan: arteriovenous malformation leading to seizures. 30.......................
Figure 2.7. MRI brain scans: various findings in TLE. Functional MRI. 31................................
Figure 2.8. [18F]FDG-PET interictal brain scan in a patent with TLE. 32..................................
Figure 2.9. SPECT imaging: findings in parieto-occipital epilepsy. SISCOM. 33......................
Figure 2.10. MEG: applications in epilepsy. 34.........................................................................
Figure 2.11. DBS vs TMS vs tES. 35.........................................................................................
Figure 2.12. Seizure foci localization using navigated TMS. 36................................................
Figure 3.1. Frequency-dependent properties of neurons. 39....................................................
Figure 3.2. TI stimulation design in a mouse brain and in a human patient. 40........................
Figure 3.3. Orientation-tunable Temporally Interfering Electric Fields (ot-TI). 42......................
Figure 3.4. Evoked SLEs in the hippocampus of freely-moving mice. 46.................................
Figure 3.5. AP: non-preferential orientation, electric field envelope versus anatomy. 47.........
Figure 3.6. Orientation-controlled stimulation of SLEs in the hippocampus with classic
implanted stimulators. 48..........................................................................................................
Figure 3.7. LFP recording of freely-moving mice after a 600μA stimulation using the
preferential ML orientation. 49..................................................................................................
Figure 3.8. Analysis of SLE characteristics (amplitude, duration, severity) in the
hippocampus. 50......................................................................................................................

Supplementary Figure S1. No induced SLE in the cortex. 53...................................................
Supplementary Figure S2. Square-wave TI. 53.........................................................................
Supplementary Figure S3. Stimulation Protocol for TI and implantable. 54.............................
Supplementary Figure S4. Field lines of TI. 54..........................................................................
Supplementary Figure S5. How NOT to make a TI field. 54.....................................................
Figure A.1. Mechanism of KA-induced neuronal damage. 61..................................................
Figure A.2. Comparison of the KA administration routes 63.....................................................
Figure A.4. The various patterns of EEG-activity during KA-induced chronic epilepsy. 73......
Figure A.5. Seizure progression over time. 74..........................................................................

List of Tables
Table 1.1. Genetic forms of epilepsy with known mutations. 15...............................................
Table 1.2. Metabolic disorders associated with epilepsy. 17....................................................
Table 1.3. Examples of autoimmune-mediated epilepsies. 18..................................................
Table 1.4. Classification of seizures by ILAE, 2017. 21.............................................................
Table 2.1. Comparison of invasive seizure foci localization techniques. 26..............................
Table 2.2. Comparison of existing EBS types and the novel method of TI stimulation. 38......
Table A.1. Systemic administration of KA in rodents. 64..........................................................
Table A.2. Intracerebroventricular administration of KA in rats. 66...........................................
Table A.3. Intrahippocampal administration of KA in rodents. 67.............................................
Table A.4. Intra-amygdaloid administration of KA in rodents. 68..............................................
Table A.5. Suprahippocampal administration of KA in mice. 71...............................................
Table A.6. Intranasal administration of KA in mice. 72..............................................................
Chapter 1
1. Epilepsy: Temporal Lobe Epilepsy
1.1. Overview
1.1.1. Definition and epidemiology
Epilepsy is a serious neurological condition affecting people of all ages and in some cases causing disability. It is characterised by recurrent seizures which should fit in any of the following criteria, defined by International League Against Epilepsy (ILAE): a) at least 2 unprovoked (or reflex) seizures occurring more than 24h apart; b) 1 unprovoked (or reflex) seizure and a probability of further seizures similar to the general recurrence risk (≥ 60%) after 2 unprovoked seizures occurring over the next 10 years; c) diagnosis of an epilepsy syndrome . An estimated amount of people affected by the disease is 1
70 million people worldwide or between 2.6 and 6 million individuals in Europe , . In 2 3
France, more than 600 thousands people suffer from epilepsy, almost half of them are younger than 20 years old . In developed countries, 40-60 new cases per 100000 4
people are registered annually, while in low/middle-income countries this number is more than twice higher (48.9 (95% CI 39.0–61.1) vs 139.0 (95% CI 69.4–278.2)) . The 5
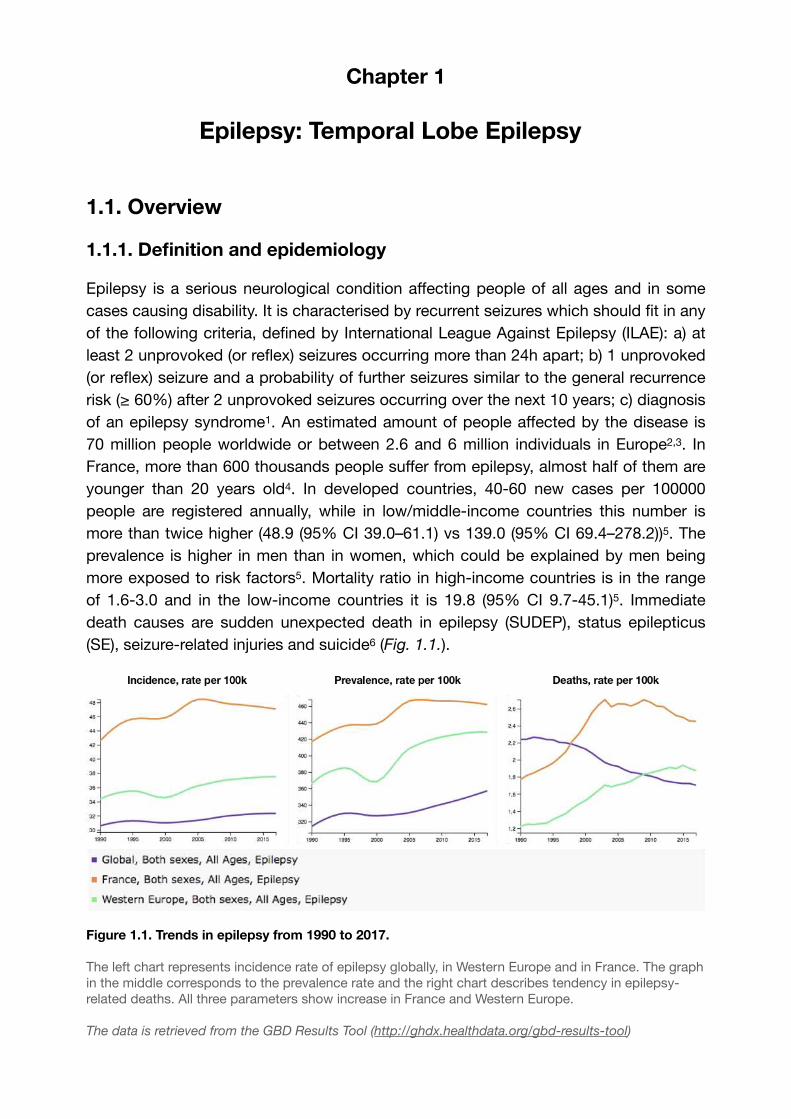
prevalence is higher in men than in women, which could be explained by men being more exposed to risk factors5. Mortality ratio in high-income countries is in the range of 1.6-3.0 and in the low-income countries it is 19.8 (95% CI 9.7-45.1)5. Immediate death causes are sudden unexpected death in epilepsy (SUDEP), status epilepticus (SE), seizure-related injuries and suicide (Fig. 1.1.).6
Figure 1.1. Trends in epilepsy from 1990 to 2017.
The left chart represents incidence rate of epilepsy globally, in Western Europe and in France. The graph in the middle corresponds to the prevalence rate and the right chart describes tendency in epilepsy-related deaths. All three parameters show increase in France and Western Europe.
The data is retrieved from the GBD Results Tool (http://ghdx.healthdata.org/gbd-results-tool)

1.1.2. Classification
The classification of epilepsies is complex and is based on several key points, using which one can establish a diagnosis. According to ILAE (2017), there are 4 general epilepsy types: focal, generalized, combined generalized & focal and unknown (Fig. 7
1.2). This should not be confused with seizure types (focal, generalized and unknown) which also represent a major diagnosis framework. All types could be further subdivided by different etiology (structural, genetic, infectious, metabolic, immune, unknown). And finally, any comorbidities should also be considered. Temporal Lobe Epilepsy (TLE), according to this classification, would most likely fit as focal epilepsy with structural etiology, if hippocampal sclerosis (HS) is confirmed. The last level of diagnosis is establishing an epilepsy syndrome, which, however, is not always possible. An epilepsy syndrome is a cluster of clinical features which tend to occur together. For instance, a seizure type, an EEG pattern, age of onset, and distinct comorbidities, taken altogether, can compose a known epilepsy syndrome. Thus, sometimes, TLE is regarded as a self-limited focal epilepsy syndrome.
Figure 1.2. Updated epilepsy classification (ILAE, 2017).
The framework for classification of epilepsy includes three levels: seizure type, epilepsy type and the conclusive diagnosis of epilepsy syndrome. All the epilepsy types vary in etiology which is listed on the right and might include any comorbidities (left), which are also important for correct diagnostics. * Denotes the onset of seizures
1.1.3. Etiology
1.1.3.1. Structural
Etiology of epilepsy is declared structural when a visible brain lesion is found during neuroimaging and is assumed to cause seizures7 (Fig. 1.3). Usually, multiple etiologic factors are combined, and thus structural etiology can be either genetic or acquired due to various reasons, such as infections, stroke, trauma (Fig. 1.3.B) etc. One of the most typical examples of structural etiology is hippocampal sclerosis in TLE (Fig. 8
1.3.A). Epilepsies of such etiology are the most likely candidates for a surgical intervention, as in most cases tissue removal is more efficient than a drug-therapy. In TLE, the possible options are hippocampectomy or temporal lobectomy.
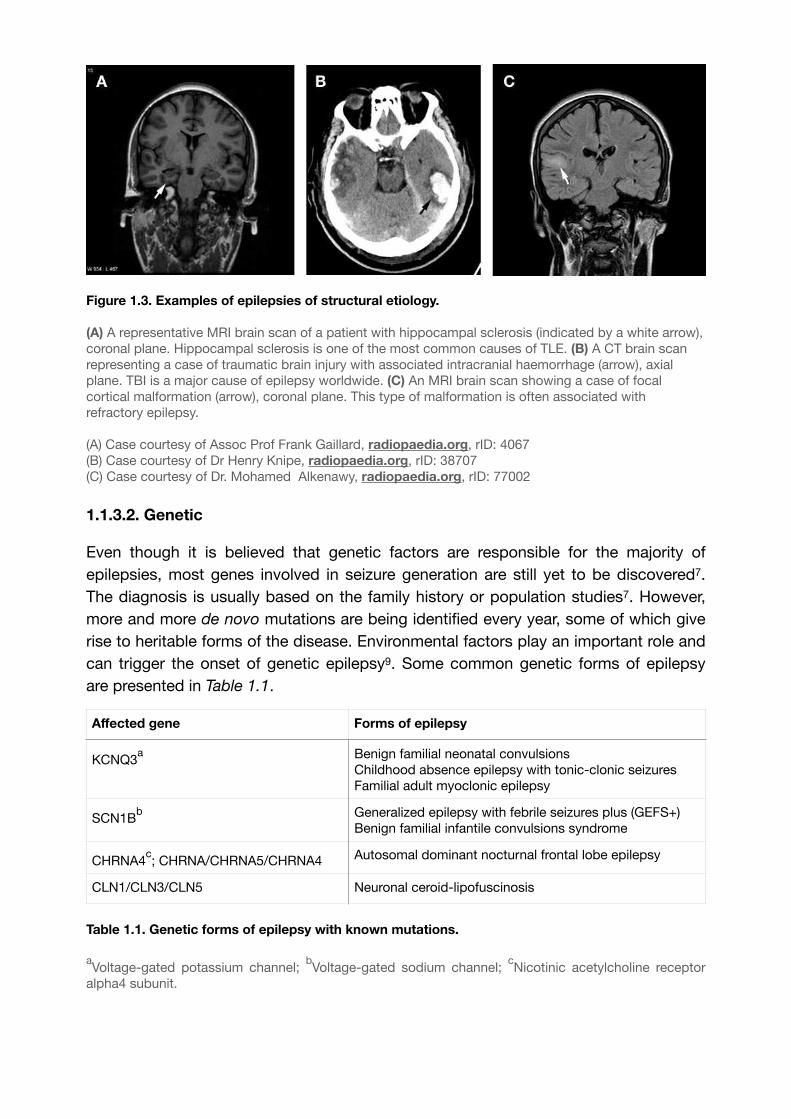
Figure 1.3. Examples of epilepsies of structural etiology.
(A) A representative MRI brain scan of a patient with hippocampal sclerosis (indicated by a white arrow), coronal plane. Hippocampal sclerosis is one of the most common causes of TLE. (B) A CT brain scan representing a case of traumatic brain injury with associated intracranial haemorrhage (arrow), axial plane. TBI is a major cause of epilepsy worldwide. (C) An MRI brain scan showing a case of focal cortical malformation (arrow), coronal plane. This type of malformation is often associated with refractory epilepsy.
(A) Case courtesy of Assoc Prof Frank Gaillard, radiopaedia.org, rID: 4067(B) Case courtesy of Dr Henry Knipe, radiopaedia.org, rID: 38707(C) Case courtesy of Dr. Mohamed Alkenawy, radiopaedia.org, rID: 77002
1.1.3.2. Genetic Even though it is believed that genetic factors are responsible for the majority of epilepsies, most genes involved in seizure generation are still yet to be discovered7. The diagnosis is usually based on the family history or population studies7. However, more and more de novo mutations are being identified every year, some of which give rise to heritable forms of the disease. Environmental factors play an important role and can trigger the onset of genetic epilepsy . Some common genetic forms of epilepsy 9
are presented in Table 1.1.
Table 1.1. Genetic forms of epilepsy with known mutations.
aVoltage-gated potassium channel;
bVoltage-gated sodium channel;
cNicotinic acetylcholine receptor
alpha4 subunit.
Affected gene Forms of epilepsy
KCNQ3a Benign familial neonatal convulsions
Childhood absence epilepsy with tonic-clonic seizuresFamilial adult myoclonic epilepsy
SCN1Bb Generalized epilepsy with febrile seizures plus (GEFS+)
Benign familial infantile convulsions syndrome
CHRNA4c; CHRNA/CHRNA5/CHRNA4 Autosomal dominant nocturnal frontal lobe epilepsy
CLN1/CLN3/CLN5 Neuronal ceroid-lipofuscinosis
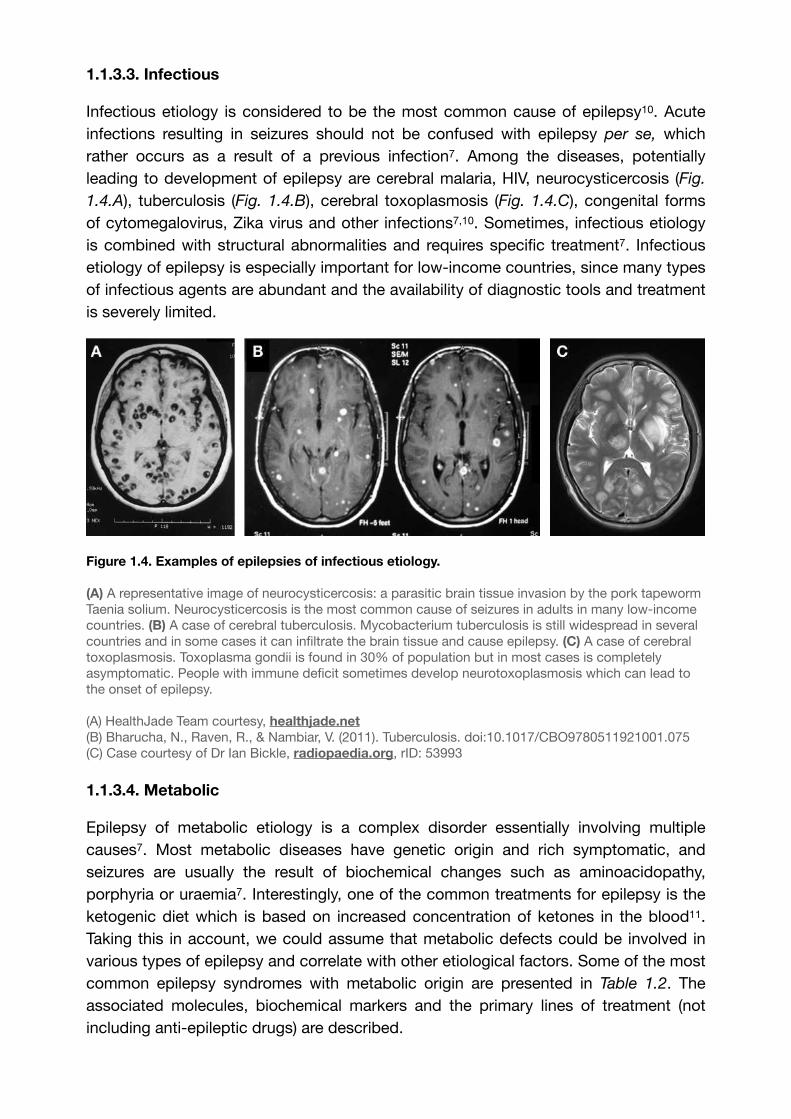
1.1.3.3. Infectious
Infectious etiology is considered to be the most common cause of epilepsy . Acute 10
infections resulting in seizures should not be confused with epilepsy per se, which rather occurs as a result of a previous infection7. Among the diseases, potentially leading to development of epilepsy are cerebral malaria, HIV, neurocysticercosis (Fig.
1.4.A), tuberculosis (Fig. 1.4.B), cerebral toxoplasmosis (Fig. 1.4.C), congenital forms of cytomegalovirus, Zika virus and other infections7,10. Sometimes, infectious etiology is combined with structural abnormalities and requires specific treatment7. Infectious etiology of epilepsy is especially important for low-income countries, since many types of infectious agents are abundant and the availability of diagnostic tools and treatment is severely limited.
Figure 1.4. Examples of epilepsies of infectious etiology.
(A) A representative image of neurocysticercosis: a parasitic brain tissue invasion by the pork tapeworm Taenia solium. Neurocysticercosis is the most common cause of seizures in adults in many low-income countries. (B) A case of cerebral tuberculosis. Mycobacterium tuberculosis is still widespread in several countries and in some cases it can infiltrate the brain tissue and cause epilepsy. (C) A case of cerebral toxoplasmosis. Toxoplasma gondii is found in 30% of population but in most cases is completely asymptomatic. People with immune deficit sometimes develop neurotoxoplasmosis which can lead to the onset of epilepsy.
(A) HealthJade Team courtesy, healthjade.net(B) Bharucha, N., Raven, R., & Nambiar, V. (2011). Tuberculosis. doi:10.1017/CBO9780511921001.075(C) Case courtesy of Dr Ian Bickle, radiopaedia.org, rID: 53993
1.1.3.4. Metabolic
Epilepsy of metabolic etiology is a complex disorder essentially involving multiple causes7. Most metabolic diseases have genetic origin and rich symptomatic, and seizures are usually the result of biochemical changes such as aminoacidopathy, porphyria or uraemia7. Interestingly, one of the common treatments for epilepsy is the ketogenic diet which is based on increased concentration of ketones in the blood . 11
Taking this in account, we could assume that metabolic defects could be involved in various types of epilepsy and correlate with other etiological factors. Some of the most common epilepsy syndromes with metabolic origin are presented in Table 1.2. The associated molecules, biochemical markers and the primary lines of treatment (not including anti-epileptic drugs) are described.
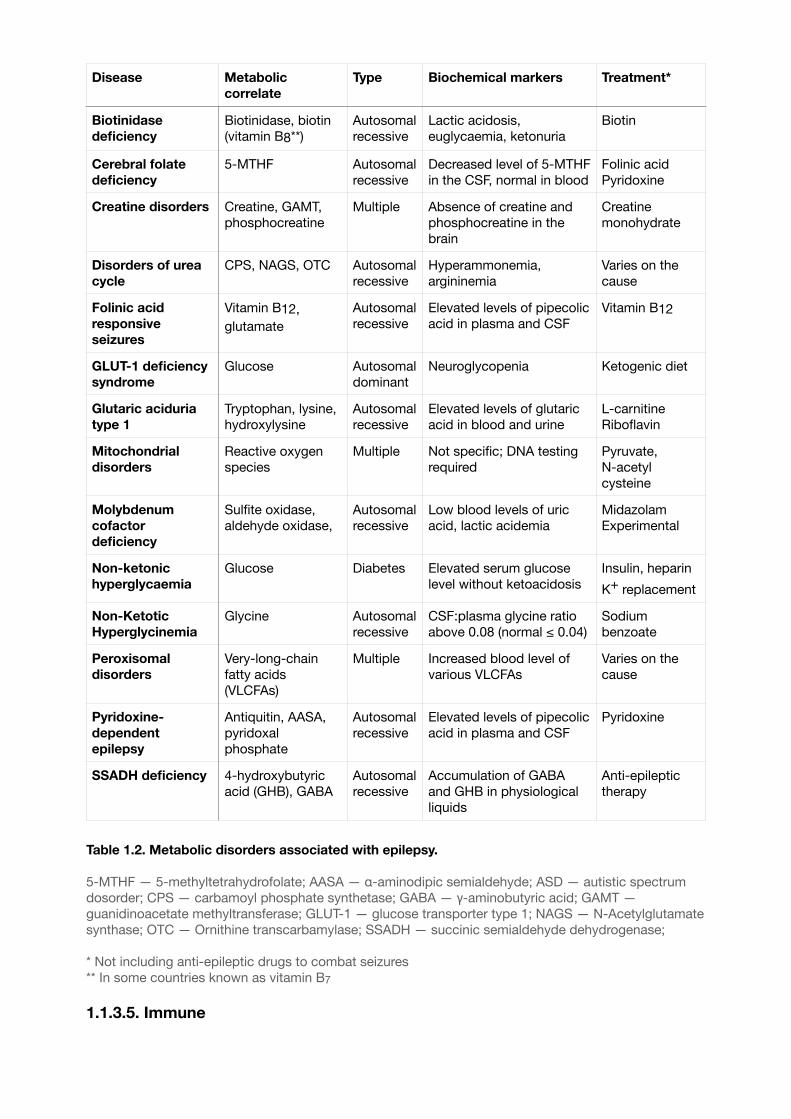
Table 1.2. Metabolic disorders associated with epilepsy.
5-MTHF — 5-methyltetrahydrofolate; AASA — α-aminodipic semialdehyde; ASD — autistic spectrum dosorder; CPS — carbamoyl phosphate synthetase; GABA — γ-aminobutyric acid; GAMT — guanidinoacetate methyltransferase; GLUT-1 — glucose transporter type 1; NAGS — N-Acetylglutamate synthase; OTC — Ornithine transcarbamylase; SSADH — succinic semialdehyde dehydrogenase;
* Not including anti-epileptic drugs to combat seizures** In some countries known as vitamin B7
1.1.3.5. Immune
Disease Metabolic correlate
Type Biochemical markers Treatment*
Biotinidase deficiency
Biotinidase, biotin (vitamin B8**)
Autosomal recessive
Lactic acidosis, euglycaemia, ketonuria
Biotin
Cerebral folate deficiency
5-MTHF Autosomal recessive
Decreased level of 5-MTHF in the CSF, normal in blood
Folinic acidPyridoxine
Creatine disorders Creatine, GAMT, phosphocreatine
Multiple Absence of creatine and phosphocreatine in the brain
Creatine monohydrate
Disorders of urea cycle
CPS, NAGS, OTC Autosomal recessive
Hyperammonemia, argininemia
Varies on the cause
Folinic acid responsive seizures
Vitamin B12, glutamate
Autosomal recessive
Elevated levels of pipecolic acid in plasma and CSF
Vitamin B12
GLUT-1 deficiency syndrome
Glucose Autosomal dominant
Neuroglycopenia Ketogenic diet
Glutaric aciduria type 1
Tryptophan, lysine, hydroxylysine
Autosomal recessive
Elevated levels of glutaric acid in blood and urine
L-carnitineRiboflavin
Mitochondrial disorders
Reactive oxygen species
Multiple Not specific; DNA testing required
Pyruvate, N-acetyl cysteine
Molybdenum cofactor deficiency
Sulfite oxidase, aldehyde oxidase,
Autosomal recessive
Low blood levels of uric acid, lactic acidemia
MidazolamExperimental
Non-ketonic hyperglycaemia
Glucose Diabetes Elevated serum glucose level without ketoacidosis
Insulin, heparin
K+ replacement
Non-Ketotic Hyperglycinemia
Glycine Autosomal recessive
CSF:plasma glycine ratio above 0.08 (normal ≤ 0.04)
Sodium benzoate
Peroxisomal disorders
Very-long-chain fatty acids (VLCFAs)
Multiple Increased blood level of various VLCFAs
Varies on the cause
Pyridoxine-dependent epilepsy
Antiquitin, AASA, pyridoxal phosphate
Autosomal recessive
Elevated levels of pipecolic acid in plasma and CSF
Pyridoxine
SSADH deficiency 4-hydroxybutyric acid (GHB), GABA
Autosomal recessive
Accumulation of GABA and GHB in physiological liquids
Anti-epileptic therapy
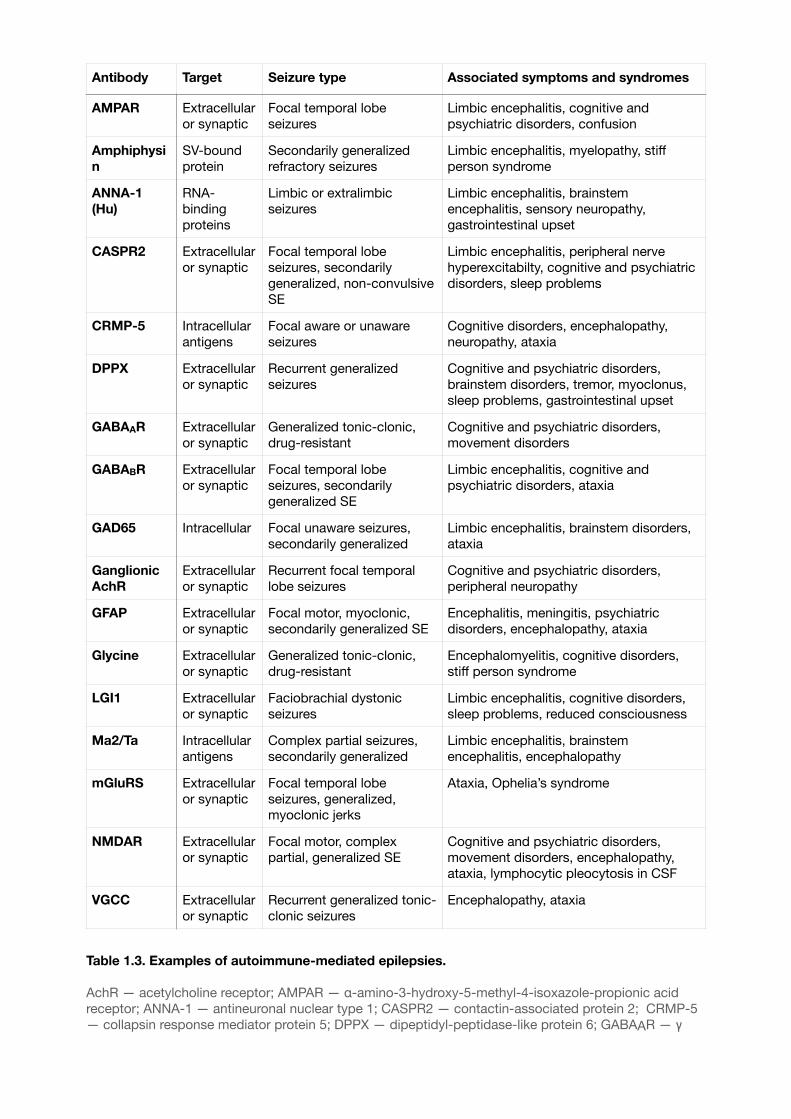
Table 1.3. Examples of autoimmune-mediated epilepsies.
AchR — acetylcholine receptor; AMPAR — α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid receptor; ANNA-1 — antineuronal nuclear type 1; CASPR2 — contactin-associated protein 2; CRMP-5 — collapsin response mediator protein 5; DPPX — dipeptidyl-peptidase-like protein 6; GABAAR — γ
Antibody Target Seizure type Associated symptoms and syndromes
AMPAR Extracellular or synaptic
Focal temporal lobe seizures
Limbic encephalitis, cognitive and psychiatric disorders, confusion
Amphiphysin
SV-bound protein
Secondarily generalized refractory seizures
Limbic encephalitis, myelopathy, stiff person syndrome
ANNA-1 (Hu)
RNA-binding proteins
Limbic or extralimbic seizures
Limbic encephalitis, brainstem encephalitis, sensory neuropathy, gastrointestinal upset
CASPR2 Extracellular or synaptic
Focal temporal lobe seizures, secondarily generalized, non-convulsive SE
Limbic encephalitis, peripheral nerve hyperexcitabilty, cognitive and psychiatric disorders, sleep problems
CRMP-5 Intracellular antigens
Focal aware or unaware seizures
Cognitive disorders, encephalopathy, neuropathy, ataxia
DPPX Extracellular or synaptic
Recurrent generalized seizures
Cognitive and psychiatric disorders, brainstem disorders, tremor, myoclonus, sleep problems, gastrointestinal upset
GABAAR Extracellular or synaptic
Generalized tonic-clonic, drug-resistant
Cognitive and psychiatric disorders, movement disorders
GABABR Extracellular or synaptic
Focal temporal lobe seizures, secondarily generalized SE
Limbic encephalitis, cognitive and psychiatric disorders, ataxia
GAD65 Intracellular Focal unaware seizures, secondarily generalized
Limbic encephalitis, brainstem disorders, ataxia
Ganglionic AchR
Extracellular or synaptic
Recurrent focal temporal lobe seizures
Cognitive and psychiatric disorders, peripheral neuropathy
GFAP Extracellular or synaptic
Focal motor, myoclonic, secondarily generalized SE
Encephalitis, meningitis, psychiatric disorders, encephalopathy, ataxia
Glycine Extracellular or synaptic
Generalized tonic-clonic, drug-resistant
Encephalomyelitis, cognitive disorders, stiff person syndrome
LGI1 Extracellular or synaptic
Faciobrachial dystonic seizures
Limbic encephalitis, cognitive disorders, sleep problems, reduced consciousness
Ma2/Ta Intracellular antigens
Complex partial seizures, secondarily generalized
Limbic encephalitis, brainstem encephalitis, encephalopathy
mGluRS Extracellular or synaptic
Focal temporal lobe seizures, generalized, myoclonic jerks
Ataxia, Ophelia’s syndrome
NMDAR Extracellular or synaptic
Focal motor, complex partial, generalized SE
Cognitive and psychiatric disorders, movement disorders, encephalopathy, ataxia, lymphocytic pleocytosis in CSF
VGCC Extracellular or synaptic
Recurrent generalized tonic-clonic seizures
Encephalopathy, ataxia
aminobutyric acid A receptor; GABABR — γ aminobutyric acid B receptor; GAD65 — glutamic acid decarboxylase 65; GFAP — glial fibrillary acid protein; LGI1 — leucine-rich glioma inactivated 1; mGluR5 — metabotropic glutamate receptor 5; NMDAR — N-methyl-D-aspartate receptor; SE — status epilepticus; SV — synaptic vesicle; VGCC — voltage-gated calcium channel.
While immune disorders are becoming more widespread, immune epilepsies are being recognized as a distinct category7 (Table 1.3). The mechanism behind is believed to be autoimmune-mediated inflammation of CNS7. Some examples of immune epilepsies are anti-NMDA encephalitis, anti-LGI1 encephalitis, celiac disease, hypoglycaemic seizures in patients with diabetes . 12
1.1.3.6. Unknown
Epilepsy of unknown etiology represents about 30% of all epilepsies in adults7. There could be multiple factors involved in the development of the disease, including yet undiscovered genetic mutations, autoimmune attacks etc. In some cases epilepsy is declared of unknown etiology due to a lack of diagnostic tools.
1.1.4. Pathophysiology
Mechanisms underlying epilepsy are still being researched and are mostly debatable. Different etiological forms of the disease vary in their pathophysiological traits and, to avoid unnecessary speculation, in this section we will focus on the pathophysiology of TLE. The common mechanism of all the epileptic syndromes is increased excitability of neurons ; in case of TLE, this attribute takes a specific form due to a unique position 13
of the hippocampus and its connections to other regions of the brain, particularly, the limbic system . Some researches claim that hyper-excitability in the hippocampus 14
may trigger a chain reaction, in which the signal travels through limbic structures, gets amplified and returns to the hippocampus, potentially inducing generalized seizures14.
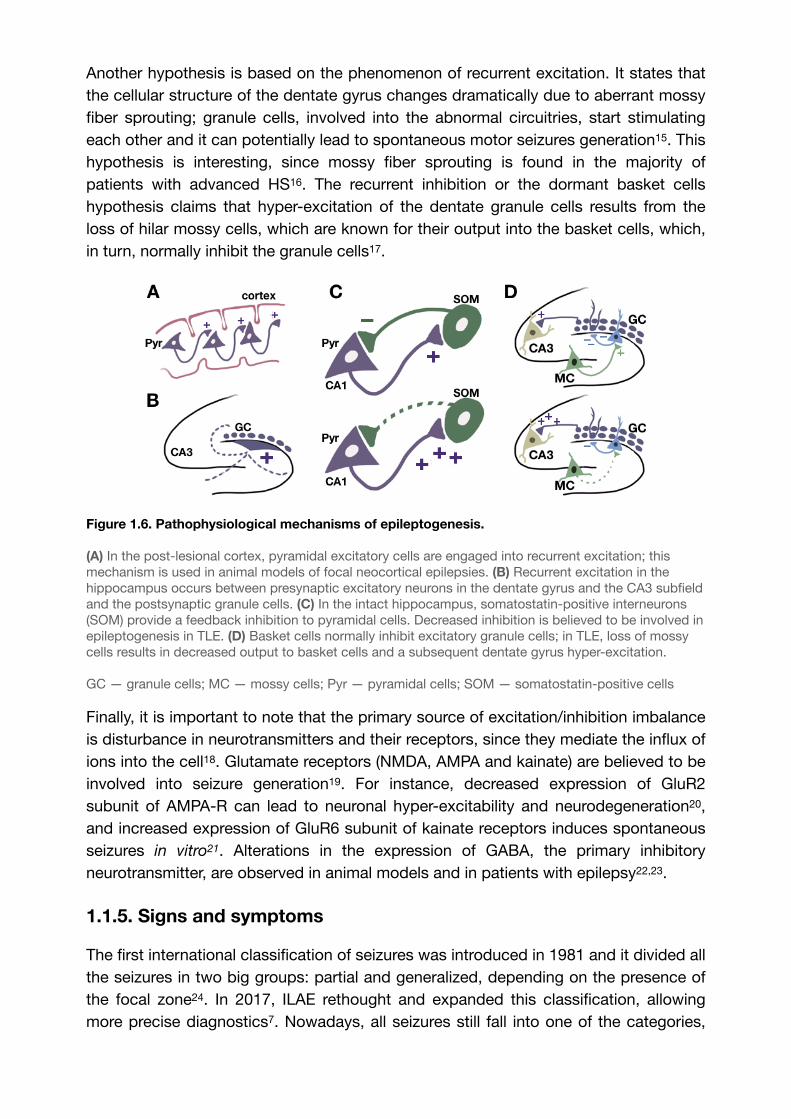
Figure 1.5. Papez circuit.
The so-called Papez circuit represents neuronal connections within the limbic system and with the neocortex. Many researchers believe that the mechanism of TLE lies within the hippocampal anatomy.From: Bear, M. F., Connors, B. W., & Paradiso, M. A. (2016). Neuroscience: Exploring the brain
Another hypothesis is based on the phenomenon of recurrent excitation. It states that the cellular structure of the dentate gyrus changes dramatically due to aberrant mossy fiber sprouting; granule cells, involved into the abnormal circuitries, start stimulating each other and it can potentially lead to spontaneous motor seizures generation . This 15
hypothesis is interesting, since mossy fiber sprouting is found in the majority of patients with advanced HS . The recurrent inhibition or the dormant basket cells 16
hypothesis claims that hyper-excitation of the dentate granule cells results from the loss of hilar mossy cells, which are known for their output into the basket cells, which, in turn, normally inhibit the granule cells .17
Figure 1.6. Pathophysiological mechanisms of epileptogenesis.
(A) In the post-lesional cortex, pyramidal excitatory cells are engaged into recurrent excitation; this mechanism is used in animal models of focal neocortical epilepsies. (B) Recurrent excitation in the hippocampus occurs between presynaptic excitatory neurons in the dentate gyrus and the CA3 subfield and the postsynaptic granule cells. (C) In the intact hippocampus, somatostatin-positive interneurons (SOM) provide a feedback inhibition to pyramidal cells. Decreased inhibition is believed to be involved in epileptogenesis in TLE. (D) Basket cells normally inhibit excitatory granule cells; in TLE, loss of mossy cells results in decreased output to basket cells and a subsequent dentate gyrus hyper-excitation.
GC — granule cells; MC — mossy cells; Pyr — pyramidal cells; SOM — somatostatin-positive cells
Finally, it is important to note that the primary source of excitation/inhibition imbalance is disturbance in neurotransmitters and their receptors, since they mediate the influx of ions into the cell . Glutamate receptors (NMDA, AMPA and kainate) are believed to be 18
involved into seizure generation . For instance, decreased expression of GluR2 19
subunit of AMPA-R can lead to neuronal hyper-excitability and neurodegeneration , 20
and increased expression of GluR6 subunit of kainate receptors induces spontaneous seizures in vitro . Alterations in the expression of GABA, the primary inhibitory 21
neurotransmitter, are observed in animal models and in patients with epilepsy , .22 23
1.1.5. Signs and symptoms
The first international classification of seizures was introduced in 1981 and it divided all the seizures in two big groups: partial and generalized, depending on the presence of the focal zone . In 2017, ILAE rethought and expanded this classification, allowing 24
more precise diagnostics7. Nowadays, all seizures still fall into one of the categories,

based on the onset: focal, generalized or unknown. Within each group, they divide by motor or non-motor onset. In case of the focal onset, they are also distinguished by degree of awareness. Full classification is presented in Table 1.4.
Table 1.4. Classification of seizures by ILAE, 2017.
Seizures differ in their onset, whether it is focal or generalized, and within each group, motor or non-motor. Focal-onset seizures also vary in the degree of awareness, which could be normal or impaired. Some seizures fall into an unclassified category, due to insufficient data or failure to determine the type.
TLE is especially interesting in its symptomatic, since its focus is localized in the hippocampus. Patients with this type of epilepsy often experience a so-called "aura", which is essentially a sensation or an intense emotion, unique for each individual . 25
Some report strong fear, for others the prevalence feeling is delight and happiness25. A classic TLE symptom is déjà vu, the feeling of a current experience already being experienced in the past . The reverse feeling, jamais vu, where familiar experience 26
perceives as something new, is also common . Sometimes, seizures can appear in 27
the form of memory flow, tactile sensations on the skin, visual, auditory, olfactory and other types of hallucinations25.
1.1.6. Diagnosis and treatment
Establishing a diagnose of an epileptic syndrome is based on three key components: visual observation, EEG-recording, unraveling the cause7. Typically, an individual with suspected epilepsy reports seizures observed by a third party. Subsequently, findings on the EEG confirm the presence of seizures. Even though epileptic abnormalities are generally accepted in the form of spikes and sharp waves, or spike-wave complexes, for each type of epilepsy there is a certain pattern of interictal EEG-activity . In TLE, 28
the typical location of interictal spikes is anterior/mid-temporal region, associated with the hippocampus . Characteristic 5-9 Hz ictal discharges accompanying seizures can 29
Focal onset Generalized onset Unknown onset
Aware Impaired awareness
Motor onset AutomatismsAtonicClonicEpileptic spasmsHyperkineticMyoclonicTonic
Tonic-clonicClonicTonicMyoclonicMyoclonic-tonic-clonicMyoclonic-atonicAtonicEpileptic spasms
Tonic-clonicEpileptic spasms
Non-motor onset AutonomicBehavior arrestCognitiveEmotionalSensory
Absence: Typical Atypical Myoclonic Eyelid myoclonia
Behavior arrest
Focal to bilateral tonic-clonic Unclassified

be also observed , . To increase the probability of interictal spikes detection, several 30 31
options are proposed: sleep EEG , photostimulation , hyperventilation , placement 32 33 34
of additional electrodes . Once the presence of seizures on the EEG is confirmed, the 35
neuroimaging is performed, to detect possible gross organic pathology. Combining all the results, the diagnosis of a certain epileptic syndrome is made.
Figure 1.7. Characteristic EEG-pattern for TLE.
(A) An example of interictal focal temporal discharges in a patient with TLE. (B) Hippocampal sclerosis: Ictal rhythmic θ discharge in a patient with TLE.
Smith, S. J. M. (2005). http://doi.org/10.1136/jnnp.2005.069245
There are 2 major lines of treatment for epilepsy, depending on its cause . Standard 36
medication therapy involves use of anticonvulsants, where a dose is carefully chosen for each individual . In many cases, establishing right type of drug is challenging and 37
requires several changes of various anticonvulsant types, until the proper medication is found37. However, for some patients conservative therapy does not yield a satisfying result; these individuals are usually subjected to surgical intervention . TLE is 38
especially known for its high resistance for drug therapy and, considering the presence of organic damage in the hippocampus, the only available treatment for such patients is surgical removal of the epileptic focus . Over the past few decades, deep brain 39
stimulation (DBS) attempts have been made in order to efficiently suppress seizures . 40
Several targets have been proposed: cerebellum , STN , SN , caudate nucleus , 41 42 43 44
anterior thalamus , centromedian nucleus , hippocampus . The results are highly 45 46 47
variable but at some extent, successful. Some researches made attempts to directly stimulate the epileptic focus in TLE and demonstrated significant decrease in seizure frequency47, In any case, DBS could be an interesting alternative to a surgical 48
treatment for patients with refractory epilepsy.

