Therapeutic Approaches to Neuroprotective Activity by Complementary and Alternative Medicines Guest Editors: Ilkay Erdogan Orhan, Monica Rosa Loizzo, and Mahmud Tareq Hassan Khan Evidence-Based Complementary and Alternative Medicine

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Therapeutic Approaches to Neuroprotective Activity by Complementary and Alternative MedicinesGuest Editors: Ilkay Erdogan Orhan, Monica Rosa Loizzo, and Mahmud Tareq Hassan Khan
Evidence-Based Complementary and Alternative Medicine

Therapeutic Approaches to NeuroprotectiveActivity by Complementary andAlternative Medicines

Evidence-Based Complementary and Alternative Medicine
Therapeutic Approaches to NeuroprotectiveActivity by Complementary andAlternative Medicines
Guest Editors: Ilkay Erdogan Orhan, Monica Rosa Loizzo,and Mahmud Tareq Hassan Khan

Copyright © 2012 Hindawi Publishing Corporation. All rights reserved.
This is a special issue published in “Evidence-Based Complementary and Alternative Medicine.” All articles are open access articlesdistributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in anymedium, provided the original work is properly cited.

Editorial Board
Terje Alraek, NorwayShrikant Anant, USASedigheh Asgary, IranHyunsu Bae, Republic of KoreaLijun Bai, ChinaSarang Bani, IndiaVassya Bankova, BulgariaWinfried Banzer, GermanyVernon A. Barnes, USADebra L. Barton, USAJairo Kenupp Bastos, BrazilDavid Baxter, New ZealandAndre-Michael Beer, GermanyAlvin J. Beitz, USAPaolo Bellavite, ItalyYong C. Boo, Republic of KoreaFrancesca Borrelli, ItalyGloria Brusotti, ItalyArndt Bssing, GermanySubhash C. Mandal, IndiaLeigh F. Callahan, USARaffaele Capasso, ItalyOpher Caspi, IsraelShun-Wan Chan, Hong KongIl-Moo Chang, Republic of KoreaChun-Tao Che, USAYunfei Chen, ChinaTzeng-Ji Chen, TaiwanKevin W. Chen, USAJuei-Tang Cheng, TaiwanEvan Paul Cherniack, USAJen-Hwey Chiu, TaiwanJae Y. Cho, Republic of KoreaWilliam C. S. Cho, Hong KongShuang-En Chuang, TaiwanEdwin L. Cooper, USAVincenzo De Feo, ItalyRocio De la Puerta, SpainAlexandra Deters, GermanyDrissa Diallo, NorwayMohamed Eddouks, MoroccoAmr E. Edris, EgyptNobuaki Egashira, JapanTobias Esch, GermanyYibin Feng, Hong Kong
Josue Fernandez-Carnero, SpainJuliano Ferreira, BrazilPeter Fisher, UKJoel J. Gagnier, CanadaM. Nabeel Ghayur, CanadaAnwarul Hassan Gilani, PakistanMichael Goldstein, USASvein Haavik, NorwayS.-H. Hong, Republic of KoreaMarkus Horneber, GermanyChing Liang Hsieh, TaiwanBenny Tan Kwong Huat, SingaporeRoman Huber, GermanyAlyson Huntley, UKAngelo Antonio Izzo, ItalyKanokwan Jarukamjorn, ThailandStefanie Joos, GermanyZ. Kain, USAOsamu Kanauchi, JapanKenji Kawakita, JapanYoun C. Kim, Republic of KoreaJongYeol Kim, Republic of KoreaCheorl-Ho Kim, Republic of KoreaYoshiyuki Kimura, JapanToshiaki Kogure, JapanChing Lan, TaiwanAlfred Langler, GermanyLixing Lao, USACharlotte Leboeuf-Yde, DenmarkTat leang Lee, SingaporeMyeong Soo Lee, Republic of KoreaJang-Hern Lee, Republic of KoreaChristian Lehmann, CanadaMarco Leonti, ItalyPing-Chung Leung, Hong KongShao Li, ChinaXiu-Min Li, USAChun Guang Li, AustraliaSabina Lim, Republic of KoreaWen Chuan Lin, ChinaChristopher G. Lis, USAGerhard Litscher, AustriaI.-Min Liu, TaiwanKe Liu, ChinaYijun Liu, USA
Gaofeng Liu, ChinaCynthia R. Long, USAIrene Lund, SwedenGail Mahady, USAJeanine L. Marnewick, South AfricaFrancesco Marotta, ItalyVirginia S. Martino, ArgentinaJames H. McAuley, AustraliaAndreas Michalsen, GermanyDavid Mischoulon, USAHyung-In Moon, Republic of KoreaAlbert Moraska, USAMark Moss, UKMark A. Moyad, USAStephen Myers, AustraliaMinKyun Na, Republic of KoreaVitaly Napadow, USAF. R. F. Nascimento, BrazilIsabella Neri, ItalyT. Benoıt Nguelefack, CameroonMartin Offenbacher, GermanyKi-Wan Oh, Republic of KoreaY. Ohta, JapanOlumayokun A. Olajide, UKThomas Ostermann, GermanyStacey A. Page, CanadaTai-Long Pan, TaiwanPatchareewan Pannangpetch, ThailandBhushan Patwardhan, IndiaBerit Smestad Paulsen, NorwayAndrea Pieroni, ItalyRichard Pietras, USAXianqin Qu, AustraliaCassandra L. Quave, USARoja Rahimi, IranKhalid Rahman, UKCheppail Ramachandran, USACesar R. Ramos-Remus, MexicoKe Ren, USAMee-Ra Rhyu, Republic of KoreaJose Luis Rıos, SpainPaolo Roberti di Sarsina, ItalyBashar Saad, Palestinian AuthorityAndreas Sandner-Kiesling, AustriaA. Roberto Soares Santos, Brazil

G. Schmeda-Hirschmann, ChileAndrew Scholey, AustraliaVeronique Seidel, UKDana Seidlova-Wuttke, GermanySenthamil R. Selvan, USATuhinadri Sen, IndiaRonald Sherman, USAKaren J. Sherman, USAKan Shimpo, JapanB.-C. Shin, Republic of KoreaJian-nan Song, ChinaRachid Soulimani, FranceElisabet S.-Victorin, SwedenMohd R. Sulaiman, MalaysiaVenil N. Sumantran, IndiaToku Takahashi, USATakashi Takahashi, JapanRabih Talhouk, LebanonJoanna Thompson-Coon, UK
Mei Tian, ChinaYao Tong, Hong KongK. V. Trinh, CanadaVolkan Tugcu, TurkeyYew-Min Tzeng, TaiwanCatherine Ulbricht, USADawn M. Upchurch, USAAlfredo Vannacci, ItalyMani Vasudevan, MalaysiaJoseph R. Vedasiromoni, IndiaCarlo Ventura, ItalyWagner Vilegas, BrazilPradeep Visen, CanadaAristo Vojdani, USADietlind Wahner-Roedler, USAChong-Zhi Wang, USAShu-Ming Wang, USAChenchen Wang, USAY. Wang, USA
Kenji Watanabe, JapanWolfgang Weidenhammer, GermanyJenny M. Wilkinson, AustraliaV. C. N. Wong, Hong KongCharlie Changli Xue, AustraliaHaruki Yamada, JapanNobuo Yamaguchi, JapanHitoshi Yamashita, JapanYong Qing Yang, ChinaKen Yasukawa, JapanE. Yesilada, TurkeyM. Yoon, Republic of KoreaHong Q. Zhang, Hong KongHong Zhang, ChinaRuixin Zhang, USABoli Zhang, ChinaHaibo Zhu, China

Contents
Therapeutic Approaches to Neuroprotective Activity by Complementary and Alternative Medicines,Ilkay Erdogan Orhan, Monica Rosa Loizzo, and Mahmud Tareq Hassan KhanVolume 2012, Article ID 376068, 2 pages
Chinese Medicine in Diabetic Peripheral Neuropathy: Experimental Research on Nerve Repair andRegeneration, Yuanlin Piao and Xiaochun LiangVolume 2012, Article ID 191632, 13 pages
Mitigation of H2O2-Induced Mitochondrial-Mediated Apoptosis in NG108-15 Cells by NovelMesuagenin C from Mesua kunstleri (King) Kosterm, Gomathi Chan,Muhamad Noor Alfarizal Kamarudin, Daniel Zin Hua Wong, Nor Hadiani Ismail, Faizuri Abdul Latif,Aurengzeb Hasan, Khalijah Awang, and Habsah Abdul KadirVolume 2012, Article ID 156521, 18 pages
Herbal Extracts Combination (WNK) Prevents Decline in Spatial Learning and Memory in APP/PS1Mice through Improvement of Hippocampal Aβ Plaque Formation, Histopathology, and Ultrastructure,Wei-hong Cong, Bin Yang, Li Xu, Xiao-xia Dong, Li-song Sheng, Jin-cai Hou, and Jian-xun LiuVolume 2012, Article ID 478190, 9 pages
A Pilot Study for the Neuroprotective Effect of Gongjin-dan on Transient Middle Cerebral ArteryOcclusion-Induced Ischemic Rat Brain, Yun-Young Sunwoo, Sang In Park, Yong-An Chung, Jisoo Lee,Moon-Seo Park, Kyung-Sool Jang, Lee-So Maeng, Dong-Kyu Jang, Ruth Im, Yu Jin Jung, Soon A. Park,Eun-Sun Kang, Min-Wook Kim, and Young-Min HanVolume 2012, Article ID 682720, 11 pages
Anxiolytic-Like Effects of Compound Zhi Zhu Xiang in Rats, Yan-Li Wang, Jin-Li Shi, Liu Yong, Zhao Ren,Yu-Jing Zhai, and Jian-You GuoVolume 2012, Article ID 701289, 7 pages
Early Blockade of TLRs MyD88-Dependent Pathway May Reduce Secondary Spinal Cord Injury in theRats, An-hui Yao, Li-yun Jia, Yu-kai Zhang, Quan-rui Ma, Peng Cheng, Ling Liu, Gong Ju, and Fang KuangVolume 2012, Article ID 591298, 13 pages
Neuroprotective Activity of Sibjeondaebo-tang on Aβ Peptide-Induced Damages, Hyeon Ju Yim,Jung Hwa Lim, Min Hee Kim, Uk Namgung, Sang Ryong Lee, and In Chul JungVolume 2012, Article ID 459894, 8 pages
Centella asiatica (L.) Urban: From Traditional Medicine to Modern Medicine with NeuroprotectivePotential, Ilkay Erdogan OrhanVolume 2012, Article ID 946259, 8 pages
Rhynchophylline Protects Cultured Rat Neurons against Methamphetamine Cytotoxicity, Dan Dan Xu,Robin Hoeven, Rong Rong, and William Chi-Shing ChoVolume 2012, Article ID 636091, 7 pages
Involvement of Heme Oxygenase-1 Induction in the Cytoprotective and Immunomodulatory Activitiesof Viola patrinii in Murine Hippocampal and Microglia Cells, Bin Li, Dong-Sung Lee, Hyun-Gyu Choi,Kyoung-Su Kim, Gil-Saeng Jeong, Ren Bo An, and Youn-Chul KimVolume 2012, Article ID 128019, 12 pages

Paeonol Protects Memory after Ischemic Stroke via Inhibiting β-Secretase and Apoptosis, han-Yu Su,Chin-Yi Cheng, Tung-Hu Tsai, and Ching-Liang HsiehVolume 2012, Article ID 932823, 11 pages
Neuroprotective Effects of San-Huang-Xie-Xin-Tang in the MPP+/MPTP Models of Parkinson’s DiseaseIn Vitro and In Vivo, Yi-Ching Lo, Yu-Tzu Shih, Yu-Ting Tseng, and Hung-Te HsuVolume 2012, Article ID 501032, 10 pages
Effect of Quercetin in the 1-Methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-Induced Mouse Model ofParkinson’s Disease, Chuanfeng Lv, Tie Hong, Zhen Yang, Yu Zhang, Lu Wang, Man Dong, Jing Zhao,Jiaye Mu, and Yixiao MengVolume 2012, Article ID 928643, 6 pages

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 376068, 2 pagesdoi:10.1155/2012/376068
Editorial
Therapeutic Approaches to Neuroprotective Activity byComplementary and Alternative Medicines
Ilkay Erdogan Orhan,1, 2 Monica Rosa Loizzo,3 and Mahmud Tareq Hassan Khan4
1 Department of Pharmacognosy, Faculty of Pharmacy, Gazi University, Ankara, Turkey2 Department of Pharmacognosy and Pharmaceutical Botany, Faculty of Pharmacy, Eastern Mediterranean University,Gazimagusa, Famagusta, The Northern Cyprus, Turkey
3 Department of Pharmaceutical Sciences, Faculty of Pharmacy, University of Calabria, Rende, Italy4 Center for Pharmaceutical Biotechnology, College of Pharmacy, University of Illinois at Chicago, Chicago, IL 60607-7173, USA
Correspondence should be addressed to Ilkay Erdogan Orhan, [email protected]
Received 20 September 2012; Accepted 20 September 2012
Copyright © 2012 Ilkay Erdogan Orhan et al. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
Neurodegeneration is a large-capacity term which can bedescribed as progressive damage on neurons in brief. Manydiseases are known to be associated with central nervoussystem such as Alzheimer’s disease (AD), Parkinson’s disease(PD), Huntington’s disease (HD), multiple sclerosis (MS),and amyotrophic lateral sclerosis (ALS/Lou Gehrig’s Dis-ease) and occur as a result of neurodegenerative processes.Actually, the neurodegenerative diseases have a quite higherprevalence among the elder population living especially inthe well-developed countries because of the higher life stan-dards. For instance, AD is estimated to affect approximately5 million people only in the USA.
Although pathogeneses of most of the neurodegenerativediseases are still continuous topics of a common researchall over the world, the principal mechanisms linked to thesediseases have been frequently suggested as genetic mutationsand intracellular mechanisms in specific brain regions suchas protein degradation pathways, misfolding or occurrence ofabnormal protein structure. Consequently, neuroprotectionis an important treatment option for such neurodegenerativedisorders. The extensive research on discovery novel drugcandidates has shown that natural products such as plantextracts and plant-originated compounds have enormouspotential to become drug leads with neuroprotective activity.One of the most impressing examples of those drugs isgalanthamine, an alkaloid isolated from the bulbs of thesnowdrop plant (Galanthus sp.) that has become a licensedand approved medicine as cholinesterase inhibitor for thetreatment of AD.
Considering the widespread research on the role ofherbal medicines in neuroprotection, 13 research or reviewarticles that contained interesting results relevant to thesubject have been included in the current special issue. Aparticular emphasis has been given to the articles dealingwith neuroprotective effect of eastern herbal medicines usedin China, Korea, Taiwan, Malaysia, and India. Among thepapers, Y. Piao and X. Liang contributed with an excellentreview about neurological activity of traditional Chinesemedicine against diabetic peripheral neuropathy (DPN)associated with chronic diabetes mellitus that causes nervedegeneration while in another review article I. E. Orhansummarized the literature relevant to neurobiological effectsof Centella asiatica “gotu kola,” a reputed plant used as braintonic in Ayurvedic medicine.
G. Chan et al. investigated neuroprotective effect of thephenylcoumarin-type of compounds isolated from the Mala-ysian plant Mesua kunstleri using neuronal cell culturetechniques, which led to identification of mesuagenin C asthe promising neuroprotective agent.
H.-C. Chang et al. reported in vivo neuroprotective effectof a Chinese formulation called Wei Na Kang (WNK),mainly consisting of Panax ginseng, Ginkgo biloba, and Crocussativus using APP/PS1 transgenic mice and concluded thatWNK has been found to reduce the decline in spatialcognition, which might be due to its effects on reducingAβ plaque formation and ameliorating histopathology andultrastructure in hippocampus of APP/PS1 mouse brain.Another study by Y.-L. Wang et al. brightened strong in vivo

2 Evidence-Based Complementary and Alternative Medicine
anxiolytic effect of the compound zhi zhu xiang (CZZX)originated from a clinical experiment that was reported tobe an effective and well-tolerated antianxiety prescriptionin China. The findings by these authors pointed out tothe statement that the anxiolytic efficacy of CZZX maybe mediated by benzodiazepine binding site modulation atγ-aminobutyric acid-A receptors. A.-H. Yao et al. alsocontributed with an article that aimed to examine therole of toll-like receptors (TLRs) myeloid differentiationfactor 88- (MyD88-) dependent pathway in the spinal cordinjury (SCI) in adult male rats. They showed that MyD88inhibitory peptide (MIP) by intramedullary applicationcaused a significant improvement in recovery of locomotorfunction. On the other hand, rhynchophylline (Rhy), a tetra-cyclic oxindole alkaloid and the active component isolatedfrom Uncaria species used for neurological conditions intraditional Chinese medicine, was investigated by D. D. Xuet al. against methamphetamine (MA) neurotoxicity usingneuronal cell culture system and demonstrated to exertinhibitory effect against MA impairment under in vitro con-ditions. S.-Y. Su et al. revealed protective role on memoryafter ischemic stroke of paeonol, the simple phenolic isolatedfrom the Chinese herb Paeonia suffruticosa via reducingamyloid precursor protein (APP), beta-site APP-cleavingenzyme (BACE), and apoptosis. Y.-C. Lo et al. investigatedneuroprotective effect of San-Huang-Xie-Xin-Tang (SHXT),another traditional Chinese medicine containing Coptidisrhizoma, Scutellariae radix, and Rhei rhizoma against PDusing the 1-methyl-4-phenylpyridinium (MPP+)/1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) models. By allparameters used in this study, the authors suggested thatSHXT exerted promising protection against MPTP-inducedneurotoxicity in PD through its antioxidative and antiapop-totic effects. In C. Lv et al.’s study, quercetin, the most studiedbioflavonoid up to date, was explicated against MPTPneurotoxicity related to PD using the similar models to Y.-C.Lo et al.’s work. C. Lv et al. revealed that quercetin treatmentevidently improved the motor balance and coordinationof MPTP-treated mice, which led to the comment thatquercetin has antiparkinsonian properties.
In another paper, Y.-Y. Sunwoo et al. evaluated neuro-protective role of gongjin-dan, a Korean herbal mixture thatcontains Moschus moschiferus, Corni fructus, Angelica gigantisradix, and Cervi parvum cornu against transient middlecerebral artery occlusion (MCAo) connected with ischemicstroke using adult male Sprague-Dawley rats to constitutecerebral ischemic model. According to their results, gongjin-dan gained the advantage over the control groups inbehavioral and immunohistological tests employed. Neu-roprotective action of another Korean herbal formulation“Sibjeondaebo-tang” (SJDBT), which has been used to treatneuropsychiatric disabilities in traditional Korean medicine,has been explicated by H. S. Lim et al. against Aβ peptide-induced damage using in vitro culture and in vivo rat brainsystems and elucidated to possess a protective role fromneuronal damage in the experimental systems used. N. Liet al. reported antioxidant and anti-inflammatory activityof NNMBS275, consisting of the ethanol extract of Violapatrinii from Korea in in murine hippocampal HT22 cells
and BV2 microglia. These authors stated that the neuropro-tective and anti-inflammatory effects of NNMBS275 werelinked to the upregulation of nuclear transcription factor-E2-related factor 2-dependent expression of heme oxygenase-1in HT22 and BV2 cells.
The aforementioned papers published in this specialissue represent quite attractive and exciting results about anumber of plant extracts and herbal compounds in neuro-protection. In most of these papers, the mechanismsunderlying their neuroprotective properties have been alsoexplained using numerous experimental parameters. As theguest editorial team, we would like to express our deepappreciation to the authors of these articles, our reviewers,and the Editor-in-Chief and Editorial Assistants of ECAMwho helped us to make this issue possible to publish.
Ilkay Erdogan OrhanMonica Rosa Loizzo
Mahmud Tareq Hassan Khan

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 191632, 13 pagesdoi:10.1155/2012/191632
Review Article
Chinese Medicine in Diabetic Peripheral Neuropathy:Experimental Research on Nerve Repair and Regeneration
Yuanlin Piao and Xiaochun Liang
Department of Traditional Chinese Medicine, Peking Union Medical College Hospital, Peking Union Medical Collegeand Chinese Academy of Medical Sciences, No. 1 Shuaifuyuan, Dongcheng District, Beijing 100730, China
Correspondence should be addressed to Xiaochun Liang, [email protected]
Received 11 February 2012; Revised 8 April 2012; Accepted 26 April 2012
Academic Editor: Monica Rosa Loizzo
Copyright © 2012 Y. Piao and X. Liang. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
Diabetic peripheral neuropathy (DPN) is one of the most common complications of chronic diabetes mellitus. Pathologicalcharacteristics of DPN include axonal atrophy, nerve demyelination, and delayed regeneration of peripheral sensory nerve fibers.The goal of treatment in DPN is not only to ameliorate neurological symptoms but also to slow or reverse the underlyingneurodegenerative process. Schwann cells and neurotrophic factors play important roles in the repair and regeneration ofperipheral nerves. The present paper reviews current studies and evidence regarding the neurological effects of traditional Chinesemedicine, with an emphasis on recent developments in the area of nerve repair and regeneration in DPN.
1. Introduction
Diabetic peripheral neuropathy (DPN) is a common com-plication of chronic diabetes. Pathological characteristicsof DPN include axonal atrophy, nerve demyelination, anddelayed regeneration of peripheral sensory nerve fibers. Toour knowledge, the pathophysiological mechanism of DPNin dysfunctional peripheral nerve repair and regeneration isnot well understood.
The symptoms associated with DPN have been men-tioned in various traditional Chinese medicine (TCM)references. Pujifang (Prescriptions for Universal Relief), anancient Chinese medicine book written in the Ming dynasty,described the following constellation of symptoms: “Thekidney pattern of diabetes consists of symptoms of thirst,dry eye, impotence, and annoying pain in the hands andfeet.” Moreover, in Wangxugaoyian (Medical Records ofWangxugao) from the Qing dynasty, there was a case of apatient with diabetes noted to have “numbness of handsand feet” and “limbs as cold as ice.” The differentiation ofDPN implicates the domains of “sinew impediment,” “bloodimpediment,” and “leg flaccidity” in Chinese medicine [1].
From the viewpoint of TCM [1], the etiology andpathogenesis of DPN are as follows: (1) with an increased
duration of disease in diabetes, a deficiency of yin burnsbody fluid and blood, resulting in empty heat. This increasesblood viscosity, resulting in blood stasis, as well as blockageof sinews and channels; (2) excessive intake of foods highin fat and sugar content results in the deficiency of spleenand stomach, resulting in the accumulation of dampnessand phlegm, which has a synergistic effect with stasis; (3)sinew and channels demonstrate poor nourishment becauseof the deficiency of liver and kidney; (4) the deficiency ofyin results in a deficiency of yang, which generates an innercold that results in microvascular coagulation. These fouraspects result in a decreased peripheral flow of qi and bloodto muscles, sinew, and channels. With regard to visceralorgan systems, DPN is related to the liver, spleen, and kidney.The nature of DPN is deficiency secondarily complicatedby excess; the deficiency is the root, and the excess is asubsequent manifestation. The root cause is deficiency inqi, yin, and yang; the subsequent complication is bloodstasis and phlegm accumulation. Common patterns andtreatments of DPN are summarized in Table 1.
Recently the effects of Schwann cells and neurotrophicfactors on the repair and regeneration of peripheral nervehave been of research interest. Recent studies have shownthat TCM medications may affect neuronal repair and

2 Evidence-Based Complementary and Alternative Medicine
Ta
ble
1:C
omm
onpa
tter
ns
and
trea
tmen
tsof
DP
Nin
Ch
ines
em
edic
ine.
Patt
ern
Clin
ical
man
ifes
tati
onT
her
apeu
tic
prin
cipl
eTr
eatm
ent
(for
mu
la)
Ori
gin
offo
rmu
laC
omp
onen
ts
Defi
cien
cyof
Yin
and
bloo
dst
asis
Nu
mbn
ess
and
burn
ing
pain
inh
ands
and
feet
grad
ual
lyex
ten
dsto
the
enti
relim
bs,
aggr
avat
edat
nig
ht;
nig
ht
swea
tin
g;sp
onta
neo
us
swea
tin
g;h
eat
sen
sati
onin
the
ches
t,pa
lms
and
sole
s;so
ren
ess
and
wea
knes
sof
the
lum
bar
regi
onan
dkn
ees
join
ts,d
rym
outh
;th
irst
y;dr
yst
ool;
dark
-red
ton
gue
wit
hpe
eled
orlit
tle
coat
ing;
fin
ean
dch
oppy
puls
e.
Nou
rish
Yin
and
rem
ove
bloo
dst
asis
Zh
ibai
Dih
uan
gW
an1
(An
emar
rhen
a-P
hel
lode
ndr
on-R
ehm
ann
iaP
ill)
toge
ther
wit
hTa
ohon
gsiw
uta
ng
2(P
ersi
ca-C
arth
amu
sFo
ur
Subs
tan
ces
Dec
octi
on)
1Ji
ngu
iyao
lue
(Syn
opsi
sof
Pre
scri
ptio
ns
ofth
eG
olde
nC
ham
ber)
;2Y
izon
gjin
jian
(Gol
den
Mir
ror
ofM
edic
ine)
Shud
ihua
ng(R
adix
Reh
man
nia
epr
epar
ata)
,Sh
anzh
uyu
(Fru
ctu
sC
orn
i),S
hany
ao(R
hiz
oma
Dio
scor
eae)
,Zex
ie(R
hiz
oma
Alis
mat
is),
Fulin
g(P
oria
),M
udan
pi(C
orte
xM
outa
n),
Zhi
mu
(Rh
izom
aA
nem
arrh
enae
),H
uang
bai(
Cor
tex
Ph
ello
den
dri)
,Dan
ggui
(Rad
ixA
nge
licae
sin
ensi
s),
Chu
anxi
ong
(Rh
izom
aC
hu
anxi
ong)
,Bai
shao
(Rad
ixPa
eon
iae
alba
),Ta
oren
(Sem
enPe
rsic
ae),
Hon
ghua
(Flo
sC
arth
amit
inct
orii
)
Defi
cien
cyof
Yan
gan
dbl
ood
stas
is
Nu
mbn
ess
pain
,agg
rava
ted
atn
igh
tor
enco
un
ters
cold
nes
s,av
ersi
onto
cold
,col
dlim
bs,s
oren
ess
and
wea
knes
sof
the
lum
bar
regi
onan
dkn
ees
join
ts,t
aste
less
nes
sin
the
mou
thw
ith
out
thir
sty,
impo
ten
ce,
prem
atu
reej
acu
lati
on,p
oor
orlo
ose
stoo
l,pa
leor
dark
ton
gue,
wh
ite
thic
kor
grea
syco
atin
g,de
epfi
ne
orde
epsl
owpu
lse.
War
man
dto
nif
yYa
ng
and
rem
ove
bloo
dst
asis
Jin
guis
hen
qiw
an1
(Gol
den
Ch
est
Kid
ney
-QiP
ill)
toge
ther
wit
hTa
ohon
gsiw
uta
ng
2(P
ersi
ca-C
arth
amu
sFo
ur
Subs
tan
ces
Dec
octi
on)
1Ji
ngu
iyao
lue
(Syn
opsi
sof
Pre
scri
ptio
ns
ofth
eG
olde
nC
ham
ber)
;2Y
izon
gjin
jian
(Gol
den
Mir
ror
ofM
edic
ine)
Fuzi
(Rad
ixA
con
itil
ater
alis
prep
arat
a),G
uizh
i(R
amu
lus
Cin
nam
omi)
,Sha
nzhu
yu(F
ruct
us
Cor
ni)
,Sh
anya
o(R
hiz
oma
Dio
scor
eae)
,Zex
ie(R
hiz
oma
Alis
mat
is),
Mud
anpi
(Cor
tex
Mou
tan
),Fu
ling
(Por
ia),
Shud
ihua
ng(R
adix
Reh
man
nia
epr
epar
ata)
,D
angg
ui(R
adix
An
gelic
aesi
nen
sis)
,Chu
anxi
ong
(Rh
izom
aC
hu
anxi
ong)
,Bai
shao
(Rad
ixPa
eon
iae
Alb
a),T
aore
n(S
emen
Pers
icae
),H
ongh
ua(F
los
Car
tham
itin
ctor
ii)
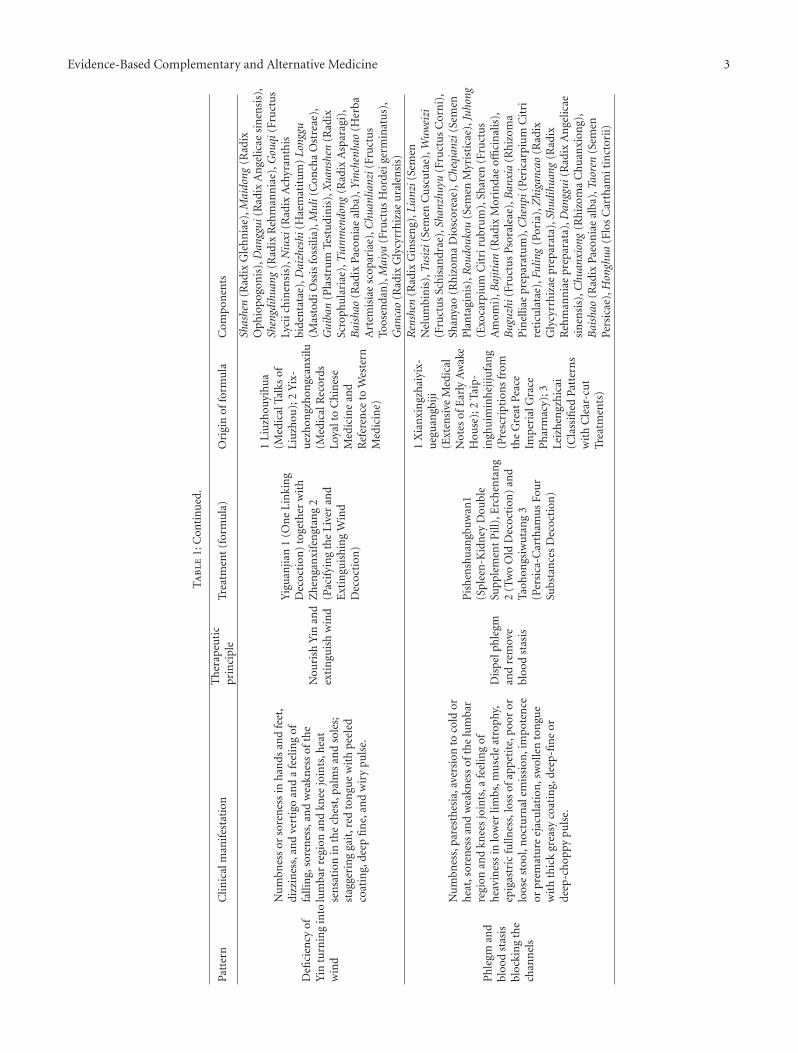
Evidence-Based Complementary and Alternative Medicine 3
Ta
ble
1:C
onti
nu
ed.
Patt
ern
Clin
ical
man
ifes
tati
onT
her
apeu
tic
prin
cipl
eTr
eatm
ent
(for
mu
la)
Ori
gin
offo
rmu
laC
omp
onen
ts
Defi
cien
cyof
Yin
turn
ing
into
win
d
Nu
mbn
ess
orso
ren
ess
inh
ands
and
feet
,di
zzin
ess,
and
vert
igo
and
afe
elin
gof
falli
ng,
sore
nes
s,an
dw
eakn
ess
ofth
elu
mba
rre
gion
and
knee
join
ts,h
eat
sen
sati
onin
the
ches
t,pa
lms
and
sole
s;st
agge
rin
gga
it,r
edto
ngu
ew
ith
peel
edco
atin
g,de
epfi
ne,
and
wir
ypu
lse.
Nou
rish
Yin
and
exti
ngu
ish
win
d
Yig
uan
jian
1(O
ne
Lin
kin
gD
ecoc
tion
)to
geth
erw
ith
Zh
enga
nxi
fen
gtan
g2
(Pac
ifyi
ng
the
Live
ran
dE
xtin
guis
hin
gW
ind
Dec
octi
on)
1Li
uzh
ouyi
hua
(Med
ical
Talk
sof
Liu
zhou
);2
Yix
-u
ezh
ongz
hon
gcan
xilu
(Med
ical
Rec
ords
Loya
lto
Ch
ines
eM
edic
ine
and
Ref
eren
ceto
Wes
tern
Med
icin
e)
Shas
hen
(Rad
ixG
leh
nia
e),M
aido
ng(R
adix
Oph
iopo
gon
is),
Dan
ggui
(Rad
ixA
nge
licae
sin
ensi
s),
Shen
gdih
uang
(Rad
ixR
ehm
ann
iae)
,Gou
qi(F
ruct
us
Lyci
ich
inen
sis)
,Niu
xi(R
adix
Ach
yran
this
bide
nta
tae)
,Dai
zhes
hi(H
aem
atit
um
)Lo
nggu
(Mas
todi
Oss
isfo
ssili
a),M
uli(
Con
cha
Ost
reae
),G
uiba
n(P
last
rum
Test
udi
nis
),X
uans
hen
(Rad
ixSc
roph
ula
riae
),T
ianm
endo
ng(R
adix
Asp
arag
i),
Bai
shao
(Rad
ixPa
eon
iae
alba
),Y
inch
enha
o(H
erba
Art
emis
iae
scop
aria
e),C
huan
lianz
i(Fr
uct
us
Toos
enda
n),
Mai
ya(F
ruct
us
Hor
deig
erm
inat
us)
,G
anca
o(R
adix
Gly
cyrr
hiz
aeu
rale
nsi
s)
Ph
legm
and
bloo
dst
asis
bloc
kin
gth
ech
ann
els
Nu
mbn
ess,
pare
sth
esia
,ave
rsio
nto
cold
orh
eat,
sore
nes
san
dw
eakn
ess
ofth
elu
mba
rre
gion
and
knee
sjo
ints
,afe
elin
gof
hea
vin
ess
inlo
wer
limbs
,mu
scle
atro
phy,
epig
astr
icfu
llnes
s,lo
ssof
appe
tite
,poo
ror
loos
est
ool,
noc
turn
alem
issi
on,i
mpo
ten
ceor
prem
atu
reej
acu
lati
on,s
wol
len
ton
gue
wit
hth
ick
grea
syco
atin
g,de
ep-fi
ne
orde
ep-c
hop
pypu
lse.
Dis
pelp
hle
gman
dre
mov
ebl
ood
stas
is
Pis
hen
shu
angb
uwan
1(S
plee
n-K
idn
eyD
oubl
eSu
pple
men
tP
ill),
Erc
hen
tan
g2
(Tw
oO
ldD
ecoc
tion
)an
dTa
ohon
gsiw
uta
ng
3(P
ersi
ca-C
arth
amu
sFo
ur
Subs
tan
ces
Dec
octi
on)
1X
ian
xin
gzh
aiyi
x-u
egu
angb
iji(E
xten
sive
Med
ical
Not
esof
Ear
lyA
wak
eH
ouse
);2
Taip
-in
ghu
imin
hej
ijufa
ng
(Pre
scri
ptio
ns
from
the
Gre
atPe
ace
Imp
eria
lGra
ceP
har
mac
y);3
Leiz
hen
gzh
icai
(Cla
ssifi
edPa
tter
ns
wit
hC
lear
-cu
tTr
eatm
ents
)
Ren
shen
(Rad
ixG
inse
ng)
,Lia
nzi(
Sem
enN
elu
mbi
nis
),Tu
sizi
(Sem
enC
usc
uta
e),W
uwei
zi(F
ruct
us
Sch
isan
drae
),Sh
anzh
uyu
(Fru
ctu
sC
orn
i),
Shan
yao
(Rh
izom
aD
iosc
orea
e),C
heqi
anzi
(Sem
enP
lan
tagi
nis
),R
oudo
ukou
(Sem
enM
yris
tica
e),J
uhon
g(E
xoca
rpiu
mC
itri
rubr
um
),Sh
aren
(Fru
ctu
sA
mom
i),B
ajit
ian
(Rad
ixM
orin
dae
offici
nal
is),
Bug
uzhi
(Fru
ctu
sPs
oral
eae)
,Ban
xia
(Rh
izom
aP
inel
liae
prep
arat
um
),C
henp
i(Pe
rica
rpiu
mC
itri
reti
cula
tae)
,Ful
ing
(Por
ia),
Zhi
ganc
ao(R
adix
Gly
cyrr
hiz
aepr
epar
ata)
,Shu
dihu
ang
(Rad
ixR
ehm
ann
iae
prep
arat
a),D
angg
ui(R
adix
An
gelic
aesi
nen
sis)
,Chu
anxi
ong
(Rh
izom
aC
hu
anxi
ong)
,B
aish
ao(R
adix
Paeo
nia
eal
ba),
Taor
en(S
emen
Pers
icae
),H
ongh
ua(F
los
Car
tham
itin
ctor
ii)
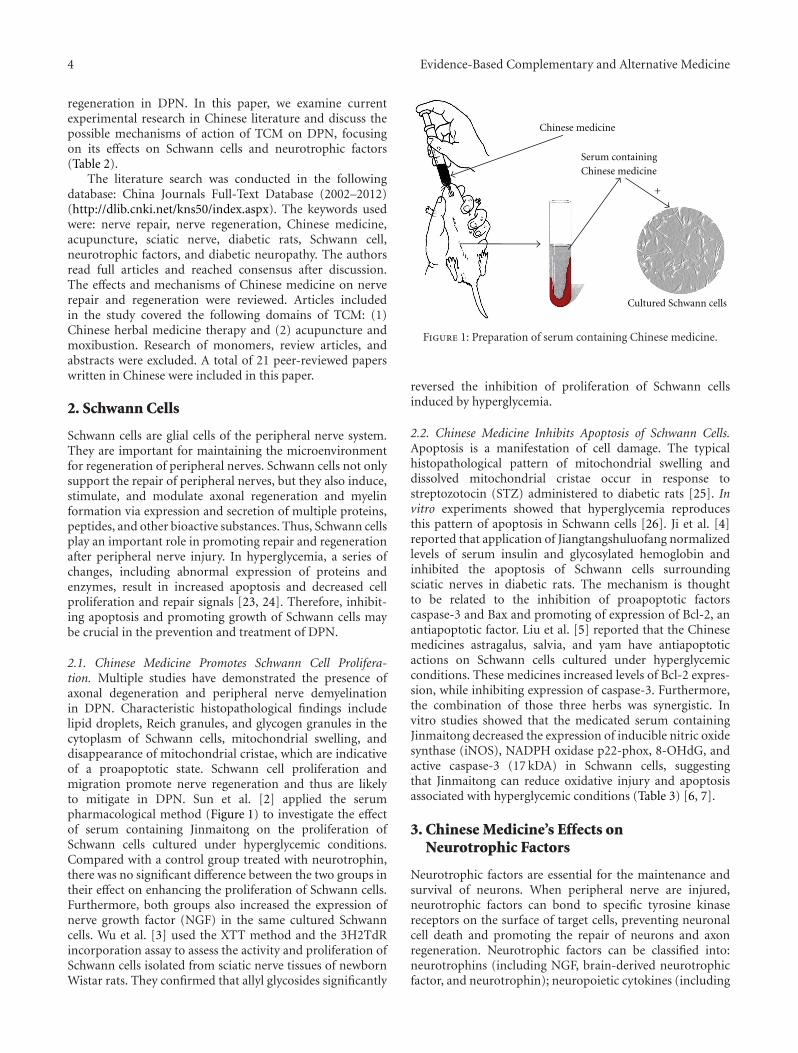
4 Evidence-Based Complementary and Alternative Medicine
regeneration in DPN. In this paper, we examine currentexperimental research in Chinese literature and discuss thepossible mechanisms of action of TCM on DPN, focusingon its effects on Schwann cells and neurotrophic factors(Table 2).
The literature search was conducted in the followingdatabase: China Journals Full-Text Database (2002–2012)(http://dlib.cnki.net/kns50/index.aspx). The keywords usedwere: nerve repair, nerve regeneration, Chinese medicine,acupuncture, sciatic nerve, diabetic rats, Schwann cell,neurotrophic factors, and diabetic neuropathy. The authorsread full articles and reached consensus after discussion.The effects and mechanisms of Chinese medicine on nerverepair and regeneration were reviewed. Articles includedin the study covered the following domains of TCM: (1)Chinese herbal medicine therapy and (2) acupuncture andmoxibustion. Research of monomers, review articles, andabstracts were excluded. A total of 21 peer-reviewed paperswritten in Chinese were included in this paper.
2. Schwann Cells
Schwann cells are glial cells of the peripheral nerve system.They are important for maintaining the microenvironmentfor regeneration of peripheral nerves. Schwann cells not onlysupport the repair of peripheral nerves, but they also induce,stimulate, and modulate axonal regeneration and myelinformation via expression and secretion of multiple proteins,peptides, and other bioactive substances. Thus, Schwann cellsplay an important role in promoting repair and regenerationafter peripheral nerve injury. In hyperglycemia, a series ofchanges, including abnormal expression of proteins andenzymes, result in increased apoptosis and decreased cellproliferation and repair signals [23, 24]. Therefore, inhibit-ing apoptosis and promoting growth of Schwann cells maybe crucial in the prevention and treatment of DPN.
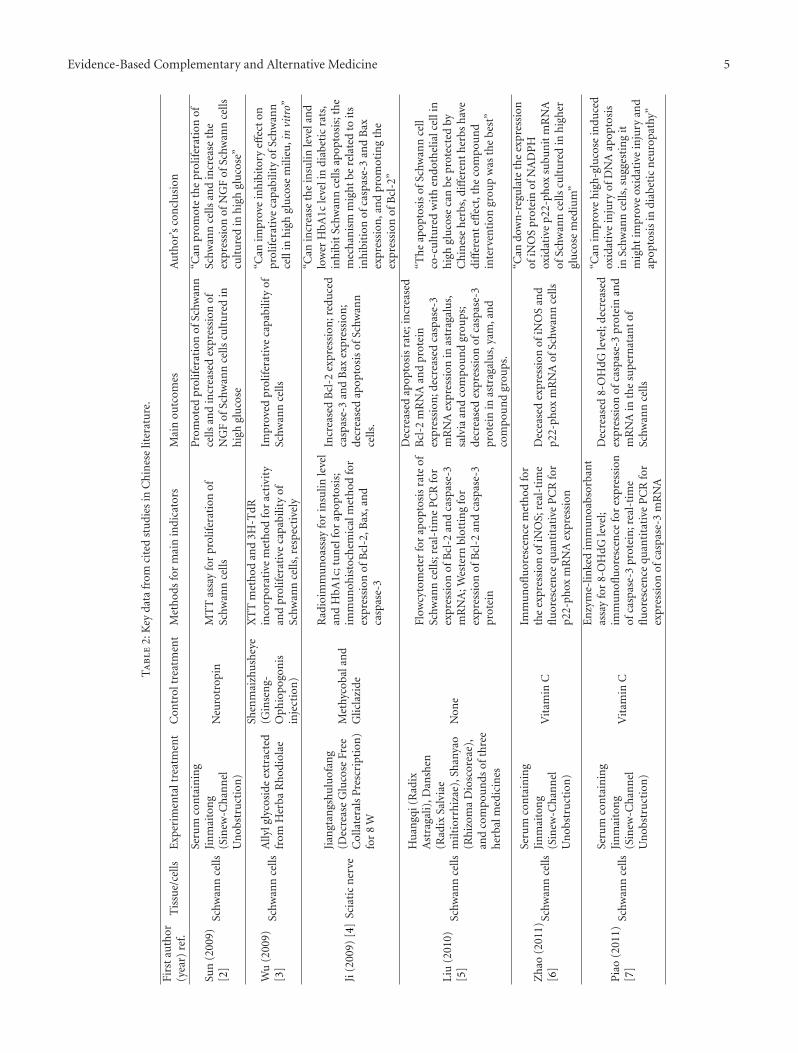
2.1. Chinese Medicine Promotes Schwann Cell Prolifera-tion. Multiple studies have demonstrated the presence ofaxonal degeneration and peripheral nerve demyelinationin DPN. Characteristic histopathological findings includelipid droplets, Reich granules, and glycogen granules in thecytoplasm of Schwann cells, mitochondrial swelling, anddisappearance of mitochondrial cristae, which are indicativeof a proapoptotic state. Schwann cell proliferation andmigration promote nerve regeneration and thus are likelyto mitigate in DPN. Sun et al. [2] applied the serumpharmacological method (Figure 1) to investigate the effectof serum containing Jinmaitong on the proliferation ofSchwann cells cultured under hyperglycemic conditions.Compared with a control group treated with neurotrophin,there was no significant difference between the two groups intheir effect on enhancing the proliferation of Schwann cells.Furthermore, both groups also increased the expression ofnerve growth factor (NGF) in the same cultured Schwanncells. Wu et al. [3] used the XTT method and the 3H2TdRincorporation assay to assess the activity and proliferation ofSchwann cells isolated from sciatic nerve tissues of newbornWistar rats. They confirmed that allyl glycosides significantly
Chinese medicine
Serum containing Chinese medicine
Cultured Schwann cells
+
Figure 1: Preparation of serum containing Chinese medicine.
reversed the inhibition of proliferation of Schwann cellsinduced by hyperglycemia.
2.2. Chinese Medicine Inhibits Apoptosis of Schwann Cells.Apoptosis is a manifestation of cell damage. The typicalhistopathological pattern of mitochondrial swelling anddissolved mitochondrial cristae occur in response tostreptozotocin (STZ) administered to diabetic rats [25]. Invitro experiments showed that hyperglycemia reproducesthis pattern of apoptosis in Schwann cells [26]. Ji et al. [4]reported that application of Jiangtangshuluofang normalizedlevels of serum insulin and glycosylated hemoglobin andinhibited the apoptosis of Schwann cells surroundingsciatic nerves in diabetic rats. The mechanism is thoughtto be related to the inhibition of proapoptotic factorscaspase-3 and Bax and promoting of expression of Bcl-2, anantiapoptotic factor. Liu et al. [5] reported that the Chinesemedicines astragalus, salvia, and yam have antiapoptoticactions on Schwann cells cultured under hyperglycemicconditions. These medicines increased levels of Bcl-2 expres-sion, while inhibiting expression of caspase-3. Furthermore,the combination of those three herbs was synergistic. Invitro studies showed that the medicated serum containingJinmaitong decreased the expression of inducible nitric oxidesynthase (iNOS), NADPH oxidase p22-phox, 8-OHdG, andactive caspase-3 (17 kDA) in Schwann cells, suggestingthat Jinmaitong can reduce oxidative injury and apoptosisassociated with hyperglycemic conditions (Table 3) [6, 7].
3. Chinese Medicine’s Effects onNeurotrophic Factors
Neurotrophic factors are essential for the maintenance andsurvival of neurons. When peripheral nerve are injured,neurotrophic factors can bond to specific tyrosine kinasereceptors on the surface of target cells, preventing neuronalcell death and promoting the repair of neurons and axonregeneration. Neurotrophic factors can be classified into:neurotrophins (including NGF, brain-derived neurotrophicfactor, and neurotrophin); neuropoietic cytokines (including
Evidence-Based Complementary and Alternative Medicine 5
Ta
ble
2:K
eyda
tafr
omci
ted
stu
dies
inC
hin
ese
liter
atu
re.
Firs
tau
thor
(yea
r)re
f.T
issu
e/ce
llsE
xper
imen
talt
reat
men
tC
ontr
oltr
eatm
ent
Met
hod
sfo
rm
ain
indi
cato
rsM
ain
outc
omes
Au
thor
’sco
ncl
usi
on
Sun
(200
9)[2
]Sc
hwan
nce
lls
Seru
mco
nta
inin
gJi
nm
aito
ng
(Sin
ew-C
han
nel
Un
obst
ruct
ion
)
Neu
rotr
opin
MT
Tas
say
for
prol
ifer
atio
nof
Schw
ann
cells
Pro
mot
edpr
olif
erat
ion
ofSc
hwan
nce
llsan
din
crea
sed
expr
essi
onof
NG
Fof
Schw
ann
cells
cult
ure
din
hig
hgl
uco
se
“Can
prom
ote
the
prol
ifer
atio
nof
Schw
ann
cells
and
incr
ease
the
expr
essi
onof
NG
Fof
Schw
ann
cells
cult
ure
din
hig
hgl
uco
se”
Wu
(200
9)[3
]Sc
hwan
nce
llsA
llylg
lyco
side
extr
acte
dfr
omH
erba
Rh
odio
lae
Shen
mai
zhu
shey
e(G
inse
ng-
Oph
iopo
gon
isin
ject
ion
)
XT
Tm
eth
odan
d3H
-TdR
inco
rpor
ativ
em
eth
odfo
rac
tivi
tyan
dpr
olif
erat
ive
capa
bilit
yof
Schw
ann
cells
,res
pec
tive
ly
Impr
oved
prol
ifer
ativ
eca
pabi
lity
ofSc
hwan
nce
lls
“Can
impr
ove
inh
ibit
ory
effec
ton
prol
ifer
ativ
eca
pabi
lity
ofSc
hwan
nce
llin
hig
hgl
uco
sem
ilieu
,in
vitr
o”
Ji(2
009)
[4]
Scia
tic
ner
ve
Jian
gtan
gshu
luof
ang
(Dec
reas
eG
luco
seFr
eeC
olla
tera
lsP
resc
ript
ion
)fo
r8
W
Met
hyco
bala
nd
Glic
lazi
de
Rad
ioim
mu
noa
ssay
for
insu
linle
vel
and
HbA
1c;t
un
elfo
rap
opto
sis;
imm
un
ohis
toch
emic
alm
eth
odfo
rex
pres
sion
ofB
cl-2
,Bax
,an
dca
spas
e-3
Incr
ease
dB
cl-2
expr
essi
on;r
edu
ced
casp
ase-
3an
dB
axex
pres
sion
;de
crea
sed
apop
tosi
sof
Schw
ann
cells
.
“Can
incr
ease
the
insu
linle
vela
nd
low
erH
bA1c
leve
lin
diab
etic
rats
,in
hib
itSc
hwan
nce
llsap
opto
sis;
the
mec
han
ism
mig
ht
bere
late
dto
its
inh
ibit
ion
ofca
spas
e-3
and
Bax
expr
essi
on,a
nd
prom
otin
gth
eex
pres
sion
ofB
cl-2
”
Liu
(201
0)[5
]Sc
hwan
nce
lls
Hu
angq
i(R
adix
Ast
raga
li),D
ansh
en(R
adix
Salv
iae
milt
iorr
hiz
ae),
Shan
yao
(Rh
izom
aD
iosc
orea
e),
and
com
pou
nds
ofth
ree
her
balm
edic
ines
Non
e
Flow
cyto
met
erfo
rap
opto
sis
rate
ofSc
hwan
nce
lls;r
eal-
tim
eP
CR
for
expr
essi
onof
Bcl
-2an
dca
spas
e-3
mR
NA
;Wes
tern
blot
tin
gfo
rex
pres
sion
ofB
cl-2
and
casp
ase-
3pr
otei
n
Dec
reas
edap
opto
sis
rate
;in
crea
sed
Bcl
-2m
RN
Aan
dpr
otei
nex
pres
sion
;dec
reas
edca
spas
e-3
mR
NA
expr
essi
onin
astr
agal
us,
salv
iaan
dco
mpo
un
dgr
oups
;de
crea
sed
expr
essi
onof
casp
ase-
3pr
otei
nin
astr
agal
us,
yam
,an
dco
mpo
un
dgr
oups
.
“Th
eap
opto
sis
ofSc
hwan
nce
llco
-cu
ltu
red
wit
hen
doth
elia
lcel
lin
hig
hgl
uco
seca
nbe
prot
ecte
dby
Ch
ines
eh
erbs
,diff
eren
th
erbs
hav
edi
ffer
ent
effec
t,th
eco
mpo
un
din
terv
enti
ongr
oup
was
the
best
”
Zh
ao(2
011)
[6]
Schw
ann
cells
Seru
mco
nta
inin
gJi
nm
aito
ng
(Sin
ew-C
han
nel
Un
obst
ruct
ion
)
Vit
amin
C
Imm
un
oflu
ores
cen
cem
eth
odfo
rth
eex
pres
sion
ofiN
OS;
real
-tim
efl
uor
esce
nce
quan
tita
tive
PC
Rfo
rp2
2-ph
oxm
RN
Aex
pres
sion
Dec
ease
dex
pres
sion
ofiN
OS
and
p22-
phox
mR
NA
ofSc
hwan
nce
lls
“Can
dow
n-r
egu
late
the
expr
essi
onof
iNO
Spr
otei
nof
NA
DP
Hox
idat
ive
p22-
phox
subu
nit
mR
NA
ofSc
hwan
nce
llscu
ltu
red
inh
igh
ergl
uco
sem
ediu
m”
Pia
o(2
011)
[7]
Schw
ann
cells
Seru
mco
nta
inin
gJi
nm
aito
ng
(Sin
ew-C
han
nel
Un
obst
ruct
ion
)
Vit
amin
C
En
zym
e-lin
ked
imm
un
oabs
orba
nt
assa
yfo
r8-
OH
dGle
vel;
imm
un
oflu
ores
cen
cefo
rex
pres
sion
ofca
spas
e-3
prot
ein
;rea
l-ti
me
flu
ores
cen
cequ
anti
tati
veP
CR
for
expr
essi
onof
casp
ase-
3m
RN
A
Dec
reas
ed8-
OH
dGle
vel;
decr
ease
dex
pres
sion
ofca
spas
e-3
prot
ein
and
mR
NA
inth
esu
pern
atan
tof
Schw
ann
cells
“Can
impr
ove
hig
h-g
luco
sein
duce
dox
idat
ive
inju
ryof
DN
Aap
opto
sis
inSc
hwan
nce
lls,s
ugg
esti
ng
itm
igh
tim
prov
eox
idat
ive
inju
ryan
dap
opto
sis
indi
abet
icn
euro
path
y”
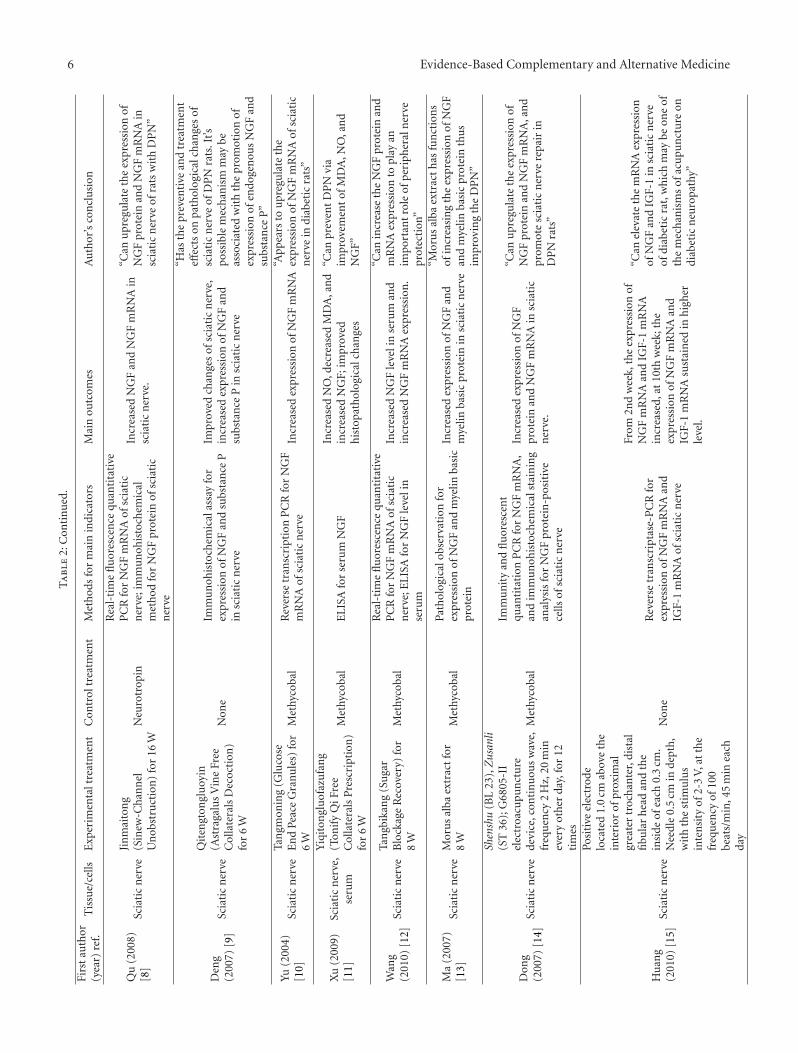
6 Evidence-Based Complementary and Alternative MedicineT
abl
e2:
Con
tin
ued
.
Firs
tau
thor
(yea
r)re
f.T
issu
e/ce
llsE
xper
imen
talt
reat
men
tC
ontr
oltr
eatm
ent
Met
hod
sfo
rm
ain
indi
cato
rsM
ain
outc
omes
Au
thor
’sco
ncl
usi
on
Qu
(200
8)[8
]Sc
iati
cn
erve
Jin
mai
ton
g(S
inew
-Ch
ann
elU
nob
stru
ctio
n)
for
16W
Neu
rotr
opin
Rea
l-ti
me
flu
ores
cen
cequ
anti
tati
veP
CR
for
NG
Fm
RN
Aof
scia
tic
ner
ve;i
mm
un
ohis
toch
emic
alm
eth
odfo
rN
GF
prot
ein
ofsc
iati
cn
erve
Incr
ease
dN
GF
and
NG
Fm
RN
Ain
scia
tic
ner
ve.
“Can
upr
egu
late
the
expr
essi
onof
NG
Fpr
otei
nan
dN
GF
mR
NA
insc
iati
cn
erve
ofra
tsw
ith
DP
N”
Den
g(2
007)
[9]
Scia
tic
ner
ve
Qit
engt
ongl
uoy
in(A
stra
galu
sV
ine
Free
Col
late
rals
Dec
octi
on)
for
6W
Non
eIm
mu
noh
isto
chem
ical
assa
yfo
rex
pres
sion
ofN
GF
and
subs
tan
ceP
insc
iati
cn
erve
Impr
oved
chan
ges
ofsc
iati
cn
erve
,in
crea
sed
expr
essi
onof
NG
Fan
dsu
bsta
nce
Pin
scia
tic
ner
ve
“Has
the
prev
enti
vean
dtr
eatm
ent
effec
tson
path
olog
ical
chan
ges
ofsc
iati
cn
erve
ofD
PN
rats
.It’s
poss
ible
mec
han
ism
may
beas
soci
ated
wit
hth
epr
omot
ion
ofex
pres
sion
ofen
doge
nou
sN
GF
and
subs
tan
ceP
”
Yu(2
004)
[10]
Scia
tic
ner
veTa
ngm
onin
g(G
luco
seE
nd
Peac
eG
ran
ule
s)fo
r6
WM
ethy
coba
lR
ever
setr
ansc
ript
ion
PC
Rfo
rN
GF
mR
NA
ofsc
iati
cn
erve
Incr
ease
dex
pres
sion
ofN
GF
mR
NA
“App
ears
tou
preg
ula
teth
eex
pres
sion
ofN
GF
mR
NA
ofsc
iati
cn
erve
indi
abet
icra
ts”
Xu
(200
9)[1
1]Sc
iati
cn
erve
,se
rum
Yiq
iton
glu
ofaz
ufa
ng
(Ton
ify
QiF
ree
Col
late
rals
Pre
scri
ptio
n)
for
6W
Met
hyco
bal
ELI
SAfo
rse
rum
NG
FIn
crea
sed
NO
,dec
reas
edM
DA
,an
din
crea
sed
NG
F;im
prov
edh
isto
path
olog
ical
chan
ges
“Can
prev
ent
DP
Nvi
aim
prov
emen
tof
MD
A,N
O,a
nd
NG
F”
Wan
g(2
010)
[12]
Scia
tic
ner
veTa
ngb
ikan
g(S
uga
rB
lock
age
Rec
over
y)fo
r8
WM
ethy
coba
l
Rea
l-ti
me
flu
ores
cen
cequ
anti
tati
veP
CR
for
NG
Fm
RN
Aof
scia
tic
ner
ve;E
LISA
for
NG
Fle
veli
nse
rum
Incr
ease
dN
GF
leve
lin
seru
man
din
crea
sed
NG
Fm
RN
Aex
pres
sion
.
“Can
incr
ease
the
NG
Fpr
otei
nan
dm
RN
Aex
pres
sion
topl
ayan
impo
rtan
tro
leof
peri
pher
aln
erve
prot
ecti
on”
Ma
(200
7)[1
3]Sc
iati
cn
erve
Mor
us
alba
extr
act
for
8W
Met
hyco
bal
Path
olog
ical
obse
rvat
ion
for
expr
essi
onof
NG
Fan
dm
yelin
basi
cpr
otei
n
Incr
ease
dex
pres
sion
ofN
GF
and
mye
linba
sic
prot
ein
insc
iati
cn
erve
“Mor
us
alba
extr
act
has
fun
ctio
ns
ofin
crea
sin
gth
eex
pres
sion
ofN
GF
and
mye
linba
sic
prot
ein
thu
sim
prov
ing
the
DP
N”
Don
g(2
007)
[14]
Scia
tic
ner
ve
Shen
shu
(BL
23),
Zus
anli
(ST
36);
G68
05-I
Iel
ectr
oacu
pun
ctu
rede
vice
,con
tin
uou
sw
ave,
freq
uen
cy2
Hz,
20m
inev
ery
othe
rda
y,fo
r12
tim
es
Met
hyco
bal
Imm
un
ity
and
flu
ores
cen
tqu
anti
tati
onP
CR
for
NG
Fm
RN
A,
and
imm
un
ohis
toch
emic
alst
ain
ing
anal
ysis
for
NG
Fpr
otei
n-p
osit
ive
cells
ofsc
iati
cn
erve
Incr
ease
dex
pres
sion
ofN
GF
prot
ein
and
NG
Fm
RN
Ain
scia
tic
ner
ve.
“Can
upr
egu
late
the
expr
essi
onof
NG
Fpr
otei
nan
dN
GF
mR
NA
,an
dpr
omot
esc
iati
cn
erve
repa
irin
DP
Nra
ts”
Hu
ang
(201
0)[1
5]Sc
iati
cn
erve
Posi
tive
elec
trod
elo
cate
d1.
0cm
abov
eth
ein
teri
orof
prox
imal
grea
ter
troc
han
ter,
dist
alfi
bula
rh
ead
and
the
insi
deof
each
0.3
cm.
Nee
dle
0.5
cmin
dept
h,
wit
hth
est
imu
lus
inte
nsi
tyof
2-3
V,a
tth
efr
equ
ency
of10
0be
ats/
min
,45
min
each
day
Non
eR
ever
setr
ansc
ript
ase-
PC
Rfo
rex
pres
sion
ofN
GF
mR
NA
and
IGF-
1m
RN
Aof
scia
tic
ner
ve
From
2nd
wee
k,th
eex
pres
sion
ofN
GF
mR
NA
and
IGF-
1m
RN
Ain
crea
sed,
at10
thw
eek;
the
expr
essi
onof
NG
Fm
RN
Aan
dIG
F-1
mR
NA
sust
ain
edin
hig
her
leve
l.
“Can
elev
ate
the
mR
NA
expr
essi
onof
NG
Fan
dIG
F-1
insc
iati
cn
erve
ofdi
abet
icra
t,w
hic
hm
aybe
one
ofth
em
ech
anis
ms
ofac
upu
nct
ure
ondi
abet
icn
euro
path
y”
Evidence-Based Complementary and Alternative Medicine 7
Ta
ble
2:C
onti
nu
ed.
Firs
tau
thor
(yea
r)re
f.T
issu
e/ce
llsE
xper
imen
talt
reat
men
tC
ontr
oltr
eatm
ent
Met
hod
sfo
rm
ain
indi
cato
rsM
ain
outc
omes
Au
thor
’sco
ncl
usi
on
Yin
(200
8)[1
6]Sc
iati
cn
erve
Mox
aed
atth
epo
ints
ofY
ishu
(Ex-
B3)
and
Zus
anli
(ST
36),
15m
inea
chpo
int,
once
daily
for
56co
nse
cuti
veda
ys
Non
eN
euro
elec
trop
hysi
olog
ical
dete
ctio
nfo
rSN
CV
;ELI
SAfo
rN
GF
ofsc
iati
cn
erve
Dec
reas
edbl
ood
glu
cose
leve
l;in
crea
sed
SNC
V;i
ncr
ease
dN
GF
con
ten
tin
scia
tic
ner
ve.
“Th
eim
prov
ing
effec
tof
mox
ibu
stio
non
diab
etic
per
iph
eral
neu
rolo
gica
lsym
ptom
sin
ara
tm
odel
ofD
PN
may
bere
late
dto
anin
crea
sein
the
NG
Fco
nte
nt
and
prom
otio
nof
per
iph
eral
neu
ropr
otec
tion
”
Wan
g(2
010)
[17]
Scia
tic
ner
veJi
nm
aito
ng
(Sin
ew-C
han
nel
Un
obst
ruct
ion
)fo
r8
WN
euro
trop
in
Th
ehy
drot
her
mal
tail-
flic
kan
dpa
inth
resh
old
tom
ech
anic
alst
imu
lati
on;S
AB
Cim
mu
noh
isto
chem
istr
ym
eth
odfo
rC
NT
Fex
pres
sion
and
real
-tim
efl
uor
esce
nce
quan
tita
tive
PC
Rfo
rC
NT
Fm
RN
Aex
pres
sion
insc
iati
cn
erve
Th
epa
inth
resh
olds
wer
era
ised
and
tail-
flic
kla
ten
cies
wer
esh
orte
ned
;th
eex
pres
sion
ofC
NT
Fm
RN
Aan
dpr
otei
nw
asin
crea
sed
“Can
obvi
ousl
yu
preg
ula
teth
eex
pres
sion
ofC
NT
Fm
RN
Aan
dpr
otei
nin
the
scia
tic
ner
veof
rats
wit
hn
euro
path
y”
Wan
g(2
010)
[18]
Schw
ann
cells
Seru
mco
nta
inin
gJi
nm
aito
ng
(Sin
ew-C
han
nel
Un
obst
ruct
ion
)
Neu
rotr
opin
SAB
Cim
mu
noh
isto
chem
istr
ym
eth
odfo
rC
NT
Fex
pres
sion
and
real
-tim
efl
uor
esce
nce
quan
tita
tive
PC
Rfo
rC
NT
Fm
RN
Aex
pres
sion
inSc
hwan
nce
lls
Incr
ease
dC
NT
Fan
dC
NT
Fm
RN
Aex
pres
sion
inSc
hwan
nce
lls
“Can
upr
egu
late
the
expr
essi
onof
CN
TF
and
CN
TF
mR
NA
ofra
tSc
hwan
nce
llsin
cult
ure
dh
igh
glu
cose
med
ium
,so
asto
impr
ove
DP
N”
Zen
g(2
005)
[19]
Scia
tic
ner
ve
Xia
okel
ingn
ongs
uoy
e(D
iabe
tes
Agi
lity
Con
cen
trat
ion
Flu
id)
for
8W
Met
hyco
bal
Rel
ativ
equ
anti
tyP
CR
for
IGF
mR
NA
Incr
ease
dex
pres
sion
ofIG
F-1
mR
NA
insc
iati
cn
erve
“Is
invo
lved
inth
ere
gula
tion
ofIG
F-1
mR
NA
expr
essi
on,a
nd
prob
ably
prev
ents
diab
etic
peri
pher
aln
euro
path
yfr
omde
teri
orat
ion”
Xu
(200
9)[2
0]Sc
iati
cn
erve
Qin
gyin
gtan
g(C
lear
ing
Nu
trit
ive
QiD
ecoc
tion
)fo
r10
WM
ethy
coba
lE
LISA
for
IGF-
1in
seru
m;I
GF-
1in
scia
tic
ner
vean
dliv
erIn
crea
sed
IGF-
1le
veli
nse
rum
,liv
eran
dsc
iati
cn
erve
“In
crea
seth
eex
pres
sion
ofIG
F-1
inti
ssu
e,an
dh
ave
effec
tsof
ner
vere
pair
inD
NP
rats
”
Wan
g(2
010)
[21]
Scia
tic
ner
veTa
ngm
onin
g(G
luco
seE
nd
Peac
eG
ran
ule
s)fo
r8
WN
one
Wes
tern
blot
tin
gfo
rex
pres
sion
ofN
GF
and
IGF-
1in
scia
tic
ner
ve;
scia
tic
ult
rast
ruct
ure
obse
rvat
ion
bytr
ansm
issi
onel
ectr
onm
icro
scop
e
Incr
ease
dex
pres
sion
ofN
GF
and
IGF-
1;pa
thol
ogic
alch
ange
sof
scia
tic
ner
vew
ere
impr
oved
bytr
ansm
issi
onel
ectr
onm
icro
scop
e
“Has
som
epr
otec
tive
effec
ton
scia
tic
ner
vein
diab
etic
rats
.Th
em
ech
anis
mm
aybe
rela
ted
toth
eu
preg
ula
tion
ofth
eex
pres
sion
ofN
GF
and
IGF-
1pr
otei
ns.”
Zh
ang
(200
8)[2
2]Se
rum
Xia
oket
ongb
ikel
i(D
iabe
tes
Free
Obs
tru
ctio
nG
ran
ula
)M
ethy
coba
lR
adio
imm
un
oass
ayfo
rIL
-1β
and
TN
F-α
,ELI
SAfo
rC
D54
Red
uce
dth
ele
velo
fIL-
1β,T
NF-α
,an
dC
D54
“Can
relie
veor
impr
ove
diab
etic
peri
pher
aln
erve
inju
ryby
inte
rfer
ing
wit
hin
flam
mat
ion
fact
ors
indi
abet
es”
MT
T:m
ethy
lth
iazo
lylt
etra
zoliu
m;E
LIS
A:e
nzy
me-
linke
dim
mu
noa
bsor
ben
tas
say;
MD
A:m
alon
dial
dehy
de;D
PN
:dia
beti
cp
erip
her
aln
euro
path
y;ST
Z:s
trep
tozo
toci
n;N
GF:
ner
vegr
owth
fact
or;A
LL
:Allo
xan
;P
CR
:pol
ymer
ase
chai
nre
acti
on;C
NT
F:ci
liary
neu
rotr
oph
icfa
ctor
;SN
CV
:sen
sory
ner
veco
ndu
ctio
nve
loci
ty.
8 Evidence-Based Complementary and Alternative MedicineT
abl
e3:
Sum
mar
yof
form
ula
sci
ted
inth
isre
view
.
Firs
tau
thor
(yea
r)re
f.Fo
rmu
laTr
adit
ion
also
urc
eof
form
ula
Trad
itio
nal
indi
cati
onC
ompo
nen
tsFu
nct
ion
Rat
ion
ale
offo
rmu
la
Ji(2
009)
[4]
Jian
gtan
gshu
luof
ang
(Dec
reas
eG
luco
seFr
eeC
olla
tera
lsP
resc
ript
ion
)
Em
piri
calf
orm
ula
ofD
r.D
ash
un
Che
n
DP
N,p
atte
rnof
defi
cien
cyof
qian
dyi
n,fl
uid
defi
cien
cyh
eat
and
bloo
dst
asis
,de
fici
ency
ofbl
ood
turn
ing
into
win
d
Shen
gdih
uang
(Rad
ixR
ehm
ann
iae)
,Sh
anzh
uyu
(Fru
ctu
sC
orn
i),G
ouqi
zi(F
ruct
us
Lyci
i),G
egen
(Rad
ixP
uer
aria
e),H
uang
qi(R
adix
Ast
raga
li),D
ansh
en(R
adix
Salv
iae
milt
iorr
hiz
ae),
Puh
uang
(Pol
len
Typh
ae),
Shui
zhi(
Hir
udo
),H
uang
lian
(Rh
izom
aC
opti
dis)
,C
hant
ui(P
erio
stra
cum
Cic
adae
),Ji
li(F
ruct
us
Trib
uli)
Nou
rish
yin
ton
ify
qi,
clea
rh
eat
and
moi
sten
dryn
ess,
acti
vate
bloo
dan
dre
mov
est
asis
,dis
pel
win
dan
dfr
eeco
llate
rals
.
Ch
ief:
shen
gdih
uang
,sha
nzhu
yu,g
ouqi
zian
dge
gen
rege
nde
rfl
uid
and
clea
rh
eat,
nou
rish
yin
ofliv
er,k
idn
eyan
dlu
ng.
Dep
uty
:hua
ngqi
ton
ifies
qi;d
ansh
en,
puhu
ang
and
shui
zhir
emov
est
asis
and
free
colla
tera
ls.
Ass
ista
nt:
huan
glia
ncl
ears
hea
t;ch
antu
ian
djil
irel
ieve
itch
ing.
Pia
o(2
011)
[7]
Jin
mai
ton
g(S
inew
-Ch
ann
elU
nob
stru
ctio
n)
Em
piri
calf
orm
ula
ofD
r.X
iaoc
hun
Lian
g
DP
N,p
atte
rnof
kidn
eyde
fici
ency
and
bloo
dst
asis
;pat
tern
ofde
fici
ency
ofbo
thyi
nan
dya
ng,
inte
rior
cold
and
stas
is
Tusi
zi(S
emen
Cu
scu
tae)
,Nuz
henz
i(F
ruct
us
Lig
ust
rilu
cidi
),Sh
uizh
i(H
iru
do),
Yanh
usuo
(Rh
izom
aC
oryd
alis
),H
uan
gqi(
Rad
ixA
stra
gali)
,Sh
engd
ihu
ang
(Rad
ixR
ehm
ann
iae)
,Gui
zhi(
Ram
ulu
sC
inn
amom
i),X
ixin
(Her
baA
sari
)
Ton
ify
kidn
ey,
acti
vate
bloo
d,w
arm
and
un
bloc
kch
ann
els
and
vess
els.
Ch
ief:
tusi
zito
nifi
esya
ng
and
nou
rish
esyi
nof
kidn
ey,s
ecu
res
esse
nce
and
impr
oves
visi
on,a
nd
chec
ksdi
arrh
ea;
nuzh
enzi
nou
rish
esyi
nof
liver
and
kidn
ey,a
nd
clea
rsem
pty-
hea
t.D
epu
ty:s
huiz
hian
dya
nhus
uobr
eak
bloo
d,ex
pels
tasi
san
dre
lieve
pain
.A
ssis
tan
t:hu
angq
ian
dsh
engd
ihua
ngto
nif
yqi
and
nou
rish
yin
;gui
zhia
nd
xixi
nw
arm
and
un
bloc
kch
ann
els
and
vess
els,
and
prom
ote
qian
dbl
ood
circ
ula
tion
.
Den
g(2
007)
[9]
Qit
engt
ongl
uoy
in(A
stra
galu
sV
ine
Free
Col
latr
als
Dec
octi
on)
Em
piri
calf
orm
ula
DP
N,p
atte
rnof
qide
fici
ency
and
bloo
dst
asis
Hua
ngqi
(Rad
ixA
stra
gali)
,Hua
ngba
i(C
orte
xP
hel
lode
ndr
i),N
iuxi
(Rad
ixA
chyr
anth
isbi
den
tata
e),J
ixue
teng
(Cau
lisSp
ath
olob
i),C
angz
hu(R
hiz
oma
Atr
acty
lodi
s),Y
iyir
en(S
emen
Coi
cis)
,Qin
gdai
(In
digo
Nat
ura
lis)
Ton
ify
qi,a
ctiv
ate
bloo
dan
dre
mov
est
asis
.
Not
men
tion
edab
out
trad
itio
nal
rati
onal
e.T
he
auth
ors
stat
edth
at“B
ased
onm
oder
nph
arm
acol
ogic
alre
sear
ch.
Hua
ngqi
stre
ngt
hen
imm
un
ity;
jixue
teng
and
niux
imod
ula
tem
icro
-cir
cula
tion
;hu
angb
ai,c
angz
hu,n
iuxi
and
yiyi
ren
hav
efu
nct
ion
sof
regu
lati
ng
glu
cose
.”
Xu
(200
9)[1
1]
Yiq
iton
glu
ofaz
ufa
ng
(Ton
ify
QiF
ree
Col
late
rals
Pre
scri
ptio
n)
Em
piri
calf
orm
ula
DP
N,p
atte
rnof
kidn
eyde
fici
ency
,qi
defi
cien
cyan
dob
stru
ctio
nof
colla
tera
lves
sels
.
Shud
ihua
ng(R
adix
Reh
man
nia
epr
aepa
rata
),H
esho
uwu
(Rad
ixPo
lygo
nim
ult
iflor
i),L
uosh
iten
g(C
aulis
Trac
hel
osp
erm
i),H
uang
qi(R
adix
Ast
raga
li),D
ilong
(Lu
mbr
icu
s),W
ugon
g(S
colo
pen
dra)
,Q
uanx
ie(S
corp
io)
Nou
rish
yin
,ton
ify
kidn
eyan
dfr
eeco
llate
rals
Ch
ief:
shud
ihua
ngan
dhe
shou
wu
nou
rish
yin
and
ton
ify
kidn
ey.
Dep
uty
:luo
shit
eng
and
huan
gqit
onif
yqi
and
free
colla
tera
ls.
Ass
ista
nt:
dilo
ng,w
ugon
gan
dqu
anxi
efr
eeco
llate
rals
and
relie
vepa
in.
Wan
g(2
010)
[12]
Tan
gbik
ang
(Su
gar
Blo
ckag
eR
ecov
ery)
Ava
riat
ion
ofH
uan
gqig
uiz
hi-
Wuw
uta
ng
(Ast
raga
lus-
Cin
nam
omu
mFi
ve-I
ngr
edie
nt
Dec
octi
on)∗
Blo
odim
pedi
men
t,ex
hib
itin
gn
um
bnes
sse
nsa
tion
insk
inan
dlim
bs,s
ligh
tly
chop
pyan
dti
ght
puls
e.
Hua
ngqi
(Rad
ixA
stra
gali)
,Nuz
henz
i(F
ruct
us
Lig
ust
rilu
cidi
),G
uizh
i(R
amu
lus
Cin
nam
omi)
,Chi
shao
(Rad
ixPa
eon
iae
rubr
a),H
uang
qin
(Rad
ixSc
ute
llari
ae),
Hua
nglia
n(R
hiz
oma
Cop
tidi
s),S
huiz
hi(H
iru
do)
Ton
ify
qian
den
rich
yin
,det
oxif
yan
dre
solv
est
asis
.
Hua
ngqi
ton
ifies
qian
dst
ren
gth
ens
defe
nce
-ext
erio
r;gu
izhi
war
ms
the
chan
nel
san
dfr
eeya
ng;
nuzh
enzi
nou
rish
esyi
nof
liver
and
kidn
ey;c
hish
aodi
ssip
ates
stas
isan
dre
lieve
spa
in;
huan
gqin
and
huan
glia
ncl
ear
hea
t,dr
yda
mpn
ess
and
deto
xify
;shu
izhi
brea
ksbl
ood
and
expe
lsst
asis
.
Evidence-Based Complementary and Alternative Medicine 9
Ta
ble
3:C
onti
nu
ed.
Firs
tau
thor
(yea
r)re
f.Fo
rmu
laTr
adit
ion
also
urc
eof
form
ula
Trad
itio
nal
indi
cati
onC
ompo
nen
tsFu
nct
ion
Rat
ion
ale
offo
rmu
la
Zen
g(2
005)
[19]
Xia
okel
ingn
ongs
uoy
e(D
iabe
tes
Agi
lity
Con
cen
trat
ion
Flu
id)
Em
piri
calf
orm
ula
DP
N,p
atte
rnof
defi
cien
cyof
qian
dyi
nan
dbl
ood
stas
is
Sang
shen
(Fru
ctu
sM
ori)
,Sh
engd
ihua
ng(R
adix
Reh
man
nia
e),
Geg
en(R
adix
Pu
erar
iae)
,Hua
ngqi
(Rad
ixA
stra
gali)
,Ren
shen
(Rad
ixG
inse
ng)
,Xua
nshe
n(R
adix
Scro
phu
lari
ae),
Shui
zhi(
Hir
udo
)
Nou
rish
kidn
eyyi
n,
ton
ify
qian
dac
tiva
tebl
ood.
Ch
ief:
sang
shen
and
shen
gdih
uang
nou
rish
yin
and
ton
ify
kidn
ey.
Dep
uty
:xua
nshe
nan
dge
gen
nou
rish
yin
and
enge
nde
rfl
uid
.A
ssis
tan
t:hu
angq
ian
dre
nshe
n,to
nif
yqi
;sh
uizh
iis
assi
stan
t,ac
tiva
tebl
ood
and
rem
ove
stas
is.
Xu
(200
9)[2
0]
Qin
gyin
gtan
g(C
lear
ing
Nu
trit
ive
Qi
Dec
octi
on)
From
Wen
bin
gtia
obia
n(S
yste
mat
ized
Iden
tifi
cati
onof
War
mPa
thog
enD
isea
ses)
Hea
tin
the
nu
tien
tas
pect
patt
ern
,ex
hib
itin
gfe
ver
wh
ich
ish
igh
erat
nig
ht,
occa
sion
alde
liriu
m,
rest
less
nes
s,in
som
nia
,th
irst
orn
oth
irst
,ski
nra
shes
,dry
tou
nge
and
fin
era
pid
puls
e.
Shui
niuj
iao
(Cor
nu
Bu
bali)
,Sh
engd
ihua
ng(R
adix
Reh
man
nia
e),
Xua
nshe
n(R
adix
Scro
phu
lari
ae),
Zhu
ye(F
oliu
mP
hyllo
stac
hys
nig
rae)
,M
aido
ng(R
adix
Oph
iopo
gon
is),
Dan
shen
(Rad
ixSa
lvia
em
iltio
rrh
izae
),H
uang
lian
(Rh
izom
aC
opti
dis)
,Jin
yinh
ua(F
los
Lon
icer
ae),
Lian
qiao
(Fru
ctus
Fors
yth
iae)
Nou
rish
yin
and
disp
elh
eat
and
acti
vate
bloo
dan
dfr
eeth
ech
ann
els.
Ch
ief:
shui
niuj
iao
and
shen
gdih
uacl
ear
the
hea
tin
nu
trie
nt
aspe
ctan
dco
olth
ebl
ood.
Dep
uty
:xua
nshe
nan
dm
aido
ngn
ouri
shyi
nan
dcl
ear
hea
t.A
ssis
tan
t:jin
yinh
ua,l
ianq
iao,
huan
glia
nan
dzh
uye
clea
rh
eat
and
deto
xify
,pr
omot
eto
disp
elth
epa
thog
enfr
omn
utr
ien
tas
pect
toqi
aspe
ct;d
ansh
enac
tiva
tes
bloo
dan
ddi
spel
stas
isan
dh
eat.
Wan
g(2
010)
[21]
Tan
gmon
ing
(Glu
cose
En
dPe
ace
Gra
nu
les)
Em
piri
calf
orm
ula
DP
N,p
atte
rnof
qide
fici
ency
and
bloo
dst
asis
Hua
ngqi
(Rad
ixA
stra
gali)
,Yan
husu
o(R
hiz
oma
Cor
ydal
is),
Sanq
i(R
adix
Not
ogin
sen
g),C
hish
ao(R
adix
Paeo
nia
eru
bra)
,Dan
shen
(Rad
ixSa
lvia
em
iltio
rrh
izae
),C
huan
xion
g(R
hiz
oma
Ch
uan
xion
g),H
ongh
ua(F
los
Car
tham
i),S
umu
(Lig
nu
mSa
ppan
),Ji
xuet
eng
(Cau
lisSp
ath
olob
i)
Ton
ify
qi,a
ctiv
ate
bloo
d,fr
eeco
llate
rals
and
relie
vepa
in.
Ch
ief:
huan
gqit
onifi
esqi
and
prom
otes
bloo
dci
rcu
lati
on.
Dep
uty
:yan
husu
oan
dch
uanx
iong
rem
ove
stas
isan
dre
gen
der
fres
hbl
ood.
Ass
ista
nt:
sanq
irem
oves
stas
isan
dre
lieve
pain
;dan
shen
,hon
ghua
and
jixue
teng
rem
ove
stas
isan
dfr
eeco
llate
rals
.
Zh
ang
(200
8)[2
2]
Xia
oket
ongb
i(D
iabe
tes
Free
Obs
tru
ctio
nG
ran
ula
)
Em
piri
calf
orm
ula
ofD
r.R
ongf
aZ
han
g
DP
N,p
atte
rnof
defi
cien
cyof
qian
dyi
nan
dbl
ood
stas
is
Hua
ngqi
(Rad
ixA
stra
gali)
,Dan
ggui
(Rad
ixA
nge
licae
sin
ensi
s),G
egen
(Rad
ixP
uer
aria
e),W
eilin
gxia
n(R
adix
Cle
mat
idis
),Z
ushi
ma
(Cor
tex
Dap
hn
egi
rald
ii),
Sang
baip
i(C
orte
xM
ori)
Ton
ify
qi,a
ctiv
ate
bloo
dan
dre
mov
est
asis
,fre
ech
ann
els
and
relie
vepa
in.
Ch
ief:
huan
gqi,
dang
guia
nd
gege
nto
nif
yqi
,act
ivat
ebl
ood
and
relie
vepa
in.
Dep
uty
:wei
lingx
ian
and
zush
ima
disp
elw
ind
and
relie
vepa
in.
Ass
ista
nt:
sang
baip
icle
ars
lun
gh
eat.
∗fr
omJi
ngu
iyao
lue
(Syn
opsi
sof
Pre
scri
ptio
ns
ofth
eG
olde
nC
ham
ber)
,wh
ich
ingr
edie
nts
are
huan
gqi(
Rad
ixA
stra
gali)
,gui
zhi(
Ram
ulu
sC
inn
amom
i),s
haoy
ao(R
adix
Paeo
nia
e),s
heng
jiang
(Rh
izom
aZ
ingi
beri
sre
cen
s)an
dda
zao
(Fru
ctu
sJu
juba
e).

10 Evidence-Based Complementary and Alternative Medicine
ciliary neurotrophic factor [CNTF] and interleukins); andthe transforming growth factor-beta (TGF-β) superfamily(which can be subdivided into acidic fibroblast growthfactors and basic fibroblast growth factors). In addition, thereare other neurotrophic factors, such as insulinlike growthfactor (IGF) and glial-derived neurotrophic factor [27].Present studies suggest that diabetes-induced dysfunction ofnerve regeneration results partially from decreased levels ofsome neurotrophic factors or their receptors.
3.1. NGF. NGF was the first discovered and most typicalneurotrophic factor. It plays an important role in neuronaldevelopment, differentiation, and the maintenance of nor-mal functions. NGF not only protects neurons and reducestheir degeneration and death, but it also promotes nerveregeneration after nerve injury. There is a deficiency of NGFin diabetes, and reduced levels or activity of NGF plays asignificant role in the pathogenesis of diabetic neuropathy[28]. Qu et al. [8] observed that 12 weeks after the successof a STZ-induced diabetic neuropathy rat model, comparedwith the normal rats, the tail-flick latency was significantlyprolonged, the pain threshold was significantly lower, andNGF protein and mRNA expression in the sciatic nerve weresignificantly reduced in the model rats. Moreover, NGF-mRNA expression level in the sciatic nerve was negativelycorrelated with the tail-flick latency and was positively cor-related with the mechanical pain threshold in the model rats.The model rats were orally administered with the Chinesemedicine compound Jinmaitong in three different dosagegroups: large, medium, and small. After the intervention, themedium-dosage group of Jinmaitong showed that the tail-flick latency was significantly reduced, the pain thresholdsignificantly increased, and sciatic NGF-mRNA and proteinexpression were significantly increased, compared with themodel control group. Furthermore, in vitro study provedthat the serum containing Jinmaitong promotes secretionof NGF in high-glucose cultured Schwann cells [2]. Dengand Zhang [9] investigated the effects of Qitengtongluoyinon protein expression of NGF and neuropeptide substanceP (SP) in sciatic nerves of STZ-induced diabetic multipleneuropathy rats, and proved that Qitengtongluoyin canprevent and treat sciatic neuropathy in STZ-induced dia-betic multiple neuropathy rats, probably via promoting theexpression of NGF and SP protein. Yu et al. [10] reported thata 6-week intervention of the Chinese medicine compoundTangmoning in STZ-induced diabetic rats resulted in asignificant increase of NGF mRNA and that the effect wassimilar to that of methycobal. Xu and Yang [11] reported thatYishentongluofazufang can increase the NGF content in theserum of STZ-induced rats. Wang and Liu [12] proved thatTangbikang can increase serum NGF level and increase theexpression of NGF mRNA of sciatic nerve in diabetic rats. Maet al. [13] testified in their experimental study that certainextracts of morus alba can improve diabetic peripheralneuropathy in alloxan-induced diabetic rats, via promotingexpression of NGF and myelin protein in sciatic nerves.
Besides the above-mentioned Chinese herbal medicinestudies, recent studies have shown that acupuncture andmoxibustion can improve DPN, possibly via their effects on
NGF. Dong et al. [14] conducted electroacupuncture (EA) onthe points of Shenshu (BL 23) and Zusanli (ST 36) in STZ-induced diabetic rats. After the intervention was done 12times, the EA group showed increased NGF-positive cells andincreased NGF mRNA expression in sciatic nerve, comparedwith the model group, suggesting EA upregulates expressionof protein and mRNA of NGF and improves nerve repair inDPN. Huang et al. [15] found that EA treatment of diabeticrats resulted in increased expression of NGF mRNA in thesciatic nerve. Yin et al. [16] applied moxibustion on STZ-induced diabetic rats, moxaed at the points of Yishu (Ex-B3) and Zusanli (ST 36), 15 min each point, once daily for56 consecutive days, found that blood glucose significantlydecreased, nerve conduction velocity significantly increased,and NGF content significantly increased in treatment group,compared with those in model group, suggesting thatmoxibustion has functions of peripheral nerve protectionwhich may be related to its promotion of NGF expressionof nerve.
3.2. CNTF. CNTF has multiple biological activities: pro-moting survival of neurons and protecting motor neurons;inhibiting degeneration of axons of motor nerves; enhanc-ing growth speed of axon; preventing muscle atrophy. Inaddition, administration of CNTF results in promotingregeneration of peripheral nerves [29]. CNTF protein andbioactivity are reduced in the peripheral nerve of diabeticrats, and CNTF treatment improved nerve regeneration andprevented nerve-conduction slowing in diabetic rats, sug-gesting CNTF plays an important role in nerve regenerationin DPN [30]. Wang et al. [17] observed the Chinese medicinecompound Jinmaitong’s effects on CNTF expression indiabetic neuropathy rats, and confirmed that Jinmaitongcan upregulate the expression of the protein and mRNA ofCNTF in the sciatic nerves of diabetic neuropathy rats. Forin vitro study, Wang et al. [18] prepared drug-containingserums with the application of 15 times the adult dosage ofboth Jinmaitong and neurotrophin and grouped as the blankcontrol group (no cells added), normal control group (addedwith normal rat serum), high-glucose group (added withglucose), Jinmaitong group (added with serum containingJinmaitong and normal rat serum), and neurotropin group(added with serum containing neurotropin). Except forthe blank control group and normal control group, the50 mmol/L glucose was added to all the groups to achievehigh-glucose Schwann cell models. The expression of CNTFand CNTF mRNA was detected by SABC immunohisto-chemistry method and real-time fluorogenetic quantitativePCR, respectively. Results showed that compared withthe normal control group, the CNTF and CNTF mRNAexpression in the high-glucose group, Jinmaitong group, andneurotropin group were lowered. Compared with the high-glucose group, the CNTF and CNTF mRNA expression ofthe Jinmaitong group and neurotropin group were increased,and the CNTF mRNA expression in the Jinmaitong groupwas higher than that in the neurotropin group. This suggeststhat Jinmaitong upregulates the expression of CNTF andCNTF mRNA of Schwann cells cultured in high-glucosemedium.

Evidence-Based Complementary and Alternative Medicine 11
Chinese medicine
Apoptosis Proliferation
Schwann cells
Caspase-3iNOSNADPH oxidase
Bcl-2 NGF CNTF IGF-1 IL-1
Neurotrophic factorsOxidative injury
Bax SP
JMT, AS
JTSL
CASY
JMT JMT JMTJMT, QT
TM, YQ
TB, MA
EA, MX
JTSL, AS
Yam, SA
CASY
XTQTXKL
QY
TM
EA
JTSL
AS: astragalus
CASY: compound of astragalus, salvia, and yam
EA: electroacupunctureJMT: jinmaitongJTSL: jiangtangshulufangMA: morus alba extractMX: moxibustionQT: qitengtongluoyin
QY: qingyingtangSA: salviaTB:tangbikangTM: tangmoningXKL: xiaokelingXT: xiaoketongbikeliYQ: yiqitongluozufang
Figure 2: Chinese medicine’s effects on Schwann cells and neurotrophic factors and its possible mechanisms of promoting nerve repair andregeneration in diabetic peripheral neuropathy.
3.3. IGF-1. IGF-1 promotes cell growth and proliferation,and it promotes the growth of axons. Recent studies haveshown that IGF-1 nourishes and supports motor andautonomic nerves. Decreased serum IGF-1 level and IGF-1 mRNA expression are shown in experimental diabeticrats, and administration of IGF-1 results in improvementof diabetic neuropathy [31]. Zeng et al. [19] applied reversetranscription polymerase chain reaction assay and confirmedthat the expression of IGF-1 mRNA was decreased in sciaticnerves in STZ-induced diabetic rats; the expression levelof IGF-1 mRNA and glucose was negatively correlated;treatment with Xiaokelingnongsuoye resulted in increasedexpression of IGF-1 mRNA in sciatic nerves. Xu et al. [20] setup a rat model of diabetic peripheral neuropathy and inves-tigated Qingyingtang’s effects on sciatic nerve conductionvelocity, histopathological changes, and the expression ofIGF-1 in tissue. They found that Qingyingtang enhanced sci-atic nerve conduction velocity, improved histopathologicalchanges, and increased the expression of IGF-1 in serum andtissue, confirming that Qingyingtang nourishes the sciaticnerve and promotes the sciatic nerve repair, possibly viaincreasing expression of IGF-1. Another study reported thatTangmoning can improve pathological changes in the sciaticnerve in rats, and it has protective effects on DPN, whichmight be related to its upregulation of the expression ofIGF-1 protein [21]. Yin et al. [16] proved that moxibustion
can upregulate IGF-1 mRNA expression of sciatic nerve indiabetic rats.
3.4. Interleukin. Interleukin 1 (IL-1) plays a central rolein the regulation of immune and inflammatory responses.It promotes cell proliferation and generation of othercytokines, and regulates metabolism. Recent studies foundthat IL-1 and IL-6 have functions of promoting regenerationof peripheral nerves; Schwann cell can secret IL-1 whichpromotes Schwann cells’ secretion of NGF; IL-6 can promotenerve regeneration of via up-regulating the expression ofCNTF mRNA [32–34]. It has not been reported that Chinesemedicine promotes nerve repair and regeneration via inter-leukin. Only a few studies reported that Chinese medicineinhibited inflammation factors in diabetes, so that improvedthe nerve impairment in DPN. Zhang et al. [22] administereddifferent dosages of Xiaoketongbi to STZ-induced diabeticrats for 2 months and found that Xiaoketongbi improvedthe peripheral neuropathy and decreased the levels of IL-1β, TNF-α, and CD54 in diabetic peripheral neuropathy rats,suggesting Xiaoketongbi relieves and improves diabetic neu-ropathy by means of inhibition of inflammation in diabetes.
4. Conclusion
Diabetic peripheral neuropathy is the result of multiplefactors, and the repair and regeneration of peripheral

12 Evidence-Based Complementary and Alternative Medicine
nerves are very complicated procedures that are regulatedby multiple factors; furthermore, the micro-environmentthat is needed for nerve repair and regeneration is notof single factor but composed of multiple related fac-tors. Recent studies have shown that Chinese medicineinhibits apoptosis, promotes proliferation in Schwann cells,and increases expression of multiple neurotrophic factors;therefore, Chinese medicine can improve nerve repair andregeneration in DPN (Figure 2). Chinese medicine has theadvantage of providing multiple therapeutic effects on mul-tiple targets, compared with Western medicine, which usesconventional chemical agents and focuses on a single target.Therefore, to a certain extent, the effective single herb orcompound of Chinese medicine might offer a more suitablemicro-environment, one that is neurologically and physical-ly needed for promoting repair and regeneration of nerves.
As we discussed in this paper, some studies investi-gated a single factor, thus, limited in explanations of themechanisms of Chinese medicine’s effects on nerve repairand regeneration. Although some Chinese medicine showedeffectiveness in vitro studies, it might not be effectiveclinically, because in vitro cultured cells independentlysurvive in an artificial environment, which is very differentfrom the environment in vivo. Because of the diversity ofpatterns of Chinese medicine, uncertainty about activitiesof various ingredients, difficulty of quality control, andunknown interactions between components in the sameChinese medicine compound, research on the mechanismsis very difficult to get further. In short, further research isneeded to clarify Chinese medicine’s clinical value and themechanisms of Chinese medicine’s functions of nerve repairand regeneration in DPN. Future studies should be carriedout with emphasis on both prevention and treatments toclarify the mechanisms by which Chinese medicine promotesnerve repair and regeneration; in the meantime, we need toexplore and block the factors that inhibit nerve repair andregeneration.
Acknowledgment
This work was supported by Natural Science Foundation ofBeijing (no. 7122147).
References
[1] X. C. Liang, “Diabetic peripheral neuropathy,” in Guidelinesfor Diagnosis and Treatment of Diabetes with Integrative Med-icine 2010, L. Lin, Ed., pp. 19–27, Military Medical SciencePress, Beijing, China, 1st edition, 2010.
[2] L. Q. Sun, X. C. Liang, H. Zhang et al., “Effects of medicatedserum of Jinmaitong capsule on proliferation and expressionof nerve growth factor in Schwann cells cultured in high glu-cose,” Zhonghua Zhong Yi Yao Zazhi, vol. 24, no. 8, pp. 1019–1022, 2009 (Chinese).
[3] B. S. Wu, W. J. Wang, and W. H. Chen, “Effect of allyl glycosideextracted from Herba Rhodiolae on activity and proliferationof cultured Schwann cells under high glucose condition,”Zhongguo Zhong Xi Yi Jie He Zazhi, vol. 29, no. 2, pp. 153–155,2009 (Chinese).
[4] J. L. Ji, D. S. Chen et al., “Experiment study of nourishing yin,invigorating qi, extinguishing wind, promoting blood circu-lation and draining collateral method of the sciatic nerveSchwann cells apoptosis of diabetic rats,” Zhongguo Zhong YiJi Zheng, vol. 18, no. 8, pp. 1304–1306, 2009 (Chinese).
[5] X. Y. Liu, Y. Y. Zhang, J. Q. Zhang et al., “Effect of astragalus,salvia, yam and its compound on apoptosis of Schwann cellsco-cultured with endothelial cells in high glucose,” Zhong YaoYao Li Yu Lin Chuang, vol. 26, no. 1, pp. 41–44, 2010 (Chinese).
[6] L. Zhao, X. C. Liang, H. Zhang et al., “Effects of Chinese herbalmedicine Jinmaitong-medicated serum on inducible nitricoxide synthase and NADPH oxidase p22-phox subunit of ratSchwann cells cultured in high-glucose medium,” Journal ofChinese Integrative Medicine, vol. 9, no. 3, pp. 299–305, 2011(Chinese).
[7] Y. L. Piao, X. C. Liang, L. Zhao et al., “Effects of medicatedserum containing Jinmaitong on 8-OHdG and active caspase3 of Schwann cells in high glucose medium,” Yi Xue Yan JiuZazhi, vol. 40, no. 10, pp. 35–39, 2011 (Chinese).
[8] L. Qu, X. C. Liang, Q. L. Wu et al., “Effects of Jinmaitong cap-sule on nerve growth factor in diabetic rats,” Zhongguo ZhongYao Za Zhi, vol. 33, no. 21, pp. 2539–2544, 2008 (Chinese).
[9] J. S. Deng and H. Zhang, “Possible preventive and therapeuticmechanism of Qitengtongluoyin on experimental diabeticneuropathy rats,” Zhongguo Lin Chuang Yao Xue Zazhi, vol.16, no. 4, pp. 242–245, 2007 (Chinese).
[10] S. J. Yu, L. Zhang, M. Wang et al., “Effect of Tangmoninggranular preparation on expression of nerve growth factorgene of sciatic nerve in diabetic rats,” Zhonghua Nei Fen MiZazhi, vol. 20, no. 2, pp. 169–170, 2004 (Chinese).
[11] Y. Xu and J. Yang, “Protective mechanisms of kidney-nour-ishing and collateral-unblocking therapy for peripheral neu-ropathy in experimental diabetic rats,” Shanghai Zhong Yi YaoZazhi, vol. 43, no. 9, pp. 67–69, 2009 (Chinese).
[12] J. Wang and T. H. Liu, “Tangbikang’s effects on serum nerverelated factor and expression of never related factor gene indiabetic rats,” Zhongguo Shi Yan Fang Ji Xue Zazhi, vol. 16, no.6, pp. 160–163, 2010 (Chinese).
[13] S. T. Ma, X. K. Zhang et al., “Effect of morus alba extract onnerve growth factor and myelin basic protein in diabetic rats,”Zhongguo Zhong Yi Ji Chu Yi Xue Zazhi, vol. 13, no. 10, pp.760–763, 2007 (Chinese).
[14] H. S. Dong, Q. J. Zhang, Q. Yang et al., “Experimental studyof nerve growth factor of sciatic nerve of rats with diabetesperipheral neuropathy accommodated by electroacupunc-ture,” Zhongguo Kang Fu Li Lun Yu Shi Jian, vol. 12, no. 8, pp.730–732, 2007 (Chinese).
[15] Z. Y. Huang, X. Zou, K. F. Wang et al., “Effect of electroacu-puncture on mRNA expression of NGF and IGF-1 in sciaticnerve of diabetic rats,” Zhong Xi Yi Jie He Yan Jiu, vol. 2, no. 2,pp. 57–60, 2010 (Chinese).
[16] H. Y. Yin, J. Wang, L. B. Xu et al., “Mechanism of peripheralneuroprotection induced by moxibustion treatment in ratswith diabetic peripheral neuropathy,” Shanghai Zhen JiuZazhi, vol. 27, no. 1, pp. 41–44, 2008 (Chinese).
[17] P. Y. Wang, X. C. Liang, L. Q. Sun et al., “Effects of Jinmaitongcapsule on expression of ciliary neurotrophic factor in diabeticrats,” Zhongguo Zhong Xi Yi Ji Jiu Zazhi, vol. 17, no. 1, pp. 3–6,2010 (Chinese).
[18] P. Y. Wang, X. C. Liang, H. Zhang et al., “Effect of Jinmaitongcapsule medicated serum on ciliary neurotrophic factor of ratSchwann cells in high glucose medium,” Zhong Yi Zazhi, vol.51, no. 8, pp. 736–739, 2010 (Chinese).

