Review Theoretical models for coronary vascular biomechanics: Progress & challenges Sarah L. Waters a, * , Jordi Alastruey b , Daniel A. Beard c , Peter H.M. Bovendeerd k , Peter F. Davies d , Girija Jayaraman e , Oliver E. Jensen f , Jack Lee g , Kim H. Parker b , Aleksander S. Popel h , Timothy W. Secomb i , Maria Siebes j , Spencer J. Sherwin k , Rebecca J. Shipley a , Nicolas P. Smith g , Frans N. van de Vosse l a Oxford Centre for Industrial and Applied mathematics, Mathematical Institute, 24-29 St Giles’, Oxford, OX1 3LB, UK b Department of Bioengineering, South Kensington Campus, Imperial College London, London SW7 2AZ, UK c Department of Physiology, Medical College of Wisconsin, 8701 Watertown Plank Road, Milwaukee, WI 53226, USA d Institute for Medicine and Engineering,1010 Vagelos Laboratories, 3340 Smith Walk, Philadelphia, PA 19104-6383, USA e Centre for Atmospheric Sciences, Indian Institute of Technology, New Delhi, India f Centre for Mathematical Medicine and Biology, School of Mathematical Sciences, University of Nottingham, University Park, Nottingham, NG7 2RD, UK g Computing Laboratory, Wolfson Building, Parks Road, Oxford OX1 3QD, UK h Department of Biomedical Engineering, School of Medicine, Johns Hopkins University, Baltimore, MD 21205, USA i Department of Physiology, University of Arizona, Tucson, AZ 85724, USA j Academic Medical Center, Department of Biomedical Engineering Physics, University of Amsterdam, P.O. Box 22660,1100 DD Amsterdam, The Netherlands k Department of Aeronautics, South Kensington Campus, Imperial College London, London, SW7 2AZ, UK l Department of Biomedical Engineering, Eindhoven University of Technology, P.O. Box 513, 5600 MB Eindhoven, The Netherlands article info Article history: Available online 30 October 2010 Keywords: Vascular structure Mechanics Haemodynamics Mass transport Regulation Adaptation Mathematical and computational model Multi-scale Cellular mechanics Integration abstract A key aim of the cardiac Physiome Project is to develop theoretical models to simulate the functional behaviour of the heart under physiological and pathophysiological conditions. Heart function is critically dependent on the delivery of an adequate blood supply to the myocardium via the coronary vasculature. Key to this critical function of the coronary vasculature is system dynamics that emerge via the interactions of the numerous constituent components at a range of spatial and temporal scales. Here, we focus on several components for which theoretical approaches can be applied, including vascular structure and mechanics, blood flow and mass transport, flow regulation, angiogenesis and vascular remodelling, and vascular cellular mechanics. For each component, we summarise the current state of the art in model development, and discuss areas requiring further research. We highlight the major challenges associated with integrating the component models to develop a computational tool that can ultimately be used to simulate the responses of the coronary vascular system to changing demands and to diseases and therapies. Ó 2010 Elsevier Ltd. All rights reserved. Contents 1. Introduction ....................................................................................................................... 50 2. Coronary vascular structure ......................................................... ................................................ 53 2.1. Introduction ............................................................. .................................................... 53 2.2. Vascular casting .............................................................................................................. 53 2.3. Structural imaging of coronary vasculature ...................................................................................... 53 2.4. Synthetic network generation .................................................................................................. 54 2.5. Optimality principles .......................................................................................................... 54 2.6. Challenges ................................................................................................................... 55 * Corresponding author. Tel.: þ44 1865 280141; fax: þ44 1865 270515. E-mail addresses: [email protected] (S.L. Waters), [email protected] (J. Alastruey), [email protected] (D.A. Beard), [email protected] (P. H.M. Bovendeerd), [email protected] (P.F. Davies), [email protected] (G. Jayaraman), [email protected] (O.E. Jensen), [email protected] (J. Lee), [email protected] (K.H. Parker), [email protected] (A.S. Popel), [email protected] (T.W. Secomb), [email protected] (M. Siebes), [email protected]. uk (S.J. Sherwin), [email protected] (R.J. Shipley), [email protected] (N.P. Smith), [email protected] (F.N. van de Vosse). Contents lists available at ScienceDirect Progress in Biophysics and Molecular Biology journal homepage: www.elsevier.com/locate/pbiomolbio 0079-6107/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.pbiomolbio.2010.10.001 Progress in Biophysics and Molecular Biology 104 (2011) 49e76

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Progress in Biophysics and Molecular Biology 104 (2011) 49e76
Contents lists avai
Progress in Biophysics and Molecular Biology
journal homepage: www.elsevier .com/locate/pbiomolbio
Review
Theoretical models for coronary vascular biomechanics: Progress & challenges
Sarah L. Waters a,*, Jordi Alastruey b, Daniel A. Beard c, Peter H.M. Bovendeerd k, Peter F. Davies d,Girija Jayaraman e, Oliver E. Jensen f, Jack Lee g, Kim H. Parker b, Aleksander S. Popel h,Timothy W. Secomb i, Maria Siebes j, Spencer J. Sherwin k, Rebecca J. Shipley a,Nicolas P. Smith g, Frans N. van de Vosse l
aOxford Centre for Industrial and Applied mathematics, Mathematical Institute, 24-29 St Giles’, Oxford, OX1 3LB, UKbDepartment of Bioengineering, South Kensington Campus, Imperial College London, London SW7 2AZ, UKcDepartment of Physiology, Medical College of Wisconsin, 8701 Watertown Plank Road, Milwaukee, WI 53226, USAd Institute for Medicine and Engineering, 1010 Vagelos Laboratories, 3340 Smith Walk, Philadelphia, PA 19104-6383, USAeCentre for Atmospheric Sciences, Indian Institute of Technology, New Delhi, IndiafCentre for Mathematical Medicine and Biology, School of Mathematical Sciences, University of Nottingham, University Park, Nottingham, NG7 2RD, UKgComputing Laboratory, Wolfson Building, Parks Road, Oxford OX1 3QD, UKhDepartment of Biomedical Engineering, School of Medicine, Johns Hopkins University, Baltimore, MD 21205, USAiDepartment of Physiology, University of Arizona, Tucson, AZ 85724, USAjAcademic Medical Center, Department of Biomedical Engineering Physics, University of Amsterdam, P.O. Box 22660, 1100 DD Amsterdam, The NetherlandskDepartment of Aeronautics, South Kensington Campus, Imperial College London, London, SW7 2AZ, UKlDepartment of Biomedical Engineering, Eindhoven University of Technology, P.O. Box 513, 5600 MB Eindhoven, The Netherlands
a r t i c l e i n f o
Article history:Available online 30 October 2010
Keywords:Vascular structureMechanicsHaemodynamicsMass transportRegulationAdaptationMathematical and computational modelMulti-scaleCellular mechanicsIntegration
* Corresponding author. Tel.: þ44 1865 280141; faxE-mail addresses: [email protected] (S.L. Wat
H.M. Bovendeerd), [email protected] (P.F. DaviesLee), [email protected] (K.H. Parker), apopel@juk (S.J. Sherwin), [email protected] (R.J. Shipley
0079-6107/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.pbiomolbio.2010.10.001
a b s t r a c t
A key aim of the cardiac Physiome Project is to develop theoretical models to simulate the functionalbehaviour of the heart under physiological and pathophysiological conditions. Heart function is criticallydependent on the delivery of an adequate blood supply to the myocardium via the coronary vasculature.Key to this critical function of the coronary vasculature is systemdynamics that emerge via the interactionsof the numerous constituent components at a range of spatial and temporal scales. Here, we focus onseveral components for which theoretical approaches can be applied, including vascular structure andmechanics, blood flow and mass transport, flow regulation, angiogenesis and vascular remodelling, andvascular cellular mechanics. For each component, we summarise the current state of the art in modeldevelopment, and discuss areas requiring further research. We highlight the major challenges associatedwith integrating the component models to develop a computational tool that can ultimately be used tosimulate the responses of the coronary vascular system to changing demands and todiseases and therapies.
� 2010 Elsevier Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 502. Coronary vascular structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53
2.1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 532.2. Vascular casting . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 532.3. Structural imaging of coronary vasculature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 532.4. Synthetic network generation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 542.5. Optimality principles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 542.6. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
: þ44 1865 270515.ers), [email protected] (J. Alastruey), [email protected] (D.A. Beard), [email protected] (P.), [email protected] (G. Jayaraman), [email protected] (O.E. Jensen), [email protected] (J.hu.edu (A.S. Popel), [email protected] (T.W. Secomb), [email protected] (M. Siebes), [email protected].), [email protected] (N.P. Smith), [email protected] (F.N. van de Vosse).
All rights reserved.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7650
3. Mechanical properties . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 553.1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 553.2. Mechanical properties of smaller vessels e the role of the myocardium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 553.3. Mechanical properties of large coronary arteries . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 563.4. Mechanical properties for the coronary venous system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 563.5. Mechanical properties for coronary pulse-wave models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 563.6. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56
4. Blood flow . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .574.1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 574.2. Blood flow in the microcirculation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
4.2.1. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 584.3. 3D flow modelling in larger vessels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
4.3.1. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 594.4. 1D pulse-wave modelling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
4.4.1. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 605. Mass transport processes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
5.1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 605.2. Oxygen transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 615.3. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61
6. Regulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 626.1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 626.2. Modelling approaches . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 626.3. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
7. Angiogenesis and vascular remodelling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 637.1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 637.2. Angiogenesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 647.3. Role of vascular endothelial growth factor family in angiogenesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 647.4. Role of mechanical factors in angiogenesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 647.5. Therapeutic angiogenesis in myocardial ischemic disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 647.6. Computational models of angiogenesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 647.7. Structural adaptation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 657.8. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
8. Vascular cellular mechanics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 658.1. The endothelial cell as a mechanotransducer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 658.2. Mechanotransduction and disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 678.3. Challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
9. Integrated model development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 699.1. Homogenisation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 699.2. Integration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
1. Introduction
The aim of the cardiac Physiome Project is to develop theoreticalmodels to simulate the functional behaviour of the heart underphysiological and pathophysiological conditions. Heart diseasecontinues to be the leading cause of morbidity and mortality inindustrialised countries and throughout much of the world, andbetter methods for cardiovascular disease management are sorelyneeded (Ricotta et al., 2008). The overall clinical goal of in silicomodelling is the development of patient-specific predictive modelsto improve diagnosis, therapy planning and treatment of cardio-vascular diseases (Siebes and Ventikos, 2010). However, theachievement of this objective will also necessarily be underpinnedby characterisation of the underlying physiological mechanismsderived from fundamental scientific investigation.
Using a multi-scale, multi-dimensional, and multi-disciplinaryapproach, theoretical modelling has the potential to predict clinicaloutcomes in order to achieve more effective healthcare. The aim isto develop individualised computer simulations that exploitpatient-specific clinical visualisation modalities and experimen-tally obtained material properties in combination with solidmechanics and fluid dynamic models. Detailed knowledgeabout physiological (control) mechanisms and pathophysiological
processes is necessary to arrive at clinically relevant decision-making tools. Ultimately, these models must account for processesoperating at different time scales, ranging from transient behav-iour of pressure and flow during a cardiac cycle, to effects ofaltered physiological demands or therapeutic interventions,through to much longer time-scale processes involving growth andremodelling due to disease progression and ageing (Lieber et al.,2005). New diagnostic methods evolving from this approachshould allow better patient selection, targetted interventions,therapy assessment and predictions of therapeutic outcomes(Ricotta et al., 2008).
Heart function is critically dependent on the availability of anadequate blood supply to the myocardium. The network ofcoronary vessels must bring oxygenated blood within a smalldistance of every point in the tissue to meet the varying meta-bolic demands of the individual myocytes. In building mathe-matical models capable of simulating functional heart behaviour,it is therefore necessary to develop models for the coronaryvasculature, in addition to models for sub-cellular function,cellular excitationecontraction coupling and cardiac tissuemechanics (see Clayton et al., 2011; Nordsletten et al., 2011). Thisarticle focuses on theoretical models for the coronary vascularsystem.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 51
The ability of the coronary vasculature to meet the metabolicneeds of heart tissue is facilitated by its dynamic structure, which isregulated on a short time scale via the active contraction anddilation of the small arteries and arterioles, and capable on a longertime scale of generating new vessels and remodelling existingvessels according to changing physiological and pathophysiologicalconditions. In addition to this wide range of temporal dynamicsthere are also many components of the coronary vasculature thatinteract over a range of spatial scales from sub-cellular to wholeorgan scale (see Fig. 1). To build mathematical models capable ofsimulating functional heart behaviour, we thus need to understandin detail the individual components of the coronary vascularsystem (including vascular structure and mechanics, fluid flow andmass transport, regulation and remodelling, and cellular biome-chanics) and how they work in an integrated way to respond to theever-changing demands placed upon the coronary vascular system.
Embedding of current physiological understanding withinmathematical frameworks is an approach that has already led toa number of important contributions for understanding coronarycirculation over many years. We start by presenting a brief synopsisof the research that has provided the foundation for the currentmodels. Within this historical summary the interested reader isreferred to a number of recent and more comprehensive reviews.However, it is important to note that the large body of work in thisarea means that, rather than providing a thorough review, our goalin the sections which follow is to examine the current status of keytheoretical models for each constituent component and theirintegration, and assess the modelling challenges that are currentlydefining the cardiac Physiome in the context of the coronarycirculation.
Unique to the coronary circulation is the continuous andrhythmical compression of the blood vessels as the heart contracts,combined with the necessity to provide continuous perfusion tomatch a wide range of metabolic rates. This squeezing effect, orsystolic flow impediment, was first proposed by Scaramucci in 1695and has subsequently been investigated by Porter (1898), Anrepet al. (1927), Downey and Kirk (1975), Spaan et al. (1981),Bruinsma et al. (1988), Krams et al. (1989a,b), among others. Inthemodels of Downey and Kirk (1975) and of Spaan et al. (1981) thecompression effects of intramyocardial pressure were assumed tobe equal to ventricular pressure at the endocardium and todecrease linearly to zero at the epicardium. Arts (1978) developedan integrated model of cardiac wall mechanics and the coronary
Fig. 1. Examples of typical length and time scales encountered in constructing modelsof the coronary vascular system.
circulation, in which the coronary microvessels were loaded by anintramyocardial pressure that was related to myofibre contractionthrough left ventricular pressure (Arts et al., 1979; Arts andReneman, 1985). Later models also considered a direct interactionbetween the myocardium and the microvasculature, throughstiffening of the myocardium during systole alone (Krams et al.,1989a, 1989b), or in combination with radial passive tissue stress(Beyar et al., 1993; Huyghe et al., 1992; Zinemanas et al., 1994; Viset al., 1997; Bovendeerd et al., 2006). In particular, Krams et al.(1989a, 1989b) suggested a more limited role of ventricular pres-sure, and applied the “time varying elastance concept” of Suga et al.(1973) to explain systolic flow impediment. This concept empha-sises the effect that time-varying ventricular wall stiffness, which isassumed to be independent of ventricular pressure, has on coro-nary blood volume. The theory of Krams et al. (1989a, 1989b) isbased on the observation that flow impediment is similar for iso-volumic (high systolic ventricular pressures) and low after loadisobaric (low systolic ventricular pressures) contractions. However,the elastance concept does not explainwhy epicardial flows are notinhibited to the same degree as endocardial flows (Goto et al., 1991;Spaan, 1995) and subsequent studies again suggested thatventricular pressures have a significant effect on time-varyingcoronary flow.
Closely linked to this issue is the role of vascular resistance andcompliance in determining coronary flow. The “vascular waterfallmechanism” proposed by Downey and Kirk (1975) sought toexplain reduced coronary inflow by the increase in resistanceresulting from the collapse of vessels embedded in the myocar-dium. However, since this throttling effect would impede botharterial inflow and venous outflow, this theory alone could notexplain the increased venous outflow during systole. The intro-duction of the “intramyocardial pump model” (Spaan et al., 1981)accounted for the role of vascular compliance. In this modelcompliant vessels are filled from the high-pressure arterial side indiastole and then discharged through the low-pressure venous sidein systole. This concept has since been extended in a number oflumped parameter mathematical models of coronary circulation,e.g. Bruinsma et al. (1988), to account for the variation in vascularresistance and compliance throughout the coronary network withthe temporal change associated with variation in intramyocardialpressure.
Key to understanding and unravelling the role of myocardialcontraction on coronary blood flow has been the development andapplication of experimental measurement techniques to determinethe temporal dynamics of myocardial blood flow across a range ofvessel sizes. These experimental observations now include flows inthemicrocirculation (see reviews of Kajiya et al. (2008) and van denAkker et al. (2010)) and flows subject to varying mechanicalconditions. These latter studies in particular have been instru-mental in quantifying phasic variations throughout the cardiaccycle in both animal models (Kimura et al., 1992; Kajiya et al., 1989,2005) and more recently humans in clinical contexts (see reviewsof Spaan et al. (2006, 2008), Knaapen et al. (2009)).
In addition to capturing the dynamic interactions of flow andmyocardial contraction, a further understanding of the control ofcoronary flow in both normal and pathological conditions isrequired; specifically the regulation of vessel resistance to matchperfusion with the metabolic demands of the heart, in spite offluctuating perfusion pressure. This tendency is termed autor-egulation, and is the result of a large number of different physio-logical mechanisms (see reviews of Rubio and Berne (1975), Feigl(1983), Jones et al. (1995), Deussen et al. (2006), Duncker andBache (2008), Zhang et al. (2008)).
A quick outline of some established concepts of autoregulationcan be summarised as follows. In the heart, microcirculation plays

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7652
a key role in regulation of flow since the majority (w70%) of theresistance, and the greatest capacity to adjust it, resides in themicrovessels (Chilian et al., 1989). Previous studies have discoveredmany different mechanisms of regulation, including the majoreffectors of myogenic response, flow-induced dilation, metaboliccontrol and conducted responses, as discussed in turn below.
The myogenic response is caused by contraction of vascularsmooth muscle cells which respond directly to distending pressurein the lumen (Bayliss, 1902), with a typical time scale in the order oftens of seconds to minutes. Under normal flow conditions, themyogenic response provides the basal tone, producing the vaso-dilatory reserves which can be exploited by other regulatorymechanisms. Reduction in perfusion pressure has been observed toproduce dilation predominantly in microvessels (Kanatsuka et al.,1989; Chilian and Layne, 1990), and graded responses wereobserved in vessels of different diameters, with the most sensitivemyogenicity found in intermediate arterioles of diameter around60 mm in pig coronary vessels (Liao and Kuo, 1997).
Flow-mediated vascular dilation occurs when the vascular wallssense fluid shear stress, leading to local release of nitric oxide (NO)and relaxation of the smooth muscle cells. It has been shown thatthe presence of intact endothelium is necessary to initiate thisprocess (Furchgott and Zawadzki, 1980; Pohl et al., 1986; Griffithet al., 1986). Although flow dependent dilation is readily observedin large coronary arteries (Hintze and Vatner, 1984) and venules(Kuo et al., 1993), studies in isolated vessels indicate that largearterioles exhibit the most sensitive response to flow stimuli (Kuoet al., 1995; Jones et al., 1995).
The metabolic control hypothesis proposes that coronary flowremains constant when subject to a fixed level of metabolicdemand, as autoregulation is governed by a myocyte-producedsubstance which diffuses to the vascular smooth muscle cells viainterstitium. Originally it was suggested that adenosine was themain substrate for metabolic control (Berne, 1963), but subsequentexperimental results have failed to confirm this. There have beenmany other mediators proposed for the role, including bradykinin,CO2 and Hþ, H2O2, potassium and endothelin. However, due to theredundant design of the metabolic control system in which block-ing of any one of these substances fails to abolish the controlmechanism, it is now widely held that the metabolic control isachieved via a combination of many different mediators (Zhanget al., 2008).
The conducted responses in flow control and the oxygen sensingmechanism of the red blood cells have been the focus of some of themore recent modelling studies. In addition, it should be noted thatcoronary autoregulation is achieved via integrated interactions ofthe aforementioned mechanisms. The quantitative investigation ofsuch a system is an ongoing challenge in the cardiac Physiomeproject, and is described in greater detail in Section 6.
A key step to characterising these autoregulator responses isa mechanistic understanding of endothelial function at the lowerspatial scale of the cell. This in turn defines a central challenge ofdeveloping theoretical multi-scale models for the coronary circu-lation, that is, to understand the role of endothelial cells lining allblood vessels in vascular physiology and pathophysiology, and howthey sense and modulate their function when exposed to changesin their local biochemical and biomechanical environment. Theendothelial cell is particularly sensitive to fluid dynamical forcessuch as shear stress and pressure, in response to which theyproduce biochemical signals during the process of mechano-transduction (Dewey et al., 1981; Davies et al., 1984, 2005; Levesqueand Nerem, 1985; Florian et al., 2003; Weinbaum et al., 2003;Mochizuki et al., 2003; Tarbell and Pahakis, 2006). The surface ofendothelial cells has two important specialisations that factor intomechanotransduction and solute transport: the glycocalyx
composed of membrane bound highly charged macromoleculesregularly distributed over the luminal surface (comprehensivelyreviewed by Reitsma et al. (2007) andWeinbaum et al. (2007)) andprimary cilia e one per cell e that can project beyond the luminalsurface as membrane bound continuations of the cytoskeleton(Kojimahara, 1990; van der Heiden et al., 2008). Both may playa role in endothelial mechanotransduction and the glycocalyx alsoacts as a transport barrier and as a porous hydrodynamic interfacein the motion of red and white cells in microvessels (Weinbaumet al., 2003). These cellular elements are more extensively out-lined in section 8.
This inherently integrative nature of coronary investigationcombines experimental measurement and modelling. Such work isfocused on understanding, arguably, one of the most complexvascular systems in terms of regulation, mechanical interaction andclinical pathologies. Below we aim to outline many of the researchchallenges faced in developing integrated mathematical models todescribe the coronary vascular systemwhich, if overcome, will alsobe invaluable in developing models to understand other organsystems.
As already highlighted, in order to build computational toolscapable of simulating functional heart behaviour, we are faced withthe challenge of integrating models for physical processes atdisparate spatial scales, e.g. incorporating micro-scale flow andmass transport processes in a macro-scale model for myocardialtissue. The challenge is to introduce small-scale information intolarger-scale models without the resulting models becomingcomputationally intractable. One approach is to use a lumpedrepresentation where fine-scale structures, e.g. blood vessels,smaller than a certain size are represented by a single compartmentwith uniform properties. A limitation of this simplified approach isthat significant spatial variations may exist within this compart-ment, which are not represented by the model. An intermediateapproach between detailed representation of fine-scale structureand a lumped approach is provided by homogenisation theory. Inthis theory, a local spatial averaging of fine-scale structure is ach-ieved by exploiting asymptotic techniques to estimate macro-scaleproperties, based on explicit solutions in smaller-scale subunits(Huyghe et al., 1989a, 1989b; Vankan et al., 1997; Chapman et al.,2008; Shipley and Chapman, 2010; Shipley et al., submitted forpublication). Although homogenisation techniques have beenused for many years in modelling the mechanical properties of themyocardium, the technique is now starting to be used more widelyin cardiovascular fluid dynamics modelling, and we highlight thismethodological approach in Section 9.