Chapter 2
2. Non-invasive seizure foci localization
2.1. Problematic in epilepsy
As it has already been mentioned before, many focal epilepsies are treatment-resistant and require surgical intervention38,39. In order to achieve the best result while removing the epileptic focus, a preoperative assessment to determine the exact epileptogenic zone location is needed. It became a major challenge for neurosurgeons, as several problems emerged. First, focal epilepsies are not always associated with structural damage, which is visible during neuroimaging7; second, to detect epileptic activity via EEG, long-lasting recording sessions are needed, and sometimes, extra techniques are required to provoke an electrographic seizure. And even visible interictal spikes cannot always spot the exact location of the epileptogenic zone and in these cases, invasive recording methods are often used. Such approach poses obvious risks of infections, haemorrhage, and other surgery-related complications. Recently, some non-invasive techniques of preoperative assessment and seizure foci detection have been proposed, but they also have their drawbacks, such as inability to reach deep brain structures, high maintenance cost and low availability. A safe, fast, non-invasive method of seizure foci localization is still a mere concept, but the technique presented in this thesis, showed interesting results and could possibly become an alternative to existing types of preoperative epileptogenic zone detection approaches.
2.2. Historical overview
2.2.1. EEG-guided localization
2.2.1.1. Invasive VS non-invasive
The most obvious way to localize a seizure focus preoperatively is by performing a simple EEG-recording. As different epilepsy types show characteristic EEG-features, sometimes it is safe to assume that the epileptogenic zone lies within the area of observation, e.g. temporal lobe spikes in case of TLE most likely would indicate a lesion in the ipsilateral hippocampus. While it was clear that such localization lacks any spatial and temporal precision, the first attempts to use EEG recordings to localize an epileptic focus preoperatively were invasive depth EEG probes, and they appeared as early as in late 1960s, when the concept of stereoelectroencephalography (sEEG) was introduced and performed (Fig. 2.1). Such method allowed a sufficient degree of 49
spacial and temporal precision and a relatively accurate estimation of epileptic activity, its origin and propagation. However, the technique was still invasive, so that further research in safer methods was needed. Next step was to place the electrodes on the brain surface bypassing the bone layer and the dura mater.

Preoperative seizure foci localization using subdural EEG recording was attempted in 1980s, when the first studies and reviews had been published , . Subdural recording 50 51
showed a relative success in foci detection and became an alternative to depth probes implanted chronically. While between 64 and 86 percent of patients were satisfied with the result of the treatment, surgeries performed without any additional visual cues, still posed risks of neurological deficits, due to extensive brain tissue removal or a failed surgery with a need of re-operation , . 52 53
Figure 2.1. Epilepsy surgery using sEEG, Sainte-Anne hospital, 1974.
One of the first attempts to localize an epileptogenic zone is documented on the photo. Alain Bonis (left) and Jean Bancaud (right) are performing "anatomo-electro-clinical correlations" using sEEG recording.
Subsequently, non-invasive EEG recording has been studied in order to perform a preoperative epileptogenic zone assessment. In case of TLE and strictly temporally localized seizures, this method was relatively efficient with 82-94% ratio of success . 54
Another review demonstrates that in comparison with depth EEG recording, which was 100% accurate in seizure localization, the scalp EEG could only identify 44% of ictal seizures and 31% of interictal seizures and in general had a false error localization rate of 10% . Such tremendous difference highlighted an existing problem in epileptogenic 55
zone localization and forced researchers all over the world to look for alternatives.
2.2.1.2. Stereoelectroencephalography
The method of stereoelectroencephalography or sEEG is widely used nowadays and is considered minimally invasive . It was designed and introduced in 1969 by French 56
neurosurgeons Jean Talairach and Jean Bancaud. They referred to the novel technique as "functional stereotactic exploration in epilepsy" and described the use of sEEG in the surgery, having performed a large set of experiments on patients with epilepsy49.

The method is based on multiple electrodes implantation in a certain area of the brain, in order to define and delineate the epileptogenic area (Fig. 2.2.A). It is used if the data acquired via non-invasive methods is insufficient, e.g. in case of discrepancy between the EEG-pattern and the neuroimaging results (for example: frontal lobe seizures in presence of HS, or confirmed diagnosis of temporal lobe epilepsy in absence of HS). The primary advantage of sEEG compared to other seizure foci localization techniques is its ability to target deep brain structures. With a high density of electrodes in the area, it is also possible to perform a 3D assessment of the epileptogenic zone by interpolation, which is especially helpful in a decision of surgical resection . 57
Figure 2.2. Invasive seizure foci localization techniques.
(A) Schematic representation of stereoelectroencephalography (sEEG) setup. Multiple 0.08mm-diameter recording electrodes are implanted into the brain tissue through little holes in the skull; such technique allows to localize seizure foci with relative precision and accurately define the borders of epileptogenic area. The procedure is minimally invasive, generally well tolerated and associated risks are extremely low. (B) Electrocorticography (ECoG) is another widely used method of seizure foci localization. A grid or a strip with up to 256 electrodes is placed on the exposed cortical surface through a wide opening in the skull - a cranial window. This manipulation is highly efficient for preoperative assessment of cortical epilepsies and for mapping of eloquent cortex, allowing great surface coverage and flexibility in use. The invasiveness of the procedure should be taken into account when the decision between two techniques is made.
Generally, sEEG implantation is well tolerated and imposes minimal risks of infection, haemorrhage or stroke57. However, the decision on performing sEEG should be taken with care and only with the certain indications (Table 2.1). The obvious disadvantages of the technique are limited surface coverage, inconvenience in electrical stimulation of the cortex, age restriction of at least two years old (due to insufficient thickness of 58
the skull) and a higher risk of intracranial haemorrhage compared to other methods .59
2.2.1.3. Electrocorticography
Electrocorticography (ECoG) was invented by American neurosurgeons Wilder Penfield and Herbert Jasper in 1954, when they published a research paper speculating on the new method . It has quickly found success in neuroscience community; nowadays, it 60
is extensively used by neurosurgeons all over the world and sometimes serves as a method of choice for preoperative epilepsy assessment.

For the ECoG application, a large craniotomy is needed, to expose the desired cortical area. Electrode arrays exist in form of grids and strips, containing up to 256 recording channels embedded into 4-5 mm diameter disks; the array is placed directly on the cortical surface, slipped underneath dura mater (Fig. 2.2.B). Such setup allows a large cortical coverage and is convenient for eloquent cortex localization. Simultaneously with the EEG recording, the electrical stimulation is usually applied . It helps to 61
localize areas responsible for speech, movement or sensory processing. Such direct cortical electrical stimulation (DCES) is also called cortical stimulation mapping and is sometimes considered as the gold standard for preoperative assessment and mapping brain functions in general , . Compared to sEEG, ECoG does not penetrate the brain 62 63
tissue, offers greater surface coverage and better access to cortical areas, and poses less risk of haemorrhage during the procedure. A brief comparison between the two methods is presented in Table 2.1.
Table 2.1. Comparison of invasive seizure foci localization techniques.
Each of the above methods has its advantages and disadvantages and in most cases of epilepsy surgery, the decision on preoperative assessment is taken between these two techniques. While sEEG uses penetrating electrodes and can reach deep brain areas, its spatial precision is low and the risk of haemorrhage is increased compared to ECoG. The latter, however, requires a generous craniotomy and is only useful in targeting shallow cortical zones.
Despite of seemingly superior nature of ECoG recording, the technique has several limitations. First, it obviously does not allow recording/stimulation of the deep brain structures, which is the primary advantage of sEEG. Second, it is highly invasive and the associated risks of wound infections, postoperative pain and other surgery-related complications are higher than ones of sEEG. Moreover, due to extensive post-surgical
Stereoelectroencephalography (sEEG) Electrocorticography (ECoG)
Indications Inconclusive results of non-invasive assessmentPrecise epileptogenic zone localizationMRI-negative epilepsyHeterotopic epileptogenic zonesPatients undergone a previous surgery
Inconclusive results of non-invasive assessmentExploring functional areasMRI-positive epilepsyCortical displasia, tuberous sclerosisNon-operated patients
Targets Deep cortical areasPreferred for bilateral recording
Shallow cortexPreferred for unilateral recording
Method Small holes in the skullMultiple probes implanted at depth128-256 channelsSampling frequency 256-512 Hz
Large cranial windowGrids/strips placed on the cortex32-256 channelsSampling frequency 1k-2k Hz
Tolerance Lower morbidityHigher tolerance
Higher morbidityLower tolerance
Risks Lower risk of infectionHigher risk of bleeding
Higher risk of infectionLower risk of bleeding
Additional (rare) risks
StrokePermanent neurological deficits
Cerebrospinal fluid leakElevated intracranial pressure

scarring of dura mater, it is very complicated to perform a second operation, so ECoG is preferred for patients which had not have invasive brain surgeries before. Bilateral recording is also challenging since it requires 2 large openings of the skull and a larger area of exposed cortex . Finally, the general tolerance and morbidity rate appears to 64
be lower for ECoG compared to sEEG57.
2.2.1.4. Hybrid EEG
In attempt to combine the strengths of both sEEG and ECoG, a hybrid technique was invented , , , , . It is based on simultaneous implantation of depth electrodes and 65 66 67 68 69
subdural grids, allowing access to a great extent of cortical area and to the structures at desired depth for recording and stimulation (Fig. 2.3). Unlike ECoG, the procedure does not require a cranial window, since subdural strips can be implanted through the burr holes slightly larger than ones used for sEEG placement67. Upon termination, the extraction of the electrodes is performed without a second surgery67. The technique is useful for cases in which non-invasive assessment points out towards non-lateralized epileptic activity but there is a strong suspicion of lateralized focus.
Figure 2.3. Hybrid EEG: a combination between sEEG and ECoG.
(A) 3D skull CT reconstruction demonstrating bilateral temporal implantation of subdural strips and Spencer’s depth electrodes, axial plane. Left side: 1,2,3 - subdural strips; 4 - Spencer’s depth electrode. (B) 3D skull CT reconstruction showing the left side. Subdural electrodes are emerging from the burr hole, sagittal plane. (C) A post-operative MRI scan visualizing two depth electrodes (1 and 4), and one subdural electrode (2) which is seen twice.
Courtesy of Mathon et al., 2015. Safety profile of intracranial electrode implantation for video-EEG recordings in drug-resistant focal epilepsy. Journal of neurology, 262(12), 2699–2712.69.
Insufficient coverage of the posterior temporal lobe and the inter-hemispheric cortex is the primary limitation of hybrid EEG57. Generally, the surgical procedure is less invasive than standard ECoG placement and the associated risks of infections and intracranial haemorrhage are low65,69; overall, the technique can be considered relatively safe. The factor which was reported to be associated with increased complication risks is MRI-negativity, which is probably due to the need in greater amount of implanted probes69. In any case, hybrid EEG is an efficient approach for preoperative diagnostics of severe intractable epilepsies.

2.2.1.5. Non-invasive EEG: electrical source imaging
Considering the main disadvantage of all the methods described above, their relative invasiveness, it is logical to assume that some developments have been made in the filed of non-invasive EEG-guided localization. Based on a traditional scalp EEG, these techniques offer a mean of preoperative assessment in epilepsy surgery and seizure foci localization, being a valuable tool among others.
The first mentions of so-called EEG source imaging technique (ESI) appeared in early 2000s. One of the first approaches was developed and introduced by Xu et al. in 2004, who used a non-recursive subspace algorithm FINE for an EEG source localization . 70
In 2007, another research team conducted an experiment on three paediatric patients, combining data obtained from scalp EEG analyzed with FINE algorithm with the data obtained from ECoG in the same patients . In all three subjects, the locations and the 71
extent of an epileptogenic zone were successfully localized, proving the efficiency of the technique. Another study on ten patients reported a similar result and provided a thorough description of the method (Fig. 2.4). It is based on 3D-modelling of a brain 72
and on high-density EEG simulation, which is combined with the real EEG data from a patient. Such process allows to accurately and non-invasively localize a seizure focus.
Figure 2.4. Spatio-temporal source localization approach (FINE).
The sophisticated algorithm allows to combine actual scalp EEG data obtained from a patient with a virtual model prediction on a seizure focus localization. A 3D reconstruction of patient’s head is made using MRI head scans. The lead-field matrix is then computed from the head model. The entire 3D brain is used a a source space for the formulae implemented in calculations. A high density EEG simulation is performed on a head model, distributed into different electrode configuration and FINE is applied. Subsequently, this data is compared with an actual patient’s EEG data and the prediction on the seizure focus location is made. Image on the bottom right shows a result from one of the patients suffering from frontal lobe epilepsy. EZ — epileptogenic zone.
Lu et al., 2012.
Another common approach is based on a technique called LORETA — a method for localizing electric activity in the brain based on the data from EEG-recording , . The 73 74
abbreviation stands for low resolution brain electromagnetic tomography; essentially, LORETA is an EEG-based neuroimaging technique which found its application in the field of epileptic surgery. In 2017, two independent studies were published, both using LORETA for a seizure foci localization , . The implementation reminds one of the FINE 75 76
algorithm; a 3D head model is built based on the MRI scans, and the EEG simulation is performed. Subsequently, the data is combined so that an accurate prediction of a seizure focus location can be made (Fig.2.5).
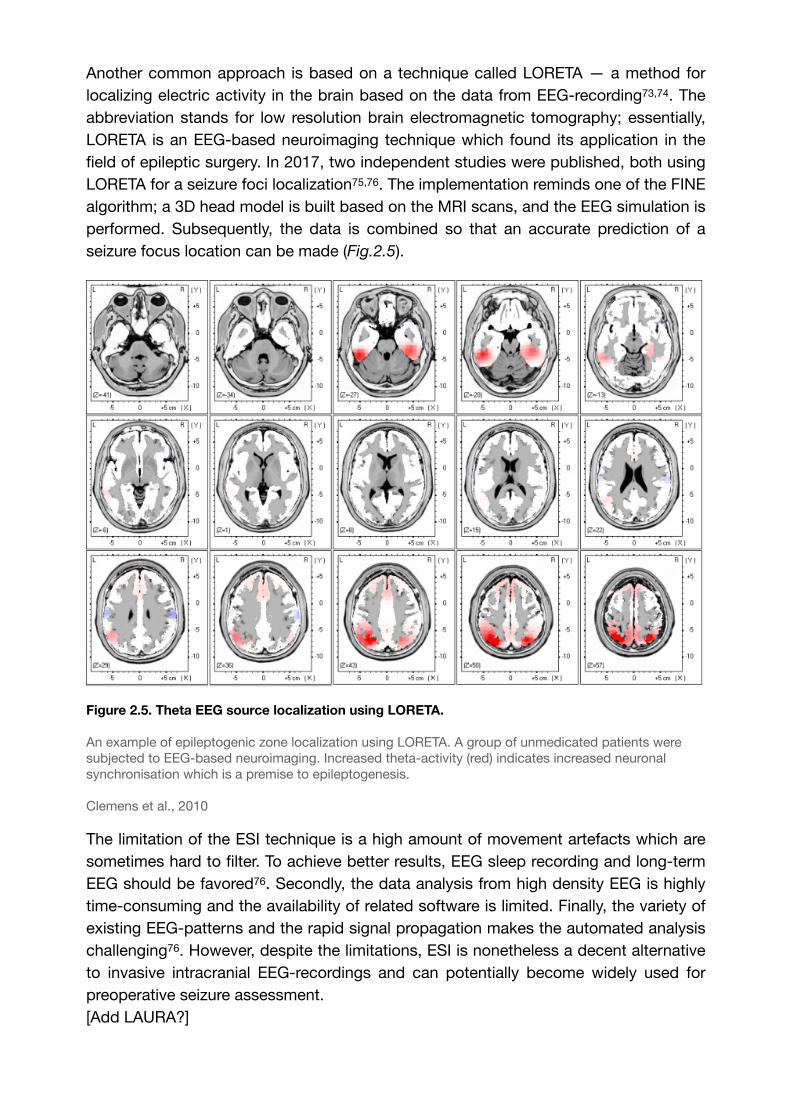
Figure 2.5. Theta EEG source localization using LORETA.
An example of epileptogenic zone localization using LORETA. A group of unmedicated patients were subjected to EEG-based neuroimaging. Increased theta-activity (red) indicates increased neuronal synchronisation which is a premise to epileptogenesis.
Clemens et al., 2010
The limitation of the ESI technique is a high amount of movement artefacts which are sometimes hard to filter. To achieve better results, EEG sleep recording and long-term EEG should be favored76. Secondly, the data analysis from high density EEG is highly time-consuming and the availability of related software is limited. Finally, the variety of existing EEG-patterns and the rapid signal propagation makes the automated analysis challenging76. However, despite the limitations, ESI is nonetheless a decent alternative to invasive intracranial EEG-recordings and can potentially become widely used for preoperative seizure assessment.[Add LAURA?]
2.2.2. Neuroimaging techniques
2.2.2.1. Computed tomography (CT)
In terms of seizure focus localization, CT cannot provide a wide range of diagnostic powers. Mostly it is used in urgent situations, such as acute seizures, and can only detect a gross organic pathology correlated to structural epilepsy . Nonetheless, it 77
has a number of advantages over MRI; it is available in most medical facilities and it can detect calcified lesions which are not specific for MRI. Generally, CT imaging has around 30% ratio of success in seizure focus detection and thus rarely serves as a 78
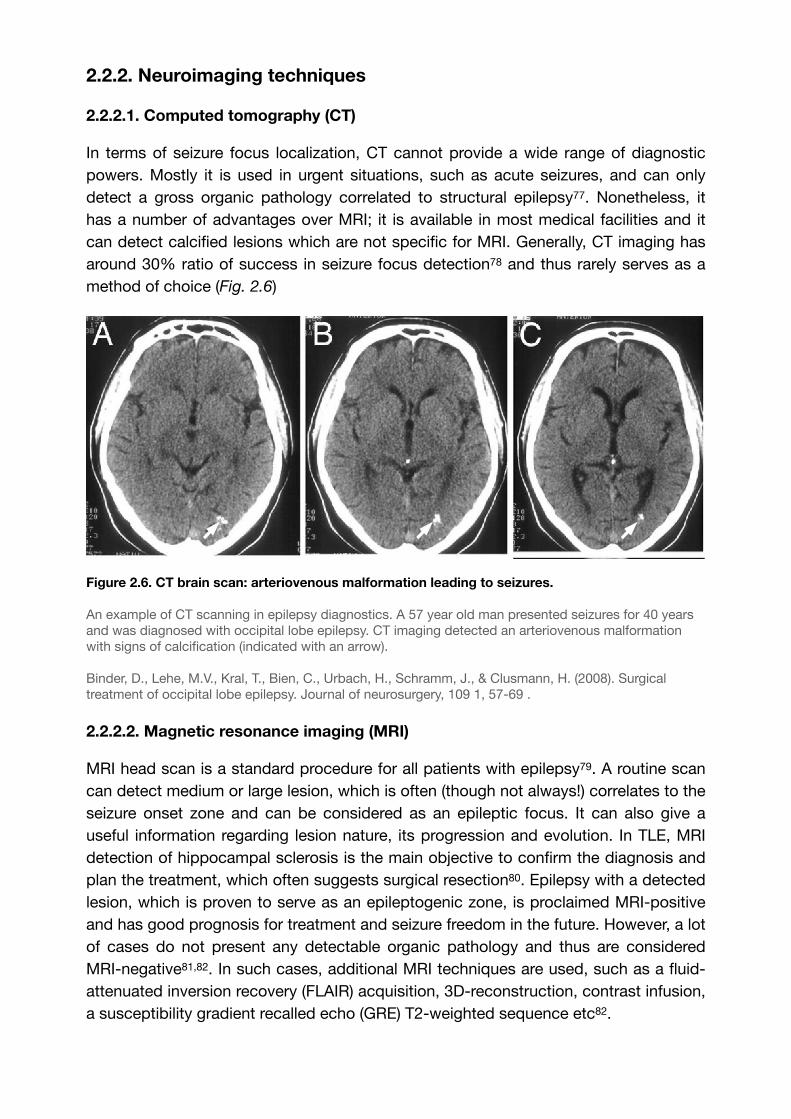
method of choice (Fig. 2.6)
Figure 2.6. CT brain scan: arteriovenous malformation leading to seizures.
An example of CT scanning in epilepsy diagnostics. A 57 year old man presented seizures for 40 years and was diagnosed with occipital lobe epilepsy. CT imaging detected an arteriovenous malformation with signs of calcification (indicated with an arrow).
Binder, D., Lehe, M.V., Kral, T., Bien, C., Urbach, H., Schramm, J., & Clusmann, H. (2008). Surgical treatment of occipital lobe epilepsy. Journal of neurosurgery, 109 1, 57-69 .
2.2.2.2. Magnetic resonance imaging (MRI)
MRI head scan is a standard procedure for all patients with epilepsy . A routine scan 79
can detect medium or large lesion, which is often (though not always!) correlates to the seizure onset zone and can be considered as an epileptic focus. It can also give a useful information regarding lesion nature, its progression and evolution. In TLE, MRI detection of hippocampal sclerosis is the main objective to confirm the diagnosis and plan the treatment, which often suggests surgical resection . Epilepsy with a detected 80
lesion, which is proven to serve as an epileptogenic zone, is proclaimed MRI-positive and has good prognosis for treatment and seizure freedom in the future. However, a lot of cases do not present any detectable organic pathology and thus are considered MRI-negative , . In such cases, additional MRI techniques are used, such as a fluid-81 82
attenuated inversion recovery (FLAIR) acquisition, 3D-reconstruction, contrast infusion, a susceptibility gradient recalled echo (GRE) T2‐weighted sequence etc82.
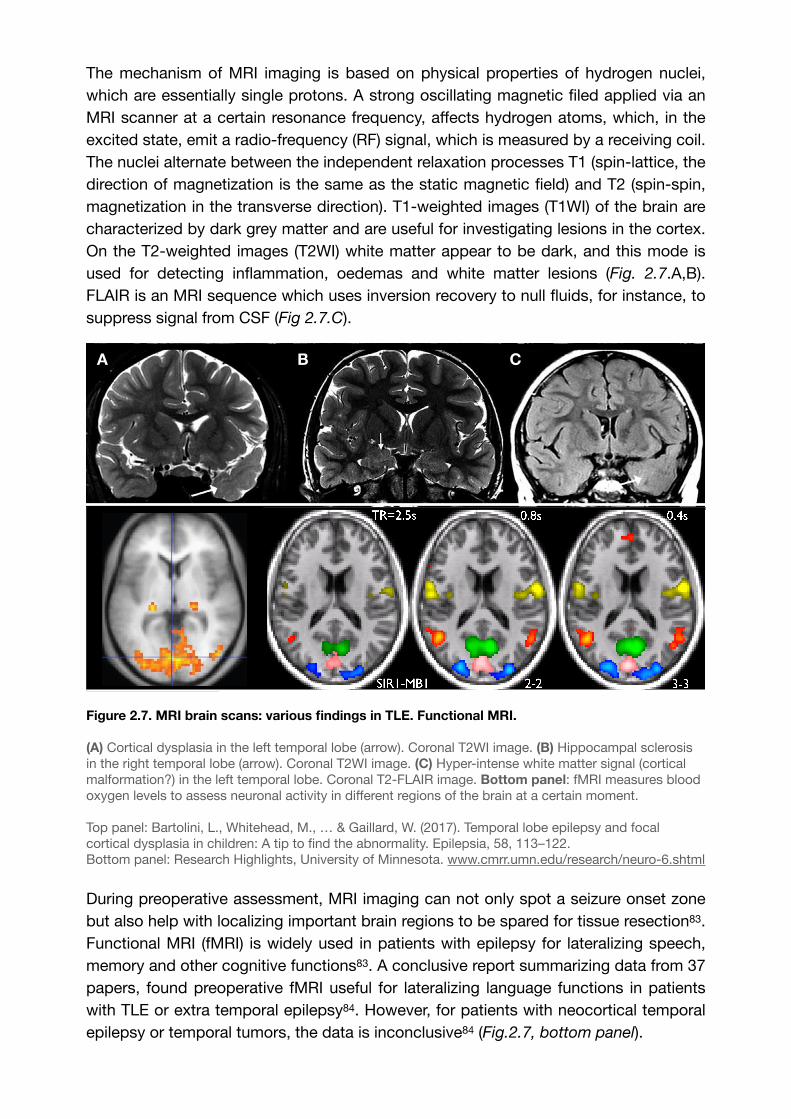
The mechanism of MRI imaging is based on physical properties of hydrogen nuclei, which are essentially single protons. A strong oscillating magnetic filed applied via an MRI scanner at a certain resonance frequency, affects hydrogen atoms, which, in the excited state, emit a radio-frequency (RF) signal, which is measured by a receiving coil. The nuclei alternate between the independent relaxation processes T1 (spin-lattice, the direction of magnetization is the same as the static magnetic field) and T2 (spin-spin, magnetization in the transverse direction). T1-weighted images (T1WI) of the brain are characterized by dark grey matter and are useful for investigating lesions in the cortex. On the T2-weighted images (T2WI) white matter appear to be dark, and this mode is used for detecting inflammation, oedemas and white matter lesions (Fig. 2.7.A,B). FLAIR is an MRI sequence which uses inversion recovery to null fluids, for instance, to suppress signal from CSF (Fig 2.7.C).
Figure 2.7. MRI brain scans: various findings in TLE. Functional MRI.
(A) Cortical dysplasia in the left temporal lobe (arrow). Coronal T2WI image. (B) Hippocampal sclerosis in the right temporal lobe (arrow). Coronal T2WI image. (C) Hyper-intense white matter signal (cortical malformation?) in the left temporal lobe. Coronal T2-FLAIR image. Bottom panel: fMRI measures blood oxygen levels to assess neuronal activity in different regions of the brain at a certain moment.
Top panel: Bartolini, L., Whitehead, M., … & Gaillard, W. (2017). Temporal lobe epilepsy and focal cortical dysplasia in children: A tip to find the abnormality. Epilepsia, 58, 113–122.Bottom panel: Research Highlights, University of Minnesota. www.cmrr.umn.edu/research/neuro-6.shtml
During preoperative assessment, MRI imaging can not only spot a seizure onset zone but also help with localizing important brain regions to be spared for tissue resection . 83
Functional MRI (fMRI) is widely used in patients with epilepsy for lateralizing speech, memory and other cognitive functions83. A conclusive report summarizing data from 37 papers, found preoperative fMRI useful for lateralizing language functions in patients with TLE or extra temporal epilepsy . However, for patients with neocortical temporal 84
epilepsy or temporal tumors, the data is inconclusive84 (Fig.2.7, bottom panel).
A B C
2.2.2.3. Positron emission tomography (PET)
PET imaging is based on radioactive isotope infusion and measuring its metabolism in different tissues and organs. In the field of epilepsy, the most common radioisotopes used is [18F]FDG (fluorine-18 fluorodeoxyglucose). Other options include [18F]MPPF (2′-methoxyphenyl-(N-2′-pyridinyl)-p-[18F]fluorobenzamidoethylpiperazine), [18F]cyclofoxy, [18F]Fallypride, [11C]Flumazenil and others . [18F]FDG is a glucose analog; it provides 85
information about cerebral glucose metabolism and is extensively used for a seizure onset zone localization85.
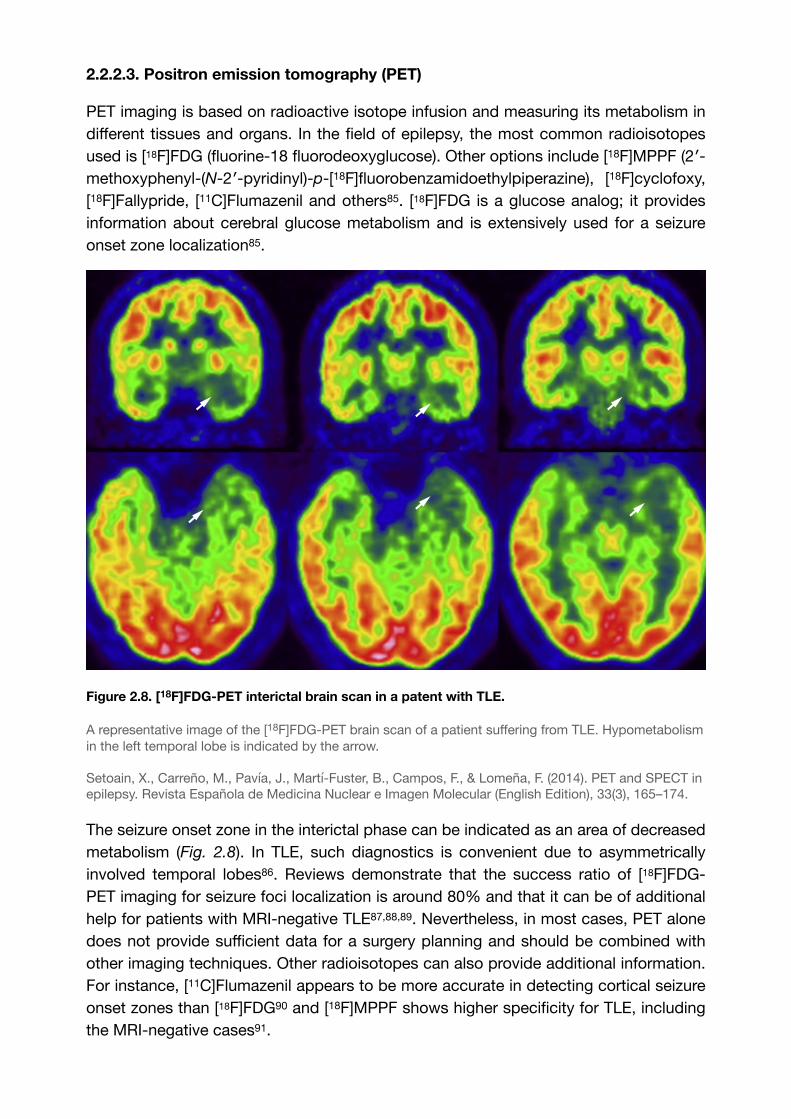
Figure 2.8. [18F]FDG-PET interictal brain scan in a patent with TLE.
A representative image of the [18F]FDG-PET brain scan of a patient suffering from TLE. Hypometabolism in the left temporal lobe is indicated by the arrow.
Setoain, X., Carreño, M., Pavía, J., Martí-Fuster, B., Campos, F., & Lomeña, F. (2014). PET and SPECT in epilepsy. Revista Española de Medicina Nuclear e Imagen Molecular (English Edition), 33(3), 165–174.
The seizure onset zone in the interictal phase can be indicated as an area of decreased metabolism (Fig. 2.8). In TLE, such diagnostics is convenient due to asymmetrically involved temporal lobes . Reviews demonstrate that the success ratio of [18F]FDG-86
PET imaging for seizure foci localization is around 80% and that it can be of additional help for patients with MRI-negative TLE , , . Nevertheless, in most cases, PET alone 87 88 89
does not provide sufficient data for a surgery planning and should be combined with other imaging techniques. Other radioisotopes can also provide additional information. For instance, [11C]Flumazenil appears to be more accurate in detecting cortical seizure onset zones than [18F]FDG and [18F]MPPF shows higher specificity for TLE, including 90
the MRI-negative cases .91
2.2.2.4. Single-photon emission computed tomography (SPECT)
Like PET, SPECT imaging uses radionuclides injected into bloodstream; actual imaging is performed via a gamma-camera which detects gamma-emission of the isotopes and measures cerebral blood flow (CBF) in various regions of the brain. It is approved for a pre-surgical evaluation of the seizure onset zone in patients with partial drug-resistant epilepsy85. The commonly used ligand for SPECT neuroimaging is technetium (99mTc) exametazime, which has a quick initial uptake and is used in both the interictal and the ictal periods.
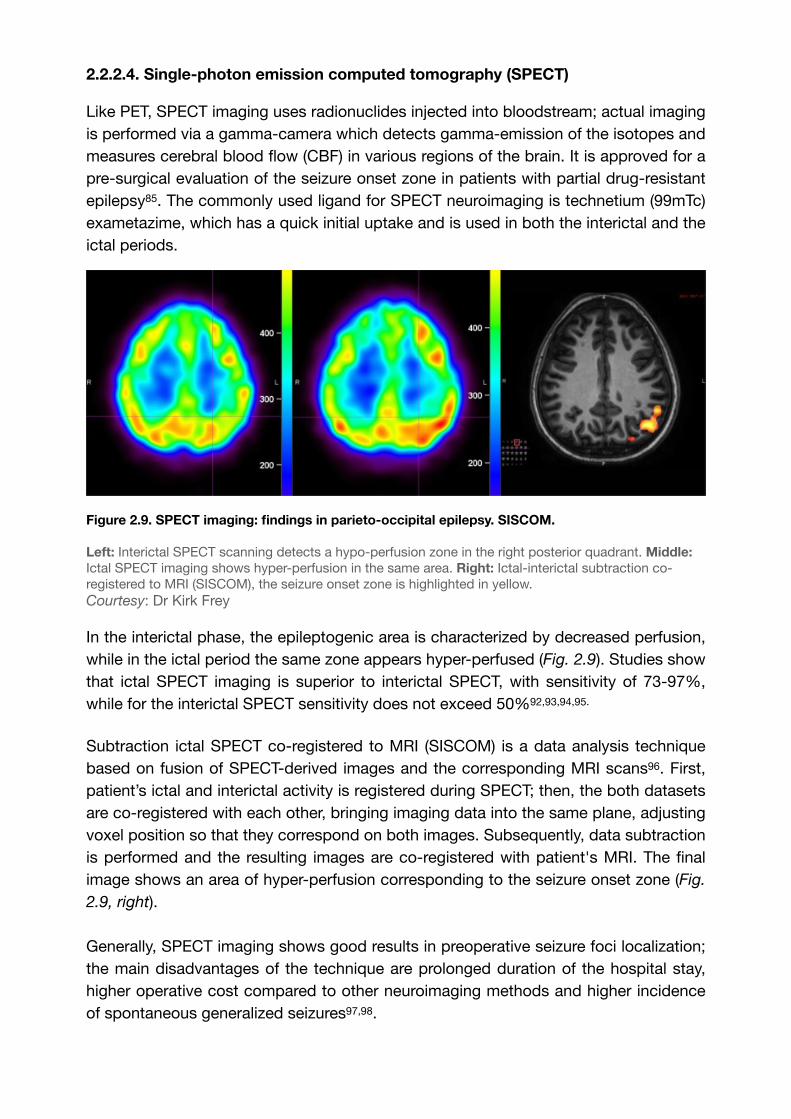
Figure 2.9. SPECT imaging: findings in parieto-occipital epilepsy. SISCOM.
Left: Interictal SPECT scanning detects a hypo-perfusion zone in the right posterior quadrant. Middle: Ictal SPECT imaging shows hyper-perfusion in the same area. Right: Ictal-interictal subtraction co-registered to MRI (SISCOM), the seizure onset zone is highlighted in yellow.Courtesy: Dr Kirk Frey
In the interictal phase, the epileptogenic area is characterized by decreased perfusion, while in the ictal period the same zone appears hyper-perfused (Fig. 2.9). Studies show that ictal SPECT imaging is superior to interictal SPECT, with sensitivity of 73-97%, while for the interictal SPECT sensitivity does not exceed 50% , , , .92 93 94 95
Subtraction ictal SPECT co-registered to MRI (SISCOM) is a data analysis technique based on fusion of SPECT-derived images and the corresponding MRI scans . First, 96
patient’s ictal and interictal activity is registered during SPECT; then, the both datasets are co-registered with each other, bringing imaging data into the same plane, adjusting voxel position so that they correspond on both images. Subsequently, data subtraction is performed and the resulting images are co-registered with patient's MRI. The final image shows an area of hyper-perfusion corresponding to the seizure onset zone (Fig.
2.9, right).
Generally, SPECT imaging shows good results in preoperative seizure foci localization; the main disadvantages of the technique are prolonged duration of the hospital stay, higher operative cost compared to other neuroimaging methods and higher incidence of spontaneous generalized seizures , .97 98
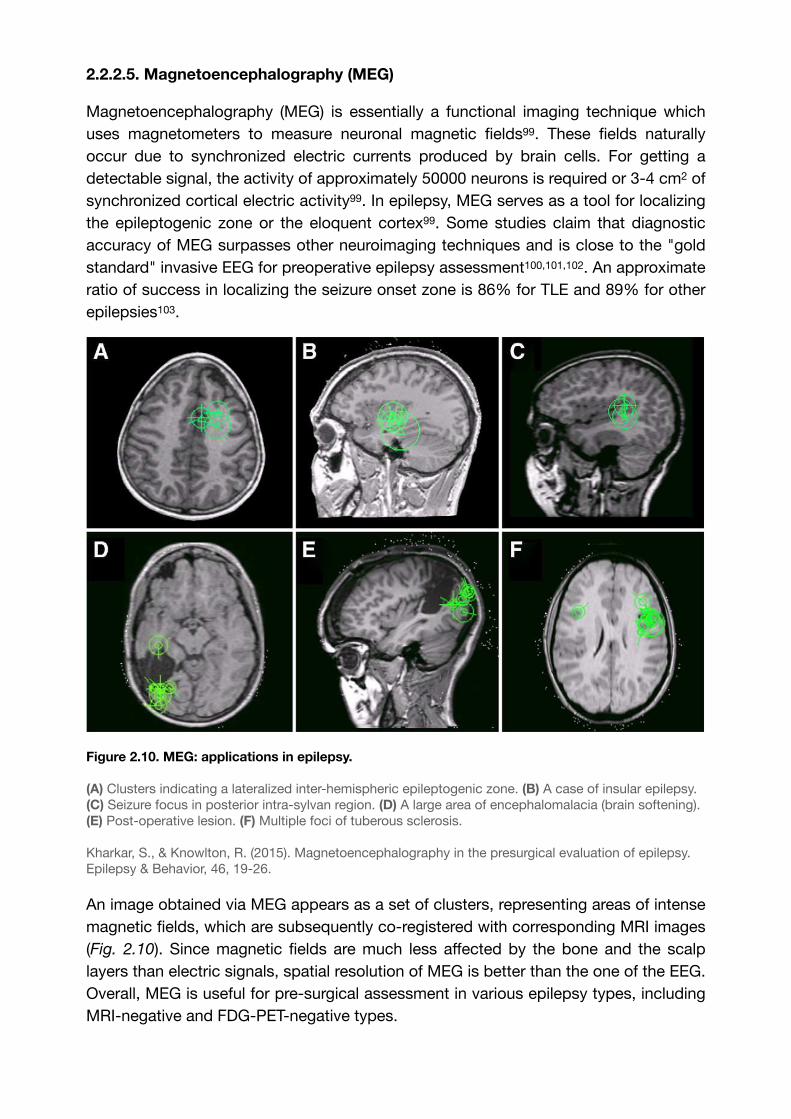
2.2.2.5. Magnetoencephalography (MEG)
Magnetoencephalography (MEG) is essentially a functional imaging technique which uses magnetometers to measure neuronal magnetic fields . These fields naturally 99
occur due to synchronized electric currents produced by brain cells. For getting a detectable signal, the activity of approximately 50000 neurons is required or 3-4 cm2 of synchronized cortical electric activity99. In epilepsy, MEG serves as a tool for localizing the epileptogenic zone or the eloquent cortex99. Some studies claim that diagnostic accuracy of MEG surpasses other neuroimaging techniques and is close to the "gold standard" invasive EEG for preoperative epilepsy assessment , , . An approximate 100 101 102
ratio of success in localizing the seizure onset zone is 86% for TLE and 89% for other epilepsies .103
Figure 2.10. MEG: applications in epilepsy.
(A) Clusters indicating a lateralized inter-hemispheric epileptogenic zone. (B) A case of insular epilepsy. (C) Seizure focus in posterior intra-sylvan region. (D) A large area of encephalomalacia (brain softening). (E) Post-operative lesion. (F) Multiple foci of tuberous sclerosis.
Kharkar, S., & Knowlton, R. (2015). Magnetoencephalography in the presurgical evaluation of epilepsy. Epilepsy & Behavior, 46, 19-26.
An image obtained via MEG appears as a set of clusters, representing areas of intense magnetic fields, which are subsequently co-registered with corresponding MRI images (Fig. 2.10). Since magnetic fields are much less affected by the bone and the scalp layers than electric signals, spatial resolution of MEG is better than the one of the EEG. Overall, MEG is useful for pre-surgical assessment in various epilepsy types, including MRI-negative and FDG-PET-negative types.
2.3. Modern non-invasive brain stimulation approach
Along with invasive EEG recording techniques and non-invasive neuroimaging tools, some other approaches have been proposed for a preoperative assessment and focal seizure localization (Fig. 2.11). An interesting idea is to use various types of electrical brain stimulation in attempt to achieve safe, non-invasive detection of the seizure onset zone with relative precision and with minimal risks for a patient. Recent studies investigated some of the brain stimulation techniques and the results seem promising.
2.3.1. Transcranial magnetic stimulation
2.3.1.1. Technical outline
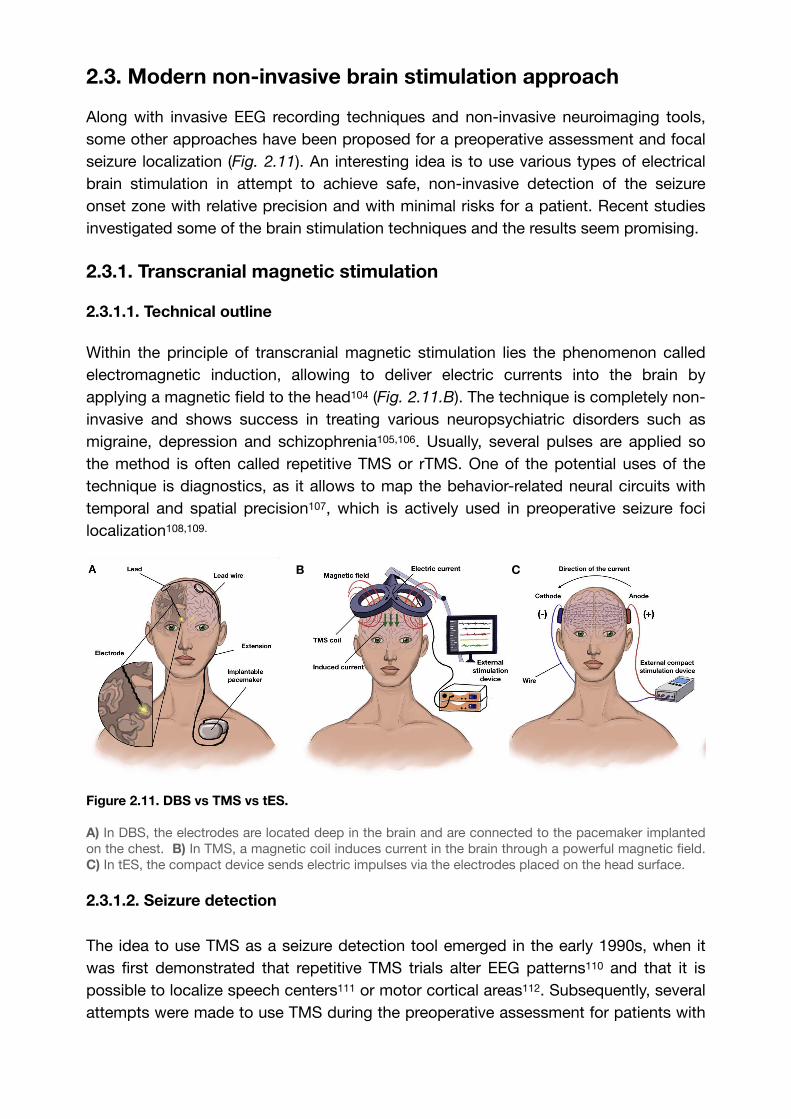
Within the principle of transcranial magnetic stimulation lies the phenomenon called electromagnetic induction, allowing to deliver electric currents into the brain by applying a magnetic field to the head (Fig. 2.11.B). The technique is completely non-104
invasive and shows success in treating various neuropsychiatric disorders such as migraine, depression and schizophrenia , . Usually, several pulses are applied so 105 106
the method is often called repetitive TMS or rTMS. One of the potential uses of the technique is diagnostics, as it allows to map the behavior-related neural circuits with temporal and spatial precision , which is actively used in preoperative seizure foci 107
localization , . 108 109
Figure 2.11. DBS vs TMS vs tES.
A) In DBS, the electrodes are located deep in the brain and are connected to the pacemaker implanted on the chest. B) In TMS, a magnetic coil induces current in the brain through a powerful magnetic field. C) In tES, the compact device sends electric impulses via the electrodes placed on the head surface.
2.3.1.2. Seizure detection
The idea to use TMS as a seizure detection tool emerged in the early 1990s, when it was first demonstrated that repetitive TMS trials alter EEG patterns and that it is 110
possible to localize speech centers or motor cortical areas . Subsequently, several 111 112
attempts were made to use TMS during the preoperative assessment for patients with
B C
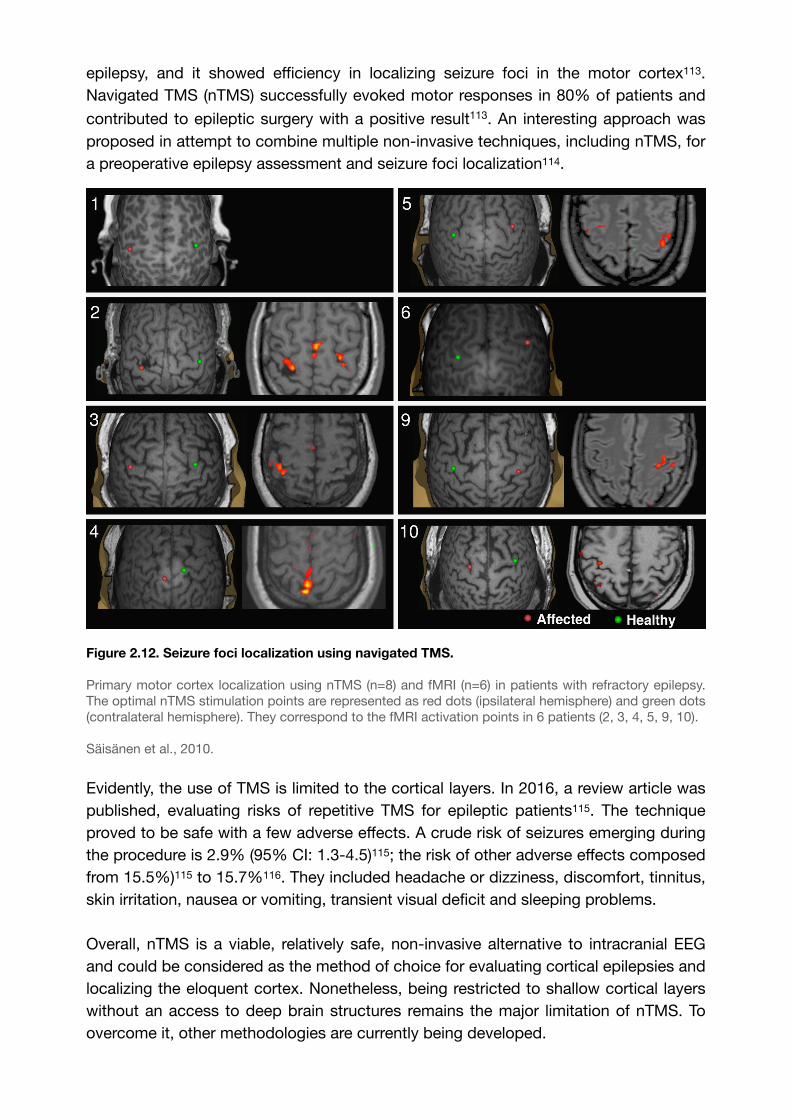
epilepsy, and it showed efficiency in localizing seizure foci in the motor cortex . 113
Navigated TMS (nTMS) successfully evoked motor responses in 80% of patients and
contributed to epileptic surgery with a positive result113. An interesting approach was proposed in attempt to combine multiple non-invasive techniques, including nTMS, for a preoperative epilepsy assessment and seizure foci localization .114
Figure 2.12. Seizure foci localization using navigated TMS.
Primary motor cortex localization using nTMS (n=8) and fMRI (n=6) in patients with refractory epilepsy. The optimal nTMS stimulation points are represented as red dots (ipsilateral hemisphere) and green dots (contralateral hemisphere). They correspond to the fMRI activation points in 6 patients (2, 3, 4, 5, 9, 10).
Säisänen et al., 2010.
Evidently, the use of TMS is limited to the cortical layers. In 2016, a review article was published, evaluating risks of repetitive TMS for epileptic patients . The technique 115
proved to be safe with a few adverse effects. A crude risk of seizures emerging during the procedure is 2.9% (95% CI: 1.3-4.5)115; the risk of other adverse effects composed from 15.5%)115 to 15.7% . They included headache or dizziness, discomfort, tinnitus, 116
skin irritation, nausea or vomiting, transient visual deficit and sleeping problems.
Overall, nTMS is a viable, relatively safe, non-invasive alternative to intracranial EEG and could be considered as the method of choice for evaluating cortical epilepsies and localizing the eloquent cortex. Nonetheless, being restricted to shallow cortical layers without an access to deep brain structures remains the major limitation of nTMS. To overcome it, other methodologies are currently being developed.