Evidence-Based Complementary and Alternative Medicine 13
[19] J. Z. Zeng, K. L. Dong, G. C. Li, L. M. Li et al., “Effect of Xi-aokelingnongsuoye on mRNA expression of insulin-likegrowth factor-1 in sciatic nerve of streptozotocin-inducedDiabetic Rats,” Zhong Nan Da Xue Xue Bao (Yi Xue Ban), vol.30, no. 1, pp. 49–52, 2005 (Chinese).
[20] X. P. Xu, J. Yang, P Zhu et al., “Experimental study on nerveprotective effect of Qingyingtang on rats with diabetic periph-eral neuropathy,” Zhongguo Zhong Yi Ji Chu Yi Xue Zazhi, vol.15, no. 11, pp. 836–838, 2009 (Chinese).
[21] W. Wang, S. J. Yu, C. N. Yu et al., “Influences of Tangmoninggranules on sciatic expression of NGF and IGF-1 proteins inrats with diabetic peripheral neuropathy,” Beijing Zhong Yi YaoDa Xue Xue Bao, vol. 33, no. 5, pp. 346–349, 2010 (Chinese).
[22] X. K. Zhang, F. B. Meng, Y. H. Duan et al., “Effects of Xi-aoketongbi granula on IL-α,TNF-β,and CD (54) of diabeticperipheral neuropathy in rats,” Shanxi Yi Xue Zazhi, vol. 37,no. 9, pp. 1136–1137, 2008 (Chinese).
[23] M. W. Kalichman, H. C. Powell, and A. P. Mizisin, “Reactive,degenerative, and proliferative Schwann cell responses inexperimental galactose and human diabetic neuropathy,” ActaNeuropathologica, vol. 95, no. 1, pp. 47–56, 1998.
[24] K. Sango, T. Suzuki, H. Yanagisawa et al., “High glucose-induced activation of the polyol pathway and changes of geneexpression profiles in immortalized adult mouse Schwanncells IMS32,” Journal of Neurochemistry, vol. 98, no. 2, pp. 446–458, 2006.
[25] A. M. Vincent, M. Brownlee, and J. W. Russell, “Oxidativestress and programmed cell death in diabetic neuropathy,”Annals of the New York Academy of Sciences, vol. 959, pp. 368–383, 2002.
[26] C. L. Delaney, J. W. Russell, H. L. Cheng, and E. L. Feldman,“Insulin-like growth factor-I and over-expression of Bcl-xLprevent glucose-mediated apoptosis in Schwann cells,” Journalof Neuropathology and Experimental Neurology, vol. 60, no. 2,pp. 147–160, 2001.
[27] Y. T. Wang and X. M. Lu, “Neurotrophic factors and theirmechanisms in repair of peripheral nerve injury,” Ren Min JunYi, vol. 49, no. 6, pp. 359–361, 2006 (Chinese).
[28] G. Pittenger and A. Vinik, “Nerve growth factor and diabeticneuropathy,” Experimental Diabesity Research, vol. 4, no. 4, pp.271–285, 2003.
[29] J. Zhang, Z. R. Chen, Z. W. Chen, G. J. Zhang et al., “Experi-mental study on enhancement of rat sciatic nerve regenerationby ciliary neurotrophic factor,” Zhonghua Chuang ShangZazhi, vol. 26, no. 4, pp. 239–242, 1999 (Chinese).
[30] A. P. Mizisin, Y. Vu, M. Shuff, and N. A. Calcutt, “Ciliary neu-rotrophic factor improves nerve conduction and amelioratesregeneration deficits in diabetic rats,” Diabetes, vol. 53, no. 7,pp. 1807–1812, 2004.
[31] M. H. Chen, P. R. Chen, M. H. Chen, S. T. Hsieh, and F. H. Lin,“Gelatin-tricalcium phosphate membranes immobilized withNGF, BDNF, or IGF-1 for peripheral nerve repair: An in vitroand in vivo study,” Journal of Biomedical Materials Research—Part A, vol. 79, no. 4, pp. 846–857, 2006.
[32] T. Shuto, H. Horie, N. Hikawa et al., “IL-6 up-regulates CNTFmRNA expression and enhances neurite regeneration,” Neu-roReport, vol. 12, no. 5, pp. 1081–1085, 2001.
[33] D. Lindholm, R. Heumann, M. Meyer, and H. Thoenen,“Interleukin-1 regulates synthesis of nerve growth factor innon-neuronal cells of rat sciatic nerve,” Nature, vol. 330, no.6149, pp. 658–659, 1987.
[34] J. Yang, L. Yu, X. Z. Qiu et al., “Experimental study on the roleof IL-1β in Schwann cells inducing and activation,” ZhonghuaXian Wei Wai Ke Zazhi, vol. 28, no. 2, pp. 142–144, 2005(Chinese).

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 156521, 18 pagesdoi:10.1155/2012/156521
Research Article
Mitigation of H2O2-Induced Mitochondrial-MediatedApoptosis in NG108-15 Cells by Novel Mesuagenin C fromMesua kunstleri (King) Kosterm
Gomathi Chan,1 Muhamad Noor Alfarizal Kamarudin,2
Daniel Zin Hua Wong,2 Nor Hadiani Ismail,3 Faizuri Abdul Latif,4
Aurengzeb Hasan,1 Khalijah Awang,1 and Habsah Abdul Kadir2
1 Department of Chemistry, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia2 Biomolecular Research Group, Biochemistry Program, Institute of Biological Sciences, Faculty of Science, University of Malaya,50603 Kuala Lumpur, Malaysia
3 Faculty of Applied Sciences, Universiti Teknologi MARA, Selangor, 40450 Shah Alam, Malaysia4 Department of Aqidah and Islamic Thoughts, Academy of Islamic Studies, University of Malaya, 50603 Kuala Lumpur, Malaysia
Correspondence should be addressed to Habsah Abdul Kadir, [email protected]
Received 29 February 2012; Revised 16 April 2012; Accepted 20 April 2012
Academic Editor: Monica Rosa Loizzo
Copyright © 2012 Gomathi Chan et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This study was aimed to isolate and evaluate neuroprotective compounds from the hexane extract of the bark of Mesua kunstleri(Clusiaceae) on H2O2-induced apoptosis in NG108-15 cells. Five 4-phenylcoumarins were isolated by using various chromato-graphic techniques via neuroprotective activity-guided fractionation and isolation from the active hexane extract. The chemicalstructures of the isolated compounds were confirmed by NMR spectroscopic data interpretation and comparison with literaturevalues. Cell viability data demonstrated that mesuagenin C 3 significantly increased cell viability. Hoechst 33342/PI staining illus-trated mesuagenin C 3 was able to abate the nuclear shrinkage, chromatin condensation and formation of apoptotic bodies. Pre-treatment with mesuagenin C 3 reduced total annexin V positive cells and increased the level of intracellular glutathione (GSH).Mesuagenin C 3 attenuated membrane potential (Δψm), reduced Bax/Bcl-2 ratio and inactivated of caspase-3/7 and -9. Theseresults indicated that mesuagenin C 3 could protect NG108-15 cells against H2O2-induced apoptosis by increasing intracellularGSH level, aggrandizing Δψm, and modulating apoptotic signalling pathway through Bcl-2 family and caspase-3/7 and -9. Thesefindings confirmed the involvement of intrinsic apoptotic pathway in H2O2-induced apoptosis and suggested that mesuagenin C 3may have potential therapeutic properties for neurodegenerative diseases.
1. Introduction
A decade after stepping into the new millennium, neuro-degenerative diseases still denote as one of world mostarduous and appalling health issues as a result of increasingdemographic fluctuation towards the aged population as wellas growing apprehension for a better quality of life. Suchalertness, together with the huge social and economic costsof the disease, has ignited in-depth research efforts to combator at least delay the onset of neurodegenerative diseases [1].In a normal cycle of tissue proliferation, the aged and oldercells are set to decease in order to give way for the generation
of new cells. Such setting of programmed cell death or betterknown as apoptosis was shown to be the prime cause inneurodegenerative diseases as a number of these diseases arecharacterized by a progressive fading away of neurons.
Apoptosis holds a cardinal role in the maturation of thenervous system as well as neural architecture through theinterplay between both anti- and proapoptotic proteins [2].Basically, neurons either will render an adaptive response orthey would initiate apoptosis when they are exposed tostress signals or apoptotic stimuli such as withdrawal of neu-rotrophic factor, ischemic stroke, misfolded proteins, mito-chondrial-complex inhibition, excessive calcium entry, and

2 Evidence-Based Complementary and Alternative Medicine
excitotoxicity [2–4]. For example, in adult neurodegenerativedisorders the formation of misfolded proteins such as β-amyloid aggregates brings upon oxidative stress and excessiveinflux of intracellular Ca2+ concentration which would even-tually lead to apoptosis [1]. Apoptosis or programmed celldeath is delineated with lucid and perspicuous morpholog-ical features, including cell shrinkage, chromatin conden-sation, loss of nuclear membrane integrity, plasma mem-brane blebbing, and eventually the breaking off of cellularfragments giving rise to the formation of apoptotic bodies[4, 5]. Naturally, apoptosis is required for normal embryonicdevelopment, sustenance of cellular homeostasis, and it hasbeen shown to be implicated in many forms of chronicdiseases. Apoptosis can take place in the body as a meanof defense mechanism in immune reactions when cells areharmed by toxic and deleterious substances [6]. Increasingsubstantial evidence and findings from stem cell model sug-gest that the fate of neural stem particularly apoptosis playsa major role in disease outcomes. Therefore, it is noteworthythat apoptosis orchestrates a crucial role in numerous neuro-degenerative diseases and targeting its pathway would confera mode of prevention and treatment [2, 4].
Generally, apoptosis is regulated by the Bcl-2 family ofproteins which are further grouped into antiapoptotic pro-teins (Bcl-2, Mcl-1) and proapoptotic proteins (Bax, Noxa),the adaptor protein Apaf-1 (apoptotic protease-activatingfactor 1), and the cysteine-aspartyl-specific proteases family(caspases) [5]. Neuronal apoptosis can occur in two commonpathways either by an extrinsic or intrinsic pathway and eachis characterized by the alterations in the mitochondria, theendoplasmic reticulum, and cascade activation of caspases.Upon apoptotic stimuli, the proapoptotic proteins willinduce the alteration in mitochondria which brings aboutthe intensification of the outer mitochondrial membranepermeability eventuating in the generation of mitochondrialpores. The subsequent efflux of cytochrome c will bind tothe Apaf-1, forming an apoptosome which triggers initiatorcaspase such as procaspase-9 and activating it to caspase-9. This in turn initiates a cascade of caspase activationespecially the effector caspases such as caspase-3 and -7which are accountable for proteolytic cleavage of proteinsand events leading to apoptosis [3, 5, 6]. Ergo, the preventionof neuronal apoptosis confers a mode of neuroprotectionin combating and delaying the onset of neurodegenerativediseases.
Nowadays, the emergence of natural products as thera-peutics agents has raised a global interest among scientists indeveloping newer but more importantly safer drugs partic-ularly for neurodegenerative diseases. We have undertakena study on the hexane extract of Mesua kunstleri as aneffort in attaining potential therapeutic compounds from theMalaysian flora [7–9]. Listed under the Clusiaceae family,M. kunstleri is a forest timber plant and locally known as“penaga.” The genus Mesua with more than 40 species isdispersed along the region of Ceylon, India, Indo-China,Thailand, Malaysia, and Queensland, centred strongly inWest Malaysia [10]. Plants that fall under the genus Mesuahave been used for various complementary medicine pur-poses such as antiallergic, rheumatism, antidiarrhoetic, and
antibacterial as they are the fruitful sources of phytochemi-cals such as phloroglucinols, xanthones, neoflavonoids, andcoumarins. These phytochemicals particularly coumarins,which are derivatives of cinnamic acid with the presence ofa benzo-α-pyrene skeleton, have been shown to exert multi-farious biological activities such as neuromodulator, antide-pressant, anti-HIV-1, anti-inflammatory, antitumor, antimi-crobial, antiviral, antifungal, and hepatoprotective effects[11–13]. The bark of M. kunstleri has been locally used tocure dyspepsis, a chronic, recurrent pain centered in theupper abdomen and renal diseases [14]. Recently, 4-phenyl-coumarins including mesuagenin C 3 from the species M.elegans have been shown to possess acetylcholinesterase(AChE) inhibitory activity [8] and cholinesterase inhibitionhas been proven to be one of the effective modes of treatmentfor Alzheimer’s diseases [15]. Withal, the medicinal use ofcoumarins has been intensified through the functionaliza-tion of its aromatic center that paves the way towards thedevelopment of neoteric analogs that are capable of inhibit-ing β-amyloid [16]. In mice model, plant-derived coumarinswere reported to confer the ability to mitigate intrac-erebroventricular injection of β-amyloid-induced memoryimpairment. Osthol from Cnidium monnieri was shown toprotect the NG108-15 by inhibiting the voltage-dependentL-type Ca2+ current [17] whereas isopentenyl-oxycoumarinfrom Citrus species salvaged the neuronal mixed cortical cellculture model against NMDA-induced neurotoxicity [18]. Inprimary cultured rat cortical model, marmesin from Angelicagigas was reported to act as neuroprotective agent in the glu-tamate-induced neurotoxicity [19].
This neuroprotective-activity-guided fractionation andisolation study has driven to the isolation of five 4-phenyl-coumarins, namely, isomammeisin (1) [11], mammea A/BA(2) [20], mesuagenin C (3) [8], 5,7-dihydroxy-8-(2-methyl-butanoyl)-6-[(E)-3,7-dimethylocta-2,6-dienyl]-4-phenyl-2H-chromen-2-one (4) [11], and 5,7-dihydroxy-8-(3-methyl-butanoyl)-6-[(E)-3,7-dimethylocta-2,6-dienyl]-4-phenyl-2H-chromen-2-one (5) [11]. Among all these compounds, com-pound 3 (mesuagenin C) showed a potent neuroprotectiveactivity against H2O2 insult, henceforth it was subjected tovarious assays to elucidate its neuroprotective mechanisms.To the best of our knowledge, this is the first report ofneuroprotective activity in NG108-15 cells by mesuageninC 3. Thus, the novel neuroprotective mesuagenin C 3 couldlead us to the discovery of novel molecules that potentiallycan act as new target sites in combating neurodegenerativediseases.
2. Material and Methods
2.1. Plant Material. The bark of M. kunstleri (King) Kostermwas collected from Rimba Teloi Forest Reserve, Kedah,Malaysia on April 1995. The sample with voucher specimennumber KL 4485 was identified by Mr. Teo Leong Eng anddeposited in the herbarium of the Department of Chemistry,Faculty of Science, University of Malaya.

Evidence-Based Complementary and Alternative Medicine 3
2.2. The Crude Extraction of M. kunstleri Bark. Dried groundbark of M. kunstleri (1.5 kg) was soaked with hexane, ethylacetate, and methanol successively (3 × 4 L, each 48 h) atroom temperature. The extracts were evaporated to drynessusing a rotary evaporator. A yellow gummy residue (110.4 g)was obtained from the hexane extract. A brown gummyresidue of ethyl acetate extract (70.1 g) was obtained whereasa brown amorphous powder (80.3 g)was attained from themethanol extract.
2.3. Neuroprotective-Activity-Guided Fractionation and Iso-lation of Mesuagenin C 3. The hexane, ethyl acetate, andmethanol extracts were tested for their neuroprotective activ-ity through MTT neuroprotective assay. Based on the resultsobtained, the hexane extract was selected for further frac-tionation since it displayed the most potent neuroprotectiveactivity and this was carried out through neuroprotective-activity-guided fractionation and isolation. A portion of thehexane crude (10.0 g) was subjected to column chromatogra-phy fractionation over silica gel (230–400 mesh) and elutedwith hexane : EtOAc (9.5 : 0.5 to 0 : 10) and EtOAc : MeOH(5 : 5) to yield fractions BH1–BH6. Fraction BH1 showed themost potent neuroprotective activity; consequently BH1 wasthen directed to silica gel chromatography and eluted withhexane-EtOAc (9.7 : 0.3 to 9.4 : 0.6) to produce subfractionsBH1-a and BH1-b. Both of these subfractions were subjectedfor neuroprotective assay and the most active subfraction(BH1-b) was subjected to HPLC analysis by using ZORBAXEclipse Plus C18, 4.6 mm i.d. × 150 mm × 3.5 μm HPLCcolumn, and for separation by using ZORBAX EclipsePlus C18, 9.4 mm i.d. × 250 mm × 3.5 μm HPLC column(3 mL/min), eluted as isocratic system with MeOH : H2O(both + 0.1% formic acid) 90 : 10 for 45 min to afford com-pounds 1, 2, 3, 4, and 5. The five compounds were then sub-jected to MTT neuroprotective assay.
2.4. General Methods of Isolation and Structural Elucidationof Compounds. JEOL LA400 FT-NMR and JEOL ECA400FT-NMR Spectrometer System (400 MHz) with CDCl3 assolvent were used to obtain the nuclear magnetic resonance(NMR) spectra. Mass spectra were obtained using AgilentTechnologies 6530 Accurate-Mass Q-TOF Liquid Chromato-graphy/Mass Spectrometry (LC-MS), with ZORBAX EclipseXDB-C18 Rapid Resolution HT 4.6 mm i.d. × 50 mm ×1.8 μm column. Column chromatography separations wereconducted by using Merck silica gel 60 (230–400 mesh) andsilica gel 60 F254 plates for thin layer chromatography (TLC)monitorings. In order to conduct high-performance liquidchromatography (HPLC) separation, Waters autopurifica-tion system was used and the HPLC was set up with BinaryGradient Module (Waters 2545), System Fluidics Organizer(Waters SFO), and Photodiode Array Detector (190–600 nm,Waters 2998) and Sample Manager (Waters 2767). HPLCanalysis and separations were performed using ZORBAXEclipse Plus C18 (4.6 mm i.d. × 150 mm × 3.5 μm) andZORBAX Eclipse Plus C18 (9.4 mm i.d.× 250 mm× 3.5 μm)HPLC columns.
2.5. Cell Culture. Mouse neuroblastoma X rat glioma hybrid-oma cell line (NG108-15) was procured from American TypeCulture Collection (ATCC). A complete medium DMEM(Dulbecco’s Modified Eagle’s Medium) (Sigma Aldrich)composed of 10% (v/v) heat inactivated foetal bovine serum(PAA Laboratories, Austria), 2% penicillin/streptomycin(PAA Laboratories, Austria), 1% amphotericin B (PAA Lab-oratories, Austria), and HAT (hypoxanthine-aminopterin-thymidine) (Sigma) was prepared and used to cultureNG108-15 hybridoma cells. The complete DMEM mediawere filter-sterilized by using a 0.22 μm filter membrane(Minisart, Sartorius Stedim). NG108-15 cells were culturedand conditioned at 5% CO2 moist atmosphere at 37◦C (CO2
incubator chamber, RSBiotech) and subcultivation was con-ducted every 3-4 days. The cells were harvested by accutase(Innovative Cell Technologies, Inc.). The cells were dailyinspected under inverted microscope (Motic) for detectionof any contamination. For experimental purpose, only cellsthat were at their exponential growth phase (60–70% con-fluency) were selected. Cell viability was determined by usingtrypan blue with a haemocytometer. DMSO (dimethyl sulf-oxide) (Sigma Aldrich) concentration in all experiments wasmaintained ≤0.5% v/v.
2.6. MTT Assay for Assessment of Neuroprotective Activity ofMesuagenin C 3. The neuroprotective effects of mesuageninC 3 on the cell viability of H2O2-stressed NG108-15 cellswere analyzed by MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay. NG108-15 cells wereraised to confluency, harvested by accutase, rinsed with PBS,and plated at a total density of 5 × 103 cells/well in a 96-well plate. The cells were left to adhere for 48 h and thenpreincubated for 2 h with mesuagenin C 3 prior to H2O2
(2 μM) exposure for subsequent 10 h. To each well 20 μLMTT solution (5 mg/mL) (Sigma Aldrich) was added andincubated at 37◦C for another 4 h. The cells were analyzed byusing a microplate reader (ASYS UVM340) at 570 nm (witha reference wavelength of 650 nm).
Cell viability was calculated based on the following for-mula:
% of cell viability
=[
absorbance of treated cells (As)absorbance of control cells (Ac)
]× 100%.
(1)
2.7. Nuclear Double Staining with Hoechst 33342/PI. A totalof 0.5 × 106 cells were plated and pretreated with mesua-genin C 3 (12.5–50 μM) for 2 h before exposure to H2O2
(400 μM) for 10 h. After treatment, cells were harvestedand washed with PBS. Hoechst 33342 (Sigma Aldrich), aDNA fluorochrome (10 μg/mL), was added, followed by PI(2.5 μg/mL), and the cells were further incubated for 15 minat 37◦C. The cells were then observed by fluorescent micro-scope (Leica Inverted Fluorescence Microscope, DM16000B)and approximately 100 cells from five random microscopicfields were counted. The use of PI enabled us to differentiatethe cells at different stages of apoptosis. Cells that werestained brightly by Hoechst 33342 were considered as early

4 Evidence-Based Complementary and Alternative Medicine
apoptotic cells. These cells exhibited reduced nuclear size,chromatin condensation, and nuclear fragmentation. On thecontrary, cells that were stained with both Hoechst 33342 andPI were considered to be at the late apoptosis.
2.8. Detection of Phosphatidylserine Externalization byAnnexin V-FITC/PI Staining. During apoptosis, the phos-phatidylserine is translocated from the inner leaflet to theouter leaflet of the plasma membrane. Thus, annexin V-FITC/PI (BD) staining was performed to aid the detectionof the apoptotic cells with translocated phosphatidylserine.Cells were stained with fluorescein-isothiocyanate- (FITC-)labelled annexin V (green fluorescence), simultaneously withdye exclusion of PI (negative for red fluorescence). Similarly,cells were plated into each 60 mm culture dishes and pre-treated with mesuagenin C 3 (12.5–50 μM) for 2 h prior toexposure to H2O2 (400 μM) for 10 h. Cells were then har-vested, gently washed, and resuspended in annexin V bindingbuffer (BD). Next, cells were stained with annexin V-FITCand PI in the dark at room temperature for 15 min. Cellswere evaluated by using flow cytometry (BD FACScalibur).The differentiation of apoptotic and necrotic cells was basedon the staining with PI.
2.9. Effect of Mesuagenin C 3 on Total Intracellular Glu-tathione (GSH) Content. Similarly, NG108-15 cells wereseeded into 60 mm culture dishes and left for 48 h to adhere.The cells were then pretreated for 2 h with mesuageninC 3 (12.5–50 μM) or dimethyl sulfoxide (DMSO, 0.5%)as untreated control and were subsequently subjected tooxidative stress by exposing to 400 μM H2O2 for 10 h. Cellswere rinsed, harvested, and pellet was collected by cen-trifugation. The pellet was washed with ice-cold PBS andresuspended in 500 μL of 5% 5-sulfosalicylic acid (SSA,Sigma Aldrich). The cell suspension was then centrifuged at10 000 rpm for 15 min and supernatant was collected tobe used in the intracellular glutathione assay using 96-wellplate format. The supernatant was added into each wellcontaining GSH standards (Sigma Aldrich), 5,5-dithio-bis(2-nitrobenzoic acid) (DTNB, Sigma Aldrich), and NADPH(Calbiochem) in phosphate buffer. The reaction was imme-diately initiated by the addition of glutathione reductase(Sigma Aldrich). The final concentrations of the reac-tion mixture were 95 mM potassium phosphate buffer(pH 7.0), 0.95 mM EDTA, 0.038 mg/mL (48 μM) NADPH,0.031 mg/mL DTNB, 0.115 units/mL glutathione reductase,and 0.24% 5-SSA. Absorbance was read at 1 min interval for15 min at 405 nm with Oasys UVM340 microplate reader.The GSH concentration in each sample was calculated andcompared with GSH standard curve.
2.10. Dissipation of Mitochondrial Membrane Potential(Δψm) Analysis. In an attempt to quantify the change inmitochondrial membrane potential (Δψm), we used JC-1kit according to the manufacturer’s protocol (Stratagene)to signal the loss of Δψm, using the lipophylic cationic
fluorescent compound JC-1 (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl benzimidazolylcarbocyanine iodide). JC-1 pos-sesses the ability to penetrate the plasma membrane andenters the cytosol of viable cell where it binds to intact mito-chondrial membranes with large Δψm to form J-aggregateswhich emit red fluorescence at 585 nm when excited. Duringmembrane depolarization these J-aggregates dissociate intomonomeric form which emits green color at 530 nm. Cellswere harvested, washed, and stained with JC-1 for 15 minat 37◦C. Cells were then washed with PBS and Δψm wasmeasured by flow cytometry (BD FACScalibur) for the detec-tion of red and green fluorescence signals. JC-1 aggregates(red fluorescence) within the mitochondria of healthy cellswere detected in the FL-2 channel whereas JC-1 monomers(green fluorescence) in the cytoplasm of apoptotic cells weredetected in the FL-1 channel.
2.11. Flow Cytometric Immunofluorescence Staining of Baxand Bcl-2 Proteins. Neuronal apoptosis is regulated by theinterplay between the pro- (Bax, Bik) and antiapoptotic (Bcl-2, Bcl-xL) proteins. The ratio of these pro- and antiapoptoticproteins will decide the fate of the neurons. We next deter-mined the protein expression of proapoptotic Bax and anti-apoptotic Bcl-2 by using flow cytometric immunofluores-cence staining according to the manufacturer protocol (SantaCruz Biotechnology, Inc.). Cells were plated and pretreatedwith mesuagenin C 3 (12.5–50 μM) for 2 h before they wereexposed to H2O2 (400 μM) for 10 h. After treatment, the cellswere harvested, washed with ice-cold PBS, fixed, and per-meabilized using intracellular flow cytometry (FCM) System(Santa Cruz Biotechnology, Inc.). Cells were resuspendedand aliquoted (1.0 × 106 cells/mL) into 100 μL using FCMwash buffer. Next, cells were either incubated with 20 μL offluorescein-isothiocyanate- (FITC-) conjugated Bax mousemonoclonal antibody (Santa Cruz Biotechnology, Inc.) orrabbit IgG isotype control (Abcam) for 1 h. Similarly, forthe expression of Bcl-2, cells were incubated with either PE-conjugated Bcl-2 mouse monoclonal antibody (Santa CruzBiotechnology, Inc.) or rabbit IgG isotype control (Abcam)for 1 h. Cells were then washed and resuspended in 500 μLFCM wash buffer. Cells were analyzed using BD Accuri C6Flow Cytometry and BD CFlow Software.
2.12. Assessment of Caspase-3/7 and -9 Activities. Caspasesare key mediators of cell death and caspase-3 is an exe-cutioner for apoptosis in cortical neurons in response tovarious insults. The dissipation of Δψm is preceded with heleakage of cytochrome c that results in the formation ofapoptosome and cascade activation of caspases. In elucidat-ing the neuroprotective mechanism of mesuagenin C 3, wedetermined caspase-3/7 and -9 activities by using Car-boxyfluorescein FLICA Apoptosis Detection Kit based on themanufacturer’s protocol (Immunochemistry Technologies,LLC). The enzyme activity was measured based on inhibitorprobes, FAM-DEVD-FMK (FLICA) and FAM-LEHD-FMK,for caspase-3/7 and -9, respectively. These probes bindcovalently and irreversibly to the active site of the activecaspase heterodimer emitting the green fluorescent signal

Evidence-Based Complementary and Alternative Medicine 5
M. kunstleri
Extraction
Hexane EtOAc MeOH(136.94 ± 2.03%) (115.47 ± 3.55%) (89.81 ± 2.31%)
BH1 BH2 BH3 BH4 BH5 BH6
(131.29 ± 3.03%) (116.72 ± 1.58%) (84.99 ± 1.61%) (77.3 ± 1.99%) (65.21 ± 1.63%) (71.92 ± 2.35%)
ColumnChromatography(Subfractionation)
97 : 3 94 : 6
BH1-a BH1-b(42.41 ± 2.35%) (101.07 ± 4.81%)
(42.41 ± 2.35%) (51.46 ± 1.42%) (78.99 ± 3.49%) (61.99 ± 1.58%) (56.47 ± 1.7%)
95 : 5 90 : 10 80 : 20
(bark)
1 2 3 4 5
50 : 5050 : 50 0 : 100Hex : EtOAc Hex : EtOAc Hex : EtOAc Hex : EtOAc Hex : EtOAc EtOAc : MeOH
HPLC
Hex : EtOAc Hex : EtOAc
Figure 1: Fractionation and isolation of the bioactive compounds from the bark of M. kunstleri.
detected by the FL-1 which is a direct measure of the numberof the active caspase enzymes. For this assay, the cells werepretreated with mesuagenin C 3 (12.5–50 μM) for 2 h andsubsequently exposed to H2O2 (400 μM) for 10 h. Likewise,cells were harvested, washed with PBS, resuspended inmedia, and stained with 30X FLICA solutions. The cells werethen incubated at 37◦C under 5% CO2 for 1 h and washedtwice with 1X FLICA washing solution. Cells were then resus-pended in a 400 μL washing buffer and fixed by the additionof 40 μL fixative solution for flow cytometry analysis. Cellswere analyzed using BD Accuri C6 Flow Cytometry and BDCFlow Software.
2.13. Statistical Analysis. All the experimental data areexpressed in mean ± standard error (S.E.). Statistical dif-ferences between groups were analyzed and calculated byone-way analysis of variance (ANOVA) from at least threeindependent experiments and this is followed by Dunnett’stest. P < 0.05 was considered to be significantly differentfrom the H2O2-treated groups.
3. Results
3.1. Hexane Fraction of M. kunstleri Exhibited the StrongestNeuroprotective Effect. We studied the concentration-depen-dent effect of H2O2 challenge on cell viability for 10 h byusing MTT cell viability assay. NG108-15 cells were treatedwith different concentrations of H2O2 (0.1–4 mM) for 10 hand a significant dose-dependent reduction in cell viabilitywas detected. After exposure to 2 mM H2O2, NG108-15
cell viability was about 45.41 ± 3.42% of the control via-bility. Pretreatment (2 h) with BH extract showed thehighest percentage cell viability of 136.94 ± 2.03%. The BHextract was then subjected to neuroprotective activity-guidedfractionation and isolation approach due to its highestneuroprotective-activity and six fractions were obtainedafter fractionation (Figure 1). Fraction BH1 possessed thestrongest neuroprotective activity among the fractions withthe percentage cell viability of 131.29± 3.03%. Fractionationof BH1 yielded two subfractions (BH1-a and BH1-b) withsub-fraction BH1-b possessing stronger neuroprotectiveeffect (101.07 ± 4.81%) as compared with fraction BH1-a.Hence, BH1-b was further chromatographed and five purecompounds were isolated and identified successfully (Figures2 and 3).
3.2. Isolation of Mesuagenin C 3 and Its Protective Effectsagainst H2O2-Induced Apoptosis in NG108-15 Cells. Amongthe five compounds obtained from the fractionation of BH1-b, mesuagenin C 3 was isolated from M. kunstleri for thefirst time and its structure was confirmed by comparison ofthe obtained spectral data with the published literature data[8]. Results were summarized in Figures 1–3. The structureof mesuagenin C 3 was further confirmed by HRESI-MS, ina positive mode, which revealed the molecular ion peak atm/z 483.1183 [M + Na]+ which corresponds to a molecularformula of C29H32O5. The comparison of 1H and 13C NMRof mesuagenin C from literature values [8] and compound3 was depicted in Table 1. In view of the mentioned data
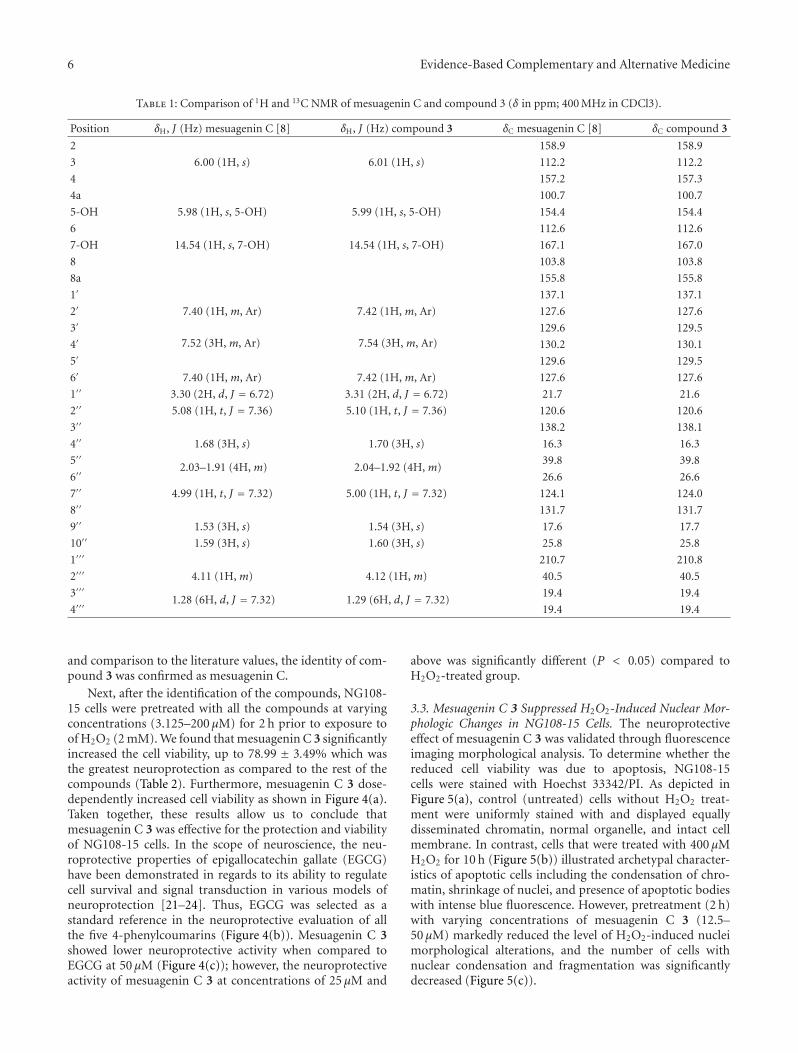
6 Evidence-Based Complementary and Alternative Medicine
Table 1: Comparison of 1H and 13C NMR of mesuagenin C and compound 3 (δ in ppm; 400 MHz in CDCl3).
Position δH, J (Hz) mesuagenin C [8] δH, J (Hz) compound 3 δC mesuagenin C [8] δC compound 3
2 158.9 158.9
3 6.00 (1H, s) 6.01 (1H, s) 112.2 112.2
4 157.2 157.3
4a 100.7 100.7
5-OH 5.98 (1H, s, 5-OH) 5.99 (1H, s, 5-OH) 154.4 154.4
6 112.6 112.6
7-OH 14.54 (1H, s, 7-OH) 14.54 (1H, s, 7-OH) 167.1 167.0
8 103.8 103.8
8a 155.8 155.8
1′ 137.1 137.1
2′ 7.40 (1H, m, Ar) 7.42 (1H, m, Ar) 127.6 127.6
3′
7.52 (3H, m, Ar) 7.54 (3H, m, Ar)129.6 129.5
4′ 130.2 130.1
5′ 129.6 129.5
6′ 7.40 (1H, m, Ar) 7.42 (1H, m, Ar) 127.6 127.6
1′′ 3.30 (2H, d, J = 6.72) 3.31 (2H, d, J = 6.72) 21.7 21.6
2′′ 5.08 (1H, t, J = 7.36) 5.10 (1H, t, J = 7.36) 120.6 120.6
3′′ 138.2 138.1
4′′ 1.68 (3H, s) 1.70 (3H, s) 16.3 16.3
5′′2.03–1.91 (4H, m) 2.04–1.92 (4H, m)
39.8 39.8
6′′ 26.6 26.6
7′′ 4.99 (1H, t, J = 7.32) 5.00 (1H, t, J = 7.32) 124.1 124.0
8′′ 131.7 131.7
9′′ 1.53 (3H, s) 1.54 (3H, s) 17.6 17.7
10′′ 1.59 (3H, s) 1.60 (3H, s) 25.8 25.8
1′′′ 210.7 210.8
2′′′ 4.11 (1H, m) 4.12 (1H, m) 40.5 40.5
3′′′1.28 (6H, d, J = 7.32) 1.29 (6H, d, J = 7.32)
19.4 19.4
4′′′ 19.4 19.4
and comparison to the literature values, the identity of com-pound 3 was confirmed as mesuagenin C.
Next, after the identification of the compounds, NG108-15 cells were pretreated with all the compounds at varyingconcentrations (3.125–200 μM) for 2 h prior to exposure toof H2O2 (2 mM). We found that mesuagenin C 3 significantlyincreased the cell viability, up to 78.99 ± 3.49% which wasthe greatest neuroprotection as compared to the rest of thecompounds (Table 2). Furthermore, mesuagenin C 3 dose-dependently increased cell viability as shown in Figure 4(a).Taken together, these results allow us to conclude thatmesuagenin C 3 was effective for the protection and viabilityof NG108-15 cells. In the scope of neuroscience, the neu-roprotective properties of epigallocatechin gallate (EGCG)have been demonstrated in regards to its ability to regulatecell survival and signal transduction in various models ofneuroprotection [21–24]. Thus, EGCG was selected as astandard reference in the neuroprotective evaluation of allthe five 4-phenylcoumarins (Figure 4(b)). Mesuagenin C 3showed lower neuroprotective activity when compared toEGCG at 50 μM (Figure 4(c)); however, the neuroprotectiveactivity of mesuagenin C 3 at concentrations of 25 μM and
above was significantly different (P < 0.05) compared toH2O2-treated group.
3.3. Mesuagenin C 3 Suppressed H2O2-Induced Nuclear Mor-phologic Changes in NG108-15 Cells. The neuroprotectiveeffect of mesuagenin C 3 was validated through fluorescenceimaging morphological analysis. To determine whether thereduced cell viability was due to apoptosis, NG108-15cells were stained with Hoechst 33342/PI. As depicted inFigure 5(a), control (untreated) cells without H2O2 treat-ment were uniformly stained with and displayed equallydisseminated chromatin, normal organelle, and intact cellmembrane. In contrast, cells that were treated with 400 μMH2O2 for 10 h (Figure 5(b)) illustrated archetypal character-istics of apoptotic cells including the condensation of chro-matin, shrinkage of nuclei, and presence of apoptotic bodieswith intense blue fluorescence. However, pretreatment (2 h)with varying concentrations of mesuagenin C 3 (12.5–50 μM) markedly reduced the level of H2O2-induced nucleimorphological alterations, and the number of cells withnuclear condensation and fragmentation was significantlydecreased (Figure 5(c)).
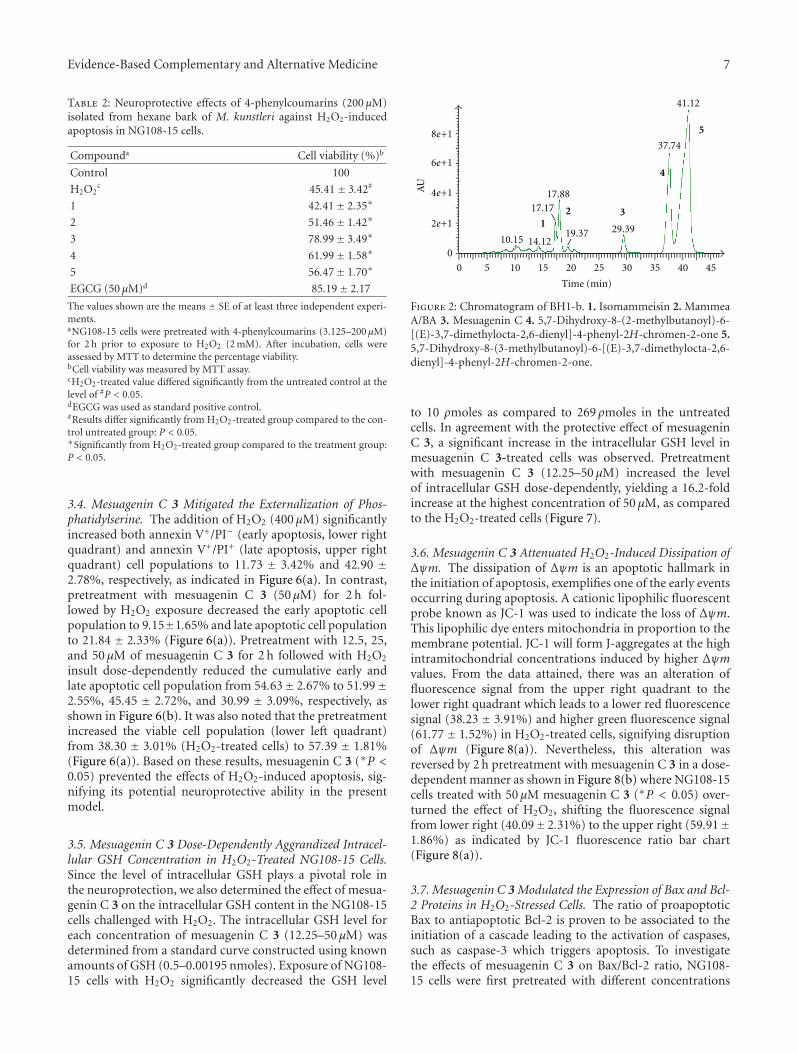
Evidence-Based Complementary and Alternative Medicine 7
Table 2: Neuroprotective effects of 4-phenylcoumarins (200 μM)isolated from hexane bark of M. kunstleri against H2O2-inducedapoptosis in NG108-15 cells.
Compounda Cell viability (%)b
Control 100
H2O2c 45.41± 3.42#
1 42.41± 2.35∗
2 51.46± 1.42∗
3 78.99± 3.49∗
4 61.99± 1.58∗
5 56.47± 1.70∗
EGCG (50 μM)d 85.19± 2.17
The values shown are the means ± SE of at least three independent experi-ments.aNG108-15 cells were pretreated with 4-phenylcoumarins (3.125–200 μM)for 2 h prior to exposure to H2O2 (2 mM). After incubation, cells wereassessed by MTT to determine the percentage viability.bCell viability was measured by MTT assay.cH2O2-treated value differed significantly from the untreated control at thelevel of #P < 0.05.dEGCG was used as standard positive control.#Results differ significantly from H2O2-treated group compared to the con-trol untreated group: P < 0.05.∗Significantly from H2O2-treated group compared to the treatment group:P < 0.05.
3.4. Mesuagenin C 3 Mitigated the Externalization of Phos-phatidylserine. The addition of H2O2 (400 μM) significantlyincreased both annexin V+/PI− (early apoptosis, lower rightquadrant) and annexin V+/PI+ (late apoptosis, upper rightquadrant) cell populations to 11.73 ± 3.42% and 42.90 ±2.78%, respectively, as indicated in Figure 6(a). In contrast,pretreatment with mesuagenin C 3 (50 μM) for 2 h fol-lowed by H2O2 exposure decreased the early apoptotic cellpopulation to 9.15±1.65% and late apoptotic cell populationto 21.84 ± 2.33% (Figure 6(a)). Pretreatment with 12.5, 25,and 50 μM of mesuagenin C 3 for 2 h followed with H2O2
insult dose-dependently reduced the cumulative early andlate apoptotic cell population from 54.63± 2.67% to 51.99±2.55%, 45.45 ± 2.72%, and 30.99 ± 3.09%, respectively, asshown in Figure 6(b). It was also noted that the pretreatmentincreased the viable cell population (lower left quadrant)from 38.30 ± 3.01% (H2O2-treated cells) to 57.39 ± 1.81%(Figure 6(a)). Based on these results, mesuagenin C 3 (∗P <0.05) prevented the effects of H2O2-induced apoptosis, sig-nifying its potential neuroprotective ability in the presentmodel.
3.5. Mesuagenin C 3 Dose-Dependently Aggrandized Intracel-lular GSH Concentration in H2O2-Treated NG108-15 Cells.Since the level of intracellular GSH plays a pivotal role inthe neuroprotection, we also determined the effect of mesua-genin C 3 on the intracellular GSH content in the NG108-15cells challenged with H2O2. The intracellular GSH level foreach concentration of mesuagenin C 3 (12.25–50 μM) wasdetermined from a standard curve constructed using knownamounts of GSH (0.5–0.00195 nmoles). Exposure of NG108-15 cells with H2O2 significantly decreased the GSH level
0 5 10 15 20 25 30 35 40 45
10.15 14.12
17.1717.88
19.37 29.39
37.74
41.12
8e+1
6e+1
4e+1
2e+1
0
12 3
4
5
AU
Time (min)
Figure 2: Chromatogram of BH1-b. 1. Isomammeisin 2. MammeaA/BA 3. Mesuagenin C 4. 5,7-Dihydroxy-8-(2-methylbutanoyl)-6-[(E)-3,7-dimethylocta-2,6-dienyl]-4-phenyl-2H-chromen-2-one 5.5,7-Dihydroxy-8-(3-methylbutanoyl)-6-[(E)-3,7-dimethylocta-2,6-dienyl]-4-phenyl-2H-chromen-2-one.
to 10 ρmoles as compared to 269 ρmoles in the untreatedcells. In agreement with the protective effect of mesuageninC 3, a significant increase in the intracellular GSH level inmesuagenin C 3-treated cells was observed. Pretreatmentwith mesuagenin C 3 (12.25–50 μM) increased the levelof intracellular GSH dose-dependently, yielding a 16.2-foldincrease at the highest concentration of 50 μM, as comparedto the H2O2-treated cells (Figure 7).
3.6. Mesuagenin C 3 Attenuated H2O2-Induced Dissipation ofΔψm. The dissipation of Δψm is an apoptotic hallmark inthe initiation of apoptosis, exemplifies one of the early eventsoccurring during apoptosis. A cationic lipophilic fluorescentprobe known as JC-1 was used to indicate the loss of Δψm.This lipophilic dye enters mitochondria in proportion to themembrane potential. JC-1 will form J-aggregates at the highintramitochondrial concentrations induced by higher Δψmvalues. From the data attained, there was an alteration offluorescence signal from the upper right quadrant to thelower right quadrant which leads to a lower red fluorescencesignal (38.23 ± 3.91%) and higher green fluorescence signal(61.77± 1.52%) in H2O2-treated cells, signifying disruptionof Δψm (Figure 8(a)). Nevertheless, this alteration wasreversed by 2 h pretreatment with mesuagenin C 3 in a dose-dependent manner as shown in Figure 8(b) where NG108-15cells treated with 50 μM mesuagenin C 3 (∗P < 0.05) over-turned the effect of H2O2, shifting the fluorescence signalfrom lower right (40.09± 2.31%) to the upper right (59.91±1.86%) as indicated by JC-1 fluorescence ratio bar chart(Figure 8(a)).
3.7. Mesuagenin C 3 Modulated the Expression of Bax and Bcl-2 Proteins in H2O2-Stressed Cells. The ratio of proapoptoticBax to antiapoptotic Bcl-2 is proven to be associated to theinitiation of a cascade leading to the activation of caspases,such as caspase-3 which triggers apoptosis. To investigatethe effects of mesuagenin C 3 on Bax/Bcl-2 ratio, NG108-15 cells were first pretreated with different concentrations

8 Evidence-Based Complementary and Alternative Medicine
1
2
3
456
7
8
OH
OH
OHOH
OH
HO
HO
HOHO
HO
O
O
O
OO
O
O
O O
O
O
O O
O O
1 2 3
4 5
Figure 3: The chemical structures of the compounds isolated from M. kunstleri via neuroprotective activity-guided approach.
of mesuagenin C 3 (12.5, 25, and 50 μM) for 2 h prior toexposure to H2O2 for 10 h. Our obtained data showed thatH2O2-treated cell profiles shifted to the right in the Baxhistograms resulting in 3.09-fold increase (Figures 9(a) and9(b)) in the expression of Bax protein, while decreasing thelevel of Bcl-2 (Figures 9(c) and 9(d)). This has resultedin a significant surge in the Bax/Bcl-2 ratio to 7.02-fold(Figure 9(e)). However, pretreatment with mesuagenin C 3(12.5, 25 and 50 μM) inhibited the H2O2-induced increaseof Bax (Figures 9(a) and 9(b)) and decrease of Bcl-2 proteinexpression (Figures 9(c) and 9(d)) dramatically when com-pared to H2O2-treated cells. This is further demonstratedby the lowering in the ratio of Bax/Bcl-2 from 5.68 to 2.12(Figure 9(e)). On that account, mesuagenin C 3 may preventthe NG108-15 cells from entering neuronal apoptosis.
3.8. Mesuagenin C 3 Attenuated H2O2-Induced Activationof Caspase-3/7 and -9 in H2O2-Treated NG108-15 Cells. Toinvestigate the involvement of caspase-dependent and cas-pase-independent pathways in H2O2-induced apoptosis, theNG108-15 cells were subjected to H2O2 treatment (400 μM)for 10 h and the caspase-3/7 and -9 activities were measuredusing flow cytometry analysis. The results in Figures 10(a)and 10(c) showed that H2O2-treated cell profiles shifted tothe right in caspase-3/7 and -9 histograms indicating signi-ficant aggrandization of caspase-3/7 (566.76 ± 13.59%) andcaspase-9 (372.97 ± 11.62%) activities when compared to
untreated control cells. We next investigated the effects ofmesuagenin C 3 on elevated caspase-3/7 and -9 activitiesinduced by H2O2. The findings demonstrated that pretreat-ment with mesuagenin C 3 (50 μM) for 2 h partially inhibitedthe elevated activities of caspase-3/7 and -9 induced byH2O2. The caspase-3/7 and -9 activities were found to besignificantly reduced to 221.59± 10.66% (Figure 10(b)) and171.39 ± 10.56% (Figure 10(d)), respectively, (∗P < 0.05),as compared to the H2O2-treated cells. The reduction ofcaspase-9 activity further explains the attenuation of thecaspase-3/7 activation which reversed the H2O2-inducedapoptosis effects. The data suggested that mesuagenin C 3may suppress caspase-9 activation followed by inhibitionof caspase-3/7 via protecting mitochondrial membraneintegrity and suppressing cytochrome c release from themitochondrial intermembrane space. The results impliedthat H2O2-induced apoptosis was mediated by caspase-dependent intrinsic apoptotic pathways.
4. Discussion
In the present study, we found that the hexane fractionof M. kunstleri prevented H2O2-induced neurotoxicityin NG108-15 cells. In order to verify the neuroprotectivecomponents of M. kunstleri, as part of a continuing studyon neuroprotection-effects of M. kunstleri, neuroprotective-activity-guided fractionation and isolation was carried out
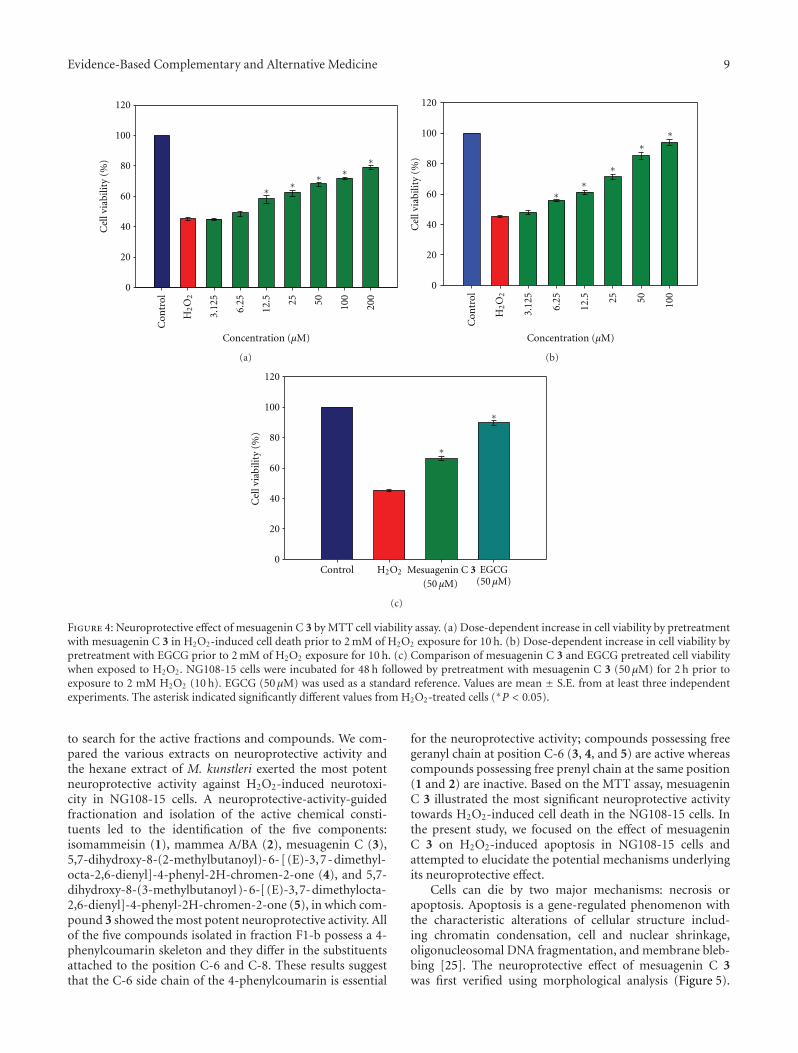
Evidence-Based Complementary and Alternative Medicine 9
Cel
l via
bilit
y (%
)
0
20
40
60
80
100
120
Concentration (μM)
Con
trol
3.12
5
6.25
12.5 25 50 100
200
∗ ∗∗ ∗
∗
H2O
2
(a)
Cel
l via
bilit
y (%
)
20
0
40
60
80
100
120
Con
trol
3.12
5
6.25
12.5 25 50 100
H2O
2
Concentration (μM)
∗∗
∗
∗∗
(b)
Cel
l via
bilit
y (%
)
0
20
40
60
80
100
120
H2O2
(50 μM)EGCG
(50 μM)Control
∗
∗
Mesuagenin C 3
(c)
Figure 4: Neuroprotective effect of mesuagenin C 3 by MTT cell viability assay. (a) Dose-dependent increase in cell viability by pretreatmentwith mesuagenin C 3 in H2O2-induced cell death prior to 2 mM of H2O2 exposure for 10 h. (b) Dose-dependent increase in cell viability bypretreatment with EGCG prior to 2 mM of H2O2 exposure for 10 h. (c) Comparison of mesuagenin C 3 and EGCG pretreated cell viabilitywhen exposed to H2O2. NG108-15 cells were incubated for 48 h followed by pretreatment with mesuagenin C 3 (50 μM) for 2 h prior toexposure to 2 mM H2O2 (10 h). EGCG (50 μM) was used as a standard reference. Values are mean ± S.E. from at least three independentexperiments. The asterisk indicated significantly different values from H2O2-treated cells (∗P < 0.05).
to search for the active fractions and compounds. We com-pared the various extracts on neuroprotective activity andthe hexane extract of M. kunstleri exerted the most potentneuroprotective activity against H2O2-induced neurotoxi-city in NG108-15 cells. A neuroprotective-activity-guidedfractionation and isolation of the active chemical consti-tuents led to the identification of the five components:isomammeisin (1), mammea A/BA (2), mesuagenin C (3),5,7-dihydroxy-8-(2-methylbutanoyl)-6-[(E)-3,7-dimethyl-octa-2,6-dienyl]-4-phenyl-2H-chromen-2-one (4), and 5,7-dihydroxy-8-(3-methylbutanoyl)-6-[(E)-3,7-dimethylocta-2,6-dienyl]-4-phenyl-2H-chromen-2-one (5), in which com-pound 3 showed the most potent neuroprotective activity. Allof the five compounds isolated in fraction F1-b possess a 4-phenylcoumarin skeleton and they differ in the substituentsattached to the position C-6 and C-8. These results suggestthat the C-6 side chain of the 4-phenylcoumarin is essential
for the neuroprotective activity; compounds possessing freegeranyl chain at position C-6 (3, 4, and 5) are active whereascompounds possessing free prenyl chain at the same position(1 and 2) are inactive. Based on the MTT assay, mesuageninC 3 illustrated the most significant neuroprotective activitytowards H2O2-induced cell death in the NG108-15 cells. Inthe present study, we focused on the effect of mesuageninC 3 on H2O2-induced apoptosis in NG108-15 cells andattempted to elucidate the potential mechanisms underlyingits neuroprotective effect.
Cells can die by two major mechanisms: necrosis orapoptosis. Apoptosis is a gene-regulated phenomenon withthe characteristic alterations of cellular structure includ-ing chromatin condensation, cell and nuclear shrinkage,oligonucleosomal DNA fragmentation, and membrane bleb-bing [25]. The neuroprotective effect of mesuagenin C 3was first verified using morphological analysis (Figure 5).
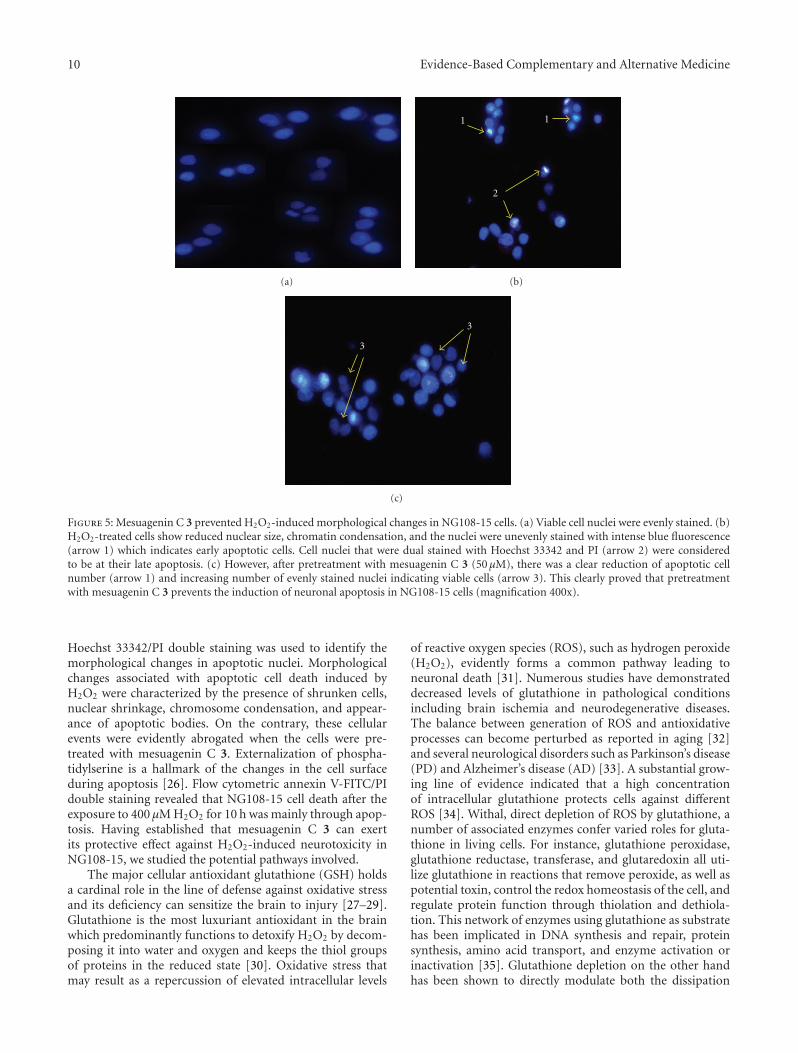
10 Evidence-Based Complementary and Alternative Medicine
(a)
11
2
(b)
3
3
(c)
Figure 5: Mesuagenin C 3 prevented H2O2-induced morphological changes in NG108-15 cells. (a) Viable cell nuclei were evenly stained. (b)H2O2-treated cells show reduced nuclear size, chromatin condensation, and the nuclei were unevenly stained with intense blue fluorescence(arrow 1) which indicates early apoptotic cells. Cell nuclei that were dual stained with Hoechst 33342 and PI (arrow 2) were consideredto be at their late apoptosis. (c) However, after pretreatment with mesuagenin C 3 (50 μM), there was a clear reduction of apoptotic cellnumber (arrow 1) and increasing number of evenly stained nuclei indicating viable cells (arrow 3). This clearly proved that pretreatmentwith mesuagenin C 3 prevents the induction of neuronal apoptosis in NG108-15 cells (magnification 400x).
Hoechst 33342/PI double staining was used to identify themorphological changes in apoptotic nuclei. Morphologicalchanges associated with apoptotic cell death induced byH2O2 were characterized by the presence of shrunken cells,nuclear shrinkage, chromosome condensation, and appear-ance of apoptotic bodies. On the contrary, these cellularevents were evidently abrogated when the cells were pre-treated with mesuagenin C 3. Externalization of phospha-tidylserine is a hallmark of the changes in the cell surfaceduring apoptosis [26]. Flow cytometric annexin V-FITC/PIdouble staining revealed that NG108-15 cell death after theexposure to 400 μM H2O2 for 10 h was mainly through apop-tosis. Having established that mesuagenin C 3 can exertits protective effect against H2O2-induced neurotoxicity inNG108-15, we studied the potential pathways involved.
The major cellular antioxidant glutathione (GSH) holdsa cardinal role in the line of defense against oxidative stressand its deficiency can sensitize the brain to injury [27–29].Glutathione is the most luxuriant antioxidant in the brainwhich predominantly functions to detoxify H2O2 by decom-posing it into water and oxygen and keeps the thiol groupsof proteins in the reduced state [30]. Oxidative stress thatmay result as a repercussion of elevated intracellular levels
of reactive oxygen species (ROS), such as hydrogen peroxide(H2O2), evidently forms a common pathway leading toneuronal death [31]. Numerous studies have demonstrateddecreased levels of glutathione in pathological conditionsincluding brain ischemia and neurodegenerative diseases.The balance between generation of ROS and antioxidativeprocesses can become perturbed as reported in aging [32]and several neurological disorders such as Parkinson’s disease(PD) and Alzheimer’s disease (AD) [33]. A substantial grow-ing line of evidence indicated that a high concentrationof intracellular glutathione protects cells against differentROS [34]. Withal, direct depletion of ROS by glutathione, anumber of associated enzymes confer varied roles for gluta-thione in living cells. For instance, glutathione peroxidase,glutathione reductase, transferase, and glutaredoxin all uti-lize glutathione in reactions that remove peroxide, as well aspotential toxin, control the redox homeostasis of the cell, andregulate protein function through thiolation and dethiola-tion. This network of enzymes using glutathione as substratehas been implicated in DNA synthesis and repair, proteinsynthesis, amino acid transport, and enzyme activation orinactivation [35]. Glutathione depletion on the other handhas been shown to directly modulate both the dissipation
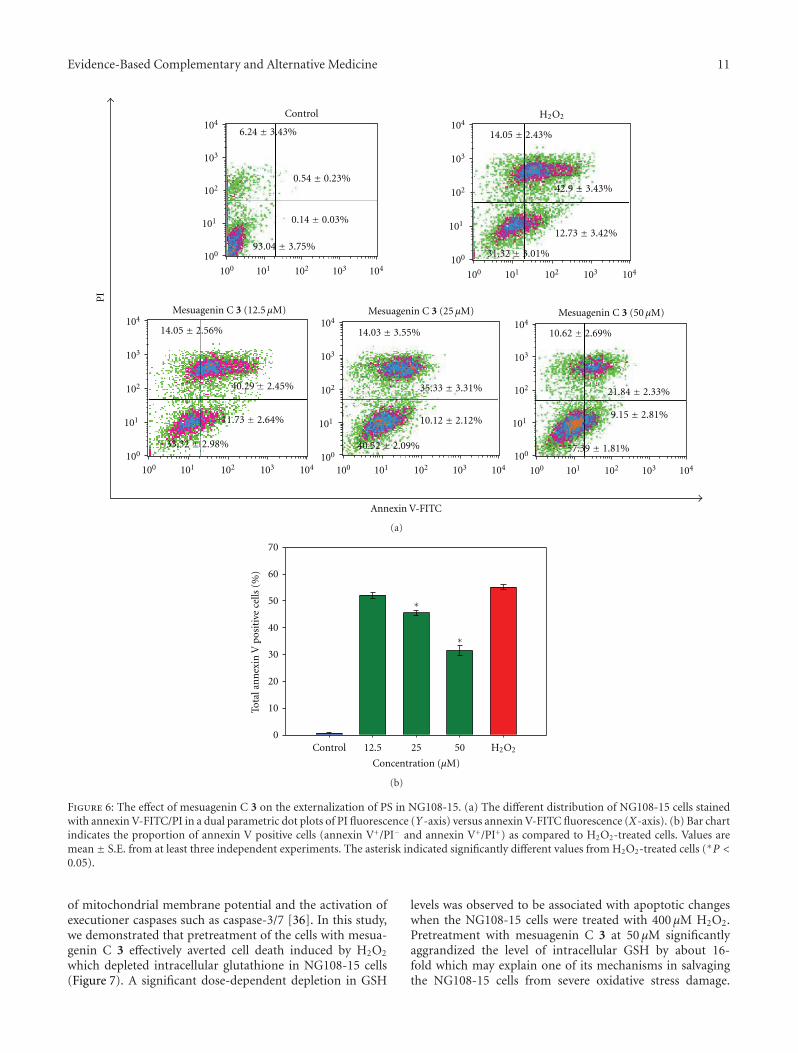
Evidence-Based Complementary and Alternative Medicine 11
100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104
100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104
PI
Control H2O2
Annexin V-FITC
9.15 ± 2.81%
0.54 ± 0.23%
6.24 ± 3.43%
0.14 ± 0.03%
93.04 ± 3.75%
14.05 ± 2.43%
14.05 ± 2.56%
40.29 ± 2.45%
11.73 ± 2.64%
40.52 ± 2.09%
10.12 ± 2.12%
35.33 ± 3.31%
10.62 ± 2.69%
21.84 ± 2.33%
57.39 ± 1.81%
14.03 ± 3.55%
33.32 ± 2.98%
42.9 ± 3.43%
31.32 ± 3.01%
12.73 ± 3.42%
Mesuagenin C 3 (12.5 μM) Mesuagenin C 3 (25 μM) Mesuagenin C 3 (50 μM)
(a)
Tota
l an
nex
in V
pos
itiv
e ce
lls (
%)
25 500
10
20
30
40
50
60
70
Concentration (μM)
H2O2Control 12.5
∗
∗
(b)
Figure 6: The effect of mesuagenin C 3 on the externalization of PS in NG108-15. (a) The different distribution of NG108-15 cells stainedwith annexin V-FITC/PI in a dual parametric dot plots of PI fluorescence (Y-axis) versus annexin V-FITC fluorescence (X-axis). (b) Bar chartindicates the proportion of annexin V positive cells (annexin V+/PI− and annexin V+/PI+) as compared to H2O2-treated cells. Values aremean ± S.E. from at least three independent experiments. The asterisk indicated significantly different values from H2O2-treated cells (∗P <0.05).
of mitochondrial membrane potential and the activation ofexecutioner caspases such as caspase-3/7 [36]. In this study,we demonstrated that pretreatment of the cells with mesua-genin C 3 effectively averted cell death induced by H2O2
which depleted intracellular glutathione in NG108-15 cells(Figure 7). A significant dose-dependent depletion in GSH
levels was observed to be associated with apoptotic changeswhen the NG108-15 cells were treated with 400 μM H2O2.Pretreatment with mesuagenin C 3 at 50 μM significantlyaggrandized the level of intracellular GSH by about 16-fold which may explain one of its mechanisms in salvagingthe NG108-15 cells from severe oxidative stress damage.
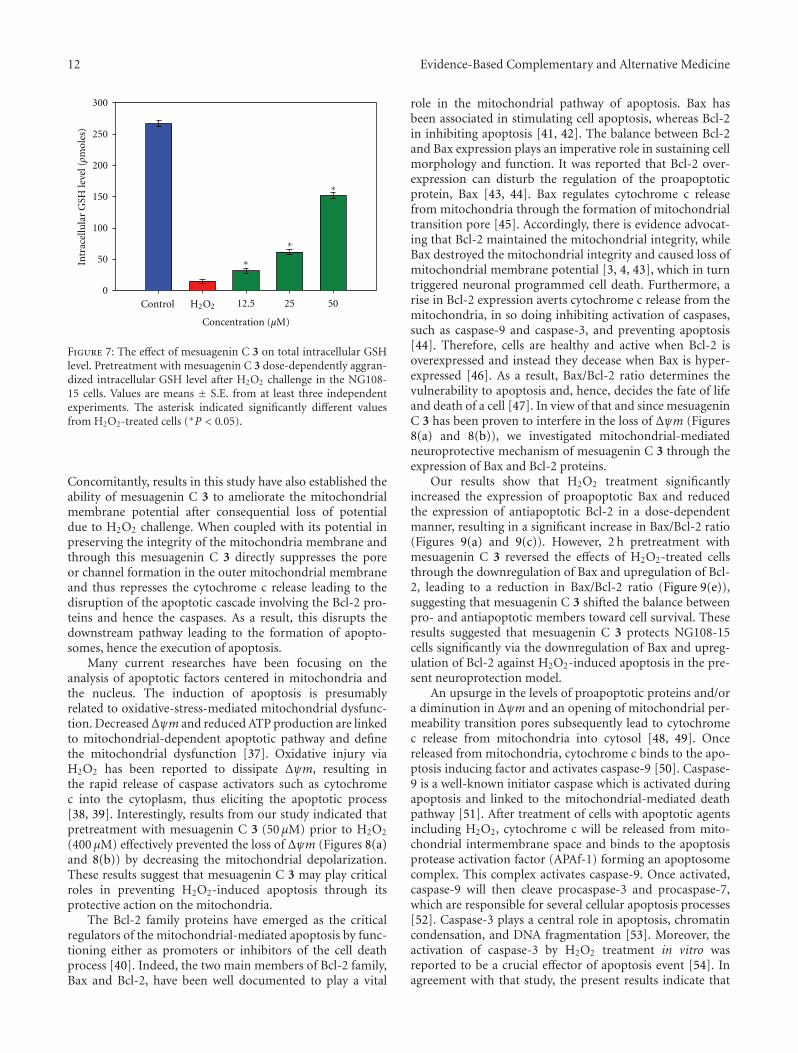
12 Evidence-Based Complementary and Alternative Medicine
Control 12.5 25 500
50
100
150
200
250
300
H2O2
∗
∗
∗
Concentration (μM)
Intr
acel
lula
r G
SH le
vel (ρm
oles
)
Figure 7: The effect of mesuagenin C 3 on total intracellular GSHlevel. Pretreatment with mesuagenin C 3 dose-dependently aggran-dized intracellular GSH level after H2O2 challenge in the NG108-15 cells. Values are means ± S.E. from at least three independentexperiments. The asterisk indicated significantly different valuesfrom H2O2-treated cells (∗P < 0.05).
Concomitantly, results in this study have also established theability of mesuagenin C 3 to ameliorate the mitochondrialmembrane potential after consequential loss of potentialdue to H2O2 challenge. When coupled with its potential inpreserving the integrity of the mitochondria membrane andthrough this mesuagenin C 3 directly suppresses the poreor channel formation in the outer mitochondrial membraneand thus represses the cytochrome c release leading to thedisruption of the apoptotic cascade involving the Bcl-2 pro-teins and hence the caspases. As a result, this disrupts thedownstream pathway leading to the formation of apopto-somes, hence the execution of apoptosis.
Many current researches have been focusing on theanalysis of apoptotic factors centered in mitochondria andthe nucleus. The induction of apoptosis is presumablyrelated to oxidative-stress-mediated mitochondrial dysfunc-tion. DecreasedΔψm and reduced ATP production are linkedto mitochondrial-dependent apoptotic pathway and definethe mitochondrial dysfunction [37]. Oxidative injury viaH2O2 has been reported to dissipate Δψm, resulting inthe rapid release of caspase activators such as cytochromec into the cytoplasm, thus eliciting the apoptotic process[38, 39]. Interestingly, results from our study indicated thatpretreatment with mesuagenin C 3 (50 μM) prior to H2O2
(400 μM) effectively prevented the loss of Δψm (Figures 8(a)and 8(b)) by decreasing the mitochondrial depolarization.These results suggest that mesuagenin C 3 may play criticalroles in preventing H2O2-induced apoptosis through itsprotective action on the mitochondria.
The Bcl-2 family proteins have emerged as the criticalregulators of the mitochondrial-mediated apoptosis by func-tioning either as promoters or inhibitors of the cell deathprocess [40]. Indeed, the two main members of Bcl-2 family,Bax and Bcl-2, have been well documented to play a vital
role in the mitochondrial pathway of apoptosis. Bax hasbeen associated in stimulating cell apoptosis, whereas Bcl-2in inhibiting apoptosis [41, 42]. The balance between Bcl-2and Bax expression plays an imperative role in sustaining cellmorphology and function. It was reported that Bcl-2 over-expression can disturb the regulation of the proapoptoticprotein, Bax [43, 44]. Bax regulates cytochrome c releasefrom mitochondria through the formation of mitochondrialtransition pore [45]. Accordingly, there is evidence advocat-ing that Bcl-2 maintained the mitochondrial integrity, whileBax destroyed the mitochondrial integrity and caused loss ofmitochondrial membrane potential [3, 4, 43], which in turntriggered neuronal programmed cell death. Furthermore, arise in Bcl-2 expression averts cytochrome c release from themitochondria, in so doing inhibiting activation of caspases,such as caspase-9 and caspase-3, and preventing apoptosis[44]. Therefore, cells are healthy and active when Bcl-2 isoverexpressed and instead they decease when Bax is hyper-expressed [46]. As a result, Bax/Bcl-2 ratio determines thevulnerability to apoptosis and, hence, decides the fate of lifeand death of a cell [47]. In view of that and since mesuageninC 3 has been proven to interfere in the loss of Δψm (Figures8(a) and 8(b)), we investigated mitochondrial-mediatedneuroprotective mechanism of mesuagenin C 3 through theexpression of Bax and Bcl-2 proteins.
Our results show that H2O2 treatment significantlyincreased the expression of proapoptotic Bax and reducedthe expression of antiapoptotic Bcl-2 in a dose-dependentmanner, resulting in a significant increase in Bax/Bcl-2 ratio(Figures 9(a) and 9(c)). However, 2 h pretreatment withmesuagenin C 3 reversed the effects of H2O2-treated cellsthrough the downregulation of Bax and upregulation of Bcl-2, leading to a reduction in Bax/Bcl-2 ratio (Figure 9(e)),suggesting that mesuagenin C 3 shifted the balance betweenpro- and antiapoptotic members toward cell survival. Theseresults suggested that mesuagenin C 3 protects NG108-15cells significantly via the downregulation of Bax and upreg-ulation of Bcl-2 against H2O2-induced apoptosis in the pre-sent neuroprotection model.
An upsurge in the levels of proapoptotic proteins and/ora diminution in Δψm and an opening of mitochondrial per-meability transition pores subsequently lead to cytochromec release from mitochondria into cytosol [48, 49]. Oncereleased from mitochondria, cytochrome c binds to the apo-ptosis inducing factor and activates caspase-9 [50]. Caspase-9 is a well-known initiator caspase which is activated duringapoptosis and linked to the mitochondrial-mediated deathpathway [51]. After treatment of cells with apoptotic agentsincluding H2O2, cytochrome c will be released from mito-chondrial intermembrane space and binds to the apoptosisprotease activation factor (APAf-1) forming an apoptosomecomplex. This complex activates caspase-9. Once activated,caspase-9 will then cleave procaspase-3 and procaspase-7,which are responsible for several cellular apoptosis processes[52]. Caspase-3 plays a central role in apoptosis, chromatincondensation, and DNA fragmentation [53]. Moreover, theactivation of caspase-3 by H2O2 treatment in vitro wasreported to be a crucial effector of apoptosis event [54]. Inagreement with that study, the present results indicate that

Evidence-Based Complementary and Alternative Medicine 13
100
101
102
103
104
100 101 102 103 104
Control H2O2
8.85 ± 2.67(%)
91.15 ± 3.03%
37.45 ± 1.68% 42.16 ± 1.10%
57.84 ± 2.43%62.55 ± 2.87%
38.23 ± 3.91%
61.77 ± 1.52%
59.91 ± 1.86%
40.09 ± 2.31%
100
101
102
103
104
100 101 102 103 104
100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104
FL-2
FL-1
JC1 JC1
JC1JC1JC1
Pro
pidi
um
iodi
de
Pro
pidi
um
iodi
de
Pro
pidi
um
iodi
de
Mesuagenin C 3 (12.5 μM) Mesuagenin C 3 (25 μM) Mesuagenin C 3 (50 μM)
(a)
Red
(58
5 n
m)/
gree
n (
530
nm
)fl
uor
esce
nce
(co
ntr
ol (
%))
0
20
40
60
80
100
120
H2O2
Concentration (μM)
Control 12.5 25 50
∗
∗
(b)
Figure 8: Flow cytometry analysis showing alterations in the Δψm of NG108-15 cells. Upon completion of treatment, the cells were stainedwith JC-1 and the alteration in Δψm was analyzed as mentioned in methods. (a) Representative dot plots of JC-1 aggregates (FL-2 Redfluorescence) versus JC-1 monomers (FL-1 green fluorescence). (b) Bar chart showing the percentages of relative fluorescence intensity ofΔψm in control, H2O2-only and varying pretreatment concentrations of mesuagenin C 3 in NG108-15 cells. Values are mean ± S.E. from atleast three independent experiments. The asterisk indicated significantly different values from H2O2-treated cells (∗P < 0.05).
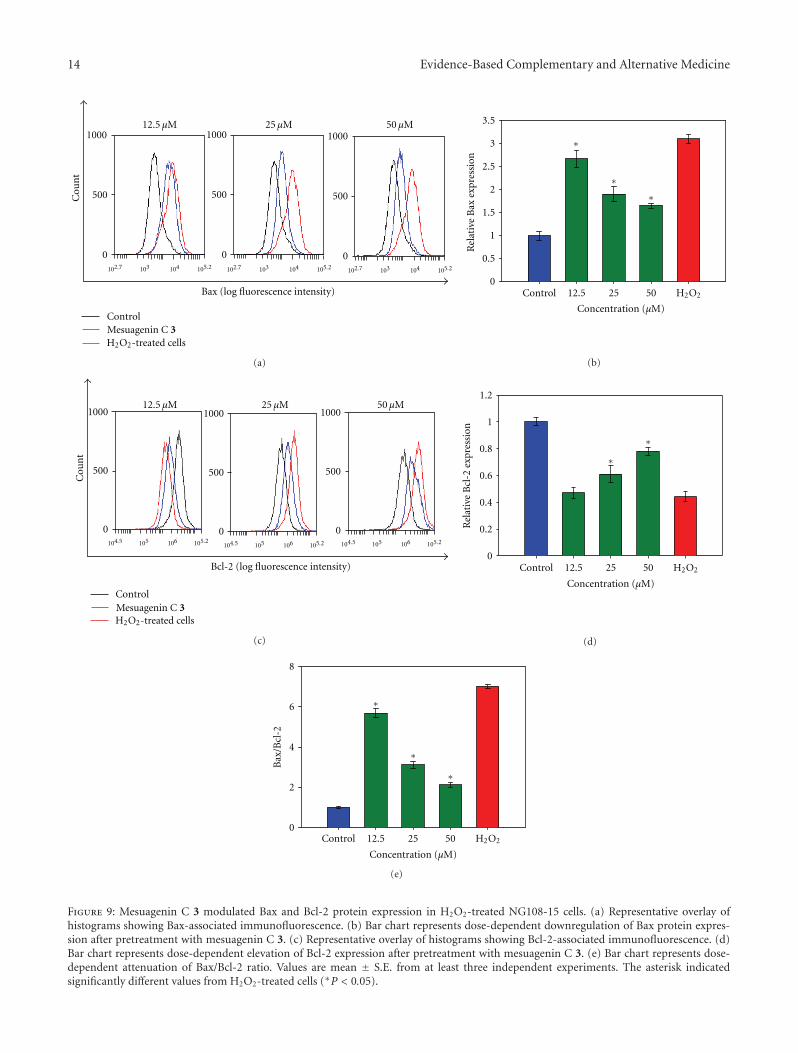
14 Evidence-Based Complementary and Alternative Medicine
1000
500
0
1000
500
0
1000
500
0102.7 105.2103 104 102.7 105.2103 104 102.7 105.2103 104
Control
H2O2-treated cells
Cou
nt
Bax (log fluorescence intensity)
Mesuagenin C 3
12.5 μM 25 μM 50 μM
(a)
Rel
ativ
e B
ax e
xpre
ssio
n
Control 12.5 25 50 H2O2
Concentration (μM)
0
0.5
1
1.5
2
2.5
3
3.5
∗
∗∗
(b)
1000
500
0
1000
500
0
1000
500
0104.5 105.2105 106
104.5 105.2105 106 104.5 105.2105 106
Control
H2O2-treated cellsMesuagenin C 3
Cou
nt
Bcl-2 (log fluorescence intensity)
12.5 μM 25 μM 50 μM
(c)
Rel
ativ
e B
cl-2
exp
ress
ion
0
0.2
0.4
0.6
0.8
1
1.2
Control 12.5 25 50 H2O2
Concentration (μM)
∗
∗
(d)
0
2
4
6
8
Bax
/Bcl
-2
Control 12.5 25 50 H2O2
Concentration (μM)
∗
∗
∗
(e)
Figure 9: Mesuagenin C 3 modulated Bax and Bcl-2 protein expression in H2O2-treated NG108-15 cells. (a) Representative overlay ofhistograms showing Bax-associated immunofluorescence. (b) Bar chart represents dose-dependent downregulation of Bax protein expres-sion after pretreatment with mesuagenin C 3. (c) Representative overlay of histograms showing Bcl-2-associated immunofluorescence. (d)Bar chart represents dose-dependent elevation of Bcl-2 expression after pretreatment with mesuagenin C 3. (e) Bar chart represents dose-dependent attenuation of Bax/Bcl-2 ratio. Values are mean ± S.E. from at least three independent experiments. The asterisk indicatedsignificantly different values from H2O2-treated cells (∗P < 0.05).
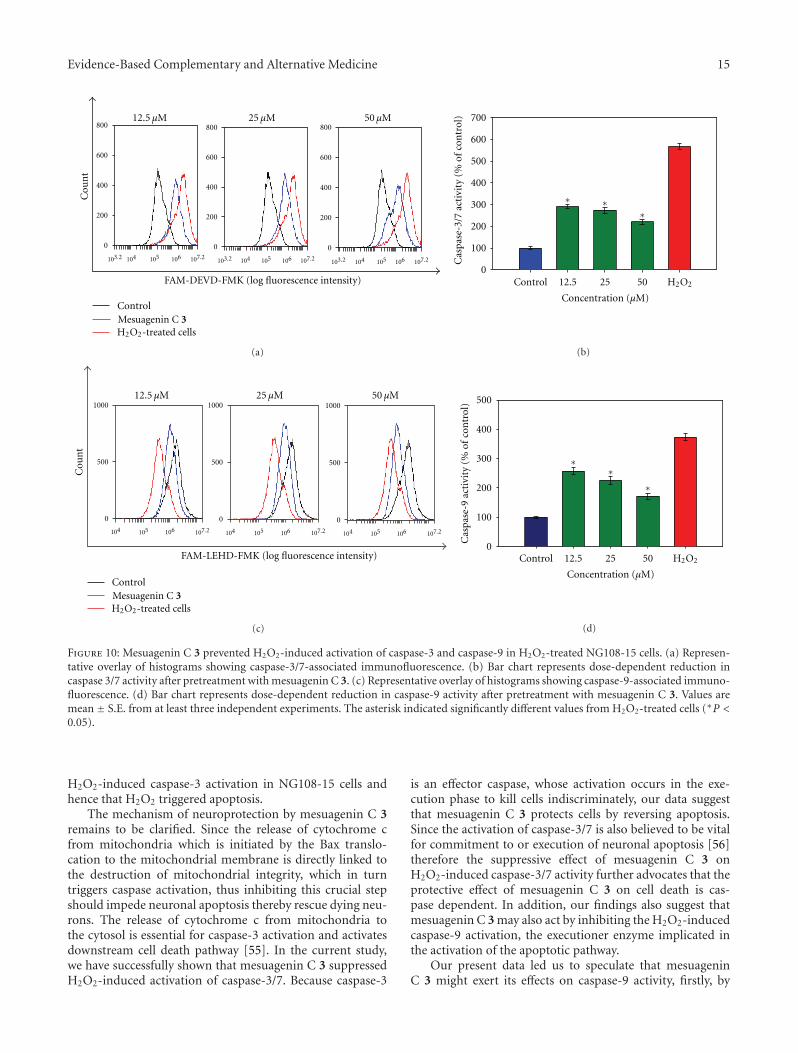
Evidence-Based Complementary and Alternative Medicine 15
0
200
400
600
800
0
200
400
600
800
0
200
400
600
800
103.2 107.2104 105 106103.2 107.2104 105 106 103.2 107.2104 105 106
Control
H2O2-treated cellsMesuagenin C 3
Cou
nt
FAM-DEVD-FMK (log fluorescence intensity)
12.5 μM 25 μM 50 μM
(a)
0
100
200
300
400
500
600
700
H2O2Control 12.5 25 50
∗ ∗∗
Concentration (μM)
Cas
pase
-3/7
act
ivit
y (%
of
con
trol
)
(b)
107.2104 105 106 107.2104 105 106 107.2104 105 106
Control
H2O2-treated cellsMesuagenin C 3
Cou
nt
0
500
1000
0
500
1000
0
500
1000
FAM-LEHD-FMK (log fluorescence intensity)
12.5 μM 25 μM 50 μM
(c)
0
100
200
300
400
500
H2O2Control 12.5 25 50
∗∗
∗
Concentration (μM)
Cas
pase
-9 a
ctiv
ity
(% o
f co
ntr
ol)
(d)
Figure 10: Mesuagenin C 3 prevented H2O2-induced activation of caspase-3 and caspase-9 in H2O2-treated NG108-15 cells. (a) Represen-tative overlay of histograms showing caspase-3/7-associated immunofluorescence. (b) Bar chart represents dose-dependent reduction incaspase 3/7 activity after pretreatment with mesuagenin C 3. (c) Representative overlay of histograms showing caspase-9-associated immuno-fluorescence. (d) Bar chart represents dose-dependent reduction in caspase-9 activity after pretreatment with mesuagenin C 3. Values aremean ± S.E. from at least three independent experiments. The asterisk indicated significantly different values from H2O2-treated cells (∗P <0.05).
H2O2-induced caspase-3 activation in NG108-15 cells andhence that H2O2 triggered apoptosis.
The mechanism of neuroprotection by mesuagenin C 3remains to be clarified. Since the release of cytochrome cfrom mitochondria which is initiated by the Bax translo-cation to the mitochondrial membrane is directly linked tothe destruction of mitochondrial integrity, which in turntriggers caspase activation, thus inhibiting this crucial stepshould impede neuronal apoptosis thereby rescue dying neu-rons. The release of cytochrome c from mitochondria tothe cytosol is essential for caspase-3 activation and activatesdownstream cell death pathway [55]. In the current study,we have successfully shown that mesuagenin C 3 suppressedH2O2-induced activation of caspase-3/7. Because caspase-3
is an effector caspase, whose activation occurs in the exe-cution phase to kill cells indiscriminately, our data suggestthat mesuagenin C 3 protects cells by reversing apoptosis.Since the activation of caspase-3/7 is also believed to be vitalfor commitment to or execution of neuronal apoptosis [56]therefore the suppressive effect of mesuagenin C 3 onH2O2-induced caspase-3/7 activity further advocates that theprotective effect of mesuagenin C 3 on cell death is cas-pase dependent. In addition, our findings also suggest thatmesuagenin C 3 may also act by inhibiting the H2O2-inducedcaspase-9 activation, the executioner enzyme implicated inthe activation of the apoptotic pathway.
Our present data led us to speculate that mesuageninC 3 might exert its effects on caspase-9 activity, firstly, by

16 Evidence-Based Complementary and Alternative Medicine
inhibiting proapoptotic proteins like p53, Bax, and Bad orinducing expression of antiapoptotic family members suchas Bcl-2 and Bcl-xL [57]. Alternatively, it is possible that itmimics the antiapoptotic family members, thus reducingheterodimerization of Bcl-2 family members which subse-quently inhibits release of cytochrome c which results inreduction of apaf-1 and procaspase-9 formation to activatethe caspase-9 activity [58–60]. Based on our findings, thealteration in the ratio of Bax/Bcl-2 protein, together with theattenuation of H2O2-induced caspase-3 and -9 activation,might be responsible for the concomitant disruption ofapoptosis by mesuagenin C 3. These observations providedfurther evidence that mesuagenin C 3 protects NG108-15cells from H2O2-induced apoptosis by blocking the caspase-dependent intrinsic apoptotic pathway.
5. Conclusion
The present study has successfully isolated and identifiedmesuagenin C 3 from the hexane fraction of M. kunstleri asthe most potent neuroprotective compound in the presentneuroprotection model. Mesuagenin C 3 significantly mit-igated the effects of H2O2-induced apoptosis on external-ization of phosphatidylserine, aggrandization of GSH level,Δψm dissipation, regulation of Bcl-2 and Bax proteins inconjunction with the attenuation of activated caspases, sug-gesting that suppression of mitochondrial-mediated apop-totic pathway could be possible mechanism underlying theneuroprotective effects exerted by mesuagenin C 3 in NG108-15 cells.
Acknowledgments
The authors are grateful for the financial support providedby Fundamental Research Grant Scheme (FP009-2010A),University of Malaya PPP Grant (PS369/2010B and PV073/2011A), University Malaya Research Grant (RP001-2012),Science Fund (02-01-01-SF0329), High Impact ResearchGrant (F000002-21001) and CNRS grant. They would like toexpress their thanks to L. E. Teo, D. M. Nor, and R. Syamsirfor the collection and identification of the plant material. Theauthors would like to show appreciation to Dr. Lee HongBoon from CARIF, Sime Darby Medical Centre, Malaysia forallowing them to use the flow cytometer.
References
[1] T. Fukushima, A. Nakamura, N. Iwakami et al., “T-817MA,a neuroprotective agent, attenuates the motor and cognitiveimpairments associated with neuronal degeneration in P301Ltau transgenic mice,” Biochemical and Biophysical ResearchCommunications, vol. 407, no. 4, pp. 730–734, 2011.
[2] J. Yuan and B. A. Yankner, “Apoptosis in the nervous system,”Nature, vol. 407, no. 6805, pp. 802–809, 2000.
[3] M. P. Mattson and T. Magnus, “Ageing and neuronal vulnera-bility,” Nature Reviews Neuroscience, vol. 7, no. 4, pp. 278–294,2006.
[4] D. E. Bredesen, R. V. Rao, and P. Mehlen, “Cell death in thenervous system,” Nature, vol. 443, no. 7113, pp. 796–802, 2006.
[5] J. F. R. Kerr, C. M. Winterfor, and B. V. Harmon, “Mor-phological criteria for identifying apoptosis,” in Cell Biology:A Laboratory Handbook, pp. 319–329, Academic Press, SanDiego, Calif, USA, 1994.
[6] C. J. Norbury and I. D. Hickson, “Cellular responses to DNAdamage,” Annual Review of Pharmacology and Toxicology, vol.41, pp. 367–401, 2001.
[7] K. Awang, S. L. Chong, K. Mohamad et al., “ErythrocarpinesA-E, new cytotoxic limonoids from Chisocheton erythrocar-pus,” Bioorganic and Medicinal Chemistry, vol. 15, no. 17, pp.5997–6002, 2007.
[8] K. Awang, G. Chan, M. Litaudon, N. H. Ismail, M. T. Martin,and F. Gueritte, “4-Phenylcoumarins from Mesua elegans withacetylcholinesterase inhibitory activity,” Bioorganic & Medici-nal Chemistry, vol. 18, no. 22, pp. 7873–7877, 2010.
[9] A. Leverrier, K. Awang, F. Gueritte, and M. Litaudon, “Pen-tacyclic polyketides from Endiandra kingiana as inhibitors ofthe Bcl-xL/Bak interaction,” Phytochemistry, vol. 72, no. 11-12,pp. 1443–1452, 2011.
[10] T. C. Whitmore, Tree Flora of the Malay Peninsula, L. Reeveand Co., London, UK, 1973.
[11] L. Verotta, E. Lovaglio, G. Vidari et al., “4-Alkyl- and 4-phenyl-coumarins from Mesua ferrea as promising multidrug resistantantibacterials,” Phytochemistry, vol. 65, no. 21, pp. 2867–2879,2004.
[12] L. Wu, X. Wang, W. Xu, F. Farzaneh, and R. Xu, “The struc-ture and pharmacological functions of coumarins and theirderivatives,” Current Medicinal Chemistry, vol. 16, no. 32, pp.4236–4260, 2009.
[13] Y. F. Li, Z. H. Gong, J. B. Cao, H. L. Wang, Z. P. Luo, andJ. Li, “Antidepressant-like effect of agmatine and its possiblemechanism,” European Journal of Pharmacology, vol. 469, no.1–3, pp. 81–88, 2003.
[14] G. Chan, K. Awang, A. H. A. Hadi, and S. W. Ng, “6-[(E)-3,7-Dimethyl-octa-2,6-dien-yl]-5,7-dihydr-oxy-8-(2-methyl-butano-yl)-4-phenyl-2H-chromen-2-one from Mesua kuns-tleri King (Kosterm),” Acta Crystallographica Section E, vol. 64,no. 7, p. 1332, 2008.
[15] B. M. McGleenon, K. B. Dynan, and A. P. Passmore, “Acetyl-cholinesterase inhibitors in Alzheimer’s disease,” British Jour-nal of Clinical Pharmacology, vol. 48, no. 4, pp. 471–480, 1999.
[16] M. J. Matos, C. Teran, Y. Perez-Castillo, E. Uriarte, L. Santana,and D. Vina, “Synthesis and study of a series of 3-arylcouma-rins as potent and selective monoamine oxidase B inhibitors,”Journal of Medicinal Chemistry, vol. 54, no. 20, pp. 7127–7137,2011.
[17] S. N. Wu, Y. K. Lo, C. C. Chen, H. F. Li, and H. T. Chiang,“Inhibitory effect of the plant-extract osthole on L-type cal-cium current in NG108-15 neuronal cells,” Biochemical Phar-macology, vol. 63, no. 2, pp. 199–206, 2002.
[18] F. Epifano, G. Molinaro, S. Genovese, R. T. Ngomba, F.Nicoletti, and M. Curini, “Neuroprotective effect of prenyloxy-coumarins from edible vegetables,” Neuroscience Letters, vol.443, no. 2, pp. 57–60, 2008.
[19] Y. K. So and C. K. Young, “Neuroprotective coumarins fromthe root of Angelica gigas: structure-activity relationships,”Archives of Pharmacal Research, vol. 30, no. 11, pp. 1368–1373,2007.
[20] L. Crombie, D. E. Games, and A. McCormick, “Isolation andstructure of mammea A/BA, A/AB and A/BB: a group of 4-aryl-coumarin extractives of Mammea americana L.,” Tetra-hedron Letters, vol. 7, no. 2, pp. 145–149, 1966.