Because of the inherently multi-scale nature of the system wehave chosen to present the research ideas by application (flow,mass transport, etc) rather than by modelling methodology, andstress that many of the same theoretical techniques are used in thedevelopment and solution of the component models. To modelcoronary flows, including the microcirculation and the largearteries and veins, we must understand the geometry of the flowdomain, and the mechanical environment withinwhich the vesselsfind themselves (determined both by the properties of the vascularwall and the surrounding myocardium). Such aspects are consid-ered in Sections 2 and 3. We then consider flow and mass transportin Sections 4 and 5. Finally, we consider how coronary vasculaturenetworks evolve on both short (regulation) and long (adaptation)time scales in Sections 6 and 7. The mechanisms by which endo-thelial cells sense fluid mechanical forces and produce biochemicalsignals in the process of mechanotransduction are considered inSection 8. Each section highlights the current state of the art ofmodelling in the field, before going on to explore open researchchallenges. In section 9 we discuss how the component modelsmay be integrated.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 53
2. Coronary vascular structure
2.1. Introduction
The human heart contains approximately 108 vessels that rangeover four orders of magnitude in calibre and length. The coronarymicrocirculation, consisting of vessels with dimensions less thanabout 200 mm (Popel and Johnson, 2005) contains more than 95% ofthe total coronary vasculature segments. These vessels areresponsible for the major resistance to vascular flow, as well asshort-term regulation (see Section 6) and long-term adaptations(see Section 7) that ensure the cardiac demands are satisfied at bothlocal and global levels. However, despite years of research,a complete and concise description of the coronary microcircula-tion remains elusivee the vessel networks exhibit a heterogeneousand anisotropic 3D mesh-like organisation, with a vascular densitythat is some 5e10 times higher compared to other organs, whichmakes quantitative characterisation of its structure technicallychallenging. Similarly, the understanding of the structure of thesmall coronary arteries and veins commonly found to run intra-murally, e.g. with regards to the extent of their collateralisation(Loukas et al., 2009c; Tayebjee et al., 2004), is incomplete.
In comparison, there are relatively complete descriptive char-acterisations of the large human coronary arterial network and itsvariations (Loukas et al., 2009a) and to a lesser extent, the venoussystem (Ho et al., 2004; Loukas et al., 2009b). At this level, thevessels largely follow a common branching pattern allowingassignments of standard anatomical ontologies and statisticalordering systems, and their quantitative and spatial distributionscan be routinely obtained by clinical imaging modalities such asmagnetic resonance imaging (MRI) or multi-detector computedtomography (MDCT) angiography.
When considering the coronary vascular network structure it isimportant to consider the spatial distribution of the segmentsembedded within the myocardium in addition to the networktopology and morphology. The significant cross-talk betweencoronary flow and cyclical muscular contraction that is particular tothe heart (Westerhof et al., 2006) gives rise to the unique pulsatilitywith out-of-phase pressure and flow waveforms and arterial/venous phasic differences in coronary flow. Thus it is not only thearterial-to-venous path travelled by an erythrocyte that matters,but also the dynamic interaction of blood with the intramyocardialstresses and deformations along the path whichmust be accountedfor. This coupling effect is particularly pronounced in the micro-vascular compartment that forms themost extensive interface withextravascular structures.
Here we discuss the state-of-the-art techniques that arecurrently used to describe structure, including vascular casting andimaging, together with synthetic network generation techniquesthat can be used when such detailed structural information is notavailable.
2.2. Vascular casting
Vascular casting has been used since the 1970s for anatomicalstudies of vascular networks. Early studies investigated the coro-narymicrovasculature usingMicrofil (Bassingthwaighte et al., 1974)or catalysed polymer resin (Van Bavel and Spaan, 1992). Althoughthis technique cannot be applied in vivo, it is still widely used forvascular anatomical studies because it preserves the complexgeometry of the vascular network at high resolution, i.e. from fullnetwork structure to the individual cellular imprints on the endo-luminal surface. More recent studies have combined radio-opaqueMicrofil with micro-computed tomography (mCT, Jorgensen et al.,1998) or fluorescent dye suspensions in the resin (Spaan et al.,
2005) to allow three-dimensional imaging, as outlined in Section2.3. Such anatomical information acquired from the combination ofcasting with advanced imaging modalities is beneficial to integra-tive modelling, as it allows building of theoretical models ofstructureefunction coupling based on multi-scale information.
A notable casting study was made in the early 1990s to obtaina comprehensive statistical characterisation of the coronary struc-ture. In a series of publications, large networks of porcine coronaryarteries (Kassab et al., 1993), veins (Kassab and Fung, 1994) andcapillaries (Kassab et al., 1994) were examined manually to deter-mine segment length and diameter, and to generate connectivitymatrices. These studies yielded detailedmorphological informationon 11 orders of arteries and 12 orders of veins (divided into sinusalor thebesian types) using the Strahler classification system(Strahler, 1957), and highlighted the patent heterogeneity presentin the coronary vasculature. A key motivation of this study was toreduce the vast amount of structural information into a formsuitable for mathematical modelling, and its outcomes are anundeniably valuable contribution to this objective. This approachhas been recently extended via non-invasive computed tomog-raphy (CT) imaging and computer-assisted analysis (Wischgollet al., 2009). However, as discussed above, the approach of quan-tifying morphological variabilities strictly within artificiallyassigned orders may be misleading as it does not account for thecoupling between structure and physiological function such asautoregulation and regional metabolic demand. The correlationbetween haemodynamic parameters and the vessel order is veryscattered, and the assumptions underlying the ordering schemesmay preclude such issues from being addressed. Moreover, due tothe destructive technique employed in this study, informationregarding the spatial distribution of segments cannot be recovered.
2.3. Structural imaging of coronary vasculature
During the past decade, vascular imaging technology and theassociated automated analysis algorithms have rapidly improved.The best resolution data are still obtained using casting techniques,and effort has been directed at constructing custom devices toimprove the resolution and coverage. Key examples include thelarge volume confocal imaging system (LVCIS) (Sands et al., 2005)and knife-edge scanning microscopy (KESM) (Mayerich et al.,2008). Both modalities operate on resin-embedded tissue byincrementally imaging a plane of tissue andmechanically removingslices to build up a three-dimensional dataset. They are capable ofachieving 300e400 nm in resolution, and can thus fully resolvevessels of the microcirculation. However, these techniques arelimited in the acquisition volume to a maximum of a few mm3 dueto the requirement the samples are fixed in resin which mustdiffuse into the tissue. Confocal microscopy has been used for dualmyocyte-vascular imaging which was used to understand theirmicrostructural relationship (Lee et al., 2007). The imaging cry-omicrotome device (Spaan et al., 2005) was purpose-built to imagelarger samples and can image the coronary network of a wholehuman sized heart with less than 25 mm voxel dimensions. Thisdevice also allows the coronary flow distribution across themyocardial wall to be quantified based on detection of fluorescentlylabelled microspheres that are injected under different conditionsin vivo (van Horssen et al., 2009, 2010).
Specialisations of more conventional modalities, such as CT orMRI, have also been used to achieve high-resolution vascularimaging. This was demonstrated in the hierarchical CT/mCT/synchrotron radiation mCT (STmCT) imaging study (Heinzer et al.,2006) and 11.7T MRI study (Burton et al., 2006). These devicesare subject to cost and availability issues, but present a promisingavenue for future investigations. The scope for automatic
S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7654
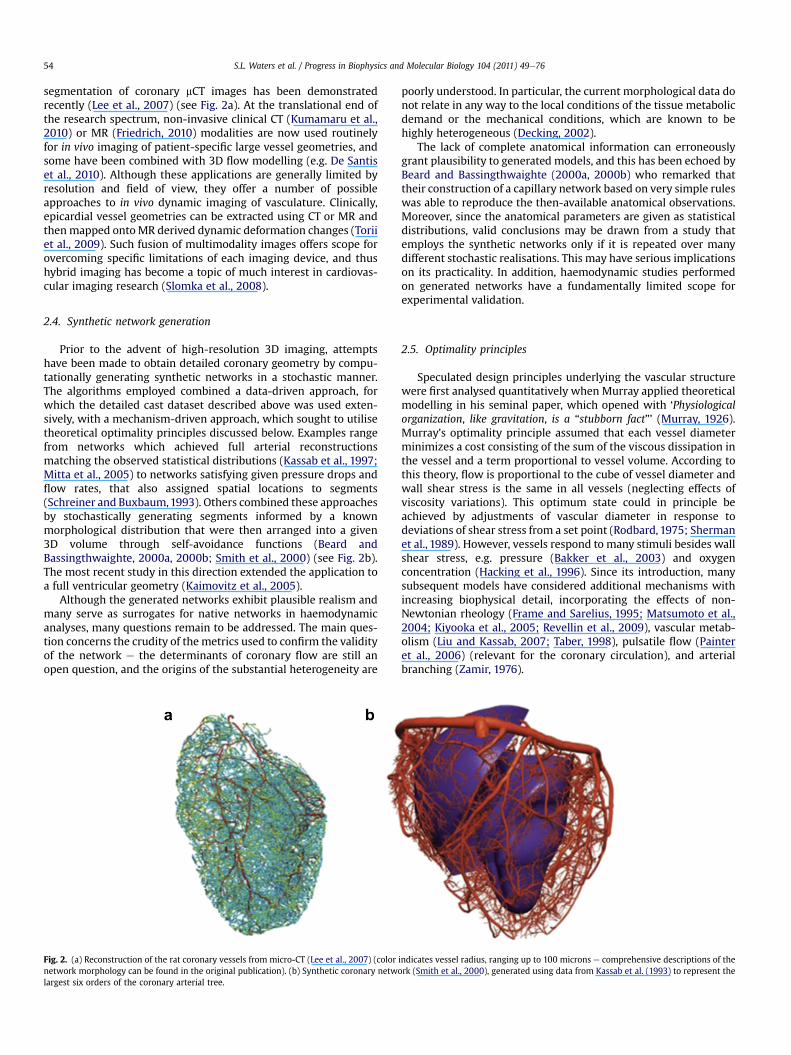
segmentation of coronary mCT images has been demonstratedrecently (Lee et al., 2007) (see Fig. 2a). At the translational end ofthe research spectrum, non-invasive clinical CT (Kumamaru et al.,2010) or MR (Friedrich, 2010) modalities are now used routinelyfor in vivo imaging of patient-specific large vessel geometries, andsome have been combined with 3D flow modelling (e.g. De Santiset al., 2010). Although these applications are generally limited byresolution and field of view, they offer a number of possibleapproaches to in vivo dynamic imaging of vasculature. Clinically,epicardial vessel geometries can be extracted using CT or MR andthenmapped ontoMR derived dynamic deformation changes (Toriiet al., 2009). Such fusion of multimodality images offers scope forovercoming specific limitations of each imaging device, and thushybrid imaging has become a topic of much interest in cardiovas-cular imaging research (Slomka et al., 2008).
2.4. Synthetic network generation
Prior to the advent of high-resolution 3D imaging, attemptshave been made to obtain detailed coronary geometry by compu-tationally generating synthetic networks in a stochastic manner.The algorithms employed combined a data-driven approach, forwhich the detailed cast dataset described above was used exten-sively, with a mechanism-driven approach, which sought to utilisetheoretical optimality principles discussed below. Examples rangefrom networks which achieved full arterial reconstructionsmatching the observed statistical distributions (Kassab et al., 1997;Mitta et al., 2005) to networks satisfying given pressure drops andflow rates, that also assigned spatial locations to segments(Schreiner and Buxbaum,1993). Others combined these approachesby stochastically generating segments informed by a knownmorphological distribution that were then arranged into a given3D volume through self-avoidance functions (Beard andBassingthwaighte, 2000a, 2000b; Smith et al., 2000) (see Fig. 2b).The most recent study in this direction extended the application toa full ventricular geometry (Kaimovitz et al., 2005).
Although the generated networks exhibit plausible realism andmany serve as surrogates for native networks in haemodynamicanalyses, many questions remain to be addressed. The main ques-tion concerns the crudity of the metrics used to confirm the validityof the network e the determinants of coronary flow are still anopen question, and the origins of the substantial heterogeneity are
Fig. 2. (a) Reconstruction of the rat coronary vessels from micro-CT (Lee et al., 2007) (colornetwork morphology can be found in the original publication). (b) Synthetic coronary netwolargest six orders of the coronary arterial tree.
poorly understood. In particular, the current morphological data donot relate in any way to the local conditions of the tissue metabolicdemand or the mechanical conditions, which are known to behighly heterogeneous (Decking, 2002).
The lack of complete anatomical information can erroneouslygrant plausibility to generated models, and this has been echoed byBeard and Bassingthwaighte (2000a, 2000b) who remarked thattheir construction of a capillary network based on very simple ruleswas able to reproduce the then-available anatomical observations.Moreover, since the anatomical parameters are given as statisticaldistributions, valid conclusions may be drawn from a study thatemploys the synthetic networks only if it is repeated over manydifferent stochastic realisations. This may have serious implicationson its practicality. In addition, haemodynamic studies performedon generated networks have a fundamentally limited scope forexperimental validation.
2.5. Optimality principles
Speculated design principles underlying the vascular structurewere first analysed quantitatively when Murray applied theoreticalmodelling in his seminal paper, which opened with ‘Physiologicalorganization, like gravitation, is a “stubborn fact”’ (Murray, 1926).Murray’s optimality principle assumed that each vessel diameterminimizes a cost consisting of the sum of the viscous dissipation inthe vessel and a term proportional to vessel volume. According tothis theory, flow is proportional to the cube of vessel diameter andwall shear stress is the same in all vessels (neglecting effects ofviscosity variations). This optimum state could in principle beachieved by adjustments of vascular diameter in response todeviations of shear stress from a set point (Rodbard, 1975; Shermanet al., 1989). However, vessels respond to many stimuli besides wallshear stress, e.g. pressure (Bakker et al., 2003) and oxygenconcentration (Hacking et al., 1996). Since its introduction, manysubsequent models have considered additional mechanisms withincreasing biophysical detail, incorporating the effects of non-Newtonian rheology (Frame and Sarelius, 1995; Matsumoto et al.,2004; Kiyooka et al., 2005; Revellin et al., 2009), vascular metab-olism (Liu and Kassab, 2007; Taber, 1998), pulsatile flow (Painteret al., 2006) (relevant for the coronary circulation), and arterialbranching (Zamir, 1976).
indicates vessel radius, ranging up to 100 microns e comprehensive descriptions of therk (Smith et al., 2000), generated using data from Kassab et al. (1993) to represent the

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 55
In reality the vascular system shows large deviations fromMurray’s law: e.g. it has been shown that in the microcirculationshear stress varies with local pressure (Pries et al., 1995). Whileoptimality principles can be helpful in understanding the overallstructure of vascular networks, they have significant limitations.Firstly, minimizing an energy cost does not necessarily take intoaccount the functional requirements and constraints that apply tothe system. No study has yet incorporated the metabolic require-ments and perfusion volume of the surrounding tissue into Mur-ray’s optimality framework. Secondly, the energy minimum can bebroad, allowing large variations in structure. It is instructive to notethat in the original Murray’s law, the “cube law” exponent may bevaried from 1.5 to 10 with only a 5% difference in the energy cost(Sherman et al., 1989), and the significant scatter observed in theanatomic branching angles correspond to only a 2% deviation fromthe optimality of minimum drag hypothesis (Zamir and Bigelow,1984). Thirdly, there is no a priori reason to assume that evolu-tionary processes have achieved a true optimum.
It is clear that considerable further effort will be required toelucidate vascular design principles, whether a stubborn fact or not.In light of the known stimuli that lead to structural adaptations(discussed in Section 7.7), incorporating such details may help toaccount for some of the physiological deviations from Murray’soptimum. In addition, it may be necessary to account for thedifferent mechanical conditions to which the vessels are exposed,within different regions of the heart (see Section 3). However, thereis no inherent mechanism for sensing optimality, except long-termevolutionary advantage, and, more recently, alternative theories forstructural adaptation have developed, which have focused onidentifying the signals and stimuli that are available to individualcomponents of the vascular system in order to understand howresponses to such signals can lead to observed vascular systembehaviours and structures. This is discussed further in Section 7.7.
2.6. Challenges
The rapid recent progresses in coronary vascular structuralcharacterisation highlights a number of major challenges for futureresearch efforts. To aid modelling studies of coronary haemody-namics and associated adaptation/mass transport processes,a concise description of the vascular structure is required. Sucha descriptionwill ideally capture the key determinants of flowwithfew parameters to aid model reduction and be physiologicallymeaningful to aid model interpretation. This information should bederived from detailed studies of coronary vascular structures,which are now possible to obtain. Also, recognising that thevasculature is a dynamically adapting system in response to shortand long-term stimuli (see Sections 6 and 7), structural studiesspanning different stages of pathology will contribute to ourfundamental understanding of the vascular structureefunction andthe etiology of the diseases. In addition, these studies shouldaddress both the topological complexities and the spatial organi-sation of the vascular network, if the coupled coronary flow-cardiaccontraction effects are to be understood.
3. Mechanical properties
3.1. Introduction
Knowledge of the mechanical properties of the coronaryvasculature and its surrounding myocardial tissues is essential forthe modelling of flow in the coronary system and its relation tocardiac muscle contraction (Zhang et al., 2004). The main charac-teristics of coronary flow can be analysed using pulse-wave velocitynetwork models (e.g. Reymond et al., 2009; Alastruey et al., 2009b;
Bessems et al., 2007). These network models form the boundaryconditions for local 3D flow phenomena in the large coronaryarteries (Quarteroni and Formaggia, 2004), and are the basis forinflow conditions for models of the myocardial microcirculation(Matsumoto and Kajiya, 2005). In the following sections, we high-light the importance of knowledge about themechanical propertiesof the different structures in the cardiac tissue and vasculature andindicate how these properties can be obtained experimentallyenabling their use in models of coronary flow, myocardial perfusionand contraction.
3.2. Mechanical properties of smaller vessels e the roleof the myocardium
The mechanical properties of the small arteries, the arteriolesand the microcirculation, which are all embedded in thesurrounding myocardial tissue, are difficult to determine. However,due to the limitedwall thickness of these vessels, thewall motion ofthese small vessels is dominated by the time-dependent dynamicsof the surrounding tissue environment (Goto et al., 1996; Soropet al., 2003). During systole, the effect of cardiac contraction onperfusion is manifested as a decrease of coronary arterial inflowand an increase of coronary venous outflow. In turn, coronarypressure affects cardiac muscle contraction and oxygen consump-tion (Gregg, 1963; Dankelman et al., 1999) and coronary fillingaffects muscle stiffness and perfusion. This two-way interactionbetween the cardiac muscle and perfusion is called mechanicalcross-talk (Westerhof et al., 2006). While fundamental to coronarydynamics, characterisation of these interactions remains a complexchallenge as evidenced by several studies that have demonstratedthat during normal contraction, intramural vessels are shieldedfrom the high systolic left ventricular pressure by the myocardiumitself (Kouwenhoven et al., 1992; VanTeeffelen et al., 1998). Thestrong interplay between cardiac contraction and vascular flow hasbeen shown to depend on the transmural location of the vessel inthe heart wall, and similar types of arteries in different myocardialpositions have different wall thickness (Choy and Kassab, 2009).
Several models have been proposed to explain aspects ofmechanical cross-talk. Some explain basic interaction mechanismsfor a representative discretely modelled blood vessel (Downey andKirk, 1975; Arts, 1978; Arts et al., 1979; Arts and Reneman, 1985;Spaan et al., 1981; Bruinsma et al., 1988; Krams et al., 1989a,1989b; Beyar et al., 1993; Huyghe et al., 1992; Zinemanas et al.,1994; Vis et al., 1997; Smith, 2004; Bovendeerd et al., 2006).Although extensions to several thousands of vessel segments havebeen presented (Lee et al., 2009) it seems unlikely that flow in thecomplete coronary vascular tree can be modelled througha discrete approach. An alternative approach which still retainslocal spatial variations is to use homogenisation theory, in whichaveraging of the fine-scale structure is achieved by exploitingasymptotic techniques to model tissue-scale properties. Thisapproach is used in the model proposed by Huyghe et al. (1992),which represents the myocardium as a two-phase mixture. Thesolid phase represents the myocardium. The fluid phase iscomposed of several compartments, representing the blood in thehierarchical structure of the coronary tree. Themodel employs a 4Dspace, consisting of the traditional 3D space augmented by a fourthdimension that describes the organisation of the vascular bed fromarteries through arterioles, microvessels and venules to veins. 4Dblood flow and pressure gradients are coupled through a 4Dpermeability tensor. The latter tensor is derived from the anatomyof the vascular bed through a formal averaging procedure (Huygheet al., 1989a, 1989b; Vankan et al., 1997). Homogenisationapproaches may be validated by comparison of the results withsimulations withmany vessels represented as discrete structures to

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7656
account for microvascular flows in cardiac tissue. The potential forhomogenisation approaches to be used in other aspects of coronaryvascular system modelling is discussed in more detail in section 9.
In both discrete and continuum models of the coronary circula-tion, models for the mechanical properties of the passive and activemyocardium are needed. To represent these constitutive propertiesa homogenisation approach is also often applied. Commonly, thepassive myocardium is considered virtually incompressible,anisotropic and non-linearly elastic. This behaviour is describedphenomenologically through a strain energy density function, witha volumetric part to control tissue volume change and an additionalpart to describe the response to deformation. Common choices forthe latter part are an exponential function or a pole zero law withstress growing unboundedly in terms of normal and shear straincomponents with respect to a local material-bound coordinatesystem (for further details see Nordsletten et al. (2011)).
Early approaches to describe the active myocardium usedphenomenological, tissue-level models. These few-parametermodels enable systematic evaluation of the force-time, force-length and forceevelocity relations on cardiac mechanics andcoronary perfusion. Obviously, they lack the coupling to (patholo-gies at) the cellular level, that is present in more recent models inwhich the complete sequence of events from depolarisation of thecell membrane, through to cross bridge formation and subsequentforce generation, is described. Pitfalls and challenges encounteredin using such latter models, related to combining detailed sub-models into on overall model, have recently been reviewed by Leeet al. (2009).
3.3. Mechanical properties of large coronary arteries
The flexibility and active properties of the large arteries stronglyinfluence the flow patterns within them. Simulation of blood flowrequires knowledge the mechanical properties of the arterial wall.For fluidestructure interaction (FSI) simulations, full 3D constitu-tive relations between tissue stress and strain tensors are required.Examples are models from the Holzapfel/Humphrey family(Holzapfel et al., 2000; Humphrey, 2001). This family includesfibre-reinforced models for the arterial wall which incorporate themicrostructural features of collagen fibres and morphologicalinformation about the different layers. Simpler phenomenologicalmodels include exponential, Truesdell-like, power law models forneo-Hookean material (Truesdell and Noll, 1965; Raghavan andVorp, 2000). The advantage of the more complex, fibre-reinforcedmodels is that the role of smooth muscle cells can be accounted forwithin the theoretical framework (Zulliger et al., 2004). The maindisadvantage of these complex models is that they contain largenumbers of parameters that are difficult to determine in a simpleexperiment. One approach to overcome this difficulty is to exploitreverse engineering techniques by using a hybrid experimental-numerical approach (Oomens et al., 1993). The main issues inobtaining these parameters are related to pre-straining and the roleof residual stresses (Zulliger et al., 2004).
3.4. Mechanical properties for the coronary venous system
In contrast to the situation in the systemic circulation, thecoronary venous system cannot be viewed as a reservoir of blood ata relatively low-pressure collecting blood from the microcircula-tion, with a quasi-static resistance to flow that is small compared tothe resistance of the microcirculation. The coronary veins areperiodically squeezed by the heart muscle contraction resulting ina flow with a significant pulsatility and a time-varying non-linearresistance and compliance. Constitutive relations between tissuestress and strain tensors of the veins and inclusion of the forces that
originate from myocardial contraction are required to model thesephenomena.