2.3.2. Transcranial electrical stimulation
2.3.2.1. Technical outline
Transcranial electrical stimulation (tES) is a type of electric brain stimulation, in which the electrodes are placed on the scalp surface (Fig. 2.11.C). There are three types of 117
tES: transcranial direct current (tDCS), alternating current (tACS) and random noise (tRNS) methods104. tDCS is performed by sending a direct current at low intensifies via the anode or the active electrode, which then returns through the cathode or the reference electrode . This technique primarily affects neurotransmitters, decreasing 118
the concentration of GABA and increasing glutamate concentration105. tACS is based on delivering current alternating between electrodes and mainly entrains neuronal firing without altering their excitability . tRNS method is similar to tACS, but while the latter 119
stimulates cells at fixed frequency, the former alternates at random frequencies and amplitudes within a certain range104. The mechanism of tRNS is poorly understood compared to other types of tES, but it is proven to be efficient in increasing cortical excitability . All three methods are experimental and are not officially approved as 120
neurological treatments, with the exception of tDCS, which is approved in EU for the treatment of major depressive disorder . A possible application of tDCS for seizure 121
foci localization is currently being researched.
2.3.2.2. Seizure detection
At the moment of composing this thesis, there are no actual reports of epileptogenic zone localization using tES. However, some studies indicate that hypothetically it might be possible to use tES for localizing brain functions , especially via introducing such 122
assets as multiple electrodes , ring configuration , or the principle of interference 123 124 125
between the electric fields . The major disadvantage of tES is its diffuse effect on the 126
brain and the lack of focality. The methods described above, potentially can increase precision of tES and allow some degree of focal stimulation. However, even taking the additional parameters into account, the obvious problem is an unwanted stimulation of multiple cortical areas. To achieve a precise focal brain stimulation without excitation of the cortex, a novel method of temporal interference (TI) stimulation was introduced.
2.3.3. TI stimulation: a potential diagnostic tool?
The revolutionary method of deep EBS using cortical electrodes was proposed by Dr Nir Grossman in 2017 . It is based on the generation of two high frequency electric 127
fields (such frequency is out of the range of neuronal response) but with a minor difference in frequencies, so that, overlapping, these fields would create an envelope oscillating at desired frequency, which is effective to stimulate neurons. This technique is very similar to tACS but, using the temporal interference (TI) principle it allows to reach brain areas well-below cortical surface, which is not possible by standard tES.
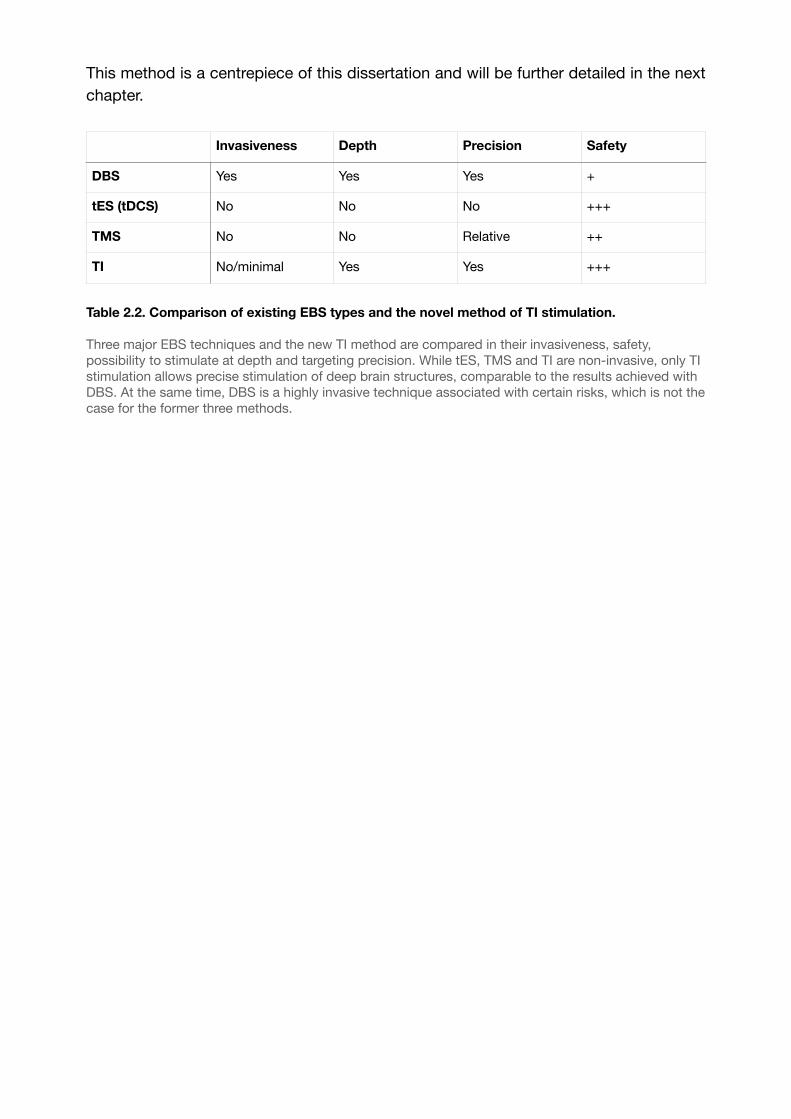
This method is a centrepiece of this dissertation and will be further detailed in the next chapter.
Table 2.2. Comparison of existing EBS types and the novel method of TI stimulation.
Three major EBS techniques and the new TI method are compared in their invasiveness, safety, possibility to stimulate at depth and targeting precision. While tES, TMS and TI are non-invasive, only TI stimulation allows precise stimulation of deep brain structures, comparable to the results achieved with DBS. At the same time, DBS is a highly invasive technique associated with certain risks, which is not the case for the former three methods.
Invasiveness Depth Precision Safety
DBS Yes Yes Yes +
tES (tDCS) No No No +++
TMS No No Relative ++
TI No/minimal Yes Yes +++

Chapter 3
3. Temporal Interference (TI) electrical stimulation
3.1. Basic concept
3.1.1. Neurophysiological basis
To speak about TI stimulation, it is important to understand how the basics of this type of stimulation lies within the physiological properties of neurons. Over the past several decades, it has been established that each neuron has its own frequency preference, meaning that it produces the best response at a certain frequency of an input current . It can be viewed as an electric circuit in which a resistor and a capacitor are 128
connected in parallel and an inductive element is added. In this case, the whole system acts as a bandpass and the impedance curve looks like a peak, which declines as the resonant frequency changes (Fig. 3.1). Even though oscillatory properties of different neurons in different parts of the brain are not the same, this system helps to find similarities between them and shed some light on neurophysiological basis of brain functioning.
Figure 3.1. Frequency-dependent properties of neurons.
(A) Input current. (B) A schematic representation of an electric circuit with an inductive element (left) and of a neuron (right). (C) The output current for the electric circuit (left) and for the neuron (right). (D) A resulting impedance in the electric circuit. (E) A resulting impedance in the neuronal cell. The impedance magnitude drops as frequency diminishes.
Generally, neurones in the mammalian brain oscillate in the range of 0.05-500 Hz, with faster frequencies confined to smaller neuronal pools and slower oscillations occurring
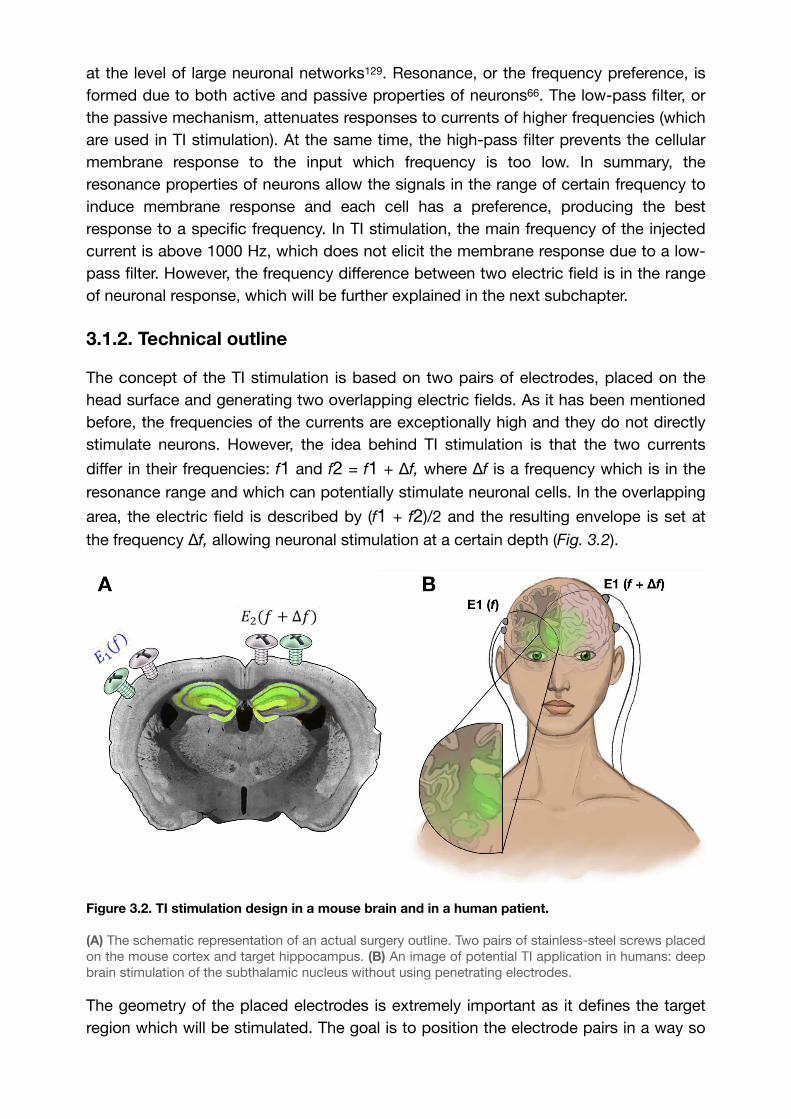
at the level of large neuronal networks . Resonance, or the frequency preference, is 129
formed due to both active and passive properties of neurons66. The low-pass filter, or the passive mechanism, attenuates responses to currents of higher frequencies (which are used in TI stimulation). At the same time, the high-pass filter prevents the cellular membrane response to the input which frequency is too low. In summary, the resonance properties of neurons allow the signals in the range of certain frequency to induce membrane response and each cell has a preference, producing the best response to a specific frequency. In TI stimulation, the main frequency of the injected current is above 1000 Hz, which does not elicit the membrane response due to a low-pass filter. However, the frequency difference between two electric field is in the range of neuronal response, which will be further explained in the next subchapter.
3.1.2. Technical outline
The concept of the TI stimulation is based on two pairs of electrodes, placed on the head surface and generating two overlapping electric fields. As it has been mentioned before, the frequencies of the currents are exceptionally high and they do not directly stimulate neurons. However, the idea behind TI stimulation is that the two currents
differ in their frequencies: f1 and f2 = f1 + Δf, where Δf is a frequency which is in the
resonance range and which can potentially stimulate neuronal cells. In the overlapping
area, the electric field is described by (f1 + f2)/2 and the resulting envelope is set at
the frequency Δf, allowing neuronal stimulation at a certain depth (Fig. 3.2).
Figure 3.2. TI stimulation design in a mouse brain and in a human patient.
(A) The schematic representation of an actual surgery outline. Two pairs of stainless-steel screws placed on the mouse cortex and target hippocampus. (B) An image of potential TI application in humans: deep brain stimulation of the subthalamic nucleus without using penetrating electrodes.
The geometry of the placed electrodes is extremely important as it defines the target region which will be stimulated. The goal is to position the electrode pairs in a way so

that the overlap between two electric fields appears in the CA3 hippocampal subfield. We used the virtual simulation software COMSOL to calculate the exact coordinates for electrode placement. Considering the anatomy of the hippocampus, it is important to follow the direction of the Schaffer collaterals, the axonal projections sending output signal to the CA1 subfield. In our case, medio-lateral electrode orientation allowed the best response to the stimulation, compared to the anterior-posterior positioning.
3.2. Research: Orientation-Tunable TI for Non-Invasive DBS in Epilepsy
3.2.1. Introduction
During pre-surgical evaluation, patients suffering from focal drug-resistant epilepsy often require invasive recordings using stereo-electroencephalography (SEEG), involving the implantation of numerous electrodes in different brain regions for the electrophysiological monitoring of seizure onset and the subsequent localization of an epileptogenic zone (EZ) , . Electrical stimulation from the intracranial electrodes is 130 131
often necessary to help define an EZ, with electrophysiological discharges and seizures triggered by different frequencies of stimulation , . In general, pathological 132 133
discharges from the EZ are characterized by several biomarkers, primarily the generation of high-frequency oscillations in the beta/gamma range, classically referred to as rapid-discharges , , . Although currently performed with invasive SEEG, 134 135 136
less-invasive methods capable of evoking such discharges and seizures in the localization of EZ tissue would be highly interesting as positive surgical outcomes are well-correlated with the removal of tissue regions which generate such high-frequency oscillations , , . 137 138 139
In the work presented here, we show the method of orientation-tunable Temporal Interference (ot-TI) for evoking seizure-like events (SLEs) having the characteristic frequencies of rapid-discharges. The method presents several clear advantages over classical brain stimulation and potential future applications in presurgical evaluation and treatment of human epilepsy. Our method of ot-TI can explore brain tissue, including sensitive regions previously unavailable for direct implantation, in a minimally-invasive way using electrodes on the cortex surface. Points of very focal stimulation at significant distances below the cortex surface are created by the envelopes of interacting electric fields applied by the surface electrodes to evoke SLEs in the hippocampus. The orientation of the electric field, as defined by the orientation of the surface electrodes, determines the effectiveness and the threshold of stimulation necessary to evoke the SLE. Temporal interference stimulation was
significantly popularized in 201765. The concept of TI stimulation exploits physiological properties of neurons, namely that the neuronal membranes filter electrical signals of frequencies more than 1 kHz, limiting depolarization and stimulation properties above these frequencies. In this work, we show that a crucial part of TI stimulation is the electrode configuration – not simply to move the spot of focal stimulation – but more

importantly to move the orientation of electric field of the TI with respect to the orientation of the underlying structure to be stimulated.
As shown in Figure 4.3.A, for classic TI, two electric fields are used with each field having a slightly different frequency, f1 and f2 = f1 + Δf, where f1 is selected above the threshold to evoke electrophysiological activity, for example 2000 Hz, and Δf is selected at a frequency typically used to evoke activity, for example 10 Hz. The two electric fields of frequency 2000 Hz and 2010 Hz create a low-frequency envelope of 10 Hz. Classically, electrode references have been placed in the chest of animals. However, this allows field lines to pass relatively arbitrarily through subcortical structures such as the hippocampus. As we show, based on the configuration of the electrode pairs it is possible to evoke dramatically different activity exploiting the orientation of the field lines with respect to the subcortical anatomy. In Figure 3.3.B, we utilize local references and orient electrode pairs with respect to axon tracts, the Schafer collaterals, of the CA field.
Figure 3.3. Orientation-tunable Temporally Interfering Electric Fields (ot-TI).
(A) Classic TI, using electrode pairs with references in the chest, a reliable method to non-invasively stimulate. However, field lines are arbitrary with respect to sub-cortical structures. (B) ot-TI, using electrode pairs with local references for controlled-orientation of electric field lines. Here, two perpendicular arrangements of ot-TI are shown, ML with field lines parallel to the alignment of axons (inset in A) in the hippocampus from CA3 to CA1 and (C) AP with field lines perpendicular to the same axons (inset in B) (D) Classic TI stimulation is performed with sine waves, however envelopes can also be created with other wave forms, for example square waves as seen here. The Ez envelope is overlaid on the mixed-frequency square wave signal to better illustrate the phenomenon. Finite-element method computation of ot-TI using square-waves, in both A and B show the maximum envelope amplitude of Ez, located at the same point of the CA3 using the two different orientations, ML and AP. These two configurations are compared experimentally to determine the minimally invasive efficacy of ot-TI to generate beta/gamma range oscillations and SLEs in the mouse hippocampus.
In the configuration shown here, we create electric fields parallel (medio-lateral, ML) and perpendicular (anterior-posterior, AP) to the axons in the mouse hippocampus.

Additionally, as shown in Figure 3.3.B, it is not necessary to exclusively utilize sine waves for stimulation, a critical detail as clinical stimulation equipment most often uses square waves (a more detailed explanation of square-wave TI in supplementary figure 1). The orientation-dependent configurations are shown with finite-element simulations in Figure 3.3.C, where the point of maximum envelope amplitude of electric field (Ez component) is located at the same point of the CA3, but using the two different
orientations, ML and AP.
3.2.2. Methodology
3.2.2.1. Animals
All experiments were performed in accordance with European Council Directive EU2010/63, and French Ethics approval (Williamson, n. APAFIS 20359 -
2019041816357133). For this study we used 4 groups of 8 male OF1 mice (Charles
Rivers Laboratories, France) aged 8-10 weeks. Animals were kept in transparent cages in groups of three to five, in a temperature-controlled room (20 ± 3°C) with a 12/12h night-day cycle. All animals had ad libitum access to food and water. Mice were divided into cohorts, differentiated by the electrode orientation: medio-lateral (ML) and anterior-posterior (AP) for both implanted and ot-TI. Additional mice were assigned for ot-TI studies with additionally implanted depth recording electrodes in the hippocampus to record the dynamics of the evoked SLE in the hippocampus vs the cortex. After the surgery, all mice were kept in separate cages to avoid fighting and to avoid damage to implanted electrodes.
Surgical procedure
The 32 mice were anaesthetized via an intraperitoneal injection of ketamine (50mg/kg) and xylazine (20mg/kg) and placed in a stereotaxic frame with the head adjusted for bregma and lambda in the same horizontal plane. After midline scalp incisions, the following stereotaxic coordinates were used for craniotomies: [AP: -1.94, ML: +0.5; -0.5; -3.9; -4.3] for the ML electrode orientation and [AP: +2.2; +1.1; -4.4; -5.5; ML: -2.04]. For the implantable twisted-pair platinum electrodes (from PlasticsOne; wire length = 2mm, individual wire diameter = 125μm), coordinates were AP: -1.94, ML: 2.8, DV: 1.57 using a 15-degree angle. All the coordinates were calculated using the Paxinos Atlas. Dura mater was gently removed and four stainless steel mini-screws (Component Supply, Miniature Stainless Steel Self-Tapping Screws: TX00- 2FH) were placed on the cortex without penetration into the brain tissue. Subsequently, dental cement (Phymep, SuperBond) was applied on the skull surface to fix the screws and the skull cap was formed using Dentalon. During the post-surgical recovery time (3 days), all the mice were observed for signs of pain, distress, and neurological complications.
3.2.2.2. TI Electrical stimulation

Electrical stimulation was performed using electrical stimulators (IntanTech, Intan 128ch Stimulation/Recording Controller) with two frequencies, 1200 Hz and 1250 Hz, and biphasic, bipolar pulses of 500 μs with the overall stimulation time of 10s
(complete details in supplementary Figure 2 and 3). Additional details on isolation of
stimulation systems is described in supplementary Figure 5. The intensity was gradually increased starting from 50 μA until seizures of stage 4 on the Racine scale were observed (ML) or until an animal exhibited intense motor response (i.e., convulsions in AP orientation). Control stimulations (1200/1200 Hz and 50/50 Hz) were performed at the threshold intensity of the TI session, with no observed SLEs. As expected, with no frequency offset, there is no envelope and thus no stimulation of the hippocampus. Each mouse received one full TI stimulation to identify the threshold and two subsequent control stimulations.
3.2.2.3. Direct stimulation
After 3 days of recovery following implantation, a protocol to induce and to characterize the AD threshold was applied using the Racine scale. Following previous work, ADs were defined as high-amplitude spikes and polyspike epileptiform events visible after the applied stimulus. We used a 10 second stimulation, bipolar, biphasic, at 50 Hz with a 500μsec pulse width, with the amplitude increased in 50 μA steps, starting from 50 μA, until reaching the seizure of stage 4 on Racine scale. The animal was given 30 min rest between attempted stimulations.
3.2.2.4. Behavioral evaluation
All mice were video monitored during stimulation. Behavioral responses were analyzed, taking care to distinguish between a motor response and an epileptic seizure. For seizure detection the following criteria were used, according to the Racine scale: Racine scale: Stage 0, no visible change in behavior; Stage 1, freezing with facial movements; Stage 2, head nodding; Stage 3, forelimb clonus; Stage 4, rearing without loss of balance; and Stage 5, rearing and loss of balance). The motor cortex response was evaluated, comparing the video recording with the simultaneous EEG recording, depth electrodes, and by extracting non-specific signs (limb twitching, vocalizing etc). In parallel to the LFP recording, video monitoring was continued during the complete stimulation/recording/rest sessions of the freely-moving animal in its cylinder open-field environment (50cm × 25cm × 25cm) to monitor the behavior and correlate it directly with the brain activity. Examples of Racine scale behavior after stimulation can be found on our Github (https://github.com/Florian139/Temporal-Interference.git).
3.2.2.5. Virtual simulation

In order to have an idea of the stimulation in terms of applied voltage and electric field, COMSOL Multiphysics, version 5.5, (www.comsol.com) was used to create Finite-element method simulations of the electrodes and environment. Prior to simulations, we created using Hexagon software a fine 3D mouse brain based on the Paxinos mouse brain atlas. We embedded the stimulation electrodes (stimulation and grounds) as platinum stimulators in an environment having the frequency-dependent complex permittivity values of brain tissue. Using electrical stimulation from our electrodes in COMSOL with our mouse brain mesh, the equipotential surfaces and electric field lines applied by the electrodes could be visualized in the hippocampus. The orientation and positioning of the electrodes could be optimized using the simulation to better target the boarder of the CA3. The physical extent of field lines with respect to the orientation of the axons of pyramidal cells in the hippocampus could be better seen. The mesh geometry utilized and the basics on the simulation physics utilized can be found on our Github (https://github.com/Florian139/Temporal-Interference.git).
3.2.2.6. Statistical analysis
All raw recordings were plotted and analyzed via Matlab. Statistical analyses (R software) of AD-thresholds were performed using the parametric one-way ANOVA and pairwise T-tests to calculate the probably significant differences in SLE amplitude, duration or severity between the 4 groups. For the TI group, the AD-threshold distribution appears to not follow a normal distribution. Therefore, all AD-thresholds were analyzed using a standard non-parametric Wilcoxon-Mann-Whitney tests. All necessary code to reproduce our figures, as lambda “.rhs” data file, can be found on our Github (https://github.com/Florian139/Temporal-Interference.git).
3.2.3. Results
We induced SLEs in the mouse hippocampus using both TI and implantable electrode protocols. The target of stimulation was the border of CA3 and CA1 (detailed coordinates in the methods section), well-below the cortex surface.
As seen in Figure 3.4.A-C for the two orientations of ot-TI, the maximum envelope is placed at the same location in the hippocampus, namely the border of the CA3 to CA1. Although the maximum is at the same point, the electric field lines are parallel (for ML) or perpendicular (for AP) to the axons of the hippocampus, the Schaffer Collaterals (SCs). In Figure 3.4.D, for all ML mice, the shape of the evoked activity, the increase in the frequency range of beta/gamma (20-40 Hz), and the behaviorally observed seizures were all consistent with a classically-described stimulation-induced focal seizure of the hippocampus, however this is the first minimally invasive evoked SLE using the method of TI. All of the 8 ML mice exhibited a behavioral seizure 4 on Racine scale (overall threshold of 700μA per electrode pair) correlated with an electrophysiological SLE.