Evidence-Based Complementary and Alternative Medicine 17
[21] W. Q. Chen, X. L. Zhao, Y. Hou et al., “Protective effects ofgreen tea polyphenols on cognitive impairments induced bypsychological stress in rats,” Behavioural Brain Research, vol.202, no. 1, pp. 71–76, 2009.
[22] S. Bastianetto, Z. X. Yao, V. Papadopoulos, and R. Quirion,“Neuroprotective effects of green and black teas and theircatechin gallate esters against β-amyloid-induced toxicity,”European Journal of Neuroscience, vol. 23, no. 1, pp. 55–64,2006.
[23] K. S. Kang, Y. Wen, N. Yamabe, M. Fukui, S. C. Bishop, andB. T. Zhu, “Dual beneficial effects of (-)-epigallocatechin-3-gallate on levodopa methylation and hippocampal neurode-generation: in vitro and in vivo studies,” PLoS ONE, vol. 5, no.8, Article ID e11951, 2010.
[24] J. W. Lee, Y. K. Lee, J. O. Ban et al., “Green tea (-)-epig-allocatechin-3-gallate inhibits β-amyloid-induced cognitivedysfunction through modification of secretase activity viainhibition of ERK and NF-κB pathways in mice,” Journal ofNutrition, vol. 139, no. 10, pp. 1987–1993, 2009.
[25] L. E. Gerschenson and R. J. Rotello, “Apoptosis: a different typeof cell death,” The FASEB Journal, vol. 6, no. 7, pp. 2450–2455,1992.
[26] M. C. Willingham, “Cytochemical methods for the detectionof apoptosis,” Journal of Histochemistry & Cytochemistry, vol.47, no. 9, pp. 1101–1109, 1999.
[27] A. Meister and M. E. Anderson, “Glutathione,” Annual Reviewof Biochemistry, vol. 52, pp. 711–760, 1983.
[28] T. Mizui, H. Kinouchi, and P. H. Chan, “Depletion of brainglutathione by buthionine sulfoximine enhances cerebralischemic injury in rats,” The Journal of Surgical Research, vol.143, no. 2, pp. 247–252, 2007.
[29] P. J. Bobyn, J. L. Franklin, C. M. Wall, J. A. Thornhill, B. H.J. Juurlink, and P. G. Paterson, “The effects of dietary sul-fur amino acid deficiency on rat brain glutathione concen-tration and neural damage in global hemispheric hypoxia—ischemia,” Nutritional Neuroscience, vol. 5, no. 6, pp. 407–416,2002.
[30] C. Mytilineou, B. C. Kramer, and J. A. Yabut, “Glutathionedepletion and oxidative stress,” Parkinsonism and Related Dis-orders, vol. 8, no. 6, pp. 385–387, 2002.
[31] B. N. Ames, M. K. Shigenaga, and T. M. Hagen, “Oxidants,antioxidants, and the degenerative diseases of aging,” Proceed-ings of the National Academy of Sciences of the United States ofAmerica, vol. 90, no. 17, pp. 7915–7922, 1993.
[32] K. B. Beckman and B. N. Ames, “The free radical theory ofaging matures,” Physiological Reviews, vol. 78, no. 2, pp. 547–581, 1998.
[33] R. Dringen, “Metabolism and functions of glutathione inbrain,” Progress in Neurobiology, vol. 62, no. 6, pp. 649–671,2000.
[34] J. M. C. Gutteridge and B. Halliwell, “Free radicals and anti-oxidants in the year 2000. A historical look to the future,”Annals of the New York Academy of Sciences, vol. 899, pp. 136–147, 2000.
[35] B. M. Lomaestro and M. Malone, “Glutathione in health anddisease: pharmacotherapeutic issues,” Annals of Pharmacother-apy, vol. 29, no. 12, pp. 1263–1273, 1995.
[36] R. Franco, M. I. Panayiotidis, and J. A. Cidlowski, “Glu-tathione depletion is necessary for apoptosis in lymphoid cellsindependent of reactive oxygen species formation,” The Jour-nal of Biological Chemistry, vol. 282, no. 42, pp. 30452–30465,2007.
[37] S. L. Hwang and G. C. Yen, “Neuroprotective effects of thecitrus flavanones against H2O2-induced cytotoxicity in PC12cells,” Journal of Agricultural and Food Chemistry, vol. 56, no.3, pp. 859–864, 2008.
[38] J. Y. Jung, C. R. Han, Y. J. Jeong et al., “Epigallocatechin gal-late inhibits nitric oxide-induced apoptosis in rat PC12 cells,”Neuroscience Letters, vol. 411, no. 3, pp. 222–227, 2007.
[39] H. B. Xu, L. Li, and G. Q. Liu, “Protection against hydrogenperoxide-induced cytotoxicity in PC12 cells by guggulsterone,”Yaoxue Xuebao, vol. 43, no. 12, pp. 1190–1197, 2008.
[40] T. Kuwana and D. D. Newmeyer, “Bcl-2-family proteins andthe role of mitochondria in apoptosis,” Current Opinion in CellBiology, vol. 15, no. 6, pp. 691–699, 2003.
[41] H. Zha and J. C. Reed, “Heterodimerization-independentfunctions of cell death regulatory proteins Bax and Bcl-2 inyeast and mammalian cells,” The Journal of Biological Che-mistry, vol. 272, no. 50, pp. 31482–31488, 1997.
[42] M. E. Haque, M. Asanuma, Y. Higashi, I. Miyazaki, K. I.Tanaka, and N. Ogawa, “Apoptosis-inducing neurotoxicity ofdopamine and its metabolites via reactive quinone generationin neuroblastoma cells,” Biochimica et Biophysica Acta, vol.1619, no. 1, pp. 39–52, 2003.
[43] I. Metrailler-Ruchonnet, A. Pagano, S. Carnesecchi, C. Ody,Y. Donati, and C. Barazzone Argiroffo, “Bcl-2 protectsagainst hyperoxia-induced apoptosis through inhibition of themitochondria-dependent pathway,” Free Radical Biology andMedicine, vol. 42, no. 7, pp. 1062–1074, 2007.
[44] S. Zhuang, Y. Yan, R. A. Daubert, J. Han, and R. G. Schnell-mann, “ERK promotes hydrogen peroxide-induced apoptosisthrough caspase-3 activation and inhibition of Akt in renalepithelial cells,” American Journal of Physiology, vol. 292, no.1, pp. F440–F447, 2007.
[45] D. T. Chao and S. J. Korsmeyer, “BCL-2 family: regulators ofcell death,” Annual Review of Immunology, vol. 16, pp. 395–419, 1998.
[46] R. Aikawa, I. Komuro, T. Yamazaki et al., “Oxidative stress acti-vates extracellular signal-regulated kinases through Src andRas in cultured cardiac myocytes of neonatal rats,” Journal ofClinical Investigation, vol. 100, no. 7, pp. 1813–1821, 1997.
[47] J. M. Adams and S. Cory, “Life-or-death decisions by the Bcl-2protein family,” Trends in Biochemical Sciences, vol. 26, no. 1,pp. 61–66, 2001.
[48] M. Crompton, “Bax, Bid and the permeabilization of the mito-chondrial outer membrane in apoptosis,” Current Opinion inCell Biology, vol. 12, no. 4, pp. 414–419, 2000.
[49] G. van Loo, X. Saelens, M. van Gurp, M. MacFarlane, S.J. Martin, and P. Vandenabeele, “The role of mitochondrialfactors in apoptosis: a Russian roulette with more than onebullet,” Cell Death and Differentiation, vol. 9, no. 10, pp. 1031–1042, 2002.
[50] S. Desagher and J. C. Martinou, “Mitochondria as the centralcontrol point of apoptosis,” Trends in Cell Biology, vol. 10, no.9, pp. 369–377, 2000.
[51] P. Caroppi, F. Sinibaldi, L. Fiorucci, and R. Santucci, “Apopto-sis and human diseases: mitochondrion damage and lethal roleof released cytochrome c as proapoptotic protein,” CurrentMedicinal Chemistry, vol. 16, no. 31, pp. 4058–4065, 2009.
[52] C. Garrido, L. Galluzzi, M. Brunet, P. E. Puig, C. Didelot, andG. Kroemer, “Mechanisms of cytochrome c release from mito-chondria,” Cell Death and Differentiation, vol. 13, no. 9, pp.1423–1433, 2006.
[53] S. J. Riedl and Y. Shi, “Molecular mechanisms of caspaseregulation during apoptosis,” Nature Reviews Molecular CellBiology, vol. 5, no. 11, pp. 897–907, 2004.

18 Evidence-Based Complementary and Alternative Medicine
[54] T. Matsura, M. Kai, Y. Fujii, H. Ito, and K. Yamada, “Hydrogenperoxide-induced apoptosis in HL-60 cells requires caspase-3 activation,” Free Radical Research, vol. 30, no. 1, pp. 73–83,1999.
[55] R. U. Janicke, M. L. Sprengart, M. R. Wati, and A. G. Porter,“Caspase-3 is required for DNA fragmentation and morpho-logical changes associated with apoptosis,” The Journal of Bio-logical Chemistry, vol. 273, no. 16, pp. 9357–9360, 1998.
[56] G. M. Cohen, “Caspases: the executioners of apoptosis,”Biochemical Journal, vol. 326, no. 1, pp. 1–16, 1997.
[57] X. Wang, “The expanding role of mitochondria in apoptosis,”Genes & Development, vol. 15, no. 22, pp. 2922–2933, 2001.
[58] B. Pettmann and C. E. Henderson, “Neuronal cell death,” Neu-ron, vol. 20, no. 4, pp. 633–647, 1998.
[59] M. O. Hengartner, “The biochemistry of apoptosis,” Nature,vol. 407, no. 6805, pp. 770–776, 2000.
[60] M. S. Ola, M. Nawaz, and H. Ahsan, “Role of Bcl-2 family pro-teins and caspases in the regulation of apoptosis,” Molecularand Cellular Biochemistry, vol. 351, no. 1-2, pp. 41–58, 2011.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 478190, 9 pagesdoi:10.1155/2012/478190
Research Article
Herbal Extracts Combination (WNK) Prevents Declinein Spatial Learning and Memory in APP/PS1 Mice throughImprovement of Hippocampal Aβ Plaque Formation,Histopathology, and Ultrastructure
Wei-hong Cong, Bin Yang, Li Xu, Xiao-xia Dong, Li-song Sheng,Jin-cai Hou, and Jian-xun Liu
Research Center, Xiyuan Hospital, China Academy of Chinese Medical Sciences, Beijing 100091, China
Correspondence should be addressed to Wei-hong Cong, [email protected]
Received 6 March 2012; Revised 10 May 2012; Accepted 10 May 2012
Academic Editor: Ilkay Erdogan Orhan
Copyright © 2012 Wei-hong Cong et al. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
To investigate the cognitive enhancement effect of WNK, an extracts combination of P. ginseng, G. biloba, and C. sativus L. andpossible mechanisms, 5-month-old APP/PS1 transgenic mice were used in this study. After 3 months of administration, all micereceived Morris water maze (MWM) training and a probe test. Mouse brain sections were detected by immunohistochemistry, HEstaining, and transmission electron microscopy. MWM results showed significant difference between transgenic mice and non-transgenic littermates (P < 0.05, P < 0.01). WNK-treated mice exhibited enhanced maze performance over the training progres-sion, especially better spatial memory retention in probe test compared to transgenic mice (P < 0.05, P < 0.01) and better spatiallearning and memory at the fourth day of MWM test compared to EGB761- (G. biloba extract-) treated ones (P < 0.05). Hippo-campal Aβ plaque burden significantly differed between APP/PS1 and littermate mice (P < 0.001), while decreased Aβ plaqueappeared in WNK- or EGB761-treated transgenic brains (P < 0.05). Neurodegenerative changes were evident from lightmicroscopic and ultrastructural observations in transgenic brains, which were improved by WNK or EGB761 treatment. Thesedata indicate WNK can reduce the decline in spatial cognition, which might be due to its effects on reducing Aβ plaque formationand ameliorating histopathology and ultrastructure in hippocampus of APP/PS1 mouse brain.
1. Introduction
Neuropathological examination of the Alzheimer’s disease(AD) brain shows extensive neuronal loss, accumulation offibrillar proteins as extracellular β-amyloid (Aβ) plaques,and as neurofibrillary tangles (NFTs) inside neurons [1].Autosomal dominant AD has been proved to be associatedwith mutations of APP, PS1, and PS2 genes [2]. These dis-coveries permitted researchers to work with some transgenicmodels of AD, such as APP and APP/PS1 transgenic mice.In the present study, APP/PS1 double transgenic mice over-expressing chimeric mouse/human APP with the Swedishdouble mutation (K595N and M596L) and human PS1-ΔE9
(deletion of presenilin 1 exon 9) mutation were used [3, 4].This line is originally maintained in a hybrid C3HeJ C57BL6/J F1 background and manifests a rapid accumulation ofamyloid plaques in the cortex and hippocampus beginning atabout 3 months of age [4–7]. Additionally, these mice exhibitother pathological featured alterations associated with AD[8, 9].
Unfortunately, there have been few safe and effectivemethods for the treatment or prevention of AD at present.Given the situation, the use of natural medicinal plants andtraditional Chinese medicines as remedies cast new light onsome chronic or complicated disorders, such as AD, whichinvolves many unknown mechanisms and various systems in

2 Evidence-Based Complementary and Alternative Medicine
the pathogenesis. WNK is an extracts combination of threetraditional Chinese herbal medicines: P. ginseng, G. biloba,and C. sativus L., which have been used as traditional Chinesetreatments for several millennia for the treatment of a varietyof conditions, including age-related memory decline. Inrecent years, they have also become increasingly popularthroughout Western society, both as “over-the-counter”herbal supplements and as prescription drugs [10, 11]. Overrecent years, especially, certain active constituents isolatedfrom them have been proved to be essential to cognitiveefficacy in several paradigms. For example, ginsenosides, gin-seng’s main active ingredient, can enhance psychomotor andcognitive performance and can benefit AD by improvingbrain cholinergic function, reducing the level of Aβ andrepairing damaged neuronal networks [12, 13]. Evidencesuggests that chronic administration of the standardized G.biloba extracts (EGB761), containing 24% of Ginkgo glyco-sides and 6% of ginkgolides-bilobalide, can ameliorate thecognitive decline that occurs during ageing and may alsoplay a beneficial role in the attenuation of the cognitivedeficits associated with a number of pathological conditions,including intermittent claudication, vascular dementia, AD,and cerebral insufficiency—a generalised condition with apresumed cerebrovascular aetiology [14–19]. C. sativus L.,commonly known as saffron, is used in traditional Chinesemedicine as a nerve sedative, antispasmodic, anticatarrhal,stimulant, and so forth. Modern pharmacological studieshave demonstrated that saffron or its active constituents hasmemory-improved, Aβ aggregation-inhibited, radical scav-enging, hypolipaemic, and antitumour effects [20–27].Improvements in memory performance have also been dem-onstrated preclinically in both young and old rats, as well asin AD rat models, and clinically in healthy middle-aged andneurasthenic human cohorts, following chronic administra-tion of ginseng, ginkgo, and their combination [28–30].
However, it seems still vague of the current overall evi-dence that saffron has a predictable and clinically significantbenefit for people with dementia or cognitive impairment.Same situation occurred to ginkgo, although its extract isamong the most widely used complementary therapies today[31, 32]. In the present study, we investigated the effect andthe possible mechanisms of action of WNK in an APP/PS1transgenic mouse model of AD. The effects of WNK andEGB761 on maze performance were compared simultane-ously.
2. Materials and Methods
2.1. Animal. Animals of both sexes were used in this study.The transgenic APP/PS1 mice, which expressed both theSwedish double mutations of APP (K670N/M671L) andmutant PS1 (PS1M146L), were provided by the Institute ofLaboratory Animal Science, Chinese Academy of MedicalSciences, and Peking Union Medical College, Beijing, China.APP695SWE/co + PS1/ΔE9 transgenic (n = 29) and litter-mate wild-type mice (n = 10), controlled for age (5 months,range 4–6), were separated into 4 groups: APP/PS1+tapwater (transgenic group), n = 9; APP/PS1 + WNK 44 mg/kg/
Table 1: Concentrations of active components of WNK.
Active components Concentrations (%)
Ginsenoside Rg1 5.5
Ginsenoside Re 3.2
Ginsenoside Rb1 13.1
Flavonoids 28.7
Total terpene lactones 11.6
Ginkgolide A 2.9
day (WNK group), n = 10; EGB761 30 mg/kg/day (EGB761group), n = 10; nontransgenic littermates + tap water (non-transgenic group), n = 10. There was no intergroup differ-ence in body weight (P > 0.05, data not shown).
Mice were fed in separate home cages with free access towater in the special animal room under a 12 : 12 h light/darkcycle and thermoregulated environment. The mice wereacclimatized to laboratory conditions before the behaviouraltest. All the procedures took place between 8 AM up to 4 PM,and each mouse was used only one time in the water maze.All experiments were conducted according to the guidelinesfor care and use of animals approved by Xiyuan Hospital,China Academy of Chinese Medical Sciences, China.
2.2. Chemicals and Reagents. EGB761 (batch no.: c02191),the G. biloba leaves extract tablets (EGb761), was product ofBeaufour Ipsen Industrie, Dreux, France. Rabbit anti-humanAβ antibody was purchased from Biosynthesis Biotechnol-ogy, Beijing, China. Goat anti-rabbit IgG antibody was fromAbcam, Cambridge, MA, USA. Ethanol, formaldehyde, andother reagents used in this study were all HPLC-gradeobtained from Fisher Scientific, New Jerseys, USA.
2.3. Formulation of WNK and Concentration Control of ActiveComponents. WNK (Wei Nao Kang) is a proprietary formula(China: ZL02131435.7; Russia: 2008144409/21(057916);Australia: 2006342350, etc.) consisting of extracts of threetraditional Chinese herbs, P. ginseng, G. biloba, and saffron,that is, total ginsenosides, total gingkgo flavonoids, and totalsaffron glycosides. WNK’s dose used in this study wasobtained according to the results of two independent orthog-onal designs with animal models of D-galactose-injectedmice and focal cerebral ischemia-reperfusion rats (data notshown). In this study, the combination of total ginsenosides(batch no.: 100909), total ginkgo flavonoids (batch no.:100914), total saffron glycosides (batch no.: 100910), wasprovided by Shineway Pharmaceutical Group, Hebei, China.HPLC and UV were used to analyze and control the activecomponents of WNK (Table 1).
2.4. Drug Administration. Mice of WNK or EGB761 groupswere administered with WNK (44 mg/kg/day) or EGB761(30 mg/kg/day) intragastrically in a volume of 10 mL/kg oncea day for 3 months. Mice of nontransgenic and transgenicgroups were given tap water with the same volume.

Evidence-Based Complementary and Alternative Medicine 3
2.5. Behavioural Assessment. Since the Morris water maze(MWM) test evaluating spatial learning has been shown to besensitive to APP/PS1 transgenic models, it was used to assesshippocampal-dependent spatial learning and memory in thisstudy [33–35].
The water maze was a white circular pool (80 cm indiameter, 50 cm deep) divided into four equal imaginaryquadrants for data analysis. The water temperature wasmaintained between 21 and 23◦C. One centimeter beneaththe surface of the water and hidden from mouse view wasa white circular platform 10 cm in diameter. The swimmingpatterns of the mice were recorded with a video cameramounted above the center of the pool and analyzed with avideo-tracking system. The water maze was located in a roomwith several visual stimuli hanging on the walls to providespatial cues. The water in the pool contained dark ink so thatthe mice could not see the platform. The time mice spent onfinding the platform was recorded as the escape latency. Everymouse has 4 times of swimming each day, released at the first,second, third, and fourth quadrant, respectively. Mice notfinding the platform within 180 s were guided to it by theexperimenter. This part of test was repeated for 4 days, andthe location of the platform was not changed. At the fifth day,the platform was removed and the mice were released at thefirst quadrant. The time spent swimming in the target quad-rant (where the platform was located during hidden platformtraining) was considered as a measure of platform locationretention.
2.6. Histopathology. One day after behavioral analysis, micein different groups were anesthetized with 5% chloral hydrateand perfused through the left ventricle with 0.9% saline. Thebrain was removed from the skull and then postfixed for 24 hwith 4% paraformaldehyde. After being progressive dehy-drated, the brain was embedded in paraffins. Four μm thickconsecutive coronal paraffin sections were collected through-out the cerebral cortex and hippocampus, according to theMouse Brain in Stereotaxic Coordinates. Each group con-sisted of three mice. Hematoxylin and eosin (HE) sectionswere obtained every 20 sections. Photomicroscopy was donewith an Olympus BX41 light microscope and Anymicro DSSImage Analysis software. Exposure settings were identicalamong sections per magnification using ×400 objectives.Once linear best-fit setting was identified, this was used forall sections per magnification and stain type. Minimal imageadjustments were applied to match the actual image whenviewed directly through the eyepiece.
2.7. Immunohistochemistry. Mice in different groups wereanesthetized with 5% chloral hydrate and perfused withsaline and then 4% paraformaldehyde through the left ven-tricle. Serial 4 μm thick paraffin sections were cut throughouthippocampus and cortex according to The Mouse BrainAtlas (Paxinos and Franklin, 2001) and mounted on APES-coated slides. Sections were deparaffinized in xylol, followedby heat-mediated antigen retrieval in 10 mM citrate buffer(pH = 6.0) and incubation with 3% H2O2 for 40 min at 37◦C.Sections were then incubated with 10% normal goat serum
for 20 min at 37◦C to block the nonspecific binding followedby incubation overnight at 4◦C with antibody against Aβ(1 : 300). Sections were washed in 0.1 M PBS, incubatedsequentially with fluorescent-labelled secondary antibody,goat anti-rabbit IgG (1 : 200) for 45 min at 37◦C. Fluorescentsignals were detected using an Olympus 1 × 81 microscopewith a Rolera-MGi Plus back-illuminated EMCCD camera(Qimaging, Surrey, Canada) using identical exposure times.Controls received identical treatment except that primaryantibody was omitted and showed no specific staining. Inten-sity was detected by Image-Pro Plus (In Vivo Version 6.0,MediaCybernetics Inc.) using ROIs.
2.8. Ultrastructure. The brains were removed immediatelyfrom the skull, pieces of selected brain regions (hippocampusCA1 area) were separated and immediately placed in 2.5%glutaraldehyde, chopped to get pieces of about 1 mm2, andkept in 2.5% glutaraldehyde for more than 12 h at room tem-perature. Samples were postfixed in 1% osmium tetroxide for1 h, dehydrated in graded increasing ethanol, and embeddedin epoxy resin. Polymerization was performed at 80◦C for24 h. Blocks were cut on a Reichert ultramicrotome intoultrathin sections (60–70 nm), which were poststained withuranyl acetate and lead citrate, and viewed under a Hitachi7100 electron microscopy.
2.9. Statistical Analysis. All data were expressed as mean ±SD, and statistical significance was evaluated by a one-wayANOVA or t-test. The data of escape latency in behavioraltesting was analysed using repeated measures of ANOVAanalyze. Values with P < 0.05 were considered significant forall the analysis.
3. Results
3.1. Learning and Memory Performance Enhancement. Themean escape latency was recorded from four quadrants inone day. During the course of the experiment, the meanlatency of nontransgenic, WNK- and EGB761-treated micein finding the hidden platform became shorter from thefirst day to the fourth day. The results showed significantdifference between transgenic mice and their nontransgeniclittermates (P < 0.05, P < 0.01), and the differences amongWNK- or EGB761-treated mice and transgenic mice werealso significant (P < 0.05, P < 0.01), especially at the final dayof the invisible platform task. Besides, the results indicatedthat the learning ability was different between WNK andEGB761-treated mice, since the latency was found decreasedto a higher degree in WNK group at the fourth day (P < 0.05;Figure 1(a)).
The time that mice swam in the first quadrant, the previ-ous platform location, at the fifth day indicates the memoryability. In this study, the time that the transgenic miceswam in the target quadrant was significantly shorter thanthat of the nontransgenic mice (P < 0.05), while it was signi-ficantly longer, compared the WNK-treated mice with thetransgenic ones (P < 0.05). No significant difference was
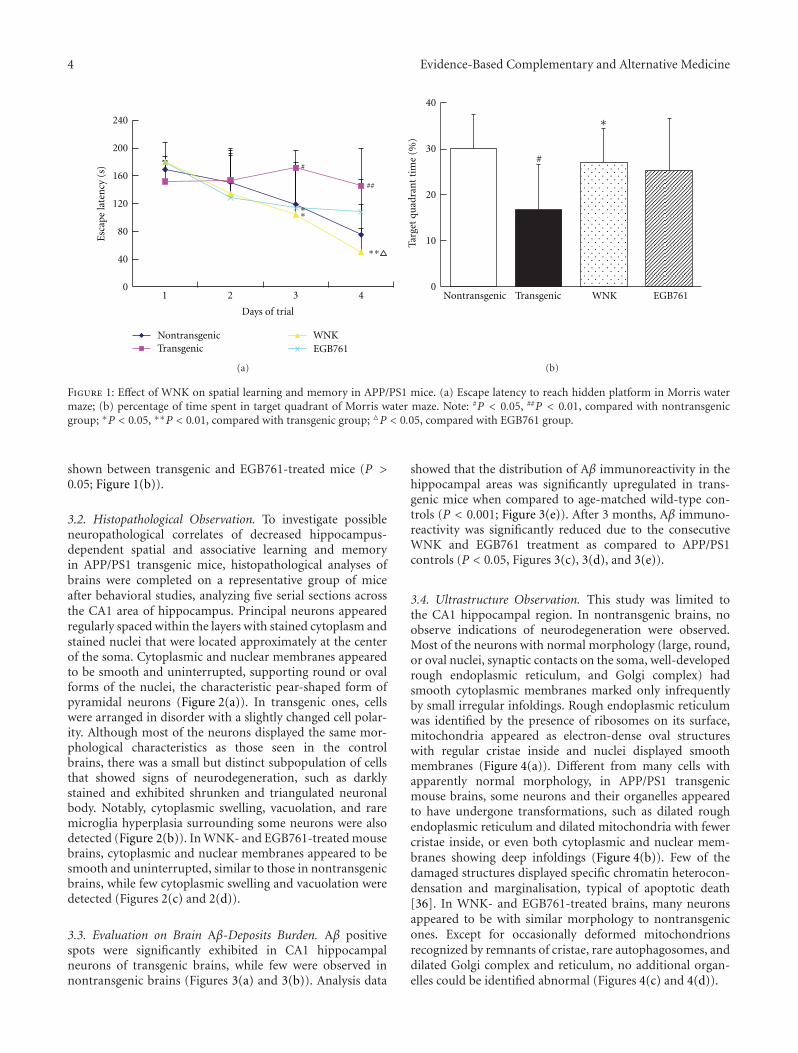
4 Evidence-Based Complementary and Alternative Medicine
1 2 3 4
Days of trial
240
0
40
80
120
160
200
TransgenicWNKEGB761
#
##
∗∗
∗∗
Nontransgenic
Esc
ape
late
ncy
(s)
(a)
0
10
20
30
40
Targ
et q
uad
ran
t ti
me
(%)
Transgenic WNK EGB761
#
∗
Nontransgenic
(b)
Figure 1: Effect of WNK on spatial learning and memory in APP/PS1 mice. (a) Escape latency to reach hidden platform in Morris watermaze; (b) percentage of time spent in target quadrant of Morris water maze. Note: #P < 0.05, ##P < 0.01, compared with nontransgenicgroup; ∗P < 0.05, ∗∗P < 0.01, compared with transgenic group; �P < 0.05, compared with EGB761 group.
shown between transgenic and EGB761-treated mice (P >0.05; Figure 1(b)).
3.2. Histopathological Observation. To investigate possibleneuropathological correlates of decreased hippocampus-dependent spatial and associative learning and memoryin APP/PS1 transgenic mice, histopathological analyses ofbrains were completed on a representative group of miceafter behavioral studies, analyzing five serial sections acrossthe CA1 area of hippocampus. Principal neurons appearedregularly spaced within the layers with stained cytoplasm andstained nuclei that were located approximately at the centerof the soma. Cytoplasmic and nuclear membranes appearedto be smooth and uninterrupted, supporting round or ovalforms of the nuclei, the characteristic pear-shaped form ofpyramidal neurons (Figure 2(a)). In transgenic ones, cellswere arranged in disorder with a slightly changed cell polar-ity. Although most of the neurons displayed the same mor-phological characteristics as those seen in the controlbrains, there was a small but distinct subpopulation of cellsthat showed signs of neurodegeneration, such as darklystained and exhibited shrunken and triangulated neuronalbody. Notably, cytoplasmic swelling, vacuolation, and raremicroglia hyperplasia surrounding some neurons were alsodetected (Figure 2(b)). In WNK- and EGB761-treated mousebrains, cytoplasmic and nuclear membranes appeared to besmooth and uninterrupted, similar to those in nontransgenicbrains, while few cytoplasmic swelling and vacuolation weredetected (Figures 2(c) and 2(d)).
3.3. Evaluation on Brain Aβ-Deposits Burden. Aβ positivespots were significantly exhibited in CA1 hippocampalneurons of transgenic brains, while few were observed innontransgenic brains (Figures 3(a) and 3(b)). Analysis data
showed that the distribution of Aβ immunoreactivity in thehippocampal areas was significantly upregulated in trans-genic mice when compared to age-matched wild-type con-trols (P < 0.001; Figure 3(e)). After 3 months, Aβ immuno-reactivity was significantly reduced due to the consecutiveWNK and EGB761 treatment as compared to APP/PS1controls (P < 0.05, Figures 3(c), 3(d), and 3(e)).
3.4. Ultrastructure Observation. This study was limited tothe CA1 hippocampal region. In nontransgenic brains, noobserve indications of neurodegeneration were observed.Most of the neurons with normal morphology (large, round,or oval nuclei, synaptic contacts on the soma, well-developedrough endoplasmic reticulum, and Golgi complex) hadsmooth cytoplasmic membranes marked only infrequentlyby small irregular infoldings. Rough endoplasmic reticulumwas identified by the presence of ribosomes on its surface,mitochondria appeared as electron-dense oval structureswith regular cristae inside and nuclei displayed smoothmembranes (Figure 4(a)). Different from many cells withapparently normal morphology, in APP/PS1 transgenicmouse brains, some neurons and their organelles appearedto have undergone transformations, such as dilated roughendoplasmic reticulum and dilated mitochondria with fewercristae inside, or even both cytoplasmic and nuclear mem-branes showing deep infoldings (Figure 4(b)). Few of thedamaged structures displayed specific chromatin heterocon-densation and marginalisation, typical of apoptotic death[36]. In WNK- and EGB761-treated brains, many neuronsappeared to be with similar morphology to nontransgenicones. Except for occasionally deformed mitochondrionsrecognized by remnants of cristae, rare autophagosomes, anddilated Golgi complex and reticulum, no additional organ-elles could be identified abnormal (Figures 4(c) and 4(d)).
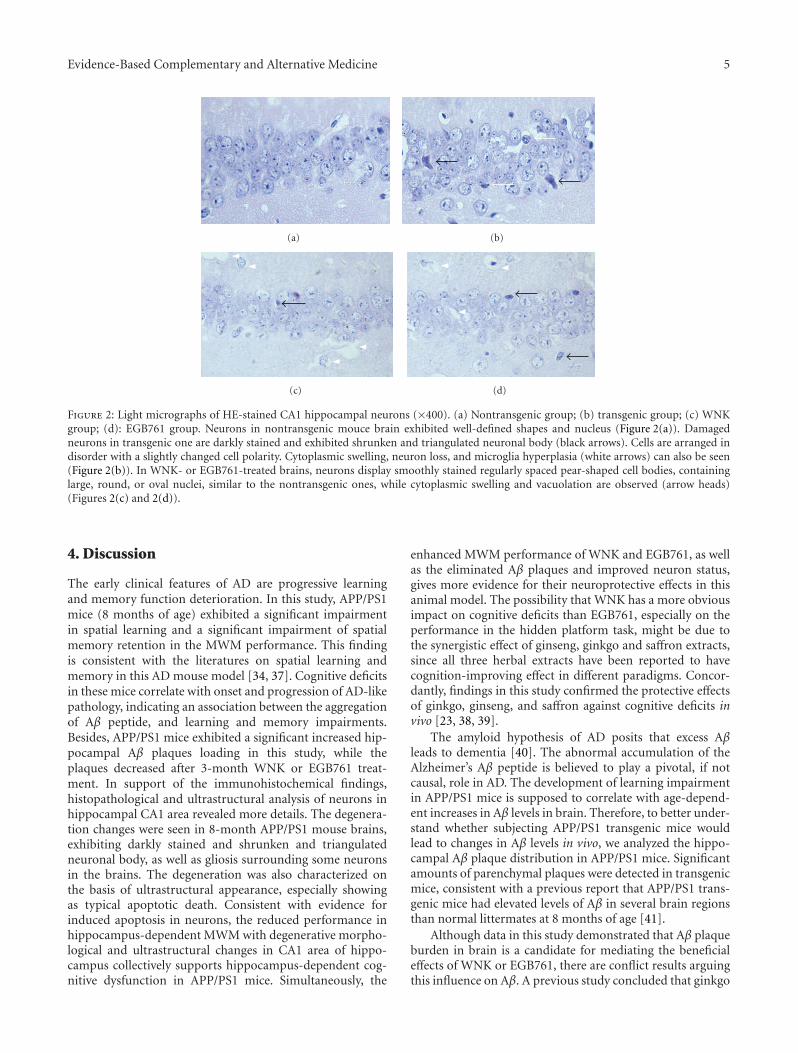
Evidence-Based Complementary and Alternative Medicine 5
(a) (b)
(c) (d)
Figure 2: Light micrographs of HE-stained CA1 hippocampal neurons (×400). (a) Nontransgenic group; (b) transgenic group; (c) WNKgroup; (d): EGB761 group. Neurons in nontransgenic mouce brain exhibited well-defined shapes and nucleus (Figure 2(a)). Damagedneurons in transgenic one are darkly stained and exhibited shrunken and triangulated neuronal body (black arrows). Cells are arranged indisorder with a slightly changed cell polarity. Cytoplasmic swelling, neuron loss, and microglia hyperplasia (white arrows) can also be seen(Figure 2(b)). In WNK- or EGB761-treated brains, neurons display smoothly stained regularly spaced pear-shaped cell bodies, containinglarge, round, or oval nuclei, similar to the nontransgenic ones, while cytoplasmic swelling and vacuolation are observed (arrow heads)(Figures 2(c) and 2(d)).
4. Discussion
The early clinical features of AD are progressive learningand memory function deterioration. In this study, APP/PS1mice (8 months of age) exhibited a significant impairmentin spatial learning and a significant impairment of spatialmemory retention in the MWM performance. This findingis consistent with the literatures on spatial learning andmemory in this AD mouse model [34, 37]. Cognitive deficitsin these mice correlate with onset and progression of AD-likepathology, indicating an association between the aggregationof Aβ peptide, and learning and memory impairments.Besides, APP/PS1 mice exhibited a significant increased hip-pocampal Aβ plaques loading in this study, while theplaques decreased after 3-month WNK or EGB761 treat-ment. In support of the immunohistochemical findings,histopathological and ultrastructural analysis of neurons inhippocampal CA1 area revealed more details. The degenera-tion changes were seen in 8-month APP/PS1 mouse brains,exhibiting darkly stained and shrunken and triangulatedneuronal body, as well as gliosis surrounding some neuronsin the brains. The degeneration was also characterized onthe basis of ultrastructural appearance, especially showingas typical apoptotic death. Consistent with evidence forinduced apoptosis in neurons, the reduced performance inhippocampus-dependent MWM with degenerative morpho-logical and ultrastructural changes in CA1 area of hippo-campus collectively supports hippocampus-dependent cog-nitive dysfunction in APP/PS1 mice. Simultaneously, the
enhanced MWM performance of WNK and EGB761, as wellas the eliminated Aβ plaques and improved neuron status,gives more evidence for their neuroprotective effects in thisanimal model. The possibility that WNK has a more obviousimpact on cognitive deficits than EGB761, especially on theperformance in the hidden platform task, might be due tothe synergistic effect of ginseng, ginkgo and saffron extracts,since all three herbal extracts have been reported to havecognition-improving effect in different paradigms. Concor-dantly, findings in this study confirmed the protective effectsof ginkgo, ginseng, and saffron against cognitive deficits invivo [23, 38, 39].
The amyloid hypothesis of AD posits that excess Aβleads to dementia [40]. The abnormal accumulation of theAlzheimer’s Aβ peptide is believed to play a pivotal, if notcausal, role in AD. The development of learning impairmentin APP/PS1 mice is supposed to correlate with age-depend-ent increases in Aβ levels in brain. Therefore, to better under-stand whether subjecting APP/PS1 transgenic mice wouldlead to changes in Aβ levels in vivo, we analyzed the hippo-campal Aβ plaque distribution in APP/PS1 mice. Significantamounts of parenchymal plaques were detected in transgenicmice, consistent with a previous report that APP/PS1 trans-genic mice had elevated levels of Aβ in several brain regionsthan normal littermates at 8 months of age [41].
Although data in this study demonstrated that Aβ plaqueburden in brain is a candidate for mediating the beneficialeffects of WNK or EGB761, there are conflict results arguingthis influence on Aβ. A previous study concluded that ginkgo
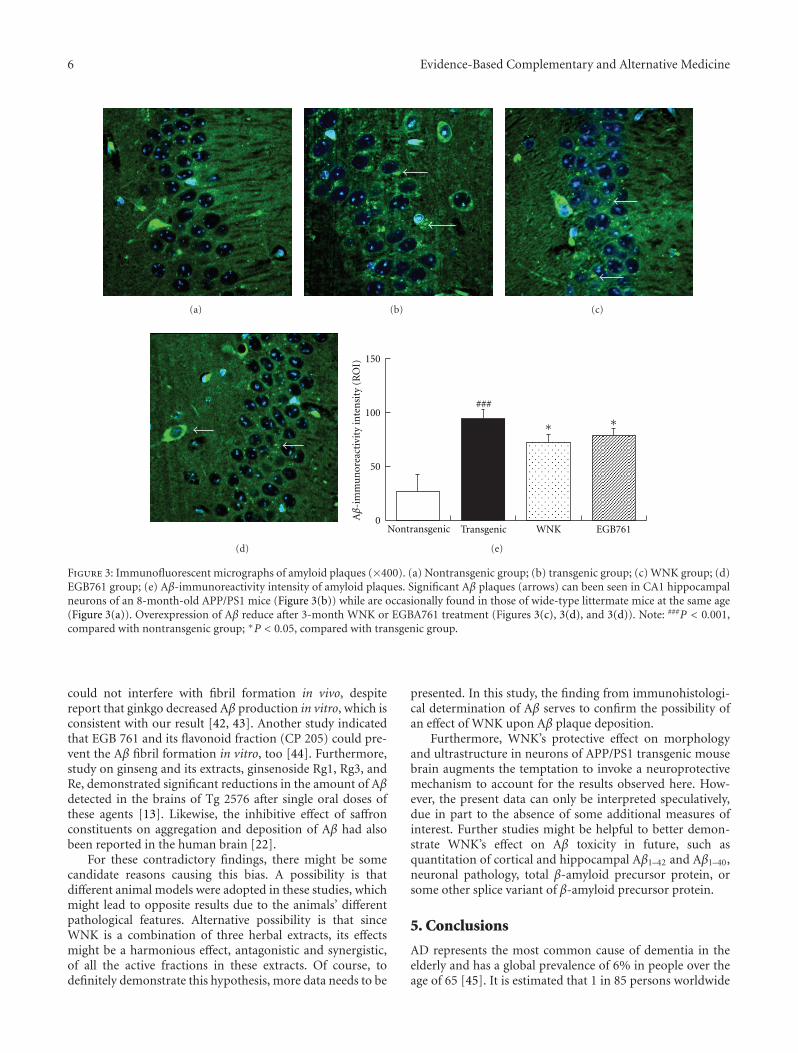
6 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d)
0
50
100
150
Transgenic WNK EGB761
Aβ
-im
mu
nor
eact
ivit
y in
ten
sity
(R
OI)
∗∗
###
Nontransgenic
(e)
Figure 3: Immunofluorescent micrographs of amyloid plaques (×400). (a) Nontransgenic group; (b) transgenic group; (c) WNK group; (d)EGB761 group; (e) Aβ-immunoreactivity intensity of amyloid plaques. Significant Aβ plaques (arrows) can been seen in CA1 hippocampalneurons of an 8-month-old APP/PS1 mice (Figure 3(b)) while are occasionally found in those of wide-type littermate mice at the same age(Figure 3(a)). Overexpression of Aβ reduce after 3-month WNK or EGBA761 treatment (Figures 3(c), 3(d), and 3(d)). Note: ###P < 0.001,compared with nontransgenic group; ∗P < 0.05, compared with transgenic group.
could not interfere with fibril formation in vivo, despitereport that ginkgo decreased Aβ production in vitro, which isconsistent with our result [42, 43]. Another study indicatedthat EGB 761 and its flavonoid fraction (CP 205) could pre-vent the Aβ fibril formation in vitro, too [44]. Furthermore,study on ginseng and its extracts, ginsenoside Rg1, Rg3, andRe, demonstrated significant reductions in the amount of Aβdetected in the brains of Tg 2576 after single oral doses ofthese agents [13]. Likewise, the inhibitive effect of saffronconstituents on aggregation and deposition of Aβ had alsobeen reported in the human brain [22].
For these contradictory findings, there might be somecandidate reasons causing this bias. A possibility is thatdifferent animal models were adopted in these studies, whichmight lead to opposite results due to the animals’ differentpathological features. Alternative possibility is that sinceWNK is a combination of three herbal extracts, its effectsmight be a harmonious effect, antagonistic and synergistic,of all the active fractions in these extracts. Of course, todefinitely demonstrate this hypothesis, more data needs to be
presented. In this study, the finding from immunohistologi-cal determination of Aβ serves to confirm the possibility ofan effect of WNK upon Aβ plaque deposition.
Furthermore, WNK’s protective effect on morphologyand ultrastructure in neurons of APP/PS1 transgenic mousebrain augments the temptation to invoke a neuroprotectivemechanism to account for the results observed here. How-ever, the present data can only be interpreted speculatively,due in part to the absence of some additional measures ofinterest. Further studies might be helpful to better demon-strate WNK’s effect on Aβ toxicity in future, such asquantitation of cortical and hippocampal Aβ1–42 and Aβ1–40,neuronal pathology, total β-amyloid precursor protein, orsome other splice variant of β-amyloid precursor protein.
5. Conclusions
AD represents the most common cause of dementia in theelderly and has a global prevalence of 6% in people over theage of 65 [45]. It is estimated that 1 in 85 persons worldwide
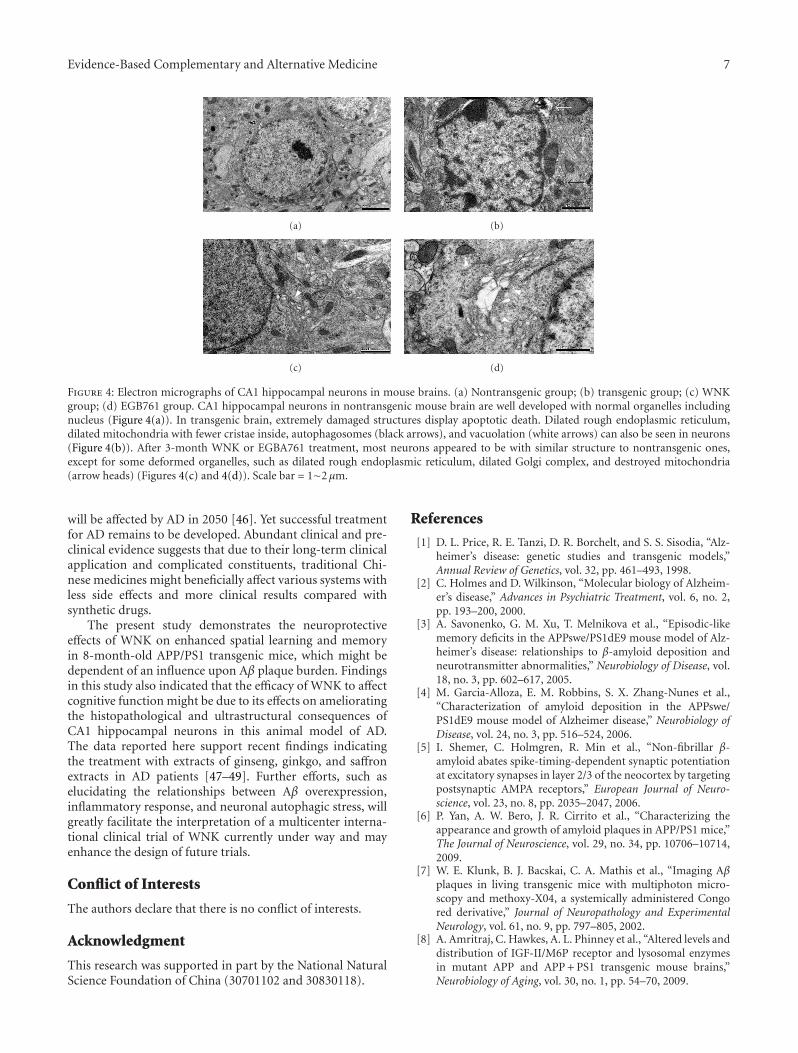
Evidence-Based Complementary and Alternative Medicine 7
(a) (b)
(c) (d)
Figure 4: Electron micrographs of CA1 hippocampal neurons in mouse brains. (a) Nontransgenic group; (b) transgenic group; (c) WNKgroup; (d) EGB761 group. CA1 hippocampal neurons in nontransgenic mouse brain are well developed with normal organelles includingnucleus (Figure 4(a)). In transgenic brain, extremely damaged structures display apoptotic death. Dilated rough endoplasmic reticulum,dilated mitochondria with fewer cristae inside, autophagosomes (black arrows), and vacuolation (white arrows) can also be seen in neurons(Figure 4(b)). After 3-month WNK or EGBA761 treatment, most neurons appeared to be with similar structure to nontransgenic ones,except for some deformed organelles, such as dilated rough endoplasmic reticulum, dilated Golgi complex, and destroyed mitochondria(arrow heads) (Figures 4(c) and 4(d)). Scale bar = 1∼2 μm.
will be affected by AD in 2050 [46]. Yet successful treatmentfor AD remains to be developed. Abundant clinical and pre-clinical evidence suggests that due to their long-term clinicalapplication and complicated constituents, traditional Chi-nese medicines might beneficially affect various systems withless side effects and more clinical results compared withsynthetic drugs.
The present study demonstrates the neuroprotectiveeffects of WNK on enhanced spatial learning and memoryin 8-month-old APP/PS1 transgenic mice, which might bedependent of an influence upon Aβ plaque burden. Findingsin this study also indicated that the efficacy of WNK to affectcognitive function might be due to its effects on amelioratingthe histopathological and ultrastructural consequences ofCA1 hippocampal neurons in this animal model of AD.The data reported here support recent findings indicatingthe treatment with extracts of ginseng, ginkgo, and saffronextracts in AD patients [47–49]. Further efforts, such aselucidating the relationships between Aβ overexpression,inflammatory response, and neuronal autophagic stress, willgreatly facilitate the interpretation of a multicenter interna-tional clinical trial of WNK currently under way and mayenhance the design of future trials.
Conflict of Interests
The authors declare that there is no conflict of interests.
Acknowledgment
This research was supported in part by the National NaturalScience Foundation of China (30701102 and 30830118).
References
[1] D. L. Price, R. E. Tanzi, D. R. Borchelt, and S. S. Sisodia, “Alz-heimer’s disease: genetic studies and transgenic models,”Annual Review of Genetics, vol. 32, pp. 461–493, 1998.
[2] C. Holmes and D. Wilkinson, “Molecular biology of Alzheim-er’s disease,” Advances in Psychiatric Treatment, vol. 6, no. 2,pp. 193–200, 2000.
[3] A. Savonenko, G. M. Xu, T. Melnikova et al., “Episodic-likememory deficits in the APPswe/PS1dE9 mouse model of Alz-heimer’s disease: relationships to β-amyloid deposition andneurotransmitter abnormalities,” Neurobiology of Disease, vol.18, no. 3, pp. 602–617, 2005.
[4] M. Garcia-Alloza, E. M. Robbins, S. X. Zhang-Nunes et al.,“Characterization of amyloid deposition in the APPswe/PS1dE9 mouse model of Alzheimer disease,” Neurobiology ofDisease, vol. 24, no. 3, pp. 516–524, 2006.
[5] I. Shemer, C. Holmgren, R. Min et al., “Non-fibrillar β-amyloid abates spike-timing-dependent synaptic potentiationat excitatory synapses in layer 2/3 of the neocortex by targetingpostsynaptic AMPA receptors,” European Journal of Neuro-science, vol. 23, no. 8, pp. 2035–2047, 2006.
[6] P. Yan, A. W. Bero, J. R. Cirrito et al., “Characterizing theappearance and growth of amyloid plaques in APP/PS1 mice,”The Journal of Neuroscience, vol. 29, no. 34, pp. 10706–10714,2009.
[7] W. E. Klunk, B. J. Bacskai, C. A. Mathis et al., “Imaging Aβplaques in living transgenic mice with multiphoton micro-scopy and methoxy-X04, a systemically administered Congored derivative,” Journal of Neuropathology and ExperimentalNeurology, vol. 61, no. 9, pp. 797–805, 2002.
[8] A. Amritraj, C. Hawkes, A. L. Phinney et al., “Altered levels anddistribution of IGF-II/M6P receptor and lysosomal enzymesin mutant APP and APP + PS1 transgenic mouse brains,”Neurobiology of Aging, vol. 30, no. 1, pp. 54–70, 2009.

8 Evidence-Based Complementary and Alternative Medicine
[9] W. Xu, Y. Q. Zhan, W. Huang, X. Wang, S. Zhang, and H. Lei,“Reduction of hippocampal N-acetyl aspartate level in agedAPPSwe/PS1dE9 transgenic mice is associated with degen-eration of CA3 pyramidal neurons,” Journal of NeuroscienceResearch, vol. 88, no. 14, pp. 3155–3160, 2010.
[10] M. O’Hara, D. Kiefer, K. Farrell, and K. Kemper, “A review of12 commonly used medicinal herbs,” Archives of Family Medi-cine, vol. 7, no. 6, pp. 523–536, 1998.
[11] C. N. Gillis, “Panax ginseng pharmacology: a nitric oxidelink?” Biochemical Pharmacology, vol. 54, no. 1, pp. 1–8, 1997.
[12] J. Liu, W. Cong, L. Xu, and J. N. Wang, “Effect of combinationof extracts of ginseng and ginkgo biloba on acetylcholine inamyloid beta-protein-treated rats determined by an improvedHPLC,” Acta Pharmacologica Sinica, vol. 25, no. 9, pp. 1118–1123, 2004.
[13] F. Chen, E. A. Eckman, and C. B. Eckman, “Reductions inlevels of the Alzheimer’s amyloid beta peptide after oral admi-nistration of ginsenosides.,” The FASEB Journal, vol. 20, no. 8,pp. 1269–1271, 2006.
[14] H. Allain, P. Raoul, A. Lieury, F. LeCoz, J. M. Gandon, andP. D’Arbigny, “Effect of two doses of Gingko biloba extract(EGb 761) on the dual-coding test in elderly subjects,” ClinicalTherapeutics, vol. 15, no. 3, pp. 549–558, 1993.
[15] G. S. Rai, C. Shovlin, and K. A. Wesnes, “A double-blind,placebo controlled study of Ginkgo biloba extract (‘Tanakan’)in elderly outpatients with mild to moderate memory impair-ment,” Current Medical Research and Opinion, vol. 12, no. 6,pp. 350–355, 1991.
[16] H. Draebeck, J. R. Petersen, N. Winberg et al., “The effect ofGinkgo biloba in patients with intermittent claudicationm,”Ugeskrif for Laeger, vol. 158, no. 27, pp. 3928–3931, 1996.
[17] S. Kanowski, W. M. Herrmann, K. Stephan, W. Wierich, andR. Horr, “Proof of efficacy of the ginkgo biloba special extractEGb 761 in outpatients suffering from mild to moderateprimary degenerative dementia of the Alzheimer type ormulti-infarct dementia,” Pharmacopsychiatry, vol. 29, no. 2,pp. 47–56, 1996.
[18] P. L. Le Bars, M. M. Katz, N. Berman, T. M. Itil, A. M. Freed-man, and A. F. Schatzberg, “A placebo-controlled, double-blind, randomized trial of an extract of Ginkgo biloba fordementia,” Journal of the American Medical Association, vol.278, no. 16, pp. 1327–1332, 1997.
[19] J. Kleijnen and P. Knipschild, “Ginkgo biloba for cerebralinsufficiency,” British Journal of Clinical Pharmacology, vol. 34,no. 4, pp. 352–358, 1992.
[20] Y. Zhang, Y. Shoyama, M. Sugiura, and H. Saito, “Effectsof Crocus sativus L. on the ethanol-induced impairment ofpassive avoidance performances in mice,” Biological and Phar-maceutical Bulletin, vol. 17, no. 2, pp. 217–221, 1994.
[21] H. Hosseinzadeh, H. R. Sadeghnia, F. A. Ghaeni, V. S.Motamedshariaty, and S. A. Mohajeri, “Effects of saffron (Cro-cus sativus L.) and its active constituent, crocin, on recognitionand spatial memory after chronic cerebral hypoperfusion inrats,” Phytotherapy Research, vol. 26, no. 3, pp. 381–386, 2011.
[22] M. A. Papandreou, C. D. Kanakis, M. G. Polissiou et al., “Inhi-bitory activity on amyloid-β aggregation and antioxidant pro-perties of Crocus sativus stigmas extract and its crocin con-stituents,” Journal of Agricultural and Food Chemistry, vol. 54,no. 23, pp. 8762–8768, 2006.
[23] B. Ghadrdoost, A. A. Vafaei, A. Rashidy-Pour et al., “Protectiveeffects of saffron extract and its active constituent crocinagainst oxidative stress and spatial learning and memory def-icits induced by chronic stress in rats,” European Journal ofPharmacology, vol. 667, no. 1–3, pp. 222–229, 2011.
[24] K. Abe and H. Saito, “Effects of saffron extract and its con-stituent crocin on learning behaviour and long-term potentia-tion,” Phytotherapy Research, vol. 14, no. 3, pp. 149–152, 2000.
[25] T. Ochiai, H. Shimeno, K. I. Mishima et al., “Protective effectsof carotenoids from saffron on neuronal injury in vitro and invivo,” Biochimica et Biophysica Acta, vol. 1770, no. 4, pp. 578–584, 2007.
[26] N. Pitsikas and N. Sakellaridis, “Crocus sativus L. extractsantagonize memory impairments in different behaviouraltasks in the rat,” Behavioural Brain Research, vol. 173, no. 1,pp. 112–115, 2006.
[27] N. Pitsikas, S. Zisopoulou, P. A. Tarantilis, C. D. Kanakis, M.G. Polissiou, and N. Sakellaridis, “Effects of the active con-stituents of Crocus sativus L., crocins on recognition andspatial rats’ memory,” Behavioural Brain Research, vol. 183, no.2, pp. 141–146, 2007.
[28] V. D. Petkov, R. Kehayov, S. Belcheva et al., “Memory effects ofstandardized extracts of Panax ginseng (G115), Ginkgo biloba(GK501) and their combination Gincosan (PHL-00701),”Planta Medica, vol. 59, no. 2, pp. 106–114, 1993.
[29] K. A. Wesnes, T. Ward, A. McGinty, and O. Petrini, “Thememory enhancing effects of a Ginkgo biloba/Panax ginsengcombination in healthy middle-aged volunteers,” Psychophar-macology, vol. 152, no. 4, pp. 353–361, 2000.
[30] K. A. Wesnes, R. A. Faleni, N. R. Hefting et al., “The cognitive,subjective, and physical effects of a Ginkgo biloba/Panaxginseng combination in healthy volunteers with neurastheniccomplaints,” Psychopharmacology Bulletin, vol. 33, no. 4, pp.677–683, 1997.
[31] M. Mazza, A. Capuano, P. Bria, and S. Mazza, “Ginkgo bilobaand donepezil: A comparison in the treatment of Alzheimer’sdementia in a randomized placebo-controlled double-blindstudy,” European Journal of Neurology, vol. 13, no. 9, pp. 981–985, 2006.
[32] B. E. Snitz, E. S. O’Meara, M. C. Carlson et al., “Ginkgo bilobafor preventing cognitive decline in older adults a randomizedtrial,” JAMA, vol. 302, no. 24, pp. 2663–2670, 2009.
[33] R. Lalonde, H. D. Kim, J. A. Maxwell, and K. Fukuchi,“Exploratory activity and spatial learning in 12-month-oldAPP 695SWE/co + PS1/ΔE9 mice with amyloid plaques,”Neuroscience Letters, vol. 390, no. 2, pp. 87–92, 2005.
[34] H. L. Liu, G. Zhao, K. Cai, H. H. Zhao, and L. D. Shi,“Treadmill exercise prevents decline in spatial learning andmemory in APP/PS1 transgenic mice through improvementof hippocampal long-term potentiation,” Behavioural BrainResearch, vol. 218, no. 2, pp. 308–314, 2011.
[35] M. Filali, R. Lalonde, and S. Rivest, “Subchronic memantineadministration on spatial learning, exploratory activity, andnest-building in an APP/PS1 mouse model of Alzheimer’sdisease,” Neuropharmacology, vol. 60, no. 6, pp. 930–936, 2011.
[36] L. Pliss, V. J. Balcar, V. Bubenıkova, J. Pokorny, T. Fitzgibbon,and F. St’astny, “Morphology and ultrastructure of rat hip-pocampal formation after i.c.v. administration of N-acetyl-L-aspartyl-L-glutamate,” Neuroscience, vol. 122, no. 1, pp. 93–101, 2003.
[37] F. Trinchese, S. Liu, F. Battaglia, S. Walter, P. M. Mathews,and O. Arancio, “Progressive age-related development of Alz-heimer-like pathology in APP/PS1 mice,” Annals of Neurology,vol. 55, no. 6, pp. 801–814, 2004.
[38] S. T. Lee, K. Chu, J. Y. Sim, J. H. Heo, and M. Kim, “Panaxginseng enhances cognitive performance in Alzheimer dis-ease,” Alzheimer Disease and Associated Disorders, vol. 22, no.3, pp. 222–226, 2008.

Evidence-Based Complementary and Alternative Medicine 9
[39] O. Shif, K. Gillette, C. M. Damkaoutis, C. Carrano, S. J.Robbins, and J. R. Hoffman, “Effects of Ginkgo biloba admi-nistered after spatial learning on water maze and radial armmaze performance in young adult rats,” Pharmacology Bio-chemistry and Behavior, vol. 84, no. 1, pp. 17–25, 2006.
[40] J. Hardy and D. J. Selkoe, “The amyloid hypothesis of Alz-heimer’s disease: Progress and problems on the road to thera-peutics,” Science, vol. 297, no. 5580, pp. 353–356, 2002.
[41] R. Minkeviclene, P. Banerjee, and H. Tanila, “Memantineimproves spatial learning in a transgenic mouse model of Alz-heimer’s disease,” Journal of Pharmacology and ExperimentalTherapeutics, vol. 311, no. 2, pp. 677–682, 2004.
[42] R. W. Stackman, F. Eckenstein, B. Frei, D. Kulhanek, J. Nowlin,and J. F. Quinn, “Prevention of age-related spatial memorydeficits in a transgenic mouse model of Alzheimer’s diseaseby chronic Ginkgo biloba treatment,” Experimental Neurology,vol. 184, no. 1, pp. 510–520, 2003.
[43] Y. Luo, J. V. Smith, V. Paramasivam et al., “Inhibition of amy-loid-β aggregation and caspase-3 activation by the Ginkgobiloba extract EGb761,” Proceedings of the National Academyof Sciences of the United States of America, vol. 99, no. 19, pp.12197–12202, 2002.
[44] F. Longpre, P. Garneau, Y. christen, and C. Ramassamy, “Pro-tection by EGb 761 against β-amyloid-induced neurotoxicity:Involvement of NF-κB, SIRT1, and MAPKs pathways andinhibition of amyloid fibril formation,” Free Radical Biologyand Medicine, vol. 41, no. 12, pp. 1781–1794, 2006.
[45] C. P. Ferri, M. Prince, C. Brayne et al., “Global prevalence ofdementia: a Delphi consensus study,” Lancet, vol. 366, no.9503, pp. 2112–2117, 2005.
[46] R. Brookmeyer, E. Johnson, K. Ziegler-Graham, and H. M.Arrighi, “Forecasting the global burden of Alzheimer’s dis-ease,” Alzheimer’s and Dementia, vol. 3, no. 3, pp. 186–191,2007.
[47] S. Akhondzadeh, M. S. Sabet, M. H. Harirchian et al., “Saffronin the treatment of patients with mild to moderate Alzheimer’sdisease: a 16-week, randomized and placebo-controlled trial,”Journal of Clinical Pharmacy and Therapeutics, vol. 35, no. 5,pp. 581–588, 2010.
[48] R. Ihl, M. Tribanek, and N. Bachinskaya, “Baseline neu-ropsychiatric symptoms are effect modifiers in Ginkgo bilobaextract (EGb 761�) treatment of dementia with neuropsy-chiatric features. Retrospective data analyses of a randomizedcontrolled trial,” Journal of the Neurological Sciences, vol. 299,no. 1-2, pp. 184–187, 2010.
[49] J. Zhong, J. Z. Tian, A. H. Zhu, and C. Z. Yang, “Clinical studyon a randomized, double-blind control of Shenwu gelatincapsule in treatment of mild cognitive impairment,” ZhongguoZhongyao Zazhi, vol. 32, no. 17, pp. 1800–1803, 2007.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 682720, 11 pagesdoi:10.1155/2012/682720
Research Article
A Pilot Study for the Neuroprotective Effect ofGongjin-dan on Transient Middle Cerebral ArteryOcclusion-Induced Ischemic Rat Brain
Yun-Young Sunwoo,1 Sang In Park,1 Yong-An Chung,1, 2 Jisoo Lee,3 Moon-Seo Park,1
Kyung-Sool Jang,1 Lee-So Maeng,1 Dong-Kyu Jang,1 Ruth Im,1 Yu Jin Jung,1 Soon A. Park,1
Eun-Sun Kang,1 Min-Wook Kim,1 and Young-Min Han1
1 Institute of Catholic Integrative Medicine (ICIM), Incheon St. Mary’s Hospital, The Catholic University of Korea,Incheon 403-720, Republic of Korea
2 Department of Radiology, Incheon St. Mary’s Hospital, The Catholic University of Korea, Incheon 403-720, Republic of Korea3 Chicago Medical School, Rosalind Franklin University of Medicine and Science, North Chicago, IL 60064, USA
Correspondence should be addressed to Yong-An Chung, [email protected]
Received 30 January 2012; Revised 30 March 2012; Accepted 31 March 2012
Academic Editor: Ilkay Erdogan Orhan
Copyright © 2012 Yun-Young Sunwoo et al. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
In this study, we investigated whether gongjin-dan improves functional recovery and has neuroprotective effects on reducingthe infarct volume after transient middle cerebral artery occlusion (MCAo). Infarct volume was measured using TTC stainingand glucose utilization by F-18 FDG PET. Functional improvement was evaluated with the Rota-rod, treadmill, Garcia scoretest, and adhesive removal test. At 14 days after MCAo, neuronal cell survival, astrocytes expansion, and apoptosis were assessedby immunohistofluorescence staining in the peri-infarct region. Also, the expression of neurotrophic factors and inflammatorycytokines such as VEGF, BDNF, Cox-2, TNF-α, IL-1β, and IL-1α was measured in ischemic hemisphere regions. The gongjin-dan-treated group showed both reduced infarct volume and increased glucose utilization. Behavior tests demonstrated a significantimprovement compared to the control. Also in the gongjin-dan treated group, NeuN-positive cells were increased and number ofastrocytes, microglia, and apoptotic cells was significantly decreased compared with the control group in the ischemic peri-infarctarea. Furthermore, the expression of VEGF and BDNF was increased and level of Cox-2, TNF-α, IL-1β, and IL-1α was decreased.These results suggest that gongjin-dan may improve functional outcome through the rapid restoration of metabolism and can beconsidered as a potential neuroprotective agent.
1. Introduction
Ischemic stroke is a second leading cause of death in indus-trialized countries and arguably the most important cause ofacquired disability [1, 2]. The initial pathophysiology due toa reduction or complete blockade of regional cerebral bloodflow (rCBF) results in deficient glucose and oxygen supply tothe affected region [3, 4]. Ischemic stroke can be separatedinto three serial phases following injury: acute (minutes tohours, metabolic stress and excitotoxicity), subacute (hoursto days, inflammation and apoptosis), and chronic (days tomonths, repair and regeneration) [5–7].
In acute stroke, the primary brain insult is largelyattributed to interruption of CBF and breakdown of theblood brain barrier (BBB). The reduction of rCBF causescerebral infarction such as middle cerebral artery occlusion(MCAo). rCBF of ischemic core was decreased 5–20%[8]. Also with rCBF recovery immediately after stroke,damaged cells in the ischemic penumbra can be rescued.Changes in rCBF and metabolism are commonly associatedwith brain disorders, such as traumatic brain injury andischemic stroke [9, 10]. Also, BBB disruption after MCAohas been implicated in hemorrhagic transformation [11].In the normal state, glucose is the predominant substrate

2 Evidence-Based Complementary and Alternative Medicine
for energy metabolism in the brain and has a tightlycontrolled relationship with rCBF. The glucose analogue F-18 FDG (2-[F-18]-fluoro-2-deoxy-D-glucose) is an indicatorof utilization and the degree of neuronal activity in the brain[12]. Furthermore, positron emission tomography (PET)detection of F-18 FDG uptake is therefore valuable in thestudy of stroke, brain injury, and specific function of livinganimals.
In recent studies, a consideration has been given to theindividual active compound alone as well as in combinationwith other herbs [2, 13, 14]. Numerous studies havesought novel neuroprotective substances and the beneficialeffects of herbal medicines against brain ischemia haveoften been reported. This is related by the inhibition ofinflammatory cytokines and microglia activation in brainischemia. In traditional oriental medicine, many herbaldrugs and prescriptions have in fact been used clinicallyfor the treatment of ischemic stroke and vascular dementia.Gongjin-dan is a multiherbal formula available in Koreaand China and is known as an antifatigue and antiagingagent. It has also been used as alternative medicine forthe treatment of various neurodegenerative disorders. Moonet al. reported that learning and memory were enhancedfollowing administration of gongjin-dan in the stress ratmodel [15]. However, experimental evidence clarifying themechanism of gongjin-dan is limited at best.
Herein, we evaluated the effects of gongjin-dan on infarctvolume and functional recovery, as well as on glucosemetabolism, apoptosis and survival of living cells, andchange of inflammatory cytokines and growth factors in anischemic stroke rat model.
2. Methods
2.1. Preparation and Administration of Gongjin-dan. For thisstudy, gongjin-dan was prepared as a mixture of Moschusmoschiferus (175 mg), Corni Fructus (1.5 g), Angelica gigantisradix (1.5 g), and Cervi Parvum Cornu (1.5 g). Gongjin-danwas extracted using reflux extraction equipment in hot waterfor an hour and concentrated using a rotary evaporatorbefore lyophilization. Gongjin-dan was treated with twice(40 mg/kg) per day for 14 days after the stroke.
2.2. Cerebral Ischemic Model. The experimental protocolused in this study was designed in compliance with theguidelines established by the Institutional Animal Care andUse Committee of Catholic University Medical School. Tran-sient MCAo was modeled as described previously [16–18],with slight modification. Adult male Sprague-Dawley rats(270–300 g) were initially anesthetized with 5% isofluranein 70% nitrous oxide and 30% oxygen using an inductionchamber and maintained by a mixture of 2% isofluraneunder temperature controlled conditions (37± 0.1◦C) usinga rectal thermometer and heating pad (Harvard ApparatusInc., Holliston, Massachusetts, USA). The right commoncarotid artery (CCA), external carotid artery (ECA), andinternal carotid artery (ICA) were exposed through a ventralmidline incision. A 4–0 monofilament nylon suture with a
rounded tip was inserted into the CCA lumen and gentlyadvanced into the ICA until it blocked the bifurcating originof the MCA. Cortical blood flow was measured continuouslyvia laser doppler (Transonic system Inc., Ithaca, NY, USA)with a photodetector probe that was stereotaxically placedthrough a burr hole in the skull overlying frontoparietalcortex (1.3 mm posterior, 2.8 mm lateral to the bregma).Ninety minutes later, animals were reanesthetized and thenreperfused by withdrawal of the suture until the tip clearedthe ECA lumen. rCBF was expressed as a percentage of pre-ischemic baseline values. MCAo animals were only includedin the present study if occlusion caused a decrease in rCBF toless than 30% of the original blood flow.
2.3. Behavior Evaluation. Following MCAo, animals wererandomly divided into two experimental groups: a controlsreceived gastric gavage with distilled water (n = 7). The othergroup received gongjin-dan (n = 7). Behavior was assessedwith the Garcia score test, adhesive removal, Rota-rod, andtreadmill test at 1, 3, 7, 10, and 14 days following MCAo byan investigator blinded to the experimental groups. Subjectswere pretrained on all tests for 3 days pre-MCAo, with onlyexception being the Garcia test.
2.3.1. Garcia Score Test. Motor behavior index was scoredblind to rat groupings by the Garcia test [19, 20]. Briefly,subjects were evaluated on six criteria for a possible totalscore ranging from 3 to 18, with a higher score indicatingbetter performance. Motor performance was evaluated onspontaneous activity, symmetry of movements, symmetry offorelimbs, and climbing the wall of wire cage, and sensoryfunction was measured (while reaction to touch on andresponse to vibrissae touch).
2.3.2. Adhesive Removal Test. For adhesive removal tests,square dots of adhesive-backed paper (113.1 mm2) were usedas bilateral tactile stimuli on the radial wrist of both distalforelimbs [21]. The time required to remove both stimulifrom each limb was recorded in five trials/day for 3 days.All animals could remove the dot within 10 sec at the endof training. So, the rats were familiarized with the testingenvironment. Resultant data are presented as the mean timerequired to removal the left dot.
2.3.3. Rota-Rod Test. Subjects were pretrained on the Rota-rod motor test three times/day for 3 days pre-MCAo. TheRota-rod cylinder (Cytec Korea Inc, Seoul, Korea) wasaccelerated from 4 to 40 rpm within 5 minutes. Only animalscapable of remaining on the Rota-rod cylinder for more than160 seconds were included in the study and the cutoff timewas 300 seconds. Data are presented as the mean durationfrom three trials.
2.3.4. Treadmill Test. Subjects were trained on a motor-driven treadmill (Panlab, Barcelona, Spain) for 20 min/day,3 days/week at a speed of 20 m/min prior to MCAo induc-tion. Rats were placed on a moving belt facing away fromthe electrified grid forcing them to run in the direction

Evidence-Based Complementary and Alternative Medicine 3
opposite of belt movement. To avoid foot shocks (with1.0 mA (intensity)), the rats had to move forward. Onlythose which learned to avoid the mild electrical shock wereincluded in this study. The maximum velocity at which therats could run on a motor-driven treadmill was recorded tomeasure 3 times [22, 23].
2.4. rCBF Measurements. rCBF was measured as describedin previous literature [24]. Rats were anesthetized (0.8%to 1.5% isoflurane, 30% O2, remainder N2O) and rectaltemperature was maintained at 37 ± 0.1◦C using a feedbackcontrolled heating system. They were then fixed to astereotaxic apparatus with a probe placed at the level of thedura directly overlying the small burr hole used to induceMCAo. rCBF was measured at the time of MCAo onset andreperfusion. Subsequent measurements were taken during15 min/day over the next 14 days. rCBF was expressed as apercentage of preischemic baseline values. Two investigatorblinded to the experimental groups performed the rCBFrecordings.
2.5. PET Imaging. PET imaging was performed on imagestaken on days 1 and 14 days after MCAo on a PET scannerusing a General Electric discovery STE (Waukesha, WI,USA). Subjects were deprived of food for about 12∼16hours preceding F-18 FDG injection but had access todrinking water at all times. F-18 FDG (1.1 ± 0.04 mCi) wasinjected intravenously into the caudal vein, followed by a30 min uptake period, during which the subjects remainedconscious in a warm environment for optimal brain F-18 FDG uptake, by placing the entire cage on a heatingpad. During brain scanning, anesthesia was maintainedwith katamine (80 mg/kg) and xyzine (10 mg/kg), and bodytemperature was kept at 37◦C with a heating pad on thescanner bed. PET imaging produces 6 coronal image planesseparated by 3 mm in stereotactic apparatus on pinpointmarker.
2.6. PET Imaging Data Analysis. PET imaging data wasanalyzed as per described in previous literature [10]. Toassess changes in metabolism induced by artery occlusion,average regions of interest (ROI) in each hemisphere wereidentified in images of the coronal brain sections. Themean standard uptake value (SUV) of F-18 FDG in ROIwas calculated as the averaged nCi/cc after calibration forboth ipsilateral and contralateral insulated areas in the sameimages. The radioactivity in the contralateral area was usedas a reference to normalize data obtained in the ipsilateralarea, and the differential uptake ratio (DUR) was calculated[10]. The total DUR for each imaging was calculated bysummation of all brain imaging. Also we confirm whetherinfarct volume was associated with elevated DUR for oneday after MCAo (see Figure 1 in Supplementary Materialsavailable online at doi:10.1155/2012/682720).
2.7. Quantitative Analysis of Infarct Volume. At 14 daysafter MCAo, all test subjects (n = 5, for each group)were deeply anesthetized with 15% urethane and sacrificed
by decapitation. The brain was immediately removed andsectioned into four equally spaced (2 mm) coronal blocksusing a rodent brain matrix. These sections were stainedwith 0.1 M PBS containing 2% 2-3-5-triphenylterazolium(TTC; Sigma, St. Louis, MO, USA) for 30 min at 37◦C. Theunstained area was considered to be the infract area. Theinjured area of brain slices was quantified using Meta-Morphimaging software (Molecular Devices Inc, Downingtown, PA,USA). The total infarct volume for each slice was calculatedby summation of all brain slices.
2.8. Immunohistofluorescence Staining. At 14 days afterMCAO, test subjects were sacrificed for histological exam-ination. Rats were deeply anesthetized with 15% urethaneand then perfused transcardially with 0.01 M PBS (pH 7.4),followed by 4% paraformaldehyde in 0.01 M PBS. The brainswere removed and post-fixed for 5 h. The postfixed tissuewas equilibrated with 30% sucrose in 0.1 M phosphatebuffer (pH 7.2) at 4◦C. The brain tissues were embedded,snap-frozen in liquid nitrogen, and stored at −70◦C untiluse. The tissue was cut into 4 μm thick coronal sectionsbetween +0.1 and 0.86 mm of the bregma from each ratfor a total of three blocks. The sections were warmed upfor 20 min and washed with 0.01 M PBS. Sections wereblocked in normal goat serum for 1 h at room tempera-ture. The sections were incubated at 4◦C overnight withthe following antibodies: mouse antiglial fibrillary acidicprotein (GFAP; Millipore, Billerica, MA, USA), mouse anti-neuronal nuclei antigen (NeuN; Millipore), and mouse anti-OX-42 (Millipore). After washing, sections were incubatedin Alexa 488-conjugated goat anti-mouse IgG (Molecularprobe, Eugene, Oregon, USA) for 1 h at room temperature.After washing, the sections were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich). Apopto-sis was detected by the terminal deoxynucleotidyl-transferasemediated d-UTP-biotin nick end (TUNEL) assay using the InSitu Cell Death Detection Kit (Roche, Mannheim, Germany)developed using the Cy3-conjugated streptavidin (JacksonImmunoResearch Laboratories, Bar Harbor, MA, USA).After washing, the sections were counterstained with DAPI,and then observed under both a fluorescence microscopeequipped with a spot digital camera and a confocal scanninglaser microscope (LSM 510, Zeiss, Oberko, Germany). UsingMeta-Morph imaging program (Molecular Devices Inc),ischemic penumbra was determined by optical density ofGFAP and OX-42 and counting of NeuN and TUNEL-positive cells.
2.9. Enzyme-Linked Immunosorbent Analysis. 14 days follow-ing MCAO, rat brains were removed, and coronal sections(200 mg) from −1.0 to 1.0 mm to bregma in the ischemichemisphere were dissected on ice and were stored at −70◦Cuntil use. Subsequently, each tissue sample was suspendedin an equal weight of homogenate buffer and homogenized.The homogenate was centrifuged (10,000 g) for 30 min at4◦C. The supernatant was collected and examined usingthe enzyme-linked immunosorbent assay (ELISA) to detectthe protein levels of neurotrophic factors and inflammatory
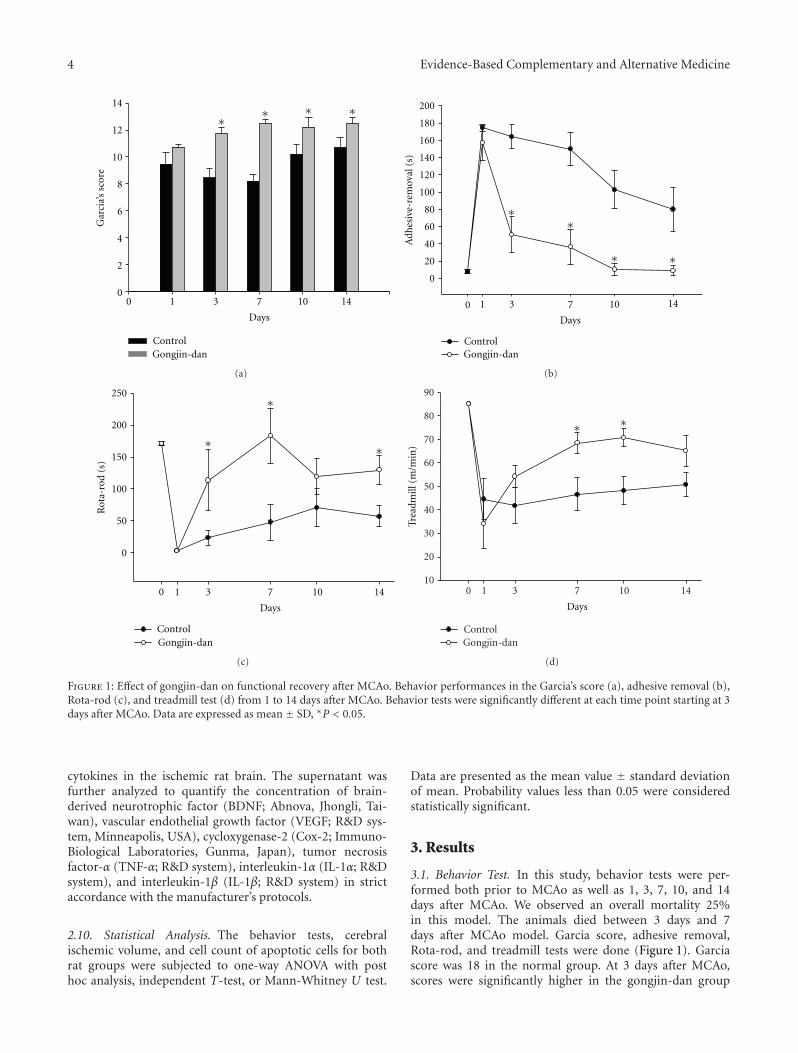
4 Evidence-Based Complementary and Alternative Medicine
0 1 3 7 10 140
2
4
6
8
10
12
14
Control Gongjin-dan
Days
∗ ∗ ∗ ∗G
arci
a’s
scor
e
(a)
0 1 3 7 10 14
0
20
40
60
80
100
120
140
160
180
200
Control Gongjin-dan
Days
Adh
esiv
e-re
mov
al (
s)
∗∗
∗ ∗
(b)
0 1 3 7 10 14
0
50
100
150
200
250
Control Gongjin-dan
Days
Rot
a-ro
d (s
)
∗
∗
∗
(c)
0 1 3 7 10 1410
20
30
40
50
60
70
80
90
Control Gongjin-dan
Days
Trea
dmill
(m
/min
)
∗ ∗
(d)
Figure 1: Effect of gongjin-dan on functional recovery after MCAo. Behavior performances in the Garcia’s score (a), adhesive removal (b),Rota-rod (c), and treadmill test (d) from 1 to 14 days after MCAo. Behavior tests were significantly different at each time point starting at 3days after MCAo. Data are expressed as mean ± SD, ∗P < 0.05.
cytokines in the ischemic rat brain. The supernatant wasfurther analyzed to quantify the concentration of brain-derived neurotrophic factor (BDNF; Abnova, Jhongli, Tai-wan), vascular endothelial growth factor (VEGF; R&D sys-tem, Minneapolis, USA), cycloxygenase-2 (Cox-2; Immuno-Biological Laboratories, Gunma, Japan), tumor necrosisfactor-α (TNF-α; R&D system), interleukin-1α (IL-1α; R&Dsystem), and interleukin-1β (IL-1β; R&D system) in strictaccordance with the manufacturer’s protocols.
2.10. Statistical Analysis. The behavior tests, cerebralischemic volume, and cell count of apoptotic cells for bothrat groups were subjected to one-way ANOVA with posthoc analysis, independent T-test, or Mann-Whitney U test.
Data are presented as the mean value ± standard deviationof mean. Probability values less than 0.05 were consideredstatistically significant.
3. Results
3.1. Behavior Test. In this study, behavior tests were per-formed both prior to MCAo as well as 1, 3, 7, 10, and 14days after MCAo. We observed an overall mortality 25%in this model. The animals died between 3 days and 7days after MCAo model. Garcia score, adhesive removal,Rota-rod, and treadmill tests were done (Figure 1). Garciascore was 18 in the normal group. At 3 days after MCAo,scores were significantly higher in the gongjin-dan group

Evidence-Based Complementary and Alternative Medicine 5
compared to the control group (11.75± 0.5 versus 8.5± 0.6,P < 0.05). Similarly, significant differences between the twogroups were also noted in the 14 days (12.5 ± 0.5 versus10.75± 0.7, P < 0.05). Performance on the adhesive removaltest was also compared between both groups. Gongjin-dan-treated group are significantly different 3 days after MCAocompared with control group (50.6±20.5 versus 163.9±13.9,P < 0.05) (Figure 1(b)). Although no difference in Rota-rod assessment was noted on day 1 after MCAo, significantbehavior recovery was demonstrated in the gongjin-dan-treated group at 3 days after MCAo (23.7 ± 12 versus113.75 ± 47, P < 0.05) (Figure 1(c)). The maximum speedat which the rats could run on a motor driven treadmillwas recorded. Three days after MCAo, the maximum velocityon the treadmill test indicated its maximum deficit. Therewas no significant difference among the groups. At 7 daysafter MCAo, the gongjin-dan treated group showed a greaterfunctional recovery than the control (68.4±4.5 versus 46.6±7, P < 0.05) (Figure 1(d)). Maximum velocity identified thegreatest velocity at 10 days after MCAo. At 14 days afterMCAo, maximum velocity was low compared with velocityat 10 days. This may be due to the weight variation betweendifferent experiments (data not shown). These results suggestthat gongjin-dan can promote the restoration of motorfunction in rat stroke model.
3.2. Infarct Volume and PET-CT. The neuroprotective effectof gongjin-dan was evaluated by measuring the infarctvolume at 14 days after MCAo. The brain of rats was stainedwith TTC to obtain the infarct volume and was calculatedby measuring the area of infarct. Figure 2 is a photographtypical of TTC-stained sections of treatment and controlgroups. As seen in Figure 2, infarct volume of the gongjin-dan group was significantly decreased compared to the con-trol (108.8±42 versus 185±43 mm3, P < 0.05) (Figure 2(d)).PET-CT has long been demonstrated as a marker of glucosemetabolism in ischemia and infarction [25, 26]. In ourstudies, PET imaging data was interpreted in terms of DUR.With the contralateral side as baseline, the metabolic changeof transient MCAo in an ipsilateral area was calculated byROI value. As seen in Figure 2(a), the quantification of theROI radioactivity was decreased in glucose uptake followingMCAo. From these control data, a DUR in the ischemicregion was significantly increased compared with normal. ADUR of gongjin-dan treatment was decreased compared tothe control (11.6±3.2 versus 17.4±7.9%, P < 0.05). This datasuggests that protected ischemic area by gongjin-dan coulduptake glucose metabolism.
3.3. Restoration of rCBF. To investigate whether gongjin-dan treatment improved rCBF, the rCBF was measured bylaser doppler. Baseline rCBF recorded just prior to MCAosurgery was applied to express values as a percentage.Following reperfusion, rCBF was maintained at 60%. 2 daysafter MCAo, the gongjin-dan treatment group improvedto near-normal rCBF level. In contrast, rCBF value in thecontrol group for 14 days was similar to the rCBF value ofreperfusion. This suggests that gongjin-dan could potentially
halt the pathological cascade of stroke. Also, it is likelythat gongjin-dan promotes behavior recovery via rCBF andglucose metabolism restoration, thereafter blocking neurosis.
3.4. Activating Astrocyte, Microglia, and Neuronal Death. Weobserved the number of astrocytes, microglia, and neuronsin the peri-infarct regions at 14 days after MCAo. In gongjin-dan group, GFAP-positive cells and OX-42-positive cellsshowed significant decrease in the optical density comparedto the control group (Figures 3(g) and 3(h)). Also NeuN-positive cells showed higher compared to the control group(Figure 3(i)). These results suggest that the inhibitory effectsof GFAP and microglial activation may contribute to itsneuroprotective effects in MCAo, thereby enabling a numberof NeuN-positive cells to survive in peri-infarct region.
3.5. Apoptotic Cell Death. We observed the number ofapoptotic cells in the peri-infarct regions at 14 days afterMCAo model. Apoptotic cells were recognized by theidentification of DNA fragmentation using the TUNELmethod. Gongjin-dan-treated groups had reduced TUNEL-positive cells compared to the control group (108±14 versus148 ± 41, P < 0.05) (Figure 4(e)). These results suggest thatthe gongjin-dan-treated group effectively prevents expansionapoptotic cell death in MCAo model.
3.6. Quantitative Analysis of Neurotrophic Factors and Inflam-matory Cytokines. We hypothesized that gongjin-dan mayprotect an injured area following a stroke by elevating neu-rotrophic factors and by reducing inflammatory cytokines.We used the ELISA to detect protein levels of neurotrophicfactors and inflammatory cytokines in the ischemic rat brain.At 14 days after MCAo, expression of BDNF and VEGF wassignificantly high compared to the control group (Figures5(a) and 5(b)). Also inflammatory cytokines such as Cox-2, IL-1α, IL-1β, and TNF-α were reduced as compared withcontrol group (Figures 5(c), 5(d), 5(e), and 5(f)). Thesedata suggest that an increment of neurotrophic factors anda decrease of inflammatory cytokines by gongjin-dan couldprotect cerebral infarction and promote behavior functionrecovery.
4. Discussion
During stroke, CBF is reduced and causes a decrease ofoxygen supply to the brain and an increase of glutamate,inflammatory mediators, and free radicals. As a result,cellular necrosis and delayed cell death occur. In this study,we examined the effect of the gongjin-dan treatment ontransient MCAo model. We investigated (1) whether theinfarct volume decreases, (2) whether blood circulation andglucose metabolism improve (laser Doppler and PET-CTwere performed for measurement), (3) whether there isfunctional recovery, and (4) whether neuronal cells in peri-infarct region can be protected from death.
We performed TTC staining in MCAo model to iden-tify the infarct volume. The gongjin-dan-treated groupshowed a more diminished infarct volume as compared
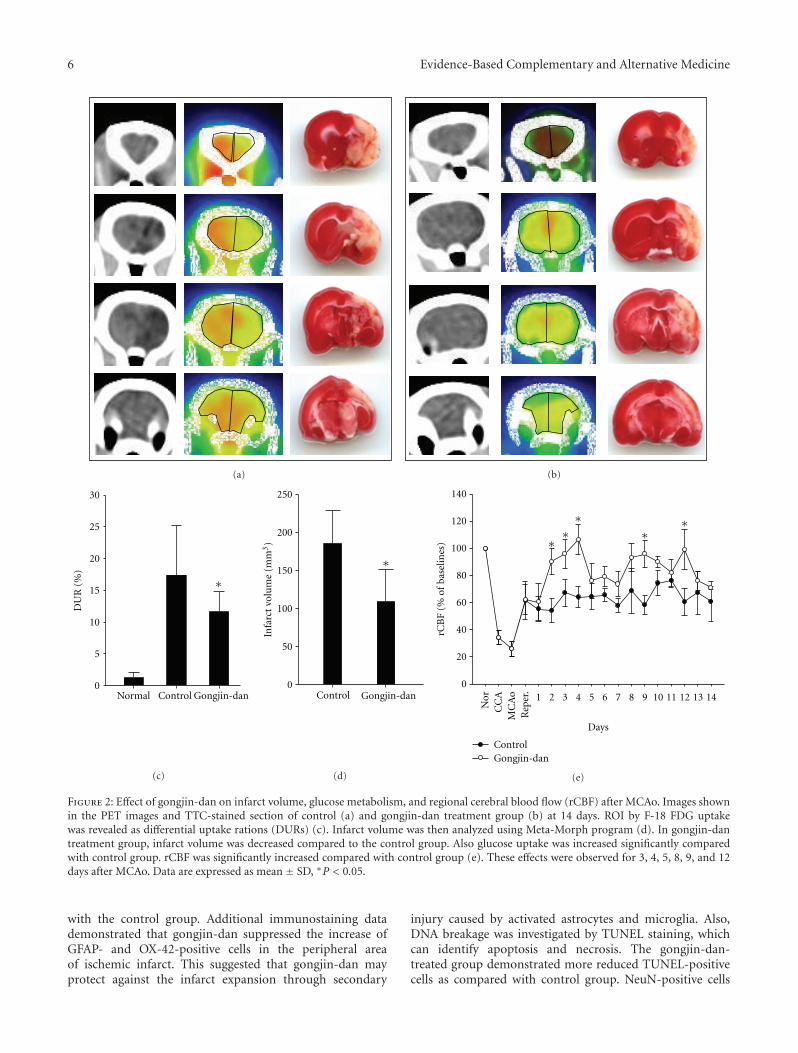
6 Evidence-Based Complementary and Alternative Medicine
(a) (b)
Normal Control Gongjin-dan
DU
R (
%)
0
5
10
15
20
25
30
∗
(c)
Control Gongjin-dan0
50
100
150
200
250
Infa
rct
volu
me
(mm
3)
∗
(d)
∗∗
∗∗
∗
Days
rCB
F (%
of
base
lines
)
0
20
40
60
80
100
120
140
ControlGongjin-dan
Nor
CC
AM
CA
oR
eper
.
1 2 3 4 5 6 7 8 9 10 11 12 13 14
(e)
Figure 2: Effect of gongjin-dan on infarct volume, glucose metabolism, and regional cerebral blood flow (rCBF) after MCAo. Images shownin the PET images and TTC-stained section of control (a) and gongjin-dan treatment group (b) at 14 days. ROI by F-18 FDG uptakewas revealed as differential uptake rations (DURs) (c). Infarct volume was then analyzed using Meta-Morph program (d). In gongjin-dantreatment group, infarct volume was decreased compared to the control group. Also glucose uptake was increased significantly comparedwith control group. rCBF was significantly increased compared with control group (e). These effects were observed for 3, 4, 5, 8, 9, and 12days after MCAo. Data are expressed as mean ± SD, ∗P < 0.05.
with the control group. Additional immunostaining datademonstrated that gongjin-dan suppressed the increase ofGFAP- and OX-42-positive cells in the peripheral areaof ischemic infarct. This suggested that gongjin-dan mayprotect against the infarct expansion through secondary
injury caused by activated astrocytes and microglia. Also,DNA breakage was investigated by TUNEL staining, whichcan identify apoptosis and necrosis. The gongjin-dan-treated group demonstrated more reduced TUNEL-positivecells as compared with control group. NeuN-positive cells
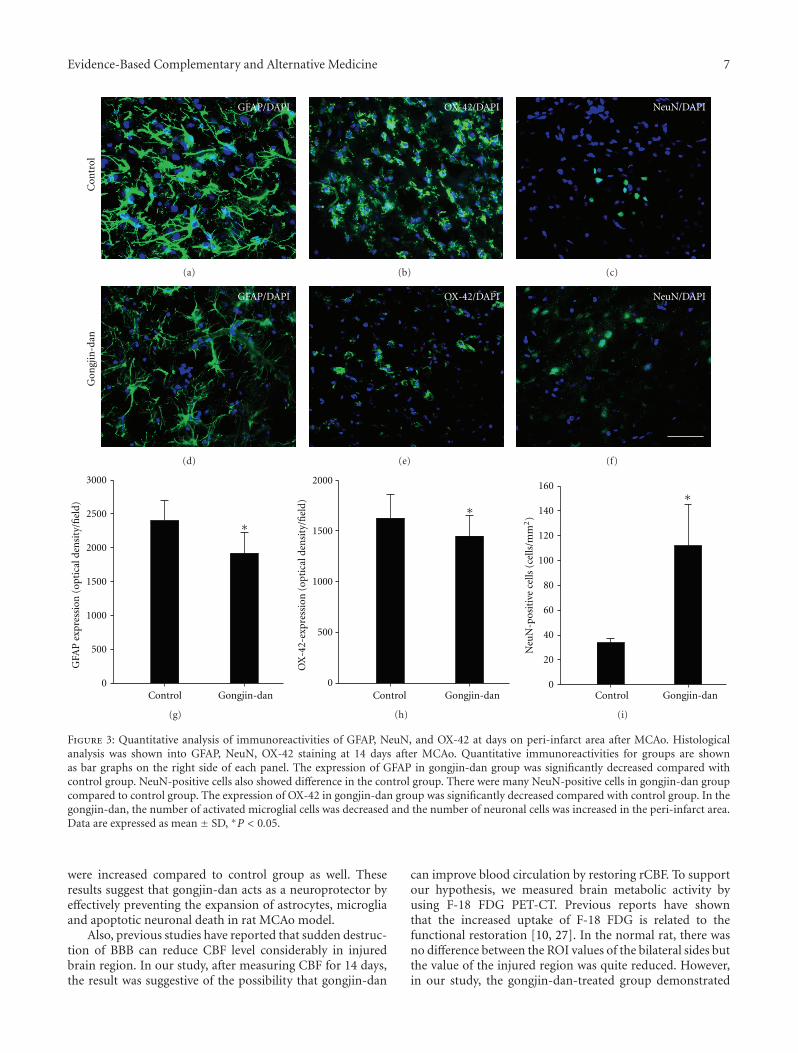
Evidence-Based Complementary and Alternative Medicine 7
GFAP/DAPI
Con
trol
(a)
OX-42/DAPI
(b)
NeuN/DAPI
(c)
GFAP/DAPI
Gon
gjin
-dan
(d)
OX-42/DAPI
(e)
NeuN/DAPI
(f)
Control Gongjin-dan
GFA
P e
xpre
ssio
n (
opti
cal d
ensi
ty/fi
eld)
0
500
1000
1500
2000
2500
3000
∗
(g)
Control Gongjin-dan
OX
-42-
expr
essi
on (
opti
cal d
ensi
ty/fi
eld)
0
500
1000
1500
2000
∗
(h)
Control Gongjin-dan0
20
40
60
80
100
120
140
160
Neu
N-p
osit
ive
cells
(ce
lls/m
m2)
∗
(i)
Figure 3: Quantitative analysis of immunoreactivities of GFAP, NeuN, and OX-42 at days on peri-infarct area after MCAo. Histologicalanalysis was shown into GFAP, NeuN, OX-42 staining at 14 days after MCAo. Quantitative immunoreactivities for groups are shownas bar graphs on the right side of each panel. The expression of GFAP in gongjin-dan group was significantly decreased compared withcontrol group. NeuN-positive cells also showed difference in the control group. There were many NeuN-positive cells in gongjin-dan groupcompared to control group. The expression of OX-42 in gongjin-dan group was significantly decreased compared with control group. In thegongjin-dan, the number of activated microglial cells was decreased and the number of neuronal cells was increased in the peri-infarct area.Data are expressed as mean ± SD, ∗P < 0.05.
were increased compared to control group as well. Theseresults suggest that gongjin-dan acts as a neuroprotector byeffectively preventing the expansion of astrocytes, microgliaand apoptotic neuronal death in rat MCAo model.
Also, previous studies have reported that sudden destruc-tion of BBB can reduce CBF level considerably in injuredbrain region. In our study, after measuring CBF for 14 days,the result was suggestive of the possibility that gongjin-dan
can improve blood circulation by restoring rCBF. To supportour hypothesis, we measured brain metabolic activity byusing F-18 FDG PET-CT. Previous reports have shownthat the increased uptake of F-18 FDG is related to thefunctional restoration [10, 27]. In the normal rat, there wasno difference between the ROI values of the bilateral sides butthe value of the injured region was quite reduced. However,in our study, the gongjin-dan-treated group demonstrated
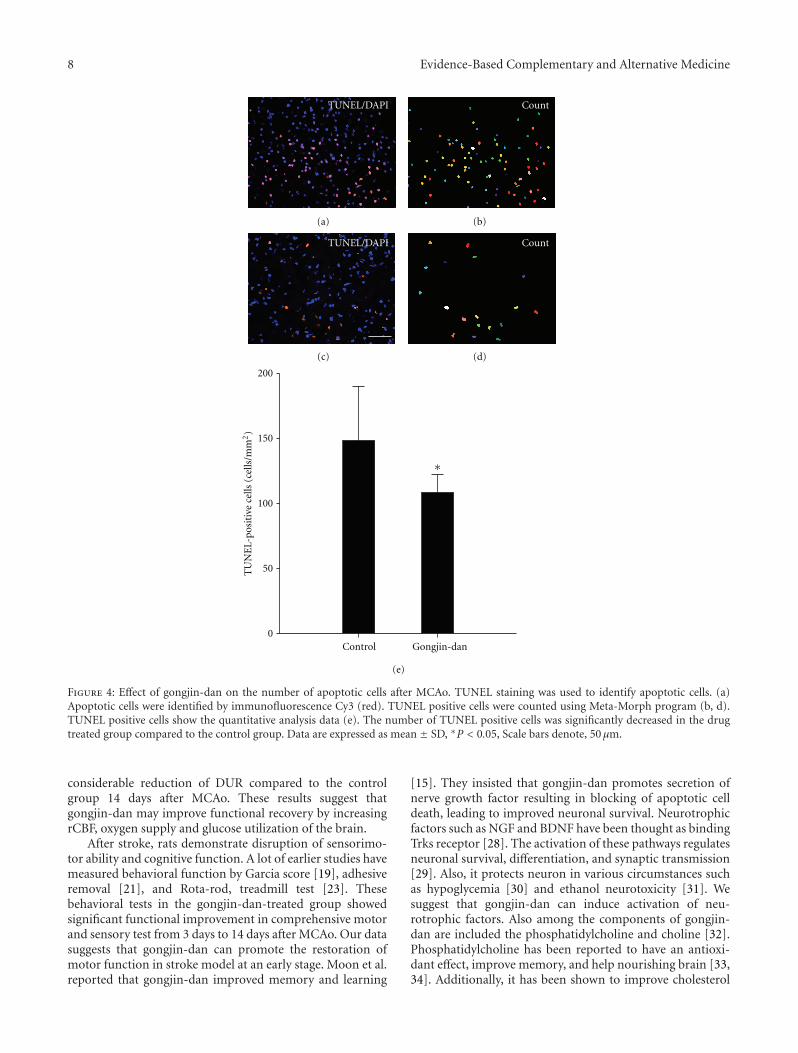
8 Evidence-Based Complementary and Alternative Medicine
TUNEL/DAPI Count
(a) (b)
(c) (d)
TUNEL/DAPI Count
Control Gongjin-dan0
50
100
150
200
TU
NE
L-po
siti
ve c
ells
(ce
lls/m
m2)
(e)
∗
Figure 4: Effect of gongjin-dan on the number of apoptotic cells after MCAo. TUNEL staining was used to identify apoptotic cells. (a)Apoptotic cells were identified by immunofluorescence Cy3 (red). TUNEL positive cells were counted using Meta-Morph program (b, d).TUNEL positive cells show the quantitative analysis data (e). The number of TUNEL positive cells was significantly decreased in the drugtreated group compared to the control group. Data are expressed as mean ± SD, ∗P < 0.05, Scale bars denote, 50 μm.
considerable reduction of DUR compared to the controlgroup 14 days after MCAo. These results suggest thatgongjin-dan may improve functional recovery by increasingrCBF, oxygen supply and glucose utilization of the brain.
After stroke, rats demonstrate disruption of sensorimo-tor ability and cognitive function. A lot of earlier studies havemeasured behavioral function by Garcia score [19], adhesiveremoval [21], and Rota-rod, treadmill test [23]. Thesebehavioral tests in the gongjin-dan-treated group showedsignificant functional improvement in comprehensive motorand sensory test from 3 days to 14 days after MCAo. Our datasuggests that gongjin-dan can promote the restoration ofmotor function in stroke model at an early stage. Moon et al.reported that gongjin-dan improved memory and learning
[15]. They insisted that gongjin-dan promotes secretion ofnerve growth factor resulting in blocking of apoptotic celldeath, leading to improved neuronal survival. Neurotrophicfactors such as NGF and BDNF have been thought as bindingTrks receptor [28]. The activation of these pathways regulatesneuronal survival, differentiation, and synaptic transmission[29]. Also, it protects neuron in various circumstances suchas hypoglycemia [30] and ethanol neurotoxicity [31]. Wesuggest that gongjin-dan can induce activation of neu-rotrophic factors. Also among the components of gongjin-dan are included the phosphatidylcholine and choline [32].Phosphatidylcholine has been reported to have an antioxi-dant effect, improve memory, and help nourishing brain [33,34]. Additionally, it has been shown to improve cholesterol
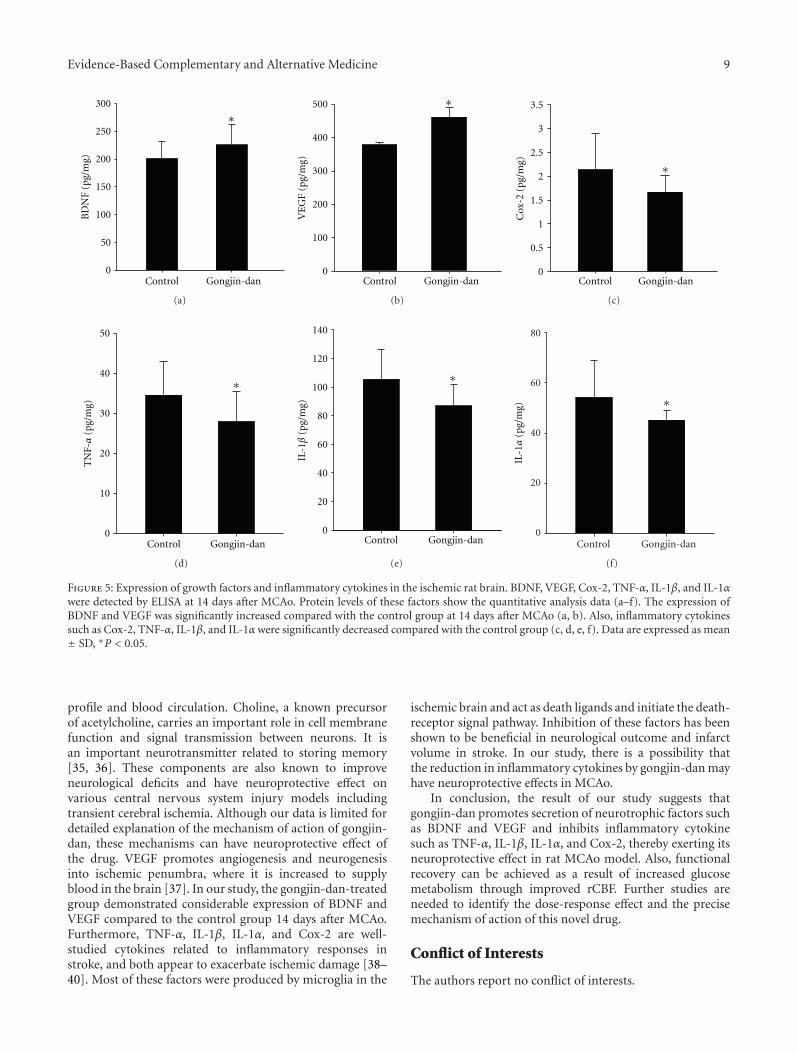
Evidence-Based Complementary and Alternative Medicine 9
∗
Control Gongjin-dan
BD
NF
(pg/
mg)
0
50
100
150
200
250
300
(a)
∗
Control Gongjin-dan0
100
200
300
400
500
VE
GF
(pg/
mg)
(b)
Control Gongjin-dan
Cox
-2 (
pg/m
g)
0
0.5
1
1.5
2
2.5
3
3.5
∗
(c)
Control Gongjin-dan0
10
20
30
40
50
TN
F-α
(pg/
mg)
∗
(d)
Control Gongjin-dan0
20
40
60
80
100
120
140
IL-1β
(pg/
mg)
∗
(e)
Control Gongjin-dan0
20
40
60
80
IL-1α
(pg/
mg) ∗
(f)
Figure 5: Expression of growth factors and inflammatory cytokines in the ischemic rat brain. BDNF, VEGF, Cox-2, TNF-α, IL-1β, and IL-1αwere detected by ELISA at 14 days after MCAo. Protein levels of these factors show the quantitative analysis data (a–f). The expression ofBDNF and VEGF was significantly increased compared with the control group at 14 days after MCAo (a, b). Also, inflammatory cytokinessuch as Cox-2, TNF-α, IL-1β, and IL-1αwere significantly decreased compared with the control group (c, d, e, f). Data are expressed as mean± SD, ∗P < 0.05.
profile and blood circulation. Choline, a known precursorof acetylcholine, carries an important role in cell membranefunction and signal transmission between neurons. It isan important neurotransmitter related to storing memory[35, 36]. These components are also known to improveneurological deficits and have neuroprotective effect onvarious central nervous system injury models includingtransient cerebral ischemia. Although our data is limited fordetailed explanation of the mechanism of action of gongjin-dan, these mechanisms can have neuroprotective effect ofthe drug. VEGF promotes angiogenesis and neurogenesisinto ischemic penumbra, where it is increased to supplyblood in the brain [37]. In our study, the gongjin-dan-treatedgroup demonstrated considerable expression of BDNF andVEGF compared to the control group 14 days after MCAo.Furthermore, TNF-α, IL-1β, IL-1α, and Cox-2 are well-studied cytokines related to inflammatory responses instroke, and both appear to exacerbate ischemic damage [38–40]. Most of these factors were produced by microglia in the
ischemic brain and act as death ligands and initiate the death-receptor signal pathway. Inhibition of these factors has beenshown to be beneficial in neurological outcome and infarctvolume in stroke. In our study, there is a possibility thatthe reduction in inflammatory cytokines by gongjin-dan mayhave neuroprotective effects in MCAo.
In conclusion, the result of our study suggests thatgongjin-dan promotes secretion of neurotrophic factors suchas BDNF and VEGF and inhibits inflammatory cytokinesuch as TNF-α, IL-1β, IL-1α, and Cox-2, thereby exerting itsneuroprotective effect in rat MCAo model. Also, functionalrecovery can be achieved as a result of increased glucosemetabolism through improved rCBF. Further studies areneeded to identify the dose-response effect and the precisemechanism of action of this novel drug.
Conflict of Interests
The authors report no conflict of interests.