3.5. Mechanical properties for coronary pulse-wave models
When considering networks of large vessels, pressure and flowdistributions can be modelled as travelling pulse waves along thevascular network (see Section 4.4). Pulse-wave velocity andattenuation are determined by geometrical properties, e.g. wallthickness and lumen diameter, and mechanical properties, e.g.anisotropic elastic or visco-elastic properties of the arterial wall,the non-Newtonian viscous properties of blood and, indirectly, bythe mechanical properties of the surrounding tissue. Consequently,lumped parameter models (0D) and wave propagation models (1D)(see section 4.4 below) that describe pressure (p), cross-sectionalarea (A), and flow (q) or velocity (v ¼ q/A) relations in the coronarynetwork require the linear or non-linear properties of the arterialwall to be defined. In the simplest case, a model for arterialcompliance in which the pressure depends on the arterial cross-sectional can be used and data can be obtained by invasive or non-invasive in vivo or ex vivo determined pressureearea relations. Thisrelation can then be used to eliminate one of the three independentvariables from the set (p,q,A) leading to either a (p,A)- or a (p,q)-formulation. However, when either non-linear or visco-elasticconstitutive relations are employed (see Reymond et al. (2009) foran overview), this elimination is not straight forward and a (p,q,A)-formulation is used (Bessems et al., 2008). The same formulationcan be used if instead of a pressureearea relation an axisymmetricmodel of a thin or thick walled tube is used to close the system ofequations (e.g. Huberts et al., 2009). Such a model requires materialgeometrical properties, e.g. wall thickness, wall composition andcomposition dependent shear and bulk modulus. More compli-cated models based on fibre-reinforced material behaviour(Holzapfel et al., 2000) are currently not applied in 1D wavepropagation models although in theory this extension is possibleonce the model parameters are known. As mentioned earlier pre-strain and residual stress are complicating factors. Considerationmust also be given to flow characteristics at arterial sites of localgeometric complexity such as branches and bifurcations of thecoronary arteries. The pulse waveforms and flow separations areextraordinarily difficult to estimate and undoubtedly are respon-sible for the biological susceptilbility to atherosclerosis at suchlocations (e.g. see Wischgoll et al., 2009).
As the coronary arteries, for a large part, are embedded in thecardiac muscle, the passive and active mechanical properties ofthe myocardium (see Section 3.2) are key to determining themechanical coupling to coronary flow (Guiot et al., 1990). Forthe large coronary vessels, the motion and deformation of themyocardium can be considered to be a boundary condition on theflow domain. For the coronary pulse-wave equations, the outflowmicrocirculation can be incorporated in the outflow boundarycondition, and in general a pressureeflow relation is specified. Thisrelation can follow from a simple windkessel model. Alternatively,more advanced models (described earlier in Section 3.2) can beemployed that incorporate the intramyocardial pressure and itsinfluence on compliance and resistance (see Zinemanas et al., 1994;Bovendeerd et al., 2006). Furthermore, by modelling the microcir-culation as a network of vessels, aspects of growth and remodellingcan be incorporated and explored.
3.6. Challenges
A challenge in the development of the continuum approach tomodel large numbers of vessel segments is to exploit detailedinformation on the anatomy of the vascular bed. A further

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 57
consideration is to incorporate effects of vascular adaptation byactive regulation of vascular smooth muscle tone and viaremodelling within the continuum approach. Additionally, thedevelopment of poroelastic continuum models (Ng et al., 2005;May-Newman and McCulloch, 1998; Huyghe et al., 1992) will notonly provide local and time-dependent pressureeflow relations,but also has the potential to describe the influence of perfusion onpassive/active properties of the myocardium. Attention should alsobe given to methods of coupling continuummodels for networks ofmicrovessels to discrete models for large arteries.
From a disease standpoint, it is important to consider the effectof cardiac contraction on the transmural distribution of blood flowand the vulnerability of the subendocardium, especially in thepresence of an epicardial stenosis (Bache and Schwartz, 1982;Hoffman and Spaan, 1990; Austin et al., 1994). From a physiolog-ical point of view, the relative duration of diastole as a function ofheart rate is an additional important parameter of subendocardialperfusion (Merkus et al., 1999, 2001; Fokkema et al., 2005; van denWijngaard et al., 2008). Ultimately, such models should endeavourto include effects of endogenous regulatory mechanisms andpharmacological responses of the coronary microcirculation thatare active in vivo (Hoffman and Spaan, 1990; Komaru et al., 2000).
Most importantly, the models should be tested by comparisonwith experimental data. Initial experiments could involve smalltissue samples, e.g. trabeculae and papillary muscle. Compared tothe in situ heart, small samples allow for easier control of experi-mental conditions and easier access for measurements. Suchexperiments can be used to test both discrete network andcontinuum models. In the future, isolated (Langendorf or beating)heart experiments would offer a testing platform (see e.g. deWegeret al., 2010). Similarly, advanced measurements to determine themechanical properties of coronary arteries and veins by applyingphysiological loads in a physiological environment (see e.g. van denBroek et al., in press) should be carried out.
4. Blood flow
4.1. Introduction
Healthy human blood is a concentrated suspension containingred blood cells (RBCs) at a concentration (haematocrit) of 40e45%.Unstressed human RBCs are biconcave discs with a diameter ofabout 8 mm. In vessels with diameters much larger than this, bloodcan be considered as a continuum with a viscosity that is approx-imately constant at normal physiological shear rates. However, thefinite size of RBCs results in non-continuum behaviour in narrowvessels. This gives rise to several effects which should be consid-ered when simulating flow in the microcirculation.
The flow characteristics exhibited in the coronary vascular treevary markedly from the microcirculation, to flow in the largecoronary arteries. The Reynolds number for blood flow in themicrocirculation is very small, typically in the range 0.001 to lessthan 1, and the flow is laminar and governed by the Stokes equa-tions. In many tissues, flow pulsatility is strongly damped byviscous effects in combination with vascular compliance before itreaches the microvessels. However, in the myocardium, thecontraction of the surrounding tissue may result in a significantpulsatile component (see Section 3.2). In this case, the flowmay beanalysed using a quasi-steady approximation.
In contrast, the Reynolds number for flow in the large arterialvessels is high, typically in the range of hundreds, and this providesparticular challenges in flowmodelling. Thewell-known sensitivityof high-Reynolds number flows to geometric features is compli-cated by the dynamic nature of the flow domain, where vesselsundergo continuous changes in shape and orientation, generating
unusual kinematic and inertial contributions to the governingdynamics (Lynch et al., 1996). Flows are additionally subject totime-dependent upstream and downstream boundary conditionsassociated with cardiac pumping and distal vessel compression (asoutlined in Section 3.5).
One-dimensional (1D) ‘reduced’ pulse-wave modelling providesa good compromise between computational cost and accuracywhen a global assessment of blood flow in the cardiovascularsystem is required. This approach can be used to simulate thechanges in blood pressure and flow in time and along the axialdirection of large (coronary and non-coronary) vessels produced bythe contraction of the heart. These changes propagate in the form ofwaves, called pulse waves, which carry valuable information aboutthe morphology and functionality of the cardiovascular system.
In the following sections, we consider in detail microcirculatoryflows, 3D flow modelling in large vessels, and reduced 1D pulse-wave models. For ease of presentation, relevant research challengesare highlighted at the end of each section.
4.2. Blood flow in the microcirculation
When blood flows in narrow tubes, the concentration of RBCswithin the tube (tube haematocrit) is less than the RBC concen-tration in the blood entering and leaving the tube (discharge hae-matocrit). This Fåhraeus effect (Fahraeus, 1928) arises from the factthat RBCs travel faster than plasma on average and therefore haveshorter transit times.
Resistance to blood flow in narrow tubes is convenientlyexpressed in terms of the apparent viscosity, defined as theviscosity which, when substituted in Poiseuille’s law, would givethe same flow rate for a given tube length, diameter and pressuredrop. In glass tubes the apparent viscosity declines substantiallywith tube diameter at diameters below 300 mm, a phenomenonknown as the FåhraeuseLindqvist effect (Fahraeus and Lindqvist,1931; Martini et al., 1930; Pries et al., 1992). At diameters neartheminimumwhich allows passage of RBCs (about 3 mm) this effectis reversed. An empirical equation (Pries et al., 1992) describes thevariation of apparent viscosity with tube diameter and dischargehaematocrit. The reduction in apparent viscosity in narrow tubesresults mainly from the presence of a layer of cell-free or cell-depleted plasma near the tube wall. A good fit to experimentalresults for diameters 30 mm and above is obtained by assuminga two-phase model of blood flow, in which a central cylindricalregion with viscosity three times plasma viscosity is surrounded bya plasma layer with a fixed width of 1.8 mm (Secomb, 1995). Itshould be noted that this width is a fitted parameter and has notbeen predicted from the mechanical properties of RBCs.
Experimental observations of distributions of flow and haema-tocrit in microvascular networks indicate that flow resistance inliving microvessels is substantially higher than in glass tubes withcorresponding diameters (Pries et al., 1990). An empirical formulafor the viscosity of blood in vivo has been developed (Pries et al.,1994), based on data obtained in the rat mesentery. The principalcause of the difference between in vitro and in vivo blood viscosityin microvessels is believed to be the presence of the relatively thickendothelial surface layer (ESL, also called glycocalyx) bound to theinner surface of endothelial cells, that may be up to 1 mm in width(Pries et al., 2000). The presence of this ESL leads to the exclusion ofred blood cells from the region near the vessel wall and a conse-quent reduction in the tube haematocrit, defined as the fraction ofthe tube volume occupied by red blood cells (Constantinescu et al.,2001; Klitzman and Duling, 1979). Studies in which the ESL wasexperimentally manipulated (Vink and Duling, 1996) yieldedinformation about the biophysical properties of the layer, whichprovided a basis for theoretical analyses of the mechanical

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7658
interactions between the layer and flowing blood (Damiano et al.,1996; Han et al., 2006; Secomb et al., 2001).
At diverging microvascular bifurcations, the partition of RBCsinto the downstream branches does not generally correspond to thepartition of total blood flow, with the result that different haema-tocrits are found in the downstream branches. Generally, thehigher-flow branch receives a higher haematocrit. Based onobservations in the rat mesentery, empirical relationships havebeen developed to describe the dependence of phase separation onthe vessel diameters and on the haematocrit of the parent vessel(Pries et al., 1989). Amore recent version gives consistent behaviourunder a wider range of conditions (Pries and Secomb, 2005).
The relationships described above provide a basis for predictingthe steady distributions of velocity, flow rate, wall shear stress,pressure and haematocrit in microvascular networks with knowntopology and geometry subject to suitable boundary conditions. Forthis analysis, the network can be considered as a set of inter-connected resistances. At each node in the network, conservation offlow leads to a linear equation in the pressures at that node and theadjacent nodes. This system can be solved efficiently for largenetworks using successive over-relaxation (Pries et al., 1990). If thenon-uniform partitioning of haematocrit at bifurcations is included,a further iterative procedure is required in which haematocrits areupdated based on current flows and the updated values are used torecompute flow resistance in each segment. Computation ofunsteady haematocrit distributions in microvascular networks ismore demanding, requiring the solution of coupled systems ofhyperbolic PDEs (Pop et al., 2007).
As described above, empirical equations provide an adequatebasis for simulating blood flow in microvascular networks.However, some limitations should be noted. The equations arebased mainly on experimental observations of blood flow in themesentery, a tissue which provides exceptionally clear visibility ofmicrovascular networks. Corresponding data for the heart are notcurrently available. While the flow behaviour of RBCs is presumablyindependent of tissue type, the ESL has not been studied in themyocardial microcirculation. In principle, the rheological behaviourof blood in microvessels should be predictable, but this will requirebetter understanding of the intrinsic mechanical properties of RBCsand their interaction with the plasma and each other at the hae-matocrits typical of microvessels. For single-file flow in capillary-sized tubes, this has been achieved (Secomb, 1987). However, thecomputational analysis of multiple interacting RBCs remains a topicof current research (Zhang et al., 2009).
The approach outlined above requires an explicit description ofnetwork geometry and topology including the smallest vessels. Asdiscussed in Section 2.1, gathering such information is challenging,not least since a complete description of the heterogeneous andanisotropic coronary microcirculation is still outstanding. Tech-niques to predict relevant flow and mass transport parametersbased on statistical properties of network structure could bevaluable in such cases. Homogenisation theory can be used toestimate tissue-scale flow and transport properties, based onexplicit solutions for flow and transport in smaller-scale subunits(Shipley and Chapman, 2010). When applied to the microcircula-tion, this approach faces difficult challenges because structural andfunctional parameters in the microcirculation show large hetero-geneity on a wide range of spatial scales (see Section 9.1).
4.2.1. ChallengesSeveral other aspects of microvascular flow should receive
future attention. As already mentioned, unsteady effects may berelevant in myocardial microcirculation (Westerhof et al., 2006).Further analysis of unsteady flow in coronary arterioles is needed toclarify the boundary conditions on simulations of coronary arterial
flow. The major function of the microcirculation is mass transport,and flow models must be linked to transport models. Regulation ofblood flow occurs mainly in the microcirculation, and both flowregulation and changes in vascular structure have fundamentaleffects on cardiac function (see Section 6 and 7 for a detaileddiscussion).
4.3. 3D flow modelling in larger vessels
Despite the challenges presented when studying high-Reynoldsnumber flows in large coronary arteries, it is important to be able toexplore the influence of detailed 3D flow features on the biologicalresponse of the arterial wall, mediated for example by shear stresstransduction or uptake at the arterial wall of transported speciessuch as low-density lipoproteins (LDL), because of the focal natureof coronary artery disease, and even the athero-susceptibility ofdisease in the physiological state (see Section 8.2). It is necessary tobe able to couple large-scale flow and luminal transport simula-tions with more detailed models of flow and transport propertiesclose to the arterial wall, accounting for example for the localtopography of the endothelium (Van Doormaal et al., 2009) (shownto lead to heterogeneous mass transfer over an individual cell) andthe detailed structure of the endothelial glycocalyx layer (Vincentet al., 2008) (where the fluctuating particulate nature of bloodwill be apparent and through which adhesive interactions betweenblood-borne cells and endothelium may take place). It is alsoimportant to integrate flow models with detailed biophysicalmodels for transport and mixing of species such as NO (Tsoukias,2008).
The ability to simulate flow and the associated stresses in largervessels with anatomically accurate geometry has seen a significantadvance over the past decade. This technology has been aided bythe availability of medical imaging techniques, e.g. MRI and CT, andsuitable computational flow modelling tools originally developedfor other disciplines such as the aeronautical industry. The simu-lation tools and computational resources now exist to simulatepulsatile, Newtonian flow in coronary arteries under a rigid wallassumption provided vessel anatomy and appropriate boundaryconditions are available. However obtaining vessel anatomy andboundary conditions is a challenging task both from an imagingand modelling perspective (Steinman, 2002). Non-Newtonianmodels are also available: however, while methods for incorpo-rating non-Newtonian models into large vessel simulations in thesystemic circulation are well developed (Leuprecht and Perktold,2001), the challenge is to determine and develop the appropriatenon-Newtonian models for complex flow environments. It is alsopossible to introduce prescribed wall motion into flow simulationsif this motion is known a priori (see Section 2.3 for details of currentdynamic imaging methods). Alternatively the technology exists tomodel fluidestructure interaction within large vessels where theblood flow and wall motion are dynamically coupled. This type ofmodelling, however, is significantly more challenging and a keylimitation to such an approach is our currently poor understandingof how the vessel is constrained by the adventitia and surroundingtissues. Furthermore, such an approach requires an accurate defi-nition of the vessel wall geometry and the wall structuralproperties.
It is often argued that vessel geometry and flow pulsatility arethe primarily determinants of the flow features and their associatedtemporal and spatial stresses within large vessels. The cycliccurvature deformations of coronary vessels have a strong effect onthe distribution of shear stress (Lynch et al., 1996; Waters andPedley, 1999; Van Meerveld and Waters, 2001; Moore et al.,2001; Weydahl and Moore, 2001). Effects such as non-Newtonianproperties and the luminal displacement in coronary arteries are

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 59
believed to have a secondary affect on the flow structure typicallychanging the magnitude rather than the character of the flow. Thekey determinants of accurate flow modelling therefore lie in theability to obtain and reconstruct accurate anatomic data andboundary conditions. Whilst the development of imaging protocolsare being advanced (see, for example, Section 2.3), the ability toreconstruct this data has been actively researched and there nowexist both commercial tools, such as Amira, and open access soft-ware such as the imaging tool kit (ITK) (http://www.itk.org) andvascular modelling tool kit (VMTK) (http://www.vmtk.org).
As well as providing insight into flows under physiological andpathological conditions, 3D flow modelling is essential to under-stand the nature of flows that arise as a result of clinical inter-vention, e.g. coronary catheterisation which uses catheters to treatcomplications like angina, atherosclerosis and heart attack (Ganzet al., 1985; Doucette et al., 1992; Di Mario et al., 1995). To accu-rately assess the influence of catheters on in vivo flow conditions, itis essential to understand arterial flow and pressure variation alongvessels when a catheter is present (Back et al., 1996; Bjorno andPattersson, 1976a, 1976b). Analytical models, coupled with in vitroexperimental evidence, have been developed to estimate the meanresistance increase due to the presence of the catheter (Back, 1994;Back et al., 1996). These studies have been extended to account formore realistic features such as artery curvature, degree of vesselstenosis, flow pulsatility and catheter flexibility (Jayaraman andDash, 2001; Dash et al., 1996, 1999; Jayaraman and Tiwari, 1995;Sarkar and Jayaraman, 1998, 2001). For translating the models astools for clinical use, modifications need to be included thatconsider the length, shape and size of the stenosis, catheter size andpositioning, coronary artery wall properties, etc. Inclusion of allthese complexities requires the use of 3D models and computa-tional fluid dynamics.
3D flow modelling can also be used to further our developmentand understanding of techniques for clinical measurements. Forexample, highly flexible sensor-equipped guide wires witha diameter of only 0.3 mm are now used relatively routinely tomeasure velocity and/or pressure dynamics in the larger coronaryvessels (Siebes et al., 2004). The effect of these wires on coronaryflow or pressure is small, especially when the eccentric location ofthe guide wire within the vessel is taken into account (Sinha Royet al., 2006; Verberne et al., 2007; Banerjee et al., 2008). Ata research level, it is also possible to obtain dynamic velocityprofiles in small epi- and endocardial vessels (Chilian and Marcus,1982; Toyota et al., 2005).
4.3.1. ChallengesThe large amount of numerical data generated from three-
dimensional unsteady flow simulations must be processed atdifferent levels of detail for different audiences. Maps of quantitiesof biological significance (such as wall shear stress distributions orwall uptake of a transported species) may be of interest to a clinicalaudience, while other quantities (for example cross-sectionallyintegrated pressure and flow distributions) must be considered ifa 3D model of a stenosis or bifurcation is to be integrated with a 1Dmodel of an arterial network (see Section 4.4). It is likely thata variety of other quantities e yet to be identified e will need to beinterrogated as new biological mechanisms are identified and newgenerations of mathematical models are developed to understandthem. The ability to generate 3D data also enables the researchcommunity to interact with asymptotic models of flow in deformedvessels where (if the underlying assumptions of these models arecorroborated) a better understanding of the important dynamics ofthe problem will ensue (Waters and Pedley, 1999).
For clinical applications, these models ultimately should incor-porate the altered material properties, haemodynamic
environment and pathophysiological conditions associated withvarious disease processes of the coronary arterial wall, as well asmicrovascular dysfunction, valvular disease and cardiomyopathiesthat affect coronaryecardiac interaction (Garcia et al., 2009;Heusch et al., 2009; Weinberg et al., in press). This will require anintegrated concerted effort of basic and clinical scientists fromvarious disciplines in order to provide experimental evidence tosupport model development or corroborate the predictions.
Such models could then be used to optimize treatment that istailored to a specific patient, pinpoint stenoses that require inter-vention, predict outcomes of minimally invasive surgical inter-ventions or optimize application of intravascular and ventricularassist devices (Moore et al., in press; Remmelink et al., 2007; Kolyvaet al., 2010; Ponzini et al., 2008; Voitl et al., 2009).
4.4. 1D pulse-wave modelling
As discussed in Section 3.5, in the large arteries pressure andflow distributions can be considered as summations of travellingpulse waves along the vascular network. As the pulse wavelengthsare (at least) three orders of magnitude larger than arterial diam-eters in normal conditions, it is reasonable to use the so-called longwave approximation. This approximation leads to the 1D governingequations for an incompressible fluid filling a compliant vessel afterintegration of the incompressible NaviereStokes equations overa generic cross section of a cylindrical domain (Canic and Kim,2003; Quarteroni and Formaggia, 2004; Smith et al., 2002). The1D equations also follow from direct application of conservation ofmass and momentum to a compliant 1D control volume ofincompressible fluid (Peiró and Veneziani, 2009; Sherwin et al.,2003). The 1D formulation has been satisfactorily tested bycomparison against in vivo (Olufsen et al., 2000; Reymond et al.,2009; Steele et al., 2003) and in vitro (Bessems et al., 2008;Matthys et al., 2007) data in large non-coronary systemic arteries.
In larger conduit arteries, reduced modelling provides valuableinsight into the development of methods for the diagnosis ofdisease (Alastruey et al., 2009a, 2009c; Franke et al., 2003;Stergiopulos et al., 1992) and the identification of anatomicalvariations (Alastruey et al., 2006, 2007) by wave analysis. Addi-tionally, it is used to improve the accuracy of the boundary condi-tions in 3D simulations of localised areas of the vasculature(Formaggia et al., 2001; Papadakis, 2009; Passerini et al., 2009;Urquiza et al., 2006). Huo and Kassab (2007), Smith et al. (2002)and Zamir (1998) applied 1D modelling to simulate pulse-wavepropagation in the coronary arteries. However, they did not accountfor the external pressure produced by the myocardium contraction(Krams et al., 1989b; Spaan et al., 1981), which was simulated in thereduced models by Guiot et al. (1990), Mynard and Nithiarasu(2008), Rammos et al. (1998). Mynard and Nithiarasu (2008) alsocoupled their coronary 1D model to a 1D model of the non-coro-nary systemic arteries through a lumped parameter model of flowin the left ventricle and aortic valve.
Any 1D model has to be truncated after a relatively smallnumber of generations of bifurcations. The downstream vascula-ture is usually simulated using lumped parameter models(Alastruey et al., 2008) or structured tree models (Olufsen et al.,2000) relating pressure to the flow at the outflow of each 1Dmodel terminal artery. Estimation of the parameters of theseterminal models can benefit from available morphometric data(Section 2). Lumped parameter models have also been used tosimulate coronary blood flow (e.g. Bruinsma et al., 1988; Scheelet al., 1989; Bovendeerd et al., 2006; Jacobs et al., 2008; Algranatiet al., 2010).
The 1D formulation has also been used to derivemethods for theanalysis of in vivo pressure and velocity measurements in both the
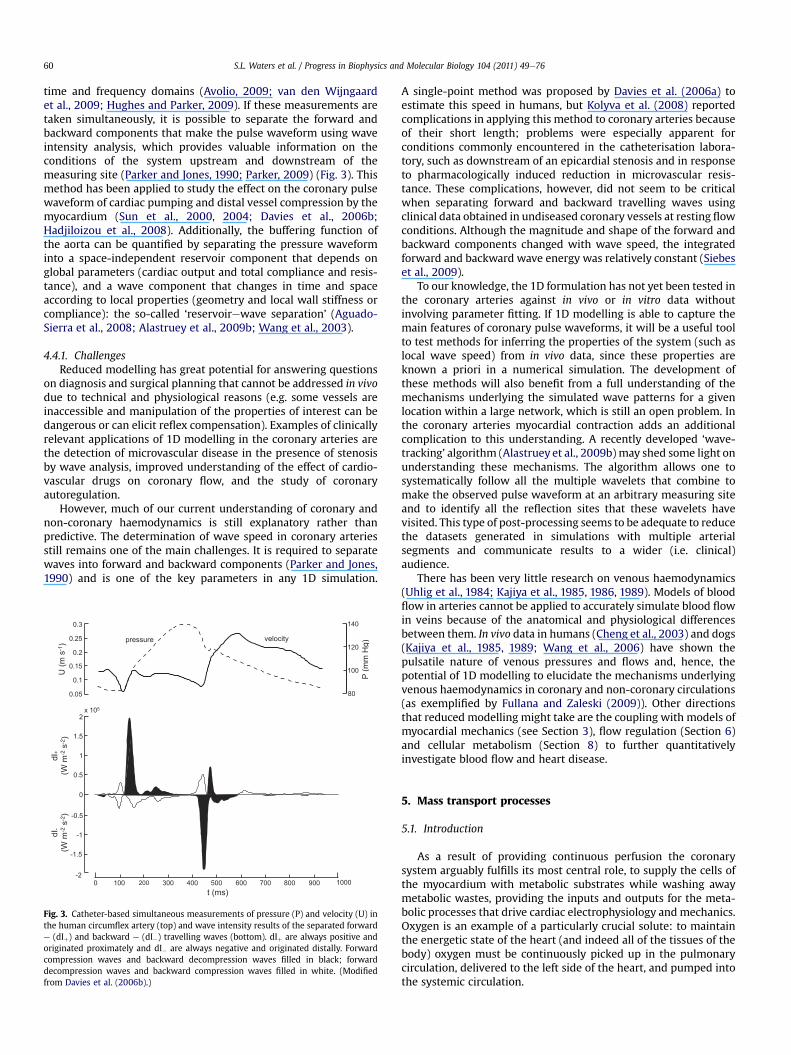
S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7660
time and frequency domains (Avolio, 2009; van den Wijngaardet al., 2009; Hughes and Parker, 2009). If these measurements aretaken simultaneously, it is possible to separate the forward andbackward components that make the pulse waveform using waveintensity analysis, which provides valuable information on theconditions of the system upstream and downstream of themeasuring site (Parker and Jones, 1990; Parker, 2009) (Fig. 3). Thismethod has been applied to study the effect on the coronary pulsewaveform of cardiac pumping and distal vessel compression by themyocardium (Sun et al., 2000, 2004; Davies et al., 2006b;Hadjiloizou et al., 2008). Additionally, the buffering function ofthe aorta can be quantified by separating the pressure waveforminto a space-independent reservoir component that depends onglobal parameters (cardiac output and total compliance and resis-tance), and a wave component that changes in time and spaceaccording to local properties (geometry and local wall stiffness orcompliance): the so-called ‘reservoirewave separation’ (Aguado-Sierra et al., 2008; Alastruey et al., 2009b; Wang et al., 2003).