Figure 3.4. Evoked SLEs in the hippocampus of freely-moving mice.
(A) and (B) depict the two ot-TI stimulation axes with electrodes to-scale. (C) The associated hotspots (maximum envelopes of electric field) are seen in the hippocampus, with the maximum amplitude placed at the CA3-CA1 junction as planned in the protocol. For ML/AP orientations, the electric field lines are parallel/perpendicular to the Schaffer collaterals (also see Figure 1). (D) Real biologically evoked SLEs in mice using ot-TI. For the ML orientation (left panels), parallel to the axons of the hippocampus, the shape of the evoked activity, the associated increase in frequencies of the beta/gamma range (20-40Hz), and observed behavioral seizures demonstrate a classic SLE, however it is the first minimally-invasive evoked SLE by using TI. The AP orientation (right panels), perpendicular to the axons of the hippocampus, showed no evoked SLEs, and for 8 mice only one showed a stage 4 behavioral seizure

on the Racine scale with a threshold above 900μA. (additional details, video, and stimulation protocol in supplementary Figure S3 as in our Github). Cortex recordings shown in supplementary Figure 1. In Figure 2D for AP mice, interesting things happen when the point of maximum envelope remains at the same coordinates, but the electric field is turned perpendicular to the axons of the hippocampus in a second group of mice. Clearly, no SLE activity is evoked with the AP orientation, although the amplitude of the envelope and its location are the same for both ML and AP. It is therefore necessary to increase the intensity of stimulation.
An explanation of stimulation using the AP orientation and its subsequent consequences in the motor cortex can be seen in Figure 3.5.A-C, where the envelope
of the electric field is compared to the underlying brain anatomy. In Figure 3.5.A, the
stimulation is equal to the stimulation applied in Figure 3.4 with no evoked SLE, and a maximum envelope of electric field is seen at the CA3-CA1 junction in the hippocampus with a small envelope in the neighboring cortex. In Figure 3.5.B, the stimulation from the electrode pairs is increased and correspondingly the maximum envelope in the hippocampus increases, however so does the envelope in the neighboring cortex. This increase in stimulation results in Figure 3.5.C, where a maximum limit of stimulation is reached as single-sided (corresponding to the implanted hemisphere) motor convulsions occur during the stimulation itself.
Figure 3.5. AP: non-preferential orientation, electric field envelope versus anatomy.
(A) Profile of the electric field and envelope for the stimulation in Figure 2B (600μA per pair), and (B) stimulation with the same orientation but with a higher stimulation amplitude (1500μA per pair). Clearly, continuously increasing the intensity of stimulation will increase the envelope of stimulation in the hippocampus, ideally leading to an SLE. However, the increase in stimulation amplitude creates a non-trivial increase of the envelope located in the cortex. (C) This envelope in the cortex is enough to evoke arbitrary activity and motor convulsions. As the stimulation intensity is increased for the AP orientation, significantly past the threshold for SLE in part B, single-sided motor convulsions are observed during the stimulation itself.
In summary, in Figure 3.5.A, corresponding to the stimulation applied in Figure 3.4, the amplitude of the electric field is clearly higher at the focal point of the coordinates in the hippocampus. However, due to the non-preferential orientation of the electric field

to the axons of the hippocampus, the threshold to evoke an SLE has not been reached. As amplitude increases to find the threshold for an SLE, a limitation of the TI method is observed with direct consequences for uncontrolled or indiscriminate orientations of TI electric fields.
Namely, as the amplitude of the focal spot increases for non-preferential orientations, so does the envelope along a radial axis moving away from the focal spot. A target structure along the radial path may have a threshold for activation, in this case convulsions in the motor cortex, which will be reached before the threshold for activation of the target, in this case seizure activity in the hippocampus. This understanding can be algorithmically formulated and used to tailor stimulation orientations, not limited to AP/ML, for other subcortical structures.
Figure 3.6. Orientation-controlled stimulation
of SLEs in the hippocampus with classic implanted stimulators.
To support the existence of orientation-dependent thresholds in TI, we subsequently demonstrate the dependence of orientation on thresholds using implantable electrodes. As seen in Figure 3.6.A and 3.6.B, we see that when using classic implanted stimulation electrodes, the threshold to evoke SLEs is highly dependent on
(A) Stimulation at the boarder of CA3 and CA1, for implantable electrodes with preferential ML orientations having equipotential surfaces of electric field paral le l to axons, and with non-preferential AP orientations having equipotential surfaces of electric field perpendicular to axons.
(B) The threshold to evoke SLEs with ML requires half the injected charge, compared to AP. This is consistent with our TI findings, that the AP orientation is more difficult for evoking SLEs
(C) As soon as the stage 4 on the Racine scale is reached, an increase in power intensity is shown with a peak in beta/gamma range oscillations between 20-40 Hz, correlating with SLEs (15s after the stimulation) for implantable orientations, however for AP (right panels) double the current is required.
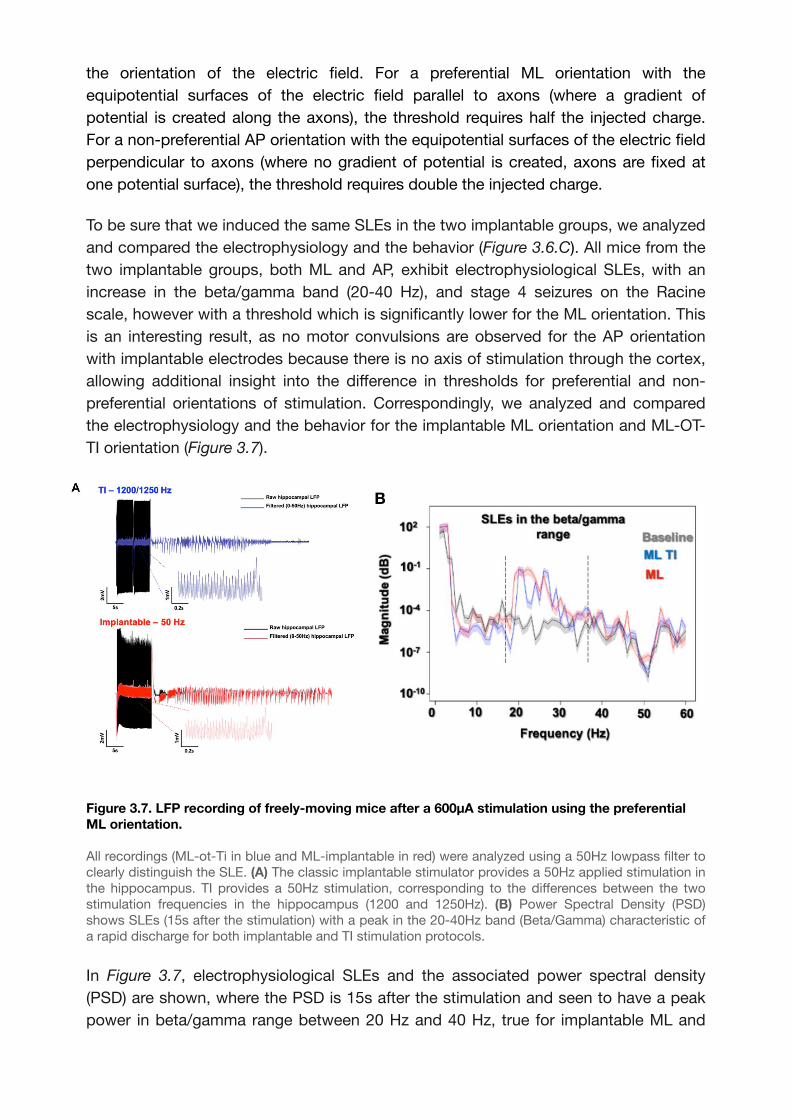
the orientation of the electric field. For a preferential ML orientation with the equipotential surfaces of the electric field parallel to axons (where a gradient of potential is created along the axons), the threshold requires half the injected charge. For a non-preferential AP orientation with the equipotential surfaces of the electric field perpendicular to axons (where no gradient of potential is created, axons are fixed at one potential surface), the threshold requires double the injected charge.
To be sure that we induced the same SLEs in the two implantable groups, we analyzed and compared the electrophysiology and the behavior (Figure 3.6.C). All mice from the two implantable groups, both ML and AP, exhibit electrophysiological SLEs, with an increase in the beta/gamma band (20-40 Hz), and stage 4 seizures on the Racine scale, however with a threshold which is significantly lower for the ML orientation. This is an interesting result, as no motor convulsions are observed for the AP orientation with implantable electrodes because there is no axis of stimulation through the cortex, allowing additional insight into the difference in thresholds for preferential and non-preferential orientations of stimulation. Correspondingly, we analyzed and compared the electrophysiology and the behavior for the implantable ML orientation and ML-OT-TI orientation (Figure 3.7).
Figure 3.7. LFP recording of freely-moving mice after a 600μA stimulation using the preferential ML orientation.
All recordings (ML-ot-Ti in blue and ML-implantable in red) were analyzed using a 50Hz lowpass filter to clearly distinguish the SLE. (A) The classic implantable stimulator provides a 50Hz applied stimulation in the hippocampus. TI provides a 50Hz stimulation, corresponding to the differences between the two stimulation frequencies in the hippocampus (1200 and 1250Hz). (B) Power Spectral Density (PSD) shows SLEs (15s after the stimulation) with a peak in the 20-40Hz band (Beta/Gamma) characteristic of a rapid discharge for both implantable and TI stimulation protocols.
In Figure 3.7, electrophysiological SLEs and the associated power spectral density (PSD) are shown, where the PSD is 15s after the stimulation and seen to have a peak power in beta/gamma range between 20 Hz and 40 Hz, true for implantable ML and
B
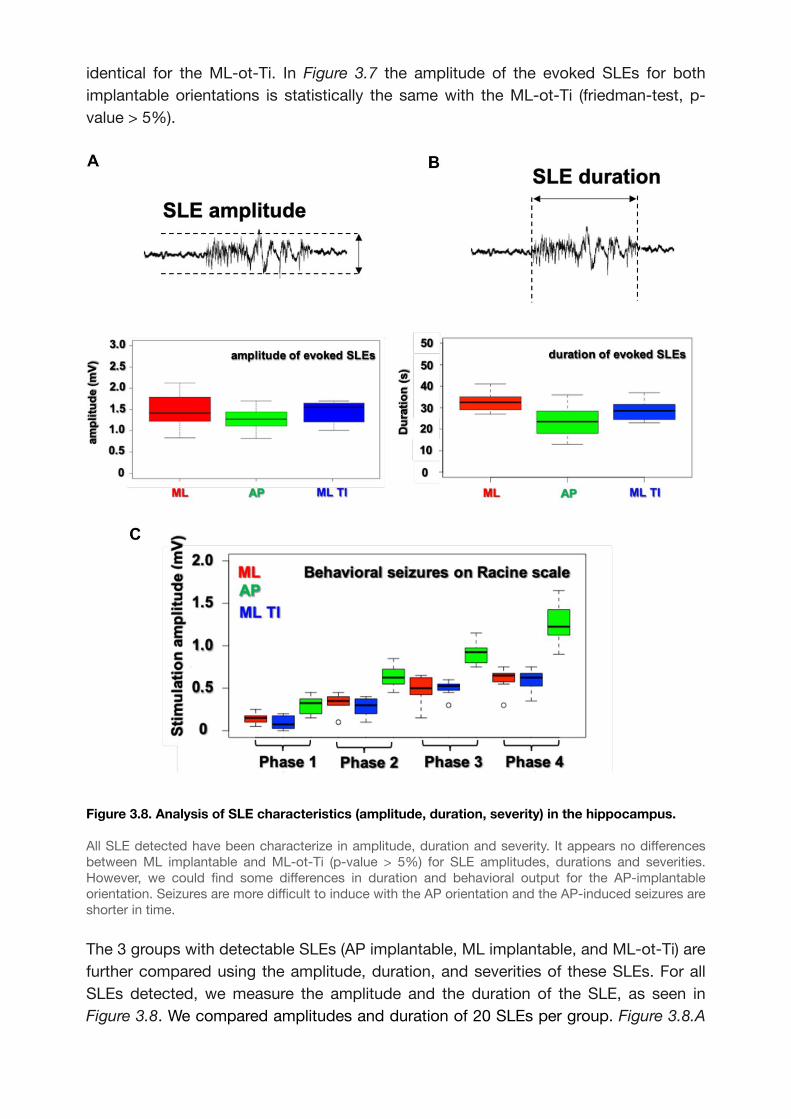
identical for the ML-ot-Ti. In Figure 3.7 the amplitude of the evoked SLEs for both implantable orientations is statistically the same with the ML-ot-Ti (friedman-test, p-value > 5%).
Figure 3.8. Analysis of SLE characteristics (amplitude, duration, severity) in the hippocampus.
All SLE detected have been characterize in amplitude, duration and severity. It appears no differences between ML implantable and ML-ot-Ti (p-value > 5%) for SLE amplitudes, durations and severities. However, we could find some differences in duration and behavioral output for the AP-implantable orientation. Seizures are more difficult to induce with the AP orientation and the AP-induced seizures are shorter in time.
The 3 groups with detectable SLEs (AP implantable, ML implantable, and ML-ot-Ti) are further compared using the amplitude, duration, and severities of these SLEs. For all SLEs detected, we measure the amplitude and the duration of the SLE, as seen in Figure 3.8. We compared amplitudes and duration of 20 SLEs per group. Figure 3.8.A

shows the SLE amplitude distribution for the 3 groups. Using first a normality test (Shapiro) and then a T-test, there were no differences between the 3 groups. SLEs durations show no differences between ML implantable and ML-ot-Ti, however these 2 groups are statistically different from the SLE durations of the AP implantable group. Consistent with the previous result, the severity of the seizure is also lower on all Racine scale stages for the AP implantable. It is simply more difficult to induce seizures with an AP orientation, and the induced seizures are also shorter and less severe than the seizures with ML orientations. However, the two ML groups, implantable and TI, are statistically identical in term of induced seizures, amplitudes, durations, and severities meaning that the minimally-invasive method is generating the same electrophysiological and behavioral event, but without the necessity of implanting the tissue.
Indeed, orientation dependence in TI is not surprising, as orientation dependent stimulation is known to exist for implantable electrodes in the field of Parkinson’s disease , . The main point is that identical evoked SLEs in the beta/gamma range 140 141
can be attained in the hippocampus using temporally interfering electric fields, without the necessity of an implanted stimulator, if care is taken to account for the orientation of the electric field with respect to the axons of the hippocampus. We created both orientations of preferential ML and non-preferential AP mice using ot-TI. Based on our calculation of geometry and field strength, we can locate the focus at the exact intersection of the CA3 and CA1 used in the implanted stimulation experiment.
3.2.4. Discussion
Clinically, comparing electrical activity from different brain regions, in particular electrically stimulated brain regions, in real-time as seizures emerge is fundamental to the identification of the EZ . Seizure onset is characterized by dramatic changes in 142
brain rhythms with several patterns of onset often observed, namely preictal epileptic spikes, trains of spikes, rapid-discharges, and slow-wave complexes with frequencies involved in the range of beta/gamma (15–30 Hz, 20-40 Hz in mesial temporal seizures) , . Typically, 8–15 implants are located in patient’s brain (generally 0.8 mm 143 144
diameter, with multiple electrode contacts 2 mm long and 1.5 mm apart), allowing stimulation at various mesial and lateral sites usually using lower frequencies (typically 1–50 Hz, for 5s) to map functional cortex or to evoke seizure onset . Unfortunately, 145
there are some parts of the brain which remain unreachable for electrode implant due to the presence of blood vessels, eloquent cortex, or other surgically-complicated implant locations and possible subsequent deficits. Clearly, there are limitations to the number and placement of intracranial electrodes and, as we have demonstrated here, ot-TI could be an interesting and additional tool for evoking and stimulating seizure onset. The possibility to explore some parts of the brain unreachable by SEEG electrodes could be very interesting during clinical investigation of, for example, the insula as it has a high density of arteries and veins and it is well-known to be a very difficult tissue area of the brain to operate . This is important because the insula is 146

also well-known to be implicated in refractory partial epilepsies and can be part of the epileptogenic zone of the patient , . Unfortunately, it is commonly described that 147 148
patients cannot undergo a proper functional mapping of this site because of the risk of haemorrhage during implant . Correspondingly, ot-TI also has the possibility to 149
reduce the overall number of SEEG electrodes implanted. There are numerous well-documented complications in SEEG electrode surgery, related infections, and complications which can results in necessary explanation of the electrode59, . A 150
reduction in SEEG electrodes would correspondingly reduce the number of SEEG-related complications.
In summary, TI is a very recently introduced concept for brain stimulation11. In the work here, we have utilized a new orientation-tunable form of TI (ot-TI) to evoke SLEs in the mouse hippocampus having the range of beta/gamma frequencies seen in MTLE, but without the necessity of implantable electrodes. Our target was the classic CA3-CA1 border structure used in epilepsy studies where the neurons of the CA and their axons, the Schaffer collaterals, are a well-known target for evoking seizures in vivo and in vitro with penetrating electrodes , , . In our alternative technique 151 152 153
relying exclusively on non-penetrating surface electrodes and ot-TI electric fields, we have shown that the SLEs generated are indeed focally evoked in the hippocampus and identical to focally evoked SLEs generated with classically implanted electrodes. We additionally demonstrated the use of square-waves with TI stimulation, as clinically, square-waves are most often used in stimulation. As we have shown, the orientation of the electric field plays a key role in evoking events, and if carefully controlled, optimal orientation dramatically lowers thresholds for both implantable electrodes and correspondingly the minimally-invasive ot-TI, replicating the implantable stimulation without the need of penetrating the cortex. This method has promise to significantly advance our capacity of probing the organization of spatiotemporal brain activity and could dramatically increase the explorable tissue for clinical definition of the EZ. Finally, it could make it possible to deliver therapeutic stimulation in a highly controlled and accurate non-invasively to deeper regions of the brain, an interesting topic for a subsequent study. Regardless, the technique undoubtedly holds great promises with potential applications in epilepsy, but also for a wide range of other brain disorders currently managed by electrical brain stimulation.
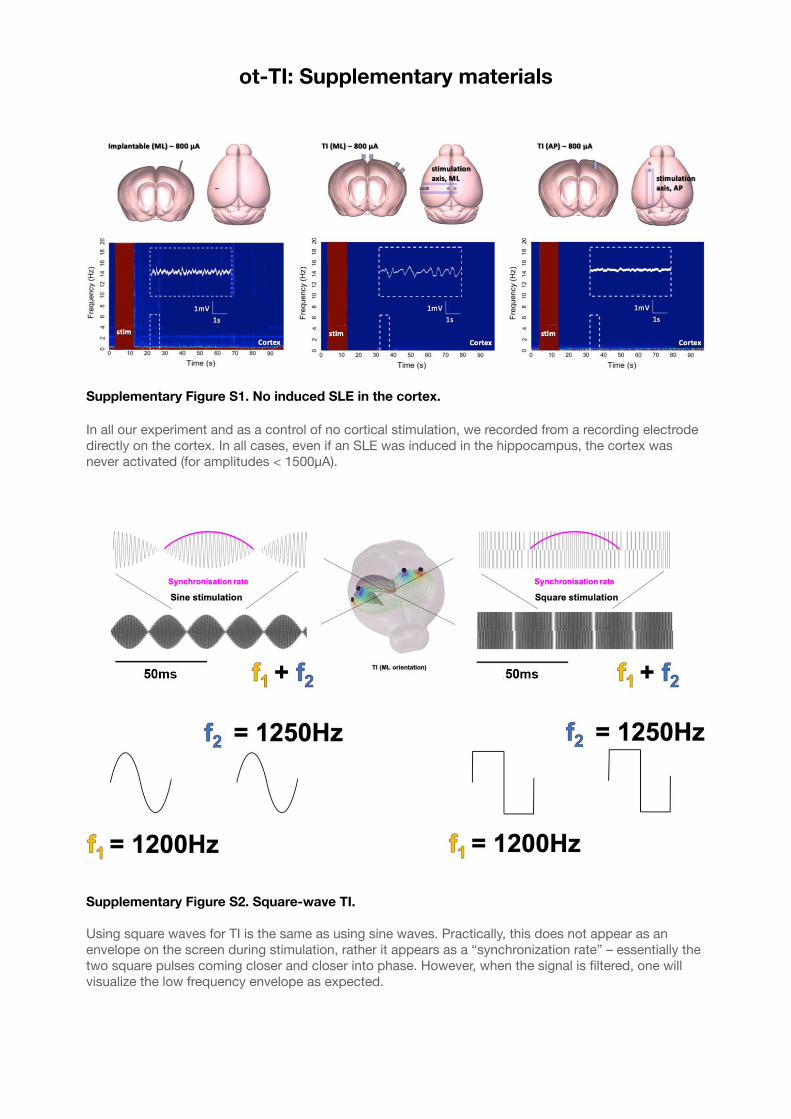
ot-TI: Supplementary materials
Supplementary Figure S1. No induced SLE in the cortex.
In all our experiment and as a control of no cortical stimulation, we recorded from a recording electrode directly on the cortex. In all cases, even if an SLE was induced in the hippocampus, the cortex was never activated (for amplitudes < 1500μA).
Supplementary Figure S2. Square-wave TI.
Using square waves for TI is the same as using sine waves. Practically, this does not appear as an envelope on the screen during stimulation, rather it appears as a “synchronization rate” – essentially the two square pulses coming closer and closer into phase. However, when the signal is filtered, one will visualize the low frequency envelope as expected.
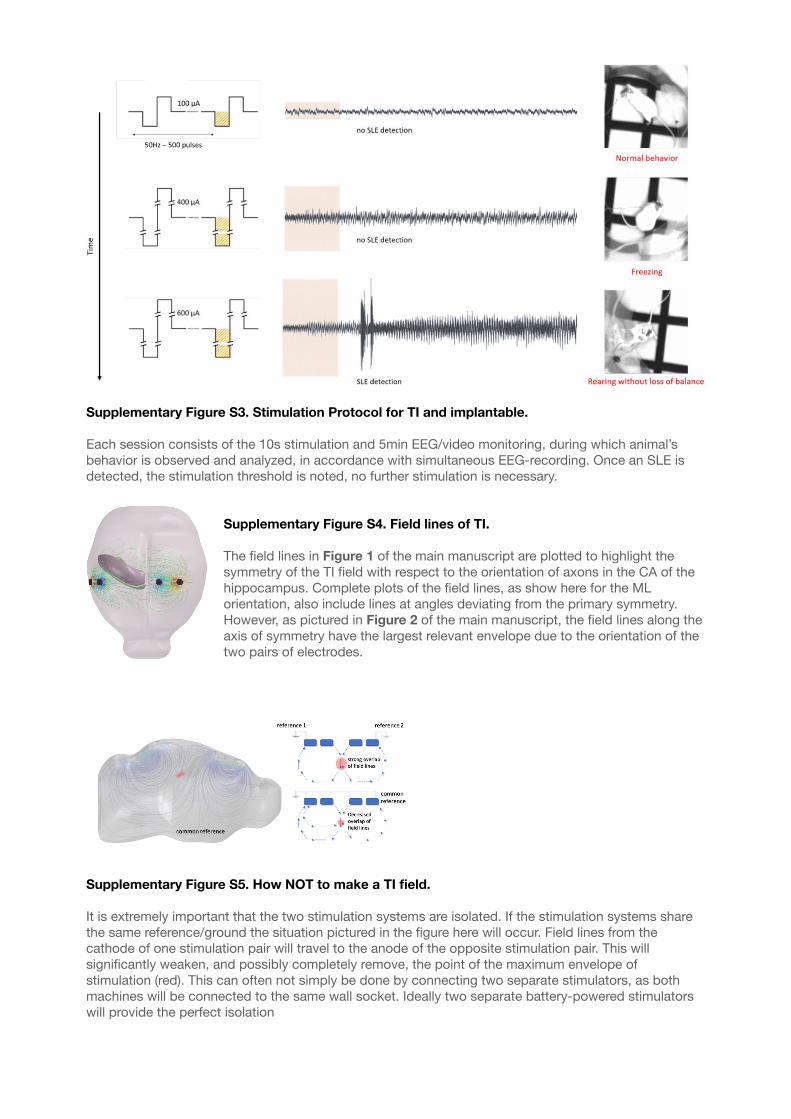
Supplementary Figure S3. Stimulation Protocol for TI and implantable.
Each session consists of the 10s stimulation and 5min EEG/video monitoring, during which animal’s behavior is observed and analyzed, in accordance with simultaneous EEG-recording. Once an SLE is detected, the stimulation threshold is noted, no further stimulation is necessary.
Supplementary Figure S4. Field lines of TI.
The field lines in Figure 1 of the main manuscript are plotted to highlight the symmetry of the TI field with respect to the orientation of axons in the CA of the hippocampus. Complete plots of the field lines, as show here for the ML orientation, also include lines at angles deviating from the primary symmetry. However, as pictured in Figure 2 of the main manuscript, the field lines along the axis of symmetry have the largest relevant envelope due to the orientation of the two pairs of electrodes.
Supplementary Figure S5. How NOT to make a TI field.
It is extremely important that the two stimulation systems are isolated. If the stimulation systems share the same reference/ground the situation pictured in the figure here will occur. Field lines from the cathode of one stimulation pair will travel to the anode of the opposite stimulation pair. This will significantly weaken, and possibly completely remove, the point of the maximum envelope of stimulation (red). This can often not simply be done by connecting two separate stimulators, as both machines will be connected to the same wall socket. Ideally two separate battery-powered stimulators will provide the perfect isolation

Chapter 4
4. Conclusion and future directions
4.1. Summary
The method described in this thesis is just one of the many attempts of influencing the brain networks via electrical stimulation. While the general concept of EBS has been used for decades, the novel paradigm of ot-TI stimulation offers hitherto impossible means to stimulate neurons at depth without using penetrating electrodes. The entire idea of non-invasive DBS seems to be somewhat science fiction but we managed to prove that it is indeed possible to achieve hippocampal stimulation using only cortical electrodes.
We have modified the initial TI design proposed by N. Grossman65, removing the bone layer, which allowed to amplify the signal, and switching the position of the reference electrodes, which produced significant difference in comparison with classic chest-positioned reference electrodes. We thoroughly explored the brain anatomy in order to better understand the underlying mechanisms of TI stimulation applied to a mouse hippocampus. We proposed the orientation-dependent model, in which the electric fields are aligned with the main axonal tracts in the CA region of the hippocampus. In this way, it is possible to achieve results with smaller current amplitude and with lower risks for an animal. Furthermore, such configuration allows relative flexibility and even placement of additional electrodes if necessary. We proved that stimulation using square waves instead of sine waves is possible. Finally, we found a dependance between the amplitude of stimulation and the behavioral response, which can be clearly distinguished as either real SLEs or the motor cortex activation.
Whether TI stimulation will find its application in clinical medicine or research, it is still interesting to speculate on the topic of non- or minimally invasive DBS. For instance, a significant cohort of epilepsy cases are drug-resistant and for such individuals surgical treatment is often the only available option. However, any surgical intervention poses certain risks, such as infections, haemorrhages, neurological deficits etc. Being able to target deep brain structures without penetrating the brain tissue could provide doctors with a safe alternative to traditional invasive techniques. Whether the aim is to localize epileptic focus or to stimulate certain brain structures, our experiments showed that both options are possible. Further research is still needed to shed more light on the exact mechanisms of how the TI stimulation affects the brain, but we can already say now that it is much safer and less labor-intense than the traditional stimulation method using implantable electrodes. Besides, positioning of the stimulators can be adjusted in a way which fits best the stimulation target, and considers anatomical features such as blood vessels, tumors and other structures to avoid.

4.2. Future directions
Having discovered the possibility to non-invasively stimulate deep brain structures, we opened a way to different opportunities and directions, each of which could be further investigated and potentially lead to new inventions.
4.2.1. Quadropole TI stimulation
One of such possibilities is to complicate the electrode geometry and electric fields interaction by including additional electrode pairs. As it has been already mentioned, stimulator placement plays a crucial role in TI stimulation, allowing careful and precise targeting. By adding an extra pair of electrodes, it is possible to enhance stimulation strategy and to make it even more precise, while reducing current amplitude and diminishing the associated risks. Currently, the first attempts of so-called quadropole TI stimulation are being made in our laboratory and the first results seem promising.
4.2.2. Therapeutic application
Another interesting application of TI stimulation is clinical use in the fields of epilepsy and other neurological conditions. In this thesis, several existing EBS techniques have been thoroughly described and further research on the TI could put it in one line with already approved methods such as DBS in PD, tES in depression and VNS for cluster headaches and migraines. Theoretically, TI stimulation could be applied to either of these conditions, since its relative safety allows to perform experiments associated with extremely low risks compared to the classic EBS with implantable electrodes. Currently, the most logical path for the TI research is epilepsy, considering that the working model for the CA3 localization is developed in our laboratory, as well as an extensive research on epilepsy models in rodents. It would be interesting to investigate how the TI stimulation interacts with an existing epileptic focus made by a neurotoxin injection. One of the many possibilities is to stimulate it directly in order to suppress seizures, another is to find a possible target among known related brain structures.
4.2.3. Diagnostics
Considering the flexibility of electrode placement and the possibility to rotate the setup during an actual stimulation session, it is safe to assume that the TI stimulation could be used as a diagnostic tool for epilepsy, helping to localize a seizure focus in the brain. In combination with EEG, it could provide a convenient way to explore seizure dynamics, its origin and propagation in a patient’s brain, without the need in invasive surgery. To conclude, we state that the TI stimulation has the potential to be a powerful tool in both diagnostics and therapy, and with advances of further research, it might happen soon.

Annex
A Future directions: The Kainic Acid models of TLE
A.1. Introduction
Mesial temporal lobe epilepsy (MTLE) is the most common type of partial epilepsy in adults . It is characterized by recurrent spontaneous seizures, which are often 154
resistant to drug treatments39. Numerous pathological alterations have been reported in patients with MTLE, including hippocampal sclerosis (HS) and cell death , mossy 155
fiber sprouting , reorganization of hippocampal interneuronal networks , , , 156 157 158 159
alterations in neuropeptide signalling , and synaptic transmission regulation , 160 161 162
granular cell dispersion and gliosis , blood-brain-barrier dysfunction and 163
angiogenesis . Whether these changes are the cause and/or the consequences of 164
seizures and comorbidities, such as cognitive deficits, anxiety, and depression, remains to be adequately addressed.
In such a context, animal models are interesting to consider because they allow detailed investigations with invasive procedures and drug testing that cannot be performed in patients. The translation of results obtained in the experimental model to patients is always debatable, if only because a rodent brain is not like a human brain. Yet, despite this intrinsic limitation, experimental models can provide hypotheses and clues relevant to humans. A clear advantage of empirical models is their relative homogeneity. The genetic variability is decreased in inbred rodents, and their housing in animal facilities provide a stable environment. If this facilitates the task of researchers, it does not reproduce the situation in humans, who are characterized by a high diversity in terms of genetic background and life experiences. Several experimental models of epilepsy have been developed, originally to mimic MTLE, including the pilocarpine model, the electrode-based kindling model, and the kainic acid (KA) model. Although homology between a rodent model and human epilepsy cannot be claimed, experimental models must reproduce the main symptom: spontaneous recurrent seizures (SRSs). If other pathological traits (e.g., mossy fiber sprouting or HS) or comorbidities (e.g., cognitive deficits and depression) exist, the relevance to the human condition may gain more weight. In this review, we will focus on the KA rodent model, its advantages and limitations, methods of administration, neuropathological, electrophysiological, histological, and behavioral outcomes.
A.2. Models of TLE: a brief overview
The main goal of creating a reliable animal model of epilepsy is to develop a chronic condition, which is consistent and efficient in generating spontaneous recurrent seizures (SRS). Several methods have been developed, all of them are based on different approaches but generally using an initial brain insult.

The administration of various chemoconvulsants either systemically or directly into the brain is commonly used to trigger epileptogenesis, the process that leads to the development of SRS. Pilocarpine and kainic acid are broadly used chemoconvulsants. Pilocarpine, which is a muscarinic acetylcholine receptor agonist, was first proposed as a way to generate experimental epilepsy in rats by W. Turski . Intraperitoneal 165
injection of 100-400mg/kg pilocarpine triggers seizures starting with the motor, olfactory, and gustatory automatisms, which evolve into status epilepticus (SE). Histological examination of the brains revealed specific alterations throughout the hippocampal formation, amygdala, thalamus, neocortex, olfactory cortex, and substantia nigra165. The most remarkable difference between the pilocarpine and the KA models is the rapidity of the pilocarpine one. Neuronal damage is visible within 3 hours after pilocarpine-induced SE, while in the case of KA, the brain lesion in the same areas is visible 8 hours after SE induction . Regarding the extension of cellular 166
damage, both KA and pilocarpine models, injected systemically, produce an extensive extrahippocampal cell loss . Mortality in the pilocarpine model appears higher, but 167
can be improved with lithium treatment 24 h prior to the injection of the convulsant . 168
Multiple reports indicate a strain difference in the pilocarpine model. For instance, cell damage and mortality rate are more significant in Long-Evans and Wistar rat strains compared to Sprague-Dawley rats . The Sprague-Dawley strain also demonstrates 169
less prominent neuronal damage compared to Wistar rats169. Overall, Long-Evans strain is the most sensitive to pilocarpine, followed by Wistar, while the Sprague-Dawley rat strain exhibits minimal sensitivity. In terms of behavioral performance, spatial memory seems to be affected to a greater extent in the pilocarpine model, although animals show reduced levels of anxiety as compared to the KA-treated rats216. The Dose-response to pilocarpine is similar in mice and rats; however, mice appear to be more sensitive and produce higher mortality rates than rats168. Also, mice treated with pilocarpine are more likely to develop SRS than the KA-treated ones167.
The injection of tetanus toxin into the hippocampus is also used to trigger epileptogenesis. The first mention of this procedure dates as early as the nineteenth century, with the demonstration that intracerebral injection of the toxin causes seizures in experimental animals . Unlike KA, tetanus toxin does not elicit SE after its 170
injection, but efficiently generates SRS within 2-21 days post-injection, which usually cease after a few weeks . A standard dose causes nearly 30% pyramidal cell loss in 171
CA1 in the unilateral hippocampus and 10% at distant sites. Moreover, loss of the dendritic spines in CA3 pyramidal neurons is also reported .172
The next common approach for modelling TLE in rodents includes various types of electrical stimulation. The most widely used model is the kindling model, in which an electrode is implanted into the brain, and a seizure focus is created by repeated electrical stimulation. The term “kindling” was first proposed by G. Goddard and his colleagues , where, after several experimental trials, they demonstrated that daily 173
electrical stimulation leads to the development of an epileptic focus in the brain, creating generalized convulsions and permanent neuropathological changes in brain

tissue . The kindling model has been extensively used targeting various regions such 174
as the hippocampal formation , , the piriform cortex , and the perirhinal cortex . 175 176 177 178
The primary hallmark of the electrical stimulation model is the gradual development of the epileptic activity, as compared to the chemical administration, in which the initial brain insult leads to the development of chronic epilepsy after a latent period. The kindling model is sometimes described as a “functional” model due to the absence of gross morphological damage, which is characteristic of chemical models . A 179
variation based on an optogenetic approach (rather than electric stimulation) called “optokindling” has been introduced recently . Although it was used to produce a 180
model for neocortical epilepsy, the same technique may be used to create a model of TLE.
One of the most common causes of TLE in patients is traumatic brain injury (TBI) . 181
Various TBI models have been developed, in particular, the fluid percussion injury model . A single severe injury produced by fluid percussion injury device is sufficient 182
to trigger epileptogenesis in rodents with some similarity with human TLE, including morphological alterations such as neuronal loss in the hippocampus and mossy fiber sprouting , .183 184
The previous models involve brain insults triggered in adult animals. Insults occurring during development can also lead to TLE later in life. For example, febrile seizures are a risk factor for the development of TLE in humans . Hyperthermia-induced seizure 185
models have been developed . Dubé et al., 2010 demonstrated that prolonged 70 186
min febrile seizures in P11 rat pups could trigger epileptogenesis .187
Neonatal hypoxia can also constitute an initial insult, which may result in TLE later in life . It is linked to the observation that hypoxic encephalopathy is the most common 188
cause of neonatal seizures . Exposing P10-P12 pups to graded global hypoxia 189
(7%-4% oxygen) for 15 minutes triggers epileptogenesis . Morphological changes 190
include mossy fiber sprouting, and electrophysiological analysis shows increased excitability, facilitated long-term potentiation induction, and longer afterdischarges.
A.3. The kainic acid model
Kainic acid, which is a cyclic analog of L-glutamate and an agonist of the ionotropic KA receptors, was first reported to damage hippocampal pyramidal neurons by Nadler et al., 1978 . However, the use of KA as a model for epilepsy was first introduced by 191
Ben-Ari and his colleagues , , who performed a unilateral intra-amygdaloid injection 192 193
of KA in unanaesthetized non-paralyzed rats and observed focal seizures evolving into SE as the dosage increased. Moreover, the histological findings revealed neuronal degeneration and gliosis in the CA3 field of the hippocampus. These, and other experiments, suggested the use of KA as a tool model TLE in rodents. The injection of KA will lead to the activation of its cognate receptors.

A.3.1. Kainic acid receptors
Extensive biomolecular research has provided us with information about the localization of kainic acid receptors (KARs) in the mammalian brain. Experiments on KAR mapping showed that these receptors could be found at different levels of expression throughout the brain, including the entorhinal cortex , cerebellum , 194 195
amygdala , basal ganglia and the hippocampus in which they are particularly 196 197
abundant . Kainic acid receptors belong to a family of ionotropic glutamate 198
receptors, along with AMPA and NMDA receptors. They can be pre- or postsynaptic. Presynaptic KARs act bidirectionally, performing an excitatory action through their ionotropic activity and inhibition via a “non-canonical” metabotropic signalling . 199
Postsynaptic receptors contribute to excitatory neurotransmission . KA receptors 200
can also control GABAergic neurotransmission, both pre- and post-synaptically , . 201 202
There are five known types of KA receptor subunits: GluR5 (GluK1), GluR6 (GluK2), GluR7 (GluK3), KA1 (GluK4) and KA2 (GluK5). Some subunits are highly expressed in the hippocampus. The GluK4 subunit is almost exclusively found in the CA3 hippocampal field, whereas its expression in CA1 is limited , . GluK5 subunits are 203 204
expressed in both CA1 and CA3 fields203, as well as in the cortex and striatum . The 205
fact that these two subunits have a high affinity for kainic acid (dissociation constant of 5-15 nM) and are mainly concentrated in CA3 may explain the pattern of excitotoxic damage found in this region .206
GluK1 are found in the CA3 field of the hippocampus203, and GluK2 are highly expressed in both CA1 and CA3198. Although both subunits have low affinity for KA (KD
of 50-100 nM), GluK1 knock-out mice show increased susceptibility to the epileptogenic effect of KA, while GluK2 ablation prevents the generation of epileptiform discharges . GluK2-KO mice are also less sensitive to the epileptogenic 207
effect of KA . Conversely, overexpression of GluK2 by HSVGluR6 viral vector 208
injection leads to seizure induction and hyperexcitabilty21. Furthermore, Ullal et al., 2005 reported that the GluR7 subunit, which has the lowest affinity to glutamate, is down-regulated by KA-induced seizures in the long term . All these findings suggest 209
that the KA administration will target all types of kainate receptors, with a unique effect in the hippocampus.
A.3.2. Mechanism of action
Excitotoxicity refers to a process in which neurons experience severe damage to the point of cell death due to overstimulation by excitatory neurotransmitters such as glutamate . The mechanism behind this includes a cascade of molecular interactions 210
that lead to osmotic imbalance, excessive depolarization, and, eventually, rupture of the postsynaptic membrane . Several mechanisms are at play. A central one involves 211
the intracellular accumulation of Ca2+, following the excessive activation of glutamate receptors (Fig. A.1.D). The rise in Ca2+ can strongly impact mitochondria and the endoplasmatic reticulum . Elimination of intracellular Ca2+ or blocking its influx into 212
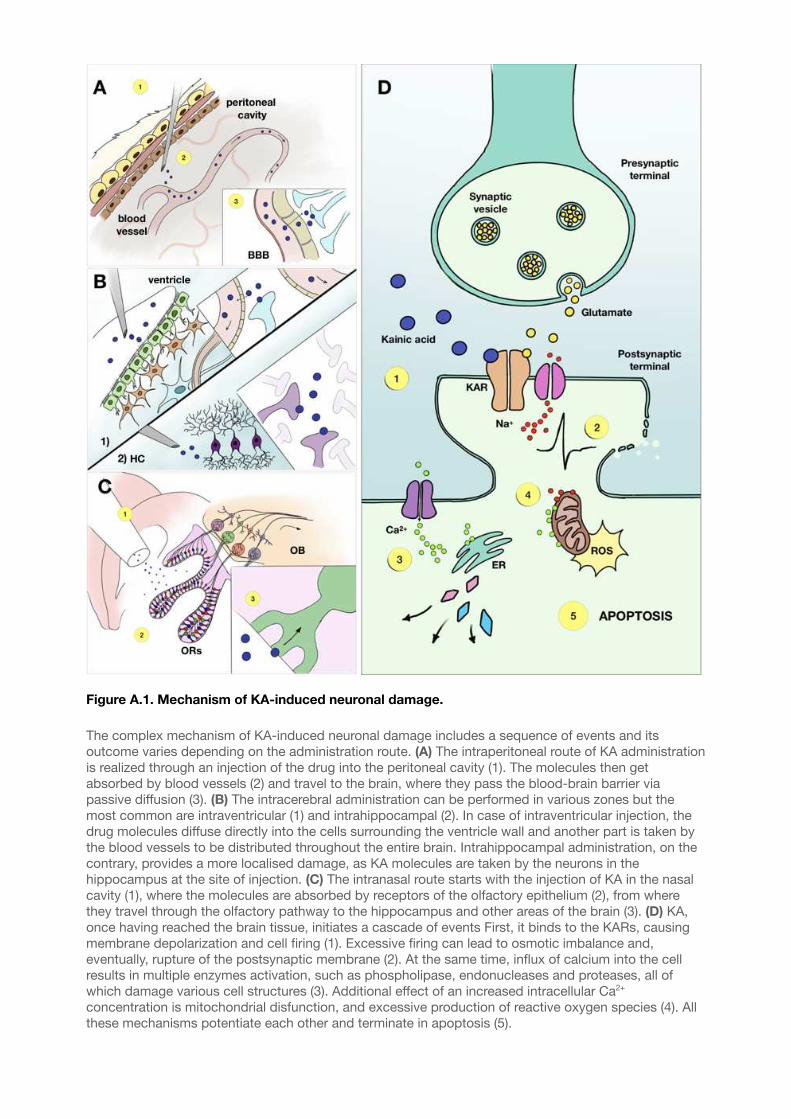
Figure A.1. Mechanism of KA-induced neuronal damage.
The complex mechanism of KA-induced neuronal damage includes a sequence of events and its outcome varies depending on the administration route. (A) The intraperitoneal route of KA administration is realized through an injection of the drug into the peritoneal cavity (1). The molecules then get absorbed by blood vessels (2) and travel to the brain, where they pass the blood-brain barrier via passive diffusion (3). (B) The intracerebral administration can be performed in various zones but the most common are intraventricular (1) and intrahippocampal (2). In case of intraventricular injection, the drug molecules diffuse directly into the cells surrounding the ventricle wall and another part is taken by the blood vessels to be distributed throughout the entire brain. Intrahippocampal administration, on the contrary, provides a more localised damage, as KA molecules are taken by the neurons in the hippocampus at the site of injection. (C) The intranasal route starts with the injection of KA in the nasal cavity (1), where the molecules are absorbed by receptors of the olfactory epithelium (2), from where they travel through the olfactory pathway to the hippocampus and other areas of the brain (3). (D) KA, once having reached the brain tissue, initiates a cascade of events First, it binds to the KARs, causing membrane depolarization and cell firing (1). Excessive firing can lead to osmotic imbalance and, eventually, rupture of the postsynaptic membrane (2). At the same time, influx of calcium into the cell results in multiple enzymes activation, such as phospholipase, endonucleases and proteases, all of which damage various cell structures (3). Additional effect of an increased intracellular Ca2+
concentration is mitochondrial disfunction, and excessive production of reactive oxygen species (4). All these mechanisms potentiate each other and terminate in apoptosis (5).