10 Evidence-Based Complementary and Alternative Medicine
Authors’ Contribution
Y.-Y. Sunwoo and S. I. Park are equally contributed to thiswork as joint first authors.
Acknowledgment
The research was supported by Grant 2011K000709 from theConverging Research Center Program through the Ministryof Education, Science, and Technology.
References
[1] W. E. Connolly, “Suffering, justice, and the politics ofbecoming,” Culture, Medicine and Psychiatry, vol. 20, no. 3, pp.251–277, 1996.
[2] X. Q. Zhou, X. N. Zeng, H. Kong, and X. L. Sun, “Neuropro-tective effects of berberine on stroke models in vitro and invivo,” Neuroscience Letters, vol. 447, no. 1, pp. 31–36, 2008.
[3] K. P. Loh, S. H. Huang, B. K. H. Tan, and Y. Z. Zhu, “Cerebralprotection of purified Herba Leonuri extract on middlecerebral artery occluded rats,” Journal of Ethnopharmacology,vol. 125, no. 2, pp. 337–343, 2009.
[4] D. Zemke, J. L. Smith, M. J. Reeves, and A. Majid, “Ischemiaand ischemic tolerance in the brain: an overview,” NeuroToxi-cology, vol. 25, no. 6, pp. 895–904, 2004.
[5] T. Yoshikawa, Y. Akiyoshi, T. Susumu et al., “GinsenosideRb1 reduces neurodegeneration in the peri-infarct area ofa thromboembolic stroke model in non-human primates,”Journal of Pharmacological Sciences, vol. 107, no. 1, pp. 32–40,2008.
[6] U. Dirnagl, R. P. Simon, and J. M. Hallenbeck, “Ischemictolerance and endogenous neuroprotection,” Trends in Neu-rosciences, vol. 26, no. 5, pp. 248–254, 2003.
[7] S. C. Fagan, D. C. Hess, E. J. Hohnadel, D. M. Pollock, andA. Ergul, “Targets for vascular protection after acute ischemicstroke,” Stroke, vol. 35, no. 9, pp. 2220–2225, 2004.
[8] M. D. Ginsberg, “Adventures in the pathophysiology of brainischemia: penumbra, gene expression, neuroprotection: the2002 Thomas Willis lecture,” Stroke, vol. 34, no. 1, pp. 214–223, 2003.
[9] M. Bergsneider, “Evolving concepts of cerebrospinal fluidphysiology,” Neurosurgery Clinics of North America, vol. 12, no.4, pp. 631–638, 2001.
[10] Y. K. Fu, C. J. Chang, K. Y. Chen et al., “Imaging of regionalmetabolic activity by 18F-FDG/PET in rats with transientcerebral ischemia,” Applied Radiation and Isotopes, vol. 67, no.10, pp. 1743–1747, 2009.
[11] R. M. Dijkhuizen, M. Asahi, O. Wu, B. R. Rosen, and E. H. Lo,“Rapid breakdown of microvascular barriers and subsequenthemorrhagic transformation after delayed recombinant tissueplasminogen activator treatment in a rat embolic strokemodel,” Stroke, vol. 33, no. 8, pp. 2100–2104, 2002.
[12] F. Gao, S. Wang, Y. Guo et al., “Protective effects of repetitivetranscranial magnetic stimulation in a rat model of transientcerebral ischaemia: a microPET study,” European Journal ofNuclear Medicine and Molecular Imaging, vol. 37, no. 5, pp.954–961, 2010.
[13] F. Pu, K. Mishima, N. Egashira et al., “Post-ischemic treatmentwith Toki-shakuyaku-san (Tang-Gui-Shao-Yao-San) preventsthe impairment of spatial memory induced by repeated cere-bral ischemia in rats,” American Journal of Chinese Medicine,vol. 33, no. 3, pp. 475–489, 2005.
[14] Y. J. Yun, B. Lee, D. H. Hahm et al., “Neuroprotective effectof Palmul-Chongmyeong-Tang on ischemia-induced learningand memory deficits in the rat,” Biological and PharmaceuticalBulletin, vol. 30, no. 2, pp. 337–342, 2007.
[15] E. Moon, Y. Her, J. B. Lee et al., “The multi-herbal medicineGongjin-dan enhances memory and learning tasks via NGFregulation,” Neuroscience Letters, vol. 466, no. 3, pp. 114–119,2009.
[16] E. Z. Longa, P. R. Weinstein, S. Carlson, and R. Cummins,“Reversible middle cerebral artery occlusion without craniec-tomy in rats,” Stroke, vol. 20, no. 1, pp. 84–91, 1989.
[17] K. S. Jang, S. I. Park, D. K. Jang et al., “Effect of combinationtherapy with sodium ozagrel and panax ginseng on transientcerebral ischemia model in rats,” Journal of Biomedicine andBiotechnology, vol. 2010, Article ID 893401, 8 pages, 2010.
[18] D. K. Jang, S. I. Park, Y. M. Han et al., “Motor-evoked potentialconfirmation of functional improvement by transplantedbone marrow mesenchymal stem cell in the ischemic ratbrain,” Journal of Biomedicine and Biotechnology, vol. 2011,Article ID 238409, 11 pages, 2011.
[19] J. H. Garcia, S. Wagner, K. F. Liu, X. J. Hu, and J. P.Mohr, “Neurological deficit and extent of neuronal necrosisattributable to middle cerebral artery occlusion in rats:statistical validation,” Stroke, vol. 26, no. 4, pp. 627–635, 1995.
[20] M. W. Kim, M. S. Bang, T. R. Han et al., “Exercise increasedBDNF and trkB in the contralateral hemisphere of theischemic rat brain,” Brain Research, vol. 1052, no. 1, pp. 16–21, 2005.
[21] J. Chen, Z. G. Zhang, Y. Li et al., “Intravenous administrationof human bone marrow stromal cells induces angiogenesis inthe ischemic boundary zone after stroke in rats,” CirculationResearch, vol. 92, no. 6, pp. 692–699, 2003.
[22] K. Komatsu, O. Honmou, J. Suzuki, K. Houkin, H. Hamada,and J. D. Kocsis, “Therapeutic time window of mesenchymalstem cells derived from bone marrow after cerebral ischemia,”Brain Research, vol. 1334, pp. 84–92, 2010.
[23] H. C. Chang, Y. R. Yang, S. G. P. Wang, and R. Y. Wang, “Effectsof treadmill training on motor performance and extracellularglutamate level in striatum in rats with or without transientmiddle cerebral artery occlusion,” Behavioural Brain Research,vol. 205, no. 2, pp. 450–455, 2009.
[24] C. V. Borlongan, J. G. Lind, O. Dillon-Carter et al., “Bonemarrow grafts restore cerebral blood flow and blood brainbarrier in stroke rats,” Brain Research, vol. 1010, no. 1-2, pp.108–116, 2004.
[25] M. Bergsneider, D. A. Hovda, D. L. McArthur et al., “Metabolicrecovery following human traumatic brain injury basedon FDG-PET: time course and relationship to neurologicaldisability,” Journal of Head Trauma Rehabilitation, vol. 16, no.2, pp. 135–148, 2001.
[26] N. Marklund, S. Sihver, B. Langstrom, M. Bergstrom, and L.Hillered, “Effect of traumatic brain injury and nitrone radicalscavengers on relative changes in regional cerebral blood flowand glucose uptake in rats,” Journal of Neurotrauma, vol. 19,no. 10, pp. 1139–1153, 2002.
[27] A. H. Moore, C. L. Osteen, A. F. Chatziioannou, D. A. Hovda,and S. R. Cherry, “Quantitative assessment of longitudinalmetabolic changes in vivo after traumatic brain injury in theadult rat using FDG-microPET,” Journal of Cerebral Blood Flowand Metabolism, vol. 20, no. 10, pp. 1492–1501, 2000.
[28] E. J. Huang and L. F. Reichardt, “Trk receptors: roles inneuronal signal transduction,” Annual Review of Biochemistry,vol. 72, pp. 609–642, 2003.

Evidence-Based Complementary and Alternative Medicine 11
[29] T. Beck, D. Lindholm, E. Castren, and A. Wree, “Brain-derivedneurotrophic factor protects against ischemic cell damagein rat hippocampus,” Journal of Cerebral Blood Flow andMetabolism, vol. 14, no. 4, pp. 689–692, 1994.
[30] B. Cheng and M. P. Mattson, “NGF and bFGF protect rat hip-pocampal and human cortical neurons against hypoglycemicdamage by stabilizing calcium homeostasis,” Neuron, vol. 7,no. 6, pp. 1031–1041, 1991.
[31] D. W. Walker, D. E. Barnes, and S. F. Zornetzer, “Neuronal lossin hippocampus induced by prolonged ethanol consumptionin rats,” Science, vol. 209, no. 4457, pp. 711–713, 1980.
[32] K. Yamasaki, M. Kikuoka, H. Nishi et al., “Contents of lecithinand choline in crude drugs,” Chemical and PharmaceuticalBulletin, vol. 42, no. 1, pp. 105–107, 1994.
[33] H. S. Levin and B. H. Peters, “Long-term administrationof oral physostigmine and lecithin improve memory inAlzheimer’s disease,” Annals of Neurology, vol. 15, no. 2, p. 210,1984.
[34] R. G. Walton, “Lecithin and physostigmine for posttraumaticmemory and cognitive deficits,” Psychosomatics, vol. 23, no. 4,pp. 435–436, 1982.
[35] M. E. Hasselmo, “The role of acetylcholine in learning andmemory,” Current Opinion in Neurobiology, vol. 16, no. 6, pp.710–715, 2006.
[36] T. Watanabe, N. Yamagata, K. Takasaki et al., “Decreasedacetylcholine release is correlated to memory impairment inthe Tg2576 transgenic mouse model of Alzheimer’s disease,”Brain Research, vol. 1249, no. C, pp. 222–228, 2009.
[37] Y. C. Gao, H. K. Lu, Q. Y. Luo, L. B. Chen, Y. Ding,and R. S. Zhu, “Comparison of free plasma metanephrinesenzyme immunoassay with 131I-MIBG scan in diagnosis ofpheochromocytoma,” Clinical and Experimental Medicine, vol.8, no. 2, pp. 87–91, 2008.
[38] Y. Yamasaki, N. Matsuura, H. Shozuhara et al., “Interleukin-1as a pathogenetic mediator of ischemic brain damage in rats,”Stroke, vol. 26, no. 4, pp. 676–681, 1995.
[39] S. J. Chan, W. S. F. Wong, P. T. H. Wong, and J. S.Bian, “Neuroprotective effects of andrographolide in a ratmodel of permanent cerebral ischaemia,” British Journal ofPharmacology, vol. 161, no. 3, pp. 668–679, 2010.
[40] C. Iadecola, K. Niwa, S. Nogawa et al., “Reduced susceptibilityto ischemic brain injury and N-methyl-D-aspartate-mediatedneurotoxicity in cyclooxygenase-2-deficient mice,” Proceedingsof the National Academy of Sciences of the United States ofAmerica, vol. 98, no. 3, pp. 1294–1299, 2001.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 701289, 7 pagesdoi:10.1155/2012/701289
Research Article
Anxiolytic-Like Effects of Compound Zhi Zhu Xiang in Rats
Yan-Li Wang,1 Jin-Li Shi,1 Liu Yong,1 Zhao Ren,2 Yu-Jing Zhai,1 and Jian-You Guo3
1 School of Chinese Materia Medica, Beijing University of Chinese Medicine, Beijing 100102, China2 Pharmaceutical Factory, Yunnan Institute of Material Medical, Yunnan 650111, China3 Key Laboratory of Mental Health, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China
Correspondence should be addressed to Jin-Li Shi, [email protected] and Jian-You Guo, [email protected]
Received 9 December 2011; Accepted 23 March 2012
Academic Editor: Ilkay Erdogan Orhan
Copyright © 2012 Yan-Li Wang et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
The purpose of this study was to determine whether compound zhi zhu xiang (CZZX) exerts anxiolytic-like effects in rats. Theanimals were orally administered CZZX (0.75, 1.5, and 3 g/kg daily) for 10 days and tested in the elevated plus maze (EPM), Vogelconflict test (VCT), and open field. Repeated treatment with CZZX (3 g/kg/day, p.o.) significantly increased the percentage ofboth entries into and time spent on the open arms of the EPM compared with saline controls. In the VCT, repeated treatment withCZZX (1.5 and 3 g/kg/day, p.o.) significantly increased the number of punished licks. The drug did not change the total entries intothe open arms of the EPM or interfere with water consumption or nociceptive threshold, discarding potential confounding factorsin the two tests. In the open field, locomotion was not reduced, discarding the possible sedative effect of CZZX. In the bindingassay, the binding of [3H] Ro 15-1788 (flumazenil) to the benzodiazepine binding site in washed crude synaptosomal membranesfrom rat cerebral cortex was affected by CZZX. These data indicate an anxiolytic-like profile of action for CZZX without sedativeside effects, and this activity may be mediated by benzodiazepine binding site modulation at γ-aminobutyric acid-A receptors.
1. Introduction
Anxiety is a widespread incapacitating psychiatric ailmentthat imposes a substantial health burden on society. Ben-zodiazepines (BZDs) are considered safe drugs and widelyprescribed for their anxiolytic, muscle relaxant, sedative-hypnotic, and anticonvulsant actions [1]. However, they mayproduce side effects, such as sedation and myorelaxation,that are considered unwanted effects of anxiolytic drugs[2]. Therefore, the search for safe and effective agents hascontinued.
Over the past decade, herbal medicines have receivedincreasing attention from the psychiatry research commu-nity. This is largely because many herbal preparations havebeen found to have beneficial effects for various psychiatricconditions in experimental animal and clinical studies [3–6].In Western societies, natural remedies for anxiety disorders,such as valerian (Valeriana officinalis), St. John’s wort (Hyper-icum perforatum), passion flower (Passiflora incarnata), hops(Humulus lupulus), and kava kava (Piper methysticum),are readily available [7]. Several herbal mixtures also haveanxiolytic-like effects in rodent models of anxiety (e.g.,
Yokukansan [8] and Xiao-Tan-Jie-Yu-Fang [9]). Moreover,substantial evidence from our laboratories indicates thatherbal medicines are beneficial in preventing mental illness,such as vanadium-enriched Cordyceps sinensis [5], gan maida zao decoction [10], and Sini tang [6].
Compound zhi zhu xiang (CZZX) originated froma clinical experimental prescription, and a clinical studyshowed that it is an effective and well-tolerated antianxietyprescription [11]. Among the constituents of CZZX, Vale-riana jatamansi Jones accounts for the largest proportionand plays the major therapeutic role. It acts as an “emperorherb” according to traditional Chinese medicine formulationtheory. The plant in India is widely known for its use inthe treatment of anxiety, insomnia, epilepsy, failing reflexes,hysteria, neurosis, and sciatica [12]. In some countries, it isused as an important sedative in herbal medicine. Moreover,large-scale studies also demonstrated that extracts of Valeri-ana officinalis L. s.l. exert anxiolytic effects [13, 14]. Ziziphusjujuba Mill. and Albizia julibrissin Durazz. play auxiliary rolesas “minister herbs” in the CZZX formulation because of theireffects on insomnia. In animal studies, Ziziphus jujuba Mill.had anxiolytic-like effects at lower doses and sedative effects

2 Evidence-Based Complementary and Alternative Medicine
at higher doses in the black and white test and elevated plusmaze [15]. Juncus effusus L. plays an assisting role in CZZXas an “assistant and messenger herb.” Clinical studies inChina revealed that it can regulate different mental disorders,such as anxiety, improve sleep, and improve gastrointestinalfunction. However, animal studies have not demonstratedthe ability of CZZX to mediate anxiety.
The purpose of the present study was to determinewhether CZZX exerts anxiolytic-like effects in rats. Theanimals were tested in the elevated plus maze, Vogel conflicttest (VCT), and open field. The binding affinity and efficacyof CZZX at the γ-aminobutyric acid-A- (GABAA-) BZDreceptor were determined to reveal the potential mechanismsthat underlie the selective behavioral profile of CZZX.
2. Material and Methods
2.1. Animals. Male Sprague-Dawley rats (160–180 g) wereobtained from the Laboratory Animal Center of the Academyof Military Medical Sciences and used for this study.The animals were housed five per cage under controlledtemperature (22 ± 1◦C) and a 12 h/12 h light/dark cycle(lights on at 07:00 AM) with free access to food and water.The experimenter handled the animals daily to acclimatethem to the manipulation. The animals were used only onceand for only one experiment. The experimental procedureswere approved by the Institutional Animal Care and UseCommittee of the Institute of Psychology of the ChineseAcademy of Sciences and in accordance with the NationalInstitutes of Health Guide for Care and Use of LaboratoryAnimals.
2.2. Drugs and Treatment. Four crude drugs (Valeriana jata-mansi, Ziziphus jujuba, Albizia julibrissin, and Juncus effusus)were purchased from the Medicinal Materials Company ofHebei Anguo and authenticated by Jin-Li Shi, Departmentof Botany, Beijing Chinese Medical University, based ontheir micro- and macroscopic characteristics. The quality ofthese crude drugs was controlled and processed accordingto the Chinese Pharmacopoeia. Drug samples were collectedas voucher specimens and kept with the records. The 35%ethanolic v/v extract of Valeriana jatamansi (200 g/2000 mL;reflux, 2 h × 3), water extracts of Ziziphus jujuba and Albiziajulibrissin (200 g/2000 mL; decoction, 3 h × 2), and 95%ethanolic v/v extract of Juncus effusus (20 g/300 mL; reflux,1 h × 2) were filtered and dried under reduced pressure at atemperature <60◦C. They were then mixed at the ratio statedin the prescription.
The doses are expressed in terms of the dried weightof the CZZX extract per unit body weight of the experi-mental animals (g/kg). Diazepam was chosen as a positivecontrol drug. For oral (p.o.) administration for 10 days,CZZX and diazepam were dissolved in saline. Each drugwas administered in a volume of 1 mL/100 g body weight.Control animals received vehicle (saline) only. In this study,the rats were administered CZZX or DZP 60 and 30 minbefore the test, respectively. The opioid agonist morphinehydrochloride was obtained from Sigma (St. Louis, MO,
USA). Morphine was administered at a dose of 5 mg/kg in asterile 0.9% NaCl solution via intraperitoneal (i.p.) injection30 min before the nociceptive test. All of the animal tests wereperformed on the 10th day of treatment.
2.3. Elevated Plus Maze. Anxiolytic activity was measuredusing the elevated plus maze [16]. The maze consisted oftwo open arms (50.8 cm × 10.2 cm × 1.3 cm) and two closedarms (50.8 cm × 10.2 cm × 40.6 cm) that extended from acentral platform (10.2 cm× 10.2 cm). The maze was elevatedto a height of 72.4 cm above the floor. The entire maze wasconstructed of clear Plexiglas. On the test days, the animalswere transported to the elevated plus maze room and leftundisturbed in a neutral box for 5 min prior to testing.Immediately after this period, each rat was placed on thecentral square facing an open arm and allowed to freelyexplore the maze for 5 min. Arm entries were defined as theentry of all four paws into an arm. A computer recordedthe time spent on and number of entries into the open andclosed arms by means of infrared photocells. The percentageof open arm entries (100× open/total entries) was calculatedfor each animal. The apparatus was wiped clean with waterand dried after each subject.
2.4. Vogel Conflict Test. The VCT was performed in aPlexiglas box (29 cm × 29 cm × 26 cm) with a stainlesssteel grid floor. The metallic spout of a drinking bottle thatcontained water projected into the box. The simultaneouscontact of the animal with the spout and the grid floorclosed an electrical circuit controlled by a sensor, producingseven pulses of water per second whenever the animal was incontact with both components. Each pulse was considereda lick. After every 20 licks, the animal received a 0.5 mAfootshock for 2 s. The sensor recorded the total number oflicks and shocks delivered during the test period. The entireapparatus was located inside a sound-attenuated cage.
2.5. Water Consumption Evaluation. The apparatus was thesame as the one used in the VCT described above, but theelectric shock-delivering system was rendered inoperative.
2.6. Acute Thermal Pain. A radiant heat apparatus wasused to induce acute pain. The thermal thresholds of therats hindpaws were measured. Each rat was placed in aplastic chamber on a glass floor, under which a radiant heatapparatus (100 W projector lamp) was located. A beam oflight through a 4 mm diameter hole in the apparatus wasfocused on the plantar surface of the left hindpaw. Thepaw withdrawal latency (PWL) was defined as the timebetween the light onset and paw withdrawal and adjustedto approximately 8 s to record the baseline. A cutoff time of22 s was used to avoid tissue damage. Four trials spaced atleast 5 min apart were conducted with each hindpaw. The lastthree trials were averaged to provide the mean latency.
2.7. Open Field Test. The open field was a 180 cm diametercylinder with 60 cm high walls. The center of the bottom ofthe apparatus had a 50 cm diameter section. The rats were

Evidence-Based Complementary and Alternative Medicine 3
placed into the field at the same point against the wall andallowed to freely explore the apparatus for 10 min. The totalpath length was recorded by an automatic video trackingsystem. Grooming time, the number of rearings, and thenumber of defecations were also recorded. After each trial,the apparatus was wiped clean with a 10% ethanol solution.
2.8. Biochemical Assay
2.8.1. Tissue Preparation. The preparation was performed at4◦C. The cerebral cortex from four rats was homogenizedfor 5 s in 20 mL of 50 mM Tris-citrate (pH 7.1) using anUltra-Turrax. The suspension was centrifuged at 27,000×gfor 15 min, and the pellet was washed three times withbuffer. The washed pellet was homogenized in 20 mL buffer.The suspension was incubated in a water bath at 37◦C for30 min to remove endogenous GABA and then centrifugedfor 10 min at 27,000×g. The final pellet was resuspended in30 mL buffer and stored in aliquots at −20◦C.
2.8.2. [3H] Ro 15-1788 (Flumazenil) Binding Assay. Thebinding assay was performed according to a previouslydescribed method [17] with modifications. The membranepreparation was thawed and washed with 20 mL of 50 mMTris-citrate (pH 7.1) at 4◦C. The suspension was thencentrifuged at 27,000×g for 10 min at 4◦C. The pelletwas resuspended in 50 mM Tris-citrate (pH 7.1) with 2 mgof the original tissue per milliliter of buffer and thenused for the binding assay. The membrane suspension(500 μL) was then added to 25 μL of test solution (CZZXextract/standard/blank) and 25 μL flumazenil (Ro 15-1788,78 Ci/mmol; Perkin-Elmer Life Sciences), mixed, and incu-bated for 40 min in an ice bath. Nonspecific binding wasdetermined using diazepam (1 μM, final concentration inassay) added to separate samples. After incubation, 5 mL ofice-cold buffer was added to the samples, and the mixturewas poured directly onto Adventic glass fiber filters (GC-50)under suction and immediately washed with 5 mL of ice-cold buffer. The amount of radioactivity was determined byconventional liquid scintillation counting. Specific bindingwas calculated as total binding minus nonspecific binding.All of the experiments were performed in triplicate.
2.9. Statistical Analysis. The data are expressed as mean ±SEM. The data from the elevated plus maze, VCT, acutethermal pain assessment, and open field were analyzedusing one-way analysis of variance (ANOVA) followed byDunnett’s test across the five groups. Probability levels lessthan 0.05 were considered statistically significant.
3. Results
3.1. Elevated Plus Maze. The one-way ANOVA indicatedsignificant differences among groups in the time spent inthe open arms of the elevated plus maze (F4,44 = 6.72,P < 0.01; Figure 1(a)) and percentage of open arm entries(F4,44 = 5.08, P < 0.01; Figure 1(b)). CZZX at a dose of 3 g/kgproduced anxiolytic-like effects, reflected by an increase in
the percentage of open arm entries (P < 0.05). Doses of 1.5and 3 g/kg also increased the time spent in the open armsof the maze (P < 0.05 and P < 0.01, resp.). No differenceswere observed in total arm entries (F4,44 = 0.88, P > 0.05;Figure 1(c)).
3.2. Vogel Conflict Test. To confirm the anxiolytic-like effectsof CZZX, we tested an independent group of rats in theVCT. In this experiment, 50 animals were used. Consistentwith the previous experiment, the one-way ANOVA revealedsignificant variance among the five groups (F4,44 = 2.03,P < 0.01). Doses of 1.5 and 3 g/kg significantly increasedthe number of punished licks compared with controls (bothP < 0.05; Figure 2).
3.3. Water Consumption. Additional experiments were per-formed to test whether CZZX increases water consumptionand nociceptive responses, which are potential confoundingfactors in the VCT. However, no differences in the numberof unpunished licks were observed among the five groups(F4,45 = 0.35, P > 0.05; Figure 3).
3.4. Acute Thermal Pain. We also investigated the effects ofsaline, CZZX, and morphine on acute thermal nociceptivethresholds measured with noxious radiant heat in normalrats. The one-way ANOVA revealed a significant effect oftreatment (F5,54 = 29.67, P < 0.05; Figure 4). Morphinesignificantly increased the PWL compared with the salinecontrol (P < 0.001). In contrast to the analgesic effectof morphine, CZZX did not have any effect on the PWLinduced by the noxious heat stimulus (P > 0.05).
3.5. Open Field Test. No difference in total path length wasobserved among the five groups (F4,44 = 1.98, P > 0.05;Figure 5), with no difference in emotional behavior (i.e.,rearing, grooming, and defecation; data not shown).
3.6. Binding Affinity of CZZX to GABAA-BZD Receptors. Inthe GABAA-BZD binding assay, CZZX was able to displaceflumazenil binding. Therefore, it may inhibit the binding ofthe specific radioligand [3H] Ro 15-1788 to the BZD receptor(Figure 6).
4. Discussion
The present study sought to analyze the behavioral effects ofCZZX, which is used to treat “nervous diseases” in traditionalChinese medicine [11]. The results showed that CZZXexhibited anxiolytic-like activity and did not induce sedativeside effects. The binding assay suggested that the anxiolyticeffects of CZZX may be attributable to the modulation ofGABAA receptors. Altogether, our results lend support tothe traditional use of CZZX in Chinese folk medicine. Thepresent findings may also provide important leads for thedevelopment of potent and selective anxiolytic agents.
When the animals were treated with the higher doses ofCZZX (1.5 and 3 g/kg) for 10 days, anxiety-like behavior inthe elevated plus maze was significantly attenuated, although
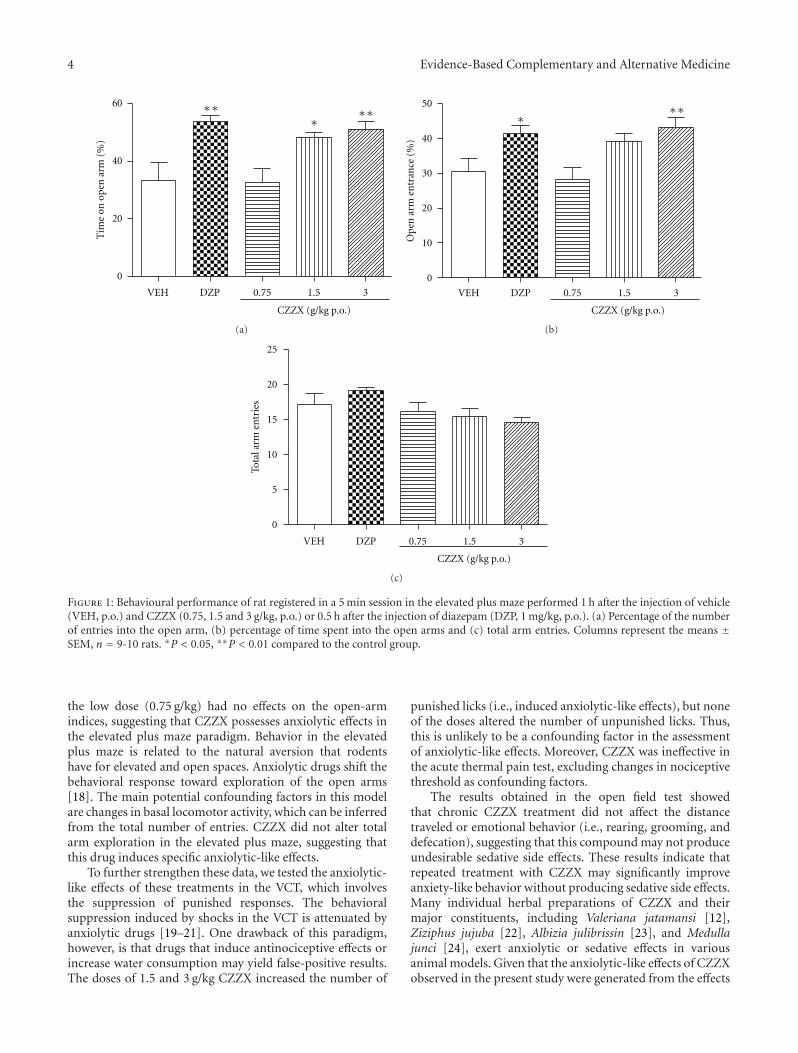
4 Evidence-Based Complementary and Alternative Medicine
VEH DZP 0.75 1.5 3
0
20
40
60 ∗∗ ∗∗
∗
CZZX (g/kg p.o.)
Tim
e on
ope
n a
rm (
%)
(a)
CZZX (g/kg p.o.)
Ope
n a
rm e
ntr
ance
(%
)
VEH DZP 0.75 1.5 3
0
10
20
30
40
50 ∗∗
∗
(b)
CZZX (g/kg p.o.)
VEH DZP 0.75 1.5 3
0
5
10
15
20
25
Tota
l arm
en
trie
s
(c)
Figure 1: Behavioural performance of rat registered in a 5 min session in the elevated plus maze performed 1 h after the injection of vehicle(VEH, p.o.) and CZZX (0.75, 1.5 and 3 g/kg, p.o.) or 0.5 h after the injection of diazepam (DZP, 1 mg/kg, p.o.). (a) Percentage of the numberof entries into the open arm, (b) percentage of time spent into the open arms and (c) total arm entries. Columns represent the means ±SEM, n = 9-10 rats. ∗P < 0.05, ∗∗P < 0.01 compared to the control group.
the low dose (0.75 g/kg) had no effects on the open-armindices, suggesting that CZZX possesses anxiolytic effects inthe elevated plus maze paradigm. Behavior in the elevatedplus maze is related to the natural aversion that rodentshave for elevated and open spaces. Anxiolytic drugs shift thebehavioral response toward exploration of the open arms[18]. The main potential confounding factors in this modelare changes in basal locomotor activity, which can be inferredfrom the total number of entries. CZZX did not alter totalarm exploration in the elevated plus maze, suggesting thatthis drug induces specific anxiolytic-like effects.
To further strengthen these data, we tested the anxiolytic-like effects of these treatments in the VCT, which involvesthe suppression of punished responses. The behavioralsuppression induced by shocks in the VCT is attenuated byanxiolytic drugs [19–21]. One drawback of this paradigm,however, is that drugs that induce antinociceptive effects orincrease water consumption may yield false-positive results.The doses of 1.5 and 3 g/kg CZZX increased the number of
punished licks (i.e., induced anxiolytic-like effects), but noneof the doses altered the number of unpunished licks. Thus,this is unlikely to be a confounding factor in the assessmentof anxiolytic-like effects. Moreover, CZZX was ineffective inthe acute thermal pain test, excluding changes in nociceptivethreshold as confounding factors.
The results obtained in the open field test showedthat chronic CZZX treatment did not affect the distancetraveled or emotional behavior (i.e., rearing, grooming, anddefecation), suggesting that this compound may not produceundesirable sedative side effects. These results indicate thatrepeated treatment with CZZX may significantly improveanxiety-like behavior without producing sedative side effects.Many individual herbal preparations of CZZX and theirmajor constituents, including Valeriana jatamansi [12],Ziziphus jujuba [22], Albizia julibrissin [23], and Medullajunci [24], exert anxiolytic or sedative effects in variousanimal models. Given that the anxiolytic-like effects of CZZXobserved in the present study were generated from the effects
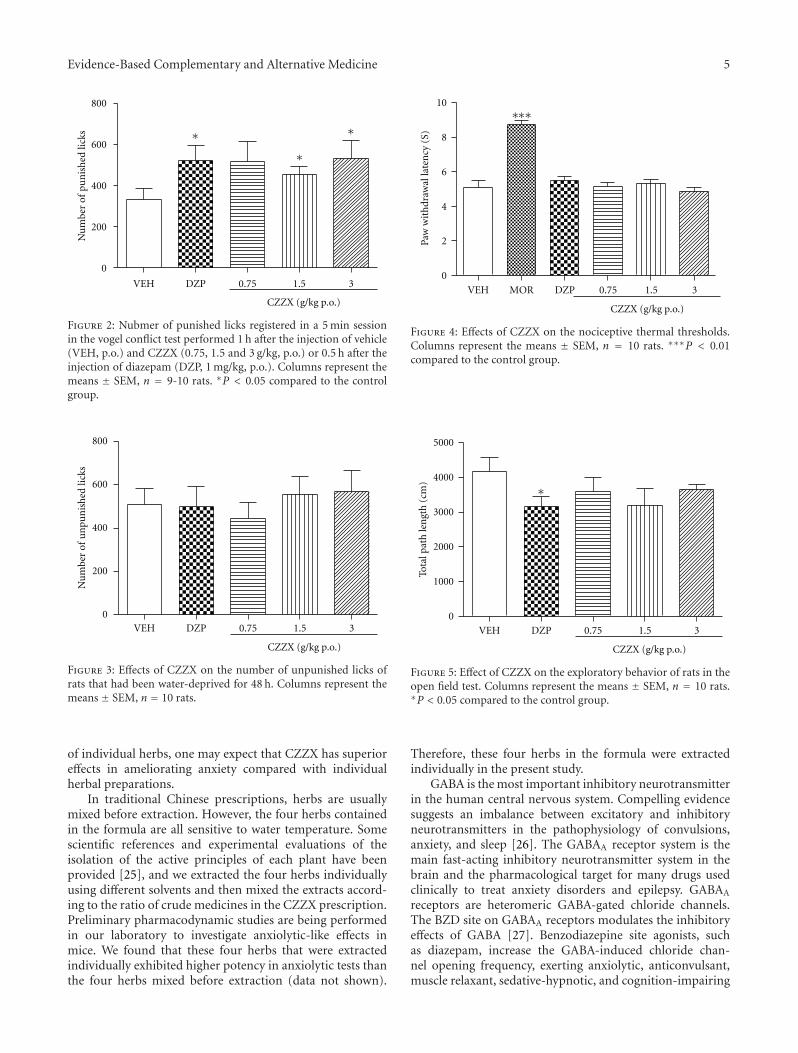
Evidence-Based Complementary and Alternative Medicine 5
CZZX (g/kg p.o.)
VEH DZP 0.75 1.5 3
0
200
400
600
800
Nu
mbe
r of
pu
nis
hed
lick
s ∗∗
∗
Figure 2: Nubmer of punished licks registered in a 5 min sessionin the vogel conflict test performed 1 h after the injection of vehicle(VEH, p.o.) and CZZX (0.75, 1.5 and 3 g/kg, p.o.) or 0.5 h after theinjection of diazepam (DZP, 1 mg/kg, p.o.). Columns represent themeans ± SEM, n = 9-10 rats. ∗P < 0.05 compared to the controlgroup.
CZZX (g/kg p.o.)
VEH DZP 0.75 1.5 30
200
400
600
800
Nu
mbe
r of
un
pun
ish
ed li
cks
Figure 3: Effects of CZZX on the number of unpunished licks ofrats that had been water-deprived for 48 h. Columns represent themeans ± SEM, n = 10 rats.
of individual herbs, one may expect that CZZX has superioreffects in ameliorating anxiety compared with individualherbal preparations.
In traditional Chinese prescriptions, herbs are usuallymixed before extraction. However, the four herbs containedin the formula are all sensitive to water temperature. Somescientific references and experimental evaluations of theisolation of the active principles of each plant have beenprovided [25], and we extracted the four herbs individuallyusing different solvents and then mixed the extracts accord-ing to the ratio of crude medicines in the CZZX prescription.Preliminary pharmacodynamic studies are being performedin our laboratory to investigate anxiolytic-like effects inmice. We found that these four herbs that were extractedindividually exhibited higher potency in anxiolytic tests thanthe four herbs mixed before extraction (data not shown).
CZZX (g/kg p.o.)
VEH DZPMOR 0.75 1.5 3
0
2
4
6
8
10
Paw
wit
hdr
awal
late
ncy
(S)
∗∗∗
Figure 4: Effects of CZZX on the nociceptive thermal thresholds.Columns represent the means ± SEM, n = 10 rats. ∗∗∗P < 0.01compared to the control group.
CZZX (g/kg p.o.)
VEH DZP 0.75 1.5 3
0
1000
2000
3000
4000
5000
Tota
l pat
h le
ngt
h (
cm) ∗
Figure 5: Effect of CZZX on the exploratory behavior of rats in theopen field test. Columns represent the means ± SEM, n = 10 rats.∗P < 0.05 compared to the control group.
Therefore, these four herbs in the formula were extractedindividually in the present study.
GABA is the most important inhibitory neurotransmitterin the human central nervous system. Compelling evidencesuggests an imbalance between excitatory and inhibitoryneurotransmitters in the pathophysiology of convulsions,anxiety, and sleep [26]. The GABAA receptor system is themain fast-acting inhibitory neurotransmitter system in thebrain and the pharmacological target for many drugs usedclinically to treat anxiety disorders and epilepsy. GABAA
receptors are heteromeric GABA-gated chloride channels.The BZD site on GABAA receptors modulates the inhibitoryeffects of GABA [27]. Benzodiazepine site agonists, suchas diazepam, increase the GABA-induced chloride chan-nel opening frequency, exerting anxiolytic, anticonvulsant,muscle relaxant, sedative-hypnotic, and cognition-impairing
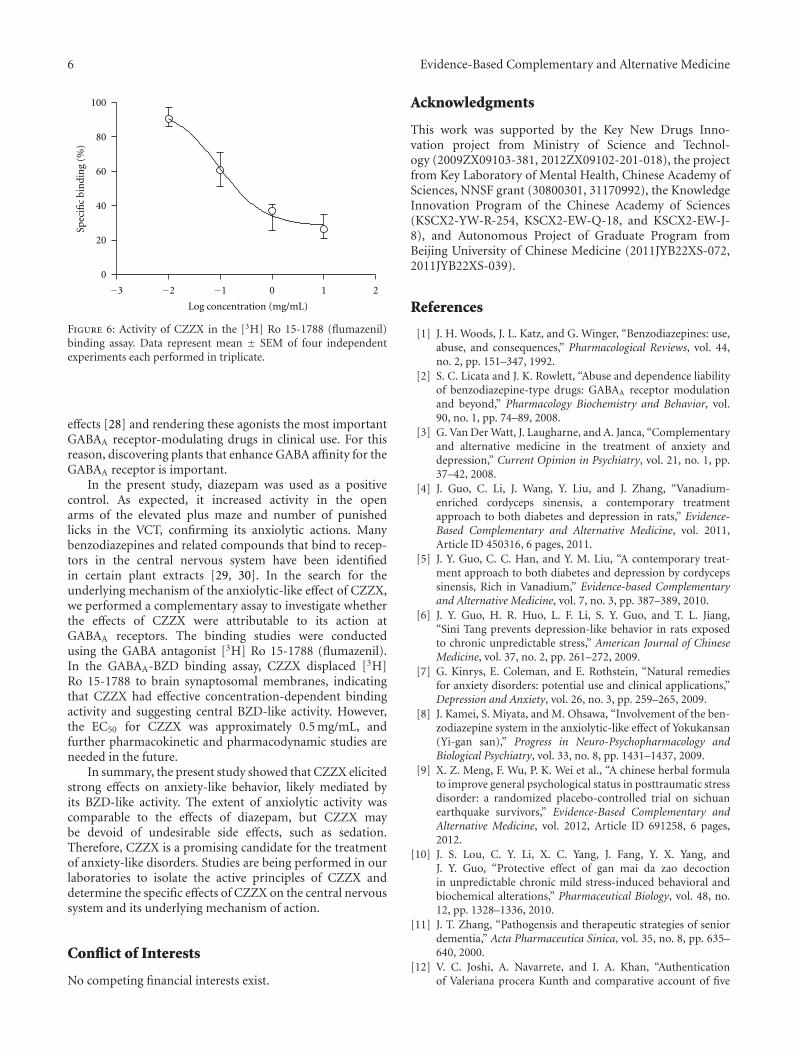
6 Evidence-Based Complementary and Alternative Medicine
0 1 2
0
20
40
60
80
100
Log concentration (mg/mL)
−3 −2 −1
Spec
ific
bin
din
g (%
)
Figure 6: Activity of CZZX in the [3H] Ro 15-1788 (flumazenil)binding assay. Data represent mean ± SEM of four independentexperiments each performed in triplicate.
effects [28] and rendering these agonists the most importantGABAA receptor-modulating drugs in clinical use. For thisreason, discovering plants that enhance GABA affinity for theGABAA receptor is important.
In the present study, diazepam was used as a positivecontrol. As expected, it increased activity in the openarms of the elevated plus maze and number of punishedlicks in the VCT, confirming its anxiolytic actions. Manybenzodiazepines and related compounds that bind to recep-tors in the central nervous system have been identifiedin certain plant extracts [29, 30]. In the search for theunderlying mechanism of the anxiolytic-like effect of CZZX,we performed a complementary assay to investigate whetherthe effects of CZZX were attributable to its action atGABAA receptors. The binding studies were conductedusing the GABA antagonist [3H] Ro 15-1788 (flumazenil).In the GABAA-BZD binding assay, CZZX displaced [3H]Ro 15-1788 to brain synaptosomal membranes, indicatingthat CZZX had effective concentration-dependent bindingactivity and suggesting central BZD-like activity. However,the EC50 for CZZX was approximately 0.5 mg/mL, andfurther pharmacokinetic and pharmacodynamic studies areneeded in the future.
In summary, the present study showed that CZZX elicitedstrong effects on anxiety-like behavior, likely mediated byits BZD-like activity. The extent of anxiolytic activity wascomparable to the effects of diazepam, but CZZX maybe devoid of undesirable side effects, such as sedation.Therefore, CZZX is a promising candidate for the treatmentof anxiety-like disorders. Studies are being performed in ourlaboratories to isolate the active principles of CZZX anddetermine the specific effects of CZZX on the central nervoussystem and its underlying mechanism of action.
Conflict of Interests
No competing financial interests exist.
Acknowledgments
This work was supported by the Key New Drugs Inno-vation project from Ministry of Science and Technol-ogy (2009ZX09103-381, 2012ZX09102-201-018), the projectfrom Key Laboratory of Mental Health, Chinese Academy ofSciences, NNSF grant (30800301, 31170992), the KnowledgeInnovation Program of the Chinese Academy of Sciences(KSCX2-YW-R-254, KSCX2-EW-Q-18, and KSCX2-EW-J-8), and Autonomous Project of Graduate Program fromBeijing University of Chinese Medicine (2011JYB22XS-072,2011JYB22XS-039).
References
[1] J. H. Woods, J. L. Katz, and G. Winger, “Benzodiazepines: use,abuse, and consequences,” Pharmacological Reviews, vol. 44,no. 2, pp. 151–347, 1992.
[2] S. C. Licata and J. K. Rowlett, “Abuse and dependence liabilityof benzodiazepine-type drugs: GABAA receptor modulationand beyond,” Pharmacology Biochemistry and Behavior, vol.90, no. 1, pp. 74–89, 2008.
[3] G. Van Der Watt, J. Laugharne, and A. Janca, “Complementaryand alternative medicine in the treatment of anxiety anddepression,” Current Opinion in Psychiatry, vol. 21, no. 1, pp.37–42, 2008.
[4] J. Guo, C. Li, J. Wang, Y. Liu, and J. Zhang, “Vanadium-enriched cordyceps sinensis, a contemporary treatmentapproach to both diabetes and depression in rats,” Evidence-Based Complementary and Alternative Medicine, vol. 2011,Article ID 450316, 6 pages, 2011.
[5] J. Y. Guo, C. C. Han, and Y. M. Liu, “A contemporary treat-ment approach to both diabetes and depression by cordycepssinensis, Rich in Vanadium,” Evidence-based Complementaryand Alternative Medicine, vol. 7, no. 3, pp. 387–389, 2010.
[6] J. Y. Guo, H. R. Huo, L. F. Li, S. Y. Guo, and T. L. Jiang,“Sini Tang prevents depression-like behavior in rats exposedto chronic unpredictable stress,” American Journal of ChineseMedicine, vol. 37, no. 2, pp. 261–272, 2009.
[7] G. Kinrys, E. Coleman, and E. Rothstein, “Natural remediesfor anxiety disorders: potential use and clinical applications,”Depression and Anxiety, vol. 26, no. 3, pp. 259–265, 2009.
[8] J. Kamei, S. Miyata, and M. Ohsawa, “Involvement of the ben-zodiazepine system in the anxiolytic-like effect of Yokukansan(Yi-gan san),” Progress in Neuro-Psychopharmacology andBiological Psychiatry, vol. 33, no. 8, pp. 1431–1437, 2009.
[9] X. Z. Meng, F. Wu, P. K. Wei et al., “A chinese herbal formulato improve general psychological status in posttraumatic stressdisorder: a randomized placebo-controlled trial on sichuanearthquake survivors,” Evidence-Based Complementary andAlternative Medicine, vol. 2012, Article ID 691258, 6 pages,2012.
[10] J. S. Lou, C. Y. Li, X. C. Yang, J. Fang, Y. X. Yang, andJ. Y. Guo, “Protective effect of gan mai da zao decoctionin unpredictable chronic mild stress-induced behavioral andbiochemical alterations,” Pharmaceutical Biology, vol. 48, no.12, pp. 1328–1336, 2010.
[11] J. T. Zhang, “Pathogensis and therapeutic strategies of seniordementia,” Acta Pharmaceutica Sinica, vol. 35, no. 8, pp. 635–640, 2000.
[12] V. C. Joshi, A. Navarrete, and I. A. Khan, “Authenticationof Valeriana procera Kunth and comparative account of five

Evidence-Based Complementary and Alternative Medicine 7
Valeriana species,” Journal of AOAC International, vol. 88, no.6, pp. 1621–1625, 2005.
[13] R. Andreatini, V. A. Sartori, M. L. V. Seabra, and J. R. Leite,“Effect of valepotriates (valerian extract) in generalized anx-iety disorder: a randomized placebo-controlled pilot study,”Phytotherapy Research, vol. 16, no. 7, pp. 650–654, 2002.
[14] M. Hattesohl, B. Feistel, H. Sievers, R. Lehnfeld, M. Hegger,and H. Winterhoff, “Extracts of Valeriana officinalis L. s.l.show anxiolytic and antidepressant effects but neither sedativenor myorelaxant properties,” Phytomedicine, vol. 15, no. 1-2,pp. 2–15, 2008.
[15] W. H. Peng, M. T. Hsieh, Y. S. Lee, Y. C. Lin, and J. Liao,“Anxiolytic effect of seed of Ziziphus jujuba in mouse modelsof anxiety,” Journal of Ethnopharmacology, vol. 72, no. 3, pp.435–441, 2000.
[16] J. Y. Guo, J. Y. Wang, and F. Luo, “Dissection of placeboanalgesia in mice: the conditions for activation of opioid andnon-opioid systems,” Journal of Psychopharmacology, vol. 24,no. 10, pp. 1561–1567, 2010.
[17] P. Kahnberg, E. Lager, C. Rosenberg et al., “Refinement andevaluation of a pharmacophore model for flavone derivativesbinding to the benzodiazepine site of the GABAA receptor,”Journal of Medicinal Chemistry, vol. 45, no. 19, pp. 4188–4201,2002.
[18] J. Y. Guo, C. Y. Li, Y. P. Ruan et al., “Chronic treat-ment with celecoxib reverses chronic unpredictable stress-induced depressive-like behavior via reducing cyclooxygenase-2 expression in rat brain,” European Journal of Pharmacology,vol. 612, no. 1–3, pp. 54–60, 2009.
[19] M. J. Millan, “The neurobiology and control of anxious states,”Progress in Neurobiology, vol. 70, no. 2, pp. 83–244, 2003.
[20] M. J. Millan and M. Brocco, “The Vogel conflict test:procedural aspects, γ-aminobutyric acid, glutamate andmonoamines,” European Journal of Pharmacology, vol. 463, no.1–3, pp. 67–96, 2003.
[21] J. Y. Guo, X. Y. Yuan, F. Sui et al., “Placebo analgesia affectsthe behavioral despair tests and hormonal secretions in mice,”Psychopharmacology, vol. 217, no. 1, pp. 83–90, 2011.
[22] J. G. Jiang, X. J. Huang, J. Chen, and Q. S. Lin, “Comparisonof the sedative and hypnotic effects of flavonoids, saponins,and polysaccharides extracted from Semen Ziziphus jujube,”Natural Product Research, vol. 21, no. 4, pp. 310–320, 2007.
[23] D. H. Yu, S. Y. Qiao, and Y. M. Zhao, “Advances in study onbark of Albizzia julibrissin,” Zhongguo Zhong Yao Za Zhi, vol.29, no. 7, pp. 619–624, 2004.
[24] Y. J. Liao, H. F. Zhai, B. Zhang, T. X. Duan, and J. M. Huang,“Anxiolytic and sedative effects of dehydroeffusol from Juncuseffusus in mice,” Planta Medica, vol. 77, no. 5, pp. 416–420,2011.
[25] D. M. Eisenberg, E. S. J. Harris, B. A. Littlefield et al., “Devel-oping a library of authenticated Traditional Chinese Medicinal(TCM) plants for systematic biological evaluation—rationale,methods and preliminary results from a Sino-Americancollaboration,” Fitoterapia, vol. 82, no. 1, pp. 17–33, 2011.
[26] H. Mohler, “GABAA receptors in central nervous systemdisease: anxiety, epilepsy, and insomnia,” Journal of Receptorsand Signal Transduction, vol. 26, no. 5-6, pp. 731–740, 2006.
[27] E. R. Korpi, G. Grunder, and H. Luddens, “Drug interactionsat GABAA receptors,” Progress in Neurobiology, vol. 67, no. 2,pp. 113–159, 2002.
[28] W. E. Haefely, J. R. Martin, J. G. Richards, and P. Schoch,“The multiplicity of actions of benzodiazepine receptor
ligands,” Canadian Journal of Psychiatry Revue Canadienne dePsychiatrie, vol. 38, supplement 4, pp. S102–S108, 1993.
[29] F. S. Duarte, M. Marder, A. A. Hoeller et al., “Anticon-vulsant and anxiolytic-like effects of compounds isolatedfrom Polygala sabulosa (Polygalaceae) in rodents: in vitroand in vivo interactions with benzodiazepine binding sites,”Psychopharmacology, vol. 197, no. 3, pp. 351–360, 2008.
[30] H. Viola, C. Wasowski, M. Levi De Stein et al., “Apigenin, acomponent of Matricaria recutita flowers, is a central ben-zodiazepine receptors-ligand with anxiolytic effects,” PlantaMedica, vol. 61, no. 3, pp. 213–216, 1995.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 591298, 13 pagesdoi:10.1155/2012/591298
Research Article
Early Blockade of TLRs MyD88-Dependent Pathway MayReduce Secondary Spinal Cord Injury in the Rats
An-hui Yao,1 Li-yun Jia,1 Yu-kai Zhang,1 Quan-rui Ma,1, 2 Peng Cheng,1
Ling Liu,1 Gong Ju,1 and Fang Kuang1
1 Institute of Neurosciences, The Fourth Military Medical University, 17 West Changle Road, Xi’an 710032, China2 Department of Human Anatomy, Ningxia Medical University, 1160 Victory Street, Yinchuan 750001, China
Correspondence should be addressed to Gong Ju, [email protected] and Fang Kuang, [email protected]
Received 10 February 2012; Revised 24 March 2012; Accepted 25 March 2012
Academic Editor: Ilkay Erdogan Orhan
Copyright © 2012 An-hui Yao et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
To determine the role of toll-like receptors (TLRs) myeloid differentiation factor 88 (MyD88) dependent pathway in the spinal cordsecondary injury, compression injury was made at T8 segment of the spinal cord in adult male Sprague-Dawley rats. Shown by RT-PCR, TLR4 mRNA in the spinal cord was quickly elevated after compression injury. Intramedullary injection of MyD88 inhibitorypeptide (MIP) resulted in significant improvement in locomotor function recovery at various time points after surgery. Meanwhile,injury area, p38 phosphorylation, and proinflammation cytokines in the injured spinal cord were significantly reduced in MIP-treated animals, compared with control peptide (CP) group. These data suggest that TLRs MyD88-dependent pathway may playan important role in the development of secondary spinal cord injury, and inhibition of this pathway at early time after primaryinjury could effectively protect cells from inflammation and apoptosis and therefore improve the functional recovery.
1. Introduction
Acute traumatic spinal cord injury (SCI) is an unexpected,catastrophic event which causes various lifelong disabilitiesof the patients. The consequences also result in graveinfluences in a row to family members and society. However,there are no fully restorative therapies for SCI up to now[1–3]. The pathological sequelae following acute SCI aredivided into two phases: primary mechanical injury andsecondary injury [4–6]. Primary SCI is caused by directmechanical trauma, and it instigates a progressive wave ofsecondary injury via activation of a series of pathophysio-logical mechanisms including alterations in microvascularperfusion, inflammation, lipid peroxidation, free radicalgeneration, apoptotic/necrotic cell death, and dysregulationof ionic homeostasis [7–11]. These mechanisms caused thedestruction of axonal tracts that were left intact after theinitial trauma, which is the major impediment to functionalrecovery after SCI. It has been proved that fibers spared as fewas 5–10% in the spinal cord are sufficient to facilitate basiclocomotion recovery following SCI in rats [12]. Therefore,
how to protect the spared fibers and reduce the secondaryinjury has become the top priority for scientists working inthe research of SCI.
Inflammatory responses are very important, even central,in the pathological process of the acute and chronic phasesof secondary injury. During the secondary injury phase,the central nervous system (CNS) evokes both innate andadaptive immunities [11, 13]. After primary SCI, residentmicroglia, invaded macrophages and dendritic cells couldwork as antigen presenting cells through toll-like receptors(TLRs) signaling [14] at and around the injury site. Atleast 12 TLRs have been identified in the mammals, andthey are involved in recognition of pathogen-associatedmolecular patterns (PAMPs) and activation/regulation ofboth innate and adaptive immunities [15, 16]. Moreover,it was recently shown that some TLRs (e.g., TLR2, TLR4)can be stimulated by endogenous molecules named danger-associated molecular patterns (DAMPs) [1]. DAMPs couldbe released from injury sites of the tissue [17, 18]. InSCI, many DAMPs including heat-shock proteins (HSPs),necrotic cells, fibronectin and hyaluronic acid, heparan

2 Evidence-Based Complementary and Alternative Medicine
sulphate, lung surfactant protein A, high-mobility groupbox 1 and mRNA would be increased at sites of spinalinjury, as accumulated studies reported [19–26]. Meanwhile,research with genetic deficient mice has indicated that TLR2or TLR4 deficiency impaired function recovery in SCI [27],while research with TLR4 loss-of-function mutation miceshowed that TLR4 deficiency protected against focal cerebralischemia and axotomy-induced neruodegeneration [28].
The signaling pathways activated by TLRs are broadlyclassified into MyD88-dependent and -independent path-ways because MyD88 is the universal adapter proteinrecruited by all TLRs except TLR3 [29, 30]. TLRs MyD88-dependent pathway activates nuclear-factor-κB (NF-κB) [31]and subsequently results in the production of inflammatorycytokines (e.g., TNF-α, IL-1β) [32–35], while MyD88-independent pathway is related to transcriptional activationof type I interferons [36–38] and also activates late phaseNF-κB via TAK which is shared by MyD88-dependentpathway. However, the exact roles of these pathways in thepathophysiology of SCI remain unclear.
NF-κB is essential for neurons survival against oxidativestress and ischemic degeneration [39], but it also contributesto inflammation and apoptosis after CNS injury [40, 41].The different phases of NF-κB may serve distinct role inthis balance. Therefore, we hypothesized that TLRs MyD88-dependent pathway correlated with the development ofspinal cord secondary injury via inflammatory reaction. Inthis study, at first we detected TLR4 mRNA expressionby RT-PCR in the spinal cord at several time points aftercompression SCI, then used inhibitory peptide to blockMyD88-dependent signaling at early time after SCI, evalu-ated motor functional recovery by Basso-Beattie-Bresnahan(BBB) score, beam-walking test, and foot print analysis,and investigated histological damage, cell apoptosis, andinflammatory reactions with immunohistochemistry (IHC)and Western blotting assay. We found that inhibition ofMyD88-dependent pathway at early time could reduce theinflammatory response, preserve more neurons, reduce thelesion size in the spinal cord, and therefore promote motorfunctional recovery from spinal cord compression injury.
2. Materials and Methods
2.1. Animals and Surgical Procedure. Male Sprague-Dawleyrats (200–220 g) were purchased from Experimental AnimalCenter of the Fourth Military Medical University. Rats werehoused in a 12/12 h light/dark cycle room with standardrodent food and water available ad libitum. Efforts were madeto minimize animal discomfort and to sacrifice the fewestanimals. All experiments were done in accordance with theguidelines established by the Animal Care Committee ofFourth Military Medical University.
All the rats were anesthetized with an intraperitonealinjection of pentobarbital sodium (50–60 mg/kg), and thenthe back region was shaved and aseptically prepared withiodophor. A laminectomy was performed at the T8 level toexpose the cord underneath without disrupting the dura.After the spinous processes of T7 and T9 were clamped tostabilize the spine, the exposed dorsal surface of the cord was
subjected to compression as described before [42]. Briefly, acompression plate attached to a 20 g copper rod was used.The compression plate was lowered down ventrally at a rateof 0.5 mm/min. It reached the bottom 5 min later and wasremoved at a constant speed.
After compression injury, 24 rats were used for RT-PCRassay to detect TLR4 mRNA expression at 0 h, 3 h, 24 h,72 h, 7 d, and 14 d after injury. Another batch of rats wasused for MyD88 blockade observation (Table 1). Locomotorfunctions were evaluated, and histological analyses wereperformed on this batch of animals.
2.2. Reverse Transcription PCR (RT-PCR) for TLR4. TLR4mRNA levels after SCI were detected by using RT-PCR.After compression SCI as previously described, rats wereanesthetized and sacrificed at 0 h, 3 h, 24 h, 72 h, 7 d, and14 d after compression injury (for every time point, n =4). Total RNA was isolated from 10 mm of thoracic spinalcord tissue centered at the injury site. RNA was purifiedusing the Trizol reagent according to the manufacturer’sinstructions. In order to remove any traces of contaminatingDNA, RNA samples were treated using the RNase-free DNaseI (TaKaRa, Co., Ltd. Dalian, China). One microgram oftotal RNA was reverse transcribed into first strand cDNA ineach 20 μL reaction mixture, according to the manufacturer’sinstructions (TaKaRa, Co., Ltd. Dalian, China). Primers usedwere as follows: TLR4, sense: 5′-GTTGGATTTTACGAA-TTCCACCT-3′, and antisense: 5′-TGCTTCTTGTTCTTC-CTCTGATG-3′; the expected size was 646 bp; β-actin, sense:5′-GAGAGGGAAATCGTGCGTGAC-3′, and antisense: 5′-CATCTGCTGGAAGGTGGACA-3′; and the expected sizewas 453 bp.
PCR was performed according to the manufacturer’sinstructions. The PCR mixture (20 μL) contained 10 μmolof each primer, 200 μM dNTPs, 25 mM MgCl2, 2 μL 10xPCR buffer, 1 U Taq DNA polymerase (TaKaRa, Co., Ltd.Dalian, China), and 10 μM purified cDNA. Denaturing,annealing and extension times were 30 seconds each, at 95,57, and 72◦C, respectively. cDNA samples were amplifiedfor 40 cycles. The specific product was separated on a 2%agarose gel (Biowest, Spain) and detected after staining withethidium bromide under UV illumination.
2.3. MyD88 Blockade Experiments. After compression injuryas described previously, rats were randomly divided intotwo groups: MIP group and CP group. Each rat of MIPgroup received intramedullary injection of 5 μL MyD88inhibitor peptide (MIP, 100 μmol/L, Imgenex, USA), andeach rat of CP group was given 5 μL control peptide (CP,100 μmol/L, Imgenex, USA), dissolved in 0.01 mol/L PBS,immediately after compression injury at the injury site. Thepeptides we used for inhibition and control contain a proteintransduction (PTD) sequence (DRQIKIWFQNRRMKWKK)derived from antennapedia, and the CP consists of only thePTD sequence, which renders the peptide cell permeability.The injections were done at both sides of the dorsal vein ata constant rate of 1 μL/min at about 1 mm below the spinalcord surface. Then the muscles and skin were closed in layers,

Evidence-Based Complementary and Alternative Medicine 3
Table 1: Animal assignment for MyD88 blockade experiments.
TotalHistological analysis Western blot assay
0 h 7 d 14 d 7 d
CP 18 3 5 5 5
MIP 18 3 5 5 5
and every two rats were placed in one cage. Manual bladderexpression was performed at least twice per day until reflexbladder emptying was established.
2.3.1. Histological Analysis. Animals were perfused via theleft cardio-ventricle with 100 mL of physiological saline andsubsequently with 400 mL of 4% paraformaldehyde in 0.1 Mphosphate buffer (PB, PH = 7.4) at 7 d and 14 d, respectively(n = 5). Three centimeters spinal cord was carefully dissectedout with the injury site in the center and was immersed in25% sucrose solution for 24–48 h at 4◦C until it sank. Sagittalsections were cut at 20 μm in thickness with a cryostat andthawed-mounted onto gelatinized slides. The sections werekept at −20◦C until used.
For Haematoxylin & Eosin (H&E) staining, sections werewashed briefly in distilled water, stained in haematoxylinsolution for 5 min. After this, sections were washed inrunning tap water for 5 min and differentiated in 1% acidalcohol for 30 sec. Then sections were washed in tap water for1 min, placed in eosin for 30 sec, dehydrated through 70%,80%, 90%, and 100% alcohol 2 min each, clear in 2 changesof xylene, 5 min each, and mounted with xylene-basedmounting medium.
For IHC staining, the sections were rinsed in PBS 3 times5 min and blocked with 1% bovine serum albumin for 1 hto eliminate nonspecific staining and then incubated withprimary antibodies 16–24 h at room temperature. Primaryantibodies were rabbit anti-Cleaved Caspase-3 (1 : 400, CellSignaling Technology, USA), rabbit anti-GFAP (1 : 4000,Sigma, USA), mouse anti-NeuN (1 : 500, Sigma, USA),and goat anti-IgG (1 : 400, Millipore, USA). Then sectionswere rinsed with PBS 3 times, 5 min for each time, andincubated with the appropriate secondary antibody for 2 hat room temperature. Sections were examined on OlympusBX 51 microscope (Olympus, Japan) or FV 1000 confocalmicroscope (Olympus, Japan).
For cell counting, five sections were chosen for analysisin each rat, namely, the section with central canal and twoadjacent sections on both sides. After pictures were taken,images were opened in Adobe Photoshop 9.0 and stucktogether as a montage. Caspase-3-positive cells (with nucleusindicated by Hoechest staining) were counted from 4 repre-sented areas randomly chosen in the sections. NeuN-positivecells within the first and second 1000 μm distance to thelesion border were counted. The mean values of the fivesections in each rat were used for statistical analysis. Forlesion area measurement, GFAP staining images were takenas mentioned previously. The boundary of the injured areawas outlined according to the differences between normaltissue and necrotic tissue or cavity. The pixels of the outlined
area were calculated and then converted into the size of injuryby using Adobe Photoshop 9.0 software.
2.3.2. Western Blot Assay. At 7 d after compression SCI, ani-mals were deeply anesthetized and sacrificed (n = 5). One-centimeter long spinal cord tissues, 5 mm rostral and 5 mmcordal to the injury site, were removed rapidly and storedin liquid nitrogen and then processed for extraction of pro-tein. Briefly, tissue samples were homogenized with 0.5 mLof ice-cold lysis buffer (20 mM Tris-HCl, pH 7.5, 1 mMEDTA, 5 mM MgCl2, 1 mM DTT, 20 μg/mL aprotinin, 1 mMPMSF, and 2 mM sodium orthovanadate). The homogenateswere centrifuged at 13,000 rpm for 10 min at 4◦C andsupernatant were removed. The protein concentration wasdetermined using Bradford method, a detergent-compatibleprotein assay with a bovine serum albumin as standard.Samples were boiled at 100◦C for 10 min and then wereelectrophoresed on 10–15% SDS-PAGE and transferred ontoa nitrocellulose membrane (Millipore, Bedford, MA, USA).The filter membranes were blocked with 5% nonfat milkfor 1.5 h at room temperature and incubated with the pri-mary antibody (phospho-p38 MAPK 1 : 1000, Cell SignalingTechnology, MA, USA; caspase-3, 1 : 1000, Cell SignalingTechnology, MA, USA; TNF-α 1 : 500, Santa, CA, USA; IL-1β, 1 : 1000, gifted by Professor Beifen Shen, Departmentof Molecular Immunology, Beijing, China) 16–24 h at 4◦C.Then the membrane was washed with TBST buffer andincubated with the secondary antibody conjugated withhorseradish peroxidase (1 : 8000; Jackson ImmunoResearch,USA) for 1 h at room temperature and visualized in ECLsolution. The density of specific bands was measured withImage J (NIH, USA) software.
2.3.3. BBB Score. BBB scales were used to detect the recoveryof the motor function every day from 0 d to 14 d after injury.Each rat was placed in an 80 × 130 × 30 cm open field andrun for 4 minutes and scaled from 0 to 21 point according tothe guidelines [43].
2.3.4. Beam-Walking Tests. Beam-walking tests were used todetect the distance from the rump to the plane to reflect therecovery of the motor function at 7 d and 14 d after injury.The apparatus used for behavioral analysis beam is 120 cmlong and 12 cm wide and supported by a 70 cm framework.We set up a video camera and recorded the animal’s behaviorwalking from one end to the other end of the beam. Then wedid video frame extraction and selected 5 represented framesto measure rump height by using Image tool 2.0 (UTHSCSA)software.
2.3.5. Footprint Analysis. Footprint analysis was conductedat 7 d and 14 d after injury. A piece of white paper (15 ×89 cm2) was put on the track and both of the hindlimbsplantar surfaces were colored blue and dorsal surfaces red[44]. Then the rats were allowed to walk from one endof the track to the other end. When foot dorsal surfacetouched the track, the white paper would be printed withred color, while when foot plantar touched the track, blue
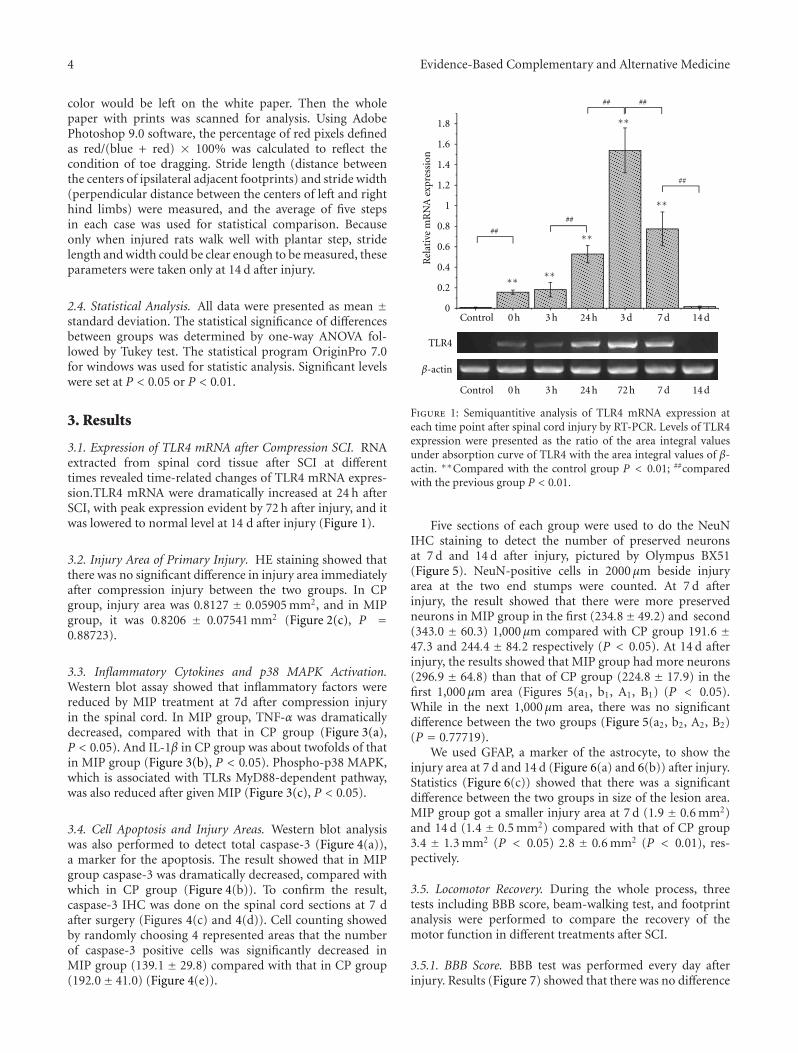
4 Evidence-Based Complementary and Alternative Medicine
color would be left on the white paper. Then the wholepaper with prints was scanned for analysis. Using AdobePhotoshop 9.0 software, the percentage of red pixels definedas red/(blue + red) × 100% was calculated to reflect thecondition of toe dragging. Stride length (distance betweenthe centers of ipsilateral adjacent footprints) and stride width(perpendicular distance between the centers of left and righthind limbs) were measured, and the average of five stepsin each case was used for statistical comparison. Becauseonly when injured rats walk well with plantar step, stridelength and width could be clear enough to be measured, theseparameters were taken only at 14 d after injury.
2.4. Statistical Analysis. All data were presented as mean ±standard deviation. The statistical significance of differencesbetween groups was determined by one-way ANOVA fol-lowed by Tukey test. The statistical program OriginPro 7.0for windows was used for statistic analysis. Significant levelswere set at P < 0.05 or P < 0.01.
3. Results
3.1. Expression of TLR4 mRNA after Compression SCI. RNAextracted from spinal cord tissue after SCI at differenttimes revealed time-related changes of TLR4 mRNA expres-sion.TLR4 mRNA were dramatically increased at 24 h afterSCI, with peak expression evident by 72 h after injury, and itwas lowered to normal level at 14 d after injury (Figure 1).
3.2. Injury Area of Primary Injury. HE staining showed thatthere was no significant difference in injury area immediatelyafter compression injury between the two groups. In CPgroup, injury area was 0.8127 ± 0.05905 mm2, and in MIPgroup, it was 0.8206 ± 0.07541 mm2 (Figure 2(c), P =0.88723).
3.3. Inflammatory Cytokines and p38 MAPK Activation.Western blot assay showed that inflammatory factors werereduced by MIP treatment at 7d after compression injuryin the spinal cord. In MIP group, TNF-α was dramaticallydecreased, compared with that in CP group (Figure 3(a),P < 0.05). And IL-1β in CP group was about twofolds of thatin MIP group (Figure 3(b), P < 0.05). Phospho-p38 MAPK,which is associated with TLRs MyD88-dependent pathway,was also reduced after given MIP (Figure 3(c), P < 0.05).
3.4. Cell Apoptosis and Injury Areas. Western blot analysiswas also performed to detect total caspase-3 (Figure 4(a)),a marker for the apoptosis. The result showed that in MIPgroup caspase-3 was dramatically decreased, compared withwhich in CP group (Figure 4(b)). To confirm the result,caspase-3 IHC was done on the spinal cord sections at 7 dafter surgery (Figures 4(c) and 4(d)). Cell counting showedby randomly choosing 4 represented areas that the numberof caspase-3 positive cells was significantly decreased inMIP group (139.1 ± 29.8) compared with that in CP group(192.0± 41.0) (Figure 4(e)).
1.8
1.6
1.4
1.2
1
0.8
0.6
0.4
0.2
0Control 0 h 3 h 24 h 3 d 7 d 14 d
Control 0 h 3 h 24 h 7 d 14 d
∗∗
∗∗
∗∗
∗∗∗∗
Rel
ativ
e m
RN
A e
xpre
ssio
n
72 h
TLR4
β-actin
## ##
##
##
##
Figure 1: Semiquantitive analysis of TLR4 mRNA expression ateach time point after spinal cord injury by RT-PCR. Levels of TLR4expression were presented as the ratio of the area integral valuesunder absorption curve of TLR4 with the area integral values of β-actin. ∗∗Compared with the control group P < 0.01; ##comparedwith the previous group P < 0.01.
Five sections of each group were used to do the NeuNIHC staining to detect the number of preserved neuronsat 7 d and 14 d after injury, pictured by Olympus BX51(Figure 5). NeuN-positive cells in 2000 μm beside injuryarea at the two end stumps were counted. At 7 d afterinjury, the result showed that there were more preservedneurons in MIP group in the first (234.8± 49.2) and second(343.0 ± 60.3) 1,000 μm compared with CP group 191.6 ±47.3 and 244.4 ± 84.2 respectively (P < 0.05). At 14 d afterinjury, the results showed that MIP group had more neurons(296.9 ± 64.8) than that of CP group (224.8 ± 17.9) in thefirst 1,000 μm area (Figures 5(a1, b1, A1, B1) (P < 0.05).While in the next 1,000 μm area, there was no significantdifference between the two groups (Figure 5(a2, b2, A2, B2)(P = 0.77719).
We used GFAP, a marker of the astrocyte, to show theinjury area at 7 d and 14 d (Figure 6(a) and 6(b)) after injury.Statistics (Figure 6(c)) showed that there was a significantdifference between the two groups in size of the lesion area.MIP group got a smaller injury area at 7 d (1.9 ± 0.6 mm2)and 14 d (1.4 ± 0.5 mm2) compared with that of CP group3.4 ± 1.3 mm2 (P < 0.05) 2.8 ± 0.6 mm2 (P < 0.01), res-pectively.
3.5. Locomotor Recovery. During the whole process, threetests including BBB score, beam-walking test, and footprintanalysis were performed to compare the recovery of themotor function in different treatments after SCI.
3.5.1. BBB Score. BBB test was performed every day afterinjury. Results (Figure 7) showed that there was no difference
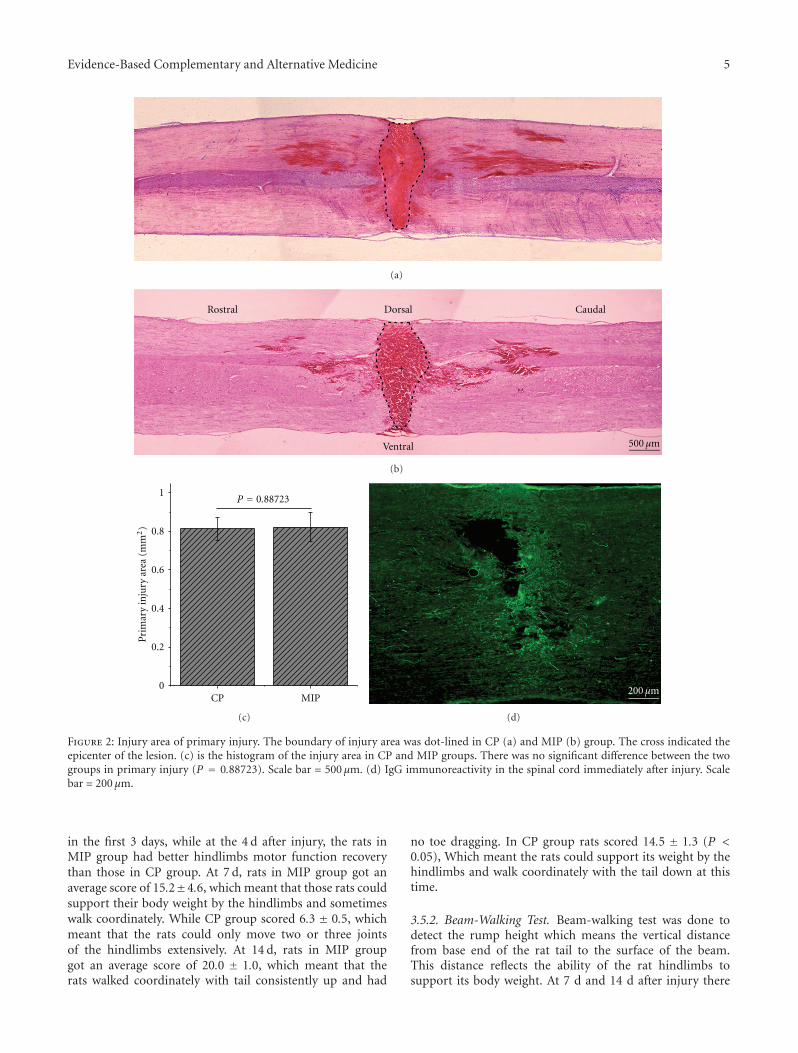
Evidence-Based Complementary and Alternative Medicine 5
+
(a)
Ventral
CaudalDorsalRostral
500 μm
+
(b)
P = 0.887231
0.8
0.6
0.4
0.2
0CP MIP
Pri
mar
y in
jury
are
a (m
m2)
(c)
200 μm
(d)
Figure 2: Injury area of primary injury. The boundary of injury area was dot-lined in CP (a) and MIP (b) group. The cross indicated theepicenter of the lesion. (c) is the histogram of the injury area in CP and MIP groups. There was no significant difference between the twogroups in primary injury (P = 0.88723). Scale bar = 500 μm. (d) IgG immunoreactivity in the spinal cord immediately after injury. Scalebar = 200 μm.
in the first 3 days, while at the 4 d after injury, the rats inMIP group had better hindlimbs motor function recoverythan those in CP group. At 7 d, rats in MIP group got anaverage score of 15.2±4.6, which meant that those rats couldsupport their body weight by the hindlimbs and sometimeswalk coordinately. While CP group scored 6.3 ± 0.5, whichmeant that the rats could only move two or three jointsof the hindlimbs extensively. At 14 d, rats in MIP groupgot an average score of 20.0 ± 1.0, which meant that therats walked coordinately with tail consistently up and had
no toe dragging. In CP group rats scored 14.5 ± 1.3 (P <0.05), Which meant the rats could support its weight by thehindlimbs and walk coordinately with the tail down at thistime.
3.5.2. Beam-Walking Test. Beam-walking test was done todetect the rump height which means the vertical distancefrom base end of the rat tail to the surface of the beam.This distance reflects the ability of the rat hindlimbs tosupport its body weight. At 7 d and 14 d after injury there
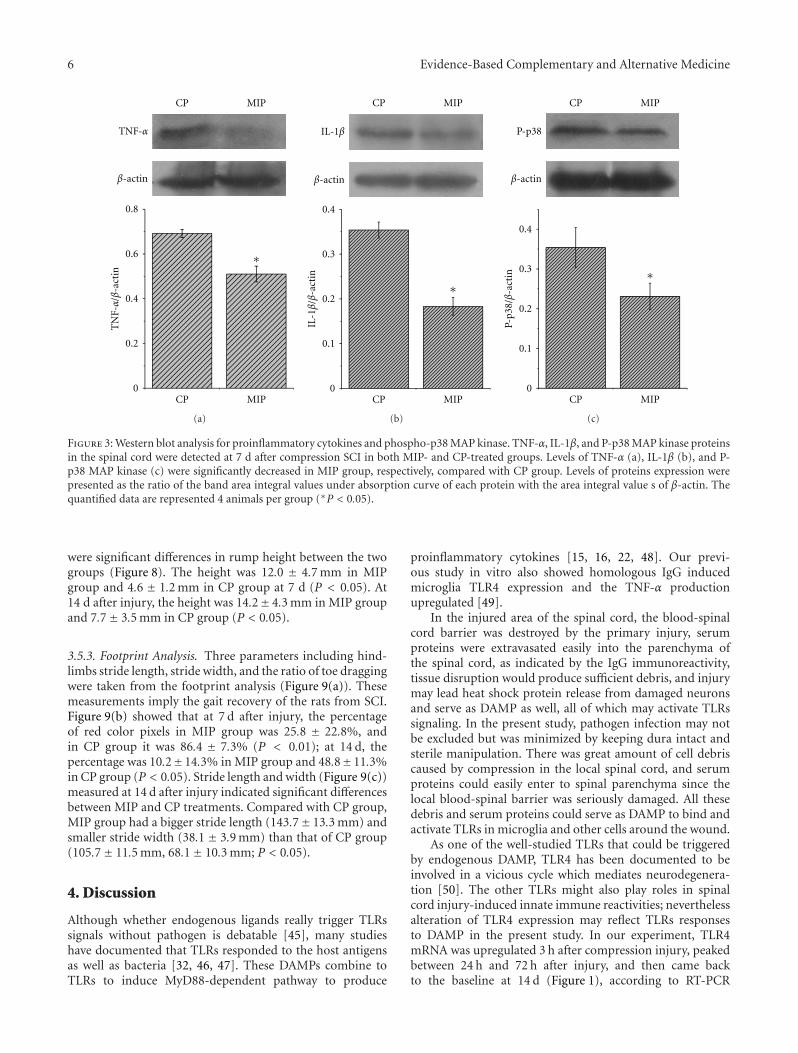
6 Evidence-Based Complementary and Alternative Medicine
0.8
0.6
0.4
0.2
0
TN
F-α
/β-a
ctin
CP MIP
CP MIP
∗
TNF-α
β-actin
(a)
0.4
0.3
0.2
0.1
0CP MIP
CP MIP
∗
IL-1β
/β-a
ctin
IL-1β
β-actin
(b)
P-p38
β-actin
0.4
0.3
0.2
0.1
0
P-p3
8/β
-act
in
CP MIP
CP MIP
∗
(c)
Figure 3: Western blot analysis for proinflammatory cytokines and phospho-p38 MAP kinase. TNF-α, IL-1β, and P-p38 MAP kinase proteinsin the spinal cord were detected at 7 d after compression SCI in both MIP- and CP-treated groups. Levels of TNF-α (a), IL-1β (b), and P-p38 MAP kinase (c) were significantly decreased in MIP group, respectively, compared with CP group. Levels of proteins expression werepresented as the ratio of the band area integral values under absorption curve of each protein with the area integral value s of β-actin. Thequantified data are represented 4 animals per group (∗P < 0.05).
were significant differences in rump height between the twogroups (Figure 8). The height was 12.0 ± 4.7 mm in MIPgroup and 4.6 ± 1.2 mm in CP group at 7 d (P < 0.05). At14 d after injury, the height was 14.2± 4.3 mm in MIP groupand 7.7± 3.5 mm in CP group (P < 0.05).
3.5.3. Footprint Analysis. Three parameters including hind-limbs stride length, stride width, and the ratio of toe draggingwere taken from the footprint analysis (Figure 9(a)). Thesemeasurements imply the gait recovery of the rats from SCI.Figure 9(b) showed that at 7 d after injury, the percentageof red color pixels in MIP group was 25.8 ± 22.8%, andin CP group it was 86.4 ± 7.3% (P < 0.01); at 14 d, thepercentage was 10.2±14.3% in MIP group and 48.8±11.3%in CP group (P < 0.05). Stride length and width (Figure 9(c))measured at 14 d after injury indicated significant differencesbetween MIP and CP treatments. Compared with CP group,MIP group had a bigger stride length (143.7± 13.3 mm) andsmaller stride width (38.1 ± 3.9 mm) than that of CP group(105.7± 11.5 mm, 68.1± 10.3 mm; P < 0.05).
4. Discussion
Although whether endogenous ligands really trigger TLRssignals without pathogen is debatable [45], many studieshave documented that TLRs responded to the host antigensas well as bacteria [32, 46, 47]. These DAMPs combine toTLRs to induce MyD88-dependent pathway to produce
proinflammatory cytokines [15, 16, 22, 48]. Our previ-ous study in vitro also showed homologous IgG inducedmicroglia TLR4 expression and the TNF-α productionupregulated [49].
In the injured area of the spinal cord, the blood-spinalcord barrier was destroyed by the primary injury, serumproteins were extravasated easily into the parenchyma ofthe spinal cord, as indicated by the IgG immunoreactivity,tissue disruption would produce sufficient debris, and injurymay lead heat shock protein release from damaged neuronsand serve as DAMP as well, all of which may activate TLRssignaling. In the present study, pathogen infection may notbe excluded but was minimized by keeping dura intact andsterile manipulation. There was great amount of cell debriscaused by compression in the local spinal cord, and serumproteins could easily enter to spinal parenchyma since thelocal blood-spinal barrier was seriously damaged. All thesedebris and serum proteins could serve as DAMP to bind andactivate TLRs in microglia and other cells around the wound.
As one of the well-studied TLRs that could be triggeredby endogenous DAMP, TLR4 has been documented to beinvolved in a vicious cycle which mediates neurodegenera-tion [50]. The other TLRs might also play roles in spinalcord injury-induced innate immune reactivities; neverthelessalteration of TLR4 expression may reflect TLRs responsesto DAMP in the present study. In our experiment, TLR4mRNA was upregulated 3 h after compression injury, peakedbetween 24 h and 72 h after injury, and then came backto the baseline at 14 d (Figure 1), according to RT-PCR
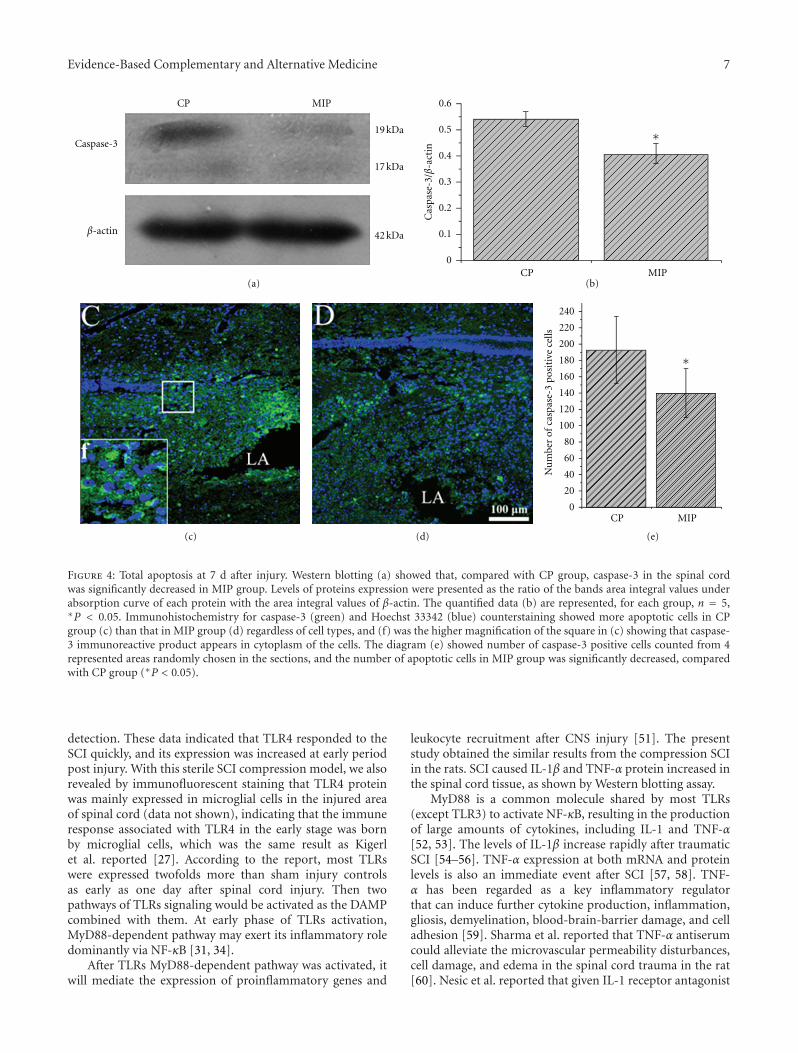
Evidence-Based Complementary and Alternative Medicine 7
240
220
200
180
160
140
120
100
80
60
40
20
0
Nu
mbe
r of
cas
pase
-3 p
osit
ive
cells
CP MIP
∗
0.6
0.5
0.4
0.3
0.2
0.1
0CP MIP
∗
Cas
pase
-3/β
-act
inCaspase-3
CP MIP
19 kDa
17 kDa
42 kDaβ-actin
(a) (b)
(c) (d) (e)
Figure 4: Total apoptosis at 7 d after injury. Western blotting (a) showed that, compared with CP group, caspase-3 in the spinal cordwas significantly decreased in MIP group. Levels of proteins expression were presented as the ratio of the bands area integral values underabsorption curve of each protein with the area integral values of β-actin. The quantified data (b) are represented, for each group, n = 5,∗P < 0.05. Immunohistochemistry for caspase-3 (green) and Hoechst 33342 (blue) counterstaining showed more apoptotic cells in CPgroup (c) than that in MIP group (d) regardless of cell types, and (f) was the higher magnification of the square in (c) showing that caspase-3 immunoreactive product appears in cytoplasm of the cells. The diagram (e) showed number of caspase-3 positive cells counted from 4represented areas randomly chosen in the sections, and the number of apoptotic cells in MIP group was significantly decreased, comparedwith CP group (∗P < 0.05).
detection. These data indicated that TLR4 responded to theSCI quickly, and its expression was increased at early periodpost injury. With this sterile SCI compression model, we alsorevealed by immunofluorescent staining that TLR4 proteinwas mainly expressed in microglial cells in the injured areaof spinal cord (data not shown), indicating that the immuneresponse associated with TLR4 in the early stage was bornby microglial cells, which was the same result as Kigerlet al. reported [27]. According to the report, most TLRswere expressed twofolds more than sham injury controlsas early as one day after spinal cord injury. Then twopathways of TLRs signaling would be activated as the DAMPcombined with them. At early phase of TLRs activation,MyD88-dependent pathway may exert its inflammatory roledominantly via NF-κB [31, 34].
After TLRs MyD88-dependent pathway was activated, itwill mediate the expression of proinflammatory genes and
leukocyte recruitment after CNS injury [51]. The presentstudy obtained the similar results from the compression SCIin the rats. SCI caused IL-1β and TNF-α protein increased inthe spinal cord tissue, as shown by Western blotting assay.
MyD88 is a common molecule shared by most TLRs(except TLR3) to activate NF-κB, resulting in the productionof large amounts of cytokines, including IL-1 and TNF-α[52, 53]. The levels of IL-1β increase rapidly after traumaticSCI [54–56]. TNF-α expression at both mRNA and proteinlevels is also an immediate event after SCI [57, 58]. TNF-α has been regarded as a key inflammatory regulatorthat can induce further cytokine production, inflammation,gliosis, demyelination, blood-brain-barrier damage, and celladhesion [59]. Sharma et al. reported that TNF-α antiserumcould alleviate the microvascular permeability disturbances,cell damage, and edema in the spinal cord trauma in the rat[60]. Nesic et al. reported that given IL-1 receptor antagonist
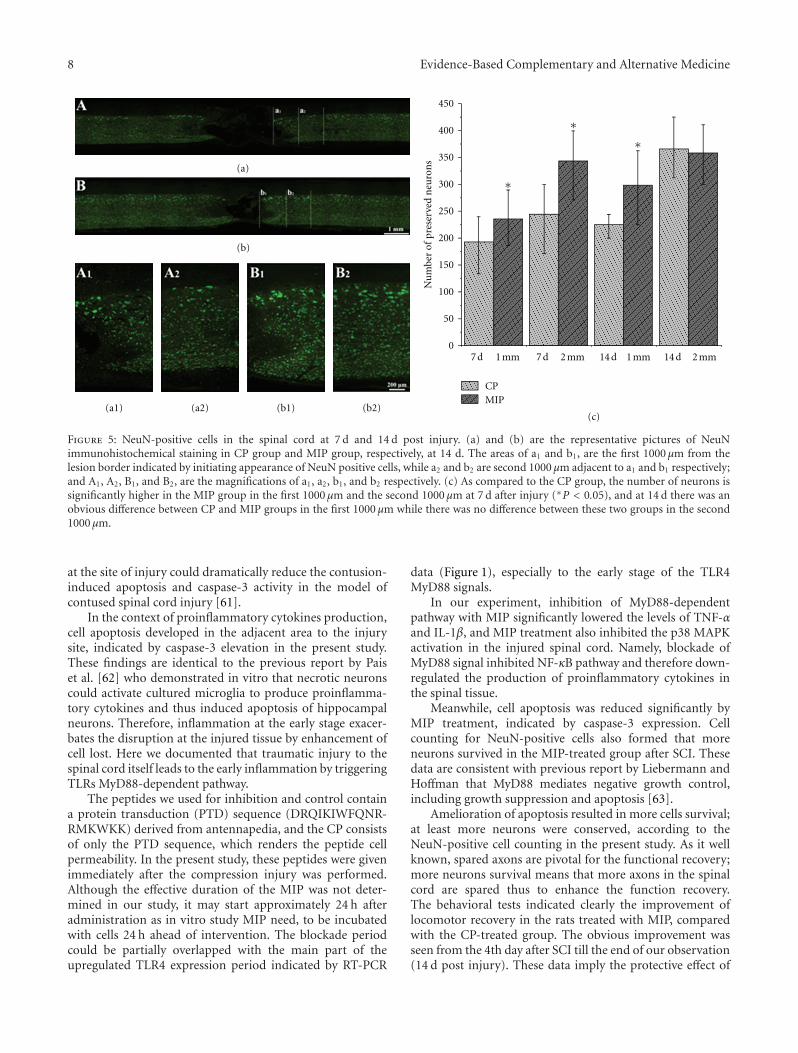
8 Evidence-Based Complementary and Alternative Medicine
(a)
(b)
(c)(a1) (a2) (b1) (b2)
450
400
350
300
250
200
150
100
50
07 d 1 mm 7 d 2 mm 14 d 1 mm 14 d 2 mm
Nu
mbe
r of
pre
serv
ed n
euro
ns
CPMIP
∗
∗∗
Figure 5: NeuN-positive cells in the spinal cord at 7 d and 14 d post injury. (a) and (b) are the representative pictures of NeuNimmunohistochemical staining in CP group and MIP group, respectively, at 14 d. The areas of a1 and b1, are the first 1000 μm from thelesion border indicated by initiating appearance of NeuN positive cells, while a2 and b2 are second 1000 μm adjacent to a1 and b1 respectively;and A1, A2, B1, and B2, are the magnifications of a1, a2, b1, and b2 respectively. (c) As compared to the CP group, the number of neurons issignificantly higher in the MIP group in the first 1000 μm and the second 1000 μm at 7 d after injury (∗P < 0.05), and at 14 d there was anobvious difference between CP and MIP groups in the first 1000 μm while there was no difference between these two groups in the second1000 μm.
at the site of injury could dramatically reduce the contusion-induced apoptosis and caspase-3 activity in the model ofcontused spinal cord injury [61].
In the context of proinflammatory cytokines production,cell apoptosis developed in the adjacent area to the injurysite, indicated by caspase-3 elevation in the present study.These findings are identical to the previous report by Paiset al. [62] who demonstrated in vitro that necrotic neuronscould activate cultured microglia to produce proinflamma-tory cytokines and thus induced apoptosis of hippocampalneurons. Therefore, inflammation at the early stage exacer-bates the disruption at the injured tissue by enhancement ofcell lost. Here we documented that traumatic injury to thespinal cord itself leads to the early inflammation by triggeringTLRs MyD88-dependent pathway.
The peptides we used for inhibition and control containa protein transduction (PTD) sequence (DRQIKIWFQNR-RMKWKK) derived from antennapedia, and the CP consistsof only the PTD sequence, which renders the peptide cellpermeability. In the present study, these peptides were givenimmediately after the compression injury was performed.Although the effective duration of the MIP was not deter-mined in our study, it may start approximately 24 h afteradministration as in vitro study MIP need, to be incubatedwith cells 24 h ahead of intervention. The blockade periodcould be partially overlapped with the main part of theupregulated TLR4 expression period indicated by RT-PCR
data (Figure 1), especially to the early stage of the TLR4MyD88 signals.
In our experiment, inhibition of MyD88-dependentpathway with MIP significantly lowered the levels of TNF-αand IL-1β, and MIP treatment also inhibited the p38 MAPKactivation in the injured spinal cord. Namely, blockade ofMyD88 signal inhibited NF-κB pathway and therefore down-regulated the production of proinflammatory cytokines inthe spinal tissue.
Meanwhile, cell apoptosis was reduced significantly byMIP treatment, indicated by caspase-3 expression. Cellcounting for NeuN-positive cells also formed that moreneurons survived in the MIP-treated group after SCI. Thesedata are consistent with previous report by Liebermann andHoffman that MyD88 mediates negative growth control,including growth suppression and apoptosis [63].
Amelioration of apoptosis resulted in more cells survival;at least more neurons were conserved, according to theNeuN-positive cell counting in the present study. As it wellknown, spared axons are pivotal for the functional recovery;more neurons survival means that more axons in the spinalcord are spared thus to enhance the function recovery.The behavioral tests indicated clearly the improvement oflocomotor recovery in the rats treated with MIP, comparedwith the CP-treated group. The obvious improvement wasseen from the 4th day after SCI till the end of our observation(14 d post injury). These data imply the protective effect of
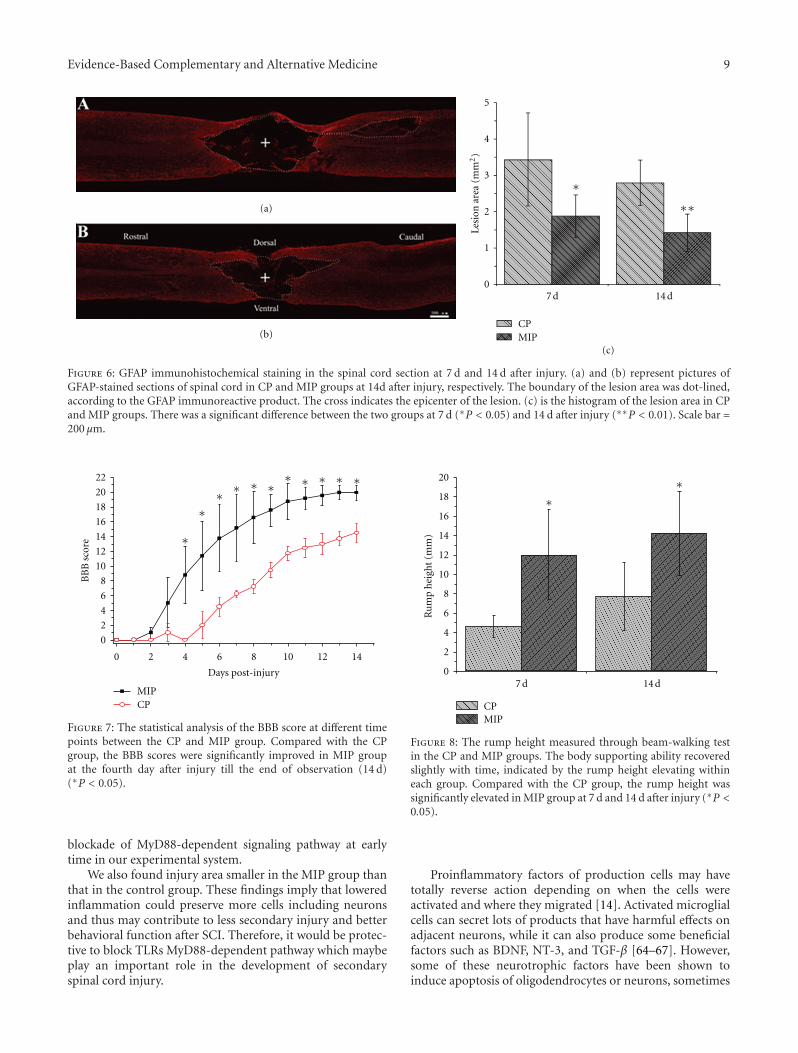
Evidence-Based Complementary and Alternative Medicine 9
Lesi
on a
rea
(mm
2)
∗
5
4
3
2
1
07 d 14 d
∗∗
CPMIP
(a)
(b)
(c)
Figure 6: GFAP immunohistochemical staining in the spinal cord section at 7 d and 14 d after injury. (a) and (b) represent pictures ofGFAP-stained sections of spinal cord in CP and MIP groups at 14d after injury, respectively. The boundary of the lesion area was dot-lined,according to the GFAP immunoreactive product. The cross indicates the epicenter of the lesion. (c) is the histogram of the lesion area in CPand MIP groups. There was a significant difference between the two groups at 7 d (∗P < 0.05) and 14 d after injury (∗∗P < 0.01). Scale bar =200 μm.
MIPCP
22
20
18
16
14
12
10
8
6
4
2
0
0 2 4 6 8 10 12 14
Days post-injury
BB
B s
core
∗∗∗∗∗∗∗∗∗
∗
∗
Figure 7: The statistical analysis of the BBB score at different timepoints between the CP and MIP group. Compared with the CPgroup, the BBB scores were significantly improved in MIP groupat the fourth day after injury till the end of observation (14 d)(∗P < 0.05).
blockade of MyD88-dependent signaling pathway at earlytime in our experimental system.
We also found injury area smaller in the MIP group thanthat in the control group. These findings imply that loweredinflammation could preserve more cells including neuronsand thus may contribute to less secondary injury and betterbehavioral function after SCI. Therefore, it would be protec-tive to block TLRs MyD88-dependent pathway which maybeplay an important role in the development of secondaryspinal cord injury.
∗∗
20
18
16
14
12
10
8
6
4
2
07 d 14 d
CPMIP
Ru
mp
hei
ght
(mm
)
Figure 8: The rump height measured through beam-walking testin the CP and MIP groups. The body supporting ability recoveredslightly with time, indicated by the rump height elevating withineach group. Compared with the CP group, the rump height wassignificantly elevated in MIP group at 7 d and 14 d after injury (∗P <0.05).
Proinflammatory factors of production cells may havetotally reverse action depending on when the cells wereactivated and where they migrated [14]. Activated microglialcells can secret lots of products that have harmful effects onadjacent neurons, while it can also produce some beneficialfactors such as BDNF, NT-3, and TGF-β [64–67]. However,some of these neurotrophic factors have been shown toinduce apoptosis of oligodendrocytes or neurons, sometimes
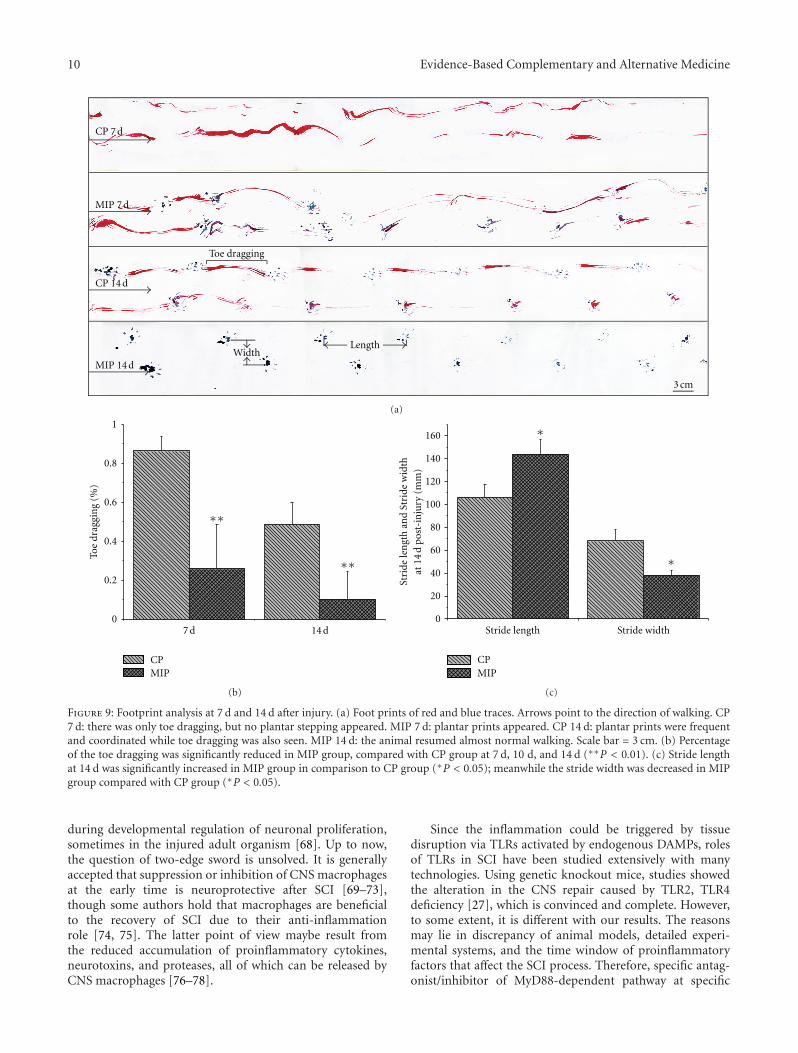
10 Evidence-Based Complementary and Alternative Medicine
CP 7 d
MIP 7 d
CP 14 d
MIP 14 d
Toe dragging
3 cm
WidthLength
(a)
∗∗
∗∗
1
0.8
0.6
0.4
0.2
07 d 14 d
CPMIP
Toe
drag
gin
g (%
)
(b)
∗
∗160
140
120
100
80
60
40
20
0Stride length Stride width
Stri
de le
ngt
h a
nd
Stri
de w
idth
at 1
4 d
post
-in
jury
(m
m)
CPMIP
(c)
Figure 9: Footprint analysis at 7 d and 14 d after injury. (a) Foot prints of red and blue traces. Arrows point to the direction of walking. CP7 d: there was only toe dragging, but no plantar stepping appeared. MIP 7 d: plantar prints appeared. CP 14 d: plantar prints were frequentand coordinated while toe dragging was also seen. MIP 14 d: the animal resumed almost normal walking. Scale bar = 3 cm. (b) Percentageof the toe dragging was significantly reduced in MIP group, compared with CP group at 7 d, 10 d, and 14 d (∗∗P < 0.01). (c) Stride lengthat 14 d was significantly increased in MIP group in comparison to CP group (∗P < 0.05); meanwhile the stride width was decreased in MIPgroup compared with CP group (∗P < 0.05).
during developmental regulation of neuronal proliferation,sometimes in the injured adult organism [68]. Up to now,the question of two-edge sword is unsolved. It is generallyaccepted that suppression or inhibition of CNS macrophagesat the early time is neuroprotective after SCI [69–73],though some authors hold that macrophages are beneficialto the recovery of SCI due to their anti-inflammationrole [74, 75]. The latter point of view maybe result fromthe reduced accumulation of proinflammatory cytokines,neurotoxins, and proteases, all of which can be released byCNS macrophages [76–78].
Since the inflammation could be triggered by tissuedisruption via TLRs activated by endogenous DAMPs, rolesof TLRs in SCI have been studied extensively with manytechnologies. Using genetic knockout mice, studies showedthe alteration in the CNS repair caused by TLR2, TLR4deficiency [27], which is convinced and complete. However,to some extent, it is different with our results. The reasonsmay lie in discrepancy of animal models, detailed experi-mental systems, and the time window of proinflammatoryfactors that affect the SCI process. Therefore, specific antag-onist/inhibitor of MyD88-dependent pathway at specific