4.4.1. ChallengesReduced modelling has great potential for answering questions
on diagnosis and surgical planning that cannot be addressed in vivodue to technical and physiological reasons (e.g. some vessels areinaccessible and manipulation of the properties of interest can bedangerous or can elicit reflex compensation). Examples of clinicallyrelevant applications of 1D modelling in the coronary arteries arethe detection of microvascular disease in the presence of stenosisby wave analysis, improved understanding of the effect of cardio-vascular drugs on coronary flow, and the study of coronaryautoregulation.
However, much of our current understanding of coronary andnon-coronary haemodynamics is still explanatory rather thanpredictive. The determination of wave speed in coronary arteriesstill remains one of the main challenges. It is required to separatewaves into forward and backward components (Parker and Jones,1990) and is one of the key parameters in any 1D simulation.
Fig. 3. Catheter-based simultaneous measurements of pressure (P) and velocity (U) inthe human circumflex artery (top) and wave intensity results of the separated forwarde (dIþ) and backward e (dI�) travelling waves (bottom). dIþ are always positive andoriginated proximately and dI� are always negative and originated distally. Forwardcompression waves and backward decompression waves filled in black; forwarddecompression waves and backward compression waves filled in white. (Modifiedfrom Davies et al. (2006b).)
A single-point method was proposed by Davies et al. (2006a) toestimate this speed in humans, but Kolyva et al. (2008) reportedcomplications in applying this method to coronary arteries becauseof their short length; problems were especially apparent forconditions commonly encountered in the catheterisation labora-tory, such as downstream of an epicardial stenosis and in responseto pharmacologically induced reduction in microvascular resis-tance. These complications, however, did not seem to be criticalwhen separating forward and backward travelling waves usingclinical data obtained in undiseased coronary vessels at resting flowconditions. Although the magnitude and shape of the forward andbackward components changed with wave speed, the integratedforward and backward wave energy was relatively constant (Siebeset al., 2009).
To our knowledge, the 1D formulation has not yet been tested inthe coronary arteries against in vivo or in vitro data withoutinvolving parameter fitting. If 1D modelling is able to capture themain features of coronary pulse waveforms, it will be a useful toolto test methods for inferring the properties of the system (such aslocal wave speed) from in vivo data, since these properties areknown a priori in a numerical simulation. The development ofthese methods will also benefit from a full understanding of themechanisms underlying the simulated wave patterns for a givenlocation within a large network, which is still an open problem. Inthe coronary arteries myocardial contraction adds an additionalcomplication to this understanding. A recently developed ‘wave-tracking’ algorithm (Alastruey et al., 2009b) may shed some light onunderstanding these mechanisms. The algorithm allows one tosystematically follow all the multiple wavelets that combine tomake the observed pulse waveform at an arbitrary measuring siteand to identify all the reflection sites that these wavelets havevisited. This type of post-processing seems to be adequate to reducethe datasets generated in simulations with multiple arterialsegments and communicate results to a wider (i.e. clinical)audience.
There has been very little research on venous haemodynamics(Uhlig et al., 1984; Kajiya et al., 1985, 1986, 1989). Models of bloodflow in arteries cannot be applied to accurately simulate blood flowin veins because of the anatomical and physiological differencesbetween them. In vivo data in humans (Cheng et al., 2003) and dogs(Kajiya et al., 1985, 1989; Wang et al., 2006) have shown thepulsatile nature of venous pressures and flows and, hence, thepotential of 1D modelling to elucidate the mechanisms underlyingvenous haemodynamics in coronary and non-coronary circulations(as exemplified by Fullana and Zaleski (2009)). Other directionsthat reduced modelling might take are the coupling with models ofmyocardial mechanics (see Section 3), flow regulation (Section 6)and cellular metabolism (Section 8) to further quantitativelyinvestigate blood flow and heart disease.
5. Mass transport processes
5.1. Introduction
As a result of providing continuous perfusion the coronarysystem arguably fulfills its most central role, to supply the cells ofthe myocardium with metabolic substrates while washing awaymetabolic wastes, providing the inputs and outputs for the meta-bolic processes that drive cardiac electrophysiology andmechanics.Oxygen is an example of a particularly crucial solute: to maintainthe energetic state of the heart (and indeed all of the tissues of thebody) oxygen must be continuously picked up in the pulmonarycirculation, delivered to the left side of the heart, and pumped intothe systemic circulation.
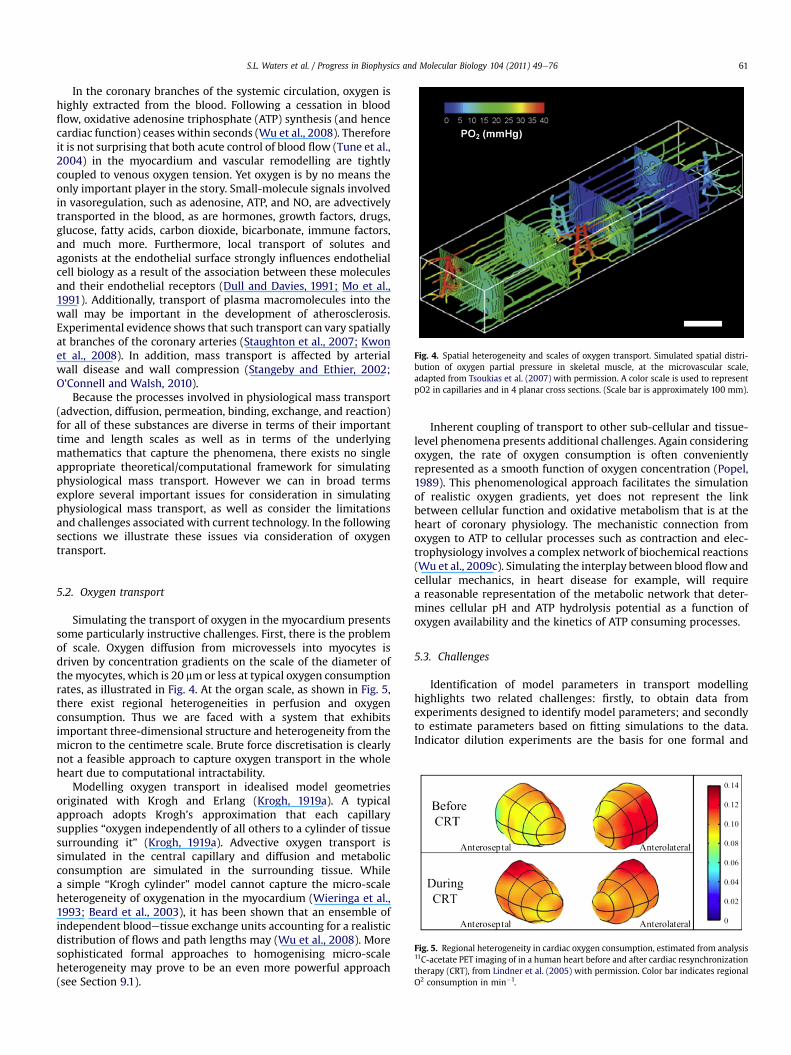
Fig. 4. Spatial heterogeneity and scales of oxygen transport. Simulated spatial distri-bution of oxygen partial pressure in skeletal muscle, at the microvascular scale,adapted from Tsoukias et al. (2007) with permission. A color scale is used to representpO2 in capillaries and in 4 planar cross sections. (Scale bar is approximately 100 mm).
S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 61
In the coronary branches of the systemic circulation, oxygen ishighly extracted from the blood. Following a cessation in bloodflow, oxidative adenosine triphosphate (ATP) synthesis (and hencecardiac function) ceases within seconds (Wu et al., 2008). Thereforeit is not surprising that both acute control of blood flow (Tune et al.,2004) in the myocardium and vascular remodelling are tightlycoupled to venous oxygen tension. Yet oxygen is by no means theonly important player in the story. Small-molecule signals involvedin vasoregulation, such as adenosine, ATP, and NO, are advectivelytransported in the blood, as are hormones, growth factors, drugs,glucose, fatty acids, carbon dioxide, bicarbonate, immune factors,and much more. Furthermore, local transport of solutes andagonists at the endothelial surface strongly influences endothelialcell biology as a result of the association between these moleculesand their endothelial receptors (Dull and Davies, 1991; Mo et al.,1991). Additionally, transport of plasma macromolecules into thewall may be important in the development of atherosclerosis.Experimental evidence shows that such transport can vary spatiallyat branches of the coronary arteries (Staughton et al., 2007; Kwonet al., 2008). In addition, mass transport is affected by arterialwall disease and wall compression (Stangeby and Ethier, 2002;O’Connell and Walsh, 2010).
Because the processes involved in physiological mass transport(advection, diffusion, permeation, binding, exchange, and reaction)for all of these substances are diverse in terms of their importanttime and length scales as well as in terms of the underlyingmathematics that capture the phenomena, there exists no singleappropriate theoretical/computational framework for simulatingphysiological mass transport. However we can in broad termsexplore several important issues for consideration in simulatingphysiological mass transport, as well as consider the limitationsand challenges associated with current technology. In the followingsections we illustrate these issues via consideration of oxygentransport.
Fig. 5. Regional heterogeneity in cardiac oxygen consumption, estimated from analysis11C-acetate PET imaging of in a human heart before and after cardiac resynchronizationtherapy (CRT), from Lindner et al. (2005) with permission. Color bar indicates regionalO2 consumption in min�1.
5.2. Oxygen transport
Simulating the transport of oxygen in the myocardium presentssome particularly instructive challenges. First, there is the problemof scale. Oxygen diffusion from microvessels into myocytes isdriven by concentration gradients on the scale of the diameter ofthemyocytes, which is 20 mmor less at typical oxygen consumptionrates, as illustrated in Fig. 4. At the organ scale, as shown in Fig. 5,there exist regional heterogeneities in perfusion and oxygenconsumption. Thus we are faced with a system that exhibitsimportant three-dimensional structure and heterogeneity from themicron to the centimetre scale. Brute force discretisation is clearlynot a feasible approach to capture oxygen transport in the wholeheart due to computational intractability.
Modelling oxygen transport in idealised model geometriesoriginated with Krogh and Erlang (Krogh, 1919a). A typicalapproach adopts Krogh’s approximation that each capillarysupplies “oxygen independently of all others to a cylinder of tissuesurrounding it” (Krogh, 1919a). Advective oxygen transport issimulated in the central capillary and diffusion and metabolicconsumption are simulated in the surrounding tissue. Whilea simple “Krogh cylinder” model cannot capture the micro-scaleheterogeneity of oxygenation in the myocardium (Wieringa et al.,1993; Beard et al., 2003), it has been shown that an ensemble ofindependent bloodetissue exchange units accounting for a realisticdistribution of flows and path lengths may (Wu et al., 2008). Moresophisticated formal approaches to homogenising micro-scaleheterogeneity may prove to be an even more powerful approach(see Section 9.1).
Inherent coupling of transport to other sub-cellular and tissue-level phenomena presents additional challenges. Again consideringoxygen, the rate of oxygen consumption is often convenientlyrepresented as a smooth function of oxygen concentration (Popel,1989). This phenomenological approach facilitates the simulationof realistic oxygen gradients, yet does not represent the linkbetween cellular function and oxidative metabolism that is at theheart of coronary physiology. The mechanistic connection fromoxygen to ATP to cellular processes such as contraction and elec-trophysiology involves a complex network of biochemical reactions(Wu et al., 2009c). Simulating the interplay between blood flowandcellular mechanics, in heart disease for example, will requirea reasonable representation of the metabolic network that deter-mines cellular pH and ATP hydrolysis potential as a function ofoxygen availability and the kinetics of ATP consuming processes.
5.3. Challenges
Identification of model parameters in transport modellinghighlights two related challenges: firstly, to obtain data fromexperiments designed to identify model parameters; and secondlyto estimate parameters based on fitting simulations to the data.Indicator dilution experiments are the basis for one formal and

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7662
powerful approach. The idea is to observe the outflow kinetics oftracer labelled substances following introduction of the labelledsubstances into an organ or tissue. Using multiple indicators, it ispossible to systematically identify a relatively large number ofparameters. For example, 5 parameters underlying intravasculardispersion of substances in the coronary vasculature in a blood-etissue transport model have been identified by fitting thewashout curve for an intravascular tracer (Kuikka et al., 1986).Parameters for permeation through endothelial clefts, acrossendothelial cell membranes, across myocyte membranes, as well asparameters involved in the kinetics of biochemical reactions maybe identified based on a series of washout curves for additionalsubstances. In all, more than 10 parameters may be estimated byexperiments with three or more tracers (Schwartz et al., 1999).
6. Regulation
6.1. Introduction
Key to efficient mass transport within the coronary circulation,tomeet the functional demands of the heart, is the regulation of thetransport processes outlined in Section 5. Under normal conditions,the rate of blood flow to tissue is regulated to meet metabolicrequirements, particularly oxygen consumption, but is relativelyunaffected by changes of arterial pressure. In human myocardium,the oxygen consumption rate varies over approximately a five-foldrange between rest and intense exercise, from about 0.1 to0.5 cm3O2/cm3/min (von Restorff et al., 1977). Perfusion increasesalmost in direct proportion to oxygen consumption over this range,and oxygen extraction, expressed relative to convective arterialinflow of oxygen, remains relatively constant in the range 70e80%.Thus, the heart exhibits remarkably efficient metabolic regulationof blood flow. Autoregulation of blood flow refers to the mainte-nance of almost constant flow in the face of changes in arterialpressure, and the normal myocardium is also very effective in thisregard. For example, perfusion is almost constant for variations ofarterial pressure in the range 60e140 mmHg (Johnson, 1980).
Local regulation of blood flow is achieved mainly through theactive contraction and dilation of small arteries and arterioles,while oxygen exchange occurs primarily in the capillaries.Much can be learned by studying blood flow to intact tissues andorgans, but a thorough understanding of flow regulation requiresconsideration of phenomena occurring at the level of themicrocirculation.
The challenge of understanding blood flow regulation has beenrecognised since the late nineteenth century (Gaskell, 1878). Overthe years, many components of flow regulation have been identi-fied and studied in depth. These include autonomic nervousstimuli, circulating substances, mechanical stimulation of vessels,myogenic responses, shear-dependent responses, metabolicresponses, conducted responses propagated along vessels, andcommunication between paired feeding and draining vessels. Suchstudies have generated a wealth of information on the structureand function of the microcirculation, and on the various mecha-nisms governing the contraction and dilation of arterioles. (SeeDavis et al. (2008) for an extensive review).
The ability of the coronary circulation to regulate blood flowaccording to spatially and temporally varying requirements isfundamental to normal cardiac function, and strongly influencesthe response of the heart to disease and other perturbations. Manydrugs aimed at improving cardiac function are vasoactive. Thedevelopment of integrated theoretical models, which will allowrealistic predictions of cardiac function and responses to variousconditions and treatments, must include a quantitative descriptionof blood flow regulation.
6.2. Modelling approaches
Progress in development of a quantitative theory of blood flowregulation has been relatively slow, despite the availability ofextensive biological data. Two major obstacles have slowed prog-ress. Firstly, key mechanisms involved in flow regulation remain tobe identified. Studies on the response of endothelial cells tochanges in wall shear stress have yielded much information aboutthe cellular processes involved, but the fundamental mechanismsby which mechanical stimuli are converted into chemical signalsremain unknown (Davies, 1995; Davies and Helmke, 2009).A similar situation holds with regard to the ability of vascularsmooth muscle to sense changes in tension or elongation (Davisand Hill, 1999). The mechanisms involved in metabolic controlhave likewise been difficult to identify (Tune et al., 2004). The lackof specific mechanistic information does not prevent the develop-ment of models based on phenomenological assumptions.However, as more mechanisms are identified and describedquantitatively, it will be possible to develop more realistic andreliable models.
A second major obstacle to progress is that the system involvesnumerous components that are coupled and interact on severallevels, making it difficult to isolate and study the componentsseparately. The circulatory system can be regarded as a network ofinterconnected variable resistances, and changes in the resistanceof any one element effect flows and pressures in all segmentsconnected in series or in parallel. Such changes alter the distri-bution of intravascular pressure and wall shear stress, which thenevoke active responses in the cells forming the vessel walls,further altering the distribution of resistance. Information aboutmetabolic and haemodynamic conditions is transferred betweentissue and vessels and among vessels by several mechanisms(Secomb and Pries, 2002). More than twenty years ago, it wasstated that “A significant remaining challenge is that of inte-grating observations on small and large vessels into a generalpicture of flow control” (Duling et al., 1987), and to a large extentthis is still true.
A logical response to this complexity is to develop theoreticalmodels for phenomena occurring on the scale of microvessels, andto use these models as a framework for integrating information onthe underlying processes and thereby predicting larger-scalebehaviour. We have already seen that the value of this approachwas recognised by August Krogh, who together with the mathe-matician Erlang, developed the classic Krogh cylinder model foroxygen transport to tissue (Krogh, 1919a, 1919b) in order to explorethe relationship between the number of capillaries in a tissue andthe rate at which oxygen can be delivered (refer also to Section 5).
Theoretical models for blood flow regulation were reviewedrecently (Secomb, 2008), and some key points from that reviewwillbe reiterated here. According to Poiseuille’s law, blood flow isproportional to the fourth power of diameter if the pressure dropand other quantities are held fixed. This implies that large changesin flow can be achieved by moderate changes in diameter, and alsothat control of blood flow requires relatively precise control ofvessel diameters. The control of blood flow is primarily achieved bycontraction and relaxation of vascular smooth muscle (VSM) invessel walls. Many factors influence the level of tone generated bya VSM cell, including its length and tension, local levels of metab-olites and signalling molecules, communications with other cellsvia gap junctions, and neural inputs.
Ideally, theoretical simulations of flow regulation should bebased on an integrated model for the responses of an individualVSM to all these stimuli. A comprehensive model of this type is notavailable, but some aspects of VSM responses have been consideredtheoretically (Gonzalez-Fernandez and Ermentrout, 1994; Yang

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 63
et al., 2003a, 2003b, 2005). The generation of nitric oxide and othersubstances by endothelial cells strongly influences VSM tone, andhas been modelled theoretically (Chen and Popel, 2006). As yet,work using this approach is at a relatively early stage.
An alternative approach is to develop models to describe thevasoactive behaviour of arteriolar segments (Miller et al., 1997).Some aspects of this behaviour have been well studied inpreparations of isolated perfused arterioles. In particular, themyogenic response of VSM to changes in vessel wall tension hasbeen extensively studied (Johnson, 1980), and several theorieshave been developed (Secomb, 2008). Increased wall shear stresscauses vasodilation of blood vessels (Pohl et al., 2000), butrelatively few theoretical models have included this effect(Carlson et al., 2008; Cornelissen et al., 2002; Fung, 1997; Liaoand Kuo, 1997).
Recently, a potential mechanism of metabolic flow regulationhas been described, based on the release of ATP by red blood cells ata rate that depends on haemoglobin saturation in the red cell.According to this mechanism, the red blood cell acts as botha carrier of oxygen and a sensor of oxygen levels (Ellsworth, 2000).Application of ATP to venules has been shown to stimulateupstream conducted responses along vessel walls via the capil-laries, leading to arteriolar vasodilation (Collins et al., 1998). Thecombination of saturation-dependent ATP release and conductedresponses initiated by ATP provides a potential mechanism formetabolic regulation of cardiac perfusion (Deussen et al., 2006),although other mechanisms may also be involved (Duncker andBache, 2008). In a recent theoretical model for metabolic flowregulation and autoregulation based on this mechanism (Arcieroet al., 2008; Carlson et al., 2008), a simplified vascular networkconsisting of seven representative segments is considered, of whichtwo segments, representing the arterioles, are assumed to bevasoactive. Length-tension characteristics of VSM are explicitlyconsidered. The total tension in the vessel wall is expressed as thesum of active and passive components. The active component isproportional to the activation which varies between 0 and 1 andhas a sigmoidal dependence on the sum of the stimuli acting tocontrol the tone of VSM, including wall tension, wall shear stressand a metabolic signal. With this model, predicted metabolicregulation and autoregulatory behaviour agree well with experi-mental data from skeletal muscle and other tissues. The model hasnot yet been tested using data from the coronary circulation.
6.3. Challenges
Despite the availability of increasing amounts of biological dataon individual mechanisms, the key challenge that remains is todevelop an integrated understanding of flow regulation. Obstacleshave been the lack ofmechanistic information on key processes andthe complexity of the system. Experimental results obtained duringthe past decade have suggested a mechanism for metabolic flowregulation in response to blood oxygen levels in venules. As moremechanisms are identified and described quantitatively, the chal-lenge will be to develop more realistic and reliable models. Suchmodels should take into account the intrinsic heterogeneity in thestructure of the microcirculation, which causes non-uniform flowdistribution. Local flow regulation must result in close matching offlow to metabolic demands despite this heterogeneity (Walley,1996). If supply and demand are not matched, oxygen extractionfrom blood is reduced and regional hypoxia may occur. A furtherchallenge in modelling flow regulation is to integrate the intrinsicmechanisms discussed above with the effects of extrinsic factors,including the effects of the autonomic nervous system and ofcirculating hormones with vasoactive properties such asadrenaline.
7. Angiogenesis and vascular remodelling
7.1. Introduction
On a significantly longer time scale than the regulatory mech-anisms discussed in Section 6, the locations of the major coronaryvessels are determined by development and remodelling mecha-nisms, During early development vascular structure is defined bygenetic programming. As the heart grows, the development of thevasculature, and especially the large number of vessels forming themicrocirculation, is increasingly determined by local responses toa number of feedback signals, including levels of metabolites andgrowth factors, andmechanical stimuli generated by the blood flowitself (Pries and Secomb, 2008). Even in adults, the vasculature isa dynamic structure, capable of generating new vessels, remodel-ling existing vessels, and regressing according to changing physi-ological and pathophysiological conditions (Carmeliet, 2005). Forexample, the growth of myocardium in response to increasedworkload must be accompanied by increases in the numbers ofmicrovessels and in the diameters of the major vessels. If someblood vessels are blocked due to atherosclerosis or thrombosis, theoutcomemay depend on the ability of remaining vessels to provideblood flow through alternative pathways, forming collaterals(Schaper, 2009).