BBB — blood-brain barrier, ER — endoplasmatic reticulum, HC — hippocampus, KAR — kainic acid receptor, OB — olfactory bulb, ORs — olfactory receptors, ROS — reactive oxygen species
mitochondria can diminish cellular sensitivity to apoptotic stimuli . The Na+ and Cl− 213
ions are also involved since their removal from the extracellular space stops neurodegeneration . Finally, extracellular K+ is also engaged in KA-induced 214
excitotoxicity . 215
Oxidative stress also plays a central in cell death in the context of excitotoxic damage. The excess of glutamate initiates reactive oxygen species (ROS) formation, and this, in turn, leads to mitochondrial dysfunction and molecular damage , . KA injection 216 217
leads to high levels of ROS , , as seen in brain tissue from TLE patients .218 219 220
A.4. Administration routes
Kainic acid, like many other drugs, can be administered in various ways, depending on the desired outcome (Fig. A.1.A-C, Fig. A.2). Each method has its advantages and drawbacks and should be chosen accordingly. The key parameters one should consider selecting the injection route are mortality rate, labor-intensity, lesion control, and, finally, age, sex, and strain of the animal. The major administration routes are systemic, intracerebral (which could be divided into intraventricular, intrahippocampal, suprahippocampal, and intra-amygdaloid), and intranasal. Each of them is described further in this chapter.
A.4.1. Systemic administration
The first experiments using systemic KA administration were performed in the late 1970s to determine whether the toxin could induce damage via axonal connections 221
(Fig. A.1.A). The studies showed that rats injected by 12 mg/kg of KA intraperitoneally experienced the onset of “wet dog shaking” seizures approximately 30-90min after injection, which eventually evolved into secondary generalized tonic-clonic convulsions in 88% of cases. Histological damage was present already at 3h post-injection and gradually increased over a period of 2 weeks. This method has been widely used with different modifications. For instance, it was shown that KA, injected subcutaneously, causes similar behavioral and histological alterations as an intraperitoneal injection and can be used as a model for TLE . The significant 222 223
advantage of systemic KA administration is its low labor-intensity, which allows the injection of numerous animals in a comparatively shorter period of time. Moreover, the absence of a surgical procedure eliminates side effects created by anesthesia, surgery invasiveness, and extra damage made by direct contact with brain tissue during the intracerebral injection. However, the obvious disadvantages of this model are 1) no control over the bioavailability of KA in the brain, and 2) high mortality rates. As it was already mentioned before, Long-Evans and Wistar rat strains have higher mortality168. In mice, C57 and CH3 strains display an increased mortality rate, while 129/SvJ and SvEms strains have a higher ratio of survival . The summarized data for systemic KA 224
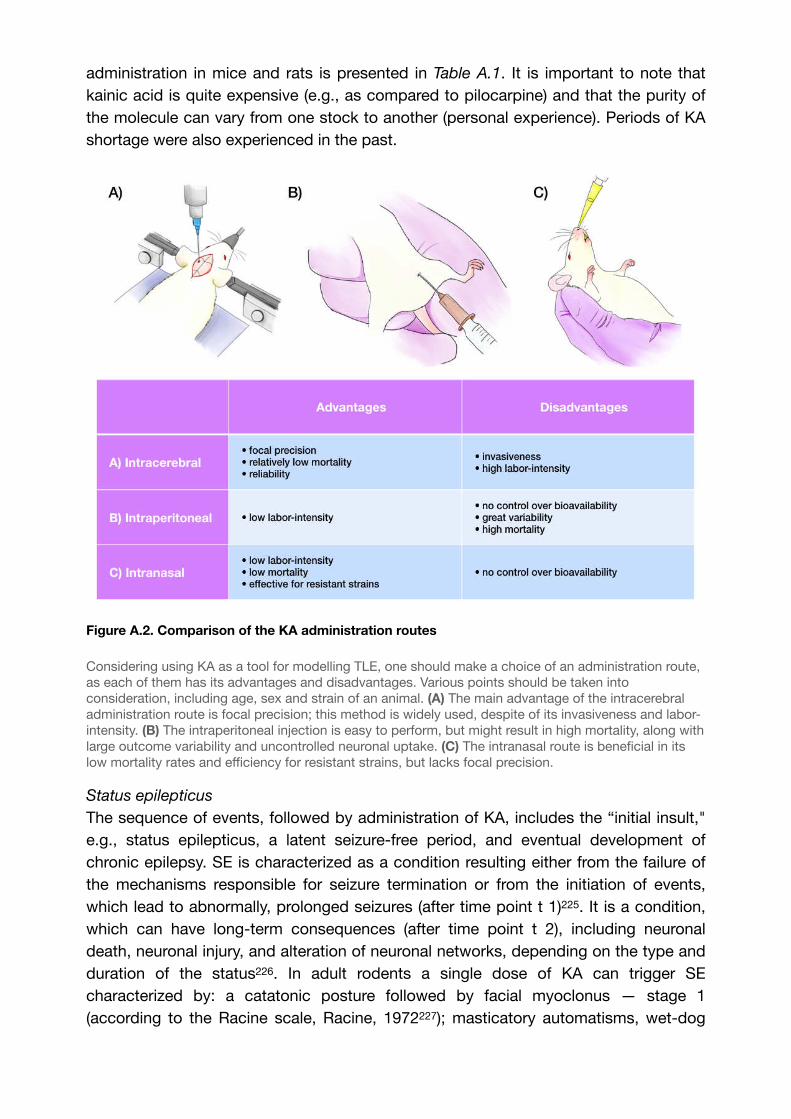
administration in mice and rats is presented in Table A.1. It is important to note that kainic acid is quite expensive (e.g., as compared to pilocarpine) and that the purity of the molecule can vary from one stock to another (personal experience). Periods of KA shortage were also experienced in the past.
Figure A.2. Comparison of the KA administration routes
Considering using KA as a tool for modelling TLE, one should make a choice of an administration route, as each of them has its advantages and disadvantages. Various points should be taken into consideration, including age, sex and strain of an animal. (A) The main advantage of the intracerebral administration route is focal precision; this method is widely used, despite of its invasiveness and labor-intensity. (B) The intraperitoneal injection is easy to perform, but might result in high mortality, along with large outcome variability and uncontrolled neuronal uptake. (C) The intranasal route is beneficial in its low mortality rates and efficiency for resistant strains, but lacks focal precision.
Status epilepticus
The sequence of events, followed by administration of KA, includes the “initial insult," e.g., status epilepticus, a latent seizure-free period, and eventual development of chronic epilepsy. SE is characterized as a condition resulting either from the failure of the mechanisms responsible for seizure termination or from the initiation of events, which lead to abnormally, prolonged seizures (after time point t 1) . It is a condition, 225
which can have long‐term consequences (after time point t 2), including neuronal death, neuronal injury, and alteration of neuronal networks, depending on the type and duration of the status . In adult rodents a single dose of KA can trigger SE 226
characterized by: a catatonic posture followed by facial myoclonus — stage 1 (according to the Racine scale, Racine, 1972 ); masticatory automatisms, wet-dog 227

shakes (characteristic for rats but not for mice) and head nodding (stage 2); rearing with facial automatisms and forelimb clonus (stages 3-4); and finally repeated rearing and falling (stage 5) , . Following systemic KA administration, SE develops within 2 228 229
hours and lasts for approximately 5-9 hours, according to multiple experiments on rats223, , , , , , . In mice, however, behavioral manifestations appear within 230 231 232 233 234 235
15-30 min post-injection , , and the duration of SE does not exceed 6 hours224, . 236 237 238
Mortality in both cases is relatively high (22% and 27%, respectively), which can be reduced with slight alterations of the protocol. Repeated injections of 5 mg/kg i/p or 239
2.5 mg/kg i/p at 30 min intervals reduce mortality down to approximately 5-6%, as 240
compared to a single full-dose injection. Likewise, to increase survival rate, SE can be terminated after a few hours by subcutaneous injection of 10mg/kg diazepam or zolazepam , . Many groups try to stop SE after 20 min. Perfect control of SE 241 242
duration requires instrumenting the animals for EEG recordings. We observed that SE starts at the electrophysiological level earlier (5 min or more) before the behavioral manifestation (unpublished). We also found a decreased mortality and accelerated recovery when timing SE with electrophysiological recordings. The protocol we are using is to instrument animals with a telemetry device for supradural recordings at least 2 weeks before KA injection. 2-3 days before KA injection, we start EEG recordings in the home cage, including following KA injection.
Table A.1. Systemic administration of KA in rodents.!
There are several characteristic features of systemic Ka administration in mice and rats. While a rat model is highly reliable in inducing SRS, in mice it poses certain challenges. The hippocampal damage is more severe in rats, while for mice cortex is affected at a greater extent.
MF — mossy fiber; SE — status epilepticus; SRS — spontaneous recurrent seizures.
Latent period and spontaneous recurrent seizures
The latent period is the time between the initial brain insult (SE in the case of the KA rodent model of TLE), and the development of chronic epilepsy manifested as the first
SE Latent period
SRS Histology Comorbidities Reference
Rats
Starts within 2hLasts for 5-9 h22% mortality
12-36 days 8 per day (± 5.4)
Bilateral damage Whole hippocampusSubiculumEntorhinal cortexMF sprouting
Cognitive impairmentAggressiveness
Stafstrom et al., 1992Lado et al., 2006Williams et al., 2009Drexel et al., 2012Bertoglio et al., 2017
Mice
Starts within 15-30 minLasts for 2-6 h27% mortality
2-3 days Rarely observedInfrequent
Bilateral damageCA3/CA1 areas of the hippocampusCortical areasMF sprouting
HyperactivityDelayed mortality
Hu et al., 1998McKhann et al., 2003Benkovic et al., 2004Puttachary et al., 2015Umpierre et al., 2016

spontaneous seizures. The latent phase is considered one of the hallmarks of human epilepsy, and it can last for many years185, . In the rat model, however, the usual 243
duration of the seizure-free period is approximately 5-30 days234,235, , although in 244
some cases it may take up to 5 months . In mice, the latent phase is significantly 245
shorter and could be estimated as 2-3 days238. During this phase, experimental animals do not exhibit any behavioral seizures, but the electrophysiological study may show some epileptic activity, e.g., non-convulsive seizures, which will be described in the next chapter. Whether a “true” latent period exists is debated, as the EEGs display evident anomalies in the few hours following SE, in particular interictal-like spikes.
The appearance of SRS can be considered as the onset of the chronic phase of epilepsy. The average daily frequency of seizures is 8 per day (±5.4) 223,230,231,232,233,234. SRS developing following KA injection in rats at different ages lasted for approximately 40 s and resembled stage 5 limbic seizures induced by electric kindling: bilateral forelimb clonus, masticatory movements, rearing and falling230. Subsequent studies demonstrated similar results, with the seizure ratio varying from 4 to 21 per day232,233. For mice, however, it is much harder to develop SRS with a systemic administration, and those detected are infrequent and highly variable224, . The F344 rat strain has 246
been demonstrated to be sensitive to KA with a much lower dose (10.5 mg/kg) needed to induce SRS . Prominent neurodegeneration in the hippocampus, extensive mossy 247
fiber sprouting, and the consistent ratio of SRS, with the same frequency over time, makes this strain interesting to obtain a stable and reproducible model. It is important to note that the high rate of SRSs may be due to the fact that animals are singly housed, which increases seizure frequency by a factor of 16, at least in the pilocarpine model , . Single housing adds stress as an independent variable, and a 248 249
confounding factor, to the epilepsy phenotype. We recommend maintaining social interaction248,249. This comment applies to all models.
Histological evaluation
Systemic administration of KA induces extensive bilateral neuronal damage throughout the brain. The first noticeable changes appear within 48 hours post-treatment and are present primarily in CA1, CA3, and CA4 hippocampal subregions20,222,235, , , . 250 251 252
Subsequently, the entire hippocampus is affected in the rat brain , . In mice, the 253 254
predominantly damaged areas are still CA3 and CA1224,236. The typical pattern of KA-induced damage includes necrosis of pyramidal cells, gliosis, and mossy fiber sprouting within the inner molecular layer of the dentate gyrus224, . Additionally, there 255
have been multiple reports indicating damage of various extrahippocampal areas, such as the amygdala , subiculum, entorhinal cortex, thalamus, caudoputamen235, 256
substantia nigra, hypothalamus166, olfactory bulb, anterior olfactory nucleus and 257
other areas. T2-weighted MRI images of Wistar and Sprague-Dawley rat strains, systemically treated with KA, showed an interesting phenomenon, contradicting previous findings on mortality. The extent of neuronal damage is higher in the Sprague-Dawley strain, while in Wistar rats, the relative volume of the hippocampus is not different from the control animals169. Confirming the MRI data, post-mortem NeuN-

staining revealed a significant pyramidal loss in CA1 and CA3 areas of the hippocampus in Sprague-Dawley rats but not in Wistar.
A.4.2. Intraventricular injection
Intracerebral KA administration manifested as an intracerebroventricular (icv) route. First of all, Nadler et al., 1978191 have demonstrated that the injection of 0.5 nmol KA intraventricularly into the rat brain leads to CA3 neurodegeneration within 1-3 days after treatment, whereas higher doses (0.8 mg and more) cause damage to both the CA1 and CA2. Similar findings were later observed by Lancaster & Wheal, 1982 , 258
who confirmed the same pattern of neuronal damage and helped to establish the intraventricular injection paradigm, which is still sometimes used as a method of choice , . The data of icv KA administration in rats is presented in Table A.2.259 260
Table A.2. Intracerebroventricular administration of KA in rats.
MF — mossy fiber; SE — status epilepticus; SRS — spontaneous recurrent seizures;
Status epilepticus
The induction of SE using the icv route of KA administration is consistent with other methods. It starts within 10-30 min post-injection and manifests as bradypnea, circling behavior, and wet-dog shakes and subsequently progresses into continuous motor seizures . The duration of SE varies between 2 and 6 h and is sometimes interrupted 261
by a diazepam injection to reduce the mortality rate260,261, . Typically, it does not 262
exceed 10% and is considered relatively low in comparison with the systemic route of administration261, .263
Latent period and spontaneous recurrent seizures
There are multiple reports regarding SRS development following icv KA administration. Having performed a 24/7 continuous video/EEG-recording, few authors reported similar findings, with the latent period duration varying between 1 and 2 weeks and the SRS appearing consistently several times per day260,263. This data suggests that the icv administration of KA serves as an efficient tool for chronic epilepsy modelling.
Histological evaluation
Morphological damage induced by an icv KA administration might seem paradoxical; despite the fact that the toxin is injected in the ventricular system and thus potentially distributes throughout the brain, the actual lesion appears to be restricted to CA3/CA4
SE Latent period
SRS Histology Comorbidities Reference
Starts within 10-30 minLasts for 2-6h<10% mortality
7-14 days 5 per day (± 3.5)
Unilateral lesionCA3/CA4 areas of the hippocampusMF sprouting
Cognitive impairment
Miyamoto et al., 1997Jing et al., 2009Gordon et al., 2014Song et al., 2016Gao et al., 2019

hippocampal subfields ipsilaterally to the injection site191,261. The contralateral hippocampus and the extrahippocampal areas are relatively spared, and in some cases, almost entirely intact260. Mossy fiber sprouting is also present in most cases260,262,263. The pathology is mostly unilateral, with occasional signs of the damage on the contralateral side258,261. The model is closer to what is found in patients with mesial TLE, as the epileptogenic network is usually limited to one hemisphere.
A.4.3. Intrahippocampal and intra-amygdaloid injections
The first experiment with intra-amygdaloid KA administration in 1978 demonstrated characteristic behavioral, electrophysiological, and neuropathological epileptic hallmarks in baboons192,193. Other studies used intrahippocampal KA injections in rats to observe the same outcome . Both methods of intracerebral administration are still 264
widely used. The principal advantage is a direct delivery into the brain tissue, bypassing the blood-brain barrier. The idea is to produce focal damage. The mortality rate appears to be lower as compared to systemic administration14. Moreover, the standardized protocol for the intrahippocampal KA administration, combined with the electrode implantation, which has been proposed recently , enables a good 265
reproducibility and allows the combination of both electrophysiological and behavioral approaches. A brief summary of both methods is presented in tables A.3 and A.4.
Table A.3. Intrahippocampal administration of KA in rodents.
Intrahippocampal KA administration produces similar results in mice and rats, with low mortality and stable SRS generation. While it is not always possible to observe behavioral seizures in a mouse model, electrographic SRS are present in most cases.
DG — dentate gyrus; eSRS — electrographic spontaneous recurrent seizures; MF — mossy fiber; SE — status epilepticus; SRS — spontaneous recurrent seizures;
Status epilepticus
SE Latent period
SRS Histology Comorbidities Reference
Rats
Starts within 5-20 minLasts for3-15 h<5% mortality
13-30 days 8 per day (± 6.3)
Unilateral lesionCA3/CA4 areas of the hippocampusDG granule cells dispersion MF sprouting
Cognitive impairmentPerseverative behaviorHyperexcitablty
Cavalheiro et al., 1982Bragin et al., 2005Arkhipov et al., 2008Rattka et al., 2013Klee et al., 2017
Mice
Starts within 30 minLasts for 3-12 h<5% mortality
2-14 days 1-2 per weekHighly variableeSRS +++
Unilateral lesionCA3/CA1 areas of the hippocampusDG granule cells dispersionMF sprouting
Cognitive impairmentHyperexcitabiityDepression (lesioned ventral hippocampus)
Bouilleret et al., 1999Riban et al., 2002Gouder et al., 2003Gröticke et al., 2008Lee et al., 2012Zeidler et al., 2018
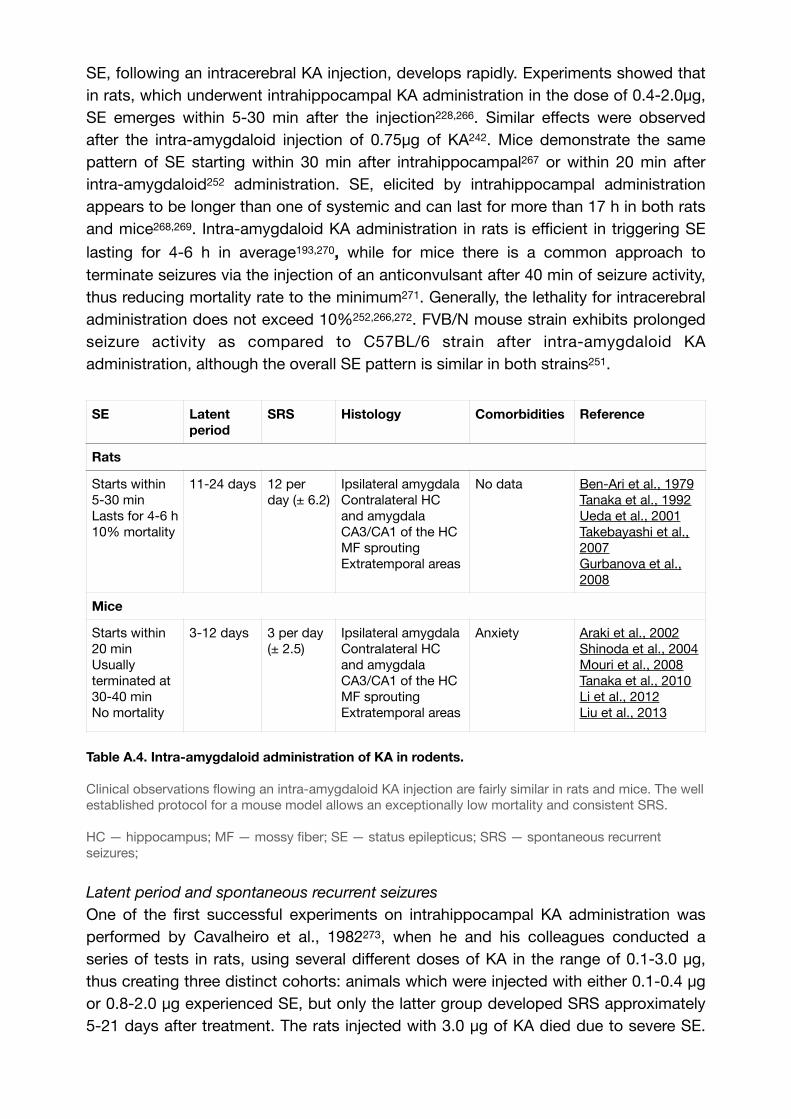
SE, following an intracerebral KA injection, develops rapidly. Experiments showed that in rats, which underwent intrahippocampal KA administration in the dose of 0.4-2.0μg, SE emerges within 5-30 min after the injection228, . Similar effects were observed 266
after the intra-amygdaloid injection of 0.75μg of KA242. Mice demonstrate the same pattern of SE starting within 30 min after intrahippocampal or within 20 min after 267
intra-amygdaloid252 administration. SE, elicited by intrahippocampal administration appears to be longer than one of systemic and can last for more than 17 h in both rats and mice , . Intra-amygdaloid KA administration in rats is efficient in triggering SE 268 269
lasting for 4-6 h in average193, , while for mice there is a common approach to 270
terminate seizures via the injection of an anticonvulsant after 40 min of seizure activity, thus reducing mortality rate to the minimum . Generally, the lethality for intracerebral 271
administration does not exceed 10%252,266, . FVB/N mouse strain exhibits prolonged 272
seizure activity as compared to C57BL/6 strain after intra-amygdaloid KA administration, although the overall SE pattern is similar in both strains251.
Table A.4. Intra-amygdaloid administration of KA in rodents.
Clinical observations flowing an intra-amygdaloid KA injection are fairly similar in rats and mice. The well established protocol for a mouse model allows an exceptionally low mortality and consistent SRS.
HC — hippocampus; MF — mossy fiber; SE — status epilepticus; SRS — spontaneous recurrent seizures;
Latent period and spontaneous recurrent seizures
One of the first successful experiments on intrahippocampal KA administration was performed by Cavalheiro et al., 1982 , when he and his colleagues conducted a 273
series of tests in rats, using several different doses of KA in the range of 0.1-3.0 μg, thus creating three distinct cohorts: animals which were injected with either 0.1-0.4 μg or 0.8-2.0 μg experienced SE, but only the latter group developed SRS approximately 5-21 days after treatment. The rats injected with 3.0 μg of KA died due to severe SE.
SE Latent period
SRS Histology Comorbidities Reference
Rats
Starts within 5-30 minLasts for 4-6 h10% mortality
11-24 days 12 per day (± 6.2)
Ipsilateral amygdala Contralateral HC and amygdala CA3/CA1 of the HCMF sprouting Extratemporal areas
No data Ben-Ari et al., 1979Tanaka et al., 1992Ueda et al., 2001Takebayashi et al., 2007Gurbanova et al., 2008
Mice
Starts within 20 minUsually terminated at 30-40 minNo mortality
3-12 days 3 per day (± 2.5)
Ipsilateral amygdalaContralateral HC and amygdalaCA3/CA1 of the HCMF sproutingExtratemporal areas
Anxiety Araki et al., 2002Shinoda et al., 2004Mouri et al., 2008Tanaka et al., 2010Li et al., 2012Liu et al., 2013
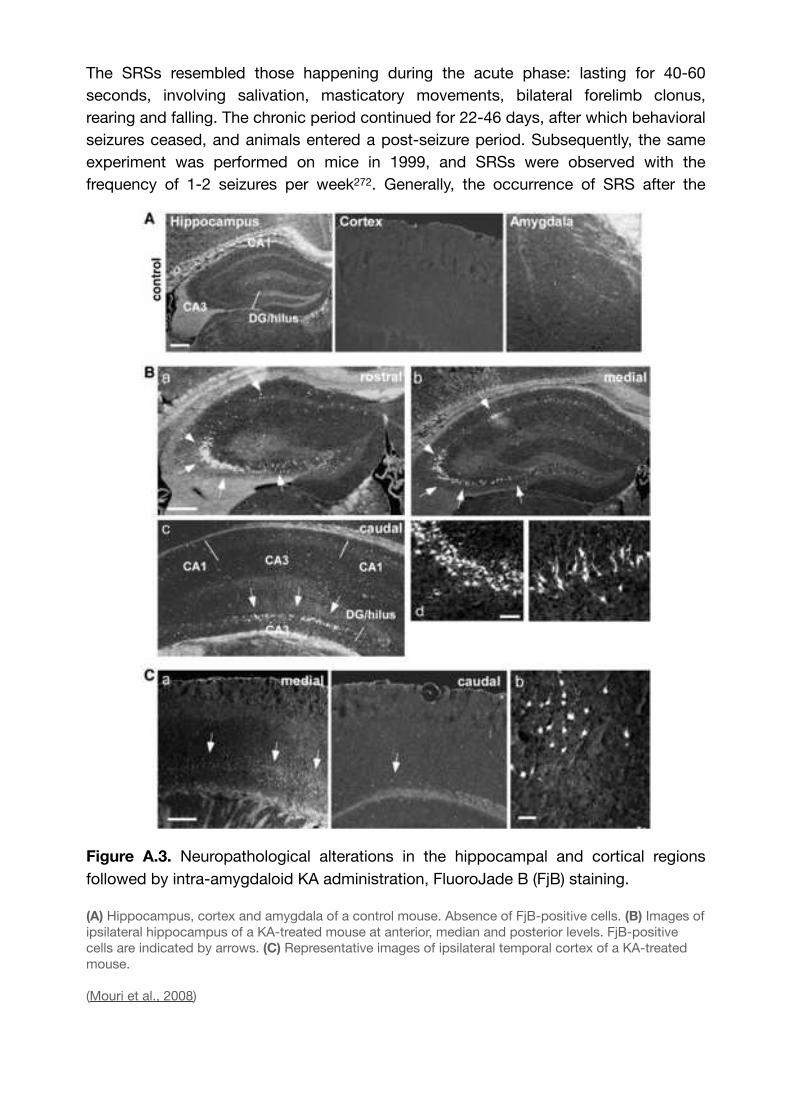
The SRSs resembled those happening during the acute phase: lasting for 40-60 seconds, involving salivation, masticatory movements, bilateral forelimb clonus, rearing and falling. The chronic period continued for 22-46 days, after which behavioral seizures ceased, and animals entered a post-seizure period. Subsequently, the same experiment was performed on mice in 1999, and SRSs were observed with the frequency of 1-2 seizures per week272. Generally, the occurrence of SRS after the
Figure A.3. Neuropathological alterations in the hippocampal and cortical regions followed by intra-amygdaloid KA administration, FluoroJade B (FjB) staining.
(A) Hippocampus, cortex and amygdala of a control mouse. Absence of FjB-positive cells. (B) Images of ipsilateral hippocampus of a KA-treated mouse at anterior, median and posterior levels. FjB-positive cells are indicated by arrows. (C) Representative images of ipsilateral temporal cortex of a KA-treated mouse.
(Mouri et al., 2008)

intrahippocampal administration of KA is consistent in rats and resembles the pattern of systemic injection266. In mice, the results are highly variable, with some authors reporting the frequency of electrographic SRSs to reach dozens per hour , , which 274 275
could be due to social isolation-induced stress. According to a recent study, this phenomenon of frequent electrographic seizures is specific to mice but not to rats . 276
Intra-amygdaloid administration is efficient for inducing chronic epilepsy in both rats and mice. If for the former the average amount of SRS is 12 per day242, for the latter the ratio is 1-5 per day252, , , . Latency to the first SRS is reported to be 1-4 277 278 279
weeks in rats273,276 and no longer than 2 weeks in mice252,267 in both intrahippocampal and intra-amygdaloid administration.
Histological evaluation
histological studies reveal similarities between the two routes of intracerebral KA administration. The intrahippocampal injection creates a focal lesion in the hippocampus, damaging predominantly the ipsilateral CA3 subfield and, to some extent, CA4 and CA1, regardless of the site of injection228,272, , . Interestingly, intra-280 281
amygdaloid administration induces massive neuron destruction in the same hippocampal regions along with a lesion in the amygdala270, . Some authors reported 282
extensive damage on the contralateral side252,282, and the resistance of the CA2 283
hippocampal subfield to the KA-induced toxicity271. Mossy fiber sprouting was also observed in both rats and mice242,252,275, . Granule cell dispersion in the dentate 284
gyrus, however, is only reported with the intrahippocampal administration265,266. Additionally, there is multiple evidence of damage in extra-temporal regions induced by intra-amygdaloid injection, such as thalamus283, entorhinal, and perirhinal cortices252,277. There is a significant strain difference in mice. C57BL/6 mouse strain is resistant to cellular damage induced by KA251, . In contrast, the FVB/N strain shows 285
increased sensitivity and extensive neurodegeneration in the CA1 and CA3 areas of the hippocampus following intra-amygdaloid KA administration251. A representative image of the FluoroJade B (FjB) staining of murine hippocampus and cortex after intra-amygdaloid KA injection, is presented in Figure A.3.
A.4.4. Suprahippocampal
One of the variations of the intrahippocampal administration route is the so-called suprahippocampal injection, where KA is being injected in the cortical area just above the hippocampus. This technique was introduced by Bedner et al., 2015 , who 286
performed an intracortical injection in order to reproduce the features of the intrahippocampal administration but also to avoid damaging the CA1 subfield of the hippocampus. The experiment was successful, and all the animals experienced SE, followed by SRS development. Subsequently, this method was tested several times by other researchers, proving to be efficient in mice (Table A.5).

Table A.5. Suprahippocampal administration of KA in mice.
DG — dentate gyrus; HC — hippocampus; SE — status epilepticus; SRS — spontaneous recurrent seizures;
Status epilepticus
Starting soon after recovery from the anesthesia, SE induced by the intracortical KA application lasts for up to 12 hours and consists of forelimb clonus and repetitive rearing and falling . The reported mortality is relatively low and ranges from 0% to 287
9.7% . As mentioned by the authors, death occurred mostly due to surgical 288
complications and not seizures286.
Latent period and spontaneous recurrent seizures
SRSs in this model appear after a brief latent period of 4-13 days and are consistent and frequent. Bedner et al., 2015286 reported a frequency of 7.5 ± 6.2 seizures per day 7-9 months post-SE and progressive nature of chronic epilepsy development, consistent with previous data on KA-induced seizure progression280. Another study demonstrated success inducing SRSs in 100% of treated animals with the seizure frequency of 1-1.5 per day288.
Histological evaluation
The lesion created by suprahippocampal injection of KA is unilateral and predominantly situated in the CA1/CA3 hippocampal subfields. Three months post-treatment, there is a complete neuronal loss in CA1, significant damage in CA3 and massive dentate granule cell dispersion286,287,288. Mossy fiber sprouting is not characteristic for this model.
A.4.5. Intranasal injection
The idea of delivering KA via nasal epithelium absorption first emerged as an attempt to trigger SE in the most common strain of transgenic laboratory mice, C57BL/6, which is known for its resistance to KA-induced neuronal death285. The first experiments showed that KA, dissolved in water and delivered intranasally in the dose of 40-60 mg/kg, causes focal seizures, which then generalize into SE, in 100% of treated C57BL/6 mice . Behavioral and histological findings were consistent with the 289
previous results, obtained via standard drug delivery protocols. Likewise, another study demonstrated that a dose of 30mg/kg KA intranasally is associated with low mortality and >90% success in developing SE . Even though the mechanism is not 290
SE Latent period
SRS Histology Comorbidities Reference
Lasts for 2-7 h<10% mortality
4-13 days 2.8 per
day (± 3.5)
Unilateral lesionCA1/CA3 of the HCDG granule cells dispersion
No data Bedner et al., 2015Jefferys et al., 2016Pitsch et al., 2019

fully understood, it is assumed that the drug is absorbed by olfactory epithelium and reaches the hippocampus (as well as other brain areas) via olfactory pathways 291
(Fig.A.1C), as the olfactory bulb has widespread connections with different regions of the brain289. Although this method has not been studied thoroughly and so far has not found widespread success in the neuroscience community, it has its own advantages, such as reproducibility, low labor-intensity and low mortality (Table A.6).
Table A.6. Intranasal administration of KA in mice.
SE — status epilepticus; SRS — spontaneous recurrent seizures;
Status epilepticus
The induction of SE appears to be consistent in all existing studies; it starts within 15-30 min after administration289, with the first symptoms being immobility and 292
staring, followed by generalized tonic-clonic seizures, lasting for 1-5 h289. The mortality rate is relatively low and can be compared to intrahippocampal routes, ranging from
0% to 12%289,290,292, , . Generally, there are no specific SE features associated with 293 294
the intranasal route of administration compared to the conventional methods.
Latent period and spontaneous recurrent seizures
Seizure progression was reported by at least one research group; Sabilallah et al.,
2016 reported latency of 15-30 days before electrographic SRSs are observed in most animals in the form of spontaneous spike activity and occasional seizures290. However, the study lacked continuous 24/7 video/EEG-recording, so the actual outcome might
slightly differ. Other authors reported increased locomotor activity289,293,294, perhaps in relationship with the development of chronic epilepsy.
Histological evaluation
Morphological damage induced by intranasal KA administration has been studied. It has been demonstrated that the brain remains mostly intact, with the exception of the
hippocampus and the olfactory bulbs289,293. The CA3 area of the hippocampus shows
the most prominent damage292,293. Mossy fiber sprouting and granule cell dispersion have not been reported. There is evidence of massive astrogliosis and microglial
response in the hippocampus289,290,292,293,294. In general, there is evidence for characteristic pathological features of “classic” KA-induced epilepsy. However, the damage seems to be localized and restricted to the CA3 area of the hippocampus.
SE Latent period
SRS Histology Comorbidities Reference
Starts within 15-30 minLasts for 1-5h~ 6.5% mortality
15-30 days Reported Bilateral damageCA3 area of the hippocampusOlfactory bulbs
Increased locomotion
Chen et al., 2002Duan et al., 2006Zhang et al., 2008Lu et al., 2008Sabilallah et al., 2016
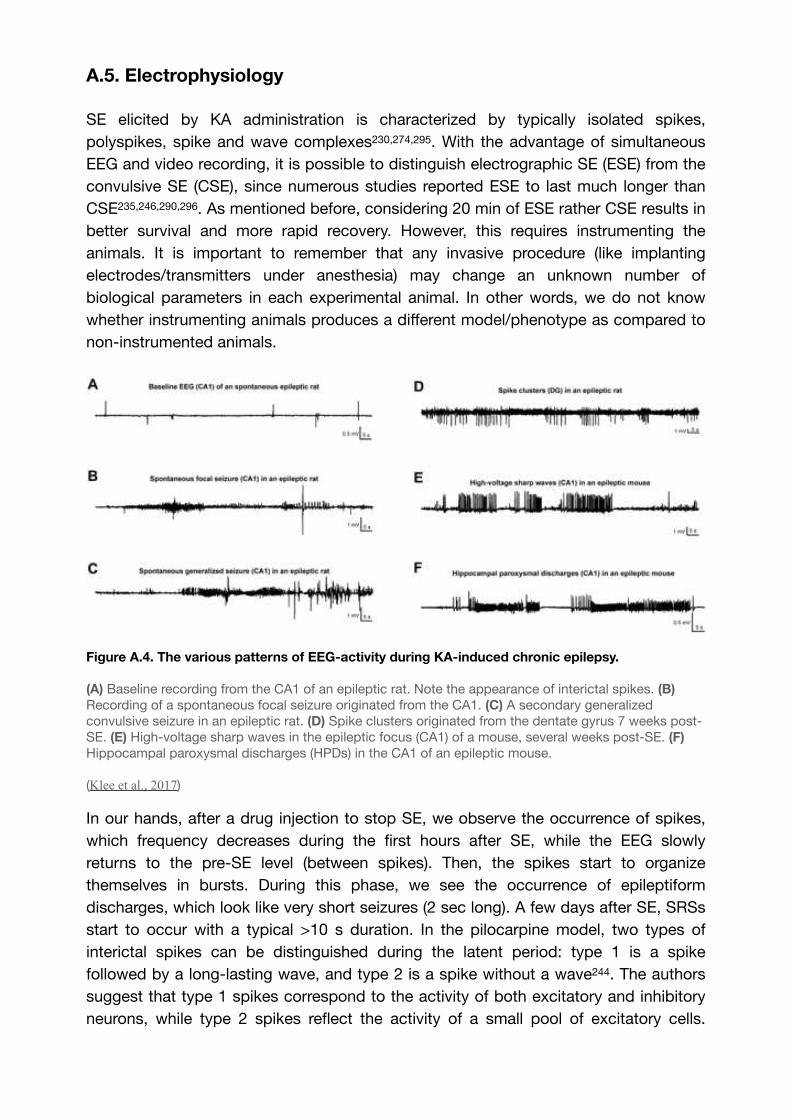
A.5. Electrophysiology
SE elicited by KA administration is characterized by typically isolated spikes, polyspikes, spike and wave complexes230,274, . With the advantage of simultaneous 295
EEG and video recording, it is possible to distinguish electrographic SE (ESE) from the convulsive SE (CSE), since numerous studies reported ESE to last much longer than CSE235,246,290, . As mentioned before, considering 20 min of ESE rather CSE results in 296
better survival and more rapid recovery. However, this requires instrumenting the animals. It is important to remember that any invasive procedure (like implanting electrodes/transmitters under anesthesia) may change an unknown number of biological parameters in each experimental animal. In other words, we do not know whether instrumenting animals produces a different model/phenotype as compared to non-instrumented animals.
Figure A.4. The various patterns of EEG-activity during KA-induced chronic epilepsy.
(A) Baseline recording from the CA1 of an epileptic rat. Note the appearance of interictal spikes. (B) Recording of a spontaneous focal seizure originated from the CA1. (C) A secondary generalized convulsive seizure in an epileptic rat. (D) Spike clusters originated from the dentate gyrus 7 weeks post-SE. (E) High-voltage sharp waves in the epileptic focus (CA1) of a mouse, several weeks post-SE. (F) Hippocampal paroxysmal discharges (HPDs) in the CA1 of an epileptic mouse.
(Klee et al., 2017)
In our hands, after a drug injection to stop SE, we observe the occurrence of spikes, which frequency decreases during the first hours after SE, while the EEG slowly returns to the pre-SE level (between spikes). Then, the spikes start to organize themselves in bursts. During this phase, we see the occurrence of epileptiform discharges, which look like very short seizures (2 sec long). A few days after SE, SRSs start to occur with a typical >10 s duration. In the pilocarpine model, two types of interictal spikes can be distinguished during the latent period: type 1 is a spike followed by a long-lasting wave, and type 2 is a spike without a wave244. The authors suggest that type 1 spikes correspond to the activity of both excitatory and inhibitory neurons, while type 2 spikes reflect the activity of a small pool of excitatory cells.
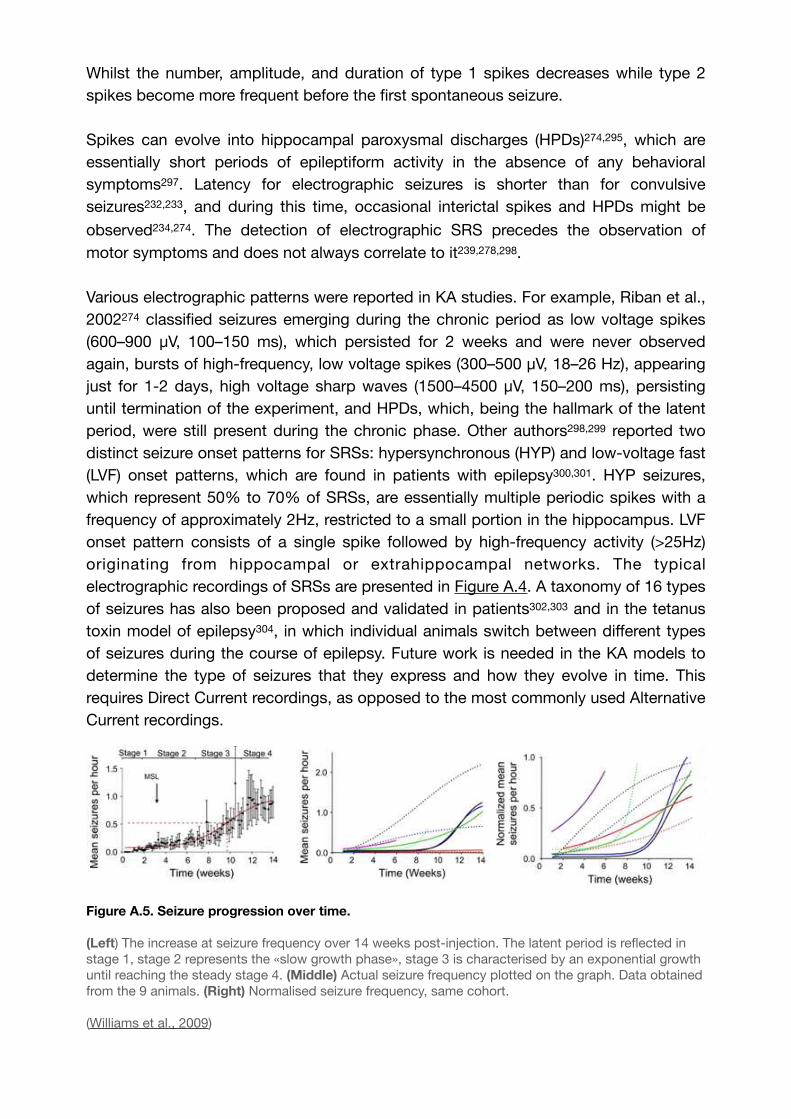
Whilst the number, amplitude, and duration of type 1 spikes decreases while type 2 spikes become more frequent before the first spontaneous seizure.
Spikes can evolve into hippocampal paroxysmal discharges (HPDs)274,295, which are essentially short periods of epileptiform activity in the absence of any behavioral symptoms . Latency for electrographic seizures is shorter than for convulsive 297
seizures232,233, and during this time, occasional interictal spikes and HPDs might be
observed234,274. The detection of electrographic SRS precedes the observation of motor symptoms and does not always correlate to it239,278, . 298
Various electrographic patterns were reported in KA studies. For example, Riban et al., 2002274 classified seizures emerging during the chronic period as low voltage spikes (600–900 μV, 100–150 ms), which persisted for 2 weeks and were never observed again, bursts of high-frequency, low voltage spikes (300–500 μV, 18–26 Hz), appearing just for 1-2 days, high voltage sharp waves (1500–4500 μV, 150–200 ms), persisting until termination of the experiment, and HPDs, which, being the hallmark of the latent period, were still present during the chronic phase. Other authors298, reported two 299
distinct seizure onset patterns for SRSs: hypersynchronous (HYP) and low-voltage fast (LVF) onset patterns, which are found in patients with epilepsy , . HYP seizures, 300 301
which represent 50% to 70% of SRSs, are essentially multiple periodic spikes with a frequency of approximately 2Hz, restricted to a small portion in the hippocampus. LVF onset pattern consists of a single spike followed by high-frequency activity (>25Hz) originating from hippocampal or extrahippocampal networks. The typical electrographic recordings of SRSs are presented in Figure A.4. A taxonomy of 16 types of seizures has also been proposed and validated in patients , and in the tetanus 302 303
toxin model of epilepsy , in which individual animals switch between different types 304
of seizures during the course of epilepsy. Future work is needed in the KA models to determine the type of seizures that they express and how they evolve in time. This requires Direct Current recordings, as opposed to the most commonly used Alternative Current recordings.
Figure A.5. Seizure progression over time.
(Left) The increase at seizure frequency over 14 weeks post-injection. The latent period is reflected in stage 1, stage 2 represents the «slow growth phase», stage 3 is characterised by an exponential growth until reaching the steady stage 4. (Middle) Actual seizure frequency plotted on the graph. Data obtained from the 9 animals. (Right) Normalised seizure frequency, same cohort.
(Williams et al., 2009)