Evidence-Based Complementary and Alternative Medicine 11
phase only may target the harmful effects, protect the sparedtissue after SCI regardless of the cell types, and promotethe recovery through inhibition of inflammation and cellapoptosis.
It is interesting that stimulated astrocytes with TLRsligands could inhibit their ability to uptake excess glutamate[79, 80]. As it is well known, SCI leads to the release ofmassive of glutamate [81]. And excitotoxicity of glutamatewas widely studied and showed significant involvement insecondary damage [81]. In 1999, Vezzani et al. reported thatglutamate could induce IL-1 synthesis [82]. These studiessuggest that TLRs may be associated with not only inflam-mation but also many other mechanisms involved in spinalcord secondary injury.
In conclusion, the present study showed with rat com-pression SCI models that traumatic injury that caused tissuedamages may trigger TLRs signals, and TLRs MyD88-dependent pathway may play crucial roles in the secondaryinjury. Inhibition of the TLRs MyD88-dependent pathway atearly time after spinal cord injury could protect the spinalcord from inflammation and apoptosis and promote thefunctional recovery.
Acknowledgments
The present work was supported by the National NaturalScience Foundation of China, the Natural Science Foun-dation of China (30500451), the Military 11th Five-yearSpecial-Purpose Program for PLA (082028), the ChinesePLA National Scientific Technological Project (06G089), theMajor Project of Military Medicine (08XJZ002), and theMilitary 11th Five-year Development of Medicine (2006K15-G1).
References
[1] K. A. Kigerl and P. G. Popovich, “Toll-like receptors in spinalcord injury,” Current Topics in Microbiology and Immunology,vol. 336, no. 1, pp. 121–136, 2009.
[2] Y. Taoka and K. Okajima, “Spinal cord injury in the rat,”Progress in Neurobiology, vol. 56, no. 3, pp. 341–358, 1998.
[3] C. H. Tator and M. G. Fehlings, “Review of the secondaryinjury theory of acute spinal cord trauma with emphasis onvascular mechanisms,” Journal of Neurosurgery, vol. 75, no. 1,pp. 15–26, 1991.
[4] W. F. Collins, “A review and update of experiment and clinicalstudies of spinal cord injury,” Paraplegia, vol. 21, no. 4, pp.204–219, 1983.
[5] E. D. Hall, P. A. Yonkers, K. L. Horan, and J. M. Braughler,“Correlation between attenuation of posttraumatic spinalcord ischemia and preservation of tissue vitamin E by the 21-aminosteroid U74006F: evidence for an in vivo antioxidantmechanism,” Journal of Neurotrauma, vol. 6, no. 3, pp. 169–176, 1989.
[6] A. N. Sandler and C. H. Tator, “Review of the effect of spinalcord trauma on the vessels and blood flow in the spinal cord,”Journal of Neurosurgery, vol. 45, no. 6, pp. 638–646, 1976.
[7] C. H. Tator, “Update on the pathophysiology and pathologyof acute spinal cord injury,” Brain Pathology, vol. 5, no. 4, pp.407–413, 1995.
[8] S. K. Agrawal and M. G. Fehlings, “Mechanisms of secondaryinjury to spinal cord axons in vitro: Role of Na+, Na+–K+–ATPase, the Na+–H+ exchanger, and the Na+–Ca2+ exchanger,”Journal of Neuroscience, vol. 16, no. 2, pp. 545–552, 1996.
[9] M. S. Beattie, G. E. Hermann, R. C. Rogers, and J. C.Bresnahan, “Cell death in models of spinal cord injury,”Progress in Brain Research, vol. 137, pp. 37–47, 2002.
[10] G. Schwartz and M. G. Fehlings, “Secondary injury mecha-nisms of spinal cord trauma: a novel therapeutic approachfor the management of secondary pathophysiology with thesodium channel blocker riluzole,” Progress in Brain Research,vol. 137, pp. 177–190, 2002.
[11] C. Profyris, S. S. Cheema, D. Zang, M. F. Azari, K. Boyle,and S. Petratos, “Degenerative and regenerative mechanismsgoverning spinal cord injury,” Neurobiology of Disease, vol. 15,no. 3, pp. 415–436, 2004.
[12] D. M. Basso, M. S. Beattie, and J. C. Bresnahan, “Gradedhistological and locomotor outcomes after spinal cord contu-sion using the NYU weight-drop device versus transection,”Experimental Neurology, vol. 139, no. 2, pp. 244–256, 1996.
[13] E. Hauben and M. Schwartz, “Therapeutic vaccination forspinal cord injury: helping the body to cure itself,” Trends inPharmacological Sciences, vol. 24, no. 1, pp. 7–12, 2003.
[14] A. Trivedi, A. D. Olivas, and L. J. Noble-Haeusslein, “Inflam-mation and spinal cord injury: infiltrating leukocytes as deter-minants of injury and repair processes,” Clinical NeuroscienceResearch, vol. 6, no. 5, pp. 283–292, 2006.
[15] M. D. Nguyen, J. P. Julien, and S. Rivest, “Innate immunity:the missing link in neuroprotection and neurodegeneration?”Nature Reviews Neuroscience, vol. 3, no. 3, pp. 216–227, 2002.
[16] A. Ozinsky, D. M. Underhill, J. D. Fontenot et al., “Therepertoire for pattern recognition of pathogens by the innateimmune system is defined by cooperation between Toll-likereceptors,” Proceedings of the National Academy of Sciences ofthe United States of America, vol. 97, no. 25, pp. 13766–13771,2000.
[17] Z. Zhang and H. J. Schluesener, “Mammalian Toll-like recep-tors: from endogenous ligands to tissue regeneration,” Cellularand Molecular Life Sciences, vol. 63, no. 24, pp. 2901–2907,2006.
[18] P. A. Carpentier, D. S. Duncan, and S. D. Miller, “Glial Toll-like receptor signaling in central nervous system infection andautoimmunity,” Brain, Behavior, and Immunity, vol. 22, no. 2,pp. 140–147, 2008.
[19] A. Asea, M. Rehli, E. Kabingu et al., “Novel signal transductionpathway utilized by extracellular HSP70. Role of Toll-likereceptor (TLR) 2 and TLR4,” The Journal of BiologicalChemistry, vol. 277, no. 17, pp. 15028–15034, 2002.
[20] S. Basu, R. J. Binder, R. Suto, K. M. Anderson, and P. K.Srivastava, “Necrotic but not apoptotic cell death releases heatshock proteins, which deliver a partial maturation signal todendritic cells and activate the NF-κB pathway,” InternationalImmunology, vol. 12, no. 11, pp. 1539–1546, 2000.
[21] G. B. Johnson, G. J. Brunn, Y. Kodaira, and J. L. Platt,“Receptor-mediated monitoring of tissue well-being via detec-tion of soluble heparan sulfate by Toll-like receptor 4,” Journalof Immunology, vol. 168, no. 10, pp. 5233–5239, 2002.
[22] M. Li, D. F. Carpio, Y. Zheng et al., “An essential role of the NF-κB/Toll-like receptor pathway in induction of inflammatoryand tissue-repair gene expression by necrotic cells,” Journal ofImmunology, vol. 166, no. 12, pp. 7128–7135, 2001.
[23] K. Ohashi, V. Burkart, S. Flohe, and H. Kolb, “Cutting edge:heat shock protein 60 is a putative endogenous ligand of the

12 Evidence-Based Complementary and Alternative Medicine
Toll-like receptor-4 complex,” Journal of Immunology, vol. 164,no. 2, pp. 558–561, 2000.
[24] Y. Okamura, M. Watari, E. S. Jerud et al., “The extra domainA of fibronectin activates Toll-like receptor 4,” The Journal ofBiological Chemistry, vol. 276, no. 13, pp. 10229–10233, 2001.
[25] T. Taguchi, J. L. Mitcham, S. K. Dower, J. E. Sims, and J. R.Testa, “Chromosomal localization of TIL, a gene encodinga protein related to the Drosophila transmembrane receptorToll, to human chromosome 4p14,” Genomics, vol. 32, no. 3,pp. 486–488, 1996.
[26] C. Termeer, F. Benedix, J. Sleeman et al., “Oligosaccharidesof hyaluronan activate dendritic cells via Toll-like receptor 4,”Journal of Experimental Medicine, vol. 195, no. 1, pp. 99–111,2002.
[27] K. A. Kigerl, W. Lai, S. Rivest, R. P. Hart, A. R. Satoskar, and P.G. Popovich, “Toll-like receptor (TLR)-2 and TLR-4 regulateinflammation, gliosis, and myelin sparing after spinal cordinjury,” Journal of Neurochemistry, vol. 102, no. 1, pp. 37–50,2007.
[28] U. Kilic, E. Kilic, C. M. Matter, C. L. Bassetti, and D. M.Hermann, “TLR-4 deficiency protects against focal cerebralischemia and axotomy-induced neurodegeneration,” Neurobi-ology of Disease, vol. 31, no. 1, pp. 33–40, 2008.
[29] T. Kawai and S. Akira, “Signaling to NF-κB by Toll-likereceptors,” Trends in Molecular Medicine, vol. 13, no. 11, pp.460–469, 2007.
[30] E. Okun, K. J. Griffioen, J. D. Lathia, S. C. Tang, M. P. Mattson,and T. V. Arumugam, “Toll-like receptors in neurodegenera-tion,” Brain Research Reviews, vol. 59, no. 2, pp. 278–292, 2009.
[31] K. D. Buchanan, S. L. Huang, H. Kim, D. D. McPherson,and R. C. MacDonald, “Encapsulation of NF-κB decoyoligonucleotides within echogenic liposomes and ultrasound-triggered release,” Journal of Controlled Release, vol. 141, no. 2,pp. 193–198, 2010.
[32] S. Akira and K. Takeda, “Toll-like receptor signalling,” NatureReviews Immunology, vol. 4, no. 7, pp. 499–511, 2004.
[33] O. Takeuchi, K. Takeda, K. Hoshino, O. Adachi, T. Ogawa, andS. Akira, “Cellular responses to bacterial cell wall componentsare mediated through MyD88-dependent signaling cascades,”International Immunology, vol. 12, no. 1, pp. 113–117, 2000.
[34] O. Takeuchi, A. Kaufmann, K. Grote et al., “Cutting edge:preferentially the R-stereoisomer of the mycoplasmal lipopep-tide macrophage-activating lipopeptide-2 activates immunecells through a Toll-like receptor 2- and MyD88-dependentsignaling pathway,” Journal of Immunology, vol. 164, no. 2, pp.554–557, 2000.
[35] H. Hacker, R. M. Vabulas, O. Takeuchi, K. Hoshino, S. Akira,and H. Wagner, “Immune cell activation by bacterial CpG-DNA through myeloid differentiation marker 88 and tumornecrosis factor receptor-associated factor (TRAF)6,” Journal ofExperimental Medicine, vol. 192, no. 4, pp. 595–600, 2000.
[36] S. E. Doyle, S. A. Vaidya, R. O’Connell et al., “IRF3 mediatesa TLR3/TLR4-specific antiviral gene program,” Immunity, vol.17, no. 3, pp. 251–263, 2002.
[37] K. Hoshino, T. Kaisho, T. Iwabe, O. Takeuchi, and S. Akira,“Differential involvement of IFN-β in Toll-like receptor-stimulated dendritic cell activation,” International Immunol-ogy, vol. 14, no. 10, pp. 1225–1231, 2002.
[38] V. Toshchakov, B. W. Jones, P. Y. Perera et al., “TLR4, but notTLR2, mediates IFN-β-induced STATIα/β-dependent geneexpression in macrophages,” Nature Immunology, vol. 3, no.4, pp. 392–398, 2002.
[39] S. Heck, F. Lezoualc’h, S. Engert, and C. Behl, “Insulin-likegrowth factor-1-mediated neuroprotection against oxidative
stress is associated with activation of nuclear factor κB,” TheJournal of Biological Chemistry, vol. 274, no. 14, pp. 9828–9835, 1999.
[40] L. Xu, Y. Zhan, Y. Wang, G. Z. Feuerstein, and X. Wang,“Recombinant adenoviral expression of dominant negativeIκBα protects brain from cerebral ischemic injury,” Biochemi-cal and Biophysical Research Communications, vol. 299, no. 1,pp. 14–17, 2002.
[41] J. R. Bethea, M. Castro, R. W. Keane, T. T. Lee, W. D. Dietrich,and R. P. Yezierski, “Traumatic spinal cord injury inducesnuclear factor-κB activation,” Journal of Neuroscience, vol. 18,no. 9, pp. 3251–3260, 1998.
[42] L. Y. Jia, A. H. Yao, F. Kuang, Y. K. Zhang, X. F. Shen, and G.Ju, “Beneficial effect of the traditional Chinese drug Shu-Xue-Tong on recovery of spinal cord injury in the rat,” Evidence-Based Complementary and Alternative Medicine, vol. 2011,Article ID 862197, 2011.
[43] D. M. Basso, M. S. Beattie, and J. C. Bresnahan, “A sensitiveand reliable locomotor rating scale for open field testing inrats,” Journal of Neurotrauma, vol. 12, no. 1, pp. 1–21, 1995.
[44] J. R. Faulkner, J. E. Herrmann, M. J. Woo, K. E. Tansey, N.B. Doan, and M. V. Sofroniew, “Reactive astrocytes protecttissue and preserve function after spinal cord injury,” Journalof Neuroscience, vol. 24, no. 9, pp. 2143–2155, 2004.
[45] M. F. Tsan and B. Gao, “Heat shock protein and innateimmunity,” Cellular and Molecular Immunology, vol. 1, no. 4,pp. 274–279, 2004.
[46] A. A. Beg, “Endogenous ligands of Toll-like receptors: Impli-cations for regulating inflammatory and immune responses,”Trends in Immunology, vol. 23, no. 11, pp. 509–512, 2002.
[47] M. F. Tsan and B. Gao, “Endogenous ligands of Toll-likereceptors,” Journal of Leukocyte Biology, vol. 76, no. 3, pp. 514–519, 2004.
[48] N. Esen, F. Y. Tanga, J. A. DeLeo, and T. Kielian, “Toll-likereceptor 2 (TLR2) mediates astrocyte activation in response tothe Gram-positive bacterium Staphylococcus aureus,” Journalof Neurochemistry, vol. 88, no. 3, pp. 746–758, 2004.
[49] R. Wu, L. Liu, Z. W. Peng et al., “Toll-like receptor 4expression and cytokine secretion in microglial cells inducedby IgG stimulation,” Chinese Journal of Cellular and MolecularImmunology, vol. 25, no. 3, pp. 201–203, 2009.
[50] S. Lehnardt, E. Schott, T. Trimbuch et al., “A vicious cycleinvolving release of heat shock protein 60 from injured cellsand activation of Toll-like receptor 4 mediates neurodegener-ation in the CNS,” Journal of Neuroscience, vol. 28, no. 10, pp.2320–2331, 2008.
[51] A. A. Babcock, H. Toft-Hansen, and T. Owens, “Signalingthrough MyD88 regulates leukocyte recruitment after braininjury,” Journal of Immunology, vol. 181, no. 9, pp. 6481–6490,2008.
[52] M. Yamamoto, S. Sato, H. Hemmi et al., “Role of adaptorTRIF in the MyD88-independent Toll-like receptor signalingpathway,” Science, vol. 301, no. 5633, pp. 640–643, 2003.
[53] E. Meylan, K. Burns, K. Hofmann et al., “RIP1 is an essentialmediator of Toll-like receptor 3-induced NF-κB activation,”Nature Immunology, vol. 5, no. 5, pp. 503–507, 2004.
[54] M. Hayashi, T. Ueyama, K. Nemoto, T. Tamaki, and E. Senba,“Sequential mRNA expression for immediate early genes,cytokines, and neurotrophins in spinal cord injury,” Journalof Neurotrauma, vol. 17, no. 3, pp. 203–218, 2000.
[55] W. J. Streit, S. L. Semple-Rowland, S. D. Hurley, R. C. Miller, P.G. Popovich, and B. T. Stokes, “Cytokine mRNA profiles incontused spinal cord and axotomized facial nucleus suggest

Evidence-Based Complementary and Alternative Medicine 13
a beneficial role for inflammation and gliosis,” ExperimentalNeurology, vol. 152, no. 1, pp. 74–87, 1998.
[56] C. X. Wang, J. A. Olschowka, and J. R. Wrathall, “Increase ofinterleukin-lβ mRNA and protein in the spinal cord followingexperimental traumatic injury in the rat,” Brain Research, vol.759, no. 2, pp. 190–196, 1997.
[57] A. G. Yakovlev and A. I. Faden, “Sequential expression of c-fos protooncogene, TNF-α, and dynorphin genes in spinalcord following experimental traumatic injury,” Molecular andChemical Neuropathology, vol. 23, no. 2-3, pp. 179–190, 1994.
[58] C. X. Wang, B. Nuttin, H. Heremans, R. Dom, and J. Gybels,“Production of tumor necrosis factor in spinal cord followingtraumatic injury in rats,” Journal of Neuroimmunology, vol. 69,no. 1-2, pp. 151–156, 1996.
[59] A. D. Kraft, C. A. McPherson, and G. J. Harry, “Heterogeneityof microglia and TNF signaling as determinants for neuronaldeath or survival,” NeuroToxicology, vol. 30, no. 5, pp. 785–793,2009.
[60] H. S. Sharma, T. Winkler, E. Stalberg, T. Gordh, P. Alm, and J.Westman, “Topical application of TNF-α antiserum attenuatesspinal cord trauma induced edema formation, microvascularpermeability disturbances and cell injury in the rat,” ActaNeurochirurgica, Supplementum, no. 86, pp. 407–413, 2003.
[61] O. Nesic, G. Y. Xu, D. McAdoo, K. W. High, C. Hulsebosch, andR. Perez-Polo, “IL-1 receptor antagonist prevents apoptosisand caspase-3 activation after spinal cord injury,” Journal ofNeurotrauma, vol. 18, no. 9, pp. 947–956, 2001.
[62] T. F. Pais, C. Figueiredo, R. Peixoto, M. H. Braz, and S. Chat-terjee, “Necrotic neurons enhance microglial neurotoxicitythrough induction of glutaminase by a MyD88-dependentpathway,” Journal of Neuroinflammation, vol. 5, article 43,2008.
[63] D. A. Liebermann and B. Hoffman, “MyD genes in negativegrowth control,” Oncogene, vol. 17, no. 25, pp. 3319–3329,1998.
[64] S. Elkabes, E. M. DiCicco-Bloom, and I. B. Black, “Brainmicroglia/macrophages express neurotrophins that selectivelyregulate microglial proliferation and function,” Journal ofNeuroscience, vol. 16, no. 8, pp. 2508–2521, 1996.
[65] T. Miwa, S. Furukawa, K. Nakajima, Y. Furukawa, and S.Kohsaka, “Lipopolysaccharide enhances synthesis of brain-derived neurotrophic factor in cultured rat microglia,” Journalof Neuroscience Research, vol. 50, no. 6, pp. 1023–1029, 1997.
[66] R. Kiefer, T. Schweitzer, S. Jung, K. V. Toyka, and H. P. Har-tung, “Sequential expression of transforming growth factor-β1 by T-cells, macrophages, and microglia in rat spinal cordduring autoimmune inflammation,” Journal of Neuropathologyand Experimental Neurology, vol. 57, no. 5, pp. 385–395, 1998.
[67] U. K. Hanisch, “Microglia as a source and target of cytokines,”Glia, vol. 40, no. 2, pp. 140–155, 2002.
[68] A. Bessis, C. Bechade, D. Bernard, and A. Roumier, “Microglialcontrol of neuronal death and synaptic properties,” Glia, vol.55, no. 3, pp. 233–238, 2007.
[69] D. Giulian and C. Robertson, “Inhibition of mononuclearphagocytes reduces ischemic injury in the spinal cord,” Annalsof Neurology, vol. 27, no. 1, pp. 33–42, 1990.
[70] A. R. Blight, “Effects of silica on the outcome from exper-imental spinal cord injury: implication of macrophages insecondary tissue damage,” Neuroscience, vol. 60, no. 1, pp.263–273, 1994.
[71] P. G. Popovich, Z. Guan, P. Wei, I. Huitinga, N. van Rooijen,and B. T. Stokes, “Depletion of hematogenous macrophagespromotes partial hindlimb recovery and neuroanatomical
repair after experimental spinal cord injury,” ExperimentalNeurology, vol. 158, no. 2, pp. 351–365, 1999.
[72] D. Gris, D. R. Marsh, M. A. Oatway et al., “Transient blockadeof the CD11d/CD18 integrin reduces secondary damage afterspinal cord injury, improving sensory, autonomic, and motorfunction,” Journal of Neuroscience, vol. 24, no. 16, pp. 4043–4051, 2004.
[73] P. J. Mabon, L. C. Weaver, and G. A. Dekaban, “Inhibitionof monocyte/macrophage migration to a spinal cord injurysite by an antibody to the integrin αD: a potential new anti-inflammatory treatment,” Experimental Neurology, vol. 166,no. 1, pp. 52–64, 2000.
[74] M. Schwartz, “‘Tissue-repairing’ blood-derived macrophagesare essential for healing of the injured spinal cord: from skin-activated macrophages to infiltrating blood-derived cells?”Brain, Behavior, and Immunity, vol. 24, no. 7, pp. 1054–1057,2010.
[75] R. Shechter, A. London, C. Varol et al., “Infiltratingblood-derived macrophages are vital cells playing an anti-inflammatory role in recovery from spinal cord injury inmice,” PLoS Medicine, vol. 6, no. 7, Article ID e1000113, 2009.
[76] R. B. Banati, J. Gehrmann, P. Schubert, and G. W. Kreutzberg,“Cytotoxicity of microglia,” Glia, vol. 7, no. 1, pp. 111–118,1993.
[77] K. Satake, Y. Matsuyama, M. Kamiya et al., “Nitric oxidevia macrophage iNOS induces apoptosis following traumaticspinal cord injury,” Molecular Brain Research, vol. 85, no. 1-2,pp. 114–122, 2000.
[78] F. Bao, Y. Chen, G. A. Dekaban, and L. C. Weaver, “Earlyanti-inflammatory treatment reduces lipid peroxidation andprotein nitration after spinal cord injury in rats,” Journal ofNeurochemistry, vol. 88, no. 6, pp. 1335–1344, 2004.
[79] A. Bal-Price and G. C. Brown, “Inflammatory neurode-generation mediated by nitric oxide from activated glia-inhibiting neuronal respiration, causing glutamate release andexcitotoxicity,” Journal of Neuroscience, vol. 21, no. 17, pp.6480–6491, 2001.
[80] P. O. Scumpia, K. M. Kelly, W. H. Reeves, and B. R. Stevens,“Double-stranded RNA signals antiviral and inflammatoryprograms and dysfunctional glutamate transport in TLR3-expressing astrocytes,” Glia, vol. 52, no. 2, pp. 153–162, 2005.
[81] D. Liu, G. Y. Xu, E. Pan, and D. J. McAdoo, “Neurotoxicityof glutamate at the concentration released upon spinal cordinjury,” Neuroscience, vol. 93, no. 4, pp. 1383–1389, 1999.
[82] A. Vezzani, M. Conti, A. de Luigi et al., “Interleukin-1βimmunoreactivity and microglia are enhanced in the rathippocampus by focal kainate application: functional evidencefor enhancement of electrographic seizures,” Journal of Neuro-science, vol. 19, no. 12, pp. 5054–5065, 1999.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 459894, 8 pagesdoi:10.1155/2012/459894
Research Article
Neuroprotective Activity of Sibjeondaebo-tang on AβPeptide-Induced Damages
Hyeon Ju Yim,1 Jung Hwa Lim,2 Min Hee Kim,1 Uk Namgung,1
Sang Ryong Lee,1 and In Chul Jung1
1 Department of Oriental Medicine, Daejeon University, Daejeon 300-716, Republic of Korea2 Department of Neuropsychiatry, Pusan National University Korean Medical Hospital, Gyeongsangnam-do,Yangsan 626-789, Republic of Korea
Correspondence should be addressed to Sang Ryong Lee, [email protected] and In Chul Jung, [email protected]
Received 7 February 2012; Accepted 12 March 2012
Academic Editor: Monica Rosa Loizzo
Copyright © 2012 Hyeon Ju Yim et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Background. Sibjeondaebo-tang (SJDBT) has been used to treat diverse disorders including neuropsychiatric disabilities intraditional Korean medicine. Objective. The present study aims to investigate the potential effects of SJDBT on neuroprotectionagainst Aβ peptide-induced damage using in vitro culture and in vivo rat brain systems. Materials and Methods. PC12 cell viabilitywas analyzed by MTT assay, and neurite arborizations and caspase 3 protein signals in cultured PC12 cells and in vivo corticalneurons were analyzed by immunofluorescence staining. Phospho-Erk1/2 protein was analyzed by immunofluorescence stainingand western blot analysis. Results. In PC12 cells, atrophied cell body and reduced neurite extension by Aβ treatment were recoveredby SJDBT treatment. Caspase 3 protein signals were increased in Aβ-treated PC12 cells, but SJDBT treatment decreased apoptoticcell death. Caspase 3 activation in cortical neurons, which was induced similarly by Aβ treatment, was reduced by SJDBT treatment.Furthermore, phospho-Erk1/2 protein levels, which had been decreased by Aβ treatment, were elevated in the cortical neurons bySJDBT treatment. Conclusion. These data show that SJDBT may play a role in protecting from damages induced by Aβ in neuronaltissue and further suggest that SJDBT can be explored as the potential therapeutic target for AD treatments in human.
1. Introduction
Alzheimer’s disease (AD) is the neurodegenerative disease,which is most common in elderly individuals throughoutthe world, particularly in the developed countries [1].Extensive studies for the last several decades have maderemarkable progress in understanding pathophysiologicalbasis on AD [2, 3]. Accumulation of β amyloid (Aβ) peptideand formation of neurofibrillary tangles are the hallmarkof AD brain in human. Thus, secretase enzyme that isinvolved in the processing of Aβ peptide from amyloidprecursor protein (APP) is one of the major targets for ADstudy [4]. Aβ aggregates can alter synaptic transmission, andlarge aggregates are known to be toxic to neurons [5]. Inaddition, numerous studies using cultured cells showed thatthe treatment of Aβ peptide induced cell death even with alow concentration, suggesting that Aβ peptide molecule itselfmay act as a trigger inducing cell death pathway [6–8]. Thus,
Aβ peptide is the major target for mechanistic studies on ADin vivo and in vitro systems.
Sibjeondaebo-tang (SJDBT) is a noted prescription in(DongEuySuSeBoWon) [9], which describes the concept of“Sasang Constitutional Medicine”. The basic theory of SasangConstitutional Medicine consists of four types: Taeyangin,Tae-eumin, Soyangin, and Soeumin. Each of Sasang Con-stitutional types classified by similar patterns has differentcharacteristics, so there are different patterns of diseasesand treatment methods for each type. It was suggestedthat the proper therapy for Soeumin, for instance, is tokeep one’s body warm and strengthen the blood and qi,because their blood and qi are weak and stagnant [10].Lee [9] showed that SJDBT is applicable to treat Soeuminexterior diseases, because it might reinforce the healthy qito individuals who are lacking qi due to the pathogenicexposure or chronic disease [11]. Consistent with thisnotion, one recent study has demonstrated that SJDBT might

2 Evidence-Based Complementary and Alternative Medicine
participate in the replenishment of declined energy andcholinergic neurotransmitter synthesis in the mouse cerebralcortex with memory impairment [12].
In the present study, possible protective effects of SJDBTwere investigated in cultured PC12 cells and Aβ-injectedmouse. Our data indicate that SJDBT is effective to a certainlevel as a protective agent against Aβ peptide-mediatedtoxicity in neuronal cells.
2. Materials and Methods
2.1. Drugs and Chemicals. SJDBT prescription used inthe present study is composed of equal amounts of thefollowing herbal ingredients: Ginseng Radix (the manufac-ture’s serial number: BH0601, production area: Geumsan-gun, Chungnam, Korea), Cynanchi Wilfordii Radix (themanufacture’s serial number: BH0816, production area:Yeongcheon-si, Gyeongbuk, Korea), Cinnamomi Cortex Inte-rior (the manufacture’s serial number: SR-0033-10, pro-duction area: Vietnam), Astragali Radix (the manufac-ture’s serial number: BH0130, production area: Jecheon-si, Chungbuk, Korea), Atractylodis Macrocephalae Rhizoma(the manufacture’s serial number: BH0715, production area:Yeongcheon-si, Gyeongbuk, Korea), Angelicae Gigantis Radix(the manufacture’s serial number: BH0615, production area:Pyeongchang-gun, Gangwon, Korea), Cnidii Rhizoma (themanufacture’s serial number: BH0801, production area:Yeongyang-gun, Gyeongbuk, Korea), Paeoniae Radix Alba(the manufacture’s serial number: BH1030, productionarea: Jinju-si, Gyeongnam, Korea), Citri Pericarpium (themanufacture’s serial number: BH1210, production area: Jeju-do, Korea), Glycyrrhizae Radix (the manufacture’s serialnumber: HPL220C-00608, production area: China), Zin-giberis Rhizoma Recens (the manufacture’s serial number:BH0910, production area: Muju-gun, Jeonbuk, Korea), andZizyphi Fructus (the manufacture’s serial number: BH1205,production area: Gyeongsan-si, Gyeongbuk, Korea).
The drug was obtained from Oriental Medical Hospitalof Daejeon University Daejeon, Korea. SJDBT (45 g dryweight) was resuspended in 1 L of water, heat extractedfor 3 hrs, and filtered three times. The filtered fluid wasdistilled using the rotary vacuum evaporator (Buchi 461,Eyela, USA). Concentrated solution was frozen at −70◦C for4 hrs and freeze dried for 24 hrs. The product for SJDBTwas 14.7 g, with 29.2% yield from the initial raw materials.The product was kept at 4◦C and dissolved in water. Thestock solution (10 mg/mL in phosphate buffered saline) wasstored at −20◦C and used for experiment by diluting withphysiological saline solution.
β-amyloid peptide was purchased from Sigma-Aldrich(USA), dissolved in H2O to 1 mM, and kept at −20◦C untiluse. Galantamine triethiodide (Sigma, USA), a competitivereversible inhibitor of acetylcholinesterase (AChE) was alsopurchased from Sigma-Aldrich and diluted with H2O toconcentrations of 0.9 mg/mL and kept at −20◦C, freezeruntil use.
2.2. Experimental Animals. Albino mice (7–9-week old,Samtago, Korea) were used in the present study. Animals
were placed in an animal room with regulated temperature(22◦C), 50% of humidity, and 12-hr-light and 12-hr-darkcycle. They were allowed to eat commercial rat chow(Samyang Co., Korea) and drink water ad libitum. Allprocedures were in strict accordance with the US guidelines(NIH publication no. 85-23 revised in 1985) for the care anduse of laboratory animals and approved by the Committeeon Use of Live Animals for Teaching and Research at DaejeonUniversity.
2.3. PC12 Cell Culture. PC12 cells were cultured in DMEM(GIBCO, USA) supplemented with 5% FBS, 5% horse serum,and 1% penicillin/streptomycin at 37◦C incubator suppliedwith 5% CO2. Cells were grown at 75 cm2 flask (SPL, Korea)for 3 days and transferred into a new medium. Cells werewashed with phosphate buffered saline (PBS) and treatedwith trypsin-versene solution (Lonza, USA) at 37◦C for5 min for trituration. Detached cells were suspended withDMEM containing 5% FBS and 5% horse serum, platedinto a new culture flask with a 1 : 4 dilution, and incu-bated at a CO2 incubator. For immunofluorescence stainingexperiments, cells (1 × 105 per coverslips) were usuallycultured on the coverslips precoated with poly-L-ornithine(0.1 mg/mL, Sigma, USA) and laminin (0.02 mg/mL, Collab-orate Research, USA) overnight at room temperature.
2.4. MTT Assay. PC12 cell viability was assayed byreduction of MTT [3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide] reagent. Cells (1 × 105/well)were plated in 96-well plate. Cells were treated with 10 μMof Aβ peptide and with different concentrations of 0.3, 0.5,and 1.0 mg/mL of SJDBT for 24 hrs. Then, the cells weretreated with MTT solution for 4 hrs, and optical density wasmeasured using spectrophotometer at 570 nm. Cell viabilitywas measured as follows:
Cell viability (%)
= optical density of cells treated with drugsoptical density of cells treated with saline vehicle
× 100.
(1)
2.5. Immunofluorescence Staining and Hoechst Nuclear Stain-ing. For immunofluorescence staining, cells or tissues onthe coverslips were fixed with 4% paraformaldehyde 4%sucrose in PBS at room temperature for 40 min, permeabi-lized with 0.5% Nonidet P-40 in PBS, and blocked with2.5% horse serum and 2.5% bovine serum albumin for4 hrs at room temperature. Cells on the coverslips wereincubated with primary antibody, washed with PBST (PBSplus 0.1% triton X-100) 3 times for 10 min each, incubatedwith fluorescein-goat anti-mouse (1 : 400 dilution, Molecularprobes, USA) or rhodamine-goat anti-rabbit secondaryantibodies (Molecular probes) in 2.5% horse serum and2.5% bovine serum albumin for 1 hr at room temperature,and cover-slipped with gelatin mount medium. For someexperimental purpose, Hoechst staining reaction for nuclearvisualization was performed between washing steps aftersecondary antibody reaction. Tissue sections were treated

Evidence-Based Complementary and Alternative Medicine 3
with 25 μg/mL of Hoechst33258 in 0.1% triton X-100 inphosphate-buffered saline solution (PBST) for 10 min. Thesecondary antibody reaction was performed in a dark place.The merged images were produced by layer blending modeoptions of the Adobe Photoshop (version 5.5). The primaryantibodies were phospho-Erk1/2 kinase antibody (1 : 800,Cell Signaling, USA), cleaved caspase 3 antibody (1 : 500, CellSignaling), and monoclonal antineurofilament 200 (1 : 400,Sigma-Aldrich).
2.6. Western Blot Analysis. Cell or tissue lysates were washedwith ice-cold PBS and sonicated under 50–200μL of tritonlysis buffer (20 mM Tris, pH 7.4, 137 mM NaCl, 25 mM β-glycerophosphate, pH 7.14, 2 mM sodium pyrophosphate,2 mM EDTA, 1 mM Na3VO4, 1% triton X-100, 10% glycerol,5 μg/mL leupeptin, 5 μg/mL aprotinin, 3 μM benzamidine,0.5 mM DTT, and 1 mM PMSF). Protein (15 μg) was resolvedin 12% SDS polyacrylamide gel and transferred to immo-bilon polyvinylidene difluoride (PVDF) membranes (Milli-pore, Bedford, USA). Blots were blocked with 5% nonfatdry milk in PBST (17 mM KH2PO4, 50 mM Na2HPO4,1.5 mM NaCl, pH 7.4, and 0.05% Tween-20) for 1 hr at roomtemperature and then incubated overnight at 4◦C in 0.1%triton X-100 in PBS plus 5% nonfat dry milk containingantibodies. Protein bands were detected using the Amer-sham ECL kit (Amersham Pharmacia Biotech, Piscataway,USA), with horseradish peroxidase-conjugated secondarygoat anti-rabbit or goat anti-mouse antibodies (Transduc-tion Laboratories, Lexington, USA). Relative intensities ofthe protein bands were analyzed by autoradiography. Theantibodies used in the present study were phospho-p44/42Erk1/2 kinase antibody (1 : 4,000, cell signaling), p44/42Erk1/2 kinase antibody (1 : 4,000, cell signaling), and cleavedcaspase-3 antibody (1 : 1,000, cell signaling).
2.7. Aβ-Administered Mouse Model. Albino ICR mice wererandomly assigned into (i) normal group, (ii) Aβ peptide-treated control group, (iii) positive control group treatedwith Aβ peptide plus galantamine, and (iv) experimentgroup treated with Aβ peptide and SJDBT. 5 μL of Aβ peptide(200 pmol) was microinjected using the glass capillaryconnected to picoinjector (Harvard Instrument, USA) intothe lateral ventricle (0.5 mm posterior to bregma, 1.5 mmlateral to midline, and 2.5 mm ventral to the brain sur-face) bilaterally. SJDBT extract (400 mg/kg) was orally (Po)administered when Aβ peptide was given, and 8 days afterAβ treatment, galantamine (3 mg/kg) was administered (Ip)on a daily basis for 3 days. The brain was then dissected, andcoronal sections (20 μm) were prepared using the cryostat forhistological analysis.
2.8. Microscopic Analysis. Images from immunofluorescencestaining were analyzed by fluorescence microscope (Nikon,Japan), and real-time images of cultured cells were analyzedby phase-contrast microscope. The images were capturedand transferred into the computer software (ACT-1). Mergedimages were analyzed under the image blend mode of theAdobe Photoshop software (version 5.5).
2.9. Statistical Analysis. Data were presented as mean ±standard error of mean (SEM). A StatView512+ computersoftware was used for statistical analysis by Student’s t-test.Statistically significant differences were reported as P < 0.05,P < 0.01, or P < 0.001 (see Figure 1(a)).
3. Results
3.1. Effects of SJDBT on Aβ-Treated PC12 Cells. To determinepossible neuroprotective activity of SJDBT in PC12 cellstreated with Aβ peptide, cell survival was measured by MTTassay. The levels of cell survival, when treated with Aβ pep-tide, were strongly decreased as much as 40% to those of nor-mal group (Figure 1(a)). Cotreatment with SJDBT extract atconcentrations of 0.3–0.5 mg/mL enhanced levels of cell sur-vival higher than 80%. However, SJDBT extract at 1.0 mg/mLslightly decreased cell survival. Thus, SJDBT extract at 0.3–0.5 mg/mL was used for the rest of the present study. Todetermine whether apoptotic signaling pathway was involvedin Aβ-induced cell death, caspase 3 activation was measuredin PC12 cells. Caspase 3 protein signals, which were notdetected in intact PC12 cells, were induced in some of the Aβ-treated cells (Figure 1(b)). It was observed that caspase 3 pro-tein signals were not localized at the central area where thenucleus was identified by Hoechst staining. In cells treatedwith SJDBT at 0.3 mg/mL, caspase 3 signals were observed inless cells compared to those in Aβ-treated cells. Furthermore,in cells treated with Aβ and 0.5 mg/mL of SJDBT, caspase 3signals were not detected in any of cultured cells.
Effects of SJDBT on morphological changes in Aβ-treated PC12 cells were investigated. In NGF-treated PC12cells, the cell body showed a morphology of spreadingand round shapes, and neurite processes were clearlyobserved (Figure 2(a)). However, cells treated with Aβrevealed shrunken cell body with no clear neurite processes.When the cells were treated with SJDBT extract, neuritegrowth processes were observed, and particularly longerprocesses were frequently observed with SJDBT treatment at0.5 mg/mL. To further examine the pattern of neurite out-growth, cells were subjected to immunofluorescence stainingwith anti-NF-200 antibody. While some neurite growthprocesses were observed in NF-200-stained normal cells, Aβtreatment resulted in overall weak NF-200 staining in PC12cells with no clear neurite processes. SJDBT treatment at0.3 mg/mL strongly induced NF-200 staining in culturedcells and showed distinct neurite processes (Figure 2(b)).Longer extension of neurite processes were observed with0.5 mg/mL of SJDBT extract though overall staining intensitywas decreased compared to those treated with 0.3 mg/mL ofSJDBT.
3.2. Effects of SJDBT on Cortical Neurons Treated with Aβ Pep-tide. To determine possible neuroprotective effects of SJDBTon Aβ-treated cerebral cortical neurons, levels of caspase 3protein were analyzed. As shown in Figure 3(a), caspase 3signals were not detected in normal tissue. Then, caspase 3-positive signals were identified in the brain tissue treated withAβ peptide though the signals were limited to a few numbersof the cells. Treatment of SJDBT or galantamine as a positive
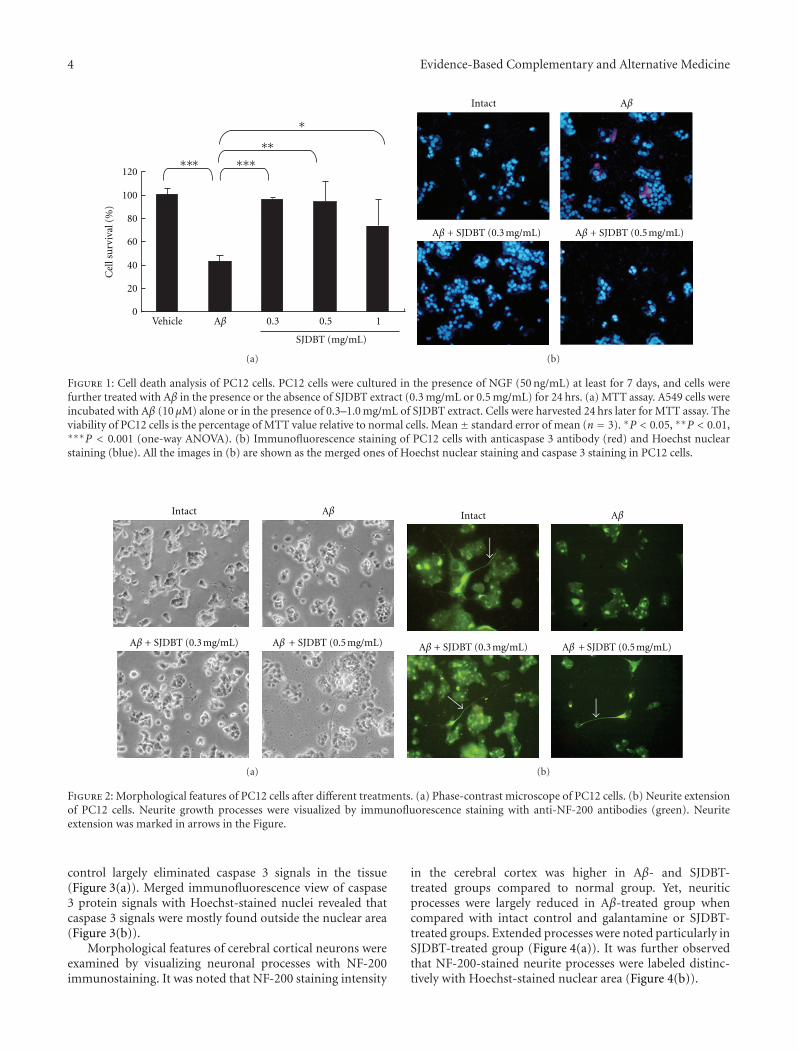
4 Evidence-Based Complementary and Alternative Medicine
120
100
80
60
40
20
0
Cel
l su
rviv
al (
%)
∗∗∗
∗∗∗∗∗∗
0.3 0.5 1Vehicle
SJDBT (mg/mL)
Aβ
(a)
Intact Aβ
Aβ + SJDBT (0.3 mg/mL) Aβ + SJDBT (0.5 mg/mL)
(b)
Figure 1: Cell death analysis of PC12 cells. PC12 cells were cultured in the presence of NGF (50 ng/mL) at least for 7 days, and cells werefurther treated with Aβ in the presence or the absence of SJDBT extract (0.3 mg/mL or 0.5 mg/mL) for 24 hrs. (a) MTT assay. A549 cells wereincubated with Aβ (10 μM) alone or in the presence of 0.3–1.0 mg/mL of SJDBT extract. Cells were harvested 24 hrs later for MTT assay. Theviability of PC12 cells is the percentage of MTT value relative to normal cells. Mean± standard error of mean (n = 3). ∗P < 0.05, ∗∗P < 0.01,∗∗∗P < 0.001 (one-way ANOVA). (b) Immunofluorescence staining of PC12 cells with anticaspase 3 antibody (red) and Hoechst nuclearstaining (blue). All the images in (b) are shown as the merged ones of Hoechst nuclear staining and caspase 3 staining in PC12 cells.
Intact Aβ
Aβ + SJDBT (0.3 mg/mL) Aβ + SJDBT (0.5 mg/mL)
(a)
Intact Aβ
Aβ + SJDBT (0.3 mg/mL) Aβ + SJDBT (0.5 mg/mL)
(b)
Figure 2: Morphological features of PC12 cells after different treatments. (a) Phase-contrast microscope of PC12 cells. (b) Neurite extensionof PC12 cells. Neurite growth processes were visualized by immunofluorescence staining with anti-NF-200 antibodies (green). Neuriteextension was marked in arrows in the Figure.
control largely eliminated caspase 3 signals in the tissue(Figure 3(a)). Merged immunofluorescence view of caspase3 protein signals with Hoechst-stained nuclei revealed thatcaspase 3 signals were mostly found outside the nuclear area(Figure 3(b)).
Morphological features of cerebral cortical neurons wereexamined by visualizing neuronal processes with NF-200immunostaining. It was noted that NF-200 staining intensity
in the cerebral cortex was higher in Aβ- and SJDBT-treated groups compared to normal group. Yet, neuriticprocesses were largely reduced in Aβ-treated group whencompared with intact control and galantamine or SJDBT-treated groups. Extended processes were noted particularly inSJDBT-treated group (Figure 4(a)). It was further observedthat NF-200-stained neurite processes were labeled distinc-tively with Hoechst-stained nuclear area (Figure 4(b)).
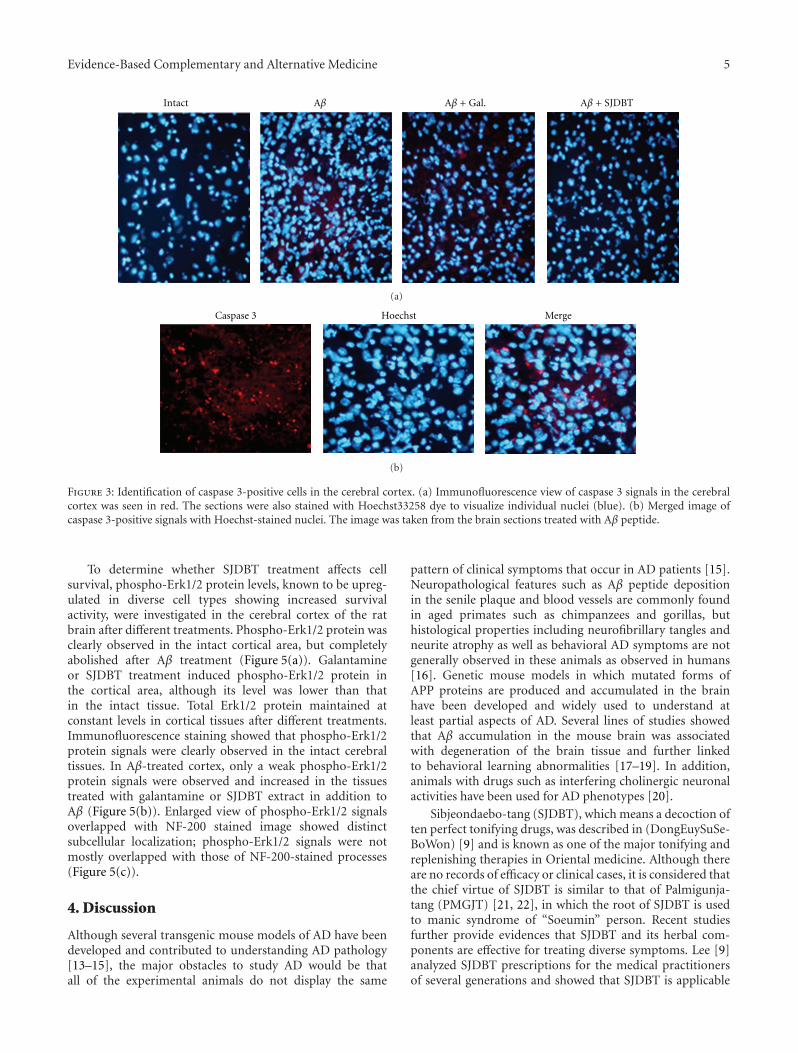
Evidence-Based Complementary and Alternative Medicine 5
Intact Aβ Aβ + Gal. Aβ + SJDBT
(a)
MergeHoechstCaspase 3
(b)
Figure 3: Identification of caspase 3-positive cells in the cerebral cortex. (a) Immunofluorescence view of caspase 3 signals in the cerebralcortex was seen in red. The sections were also stained with Hoechst33258 dye to visualize individual nuclei (blue). (b) Merged image ofcaspase 3-positive signals with Hoechst-stained nuclei. The image was taken from the brain sections treated with Aβ peptide.
To determine whether SJDBT treatment affects cellsurvival, phospho-Erk1/2 protein levels, known to be upreg-ulated in diverse cell types showing increased survivalactivity, were investigated in the cerebral cortex of the ratbrain after different treatments. Phospho-Erk1/2 protein wasclearly observed in the intact cortical area, but completelyabolished after Aβ treatment (Figure 5(a)). Galantamineor SJDBT treatment induced phospho-Erk1/2 protein inthe cortical area, although its level was lower than thatin the intact tissue. Total Erk1/2 protein maintained atconstant levels in cortical tissues after different treatments.Immunofluorescence staining showed that phospho-Erk1/2protein signals were clearly observed in the intact cerebraltissues. In Aβ-treated cortex, only a weak phospho-Erk1/2protein signals were observed and increased in the tissuestreated with galantamine or SJDBT extract in addition toAβ (Figure 5(b)). Enlarged view of phospho-Erk1/2 signalsoverlapped with NF-200 stained image showed distinctsubcellular localization; phospho-Erk1/2 signals were notmostly overlapped with those of NF-200-stained processes(Figure 5(c)).
4. Discussion
Although several transgenic mouse models of AD have beendeveloped and contributed to understanding AD pathology[13–15], the major obstacles to study AD would be thatall of the experimental animals do not display the same
pattern of clinical symptoms that occur in AD patients [15].Neuropathological features such as Aβ peptide depositionin the senile plaque and blood vessels are commonly foundin aged primates such as chimpanzees and gorillas, buthistological properties including neurofibrillary tangles andneurite atrophy as well as behavioral AD symptoms are notgenerally observed in these animals as observed in humans[16]. Genetic mouse models in which mutated forms ofAPP proteins are produced and accumulated in the brainhave been developed and widely used to understand atleast partial aspects of AD. Several lines of studies showedthat Aβ accumulation in the mouse brain was associatedwith degeneration of the brain tissue and further linkedto behavioral learning abnormalities [17–19]. In addition,animals with drugs such as interfering cholinergic neuronalactivities have been used for AD phenotypes [20].
Sibjeondaebo-tang (SJDBT), which means a decoction often perfect tonifying drugs, was described in (DongEuySuSe-BoWon) [9] and is known as one of the major tonifying andreplenishing therapies in Oriental medicine. Although thereare no records of efficacy or clinical cases, it is considered thatthe chief virtue of SJDBT is similar to that of Palmigunja-tang (PMGJT) [21, 22], in which the root of SJDBT is usedto manic syndrome of “Soeumin” person. Recent studiesfurther provide evidences that SJDBT and its herbal com-ponents are effective for treating diverse symptoms. Lee [9]analyzed SJDBT prescriptions for the medical practitionersof several generations and showed that SJDBT is applicable
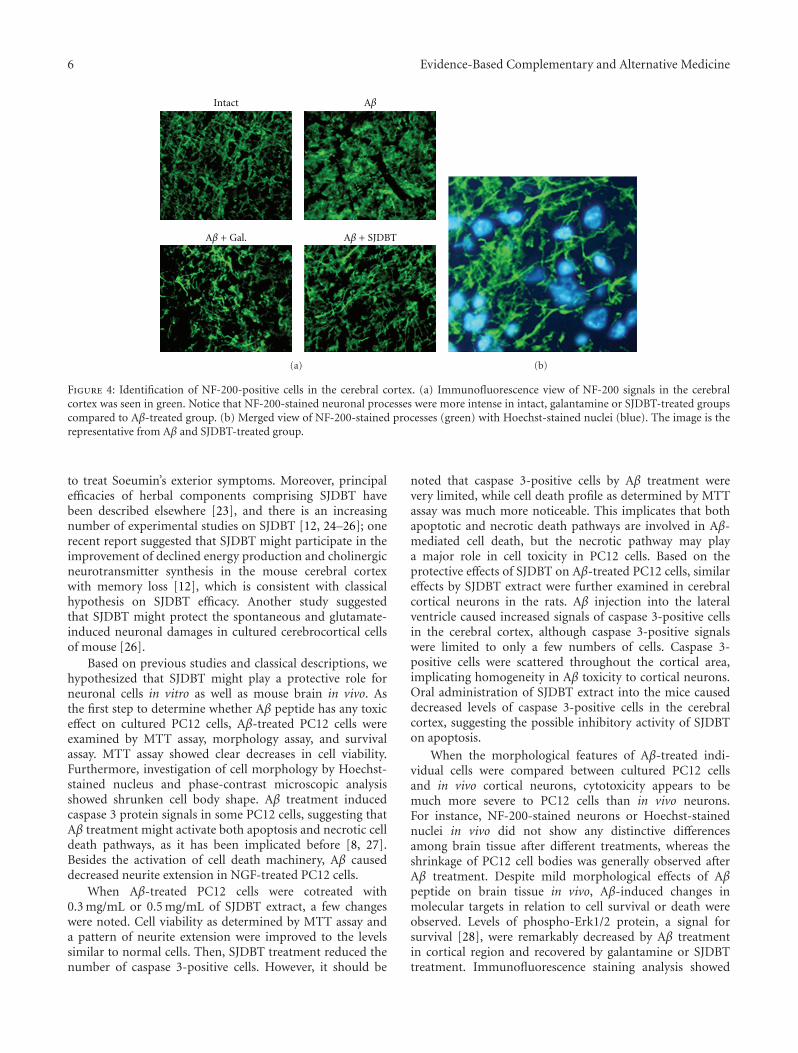
6 Evidence-Based Complementary and Alternative Medicine
Intact Aβ
Aβ + Gal. Aβ + SJDBT
(a) (b)
Figure 4: Identification of NF-200-positive cells in the cerebral cortex. (a) Immunofluorescence view of NF-200 signals in the cerebralcortex was seen in green. Notice that NF-200-stained neuronal processes were more intense in intact, galantamine or SJDBT-treated groupscompared to Aβ-treated group. (b) Merged view of NF-200-stained processes (green) with Hoechst-stained nuclei (blue). The image is therepresentative from Aβ and SJDBT-treated group.
to treat Soeumin’s exterior symptoms. Moreover, principalefficacies of herbal components comprising SJDBT havebeen described elsewhere [23], and there is an increasingnumber of experimental studies on SJDBT [12, 24–26]; onerecent report suggested that SJDBT might participate in theimprovement of declined energy production and cholinergicneurotransmitter synthesis in the mouse cerebral cortexwith memory loss [12], which is consistent with classicalhypothesis on SJDBT efficacy. Another study suggestedthat SJDBT might protect the spontaneous and glutamate-induced neuronal damages in cultured cerebrocortical cellsof mouse [26].
Based on previous studies and classical descriptions, wehypothesized that SJDBT might play a protective role forneuronal cells in vitro as well as mouse brain in vivo. Asthe first step to determine whether Aβ peptide has any toxiceffect on cultured PC12 cells, Aβ-treated PC12 cells wereexamined by MTT assay, morphology assay, and survivalassay. MTT assay showed clear decreases in cell viability.Furthermore, investigation of cell morphology by Hoechst-stained nucleus and phase-contrast microscopic analysisshowed shrunken cell body shape. Aβ treatment inducedcaspase 3 protein signals in some PC12 cells, suggesting thatAβ treatment might activate both apoptosis and necrotic celldeath pathways, as it has been implicated before [8, 27].Besides the activation of cell death machinery, Aβ causeddecreased neurite extension in NGF-treated PC12 cells.
When Aβ-treated PC12 cells were cotreated with0.3 mg/mL or 0.5 mg/mL of SJDBT extract, a few changeswere noted. Cell viability as determined by MTT assay anda pattern of neurite extension were improved to the levelssimilar to normal cells. Then, SJDBT treatment reduced thenumber of caspase 3-positive cells. However, it should be
noted that caspase 3-positive cells by Aβ treatment werevery limited, while cell death profile as determined by MTTassay was much more noticeable. This implicates that bothapoptotic and necrotic death pathways are involved in Aβ-mediated cell death, but the necrotic pathway may playa major role in cell toxicity in PC12 cells. Based on theprotective effects of SJDBT on Aβ-treated PC12 cells, similareffects by SJDBT extract were further examined in cerebralcortical neurons in the rats. Aβ injection into the lateralventricle caused increased signals of caspase 3-positive cellsin the cerebral cortex, although caspase 3-positive signalswere limited to only a few numbers of cells. Caspase 3-positive cells were scattered throughout the cortical area,implicating homogeneity in Aβ toxicity to cortical neurons.Oral administration of SJDBT extract into the mice causeddecreased levels of caspase 3-positive cells in the cerebralcortex, suggesting the possible inhibitory activity of SJDBTon apoptosis.
When the morphological features of Aβ-treated indi-vidual cells were compared between cultured PC12 cellsand in vivo cortical neurons, cytotoxicity appears to bemuch more severe to PC12 cells than in vivo neurons.For instance, NF-200-stained neurons or Hoechst-stainednuclei in vivo did not show any distinctive differencesamong brain tissue after different treatments, whereas theshrinkage of PC12 cell bodies was generally observed afterAβ treatment. Despite mild morphological effects of Aβpeptide on brain tissue in vivo, Aβ-induced changes inmolecular targets in relation to cell survival or death wereobserved. Levels of phospho-Erk1/2 protein, a signal forsurvival [28], were remarkably decreased by Aβ treatmentin cortical region and recovered by galantamine or SJDBTtreatment. Immunofluorescence staining analysis showed
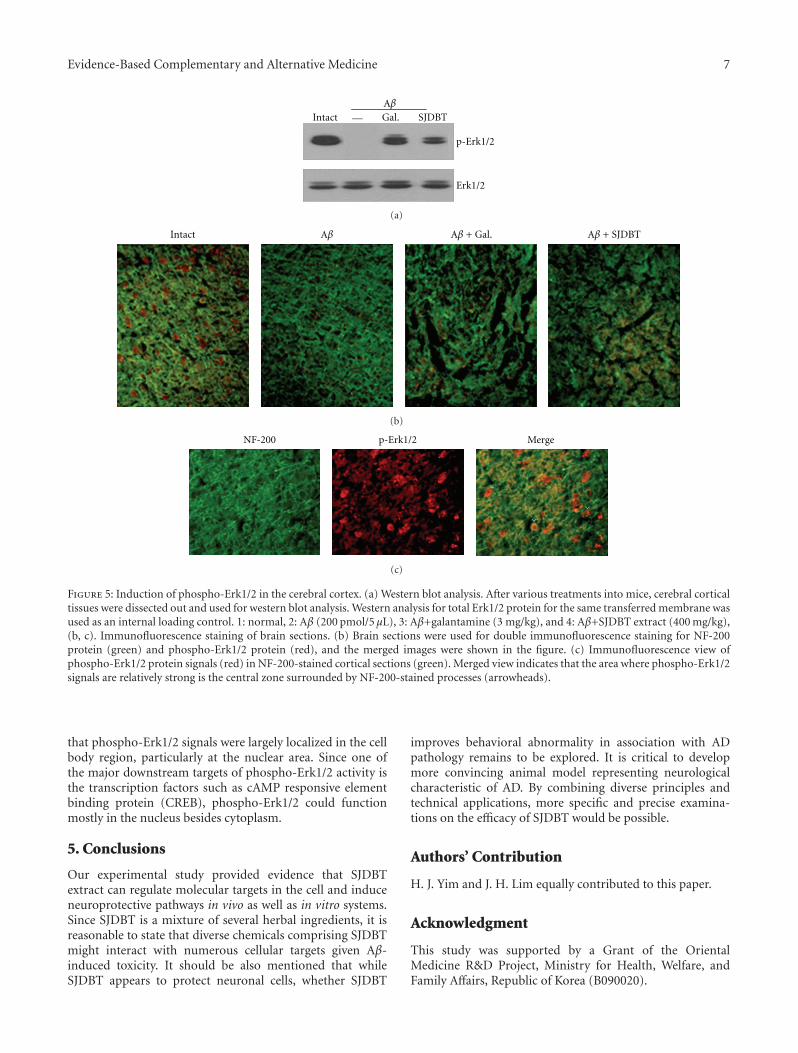
Evidence-Based Complementary and Alternative Medicine 7
IntactAβGal. SJDBT
p-Erk1/2
Erk1/2
—
(a)
Intact Aβ Aβ + Gal. Aβ + SJDBT
(b)
p-Erk1/2 MergeNF-200
(c)
Figure 5: Induction of phospho-Erk1/2 in the cerebral cortex. (a) Western blot analysis. After various treatments into mice, cerebral corticaltissues were dissected out and used for western blot analysis. Western analysis for total Erk1/2 protein for the same transferred membrane wasused as an internal loading control. 1: normal, 2: Aβ (200 pmol/5 μL), 3: Aβ+galantamine (3 mg/kg), and 4: Aβ+SJDBT extract (400 mg/kg),(b, c). Immunofluorescence staining of brain sections. (b) Brain sections were used for double immunofluorescence staining for NF-200protein (green) and phospho-Erk1/2 protein (red), and the merged images were shown in the figure. (c) Immunofluorescence view ofphospho-Erk1/2 protein signals (red) in NF-200-stained cortical sections (green). Merged view indicates that the area where phospho-Erk1/2signals are relatively strong is the central zone surrounded by NF-200-stained processes (arrowheads).
that phospho-Erk1/2 signals were largely localized in the cellbody region, particularly at the nuclear area. Since one ofthe major downstream targets of phospho-Erk1/2 activity isthe transcription factors such as cAMP responsive elementbinding protein (CREB), phospho-Erk1/2 could functionmostly in the nucleus besides cytoplasm.
5. Conclusions
Our experimental study provided evidence that SJDBTextract can regulate molecular targets in the cell and induceneuroprotective pathways in vivo as well as in vitro systems.Since SJDBT is a mixture of several herbal ingredients, it isreasonable to state that diverse chemicals comprising SJDBTmight interact with numerous cellular targets given Aβ-induced toxicity. It should be also mentioned that whileSJDBT appears to protect neuronal cells, whether SJDBT
improves behavioral abnormality in association with ADpathology remains to be explored. It is critical to developmore convincing animal model representing neurologicalcharacteristic of AD. By combining diverse principles andtechnical applications, more specific and precise examina-tions on the efficacy of SJDBT would be possible.
Authors’ Contribution
H. J. Yim and J. H. Lim equally contributed to this paper.
Acknowledgment
This study was supported by a Grant of the OrientalMedicine R&D Project, Ministry for Health, Welfare, andFamily Affairs, Republic of Korea (B090020).

8 Evidence-Based Complementary and Alternative Medicine
References
[1] L. M. Bierer, V. Haroutunian, S. Gabriel et al., “Neurochemicalcorrelates of dementia severity in Alzheimer’s disease: relativeimportance of the cholinergic deficits,” Journal of Neurochem-istry, vol. 64, no. 2, pp. 749–760, 1995.
[2] K. R. Brunden, J. Q. Trojanowski, and V. M. Y. Lee, “Advancesin tau-focused drug discovery for Alzheimer’s disease andrelated tauopathies,” Nature Reviews Drug Discovery, vol. 8, no.10, pp. 783–793, 2009.
[3] C. A. Lemere, “Developing novel immunogens for an effective,safe Alzheimer’s disease vaccine,” Neurodegenerative Diseases,vol. 2, no. 5, pp. 267–272, 2006.
[4] S. E. Gandy, J. D. Buxbaum, T. Suzuki et al., “The nature andmetabolism of potentially amyloidogenic carboxyl-terminalfragments of the Alzheimer β/A4-amyloid precursor protein:some technical notes,” Neurobiology of Aging, vol. 13, no. 5,pp. 601–603, 1992.
[5] T. A. Kokjohn and A. E. Roher, “Amyloid precursor proteintransgenic mouse models and Alzheimer’s disease: under-standing the paradigms, limitations, and contributions,”Alzheimer’s and Dementia, vol. 5, no. 4, pp. 340–347, 2009.
[6] S. J. Choi, C. H. Jeong, S. G. Choi et al., “Zeatin preventsamyloid β-induced neurotoxicity and scopolamine-inducedcognitive deficits,” Journal of Medicinal Food, vol. 12, no. 2, pp.271–277, 2009.
[7] W. W. Smith, D. D. Norton, M. Gorospe et al., “Phosphory-lation of p66Shc and forkhead proteins mediates Aβ toxicity,”Journal of Cell Biology, vol. 169, no. 2, pp. 331–339, 2005.
[8] A. Eckert, C. A. Marques, U. Keil, K. Schussel, and W. E.Muller, “Increased apoptotic cell death in sporadic and geneticAlzheimer’s disease,” Annals of the New York Academy ofSciences, vol. 1010, pp. 604–609, 2003.
[9] J. M. Lee, DongEuySuSeBoWon, Haingrim Press, Seoul, Korea,1986.
[10] I. B. Song, J. K. Kim, and M. H. Huh, An Introduction to SasangConstitutional Medicine, Jimoondang, Seoul, Korea, 2005.
[11] B. H. Koh and I. B. Song, “An experimental study on theeffects of deer horn, rehmannia glutiosa, panax ginseng andacanthopanax sessiliflorum on immune reaponse & nk cellactivity in mice,” Journal of Korean Oriental Medicine, vol. 7,pp. 157–173, 1986.
[12] J. Y. Ma, J. M. Yang, H. J. Joo, M. Y. Lee, J. H. Park, and K.Y. Jung, “Improvement of cerebral ATP and choline deficien-cies by Shao-Yin-Ren Shi-Quang-Da-Bu-Tang in senescence-accelerated mouse prone 8,” Journal of Ethnopharmacology,vol. 67, no. 3, pp. 297–305, 1999.
[13] D. Quon, Y. Wang, R. Catalano, J. M. Scardina, K. Murakami,and B. Cordell, “Formation of β-amyloid protein deposits inbrains of transgenic mice,” Nature, vol. 352, no. 6332, pp. 239–241, 1991.
[14] D. Games, D. Adams, R. Alessandrini et al., “Alzheimer-typeneuropathology in transgenic mice overexpressing V717F β-amyloid precursor protein,” Nature, vol. 373, no. 6514, pp.523–527, 1995.
[15] K. K. Hsiao, D. R. Borchelt, K. Olson et al., “Age-related CNSdisorder and early death in transgenic FVB/N mice overex-pressing Alzheimer amyloid precursor proteins,” Neuron, vol.15, no. 5, pp. 1203–1218, 1995.
[16] M. Gearing, G. W. Rebeck, B. T. Hyman, J. Tigges, and S. S.Mirra, “Neuropathology and apolipoprotein E profile of agedchimpanzees: implications for Alzheimer disease,” Proceedingsof the National Academy of Sciences of the United States ofAmerica, vol. 91, no. 20, pp. 9382–9386, 1994.
[17] C. Janus and D. Westaway, “Transgenic mouse models ofAlzheimer’s disease,” Physiology and Behavior, vol. 73, no. 5,pp. 873–886, 2001.
[18] L. Gimenez-Llort, G. Blazquez, T. Canete et al., “Modelingbehavioral and neuronal symptoms of Alzheimer’s diseasein mice: a role for intraneuronal amyloid,” Neuroscience andBiobehavioral Reviews, vol. 31, no. 1, pp. 125–147, 2007.
[19] J. C. Dodart, C. Mathis, K. R. Bales, and S. M. Paul, “Does mymouse have Alzheimer’s disease?” Genes, Brain and Behavior,vol. 1, no. 3, pp. 142–155, 2002.
[20] A. Bellucci, I. Luccarini, C. Scali et al., “Cholinergic dysfunc-tion, neuronal damage and axonal loss in TgCRND8 mice,”Neurobiology of Disease, vol. 23, no. 2, pp. 260–272, 2006.
[21] I. S. Park, DongEuySaSangYoGyul, vol. 198, Sonamoo, Seoul,Korea, 1992.
[22] J. S. Won, JeungBoDongEuySaSangShinPyun, Tonghab-euywonsa, Seoul, Korea, 1974.
[23] S. I. Lee, Herbal Medicine, Sungbo-sa, Seoul, Korea, 1975.[24] H. S. Kim, “A study on the effect of SibZunDaeBoTang
administered in rabbit with hepatic disfunction on the rate ofrecovery,” Journal of Korean Oriental Internal Medicine, vol. 1,pp. 78–84, 1976.
[25] Y. G. Chang, I. B. Song, and B. H. Ko, “The experimental studyof soum-in sipjondaebotang and gukbang sipjondaebotang onyang insufficient syndrome,” Journal of Sasang ConstitutionalMedicine, vol. 7, pp. 281–293, 1995.
[26] M. Y. Lee, T. Y. Ma, Y .K. Choo, and K. Y. Jung, “Protectionof spontaneous and glutamate-induced neuronal damagesby Soeumin Sibjeundaibo-tang and Soyangin Sibimijihwang-tang in cultured mice cerebrocortical cells,” Oriental Pharmacyand Experimental Medicine, vol. 1, pp. 55–63, 2000.
[27] R. Vassar, “Caspase-3 cleavage of GGA3 stabilizes BACE:implications for Alzheimer’s disease,” Neuron, vol. 54, no. 5,pp. 671–673, 2007.
[28] Z. Xia, M. Dickens, J. Raingeaud, R. J. Davis, and M. E.Greenberg, “Opposing effects of ERK and JNK-p38 MAPkinases on apoptosis,” Science, vol. 270, no. 5240, pp. 1326–1331, 1995.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 946259, 8 pagesdoi:10.1155/2012/946259
Review Article
Centella asiatica (L.) Urban: From Traditional Medicine toModern Medicine with Neuroprotective Potential
Ilkay Erdogan Orhan1, 2
1 Department of Pharmacognosy, Faculty of Pharmacy, Gazi University, 06330 Ankara, Turkey2 Department of Pharmacognosy and Pharmaceutical Botany, Faculty of Pharmacy, Eastern Mediterranean University,Gazimagosa, (Famagusta), Cyprus
Correspondence should be addressed to Ilkay Erdogan Orhan, [email protected]
Received 14 January 2012; Revised 27 February 2012; Accepted 6 March 2012
Academic Editor: Mahmud Tareq Hassan Khan
Copyright © 2012 Ilkay Erdogan Orhan. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
This paper covers the studies relevant to neuroprotective activity of Centella asiatica (L.) Urban, also known as “Gotu Kola.” Theplant is native to the Southeast Asia and has been used traditionally as brain tonic in ayurvedic medicine. The neuroprotectiveeffect of C. asiatica has been searched using the key words “Centella, Centella asiatica, gotu kola, Asiatic pennywort, neuroprotection,and memory” through the electronic databases including Sciencedirect, Web of Science, Scopus, Pubmed, and Google Scholar.According to the literature survey, C. asiatica (gotu kola) has been reported to have a comprehensive neuroprotection bydifferent modes of action such as enzyme inhibition, prevention of amyloid plaque formation in Alzheimer’s disease, dopamineneurotoxicity in Parkinson’s disease, and decreasing oxidative stress. Therefore, C. asiatica could be suggested to be a desiredphytopharmaceutical with neuroprotective effect emerged from traditional medicine.
1. Introduction
Centella asiatica (L.) Urban (Syn. Centella coriacea Nannfd.,Hydrocotyle asiatica L., Hydrocotyle lunata Lam., and Trisan-thus cochinchinensis Lour.) is a tropical medicinal plant fromApiaceae family native to Southeast Asian countries suchas India, Sri Lanka, China, Indonesia, and Malaysia as wellas South Africa and Madagascar [1]. C. asiatica, commonlyknown as “Gotu kola, Asiatic pennywort, Indian pennywort,Indian water navelwort, wild violet, and tiger herb” in Eng-lish, is a tropical plant, which has been also cultivated suc-cessfully due to its medical importance in some countriesincluding Turkey, and it has a long history of utilization inayurvedic and Chinese traditional medicines since centuries[2]. The leaves, which are edible, are in yellowish-green color,thin, alternate with long petioles, and quite characteristicreniform, orbicular, or oblong-elliptic shapes with sevenveins [3] (Figure 1). The plant grows horizontally throughits green to red stolones which combine to each other androots in underground. Monographs of the plant describingmainly its wound healing and memory enhancement effects
exist in the European Pharmacopeia, Commission E of theGerman Ministry of Health, and World Health Organization(WHO) [4]. In addition to neuroprotective effect of C. asi-atica, it has been reported to own a wide range of biologicalactivities desired for human health such as wound healing[5–7], anti-inflammatory [8, 9], antipsoriatic [10], antiulcer[11, 12], hepatoprotective [13], anticonvulsant [14], sedative[15], immunostimulant [16], cardioprotective [17, 18],antidiabetic [19], cytotoxic and antitumor [20, 21], antiviral[22], antibacterial [23], insecticidal [24], antifungal [25],antioxidant [26–28], and for lepra [29] and venous defi-ciency treatments [30, 31].
Numerous preparations of this plant in various phar-maceutical forms recommended for several indications in-cluding neurological disorders are available allover the world.Taking this fact into consideration, many researchers havefocused on neuroprotective effect of C. asiatica in orderto confirm its traditional use on scientific base. For thispurpose, a literature survey has been performed using thedatabases searched up to the year 2012 for the latestinformation on C. asiatica. This paper aims to cover up

2 Evidence-Based Complementary and Alternative Medicine
Figure 1: Centella asiatica (L.) Urban (Apiaceae).
HO
HO
O
OHOH
OH
R2
R2
R1
R1
OR3
R3
H H
HH
H
CH2OHCH2OH
CH2OHCH2OH
CH3
Asiatic acidMadecassic acidMadasiatic acidAsiaticosideMadecassoside
Glucose-glucose-rhamnoseGlucose-glucose-rhamnose
Figure 2: The major triterpene saponoside derivatives found inCentella asiatica.
in vitro, in vivo, and clinical studies reporting the results rele-vant to neuroprotective effect of this plant.
2. Phytochemical Content of C. asiatica
C. asiatica has been reported to contain a vast numberof compounds belonging to different chemical classes. Themajor chemical class found in this plant is triterpene sapono-sides. The major ones are known as asiatic acid, madecassicacid (6-hydroxy-asiatic acid), asiaticoside, madecassoside,and madasiatic acid (Figure 2), betulinic acid, thankunicacid, and isothankunic acid [32, 33]. Moreover, There aresome other triterpenes such as brahmic acid, centellin,centellicin, asiaticin, bayogenin, terminolic acid, 3β,6β,23-trihydroxyolean-12-en-28-oic acid, 3β,6β,23-trihydroxyurs-12-en-28-oic acid, 3-O-[α-L-arabinofuranosyl] 2α,3β,6β,23-α tetrahydroxyurs-12-en-28-oic acid, centellasapogenol A,
HO O
O
O
O
OH
OH
OH
Figure 3: Castilliferol.
HO
HO
HO
O
O
O
OOH
OH
OH
COOH
Figure 4: Isochlorogenic acid.
centellasaponins A-D, ursolic acid, pomolic acid, 3-epimas-linic acid, 23-O-acetylmadecassoside, and 23-O-acetylasiat-icoside B [34–41].
Presence of several flavonoid derivatives such as quer-cetin, kaempferol, patuletin, rutin, apigenin, castilliferol(Figure 3), castillicetin, and myricetin has been reported inC. asiatica [35, 39, 42], while isolation of polysaccharides(e.g., centellose) [43], polyacetylenes (e.g., cadinol, acetox-ycentellinol, centellin, centellicin, and asiaticin) [36, 44],sterols (e.g., 11-oxoheneicosanil-cyclohexane, dotriacont-8-en-1-oic acid, sitosterol 3-O-β-glucoside, stigmasterol 3-O-β-glucoside, and castasterone) [41, 45, 46], and phenolicacids (e.g., rosmarinic acid, 3,5-di-O-caffeoil quinic acid, 1,5-di-O-caffeoil quinic acid, 3,4-di-O-caffeoil quinic acid, 4,5-di-O-caffeoil quinic acid, ettacrynic acid, chlorogenic acid,and isochlorogenic acid [Figure 4]) [40, 42, 47] has beenalso identified in this species. In our quantitative study on C.asiatica of Turkish origin by HPLC, we reported existence ofseveral phenolic acids, for example, p-hydroxybenzoic acid,vanillic acid, p-coumaric acid, o-coumaric acid, and trans-cinnamic acid [48].
On the other hand, only a few studies have describedthe chemical composition of the essential oils obtained fromC. asiatica from Japan, South Africa, and Thailand, whichmainly consisted of monoterpene and sesquiterpene deriva-tives [49–51]. In our work, we examined the essential oilcomposition of C. asiatica cultivated in Turkey by GC-MSfor the first time and identified α-copaene as the majorcomponent [48].
3. Neuroprotective Activity of C. asiatica
3.1. In Vitro Studies. C. asiatica (gotu kola) is a reputedplant species for its traditional use in ayurvedic and Chinesemedicines [52], and its positive effects on brain aging havebeen generally attributed to its two major triterpene sapono-sides; asiatic and madecassic acids as well as their heterosides;asiaticoside and madecassoside, respectively. For instance,

Evidence-Based Complementary and Alternative Medicine 3
the hydroalcoholic extract of the plant was tested in vitroagainst acetylcholinesterase (AChE), the key enzyme taking acritical role in the pathogenesis of Alzheimers disease (AD).Since deficit in the level of acetylcholine (ACh), which ishydrolyzed by AChE, has been identified in the brains ofAD patients, inhibition of AChE as well as its sister enzymebutyrylcholinesterase (BChE) has become a rational target indrug development against AD [53]. The extract was foundto inhibit AChE with 50% of inhibition rate at 150 μg/mLconcentration by the spectrophotometric method of Ellman[54]. In our study on the ethanol extracts prepared from theaerial parts of C. asiatica of both Turkish and Indian originsalong with the standardized gotu kola extract (containing10.78% of total asiaticoside and madecassoside) importedfrom China, we comparatively examined inhibitory potentialof these three extracts against AChE, BChE, and tyrosinase(TYRO) at 50, 100, and 200 μg/mL concentrations [48].As aforementioned that cholinesterases are the importantenzymes for AD treatment, TYRO has become an importanttarget for Parkinson’s disease (PD) since this enzyme playsa role in neuromelanin formation in the human brain andcould be significant in occurrence of dopamine neurotoxi-city associated with neurodegeneration linked to PD [55].According to our results obtained at 200 μg/mL, only thestandardized extract was found to inhibit AChE (48.28 ±1.64%), whereas the ethanol extracts of the plant samplesfrom Turkey and India exerted 46.95 ± 0.94% and 70.30 ±3.77% against BChE, respectively, and a notable inhibitionagainst TYRO (42.83± 4.21% and 56.20± 3.17%, resp.).
Awad et al. investigated inhibitory property of C. asiaticaextract towards glutamic acid decarboxylase (GAD) and γ-aminobutyric acid transaminase (GABA-T), which are theenzymes responsible for GABA metabolism and found outthat the extract stimulated the activity of GAD over 40%[56]. On the other hand, the leaf extract of C. asiatica grow-ing in China was shown to display neuroprotection throughenhancing phosphorylation of cyclic AMP response elementbinding protein (CREB) in neuroblastoma cells in Aβ(1–42) proteins found within the amyloid plaques occurring inthe brains of AD patients [57]. In another study [58], effectof the aqueous leaf extract of the plant on monomers oroligomers that lead to formation of Aβ(1–42) proteins inAD via aggregation was examined using both thioflavin-Ttest and transmission electron microscope; however, it wasobserved not to cause any inhibition on aggregation of themonomers and oligomers. Inhibitory activity of the aqueousextract of C. asiatica that contained 84% of asiaticoside wastested by the radioenzymatic assay against phospholipaseA2 (PLA2), which play role in neuropsychiatric diseases.The findings pointed out to the fact that the extract couldinhibit Ca2+-independent PLA2 and cytosolic PLA2 [59].The ethanol extract of the plant was observed to cause anincrease in neurite development in human SH-SY5Y celllines at 100 μg/mL concentration, whereas its aqueous extractdid not lead any increase in the same cells [60]. Then, thesubfractions of the ethanol extract were also tested furtherin the same assay for neuritic development, and the mosteffective subfraction was demonstrated to have a nonpolarchemical nature. According to the results of that study, the
authors concluded that C. asiatica extract might be beneficialin prevention of neuronal damage.
Lee et al. studied neuroprotective potential of thirty sixderivatives of asiatic acid prepared by various structuralmodifications and tested in primary cell culture consisting ofrat cortical neurons exposed to glutamate, which is knownas a neurotoxin [61]. Three of the compounds displayedhigher protective activity than asiatic acid per se and alsosignificantly reduced production of glutamate-induced nitricoxide (NO) as well as levels of glutathione, glutathioneperoxidase, and some other related enzymes.
3.2. In Vivo Studies. Neuroprotective effect of C. asiatica andits major triterpene saponosides has been extensively studiedthrough different experimental models on animals such aspassive avoidance and elevated-plus labyrinth tests for mem-ory enhancing effect [62]. A research was carried out in ratsto determine effect of the aqueous extract of C. asiatica onintracerebrovascular streptozocin-induced memory associ-ated with sporadic type of AD by applying the extract atdoses of 100, 200, and 300 mg/kg (b.w.) and measuring someoxidative stress parameters such as glutathione, superoxidedismutase (SOD), and catalase (CAT) [63]. While a cleardose-dependent improvement was observed in memory-related behaviors in the rat group administered the extract at200 mg/kg (b.w.) dose, a serious decrease in malondialdehyde(MDA) and an increase in glutathione and CAT levels wererecorded, which led to a final suggestion by the authors thatC. asiatica extract has a positive effect on memory that isalso related to its remarkable antioxidant effect. The sameresearch group subjected this extract to passive avoidanceand spontaneous locomotor activity behavioral tests usingpentylenetetrazole-(PTZ-) induced memory loss in rats at100 and 300 mg/kg (b.w.) doses [64]. Following the behav-ioral tests, MDA and glutathione levels were determined inthe rat brains as oxidative stress markers, which significantlycontribute to neurodegeneration. Accordingly, the extractsat the tested doses caused a notable improvement in all testparameters.
In another study by Rao et al. [65], enhancing effect ofC. asiatica extract on learning and memory was examinedduring 15 days at 200, 500, 700, and 1000 mg/kg (b.w.)doses by oral administration to mice. Open area, light/darkcompartment, and radial-armed labyrinth tests were appliedas experimental models, while AChE activity and dendriticarborization development were taken into consideration asbiochemical markers. According to the findings, the extractdisplayed improving effect in radial-armed labyrinth test,whereas it did not cause any change in locomotor activity. Onthe other hand, extract administration resulted in an increasein AChE activity and dendritic arborization in CA3 neuronslocated in hippocampus. Thus, the authors concluded thatthe extracts may positively influence neuronal morphology,especially in young adult mice. In a similar study performedby the same researchers, the fresh leaf extract of C. asiaticawas given to adult mice at 2, 4, and 6 mL/kg doses during2, 4, and 6 weeks, respectively [66]. After these periods, theremoved brains of mice were investigated under microscope,

4 Evidence-Based Complementary and Alternative Medicine
which pointed out to the evidence that the extract givenat 6 mL/kg dose during 6 weeks caused a significant aug-ment in dendritic arborization in neurons. These authorscame to another similar conclusion that the juice obtainedby pressing the fresh leaves of C. asiatica tested in thesame experimental model in mice also enhanced dendriticarborization [67]. Besides, C. asiatica extract was shown toreduce levels of β-amyloid plaques in hippocampus in mice[68].
Shinomol and Muralidhara investigated effect of C. asi-atica extract against oxidative stress and mitochondrial dys-function induced by 3-nitropropionic acid, a fungal-derivedneurotoxin, in the brains of male prepubertal mice, and theextract was found to diminish oxidative stress remarkablythrough influencing the parameters such as MDA and rad-ical oxygen species [69]. In a related study on rats, C.asiatica extract was reported to have a protective effectagainst mitochondrial damage occurred in PD by means ofimproving oxidative stress parameters [70]. Anticonvulsanteffect of the crude material and extracts prepared from C.asiatica, also known as “brahmi” in Hindu, was determinedin PTZ-induced convulsion model in rats and comparedwith fenitoin as the reference drug [71]. The data indicatedthat the crude material of the plant exerted a mild level ofanticonvulsant effect at 500 mg/kg dose, while the methanolextract had superior effect to that of the crude material at3rd and 6th hs. The extract prepared with propylene glycolalso produced a dose-dependent anticonvulsant activity at500 and 1000 mg/kg (b.w.) doses. Similarly, Ganachari et al.demonstrated in vivo anticonvulsant effect of the hydroal-coholic extract of C. asiatica against PTZ- and strychnine-induced opistotonus convulsions at 100 mg/kg (b.w.) [72].Moreover, the extract was observed to reduce lipid per-oxidation and spontaneous locomotor activity, whilst itpotentiated pentobarbital-induced sleeping duration anddiazepam-induced hyperactivity. In another paper [73], theethyl acetate fraction of C. asiatica as well as combination ofthe fraction with some antiepileptic drugs including fenitoin,valproate, and gabapentin individually was administeredintraperitoneally to the mice with convulsion induced byPTZ and found that the combinations caused an additiveeffect producing a higher anticonvulsant activity than eachof the drugs. Additionally, neurotoxicity of the fractionand each combination was established by rotarod test,and combination of the extract with gabapentin was lessneurotoxic. In the light of this evidence, the authors statedthat conjoint use of the ethyl acetate fraction of C. asiaticawith epileptic drugs might be beneficial for epileptic patients.In another study [74], De Lucia et al. reported anticonvulsantand sedative activities of the hydroalcoholic extract of C.asiatica in rats using elevated-plus labyrinth and PTZ-induced convulsion models, and the extract was also shownto exert low toxicity by chronic application with the LD50
value of 675 mg/kg (b.w.). Anticonvulsant activity of thehexane, chloroform, ethyl acetate, water, and n-butanol ex-tracts prepared from C. asiatica was determined using PTZ-induced convulsion model in male Wistar rats, and effect ofthe extracts was also searched on Na+/K+, Mg2+, and Ca2+-ATPase activity [75]. The results pointed out to an increase in
activity of three types of ATPases in the extract-administeredgroups accompanied by anticonvulsant activity. Anxiolyticactivity of the hexane, ethyl acetate, and methanol extractsof C. asiatica and asiaticoside was tested using elevated-pluslabyrinth, open area, social interaction, locomotor activity,and new cage models in rats [76]. The results indicatedthat only the methanol and ethyl acetate extracts of theplant along with asiaticoside displayed anxiolytic activity inelevated-plus labyrinth test. In another paper [77], sedativeeffect of C. asiatica was mainly attributed to brahmoside andbrahminoside, the triterpene derivatives, whereas anxiolyticactivity was suggested to be partly resulted from the inter-action with cholecystokinin receptors (CCKB), a group ofG protein-coupled receptors which are considered to take apotential place in modulation of anxiety, nociception, andmemory.
C. asiatica extract was administered orally to old ratsduring 60 days at 300 mg/kg (b.w.) dose per day, and thecortex, hypothalamus, striatum, cerebellum, and hippocam-pus regions of the rat brains were investigated in termsof lipid peroxidation and protein carbonyl (PCO) contents[78]. The researchers made a statement that the extractmay be showing a neuroprotective effect in old rats by wayof bringing about a significant decrease in PCO contentsand lipid peroxidation. Radical scavenging effect of thechloroform-methanol (4 : 1) extract of the plant was exam-ined in monosodium glutamate-treated Sprague-Dawleyfemale rats at 100 and 200 mg/kg doses [79]. Following theextract administration, a significant increase was observedin SOD and CAT levels, whereas glutathione level was notinfluenced. Flora and Gupta reported that the flavonoidfraction of C. asiatica demonstrated a protecting effectagainst lead acetate-induced neurotoxicity in mice throughantioxidant mechanisms [80]. In another paper, asiatic acid,one of the major triterpene derivatives in C. asiatica, ad-ministered orally at 30, 75, and 165 mg/kg (b.w.) doses,was shown to have neuroprotective property in mice withpermanent cerebral ischemia by evaluating infarct volumeand behavioral changes between 1st and 7th days [81]. Inthe same study, the compound was additionally investigatedin HT-22 cells exposed to oxygen glucose in terms of cellviability and mitochondrial membrane potential. Asiatic acidconsiderably diminished the infarct volume by 60% and26% at the 1st and 7th days, respectively, which improvedneurological status at 24 h after ischemia. The authors con-cluded that asiatic acid, which might be mediated to someextent by decreasing blood-brain barrier permeability as wellas reduction in mitochondrial damage, could be useful forcerebral ischemia treatment.
Probable improving effect of C. asiatica extract at 150and 300 mg/kg (p.o.) doses was assessed against colchicine-induced memory using Morris water maze and plus-mazeperformance tests in male Wistar rats as well as oxidativedamage parameters such as lipid peroxidation, nitrite, re-duced glutathione, glutathione-S-transferase, SOD, and asa biochemical parameter, AChE activity [82]. The 25-daychronic administration of the extract caused a significantimprovement in memory and oxidative damage parametersalong with AChE activity. On the other hand, asiaticoside

Evidence-Based Complementary and Alternative Medicine 5
from C. asiatica exerted a neuroprotective effect against PDby reversing neurotoxicity induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in rats via balancingdopamine and antioxidant mechanism [83].
Antidepressant activity of C. asiatica was evaluated usingits triterpene fraction in cortex, hippocampus, and thalamusregions of rat brains by determining the corticosterone lev-els [84]. The triterpene fraction created a momentous dim-inution in corticosterone level and a notable increase inamount of monoamine-related neurotransmitters.
3.3. Clinical Studies. Although many in vivo studies havebeen carried out on central-nervous-system-(CNS-) relatedeffects of C. asiatica, the literature survey has revealedpresence of only a limited number of clinical studies with thisspecies. The results of an early double-blind clinical studyon the children with mental deficiency in 1977 showed thata statistically significant improvement was recorded in thechildren in 3rd and 6th months following administration ofC. asiatica [85].
Possible effect of the capsulated aqueous extract ofC. asiatica standardized to contain 29.9 mg/g tannic acid,1.09 mg/g asiaticoside, and 48.89 mg/g asiatic acid was de-termined in a randomized, double-blind, and placebo-con-trolled clinical study carried out on 28 healthy and eldervolunteers consisting of 4 men and 24 women with theaverage age of 65.05 ± 3.56 in Thailand [86]. The extractwas given to the subjects once a day at 250, 500, and 750 mgdoses during 2 months, and their cognitive performancewas evaluated by a variety of parameters using computer-assisted techniques. The findings revealed that the highestdose of C. asiatica extract tested in this study possessed acognitive enhancing effect. In a similar study [87], Dev etal. investigated effect of the capsulated C. asiatica extracton cognitive performance conducted with a total 41 ofmiddle-age healthy subjects consisting of 22 women and19 men. The extract was given to the subjects in a capsuleonce a day during 2 months. The cognitive performancewas measured using Woodcock-Johnson Cognitive AbilitiesTest III (WJCAT III), and the extract was found to havea remarkably positive influence on all of the subjects. Arecent clinical study consisting of 60 elderly subjects withaverage age of 65 with mild cognitive deficiency indicatedthat C. asiatica extract administered at 500 mg dose twiceper day during 6 months led to a significant cognitiveimprovement according to Mini Mental State Examination(MMSE) scoring [88].
4. Precautions
Although C. asiatica is one of the top-selling herbal medi-cines due to its remarkable pharmacological effects, someprecautions should be taken for this plant. It has been knownto be safe when taken at the recommended doses; however,skin irritation and contact dermatitis have been reportedin some cases [89–91]. In a very early paper in 1969 [92],the total saponoside fraction containing brahmic acid andits derivatives of the plant was stated to cause infertilityin an experiment conducted on human and rat sperms. In
consistency with this paper, Newall et al. also affirmed thatinfertility was observed in female mice after oral admin-istration of C. asiatica [93]. Another result pointed out tothe fact that chronic treatment of C. asiatica might inducea spontaneous abortion in pregnant women [94]. Since theplant may bring about a raise in blood sugar and lipidlevels, diabetic and hyperlipidemic patients should considertaking preparations of C. asiatica [93]. Briefly, maximumduration suggested for the use of C. asiatica preparations is6 weeks, and at least, a 2-week break is needed after everylong duration use. Even though no drug interaction hasbeen reported for this plant up to date, pregnant and breast-feeding women are suggested to avoid using this herbalmedicine.
5. Conclusion
C. asiatica, widely known as “gotu kola,” is a reputed me-dicinal plant for its various pharmacological effects favorablefor human health. Besides its potent wound healing property,a number of studies described the noteworthy protectiveeffect of the plant against several diseases of CNS. Biologicaleffects of C. asiatica have been generally attributed to themajor triterpene derivatives including asiatic acid, made-cassic acid, asiaticoside, madecassoside, and brahmic acid.The neuroprotective effect of the plant has been suggestedto result from different mechanisms, most of which havereferred to positive influences on oxidative stress parameters.
References
[1] S. S. Jamil, Q. Nizami, and M. Salam, “Centella asiatica (Linn.)Urban: a review,” Natural Product Radiance, vol. 6, no. 2, pp.158–170, 2007.
[2] G. J. Meulenbeld and D. Wujastyk, Studies on Indian MedicalHistory, Motilal Banarsidas, New Delhi, India, 2001.
[3] R. N. Chopra, S. L. Nayar, and I. C. Chopra, Glossary ofIndian Medicinal Plants (Including the Supplement), Councilof Scientific and Industrial Research, New Delhi, India, 1986.
[4] M. J. R. Howes and P. Houghton, “Plants used in Chinese andIndian traditional medicine for improvement of memory andcognitive function,” Pharmacology Biochemistry and Behavior,vol. 75, no. 3, pp. 513–527, 2003.
[5] R. Tenni, G. Zanaboni, M. P. De Agostini, A. Rossi, C. Bend-otti, and G. Cetta, “Effect of the triterpenoid fraction ofCentella asiatica on macromolecules of the connective matrixin human skin fibroblast cultures,” Italian Journal of Biochem-istry, vol. 37, no. 2, pp. 69–77, 1988.
[6] L. Suguna, P. Sivakumar, and G. Chandrakasan, “Effects ofCentella asiatica extract on dermal wound healing in rats,”Indian Journal of Experimental Biology, vol. 34, no. 12, pp.1208–1211, 1996.
[7] B. S. Shetty, S. L. Udupa, and A. L. Udupa, “Biochemicalanalysis of granulation tissue in steroid and Centella asiatica(Linn) treated rats,” Pharmacologyonline, vol. 2, pp. 624–632,2008.
[8] M. N. Somchit, M. R. Sulaiman, A. Zuraini et al., “Antinoci-ceptive and antiinflammatory effects of Centella asiatica,”Indian Journal of Pharmacology, vol. 36, no. 6, pp. 377–380,2004.