Improved understanding and quantitative simulation of angio-genesis (the growth of new blood vessels from pre-existingmicrovasculature) and vascular remodelling (structural changes ofexisting vessels) including arteriogenesis contributes in manyimportant ways. Quantitative information on the structure of thevasculature is needed as a basis for analysing the haemodynamics ofcoronary circulation, and also for predicting mass transport char-acteristics. A number of experimental techniques for determiningnetwork structure have been developed (Cassot et al., 2006;Jorgensen et al., 1998; Kassab et al., 1993; Sands et al., 2005): seealso the detailed discussion of vascular casting (Section 2.2) andstructural imaging of coronary vasculature (in Section 2.3) Withthese techniques, increasingly detailed maps of vascular structurehave been obtained, extending to themicrovascular level. However,such studies are difficult and laborious, and are specific to a partic-ular tissue sample. Their value and generality could be enhancedwith better understanding of the principles and processes under-lying the development of vascular structures. Furthermore, topredict the outcome of diseases and the effects of treatments in thecoronary circulation, it will be necessary to simulate structuralresponses of the vasculature to perturbations and to surgical orpharmacological interventions. For example, with the growingclinical importance of anti-angiogenic drugs comes the need tobetter understand the cardiovascular side effects of such drugs.
Numerous biochemical and physical factors govern angiogen-esis and vascular remodelling. Vascular remodelling includesprocesses of arteriogenesis and collateral growth important forcoronary circulation. Among the biochemical factors are severalfamilies of growth factors that stimulate vascular growth: vascularendothelial growth factors (VEGF), fibroblast growth factors (FGF),hepatocyte growth factors (HGF), angiopoietins (Ang), platelet-derived growth factor (PDGF), and transforming growth factor(TGF-b). Other protein families play an important role in angio-genesis and vascular remodelling, e.g. matrix metalloproteinases(MMPs) that have the capacity to proteolyze the extracellularmatrix (ECM) and enable new or existing vessels to grow. Hundredsof other molecules have been identified as important participantsin the processes of angiogenesis and vascular remodelling. Thus,developing a comprehensive set of computational models at themolecular level describing even the major factors and their inter-actions is challenging.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7664
Physical forces also play an important role in both angiogenesisand vascular remodelling. The physical forces within the vascularwall include time-dependent shear and pressure forces exerted bythe flowing blood, and rhythmic stretching and compressing of thevasculature by the contracting myocardium. Fluid mechanicalforces acting on the endothelium are transduced and result incomplex signalling cascades in the endothelium and smoothmuscle (Valentin et al., 2009; Valentin and Humphrey, 2009a,2009b). Experiments demonstrate that physical forces differen-tially affect vascular growth; e.g. elevated static stretching leads tosprouting angiogenesis whereas shear forces lead to splittingangiogenesis (splitting of existing capillary vessels into two)(Hudlicka and Brown, 2009).
Vessels can grow by proliferation of vascular cells and secretionof extracellular matrix components, and by either incorporation ofcirculating endothelial progenitor cells (EPCs) (Jujo et al., 2008) orrecruitment of stromal cells that differentiate into vascular peri-cytes or smooth muscle cells upon contacting the vessel (Peirceet al., 2004).
7.2. Angiogenesis
Coronary angiogenesis may be a normal physiological process(e.g. as a result of exercise) or pathological (e.g. in a developingatherosclerotic plaque). In many cases, including diseases ofischemic heart and skeletal muscle, angiogenesis occurs inresponse to hypoxia. A transcription factor, hypoxia-induciblefactor, HIF1a, acts as an oxygen sensor whose activity leads toupregulation of VEGF secretion from myocytes and stromal cells(Semenza, 2004) that diffuse through the interstitial space and canbind to the endothelial cell receptors VEGFR1 and VEGFR2, initi-ating a signal transduction cascade that leads to endothelial cellproliferation and migration. The production of matrix metal-loproteinases, MMPs, by the endothelial cells is necessary forselective proteolysis of the capillary basement membrane and theECM, which constitute physical barriers to capillary sprouting. Inaddition, MMPs release growth factors sequestered in the ECM;they may also uncover cryptic sites of the ECM proteins, a numberof which have been identified as anti-angiogenic. Once the catalyticcascade is initiated, a single endothelial cell breaks through thebasement membrane and invades the ECM, serving as the tip of anemerging capillary sprout that is followed by proliferating “stalk”ECs (Gerhardt et al., 2003). VEGF also serves as a chemoattractantfor the nascent sprout. Multiple capillary sprouts may interconnectleading to the formation of a neovascular plexus. Many of thesprouts will retract; the surviving sprouts undergo maturation andremodelling by recruiting precursors of supporting vascular cells,pericytes and smooth muscle cells, via platelet-derived growthfactor, PDGF, secreted by endothelial cells. When the precursor cellsmake contact with the nascent sprout, transforming growth factorTGF-b acts by suppressing proliferation and migration of theendothelial cell and stimulating differentiation of the precursorcells into pericytes and smooth muscle cells.
7.3. Role of vascular endothelial growth factor familyin angiogenesis
The discovery of VEGF-A has led to an exponential growth ofpublications on angiogenesis and VEGF. Currently, six humanalternate splice-isoforms of VEGF-A have been identified: VEGF121,VEGF145, VEGF165, VEGF183, VEGF189, and VEGF209. Binding ofligands to the VEGF-receptors on the surface of endothelial cellsoriginates signal transduction cascades leading to receptor phos-phorylation and the resulting increase of vascular permeability, ECcell proliferation, migration, protease secretion and stabilisation of
neovessels. The VEGF family members are critical angiogenicgrowth factors, and are expressed in cardiac muscle, yet the preciserole that each of these factors plays is not completely understood.VEGF is necessary for the embryonic development of coronaryvessels. Increases in VEGF expression in muscle are linked to, andprecede, the increases in vascular density that occur when cardiacmuscle is faced with increases in functional demand. VEGF alsopromotes the activation of endothelial nitric oxide synthase (eNOS)and NO production. Activation of eNOS and generation of NO havebeen shown to be essential for angiogenesis, and NO productionmay be a critical step in the mobilisation of repair cells from thebone marrow, which can contribute to angiogenesis and vascularremodelling. Mathematical and computational models of VEGF-receptor interactions in vitro and in vivo have been formulated ina series of recent studies (Mac Gabhann and Popel, 2008; Stefaniniet al., 2008; Wu et al., 2009a, 2009b).
7.4. Role of mechanical factors in angiogenesis
These biochemically regulated pathways are in turn regulatedby mechanical forces which are a major determinant of angiogen-esis (Ingber, 2002; Milkiewicz et al., 2006; Shiu et al., 2005). Effectsof shear forces and stretching have been amply demonstrated at thelevel of endothelial cells. The forces affect gene expression andintra- and inter-cellular signalling. The interaction of the endo-thelial cell with its physical microenvironment, particularly theextracellular matrix, impacts the cell function. Mathematical andcomputational models of endothelial cell mechanics are underdevelopment (Ferko et al., 2007). These developments have yet tobe integrated into a molecular- and cellular-based model ofa microvessels and its growth.
7.5. Therapeutic angiogenesis in myocardial ischemic disease
Myocardial ischemia is in most cases a consequence of thecoronary artery disease; it can be ameliorated by increased bloodperfusion of the ischemic tissue. Increased blood perfusion requiresboth angiogenesis and vascular remodelling. Therapeutic angio-genesis is the induction of microvascular growth aimed at treatingpatients with inadequate tissue perfusion. Numerous clinical trialsfor different modes of therapeutic angiogenesis have been con-ducted in the last two decades; however, at present no therapiesare available to increase blood flow to the ischemic myocardium. Insome cases, transient increases of perfusion have been achieved,but over time the improvements are lost. Theoretical studiesaddressing some aspects of therapeutic angiogenesis are emerging(Mac Gabhann et al., in press-a, in press-b).
7.6. Computational models of angiogenesis
Computational and mathematical models of angiogenesis haverecently been reviewed (Peirce, 2008; Qutub et al., 2009a, 2009b).Most of the applications of published models deal with tumour-induced angiogenesis (Bauer et al., 2007; Macklin et al., 2009;Owen et al., 2009) and skeletal muscle (Mac Gabhann et al.,2007a, 2007b; Mac Gabhann and Popel, 2007; Peirce et al., 2004).Even though many of the model elements are similar in differentorgans and tissues, specific modelling of the important area ofcoronary angiogenesis awaits development. Models at the vesseland network levels consider sprouting angiogenesis; splittingangiogenesis and intussusception as models of angiogenesis havereceived little attention in theoretical studies.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 65
7.7. Structural adaptation
An early theoretical approach to the problem of structuraladaptation was based on an optimality principle (Murray, 1926). Asdiscussed in Section 2.5, the vascular system shows large deviationsfrom Murray’s law, and optimality principles, based on minimizingan energy cost, do not necessarily take into account the functionalrequirements and constraints that apply to the system (Renemanand Hoeks, 2008). Murray’s law suggests that the vascular systemcould achieve an optimum state by adjustments of vascular diam-eter in response to deviations of shear stress from a set point(Rodbard, 1975). According to this theory, an increase in blood flowin a given vessel triggers increased wall shear stress, leading toenlargement of the vessel, and a return of the wall shear stressto the set point value. Central to this model is the primary regula-tory function of the endothelium, the presence of which is essentialfor physiological acute vasoregulation (Pohl et al., 1986) and criticalfor (chronic) adaptive structural remodelling (Langille andO’Donnel, 1986).
However, vessels respond structurally to several stimuli besideswall shear stress. Theories including the effects of multiple stimulihave been developed (Pries et al., 1998, 2001, 2005), and recentlyreviewed (Pries and Secomb, 2008). In these theories, the diameterand wall thickness of each vessel in a network are treated asdynamic variables that change in response to several stimuliincluding wall shear stress, circumferential wall tension, and levelsof metabolites (Hacking et al., 1996). In particular, an increase inintravascular pressure leads to increased tension in the vessel wall.The structural response involves an increase in wall mass throughgrowth and proliferation of smooth muscle cells, and a decrease inthe internal diameter of the vessel. An increase in peripheralresistance is a key aspect in the development of hypertension, andreflects inward remodelling of the small arteries and arterioles(Mulvany, 2008). This structural response to increased pressureproduces a positive feedback mechanism, since an increase inperipheral resistance requires increased arterial pressure to main-tain flow. Such a mechanism may play an important role in theprogressive development of hypertension (Pries et al., 1999).Conversely, prolonged exposure to reduced pressure and low VSMtone, e.g. downstream of a coronary stenosis, may induce outwardremodelling (Li and Fung, 2002), which in turn gives rise toimpaired autoregulation when distending pressure is suddenlyrestored by treatment of the stenosis (Verhoeff et al., 2005).
Theories for structural adaptation must also include responsesto changing metabolic demands. For instance, chronic hypoxialeads to enlargement of existing vessels, and therefore to increasedblood flow. This response is also necessary to stabilise parallel flowpathways which otherwise lead to unstable behaviour (Hackinget al., 1996). A key element is the inclusion of effects of con-ducted responses that allow propagation of information abouttissue metabolic needs upstream to feeding vessels. Without con-ducted responses, the network is unable to appropriately balanceflows in long and short flow pathways (Pries et al., 1998).
7.8. Challenges
The theory for angiogenesis is not at the stage where theformation of microvascular networks can be reliably predicted.Progress has been made in predicting events at the molecular level,e.g. for the VEGF, FGF2, MMP, and HIF1a families, but much remainsto be done at both the molecular and cellular levels in describingthe effects of other important molecules as well as mechanicalforces. Above the cellular level, the theory of network formation hasyet to be developed; such a theory will use a combination of the
molecular and cellular information and experimentally-derivedrelationships.
The theories for structural adaptation described above aresuitable for inclusion in integrated network-level models for thelong-term remodelling of the coronary vasculature. For example, ifvascular network structure is known in sufficient detail, it ispossible in principle to predict the formation of collateral pathwaysfollowing the occlusion of an artery (Gruionu et al., 2005a).However, this approach presents several obstacles and challenges.Experimental information on the dynamics of structural adaptationis difficult to obtain, because experimental preparations allowingobservation of vascular structure cannot generally be maintainedfor periods longer than a few hours, with the exception of windowchamber preparations. The direct observation of long-term adap-tation in cardiac tissue is particularly challenging, except for largervessels that can be imaged with x-ray contrast media. The theo-retical models developed so far do not cover the full range ofphysiological and pathophysiological states. For instance, ischemiaand inflammation may evoke a different set of responses fromthose occurring in normal tissue (Gruionu et al., 2005b). In vesselswith active control of vascular smooth muscle tone, remodelling isclosely intertwined with regulation of blood flow (Bakker et al.,2008; Valentin and Humphrey, 2009b). Theories integrating thesetwo processes are at an early stage of development (Jacobsen et al.,2008). In the myocardium, vessels are subjected to a pulsatilemechanical environment unlike that in other tissues, and this mayinfluence remodelling (see Section 3). As yet, available theories donot explicitly link the dynamics of remodelling with effects occur-ring at the sub-cellular level. Such linking of theories acrossdifferent physical scales is a particular challenge here as in otheraspects of the Cardiac Physiome project (Bassingthwaighte et al.,2009).
8. Vascular cellular mechanics
The endothelial cell, located at the blood/vessel interface, hasmany critical functions which include the maintenance of anti-coagulant properties, the physiological control of lumen diameter(vasoregulation), the regulation of vascular permeability, andmediation of both pathological and protective responses associatedwith acute and chronic inflammation, wound healing, and majorcardiovascular disorders such as atherogenesis (Aird, 2007).Throughout the coronary vascular system the endothelial cell is keyto sensing the local fluid mechanical environment and transducingthe associated fluid mechanical forces into (bio)chemical signals.Here we focus on the response of the endothelial cell to the localflow environment, and describe how in some arterial sites the flowof non-Newtonian fluid through complex geometries that featureflow separation, oscillatory flow with a reversed component, etc,can initiate a series of events resulting in arterial disease.
8.1. The endothelial cell as a mechanotransducer
The endothelial cell is sensitive to fluid dynamical forces such asshear stress (acting in the direction of the flow) and pressure(acting normal to the wall). Endothelial sensitivity to shear stress isparticularly important because communication with underlyingsmooth muscle cells regulates their vasoactivity, migration, prolif-eration and synthetic rates. Early studies of endothelial mechano-transduction considered changes in endothelial cell morphologywhen cultured endothelial cells were first subjected to defined flow(Dewey et al., 1981; Levesque and Nerem, 1985). Many studiesquickly followed showing that endothelial signalling pathwaysrespond to changes in fluid shear stress (Davies et al., 1984; Olesenet al., 1988). Major reviews of endothelial mechanotransduction

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7666
include Davies (1995), Nerem et al. (1998), Shyy and Chien (2002),Resnick et al. (2003), Helmke (2005), Lehoux et al. (2006), Hahnand Schwartz (2009), and Davies and Helmke (2009). Flow influ-ences endothelial biology both by the direct action of haemody-namic deformation forces (principally due to shear stress) on theendothelium itself, and by modifying the local transport of solutesand agonists at the endothelial surface and hence the associationbetween these molecules and their endothelial receptors (Dullet al., 1992; Choi et al., 2007).
The luminal side of the endothelium is exposed to flowingblood, the cell edges are anchored to adjacent cells through junc-tional complexes, and the abluminal surface away from the flow isattached to a sub-cellular connective tissue matrix through highlyregulated transmembrane integrin proteins. The cell cytoskeleton isconnected to many cell membrane proteins, and in attached cellsexists in a state of intracellular tension or prestress (Ingber, 2008)that is changed by external loading, e.g. flow changes. A localdeformation of the luminal cell surface may be propagated near-instantaneously to distant cellular sites via membrane proteinsattached to the cytoskeleton and the cytoskeleton itself (Helmkeet al., 2001). This process of mechanotransmission results in theheterogeneous distribution of stress to locations of strain concen-tration in the cell, most notably at cell junctions and sub-cellularadhesion sites (Helmke et al., 2003), known hot-spots of mecha-notransduction (Davies, 1995; Shyy and Chien, 2002). Sincecytoskeletal deformations are accompanied by instantaneousbiochemical signalling at multiple sites of strain concentrationthroughout the cell, a compelling argument can be made forstructural continuity originating at the sites of initial haemody-namics-imposed deformation representing a decentralisedmechanotransduction model (Davies, 1995).
Two structures potentially play a key role in the endothelial cellresponse to its local haemodynamic environment: the glycocalyxsurface coating of the cell (Vink and Duling, 1996, 2000; Weinbaumet al., 2007; Reitsma et al., 2007) and the primary ciliumwhich canextend up to several microns into the vascular lumen.
The primary structural components of the vascular endothelialglycocalyx are proteoglycans. The primary core proteins are syn-decans and glypicans. Dense arrays of glycosaminoglycan (GAG)polymer side chains are bound to the core proteins. Although it iswidely accepted that the structure of the glycocalyx is a densepolymer mesh, direct measurements of material properties rele-vant to mechanotransmission and mechanotransduction remainelusive. Weinbaum et al. (2003) proposed that the material prop-erties are determined primarily by the flexural rigidity of the coreproteins, which they estimated from the recovery time of the gly-cocalyx thickness after “crushing” by a passing leukocyte ina capillary in vivo. Weinbaum et al. (2003) predict that a wall shearstress of 1 Pa is predicted to cause a 6-nm lateral displacement ofactin filaments in the submembranous cortical web. Thisdisplacement magnitude is similar to that of the myosin motor stepsize, implying that the force transmitted fromwall shear stress is ofsimilar magnitude to individual actomyosin contraction events.More recently this hypothesis has been extended to propose thatthe forces transmitted from shear stress cause sufficient deforma-tion that the actin dense peripheral bands in regions near cellecelljunctions interact mechanically like the rubber fenders on bumpercars (Thi et al., 2004). In this model, the submembranous actincortical web and the dense peripheral bands act as a functionalmechanical unit, and force transmission from the cell surface tojunctional regions occurs through the cytoskeleton in a mannerconsistent with a decentralisation distribution throughout the cell.
A large body of experimental data support the hypothesis thatthe glycocalyx serves to influence force transmission to either localsignal transduction complexes associated with the luminal
membrane or to dense peripheral bands in the actin cytoskeleton(Vink et al., 2000; Hecker et al., 1993; Pahakis et al., 2007; Florianet al., 2003; Gouverneur et al., 2006; Thi et al., 2004; Yao et al.,2007). Despite criticism of the specificity of enzymes used inmany experiments to degrade glycocalyx components, both in vitroand in vivo analyses place the glycocalyx near the initiation ofphysiologically relevant endothelial mechanotransmission andmechanotransduction mechanisms. In the near future, inhibition ofgene and protein expression of glycocalyx components usinggenetic intervention is likely to be more revealing of the extent towhich this complex surface coat is involved in endothelialmechanotransduction.
Endothelial primary cilia have received much less attention.Mammalian cells including endothelium are capable of expressinga single intermediate filament-rich primary cilium of undeter-mined function (Kojimahara, 1990). The structures are not obviousin cultured endothelium observed by light microscopy and have notbeen reported in AFM scans of living and fixed cells. However, theyhave been identified in corneal vessels by electron microscopy andin cultured human umbilical vein endothelial cells by antibodystaining of acetyl-a-tubulin and vimentin. Although the glycocalyxmay extend the cell boundary by up to 1 mm, its distributiongenerally follows the topography of the cell membrane. In contrast,erect cilia may extend several micrometers into the flow field,where higher velocities will impose a greater drag force. Primarycilia may act as amplifiers to open transient receptor potential (TRP)channels or to transmit stress to cortical (and hence cytoplasmic)cytoskeleton. Exposure of human umbilical vein endothelial cells tolaminar shear stress resulted in primary cilium disassembly (Iominiet al., 2004). In vivo, endothelial cilia have been mapped to highshear locations in the embryonic chicken cardiovascular system(van der Heiden et al., 2006). Recent in situ immunostaining of largevessel endothelium in adult mice has revealed expression ofendothelial cilia restricted to sites of flow distrurbance (van derHeiden et al., 2008). This distribution and the induction of arte-rial endothelial cilia by the experimental creation of flow distur-bance (low and disturbed shear stresses) have led to the proposalthat primary cilia act as fluid shear stress sensors in endothelium.The mechanical properties of cilia, a critical consideration for flowmechanotransduction, are unstudied. Further mechanisticevidence of cilium-mediated mechanotransduction using geneticmanipulation will help evaluate the role of these structures inarteries to provide a better basis for modelling at the sub-cellularlevel.
In developing theoretical models for mechanotransduction, thematerial properties of the endothelium can be described byconstitutive equations. Stressestrain relationships range fromlinear visco-elasticity models of individual elements in series or inparallel to complex moduli that include changes in material prop-erties as a function of stress or strain history (Davies and Helmke,2009). Measurements of stressestrain relationships are ideallymade in living cells, although measurements using gels of cyto-skeletal proteins are also instructive (Janmey et al., 1991, 1998). Forexample, filamentous actin, a prominent cytoskeletal element, canbe represented to a first approximation as elastic, whereas inter-mediate filaments exhibit both creep and strain hardening prop-erties, while microtubules exhibit slow creep.
Many potent labile molecules are present in the cell-free fluidphase close to the endothelial surface (referred to as the boundarylayer). These molecules are subject to convective transport thataffects their integrity and receptor binding. In vivo, bulk flow ofblood plasma is excluded from the glycocalyx, as demonstrated bynear-wall microparticle image velocimetry (m-PIV; Smith et al.,2003), and the presence of the endothelial cell glycocalyx furthercomplicates modelling solute transport to receptors at the

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 67
endothelial plasma membrane (Pries et al., 2000; Weinbaum et al.,2007). The best studied example are probably adenosine nucleo-tides (ATP, ADP, adenosine) which together with flow have beenshown to influence endothelial calcium responses (Dull and Davies,1991) and autocrine signalling in epithelial cells (Tschumperlinet al., 2004); the glycocalyx has been lumped into these andsimilar transport models. Since potent degradative enzymes at theendothelial cell surface can destroy ATP, a combination of flow andenzyme activity regulates the concentration of ATP in the boundarylayer and limits access to its receptor. The concentration of ATP inthe boundary layer is likely to vary widely as a result of the complexflow fields that are encountered in vivo and this, plus differentialtransport in a heterogeneous glycocalyx, may account for thedifferential signal transduction responses that are observed in theendothelium.
8.2. Mechanotransduction and disease
Within the normal physiology of the heart, structural andfunctional cell and tissue heterogeneities can increase the proba-bility of later cardiovascular disease. However, beyond overtstructural defects are more subtle heterogeneities of arterialgeometry that, by creating local flow disturbances, promote athero-susceptible pro-pathological endothelial cell phenotypes beforeany pathological changes are detectable.
As discussed from a fluid dynamic perspective in Section 4.3, thegeometry of the major arteries is key in determining coronaryartery flow dynamics. Flow modelling studies link regions of flow
Fig. 6. Arterial endothelial morphologies in situ reflect local haemodynamics. A: In pulsatilealigned in the direction of flow (arrow), a morphology that is present in most regions ofalignment. C: Cell shape transition region adjacent to an intercostal branch of the aorta whecorner of image) changes to unaligned polygonal-shaped endothelial cells (right corner). Therandomly oriented cells are associated with arterial branch sites, curvatures and bifurcatiocurvature of aortic arch, endothelial nuclear shape is representative of a similar disturbed fl
observed by scanning electron microscopy (Davies and Bowyer, 1975). In B and D, the nucleallows rapid evaluation of cell alignment (within several minutes after opening fresh arteriFrom: Davies, P.F., 2007. In: Aird, W.C. (Eds.) ‘Endothelial Biomedicine; A Comprehensive Tr
separation and flow reversal to the preferential susceptibility forthe development of atherosclerotic lesions, the major underlyingcause of most coronary vascular disease. Fig. 6 shows endothelialcell 2-D morphology in locations where there is unidirectionallaminar flow (most arterial regions; Fig. 6A,B) and where there issignificant flow separation with transient flow reversals (branches,curves; Fig. 6C,D). In the latter regions, the cell morphologiesundergo a distinct change from normally aligned endothelial shapeto a polygonal morphology without preferential orientation. Thisreveals important changes in the local flow characteristics. Forexample in the aortic arch inner curvature and regions of thecoronary arteries proximal to branches there are significant regionsof polygonal-shaped endothelial cells also observable by simplenuclear staining (Fig. 5D). Downstream and distal to the branchpoints, a transition back to aligned cells defines regional morpho-logical boundaries that likely reflect distinct haemodynamic char-acteristics (and regions of differential endothelial phenotypes). Instudies of cultured endothelium that was induced to assumesimilar 2-D outlines by flow, Barbee et al. (1994) demonstrated byatomic force microscopy that the topography of the cells is flat-tened when the cells are aligned. Since the 3D topography is theflow interface, these surface topography differences have impor-tant implications for mechanotransduction at the sub-cellular scale(Davies et al., 1995) and perhaps also for convective mass transfercharacteristics (Hillsley and Tarbell, 2002).