The arguments developed previously clearly show complex dynamical phenomena that occur during the latent period. As soon as SRSs start to occur, it is essential to remember that a steady state is never reached. Seizures tend to progress over time. Williams et al., 2009233 proposed that the evolution of seizures consists of 4 stages, where stage 1 represents a latent period, stage 2 is characterized by a slow increase in seizure frequency, e.g., “slow growth phase," stage 3 is marked as the “exponential growth phase” and stage 4 is a final steady-state plateau phase, which, however, was not observed in all the animals (Fig.A.5). This result agrees with other findings, most of which described a progressive increase of SRSs during the chronic period of KA-induced epilepsy234,266, , . Thus, the evolution of seizures can perhaps be best 305 306
represented by a sigmoid Boltzmann function, showing the exponential growth phase. Recent works depict a more complex picture as circadian and multidien rhythms need to be considered.
Human studies clearly show that seizure activity follows various rhythms: circadian, ultradian, multidien or multi-day, seasonal etc. , , , , . The circadian rhythmicity 307 308 309 310 311
has been recognized for millennia in humans, but multidien rhythms have been identified recently in patients311. The authors found that interictal activity, in addition to its circadian rhythmicity, has a slow envelope of several days period, which is patient-specific. Interestingly, seizures tend to occur during the rising phase of this low rhythm. In addition, clusters of seizures are also found during the rising phase. Similar features have been found in the KA model with a robust periodicity of 5-7 days, each animal following its own cycle . Although rendering data analysis and interpretation more 312
complex, it is important to take circadian and multidien cycles into account.
Finally, as mentioned above, the way animals are housed appears a key determinant of SRS frequency. When animals are singly housed, they are highly stressed, and their seizure frequency is 16 times greater than in animals maintaining social interaction 248,249. Solutions have been developed to maintain social interaction when animals are instrumented248,249.
A.6. Age, sex and strain specificity
The outcome of KA administration depends not only upon the used species and strains but also on the sex and age of the animal. This diversity is an advantage as it may allow us to reproduce the diversity of phenotypes found in patients with TLE. For instance, C57BL/6 mice, the most widely used transgenic strain, are resistant to KA-induced damage, which can be, however, compromised by intranasal drug administration. Other strains with known resistance are BALB/c285, C3HeB/FeJ, 129/SvEms, 129/SvJ224, ICR and FVB/N . It has also been proposed to differentiate 313
between behavioral resistance (129/SvEms), histological resistance (C57, C3H), and combined resistance (129/SvJ)224. In contrast, other mouse strains have increased sensitivity to KA, such as C57BL/10, DBA/2J, and F1 C57BL/6*CBA/J313. In addition,

DBA/2J and FVB/N mouse strains exhibit high seizure-induced mortality as compared to C57BL/6J , .314 315
Another important factor one should consider when working with a rodent model of TLE induced by KA administration is the age of the animals. The first indications of different seizure susceptibility in younger and older rats appeared in the late 1980s when it was demonstrated that an i/p injection of KA in P12 rat pups causes more severe SE as compared to P27 adult rats . Subsequently, other authors reported 316
similar results: shorter latency before developing SE in younger (P5-10) rats compared to P20-60230, and higher mortality in P15 rats as compared to P53 after an i/p KA injection .317
Aged rodents are more prone to seizure activity than adults , and that this 318 319
tendency does not depend on a strain319. Aged rats have a shorter latency to SE onset and exacerbation of pre-seizure behavioral manifestations as compared to middle-aged animals . The reasons behind this are debated and include multiple theories 320
including age-related changes in synaptic connectivity , electrotonic coupling , 321 322
number and type of neurons and diminished density of glutamate receptors in the 323
aged brain , . Overall, this data seems to describe a parabolic pattern of KA-324 325
induced seizure susceptibility, where rodents of young and old age are more vulnerable than middle-aged animals. This correlates well with findings in human epilepsy, where age-specific incidence peaks in childhood and then, after a decline in middle age, raises again at the age of 60 years and older .326
It is remarkable to notice that the vast majority of epilepsy studies have been and are being performed in male rodents. We do not know whether the wealth of data obtained in males applies to females. This key issue remains to addressed. Differences are expected, if only when considering structural arguments. The rodent hippocampus, particularly the CA3 area, contains various receptors for gonadal steroids. For instance, the concentration of androgen receptors in stratum lucidum of the CA3 subfield appears to be higher than in any other hippocampal region , and mossy 327
fibers express ERα and ERβ estrogen receptors . A number of studies have shown 328
that mossy fiber pathway stimulation evokes different levels of BDNF protein expression response in males, females in various stages of oestrus cycle and ovariectomized female rats , , . Only females in proestrus and estrous stages of 329 330 331
menstrual cycles exhibit epileptiform activity after a 10 s 1 Hz train of paired pulses and exhibit a strong BDNF expression330,331. Furthermore, in C57BL/6 mice, KA injected at 20mg/kg or 30mg/kg causes SE in ~100% of aged female mice, as compared to the young females and males of both groups, as well as the highest BDNF expression293. Altogether, this data provides a valuable insight into the influence of sex on KA administration and susceptibility to seizures, showing that the oestrus cycle should be taken into consideration while modeling epilepsy in female mice. However, a tremendous amount of research effort is required to study epilepsy in experimental female models of TLE.

A.7. Concluding remarks
In this review, we have described the different versions of the kainic acid model. This model has been extensively used for decades and is proven to be a reliable tool to mimic numerous behavioral, electrophysiological, and neuropathological features of epilepsy. In our opinion, whether or not the KA models are mimicking human TLE is not a relevant question if only because a rodent brain is not a human brain. What is important are the questions addressed in the rodent models and the hypotheses being tested. We argue that all models are interesting as long as they are characterized by spontaneous seizures.
Diversity is a hallmark of human epilepsy, even for some specific types such as TLE. Interestingly, different kainic acid models are also very diverse. The results vary as a function of the way kainic acid is administered, species, strain, sex, and age. We argue that this is a strength as the different models may cover some of the diversity found in patients.
Clearly, each model only revealed the tip of the iceberg. Many important issues remain unaddressed. Perhaps the most important one is to determine the seizure onset zone(s) and the propagation pattern in the brain. This requires multi-site recordings, which are regularly done in patients during presurgical evaluation. Such a study has been performed in the pilocarpine model . The same approach should now be used 332
in the various KA models. Another pressing question is sex. As mentioned, the number of studies performed in males far outweigh those done in females. It is like half of the field is missing. Although more work is done during development and ageing, more work is also clearly needed as the mechanisms are likely to be age-dependent. Finally, recent studies highlight the necessity to consider circadian and multidien rhythms, the existence of different classes of seizures, as well as the impact of housing (single or colony), which can add the confounding factor of stress. Perhaps accepting the diversity of what the different models and experimental conditions have to offer is the best way to understand the diversity of human phenotypes.

Bibliography
Fisher RS, Acevedo C, Arzimanoglou A, Bogacz A, Cross JH, … & Wiebe S (April 2014). "ILAE official 1
report: a practical clinical definition of epilepsy". Epilepsia. 55 (4): 475–82. doi:10.1111/epi.12550
Tang, F., Hartz, A. M. S., & Bauer, B. (2017). Drug-Resistant Epilepsy: Multiple Hypotheses, Few 2
Answers. Frontiers in Neurology. JOUR. https://www.frontiersin.org/article/10.3389/fneur.2017.00301
Behr, C., Goltzene, M. A., Kosmalski, G., Hirsch, E., & Ryvlin, P. (2016). Epidemiology of epilepsy. 3
Revue Neurologique, 172(1), 27–36. JOUR. https://doi.org/10.1016/j.neurol.2015.11.003
Goltzene, M.-A., Kleinlogel, S., Ellero, C., Valenti-Hirsch, M.-P., Hirsch, É., & Gonzalez, M. (2018). 4
Intégration dans le milieu professionnel des patients atteints d’épilepsie : de l’importance d’une réunion de concertation pluridisciplinaire. Archives Des Maladies Professionnelles et de l’Environnement, 79(3), 242–243. JOUR. https://doi.org/10.1016/j.admp.2018.03.127
Beghi, E. (2020). The Epidemiology of Epilepsy. Neuroepidemiology, 54(2), 185–191. JOUR. http://5
doi.org/10.1159/000503831
Thurman, D.J., Logroscino, G., Beghi, E., Hauser, W.A., Hesdorffer, D.C., Newton, C.R., Scorza, F.A., 6
Sander, J.W., Tomson, T. and (2017), The burden of premature mortality of epilepsy in high‐income countries: A systematic review from the Mortality Task Force of the International League Against Epilepsy. Epilepsia, 58: 17-26. doi:10.1111/epi.13604
Fisher, RS; Cross, JH; French, JA; Higurashi, N; Hirsch, E; … & Sameer M. (April 2017). "Operational 7
classification of seizure types by the International League Against Epilepsy: Position Paper of the ILAE Commission for Classification and Terminology". Epilepsia. 58 (4): 522–530. doi:10.1111/epi.13670.
Blumcke, I., Spreafico, R., Haaker, G., Coras, R., Kobow, K., Bien, C. G., … Avanzini, G. (2017). 8
Histopathological Findings in Brain Tissue Obtained during Epilepsy Surgery. New England Journal of
Medicine, 377(17), 1648–1656. JOUR. http://doi.org/10.1056/NEJMoa1703784
Chen, T., Giri, M., Xia, Z., Subedi, Y. N., & Li, Y. (2017). Genetic and epigenetic mechanisms of 9
epilepsy: a review. Neuropsychiatric disease and treatment, 13, 1841–1859. https://doi.org/10.2147/NDT.S142032
Vezzani, A., Fujinami, R. S., White, H. S., Preux, P. M., Blümcke, I., Sander, J. W., & Löscher, W. (2016). 10
Infections, inflammation and epilepsy. Acta neuropathologica, 131(2), 211–234. https://doi.org/10.1007/s00401-015-1481-5
Neal, E. G., Chaffe, H., Schwartz, R. H., Lawson, M. S., Edwards, N., Fitzsimmons, G., … Cross, J. H. 11
(2008). The ketogenic diet for the treatment of childhood epilepsy: a randomised controlled trial. The
Lancet Neurology, 7(6), 500–506. JOUR. https://doi.org/10.1016/S1474-4422(08)70092-9
Lancaster, E., & Dalmau, J. (2012). Neuronal autoantigens—pathogenesis, associated disorders and 12
antibody testing. Nature Reviews Neurology, 8(7), 380.
Devinsky, O., Vezzani, A., Najjar, S., De Lanerolle, N. C., & Rogawski, M. A. (2013). Glia and epilepsy: 13
excitability and inflammation. Trends in Neurosciences, 36(3), 174–184. JOUR. https://doi.org/10.1016/j.tins.2012.11.008
Sharma, A. K., Reams, R. Y., Jordan, W. H., Miller, M. A., Thacker, H. L., & Snyder, P. W. (2007). Mesial 14
Temporal Lobe Epilepsy: Pathogenesis, Induced Rodent Models and Lesions. Toxicologic Pathology, 35(7), 984–999. https://doi.org/10.1080/01926230701748305
Sutula, T. P., & Dudek, F. E. (2007). Unmasking recurrent excitation generated by mossy fiber 15
sprouting in the epileptic dentate gyrus: an emergent property of a complex system. In H. E. B. T.-P. in B. R. Scharfman (Ed.), The Dentate Gyrus: A Comprehensive Guide to Structure, Function, and Clinical
Implications (Vol. 163, pp. 541–563). CHAP, Elsevier. https://doi.org/10.1016/S0079-6123(07)63029-5
Schmeiser, B., Zentner, J., Prinz, M., Brandt, A., & Freiman, T. M. (2017). Extent of mossy fiber 16
sprouting in patients with mesiotemporal lobe epilepsy correlates with neuronal cell loss and granule cell dispersion. Epilepsy Research, 129, 51–58. JOUR. https://doi.org/10.1016/j.eplepsyres.2016.11.011

Sloviter, R.S., Zappone, C.A., Harvey, B.D., Bumanglag, A.V., Bender, R.A. and Frotscher, M. (2003), “17
Dormant basket cell” hypothesis revisited: Relative vulnerabilities of dentate gyrus mossy cells and inhibitory interneurons after hippocampal status epilepticus in the rat. J. Comp. Neurol., 459: 44-76. doi:10.1002/cne.10630
Engelborghs, S., D’hooge, R., & De Deyn, P. P. (2000). Pathophysiology of epilepsy. Acta neurologica 18
belgica, 100(4), 201-213.
Chapman, A. G. (2000). Glutamate and Epilepsy. The Journal of Nutrition, 130(4), 1043S–1045S. 19
JOUR. http://doi.org/10.1093/jn/130.4.1043S
Sommer, C., Roth, S. U., & Kiessling, M. (2001). Kainate-induced epilepsy alters protein expression of 20
AMPA receptor subunits GluR1, GluR2 and AMPA receptor binding protein in the rat hippocampus. Acta
Neuropathologica, 101(5), 460–468. JOUR. http://doi.org/10.1007/s004010000310
Telfeian, A. E., Federoff, H. J., Leone, P., During, M. J., & Williamson, A. (2000). Overexpression of 21
GluR6 in rat hippocampus produces seizures and spontaneous nonsynaptic bursting in vitro. Neurobiology of disease, 7(4), 362–374. https://doi.org/10.1006/nbdi.2000.0294
During, M. J., Ryder, K. M., & Spencer, D. D. (1995). Hippocampal GABA transporter function in 22
temporal-lobe epilepsy. Nature, 376(6536), 174–177. JOUR. http://doi.org/10.1038/376174a0
Kaila, K., Ruusuvuori, E., Seja, P., Voipio, J., & Puskarjov, M. (2014). GABA actions and ionic plasticity 23
in epilepsy. Current Opinion in Neurobiology, 26, 34–41. https://doi.org/10.1016/j.conb.2013.11.004
"Proposal for revised clinical and electroencephalographic classification of epileptic seizures. From 24
the Commission on Classification and Terminology of the International League Against Epilepsy". Epilepsia. 22 (4): 489–501. 1981. doi:10.1111/j.1528-1157.1981.tb06159.x
Foldvary-Schaefer, N., & Unnwongse, K. (2011). Localizing and lateralizing features of auras and 25
seizures. Epilepsy & Behavior, 20(2), 160–166. JOUR. https://doi.org/10.1016/j.yebeh.2010.08.034
Guedj, E., Aubert, S., McGonigal, A., Mundler, O., & Bartolomei, F. (2010). Déjà-vu in temporal lobe 26
epilepsy: Metabolic pattern of cortical involvement in patients with normal brain MRI. Neuropsychologia, 48(7), 2174–2181. JOUR. https://doi.org/10.1016/j.neuropsychologia.2010.04.009
Illman, N. A., Butler, C. R., Souchay, C., & Moulin, C. J. A. (2012). Déjà Experiences in Temporal Lobe 27
Epilepsy. Epilepsy Research and Treatment, 2012, 539567. JOUR. http://doi.org/10.1155/2012/539567
Smith, S. J. M. (2005). EEG in the diagnosis, classification, and management of patients with epilepsy. 28
Journal of Neurology, Neurosurgery & Psychiatry, 76. http://doi.org/10.1136/jnnp.2005.069245
Lin, Y. Y., Shih, Y. H., Hsieh, J. C., Yu, H. Y., Yiu, C. H., Wong, T. T., … Chang, M. S. (2003). 29
Magnetoencephalographic yield of interictal spikes in temporal lobe epilepsy: Comparison with scalp EEG recordings. NeuroImage, 19(3), 1115–1126. JOUR. https://doi.org/10.1016/S1053-8119(03)00181-2
Di Gennaro, G., Quarato, P. P., Sebastiano, F., Esposito, V., Onorati, P., Grammaldo, L. G., … Cantore, 30
G. (2004). Ictal heart rate increase precedes EEG discharge in drug-resistant mesial temporal lobe seizures. Clinical Neurophysiology, 115(5), 1169–1177. https://doi.org/10.1016/j.clinph.2003.12.016
Jung, K.-Y., Kang, J.-K., Kim, J. H., Im, C.-H., Kim, K. H., & Jung, H.-K. (2009). Spatiotemporospectral 31
characteristics of scalp ictal EEG in mesial temporal lobe epilepsy with hippocampal sclerosis. Brain
Research, 1287, 206–219. https://doi.org/10.1016/j.brainres.2009.06.071
Ferrillo, F., Beelke, M., & Nobili, L. (2000). Sleep EEG synchronization mechanisms and activation of 32
interictal epileptic spikes. Clinical Neurophysiology, 111, S65–S73. JOUR. https://doi.org/10.1016/S1388-2457(00)00404-1
Kasteleijn‐Nolst Trenité, D., Rubboli, G., Hirsch, E., Martins da Silva, A., Seri, S., Wilkins, A., Parra, J., 33
Covanis, A., Elia, M., Capovilla, G., Stephani, U. and Harding, G. (2012), Methodology of photic stimulation revisited: Updated European algorithm for visual stimulation in the EEG laboratory. Epilepsia, 53: 16-24. doi:10.1111/j.1528-1167.2011.03319.x
Guaranha, M.S.B., Garzon, E., Buchpiguel, C.A., Tazima, S., Yacubian, E.M.T. and Sakamoto, A.C. 34
(2005), Hyperventilation Revisited: Physiological Effects and Efficacy on Focal Seizure Activation in the Era of Video‐EEG Monitoring. Epilepsia, 46: 69-75. doi:10.1111/j.0013-9580.2005.11104.x

Sohrabpour, A., Lu, Y., Kankirawatana, P., Blount, J., Kim, H., & He, B. (2015). Effect of EEG electrode 35
number on epileptic source localization in pediatric patients. Clinical Neurophysiology, 126(3), 472–480. JOUR. https://doi.org/10.1016/j.clinph.2014.05.038
Shorvon, S., Perucca, E., & Engel Jr, J. (Eds.). (2015). The treatment of epilepsy. John Wiley & Sons.36
National Clinical Guideline Centre (January 2012). The Epilepsies: The diagnosis and management of 37
the epilepsies in adults and children in primary and secondary care. National Institute for Health and Clinical Excellence. pp. 21–28.
Kral, T., Clusmann, H., Urbach, J., Schramm, J., Elger, C. E., Kurthen, M., & Grunwald, T. (2002). 38
Preoperative evaluation for epilepsy surgery (Bonn Algorithm). Zentralblatt für Neurochirurgie, 63(03), 106-110
Engel, J., McDermott, M. P., Wiebe, S., Langfitt, J. T., Stern, J. M., Dewar, S., … Early Randomized 39
Surgical Epilepsy Trial (ERSET) Study Group, for the. (2012). Early Surgical Therapy for Drug-Resistant Temporal Lobe Epilepsy: A Randomized Trial. JAMA, 307(9), 922–930. JOUR. http://doi.org/10.1001/jama.2012.220
Boon, P., Vonck, K., De Herdt, V., Van Dycke, A., Goethals, M., Goossens, L., Van Zandijcke, M., De 40
Smedt, T., Dewaele, I., Achten, R., Wadman, W., Dewaele, F., Caemaert, J. and Van Roost, D. (2007), Deep Brain Stimulation in Patients with Refractory Temporal Lobe Epilepsy. Epilepsia, 48: 1551-1560. doi:10.1111/j.1528-1167.2007.01005.x
Kros, L., Eelkman Rooda, O. H. J., De Zeeuw, C. I., & Hoebeek, F. E. (2015). Controlling Cerebellar 41
Output to Treat Refractory Epilepsy. Trends in Neurosciences, 38(12), 787–799. JOUR. https://doi.org/10.1016/j.tins.2015.10.002
Charbades, S., Kahane, P., Minotti, L., Koudsie, A., Hirsh, E., & Benabid, A. L. (2002). Deep brain 42
stimulation in epilepsy with particular reference to the subthalamic nucleus. Epileptic disorders, 4, S83-S93.
Shi, L.-H., Luo, F., Woodward, D., & Chang, J.-Y. (2006). Deep brain stimulation of the substantia nigra 43
pars reticulata exerts long lasting suppression of amygdala-kindled seizures. Brain Research, 1090(1), 202–207. JOUR. https://doi.org/10.1016/j.brainres.2006.03.050
La Grutta, V., & Sabationo, M. (1988). Focal hippocampal epilepsy: Effect of caudate stimulation. 44
Experimental Neurology, 99(1), 38–49. JOUR. https://doi.org/10.1016/0014-4886(88)90125-2
Hodaie, M., Wennberg, R., Dostrovsky, J. and Lozano, A. (2002), Chronic Anterior Thalamus 45
Stimulation for Intractable Epilepsy. Epilepsia, 43: 603-608. doi:10.1046/j.1528-1157.2002.26001.x
Velasco, F., Velasco, A. L., Velasco, M., Jiménez, F., Carrillo-Ruiz, J. D., & Castro, G. (2007). Deep 46
brain stimulation for treatment of the epilepsies: the centromedian thalamic target BT - Operative Neuromodulation: Volume 2: Neural Networks Surgery. In D. E. Sakas & B. A. Simpson (Eds.) (pp. 337–342). CHAP, Vienna: Springer Vienna. http://doi.org/10.1007/978-3-211-33081-4_38
Velasco, A.L., Velasco, F., Velasco, M., Trejo, D., Castro, G. and Carrillo‐Ruiz, J.D. (2007), Electrical 47
Stimulation of the Hippocampal Epileptic Foci for Seizure Control: A Double‐Blind, Long‐Term Follow‐Up Study. Epilepsia, 48: 1895-1903. doi:10.1111/j.1528-1167.2007.01181.x
Chkhenkeli, S. A., Šramka, M., Lortkipanidze, G. S., Rakviashvili, T. N., Bregvadze, E. S., Magalashvili, 48
G. E., … Chkhenkeli, I. S. (2004). Electrophysiological effects and clinical results of direct brain stimulation for intractable epilepsy. Clinical Neurology and Neurosurgery, 106(4), 318–329. JOUR. https://doi.org/10.1016/j.clineuro.2004.01.009
Bancaud, J., Angelergues, R., Bernouilli, C., Bonis, A., Bordas-Ferrer, M., Bresson, M., ... & Takeda, A. 49
(1969). Functional stereotaxic exploration (stereo-electroencephalography) in epilepsies. Revue
neurologique, 120(6), 448-448.
Rosenbaum, T. J., Laxer, K. D., Vessely, M., & Smith, W. B. (1986). Subdural Electrodes for Seizure 50
Focus Localization. Neurosurgery, 19(1), 73–81. http://doi.org/10.1227/00006123-198607000-00011
Goldring, S., & Gregorie, E. M. (1984). Surgical management of epilepsy using epidural recordings to 51
localize the seizure focus. Journal of Neurosurgery, 60(3), 457–466. JOUR. http://doi.org/10.3171/jns.1984.60.3.0457

Pilcher, W. H., & Rusyniak, W. G. (1993). Complications of Epilepsy Surgery. Neurosurgery Clinics of 52
North America, 4(2), 311–325. JOUR. https://doi.org/10.1016/S1042-3680(18)30597-7
Salanova, V., Quesney, L.F., Rasmussen, T., Andermann, F. and Olivier, A. (1994), Reevaluation of 53
Surgical Failures and the Role of Reoperation in 39 Patients with Frontal Lobe Epilepsy. Epilepsia, 35: 70-80. doi:10.1111/j.1528-1157.1994.tb02914.x
Risinger, M. W., Engel, J., Van Ness, P. C., Henry, T. R., & Crandall, P. H. (1989). Ictal localization of 54
temporal lobe seizures with scalp/sphenoidal recordings. Neurology, 39(10), 1288 LP-1288. JOUR. http://doi.org/10.1212/WNL.39.10.1288
Wyler, A. R., Richey, E. T., & Hermann, B. P. (1989). Comparison of scalp to subdural recordings for 55
localizing epileptogenic foci. Journal of Epilepsy, 2(2), 91–96. JOUR. https://doi.org/10.1016/0896-6974(89)90043-1
Cossu, M., Cardinale, F., Castana, L., Citterio, A., Francione, S., Tassi, L., Benabid, A. L., & Lo Russo, 56
G. (2005). Stereoelectroencephalography in the presurgical evaluation of focal epilepsy: a retrospective analysis of 215 procedures. Neurosurgery, 57(4), 706–718.
Jayakar, P., Gotman, J., Harvey, A.S., Palmini, A., Tassi, L., Schomer, D., Dubeau, F., Bartolomei, F., 57
Yu, A., Kršek, P., Velis, D. and Kahane, P. (2016), Diagnostic utility of invasive EEG for epilepsy surgery: Indications, modalities, and techniques. Epilepsia, 57: 1735-1747. doi:10.1111/epi.13515
Taussig, D., Chipaux, M., Fohlen, M., Dorison, N., Bekaert, O., Ferrand-Sorbets, S., & Dorfmüller, G. 58
(2020). Invasive evaluation in children (SEEG vs subdural grids). Seizure, 77, 43–51. https://doi.org/10.1016/j.seizure.2018.11.008
Mullin, J.P., Shriver, M., Alomar, S., Najm, I., Bulacio, J., Chauvel, P. and Gonzalez‐Martinez, J. (2016), 59
Is SEEG safe? A systematic review and meta‐analysis of stereo‐electroencephalography–related complications. Epilepsia, 57: 386-401. doi:10.1111/epi.13298
Penfield, W., & Jasper, H. (1954). Electrocorticography. Epilepsy and the functional anatomy of the 60
human brain, 692-738.
Schuh, L., & Drury, I. (1997). Intraoperative electrocorticography and direct cortical electrical 61
stimulation. Seminars in Anesthesia, Perioperative Medicine and Pain, 16(1), 46–55. https://doi.org/10.1016/S0277-0326(97)80007-4
Borchers, S., Himmelbach, M., Logothetis, N., & Karnath, H.-O. (2012). Direct electrical stimulation of 62
human cortex — the gold standard for mapping brain functions? Nature Reviews Neuroscience, 13(1), 63–70. https://doi.org/10.1038/nrn3140
Miller, K. J., denNijs, M., Shenoy, P., Miller, J. W., Rao, R. P. N., & Ojemann, J. G. (2007). Real-time 63
functional brain mapping using electrocorticography. NeuroImage, 37(2), 504–507. https://doi.org/10.1016/j.neuroimage.2007.05.029
Hamer, H. M., Morris, H. H., Mascha, E. J., Karafa, M. T., Bingaman, W. E., Bej, M. D., … Lüders, H. 64
O. (2002). Complications of invasive video-EEG monitoring with subdural grid electrodes. Neurology, 58(1), 97 LP – 103. https://doi.org/10.1212/WNL.58.1.97
Placantonakis, D. G., Shariff, S., Lafaille, F., Labar, D., Harden, C., Hosain, S., Kandula, P., Schaul, N., 65
Kolesnik, D., & Schwartz, T. H. (2010). Bilateral intracranial electrodes for lateralizing intractable epilepsy: efficacy, risk, and outcome. Neurosurgery, 66(2), 274–283. https://doi.org/10.1227/01.NEU.0000363184.43723.94
Spencer, S. S., Spencer, D. D., Williamson, P. D., & Mattson, R. (1990). Combined depth and subdural 66
electrode investigation in uncontrolled epilepsy. Neurology, 40(1), 74–79. https://doi.org/10.1212/wnl.40.1.74
van Veelen, C. W., Debets, R. M., van Huffelen, A. C., van Emde Boas, W., Binnie, C. D., Storm van 67
Leeuwen, W., Velis, D. N., & van Dieren, A. (1990). Combined use of subdural and intracerebral electrodes in preoperative evaluation of epilepsy. Neurosurgery, 26(1), 93–101. https://doi.org/10.1097/00006123-199001000-00013
Guerrini, R., Scerrati, M., Rubboli, G., … Commission for Epilepsy Surgery of the Italian League 68
Against Epilepsy (2013). Overview of presurgical assessment and surgical treatment of epilepsy from the Italian League Against Epilepsy. Epilepsia, 54 Suppl 7, 35–48. https://doi.org/10.1111/epi.12308

Mathon, B., Clemenceau, S., Hasboun, D., Habert, M. O., Belaid, H., Nguyen-Michel, V. H., Lambrecq, 69
V., Navarro, V., Dupont, S., Baulac, M., Cornu, P., & Adam, C. (2015). Safety profile of intracranial electrode implantation for video-EEG recordings in drug-resistant focal epilepsy. Journal of neurology, 262(12), 2699–2712. https://doi.org/10.1007/s00415-015-7901-6
Xu, X.-L., Xu, B., & He, B. (2004). An alternative subspace approach to EEG dipole source localization. 70
Physics in Medicine and Biology, 49(2), 327–343. https://doi.org/10.1088/0031-9155/49/2/010
Ding, L., Wilke, C., Xu, B., Xu, X., van Drongelen, W., Kohrman, M., & He, B. (2007). EEG Source 71
Imaging: Correlating Source Locations and Extents With Electrocorticography and Surgical Resections in Epilepsy Patients. Journal of Clinical Neurophysiology, 24(2). https://journals.lww.com/clinicalneurophys/Fulltext/2007/04000/EEG_Source_Imaging__Correlating_Source_Locations.6.aspx
Lu, Y., Yang, L., Worrell, G. A., & He, B. (2012). Seizure source imaging by means of FINE spatio-72
temporal dipole localization and directed transfer function in partial epilepsy patients. Clinical
Neurophysiology, 123(7), 1275–1283. https://doi.org/10.1016/j.clinph.2011.11.007
Pascual-Marqui, R. D., Michel, C. M., & Lehmann, D. (1994). Low resolution electromagnetic 73
tomography: a new method for localizing electrical activity in the brain. International journal of
psychophysiology : official journal of the International Organization of Psychophysiology, 18(1), 49–65. https://doi.org/10.1016/0167-8760(84)90014-x
Clemens, B., Bessenyei, M., Fekete, I., Puskás, S., Kondákor, I., Tóth, M., & Hollódy, K. (2010). Theta 74
EEG source localization using LORETA in partial epilepsy patients with and without medication. Clinical
Neurophysiology, 121(6), 848–858. https://doi.org/10.1016/j.clinph.2010.01.020
Staljanssens, W., Strobbe, G., Van Holen, R., Keereman, V., Gadeyne, S., Carrette, E., … van Mierlo, P. 75
(2017). EEG source connectivity to localize the seizure onset zone in patients with drug resistant epilepsy. NeuroImage: Clinical, 16, 689–698. https://doi.org/10.1016/j.nicl.2017.09.011
Nemtsas, P., Birot, G., Pittau, F., Michel, C.M., Schaller, K., Vulliemoz, S., Kimiskidis, V.K. and Seeck, 76
M. (2017), Source localization of ictal epileptic activity based on high‐density scalp EEG data. Epilepsia, 58: 1027-1036. doi:10.1111/epi.13749
Kvam, K. A., Douglas, V. C., Whetstone, W. D., Josephson, S. A., & Betjemann, J. P. (2019). Yield of 77
Emergent CT in Patients With Epilepsy Presenting With a Seizure. The Neurohospitalist, 9(2), 71–78. https://doi.org/10.1177/1941874418808676
Bronen, R. A., Fulbright, R. K., Spencer, D. D., Spencer, S. S., Kim, J. H., Lange, R. C., & Sutilla, C. 78
(1996). Refractory epilepsy: comparison of MR imaging, CT, and histopathologic findings in 117 patients. Radiology, 201(1), 97–105. https://doi.org/10.1148/radiology.201.1.8816528
Rüber, T., David, B., & Elger, C. E. (2018). MRI in epilepsy: clinical standard and evolution. Current 79
opinion in neurology, 31(2), 223-231.
Bernasconi, N., Bernasconi, A., Caramanos, Z., Antel, S. B., Andermann, F., & Arnold, D. L. (2003). 80
Mesial temporal damage in temporal lobe epilepsy: a volumetric MRI study of the hippocampus, amygdala and parahippocampal region. Brain, 126(2), 462–469. https://doi.org/10.1093/brain/awg034
Wang, Z. I., Alexopoulos, A. V, Jones, S. E., Jaisani, Z., Najm, I. M., & Prayson, R. A. (2013). The 81
pathology of magnetic-resonance-imaging-negative epilepsy. Modern Pathology, 26(8), 1051–1058. https://doi.org/10.1038/modpathol.2013.52
Vézina, L.‐G. (2011), MRI‐negative epilepsy: Protocols to optimize lesion detection. Epilepsia, 52: 82
25-27. doi:10.1111/j.1528-1167.2011.03147.x
Binder, J.R., Sabsevitz, D.S., Swanson, S.J., Hammeke, T.A., Raghavan, M. and Mueller, W.M. (2008), 83
Use of preoperative functional MRI to predict verbal memory decline after temporal lobe epilepsy surgery. Epilepsia, 49: 1377-1394. doi:10.1111/j.1528-1167.2008.01625.x
Szaflarski, J. P., Gloss, D., Binder, J. R., Gaillard, W. D., Golby, A. J., Holland, S. K., … Theodore, W. 84
H. (2017). Practice guideline summary: Use of fMRI in the presurgical evaluation of patients with epilepsy. Neurology, 88(4), 395 LP – 402. https://doi.org/10.1212/WNL.0000000000003532
la Fougère, C., Rominger, A., Förster, S., Geisler, J., & Bartenstein, P. (2009). PET and SPECT in 85
epilepsy: A critical review. Epilepsy & Behavior, 15(1), 50–55. https://doi.org/10.1016/j.yebeh.2009.02.025

Vanli-Yavuz, E. N., Baykan, B., Sencer, S., Sencer, A., Baral-Kulaksizoglu, I., Bebek, N., Gurses, C., & 86
Gokyigit, A. (2017). How Different Are the Patients With Bilateral Hippocampal Sclerosis From the Unilateral Ones Clinically?. Clinical EEG and neuroscience, 48(3), 209–216. https://doi.org/10.1177/1550059416653900
LoPinto‐Khoury, C., Sperling, M.R., Skidmore, C., Nei, M., Evans, J., Sharan, A. and Mintzer, S. 87
(2012), Surgical outcome in PET‐positive, MRI‐negative patients with temporal lobe epilepsy. Epilepsia, 53: 342-348. doi:10.1111/j.1528-1167.2011.03359.x
O’Brien, T. J., Hicks, R. J., Ware, R., Binns, D. S., Murphy, M., & Cook, M. J. (2001). The utility of a 3-88
dimensional, large-field-of-view, sodium iodide crystal–based PET scanner in the presurgical evaluation of partial epilepsy. Journal of Nuclear Medicine, 42(8), 1158-1165.
Won, H. J., Chang, K.-H., Cheon, J.-E., Kim, H. D., Lee, D. S., Han, M. H., … Chung, C.-K. (1999). 89
Comparison of MR Imaging with PET and IctalSPECT in 118 Patients with Intractable Epilepsy. American Journal of Neuroradiology, 20(4), 593 LP – 599. http://www.ajnr.org/content/20/4/593.abstract
Muzik, O., da Silva, E. A., Juhasz, C., Chugani, D. C., Shah, J., Nagy, F., … Chugani, H. T. (2000). 90
Intracranial EEG versus flumazenil and glucose PET in children with extratemporal lobe epilepsy. Neurology, 54(1), 171 LP – 171. https://doi.org/10.1212/WNL.54.1.171
Didelot, A., Ryvlin, P., Lothe, A., Merlet, I., Hammers, A., & Mauguière, F. (2008). PET imaging of brain 91
5-HT1A receptors in the preoperative evaluation of temporal lobe epilepsy. Brain, 131(10), 2751–2764. https://doi.org/10.1093/brain/awn220
Spanaki, M. V., Spencer, S. S., Corsi, M., MacMullan, J., Seibyl, J., & Zubal, I. G. (1999). Sensitivity 92
and specificity of quantitative difference SPECT analysis in seizure localization. Journal of Nuclear
Medicine, 40(5), 730-736.
Devous, M. D., Thisted, R. A., Morgan, G. F., Leroy, R. F., & Rowe, C. C. (1998). SPECT brain imaging 93
in epilepsy: a meta-analysis. In Database of Abstracts of Reviews of Effects (DARE): Quality-assessed
Reviews [Internet]. Centre for Reviews and Dissemination (UK).
WEIL, S., NOACHTAR, S., ARNOLD, S., YOUSRY, T. A., WINKLER, P. A., & TATSCH, K. (2001). Ictal 94
ECD-SPECT differentiates between temporal and extratemporal epilepsy: confirmation by excellent postoperative seizure control. Nuclear Medicine Communications, 22(2).
Zaknun, J. J., Bal, C., Maes, A., Tepmongkol, S., Vazquez, S., Dupont, P., & Dondi, M. (2008). 95
Comparative analysis of MR imaging, Ictal SPECT and EEG in temporal lobe epilepsy: a prospective IAEA multi-center study. European Journal of Nuclear Medicine and Molecular Imaging, 35(1), 107–115. https://doi.org/10.1007/s00259-007-0526-y
O’Brien, T. J., O’Connor, M. K., Mullan, B. P., Brinkmann, B. H., Hanson, D., Jack, C. R., & So, E. L. 96
(1998). Subtraction ictal SPET co-registered to MRI in partial epilepsy: Description and technical validation of the method with phantom and patient studies. Nuclear Medicine Communications, 19(1).
Velasco, T. R., Wichert-Ana, L., Mathern, G. W., Araújo, D., Walz, R., Bianchin, M. M., … Sakamoto, A. 97
C. (2011). Utility of Ictal Single Photon Emission Computed Tomography in Mesial Temporal Lobe Epilepsy With Hippocampal Atrophy: A Randomized Trial. Neurosurgery, 68(2), 431–436. https://doi.org/10.1227/NEU.0b013e318201c293
Sun, P. Y., Wyatt, K., Nickels, K. C., Wong-Kisiel, L. C., Mandrekar, J., & Wirrell, E. (2015). Predictors of 98
Length of Stay in Children Admitted for Presurgical Evaluation for Epilepsy Surgery. Pediatric Neurology, 53(3), 207–210. https://doi.org/10.1016/j.pediatrneurol.2015.05.016
Singh S. P. (2014). Magnetoencephalography: Basic principles. Annals of Indian Academy of 99
Neurology, 17(Suppl 1), S107–S112. https://doi.org/10.4103/0972-2327.128676
Papanicolaou, A. C., Pataraia, E., Billingsley-Marshall, R., Castillo, E. M., Wheless, J. W., Swank, P., 100
Breier, J. I., Sarkari, S., & Simos, P. G. (2005). Toward the Substitution of Invasive Electroencephalography in Epilepsy Surgery. Journal of Clinical Neurophysiology, 22(4). https://journals.lww.com/clinicalneurophys/Fulltext/2005/08000/Toward_the_Substitution_of_Invasive.1.aspx
Lau, M., Yam, D., & Burneo, J. G. (2008). A systematic review on MEG and its use in the presurgical 101
evaluation of localization-related epilepsy. Epilepsy Research, 79(2), 97–104. https://doi.org/10.1016/j.eplepsyres.2008.01.004