6 Evidence-Based Complementary and Alternative Medicine
[9] M. George, L. Joseph, and Ramaswamy, “Anti-allergic, anti-pruritic, and anti-inflammatory activities of Centella asiaticaextracts,” African Journal of Traditional, Complementary andAlternative Medicines, vol. 6, no. 4, pp. 554–559, 2009.
[10] J. H. Sampson, A. Raman, G. Karlsen, H. Navsaria, and I.Leigh, “In vitro keratinocyte antiproliferant effect of Centellaasiatica extract and triterpenoid saponins,” Phytomedicine, vol.8, no. 3, pp. 230–235, 2001.
[11] C. L. Cheng and M. W. L. Koo, “Effects of Centella asiatica onethanol induced gastric mucosal lesions in rats,” Life Sciences,vol. 67, no. 21, pp. 2647–2653, 2000.
[12] C. L. Cheng, J. S. Guo, J. Luk, and M. W. L. Koo, “The heal-ing effects of Centella extract and asiaticoside on acetic acidinduced gastric ulcers in rats,” Life Sciences, vol. 74, no. 18, pp.2237–2249, 2004.
[13] S. S. Pingale, “Evaluation of effect of Centella asiatica on CCL4
induced rat liver damage,” Pharmacologyonline, vol. 3, pp.537–543, 2008.
[14] S. Sudha, S. Kumaresan, A. Amit, J. David, and B. V. Ven-kataraman, “Anti-convulsant activity of different extracts ofCentella asiatica and Bacopa monnieri in animals,” Journal ofNatural Remedies, vol. 2, no. 1, pp. 33–41, 2002.
[15] P. Wijeweera, J. T. Arnason, D. Koszycki, and Z. Merali,“Evaluation of anxiolytic properties of Gotukola—(Centellaasiatica) extracts and asiaticoside in rat behavioral models,”Phytomedicine, vol. 13, no. 9-10, pp. 668–676, 2006.
[16] X. S. Wang, Q. Dong, J. P. Zuo, and J. N. Fang, “Structureand potential immunological activity of a pectin from Centellaasiatica (L.) Urban,” Carbohydrate Research, vol. 338, no. 22,pp. 2393–2402, 2003.
[17] A. Gnanapragasam, K. Kumar Ebenezar, V. Sathish, P. Govin-daraju, and T. Devaki, “Protective effect of Centella asiatica onantioxidant tissue defense system against adriamycin inducedcardiomyopathy in rats,” Life Sciences, vol. 76, no. 5, pp. 585–597, 2004.
[18] M. Raghavendra, R. Maiti, S. Kumar, A. Trigunayat, S. Mitra,and S. Acharya, “Role of Centella asiatica on cerebral post-ischemic reperfusion and long-term hypoperfusion in rats,”International Journal of Green Pharmacy, vol. 3, no. 2, pp. 88–96, 2009.
[19] M. L. Venu Gopal Rao and S. A. Mastan, “Antidiabetic effectsof methanolic extract of Centella asiatica (Linn.) on inducedhyperglycemic rats,” Biosciences Biotechnology Research Asia,vol. 4, no. 2, pp. 721–724, 2007.
[20] Y. S. Lee, D. Q. Jin, E. J. Kwon et al., “Asiatic acid, a triterpene,induces apoptosis through intracellular Ca2+ release andenhanced expression of p53 in HepG2 human hepatoma cells,”Cancer Letters, vol. 186, no. 1, pp. 83–91, 2002.
[21] P. Bunpo, K. Kataoka, H. Arimochi et al., “Inhibitory effectsof Centella asiatica on azoxymethane-induced aberrant cryptfocus formation and carcinogenesis in the intestines of F344rats,” Food and Chemical Toxicology, vol. 42, no. 12, pp. 1987–1997, 2004.
[22] C. Yoosook, N. Bunyapraphatsara, Y. Boonyakiat, and C. Kant-asuk, “Anti-herpes simplex virus activities of crude waterextracts of Thai medicinal plants,” Phytomedicine, vol. 6, no.6, pp. 411–419, 2000.
[23] M. R. Zaidan, A. Noor Rain, A. R. Badrul, A. Adlin, A.Norazah, and I. Zakiah, “In vitro screening of five local me-dicinal plants for antibacterial activity using disc diffusionmethod,” Tropical Biomedicine, vol. 22, no. 2, pp. 165–170,2005.
[24] N. Senthilkumar, P. Varma, and G. Gurusubramanian, “Lar-vicidal and adulticidal activities of some medicinal plants
against the Malarial Vector, Anopheles stephensi (Liston),”Parasitology Research, vol. 104, no. 2, pp. 237–244, 2009.
[25] E. Naz and M. Ahmad, “Evaluation of five indigenous medic-inal plants of Sindh, Pakistan for their antifungal potential,”Pakistan Journal of Scientific and Industrial Research, vol. 52,no. 6, pp. 328–333, 2009.
[26] A. A. Hamid, Z. Shah, R. Muse, and S. Mohamed, “Characteri-sation of antioxidative activities of various extracts of Centellaasiatica (L) Urban,” Food Chemistry, vol. 77, no. 4, pp. 465–469, 2002.
[27] G. Jayashree, G. Kurup Muraleedhara, S. Sudarslal, and V. B.Jacob, “Anti-oxidant activity of Centella asiatica on lympho-ma-bearing mice,” Fitoterapia, vol. 74, no. 5, pp. 431–434,2003.
[28] M. Bajpai, A. Pande, S. K. Tewari, and D. Prakash, “Phenoliccontents and antioxidant activity of some food and medicinalplants,” International Journal of Food Sciences and Nutrition,vol. 56, no. 4, pp. 287–291, 2005.
[29] S. Chaudhuri, S. Ghosh, and T. Chakraborty, “Use of acommon Indian herb “Mandukaparni” in the treatment ofleprosy. (Preliminary report),” Journal of the Indian MedicalAssociation, vol. 70, no. 8, pp. 177–180, 1978.
[30] J. P. Pointel, H. Boccalon, and M. Cloarec, “Titrated extract ofCentella asiatica (TECA) in the treatment of venous insuffi-ciency of the lower limbs,” Angiology, vol. 38, no. 1, pp. 46–50,1987.
[31] M. R. Cesarone, G. Belcaro, A. Rulo et al., “Microcirculatoryeffects of total triterpenic fraction of Centella asiatica inchronic venous hypertension: measurement by laser Doppler,TcPo2-co2, and leg volumetry,” Angiology, vol. 52, no. 10, pp.S45–S48, 2001.
[32] E. Williamson, “Centella asiatica (L.) Urb,” in Major Herbs ofAyurveda, E. Williamson, Ed., pp. 102–110, Elsevier Science,London, UK, 2002.
[33] J. Pan, G. Kai, C. Yuan, B. Zhou, R. Jin, and Y. Yuan,“Separation and determination of madecassic acid in extractsof Centella asiatica using high performance liquid chromatog-raphy with β-cyclodextrin as mobile phase additive,” ChineseJournal of Chromatography, vol. 25, no. 3, pp. 316–318, 2007.
[34] N. P. Sahu, S. K. Roy, and S. B. Mahato, “Spectroscopicdetermination of structures of triterpenoid trisaccharidesfrom Centella asiatica,” Phytochemistry, vol. 28, no. 10, pp.2852–2854, 1989.
[35] M. Kuroda, Y. Mimaki, H. Harada, H. Sakagami, and Y.Sashida, “Five new triterpene glycosides from Centella asiat-ica,” Natural Medicines, vol. 55, no. 3, pp. 134–138, 2001.
[36] B. S. Siddiqui, H. Aslam, S. T. Ali, S. Khan, and S. Begum,“Chemical constituents of Centella asiatica,” Journal of AsianNatural Products Research, vol. 9, no. 4, pp. 407–414, 2007.
[37] Y. N. Shukla, R. Srivastava, A. K. Tripathi, and V. Prajapati,“Characterization of an ursane triterpenoid from Centellaasiatica with growth inhibitory activity against Spilarctia obli-qua,” Pharmaceutical Biology, vol. 38, no. 4, pp. 262–267, 2000.
[38] H. Matsuda, T. Morikawa, H. Ueda, and M. Yoshikawa, “Me-dicinal foodstuffs. XXVI. Inhibitors of aldose reductase andnew triterpene and its oligoglycoside, centellasapogenol Aand centellasaponin A, from Centella asiatica (Gotu Kola),”Heterocycles, vol. 55, no. 8, pp. 1499–1504, 2001.
[39] H. Matsuda, T. Morikawa, H. Ueda, and M. Yoshikawa, “Me-dicinal foodstuffs. XXVII. Saponin constituents of gotu kola(2): structures of new ursane- and oleanane-type triterpeneoligoglycosides, centellasaponins B, C, and D, from Centellaasiatica cultivated in Sri Lanka,” Chemical and PharmaceuticalBulletin, vol. 49, no. 10, pp. 1368–1371, 2001.

Evidence-Based Complementary and Alternative Medicine 7
[40] M. Yoshida, M. Fuchigami, T. Nagao et al., “Antiproliferativeconstituents from umbelliferae plants VII. Active triterpenesand rosmarinic acid from Centella asiatica,” Biological andPharmaceutical Bulletin, vol. 28, no. 1, pp. 173–175, 2005.
[41] C. S. Rumalla, Z. Ali, A. D. Weerasooriya, T. J. Smillie, and I. A.Khan, “Two new triterpene glycosides from Centella asiatica,”Planta Medica, vol. 76, no. 10, pp. 1018–1021, 2010.
[42] R. Subban, A. Veerakumar, R. Manimaran, K. M. Hashim, andI. Balachandran, “Two new flavonoids from Centella asiatica(Linn.),” Journal of Natural Medicines, vol. 62, no. 3, pp. 369–373, 2008.
[43] X. S. Wang, J. Y. Duan, and J. N. Fang, “Structural featuresof a polysaccharide from Centella asiatica,” Chinese ChemicalLetters, vol. 15, no. 2, pp. 187–190, 2004.
[44] G. Govindan, T. G. Sambandan, M. Govindan et al., “Abioactive polyacetylene compound isolated from Centellaasiatica,” Planta Medica, vol. 73, no. 6, pp. 597–599, 2007.
[45] R. Srivastava and Y. N. Shukla, “Some chemical constituentsfrom Centella asiatica,” Indian Drugs, vol. 33, no. 5, pp. 233–234, 1996.
[46] N. Sondhi, R. Bhardwaj, S. Kaur, M. Chandel, N. Kumar, andB. Singh, “Inhibition of H2O2-induced DNA damage in singlecell gel electrophoresis assay (comet assay) by castasteroneisolated from leaves of Centella asiatica,” Health, vol. 2, no. 6,pp. 595–602, 2010.
[47] L. Suntornsuk and O. Anurukvorakun, “Precision improve-ment for the analysis of flavonoids in selected Thai plants bycapillary zone electrophoresis,” Electrophoresis, vol. 26, no. 3,pp. 648–660, 2005.
[48] I. Erdogan Orhan, E. Atasu, F. S. Senol et al., “High-through-put bioactivity screening of the Southeast Asian vegetableCentella asiatica (L.) Urban (gotu kola) and its phytochemicalanalysis,” Food Chemistry . Under review.
[49] Y. Asakawa, R. Matsuda, and T. Takemoto, “Mono- and ses-quiterpenoids from Hydrocotyle and Centella species,” Phyto-chemistry, vol. 21, no. 10, pp. 2590–2592, 1982.
[50] O. A. Oyedeji and A. J. Afolayan, “Chemical composition andantibacterial activity of the essential oil of Centella asiaticagrowing in South Africa,” Pharmaceutical Biology, vol. 43, no.3, pp. 249–252, 2005.
[51] P. Wongfhun, M. H. Gordon, and A. Apichartsrangkoon,“Flavour characterisation of fresh and processed pennywort(Centella asiatica L.) juices,” Food Chemistry, vol. 119, no. 1,pp. 69–74, 2010.
[52] M. J. R. Howes and P. J. Houghton, “Plants used in Chineseand Indian traditional medicine for improvement of mem-ory and cognitive function,” Pharmacology Biochemistry andBehavior, vol. 75, no. 3, pp. 513–527, 2003.
[53] G. Orhan, I. Orhan, and B. Sener, “Recent developments innatural and synthetic drug research for Alzheimer’s Disease,”Letters in Drug Design and Discovery, vol. 3, no. 4, pp. 268–274,2006.
[54] P. K. Mukherjee, V. Kumar, and P. J. Houghton, “Screeningof Indian medicinal plants for acetylcholinesterase inhibitoryactivity,” Phytotherapy Research, vol. 21, no. 12, pp. 1142–1145,2007.
[55] M. T. H. Khan, “Molecular design of tyrosinase inhibitors: acritical review of promising novel inhibitors from syntheticorigins,” Pure and Applied Chemistry, vol. 79, no. 12, pp. 2277–2295, 2007.
[56] R. Awad, D. Levac, P. Cybulska, Z. Merali, V. L. Trudeau,and J. T. Arnason, “Effects of traditionally used anxiolyticbotanicals on enzymes of the γ-aminobutyric acid (GABA)
system,” Canadian Journal of Physiology and Pharmacology,vol. 85, no. 9, pp. 933–942, 2007.
[57] Y. Xu, Z. Cao, I. Khan, and Y. Luo, “Gotu Kola (Centella Asiat-ica) extract enhances phosphorylation of cyclic AMP responseelement binding protein in neuroblastoma cells expressingamyloid beta peptide,” Journal of Alzheimer’s Disease, vol. 13,no. 3, pp. 341–349, 2008.
[58] B. N. Ramesh, S. S. Indi, and K. S. J. Rao, “Studies to under-stand the effect of Centella asiatica on Aβ(42) aggregation invitro,” Current Trends in Biotechnology and Pharmacy, vol. 4,no. 2, pp. 716–724, 2010.
[59] N. R. Barbosa, F. Pittella, and W. F. Gattaz, “Centella asiaticawater extract inhibits iPLA2 and cPLA2 activities in ratcerebellum,” Phytomedicine, vol. 15, no. 10, pp. 896–900, 2008.
[60] A. Soumyanath, Y. P. Zhong, S. A. Gold et al., “Centella asiaticaaccelerates nerve regeneration upon oral administration andcontains multiple active fractions increasing neurite elonga-tion in-vitro,” Journal of Pharmacy and Pharmacology, vol. 57,no. 9, pp. 1221–1229, 2005.
[61] M. K. Lee, S. R. Kim, S. H. Sung et al., “Asiatic acid derivativesprotect cultured cortical neurons from glutamate-inducedexcitotoxicity,” Research Communications in Molecular Pathol-ogy and Pharmacology, vol. 108, no. 1-2, pp. 75–86, 2000.
[62] M. H. Veerendra Kumar and Y. K. Gupta, “Effect of differentextracts of Centella asiatica on cognition and markers ofoxidative stress in rats,” Journal of Ethnopharmacology, vol. 79,no. 2, pp. 253–260, 2002.
[63] M. H. Veerendra Kumar and Y. K. Gupta, “Effect of Centellaasiatica on cognition and oxidative stress in an intracere-broventricular streptozotocin model of Alzheimer’s disease inrats,” Clinical and Experimental Pharmacology and Physiology,vol. 30, no. 5-6, pp. 336–342, 2003.
[64] Y. K. Gupta, M. H. V. Kumar, and A. K. Srivastava, “Effect ofCentella asiatica on pentylenetetrazole-induced kindling, cog-nition and oxidative stress in rats,” Pharmacology Biochemistryand Behavior, vol. 74, no. 3, pp. 579–585, 2003.
[65] S. B. Rao, M. Chetana, and P. Uma Devi, “Centella asiaticatreatment during postnatal period enhances learning andmemory in mice,” Physiology and Behavior, vol. 86, no. 4, pp.449–457, 2005.
[66] M. R. Gadahad, M. Rao, and G. Rao, “Enhancement ofhippocampal CA3 neuronal dendritic arborization by Centellaasiatica (Linn) fresh leaf extract treatment in adult rats,”Journal of the Chinese Medical Association, vol. 71, no. 1, pp.6–13, 2008.
[67] K. G. M. Rao, S. Muddanna Rao, and S. Gurumadhva Rao,“Enhancement of amygdaloid neuronal dendritic arborizationby fresh leaf juice of Centella asiatica (Linn) during growthspurt period in rats,” Evidence-based Complementary andAlternative Medicine, vol. 6, no. 2, pp. 203–210, 2009.
[68] M. Dhanasekaran, L. A. Holcomb, A. R. Hitt et al., “Centellaasiatica extract selectively decreases amyloid β levels in hip-pocampus of alzheimer’s disease animal model,” PhytotherapyResearch, vol. 23, no. 1, pp. 14–19, 2009.
[69] G. K. Shinomol and K. Muralidhara, “Effect of Centella asi-atica leaf powder on oxidative markers in brain regions ofprepubertal mice in vivo and its in vitro efficacy to amel-iorate 3-NPA-induced oxidative stress in mitochondria,” Phy-tomedicine, vol. 15, no. 11, pp. 971–984, 2008.
[70] N. Haleagrahara and K. Ponnusamy, “Neuroprotective effectof Centella asiatica extract (CAE) on experimentally inducedparkinsonism in aged Sprague-Dawley rats,” Journal of Toxico-logical Sciences, vol. 35, no. 1, pp. 41–47, 2010.

8 Evidence-Based Complementary and Alternative Medicine
[71] S. Sudha, S. Kumaresan, A. Amit, J. David, and B. V. Ven-kataraman, “Anti-convulsant activity of different extracts ofCentella asiatica and Bacopa monnieri in animals,” Journal ofNatural Remedies, vol. 2, no. 1, pp. 33–41, 2002.
[72] M. S. Ganachari, S. V. Veeresh Babu, and S. S. Katare, “Neuro-pharmacology of an extract derived from Centella asiatica,”Pharmaceutical Biology, vol. 42, no. 3, pp. 246–252, 2004.
[73] A. Vattanajun, H. Watanabe, M. H. Tantisira, and B. Tantisira,“Isobolographically additive anticonvulsant activity betweenCentella asiatica’s ethyl acetate fraction and some antiepilepticdrugs,” Journal of the Medical Association of Thailand, vol. 88,supplement 3, pp. S131–140, 2005.
[74] R. De Lucia, J. A. A. Sertie, E. A. Camargo, and S. Panizza,“Pharmacological and toxicological studies on Centella asiat-ica extract,” Fitoterapia, vol. 68, no. 5, pp. 413–416, 1997.
[75] G. Visweswari, K. Siva Prasad, V. Lokanatha, and W. Rajendra,“The antiepileptic effect of Centella asiatica on the activities ofNa+ /K+ , Mg2+ and Ca2+ -ATPases in rat brain during penty-lenetetrazol-induced epilepsy,” Indian Journal of Pharmacol-ogy, vol. 42, no. 2, pp. 82–86, 2010.
[76] P. Wijeweera, J. T. Arnason, D. Koszycki, and Z. Merali,“Evaluation of anxiolytic properties of Gotukola—(Centellaasiatica) extracts and asiaticoside in rat behavioral models,”Phytomedicine, vol. 13, no. 9-10, pp. 668–676, 2006.
[77] A. S. Ramaswamy, S. M. Pariyaswami, and N. Basu, “Pharma-cological studies on Centella asiatica.Linn,” Indian Journal ofMedicinal Research, vol. 4, pp. 160–164, 1970.
[78] M. Bajpai, A. Pande, S. K. Tewari, and D. Prakash, “Phenoliccontents and antioxidant activity of some food and medicinalplants,” International Journal of Food Sciences and Nutrition,vol. 56, no. 4, pp. 287–291, 2005.
[79] M. Hussin, A. Abdul-Hamid, S. Mohamad, N. Saari, M. Ismail,and M. H. Bejo, “Protective effect of Centella asiatica extractand powder on oxidative stress in rats,” Food Chemistry, vol.100, no. 2, pp. 535–541, 2007.
[80] S. J. S. Flora and R. Gupta, “Beneficial effects of Centellaasiatica aqueous extract against arsenic-induced oxidativestress and essential metal status in rats,” Phytotherapy Research,vol. 21, no. 10, pp. 980–988, 2007.
[81] R. G. Krishnamurthy, M. C. Senut, D. Zemke et al., “Asiaticacid, a pentacyclic triterpene from Centella asiatica, is neu-roprotective in a mouse model of focal cerebral ischemia,”Journal of Neuroscience Research, vol. 87, no. 11, pp. 2541–2550, 2009.
[82] A. Kumar, S. Dogra, and A. Prakash, “Neuroprotective effectsof Centella asiatica against Intracerebroventricular colchicine-induced cognitive impairment and oxidative stress,” Interna-tional Journal of Alzheimer’s Disease, vol. 2009, Article ID972178, 8 pages, 2009.
[83] C. L. Xu, Q. Z. Wang, L. M. Sun et al., “Asiaticoside: at-tenuation of neurotoxicity induced by MPTP in a rat modelof Parkinsonism via maintaining redox balance and up-regulating the ratio of Bcl-2/Bax,” Pharmacology Biochemistryand Behavior, vol. 100, pp. 413–418, 2012.
[84] Y. Chen, T. Han, Y. Rui, M. Yin, L. Qin, and H. Zheng, “Effectsof total triterpenes of Centella asiatica on the corticosteronelevels in serum and contents of monoamine in depression ratbrain,” Zhong yao Cai, vol. 28, no. 6, pp. 492–496, 2005.
[85] M. V. R. Appa Rao, K. Srinivasan, and T. Koteswara Rao,“The effect of Centella asiatica on the general mental ability ofmentally retarded children,” Indian Journal of Psychiatry, vol.19, no. 4, pp. 54–59, 1977.
[86] J. Wattanathorn, L. Mator, S. Muchimapura et al., “Positivemodulation of cognition and mood in the healthy elderly
volunteer following the administration of Centella asiatica,”Journal of Ethnopharmacology, vol. 116, no. 2, pp. 325–332,2008.
[87] R. D. O. Dev, S. Mohamed, Z. Hambali, and B. A. Samah,“Comparison on cognitive effects of Centella asiatica in health-y middle age female and male volunteers,” European Journal ofScientific Research, vol. 31, no. 4, pp. 553–565, 2009.
[88] S. Tiwari, S. Singh, K. Patwardhan, S. Gehlot, and I. S. Gamb-hir, “Effect of Centella asiatica on mild cognitive impairment(MCI) and other common age-related clinical problems,”Digest Journal of Nanomaterials and Biostructures, vol. 3, pp.215–220, 2008.
[89] I. Bilbao, A. Aguirre, R. Zabala, R. Gonzalez, J. Raton, and J. L.D. Perez, “Allergic contact dermatitis from butoxyethyl nico-tinic acid and Centella asiatica extract,” Contact Dermatitis,vol. 33, no. 6, pp. 435–436, 1995.
[90] M. A. Gonzalo-Garijo, F. Revenga-Arranz, and P. Bobadilla-Gonzalez, “Allergic contact dermatitis due to Centella asiatica:a new case,” Allergy and Immunopathology, vol. 24, pp. 132–134, 1996.
[91] J. Gomes, T. Pereira, C. Vilarinho, M. D. L. Duarte, and C.Brito, “Contact dermatitis due to Centella asiatica,” ContactDermatitis, vol. 62, no. 1, pp. 54–55, 2010.
[92] B. Singh and R. P. Rastogi, “Chemical examination of Centellaasiatica linn-III. Constitution of brahmic acid,” Phytochem-istry, vol. 7, no. 8, pp. 1385–1393, 1968.
[93] C. A. Newall, L. A. Anderson, and J. D. Phillipson, HerbalMedicines, Pharmaceutical Press, London, UK, 1996.
[94] T. Dutta and U. P. Basu, “Crude extract of Centella asiaticaand products derived from its glycosides as oral antifertilityagents.,” Indian Journal of Experimental Biology, vol. 6, no. 3,pp. 181–182, 1968.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 636091, 7 pagesdoi:10.1155/2012/636091
Research Article
Rhynchophylline Protects Cultured Rat Neurons againstMethamphetamine Cytotoxicity
Dan Dan Xu,1, 2 Robin Hoeven,3 Rong Rong,4 and William Chi-Shing Cho5
1 Faculty of Science, The Chinese University of Hong Kong, Hong Kong2 Jiujianpeng Technology R&D Center, Linyi, China3 Faculty of Science, Katholieke Universiteit, Leuven, Belgium4 School of Pharmacy, Shandong University of Traditional Chinese Medicine, Jinan, China5 Department of Clinical Oncology, Queen Elizabeth Hospital, Hong Kong
Correspondence should be addressed to William Chi-Shing Cho, [email protected]
Received 19 November 2011; Accepted 7 February 2012
Academic Editor: Ilkay Erdogan Orhan
Copyright © 2012 Dan Dan Xu et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Rhynchophylline (Rhy) is an active component isolated from species of the genus Uncaria which has been used for the treatment ofailments to the central nervous system in traditional Chinese medicine. Besides acting as a calcium channel blocker, Rhy was alsoreported to be able to protect against glutamate-induced neuronal death. We thus hypothesize that Rhy may have neuroprotectiveactivity against methamphetamine (MA). The primary neurons were cultured directly from the cerebral cortex of neonatal rats,acting as in vitro model in the present study. The neurotoxicity of MA and the protective effect of Rhy were evaluated by MTT assay.The effects of MA, Rhy or their combination on intracellular free calcium concentration ([Ca2+]i) were determined in individualneocortical neurons by the Fluo-3/AM tracing method. The MTT assay demonstrated that MA has a dose-dependent neurotoxicityin neuronal cultures. The addition of Rhy prior to the exposure to MA prevented neuronal death. Time course studies with theFluo-3/AM probe showed that Rhy significantly decreased neuronal [Ca2+]i which was elevated by the exposure to MA. Our resultssuggested that Rhy can protect the neuronal cultures against MA exposure and promptly attenuate intracellular calcium overloadtriggered by MA challenge. This is the first report demonstrating an inhibitory effect of Rhy against MA impairment in culturedneurons in vitro.
1. Introduction
Rhynchophylline (Rhy) is a major tetracyclic oxindole alka-loid (the chemical structure of Rhy is shown in Figure 1),originally isolated from species of the genus Uncaria whichis widely used in traditional Chinese medicine prescribedmainly to treat ailments to the central nervous and car-diovascular systems, such as lightheadedness, convulsions,numbness, and hypertension [1].
In the cardiovascular system, it is well documented thatthe vasodilative effect of Rhy is mainly due to the dysfunctionof Ca2+ transport, including influx of extracellular calciumand release of intracellular calcium by blocking the voltage-dependent calcium channel and the receptor-regulation cal-cium channel [2]. There are records that an extraction withmethanol from Uncaria rhynchophylla exhibits a significant
neuroprotective effect in vitro, in which Rhy appears to be theactive pharmacological component [3]. Further research onrat cerebellar granule cells showed Rhy to be able to protectagainst glutamate-induced neuronal death [4]. Moreover,Rhy is thought to have calcium channel blocking functions,which can further explain the neuroprotective functions.Calcium channel blockage can potentially have a protectiveeffect because of the positive relation between increasedcalcium influx and cell death [1]. For this reason, weexpect Rhy to have neuroprotective functions for the centralnervous system.
However, the mechanistic study for Rhy is limited,especially for the neural impairment induced by drugabuse. Methamphetamine (MA) is a commonly abusedpsychostimulant in the world, which leads to severelyneurodegenerative changes in the human brain [5]. It
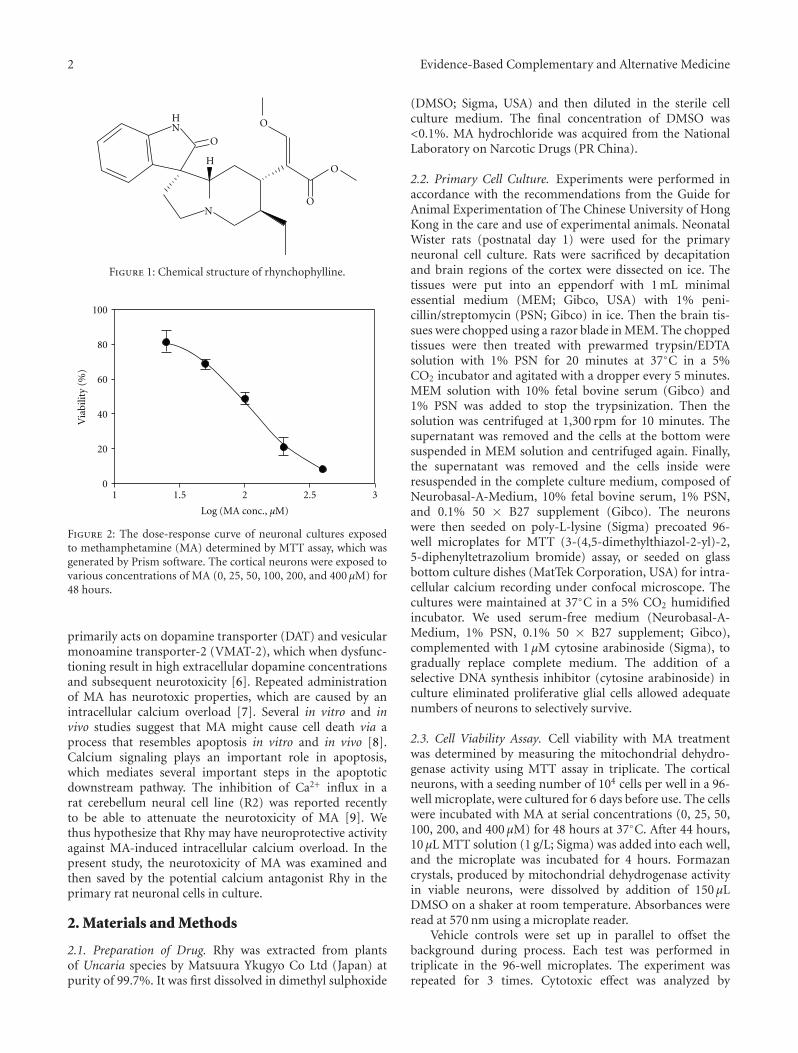
2 Evidence-Based Complementary and Alternative Medicine
NO
O
O
ONH
H
Figure 1: Chemical structure of rhynchophylline.
100
80
60
40
20
0
Via
bilit
y (%
)
1 1.5 2 2.5 3
Log (MA conc., µM)
Figure 2: The dose-response curve of neuronal cultures exposedto methamphetamine (MA) determined by MTT assay, which wasgenerated by Prism software. The cortical neurons were exposed tovarious concentrations of MA (0, 25, 50, 100, 200, and 400 µM) for48 hours.
primarily acts on dopamine transporter (DAT) and vesicularmonoamine transporter-2 (VMAT-2), which when dysfunc-tioning result in high extracellular dopamine concentrationsand subsequent neurotoxicity [6]. Repeated administrationof MA has neurotoxic properties, which are caused by anintracellular calcium overload [7]. Several in vitro and invivo studies suggest that MA might cause cell death via aprocess that resembles apoptosis in vitro and in vivo [8].Calcium signaling plays an important role in apoptosis,which mediates several important steps in the apoptoticdownstream pathway. The inhibition of Ca2+ influx in arat cerebellum neural cell line (R2) was reported recentlyto be able to attenuate the neurotoxicity of MA [9]. Wethus hypothesize that Rhy may have neuroprotective activityagainst MA-induced intracellular calcium overload. In thepresent study, the neurotoxicity of MA was examined andthen saved by the potential calcium antagonist Rhy in theprimary rat neuronal cells in culture.
2. Materials and Methods
2.1. Preparation of Drug. Rhy was extracted from plantsof Uncaria species by Matsuura Ykugyo Co Ltd (Japan) atpurity of 99.7%. It was first dissolved in dimethyl sulphoxide
(DMSO; Sigma, USA) and then diluted in the sterile cellculture medium. The final concentration of DMSO was<0.1%. MA hydrochloride was acquired from the NationalLaboratory on Narcotic Drugs (PR China).
2.2. Primary Cell Culture. Experiments were performed inaccordance with the recommendations from the Guide forAnimal Experimentation of The Chinese University of HongKong in the care and use of experimental animals. NeonatalWister rats (postnatal day 1) were used for the primaryneuronal cell culture. Rats were sacrificed by decapitationand brain regions of the cortex were dissected on ice. Thetissues were put into an eppendorf with 1 mL minimalessential medium (MEM; Gibco, USA) with 1% peni-cillin/streptomycin (PSN; Gibco) in ice. Then the brain tis-sues were chopped using a razor blade in MEM. The choppedtissues were then treated with prewarmed trypsin/EDTAsolution with 1% PSN for 20 minutes at 37◦C in a 5%CO2 incubator and agitated with a dropper every 5 minutes.MEM solution with 10% fetal bovine serum (Gibco) and1% PSN was added to stop the trypsinization. Then thesolution was centrifuged at 1,300 rpm for 10 minutes. Thesupernatant was removed and the cells at the bottom weresuspended in MEM solution and centrifuged again. Finally,the supernatant was removed and the cells inside wereresuspended in the complete culture medium, composed ofNeurobasal-A-Medium, 10% fetal bovine serum, 1% PSN,and 0.1% 50 × B27 supplement (Gibco). The neuronswere then seeded on poly-L-lysine (Sigma) precoated 96-well microplates for MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay, or seeded on glassbottom culture dishes (MatTek Corporation, USA) for intra-cellular calcium recording under confocal microscope. Thecultures were maintained at 37◦C in a 5% CO2 humidifiedincubator. We used serum-free medium (Neurobasal-A-Medium, 1% PSN, 0.1% 50 × B27 supplement; Gibco),complemented with 1 µM cytosine arabinoside (Sigma), togradually replace complete medium. The addition of aselective DNA synthesis inhibitor (cytosine arabinoside) inculture eliminated proliferative glial cells allowed adequatenumbers of neurons to selectively survive.
2.3. Cell Viability Assay. Cell viability with MA treatmentwas determined by measuring the mitochondrial dehydro-genase activity using MTT assay in triplicate. The corticalneurons, with a seeding number of 104 cells per well in a 96-well microplate, were cultured for 6 days before use. The cellswere incubated with MA at serial concentrations (0, 25, 50,100, 200, and 400 µM) for 48 hours at 37◦C. After 44 hours,10 µL MTT solution (1 g/L; Sigma) was added into each well,and the microplate was incubated for 4 hours. Formazancrystals, produced by mitochondrial dehydrogenase activityin viable neurons, were dissolved by addition of 150 µLDMSO on a shaker at room temperature. Absorbances wereread at 570 nm using a microplate reader.
Vehicle controls were set up in parallel to offset thebackground during process. Each test was performed intriplicate in the 96-well microplates. The experiment wasrepeated for 3 times. Cytotoxic effect was analyzed by
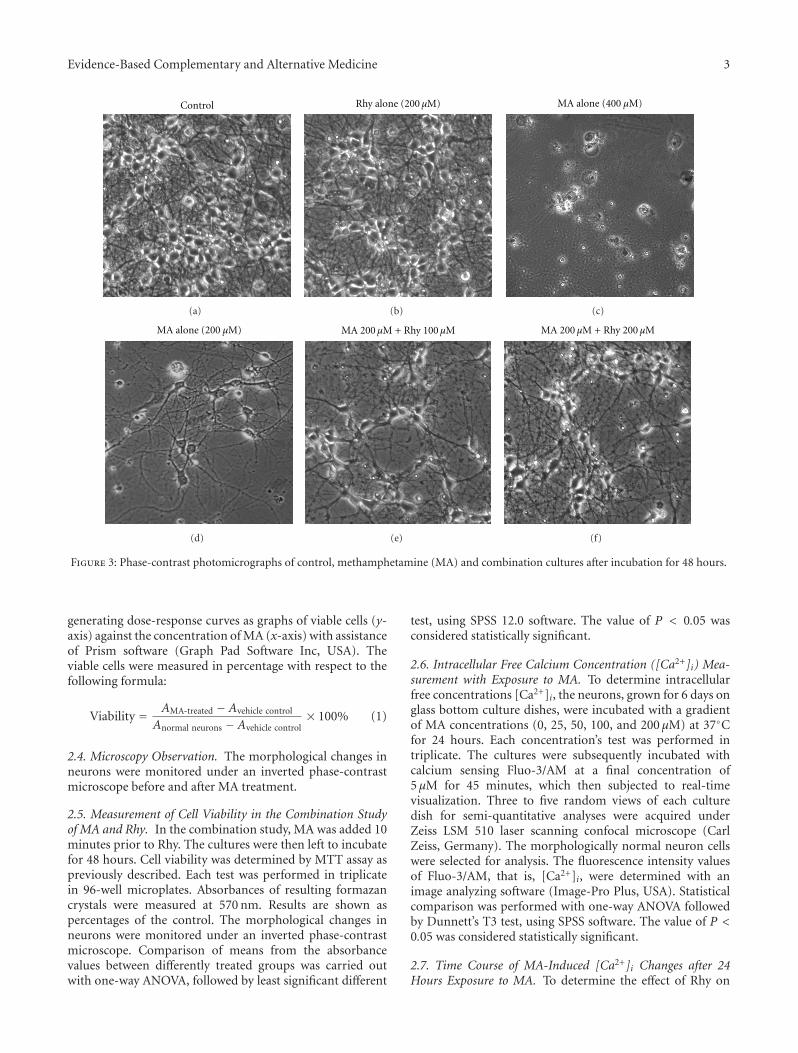
Evidence-Based Complementary and Alternative Medicine 3
Control
(a)
Rhy alone (200 µM)
(b)
MA alone (400 µM)
(c)
MA alone (200 µM)
(d)
MA 200 µM + Rhy 100 µM
(e)
MA 200 µM + Rhy 200 µM
(f)
Figure 3: Phase-contrast photomicrographs of control, methamphetamine (MA) and combination cultures after incubation for 48 hours.
generating dose-response curves as graphs of viable cells (y-axis) against the concentration of MA (x-axis) with assistanceof Prism software (Graph Pad Software Inc, USA). Theviable cells were measured in percentage with respect to thefollowing formula:
Viability = AMA-treated − Avehicle control
Anormal neurons − Avehicle control× 100% (1)
2.4. Microscopy Observation. The morphological changes inneurons were monitored under an inverted phase-contrastmicroscope before and after MA treatment.
2.5. Measurement of Cell Viability in the Combination Studyof MA and Rhy. In the combination study, MA was added 10minutes prior to Rhy. The cultures were then left to incubatefor 48 hours. Cell viability was determined by MTT assay aspreviously described. Each test was performed in triplicatein 96-well microplates. Absorbances of resulting formazancrystals were measured at 570 nm. Results are shown aspercentages of the control. The morphological changes inneurons were monitored under an inverted phase-contrastmicroscope. Comparison of means from the absorbancevalues between differently treated groups was carried outwith one-way ANOVA, followed by least significant different
test, using SPSS 12.0 software. The value of P < 0.05 wasconsidered statistically significant.
2.6. Intracellular Free Calcium Concentration ([Ca2+]i) Mea-surement with Exposure to MA. To determine intracellularfree concentrations [Ca2+]i, the neurons, grown for 6 days onglass bottom culture dishes, were incubated with a gradientof MA concentrations (0, 25, 50, 100, and 200 µM) at 37◦Cfor 24 hours. Each concentration’s test was performed intriplicate. The cultures were subsequently incubated withcalcium sensing Fluo-3/AM at a final concentration of5 µM for 45 minutes, which then subjected to real-timevisualization. Three to five random views of each culturedish for semi-quantitative analyses were acquired underZeiss LSM 510 laser scanning confocal microscope (CarlZeiss, Germany). The morphologically normal neuron cellswere selected for analysis. The fluorescence intensity valuesof Fluo-3/AM, that is, [Ca2+]i, were determined with animage analyzing software (Image-Pro Plus, USA). Statisticalcomparison was performed with one-way ANOVA followedby Dunnett’s T3 test, using SPSS software. The value of P <0.05 was considered statistically significant.
2.7. Time Course of MA-Induced [Ca2+]i Changes after 24Hours Exposure to MA. To determine the effect of Rhy on
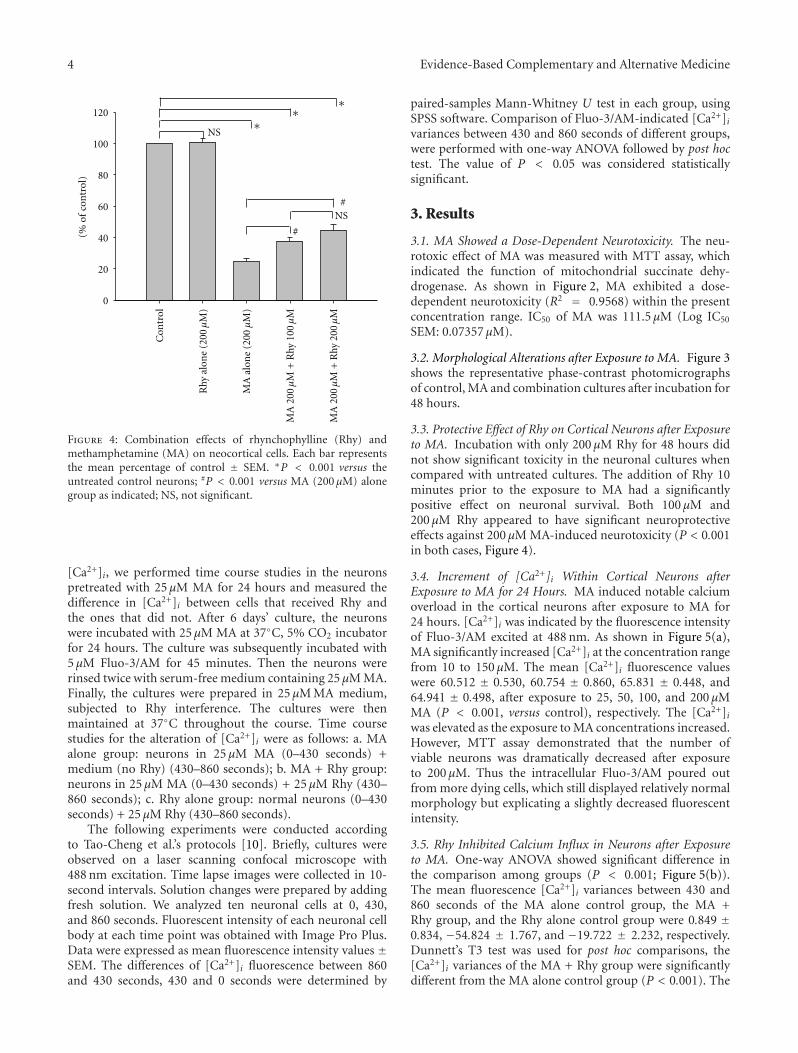
4 Evidence-Based Complementary and Alternative Medicine(%
of
con
trol
)
120
100
80
60
40
20
0
#
#
NS
NS
Con
trol
Rhy
alo
ne
(200
µM
)
MA
alo
ne
(200
µM
)
MA
200
µM
+R
hy 1
00 µ
M
MA
200
µM
+R
hy 2
00 µ
M
∗∗
∗
Figure 4: Combination effects of rhynchophylline (Rhy) andmethamphetamine (MA) on neocortical cells. Each bar representsthe mean percentage of control ± SEM. ∗P < 0.001 versus theuntreated control neurons; #P < 0.001 versus MA (200 µM) alonegroup as indicated; NS, not significant.
[Ca2+]i, we performed time course studies in the neuronspretreated with 25 µM MA for 24 hours and measured thedifference in [Ca2+]i between cells that received Rhy andthe ones that did not. After 6 days’ culture, the neuronswere incubated with 25 µM MA at 37◦C, 5% CO2 incubatorfor 24 hours. The culture was subsequently incubated with5 µM Fluo-3/AM for 45 minutes. Then the neurons wererinsed twice with serum-free medium containing 25 µM MA.Finally, the cultures were prepared in 25 µM MA medium,subjected to Rhy interference. The cultures were thenmaintained at 37◦C throughout the course. Time coursestudies for the alteration of [Ca2+]i were as follows: a. MAalone group: neurons in 25 µM MA (0–430 seconds) +medium (no Rhy) (430–860 seconds); b. MA + Rhy group:neurons in 25 µM MA (0–430 seconds) + 25 µM Rhy (430–860 seconds); c. Rhy alone group: normal neurons (0–430seconds) + 25 µM Rhy (430–860 seconds).
The following experiments were conducted accordingto Tao-Cheng et al.’s protocols [10]. Briefly, cultures wereobserved on a laser scanning confocal microscope with488 nm excitation. Time lapse images were collected in 10-second intervals. Solution changes were prepared by addingfresh solution. We analyzed ten neuronal cells at 0, 430,and 860 seconds. Fluorescent intensity of each neuronal cellbody at each time point was obtained with Image Pro Plus.Data were expressed as mean fluorescence intensity values ±SEM. The differences of [Ca2+]i fluorescence between 860and 430 seconds, 430 and 0 seconds were determined by
paired-samples Mann-Whitney U test in each group, usingSPSS software. Comparison of Fluo-3/AM-indicated [Ca2+]ivariances between 430 and 860 seconds of different groups,were performed with one-way ANOVA followed by post hoctest. The value of P < 0.05 was considered statisticallysignificant.
3. Results
3.1. MA Showed a Dose-Dependent Neurotoxicity. The neu-rotoxic effect of MA was measured with MTT assay, whichindicated the function of mitochondrial succinate dehy-drogenase. As shown in Figure 2, MA exhibited a dose-dependent neurotoxicity (R2 = 0.9568) within the presentconcentration range. IC50 of MA was 111.5 µM (Log IC50
SEM: 0.07357 µM).
3.2. Morphological Alterations after Exposure to MA. Figure 3shows the representative phase-contrast photomicrographsof control, MA and combination cultures after incubation for48 hours.
3.3. Protective Effect of Rhy on Cortical Neurons after Exposureto MA. Incubation with only 200 µM Rhy for 48 hours didnot show significant toxicity in the neuronal cultures whencompared with untreated cultures. The addition of Rhy 10minutes prior to the exposure to MA had a significantlypositive effect on neuronal survival. Both 100 µM and200 µM Rhy appeared to have significant neuroprotectiveeffects against 200 µM MA-induced neurotoxicity (P < 0.001in both cases, Figure 4).
3.4. Increment of [Ca2+]i Within Cortical Neurons afterExposure to MA for 24 Hours. MA induced notable calciumoverload in the cortical neurons after exposure to MA for24 hours. [Ca2+]i was indicated by the fluorescence intensityof Fluo-3/AM excited at 488 nm. As shown in Figure 5(a),MA significantly increased [Ca2+]i at the concentration rangefrom 10 to 150 µM. The mean [Ca2+]i fluorescence valueswere 60.512 ± 0.530, 60.754 ± 0.860, 65.831 ± 0.448, and64.941 ± 0.498, after exposure to 25, 50, 100, and 200 µMMA (P < 0.001, versus control), respectively. The [Ca2+]iwas elevated as the exposure to MA concentrations increased.However, MTT assay demonstrated that the number ofviable neurons was dramatically decreased after exposureto 200 µM. Thus the intracellular Fluo-3/AM poured outfrom more dying cells, which still displayed relatively normalmorphology but explicating a slightly decreased fluorescentintensity.
3.5. Rhy Inhibited Calcium Influx in Neurons after Exposureto MA. One-way ANOVA showed significant difference inthe comparison among groups (P < 0.001; Figure 5(b)).The mean fluorescence [Ca2+]i variances between 430 and860 seconds of the MA alone control group, the MA +Rhy group, and the Rhy alone control group were 0.849 ±0.834, −54.824 ± 1.767, and −19.722 ± 2.232, respectively.Dunnett’s T3 test was used for post hoc comparisons, the[Ca2+]i variances of the MA + Rhy group were significantlydifferent from the MA alone control group (P < 0.001). The
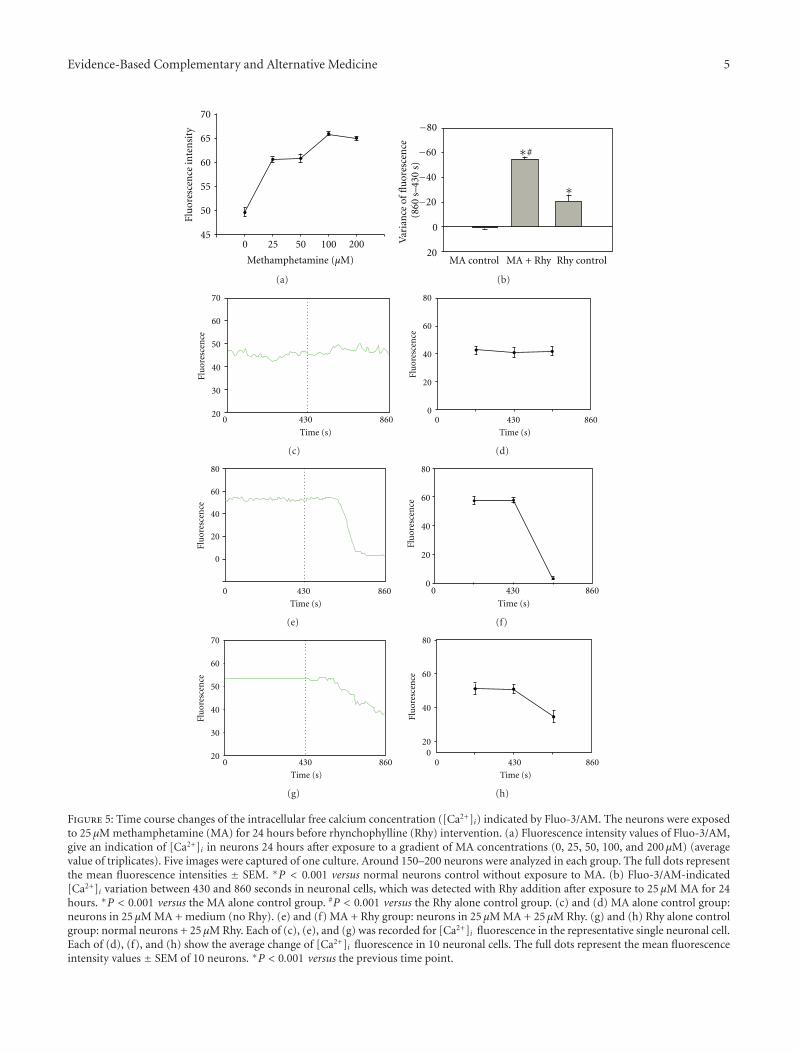
Evidence-Based Complementary and Alternative Medicine 5
70
65
60
55
50
450 25 50 100 200
Flu
ores
cen
ce in
ten
sity
Methamphetamine (µM)
(a)
0
20
−80
−60
−40
−20
MA control MA + Rhy Rhy control
Var
ian
ce o
f fl
uor
esce
nce
(860
s–4
30 s
)
#∗
∗
(b)
70
60
50
40
30
20
Time (s)
Flu
ores
cen
ce
0 430 860
(c)
80
60
40
20
0Fl
uor
esce
nce
Time (s)
0 430 860
(d)
80
60
40
20
0
Flu
ores
cen
ce
Time (s)
0 430 860
(e)
80
60
40
20
0
Flu
ores
cen
ce
Time (s)
0 430 860
(f)
60
50
40
30
20
Flu
ores
cen
ce
70
Time (s)
0 430 860
(g)
80
60
40
200
Flu
ores
cen
ce
Time (s)
0 430 860
(h)
Figure 5: Time course changes of the intracellular free calcium concentration ([Ca2+]i) indicated by Fluo-3/AM. The neurons were exposedto 25 µM methamphetamine (MA) for 24 hours before rhynchophylline (Rhy) intervention. (a) Fluorescence intensity values of Fluo-3/AM,give an indication of [Ca2+]i in neurons 24 hours after exposure to a gradient of MA concentrations (0, 25, 50, 100, and 200 µM) (averagevalue of triplicates). Five images were captured of one culture. Around 150–200 neurons were analyzed in each group. The full dots representthe mean fluorescence intensities ± SEM. ∗P < 0.001 versus normal neurons control without exposure to MA. (b) Fluo-3/AM-indicated[Ca2+]i variation between 430 and 860 seconds in neuronal cells, which was detected with Rhy addition after exposure to 25 µM MA for 24hours. ∗P < 0.001 versus the MA alone control group. #P < 0.001 versus the Rhy alone control group. (c) and (d) MA alone control group:neurons in 25 µM MA + medium (no Rhy). (e) and (f) MA + Rhy group: neurons in 25 µM MA + 25 µM Rhy. (g) and (h) Rhy alone controlgroup: normal neurons + 25 µM Rhy. Each of (c), (e), and (g) was recorded for [Ca2+]i fluorescence in the representative single neuronal cell.Each of (d), (f), and (h) show the average change of [Ca2+]i fluorescence in 10 neuronal cells. The full dots represent the mean fluorescenceintensity values ± SEM of 10 neurons. ∗P < 0.001 versus the previous time point.

6 Evidence-Based Complementary and Alternative Medicine
inhibition of the intracellular calcium influx by Rhy alone(Rhy control group) is significantly stronger in neurons afterexposure to MA than in untreated normal neurons (P <0.001).
Figures 5(c)–5(h) also shows time course studies of the[Ca2+]i of the cultures exposed to 25 µM MA for 24 hoursbefore Rhy intervention. These results were compared to theeffect of the addition of medium (without Rhy) (as reference)on the [Ca2+]i, which showed no change in [Ca2+]i (Figures5(c) and 5(d)). As seen in Figures 5(e) and 5(f), the paired-samples t-tests between 430 and 860 seconds showed that theaddition of Rhy at 430 seconds dramatically attenuated thefluorescence intensity of Fluo-3/AM calcium indicators (P <0.001). In the Rhy alone control group, the normal neuronswere stimulated with 25 µM Rhy for 430 seconds after normalcell recording. Addition of Rhy alone significantly decreasedthe fluorescence intensity of intracellular Fluo-3/AM (P <0.01), which is consistent with the calcium channel blockingproperties of Rhy (Figures 5(g) and 5(h)).
4. Discussion
The majority of researches have been limited to MA neuro-toxicity to dopaminergic neurons, in which the dopaminereuptake is inhibited by MA resulting in intracellular high-concentration dopamine [11]. Actually, MA can induceneural impairment in other brain areas too. For example,neurodegeneration has been found in the piriform andparietal cortex, thalamus and hippocampus, areas whichreceive only a sparse dopaminergic input, and in non-dopamine striatal elements [7]. The main objective ofthe present study is to determine whether the neurotoxiceffects of MA as a psychostimulatory drug, which causesneural death on rodent cortical cells in vitro, can beattenuated by the pretreatment with Rhy in cell cultures.The neurotoxicity is thought to be linked to intracellularcalcium levels, which appear to correlate with MA incubationin a dose-dependent manner. The significant neurotoxicitycaused by several types of amphetamines in rat neocorticalneurons was previously reported to be accompanied byinternucleosomal DNA cleavage and nuclear breakdown, aswell as differential expression of the anti- and proapoptoticbcl-xL/S splice variants, indicating an involvement of theapoptotic pathways in amphetamine neurotoxicity [12]. Ithas been confirmed that MA can be neurotoxic to corticalcells directly, with the increase in calcium load. The [Ca2+]i istherefore suggested to play a pivotal role in MA-induced celldeath. MA is a cationic lipophilic molecule that can diffuseinto the mitochondria and affects the mitochondrial calciumATPase, which is responsible for pumping Ca2+ into theinner mitochondrial space for storage purposes. The releaseof these stores, through the mitochondrial membrane thatis made permeable by formation of transition pores, triggersthe activation of the cell death pathway. Cell death requiresthe involvement of cysteine proteases and caspases, whichare normally associated with the mitochondrial envelope intheir inactive state [7]. Consequently, Ca2+ influx is followedby mitochondrial damage and reactive species formation. Itis well documented that MA induced neurological damage
caused by the above described mechanisms, because it isresponsible for Ca2+ influx due to its effect on ionotropicglutamate receptors [11].
Rhy has been proven to be a potent calcium channelantagonist in blood vessels and it is assumed to act as one ofthe noncompetitive antagonists for glutamate receptors [2].Therefore Rhy is thought to contribute to the neuroprotec-tive and anticonvulsant activity of plant extracts of Uncariaspecies [13]. Moreover, another in vivo behavioral studyindicates that Rhy has protective features against harmfuleffects of MA [14]. To conclude, our study demonstrates thatRhy decreases the high level of intracellular calcium inducedby MA in cultured neurons, through the direct blockadeof calcium channels and/or the inhibition of ionotropicglutamate receptors.
This is the first report demonstrating an inhibitory effectof Rhy against MA impairment in cultured neurons in vitro.Our results suggested that Rhy can protect the neuronalcultures against MA exposure and promptly attenuate intra-cellular calcium overload triggered by the MA toxin.
Conflict of Interests
The authors declare that they have no conflict of interests.
References
[1] J. S. Shi, J. X. Yu, X. P. Chen, and R. X. Xu, “Pharmacologicalactions of Uncaria alkaloids, rhynchophylline and isorhyn-chophylline,” Acta Pharmacologica Sinica, vol. 24, no. 2, pp.97–101, 2003.
[2] J. Zhou and S. Zhou, “Antihypertensive and neuroprotectiveactivities of rhynchophylline: the role of rhynchophyllinein neurotransmission and ion channel activity,” Journal ofEthnopharmacology, vol. 132, no. 1, pp. 15–27, 2010.
[3] K. Suk, S. Y. Kim, K. Leem et al., “Neuroprotection bymethanol extract of Uncaria rhynchophylla against globalcerebral ischemia in rats,” Life Sciences, vol. 70, no. 21, pp.2467–2480, 2002.
[4] Y. Shimada, H. Goto, T. Itoh et al., “Evaluation of theprotective effects of alkaloids isolated from the hooks andstems of Uncaria sinensis on glutamate-induced neuronaldeath in cultured cerebellar granule cells from rats,” Journal ofPharmacy and Pharmacology, vol. 51, no. 6, pp. 715–722, 1999.
[5] I. N. Krasnova and J. L. Cadet, “Methamphetamine toxicityand messengers of death,” Brain Research Reviews, vol. 60, no.2, pp. 379–407, 2009.
[6] N. D. Volkow, L. Chang, G. J. Wang et al., “Loss of dopaminetransporters in methamphetamine abusers recovers withprotracted abstinence,” Journal of Neuroscience, vol. 21, no. 23,pp. 9414–9418, 2001.
[7] C. Davidson, A. J. Gow, T. H. Lee, and E. H. Ellinwood,“Methamphetamine neurotoxicity: necrotic and apoptoticmechanisms and relevance to human abuse and treatment,”Brain Research Reviews, vol. 36, no. 1, pp. 1–22, 2001.
[8] J. L. Cadet, S. Jayanthi, and X. Deng, “Speed kills: cellular andmolecular bases of methamphetamine-induced nerve termi-nal degeneration and neuronal apoptosis,” FASEB Journal, vol.17, no. 13, pp. 1775–1788, 2003.
[9] J. L. Zhou, J. H. Liang, and C. L. Li, “Inhibition ofmethamphetamine-induced apoptosis by the calcium channel

Evidence-Based Complementary and Alternative Medicine 7
blocker verapamil in rat cerebellar neurons,” Beijing Da XueXue Bao, vol. 36, no. 4, pp. 361–365, 2004.
[10] J. H. Tao-Cheng, L. Vinade, C. Smith et al., “Sustained eleva-tion of calcium induces Ca2+/calmodulin-dependent proteinkinase II clusters in hippocampal neurons,” Neuroscience, vol.106, no. 1, pp. 69–78, 2001.
[11] E. L. Riddle, A. E. Fleckenstein, and G. R. Hanson, “Mech-anisms of methamphetamine-induced dopaminergic neuro-toxicity,” AAPS Journal, vol. 8, no. 2, pp. E413–E418, 2006.
[12] G. Stumm, J. Schlegel, T. Schafer et al., “Amphetamines induceapoptosis and regulation of bcl-x splice variants in neocorticalneurons,” FASEB Journal, vol. 13, no. 9, pp. 1065–1072, 1999.
[13] T. H. Kang, Y. Murakami, K. Matsumoto et al., “Rhyn-chophylline and isorhynchophylline inhibit NMDA receptorsexpressed in Xenopus oocytes,” European Journal of Pharma-cology, vol. 455, no. 1, pp. 27–34, 2002.
[14] J. Y. Zhou, Z. X. Mo, and S. W. Zhou, “Rhynchophyllinedown-regulates NR2B expression in cortex and hippocampalCA1 area of amphetamine-induced conditioned place ratpreference,” Archives of Pharmacal Research, vol. 33, no. 4, pp.557–565, 2010.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 128019, 12 pagesdoi:10.1155/2012/128019
Research Article
Involvement of Heme Oxygenase-1 Induction in theCytoprotective and Immunomodulatory Activities of Violapatrinii in Murine Hippocampal and Microglia Cells
Bin Li,1 Dong-Sung Lee,1 Hyun-Gyu Choi,1 Kyoung-Su Kim,1 Gil-Saeng Jeong,2
Ren Bo An,3 and Youn-Chul Kim1
1 Standardized Material Bank for New Botanical Drugs, College of Pharmacy, Wonkwang University, Iksan 570-749, Republic of Korea2 College of Pharmacy, Keimyung University, Daegu 704-701, Republic of Korea3 Key Laboratory of Natural Resources of Changbai Mountain & Functional Molecules, Yanbian University, Ministry of Education,Yanji, Jilin 133-000, China
Correspondence should be addressed to Youn-Chul Kim, [email protected]
Received 18 November 2011; Revised 4 January 2012; Accepted 10 January 2012
Academic Editor: Mahmud Tareq Hassan Khan
Copyright © 2012 Bin Li et al. This is an open access article distributed under the Creative Commons Attribution License, whichpermits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
A number of diseases that lead to injury of the central nervous system are caused by oxidative stress and inflammation in the brain.In this study, NNMBS275, consisting of the ethanol extract of Viola patrinii, showed potent antioxidative and anti-inflammatoryactivity in murine hippocampal HT22 cells and BV2 microglia. NNMBS275 increased cellular resistance to oxidative injury causedby glutamate-induced neurotoxicity and reactive oxygen species generation in HT22 cells. In addition, the anti-inflammatoryeffects of NNMBS275 were demonstrated by the suppression of proinflammatory mediators, including proinflammatory enzymes(inducible nitric oxide synthase and cyclooxygenase-2) and cytokines (tumor necrosis factor-α and interleukin-1β). Furthermore,we found that the neuroprotective and anti-inflammatory effects of NNMBS275 were linked to the upregulation of nucleartranscription factor-E2-related factor 2-dependent expression of heme oxygenase-1 in HT22 and BV2 cells. These results suggestthat NNMBS275 possesses therapeutic potential against neurodegenerative diseases that are induced by oxidative stress andneuroinflammation.
1. Introduction
Brain tissues are vulnerable to oxidative stress, which mayoccur physiologically as a component of the aging process, orpathologically as a result of neurodegenerative disease [1, 2].Oxidative stress, or the accumulation of reactive oxygenspecies (ROS), not only results in accidental damage to cells,but also actively triggers intracellular signaling pathways thatlead to cell death. Neuronal oxidative stress has been pos-tulated to be the underlying basis for neuronal cell death inneurodegenerative diseases such as Alzheimer’s disease andParkinson’s disease [3–5]. Glutamate is the main excitatoryneurotransmitter in the central nervous system (CNS).However, at high concentrations, glutamate has been shownto induce neuronal cell death via both receptor-initiatedexcitotoxicity and non-receptor-mediated oxidative stress
[6, 7]. In this study, immortalized mouse hippocampalHT22 cells were used as in vitro models for studyingthe mechanism of oxidative glutamate toxicity. Glutamate-induced neurotoxicity in HT22 cells has been characterizedas an intracellular process attributable to the inhibitionof cystine uptake via the cystine/glutamate antiporter sys-tem Xc−, eventually leading to programmed cell deathdue to glutathione depletion and accumulation of ROS[8, 9].
Heme oxygenase 1 (HO-1), also referred to as HSP32,belongs to the HSP family and protects mammalian cellsfrom oxidative stress by degrading toxic heme into freeiron, carbon monoxide, and biliverdin/bilirubin. AlthoughHO-1 does not directly catalyze an antioxidant reaction, itsinduction is generally considered an adaptive cytoprotectiveresponse against the toxicity of oxidative stress [10–12]. The

2 Evidence-Based Complementary and Alternative Medicine
induction of HO-1 is primarily regulated at the transcrip-tional level, and its induction by various inducers is relatedto the nuclear transcription factor-E2-related factor 2 (Nrf2)[13]. Nrf-2 is a basic leucine zipper transcription factor thatresides in the cytoplasm bound to its inhibitor protein, Keap1, and translocates to the nucleus after stimulation. It thenforms heterodimers with small oncogene family proteins forthe selective recognition of the antioxidant responsive ele-ment on target genes, resulting in the regulation of gene ex-pression of phase II detoxifying enzymes, including HO-1[14, 15]. HO-1 induction occurs in a wide variety of cells, in-cluding neuronal and nonneuronal brain cells. Previousstudies have reported that the cerebellar granule cells harvest-ed from transgenic mice designed to overexpress HO-1 inneurons appear to be relatively resistant to H2O2-mediat-ed oxidative damage in vitro [16]. Furthermore, several re-searchers have reported that phytochemicals, such as resver-atrol, curcumin, and epigallocatechin-3-gallate, exhibit thesignificant therapeutic benefit in the case of oxidative stress-induced neuronal damage via the induction of HO-1 [17–19].
Along with neuronal oxidative stress, neuroinflammationalso contributes to the pathogenesis of several neurodegen-erative disorders [20, 21]. Microglia, the immunocompetentresident cells of the brain, play the main role in innateimmune defense in the CNS. Several studies have shownthat excessive activation of microglia increases neurotoxicityand, therefore, contributes to neuronal injury through therelease of various proinflammatory cytokines and inflamma-tory mediators such as nitric oxide (NO), prostaglandins,tumor necrosis factor (TNF)-α, and interleukin (IL)-1β[22–25]. In addition to its antioxidative effects, recentstudies have also demonstrated anti-inflammatory effectsof HO-1 in a number of inflammation models [26–29].The anti-inflammatory action of HO-1 is mediated byinhibiting the production of proinflammatory cytokines andchemokines such as TNF-α, IL-1β, and IL-6 in activatedmacrophages. HO-1 and its product, carbon monoxide, canalso suppress the expression of proinflammatory COX-2 andiNOS, thereby reducing the production of COX-2-derivedprostaglandin E2 (PGE2) and iNOS-derived NO [30–33].The anti-inflammatory action of HO-1 in microglia has alsobeen reported. Therefore, regulating the degree of generationof oxidative stress and microglial activation via the upregula-tion of HO-1 may be considered an important interventionin formulating a strategy for the treatment of neurologicaldiseases.
Violae herba has been widely used in traditional Chinesemedicine to treat carbuncles, boils, and other cutaneous andsubcutaneous pyogenic infections. Sources of violae herbainclude Viola patrinii, V. hederacea, V. arvensis, and V. odor-ata. Previous phytochemical studies of Viola species havereported the isolation of cyclotides and several flavonoid gly-cosides [34–37]. However, little is known about the biologicalactivities of these plants. As a part of ongoing research toidentify natural products that can induce HO-1 expressionin vitro, we have shown here that the ethanol extract of V.patrinii (NNMBS275) was shown to significantly increase thelevels of HO-1 in both BV2 microglia and HT22 cells.
2. Materials and Methods
2.1. Chemicals and Reagents. Dulbecco’s modified Eagle’smedium (DMEM), fetal bovine serum (FBS), and othertissue culture reagents were purchased from GIBCO BRLCo. (Grand Island, NY, USA). Tin protoporphyrin IX (SnPPIX; inhibitor of HO activity) was obtained from PorphyrinProducts (Logan, UT, USA). Cobalt protoporphyrin IX(CoPP; HO-1 inducer), Trolox, and all other chemicals wereobtained from Sigma Chemical Co. (St. Louis, MO, USA).TRIzol reagent and a polymerase chain reaction (PCR)kit were purchased from Invitrogen Corporation (Carlsbad,CA, USA). Primary antibodies, including mouse/goat/rabbitanti-HO-1, anti-COX-2, and anti-iNOS, and secondaryantibodies were purchased from Santa Cruz Biotechnology(Santa Cruz, CA, USA). Enzyme-linked immunosorbentassay (ELISA) kits for PGE2, TNF-α, and IL-1β werepurchased from R & D Systems, Inc. (Minneapolis, MN,USA).
2.2. Plant Material and Preparation of NNMBS275. Theaerial parts of Viola patrinii DC (Violaceae) were collected inthe Botanical Garden of Wonkwang University, Iksan, Korea,in June 2010. A voucher specimen (no. WP2010-26) wasauthenticated by Dr. Kyu-Kwan Jang, the Botanical Gardenof Wonkwang University, and deposited at the herbariumof the College of Pharmacy, Wonkwang University. Driedand pulverized V. patrinii (50 g) was extracted twice withethanol (300 mL) at room temperature for 3 days, and theextract was concentrated in vacuo to obtain an ethanolextract (NNMBS275, 720 mg). NNMBS275 was deposited atthe Standardized Material Bank for New Botanical Drugs,Wonkwang University. For each experiment, NNMBS275was dissolved in dimethylsulfoxide (final culture concen-tration, 0.05%). Serum-free medium was used as a vehiclecontrol. Preliminary studies indicated that the solvent hadno effect on cell viability at the concentration used.
2.3. High-Performance Liquid Chromatography (HPLC).Chromatographic experiments were performed on aSNKNM series HPLC instrument equipped with a sampleinjector and a diode-assay UV/Vis detector (DAD) (SYKAM,Germany). For all experiments, an Inertsil ODS C-18column (4.6 × 150 nm; 5 μm, GL Science Inc., Tokyo, Japan)was used as the stationary phase, and the injection volumewas 20 μL. The mobile phase was composed of water (A)and acetonitrile (B), with an applied gradient of 20% Bincreasing to 60% B in 40 min. The column was cleanedwith 10% B for 20 min, and then the system was equilibratedfor 20 min with the starting conditions. Flow rate was0.7 mL/min, and the detection wavelength was adjusted to210 nm.
2.4. Cell Culture and Viability Assay. Mouse hippocampalHT22 cells were received from Prof. In-Hee Mook at SeoulNational University (Seoul, Korea), and BV2 microglia cellswere received from Prof. Hyun Park at Wonkwang University.The cells were maintained at 5 × 105 cells/mL in DMEM
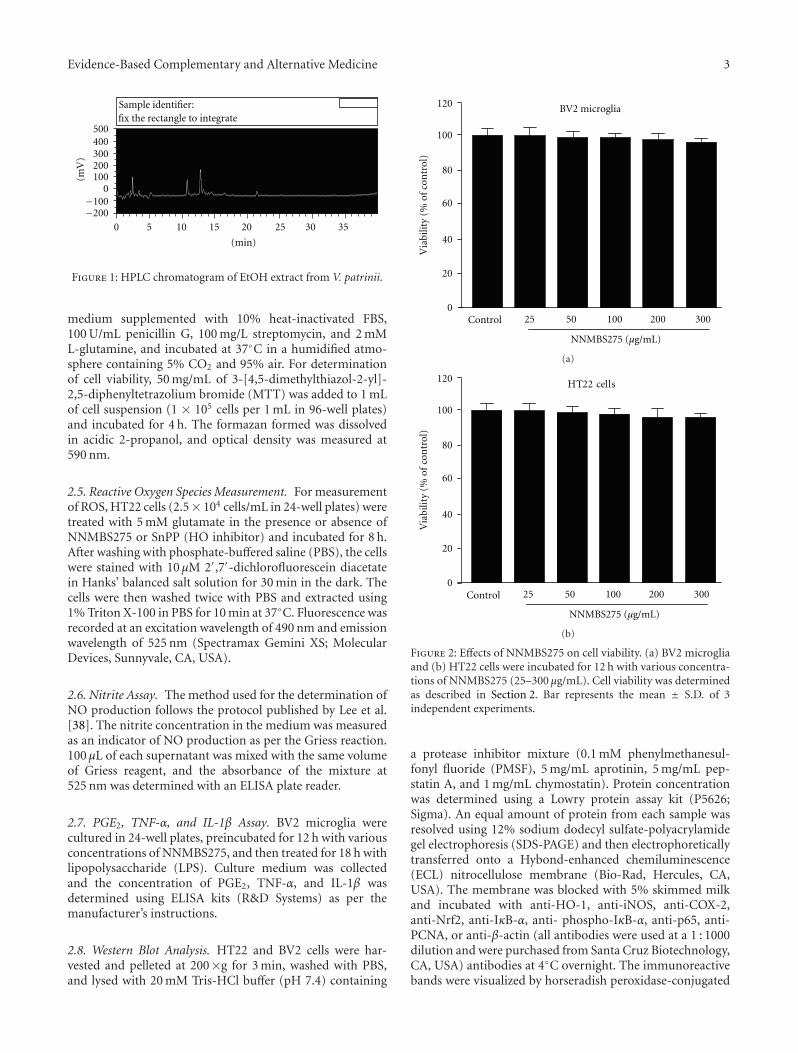
Evidence-Based Complementary and Alternative Medicine 3
0 5 10 15 20 25 30 35−200−100
0100200300400500
(mV
)
Sample identifier:fix the rectangle to integrate
(min)
Figure 1: HPLC chromatogram of EtOH extract from V. patrinii.
medium supplemented with 10% heat-inactivated FBS,100 U/mL penicillin G, 100 mg/L streptomycin, and 2 mML-glutamine, and incubated at 37◦C in a humidified atmo-sphere containing 5% CO2 and 95% air. For determinationof cell viability, 50 mg/mL of 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) was added to 1 mLof cell suspension (1 × 105 cells per 1 mL in 96-well plates)and incubated for 4 h. The formazan formed was dissolvedin acidic 2-propanol, and optical density was measured at590 nm.
2.5. Reactive Oxygen Species Measurement. For measurementof ROS, HT22 cells (2.5× 104 cells/mL in 24-well plates) weretreated with 5 mM glutamate in the presence or absence ofNNMBS275 or SnPP (HO inhibitor) and incubated for 8 h.After washing with phosphate-buffered saline (PBS), the cellswere stained with 10 μM 2′,7′-dichlorofluorescein diacetatein Hanks’ balanced salt solution for 30 min in the dark. Thecells were then washed twice with PBS and extracted using1% Triton X-100 in PBS for 10 min at 37◦C. Fluorescence wasrecorded at an excitation wavelength of 490 nm and emissionwavelength of 525 nm (Spectramax Gemini XS; MolecularDevices, Sunnyvale, CA, USA).
2.6. Nitrite Assay. The method used for the determination ofNO production follows the protocol published by Lee et al.[38]. The nitrite concentration in the medium was measuredas an indicator of NO production as per the Griess reaction.100 μL of each supernatant was mixed with the same volumeof Griess reagent, and the absorbance of the mixture at525 nm was determined with an ELISA plate reader.
2.7. PGE2, TNF-α, and IL-1β Assay. BV2 microglia werecultured in 24-well plates, preincubated for 12 h with variousconcentrations of NNMBS275, and then treated for 18 h withlipopolysaccharide (LPS). Culture medium was collectedand the concentration of PGE2, TNF-α, and IL-1β wasdetermined using ELISA kits (R&D Systems) as per themanufacturer’s instructions.
2.8. Western Blot Analysis. HT22 and BV2 cells were har-vested and pelleted at 200×g for 3 min, washed with PBS,and lysed with 20 mM Tris-HCl buffer (pH 7.4) containing
0
20
40
80
100
60
Control 25 10050 200 300
120
NNMBS275 (μg/mL)
Via
bilit
y (%
of c
ontr
ol)
BV2 microglia
(a)
HT22 cells
Control 25 10050 200 300
NNMBS275 (μg/mL)
0
20
40
80
100
60
120
Via
bilit
y (%
of
con
trol
)
(b)
Figure 2: Effects of NNMBS275 on cell viability. (a) BV2 microgliaand (b) HT22 cells were incubated for 12 h with various concentra-tions of NNMBS275 (25–300 μg/mL). Cell viability was determinedas described in Section 2. Bar represents the mean ± S.D. of 3independent experiments.
a protease inhibitor mixture (0.1 mM phenylmethanesul-fonyl fluoride (PMSF), 5 mg/mL aprotinin, 5 mg/mL pep-statin A, and 1 mg/mL chymostatin). Protein concentrationwas determined using a Lowry protein assay kit (P5626;Sigma). An equal amount of protein from each sample wasresolved using 12% sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) and then electrophoreticallytransferred onto a Hybond-enhanced chemiluminescence(ECL) nitrocellulose membrane (Bio-Rad, Hercules, CA,USA). The membrane was blocked with 5% skimmed milkand incubated with anti-HO-1, anti-iNOS, anti-COX-2,anti-Nrf2, anti-IκB-α, anti- phospho-IκB-α, anti-p65, anti-PCNA, or anti-β-actin (all antibodies were used at a 1 : 1000dilution and were purchased from Santa Cruz Biotechnology,CA, USA) antibodies at 4◦C overnight. The immunoreactivebands were visualized by horseradish peroxidase-conjugated
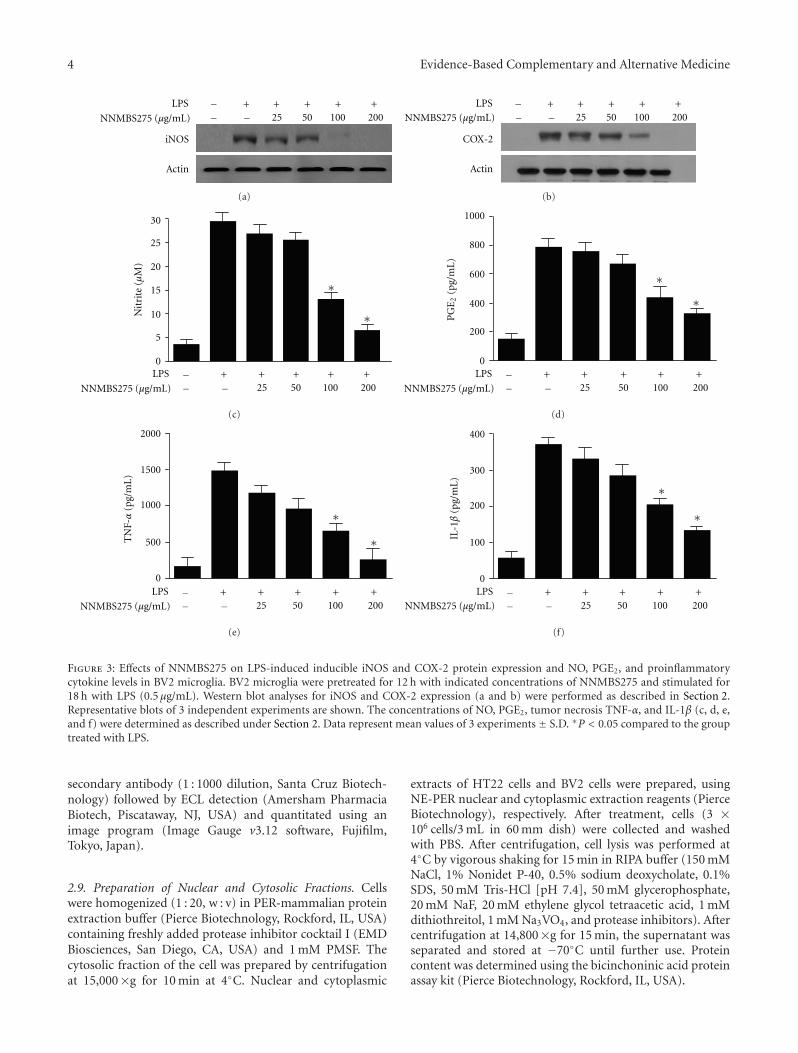
4 Evidence-Based Complementary and Alternative Medicine
iNOS
Actin
+ + + + +25 50 100 200
LPS
NNMBS275 − −−
(μg/mL)
(a)
Actin
+ + + + +25 50 100 200
LPS
− −COX-2
NNMBS275 (μg/mL)−
(b)
30
0
20
15
10
5
25
∗
∗Nit
rite
(μ
M)
+ + + + +25 50 100 200
LPS −− −NNMBS275 (μg/mL)
(c)
∗
∗
+ + + + +25 50 100 200
LPS −− −
0
1000
800
600
400
200
PG
E2
(pg/
mL
)
NNMBS275 (μg/mL)
(d)
∗
∗
+ + + + +25 50 100 200
LPS −− −
0
2000
1500
1000
500TN
F-α
(pg/
mL
)
NNMBS275 (μg/mL)
(e)
∗
∗
+ + + + +25 50 100 200
LPS −− −
0
400
300
200
100
IL-1β
(pg/
mL
)
NNMBS275 (μg/mL)
(f)
Figure 3: Effects of NNMBS275 on LPS-induced inducible iNOS and COX-2 protein expression and NO, PGE2, and proinflammatorycytokine levels in BV2 microglia. BV2 microglia were pretreated for 12 h with indicated concentrations of NNMBS275 and stimulated for18 h with LPS (0.5 μg/mL). Western blot analyses for iNOS and COX-2 expression (a and b) were performed as described in Section 2.Representative blots of 3 independent experiments are shown. The concentrations of NO, PGE2, tumor necrosis TNF-α, and IL-1β (c, d, e,and f) were determined as described under Section 2. Data represent mean values of 3 experiments ± S.D. ∗P < 0.05 compared to the grouptreated with LPS.
secondary antibody (1 : 1000 dilution, Santa Cruz Biotech-nology) followed by ECL detection (Amersham PharmaciaBiotech, Piscataway, NJ, USA) and quantitated using animage program (Image Gauge v3.12 software, Fujifilm,Tokyo, Japan).
2.9. Preparation of Nuclear and Cytosolic Fractions. Cellswere homogenized (1 : 20, w : v) in PER-mammalian proteinextraction buffer (Pierce Biotechnology, Rockford, IL, USA)containing freshly added protease inhibitor cocktail I (EMDBiosciences, San Diego, CA, USA) and 1 mM PMSF. Thecytosolic fraction of the cell was prepared by centrifugationat 15,000×g for 10 min at 4◦C. Nuclear and cytoplasmic
extracts of HT22 cells and BV2 cells were prepared, usingNE-PER nuclear and cytoplasmic extraction reagents (PierceBiotechnology), respectively. After treatment, cells (3 ×106 cells/3 mL in 60 mm dish) were collected and washedwith PBS. After centrifugation, cell lysis was performed at4◦C by vigorous shaking for 15 min in RIPA buffer (150 mMNaCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1%SDS, 50 mM Tris-HCl [pH 7.4], 50 mM glycerophosphate,20 mM NaF, 20 mM ethylene glycol tetraacetic acid, 1 mMdithiothreitol, 1 mM Na3VO4, and protease inhibitors). Aftercentrifugation at 14,800×g for 15 min, the supernatant wasseparated and stored at −70◦C until further use. Proteincontent was determined using the bicinchoninic acid proteinassay kit (Pierce Biotechnology, Rockford, IL, USA).
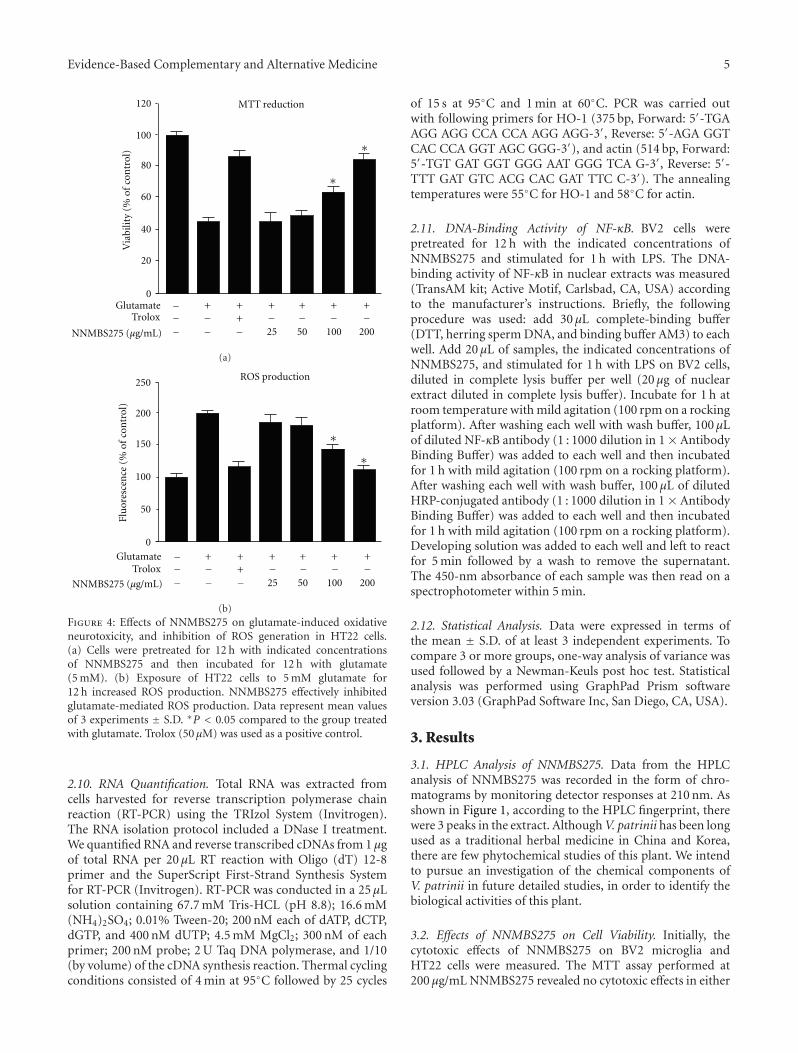
Evidence-Based Complementary and Alternative Medicine 5
0
20
40
80
100
60
GlutamateTrolox
120 MTT reduction
Via
bilit
y (%
of
con
trol
) ∗
∗
+ ++
+ + +
25 50 100 200
−−−
− − − −−
+−
−NNMBS275 (μg/mL)
(a)
Flu
ores
cen
ce (
% o
f co
ntr
ol)
0
50
100
200
250
150
ROS production
∗∗
GlutamateTrolox
+ ++
+ + +
25 50 100 200
−−−
− − − −−
+−
−NNMBS275 (μg/mL)
(b)
Figure 4: Effects of NNMBS275 on glutamate-induced oxidativeneurotoxicity, and inhibition of ROS generation in HT22 cells.(a) Cells were pretreated for 12 h with indicated concentrationsof NNMBS275 and then incubated for 12 h with glutamate(5 mM). (b) Exposure of HT22 cells to 5 mM glutamate for12 h increased ROS production. NNMBS275 effectively inhibitedglutamate-mediated ROS production. Data represent mean valuesof 3 experiments ± S.D. ∗P < 0.05 compared to the group treatedwith glutamate. Trolox (50 μM) was used as a positive control.
2.10. RNA Quantification. Total RNA was extracted fromcells harvested for reverse transcription polymerase chainreaction (RT-PCR) using the TRIzol System (Invitrogen).The RNA isolation protocol included a DNase I treatment.We quantified RNA and reverse transcribed cDNAs from 1 μgof total RNA per 20 μL RT reaction with Oligo (dT) 12-8primer and the SuperScript First-Strand Synthesis Systemfor RT-PCR (Invitrogen). RT-PCR was conducted in a 25 μLsolution containing 67.7 mM Tris-HCL (pH 8.8); 16.6 mM(NH4)2SO4; 0.01% Tween-20; 200 nM each of dATP, dCTP,dGTP, and 400 nM dUTP; 4.5 mM MgCl2; 300 nM of eachprimer; 200 nM probe; 2 U Taq DNA polymerase, and 1/10(by volume) of the cDNA synthesis reaction. Thermal cyclingconditions consisted of 4 min at 95◦C followed by 25 cycles
of 15 s at 95◦C and 1 min at 60◦C. PCR was carried outwith following primers for HO-1 (375 bp, Forward: 5′-TGAAGG AGG CCA CCA AGG AGG-3′, Reverse: 5′-AGA GGTCAC CCA GGT AGC GGG-3′), and actin (514 bp, Forward:5′-TGT GAT GGT GGG AAT GGG TCA G-3′, Reverse: 5′-TTT GAT GTC ACG CAC GAT TTC C-3′). The annealingtemperatures were 55◦C for HO-1 and 58◦C for actin.
2.11. DNA-Binding Activity of NF-κB. BV2 cells werepretreated for 12 h with the indicated concentrations ofNNMBS275 and stimulated for 1 h with LPS. The DNA-binding activity of NF-κB in nuclear extracts was measured(TransAM kit; Active Motif, Carlsbad, CA, USA) accordingto the manufacturer’s instructions. Briefly, the followingprocedure was used: add 30 μL complete-binding buffer(DTT, herring sperm DNA, and binding buffer AM3) to eachwell. Add 20 μL of samples, the indicated concentrations ofNNMBS275, and stimulated for 1 h with LPS on BV2 cells,diluted in complete lysis buffer per well (20 μg of nuclearextract diluted in complete lysis buffer). Incubate for 1 h atroom temperature with mild agitation (100 rpm on a rockingplatform). After washing each well with wash buffer, 100 μLof diluted NF-κB antibody (1 : 1000 dilution in 1× AntibodyBinding Buffer) was added to each well and then incubatedfor 1 h with mild agitation (100 rpm on a rocking platform).After washing each well with wash buffer, 100 μL of dilutedHRP-conjugated antibody (1 : 1000 dilution in 1 × AntibodyBinding Buffer) was added to each well and then incubatedfor 1 h with mild agitation (100 rpm on a rocking platform).Developing solution was added to each well and left to reactfor 5 min followed by a wash to remove the supernatant.The 450-nm absorbance of each sample was then read on aspectrophotometer within 5 min.
2.12. Statistical Analysis. Data were expressed in terms ofthe mean ± S.D. of at least 3 independent experiments. Tocompare 3 or more groups, one-way analysis of variance wasused followed by a Newman-Keuls post hoc test. Statisticalanalysis was performed using GraphPad Prism softwareversion 3.03 (GraphPad Software Inc, San Diego, CA, USA).
3. Results
3.1. HPLC Analysis of NNMBS275. Data from the HPLCanalysis of NNMBS275 was recorded in the form of chro-matograms by monitoring detector responses at 210 nm. Asshown in Figure 1, according to the HPLC fingerprint, therewere 3 peaks in the extract. Although V. patrinii has been longused as a traditional herbal medicine in China and Korea,there are few phytochemical studies of this plant. We intendto pursue an investigation of the chemical components ofV. patrinii in future detailed studies, in order to identify thebiological activities of this plant.
3.2. Effects of NNMBS275 on Cell Viability. Initially, thecytotoxic effects of NNMBS275 on BV2 microglia andHT22 cells were measured. The MTT assay performed at200 μg/mL NNMBS275 revealed no cytotoxic effects in either
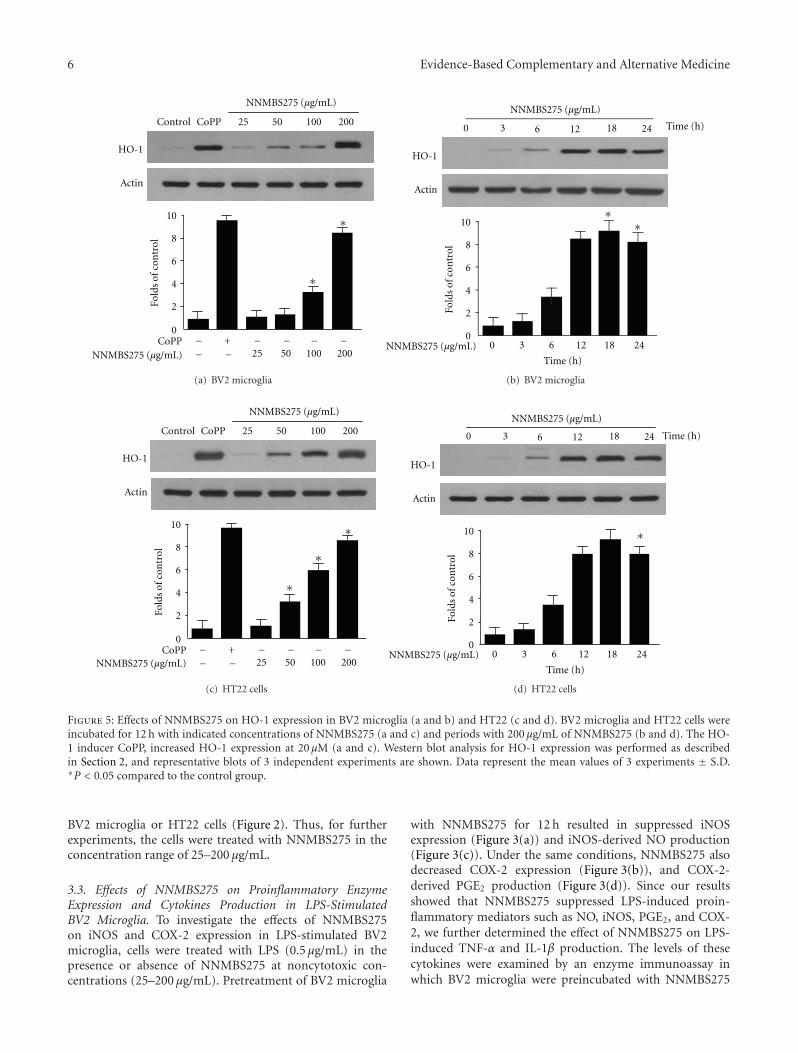
6 Evidence-Based Complementary and Alternative Medicine
Control CoPP 25 10050 200
HO-1
Actin
CoPP0
2
4
6
8
10
Fold
s of
con
trol
+25 50 100 200
− −−−−− −
∗
∗
NNMBS275 (μg/mL)
NNMBS275 (μg/mL)
(a) BV2 microglia
HO-1
Actin
0
2
4
6
8
10
Fold
s of
con
trol
∗∗
0 3 6 12 18 24
0 3 6 12 18 24
Time (h)
Time (h)
NNMBS275 (μg/mL)
NNMBS275 (μg/mL)
(b) BV2 microglia
Control CoPP 25 10050 200
HO-1
Actin
CoPP0
2
4
6
8
10
Fold
s of
con
trol
+25 50 100 200
− −−−−− −
∗
∗
∗
NNMBS275 (μg/mL)
NNMBS275 (μg/mL)
(c) HT22 cells
HO-1
Actin
0
2
4
6
8
10
Fold
s of
con
trol
∗
0 3 6 12 18 24
0 3 6 12 18 24
Time (h)
Time (h)
NNMBS275 (μg/mL)
NNMBS275 (μg/mL)
(d) HT22 cells
Figure 5: Effects of NNMBS275 on HO-1 expression in BV2 microglia (a and b) and HT22 (c and d). BV2 microglia and HT22 cells wereincubated for 12 h with indicated concentrations of NNMBS275 (a and c) and periods with 200 μg/mL of NNMBS275 (b and d). The HO-1 inducer CoPP, increased HO-1 expression at 20 μM (a and c). Western blot analysis for HO-1 expression was performed as describedin Section 2, and representative blots of 3 independent experiments are shown. Data represent the mean values of 3 experiments ± S.D.∗P < 0.05 compared to the control group.
BV2 microglia or HT22 cells (Figure 2). Thus, for furtherexperiments, the cells were treated with NNMBS275 in theconcentration range of 25–200 μg/mL.
3.3. Effects of NNMBS275 on Proinflammatory EnzymeExpression and Cytokines Production in LPS-StimulatedBV2 Microglia. To investigate the effects of NNMBS275on iNOS and COX-2 expression in LPS-stimulated BV2microglia, cells were treated with LPS (0.5 μg/mL) in thepresence or absence of NNMBS275 at noncytotoxic con-centrations (25–200 μg/mL). Pretreatment of BV2 microglia
with NNMBS275 for 12 h resulted in suppressed iNOSexpression (Figure 3(a)) and iNOS-derived NO production(Figure 3(c)). Under the same conditions, NNMBS275 alsodecreased COX-2 expression (Figure 3(b)), and COX-2-derived PGE2 production (Figure 3(d)). Since our resultsshowed that NNMBS275 suppressed LPS-induced proin-flammatory mediators such as NO, iNOS, PGE2, and COX-2, we further determined the effect of NNMBS275 on LPS-induced TNF-α and IL-1β production. The levels of thesecytokines were examined by an enzyme immunoassay inwhich BV2 microglia were preincubated with NNMBS275