In flow separation a volume of blood separates from the bulkflow to form a vortex because of a pressure gradient induced bylocal changes in the vessel geometry. Because blood flow is
but unidirectional laminar flow in the descending thoracic aorta, endothelial cells arethe arterial circulation. B: The nuclear orientation in a similar region also indicatesre flow separates and reverses during part of the cardiac cycle. Cell alignment (top leftcurved arrows mark a haemodynamic transition zone a few cells wide. Polygonal andns that are regions susceptible to atherosclerosis. D: At the athero-susceptible innerow region. In A and C the cell shapes are outlined by silver proteinate deposition andus is stained with DAPI and observed by fluorescence microscopy. The latter techniqueal tissue). Nuclear orientation and cell shape are closely correlated. Scale bar ¼ 20 mm.eatise’, Cambridge University Press, pp. 230e245, with permission.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7668
pulsatile in large elastic and muscular arteries, the vortex maycollapse in diastole and reform during systole Steinman and Taylor(2005). The important haemodynamic characteristics of suchcomplex separation regions are that there is a phase of flowreversal with steep temporal and spatial gradients that createsoscillatory haemodynamic forces; the Oscillatory Shear Index, OSI,is a composite measurement of this. OSI is the fraction of total flowvolume (or 2-D section thereof) that reverses during a singlecardiac cycle (Suzuki et al., 1998). In flow separation regions theaverage shear stress is lower and the particle (and solute and bloodcell) residence time is increased. Consequently the mechanical andfluid transport environment of endothelium at OSI sites issubstantially different; this is reflected in the local endothelialmorphology and the athero-susceptible biological phenotype.
Obstruction of blood flow in the coronary arteries, caused byatherosclerosis and/or atherothrombosis, has devastating effectsupon large downstream regions of coronary perfusion resulting inangina, vasospasm, and heart attack. Regions proximal to branchesare more susceptible to atherosclerosis than more distant sites; forexample the proximal flow entry regions of the left and rightcoronary arteries, the left coronary artery branching region to thecircumflex and left anterior descending (LAD) arteries, and majorbifurcations within these arteries. The regions where the epicardialportions of the coronary arteries enter the deeper myocardium arealso susceptible regions. Arterial geometry at most of these sitesresults in flow separation during at least part of the cardiac cycle.Most experimental investigations have been conducted on moreaccessible flow disturbance-vascular susceptibility relationships inarteries outside of the heart (Ku and Giddens, 1983; Davies et al.,2002) but recent genomic investigations of the endothelium(Passerini et al., 2004; Davies, 2009; Civelek et al., in press) indicatethat mechanisms uncovered in the non-cardiac circulation are alsoapplicable to the coronary circulation.
Separated flow generates complicated spatio-temporalshearestress gradients that directly affect the biology of theendothelium. The local haemodynamic environment influencesendothelial gene and protein expression, post-translational events,and cell function through a combination of biomechanical andtransport mechanisms. Passerini et al. (2004) reported the first highthroughput genomic analyses of endothelial cells in the aortic arch(which exhibits, for example, flow separation, flow reversal, lowaverage shear stress, OSI) compared with endothelial cells subjectto undisturbed flow in the nearby descending thoracic aorta. Therewas differential expression of many genes associated with bothpro-inflammatory (athero-susceptible) and anti-inflammatorypathways, pro- and anti-coagulation mechanisms, and cholesterolbalance, revealing a heightened upregulation of checks andbalances in the endothelium of athero-susceptible regions. Therewas no evidence of pathological changes in the vessel wall in theseregions of susceptibility nor were any of the cardinal markers ofinflammation expressed. A working interpretation is that cells indisturbed flow regions may be sensitised to additional factors thatpromote atherogenesis (hypercholesterolemia, hypertension, dia-betes, smoking, etc) compared to cells in protected locations. Someinsight into the underlying mechanisms that may promote thephenotype differences has recently come to light from studies ofthe endoplasmic reticulum (ER)-stress response in endothelial cells(Feaver et al., 2008; Zhang et al., 2009; Civelek et al., in press). Thesestudies implicate disturbed flow as a regulatory factor in which theendothelium appears to be subject to a biomechanical form ofstress (as compared with other forms of stress) that results in thechronic activation of the ER-stress response and the adaptiveunfolded protein response mechanism. Indeed complex shearstress, such as occurs in flow separation regions, invokes intracel-lular metabolic stress as a coping mechanism and this state is
related to athero-susceptibility. A post-transcriptional pathwaymediated by small microRNAs has recently been reported inathero-susceptible endothelium to regulate upstream elements ofthe pro-inflammatory NFkappaB pathway (Fang et al., 2010).
The association of disturbed flow and reactive oxygen species(ROS)-related genes is a recurrent finding in endothelial genomicanalyses. For example, the residence time of superoxide anion,a damaging free radical, is increased at sites of disturbed flow. ROScan induce ER-stress but it is unclear how these mechanisms arelinked together in disturbed flow. ROS is clearly present in higherconcentrations in disturbed flow arterial regions (Davies andCivelek, in press), and this may impact glycocalyx integrity (vanden Berg et al., 2006; Vink et al., 2000).
Another signature of flow disturbance in vivo is a decreasedavailability of nitric oxide caused by downregulation of endothelialnitric oxide synthase (eNOS, NOS3) (de Nigris et al., 2003; Passeriniet al., 2004). Since steady undisturbed flow is known to upregulateeNOS (Boo et al., 2002), it is likely that the disturbed flow charac-teristics are responsible for eNOS downregulation in these regionsin vivo.
8.3. Challenges
The incorporation of cell-scale modelling into models for thecoronary vasculature is an exciting challenge that will benefit froma number of developing ideas and technologies.
A wealth of descriptive detail is emerging that better definesendothelial heterogeneity based on histology and immunohisto-chemistry in which care is taken to retain spatial relationships;however, information available by this approach is at best semi-quantitative. High throughput approaches are increasingly usefulbecause they carry a large bioinformatics data load that can morereadily be imported into systems modelling. Other quantitativeapproaches might be considered include: (i) the refinement ofregional heterogeneity to single cell measurement of candidategene expression; (ii) site-specific endothelial ROS levels (visualisedby dihydroethidium); (iii) activities of key endothelial enzymes ingeometrically-defined locations of coronary arteries; and (iv) moresophisticated proteomic/mass spectrometry technologies of spatialquantitation. In pathogenesis, a major challenge is the accuratemeasurement of endothelial phenotype(s) overlying developinglesions, which presents a non-trivial problem of accessibility, cellisolation and nucleic acid purification.
In addition to determining the effects of heterogeneity higherresolution imaging of flow characteristics is also required foradvancing both models and understanding of endothelial function.The localisation of flow disturbances needs to be better defined andat higher resolution. Current methods range from model simula-tions based upon vascular casting, imaging by MRI and ultrasound,and by directly looking at the morphology of the endothelial cells(e.g. Fig. 6). The latter method is simple but informative (Davies andBowyer, 1975; Nerem et al., 1981). The latter method is simple butinformative. In locations where there is significant flow separationwith flow reversals, the normally aligned endothelial shape is lostand a polygonal morphology exists; i.e. the cell reveals importantchanges in the local flowcharacteristics (see Fig. 6). For example theaortic arch inner curvature and regions of the coronary arteriesproximal to branches there are significant regions of polygonal-shaped endothelial cells (observable by simple nuclear staining fororientation). Downstream and distal to branch points, a transitionback to aligned cells clearly defines morphological boundaries thatlikely reflect distinct haemodynamic characteristics (and regions ofdifferential endothelial phenotypes). While great improvements in3D imaging and computational flow modelling of arteries isoccurring (e.g. Steinman and Taylor, 2005; Markl et al., 2009), the

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 69
resolution falls short of that required for cellular/sub-cellularmodelling.
Finally, we also need a better characterisation of stenotic lesionsand their haemodynamic impact. While the fundamental rela-tionship between arterial stenosis geometry and the resultingpressure dropeflow relationship developed by Young (1979) wasalso shown to hold for coronary arteries in animals and patients(Gould, 1985), assessing the physiological severity of an obstructivelesion remains a key problem in clinical practice. Cardiologists arefrequently faced with multiple non-obstructive lesions and need todecide on the optimal treatment strategy. Modelling of fluiddynamics associated with serial lesions, their location andmorphology has the potential to help with decision-making in thecatheterisation laboratory. In addition, plaque composition andconfiguration has been shown to directly influence the risk ofrupture, leading to acute and often devastating coronary events.Novel imaging methods such as optical coherence tomography andnear infrared spectroscopy can support the clinical assessment ofsuch vulnerable plaques (Narula and Strauss, 2007; Schuijf et al.,2009). Risk stratification and patient management may alsosubstantially be aided by FSI models of unstable plaques. Finally,the perfusion provided by collateral vessels is not well understoodand largely dependent on indirect assessment that is not yet wellsupported by appropriate models (Spaan et al., 2006).
9. Integrated model development
The aim of the Cardiac Physiome project is to simulate thenormal and pathophysiological function of the heart, and tounderstand how clinical interventions and drug therapies may beused to treat the diseased heart. Models are needed for the keycomponents of the coronary vascular system, including structure,biomechanics, flow, mass transport, regulation and remodelling,and cellular responses. However, physiological processes involvemany interactions between these basic components, as illustratedby the preceding sections. The challenge then is to developmodelling frameworks within which these components can beintegrated, to allow predictions of cardiac function, (compensatory)responses to disease conditions, and optimal treatment strategies.
The models for these components involve descriptions ofbiochemical and biomechanical phenomena occurring at multipletemporal and spatial scales, including the molecular, cellular,vascular, tissue and organ levels. In integrating these componentswe are therefore faced with the challenge of combining models atdisparate scales. A wide range of theoretical and numerical toolscan be used in the development of such multi-scale models,including cellular automata, agent based modelling, stochasticprocesses, continuum mechanics, compartmental models, multi-phase flows, computational fluid and solid dynamics, many ofwhich have been discussed above.
The challenge is to introduce small-scale information intolarger-scale models without the resulting models becomingcomputationally intractable. One approach is to use a lumpedrepresentation of where vessels smaller than a certain size arerepresented by a single compartment of fixed resistance anduniform properties. This approach is appropriate inmany situationsand has the advantage of simplicity. However, a limitation of thisapproach is that significant spatial variations may exist within thiscompartment, which are not represented by the model.
An intermediate approach between detailed representation ofevery vessel and a lumped approach is provided by homogenisationtheory. In this theory, a local spatial averaging of fine-scale struc-ture is achieved by exploiting asymptotic techniques to estimatetissue-scale properties, based on explicit solutions in smaller-scalesubunits. The approach relies on the separation of the system into
disparate length scales; in the simplest case this could bea repeating micro-scale where the structural detail is visible, anda tissue-scale where only the averaged properties are identifiable.
9.1. Homogenisation
Homogenisation theory is in the early stages of being applied toanswer specific questions on cardiac function or treatment strate-gies (see Section 3.2). To illustrate the potential of the homogeni-sation approach, it is relevant to consider related work modellingperfusion in vascular tumours (Chapman et al., 2008; Shipley andChapman, 2010). The approach assumes a periodic micro-struc-ture with Stokes flow in the capillaries, Darcy flow in the inter-stitium, and leakage through the vascular walls; the resultingequations comprise a double porous medium with coupled Darcyflow through the interstitium and vasculature. The Darcy fluidpermeability tensors capture the dependence of tissue-scaleperfusion on the micro-scale flow characteristics. One advantage isthat the effective equations are derived without prescribing themicro-scale structure so that the impact of different networktopologies can be tested a priori. Also, the final equations arecomputationally efficient to solve compared to numerical solutionsof the full capillary network.
In addition to fluid transport, such an approach to homogenisingmicro-scale structures may prove powerful in describing transportof oxygen (and other solutes) within the heart. In the context oftumour modelling, Shipley and Chapman (2010) illustrate howsuch techniques can be used to model substrate transport, fora range of micro-scale transport properties. When employed inheart modelling, such methods have the potential to overcome thelimitation of “Krogh cylinder” models (Section 5.2).
However, the current framework faces significant challenges if itis going to capture the cardiac vasculature in a physiologicallyrealistic way. First of all, the vasculature is a continuous hierarchy ofvessels arranged in a complex 3D topology that is not captured bymicro and tissue length scales in real space. One possible approachis to discretise the vasculature into units categorised by vesseldimensions, and to move up the length scales using homogenisa-tion: for example, Shipley et al. (submitted for publication) dividesthe vasculature into capillaries, arterioles and venules. It is essentialthat any such approach is motivated directly by vascular imagingdata, validated against perfusion data, and is applied in topologicalspace to realistically account for the intricate structure of thecoronary vascular tree. Secondly, the current framework dependson the leakiness of vessel walls in tumours to correlate vessel andinterstitial pressure fields at nearby spatial locations; this vesselleakiness makes spatial averaging a natural choice for tumours.However, the vessels of the myocardium are non-leaky, and thisexacerbates the challenge of identifying a meaningful averagingapproach. Finally, the models thus far are fixed in time, whereasrealistic models of cardiac behaviour must account for time-dependent changes in the vascular structure in response to regu-lation, angiogenesis, vascular remodelling and myocardialcontraction. The functional parameters characterising theseprocesses show large heterogeneity on a wide range of spatialscales, and homogenisation theory will require further develop-ment to take account of the interplay between these spatial andtemporal effects.
9.2. Integration
Coronary artery disease is a multifaceted problem that involvesnot only the micro-scale environment of the arterial wall but hasfar-reaching consequences for perfusion of the downstreammyocardial tissue, which eventually degrades myocardial function

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7670
and cardiac performance. This in turn can negatively influence theconduction system, cardiac contraction patterns, and therebycoronary flow. Compensatory physiological mechanisms are anintegral part of the disease process that need to be taken intoaccount in order to arrive at a complete picture of coronary vasculardisease. The link to clinical applications invariably needs to includepatient-specific anatomical models and additional structural andbiochemical parameters that are as yet difficult to acquire in vivo.A key challenge for the near future is to develop a suite of tools thatcan be adopted in a healthcare environment. The translation ofcomputational physiological models to clinical applications is a keygoal of the physiome approach (Brook and Waters, 2008; Hunterand Viceconti, 2009; Popel and Hunter, 2009; Miller, 2010).
The integration of models for multiple processes, presentsadditional challenges in terms of handling and processing largeheterogeneous datasets, and managing and interfacing softwareimplementations of model components. The wider researchcommunity will benefit from the establishment of an effectiveinfrastructure for sharing and disseminating the acquired researchoutputs. Future efforts should be aimed at developing commonimage and model data standards, tools for processing, visualisationand analysis, and publicly-accessible repositories that will facilitatequantitative modelling investigations. Due to the rapid advance-ments in acquisition speed and the size of the datasets, e.g. high-resolution imaging data may typically be several gigabytes in size,manual analysis has become impractical. The design of efficient andreliable automated analysis and visualisation algorithms remainsa constant challenge for the future, and will benefit from cross-disciplinary research and application of emerging technologiessuch as GPU computing. In addition, public databases for sharingthese raw data and reconstructed models with the widerresearch community must be established. Some initiatives havebeen made in this direction (e.g. euHeartDB, https://euheartdb.physiomeproject.org/welcome.do Gianni et al. (2010)). Theseissues of integration and multi-scale modelling are recurrent issuesencountered with the Physiome framework, and are not unique tothe coronary vasculature (Brook and Waters, 2008).
As illustrated in this review, progress has been made in manyareas of theoretical modelling of coronary vascular biomechanics.To realize the full potential of these advances, and to contributesubstantially tomedical progress, muchmorework is needed in thedevelopment of models for various aspects of the coronary circu-lation and their integration into models with predictive capabil-ities. These areas offer many exciting avenues and challenges forfuture research.
Acknowledgements
This paper arose from discussion at theworkshop on the CardiacPhysiome Project held at and funded by the Newton Institute forMathematical Sciences at the University of Cambridge, UK in July2009. SLW would like to thank the UK Engineering and PhysicalSciences Research Council (EPSRC) for funding in the form of anAdvanced Research Fellowship (EP/D070635/2). NPS would like toacknowledge support from the EPSRC (FP/F059361/1), the UKBiotechnology and Biological Sciences Research Council (BB/F01080X/1) and the European Commission (FP7-ICT-2007-224495:euHeart). JA would like to thank the British Heart Foundation forfunding in the form of an Intermediate Basic Science ResearchFellowship (FS/09/030/27812). PFD gratefully acknowledgesresearch grant support from the US National Institute of Health andthe American Heart Association, MS would like to acknowledgesupport from the Netherlands Heart Foundation (2006B186,2006B226) and the European Commission (FP7-ICT-2007-224495:
euHeart). We would like to thank E.S.Waters for compiling thecomprehensive list of references.
References
Aguado-Sierra, J., Alastruey, J., Wang, J.J., Hadjiloizou, N., Davies, J.E., Parker, K.H.,2008. Separation of the reservoir and wave pressure and velocity frommeasurements at an arbitrary location in arteries. Proc. Inst. Mech. Eng. Part H:J. Eng. Med. 222, 403e416.
Aird, W.C., 2007. Endothelial Biomedicine. Cambridge University Press, New York.Alastruey, J., Parker, K.H., Peiró, J., Sherwin, S.J., 2006. Can the modified Allen’s test
always detect sufficient collateral flow in the hand? A computational study.Comp. Meths. Biomech. Biomed. Eng. 9, 353e361.
Alastruey, J., Parker, K.H., Peiró, J., Sherwin, S.J., 2007. Modelling the circle of Willisto assess the effects of anatomical variations and occlusions on cerebral flows. J.Biomech. 40, 1794e1805.
Alastruey, J., Parker, K.H., Peiró, J., Sherwin, S.J., 2008. Lumped parameter outflowmodels for 1-D blood flow simulations: effect on pulse waves and parameterestimation. Commun. Comput. Phys. 4, 317e336.
Alastruey, J., Nagel, S.R., Nier, B., Hunt, A.A.E., Weinberg, P.D., Peiró, J., 2009a.Modelling pulse wave propagation in the rabbit systemic circulation to assessthe effects of altered nitric oxide synthesis. J. Biomech. 42, 2116e2123.
Alastruey, J., Parker, K.H., Peiró, J., Sherwin, S.J., 2009b. Analysing the pattern ofpulse waves in arterial networks: a timeedomain study. J. Eng. Math. 64,331e351.
Alastruey, J., Sherwin, S.J., Parker, K.H., Rubens, D.D., 2009c. Placental transfusioninsult in the predisposition for SIDS: a mathematical study. Early Hum. Devel.85, 455e459.
Algranati, D., Kassab, G.S., Lanir, Y., 2010. Mechanisms of myocardiumecoronaryvessel interaction. Am. J. Physiol. Heart Circ. Physiol. 298, H861eH873.
Anrep, G.V., Cruickshank, E.W.H., Downing, A.C., 1927. The coronary circulation inrelation to the cardiac cycle. Heart 14, 111e133.
Arciero, J.C., Carlson, B.E., Secomb, T.W., 2008. Theoretical model of metabolic bloodflow regulation: roles of ATP release by red blood cells and conductedresponses. Am. J. Physiol. Heart Circ. Physiol. 295, H1562eH1571.
Arts, T., Reneman, R.S., 1985. Interaction between intramyocardial pressure (IMP)and myocardial circulation. J. Biomech. Eng. 107, 51e56.
Arts, T., Reneman, R.S., Veenstra, P.C., 1979. A model of the mechanics of the leftventricle. Ann. Biomed. Eng. 7, 299e318.
Arts, M.G.J., 1978. A Mathematical Model of the Dynamics of the Left Ventricle andthe Coronary Circulation. PhD thesis, University of Limburg, Maastricht, TheNetherlands.
Austin Jr., R.E., Smedira, N.G., Squiers, T.M., Hoffman, J.I., 1994. Influence of cardiaccontraction and coronary vasomotor tone on regional myocardial blood flow.Am. J. Physiol. Heart Circ. Physiol. 266, H2542eH2553.
Avolio, A., 2009. Input impedance of distributed arterial structures as used ininvestigations of underlying concepts in arterial haemodynamics. Med. Biol.Eng. Comput. 47, 143e151.
Bache, R.J., Schwartz, J.S., 1982. Effect of perfusion pressure distal to coronarystenosis on transmural myocardial blood flow. Circ 65, 928e932.
Back, L.H., Kwack, E.Y., Back, M.R., 1996. Flow rateepressure drop relation in coro-nary angioplasty: catheter obstruction effect. J. Biomech. Eng. 118, 83e89.
Back, L.H., 1994. Estimated mean flow resistance increase during coronary arterycatheterization. J. Biomech. 27, 169e175.
Bakker, E.N., Versluis, J.P., Sipkema, P., VanTeeffelen, J.W., Rolf, T.M., Spaan, J.A.,VanBavel, E., 2003. Differential structural adaptation to haemodynamics alongsingle rat cremaster arterioles. J. Physiol. 548, 549e555.
Bakker, E.N., Matlung, H.L., Bonta, P., de Vries, C.J., van Rooijen, N., VanBavel, E.,2008. Blood flow-dependent arterial remodelling is facilitated by inflammationbut directed by vascular tone. Cardiovasc. Res. 78, 341e348.
Banerjee, R., Ashtekar, K., Helmy, T., Effat, M., Back, L., Khoury, S., 2008. Hemody-namic diagnostics of epicardial coronary stenoses: in-vitro experimental andcomputational study. Biomed. Eng. Online 7, 24.
Barbee, K.A., Davies, P.F., Lal, R., 1994. Shear stress-induced reorganization of thesurface topography of living endothelial cells imaged by atomic force micros-copy. Circ. Res. 74, 163e171.
Bassingthwaighte, J.B., Yipintsoi, T., Harvey, R.B., 1974. Microvasculature of the dogleft ventricular myocardium. Microvasc. Res. 7 (2), 229e249.
Bassingthwaighte, J., Hunter, P., Noble, D., 2009. The cardiac Physiome: perspectivesfor the future. Exp. Physiol. 94, 597e605.
Bauer, A.L., Jackson, T.L., Jiang, Y., 2007. A cell-based model exhibiting branching andanastomosis during tumor-induced angiogenesis. Biophys. J. 92, 3105e3121.
Bayliss, W.M., 1902. On the local reactions of the arterial wall to changes of internalpressure. J. Physiol. 28, 220e231.
Beard, D.A., Bassingthwaighte, J.B., 2000a. The fractal nature of myocardial bloodflow emerges from a whole-organ model of arterial network. J. Vasc. Res. 37 (4),282e296.
Beard, D.A., Bassingthwaighte, J.B., 2000b. Advection and diffusion of substances inbiological tissues with complex vascular networks. Ann. Biomed. Eng. 28 (3),253e268.
Beard, D.A., Schenkman, K.A., Feigl, E.O., 2003. Myocardial oxygenation in isolatedhearts predicted by an anatomically realistic microvascular transport model.Am. J. Physiol. 285, H1826eH1836.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 71
Berne, R.M., 1963. Cardiac nucleotides in hypoxia: possible role in regulation ofcoronary blood flow. Am. J. Physiol. 204, 317e322.
Bessems, D., Rutten, M.C.M., van de Vosse, F.N., 2007. A wave propagation model ofblood flow in large vessels using an approximate velocity profile function. J.Fluid Mech. 580, 145e168.