Knowlton, R.C., Elgavish, R., Howell, J., Blount, J., Burneo, J.G., Faught, E., Kankirawatana, P., Riley, 102
K., Morawetz, R., Worthington, J. and Kuzniecky, R.I. (2006), Magnetic source imaging versus intracranial electroencephalogram in epilepsy surgery: A prospective study. Ann Neurol., 59: 835-842. doi:10.1002/ana.20857
Stefan, H., Hummel, C., Scheler, G., Genow, A., Druschky, K., Tilz, C., Kaltenhäuser, M., 103
Hopfengärtner, R., Buchfelder, M., & Romstöck, J. (2003). Magnetic brain source imaging of focal epileptic activity: a synopsis of 455 cases. Brain, 126(11), 2396–2405. https://doi.org/10.1093/brain/awg239
Hallett, M. (2000). Transcranial magnetic stimulation and the human brain. Nature, 406(6792), 147–104
150. JOUR. http://doi.org/10.1038/35018000
Lipton, R. B., Dodick, D. W., Silberstein, S. D., Saper, J. R., Aurora, S. K., Pearlman, S. H., … 105
Goadsby, P. J. (2010). Single-pulse transcranial magnetic stimulation for acute treatment of migraine with aura: a randomised, double-blind, parallel-group, sham-controlled trial. The Lancet Neurology, 9(4), 373–380. JOUR. https://doi.org/10.1016/S1474-4422(10)70054-5
Hovington, C. L., McGirr, A., Lepage, M., & Berlim, M. T. (2013). Repetitive transcranial magnetic 106
stimulation (rTMS) for treating major depression and schizophrenia: a systematic review of recent meta-analyses. Annals of Medicine, 45(4), 308–321. JOUR. http://doi.org/10.3109/07853890.2013.783993
Groppa, S., Oliviero, A., Eisen, A., Quartarone, A., Cohen, L. G., Mall, V., … Siebner, H. R. (2012). A 107
practical guide to diagnostic transcranial magnetic stimulation: Report of an IFCN committee. Clinical
Neurophysiology, 123(5), 858–882. JOUR. https://doi.org/10.1016/j.clinph.2012.01.010
Picht, T., Mularski, S., Kuehn, B., Vajkoczy, P., Kombos, T., & Suess, O. (2009). Navigated 108
Transcranial Magnetic Stimulation for Preoperative Functional Diagnostics in Brain Tumor Surgery. Operative Neurosurgery, 65. JOUR. http://doi.org/10.1227/01.NEU.0000348009.22750.59
Säisänen, L., Könönen, M., Julkunen, P., Määttä, S., Vanninen, R., Immonen, A., Jutila, L., Kälviäinen, 109
R., Jääskeläinen, J. E., & Mervaala, E. (2010). Non-invasive preoperative localization of primary motor cortex in epilepsy surgery by navigated transcranial magnetic stimulation. Epilepsy research, 92(2-3), 134–144. https://doi.org/10.1016/j.eplepsyres.2010.08.013
Jennum, P., Winkel, H., Fuglsang-Frederiksen, A., & Dam, M. (1994). EEG changes following 110
repetitive transcranial magnetic stimulation in patients with temporal lobe epilepsy. Epilepsy Research, 18(2), 167–173. JOUR. https://doi.org/10.1016/0920-1211(94)90009-4
Jennum, P., Friberg, L., Fuglsang-Frederiksen, A., & Dam, M. (1994). Speech localization using 111
repetitive transcranial magnetic stimulation. Neurology, 44(2), 269 LP-269. JOUR. http://doi.org/10.1212/WNL.44.2.269
Macdonell, R. A. L., Jackson, G. D., Curatolo, J. M., Abbott, D. F., Berkovic, S. F., Carey, L. M., … 112
Scheffer, I. E. (1999). Motor cortex localization using functional MRI and transcranial magnetic stimulation. Neurology, 53(7), 1462 LP-1462. JOUR. http://doi.org/10.1212/WNL.53.7.1462
Säisänen, L., Könönen, M., Julkunen, P., Määttä, S., Vanninen, R., Immonen, A., Jutila, L., Kälviäinen, 113
R., Jääskeläinen, J. E., & Mervaala, E. (2010). Non-invasive preoperative localization of primary motor cortex in epilepsy surgery by navigated transcranial magnetic stimulation. Epilepsy research, 92(2-3), 134–144. https://doi.org/10.1016/j.eplepsyres.2010.08.013
Vitikainen, A. M., Lioumis, P., Paetau, R., Salli, E., Komssi, S., Metsähonkala, L., Paetau, A., Kicić, D., 114
Blomstedt, G., Valanne, L., Mäkelä, J. P., & Gaily, E. (2009). Combined use of non-invasive techniques for improved functional localization for a selected group of epilepsy surgery candidates. NeuroImage, 45(2), 342–348. https://doi.org/10.1016/j.neuroimage.2008.12.026
Pereira, L. S., Müller, V. T., da Mota Gomes, M., Rotenberg, A., & Fregni, F. (2016). Safety of repetitive 115
transcranial magnetic stimulation in patients with epilepsy: A systematic review. Epilepsy & Behavior, 57, 167–176. JOUR. https://doi.org/10.1016/j.yebeh.2016.01.015
Bae, E. H., Schrader, L. M., Machii, K., Alonso-Alonso, M., Riviello, J. J., Jr, Pascual-Leone, A., & 116
Rotenberg, A. (2007). Safety and tolerability of repetitive transcranial magnetic stimulation in patients with epilepsy: a review of the literature. Epilepsy & behavior : E&B, 10(4), 521–528. https://doi.org/10.1016/j.yebeh.2007.03.004

Paulus, W. (2011). Transcranial electrical stimulation (tES – tDCS; tRNS, tACS) methods. 117
Neuropsychological Rehabilitation, 21(5), 602–617.JOUR. http://doi.org/10.1080/09602011.2011.557292
Stagg, C. J., & Nitsche, M. A. (2011). Physiological Basis of Transcranial Direct Current Stimulation. 118
The Neuroscientist, 17(1), 37–53. JOUR. http://doi.org/10.1177/1073858410386614
Helfrich, R. F., Schneider, T. R., Rach, S., Trautmann-Lengsfeld, S. A., Engel, A. K., & Herrmann, C. S. 119
(2014). Entrainment of Brain Oscillations by Transcranial Alternating Current Stimulation. Current
Biology, 24(3), 333–339. JOUR. https://doi.org/10.1016/j.cub.2013.12.041
Chaieb, L., Paulus, W., & Antal, A. (2011). Evaluating aftereffects of short-duration transcranial 120
random noise stimulation on cortical excitability. Neural plasticity, 2011.
Shiozawa, P., Fregni, F., Benseñor, I. M., Lotufo, P. A., Berlim, M. T., Daskalakis, J. Z., … Brunoni, A. 121
R. (2014). Transcranial direct current stimulation for major depression: an updated systematic review and meta-analysis. International Journal of Neuropsychopharmacology, 17(9), 1443–1452. JOUR. http://doi.org/10.1017/S1461145714000418
Karabanov, A. N., Saturnino, G. B., Thielscher, A., & Siebner, H. R. (2019). Can Transcranial Electrical 122
Stimulation Localize Brain Function? . In Frontiers in Psychology (Vol. 10, p. 213). https://www.frontiersin.org/article/10.3389/fpsyg.2019.00213
Dmochowski, J. P., Datta, A., Bikson, M., Su, Y., & Parra, L. C. (2011). Optimized multi-electrode 123
stimulation increases focality and intensity at target. Journal of Neural Engineering, 8(4), 46011. https://doi.org/10.1088/1741-2560/8/4/046011
Edwards, D., Cortes, M., Datta, A., Minhas, P., Wassermann, E. M., & Bikson, M. (2013). Physiological 124
and modeling evidence for focal transcranial electrical brain stimulation in humans: A basis for high-definition tDCS. NeuroImage, 74, 266–275. https://doi.org/10.1016/j.neuroimage.2013.01.042
Villamar, M. F., Wivatvongvana, P., Patumanond, J., … ., & Fregni, F. (2013). Focal Modulation of the 125
Primary Motor Cortex in Fibromyalgia Using 4x1Ring High-Definition Transcranial Direct Current Stimulation (HD-tDCS): Immediate and Delayed Analgesic Effects of Cathodal and Anodal Stimulation. The Journal of Pain, 14(4), 371–383. https://doi.org/10.1016/j.jpain.2012.12.007
Vöröslakos, M., Takeuchi, Y., Brinyiczki, K., Zombori, T., Oliva, A., Fernández-Ruiz, A., Kozák, G., 126
Kincses, Z. T., Iványi, B., Buzsáki, G., & Berényi, A. (2018). Direct effects of transcranial electric stimulation on brain circuits in rats and humans. Nature Communications, 9(1), 483. https://doi.org/10.1038/s41467-018-02928-3
Grossman, N., Bono, D., Dedic, N., Kodandaramaiah, S. B., Rudenko, A., Suk, H. J., Cassara, A. M., 127
Neufeld, E., Kuster, N., Tsai, L. H., Pascual-Leone, A., & Boyden, E. S. (2017). Noninvasive Deep Brain Stimulation via Temporally Interfering Electric Fields. Cell, 169(6), 1029–1041.e16. https://doi.org/10.1016/j.cell.2017.05.024
Hutcheon, B., & Yarom, Y. (2000). Resonance, oscillation and the intrinsic frequency preferences of 128
neurons. Trends in neurosciences, 23(5), 216–222. https://doi.org/10.1016/s0166-2236(00)01547-2
Buzsáki, G., & Draguhn, A. (2004). Neuronal Oscillations in Cortical Networks. Science, 304(5679), 129
1926 LP-1929. JOUR. http://doi.org/10.1126/science.1099745
Koessler, L., Benar, C., Maillard, L., Badier, J.-M., Vignal, J. P., Bartolomei, F., … Gavaret, M. (2010). 130
Source localization of ictal epileptic activity investigated by high resolution EEG and validated by SEEG. NeuroImage, 51(2), 642–653. JOUR. https://doi.org/10.1016/j.neuroimage.2010.02.067
Bartolomei, F., Chauvel, P., & Wendling, F. (2008). Epileptogenicity of brain structures in human 131
temporal lobe epilepsy: a quantified study from intracerebral EEG. Brain, 131(7), 1818–1830. JOUR. http://doi.org/10.1093/brain/awn111
Guye, M., Régis, J., Tamura, M., Wendling, F., Gonigal, A. M., Chauvel, P., & Bartolomei, F. (2006). The 132
role of corticothalamic coupling in human temporal lobe epilepsy. Brain, 129(7), 1917–1928. JOUR. http://doi.org/10.1093/brain/awl151
Munari, C., Kahane, P., Tassi, L., Francione, S., Hoffmann, D., Russo, G. Lo, & Benabid, A. L. (1993). 133
Intracerebral Low Frequency Electrical Stimulation: a New Tool for the Definition of the “Epileptogenic Area”? BT - Advances in Stereotactic and Functional Neurosurgery 10. In B. A. Meyerson, G. Broggi, J. Martin-Rodriguez, C. Ostertag, & M. Sindou (Eds.) (pp. 181–185). CONF, Vienna: Springer Vienna.

Bartolomei, F., Wendling, F., Bellanger, J.-J., Régis, J., & Chauvel, P. (2001). Neural networks involving 134
the medial temporal structures in temporal lobe epilepsy. Clinical Neurophysiology, 112(9), 1746–1760. JOUR. https://doi.org/10.1016/S1388-2457(01)00591-0
Allen, P. J., Fish, D. R., & Smith, S. J. M. (1992). Very high-frequency rhythmic activity during SEEG 135
suppression in frontal lobe epilepsy. Electroencephalography and Clinical Neurophysiology, 82(2), 155–159. JOUR. https://doi.org/10.1016/0013-4694(92)90160-J
Wendling, F., Bartolomei, F., Bellanger, J. J., Bourien, J., & Chauvel, P. (2003). Epileptic fast 136
intracerebral EEG activity: evidence for spatial decorrelation at seizure onset. Brain, 126(6), 1449–1459. JOUR. http://doi.org/10.1093/brain/awg144
Frauscher, B., Bartolomei, F., Kobayashi, K., Cimbalnik, J., van ‘t Klooster, M.A., Rampp, S., Otsubo, 137
H., Höller, Y., Wu, J.Y., Asano, E., Engel, J., Jr., Kahane, P., Jacobs, J. and Gotman, J. (2017), High‐frequency oscillations: The state of clinical research. Epilepsia, 58: 1316-1329. doi:10.1111/epi.13829
Alarcon, G., Binnie, C. D., Elwes, R. D. C., & Polkey, C. E. (1995). Power spectrum and intracranial 138
EEG patterns at seizure onset in partial epilepsy. Electroencephalography and Clinical Neurophysiology, 94(5), 326–337. JOUR. https://doi.org/10.1016/0013-4694(94)00286-T
Cuello Oderiz, C., von Ellenrieder, N., Dubeau, F., Eisenberg, A., Gotman, J., Hall, J., … Frauscher, B. 139
(2019). Association of Cortical Stimulation–Induced Seizure With Surgical Outcome in Patients With Focal Drug-Resistant Epilepsy. JAMA Neurology, 76(9), 1070–1078. JOUR. http://doi.org/10.1001/jamaneurol.2019.1464
Reinacher, P. C., Krüger, M. T., Coenen, V. A., Shah, M., Roelz, R., Jenkner, C., & Egger, K. (2017). 140
Determining the Orientation of Directional Deep Brain Stimulation Electrodes Using 3D Rotational Fluoroscopy. American Journal of Neuroradiology, 38(6), 1111 LP-1116. JOUR. http://doi.org/10.3174/ajnr.A5153
Lehto, L. J., Slopsema, J. P., Johnson, M. D., Shatillo, A., Teplitzky, B. A., Utecht, L., … Michaeli, S. 141
(2017). Orientation selective deep brain stimulation. Journal of Neural Engineering, 14(1), 16016. JOUR. http://doi.org/10.1088/1741-2552/aa5238
Bancaud, J., & Talairach, J. (1992). Clinical semiology of frontal lobe seizures. Advances in neurology, 142
57, 3–58.
Lagarde, S., Bonini, F., McGonigal, A., Chauvel, P., Gavaret, M., Scavarda, D., Carron, R., Régis, J., 143
Aubert, S., Villeneuve, N., Giusiano, B., Figarella‐Branger, D., Trebuchon, A. and Bartolomei, F. (2016), Seizure‐onset patterns in focal cortical dysplasia and neurodevelopmental tumors: Relationship with surgical prognosis and neuropathologic subtypes. Epilepsia, 57: 1426-1435. doi:10.1111/epi.13464
Perucca, P., Dubeau, F., & Gotman, J. (2014). Intracranial electroencephalographic seizure-onset 144
patterns: effect of underlying pathology. Brain : a journal of neurology, 137(Pt 1), 183–196. https://doi.org/10.1093/brain/awt299
Bartolomei, F., Lagarde, S., Wendling, F., McGonigal, A., Jirsa, V., Guye, M. and Bénar, C. (2017), 145
Defining epileptogenic networks: Contribution of SEEG and signal analysis. Epilepsia, 58: 1131-1147. doi:10.1111/epi.13791
Uddin, L. Q., Nomi, J. S., Hébert-Seropian, B., Ghaziri, J., & Boucher, O. (2017). Structure and 146
Function of the Human Insula. Journal of Clinical Neurophysiology, 34(4). https://journals.lww.com/clinicalneurophys/Fulltext/2017/07000/Structure_and_Function_of_the_Human_Insula.2.aspx
Nguyen, D. K., Nguyen, D. B., Malak, R., Leroux, J. M., Carmant, L., Saint-Hilaire, J. M., Giard, N., 147
Cossette, P., & Bouthillier, A. (2009). Revisiting the role of the insula in refractory partial epilepsy. Epilepsia, 50(3), 510–520. https://doi.org/10.1111/j.1528-1167.2008.01758.x
Isnard, J., & Mauguière, F. (2005). Le lobe de l'insula et les épilepsies partielles [The insula in partial 148
epilepsy]. Revue neurologique, 161(1), 17–26. https://doi.org/10.1016/s0035-3787(05)84970-7
Ryvlin, P., Minotti, L., Demarquay, G., Hirsch, E., Arzimanoglou, A., Hoffman, D., Guénot, M., Picard, 149
F., Rheims, S., & Kahane, P. (2006). Nocturnal hypermotor seizures, suggesting frontal lobe epilepsy, can originate in the insula. Epilepsia, 47(4), 755–765. https://doi.org/10.1111/j.1528-1167.2006.00510.x

Bourdillon, P., Ryvlin, P., Isnard, J., Montavont, A., Catenoix, H., Mauguière, F., … Guénot, M. (2017). 150
Stereotactic Electroencephalography Is a Safe Procedure, Including for Insular Implantations. World
Neurosurgery, 99, 353–361. JOUR. https://doi.org/10.1016/j.wneu.2016.12.025
Schultz, S.R. and Rolls, E.T. (1999), Analysis of information transmission in the schaffer collaterals. 151
Hippocampus, 9: 582-598. doi:10.1002/(SICI)1098-1063(1999)9:5<582::AID-HIPO12>3.0.CO;2-P
Barbarosie, M., & Avoli, M. (1997). CA3-Driven Hippocampal-Entorhinal Loop Controls Rather than 152
Sustains <em>In Vitro</em> Limbic Seizures. The Journal of Neuroscience, 17(23), 9308 LP-9314. JOUR. http://doi.org/10.1523/JNEUROSCI.17-23-09308.1997
Pelletier, M. R., Wadia, J. S., Mills, L. R., & Carlen, P. L. (1999). Seizure-induced cell death produced 153
by repeated tetanic stimulation in vitro: possible role of endoplasmic reticulum calcium stores. Journal of
neurophysiology, 81(6), 3054–3064. https://doi.org/10.1152/jn.1999.81.6.3054
Engel, J. (2001). Mesial Temporal Lobe Epilepsy: What Have We Learned? The Neuroscientist, 7(4), 154
340–352. JOUR. http://doi.org/10.1177/107385840100700410
Berkovic, S. F., Andermann, F., Olivier, A., Ethier, R., Melanson, D., Robitaille, Y., … Feindel, W. (1991). 155
Hippocampal sclerosis in temporal lobe epilepsy demonstrated by magnetic resonance imaging. Annals of Neurology, 29(2), 175–182. JOUR. http://doi.org/10.1002/ana.410290210
Sutula, T., Cascino, G., Cavazos, J., Parada, I., & Ramirez, L. (1989). Mossy fiber synaptic 156
reorganization in the epileptic human temporal lobe. Annals of Neurology, 26(3), 321–330. JOUR. http://doi.org/10.1002/ana.410260303
Muñoz, A., Ballesteros‐Yáñez, I., DeFelipe, J., Arellano, J. I., & Sola, R. G. (2004). Histopathology and 157
reorganization of chandelier cells in the human epileptic sclerotic hippocampus. Brain, 127(1), 45–64. JOUR. http://doi.org/10.1093/brain/awh004
Tóth, K., Eross, L., Vajda, J., Halász, P., Freund, T. F., & Maglóczky, Z. (2010). Loss and reorganization 158
of calretinin-containing interneurons in the epileptic human hippocampus. Brain : A Journal of Neurology, 133(9), 2763–2777. JOUR. http://doi.org/10.1093/brain/awq149
Sloviter, R. S., Sollas, A. L., Barbaro, N. M., & Laxer, K. D. (1991). Calcium-binding protein (calbindin-159
D28K) and parvalbumin immunocytochemistry in the normal and epileptic human hippocampus. Journal of Comparative Neurology, 308(3), 381–396. JOUR. http://doi.org/10.1002/cne.903080306
Vezzani, A., & Sperk, G. (2004). Overexpression of NPY and Y2 receptors in epileptic brain tissue: an 160
endogenous neuroprotective mechanism in temporal lobe epilepsy? Neuropeptides, 38(4), 245–252. JOUR. https://doi.org/10.1016/j.npep.2004.05.004
Kharlamov, E. A., Kharlamov, A., & Kelly, K. M. (2007). Changes in neuropeptide Y protein expression 161
following photothrombotic brain infarction and epileptogenesis. Brain Research, 1127(1), 151–162. JOUR. http://doi.org/10.1016/j.brainres.2006.09.107
Casillas-Espinosa, P. M., Powell, K. L., & O’Brien, T. J. (2012). Regulators of synaptic transmission: 162
Roles in the pathogenesis and treatment of epilepsy. Epilepsia, 53(s9), 41–58. JOUR. http://doi.org/10.1111/epi.12034
MD, I. B., MRCPath, M. T., & Wiestler, O. D. (2002). Ammon’s Horn Sclerosis: A Maldevelopmental 163
Disorder Associated with Temporal Lobe Epilepsy. Brain Pathology, 12(2), 199–211. JOUR. http://doi.org/10.1111/j.1750-3639.2002.tb00436.x
Crespel, A., Lebrun, A., de Bock, F., Bockaert, J., Picot, M.-C., Rousset, M.-C., … Rigau, V. (2007). 164
Angiogenesis is associated with blood–brain barrier permeability in temporal lobe epilepsy. Brain, 130(7), 1942–1956. JOUR. http://doi.org/10.1093/brain/awm118
Turski, W. A., Cavalheiro, E. A., Schwarz, M., Czuczwar, S. J., Kleinrok, Z., & Turski, L. (1983). Limbic 165
seizures produced by pilocarpine in rats: Behavioural, electroencephalographic and neuropathological study. Behavioural Brain Research, 9(3), 315–335. JOUR. http://doi.org/https://doi.org/10.1016/0166-4328(83)90136-5
Covolan, L., & Mello, L. E. A. . (2000). Temporal profile of neuronal injury following pilocarpine or 166
kainic acid-induced status epilepticus. Epilepsy Research, 39(2), 133–152. JOUR. https://doi.org/10.1016/S0920-1211(99)00119-9

Lévesque, M., & Avoli, M. (2013). The kainic acid model of temporal lobe epilepsy. Neuroscience & 167
Biobehavioral Reviews, 37(10, Part 2), 2887–2899. https://doi.org/10.1016/j.neubiorev.2013.10.011
Curia, G., Longo, D., Biagini, G., Jones, R. S. G., & Avoli, M. (2008). The pilocarpine model of 168
temporal lobe epilepsy. Journal of Neuroscience Methods, 172(2), 143–157. JOUR. https://doi.org/10.1016/j.jneumeth.2008.04.019
Inostroza, M., Cid, E., Brotons-Mas, J., Gal, B., Aivar, P., Uzcategui, Y. G., … Menendez de la Prida, 169
L. (2011). Hippocampal-Dependent Spatial Memory in the Water Maze is Preserved in an Experimental Model of Temporal Lobe Epilepsy in Rats. PLOS ONE, 6(7), e22372. JOUR. https://doi.org/10.1371/journal.pone.0022372
Roux, E., & Borrel, A. (1898). Tétanos cérébral et immunité contre le tétanos. Ann. Inst. Pasteur, 12, 170
225–239.
JEFFERYS, J. G. R., & WALKER, M. C. (2006). Tetanus toxin model of focal epilepsy. In Models of 171
seizures and epilepsy (pp. 407–414). CHAP, Elsevier.
Benke, T. A., & Swann, J. (2004). The Tetanus Toxin Model of Chronic Epilepsy BT - Recent 172
Advances in Epilepsy Research. In D. K. Binder & H. E. Scharfman (Eds.) (pp. 226–238). CHAP, Boston, MA: Springer US. http://doi.org/10.1007/978-1-4757-6376-8_16
GODDARD, G. V. (1967). Development of Epileptic Seizures through Brain Stimulation at Low 173
Intensity. Nature, 214(5092), 1020–1021. JOUR. http://doi.org/10.1038/2141020a0
Goddard, G. V, McIntyre, D. C., & Leech, C. K. (1969). A permanent change in brain function resulting 174
from daily electrical stimulation. Experimental Neurology, 25(3), 295–330. JOUR. https://doi.org/10.1016/0014-4886(69)90128-9
Kairiss, E. W., Racine, R. J., & Smith, G. K. (1984). The development of the interictal spike during 175
kindling in the rat. Brain Research, 322(1), 101–110. https://doi.org/10.1016/0006-8993(84)91185-5
Palizvan, M. R., Fathollahi, Y., Semnanian, S., Hajezadeh, S., & Mirnajafizadh, J. (2001). Differential 176
effects of pentylenetetrazol-kindling on long-term potentiation of population excitatory postsynaptic potentials and population spikes in the CA1 region of rat hippocampus. Brain Research, 898(1), 82–90. JOUR. https://doi.org/10.1016/S0006-8993(01)02146-1
LÖSCHER, W., & EBERT, U. (1996). THE ROLE OF THE PIRIFORM CORTEX IN KINDLING. Progress 177
in Neurobiology, 50(5), 427–481. JOUR. https://doi.org/10.1016/S0301-0082(96)00036-6
McIntyre, D. C., Kelly, M. E., & Armstrong, J. N. (1993). Kindling in the perirhinal cortex. Brain 178
Research, 615(1), 1–6. JOUR. https://doi.org/10.1016/0006-8993(93)91108-5
Morimoto, K., Fahnestock, M., & Racine, R. J. (2004). Kindling and status epilepticus models of 179
epilepsy: rewiring the brain. Progress in Neurobiology, 73(1), 1–60. JOUR. https://doi.org/10.1016/j.pneurobio.2004.03.009
Cela, E., McFarlan, A. R., Chung, A. J., Wang, T., Chierzi, S., Murai, K. K., & Sjöström, P. J. (2019). An 180
Optogenetic Kindling Model of Neocortical Epilepsy. Scientific Reports, 9(1), 5236. JOUR. http://doi.org/10.1038/s41598-019-41533-2
Pitkänen, A., & Mcintosh, T. K. (2006). Animal Models of Post-Traumatic Epilepsy. Journal of 181
Neurotrauma, 23(2), 241–261. JOUR. http://doi.org/10.1089/neu.2006.23.241
D’Ambrosio, R., Fender, J. S., Fairbanks, J. P., Simon, E. A., Born, D. E., Doyle, D. L., & Miller, J. W. 182
(2004). Progression from frontal–parietal to mesial–temporal epilepsy after fluid percussion injury in the rat. Brain, 128(1), 174–188. JOUR. http://doi.org/10.1093/brain/awh337
Kharatishvili, I., Nissinen, J. P., McIntosh, T. K., & Pitkänen, A. (2006). A model of posttraumatic 183
epilepsy induced by lateral fluid-percussion brain injury in rats. Neuroscience, 140(2), 685–697. JOUR. https://doi.org/10.1016/j.neuroscience.2006.03.012
Pitkänen, A., & Immonen, R. (2014). Epilepsy Related to Traumatic Brain Injury. Neurotherapeutics, 184
11(2), 286–296. JOUR. http://doi.org/10.1007/s13311-014-0260-7

French, J. A., Williamson, P. D., Thadani, V. M., Darcey, T. M., Mattson, R. H., Spencer, S. S., & 185
Spencer, D. D. (1993). Characteristics of medial temporal lobe epilepsy: I. Results of history and physical examination. Annals of Neurology, 34(6), 774–780. JOUR. http://doi.org/10.1002/ana.410340604
Holtzman, D., Obana, K., & Olson, J. (1981). Hyperthermia-induced seizures in the rat pup: a model 186
for febrile convulsions in children. Science, 213(4511), 1034 LP-1036. JOUR. http://doi.org/10.1126/science.7268407
Dubé, C. M., Ravizza, T., Hamamura, M., Zha, Q., Keebaugh, A., Fok, K., … Baram, T. Z. (2010). 187
Epileptogenesis Provoked by Prolonged Experimental Febrile Seizures: Mechanisms and Biomarkers. The Journal of Neuroscience, 30(22), 7484 LP-7494. http://doi.org/10.1523/JNEUROSCI.0551-10.2010
Jensen, F. E., Applegate, C. D., Holtzman, D., Belin, T. R., & Burchfiel, J. L. (1991). Epileptogenic 188
effect of hypoxia in the immature rodent brain. Annals of Neurology, 29(6), 629–637. JOUR. http://doi.org/10.1002/ana.410290610
AICARDI, J., & CHEVRIE, J. J. (1970). Convulsive Status Epilepticus in Infants and Children. 189
Epilepsia, 11(2), 187–197. JOUR. http://doi.org/10.1111/j.1528-1157.1970.tb03880.x
Rakhade, S. N., Klein, P. M., Huynh, T., Hilario-Gomez, C., Kosaras, B., Rotenberg, A., & Jensen, F. E. 190
(2011). Development of later life spontaneous seizures in a rodent model of hypoxia-induced neonatal seizures. Epilepsia, 52(4), 753–765. JOUR. http://doi.org/10.1111/j.1528-1167.2011.02992.x
NADLER, J. V., PERRY, B. W., & COTMAN, C. W. (1978). Intraventricular kainic acid preferentially 191
destroys hippocampal pyramidal cells. Nature, 271(5646), 676–677. http://doi.org/10.1038/271676a0
Ben-Ari, Y., & Lagowska, J. (1978). [Epileptogenic action of intra-amygdaloid injection of kainic acid]. 192
Comptes rendus hebdomadaires des seances de l’Academie des sciences. Serie D: Sciences naturelles, 287(8), 813–816. JOUR. http://europepmc.org/abstract/MED/103652
Ben-Ari, Y., Lagowska, J., Tremblay, E., & Le Gal La Salle, G. (1979). A new model of focal status 193
epilepticus: intra-amygdaloid application of kainic acid elicits repetitive secondarily generalized convulsive seizures. Brain Research, 163(1), 176–179. https://doi.org/10.1016/0006-8993(79)90163-X
Patel, S., Meldrum, B. S., & Collins, J. F. (1986). Distribution of [3H]kainic acid and binding sites in the 194
rat brain: in vivo and in vitro receptor autoradiography. Neuroscience Letters, 70(3), 301–307. JOUR. https://doi.org/10.1016/0304-3940(86)90569-0
Wisden, W., & Seeburg, P. H. (1993). A complex mosaic of high-affinity kainate receptors in rat brain. 195
The Journal of Neuroscience, 13(8), 3582 LP-3598. JOUR. http://doi.org/10.1523/JNEUROSCI.13-08-03582.1993
ROGAWSKI, M. A., GRYDER, D., CASTANEDA, D., YONEKAWA, W., BANKS, M. K., & LI, H. E. 196
(2003). GluR5 Kainate Receptors, Seizures, and the Amygdala. Annals of the New York Academy of Sciences, 985(1), 150–162. JOUR. http://doi.org/10.1111/j.1749-6632.2003.tb07079.x
Jin, X.-T., & Smith, Y. (2011). Localization and functions of kainate receptors in the basal ganglia. 197
Advances in Experimental Medicine and Biology, 717, 27–37. JOUR. http://doi.org/10.1007/978-1-4419-9557-5_3
Bloss, E. B., & Hunter, R. G. (2010). Chapter 9 - Hippocampal Kainate Receptors. In G. B. T.-V. & H. 198
Litwack (Ed.), Hormones of the Limbic System (Vol. 82, pp. 167–184). CHAP, Academic Press. https://doi.org/10.1016/S0083-6729(10)82009-6
Lerma, J., & Marques, J. M. (2013). Kainate Receptors in Health and Disease. Neuron, 80(2), 292–199
311. JOUR. https://doi.org/10.1016/j.neuron.2013.09.045
Huettner, J. E. (2003). Kainate receptors and synaptic transmission. Progress in Neurobiology, 70(5), 200
387–407. JOUR. https://doi.org/10.1016/S0301-0082(03)00122-9
Cossart, R., Dinocourt, C., Hirsch, J. C., Merchan-Perez, A., De Felipe, J., Ben-Ari, Y., … Bernard, C. 201
(2001). Dendritic but not somatic GABAergic inhibition is decreased in experimental epilepsy. Nature Neuroscience, 4(1), 52–62. JOUR. http://doi.org/10.1038/82900
Cossart, R., Tyzio, R., Dinocourt, C., Esclapez, M., Hirsch, J. C., Ben-Ari, Y., & Bernard, C. (2001). 202
Presynaptic Kainate Receptors that Enhance the Release of GABA on CA1 Hippocampal Interneurons. Neuron, 29(2), 497–508. JOUR. https://doi.org/10.1016/S0896-6273(01)00221-5

Bahn, S., Volk, B., & Wisden, W. (1994). Kainate receptor gene expression in the developing rat brain. 203
The Journal of Neuroscience, 14(9), 5525 LP-5547. JOUR. http://doi.org/10.1523/JNEUROSCI.14-09-05525.1994
Darstein, M., Petralia, R. S., Swanson, G. T., Wenthold, R. J., & Heinemann, S. F. (2003). Distribution 204
of Kainate Receptor Subunits at Hippocampal Mossy Fiber Synapses. The Journal of Neuroscience, 23(22), 8013 LP-8019. JOUR. http://doi.org/10.1523/JNEUROSCI.23-22-08013.2003
Gallyas Jr, F., Ball, S. M., & Molnar, E. (2003). Assembly and cell surface expression of KA-2 subunit-205
containing kainate receptors. Journal of Neurochemistry, 86(6), 1414–1427. JOUR. http://doi.org/10.1046/j.1471-4159.2003.01945.x
Fernandes, H. B., Catches, J. S., Petralia, R. S., Copits, B. A., Xu, J., Russell, T. A., … Contractor, A. 206
(2009). High-Affinity Kainate Receptor Subunits Are Necessary for Ionotropic but Not Metabotropic Signaling. Neuron, 63(6), 818–829. JOUR. https://doi.org/10.1016/j.neuron.2009.08.010
Fisahn, A., Contractor, A., Traub, R. D., Buhl, E. H., Heinemann, S. F., & McBain, C. J. (2004). Distinct 207
Roles for the Kainate Receptor Subunits GluR5 and GluR6 in Kainate-Induced Hippocampal Gamma Oscillations. The Journal of Neuroscience, 24(43), 9658 LP-9668. JOUR. http://doi.org/10.1523/JNEUROSCI.2973-04.2004
Mulle, C., Sailer, A., Pérez-Otaño, I., Dickinson-Anson, H., Castillo, P. E., Bureau, I., … Heinemann, S. 208
F. (1998). Altered synaptic physiology and reduced susceptibility to kainate-induced seizures in GluR6-deficient mice. Nature, 392(6676), 601–605. JOUR. http://doi.org/10.1038/33408
Ullal, G., Fahnestock, M., & Racine, R. (2005). Time-dependent Effect of Kainate-induced Seizures on 209
Glutamate Receptor GluR5, GluR6, and GluR7 mRNA and Protein Expression in Rat Hippocampus. Epilepsia, 46(5), 616–623. JOUR. http://doi.org/10.1111/j.1528-1167.2005.49604.x
Dong, X., Wang, Y., & Qin, Z. (2009). Molecular mechanisms of excitotoxicity and their relevance to 210
pathogenesis of neurodegenerative diseases. Acta Pharmacologica Sinica, 30(4), 379–387. JOUR. http://doi.org/10.1038/aps.2009.24
Beck, J., Lenart, B., Kintner, D. B., & Sun, D. (2003). Na-K-Cl Cotransporter Contributes to 211
Glutamate-Mediated Excitotoxicity. The Journal of Neuroscience, 23(12), 5061 LP-5068. JOUR. http://doi.org/10.1523/JNEUROSCI.23-12-05061.2003
Friedman, L. K. (2006). Calcium: a role for neuroprotection and sustained adaptation. Molecular 212
Interventions, 6(6), 315–329. JOUR. http://doi.org/10.1124/mi.6.6.5
Lee, B. K., Lee, D. H., Park, S., Park, S. L., Yoon, J.-S., Lee, M. G., … Jung, Y.-S. (2009). Effects of 213
KR-33028, a novel Na+/H+ exchanger-1 inhibitor, on glutamate-induced neuronal cell death and ischemia-induced cerebral infarct. Brain Research, 1248, 22–30. JOUR. https://doi.org/10.1016/j.brainres.2008.10.061
Chen, Q., Olney, J. W., Lukasiewicz, P. D., Almli, T., & Romano, C. (1998). Ca+-Independent 214
Excitotoxic Neurodegeneration in Isolated Retina, an Intact Neural Net: A Role for Cl− and Inhibitory Transmitters. Molecular Pharmacology, 53(3), 564 LP-572. JOUR. http://doi.org/10.1124/mol.53.3.564
Ha, B. K., Vicini, S., Rogers, R. C., Bresnahan, J. C., Burry, R. W., & Beattie, M. S. (2002). Kainate-215
induced excitotoxicity is dependent upon extracellular potassium concentrations that regulate the activity of AMPA/KA type glutamate receptors. Journal of Neurochemistry, 83(4), 934–945. JOUR. http://doi.org/10.1046/j.1471-4159.2002.01203.x
Murphy, T. H., Miyamoto, M., Sastre, A., Schnaar, R. L., & Coyle, J. T. (1989). Glutamate toxicity in a 216
neuronal cell line involves inhibition of cystine transport leading to oxidative stress. Neuron, 2(6), 1547–1558. JOUR. https://doi.org/10.1016/0896-6273(89)90043-3
Nguyen, D., Alavi, M. V, Kim, K.-Y., Kang, T., Scott, R. T., Noh, Y. H., … Ju, W.-K. (2011). A new 217
vicious cycle involving glutamate excitotoxicity, oxidative stress and mitochondrial dynamics. Cell Death & Disease, 2(12), e240–e240. JOUR. http://doi.org/10.1038/cddis.2011.117
Chuang, Y.-C., Chang, A. Y. W., Lin, J.-W., Hsu, S.-P., & Chan, S. H. H. (2004). Mitochondrial 218
Dysfunction and Ultrastructural Damage in the Hippocampus during Kainic Acid–induced Status Epilepticus in the Rat. Epilepsia, 45(10), 1202–1209. http://doi.org/10.1111/j.0013-9580.2004.18204.x