Evidence-Based Complementary and Alternative Medicine 7
Con
trol
25 10050 200
HO-1
Actin
NNMBS275 (μg/mL)
(a) BV2 microglia
HO-1
Actin
Con
trol
25 10050 200
NNMBS275 (μg/mL)
(b) HT22 cells
Figure 6: Effects of NNMBS275 on mRNA expression of HO-1 in BV2 microglia and HT22. (a) BV2 microglia and (b) HT22cells were incubated for 6 h with indicated concentrations ofNNMBS275. RNA quantification was performed as described inSection 2, and representative blots of three independent experi-ments are shown.
for 12 h followed by LPS stimulation. As shown in Figures3(e) and 3(f), pretreatment with NNMBS275 resulted in asignificant and concentration-dependent decrease in TNF-αand IL-1β production.
3.4. Effect of NNMBS275 on Glutamate-Induced Cytotoxicityand Inhibition of ROS Generation in HT22 Cells. To evaluatethe in vitro neuroprotective effect of NNMBS275, we testedits protective effect on glutamate-induced cytotoxicity inHT22 cells. Treatment with glutamate for 12 h increasedHT22 cell death by 55% compared to untreated cells, whereasNNMBS275, at noncytotoxic concentrations (25, 50, 100,and 200 μg/mL), caused a dose-dependent increase in theviability of these cells (Figure 4(a)). Glutamate also doubledROS production, and NNMBS275 effectively suppressedthis induction (Figure 4(b)). Trolox, well known for itsantioxidative efficiency, was used as a positive control,and it showed a significant cytoprotective effect and ROS-scavenging activity at a concentration of 50 μM.
3.5. Effects of NNMBS275 on HO-1 Protein and mRNAExpression. We examined whether noncytotoxic concentra-tions (25–200 μg/mL) of NNMBS275 affected HO-1 proteinand mRNA expression, by treating BV2 microglia and HT22cells with this agent for 12 h. NNMBS275 caused a dose-dependent increase in HO-1 expression in BV2 microglia(Figure 5(a)) and HT22 cells (Figure 5(c)). The inductionof HO-1 by NNMBS275 reached a peak at 200 μg/mL. TheHO-1 inducer CoPP, used as a positive control, increasedHO-1 expression at a dose of 20 μM. At an NNMBS275concentration of 200 μg/mL, HO-1 expression showed anincrease at 6 h, peaked at around 18 h, and decreased after24 h in BV2 microglia and HT22 cells (Figures 5(b) and
5(d)). Consistently, NNMBS275-induced mRNA expressionlevels of HO-1 also increased in a dose-dependent manner inboth BV2 microglia and HT22 cells, as assessed by RT-PCRanalysis (Figure 6).
3.6. Effects of HO-1 Expression on Glutamate-Induced Oxida-tive Neurotoxicity and Inhibition of Proinflammatory Medi-ators by NNMBS275. Recent reports have described theexpression of HO-1 as an adaptive and protective responseagainst inflammation and oxidative insult in a wide varietyof cells, including glial and neuronal cells, such as BV2 andHT22. We examined whether NNMBS275-induced HO-1expression mediated its cytoprotective effects in HT22 cells.HT22 cells were cotreated with 200 μg/mL of NNMBS275 for12 h in the absence or presence of SnPP, an inhibitor of HO-1 activity. SnPP significantly inhibited NNMBS275-mediatedcytoprotection (Figure 7(a)). NNMBS275-induced HO-1expression was also required for suppressing glutamate-induced ROS generation (Figure 7(b)). On the basis ofour findings that preincubation of BV2 microglia withNNMBS275 markedly inhibited LPS-induced proinflamma-tory enzymes and proinflammatory cytokines (Figure 3),and that NNMBS275 was able to induce HO-1 expres-sion (Figure 5) in BV2 microglia, we examined whetherNNMBS275-mediated HO-1 induction might be responsiblefor the inhibition of iNOS-derived NO, COX-2-derivedPGE2, TNF-α, and IL-1β production (Figures 7(c), 7(d), 7(e),and 7(f)). BV2 microglia were pretreated with NNMBS275for 12 h in the presence of SnPP, followed by LPS stimulation.As shown in Figure 7, SnPP treatment partially reversed theinhibitory effects of NNMBS275 on NO, PGE2, TNF-α, andIL-1β production.
3.7. Effects of NNMBS275 on Nrf2 Nuclear Translocation.Several studies have reported that nuclear translocationof activated Nrf2 is an important upstream contributorto the mechanism of HO-1 expression. Therefore, weinvestigated whether treatment of BV2 microglia and HT22cells with NNMBS275 induced the translocation of Nrf2to the nucleus. Using western blot analysis, we tested thepresence of Nrf2 proteins in nuclear compartments ofBV2 microglia and HT22 cells. Cells were incubated with200 μg/mL NNMBS275 for 0.5, 1, and 1.5 h. As shown inFigure 8, the nuclear fractions of NNMBS275-treated BV2microglia and HT22 cells showed a gradual increase in Nrf2levels, whereas Nrf2 levels decreased concomitantly in thecytoplasmic fractions.
3.8. Effects of NNMBS275 on the Phosphorylation andDegradation of IκB-α and NF-κB Activation. NF-κB hasbeen implicated in the transcriptional regulation of LPS-induced iNOS expression. Therefore, we examined the effectsof NNMBS275 on the protein expression level of IκB-αphosphorylation and degradation of IκB-α, an inhibitorassociated with NF-κB in the cytoplasm. As shown inFigure 9(a), IκB-α was degraded after treatment with LPSin BV2 cells for 1 h, and this degradation was markedly
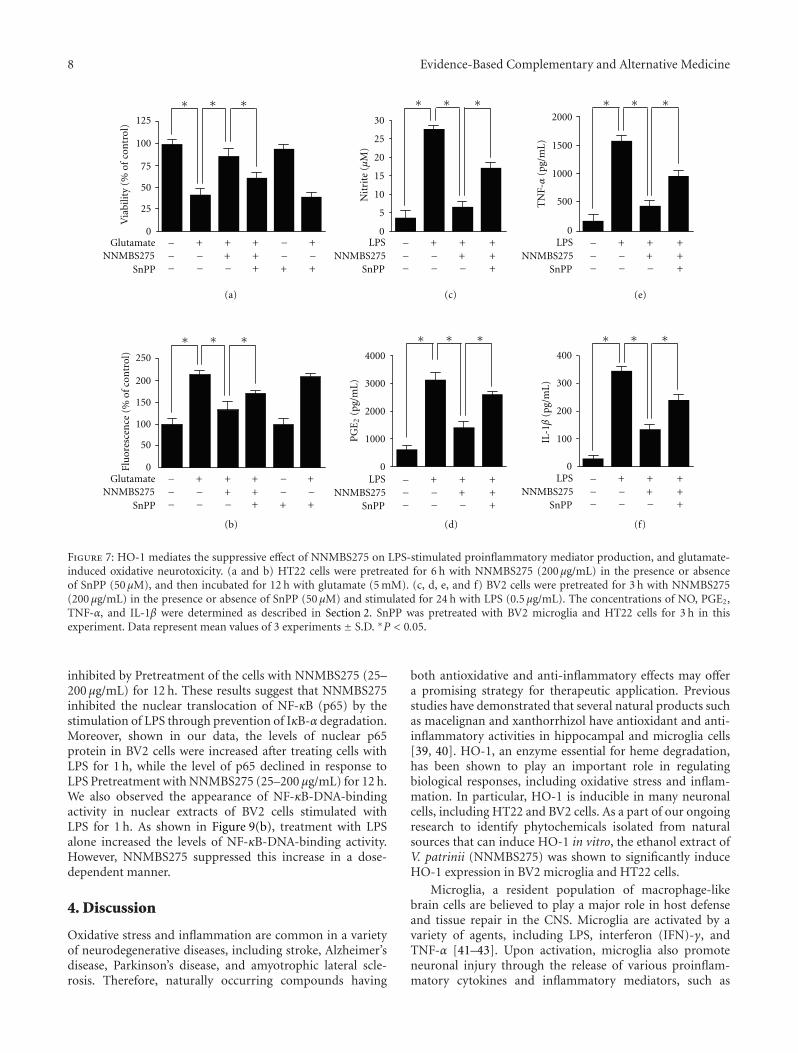
8 Evidence-Based Complementary and Alternative Medicine
1000
2000
3000
4000
0 0
400
300
200
100
500
1500
0
2000
1000
LPSNNMBS275
SnPP
0
30
20
15
10
5
25Fl
uor
esce
nce
(%
of c
ontr
ol)
0
250
200
150
100
50
0
125
100
75
50
25
Glutamate ++
+
+
+NNMBS275 −
−−
−−++
+
+++
+−−−
−−
+−−
−−
++−
LPSNNMBS275
SnPP++
+−−
−
+−−
++−
LPSNNMBS275
SnPP++
+−−
−
+−−
++−
LPSNNMBS275
SnPP++
+−−
−
+−−
++−
SnPP
Glutamate ++
+
+
+NNMBS275 −
−−
−−++
+
+− −−
−SnPP
Via
bilit
y (%
of
con
trol
)
∗∗∗
∗∗∗ ∗∗∗ ∗∗∗
∗∗∗ ∗∗∗
Nit
rite
(μ
M)
TN
F-α
(pg/
mL
)
PG
E2
(pg/
mL
)
IL-1β
(pg/
mL
)
(a) (c) (e)
(b) (d) (f)
Figure 7: HO-1 mediates the suppressive effect of NNMBS275 on LPS-stimulated proinflammatory mediator production, and glutamate-induced oxidative neurotoxicity. (a and b) HT22 cells were pretreated for 6 h with NNMBS275 (200 μg/mL) in the presence or absenceof SnPP (50 μM), and then incubated for 12 h with glutamate (5 mM). (c, d, e, and f) BV2 cells were pretreated for 3 h with NNMBS275(200 μg/mL) in the presence or absence of SnPP (50 μM) and stimulated for 24 h with LPS (0.5 μg/mL). The concentrations of NO, PGE2,TNF-α, and IL-1β were determined as described in Section 2. SnPP was pretreated with BV2 microglia and HT22 cells for 3 h in thisexperiment. Data represent mean values of 3 experiments ± S.D. ∗P < 0.05.
inhibited by Pretreatment of the cells with NNMBS275 (25–200 μg/mL) for 12 h. These results suggest that NNMBS275inhibited the nuclear translocation of NF-κB (p65) by thestimulation of LPS through prevention of IκB-α degradation.Moreover, shown in our data, the levels of nuclear p65protein in BV2 cells were increased after treating cells withLPS for 1 h, while the level of p65 declined in response toLPS Pretreatment with NNMBS275 (25–200 μg/mL) for 12 h.We also observed the appearance of NF-κB-DNA-bindingactivity in nuclear extracts of BV2 cells stimulated withLPS for 1 h. As shown in Figure 9(b), treatment with LPSalone increased the levels of NF-κB-DNA-binding activity.However, NNMBS275 suppressed this increase in a dose-dependent manner.
4. Discussion
Oxidative stress and inflammation are common in a varietyof neurodegenerative diseases, including stroke, Alzheimer’sdisease, Parkinson’s disease, and amyotrophic lateral scle-rosis. Therefore, naturally occurring compounds having
both antioxidative and anti-inflammatory effects may offera promising strategy for therapeutic application. Previousstudies have demonstrated that several natural products suchas macelignan and xanthorrhizol have antioxidant and anti-inflammatory activities in hippocampal and microglia cells[39, 40]. HO-1, an enzyme essential for heme degradation,has been shown to play an important role in regulatingbiological responses, including oxidative stress and inflam-mation. In particular, HO-1 is inducible in many neuronalcells, including HT22 and BV2 cells. As a part of our ongoingresearch to identify phytochemicals isolated from naturalsources that can induce HO-1 in vitro, the ethanol extract ofV. patrinii (NNMBS275) was shown to significantly induceHO-1 expression in BV2 microglia and HT22 cells.
Microglia, a resident population of macrophage-likebrain cells are believed to play a major role in host defenseand tissue repair in the CNS. Microglia are activated by avariety of agents, including LPS, interferon (IFN)-γ, andTNF-α [41–43]. Upon activation, microglia also promoteneuronal injury through the release of various proinflam-matory cytokines and inflammatory mediators, such as
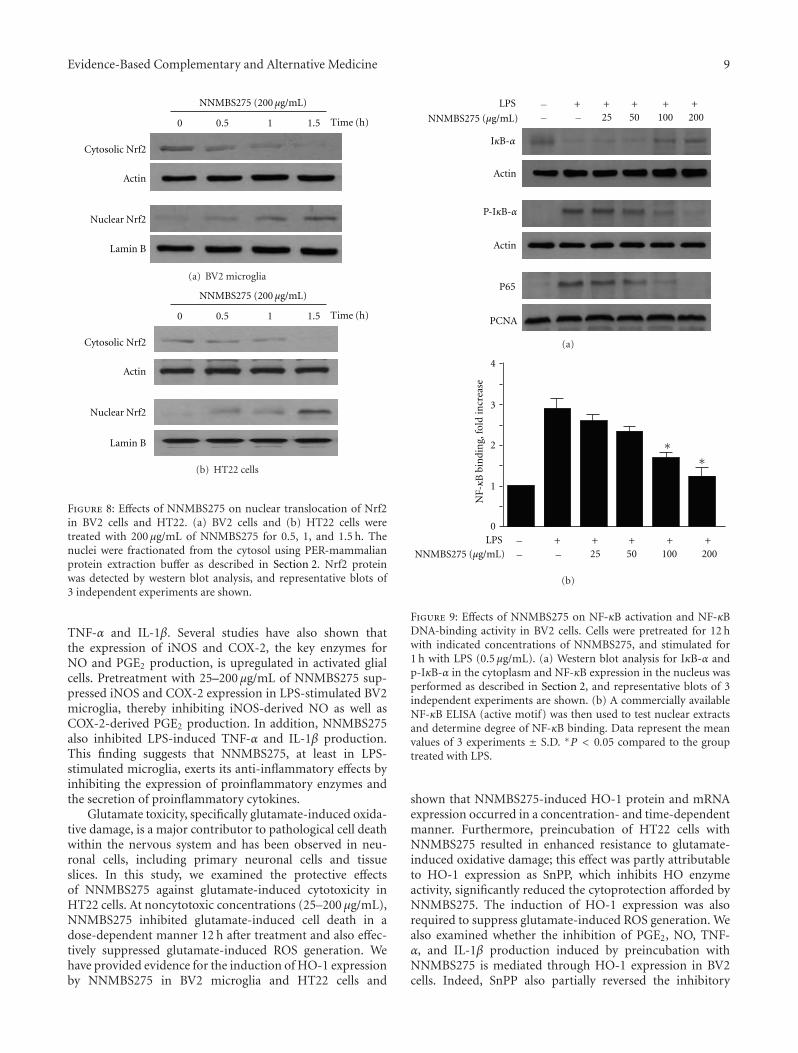
Evidence-Based Complementary and Alternative Medicine 9
0.5 1 1.5
Cytosolic Nrf2
Actin
Nuclear Nrf2
Lamin B
Time (h)0
NNMBS275 (200 μg/mL)
(a) BV2 microglia
0.5 1 1.5
Cytosolic Nrf2
Actin
Nuclear Nrf2
Lamin B
Time (h)0
NNMBS275 (200 μg/mL)
(b) HT22 cells
Figure 8: Effects of NNMBS275 on nuclear translocation of Nrf2in BV2 cells and HT22. (a) BV2 cells and (b) HT22 cells weretreated with 200 μg/mL of NNMBS275 for 0.5, 1, and 1.5 h. Thenuclei were fractionated from the cytosol using PER-mammalianprotein extraction buffer as described in Section 2. Nrf2 proteinwas detected by western blot analysis, and representative blots of3 independent experiments are shown.
TNF-α and IL-1β. Several studies have also shown thatthe expression of iNOS and COX-2, the key enzymes forNO and PGE2 production, is upregulated in activated glialcells. Pretreatment with 25–200 μg/mL of NNMBS275 sup-pressed iNOS and COX-2 expression in LPS-stimulated BV2microglia, thereby inhibiting iNOS-derived NO as well asCOX-2-derived PGE2 production. In addition, NNMBS275also inhibited LPS-induced TNF-α and IL-1β production.This finding suggests that NNMBS275, at least in LPS-stimulated microglia, exerts its anti-inflammatory effects byinhibiting the expression of proinflammatory enzymes andthe secretion of proinflammatory cytokines.
Glutamate toxicity, specifically glutamate-induced oxida-tive damage, is a major contributor to pathological cell deathwithin the nervous system and has been observed in neu-ronal cells, including primary neuronal cells and tissueslices. In this study, we examined the protective effectsof NNMBS275 against glutamate-induced cytotoxicity inHT22 cells. At noncytotoxic concentrations (25–200 μg/mL),NNMBS275 inhibited glutamate-induced cell death in adose-dependent manner 12 h after treatment and also effec-tively suppressed glutamate-induced ROS generation. Wehave provided evidence for the induction of HO-1 expressionby NNMBS275 in BV2 microglia and HT22 cells and
Actin
Actin
P65
PCNA
+ + + + +25 50 100 200
LPS −− −
IκB-α
P-IκB-α
NNMBS275 (μg/mL)
(a)
bin
din
g, fo
ld in
crea
se
0
3
4
2
1N
F-κB
∗∗
+ + + + +25 50 100 200
LPS −− −NNMBS275 (μg/mL)
(b)
Figure 9: Effects of NNMBS275 on NF-κB activation and NF-κBDNA-binding activity in BV2 cells. Cells were pretreated for 12 hwith indicated concentrations of NNMBS275, and stimulated for1 h with LPS (0.5 μg/mL). (a) Western blot analysis for IκB-α andp-IκB-α in the cytoplasm and NF-κB expression in the nucleus wasperformed as described in Section 2, and representative blots of 3independent experiments are shown. (b) A commercially availableNF-κB ELISA (active motif) was then used to test nuclear extractsand determine degree of NF-κB binding. Data represent the meanvalues of 3 experiments ± S.D. ∗P < 0.05 compared to the grouptreated with LPS.
shown that NNMBS275-induced HO-1 protein and mRNAexpression occurred in a concentration- and time-dependentmanner. Furthermore, preincubation of HT22 cells withNNMBS275 resulted in enhanced resistance to glutamate-induced oxidative damage; this effect was partly attributableto HO-1 expression as SnPP, which inhibits HO enzymeactivity, significantly reduced the cytoprotection afforded byNNMBS275. The induction of HO-1 expression was alsorequired to suppress glutamate-induced ROS generation. Wealso examined whether the inhibition of PGE2, NO, TNF-α, and IL-1β production induced by preincubation withNNMBS275 is mediated through HO-1 expression in BV2cells. Indeed, SnPP also partially reversed the inhibitory

10 Evidence-Based Complementary and Alternative Medicine
LPS
Neuronal cell damage
Glutamate
ROS production
Oxidative c cytotoxicity
NNMBS275
Nrf2 activation
Heme oxygenase-1
Neuroinflammation Oxidative stress
NF-κB activation
NO, PGF2, IL-1βand
TNF-α production
Figure 10: The pathway scheme of the anti-inflammatory and cytoprotective effects of NNMBS275 in BV2 microglia and HT22cells. NNMBS275 increased cellular resistance to oxidative injury caused by glutamate-induced oxidative cytotoxicity in HT22 cells,via Nrf2-dependent HO-1 expression. In BV2 cells, NNMBS275 suppressed the LPS-induced expression of proinflammatory enzymesand inflammatory mediators through its interference of NF-κB activation and Nrf2-dependent HO-1 expression. This study indicatesthat NNMBS275 effectively modulates the regulation of antioxidative and anti-inflammatory action, via Nrf2 pathway dependent HO-1expression in BV2 microglia and HT22 cells.
effects of NNMBS275 on NO, PGE2, TNF-α, and IL-1βproduction suggesting that HO-1 expression is involved inglutamate- or LPS-induced cell responses.
HO-1 induction by low-molecular weight compoundsmight play an important role in the neuroprotection of CNSneurons [44, 45]. Recent reports have suggested that phy-tochemicals can activate Nrf2 by directly binding to Keap1through covalent linkages, resulting in the induction ofsome cytoprotective proteins, including HO-1. In the presentstudy, we showed that NNMBS275 significantly increasedNrf2 levels and efficiently promoted its translocation intothe nucleus in both BV2 microglia and HT22 cells. Thissuggests that Nrf2 may play an essential role in NNMBS275-induced HO-1 expression. Since NNMBS275, as shown inthis study, can actively induce the expression of HO-1 inmouse hippocampal HT22 cells and BV2 microglia, it ispossible that the antioxidative and anti-inflammatory effectsof NNMBS275 are partly mediated by the products ofthe HO-1 enzyme reaction, namely, CO, bilirubin and/orbiliverdin.
It is also well known that NF-κB is a significant rolefactor regulating the expression of inflammation-associatedenzymes and cytokine genes, such as iNOS, COX-2, TNF-α,and IL-1β, which contain NF-κB-binding motifs within theirrespective promoters [46]. Many anti-inflammatory agentsexhibit their potency by suppressing NF-κB signaling [47,48]. In this study, we examined the effects of NNMBS275 onthe phosphorylation of IκBα and the nuclear translocation ofa NF-κB subunit, p65, in BV2 cells. As shown in Figure 9, thephosphorylation and degradation of IκBα and the nucleartranslocation of p65 induced by LPS were significantlyreduced after pretreatment with NNMBS275. Furthermore,we also observed the appearance of NF-κB-DNA-bindingactivity of NF-κB in nuclear extracts of BV2 cells stimulatedwith LPS. These results suggest that NNMBS275, at leastin LPS-stimulated BV2 cells, exerted its anti-inflammatoryeffects by suppressing the expression of the proinflammatoryenzymes as well as the production of the proinflammatorycytokines.
In conclusion, NNMBS275, the ethanol extract of V.patrinii, showed potent anti-inflammatory effects on mi-croglial activation by LPS, possibly through the Nrf2 path-way-dependent expression of HO-1 and suppression of NF-κB signaling. Furthermore, we investigated the cytoprotec-tive effects of NNMBS275 on glutamate-induced neuro-toxicity in mouse hippocampal HT22 cells via the HO-1expression (Figure 10). The results of our study demonstratethe importance of HO-1 in mediating the antioxidativeand anti-inflammatory effects of NNMBS275 in mousemicroglial and hippocampal cells and reveal the possibletherapeutic value of NNMBS275 in preventing the progres-sion of neurodegenerative disease. Further detailed chemicalstudies of NNMBS275 would help to clarify its therapeuticpotential.
Authors’ Contribution
B. Li and D.-S. Lee contributed equally to this work.
Acknowledgments
The research was supported by Basic Science ResearchProgram through the National Research Foundation of Korea(NRF) funded by the Ministry of Education, Science andTechnology (MEST) (no. 2010-0024029).
References
[1] A. Hald and J. Lotharius, “Oxidative stress and inflammationin Parkinson’s disease: is there a causal link?” ExperimentalNeurology, vol. 193, no. 2, pp. 279–290, 2005.
[2] G. E. Gibson and H. Zhang, “Abnormalities in oxidative pro-cesses in non-neuronal tissues from patients with Alzheimer’sdisease,” Journal of Alzheimer’s Disease, vol. 3, no. 3, pp. 329–338, 2001.
[3] J. T. Coyle and P. Puttfarcken, “Oxidative stress, glutamate, andneurodegenerative disorders,” Science, vol. 262, no. 5134, pp.689–695, 1993.

Evidence-Based Complementary and Alternative Medicine 11
[4] T. Satoh, Y. Enokido, T. Kubo, M. Yamada, and H. Hatanaka,“Oxygen toxicity induces apoptosis in neuronal cells,” Cellularand Molecular Neurobiology, vol. 18, no. 6, pp. 649–666, 1998.
[5] T. Satoh and S. A. Lipton, “Redox regulation of neuronalsurvival mediated by electrophilic compounds,” Trends inNeurosciences, vol. 30, no. 1, pp. 37–45, 2007.
[6] O. G. Rossler, I. Bauer, H. Y. Chung, and G. Thiel, “Glutamate-induced cell death of immortalized murine hippocampalneurons: neuroprotective activity of heme oxygenase-1, heatshock protein 70, and sodium selenite,” Neuroscience Letters,vol. 362, no. 3, pp. 253–257, 2004.
[7] M. P. Mattson, “Apoptosis in neurodegenerative disorders,”Nature Reviews Molecular Cell Biology, vol. 1, no. 2, pp. 120–129, 2000.
[8] P. Maher and J. B. Davis, “The role of monoamine metabolismin oxidative glutamate toxicity,” Journal of Neuroscience, vol.16, no. 20, pp. 6394–6401, 1996.
[9] G. S. Jeong, R. B. An, H. O. Pae et al., “Cudratricusxanthone aprotects mouse hippocampal cells against glutamate-inducedneurotoxicity via the induction of heme oxygenase-1,” PlantaMedica, vol. 74, no. 11, pp. 1368–1373, 2008.
[10] S. W. Ryter, L. E. Otterbein, D. Morse, and A. M. K. Choi,“Heme oxygenase/carbon monoxide signaling pathways: reg-ulation and functional significance,” Molecular and CellularBiochemistry, vol. 234-235, pp. 249–263, 2002.
[11] J. Kapitulnik, “Bilirubin: an endogenous product of heme deg-radation with both cytotoxic and cytoprotective properties,”Molecular Pharmacology, vol. 66, no. 4, pp. 773–779, 2004.
[12] K. A. Kirkby and C. A. Adin, “Products of heme oxygenase andtheir potential therapeutic applications,” American Journal ofPhysiology, vol. 290, no. 3, pp. F563–F571, 2006.
[13] K. Itoh, T. Chiba, S. Takahashi et al., “An Nrf2/small Maf het-erodimer mediates the induction of phase II detoxifying en-zyme genes through antioxidant response elements,” Biochem-ical and Biophysical Research Communications, vol. 236, no. 2,pp. 313–322, 1997.
[14] W. Qiang, J. M. Cahill, J. Liu et al., “Activation of transcriptionfactor Nrf-2 and its downstream targets in response toMoloney murine leukemia virus ts1-induced thiol depletionand oxidative stress in astrocytes,” Journal of Virology, vol. 78,no. 21, pp. 11926–11938, 2004.
[15] K. M. Kim, H. O. Pae, M. Zheng, R. Park, Y. M. Kim, and H.T. Chung, “Carbon monoxide induces heme oxygenase-1 viaactivation of protein kinase R-like endoplasmic reticulumkinase and inhibits endothelial cell apoptosis triggered byendoplasmic reticulum stress,” Circulation Research, vol. 101,no. 9, pp. 919–927, 2007.
[16] S. Guzman-Beltran, S. Espada, M. Orozco-Ibarra, J. Pedraza-Chaverri, and A. Cuadrado, “Nordihydroguaiaretic acid acti-vates the antioxidant pathway Nrf2/HO-1 and protects cere-bellar granule neurons against oxidative stress,” NeuroscienceLetters, vol. 447, no. 2-3, pp. 167–171, 2008.
[17] C. Y. Chen, J. H. Jang, M. H. Li, and Y. J. Surh, “Resveratrolupregulates heme oxygenase-1 expression via activation ofNF-E2-related factor 2 in PC12 cells,” Biochemical and Bio-physical Research Communications, vol. 331, no. 4, pp. 993–1000, 2005.
[18] C. Mancuso, G. Scapagnini, D. Curro et al., “Mitochondri-al dysfunction, free radical generation and cellular stress re-sponse in neurodegenerative disorders,” Frontiers in Bio-science, vol. 12, no. 3, pp. 1107–1123, 2007.
[19] L. Romeo, M. Intrieri, V. D’Agata et al., “The major greentea polyphenol, (-)-epigallocatechin-3-gallate, induces heme
oxygenase in rat neurons and acts as an effective neuropro-tective agent against oxidative stress,” Journal of the AmericanCollege of Nutrition, vol. 28, no. 4, 2009.
[20] F. Gonzalez-Scarano and G. Baltuch, “Microglia as mediatorsof inflammatory and degenerative diseases,” Annual Review ofNeuroscience, vol. 22, pp. 219–240, 1999.
[21] M. Vila, V. Jackson-Lewis, C. Guegan et al., “The role of glialcells in Parkinson’s disease,” Current Opinion in Neurology, vol.14, no. 4, pp. 483–489, 2001.
[22] G. W. Kreutzberg, “Microglia: a sensor for pathological eventsin the CNS,” Trends in Neurosciences, vol. 19, no. 8, pp. 312–318, 1996.
[23] C. C. Chao, S. Hu, T. W. Molitor, E. G. Shaskan, and P. K.Peterson, “Activated microglia mediate neuronal cell injury viaa nitric oxide mechanism,” Journal of Immunology, vol. 149,no. 8, pp. 2736–2741, 1992.
[24] L. Minghetti and G. Levi, “Induction of prostanoid biosyn-thesis by bacterial lipopolysaccharide and isoproterenol in ratmicroglial cultures,” Journal of Neurochemistry, vol. 65, no. 6,pp. 2690–2698, 1995.
[25] L. Meda, M. A. Cassatella, G. I. Szendrei et al., “Activation ofmicroglial cells by β-amyloid protein and interferon-γ,”Nature, vol. 374, no. 6523, pp. 647–650, 1995.
[26] F. Gueler, J. K. Park, S. Rong et al., “Statins attenuate ischemia-reperfusion injury by inducing heme oxygenase-1 in infiltrat-ing macrophages,” American Journal of Pathology, vol. 170, no.4, pp. 1192–1199, 2007.
[27] K. J. Min, J. H. Kim, I. Jou, and E. H. Joe, “Adenosine induceshemeoxygenase-1 expression in microglia through the activa-tion of phosphatidylinositol 3-kinase and nuclear factor E2-related factor 2,” GLIA, vol. 56, no. 9, pp. 1028–1037, 2008.
[28] A. Yachie, T. Toma, K. Mizuno et al., “Heme oxygenase-1production by peripheral blood monocytes during acuteinflammatory illnesses of children,” Experimental Biology andMedicine, vol. 228, no. 5, pp. 550–556, 2003.
[29] A. Zampetaki, T. Minamino, S. A. Mitsialis, and S. Kourem-banas, “Effect of heme oxygenase-1 overexpression in twomodels of lung inflammation,” Experimental Biology andMedicine, vol. 228, no. 5, pp. 442–446, 2003.
[30] L. E. Otterbein, F. H. Bach, J. Alam et al., “Carbon monoxidehas anti-inflammatory effects involving the mitogen- activatedprotein kinase pathway,” Nature Medicine, vol. 6, no. 4, pp.422–428, 2000.
[31] P. Wiesel, L. C. Foster, A. Pellacani et al., “Thioredoxin facil-itates the induction of heme oxygenase-1 in response to in-flammatory mediators,” Journal of Biological Chemistry, vol.275, no. 32, pp. 24840–24846, 2000.
[32] G. Y. Suh, Y. Jin, A. K. Yi, X. M. Wang, and A. M. K. Choi,“CCAAT/enhancer-binding protein mediates carbon monox-ide-induced suppression of cyclooxygenase-2,” American Jour-nal of Respiratory Cell and Molecular Biology, vol. 35, no. 2, pp.220–226, 2006.
[33] G. S. Oh, H. O. Pae, B. S. Lee et al., “Hydrogen sulfide inhibitsnitric oxide production and nuclear factor-κB via hemeoxygenase-1 expression in RAW264.7 macrophages stimulatedwith lipopolysaccharide,” Free Radical Biology and Medicine,vol. 41, no. 1, pp. 106–119, 2006.
[34] K. S. Kim, Y. J. Kwak, K. M. Kim et al., “Purification and struc-ture determination of gelatinase and collagenase inhibitorsfrom Viola patrinii fermentation extracts,” Immunopharma-cology and Immunotoxicology, vol. 32, no. 4, pp. 614–616, 2010.
[35] E. Svangard, U. Goransson, D. Smith et al., “Primary and 3-D modelled structures of two cyclotides from Viola odorata,”Phytochemistry, vol. 64, no. 1, pp. 135–142, 2003.

12 Evidence-Based Complementary and Alternative Medicine
[36] A. P. Carnat, A. Carnat, D. Fraisse, and J. L. Lamaison, “Vio-larvensin, a new flavone di-C-glycoside from Viola arvensis,”Journal of Natural Products, vol. 61, no. 2, pp. 272–274, 1998.
[37] B. Chen, M. L. Colgrave, N. L. Daly, K. J. Rosengren, K. R.Gustafson, and D. J. Craik, “Isolation and characterization ofnovel cyclotides from Viola hederaceae: solution structure andanti-HIV activity of vhl-1, a leaf-specific expressed cyclotide,”Journal of Biological Chemistry, vol. 280, no. 23, pp. 22395–22405, 2005.
[38] D. S. Lee, G. S. Jeong, B. Li, H. Park, and Y. C. Kim, “Anti-inflammatory effects of sulfuretin from Rhus vernicifluaStokes via the induction of heme oxygenase-1 expression inmurine macrophages,” International Immunopharmacology,vol. 10, no. 8, pp. 850–858, 2010.
[39] C. S. Lim, D. Q. Jin, H. Mok et al., “Antioxidant and anti-inflammatory activities of xanthorrhizol in hippocampal neu-rons and primary cultured microglia,” Journal of NeuroscienceResearch, vol. 82, no. 6, pp. 831–838, 2005.
[40] D. Q. Jin, C. S. Lim, J. K. Hwang, I. Ha, and J. S. Han, “Anti-oxidant and anti-inflammatory activities of macelignan inmurine hippocampal cell line and primary culture of ratmicroglial cells,” Biochemical and Biophysical Research Com-munications, vol. 331, no. 4, pp. 1264–1269, 2005.
[41] Y. Nakamura, Q. S. Si, and K. Kataoka, “Lipopolysaccharide-induced microglial activation in culture: temporal profilesof morphological change and release of cytokines and nitricoxide,” Neuroscience Research, vol. 35, no. 2, pp. 95–100, 1999.
[42] L. I. Romero, J. B. Tatro, J. A. Field, and S. Reichlin, “Rolesof IL-1 and TNF-α in endotoxin-induced activation of nitricoxide synthase in cultured rat brain cells,” American Journal ofPhysiology, vol. 270, no. 2, pp. R326–R332, 1996.
[43] B. Popko, J. G. Corbin, K. D. Baerwald, J. Dupree, and A. M.Garcia, “The effects of interferon-γ on the central nervoussystem,” Molecular Neurobiology, vol. 14, no. 1-2, pp. 19–35,1997.
[44] T. Satoh, M. Baba, D. Nakatsuka et al., “Role of heme oxygen-ase-1 protein in the neuroprotective effects of cyclopentenoneprostaglandin derivatives under oxidative stress,” EuropeanJournal of Neuroscience, vol. 17, no. 11, pp. 2249–2255, 2003.
[45] S. Dore, M. Takahashi, C. D. Ferris, L. D. Hester, D. Guastella,and S. H. Snyder, “Bilirubin, formed by activation of hemeoxygenase-2, protects neurons against oxidative stress injury,”Proceedings of the National Academy of Sciences of the UnitedStates of America, vol. 96, no. 5, pp. 2445–2450, 1999.
[46] D. Thanos and T. Maniatis, “Identification of the rel familymembers required for virus induction of the human betainterferon gene,” Molecular and Cellular Biology, vol. 15, no.1, pp. 152–164, 1995.
[47] M. S. Choi, S. H. Lee, H. S. Cho et al., “Inhibitory effectof obovatol on nitric oxide production and activation ofNF-κB/MAP kinases in lipopolysaccharide-treated RAW264.7cells,” European Journal of Pharmacology, vol. 556, no. 1–3, pp. 181–189, 2007.
[48] H. K. Byung, M. C. Sung, A. M. Reddy, S. K. Yeong, R. M.Kyung, and Y. Kim, “Down-regulatory effect of quercitringallate on nuclear factor-κB-dependent inducible nitric ox-idesynthase expression in lipopolysaccharide-stimulated macro-phages RAW 264.7,” Biochemical Pharmacology, vol. 69, no. 11,pp. 1577–1583, 2005.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 932823, 11 pagesdoi:10.1155/2012/932823
Research Article
Paeonol Protects Memory after Ischemic Stroke via Inhibitingβ-Secretase and Apoptosis
Shan-Yu Su,1, 2 Chin-Yi Cheng,3 Tung-Hu Tsai,4 and Ching-Liang Hsieh1, 3
1 Department of Chinese Medicine, China Medical University Hospital, Taichung 40447, Taiwan2 School of Post-Baccalaureate Chinese Medicine, College of Chinese Medicine, China Medical University, Taichung 40402, Taiwan3 Graduate Institute of Acupuncture Science, College of Chinese Medicine, China Medical University, 91 Hsueh-Shih Road,Taichung 40402, Taiwan
4 Institute of Traditional Medicine, National Yang-Ming University, Taipei 11221, Taiwan
Correspondence should be addressed to Ching-Liang Hsieh, [email protected]
Received 30 September 2011; Revised 23 November 2011; Accepted 24 November 2011
Academic Editor: Monica Rosa Loizzo
Copyright © 2012 Shan-Yu Su et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Poststroke dementia commonly occurs following stroke, with its pathogenesis related to β-amyloid production and apoptosis.The present study evaluate the effects of paeonol, one of the phenolic phytochemicals isolated from the Chinese herb Paeoniasuffruticosa Andrews (MC), on protection from memory loss after ischemic stroke in the subacute stage. Rats were subjected totransient middle cerebral artery occlusion (tMCAo) with 10 min of ischemia. The data revealed that paeonol recovered the step-through latency in the retrieval test seven days after tMCAo, but did not improve the neurological deficit induced by tMCAo.Levels of Amyloid precursor protein (APP)- and beta-site APP cleaving enzyme (BACE; β-secretase)-immunoreactive cells, andterminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL)-positive cells decreased in the paeonol-administered group. Western blotting revealed decreased levels of Bax protein in mitochondria and apoptosis-inducing factor(AIF) in cytosol following paeonol treatment. In conclusion, we speculate that paeonol protected memory after ischemic strokevia reducing APP, BACE, and apoptosis. Supression the level of Bax and blocking the release of AIF into cytosol might participatein the anti-apoptosis provided by paeonol.
1. Introduction
Cognitive impairment is a commonly occurring sequelafollowing stroke and is the second leading cause of dementiain the elderly, after Alzheimer’s disease [1, 2]. The majority ofstroke survivors suffer from various cognitive dysfunctions,including dementia (which exists in 25% of stroke patients),linked with disability, poor functional outcome, and lifedissatisfaction [3]. With recent improvements in the medicaltreatment of stroke, survival rates have increased, and theprevention and treatment of vascular cognitive impairmentand vascular dementia after stroke have become increasinglyimportant [3].
Although there are differences in the causes of thediseases, poststroke dementia shares common mechanismswith Alzheimer’s disease, including the augmentation of β-amyloid production and tau protein phosphorylation [4,
5]. Following ischemic episode, the expression of amyloidprecursor protein (APP) upregulates β-amyloid oligomers inthe brain’s extracellular spaces [6–8], and amyloid precursorprotein production increases in astrocytes [9]. Then, theinteractions between β-amyloid and several factors, includ-ing apolipoproteins, presenilins, tau protein, α-synuclein,inflammation factors, and neuronal survival/death decisionsin the brain, contribute to ischemic brain degeneration,leading to white matter damage and neuronal cell death[5, 7]. Excepting necrotic cell death which happens withinminutes, neuronal death (including apoptosis) commencesseveral hours after ischemic stroke and lasts several days [10].Therapies which can salvage neuronal cells from apoptosisin ischemic penumbra might improve patient outcomes afterischemic stroke [11].
Paeonol (2′-hydroxy-4′-methoxyacetophenone) is one ofthe phenolic phytochemicals isolated from the Chinese herb

2 Evidence-Based Complementary and Alternative Medicine
Paeonia suffruticosa Andrews (MC), and widely consumed asa nutrient supplement in a Chinese medical formula whichhad the third highest sales volumes in Beijing during 2007and 2009 [12]. In an animal study, paeonol attenuated neu-rotoxicity and ameliorated cognitive impairment inducedby d-galactose [13]. The present group has previouslydemonstrated that paeonol reduced the infarct area in atransient middle cerebral artery occlusion (tMCAo) ratmodel [14]. It also attenuated oxidative stress-induced APPexpression in a cell model [15].
To investigate the validity of paeonol in protectionof memory after stroke, rat model of tMCAo was used.Neurological status and memory was tested seven daysafter tMCAo. Levels of APP, beta-site APP cleaving enzyme(BACE), and apoptosis in brain tissue were evaluated. Then,the levels of proteins that participate in apoptotic intrinsicand extrinsic pathways were also measured. The data revealedthat paeonol protected memory after ischemic stroke viareducing APP, BACE, and apoptosis. Suppression of level ofmitochondrial Bax and cytosolic AIF might also participatein the protective effect.
2. Materials and Methods
2.1. Animals and Chemicals. All experimental procedureswere performed on adult male Sprague-Dawley rats, weigh-ing 300 to 350 g, according to the guidelines approved by theCare and Use of Laboratory Animals Committee of ChinaMedical University. Adequate measures were taken to min-imize animals’ pain or discomfort. Rats were housed underlight-dark—(12 h/12 h) and room temperature—controlledconditions.
Paeonol was isolated and purified from the root barkof Paeonia suffruticosa as described previously [14]. Freshlyprepared paeonol was first dissolved in tetraglycol and thendiluted in phosphate-buffered saline (PBS; 137 mM NaCl,1.4 mM KH2PO4, 4.3 mM Na2HPO4, 2.7 mM KCl, pH 7.2)to reach a final concentration of 2 mg/mL in a 5% solutionof tetraglycol in PBS. Chloral hydrate (Merck, Darmstadt,Germany) was dissolved in water to a stock concentration of400 mg/mL.
2.2. Occlusion Model. Ischemia was induced via intraluminalsuture occlusion of the middle cerebral artery (MCA) asdescribed previously [16]. Briefly, rats were anesthetized withchloral hydrate (400 mg/kg, i.p.). The right common carotidartery (CCA) and internal carotid artery (ICA) were exposedvia a midline incision in the neck. The pterygopalatine arterywas ligated close to its origin. A 3/0 nylon filament suture,blunted at the tip by a flame and coated with poly-L-lysine(Sigma, USA), was advanced from the right external carotidartery through the CCA and up to the ICA for a distance of20 to 25 mm to block the origin of the MCA. After 10 min ofischemia, the nylon suture was removed to allow reperfusion.
2.3. Grouping and Experiment. A total of 36 rats wererandomly divided into three groups: paeonol group, vehiclegroup, and sham group. Rats in the paeonol group werepre-administered paeonol one hour before introduction of
tMCAo. A dose of 20 mg/kg (i.p.) was chosen according toour previous study, which revealed that 20 mg/kg of paeonolexhibits the best neuroprotective effect [14]. After 10 minof ischemia, rats were subjected to reperfusion. Rats in thevehicle group were subjected to the same procedure as ratsin the paeonol group, but PBS was administered insteadof paeonol. Rats in the sham group underwent the sameprocedure as rats in the vehicle group, but the origin of theMCA was not occluded.
One day before tMCAo, rats underwent habituation andtraining for the passive avoidance trial. One hour beforetMCAo, rats underwent the retention trial. Twenty-fourhours after tMCAo, the neurological status were measured.The second retention trial and evaluation of neurologicalstatus were performed on the seventh day after tMCAo.Rats were then sacrificed for Western blotting and termi-nal deoxynucleotidyl transferase-mediated dUTP-biotin nickend labeling (TUNEL) and immunohistochemical (IHC)staining.
2.4. Measurement of Neurological Status. The neurologicalstatus of each rat was measured using Modified NeurologicalSeverity Score, 24 hours after reperfusion, by an investigatorblind to the treatment group. Motor, sensory, balance, andreflex functions were assessed based on a neurological deficitscore (18-point scale) described by Chen et al. [17]. Briefly,motor tests included placing the rat on the floor (inabilityto walk straight was scored as 1, circling toward paretic sidewas scored as 2, and falling down to the paretic side wasscored as 3) and raising each rat by its tail (flexion of forelimbwas scored as 1, flexion of hindlimb was scored as 1, andhead moving >10◦ was scored as 1). Sensory tests includedtactile (deficiency, 1) and pushing paw against table edge(deficiency, 1) subtests. Ability to balance on the beam wasscored as follows: rats grasped onto side of a beam, 1; huggedthe beam and one limb slipped off the beam, 2; hugged thebeam and two limbs slipped off the beam, 3; attempted tobalance but fell off (>40 s), 4; attempted to balance but fell off(<20 s), 5; fell off the beam without any attempt to balance,6. Reflex tests included pinna reflex (deficiency, 1), cornealreflex (deficiency, 1), and startle reflex subtests (deficiency,1). Abnormal movement was scored (seizure, 1).
2.5. Passive Avoidance Test. The passive avoidance apparatusconsisted of two chambers of the same size (25 × 20 ×17 cm high) connected via a guillotine door (9 × 7 cm).The floor of each chamber was made of 14 stainless steelrods (6 mm in diameter), spaced 1.8 cm center to center, andwired to a shock scrambler (Gemini Avoidance System, SanDiego Instruments, San Diego, CA, USA). For habituationthe rats were placed in the right chamber of the apparatusand, 5 sec later, the house light was turned on and theguillotine door was raised. Upon entering the dark chamber,the guillotine door was closed and 30 sec later the ratswere taken out from the dark chamber and put into theirhome cages. Entrance latency into the dark compartmentwas recorded when the animal had placed all four pawsinto the dark chamber. If the animal waited for more than100 sec to enter the dark chamber, it was eliminated from

Evidence-Based Complementary and Alternative Medicine 3
the experiment. The habituation was repeated 30 min laterand followed after the same interval by a training session.During the training session the guillotine door was closedand an intermittent electric shock (50 Hz, 3 s, 0.5 mA) wasdelivered to the floor of the dark chamber immediately afterthe animals had enter the dark chamber. After 30 sec, the ratwas taken from the dark chamber and placed into its homecage. Then after 2 min, the training session was repeated.The rat received a footshock each time it reentered the dark.Training was terminated when the rat remained in the lightcompartment for 120 sec. The numbers of trials (enteringthe dark chamber) were recorded. The retention trial wasperformed 24 hours and seven days after tMCAo. Afterthe house light was on and the guillotine door was open,the step-through latency (STL) into the dark chamber wasrecorded for up to 300 sec. If the rat did not enter the darkchamber after 300 sec, the retention trial was terminated anda ceiling score of 300 sec was assigned.
2.6. IHC Assay. For ICH and TUNEL staining, rats weretranscardially perfused with 200 mL of 0.9% saline and200 mL of 4% paraformaldehyde (PFA, pH 7.4). Rat brainswere removed quickly and postfixed in 4% PFA followedby 30% sucrose (wt/vol) for three days and then cutinto 15 μm sections using a cryostat. Brain sections wererinsed with Dulbecco’s phosphate-buffered saline (DPBS,Sigma, USA) containing 0.01% Tween-20 and immersedin 3% H2O2/methanol for 15 min to inhibit endogenousperoxidase activity. Thereafter, sections were incubated with10% normal animal serum (Zymed, CA, USA) for 20 minat room temperature. The sections were incubated in moistchambers with primary anti-APP (1 : 100 dilution, mousemonoclonal, 22C11, Chemicon, Billerica, MA, USA) andanti-BACE (1 : 100 dilution, Chemicon) for one hour at37◦C. Following incubation with secondary antibody andavidine-biotin peroxidase complex (ABC kit, Zymed, CA,USA) sections were colored using a 3,3′-diaminobenzidine(DAB) kit (Scytek Laboratories, Logan, U.T, USA), andthen counterstained with hematoxylin. The stained sectionswere mounted in mounting media (Assistant-Histokitt,Germany) and immunoreactive cells were counted under themicroscope (Axioskop 40, Zeiss). Immunoreactive cells werecounted for nine consecutive high power fields (HPFs) alongthe CA1 region and nine HPF within a square in the MCAterritory of the cortex (Figure 3(a)). Negative control stainswere performed on adjacent sections in the control groupand subjected to the same IHC assay procedures, but withoutprimary antibodies.
2.7. TUNEL Assay. TUNEL staining was performed accord-ing to the manufacturer’s instructions of a commercialkit (Merck KGaA, Darmstadt, Germany) to identify cellswith nuclear DNA fragmentation. Briefly, the brain sec-tions, which were chosen adjacent to those used for IHCwere incubated with proteinase K (20 μg/mL) for 20 minand then incubated with 1X TdT equilibration buffer for30 min at room temperature, followed by incubation withTdT labeling reaction mixture for 1.5 h at 37◦C. Afteraddition of stop solution and blocking buffer, sections
were incubated with conjugate solution for 30 min at roomtemperature and TUNEL positive cells were visualized usingDAB kit. Finally, sections were counterstained with methylgreen.
2.8. Western Blotting Analysis. For Western blotting, ratswere anesthetized using choral hydrate and perfused tran-scardially with 400 mL of 0.9% saline. Brains were removed,coronally sectioned from −4.3 to +1.7 mm bregma, andseparated into the right cortex, right striatum, left cortex, andleft striatum. The right cortex was weighed and the cytosolicand mitochondrial proteins were separated using a commer-cial kit (BioVision, Mountain View, CA, USA). Protein con-centrations of the cytosolic and mitochondrial fractions weredetermined using Bio-Rad assay. The protein extracts (10 μg)were then separated using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, transferred to nitrocel-lulose membranes, probed with antibodies, and detectedusing peroxidase-conjugated anti-rabbit antibody followedby chemiluminescence as described previously [18]. Theantibody against actin (dilution 1 : 5000) was purchased fromChemicon. Antibodies against cytochrome c (1 : 1000 dilu-tion), Bax (1 : 1000 dilution), B-cell leukemia/lymphoma-2 (Bcl-2) (1 : 1000 dilution), and tumor necrosis factorreceptor type 1-associated death domain (TRADD) (1 : 1000dilution) were purchased from Cell Signaling (Beverly,MA, USA). Antibodies against apoptosis-inducing factor(AIF) (1 : 100 dilution) and Fas-associated death domain(FADD) (1 : 1000 dilution) were purchased from Calbiochem(San Diego, CA, USA), antibody against cleaved caspase-8(1 : 1000 dilution) was purchased from BioVision (Moun-tain View, CA, USA), and that against Cox4 (1 : 5000)was purchased from Abcam (Cambridge, MA, USA). Theintensities of bands on the gel were calculated usingGel-Pro Analyzer (Media Cybernetics Inc., Bethesda, MD,USA).
2.9. Statistical Analysis. Data are expressed as mean ± SD.Data of sham, vehicle, and paeonol groups were comparedusing one-way ANOVA followed by posthoc Scheffe’s test. Aprobability value of less than 0.05 was considered statisticallysignificant.
3. Results
3.1. Effects of Paeonol on Neurological Deficit Induced bytMCAo. There were three groups of rats: sham, vehicle, andpaeonol. Rats in the vehicle and paeonol groups receivedtMCAo via a 10 min occlusion of the MCA, while rats in thesham group received a sham operation. Rats in the paeonolgroup were pre-administered paeonol one hour beforetMCAo. Twenty-four hours after tMCAo, rats in the vehicleand paeonol groups presented neurological deficit scores of7.0±0.8 (range, 6–8) and 5.5±2.5 (range, 5–8), respectively.Seven days after tMACo the neurological deficit score was5.4 ± 2.0 in the vehicle group and 4.3 ± 2.7 in the paeonolgroup. There were no significant differences in neurologicaldeficit score between vehicle and paeonol groups either 24hours or seven days after tMCAo (Figure 1).
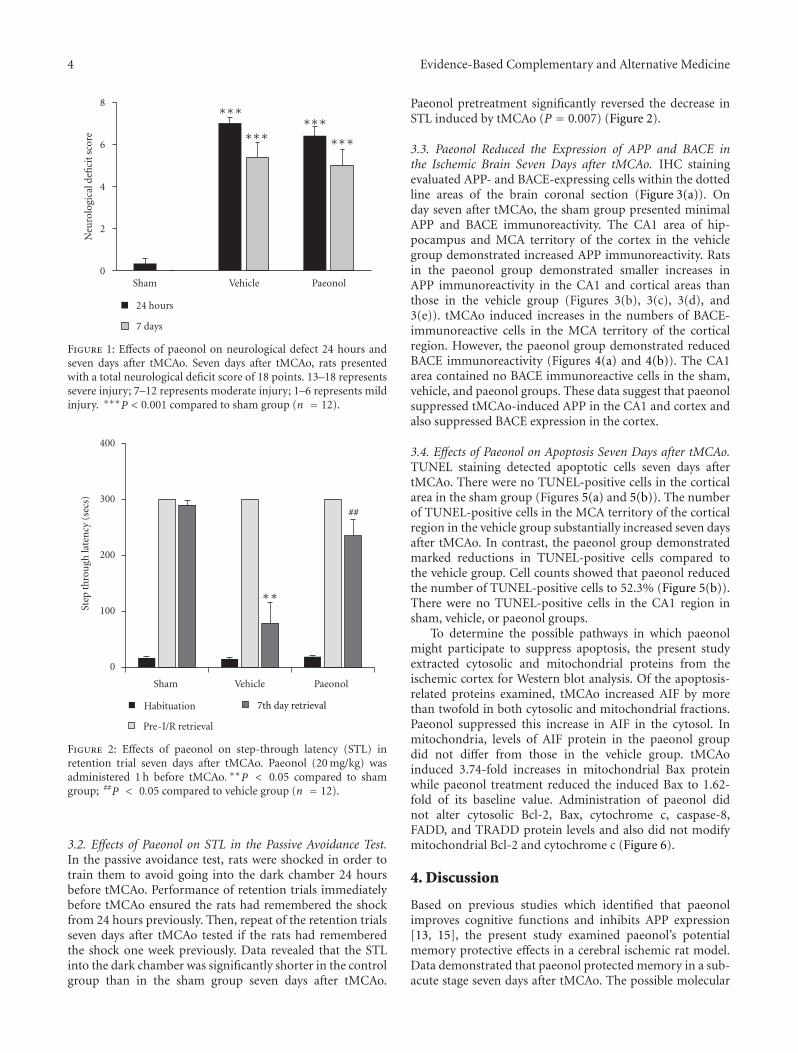
4 Evidence-Based Complementary and Alternative Medicine
0
2
4
6
8
Sham Vehicle Paeonol
24 hours
7 days
Neu
rolo
gica
l defi
cit s
core
∗∗∗
∗∗∗∗∗∗
∗∗∗
Figure 1: Effects of paeonol on neurological defect 24 hours andseven days after tMCAo. Seven days after tMCAo, rats presentedwith a total neurological deficit score of 18 points. 13–18 representssevere injury; 7–12 represents moderate injury; 1–6 represents mildinjury. ∗∗∗P < 0.001 compared to sham group (n = 12).
0
100
200
300
400
Sham Vehicle
Habituation
Pre-I/R retrieval
Step
thro
ugh
late
ncy
(se
cs)
##
∗∗
7th day retrieval
Paeonol
Figure 2: Effects of paeonol on step-through latency (STL) inretention trial seven days after tMCAo. Paeonol (20 mg/kg) wasadministered 1 h before tMCAo. ∗∗P < 0.05 compared to shamgroup; ##P < 0.05 compared to vehicle group (n = 12).
3.2. Effects of Paeonol on STL in the Passive Avoidance Test.In the passive avoidance test, rats were shocked in order totrain them to avoid going into the dark chamber 24 hoursbefore tMCAo. Performance of retention trials immediatelybefore tMCAo ensured the rats had remembered the shockfrom 24 hours previously. Then, repeat of the retention trialsseven days after tMCAo tested if the rats had rememberedthe shock one week previously. Data revealed that the STLinto the dark chamber was significantly shorter in the controlgroup than in the sham group seven days after tMCAo.
Paeonol pretreatment significantly reversed the decrease inSTL induced by tMCAo (P = 0.007) (Figure 2).
3.3. Paeonol Reduced the Expression of APP and BACE inthe Ischemic Brain Seven Days after tMCAo. IHC stainingevaluated APP- and BACE-expressing cells within the dottedline areas of the brain coronal section (Figure 3(a)). Onday seven after tMCAo, the sham group presented minimalAPP and BACE immunoreactivity. The CA1 area of hip-pocampus and MCA territory of the cortex in the vehiclegroup demonstrated increased APP immunoreactivity. Ratsin the paeonol group demonstrated smaller increases inAPP immunoreactivity in the CA1 and cortical areas thanthose in the vehicle group (Figures 3(b), 3(c), 3(d), and3(e)). tMCAo induced increases in the numbers of BACE-immunoreactive cells in the MCA territory of the corticalregion. However, the paeonol group demonstrated reducedBACE immunoreactivity (Figures 4(a) and 4(b)). The CA1area contained no BACE immunoreactive cells in the sham,vehicle, and paeonol groups. These data suggest that paeonolsuppressed tMCAo-induced APP in the CA1 and cortex andalso suppressed BACE expression in the cortex.
3.4. Effects of Paeonol on Apoptosis Seven Days after tMCAo.TUNEL staining detected apoptotic cells seven days aftertMCAo. There were no TUNEL-positive cells in the corticalarea in the sham group (Figures 5(a) and 5(b)). The numberof TUNEL-positive cells in the MCA territory of the corticalregion in the vehicle group substantially increased seven daysafter tMCAo. In contrast, the paeonol group demonstratedmarked reductions in TUNEL-positive cells compared tothe vehicle group. Cell counts showed that paeonol reducedthe number of TUNEL-positive cells to 52.3% (Figure 5(b)).There were no TUNEL-positive cells in the CA1 region insham, vehicle, or paeonol groups.
To determine the possible pathways in which paeonolmight participate to suppress apoptosis, the present studyextracted cytosolic and mitochondrial proteins from theischemic cortex for Western blot analysis. Of the apoptosis-related proteins examined, tMCAo increased AIF by morethan twofold in both cytosolic and mitochondrial fractions.Paeonol suppressed this increase in AIF in the cytosol. Inmitochondria, levels of AIF protein in the paeonol groupdid not differ from those in the vehicle group. tMCAoinduced 3.74-fold increases in mitochondrial Bax proteinwhile paeonol treatment reduced the induced Bax to 1.62-fold of its baseline value. Administration of paeonol didnot alter cytosolic Bcl-2, Bax, cytochrome c, caspase-8,FADD, and TRADD protein levels and also did not modifymitochondrial Bcl-2 and cytochrome c (Figure 6).
4. Discussion
Based on previous studies which identified that paeonolimproves cognitive functions and inhibits APP expression[13, 15], the present study examined paeonol’s potentialmemory protective effects in a cerebral ischemic rat model.Data demonstrated that paeonol protected memory in a sub-acute stage seven days after tMCAo. The possible molecular
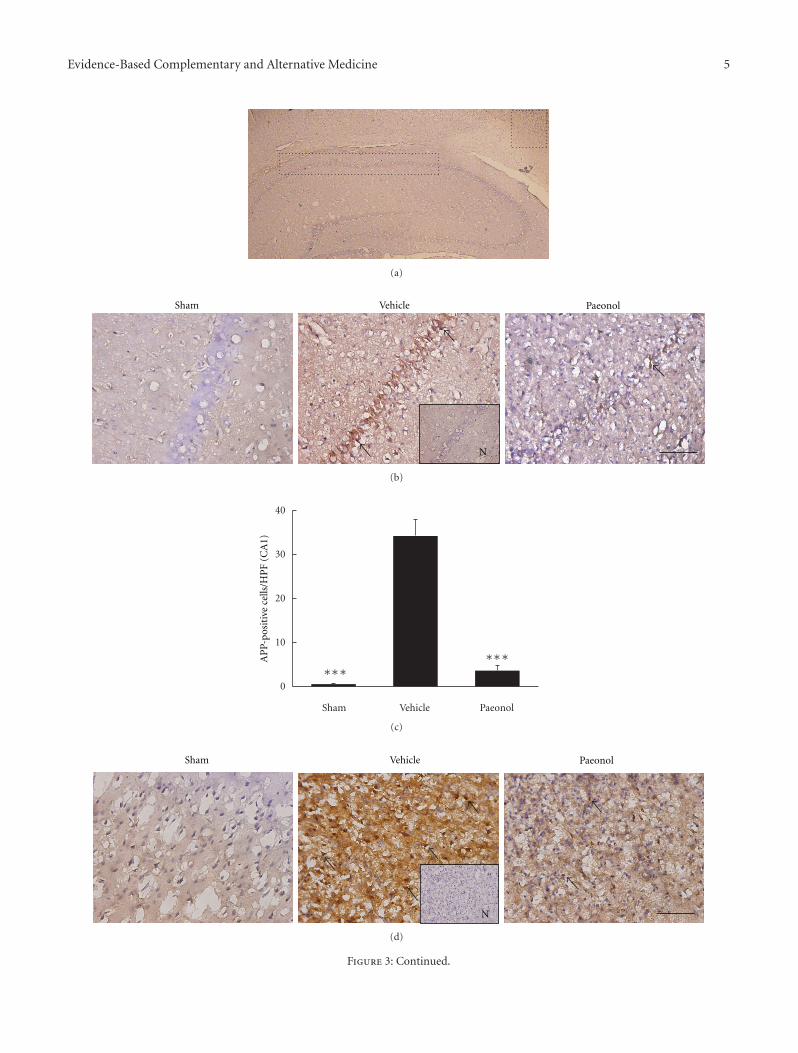
Evidence-Based Complementary and Alternative Medicine 5
(a)
Sham Vehicle Paeonol
N
(b)
0
10
20
30
40
Sham Vehicle
AP
P-po
siti
ve c
ells
/HP
F (C
A1)
∗∗∗∗∗∗
Paeonol
(c)
Sham Vehicle Paeonol
N
(d)
Figure 3: Continued.
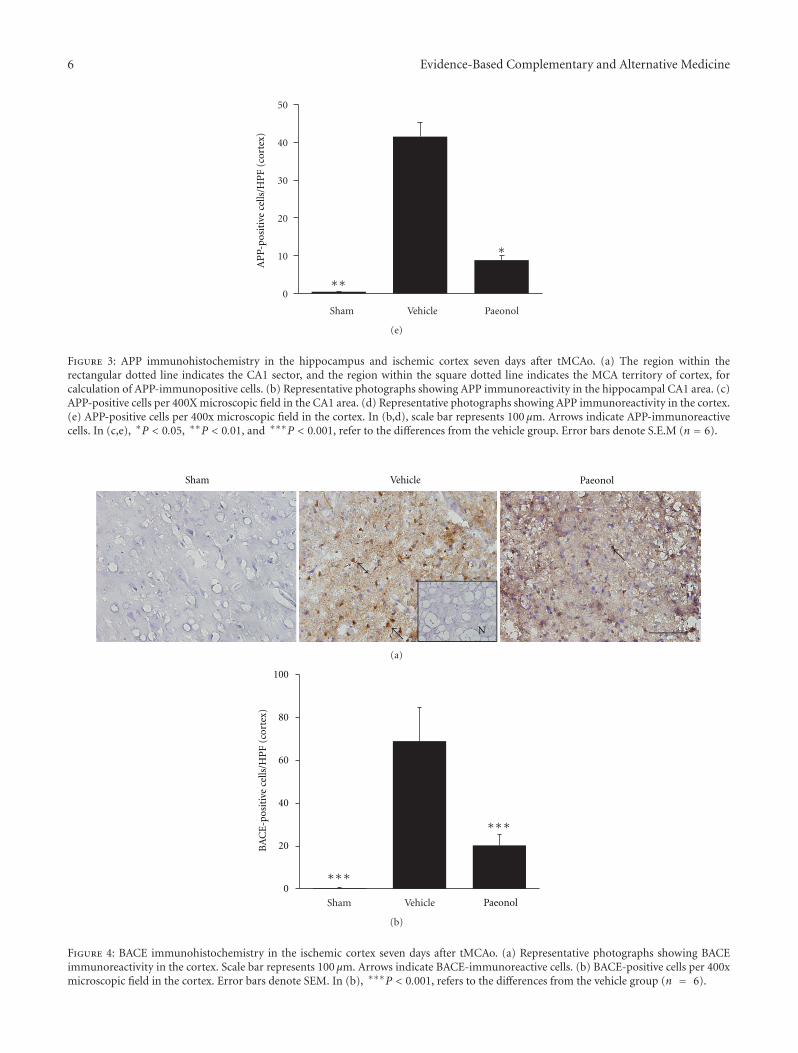
6 Evidence-Based Complementary and Alternative Medicine
0
10
20
30
40
50
Sham Vehicle
AP
P-po
siti
ve c
ells
/HP
F (c
orte
x)
∗∗
∗
Paeonol
(e)
Figure 3: APP immunohistochemistry in the hippocampus and ischemic cortex seven days after tMCAo. (a) The region within therectangular dotted line indicates the CA1 sector, and the region within the square dotted line indicates the MCA territory of cortex, forcalculation of APP-immunopositive cells. (b) Representative photographs showing APP immunoreactivity in the hippocampal CA1 area. (c)APP-positive cells per 400X microscopic field in the CA1 area. (d) Representative photographs showing APP immunoreactivity in the cortex.(e) APP-positive cells per 400x microscopic field in the cortex. In (b,d), scale bar represents 100 μm. Arrows indicate APP-immunoreactivecells. In (c,e), ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001, refer to the differences from the vehicle group. Error bars denote S.E.M (n = 6).
Sham Vehicle Paeonol
N
(a)
0
20
40
60
80
100
BA
CE
-pos
itiv
e ce
lls/H
PF
(cor
tex)
Sham Vehicle
∗∗∗
∗∗∗
Paeonol
(b)
Figure 4: BACE immunohistochemistry in the ischemic cortex seven days after tMCAo. (a) Representative photographs showing BACEimmunoreactivity in the cortex. Scale bar represents 100 μm. Arrows indicate BACE-immunoreactive cells. (b) BACE-positive cells per 400xmicroscopic field in the cortex. Error bars denote SEM. In (b), ∗∗∗P < 0.001, refers to the differences from the vehicle group (n = 6).
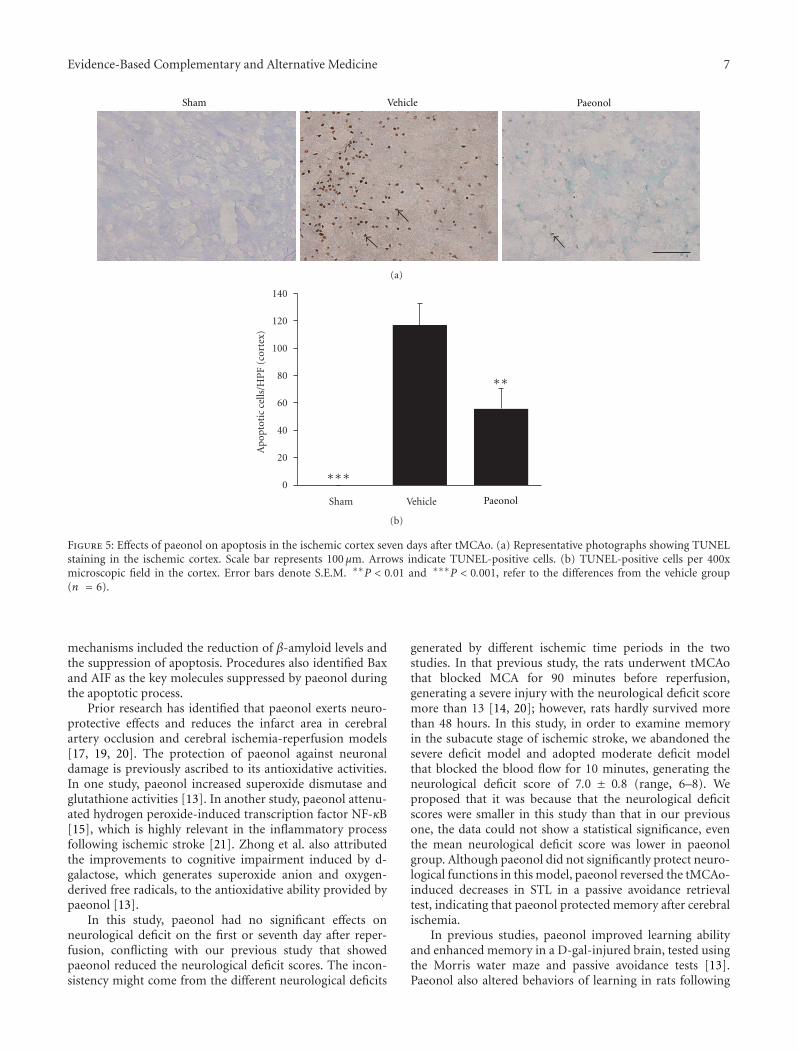
Evidence-Based Complementary and Alternative Medicine 7
Sham Vehicle Paeonol
(a)
0
20
40
60
80
100
120
140
Ap
opto
tic
cells
/HP
F (c
orte
x)
Sham Vehicle
∗∗∗
∗∗
Paeonol
(b)
Figure 5: Effects of paeonol on apoptosis in the ischemic cortex seven days after tMCAo. (a) Representative photographs showing TUNELstaining in the ischemic cortex. Scale bar represents 100 μm. Arrows indicate TUNEL-positive cells. (b) TUNEL-positive cells per 400xmicroscopic field in the cortex. Error bars denote S.E.M. ∗∗P < 0.01 and ∗∗∗P < 0.001, refer to the differences from the vehicle group(n = 6).
mechanisms included the reduction of β-amyloid levels andthe suppression of apoptosis. Procedures also identified Baxand AIF as the key molecules suppressed by paeonol duringthe apoptotic process.
Prior research has identified that paeonol exerts neuro-protective effects and reduces the infarct area in cerebralartery occlusion and cerebral ischemia-reperfusion models[17, 19, 20]. The protection of paeonol against neuronaldamage is previously ascribed to its antioxidative activities.In one study, paeonol increased superoxide dismutase andglutathione activities [13]. In another study, paeonol attenu-ated hydrogen peroxide-induced transcription factor NF-κB[15], which is highly relevant in the inflammatory processfollowing ischemic stroke [21]. Zhong et al. also attributedthe improvements to cognitive impairment induced by d-galactose, which generates superoxide anion and oxygen-derived free radicals, to the antioxidative ability provided bypaeonol [13].
In this study, paeonol had no significant effects onneurological deficit on the first or seventh day after reper-fusion, conflicting with our previous study that showedpaeonol reduced the neurological deficit scores. The incon-sistency might come from the different neurological deficits
generated by different ischemic time periods in the twostudies. In that previous study, the rats underwent tMCAothat blocked MCA for 90 minutes before reperfusion,generating a severe injury with the neurological deficit scoremore than 13 [14, 20]; however, rats hardly survived morethan 48 hours. In this study, in order to examine memoryin the subacute stage of ischemic stroke, we abandoned thesevere deficit model and adopted moderate deficit modelthat blocked the blood flow for 10 minutes, generating theneurological deficit score of 7.0 ± 0.8 (range, 6–8). Weproposed that it was because that the neurological deficitscores were smaller in this study than that in our previousone, the data could not show a statistical significance, eventhe mean neurological deficit score was lower in paeonolgroup. Although paeonol did not significantly protect neuro-logical functions in this model, paeonol reversed the tMCAo-induced decreases in STL in a passive avoidance retrievaltest, indicating that paeonol protected memory after cerebralischemia.
In previous studies, paeonol improved learning abilityand enhanced memory in a D-gal-injured brain, tested usingthe Morris water maze and passive avoidance tests [13].Paeonol also altered behaviors of learning in rats following
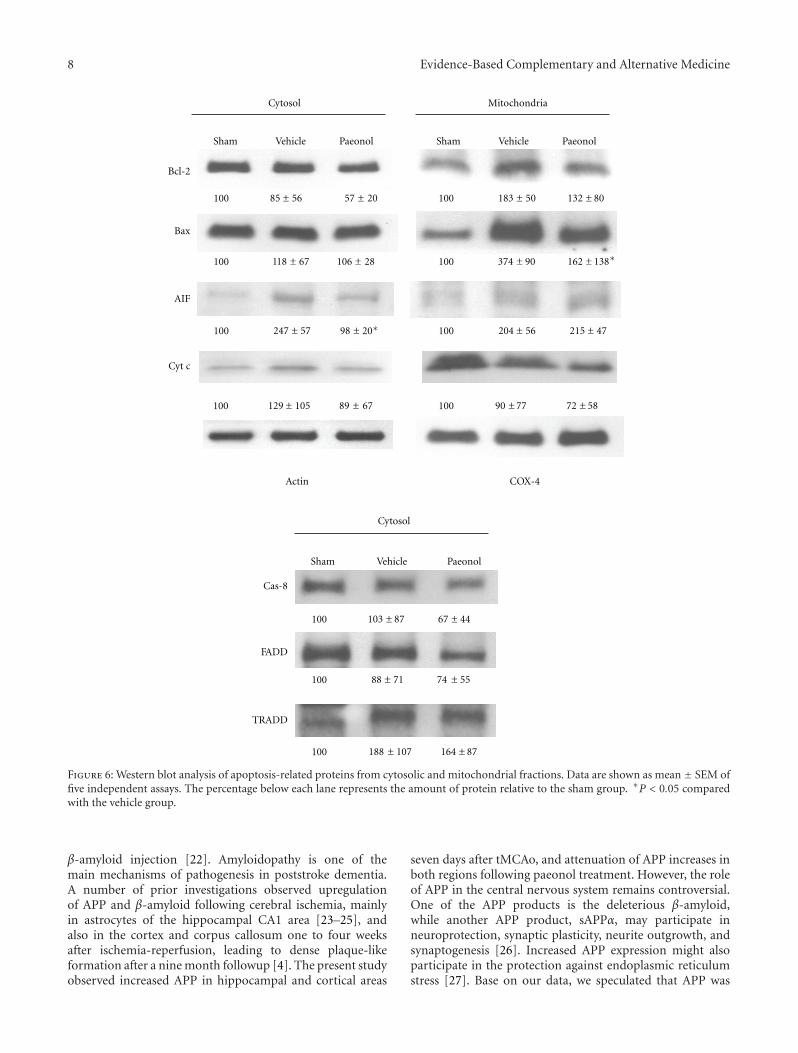
8 Evidence-Based Complementary and Alternative Medicine
100 118 ± 67 106 ± 28 100 374 ± 90 162 ± 138
Sham
Cytosol Mitochondria
Bcl-2
Bax
100 85 ± 56 57 ± 20 100 183 ± 50 132 ± 80
100 247 ± 57 98 ± 100 204 ± 56 215 ±
AIF
Cyt c
100 129 ± 105 89 ± 67 100 90 ± 77 72 ± 58
Cytosol
100 103 ± 87 67 ± 44
Cas-8
FADD
100 88 ± 71 74 ± 55
TRADD
100 188 ± 107 164 ± 87
Actin COX-4
Vehicle Paeonol Sham Vehicle Paeonol
Sham Vehicle Paeonol
20∗ 47
∗
Figure 6: Western blot analysis of apoptosis-related proteins from cytosolic and mitochondrial fractions. Data are shown as mean ± SEM offive independent assays. The percentage below each lane represents the amount of protein relative to the sham group. ∗P < 0.05 comparedwith the vehicle group.
β-amyloid injection [22]. Amyloidopathy is one of themain mechanisms of pathogenesis in poststroke dementia.A number of prior investigations observed upregulationof APP and β-amyloid following cerebral ischemia, mainlyin astrocytes of the hippocampal CA1 area [23–25], andalso in the cortex and corpus callosum one to four weeksafter ischemia-reperfusion, leading to dense plaque-likeformation after a nine month followup [4]. The present studyobserved increased APP in hippocampal and cortical areas
seven days after tMCAo, and attenuation of APP increases inboth regions following paeonol treatment. However, the roleof APP in the central nervous system remains controversial.One of the APP products is the deleterious β-amyloid,while another APP product, sAPPα, may participate inneuroprotection, synaptic plasticity, neurite outgrowth, andsynaptogenesis [26]. Increased APP expression might alsoparticipate in the protection against endoplasmic reticulumstress [27]. Base on our data, we speculated that APP was

Evidence-Based Complementary and Alternative Medicine 9
not the key factor for neuronal damage seven days aftertMCAo, because there was no apoptosis and no infarctionobserved in hippocampus (data not shown), even APP levelwas up-regulated in the same area. On the other hand,the coinduction and colocalization of BACE with apoptosisafter tMCAo in cortex suggested that BACE played a moredeleterious role than APP.
The three types of proteases which cleave APP to generateβ-amyloid are α-, β-, and γ-secretase [28]. Among the threesecretases, BACE coexpresses most highly with β-amyloid[29, 30], and controls the rate-limiting step in the productionof β-amyloid [31]. The high correlation between BACEand β-amyloid renders BACE being considered one of theeffective therapeutic targets to treat β-amyloid-related dis-eases [32, 33]. In previous studies, focal ischemia increasedthe activity of BACE, especially BACE1, in the ischemiccortex [7] and thalamus [34]. Consistent with a previousstudy by Wen et al. [7], the present study observed BACEinduction and apoptosis occurred in the same location, thatis, ischemic cortex, suggesting the high relevance of BACEand apoptosis in ischemic stroke. Paeonol treatment limitedthe induction of BACE and suppressed apoptosis, indicatingthat suppression of β-amyloid production by paeonol mightclosely link to the suppression of apoptosis, and mightparticipate in the memory protection after cerebral ischemia.
After cerebral ischemia, apoptosis occurs in CA1 withinthree days of cerebral ischemia, disappears after seven days,and do not reoccur after a further seven days [35, 36]. Inthe present study, apoptosis was observed in the ischemiccortex, but not in the CA1 region, seven days after tMCAo.The suppression of apoptosis in the ischemic cortex indicatedthat paeonol protected neuronal cells from tMCAo injury.In recent research, paeonol also has been shown to inhibitglutamate-induced apoptosis in PC12 cells [37] and in ratbrains receiving β-amyloid injection [22], while the detailedmechanisms relating to the suppression of apoptosis inneuronal cells have yet to be identified. There are twogeneral pathways of apoptosis following cerebral ischemia:the intrinsic and extrinsic pathways. The intrinsic pathway isinitiated by the accumulation of intracellular Ca2+ followingthe depletion of energy after ischemia [10]. Activation ofcalpains by Ca2+ results in cleaving of the Bcl-2 interactingdomain (BID) to its truncated form (tBID), which targets theouter mitochondrial membrane and induces conformationalchanges in proapoptotic proteins, such as Bak, Bax, Bad, andBcl-XS. These pro-apoptotic proteins can also heterdimerisewith antiapoptotic proteins, including Bcl-2 or Bcl-XL[38]. The binding of tBID to pro-apoptotic proteins opensthe mitochondrial transition pore, leading to the releaseof cytochrome c and AIF into the cytosol [39]. Oncereleased from mitochondria, cytochrome c binds to andactivates caspase-9, and then caspase-3, which cleaves poly(ADP-ribose) polymerase (PARP), leading to DNA injuryand caspase-dependent apoptotic cell death [40]. CytosolicAIF further translocates into the nucleus and stimulatescaspase-independent DNA fragmentation [41]. The extrinsicpathway starts from the activation of the cell surface Fas andtumor necrosis receptors. The Fas receptor triggers FADDdirectly and tumor necrosis receptor triggers FADD via
TRADD, activating caspase-8, and then caspase-3, resultingin PARP cleavage and DNA damage [42]. The present studyis the first to identify Bax and AIF as the two main moleculesmodified by paeonol which paeonol can adjust during thesuppression of apoptosis following cerebral ischemia.
The reduction of Bax by paeonol suggests that paeonolmight suppress the apoptosis by inhibiting the releaseof mitochondrial factor into cytosol. The simultaneoussuppression of cytosolic AIF supported this assumption.However, level of cytochrome c was not significantly changedby paeonol, although the mean value of cytochrome c waslower in the paeonol group (0.89-fold sham) than that inthe vehicle group (1.29-fold sham). We proposed that thesmall enhancement of cytochrome c decreased the statisticalsignificance. A report by Zhu et al. described that aftercerebral ischemia, AIF expression was more pronouncedin neurons of the male brain, while female brain neuronsshowed marked increases in expression of caspase-3 [43].The present study with male rats induced a 2.47-fold ofelevated AIF in cytosol, which was reversed to almostbaseline levels. There are only a few herbs which are knownto regulate the expression of AIF. Isatis indigotica [44]and polyphyllin D [45] increase the release of AIF frommitochondria, causing apoptosis, while berberine inhibitsAIF and therefore suppresses apoptosis [46].
Medicinal plant extracts and natural antioxidants mighthave potential use in the prevention and treatment ofdementia [2, 47]. Results from the present study indicate thatpaeonol, which derives from a widely administered medici-nal herb, protects the memory following cerebral ischemia.Paeonol treatment reduced APP and BACE expression, andalso the numbers of apoptotic cells. Suppression of the levelof Bax protein and blocking AIF from releasing to the cytosolmight also be the mechanisms in which paeonol exerted itseffect on anti-apoptosis and memory protection.
Abbreviations
AIF: Apoptosis-inducing factorAPP: Amyloid precursor proteinBACE: Beta-site APP cleaving enzymeBcl-2: B-cell leukemia/lymphoma-2CCA: Common carotid arteryDAB: 3,3′-DiaminobenzidineHPF: High power fieldICA: Internal carotid arteryIHC: ImmunohistochemistryMCA: Middle cerebral arterytMCAo: Transient middle cerebral artery occlusionPBS: Phosphate-buffered salineSTL: Step-through latencyTUNEL: Terminal deoxynucleotidyl transferase-mediated
dUTP-biotin nick end labelling.
Acknowledgments
This study was supported by grants from the China MedicalUniversity Hospital, Taiwan (DMR-99-002), and in part by

10 Evidence-Based Complementary and Alternative Medicine
the Taiwan Department of Health Clinical Trial and ResearchCenter of Excellence (DOH101-TD-B-111-004).
References
[1] V. Solfrizzi, F. Panza, A. M. Colacicco et al., “Vascular riskfactors, incidence of MCI, and rates of progression to demen-tia,” Neurology, vol. 63, no. 10, pp. 1882–1891, 2004.
[2] R. N. Kalaria, G. E. Maestre, R. Arizaga et al., “Alzheimer’sdisease and vascular dementia in developing countries: preva-lence, management, and risk factors,” The Lancet Neurology,vol. 7, no. 9, pp. 812–826, 2008.
[3] M. Lesniak, T. Bak, W. Czepiel, J. Seniow, and A. Członkowska,“Frequency and prognostic value of cognitive disorders instroke patients,” Dementia and Geriatric Cognitive Disorders,vol. 26, no. 4, pp. 356–363, 2008.
[4] R. Pluta, “From brain ischemia-reperfusion injury to possiblesporadic Alzheimer’s disease,” Current Neurovascular Research,vol. 1, no. 5, pp. 441–453, 2004.
[5] R. Pluta, M. Ułamek, and M. Jabłonski, “Alzheimer’s mecha-nisms in ischemic brain degeneration,” Anatomical Record, vol.292, no. 12, pp. 1863–1881, 2009.
[6] M. Sastre, I. Dewachter, G. E. Landreth et al., “Nonsteroidalanti-inflammatory drugs and peroxisome proliferator-activated receptor-γ agonists modulate immunostimulatedprocessing of amyloid precursor protein through regulationof β-secretase,” Journal of Neuroscience, vol. 23, no. 30, pp.9796–9804, 2003.
[7] Y. Wen, O. Onyewuchi, S. Yang, R. Liu, and J. W. Simpkins,“Increased β-secretase activity and expression in rats followingtransient cerebral ischemia,” Brain Research, vol. 1009, no. 1-2,pp. 1–8, 2004.
[8] T. Van Groen, K. Puurunen, H. M. Maki, J. Sivenius, and J.Jolkkonen, “Transformation of diffuse β-amyloid precursorprotein and β-amyloid deposits to plaques in the thalamusafter transient occlusion of the middle cerebral artery in rats,”Stroke, vol. 36, no. 7, pp. 1551–1556, 2005.
[9] J. T. Rogers, L. M. Leiter, J. McPhee et al., “Translation of theAlzheimer amyloid precursor protein mRNA is up- regulatedby interleukin-1 through 5’-untranslated region sequences,”Journal of Biological Chemistry, vol. 274, no. 10, pp. 6421–6431, 1999.
[10] B. R. S. Broughton, D. C. Reutens, and C. G. Sobey, “Apoptoticmechanisms after cerebral ischemia,” Stroke, vol. 40, no. 5, pp.e331–e339, 2009.
[11] R. C. Taylor, S. P. Cullen, and S. J. Martin, “Apoptosis: con-trolled demolition at the cellular level,” Nature Reviews Molec-ular Cell Biology, vol. 9, no. 3, pp. 231–241, 2008.
[12] X. F. Sun, “Utilization of traditional chinese medicines (tcm)for strengthening body resistance during 2007–2009 period,”Evaluation and Analysis of Drug-Use in Hospitals of China, vol.3, pp. 244–246, 2011.
[13] S. Z. Zhong, Q. H. Ge, R. Qu, Q. Li, and S. P. Ma, “Paeonolattenuates neurotoxicity and ameliorates cognitive impair-ment induced by d-galactose in ICR mice,” Journal of theNeurological Sciences, vol. 277, no. 1-2, pp. 58–64, 2009.
[14] C. L. Hsieh, C. Y. Cheng, T. H. Tsai et al., “Paeonol re-duced cerebral infarction involving the superoxide anion andmicroglia activation in ischemia-reperfusion injured rats,”Journal of Ethnopharmacology, vol. 106, no. 2, pp. 208–215,2006.
[15] S.-Y. Su, C.-Y. Cheng, T.-H. Tsai, C.-Y. Hsiang, T.-Y. Ho,and C.-L. Hsieh, “Paeonol attenuates H2O2-induced NF-κB-
associated amyloid precursor protein expression,” AmericanJournal of Chinese Medicine, vol. 38, no. 6, pp. 1171–1192,2010.
[16] C. Y. Cheng, S. Y. Su, N. Y. Tang, T. Y. Ho, S. Y. Chiang,and C. L. Hsieh, “Ferulic acid provides neuroprotectionagainst oxidative stress-related apoptosis after cerebral ische-mia/reperfusion injury by inhibiting ICAM-1 mRNA expres-sion in rats,” Brain Research, vol. 1209, pp. 136–150, 2008.
[17] J. Chen, P. R. Sanberg, Y. Li et al., “Intravenous administrationof human umbilical cord blood reduces behavioral deficitsafter stroke in rats,” Stroke, vol. 32, no. 11, pp. 2682–2688,2001.
[18] C. Y. Hsiang, S. L. Wu, and T. Y. Ho, “Morin inhibits12-O-tetradecanoylphorbol-13-acetate-induced hepatocellu-lar transformation via activator protein 1 signaling pathwayand cell cycle progression,” Biochemical Pharmacology, vol. 69,no. 11, pp. 1603–1611, 2005.
[19] J. M. Xiao, W. C. Si, J. W. Wen et al., “Anxiolytic-like effectof paeonol in mice,” Pharmacology Biochemistry and Behavior,vol. 81, no. 3, pp. 683–687, 2005.
[20] C. Y. Cheng, T. Y. Ho, E. J. Lee, S. Y. Su, N. Y. Tang, and C.L. Hsieh, “Ferulic acid reduces cerebral infarct through its an-tioxidative and anti-Inflammatory effects following transientfocal cerebral ischemia in rats,” American Journal of ChineseMedicine, vol. 36, no. 6, pp. 1105–1119, 2008.
[21] Q. Wang, X. N. Tang, and M. A. Yenari, “The inflammatoryresponse in stroke,” Journal of Neuroimmunology, vol. 184, no.1-2, pp. 53–68, 2007.
[22] J. Zhou, L. Zhou, D. Hou, J. Tang, J. Sun, and S. C. Bondy,“Paeonol increases levels of cortical cytochrome oxidase andvascular actin and improves behavior in a rat model of Alz-heimer’s disease,” Brain Research, vol. 1388, pp. 141–147, 2011.
[23] T. Nihashi, S. Inao, Y. Kajita et al., “Expression and distri-bution of beta amyloid precursor protein and beta amyloidpeptide in reactive astrocytes after transient middle cerebralartery occlusion,” Acta Neurochirurgica, vol. 143, no. 3, pp.287–295, 2001.
[24] R. Pluta, “Astroglial expression of the β-amyloid in ischemia-reperfusion brain injury,” Annals of the New York Academy ofSciences, vol. 977, pp. 102–108, 2002.
[25] A. Shiino, M. Matsuda, J. Handa, and B. Chance, “Poor re-covery of mitochondrial redox state in CA1 after transientforebrain ischemia in gerbils,” Stroke, vol. 29, no. 11, pp. 2421–2425, 1998.
[26] D. Kogel, T. Deller, and C. Behl, “Roles of amyloid precursorprotein family members in neuroprotection, stress signalingand aging,” Experimental Brain Research. In press.
[27] D. Kogel, R. Schomburg, T. Schurmann et al., “The amyloidprecursor protein protects PC12 cells against endoplasmicreticulum stress-induced apoptosis,” Journal of Neurochem-istry, vol. 87, no. 1, pp. 248–256, 2003.
[28] M. Citron, “Secretases as targets for the treatment of Alzheim-er’s disease,” Molecular Medicine Today, vol. 6, no. 10, pp. 392–397, 2000.
[29] U. Bodendorf, S. Danner, F. Fischer et al., “Expression ofhuman β-secretase in the mouse brain increases the steady-state level of β-amyloid,” Journal of Neurochemistry, vol. 80,no. 5, pp. 799–806, 2002.
[30] R. Vassar, “The β-secretase, BACE: a prime drug target forAlzheimer’s disease,” Journal of Molecular Neuroscience, vol.17, no. 2, pp. 157–170, 2001.
[31] D. Fleck, A. N. Garratt, C. Haass, and M. Willem, “Ba-ce1 dependent neuregulin proteolysis,” Current AlzheimerResearch. In press.