Bessems, D., Giannopapa, C.G., Rutten, M.C.M., van de Vosse, F.N., 2008. Experi-mental validation of a time-domain-based wave propagation model of bloodflow in viscoelastic vessels. J. Biomech. 41, 284e291.
Beyar, R., Ben-Ari, R., Gibbons-Kroeker, C.A., Tyberg, J.V., Sideman, S., 1993. Effect ofinterconnecting collagen fibers on left ventricular function and intramyocardialcompression. Cardiovasc. Res. 27, 2252e2263.
Bjorno, L., Pattersson, H., 1976a. Hydro- and hemodynamic effects of catheterizationof vessels e I. An experimental model. Acta Radiol. Diag. 17, 511.
Bjorno, L., Pattersson, H., 1976b. Hydro- and hemodynamic effects of catheterizationof vessels e II. Model experiments comparing circular and annular human areareduction. Acta Radiol. Diag 17, 749e762.
Boo, Y.C., Sorescu, G., Boyd, N., Shiojima, I., Walsh, K., Du, J., Jo, H., 2002. Shear stressstimulates phosphorylation of endothelial nitric-oxide synthase at Ser1179 byAkt-independent mechanisms: role of protein kinase A. J. Biol. Chem. 277,3388e3396.
Bovendeerd, P.H.M., Borsje, P., Arts, T., van de Vosse, F.N., 2006. Dependence ofintramyocardial pressure and coronary flow on ventricular loading andcontractility: a model study. Ann. Biomed. Eng. 34 (12), 1833e1845.
Brook, B.S., Waters, S.L., 2008. Mathematical challenges in integrative physiology. J.Math. Biol. 56, 893e896.
Bruinsma, P., Arts, T., Dankelman, J., Spaan, J.A., 1988. Model of the coronarycirculation based on pressure dependence of coronary resistance and compli-ance. Basic Res. Cardiol. 83, 510e524.
Burton, R.A., Plank, G., Schneider, J.E., Grau, V., Ahammer, H., Keeling, S.L., Lee, J.,Smith, N.P., Gavaghan, D., Trayanova, N., Kohl, P., 2006. Three-dimensionalmodels of individual cardiac histoanatomy: tools and challenges. Ann. N. Y.Acad. Sci. 1080, 301e319.
Canic, S., Kim, E.H., 2003. Mathematical analysis of the quasilinear effects ina hyperbolic model blood flow through compliant axi-symmetric vessels. Math.Meth. Appl. Sci. 26, 1161e1186.
Carlson, B.E., Arciero, J.C., Secomb, T.W., 2008. Theoretical model of blood flowautoregulation: roles of myogenic, shear-dependent, and metabolic responses.Am. J. Physiol. Heart Circ. Physiol. 295, H1572eH1579.
Carmeliet, P., 2005. Angiogenesis in life, disease and medicine. Nature 438,932e936.
Cassot, F., Lauwers, F., Fouard, C., Prohaska, S., Lauwers-Cances, V., 2006. A novelthree-dimensional computer-assisted method for a quantitative study ofmicrovascular networks of the human cerebral cortex. Microcirc 13, 1e18.
Chapman, S.J., Shipley, R.J., Jawad, R., 2008. Multiscale modelling of fluid transportin tumors. Bull. Math. Biol. 70, 2334e2357.
Chen, K., Popel, A.S., 2006. Theoretical analysis of biochemical pathways of nitricoxide release from vascular endothelial cells. Free Radic. Biol. Med. 41,668e680.
Cheng, C.P., Herfkens, R.J., Taylor, C.A., 2003. Inferior vena cava hemodynamicsquantified in vivo at rest and during lower limb exercise using magnetic reso-nance imaging. Am. J. Physiol. Heart Circ. Physiol. 284, H1161eH1167.
Chilian, W.M., Layne, S.M., 1990. Coronary microvascular responses to reductions inperfusion pressure. Evidence for persistent arteriolar vasomotor tone duringcoronary hypoperfusion. Circ. Res. 66, 1227e1238.
Chilian, W.M., Marcus, M.L., 1982. Phasic coronary blood flow velocity in intramuraland epicardial coronary arteries. Circ. Res. 50 (6), 775e781.
Chilian, W.M., Layne, S.M., Klausner, E.C., Eastham, C.L., Marcus, M.L., 1989. Redis-tribution of coronary microvascular resistance produced by dipyridamole. Am.J. Physiol. 256, H383eH390.
Choi, H., Ferrara, K., Barakat, A., 2007. Modulation of ATP/ADP concentration at theendothelial surface by shear stress: effect of flow recirculation. Ann. Biomed.Eng. 35, 505e516.
Choy, J.S., Kassab, G.S., 2009. Wall thickness of coronary vessels varies transmuralyin the LV but notthe RV: implications for local stress distribution. Am. J. Physiol.Heart Circ. Physiol. 297, H750eH758.
Civelek, M., Manduchi, E., Grant, G., Karanian, J.W., Pritchard, W.P., Guerraty, M.,Davies, P.F., Chronic endoplasmic reticulum stress partially activates endothelialunfolded protein response (UPR) in arterial regions susceptible to atheroscle-rosis. Circ. Res., in press.
Clayton, R.H., Bernus, O.V., Cherry, E.M., Dierckx, H., Fenton, F.H., Mirabella, L.,Panfilov, A.V., Sachse, F.B., Seeman, G., Zhang, H., 2011. Models of cardiac tissueelectrophysiology: Progress, challenges and open questions. Prog. Biophys. Mol.Biol. 104, 22e48.
Collins, D.M., McCullough, W.T., Ellsworth, M.L., 1998. Conducted vascularresponses: communication across the capillary bed. Microvasc. Res. 56,43e53.
Constantinescu, A.A., Vink, H., Spaan, J.A., 2001. Elevated capillary tube hematocritreflects degradation of endothelial cell glycocalyx by oxidised LDL. Am. J.Physiol. Heart Circ. Physiol. 280 (3), H1051eH1057.
Cornelissen, A.J., Dankelman, J., VanBavel, E., Spaan, J.A., 2002. Balance betweenmyogenic, flow-dependent, and metabolic flow control in coronary arterialtree: a model study. Am. J. Physiol. Heart Circ. Physiol. 282, H2224eH2237.
Damiano, E.R., Duling, B.R., Ley, K., Skalak, T.C., 1996. Axisymmetric pressure-drivenflow of rigid pellets through a cylindrical tube lined with a deformable porouswall layer. J. Fluid Mech. 314, 163e189.
Dankelman, J., Stassen, H.G., Spaan, J.A., 1999. Interaction between Gregg’sphenomenon and coronary flow. Med. Biol. Eng. Comput. 37 (6), 742e749.
Dash, R.K., Jayaraman, G., Mehta, K.N., 1996. Estimation of increased flow resistancein a narrow catheterized artery e a theoretical model. J. Biomech. 29,917e930.
Dash, R.K., Jayaraman, G., Mehta, K.N., 1999. Flow in a catheterised curved arterywith stenosis. J. Biomech. 32, 49e61.
Davies, P.F., Bowyer, D.E., 1975. Scanning electron microscopy: arterial endothelialintegrity after fixation at physiological pressure. Atherosclerosis 21, 463e469.
Davies, P.F., Civelek, M., Endothelial reticulum stress, redox and a pro-inflammatoryenvironment in athero-susceptible endothelium in vivo at sites of complexhemodynamic shear stress, Antioxid. Redox Signal, in press.
Davies, P., Helmke, B.P., 2009. Endothelial mechanotransduction. In: Mofrad, R.K.,Kamm, R.D. (Eds.), Cellular Mechanotransduction. Cambridge University Press,pp. 20e60.
Davies, P.F., Dewey, C.F., Bussolari, S.R., Gordon, E.J., Gimbrone Jr., M.A., 1984.Influence of hemodynamic forces on vascular endothelial function: in vitrostudies of shear stress and pinocytosis in cul-tured bovine aortic ECs. J. Clin.Invest 73, 1121e1129.
Davies, P.F., Mundel, T., Barbee, K.A., 1995. A mechanism for heterogeneous endo-thelial responses to flow in vivo and in vitro. J. Biomech. 28 (12), 1553e1560.
Davies, P.F., Polacek, D.C., Shi, C., Helmke, B.P., 2002. The convergence of haemo-dynamics, genomics, and endothelial structure, in studies of the focal origin ofatherosclerosis. Biorheol 39, 299e307.
Davies, P.F., Spaan, J.A., Krams, R., 2005. Shear stress biology of the endothelium.Ann. Biomed. Eng. 33 (12), 1714e1718.
Davies, J.E., Francis, D.P., Hughes, A.D., Parker, K.H., Mayet, J., 2006a. Use ofsimultaneous pressure and velocity measurements to estimate arterial wavespeed at a single site in humans. Am. J. Physiol. Heart Circ. Physiol. 290,H878eH885.
Davies, J.E., Whinnett, Z.I., Francis, D.P., Manisty, C.H., Aguado-Sierra, J., Willson, K.,Foale, R.A., Malik, I.S., Hughes, A.D., Parker, K.H., Mayet, J., 2006b. Evidence ofa dominant backward-propagating “suction” wave responsible for diastoliccoronary filling in humans, attenuated in left ventricular hypertrophy. Circ 113,1768e1778.
Davies, P.F., 1995. Flow-mediated endothelial mechanotransduction. Physiol. Rev.75, 519e560.
Davies, P.F., 2009. Hemodynamic shear stress and the endothelium in cardiovas-cular pathophysiology. Nat. Clin. Prac, Cardiovasc. Med. 6, 16e26.
Davis, M.J., Hill, M.A., 1999. Signaling mechanisms underlying the vascularmyogenic response. Physiol. Rev. 79, 387e423.
Davis, M.J., Hill, M.A., Kuo, L., 2008. Local regulation of microvascular perfusion. In:Tuma, R.F., Duran, W.N., Ley, K. (Eds.), Handbook of Physiology: Microcircula-tion, second ed. Academic Press, San Diego, pp. 161e284.
de Nigris, F., Lerman, L.O., Ignarro, S.W., Sica, G., Lerman, A., Palinski, W., Ignarro, L.J.,Napoli, C., 2003. Beneficial effects of antioxidants and L-arginine on oxidation-sensitive gene expression and endothelial NO synthase activity at sites ofdisturbed shear stress. Proc. Natl. Acad. Sci. USA 100, 1420e1425.
De Santis, G., Mortier, P., De Beule, M., Segers, P., Verdonck, P., Verhegghe, B., 2010.Patient-specific computational fluid dynamics: structured mesh generationfrom coronary angiography. Med. Biol. Eng. Comput. 48, 371e380.
de Weger, A., van Tuijl, S., Stijnen, M., Steendijk, P., de Hart, J., 2010. Direct endo-scopic visual assessment of a transcatheter aortic valve implantation andperformance in the physioheart, an isolated working heart platform. Circualtion121 (13), e261ee262.
Decking, U.K., 2002. Spatial heterogeneity in the heart: recent insights and openquestions. News Physiol. Sci. 17, 246e250.
Deussen, A., Brand, M., Pexa, A., Weichsel, J., 2006. Metabolic coronary flow regu-lationecurrent concepts. Basic Res. Cardiol. 101, 453e464.
Dewey, C.F., Gimbrone Jr., M.A., Bussolari, S.R., Davies, P.F., 1981. The dynamicresponse of vascular ECs to fluid shear stress. J. Biomech. Eng. 103, 177e185.
Di Mario, C., Gil, R., Sunamura, M., Serruys, W., 1995. New concepts for interpre-tation of intracoronary velocity and pressure tracings. Br. Heart J. 74, 485e492.
Doucette, J.W., Corl, P.D., Payne, H.M., Flynn, A.E., Goto, M., Nassi, M., Segal, J., 1992.Validation of a Doppler guide wire for intravascular measurement of coronaryartery flow velocity. Circulation 85, 1899e1911.
Downey, J.M., Kirk, E.S., 1975. Inhibition of coronary blood flow by a vascularwaterfall mechanism. Circ. Res. 36, 753e760.
Duling, B.R., Hogan, R.D., Langille, B.L., Lelkes, P., Segal, S.S., Vatner, S.F., Weigelt, H.,Young, M.A., 1987. Vasomotor control: functional hyperemia and beyond. Fed.Proc. 46, 251e263.
Dull, R.O., Davies, P.F., 1991. Flow modulation of agonist (ATP)-response (Ca2þ)coupling in vascular endothelial cells. Am. J. Physiol. 261, H149eH154.
Dull, R.O., Tarbell, J.M., Davies, P.F., 1992. Mechanisms of flow-mediated signaltransduction in endothelial cells: kinetics of ATP surface concentrations. J. Vasc.Res. 29, 410e419.
Duncker, D.J., Bache, R.J., 2008. Regulation of coronary blood flow during exercise.Physiol. Rev. 88, 1009e1086.
Ellsworth, M.L., 2000. The red blood cell as an oxygen sensor: what is the evidence?Acta Physiol. Scand. 168, 551e559.
Fahraeus, R., Lindqvist, T., 1931. The viscosity of the blood in narrow capillary tubes.Am. J. Physiol. 96, 562e568.
Fahraeus, R., 1928. Die Strömungsverhältnisse und die Verteilung der Blutzellen imGefäbsystem. Zur Frage der Bedeutung der intravasculären Erythrocytenag-gregation. Klinische Wochenschrift 7, 100e106.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7672
Fang, Y., Shi, C., Manduchi, E., Civelek, M., Davies, P.F., 2010. MicroRNA-10a regu-lation of pro-inflammatory phenotype in athero-susceptible endothelium invivo and in vitro. Proc. Natl. Acad. Sci. USA 107, 13450e13455.
Feaver, R.E., Hastings, N.E., Pryor, A., Blackman, B.R., 2008. GRP78 upregulation byatheroprone shear stress via p38-, alpha2beta1-dependent mechanism inendothelial cells. Arterioscler Thromb. Vasc. Biol. 28 (8), 1534e1541.
Feigl, E.O., 1983. Coronary physiology. Physiol. Rev. 63, 1e205.Ferko, M.C., Bhatnagar, A., Garcia, M.B., Butler, P.J., 2007. Finite-element stress
analysis of a multicomponent model of sheared and focally-adhered endothe-lial cells. Ann. Biomed. Eng. 35, 208e223.
Florian, J.A., Kosky, J.R., Ainslie, K., Pang, Z., Dull, R.O., Tarbell, J.M., 2003. Heparansulfate proteoglycan is a mechanosensor on endothelial cells. Circ. Res. 93 (10),e136e42.
Fokkema, D.S., VanTeeffelen, J.W.G.E., Dekker, S., Vergroesen, I., Reitsma, J.B.,Spaan, J.A.E., 2005. Diastolic time fraction as a determinant of subendocardialperfusion. Am. J. Physiol. Heart Circ. Physiol. 288, H2450eH2456.
Formaggia, L., Gerbeau, J.F., Nobile, F., Quarteroni, A., 2001. On the coupling of 3Dand 1D NaviereStokes equations for flow problems in compliant vessels. Comp.Meth. App. Mech. Eng. 191, 561e582.
Frame, M.D., Sarelius, I.H., 1995. Energy optimization and bifurcation angles in themicrocirculation. Microvasc. Res. 50 (3), 301e310.
Franke, V.E., Parker, K.H., Wee, L.Y., Fisk, N.M., Sherwin, S.J., 2003. Time domaincomputational modelling of 1D arterial networks in monochorionic placentas.ESAIM: math. Mod. Num. Anal. 37 (4), 557e580.
Friedrich, M.G., 2010. Current status of cardiovascular magnetic resonance imagingin the assessment of coronary vasculature. Can. J. Cardiol. 26, 51Ae55A.
Fullana, J.M., Zaleski, S., 2009. A branched one-dimensional model of vesselnetworks. J. Fluid Mech. 621, 183e204.
Fung, Y.C., 1997. Biomechanics: Circulation, second ed. Springer-Verlag, New York.Furchgott, R.F., Zawadzki, J.V., 1980. The obligatory role of ECs in the relaxation of
arterial smooth muscle by acetylcholine. Nature 288, 373e376.Ganz, P., Abben, R., Friedman, P.L., Garnic, J.D., Barry, W.H., Levin, D.C., 1985.
Usefulness of transstenotic coronary pressure gradient measurements duringdiagnostic catheterization. Am. J. Cardiol. 55, 910e914.
Garcia, D., Camici, P.G., Durand, L.-G., Rajappan, K., Gaillard, E., Rimoldi, O.E.,Pibarot, P., 2009. Impairment of coronary flow reserve in aortic stenosis. J. Appl.Physiol. 106, 113e121.
Gaskell, W.H., 1878. Further researches on the vasomotor nerves of ordinarymuscles. J. Physiol. 1, 262e426.
Gerhardt, H., Golding, M., Fruttiger, M., Ruhrberg, C., Lundkvist, A., Abramsson, A.,Jeltsch, M., Mitchell, C., Alitalo, K., Shima, D., Betsholtz, C., 2003. VEGF guidesangiogensic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 161 (8),1163e1177.
Gianni, D., McKeever, S., Britten, R., Yu, T., Hunter, P.J., Smith, N.P., 2010. A steptowards the implementation of the Virtual Physiological Human Project:sharing and reusing geometrical models of the heart over the Web. Phil. Trans.368 (1921), 3039e3056.
Gonzalez-Fernandez, J.M., Ermentrout, B., 1994. On the origin and dynamics of thevasomotion of small arteries. Math. Biosci. 119, 127e167.
Goto, M., Flynn, A.E., Doucette, J.W., Jansen, C.M.A., Stork, M.M., Coggins, D.L.,Muehrcke, D.D., Husseini, W.K., Hoffman, J.I.E., 1991. Cardiac contraction affectsdeep myocardial vessels predominantly. Am. J. Physiol. 261, H1417eH1429.
Goto, M., VanBavel, E., Giezeman, M.J., Spaan, J.A., 1996. Vasodilatory effect ofpulsatile pressure on coronary resistance vessels. Circ. Res. 79 (5), 1039e1045.
Gould, K.L., 1985. Quantification of coronary artery stenosis in vivo. Circ. Res. 57,341e353.
Gouverneur, M., Spaan, J.A.E., Pannekoek, H., Fontijn, R.D., Vink, H., 2006. Fluidshear stress stimulates incorporation of hyaluronan into endothelial cell gly-cocalyx. Am. J. Physiol. 290, H458eH462.
Gregg, D.E., 1963. Effect of coronary perfusion pressure or coronary flow on oxygenusage of the myocardium. Circ. Res. 13, 497e500.
Griffith, T.M., Edwards, D.H., Davies, R., Harrison, T.J., Evans, K.T., 1986. EDRF coor-dinates the behaviour of vascular resistance vessels. Nature 329, 442e445.
Gruionu, G., Hoying, J.B., Gruionu, L.G., Laughlin, M.H., Secomb, T.W., 2005a.Structural adaptation increases predicted perfusion capacity after vesselobstruction in arteriolar arcade network of pig skeletal muscle. Am. J. Physiol.Heart Circ. Physiol. 288, H2778eH2784.
Gruionu, G., Hoying, J.B., Pries, A.R., Secomb, T.W., 2005b. Structural remodeling ofmouse gracilis artery after chronic alteration in blood supply. Am. J. Physiol.Heart Circ. Physiol. 288, H2047eH2054.
Guiot, C., Pianta, G., Cancelli, C., Pedley, T.J., 1990. Prediction of coronary blood flowwith a numerical model based on collapsible tube dynamics. Am. J. Phys. 258,H1606eH1614.
Hacking,W.J.G., VanBavel, E., Spaan, J.A.E.,1996. Shear stress is not sufficient to controlgrowth of vascular networks: a model study. Am. J. Phys. 270, H364eH375.
Hadjiloizou, N., Davies, J.E., Malik, I.S., Aguado-Sierra, J., Willson, K., Foale, R.A.,Parker, K.H., Hughes, A.D., Francis, D.P., Mayet, J., 2008. Differences in cardiacmicrocirculatory wave patterns between the proximal left mainstem andproximal right coronary artery. Am. J. Physiol. Heart Circ. Physiol. 295,H1198eH1205.
Hahn, C., Schwartz, M.A., 2009. Mechanotransduction in vascular physiology andatherogenesis. Nat. Rev. Mol. Cell. Biol. 10, 53e62.
Han, Y.F., Weinbaum, S., Spaan, J.A.E., Vink, H., 2006. Large deformation analysis ofthe elastic recoil of fibre layers in a Brinkman medium with application to theendothelial glycocalyx. J. Fluid Mech. 554, 217e235.
Hecker, M., Mulsch, A., Bassenge, E., Busse, R., 1993. Vasoconstriction and increasedflow: two principal mechanisms of shear stress-dependent endothelial autacoidrelease. Am. J. Physiol. 265, H828eH833.
Heinzer, S., Krucker, T., Stampanoni, M., Abela, R., Meyer, E.P., Schuler, A.,Schneider, P., Müller, R., 2006. Hierarchical microimaging for multiscale analysisof large vascular networks. Neuroimage 32 (2), 626e636.
Helmke, B.P., Thakker, D.B., Goldman, R.D., Davies, P.F., 2001. Spatiotemporal anal-ysis of flow-induced intermediate filament displacement in living endothelialcells. Biophys. J. 80, 184e194.
Helmke, B.P., Rosen, A.B., Davies, P.F., 2003. Mapping mechanical strain of anendogenous cytoskeletal network in living endothelial cells. Biophys. J. 84,2691e2699.
Helmke, B.P., 2005. Molecular control of cytoskeletal mechanics by hemodynamicforces. Physiology (Bethesda) 20, 43e53.
Heusch, G., Kleinbongard, P., Bose, D., Levkau, B., Haude, M., Schulz, R., Erbel, R.,2009. Coronary microembolization: from bedside to bench and back to bedside.Circ 120, 1822e1836.
Hillsley, M.V., Tarbell, J.M., 2002. Oscillatory shear alters endothelial hydraulicconductivity and nitric oxide levels. Biochem. Biophys. Res. Commun. 293,1466e1471.
Hintze, T.H., Vatner, S.F., 1984. Reactive dilation of large coronary arteries inconscious dogs. Circ. Res. 54, 50e57.
Ho, S.Y., Sánchez-Quintana, D., Becker, A.E., 2004. A review of the coronary venoussystem: a road less travelled. Heart Rhythm 1 (1), 107e112.
Hoffman, J.I.E., Spaan, J.A.E., 1990. Pressure-flow relations in coronary circulation.Physiol. Rev. 70, 331e390.
Holzapfel, G.A., Gasser, T.C., Ogden, R.W., 2000. A new constitutive framework forarterial wall mechanics and a comparative study of material models. J. Elas. 61,1e48.
Huberts, W., Bosboom, E.M.H., van de Vosse, F.N., 2009. A lumped model for bloodflow and pressure in the systemic arteries based on an approximate velocityprofile function. Math. Biosci. Eng. 6 (1), 27e40.
Hudlicka, O., Brown, M.D., 2009. Adaptation of skeletal muscle microvasculature toincreased or decreased blood flow: role of shear stress, nitric oxide and vascularendothelial growth factor. J. Vasc. Res. 46, 504e512.
Hughes, A., Parker, K., 2009. Forward and backward waves in the arterial system:impedance or wave intensity analysis? Med. Biol. Eng. Comput. 47, 207e210.
Humphrey, J.D., 2001. Cardiovascular Solid Mechanics: Cells, Tissues, and Organs.Springer, New York.
Hunter, P.J., Viceconti, M., 2009. The VPH-Physiome project: standards and tools formultiscale modeling in clinical applications. IEEE Rev. Biomed. Eng. 2, 40e53.
Huo, Y., Kassab, G.S., 2007. A hybrid one-dimensional/Womersley model of pulsatileblood flow in the entire coronary arterial tree. Am. J. Phys. 292, H2623eH2633.
Huyghe, J.M.R.J., Oomens, C.W.J., van Campen, D.H., Heethaar, R.M., 1989a. LowReynolds number steady state flow through a branching network of rigidvessels: I. a mixture theory. Biorheol 26, 55e71.