Gano, L. B., Liang, L.-P., Ryan, K., Michel, C. R., Gomez, J., Vassilopoulos, A., … Patel, M. (2018). 219
Altered mitochondrial acetylation profiles in a kainic acid model of temporal lobe epilepsy. Free Radical Biology and Medicine, 123, 116–124. JOUR. https://doi.org/10.1016/j.freeradbiomed.2018.05.063
Rowley, S., & Patel, M. (2013). Mitochondrial involvement and oxidative stress in temporal lobe 220
epilepsy. Free Radical Biology & Medicine, 62, 121–131. JOUR. http://doi.org/10.1016/j.freeradbiomed.2013.02.002
Schwob, J. E., Fuller, T., Price, J. L., & Olney, J. W. (1980). Widespread patterns of neuronal damage 221
following systemic or intracerebral injections of kainic acid: A histological study. Neuroscience, 5(6), 991–1014. JOUR. https://doi.org/10.1016/0306-4522(80)90181-5
Sperk, G., Lassmann, H., Baran, H., Kish, S. J., Seitelberger, F., & Hornykiewicz, O. (1983). Kainic 222
acid induced seizures: Neurochemical and histopathological changes. Neuroscience, 10(4), 1301–1315. JOUR. https://doi.org/10.1016/0306-4522(83)90113-6
Bertoglio, D., Amhaoul, H., Van Eetveldt, A., Houbrechts, R., Van De Vijver, S., Ali, I., & 223
Dedeurwaerdere, S. (2017). Kainic Acid-Induced Post-Status Epilepticus Models of Temporal Lobe Epilepsy with Diverging Seizure Phenotype and Neuropathology. Frontiers in Neurology, 8, 588. JOUR. http://doi.org/10.3389/fneur.2017.00588
McKhann, G. M., Wenzel, H. J., Robbins, C. A., Sosunov, A. A., & Schwartzkroin, P. A. (2003). Mouse 224
strain differences in kainic acid sensitivity, seizure behavior, mortality, and hippocampal pathology. Neuroscience, 122(2), 551–561. JOUR. https://doi.org/10.1016/S0306-4522(03)00562-1
El Houssaini, K., Ivanov, A. I., Bernard, C., & Jirsa, V. K. (2015). Seizures, refractory status epilepticus, 225
and depolarization block as endogenous brain activities. Physical review. E, Statistical, nonlinear, and
soft matter physics, 91(1), 010701. https://doi.org/10.1103/PhysRevE.91.010701
Trinka, E., Cock, H., Hesdorffer, D., Rossetti, A. O., Scheffer, I. E., Shinnar, S., … Lowenstein, D. H. 226
(2015). A definition and classification of status epilepticus – Report of the ILAE Task Force on Classification of Status Epilepticus. Epilepsia, 56(10), 1515–1523. http://doi.org/10.1111/epi.13121
Racine, R. J. (1972). Modification of seizure activity by electrical stimulation: II. Motor seizure. 227
Electroencephalography and Clinical Neurophysiology, 32(3), 281–294. JOUR. https://doi.org/10.1016/0013-4694(72)90177-0
Raedt, R., Van Dycke, A., Van Melkebeke, D., De Smedt, T., Claeys, P., Wyckhuys, T., … Boon, P. 228
(2009). Seizures in the intrahippocampal kainic acid epilepsy model: characterization using long-term video-EEG monitoring in the rat. Acta Neurologica Scandinavica, 119(5), 293–303. JOUR. http://doi.org/10.1111/j.1600-0404.2008.01108.x
Pernot, F., Heinrich, C., Barbier, L., Peinnequin, A., Carpentier, P., Dhote, F., … Dorandeu, F. (2011). 229
Inflammatory changes during epileptogenesis and spontaneous seizures in a mouse model of mesiotemporal lobe epilepsy. Epilepsia, 52(12), 2315–2325. JOUR. http://doi.org/10.1111/j.1528-1167.2011.03273.x
Stafstrom, C. E., Thompson, J. L., & Holmes, G. L. (1992). Kainic acid seizures in the developing 230
brain: status epilepticus and spontaneous recurrent seizures. Developmental Brain Research, 65(2), 227–236. JOUR. https://doi.org/10.1016/0165-3806(92)90184-X
Giorgi, F. S., Malhotra, S., Hasson, H., Velíšková, J., Rosenbaum, D. M., & Moshé, S. L. (2005). 231
Effects of Status Epilepticus Early in Life on Susceptibility to Ischemic Injury in Adulthood. Epilepsia, 46(4), 490–498. JOUR. http://doi.org/10.1111/j.0013-9580.2005.42304.x
Lado, F. A. (2006). Chronic Bilateral Stimulation of the Anterior Thalamus of Kainate-treated Rats 232
Increases Seizure Frequency. Epilepsia, 47(1), 27–32. http://doi.org/10.1111/j.1528-1167.2006.00366.x
Williams, P. A., White, A. M., Clark, S., Ferraro, D. J., Swiercz, W., Staley, K. J., & Dudek, F. E. (2009). 233
Development of Spontaneous Recurrent Seizures after Kainate-Induced Status Epilepticus. The Journal of Neuroscience, 29(7), 2103 LP-2112. JOUR. http://doi.org/10.1523/JNEUROSCI.0980-08.2009
White, A., Williams, P. A., Hellier, J. L., Clark, S., Dudek, F. E., & Staley, K. J. (2010). EEG spike activity 234
precedes epilepsy after kainate-induced status epilepticus. Epilepsia, 51(3), 371–383. JOUR. http://doi.org/10.1111/j.1528-1167.2009.02339.x

Drexel, M., Preidt, A. P., & Sperk, G. (2012). Sequel of spontaneous seizures after kainic acid-induced 235
status epilepticus and associated neuropathological changes in the subiculum and entorhinal cortex. Neuropharmacology, 63(5), 806–817. JOUR. http://doi.org/10.1016/j.neuropharm.2012.06.009
Hu, R. Q., Koh, S., Torgerson, T., & Cole, A. J. (1998). Neuronal stress and injury in C57/BL mice after 236
systemic kainic acid administration. Brain Research, 810(1), 229–240. JOUR. http://doi.org/10.1016/S0006-8993(98)00863-4
Benkovic, S. A., O’Callaghan, J. P., & Miller, D. B. (2004). Sensitive indicators of injury reveal 237
hippocampal damage in C57BL/6J mice treated with kainic acid in the absence of tonic–clonic seizures. Brain Research, 1024(1), 59–76. JOUR. http://doi.org/10.1016/j.brainres.2004.07.021
Puttachary, S., Sharma, S., Tse, K., Beamer, E., Sexton, A., Crutison, J., & Thippeswamy, T. (2015). 238
Immediate Epileptogenesis after Kainate-Induced Status Epilepticus in C57BL/6J Mice: Evidence from Long Term Continuous Video-EEG Telemetry. PLOS ONE, 10(7), e0131705. JOUR. https://doi.org/10.1371/journal.pone.0131705
Tse, K., Puttachary, S., Beamer, E., Sills, G. J., & Thippeswamy, T. (2014). Advantages of repeated 239
low dose against single high dose of kainate in C57BL/6J mouse model of status epilepticus: behavioral and electroencephalographic studies. PloS One, 9(5), e96622–e96622. JOUR. http://doi.org/10.1371/journal.pone.0096622
Hellier, J. L., & Dudek, F. E. (2005). Chemoconvulsant Model of Chronic Spontaneous Seizures. 240
Current Protocols in Neuroscience, 31(1), 9.19.1-9.19.12. http://doi.org/10.1002/0471142301.ns0919s31
Sharma, A. K., Jordan, W. H., Reams, R. Y., Hall, D. G., & Snyder, P. W. (2008). Temporal Profile of 241
Clinical Signs and Histopathologic Changes in an F-344 Rat Model of Kainic Acid–induced Mesial Temporal Lobe Epilepsy. Toxicologic Pathology, 36(7), 932–943. JOUR. http://doi.org/10.1177/0192623308326093
Gurbanova, A. A., Aker, R. G., Sirvanci, S., Demiralp, T., & Onat, F. Y. (2008). Intra-Amygdaloid 242
Injection of Kainic Acid in Rats with Genetic Absence Epilepsy: The Relationship of Typical Absence Epilepsy and Temporal Lobe Epilepsy. The Journal of Neuroscience, 28(31), 7828 LP-7836. JOUR. http://doi.org/10.1523/JNEUROSCI.1097-08.2008
Mathern, G. W., Babb, T. L., Vickrey, B. G., Melendez, M., & Pretorius, J. K. (1995). The clinical-243
pathogenic mechanisms of hippocampal neuron loss and surgical outcomes in temporal lobe epilepsy. Brain, 118(1), 105–118. JOUR. http://doi.org/10.1093/brain/118.1.105
Chauvière, L., Doublet, T., Ghestem, A., Siyoucef, S. S., Wendling, F., Huys, R., … Bernard, C. (2012). 244
Changes in interictal spike features precede the onset of temporal lobe epilepsy. Annals of Neurology, 71(6), 805–814. JOUR. http://doi.org/10.1002/ana.23549
Williams, P., White, A., Ferraro, D., Clark, S., Staley, K., & Dudek, F. E. (2006). The use of 245
radiotelemetry to evaluate electrographic seizures in rats with kainate-induced epilepsy. Journal of Neuroscience Methods, 155(1), 39–48. JOUR. https://doi.org/10.1016/j.jneumeth.2005.12.035
Umpierre, A. D., Bennett, I. V, Nebeker, L. D., Newell, T. G., Tian, B. B., Thomson, K. E., … Wilcox, K. 246
S. (2016). Repeated low-dose kainate administration in C57BL/6J mice produces temporal lobe epilepsy pathology but infrequent spontaneous seizures. Experimental Neurology, 279, 116–126. JOUR. http://doi.org/10.1016/j.expneurol.2016.02.014
Rao, M. S., Hattiangady, B., Reddy, D. S., & Shetty, A. K. (2006). Hippocampal neurodegeneration, 247
spontaneous seizures, and mossy fiber sprouting in the F344 rat model of temporal lobe epilepsy. Journal of Neuroscience Research, 83(6), 1088–1105. JOUR. http://doi.org/10.1002/jnr.20802
Manouze, H., Ghestem, A., Poillerat, V., Bennis, M., Ba-M’hamed, S., Benoliel, J. J., … Bernard, C. 248
(2019). Effects of Single Cage Housing on Stress, Cognitive, and Seizure Parameters in the Rat and Mouse Pilocarpine Models of Epilepsy. Eneuro, 6(4), ENEURO.0179-18.2019. JOUR. http://doi.org/10.1523/ENEURO.0179-18.2019
Bernard, C. (2019). On the interpretation of results obtained in singly housed animals. Epilepsia, 249
60(10), 2013–2015. JOUR. http://doi.org/10.1111/epi.16347

Haas, K. Z., Sperber, E. F., Opanashuk, L. A., Stanton, P. K., & Moshé, S. L. (2001). Resistance of 250
immature hippocampus to morphologic and physiologic alterations following status epilepticus or kindling. Hippocampus, 11(6), 615–625. JOUR. http://doi.org/10.1002/hipo.1076
Kasugai, M., Akaike, K., Imamura, S., Matsukubo, H., Tojo, H., Nakamura, M., … Sano, A. (2007). 251
Differences in two mice strains on kainic acid-induced amygdalar seizures. Biochemical and Biophysical Research Communications, 357(4), 1078–1083. JOUR. https://doi.org/10.1016/j.bbrc.2007.04.067
Mouri, G., Jimenez-Mateos, E., Engel, T., Dunleavy, M., Hatazaki, S., Paucard, A., … Henshall, D. C. 252
(2008). Unilateral hippocampal CA3-predominant damage and short latency epileptogenesis after intra-amygdala microinjection of kainic acid in mice. Brain Research, 1213, 140–151. JOUR. https://doi.org/10.1016/j.brainres.2008.03.061
Kar, S., Seto, D., Doré, S., Chabot, J.-G., & Quirion, R. (1997). Systemic administration of kainic acid 253
induces selective time dependent decrease in [125I]insulin-like growth factor I, [125I]insulin-like growth factor II and [125I]insulin receptor binding sites in adult rat hippocampal formation. Neuroscience, 80(4), 1041–1055. JOUR. https://doi.org/10.1016/S0306-4522(97)00185-1
Suárez, L. M., Cid, E., Gal, B., Inostroza, M., Brotons-Mas, J. R., Gómez-Domínguez, D., … Solís, J. 254
M. (2012). Systemic Injection of Kainic Acid Differently Affects LTP Magnitude Depending on its Epileptogenic Efficiency. PLOS ONE, 7(10), e48128. https://doi.org/10.1371/journal.pone.0048128
Tauck, D. L., & Nadler, J. V. (1985). Evidence of functional mossy fiber sprouting in hippocampal 255
formation of kainic acid-treated rats. The Journal of Neuroscience, 5(4), 1016 LP-1022. JOUR. http://doi.org/10.1523/JNEUROSCI.05-04-01016.1985
Fritsch, B., Qashu, F., Figueiredo, T. H., Aroniadou-Anderjaska, V., Rogawski, M. A., & Braga, M. F. M. 256
(2009). Pathological alterations in GABAergic interneurons and reduced tonic inhibition in the basolateral amygdala during epileptogenesis. Neuroscience, 163(1), 415–429. JOUR. http://doi.org/10.1016/j.neuroscience.2009.06.034
Anthony Altar, C., & Baudry, M. (1990). Systemic injection of kainic acid: Gliosis in olfactory and 257
limbic brain regions quantified with [3H]PK 11195 binding autoradiography. Experimental Neurology, 109(3), 333–341. JOUR. https://doi.org/10.1016/S0014-4886(05)80024-X
Lancaster, B., & Wheal, H. V. (1982). A comparative histological and electrophysiological study of 258
some neurotoxins in the rat hippocampus. Journal of Comparative Neurology, 211(2), 105–114. JOUR. http://doi.org/10.1002/cne.902110202
Luo, L., Jin, Y., Kim, I.-D., & Lee, J.-K. (2013). Glycyrrhizin Attenuates Kainic Acid-Induced Neuronal 259
Cell Death in the Mouse Hippocampus. Exp Neurobiol, 22(2), 107–115. JOUR. http://synapse.koreamed.org/DOIx.php?id=10.5607%2Fen.2013.22.2.107
Song, C., Xu, W., Zhang, X. et al. (2016). CXCR4 Antagonist AMD3100 Suppresses the Long-Term 260
Abnormal Structural Changes of Newborn Neurons in the Intraventricular Kainic Acid Model of Epilepsy. Mol Neurobiol, 53: 1518. https://doi.org/10.1007/s12035-015-9102-9
Miyamoto, M., Ishida, M., & Shinozaki, H. (1997). Anticonvulsive and neuroprotective actions of a 261
potent agonist (DCG-IV) for Group II metabotropic glutamate receptors against intraventricular kainate in the rat. Neuroscience, 77(1), 131–140. JOUR. https://doi.org/10.1016/S0306-4522(96)00442-3
Jing, M., Shingo, T., Yasuhara, T., Kondo, A., Morimoto, T., Wang, F., … Date, I. (2009). The combined 262
therapy of intrahippocampal transplantation of adult neural stem cells and intraventricular erythropoietin-infusion ameliorates spontaneous recurrent seizures by suppression of abnormal mossy fiber sprouting. Brain Research, 1295, 203–217. JOUR. https://doi.org/10.1016/j.brainres.2009.07.079
Gao, X., Guo, M., Meng, D., Sun, F., Guan, L., Cui, Y., … Qi, S. (2019). Silencing MicroRNA-134 263
Alleviates Hippocampal Damage and Occurrence of Spontaneous Seizures After Intraventricular Kainic Acid-Induced Status Epilepticus in Rats. Frontiers in Cellular Neuroscience, 13, 145. JOUR. http://doi.org/10.3389/fncel.2019.00145
Fonnum, F., & Walaas, I. (1978). THE EFFECT OF INTRAHIPPOCAMPAL KAINIC ACID INJECTIONS 264
AND SURGICAL LESIONS ON NEUROTRANSMITTERS IN HIPPOCAMPUS AND SEPTUM. Journal of Neurochemistry, 31(5), 1173–1181. JOUR. http://doi.org/10.1111/j.1471-4159.1978.tb06241.x

Bielefeld, P., Sierra, A., Encinas, J. M., Maletic-Savatic, M., Anderson, A., & Fitzsimons, C. P. (2017). 265
A Standardized Protocol for Stereotaxic Intrahippocampal Administration of Kainic Acid Combined with Electroencephalographic Seizure Monitoring in Mice. Frontiers in Neuroscience, 11, 160. JOUR. http://doi.org/10.3389/fnins.2017.00160
Bragin, A., Engel Jr., J., Wilson, C. L., Vizentin, E., & Mathern, G. W. (1999). Electrophysiologic 266
Analysis of a Chronic Seizure Model After Unilateral Hippocampal KA Injection. Epilepsia, 40(9), 1210–1221. JOUR. http://doi.org/10.1111/j.1528-1157.1999.tb00849.x
Lee, D. J., Hsu, M. S., Seldin, M. M., Arellano, J. L., & Binder, D. K. (2012). Decreased expression of 267
the glial water channel aquaporin-4 in the intrahippocampal kainic acid model of epileptogenesis. Experimental Neurology, 235(1), 246–255. JOUR. https://doi.org/10.1016/j.expneurol.2012.02.002
Rattka, M., Brandt, C., & Löscher, W. (2013). The intrahippocampal kainate model of temporal lobe 268
epilepsy revisited: Epileptogenesis, behavioral and cognitive alterations, pharmacological response, and hippoccampal damage in epileptic rats. Epilepsy Research, 103(2), 135–152. JOUR. https://doi.org/10.1016/j.eplepsyres.2012.09.015
Gröticke, I., Hoffmann, K., & Löscher, W. (2008). Behavioral alterations in a mouse model of temporal 269
lobe epilepsy induced by intrahippocampal injection of kainate. Experimental Neurology, 213(1), 71–83. JOUR. https://doi.org/10.1016/j.expneurol.2008.04.036
Tanaka, T., Tanaka, S., Fujita, T., Takano, K., Fukuda, H., Sako, K., & Yonemasu, Y. (1992). 270
Experimental complex partial seizures induced by a microinjection of kainic acid into limbic structures. Progress in Neurobiology, 38(3), 317–334. JOUR. https://doi.org/10.1016/0301-0082(92)90023-8
Shinoda, S., Araki, T., Lan, J.-Q., Schindler, C. K., Simon, R. P., Taki, W., & Henshall, D. C. (2004). 271
Development of a model of seizure-induced hippocampal injury with features of programmed cell death in the BALB/c mouse. Journal of Neuroscience Research, 76(1), 121–128. JOUR. http://doi.org/10.1002/jnr.20064
Bouilleret, V., Ridoux, V., Depaulis, A., Marescaux, C., Nehlig, A., & Le Gal La Salle, G. (1999). 272
Recurrent seizures and hippocampal sclerosis following intrahippocampal kainate injection in adult mice: electroencephalography, histopathology and synaptic reorganization similar to mesial temporal lobe epilepsy. Neuroscience, 89(3), 717–729. JOUR. https://doi.org/10.1016/S0306-4522(98)00401-1
Cavalheiro, E. A., Riche, D. A., & Le Gal La Salle, G. (1982). Long-term effects of intrahippocampal 273
kainic acid injection in rats: A method for inducing spontaneous recurrent seizures. Electroencephalography and Clinical Neurophysiology, 53(6), 581–589. JOUR. https://doi.org/10.1016/0013-4694(82)90134-1
Riban, V., Bouilleret, V., Pham-Lê, B. T., Fritschy, J.-M., Marescaux, C., & Depaulis, A. (2002). 274
Evolution of hippocampal epileptic activity during the development of hippocampal sclerosis in a mouse model of temporal lobe epilepsy. Neuroscience, 112(1), 101–111. JOUR. https://doi.org/10.1016/S0306-4522(02)00064-7
Gouder, N., Fritschy, J.-M., & Boison, D. (2003). Seizure Suppression by Adenosine A1 Receptor 275
Activation in a Mouse Model of Pharmacoresistant Epilepsy. Epilepsia, 44(7), 877–885. JOUR. http://doi.org/10.1046/j.1528-1157.2003.03603.x
Klee, R., Brandt, C., Töllner, K., & Löscher, W. (2017). Various modifications of the intrahippocampal 276
kainate model of mesial temporal lobe epilepsy in rats fail to resolve the marked rat-to-mouse differences in type and frequency of spontaneous seizures in this model. Epilepsy & Behavior, 68, 129–140. JOUR. https://doi.org/10.1016/j.yebeh.2016.11.035
Tanaka, K., Jimenez-Mateos, E. M., Matsushima, S., Taki, W., & Henshall, D. C. (2010). Hippocampal 277
damage after intra-amygdala kainic acid-induced status epilepticus and seizure preconditioning-mediated neuroprotection in SJL mice. Epilepsy Research, 88(2), 151–161. JOUR. https://doi.org/10.1016/j.eplepsyres.2009.10.012
Li, T., Lytle, N., Lan, J.-Q., Sandau, U. S., & Boison, D. (2012). Local disruption of glial adenosine 278
homeostasis in mice associates with focal electrographic seizures: a first step in epileptogenesis? Glia, 60(1), 83–95. JOUR. http://doi.org/10.1002/glia.21250

Liu, G., Gu, B., He, X.-P., Joshi, R. B., Wackerle, H. D., Rodriguiz, R. M., … McNamara, J. O. (2013). 279
Transient Inhibition of TrkB Kinase after Status Epilepticus Prevents Development of Temporal Lobe Epilepsy. Neuron, 79(1), 31–38. JOUR. https://doi.org/10.1016/j.neuron.2013.04.027
French, E. D., Aldinio, C., & Schwarcz, R. (1982). Intrahippocampal kainic acid, seizures and local 280
neuronal degeneration: Relationships assessed in unanesthetized rats. Neuroscience, 7(10), 2525–2536. JOUR. https://doi.org/10.1016/0306-4522(82)90212-3
Twele, F., Bankstahl, M., Klein, S., Römermann, K., & Löscher, W. (2015). The AMPA receptor 281
antagonist NBQX exerts anti-seizure but not antiepileptogenic effects in the intrahippocampal kainate mouse model of mesial temporal lobe epilepsy. Neuropharmacology, 95, 234–242. JOUR. https://doi.org/10.1016/j.neuropharm.2015.03.014
Araki, T., Simon, R. P., Taki, W., Lan, J.-Q., & Henshall, D. C. (2002). Characterization of neuronal 282
death induced by focally evoked limbic seizures in the C57BL/6 mouse. Journal of Neuroscience Research, 69(5), 614–621. JOUR. http://doi.org/10.1002/jnr.10356
Ben-Ari, Y., Tremblay, E., Ottersen, O. P., & Meldrum, B. S. (1980). The role of epileptic activity in 283
hippocampal and “remote” cerebral lesions induced by kainic acid. Brain Research, 191(1), 79–97. JOUR. https://doi.org/10.1016/0006-8993(80)90316-9
Zeidler, Z., Brandt-Fontaine, M., Leintz, C., Krook-Magnuson, C., Netoff, T., & Krook-Magnuson, E. 284
(2018). Targeting the Mouse Ventral Hippocampus in the Intrahippocampal Kainic Acid Model of Temporal Lobe Epilepsy. eNeuro, 5(4), ENEURO.0158-18.2018. JOUR. http://doi.org/10.1523/ENEURO.0158-18.2018
Schauwecker, P. E., & Steward, O. (1997). Genetic determinants of susceptibility to excitotoxic cell 285
death: implications for gene targeting approaches. Proceedings of the National Academy of Sciences of the United States of America, 94(8), 4103–4108. JOUR. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/9108112
Bedner, P., Dupper, A., Hüttmann, K., Müller, J., Herde, M. K., Dublin, P., … Steinhäuser, C. (2015). 286
Astrocyte uncoupling as a cause of human temporal lobe epilepsy. Brain, 138(5), 1208–1222. JOUR. http://doi.org/10.1093/brain/awv067
Jefferys, J., Steinhäuser, C., & Bedner, P. (2016). Chemically-induced TLE models: Topical 287
application. Journal of Neuroscience Methods, 260, 53–61. JOUR. https://doi.org/10.1016/j.jneumeth.2015.04.011
Pitsch, J., Kuehn, J. C., Gnatkovsky, V., Müller, J. A., van Loo, K. M. J., de Curtis, M., … Becker, A. J. 288
(2019). Anti-epileptogenic and Anti-convulsive Effects of Fingolimod in Experimental Temporal Lobe Epilepsy. Molecular Neurobiology, 56(3), 1825–1840. JOUR. http://doi.org/10.1007/s12035-018-1181-y
Chen, Z., Ljunggren, H.-G., Bogdanovic, N., Nennesmo, I., Winblad, B., & Zhu, J. (2002). Excitotoxic 289
neurodegeneration induced by intranasal administration of kainic acid in C57BL/6 mice. Brain Research, 931(2), 135–145.JOUR.https://doi.org/10.1016/S0006-8993(02)02268-0
Sabilallah, M., Fontanaud, P., Linck, N., Boussadia, B., Peyroutou, R., Lasgouzes, T., … Hirbec, H. E. 290
(2016). Evidence for Status Epilepticus and Pro-Inflammatory Changes after Intranasal Kainic Acid Administration in Mice. PLOS ONE, 11(3), e0150793. JOUR. https://doi.org/10.1371/journal.pone.0150793
Frey, W. H., Liu, J., Chen, X., Thorne, R. G., Fawcett, J. R., Ala, T. A., & Rahman, Y.-E. (1997). Delivery 291
of 125I-NGF to the Brain via the Olfactory Route. Drug Delivery, 4(2), 87–92. JOUR. http://doi.org/10.3109/10717549709051878
Duan, R.-S., Chen, Z., Dou, Y.-C., Concha Quezada, H., Nennesmo, I., Adem, A., … Zhu, J. (2006). 292
Apolipoprotein E deficiency increased microglial activation/CCR3 expression and hippocampal damage in kainic acid exposed mice. Experimental Neurology, 202(2), 373–380. JOUR. https://doi.org/10.1016/j.expneurol.2006.06.013
Zhang, X.-M., Zhu, S.-W., Duan, R.-S., Mohammed, A. H., Winblad, B., & Zhu, J. (2008). Gender 293
differences in susceptibility to kainic acid-induced neurodegeneration in aged C57BL/6 mice. NeuroToxicology, 29(3), 406–412. JOUR. https://doi.org/10.1016/j.neuro.2008.01.006

Lu, M.-O., Zhang, X.-M., Mix, E., Quezada, H. C., Jin, T., Zhu, J., & Adem, A. (2008). TNF-α receptor 294
1 deficiency enhances kainic acid–induced hippocampal injury in mice. Journal of Neuroscience Research, 86(7), 1608–1614. JOUR. http://doi.org/10.1002/jnr.21600
Jagirdar, R., Drexel, M., Kirchmair, E., Tasan, R. O., & Sperk, G. (2015). Rapid changes in expression 295
of class I and IV histone deacetylases during epileptogenesis in mouse models of temporal lobe epilepsy. Experimental Neurology, 273, 92–104. JOUR. https://doi.org/10.1016/j.expneurol.2015.07.026
Fritsch, B., Stott, J. J., Joelle Donofrio, J., & Rogawski, M. A. (2010). Treatment of early and late 296
kainic acid-induced status epilepticus with the noncompetitive AMPA receptor antagonist GYKI 52466. Epilepsia, 51(1), 108–117. JOUR. http://doi.org/10.1111/j.1528-1167.2009.02205.x
Pallud, J., Häussler, U., Langlois, M., Hamelin, S., Devaux, B., Deransart, C., & Depaulis, A. (2011). 297
Dentate gyrus and hilus transection blocks seizure propagation and granule cell dispersion in a mouse model for mesial temporal lobe epilepsy. Hippocampus, 21(3), 334–343. JOUR. http://doi.org/10.1002/hipo.20795
Bragin, A., Azizyan, A., Almajano, J., Wilson, C. L., & Engel Jr., J. (2005). Analysis of Chronic Seizure 298
Onsets after Intrahippocampal Kainic Acid Injection in Freely Moving Rats. Epilepsia, 46(10), 1592–1598. JOUR. http://doi.org/10.1111/j.1528-1167.2005.00268.x
Lévesque, M., Salami, P., Gotman, J., & Avoli, M. (2012). Two seizure-onset types reveal specific 299
patterns of high-frequency oscillations in a model of temporal lobe epilepsy. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 32(38), 13264–13272. JOUR. http://doi.org/10.1523/JNEUROSCI.5086-11.2012
Velasco, A. L., Wilson, C. L., Babb, T. L., & Engel Jr, J. (2000). Functional and anatomic correlates of 300
two frequently observed temporal lobe seizure-onset patterns. Neural Plasticity, 7(1–2), 49–63. JOUR. http://doi.org/10.1155/NP.2000.49
Ogren, J. A., Wilson, C. L., Bragin, A., Lin, J. J., Salamon, N., Dutton, R. A., … Staba, R. J. (2009). 301
Three-dimensional surface maps link local atrophy and fast ripples in human epileptic hippocampus. Annals of Neurology, 66(6), 783–791. JOUR. http://doi.org/10.1002/ana.21703
Jirsa, V. K., Stacey, W. C., Quilichini, P. P., Ivanov, A. I., & Bernard, C. (2014). On the nature of seizure 302
dynamics. Brain : a journal of neurology, 137(Pt 8), 2210–2230. https://doi.org/10.1093/brain/awu133
Saggio, M. L., Crisp, D., Scott, J. M., Karoly, P., Kuhlmann, L., Nakatani, M., … Stacey, W. C. (2020). 303
A taxonomy of seizure dynamotypes. eLife, 9, e55632. JOUR. http://doi.org/10.7554/eLife.55632
Crisp, D. N., Cheung, W., Gliske, S. V, Lai, A., Freestone, D. R., Grayden, D. B., … Stacey, W. C. 304
(2020). Quantifying epileptogenesis in rats with spontaneous and responsive brain state dynamics. Brain Communications, 2(1). JOUR. http://doi.org/10.1093/braincomms/fcaa048
Gorter, J. A., Van Vliet, E. A., Aronica, E., & Da Silva, F. H. L. (2001). Progression of spontaneous 305
seizures after status epilepticus is associated with mossy fibre sprouting and extensive bilateral loss of hilar parvalbumin and somatostatin-immunoreactive neurons. European Journal of Neuroscience, 13(4), 657–669. JOUR. http://doi.org/10.1046/j.1460-9568.2001.01428.x
Van Nieuwenhuyse, B., Raedt, R., Sprengers, M., Dauwe, I., Gadeyne, S., Carrette, E., … Vonck, K. 306
(2015). The systemic kainic acid rat model of temporal lobe epilepsy: Long-term EEG monitoring. Brain Research, 1627, 1–11. JOUR. https://doi.org/10.1016/j.brainres.2015.08.016
Quigg, M. (2000). Circadian rhythms: interactions with seizures and epilepsy. Epilepsy Research, 307
42(1), 43–55. JOUR. https://doi.org/10.1016/S0920-1211(00)00157-1
Manfredini, R., Vergine, G., Boari, B., Faggioli, R., & Borgna-Pignatti, C. (2004). Circadian and 308
seasonal variation of first febrile seizures. The Journal of Pediatrics, 145(6), 838–839. JOUR. https://doi.org/10.1016/j.jpeds.2004.06.079
Spencer, D. C., Sun, F. T., Brown, S. N., Jobst, B. C., Fountain, N. B., Wong, V. S. S., … Quigg, M. 309
(2016). Circadian and ultradian patterns of epileptiform discharges differ by seizure-onset location during long-term ambulatory intracranial monitoring. Epilepsia, 57(9), 1495–1502. JOUR. http://doi.org/10.1111/epi.13455

Karoly, P. J., Goldenholz, D. M., Freestone, D. R., Moss, R. E., Grayden, D. B., Theodore, W. H., & 310
Cook, M. J. (2018). Circadian and circaseptan rhythms in human epilepsy: a retrospective cohort study. The Lancet Neurology, 17(11), 977–985. JOUR. https://doi.org/10.1016/S1474-4422(18)30274-6
Baud, M. O., Kleen, J. K., Mirro, E. A., Andrechak, J. C., King-Stephens, D., Chang, E. F., & Rao, V. R. 311
(2018). Multi-day rhythms modulate seizure risk in epilepsy. Nature Communications, 9(1), 88. JOUR. http://doi.org/10.1038/s41467-017-02577-y
Baud, M. O., Ghestem, A., Benoliel, J.-J., Becker, C., & Bernard, C. (2019). Endogenous multidien 312
rhythm of epilepsy in rats. Experimental Neurology, 315, 82–87. JOUR. https://doi.org/10.1016/j.expneurol.2019.02.006
McLin, J. P., & Steward, O. (2006). Comparison of seizure phenotype and neurodegeneration induced 313
by systemic kainic acid in inbred, outbred, and hybrid mouse strains. European Journal of Neuroscience, 24(8), 2191–2202. JOUR. http://doi.org/10.1111/j.1460-9568.2006.05111.x
Ferraro, T. N., Golden, G. T., Smith, G. G., & Berrettini, W. H. (1995). Differential Susceptibility to 314
Seizures Induced by Systemic Kainic Acid Treatment in Mature DBA/2J and C57BLl6J Mice. Epilepsia, 36(3), 301–307. JOUR. http://doi.org/10.1111/j.1528-1157.1995.tb00999.x
Royle, S. J., Collins, F. C., Rupniak, H. T., Barnes, J. C., & Anderson, R. (1999). Behavioural analysis 315
and susceptibility to CNS injury of four inbred strains of mice. Brain Research, 816(2), 337–349. JOUR. https://doi.org/10.1016/S0006-8993(98)01122-6
Holmes, G. L., & Thompson, J. L. (1988). Effects of kainic acid on seizure susceptibility in the 316
developing brain. Developmental Brain Research, 39(1), 51–59. JOUR. https://doi.org/10.1016/0165-3806(88)90066-1
Mikati, M. A., Werner, S., Shehab, L., Shamseddine, A., Simaan, E., Liu, Z., … Holmes, G. (2003). 317
Stages of status epilepticus in the developing brain. Epilepsy Research, 55(1), 9–19. JOUR. https://doi.org/10.1016/S0920-1211(03)00089-5
Wozniak, D. F., Stewart, G. R., Philip Miller, J., & Olney, J. W. (1991). Age-related sensitivity to kainate 318
neurotoxicity. Experimental Neurology, 114(2), 250–253. JOUR. https://doi.org/10.1016/0014-4886(91)90042-B
McCord, M. C., Lorenzana, A., Bloom, C. S., Chancer, Z. O., & Schauwecker, P. E. (2008). Effect of 319
age on kainate-induced seizure severity and cell death. Neuroscience, 154(3), 1143–1153. JOUR. http://doi.org/10.1016/j.neuroscience.2008.03.082
Darbin, O., Naritoku, D., & Patrylo, P. R. (2004). Aging Alters Electroencephalographic and Clinical 320
Manifestations of Kainate-induced Status Epilepticus. Epilepsia, 45(10), 1219–1227. JOUR. http://doi.org/10.1111/j.0013-9580.2004.66103.x
Rapp, P. R., Stack, E. C., & Gallagher, M. (1999). Morphometric studies of the aged hippocampus: I. 321
Volumetric analysis in behaviorally characterized rats. Journal of Comparative Neurology, 403(4), 459–470. JOUR. http://doi.org/10.1002/(SICI)1096-9861(19990125)403:4<459::AID-CNE3>3.0.CO;2-9
Barnes, C. A., Rao, G., & McNaughton, B. L. (1987). Increased electrotonic coupling in aged rat 322
hippocampus: A possible mechanism for cellular excitability changes. Journal of Comparative Neurology, 259(4), 549–558. JOUR. http://doi.org/10.1002/cne.902590405
Stanley, D. P., & Shetty, A. K. (2004). Aging in the rat hippocampus is associated with widespread 323
reductions in the number of glutamate decarboxylase-67 positive interneurons but not interneuron degeneration. Journal of Neurochemistry, 89(1), 204–216. JOUR. http://doi.org/10.1111/j.1471-4159.2004.02318.x
Wenk, G. L., & Barnes, C. A. (2000). Regional changes in the hippocampal density of AMPA and 324
NMDA receptors across the lifespan of the rat. Brain Research, 885(1), 1–5. JOUR. https://doi.org/10.1016/S0006-8993(00)02792-X
Lerma, J., Paternain, A. V, Rodríguez-Moreno, A., & López-García, J. C. (2001). Molecular Physiology 325
of Kainate Receptors. Physiological Reviews, 81(3), 971–998. JOUR. http://doi.org/10.1152/physrev.2001.81.3.971

Kotsopoulos, I. A. W., Van Merode, T., Kessels, F. G. H., De Krom, M. C. T. F. M., & Knottnerus, J. A. 326
(2002). Systematic Review and Meta-analysis of Incidence Studies of Epilepsy and Unprovoked Seizures. Epilepsia, 43(11), 1402–1409. JOUR. http://doi.org/10.1046/j.1528-1157.2002.t01-1-26901.x
Tabori, N. E., Stewart, L. S., Znamensky, V., Romeo, R. D., Alves, S. E., McEwen, B. S., & Milner, T. A. 327
(2005). Ultrastructural evidence that androgen receptors are located at extranuclear sites in the rat hippocampal formation. Neuroscience, 130(1), 151–163. JOUR. https://doi.org/10.1016/j.neuroscience.2004.08.048
Torres-Reveron, A., Khalid, S., Williams, T. J., Waters, E. M., Jacome, L., Luine, V. N., … Milner, T. A. 328
(2009). Hippocampal dynorphin immunoreactivity increases in response to gonadal steroids and is positioned for direct modulation by ovarian steroid receptors. Neuroscience, 159(1), 204–216. JOUR. http://doi.org/10.1016/j.neuroscience.2008.12.023
Skucas, V. A., Duffy, A. M., Harte-Hargrove, L. C., Magagna-Poveda, A., Radman, T., Chakraborty, G., 329
… Scharfman, H. E. (2013). Testosterone depletion in adult male rats increases mossy fiber transmission, LTP, and sprouting in area CA3 of hippocampus. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 33(6), 2338–2355. JOUR. http://doi.org/10.1523/JNEUROSCI.3857-12.2013
Scharfman, H. E., Mercurio, T. C., Goodman, J. H., Wilson, M. A., & MacLusky, N. J. (2003). 330
Hippocampal excitability increases during the estrous cycle in the rat: a potential role for brain-derived neurotrophic factor. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 23(37), 11641–11652.
Scharfman, H. E., Hintz, T. M., Gomez, J., Stormes, K. A., Barouk, S., Malthankar-Phatak, G. H., … 331
Maclusky, N. J. (2007). Changes in hippocampal function of ovariectomized rats after sequential low doses of estradiol to simulate the preovulatory estrogen surge. The European Journal of Neuroscience, 26(9), 2595–2612. http://doi.org/10.1111/j.1460-9568.2007.05848.x
Toyoda, I., Fujita, S., Thamattoor, A. K., & Buckmaster, P. S. (2015). Unit Activity of Hippocampal 332
Interneurons before Spontaneous Seizures in an Animal Model of Temporal Lobe Epilepsy. The Journal of Neuroscience, 35(16), 6600 LP-6618. JOUR. http://doi.org/10.1523/JNEUROSCI.4786-14.2015
Related Documents