Evidence-Based Complementary and Alternative Medicine 11
[32] E. Tamagno, M. Guglielmotto, D. Monteleone, and M. Taba-ton, “Amyloid-β production: major link between oxidativestress and BACE1,” Neurotoxicity Research. In press.
[33] P. Giusti-Rodrıguez, J. Gao, J. Graff, D. Rei, T. Soda, and L.-H.Tsai, “Synaptic deficits are rescued in the p25/Cdk5 model ofneurodegeneration by the reduction of β-Secretase (BACE1),”Journal of Neuroscience, vol. 31, no. 44, pp. 15751–15756, 2011.
[34] M. Hiltunen, P. Makinen, S. Peraniemi et al., “Focal cerebralischemia in rats alters APP processing and expression of Aβpeptide degrading enzymes in the thalamus,” Neurobiology ofDisease, vol. 35, no. 1, pp. 103–113, 2009.
[35] T. Sugawara, M. Kawase, A. Lewen et al., “Effect of hypoten-sion severity on hippocampal CA1 neurons in a rat globalischemia model,” Brain Research, vol. 877, no. 2, pp. 281–287,2000.
[36] D. Yin, C. Zhou, I. Kusaka et al., “Inhibition of apoptosis byhyperbaric oxygen in a rat focal cerebral ischemic model,”Journal of Cerebral Blood Flow and Metabolism, vol. 23, no. 7,pp. 855–864, 2003.
[37] X. Wang, G. Zhu, S. Yang et al., “Paeonol prevents excitotoxi-city in rat pheochromocytoma PC12 cells via downregulationof ERK activation and inhibition of apoptosis,” Planta Medica,vol. 77, no. 15, pp. 1695–1701, 2011.
[38] T. Sugawara, M. Fujimura, N. Noshita et al., “Neuronaldeath/survival signaling pathways in cerebral ischemia,” Neu-roRx, vol. 1, no. 1, pp. 17–25, 2004.
[39] R. A. Kirkland, J. A. Windelborn, J. M. Kasprzak, and J.L. Franklin, “A bax-induced pro-oxidant state is critical forcytochrome c release during programmed neuronal death,”Journal of Neuroscience, vol. 22, no. 15, pp. 6480–6490, 2002.
[40] S. Namura, J. Zhu, K. Fink et al., “Activation and cleavageof caspase-3 in apoptosis induced by experimental cerebralischemia,” Journal of Neuroscience, vol. 18, no. 10, pp. 3659–3668, 1998.
[41] B. B. Cho and L. H. Toledo-Pereyra, “Caspase-independentprogrammed cell death following ischemic stroke,” Journal ofInvestigative Surgery, vol. 21, no. 3, pp. 141–147, 2008.
[42] S. Love, “Apoptosis and brain ischaemia,” Progress in Neuro-Psychopharmacology and Biological Psychiatry, vol. 27, no. 2,pp. 267–282, 2003.
[43] C. Zhu, X. Wang, Z. Huang et al., “Apoptosis-inducing factoris a major contributor to neuronal loss induced by neonatalcerebral hypoxia-ischemia,” Cell Death and Differentiation,vol. 14, no. 4, pp. 775–784, 2007.
[44] Y.-C. Chung, F.-Y. Tang, J.-W. Liao et al., “Isatis indig-otica induces hepatocellular cancer cell death via caspase-independent apoptosis-inducing factor translocation apop-totic pathway in vitro and in vivo,” Integrative Cancer Ther-apies, vol. 10, no. 2, pp. 201–214, 2011.
[45] J. Yuen-Nei Cheung, R. Chik-Ying Ong, Y. K. Suen et al.,“Polyphyllin D is a potent apoptosis inducer in drug-resistantHepG2 cells,” Cancer Letters, vol. 217, no. 2, pp. 203–211, 2005.
[46] X. Q. Zhou, X. N. Zeng, H. Kong, and X. L. Sun, “Neuropro-tective effects of berberine on stroke models in vitro and invivo,” Neuroscience Letters, vol. 447, no. 1, pp. 31–36, 2008.
[47] B. Zhao, “Natural antioxidants protect neurons in Alzheimer’sdisease and parkinson’s disease,” Neurochemical Research, vol.34, no. 4, pp. 630–638, 2009.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 501032, 10 pagesdoi:10.1155/2012/501032
Research Article
Neuroprotective Effects of San-Huang-Xie-Xin-Tang in theMPP+/MPTP Models of Parkinson’s Disease In Vitro and In Vivo
Yi-Ching Lo,1, 2, 3 Yu-Tzu Shih,1 Yu-Ting Tseng,2 and Hung-Te Hsu3, 4
1 Department of Pharmacology, School of Medicine, College of Medicine, Kaohsiung Medical University, 100 Shih-Chuan 1st Road,Kaohsiung 80708, Taiwan
2 Graduate Institute of Natural Products, Kaohsiung Medical University, Kaohsiung 80708, Taiwan3 Graduate Institute of Medicine, Kaohsiung Medical University, Kaohsiung 80708, Taiwan4 Department of Anesthesia, Kaohsiung Medical University Hospital, Kaohsiung 80708, Taiwan
Correspondence should be addressed to Yi-Ching Lo, [email protected]
Received 16 November 2011; Accepted 17 December 2011
Academic Editor: Ilkay Erdogan Orhan
Copyright © 2012 Yi-Ching Lo et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
San-Huang-Xie-Xin-Tang (SHXT), composed of Coptidis rhizoma, Scutellariae radix, and Rhei rhizoma, is a traditional Chinesemedicine used for complementary and alternative therapy of cardiovascular and neurodegenerative diseases via its anti-inflammatory and antioxidative effects. The aim of this study is to investigate the protective effects of SHXT in the 1–methyl–4–phenylpyridinium (MPP+)/1–methyl–4–phenyl–1,2,3,6–tetrahydropyridine (MPTP) models of Parkinson’s disease. Rat primarymesencephalic neurons and mouse Parkinson disease model were used in this study. Oxidative stress was induced by MPP+ in vitroand MPTP in vivo. In MPP+-treated mesencephalic neuron cultures, SHXT significantly increased the numbers of TH-positiveneurons. SHXT reduced apoptotic signals (cytochrome and caspase) and apoptotic death. MPP+-induced gp91phox activation andROS production were attenuated by SHXT. In addition, SHXT increased the levels of GSH and SOD in MPP+-treated neurons. InMPTP animal model, SHXT markedly increased TH-positive neurons in the substantia nigra pars compacta (SNpc) and improvedmotor activity of mice. In conclusion, the present results reveal the evidence that SHXT possesses beneficial protection againstMPTP-induced neurotoxicity in this model of Parkinson’s disease via its antioxidative and antiapoptotic effects. SHXT might be apotentially alternative and complementary medicine for neuroprotection.
1. Introduction
San-Huang-Xie-Xin-Tang (SHXT) is a traditional Chinesemedicinal formula, containing Coptidis rhizoma (Coptischinesis Franch), Scutellariae radix (Scutellaria baicalensisGeorgi), and Rhei rhizoma (Rheum officinale Baill). SHXTis traditionally used to treat various diseases via its anti-inflammatory effects. Some of the major components (e.g.,baicalin, baicalein, wogonin, and emodin) in SHXT haveidentified their potential protection on neuron. Baicalinprotects against cerebral ischemia/reperfusion in vivo andoxygen-glucose deprivation in vitro [1, 2]. Baicalein prevents6-OHDA-induced experimental parkinsonism [3], ischemiainjury [4], and traumatic brain injury [5]. Baicalein [6]and wogonin [7] attenuate inflammation-mediated neurode-generation. Emodin, an anthraquinone derivative extractedfrom Rhei rhizoma, decreased glutamate excitotoxicity [8]
and β-amyloid-induced neurotoxicity [9]. Accordingly,SHXT is standardized and pharmacologically investigated inour lab. We demonstrate that SHXT attenuates lipopolysac-charide (LPS) and Helicobacter pylori-induced inflammatoryresponses [10–12]. We also reveal the potential benefits ofSHXT used in the treatment of pulmonary hypertension[13]. We further prove the potential protective mechanismsof SHXT on activated microglia- and 6-OHDA-induced neu-rotoxicity via its anti-inflammatory and antioxidative prop-erties [14].
Oxidative stress has been implicated in the progressionand cellular damage in many neurodegenerative disorders,such as Parkinson’s disease (PD), Alzheimer’s disease (AD),and amyotrophic lateral sclerosis (ALS) [15]. PD is aprogressive neurodegenerative disease characterized by a lossof dopaminergic neurons in substantia nigra pars compacta

2 Evidence-Based Complementary and Alternative Medicine
(SNpc) and can be modeled by the neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [16]. Oxidativestress-induced mitochondrial dysfunction is recognized as akey player in the pathogenesis of PD [17]. MPTP can crossthe blood-brain barrier and converts to its active metabolite1-methyl-4-phenylpyridinium (MPP+) [18], which is selec-tively accumulated by high-affinity dopamine transportersinto the mitochondria of dopaminergic neurons and induceddopaminergic neuron death [19]. Since SHXT possessesnovel anti-inflammatory and antioxidative effects, therefore,we aimed to investigate the protective effects and mecha-nisms of SHXT on oxidative stress-induced neurotoxicityby using MPP+ in vitro model and MPTP mouse model ofParkinson’s disease.
2. Materials and Methods
2.1. Materials. The blended mixture of Coptidis rhizoma,root of Scutellariae radix, and Rhei rhizoma was preparedin a ratio of 1 : 1 : 2, respectively. The voucher specimen,method for extraction and contents of each component inSHXT were described in our previous study [11]. Briefly, themixture was extracted with 5 parts distilled water for 1 h,centrifuged at 1500×g at room temperature to obtain thesupernatant and then concentrated under reduced pressureat 65◦C to obtain the semisolid form of SHXT (yield: 23.2%),which was made up by distilled water to contain 60%water. The content (μg/mL) of each component in SHXTanalyzed by HPLC is as follows: baicalin 1153.07 ± 56.36,baicalein 82.81±3.74, emodin 11.15±1.22, wogonin 19.55±0.83, rhein 126.12 ± 3.84, berberine 62.14 ± 4.27, coptisine6.15 ± 0.34, palmatine 25.11 ± 3.78, sennoside A 128.02 ±13.56, and sennoside B 95.90 ± 3.59. Minimum essentialmedium (MEM), fetal bovine serum (FBS), horse serum,glutamine, B27, nonessential aminoacids, sodium pyruvate,penicillin, amphotericin B, streptomycin, and Alexa Fluor488 goat anti-rabbit IgG (H + L) were obtained fromInvitrogen (Carlsbad, CA, USA). All materials for SDS-PAGE were obtained from Bio-Rad (Hercules, CA, USA).Mouse antibodies against cytochrome c, rabbit antibodyagainst TH and caspase-3, and all horseradish peroxidase-conjugated secondary antibodies were obtained from SantaCruz Biotechnology (Santa Cruz, CA, USA). Mouse antibodyagainst gp91phox was obtained from BD Bioscience (SanJose, CA, USA). Mouse anticaspase-9 was obtained fromMillipore (Bedford, MA, USA). Enhanced chemilumines-cence reagent and polyvinylidene difluoride (PVDF) mem-brane were purchased from PerkinElmer Life and AnalyticalSciences (Boston, MA, USA). LDH cytotoxicity assay kitwas purchased from G-Biosciences (St. Louis, MO, USA).Annexin-V-FITC assay kit was obtained from Strong BiotechCorporation (Taipei, Taiwan). Glutathione measurement kitand superoxide dismutase activity kit were purchased fromAssay Designs (Ann Arbor, MI, USA). Simple stain mouseMAX PO was purchased from Nichirei Biosciences (Tokyo,Japan). Diaminobenzidine (DAB) kit was obtained fromBioGenex (San Ramon, CA, USA). All other materials werepurchased from Sigma Chemical Company (St. Louis, MO,USA).
2.2. Animals. All animals used for this study were approvedby the Animal Care and Use Committee at the KaohsiungMedical University. Pregnant Sprague-Dawley rats and maleC57BL/6 mice (7-8 weeks old, 20–25 g) were purchasedfrom the National Laboratory Animal Breeding and ResearchCenter (Taipei, Taiwan). They were housed under conditionsof constant temperature and controlled illumination (lightson between 7 : 30 and 19:30).
2.3. MPTP Mouse Model. Experimental Parkinson’s diseasemodel was established by intraperitoneal (i.p.) injection ofMPTP (20 mg/kg four times at 2 h interval). Mice receivedSHXT (10 mg/kg or 20 mg/kg, i.p., once per day) for 7 dayswhile MPTP was given on the 8th day. Control animalsreceived saline only. Mice have to undergo locomotor activityassay 7 days after MPTP treatment and sacrificed for THimmunohistochemistry.
2.4. Primary Mesencephalic Neuron Cultures. Primary mes-encephalic neuron cultures were prepared from gestationday 15-16 rat embryos (E15-16). In brief, meninges-freeventral mesencephalon were isolated and suspended. Cellswere plated onto 6-, 24-, or 96- well plates precoatedwith 20 μg/mL poly-L-lysine in MEM containing 10% FBS,10% horse serum, 1 g/l glucose, 2 mM L-glutamine, 1 mMsodium pyruvate, 100 μM nonessential aminoacids, 100U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mLamphotericin B at 37◦C in a humidified incubator under 5%CO2 and 95% air. After 24 h incubation, culture mediumwas replaced by MEM supplemented with 2% B27 and10 μM cytosine arabinoside to control glia proliferation.After further 48 h, the medium was replaced with MEMcontaining 2% B27 and the cells were allowed to develop invitro for 6 days.
2.5. MTT Assay. Cell viability was measured by a quantita-tive colorimetric assay with MTT, showing the mitochondrialactivity of living cells. After indicated treatments, cells wereincubated with 0.1 mg/mL MTT for 3 h at 37◦C. The reactionwas determined by addition of 100 μL DMSO. The amountof MTT formazan product was determined by measuring theabsorbance at 560 nm using a microplate reader.
2.6. LDH Assay. Cytotoxicity was evaluated by colorimetricassay based on the measurement of lactate dehydrogenase(LDH) activity release from damaged cells into the super-natant. The release of LDH is measured with a coupledenzymatic reaction using a cytotoxicity detection kit thatresults in the conversion of a tetrazolium salt into a red colorformazan. The amount of formazan formed correlated withLDH activity. The formazan product was measured with amicroplate reader at 490 nm.
2.7. TH Immunocytochemistry. Immunocytochemistry assaywas used to detect the expression of tyrosine hydroxylase(TH) and used to estimate the number of dopaminergicneurons in neuron cultures. After the cells were pretreatedwith SHXT for 1 h in 24-well plates and followed by exposure
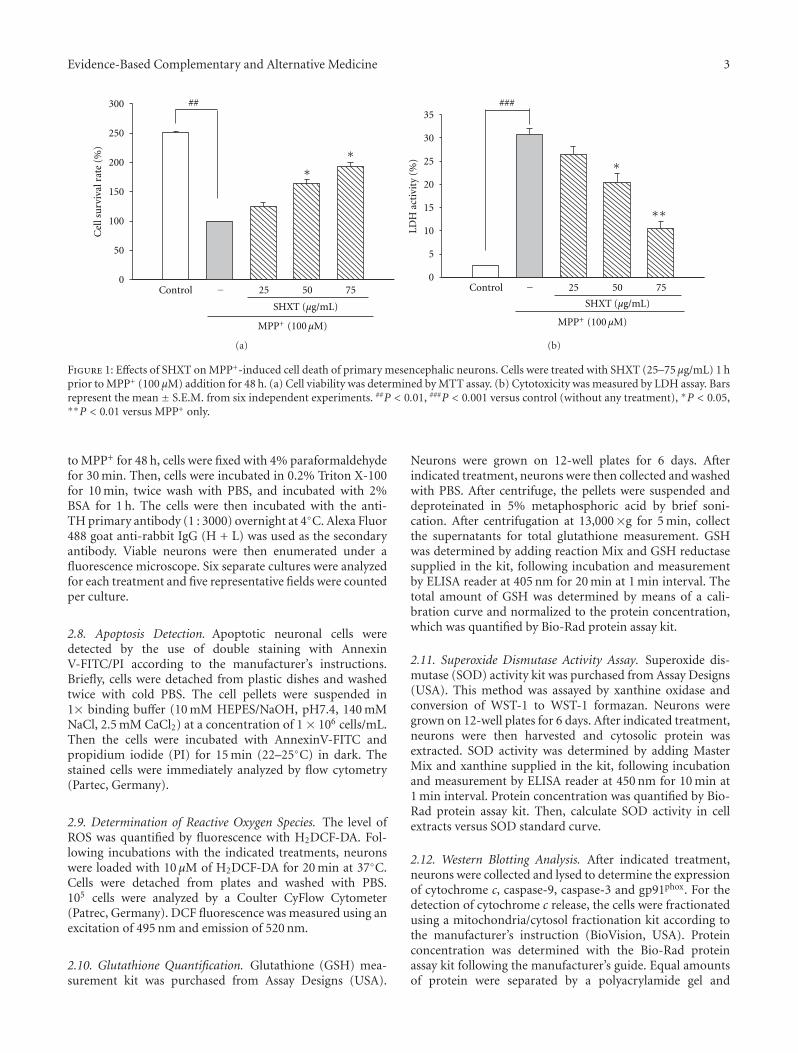
Evidence-Based Complementary and Alternative Medicine 3
##C
ell s
urv
ival
rat
e (%
)
SHXT (μg/mL)
MPP+ (100 μM)
∗∗
0
50
100
150
200
250
300
Control − 25 50 75
(a)
LD
H a
ctiv
ity
(%)
SHXT (μg/mL)
MPP+ (100 μM)
∗
###
∗∗
0
5
10
15
20
25
30
35
Control − 25 50 75
(b)
Figure 1: Effects of SHXT on MPP+-induced cell death of primary mesencephalic neurons. Cells were treated with SHXT (25–75 μg/mL) 1 hprior to MPP+ (100 μM) addition for 48 h. (a) Cell viability was determined by MTT assay. (b) Cytotoxicity was measured by LDH assay. Barsrepresent the mean ± S.E.M. from six independent experiments. ##P < 0.01, ###P < 0.001 versus control (without any treatment), ∗P < 0.05,∗∗P < 0.01 versus MPP+ only.
to MPP+ for 48 h, cells were fixed with 4% paraformaldehydefor 30 min. Then, cells were incubated in 0.2% Triton X-100for 10 min, twice wash with PBS, and incubated with 2%BSA for 1 h. The cells were then incubated with the anti-TH primary antibody (1 : 3000) overnight at 4◦C. Alexa Fluor488 goat anti-rabbit IgG (H + L) was used as the secondaryantibody. Viable neurons were then enumerated under afluorescence microscope. Six separate cultures were analyzedfor each treatment and five representative fields were countedper culture.
2.8. Apoptosis Detection. Apoptotic neuronal cells weredetected by the use of double staining with AnnexinV-FITC/PI according to the manufacturer’s instructions.Briefly, cells were detached from plastic dishes and washedtwice with cold PBS. The cell pellets were suspended in1× binding buffer (10 mM HEPES/NaOH, pH7.4, 140 mMNaCl, 2.5 mM CaCl2) at a concentration of 1 × 106 cells/mL.Then the cells were incubated with AnnexinV-FITC andpropidium iodide (PI) for 15 min (22–25◦C) in dark. Thestained cells were immediately analyzed by flow cytometry(Partec, Germany).
2.9. Determination of Reactive Oxygen Species. The level ofROS was quantified by fluorescence with H2DCF-DA. Fol-lowing incubations with the indicated treatments, neuronswere loaded with 10 μM of H2DCF-DA for 20 min at 37◦C.Cells were detached from plates and washed with PBS.105 cells were analyzed by a Coulter CyFlow Cytometer(Patrec, Germany). DCF fluorescence was measured using anexcitation of 495 nm and emission of 520 nm.
2.10. Glutathione Quantification. Glutathione (GSH) mea-surement kit was purchased from Assay Designs (USA).
Neurons were grown on 12-well plates for 6 days. Afterindicated treatment, neurons were then collected and washedwith PBS. After centrifuge, the pellets were suspended anddeproteinated in 5% metaphosphoric acid by brief soni-cation. After centrifugation at 13,000×g for 5 min, collectthe supernatants for total glutathione measurement. GSHwas determined by adding reaction Mix and GSH reductasesupplied in the kit, following incubation and measurementby ELISA reader at 405 nm for 20 min at 1 min interval. Thetotal amount of GSH was determined by means of a cali-bration curve and normalized to the protein concentration,which was quantified by Bio-Rad protein assay kit.
2.11. Superoxide Dismutase Activity Assay. Superoxide dis-mutase (SOD) activity kit was purchased from Assay Designs(USA). This method was assayed by xanthine oxidase andconversion of WST-1 to WST-1 formazan. Neurons weregrown on 12-well plates for 6 days. After indicated treatment,neurons were then harvested and cytosolic protein wasextracted. SOD activity was determined by adding MasterMix and xanthine supplied in the kit, following incubationand measurement by ELISA reader at 450 nm for 10 min at1 min interval. Protein concentration was quantified by Bio-Rad protein assay kit. Then, calculate SOD activity in cellextracts versus SOD standard curve.
2.12. Western Blotting Analysis. After indicated treatment,neurons were collected and lysed to determine the expressionof cytochrome c, caspase-9, caspase-3 and gp91phox. For thedetection of cytochrome c release, the cells were fractionatedusing a mitochondria/cytosol fractionation kit according tothe manufacturer’s instruction (BioVision, USA). Proteinconcentration was determined with the Bio-Rad proteinassay kit following the manufacturer’s guide. Equal amountsof protein were separated by a polyacrylamide gel and
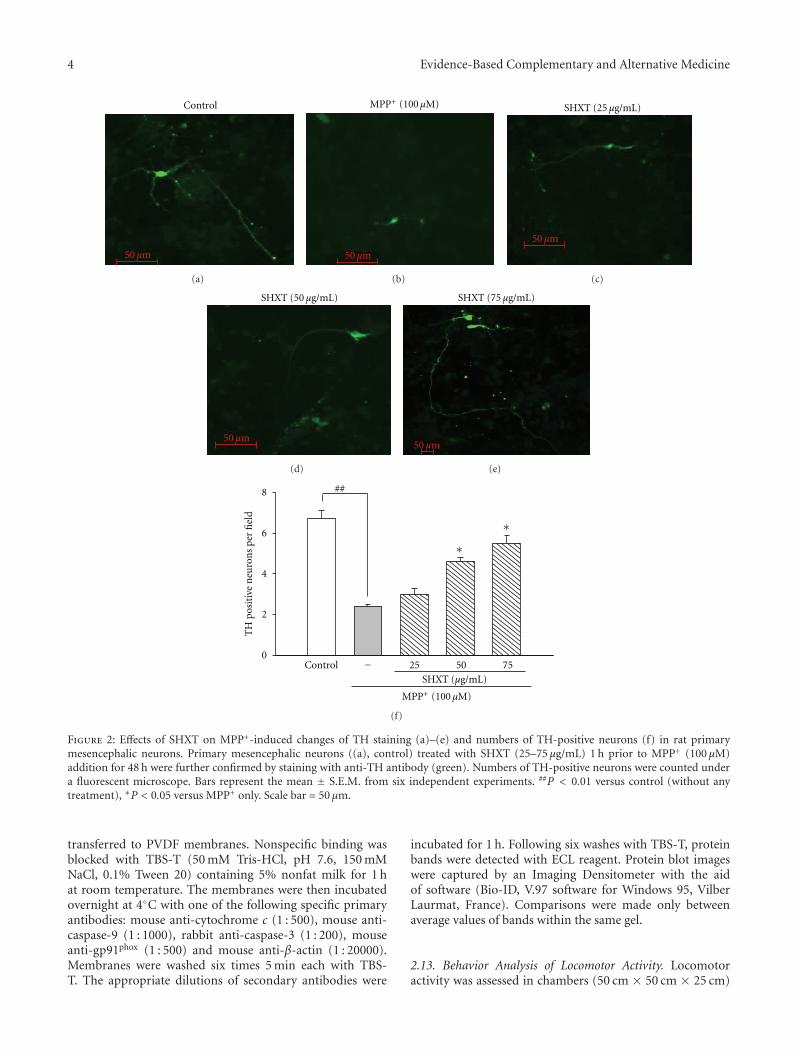
4 Evidence-Based Complementary and Alternative Medicine
Control
50 μm
(a)
MPP+ (100 μM)
50 μm
(b)
SHXT (25 μg/mL)
50 μm
(c)
SHXT (50 μg/mL)
50 μm
(d)
SHXT (75 μg/mL)
50 μm
(e)
TH
pos
itiv
e n
euro
ns
per
fiel
d
0
2
4
6
8
SHXT (μg/mL)
MPP+ (100 μM)
##
∗
∗
Control − 25 50 75
(f)
Figure 2: Effects of SHXT on MPP+-induced changes of TH staining (a)–(e) and numbers of TH-positive neurons (f) in rat primarymesencephalic neurons. Primary mesencephalic neurons ((a), control) treated with SHXT (25–75 μg/mL) 1 h prior to MPP+ (100 μM)addition for 48 h were further confirmed by staining with anti-TH antibody (green). Numbers of TH-positive neurons were counted undera fluorescent microscope. Bars represent the mean ± S.E.M. from six independent experiments. ##P < 0.01 versus control (without anytreatment), ∗P < 0.05 versus MPP+ only. Scale bar = 50 μm.
transferred to PVDF membranes. Nonspecific binding wasblocked with TBS-T (50 mM Tris-HCl, pH 7.6, 150 mMNaCl, 0.1% Tween 20) containing 5% nonfat milk for 1 hat room temperature. The membranes were then incubatedovernight at 4◦C with one of the following specific primaryantibodies: mouse anti-cytochrome c (1 : 500), mouse anti-caspase-9 (1 : 1000), rabbit anti-caspase-3 (1 : 200), mouseanti-gp91phox (1 : 500) and mouse anti-β-actin (1 : 20000).Membranes were washed six times 5 min each with TBS-T. The appropriate dilutions of secondary antibodies were
incubated for 1 h. Following six washes with TBS-T, proteinbands were detected with ECL reagent. Protein blot imageswere captured by an Imaging Densitometer with the aidof software (Bio-ID, V.97 software for Windows 95, VilberLaurmat, France). Comparisons were made only betweenaverage values of bands within the same gel.
2.13. Behavior Analysis of Locomotor Activity. Locomotoractivity was assessed in chambers (50 cm × 50 cm × 25 cm)
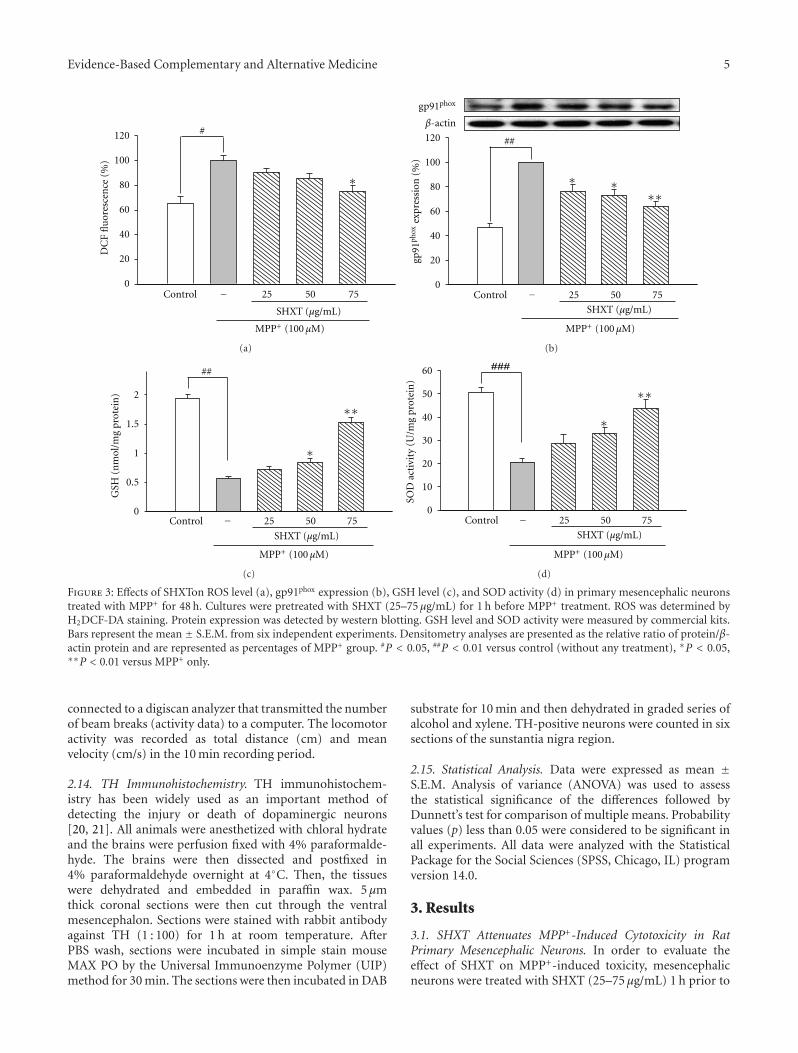
Evidence-Based Complementary and Alternative Medicine 5
#
SHXT (μg/mL)
MPP+ (100 μM)
∗
0
20
40
60
80
100
120
DC
F fl
uor
esce
nce
(%
)
Control − 25 50 75
(a)
SHXT (μg/mL)
MPP+ (100 μM)
0
20
40
60
80
100
120
gp91
phox
expr
essi
on (
%)
gp91phox
β-actin
∗∗∗
##
∗
Control − 25 50 75
(b)
SHXT (μg/mL)
MPP+ (100 μM)
∗∗
##
∗
GSH
(n
mol
/mg
prot
ein
)
0
0.5
1
1.5
2
Control − 25 50 75
(c)
###
∗∗
∗
SHXT (μg/mL)
MPP+ (100 μM)
0
10
20
30
40
50
60
SOD
act
ivit
y (U
/mg
prot
ein
)
Control − 25 50 75
(d)
Figure 3: Effects of SHXTon ROS level (a), gp91phox expression (b), GSH level (c), and SOD activity (d) in primary mesencephalic neuronstreated with MPP+ for 48 h. Cultures were pretreated with SHXT (25–75 μg/mL) for 1 h before MPP+ treatment. ROS was determined byH2DCF-DA staining. Protein expression was detected by western blotting. GSH level and SOD activity were measured by commercial kits.Bars represent the mean ± S.E.M. from six independent experiments. Densitometry analyses are presented as the relative ratio of protein/β-actin protein and are represented as percentages of MPP+ group. #P < 0.05, ##P < 0.01 versus control (without any treatment), ∗P < 0.05,∗∗P < 0.01 versus MPP+ only.
connected to a digiscan analyzer that transmitted the numberof beam breaks (activity data) to a computer. The locomotoractivity was recorded as total distance (cm) and meanvelocity (cm/s) in the 10 min recording period.
2.14. TH Immunohistochemistry. TH immunohistochem-istry has been widely used as an important method ofdetecting the injury or death of dopaminergic neurons[20, 21]. All animals were anesthetized with chloral hydrateand the brains were perfusion fixed with 4% paraformalde-hyde. The brains were then dissected and postfixed in4% paraformaldehyde overnight at 4◦C. Then, the tissueswere dehydrated and embedded in paraffin wax. 5 μmthick coronal sections were then cut through the ventralmesencephalon. Sections were stained with rabbit antibodyagainst TH (1 : 100) for 1 h at room temperature. AfterPBS wash, sections were incubated in simple stain mouseMAX PO by the Universal Immunoenzyme Polymer (UIP)method for 30 min. The sections were then incubated in DAB
substrate for 10 min and then dehydrated in graded series ofalcohol and xylene. TH-positive neurons were counted in sixsections of the sunstantia nigra region.
2.15. Statistical Analysis. Data were expressed as mean ±S.E.M. Analysis of variance (ANOVA) was used to assessthe statistical significance of the differences followed byDunnett’s test for comparison of multiple means. Probabilityvalues (p) less than 0.05 were considered to be significant inall experiments. All data were analyzed with the StatisticalPackage for the Social Sciences (SPSS, Chicago, IL) programversion 14.0.
3. Results
3.1. SHXT Attenuates MPP+-Induced Cytotoxicity in RatPrimary Mesencephalic Neurons. In order to evaluate theeffect of SHXT on MPP+-induced toxicity, mesencephalicneurons were treated with SHXT (25–75 μg/mL) 1 h prior to
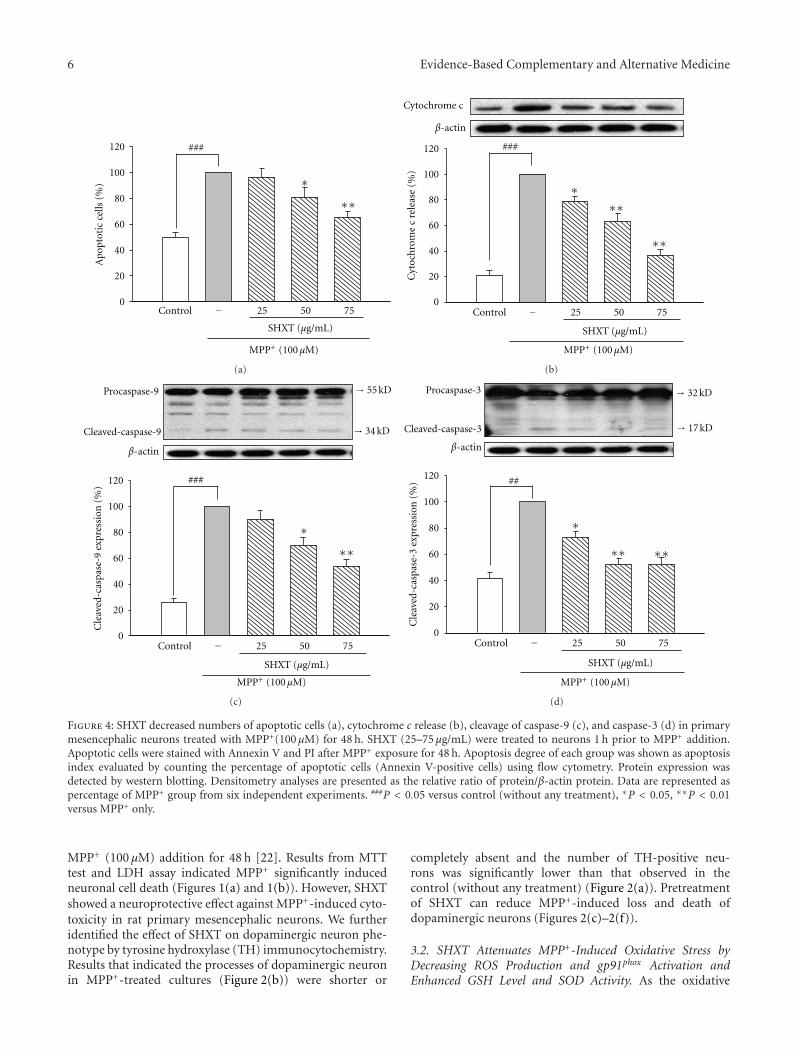
6 Evidence-Based Complementary and Alternative Medicine
Apo
ptot
ic c
ells
(%
)
SHXT (μg/mL)
MPP+ (100 μM)
0
20
40
60
80
100
120 ###
∗∗∗
Control − 25 50 75
(a)
Cyt
och
rom
e c
rele
ase
(%)
SHXT (μg/mL)
MPP+ (100 μM)
0
20
40
60
80
100
120
Cytochrome c
β-actin
###
∗∗∗
∗∗
Control − 25 50 75
(b)
Cle
aved
-cas
pase
-9 e
xpre
ssio
n (
%)
→ 55 kD
→ 34 kD
0
20
40
60
80
100
120 ###
∗∗∗
Cleaved-caspase-9
Procaspase-9
β-actin
SHXT (μg/mL)
MPP+ (100 μM)
Control − 25 50 75
(c)
∗∗
##
Cle
aved
-cas
pase
-3 e
xpre
ssio
n (
%)
→ 32 kD
→ 17 kD
SHXT (μg/mL)
MPP+ (100 μM)
Cleaved-caspase-3
Procaspase-3
β-actin
∗∗
∗
0
20
40
60
80
100
120
Control − 25 50 75
(d)
Figure 4: SHXT decreased numbers of apoptotic cells (a), cytochrome c release (b), cleavage of caspase-9 (c), and caspase-3 (d) in primarymesencephalic neurons treated with MPP+(100 μM) for 48 h. SHXT (25–75 μg/mL) were treated to neurons 1 h prior to MPP+ addition.Apoptotic cells were stained with Annexin V and PI after MPP+ exposure for 48 h. Apoptosis degree of each group was shown as apoptosisindex evaluated by counting the percentage of apoptotic cells (Annexin V-positive cells) using flow cytometry. Protein expression wasdetected by western blotting. Densitometry analyses are presented as the relative ratio of protein/β-actin protein. Data are represented aspercentage of MPP+ group from six independent experiments. ###P < 0.05 versus control (without any treatment), ∗P < 0.05, ∗∗P < 0.01versus MPP+ only.
MPP+ (100 μM) addition for 48 h [22]. Results from MTTtest and LDH assay indicated MPP+ significantly inducedneuronal cell death (Figures 1(a) and 1(b)). However, SHXTshowed a neuroprotective effect against MPP+-induced cyto-toxicity in rat primary mesencephalic neurons. We furtheridentified the effect of SHXT on dopaminergic neuron phe-notype by tyrosine hydroxylase (TH) immunocytochemistry.Results that indicated the processes of dopaminergic neuronin MPP+-treated cultures (Figure 2(b)) were shorter or
completely absent and the number of TH-positive neu-rons was significantly lower than that observed in thecontrol (without any treatment) (Figure 2(a)). Pretreatmentof SHXT can reduce MPP+-induced loss and death ofdopaminergic neurons (Figures 2(c)–2(f)).
3.2. SHXT Attenuates MPP+-Induced Oxidative Stress byDecreasing ROS Production and gp91phox Activation andEnhanced GSH Level and SOD Activity. As the oxidative
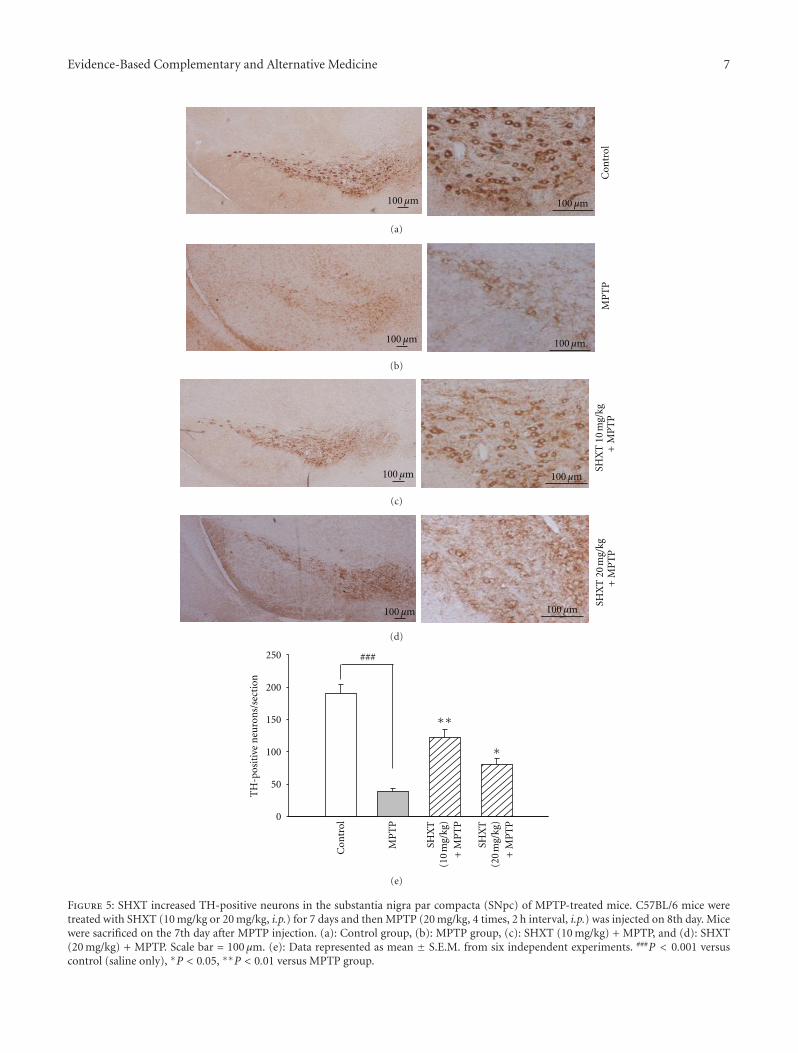
Evidence-Based Complementary and Alternative Medicine 7
100 μm 100 μm
Con
trol
(a)
100 μm 100 μm
MP
TP
(b)
100 μm 100 μm
SHX
T 1
0 m
g/kg
+M
PT
P
(c)
100 μm 100 μmSH
XT
20
mg/
kg+
MP
TP
(d)
TH
-pos
itiv
e n
euro
ns/
sect
ion
0
50
100
150
200
250
Con
trol
MP
TP
SH
XT
(1
0 m
g/kg
)+
MP
TP
SH
XT
(2
0 m
g/kg
)+
MP
TP
###
∗∗
∗
(e)
Figure 5: SHXT increased TH-positive neurons in the substantia nigra par compacta (SNpc) of MPTP-treated mice. C57BL/6 mice weretreated with SHXT (10 mg/kg or 20 mg/kg, i.p.) for 7 days and then MPTP (20 mg/kg, 4 times, 2 h interval, i.p.) was injected on 8th day. Micewere sacrificed on the 7th day after MPTP injection. (a): Control group, (b): MPTP group, (c): SHXT (10 mg/kg) + MPTP, and (d): SHXT(20 mg/kg) + MPTP. Scale bar = 100 μm. (e): Data represented as mean ± S.E.M. from six independent experiments. ###P < 0.001 versuscontrol (saline only), ∗P < 0.05, ∗∗P < 0.01 versus MPTP group.
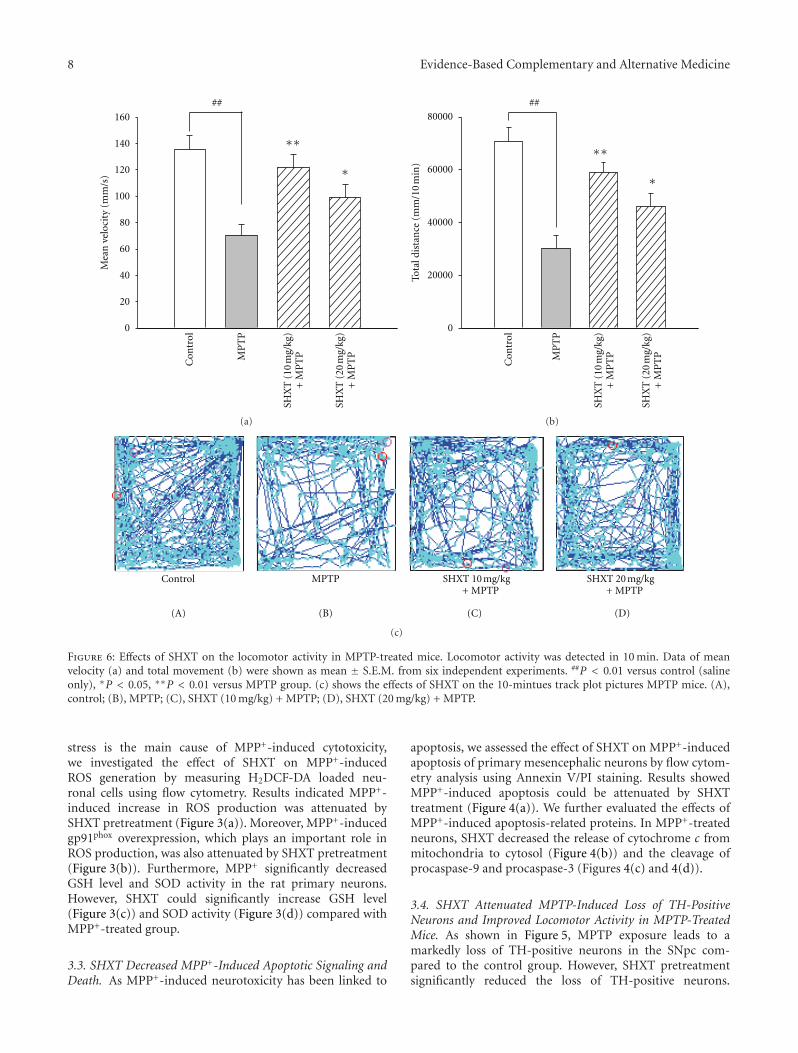
8 Evidence-Based Complementary and Alternative Medicine
Mea
n v
eloc
ity
(mm
/s)
Con
trol
MP
TP
SHX
T (
10 m
g/kg
)+
MP
TP
SHX
T (
20 m
g/kg
)+
MP
TP
∗∗
∗
0
20
40
60
80
100
120
140
160
##
(a)To
tal d
ista
nce
(m
m/1
0 m
in)
Con
trol
MP
TP
SHX
T (
10 m
g/kg
)+
MP
TP
SHX
T (
20 m
g/kg
)+
MP
TP
∗∗
∗
##
0
20000
40000
60000
80000
(b)
Control MPTP SHXT 10 mg/kg+ MPTP
SHXT 20 mg/kg+ MPTP
(A) (B) (C) (D)
(c)
Figure 6: Effects of SHXT on the locomotor activity in MPTP-treated mice. Locomotor activity was detected in 10 min. Data of meanvelocity (a) and total movement (b) were shown as mean ± S.E.M. from six independent experiments. ##P < 0.01 versus control (salineonly), ∗P < 0.05, ∗∗P < 0.01 versus MPTP group. (c) shows the effects of SHXT on the 10-mintues track plot pictures MPTP mice. (A),control; (B), MPTP; (C), SHXT (10 mg/kg) + MPTP; (D), SHXT (20 mg/kg) + MPTP.
stress is the main cause of MPP+-induced cytotoxicity,we investigated the effect of SHXT on MPP+-inducedROS generation by measuring H2DCF-DA loaded neu-ronal cells using flow cytometry. Results indicated MPP+-induced increase in ROS production was attenuated bySHXT pretreatment (Figure 3(a)). Moreover, MPP+-inducedgp91phox overexpression, which plays an important role inROS production, was also attenuated by SHXT pretreatment(Figure 3(b)). Furthermore, MPP+ significantly decreasedGSH level and SOD activity in the rat primary neurons.However, SHXT could significantly increase GSH level(Figure 3(c)) and SOD activity (Figure 3(d)) compared withMPP+-treated group.
3.3. SHXT Decreased MPP+-Induced Apoptotic Signaling andDeath. As MPP+-induced neurotoxicity has been linked to
apoptosis, we assessed the effect of SHXT on MPP+-inducedapoptosis of primary mesencephalic neurons by flow cytom-etry analysis using Annexin V/PI staining. Results showedMPP+-induced apoptosis could be attenuated by SHXTtreatment (Figure 4(a)). We further evaluated the effects ofMPP+-induced apoptosis-related proteins. In MPP+-treatedneurons, SHXT decreased the release of cytochrome c frommitochondria to cytosol (Figure 4(b)) and the cleavage ofprocaspase-9 and procaspase-3 (Figures 4(c) and 4(d)).
3.4. SHXT Attenuated MPTP-Induced Loss of TH-PositiveNeurons and Improved Locomotor Activity in MPTP-TreatedMice. As shown in Figure 5, MPTP exposure leads to amarkedly loss of TH-positive neurons in the SNpc com-pared to the control group. However, SHXT pretreatmentsignificantly reduced the loss of TH-positive neurons.

Evidence-Based Complementary and Alternative Medicine 9
Furthermore, in comparison with the control group, theMPTP-treated mice displayed a significant decrease inlocomotor activity by measuring mean velocity and totalmovement distance (Figures 6(a) and 6(b)), which werereversed by SHXT treatment. Figure 6(c) showed the effectsof SHXT on the 10 min track plot pictures of MPTP mice.
4. Discussion
PD is the second most prevalent age-related neurodegen-erative diseases, primarily affecting people of ages over 55years with physiological manifestations [23]. In the presentstudy, we provide the evidence that the traditional Chinesemedicine SHXT possesses novel protective effects in theMPP+/MPTP models of PD. In vitro study indicates SHXTprotects dopaminergic neurons from the damage induced byMPP+. SHXT attenuates MPP+-induced oxidative stress bydecreasing ROS production and gp91phox expression. SHXTalso enhances antioxidative defense by increasing GSH leveland SOD activity. SHXT also decreases apoptosis-relatedsignal and apoptotic death induced by MPP+. In vivo studyshows SHXT treatment significantly increased TH-positiveneurons in the SNpc and improved motor activity in MPTPmice.
The antioxidant and anti-inflammatory effects of SHXThave been demonstrated in experimental models of variousdiseases [10–14]. Oxidative damage may occur in theparkinsonian brain and is responsible for neurodegenera-tion. Activation of NADPH oxidase is regarded as a majorsource of superoxide in a number of neurodegenerativediseases, including PD. The neurotoxicity of MPTP wasdiminished in mice lacking functional NADPH oxidase[24]. Furthermore, antioxidant status is contributive to theprotection of MPTP-induced dopaminergic neurons loss.For instance, MPTP depletes striatal GSH in mice, and thiseffect may make dopaminergic neurons more susceptible tooxidative stress [25]. SOD is the most important enzyme forthe detoxification of ROS and protection against oxidativestress, and it can thus help prevent neuronal cells fromapoptosis [26]. Our previous study revealed SHXT possessneuroprotective effect against activated microglia- and 6-OHDA-induced toxicity [14]. The present results furthershowed SHXT ameliorated MPP+-induced oxidative stressby inhibiting ROS production and gp91phox expression. Italso upregulated antioxidant protective systems by increasingboth levels of GSH and SOD. Moreover, ROS productioncontributes to the apoptotic processes found in PD [27]. Therelease of cytochrome c from mitochondria to cytosol couldform apoptosome with apoptosis-activating factor (Apaf-1) and procaspase-9, leading to the activation of caspase-9 and caspase-3 [28]. Caspase-3 has been demonstratedto participate in MPP+-induced apoptosis and is regulatedby mitochondria dysfunction-mediated ROS overproduction[29]. The present results indicated that SHXT not onlyattenuated MPP+-induced oxidative stress, but it also down-regulated MPP+-induced apoptotic death and molecularevents including cytochrome c release and caspase cascadeactivation.
The most debilitating symptom of PD is the loss ofmotor control. The MPTP mouse model caused patho-physiology similar to patients with PD [30]. The ability toreflect behavior abnormalities of PD is one of the mostuseful characteristics of this animal model. According toour results, mice indeed display locomotor abnormalities7 days after MPTP injection (20 mg/kg, 4 times, 2 h inter-val). SHXT pretreatment improved locomotor activity inMPTP mice. Moreover, TH is the rate-limiting enzyme inthe synthesis of catecholamine neurotransmitters such asdopamine, epinephrine, and norepinephrine. TH activityis progressively decreased following the loss of dopamineneurons in the substantia nigra in the patients with PD[20, 21]. Our results showed that SHXT significantly reducedthe loss of TH-positive neurons not only in MPP+-treatedmesencephalic neurons in vitro but also in substantia nigraof MPTP mice in vivo. These results suggested that SHXTcould protect dopaminergic neurons against MPTP-inducedtoxicity.
In conclusion, SHXT provides novel neuroprotectiveeffects, at least in part, via enhancing antioxidative defense,attenuating oxidative stress and decreasing apoptotic deathin MPP+-treated mesencephalic neuron and in MPTP-mouse model of PD. Therefore, SHXT might be a potentiallyalternative and complementary medicine used in the treat-ment of PD.
Acknowledgments
The authors would like to thank Mr. Lue Sheng-I atDepartment of Physiology and Master’s Program, School ofMedicine, Kaohsiung Medical University, Kaohsiung, Taiwanfor his technological assistance. This work was supported bythe National Science Council of Taiwan [NSC 96–2320-B-037-039-MY3 and NSC 99–2320-B- 037-023-MY3].
References
[1] H. Li, J. Hu, L. Ma et al., “Comprehensive study of baicalindown-regulating NOD2 receptor expression of neurons withoxygen-glucose deprivation in vitro and cerebral ischemia-reperfusion in vivo,” European Journal of Pharmacology, vol.649, no. 1–3, pp. 92–99, 2010.
[2] X. K. Tu, W. Z. Yang, S. S. Shi, C. H. Wang, and C. M.Chen, “Neuroprotective effect of baicalin in a rat model ofpermanent focal cerebral ischemia,” Neurochemical Research,vol. 34, no. 9, pp. 1626–1634, 2009.
[3] X. Mu, G. He, Y. Cheng, X. Li, B. Xu, and G. Du, “Baicaleinexerts neuroprotective effects in 6-hydroxydopamine-inducedexperimental parkinsonism in vivo and in vitro,” Pharmacol-ogy Biochemistry and Behavior, vol. 92, no. 4, pp. 642–648,2009.
[4] C. Liu, J. Wu, K. Xu et al., “Neuroprotection by baicalein inischemic brain injury involves PTEN/AKT pathway,” Journalof Neurochemistry, vol. 112, no. 6, pp. 1500–1512, 2010.
[5] S. F. Chen, C. W. Hsu, W. H. Huang, and J. Y. Wang, “Post-injury baicalein improves histological and functional out-comes and reduces inflammatory cytokines after experimentaltraumatic brain injury,” British Journal of Pharmacology, vol.155, no. 8, pp. 1279–1296, 2008.

10 Evidence-Based Complementary and Alternative Medicine
[6] F. Q. Li, T. Wang, Z. Pei, B. Liu, and J. S. Hong, “Inhibition ofmicroglial activation by the herbal flavonoid baicalein atten-uates inflammation-mediated degeneration of dopaminergicneurons,” Journal of Neural Transmission, vol. 112, no. 3, pp.331–347, 2005.
[7] H. Lee, Y. O. Kim, H. Kim et al., “Flavonoid wogonin frommedicinal herb is neuroprotective by inhibiting inflammatoryactivation of microglia,” The FASEB Journal, vol. 17, no. 13,pp. 1943–1944, 2003.
[8] J. W. Gu, H. Hasuo, M. Takeya, and T. Akasu, “Effects ofemodin on synaptic transmission in rat hippocampal CA1pyramidal neurons in vitro,” Neuropharmacology, vol. 49, no.1, pp. 103–111, 2005.
[9] T. Liu, H. Jin, Q. R. Sun, J. H. Xu, and H. T. Hu, “Neuro-protective effects of emodin in rat cortical neurons against β-amyloid-induced neurotoxicity,” Brain Research, vol. 1347, no.C, pp. 149–160, 2010.
[10] Y. C. Lo, Y. L. Lin, K. L. Yu et al., “San-Huang-Xie-Xin-Tangattenuates inflammatory responses in lipopolysaccharide-exposed rat lungs,” Journal of Ethnopharmacology, vol. 101, no.1–3, pp. 68–74, 2005.
[11] Y. C. Lo, P. L. Tsai, Y. B. Huang et al., “San-Huang-Xie-Xin-Tang reduces lipopolysaccharides-induced hypotension andinflammatory mediators,” Journal of Ethnopharmacology, vol.96, no. 1-2, pp. 99–106, 2005.
[12] Y. T. Shih, D. C. Wu, C. M. Liu, Y. C. Yang, I. J. Chen, and Y. C.Lo, “San-Huang-Xie-Xin-Tang inhibits Helicobacter pylori-induced inflammation in human gastric epithelial AGS cells,”Journal of Ethnopharmacology, vol. 112, no. 3, pp. 537–544,2007.
[13] H. H. Tsai, I. J. Chen, and Y. C. Lo, “Effects of San-Huang-Xie-Xin-Tang on U46619-induced increase in pulmonary arterialblood pressure,” Journal of Ethnopharmacology, vol. 117, no. 3,pp. 457–462, 2008.
[14] Y.-T. Shih, I.-J. Chen, Y.-C. Wu, and Y.-C. Lo, “San-Huang-Xie-Xin-Tang protects against activated microglia- and 6-OHDA-induced toxicity in neuronal SH-SY5Y cells,” Evidence-Based Complementary and Alternative Medicine, vol. 2011,Article ID 429384, 11 pages, 2011.
[15] K. J. Barnham, C. L. Masters, and A. I. Bush, “Neurodegen-erative diseases and oxidatives stress,” Nature Reviews DrugDiscovery, vol. 3, no. 3, pp. 205–214, 2004.
[16] J. W. Langston and I. Irwin, “MPTP: current concepts andcontroversies,” Clinical Neuropharmacology, vol. 9, no. 6, pp.485–507, 1986.
[17] C. Mancuso, G. Scapagnini, D. Curro et al., “Mitochon-drial dysfunction, free radical generation and cellular stressresponse in neurodegenerative disorders,” Frontiers in Bio-science, vol. 12, no. 3, pp. 1107–1123, 2007.
[18] K. F. Tipton and T. P. Singer, “Advances in our understandingof the mechanisms of the neurotoxicity of MPTP and relatedcompounds,” Journal of Neurochemistry, vol. 61, no. 4, pp.1191–1206, 1993.
[19] M. Alcaraz-Zubeldia, P. Rojas, C. Boll, and C. Rıos, “Neuro-protective effect of acute and chronic administration of copper(II) sulfate against MPP,” Neurochemical Research, vol. 26, no.1, pp. 59–64, 2001.
[20] J. Haavik and K. Toska, “Tyrosine hydroxylase and Parkinson’sdisease,” Molecular Neurobiology, vol. 16, no. 3, pp. 285–309,1998.
[21] Y. Oiwa, R. Yoshimura, K. Nakai, and T. Itakura, “Dopamin-ergic neuroprotection and regeneration by neurturin assessed
by using behavioral, biochemical and histochemical measure-ments in a model of progressive Parkinson’s disease,” BrainResearch, vol. 947, no. 2, pp. 271–283, 2002.
[22] Y.-Y. Li, J.-H. Lu, Q. Li, Y.-Y. Zhao, and X.-P. Pu, “Pediculario-side A from Buddleia lindleyana inhibits cell death inducedby 1-methyl-4-phenylpyridinium ions (MPP+) in primarycultures of rat mesencephalic neurons,” European Journal ofPharmacology, vol. 579, no. 1–3, pp. 134–140, 2008.
[23] R. L. Miller, M. James-Kracke, G. Y. Sun, and A. Y. Sun,“Oxidative and inflammatory pathways in parkinson’s dis-ease,” Neurochemical Research, vol. 34, no. 1, pp. 55–65, 2009.
[24] D. C. Wu, P. Teismann, K. Tieu et al., “NADPH oxidasemediates oxidative stress in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson’s disease,” Proceedingsof the National Academy of Sciences of the United States ofAmerica, vol. 100, no. 10, pp. 6145–6150, 2003.
[25] K. Aoyama, N. Matsumura, M. Watabe, and T. Nakaki,“Oxidative stress on EAAC1 is involved in MPTP-inducedglutathione depletion and motor dysfunction,” EuropeanJournal of Neuroscience, vol. 27, no. 1, pp. 20–30, 2008.
[26] L. J. S. Greenlund, T. L. Deckwerth, and E. M. JohnsonJr., “Superoxide dismutase delays neuronal apoptosis: a rolefor reactive oxygen species in programmed neuronal death,”Neuron, vol. 14, no. 2, pp. 303–315, 1995.
[27] D. Di Monte, M. S. Sandy, G. Ekstrom, and M. T. Smith,“Comparative studies on the mechanisms of paraquat and1-methyl-4-phenylpyridine (MPP+) cytotoxicity,” Biochemicaland Biophysical Research Communications, vol. 137, no. 1, pp.303–309, 1986.
[28] N. A. Thornberry and Y. Lazebnik, “Caspases: enemieswithin,” Science, vol. 281, no. 5381, pp. 1312–1316, 1998.
[29] J. I. Kakimura, Y. Kitamura, K. Takata, Y. Kohno, Y. Nomura,and T. Taniguchi, “Release and aggregation of cytochrome cand α-synuclein are inhibited by the antiparkinsonian drugs,talipexole and pramipexole,” European Journal of Pharmacol-ogy, vol. 417, no. 1-2, pp. 59–67, 2001.
[30] R. Camicioli, S. J. Grossmann, P. S. Spencer, K. Hudnell, andW. Kent Anger, “Discriminating mild parkinsonism: methodsfor epidemiological research,” Movement Disorders, vol. 16, no.1, pp. 33–40, 2001.

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2012, Article ID 928643, 6 pagesdoi:10.1155/2012/928643
Research Article
Effect of Quercetin inthe 1-Methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-InducedMouse Model of Parkinson’s Disease
Chuanfeng Lv,1 Tie Hong,1 Zhen Yang,2 Yu Zhang,1 Lu Wang,3 Man Dong,1 Jing Zhao,1
Jiaye Mu,3 and Yixiao Meng3
1 Department of Pharmacology, School of Pharmacy, Jilin University, Changchun 130021, China2 Institute of Changbai Mountain Natural Resources, Jilin Academy of Chinese Medicine Sciences, Changchun 130012, China3 College of Pharmacy, Yanbian University, Yanji 133002, China
Correspondence should be addressed to Tie Hong, [email protected]
Received 17 September 2011; Revised 5 November 2011; Accepted 7 November 2011
Academic Editor: Mahmud Tareq Hassan Khan
Copyright © 2012 Chuanfeng Lv et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
In this paper, the protective effect of the bioflavonoid quercetin on behaviors, antioxidases, and neurotransmitters in 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-(MPTP-) induced Parkinson’s disease (PD) was investigated. Quercetin treatment (50 mg/kg,100 mg/kg and 200 mg/kg body weight) was orally administered for 14 consecutive days. The results show that quercetin treatmentmarkedly improves the motor balance and coordination of MPTP-treated mice. Significant increases were observed in the activitiesof glutathione peroxidase (GPx), superoxide dismutase (SOD), and Na+, K+-ATPase, AchE, the content of dopamine (DA) in thequercetin plus MPTP groups compared to those in the MPTP group. Significant reduction the 4-hydroxy-2-nonenal (4-HNE)immunoreactivity in striatum of brains was observed in the quercetin plus MPTP groups in comparison to the MPTP group.Taken together, we propose that quercetin has shown antiparkinsonian properties in our studies. More work is needed to exploredetailed mechanisms of action.
1. Introduction
Parkinson’s disease (PD), which is a type of regressive diseaseof the central nervous system, is the second most commondisorder of the aging brain after Alzheimer’s disease. Theclinical manifestations are static tremors, myotonia, bradyki-nesia, and poor balance. The main drug families that areuseful for treating motor symptoms are levodopa, dopamineagonists, and MAO-B inhibitors [1].
PD models are divided into two broad categories: geneticand toxic [2, 3]. Most of the data that addresses the effectsof oxidative stress have been generated using toxic modelssuch as those produced by 1-methyl-4-phenyl-1, 2, 3, 6-hydroxydopamine (MPTP), which damages the nigrostriataldopaminergic system.
The antioxidative activity of quercetin, which is abioflavonoid, has been studied [4]. Recent studies haveshown that quercetin crosses the blood-brain barrier (BBB)[5] and protects HT-22 cells by prohibiting the formation
of reactive oxygen species (ROS), which is generated by glu-tamic acid-induced oxidation toxicity and lipid peroxidation[6].
In the present study, we investigated the neuropro-tective effects of quercetin to modify glutathione peroxi-dase (GPx), superoxide dismutase (SOD), Na+, K+-ATPase,immunoreactivity of 4-hydroxy-2-nonenal (4-HNE), acetyl-cholinesterase (AChE) activities, and the level of dopamine(DA) in the brain tissue in the MPTP-induced mouse modelof Parkinson’s disease.
2. Materials and Methods
2.1. Animals. Specific pathogen-free adult male C57BL/6mice (25 ± 2 g, body weight; 2–2.5 months old) werehoused in standard cages with wood shavings. Ten animalsper cage were kept in a room with a carefully controlledambient temperature (25◦C) and artificially illumination
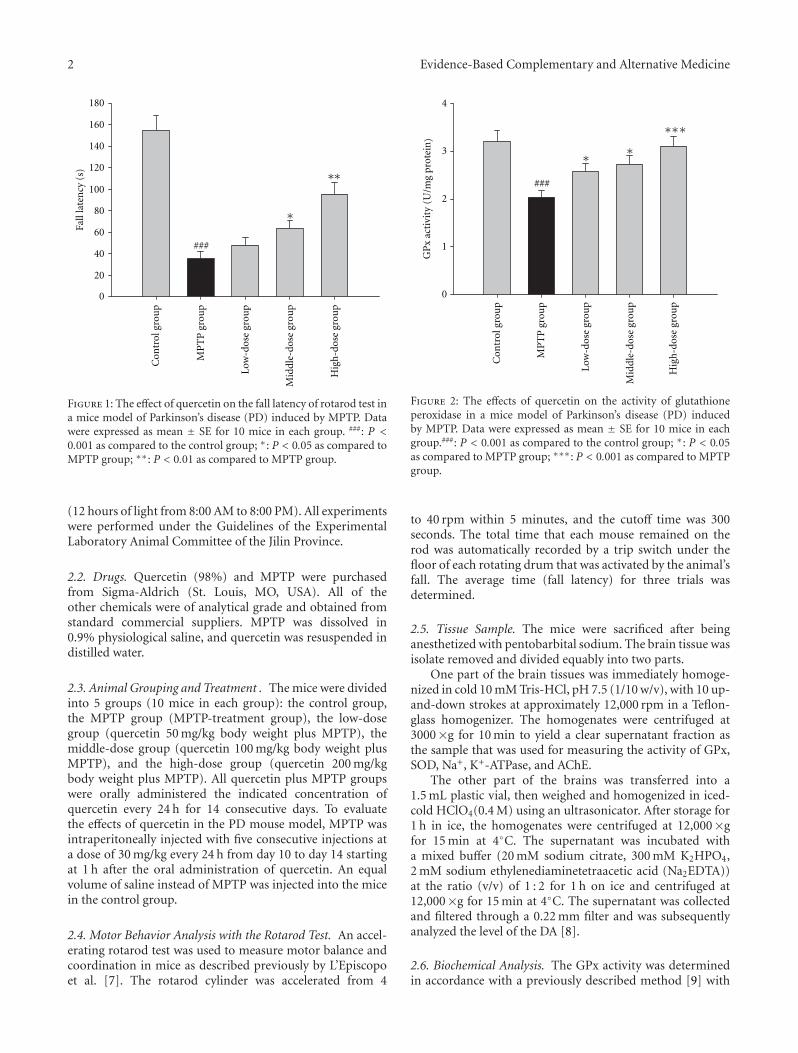
2 Evidence-Based Complementary and Alternative Medicine
∗
###
0
20
40
60
80
100
120
140
160
180
Fall
late
ncy
(s)
Con
trol
gro
up
MP
TP
gro
up
Low
-dos
e gr
oup
Mid
dle-
dose
gro
up
Hig
h-d
ose
grou
p
∗∗
Figure 1: The effect of quercetin on the fall latency of rotarod test ina mice model of Parkinson’s disease (PD) induced by MPTP. Datawere expressed as mean ± SE for 10 mice in each group. ###: P <0.001 as compared to the control group; ∗: P < 0.05 as compared toMPTP group; ∗∗: P < 0.01 as compared to MPTP group.
(12 hours of light from 8:00 AM to 8:00 PM). All experimentswere performed under the Guidelines of the ExperimentalLaboratory Animal Committee of the Jilin Province.
2.2. Drugs. Quercetin (98%) and MPTP were purchasedfrom Sigma-Aldrich (St. Louis, MO, USA). All of theother chemicals were of analytical grade and obtained fromstandard commercial suppliers. MPTP was dissolved in0.9% physiological saline, and quercetin was resuspended indistilled water.
2.3. Animal Grouping and Treatment . The mice were dividedinto 5 groups (10 mice in each group): the control group,the MPTP group (MPTP-treatment group), the low-dosegroup (quercetin 50 mg/kg body weight plus MPTP), themiddle-dose group (quercetin 100 mg/kg body weight plusMPTP), and the high-dose group (quercetin 200 mg/kgbody weight plus MPTP). All quercetin plus MPTP groupswere orally administered the indicated concentration ofquercetin every 24 h for 14 consecutive days. To evaluatethe effects of quercetin in the PD mouse model, MPTP wasintraperitoneally injected with five consecutive injections ata dose of 30 mg/kg every 24 h from day 10 to day 14 startingat 1 h after the oral administration of quercetin. An equalvolume of saline instead of MPTP was injected into the micein the control group.
2.4. Motor Behavior Analysis with the Rotarod Test. An accel-erating rotarod test was used to measure motor balance andcoordination in mice as described previously by L’Episcopoet al. [7]. The rotarod cylinder was accelerated from 4
0
1
2
3
4
Con
trol
gro
up
MP
TP
gro
up
Low
-dos
e gr
oup
Mid
dle-
dose
gro
up
Hig
h-d
ose
grou
p
###
∗
∗∗∗∗
GP
x ac
tivi
ty(U
/mg
prot
ein )
Figure 2: The effects of quercetin on the activity of glutathioneperoxidase in a mice model of Parkinson’s disease (PD) inducedby MPTP. Data were expressed as mean ± SE for 10 mice in eachgroup.###: P < 0.001 as compared to the control group; ∗: P < 0.05as compared to MPTP group; ∗∗∗: P < 0.001 as compared to MPTPgroup.
to 40 rpm within 5 minutes, and the cutoff time was 300seconds. The total time that each mouse remained on therod was automatically recorded by a trip switch under thefloor of each rotating drum that was activated by the animal’sfall. The average time (fall latency) for three trials wasdetermined.
2.5. Tissue Sample. The mice were sacrificed after beinganesthetized with pentobarbital sodium. The brain tissue wasisolate removed and divided equably into two parts.
One part of the brain tissues was immediately homoge-nized in cold 10 mM Tris-HCl, pH 7.5 (1/10 w/v), with 10 up-and-down strokes at approximately 12,000 rpm in a Teflon-glass homogenizer. The homogenates were centrifuged at3000×g for 10 min to yield a clear supernatant fraction asthe sample that was used for measuring the activity of GPx,SOD, Na+, K+-ATPase, and AChE.
The other part of the brains was transferred into a1.5 mL plastic vial, then weighed and homogenized in iced-cold HClO4(0.4 M) using an ultrasonicator. After storage for1 h in ice, the homogenates were centrifuged at 12,000×gfor 15 min at 4◦C. The supernatant was incubated witha mixed buffer (20 mM sodium citrate, 300 mM K2HPO4,2 mM sodium ethylenediaminetetraacetic acid (Na2EDTA))at the ratio (v/v) of 1 : 2 for 1 h on ice and centrifuged at12,000×g for 15 min at 4◦C. The supernatant was collectedand filtered through a 0.22 mm filter and was subsequentlyanalyzed the level of the DA [8].
2.6. Biochemical Analysis. The GPx activity was determinedin accordance with a previously described method [9] with
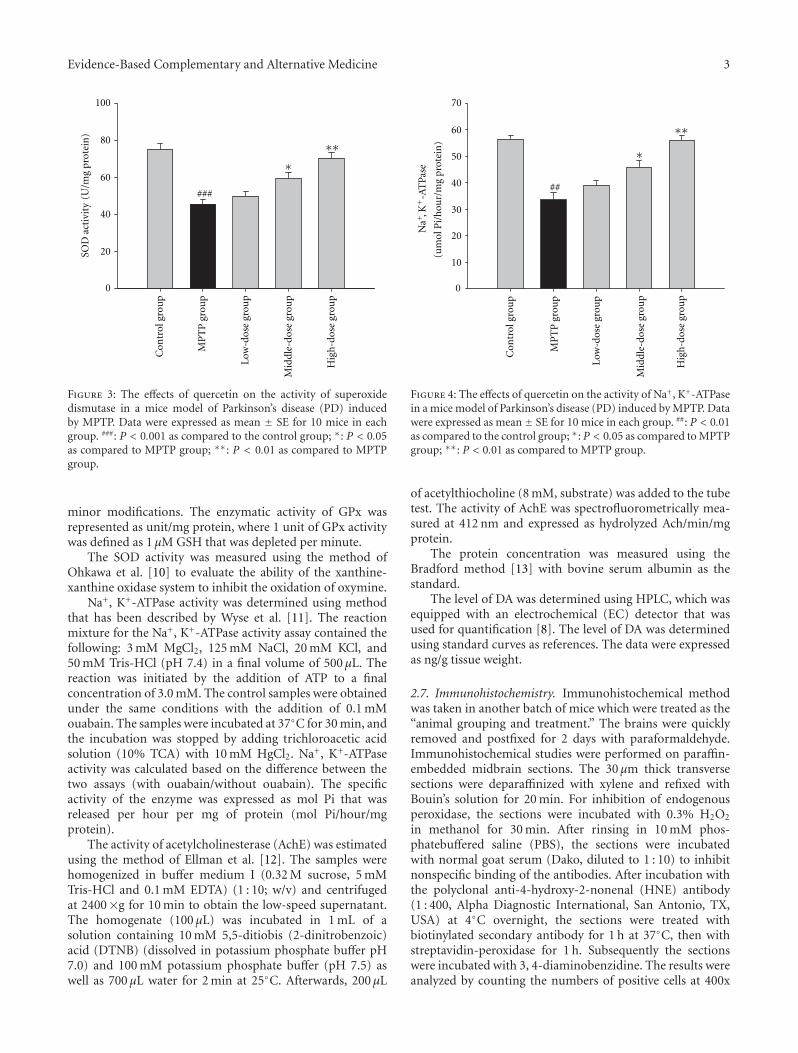
Evidence-Based Complementary and Alternative Medicine 3
Con
trol
gro
up
MP
TP
gro
up
Low
-dos
e gr
oup
Mid
dle-
dose
gro
up
Hig
h-d
ose
grou
p
SOD
act
ivit
y(U
/mg
prot
ein
)
0
20
40
60
80
100
###
∗∗∗
Figure 3: The effects of quercetin on the activity of superoxidedismutase in a mice model of Parkinson’s disease (PD) inducedby MPTP. Data were expressed as mean ± SE for 10 mice in eachgroup. ###: P < 0.001 as compared to the control group; ∗: P < 0.05as compared to MPTP group; ∗∗: P < 0.01 as compared to MPTPgroup.
minor modifications. The enzymatic activity of GPx wasrepresented as unit/mg protein, where 1 unit of GPx activitywas defined as 1 µM GSH that was depleted per minute.
The SOD activity was measured using the method ofOhkawa et al. [10] to evaluate the ability of the xanthine-xanthine oxidase system to inhibit the oxidation of oxymine.
Na+, K+-ATPase activity was determined using methodthat has been described by Wyse et al. [11]. The reactionmixture for the Na+, K+-ATPase activity assay contained thefollowing: 3 mM MgCl2, 125 mM NaCl, 20 mM KCl, and50 mM Tris-HCl (pH 7.4) in a final volume of 500 µL. Thereaction was initiated by the addition of ATP to a finalconcentration of 3.0 mM. The control samples were obtainedunder the same conditions with the addition of 0.1 mMouabain. The samples were incubated at 37◦C for 30 min, andthe incubation was stopped by adding trichloroacetic acidsolution (10% TCA) with 10 mM HgCl2. Na+, K+-ATPaseactivity was calculated based on the difference between thetwo assays (with ouabain/without ouabain). The specificactivity of the enzyme was expressed as mol Pi that wasreleased per hour per mg of protein (mol Pi/hour/mgprotein).
The activity of acetylcholinesterase (AchE) was estimatedusing the method of Ellman et al. [12]. The samples werehomogenized in buffer medium I (0.32 M sucrose, 5 mMTris-HCl and 0.1 mM EDTA) (1 : 10; w/v) and centrifugedat 2400×g for 10 min to obtain the low-speed supernatant.The homogenate (100 µL) was incubated in 1 mL of asolution containing 10 mM 5,5-ditiobis (2-dinitrobenzoic)acid (DTNB) (dissolved in potassium phosphate buffer pH7.0) and 100 mM potassium phosphate buffer (pH 7.5) aswell as 700 µL water for 2 min at 25◦C. Afterwards, 200 µL
0
10
20
30
40
50
60
70
##
Na+ , K
+-A
TPa
se
(um
ol P
i/h
our/
mg
prot
ein
)
Con
trol
gro
up
MP
TP
gro
up
Low
-dos
e gr
oup
Mid
dle-
dose
gro
up
Hig
h-d
ose
grou
p
∗
∗∗
Figure 4: The effects of quercetin on the activity of Na+, K+-ATPasein a mice model of Parkinson’s disease (PD) induced by MPTP. Datawere expressed as mean ± SE for 10 mice in each group. ##: P < 0.01as compared to the control group; ∗: P < 0.05 as compared to MPTPgroup; ∗∗: P < 0.01 as compared to MPTP group.
of acetylthiocholine (8 mM, substrate) was added to the tubetest. The activity of AchE was spectrofluorometrically mea-sured at 412 nm and expressed as hydrolyzed Ach/min/mgprotein.
The protein concentration was measured using theBradford method [13] with bovine serum albumin as thestandard.
The level of DA was determined using HPLC, which wasequipped with an electrochemical (EC) detector that wasused for quantification [8]. The level of DA was determinedusing standard curves as references. The data were expressedas ng/g tissue weight.
2.7. Immunohistochemistry. Immunohistochemical methodwas taken in another batch of mice which were treated as the“animal grouping and treatment.” The brains were quicklyremoved and postfixed for 2 days with paraformaldehyde.Immunohistochemical studies were performed on paraffin-embedded midbrain sections. The 30 µm thick transversesections were deparaffinized with xylene and refixed withBouin’s solution for 20 min. For inhibition of endogenousperoxidase, the sections were incubated with 0.3% H2O2
in methanol for 30 min. After rinsing in 10 mM phos-phatebuffered saline (PBS), the sections were incubatedwith normal goat serum (Dako, diluted to 1 : 10) to inhibitnonspecific binding of the antibodies. After incubation withthe polyclonal anti-4-hydroxy-2-nonenal (HNE) antibody(1 : 400, Alpha Diagnostic International, San Antonio, TX,USA) at 4◦C overnight, the sections were treated withbiotinylated secondary antibody for 1 h at 37◦C, then withstreptavidin-peroxidase for 1 h. Subsequently the sectionswere incubated with 3, 4-diaminobenzidine. The results wereanalyzed by counting the numbers of positive cells at 400x
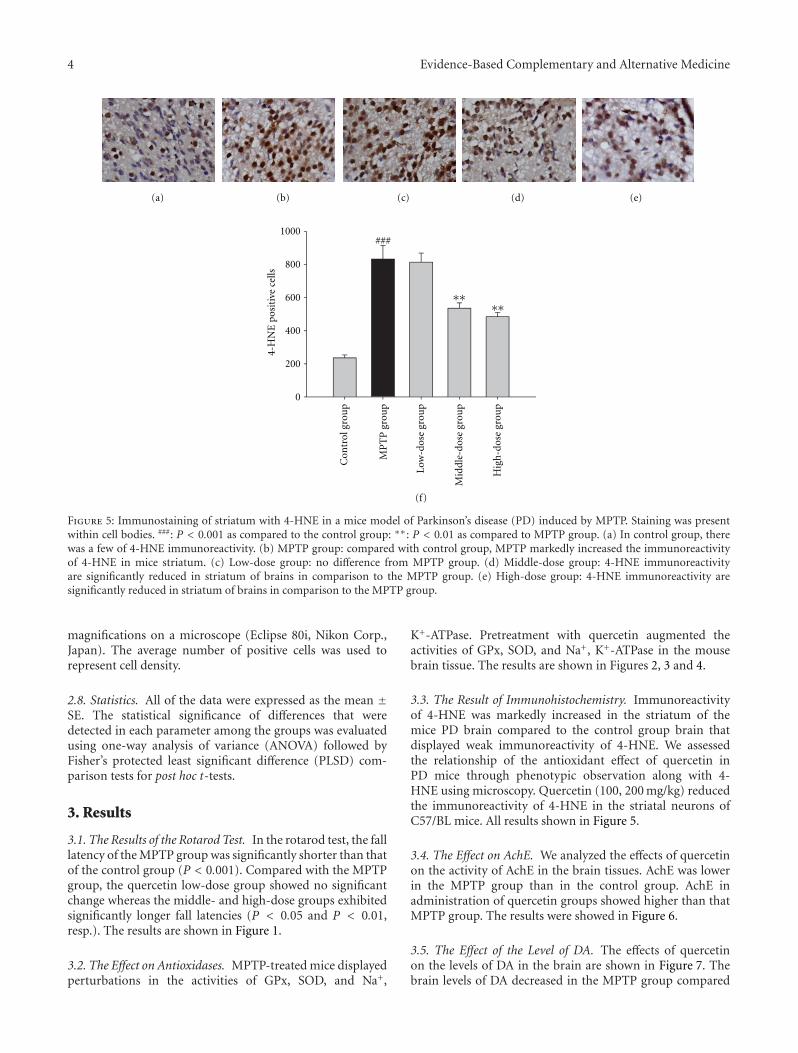
4 Evidence-Based Complementary and Alternative Medicine
4-H
NE
pos
itiv
e ce
lls
0
200
400
600
800
1000###
Con
trol
gro
up
MP
TP
gro
up
Low
-dos
e gr
oup
Mid
dle-
dose
gro
up
Hig
h-d
ose
grou
p
∗∗∗∗
(a) (b) (c) (d) (e)
(f)
Figure 5: Immunostaining of striatum with 4-HNE in a mice model of Parkinson’s disease (PD) induced by MPTP. Staining was presentwithin cell bodies. ###: P < 0.001 as compared to the control group: ∗∗: P < 0.01 as compared to MPTP group. (a) In control group, therewas a few of 4-HNE immunoreactivity. (b) MPTP group: compared with control group, MPTP markedly increased the immunoreactivityof 4-HNE in mice striatum. (c) Low-dose group: no difference from MPTP group. (d) Middle-dose group: 4-HNE immunoreactivityare significantly reduced in striatum of brains in comparison to the MPTP group. (e) High-dose group: 4-HNE immunoreactivity aresignificantly reduced in striatum of brains in comparison to the MPTP group.
magnifications on a microscope (Eclipse 80i, Nikon Corp.,Japan). The average number of positive cells was used torepresent cell density.
2.8. Statistics. All of the data were expressed as the mean ±SE. The statistical significance of differences that weredetected in each parameter among the groups was evaluatedusing one-way analysis of variance (ANOVA) followed byFisher’s protected least significant difference (PLSD) com-parison tests for post hoc t-tests.
3. Results
3.1. The Results of the Rotarod Test. In the rotarod test, the falllatency of the MPTP group was significantly shorter than thatof the control group (P < 0.001). Compared with the MPTPgroup, the quercetin low-dose group showed no significantchange whereas the middle- and high-dose groups exhibitedsignificantly longer fall latencies (P < 0.05 and P < 0.01,resp.). The results are shown in Figure 1.
3.2. The Effect on Antioxidases. MPTP-treated mice displayedperturbations in the activities of GPx, SOD, and Na+,
K+-ATPase. Pretreatment with quercetin augmented theactivities of GPx, SOD, and Na+, K+-ATPase in the mousebrain tissue. The results are shown in Figures 2, 3 and 4.
3.3. The Result of Immunohistochemistry. Immunoreactivityof 4-HNE was markedly increased in the striatum of themice PD brain compared to the control group brain thatdisplayed weak immunoreactivity of 4-HNE. We assessedthe relationship of the antioxidant effect of quercetin inPD mice through phenotypic observation along with 4-HNE using microscopy. Quercetin (100, 200 mg/kg) reducedthe immunoreactivity of 4-HNE in the striatal neurons ofC57/BL mice. All results shown in Figure 5.
3.4. The Effect on AchE. We analyzed the effects of quercetinon the activity of AchE in the brain tissues. AchE was lowerin the MPTP group than in the control group. AchE inadministration of quercetin groups showed higher than thatMPTP group. The results were showed in Figure 6.
3.5. The Effect of the Level of DA. The effects of quercetinon the levels of DA in the brain are shown in Figure 7. Thebrain levels of DA decreased in the MPTP group compared
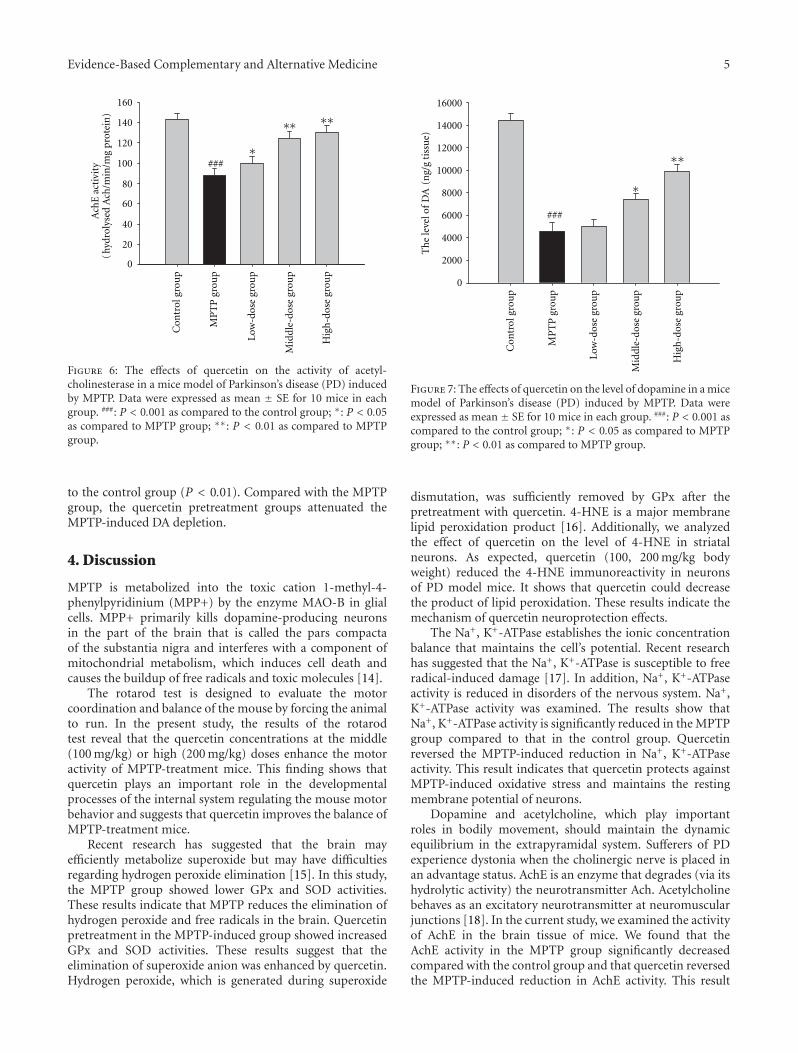
Evidence-Based Complementary and Alternative Medicine 5
0
20
40
60
80
100
120
140
160
###
Ach
E a
ctiv
ity
hydr
olys
ed A
ch/m
in/m
g pr
otei
n)
(
Con
trol
gro
up
MP
TP
gro
up
Low
-dos
e gr
oup
Mid
dle-
dose
gro
up
Hig
h-d
ose
grou
p
∗
∗∗ ∗∗
Figure 6: The effects of quercetin on the activity of acetyl-cholinesterase in a mice model of Parkinson’s disease (PD) inducedby MPTP. Data were expressed as mean ± SE for 10 mice in eachgroup. ###: P < 0.001 as compared to the control group; ∗: P < 0.05as compared to MPTP group; ∗∗: P < 0.01 as compared to MPTPgroup.
to the control group (P < 0.01). Compared with the MPTPgroup, the quercetin pretreatment groups attenuated theMPTP-induced DA depletion.
4. Discussion
MPTP is metabolized into the toxic cation 1-methyl-4-phenylpyridinium (MPP+) by the enzyme MAO-B in glialcells. MPP+ primarily kills dopamine-producing neuronsin the part of the brain that is called the pars compactaof the substantia nigra and interferes with a component ofmitochondrial metabolism, which induces cell death andcauses the buildup of free radicals and toxic molecules [14].
The rotarod test is designed to evaluate the motorcoordination and balance of the mouse by forcing the animalto run. In the present study, the results of the rotarodtest reveal that the quercetin concentrations at the middle(100 mg/kg) or high (200 mg/kg) doses enhance the motoractivity of MPTP-treatment mice. This finding shows thatquercetin plays an important role in the developmentalprocesses of the internal system regulating the mouse motorbehavior and suggests that quercetin improves the balance ofMPTP-treatment mice.
Recent research has suggested that the brain mayefficiently metabolize superoxide but may have difficultiesregarding hydrogen peroxide elimination [15]. In this study,the MPTP group showed lower GPx and SOD activities.These results indicate that MPTP reduces the elimination ofhydrogen peroxide and free radicals in the brain. Quercetinpretreatment in the MPTP-induced group showed increasedGPx and SOD activities. These results suggest that theelimination of superoxide anion was enhanced by quercetin.Hydrogen peroxide, which is generated during superoxide
Th
e le
vel o
f D
A(n
g/g
tiss
ue)
0
2000
4000
6000
8000
10000
12000
14000
16000
###
Con
trol
gro
up
MP
TP
gro
up
Low
-dos
e gr
oup
Mid
dle-
dose
gro
up
Hig
h-d
ose
grou
p
∗
∗∗
Figure 7: The effects of quercetin on the level of dopamine in a micemodel of Parkinson’s disease (PD) induced by MPTP. Data wereexpressed as mean ± SE for 10 mice in each group. ###: P < 0.001 ascompared to the control group; ∗: P < 0.05 as compared to MPTPgroup; ∗∗: P < 0.01 as compared to MPTP group.
dismutation, was sufficiently removed by GPx after thepretreatment with quercetin. 4-HNE is a major membranelipid peroxidation product [16]. Additionally, we analyzedthe effect of quercetin on the level of 4-HNE in striatalneurons. As expected, quercetin (100, 200 mg/kg bodyweight) reduced the 4-HNE immunoreactivity in neuronsof PD model mice. It shows that quercetin could decreasethe product of lipid peroxidation. These results indicate themechanism of quercetin neuroprotection effects.
The Na+, K+-ATPase establishes the ionic concentrationbalance that maintains the cell’s potential. Recent researchhas suggested that the Na+, K+-ATPase is susceptible to freeradical-induced damage [17]. In addition, Na+, K+-ATPaseactivity is reduced in disorders of the nervous system. Na+,K+-ATPase activity was examined. The results show thatNa+, K+-ATPase activity is significantly reduced in the MPTPgroup compared to that in the control group. Quercetinreversed the MPTP-induced reduction in Na+, K+-ATPaseactivity. This result indicates that quercetin protects againstMPTP-induced oxidative stress and maintains the restingmembrane potential of neurons.
Dopamine and acetylcholine, which play importantroles in bodily movement, should maintain the dynamicequilibrium in the extrapyramidal system. Sufferers of PDexperience dystonia when the cholinergic nerve is placed inan advantage status. AchE is an enzyme that degrades (via itshydrolytic activity) the neurotransmitter Ach. Acetylcholinebehaves as an excitatory neurotransmitter at neuromuscularjunctions [18]. In the current study, we examined the activityof AchE in the brain tissue of mice. We found that theAchE activity in the MPTP group significantly decreasedcompared with the control group and that quercetin reversedthe MPTP-induced reduction in AchE activity. This result

6 Evidence-Based Complementary and Alternative Medicine
suggests that quercetin reduces the level of acetylcholine byenhancing the AchE activity, which may mediate quercetin-induced improvements in motor balance and coordinationof MPTP-treated mice.
Dopamine is the primary neurotransmitter that isinvolved in motor functions. The loss of dopamine directlyaffects physical movements and is considered a cardinalfeature of PD in humans or in animal models of thedisease [19]. MPTP causes a significant reduction in braindopamine levels. The results of our present study showthat the administration of quercetin markedly improvesMPTP-induced dopamine depletion in the brain tissuewhich is significantly altered following MPTP treatment. Theenhancement of dopamine content by quercetin may restorethe changes in locomotor activity.
Conflict of Interests
All authors manifest that there is no conflict of interests.
References
[1] D. Heisters, “Parkinson’s: symptoms, treatments andresearch,” British Journal of Nursing, vol. 20, no. 9, pp.548–554, 2011.
[2] R. D. Prediger, A. S. Aguiar Jr., E. L.G. Moreira et al.,“The intranasal administration of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (mptp): a new rodent model to testpalliative and neuroprotective agents for parkinson’s disease,”Current Pharmaceutical Design, vol. 17, no. 5, pp. 489–507,2011.
[3] F. Cicchetti, J. Drouin-Ouellet, and R. E. Gross, “Environmen-tal toxins and Parkinson’s disease: what have we learned frompesticide-induced animal models?” Trends in PharmacologicalSciences, vol. 30, no. 9, pp. 475–483, 2009.
[4] A. Ishisaka, S. Ichikawa, H. Sakakibara et al., “Accumulationof orally administered quercetin in brain tissue and itsantioxidative effects in rats,” Free Radical Biology and Medicine,vol. 51, no. 7, pp. 1329–1336, 2011.
[5] K. A. Youdim, M. Z. Qaiser, D. J. Begley, C. A. Rice-Evans, andN. J. Abbott, “Flavonoid permeability across an in situ modelof the blood-brain barrier,” Free Radical Biology and Medicine,vol. 36, no. 5, pp. 592–604, 2004.
[6] K. Ishige, D. Schubert, and Y. Sagara, “Flavonoids protect neu-ronal cells from oxidative stress by three distinct mechanisms,”Free Radical Biology and Medicine, vol. 30, no. 4, pp. 433–446,2001.
[7] F. L’Episcopo, C. Tirolo, S. Caniglia et al., “Combin-ing nitric oxide release with anti-inflammatory activitypreserves nigrostriatal dopaminergic innervation and pre-vents motor impairment in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson’s disease,” Journal ofNeuroinflammation, vol. 7, article 83, 2010.
[8] S. Li and X. P. Pu, “Neuroprotective effect of kaempferolagainst a 1-methyl-4-phenyl-1,2,3,6- tetrahydropyridine-induced mouse model of Parkinson’s disease,” Biological andPharmaceutical Bulletin, vol. 34, no. 8, pp. 1291–1296, 2011.
[9] J. T. Rotruck, A. L. Pope, H. E. Ganther, A. B. Swanson, D. G.Hafeman, and W. G. Hoekstra, “Selenium: biochemical role asa component of glatathione peroxidase,” Science, vol. 179, no.4073, pp. 588–590, 1973.
[10] H. Ohkawa, N. Ohishi, and K. Yagi, “Assay for lipid peroxidesin animal tissues by thiobarbituric acid reaction,” AnalyticalBiochemistry, vol. 95, no. 2, pp. 351–358, 1979.
[11] A. T. S. Wyse, M. Wajner, A. Brusque, and C. M. D.Wannmacher, “Alanine reverses the inhibitory effect of pheny-lalanine and its metabolites on Na+,K+-ATPase in synapticplasma membranes from cerebral cortex of rats,” BiochemicalSociety Transactions, vol. 23, no. 2, p. 227, 1995.
[12] G. L. Ellman, K. D. Courtney, V. Andres, and R. M.Featherstone, “A new and rapid colorimetric determination ofacetylcholinesterase activity,” Biochemical Pharmacology, vol.7, no. 2, pp. 88–95, 1961.
[13] Z. Q. Su, S. H. Wu, H. L. Zhang, and Y. F. Feng, “Developmentand validation of an improved Bradford method for deter-mination of insulin from chitosan nanoparticulate systems,”Pharmaceutical Biology, vol. 48, no. 9, pp. 966–973, 2010.
[14] C. Rohl, E. Armbrust, E. Herbst et al., “Mechanisms involvedin the modulation of astroglial resistance to oxidative stressinduced by activated microglia: antioxidative systems, per-oxide elimination, radical generation, lipid peroxidation,”Neurotoxicity Research, vol. 17, no. 4, pp. 317–331, 2010.
[15] T. Ito, K. Suzuki, K. Uchida, and H. Nakayama, “Different sus-ceptibility to 1-methyl-4-phenylpyridium (MPP+)-inducednigro-striatal dopaminergic cell loss between C57BL/6 andBALB/c mice is not related to the difference of monoamineoxidase-B (MAO-B),” Experimental and Toxicologic Pathology.In press.
[16] J. Lee, B. Kosaras, S. J. Del Signore et al., “Modulationof lipid peroxidation and mitochondrial function improvesneuropathology in Huntington’s disease mice,” Acta Neu-ropathologica, vol. 121, no. 4, pp. 487–498, 2010.
[17] G. J. Lees, A. Lehmann, M. Sandberg, and A. Hamberger,“The neurotoxicity of ouabain, a sodium-potassium ATPaseinhibitor, in the rat hippocampus,” Neuroscience Letters, vol.120, no. 2, pp. 159–162, 1990.
[18] P. Mohseni, H. K. Sung, A. J. Murphy et al., “Nestin isnot essential for development of the CNS but required fordispersion of acetylcholine receptor clusters at the area ofneuromuscular junctions,” Journal of Neuroscience, vol. 31, no.32, pp. 11547–11552, 2011.
[19] Y. S. Lau, G. Patki, K. Das-Panja, W. -D. Le, and S. O. Ahmad,“Neuroprotective effects and mechanisms of exercise in achronic mouse model of Parkinson’s disease with moderateneurodegeneration,” European Journal of Neuroscience, vol. 33,no. 7, pp. 1264–1274, 2011.
Related Documents