Huyghe, J.M.R.J., Oomens, C.W.J., van Campen, D.H., Heethaar, R.M., 1989b. LowReynolds number steady state flow through a branching network of rigidvessels: II. A finite element mixture model. Biorheol 26, 73e84.
Huyghe, J.M., Arts, T., van Campen, D.H., Reneman, R.S., 1992. Porous medium finiteelement model of the beating left ventricle. Am. J. Physiol. 262, H1256eH1267.
Ingber, D.E., 2002. Mechanical signaling and the cellular response to extracellularmatrix in angiogenesis and cardiovascular physiology. Circ. Res. 91, 877e887.
Ingber, D.E., 2008. Tensegrity-based mechanosensing from macro to micro. Prog.Biophys. Mol. Biol. 97, 163e179.
Iomini, C., Tejada, K., Mo, W., Vaananen, H., Piperno, G., 2004. Primary cilia ofhuman endothelial cells disassemble under laminar shear stress. J. Cell Biol. 164,811e817.
Jacobs, J., Algranati, D., Lanir, Y., 2008. Lumped flowmodeling in dynamically loadedcoronary vessels. J. Biomech. Eng. 130, 054504.
Jacobsen, J.C., Mulvany, M.J., Holstein-Rathlou, N.H., 2008. A mechanism for arte-riolar remodeling based on maintenance of smooth muscle cell activation. Am.J. Physiol. Regul. Integr. Comp. Physiol. 294, R1379eR1389.
Janmey, P.A., Euteneuer, U., Traub, P., Schliwa, M., 1991. Viscoelastic properties ofvimentin compared with other filamentous biopolymer networks. J. Cell Biol.113, 155e160.
Janmey, P.A., Euteneuer, U., Traub, P., Schliwa, M., 1998. Viscoelasticity of interme-diate filament networks. Sub-Cellular Biochem. 31, 381e397.
Jayaraman, G., Dash, R.K., 2001. Numerical study of flow in a constricted curvedannulus: an application to flow in a catheterized artery. J. Eng. Maths 40,355e376.
Jayaraman, G., Tiwari, K., 1995. Flow in a catheterized curved artery. Med. Biol.Engng. Compt 33, 720e724.
Johnson, P.C., 1980. The myogenic response. In: Bohr, D.F., Somlyo, A.P.,Sparks Jr., H.V. (Eds.), Handbook of Physiology, Section 2, the CardiovascularSystem. Vascular Smooth Muscle, vol. II. American Physiological Society,Bethesda, MD, pp. 409e442.
Jones, C.J., Kuo, L., Davis, M.J., Chilian, W.M., 1995. Regulation of coronary bloodflow: coordination of heterogeneous control mechanisms in vascular micro-domains. Cardiovasc. Res. 29, 585e596.
Jorgensen, S.M., Demirkaya, O., Ritman, E.L., 1998. Three-dimensional imaging ofvasculature and parenchyma in intact rodent organs with X-ray micro-CT. Am. J.Physiol. 275, H1103eH1114.
Jujo, K., Ii, M., Losordo, D.W., 2008. Endothelial progenitor cells in neo-vascularization of infarcted myocardium. J. Mol. Cell. Cardiol. 45, 530e544.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 73
Kaimovitz, B., Lanir, Y., Kassab, G.S., 2005. Large-scale 3-D geometric reconstructionof the porcine coronary arterial vasculature based on detailed anatomical data.Ann. Biomed. Eng. 33 (11), 1517e1535.
Kajiya, F., Tsujioka, K., Goto, M., Wada, Y., Tadaoka, S., Nakai, M., Hiramatsu, O.,Ogasawara, Y., Mito, K., Hoki, N., 1985. Evaluation of phasic blood flow velocityin the great cardiac vein by a laser Doppler method. Heart Vess 1, 16e23.
Kajiya, F., Tsujioka, K., Goto, M., Wada, Y., Chen, X.L., Nakai, Mm, Tadaoka, S.,Hiramatsu, O., Ogasawara, Y., Mito, K., 1986. Functional characteristics ofintramyocardial capacitance vessels during diastole in the dog. Circ. Res. 198658(4), 476e485.
Kajiya, F., Tsujioka, K., Ogasawara, Y., Hiramatsu, O., Wada, Y., Goto, M., Yanak, M.,1989. Analysis of the characteristics of the flow velocity waveforms in left atrialsmall arteries and veins in the dog. Circ. Res. 65, 1172e1181.
Kajiya, F., Zamir, M., Carlier, S., 2005. Cardiac hemodynamics, coronary circulationand interventional cardiology. Ann. Biomed. Eng. 33 (12), 1728e1734.
Kajiya, F., Yada, T., Hiramatsu, O., Ogasawara, Y., Inai, Y., Kajiya, M., 2008. Coronarymicrocirculation in the beating heart. Med. Biol. Eng. Comput. 46 (5), 411e419.
Kanatsuka, H., Lamping, K.G., Eastham, C.L., Dellsperger, K.C., Marcus, M.L., 1989.Comparison of the effects of increased myocardial oxygen consumption andadenosine on the coronary microvascular resistance. Circ. Res. 65, 1296e1305.
Kassab, G.S., Fung, Y.C., 1994. Topology and dimensions of pig coronary capillarynetwork. Am. J. Physiol. 267 (1), H319eH325.
Kassab, G.S., Rider, C.A., Tang, N.J., Fung, Y.C.B., 1993. Morphometry of pig coronaryarterial trees. Am. J. Physiol. Heart Circ. Physiol. 265, H350eH365.
Kassab, G.S., Lin, D.H., Fung, Y.C., 1994. Morphometry of pig coronary venoussystem. Am. J. Physiol. 267 (6), H2100eH2113.
Kassab, G.S., Berkley, J., Fung, Y.C., 1997. Analysis of pig’s coronary arterial blood flowwith detailed anatomical data. Ann. Biomed. Eng. 25 (1), 204e217.
Kimura, A., Hiramatsu, O., Wada, Y., Yada, T., Yamamoto, T., Goto, M., Ogasawara, Y.,Tsujioka, K., Kajiya, F., 1992. Atrial contractility affects phasic blood flow velocityof atrial small vessels in the dog. Cardiovasc. Res. 26 (12), 1219e1225.
Kiyooka, T., Hiramatsu, O., Shigeto, F., Nakamoto, H., Tachibana, H., Yada, T.,Ogasawara, Y., Kajiya, M., Morimoto, T., Morizane, Y., Mohri, S., Shimizu, J.,Ohe, T., Kajiya, F., 2005. Direct observation of epicardial coronary capillaryhemodynamics during reactive hyperemia and during adenosine administra-tion by intravital video microscopy. Am. J. Physiol. Heart Circ. Physiol. 288,H1437eH1443.
Klitzman, B., Duling, B.R., 1979. Microvascular hematocrit and red cell flow inresting and contracting striated muscle. Am. J. Physiol. 237, H481eH490.
Knaapen, P., Camici, P.G., Marques, K.M., Nijveldt, R., Bax, J.J., Westerhof, N.,Götte, M.J., Jerosch-Herold, M., Schelbert, H.R., Lammertsma, A.A., vanRossum, A.C., 2009. Coronary microvascular resistance: methods for its quan-tification in humans. Basic Res. Cardiol. 104 (5), 485e498.
Kojimahara, M., 1990. Endothelial cilia in rat mesenteric arteries and intra-myocardial capillaries. Z. Mikrosk. Anat. Forsch 104, 412e416.
Kolyva, C., Spaan, J.A.E., Piek, J.J., Siebes, M., 2008. Windkesselness of coronaryarteries hampers assessment of human coronary wave speed by single-pointtechnique. Am. J. Physiol. Heart Circ. Physiol. 295, H482eH490.
Kolyva, C., Pantalos, G.M., Pepper, J.R., Khir, A.W., 2010. How much of the intraaorticballoon volume is displaced toward the coronary circulation? J. Thorac. Car-diovasc. Surg. 140, 110e116.
Komaru, T., Kanatsuka, H., Shirato, K., 2000. Coronary microcirculation: physiologyand pharmacology. Pharmacol. Ther. 86, 217e261.
Kouwenhoven, E., Vergroesen, I., Han, Y., Spaan, J.A., 1992. Retrograde coronary flowis limited by time-varying elastance. Am. J. Physiol. Heart Circ. Physiol. 263,H484eH490.
Krams, R., van Haelst, A.C., Sipkema, P., Westerhof, N., 1989a. Can coronary systo-licediastolic flow differences be predicted by left ventricular pressure or time-varying intramyocardial elastance? Basic Res. Cardiol. 84, 149e159.
Krams, R., Sipkema, P., Westerhof, N., 1989b. Varying elastance concept may explaincoronary systolic flow impediment. Am. J. Physiol. 257, H1471eH1479.
Krogh, A., 1919a. The number and distribution of capillaries in muscle with calcu-lations of the oxygen pressure head necessary for supplying the tissue. J.Physiol. (Lond.) 52, 409e415.
Krogh, A., 1919b. The supply of oxygen to the tissues and the regulation of thecapillary circulation. J. Physiol. 52, 457e474.
Ku, D.N., Giddens, D.P., 1983. Pulsatile flow in a model carotid bifurcation. Arterioscl3 331e319.
Kuikka, J., Levin, M., Bassingthwaighte, J.B., 1986. Multiple tracer dilution estimatesof D- and 2-deoxy-D-glucose uptake by the heart. Am. J. Physiol. 250, H29eH42.
Kumamaru, K.K., Hoppel, B.E., Mather, R.T., Rybicki, F.J., 2010. CT angiography:current technology and clinical use. Radio. Clin. North Am. 48, 213e235.
Kuo, L., Arko, F., Chilian, W.M., Davis, M.J., 1993. Coronary venular responses to flowand pressure. Circ. Res. 72, 607e615.
Kuo, L., Davies, M.J., Chilian, W.M., 1995. Longitudinal gradients for endothelium-dependent and independent vascular responses in the coronary microcircula-tion. Circualtion 92, 518e525.
Kwon, G.P., Schroeder, J.L., Amar, M.J., Remaley, A.T., Balaban, R.S., 2008. Contribu-tion of macromolecular structure to the retention of low-density lipoprotein atarterial branch points. Circ 117, 2919e2927.
Langille, B.L., O’Donnel, F., 1986. Reductions in arterial diameter pro-duced bychronic decreases in blood flow are endothelium-dependent. Science 231,405e407.
Lee, J., Beighley, P., Ritman, E., Smith, N., 2007. Automatic segmentation of 3D micro-CT coronary vascular images. Med. Image. Anal. 11 (6), 630e647.
Lee, J., Niederer, S., Nordsletten, D., Le Grice, I., Smail, B., Kay, D., Smith, N., 2009.Coupling contraction, excitation, ventricular and coronary blood flow across scaleandphysics in theheart. Phil. Trans. A.Math. Phys. Eng. Sci. 367 (1896), 2311e2331.
Lehoux, S., Castier, Y., Tedgui, A., 2006. Molecular mechanisms of the vascularresponses to haemodynamic forces. J. Intern. Med. 259, 381e392.
Leuprecht, A., Perktold, K., 2001. Computer simulation of non-Newtonian effects ofblood flow in large arteries. Comp. Meth. Biomech. Biomed. Engg. 4 (2),149e163.
Levesque, M.J., Nerem, R.M., 1985. The elongation and orientation of culturedendothelial cells in response to shear stress. J. Biomech. Eng. 107, 341e347.
Li, Z.J., Fung, Y.C., 2002. Changes of zero-bending-moment states and structures ofrat arteries in response to a step lowering of the blood pressure. Ann. Biomed.Eng. 30, 379e391.
Liao, J.C., Kuo, L., 1997. Interaction between adenosine and flow-induced dilation incoronary microvascular network. Am. J. Physiol. 272, H1571eH1581.
Lieber, B., Siebes, M., Yamaguchi, T., 2005. Correlation of hemodynamic events withclinical and pathological observations. Ann. Biomed. Eng. 33, 1695e1703.
Lindner, O., Vogt, J., Kammeier, A., Wielepp, P., Holzinger, J., Baller, D., Lamp, B.,Hansky, B., Körfer, R., Horstkotte, D., Burchert, W., 2005. Effect of cardiacresynchronization therapy on global and regional oxygen consumption andmyocardial blood flow in patients with non-ischaemic and ischaemic cardio-myopathy. Eur. Heart J. 26 (1), 70e76.
Liu, Y., Kassab, G.S., 2007. Vascular metabolic dissipation in Murray’s law. Am. J.Physiol. Heart Circ. Physiol. 292 (3), H1336eH1339.
Loukas, M., Bilinsky, S., Bilinsky, E., Petru, M., Anderson, R.H., 2009a. The clinicalanatomy of the coronary collateral circulation. Clin. Anat. 22 (1), 146e160.
Loukas, M., Bilinsky, S., el-Sedfy, A., Anderson, R.H., 2009b. Cardiac veins: a reviewof the literature. Clin. Anat. 22 (1), 129e145.
Loukas, M., Groat, C., Khangura, R., Owens, D.G., Anderson, R.H., 2009c. Thenormal and abnormal anatomy of the coronary arteries. Clin. Anat. 22 (1),114e128.
Lynch, D.G., Waters, S.L., Pedley, T.J., 1996. Flow in a tube with non-uniform, time-dependent curvature: governing equations and simple examples. J. Fluid Mech.323, 237e265.
Mac Gabhann, F., Popel, A.S., 2007. Interactions of VEGF isoforms with VEGFR-1,VEGFR-2, and neuropilin in vivo: a computational model of human skeletalmuscle. Am. J. Physiol. Heart Circ. Physiol. 292, H459eH474.
Mac Gabhann, F., Popel, A.S., 2008. Systems biology of vascular endothelial growthfactors. Microcirc 15, 715e738.
Mac Gabhann, F., Ji, J.W., Popel, A.S., 2007a. Multi-scale computational models ofpro-angiogenic treatments in peripheral arterial disease. Ann. Biomed. Eng. 35,982e994.
Mac Gabhann, F., Ji, J.W., Popel, A.S., 2007b. VEGF gradients, receptor activation, andsprout guidance in resting and exercising skeletal muscle. J. Appl. Physiol. 102,722e734.
Mac Gabhann, F, Annex, B.H., Popel, A.S., Gene Therapy from the perspective ofSystems Biology. Curr. Opinion Mol. Therap., in press-a.
Mac Gabhann, F., Qutub, A.A., Annex, B.H., Popel, A.S., Systems biology of pro-angiogenic therapies targeting the VEGF system. Wiley InterdisciplinaryReviews: Systems Biology & Medicine, in press-b.
Macklin, P., McDougall, S., Anderson, A.R., Chaplain, M.A., Cristini, V., Lowengrub, J.,2009. Multiscale modelling and nonlinear simulation of vascular tumourgrowth. J. Math. Biol. 58, 765e798.
Markl, M., Arnold, R., Hirtler, D., von Zur Muhlen, C., Harloff, A., Langer, M., Hennig, J.,Frydrychowicz, A., 2009. Three-dimensional flow characteristics in aorticcoarctation and poststenotic dilatation. J. Comput. Assist. Tomogr. 33, 776e778.
Martini, P., Pierach, A., Schreyer, E., 1930. Die Strömung des Blutes in engen Gefäben.Eine Abweichung vom Poiseuille’schen Gesetz. Dt. Arch. Klin. Med. 169,212e222.
Matsumoto, T., Kajiya, F., 2005. Coronary microcirculation: physiology andmechanics. Fluid Dyn. Res. 37, 60e81.
Matsumoto, T., Tachibana, H., Asano, T., Takemoto, M., Ogasawara, Y., Umetani, K.,Kajiya, F., 2004. Pattern differences between distributions of microregionalmyocardial flows in crystalloid- and blood-perfused rat hearts. Am. J. Physiol.Heart Circ. Physiol. 286, H1331eH1338.
Matthys, K.S., Alastruey, J., Peiró, J., Khir, A.W., Segers, P., Verdonck, P.R., Parker, K.H.,Sherwin, S.J., 2007. Pulse wave propagation in a model human arterial network:assessment of 1-D numerical simulations against in vitro measurements. J.Biomech. 40, 3476e3486.
Mayerich, D., Abbott, L., McCormick, B., 2008. Knife-edge scanning microscopy forimaging and reconstruction of three-dimensional anatomical structures of themouse brain. J. Microsc. 231 (Pt 1), 134e143.
May-Newman, K., McCulloch, A.D., 1998. Homogenization modeling for themechanics of perfused myocardium. Prog. Biophys. Mol. Biol. 69 (2e3),463e481.
Merkus, D., Kajiya, F., Vink, H., Vergroesen, I., Dankelman, J., Goto, M., Spaan, J.A.,1999. Prolonged diastolic time fraction protects myocardial perfusion whencoronary blood flow is reduced. Circulation 100, 75e81.
Merkus, D., Vergroesen, I., Hiramatsu, O., Tachibana, H., Nakamoto, H., Toyota, E.,Goto, M., Ogasawara, Y., Spaan, J.A.E., Kajiya, F., 2001. Stenosis differentiallyaffects subendocardial and subepicardial arterioles in vivo. Am. J. Physiol. HeartCirc. Physiol. 280, H1674eH1682.
Milkiewicz, M., Ispanovic, E., Doyle, J.L., Haas, T.L., 2006. Regulators of angiogenesisand strategies for their therapeutic manipulation. Int. J. Biochem. Cell Biol. 38,333e357.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7674
Miller Jr., F.J., Dellsperger, K.C., Gutterman, D.D., 1997. Myogenic constriction ofjuman coronary arterioles. Am. J. Physiol. Heart Circ. Physiol. 273, H257eH264.
Miller, K., 2010. The Physiome, a mission imperative. Biomed. Comput. Rev., 8e15.Summer 2010. www.biomedicalcomputationreview.org.
Mitta, I.N., Zhou, Y., Ung, S., Linares, C., Molloi, S., Kassab, G.S., 2005. A computerreconstruction of the entire coronary arterial tree based on detailed morpho-metric data. Ann. Biomed. Eng. 33 (8), 1015e1026.
Mo, M., Eskin, S.G., Schilling, W.P., 1991. Flow-induced changes in Ca2 signaling ofvascular endothelial cells: effects of flow and ATP. Am. J. Physiol. 260,H1698eH1707.
Mochizuki, S., Vink, H., Hiramatsu, O., Kajita, T., Shigeto, F., Spaan, J.A., Kajiya, F.,2003. Role of hyaluronic acid glycosaminoglycans in shear-induced endothe-lium-derived nitric oxide release. Am. J. Physiol. Heart Circ. Physiol. 285 (2),H722eH726.
Moore, J.E., Weydahl, E.S., Santamarina, A., 2001. Frequency dependence of dynamiccurvature effects on flow through coronary arteries. J. Biomech. Eng. 123,129e133.
Moore, J.E., Timmins, L.H., Ladisa, J.F., Coronary artery bifurcation biomechanics andimplications for interventional strategies. Catheter Cardiovasc. Interv., in press.
Mulvany, M.J., 2008. Small artery remodelling in hypertension: causes, conse-quences and therapeutic implications. Med. Biol. Eng. Comput. 46, 461e467.
Murray, C.D., 1926. The physiological principle of minimum work: I. The vascularsystem and the cost of blood volume. Proc. Natl. Acad. Sci. U S A. 12 (3), 207e214.
Mynard, J.P., Nithiarasu, P., 2008. A 1D arterial blood flow model incorporatingventricular pressure, aortic valve and regional coronary flow using the locallyconservative Galerkin (LCG) method. Com. Num. Meth. Eng. 24, 367e417.
Narula, J., Strauss, H.W., 2007. The popcorn plaques. Nat. Med. 13, 532e534.Nerem, R.M., Levesque, M.J., Cornhill, J.F., 1981. Vascular endothelial morphology as
an indicator of the pattern of blood flow. J. Biomech. Eng. 103, 172e176.Nerem, R.M., Alexander, R.W., Chappell, D.C., Medford, R.M., Varner, S.E.,
Taylor, W.R., 1998. Study of the influence of flow on vascular endothelialbiology. Am. J. Med. Sci. 316, 169e175.
Ng, E.Y.K., Ghista, D.N., Jegathese, R.C., 2005. Perfusion studies of steady flow inporoelastic myocardium tissue. Comp. Meth. Biomech. Biomed. Eng. 8 (6),349e357.
Nordsletten, D.A., Niederer, S.A., Nash, M.P., Hunter, P.J., Smith, N.P., 2011. CouplingMulti-Physics Models to Cardiac Mechanics. Prog. Bio. Phys. Mol. Bio 104, 77e87.
O’Connell, B., Walsh, M., 2010. Demonstrating the influence of compression onartery wall mass transport. Ann. Biomed. Eng. 38, 1354e1366.
Olesen, S.P., Clapham, D.E., Davies, P.F., 1988. Hemodynamic shear stress activitiesa Kþ current in vascular endothelial cells. Nature 331, 168e170.
Olufsen, M.S., Peskin, C.S., Kim, W.Y., Pedersen, E.M., Nadim, A., Larsen, J., 2000.Numerical simulation and experimental validation of blood flow in arterieswith structured-tree outflow conditions. Ann. Biomed. Eng. 28, 1281e1299.
Oomens, C.W.J., Hendriks, M.A.N., van Ratingen, M.R., Janssen, J.D., Kok, J.J., 1993. ANumerical e experimental method for a mechanical characterization of bio-logical materials. J. Biomech. 26 (4/5), 617e621.
Owen, M.R., Alarcon, T., Maini, P.K., Byrne, H.M., 2009. Angiogenesis and vascularremodelling in normal and cancerous tissues. J. Math. Biol. 58, 689e721.
Pahakis, M.Y., Kosky, J.R., Dull, R.O., Tarbell, J.M., 2007. The role of endothelial gly-cocalyx components in mechanotransduction of fluid shear stress. Biochem.Biophys. Res. Comm. 355, 228e233.
Painter, P.R., Edén, P., Bengtsson, H.U., 2006. Pulsatile blood flow, shear force, energydissipation and Murray’s Law. Theor. Biol. Med. Model. 3, 31.
Papadakis, G., 2009. Coupling 3D and 1D fluidestructure-interaction models forwave propagation in flexible vessels using a finite volume pressure-correctionscheme. Commun. Numer. Meth. Eng. 25, 533e551.
Parker, K.H., Jones, C.J., 1990. Forward and backward running waves in the arteries:analysis using the method of characteristics. J. Biomech. Eng. 112, 322e326.
Parker, K., 2009. An introduction to wave intensity analysis. Med. Biol. Eng. Comput.47, 175e188.
Passerini, A., Polacek, D.C., Shi, C., Francesco, N.M., Manduchi, E., Grant, G.,Pritchard, W.P., Powell, S.J., Chang, G., Stoeckert, C., Davies, P.F., 2004. Coexistingpro-inflammatory and anti-oxidative endothelial transcription profiles ina disturbed flow region of the adult porcine aorta. Proc. Natl. Acad. Sci. 101,2482e2487.
Passerini, T., De Luca, M.R., Formaggia, L., Quarteroni, A., Veneziani, A., 2009. A3D/1D geometrical multiscale model of cerebral vasculature. J. Eng. Math. 64,319e330.
Peirce, S.M., Van Gieson, E.J., Skalak, T.C., 2004. Multicellular simulation predictsmicrovascular patterning and in silico tissue assembly. FASEB. J. 18, 731e733.
Peirce, S.M., 2008. Computational and mathematical modeling of angiogenesis.Microcirc 15, 739e751.
Peiró, J., Veneziani, A., 2009. Reduced models of the cardiovascular system. In:Formaggia, L., Quarteroni, A., Veneziani, A. (Eds.), Cardiovascular Mathe-matics. Modelling and Simulation of the Circulatory System. Springer, Milan,pp. 347e394.
Pohl, U., Holtz, J., Busse, R., Bassenge, E., 1986. Crucial role of endothelium in thevasodilator response to increased flow in vivo. Hypertension 8, 37e44.
Pohl, U., de Wit, C., Gloe, T., 2000. Large arterioles in the control of blood flow: roleof endothelium- dependent dilation. Acta Physiol. Scand. 168, 505e510.
Ponzini, R., Lemma, M., Morbiducci, U., Montevecchi, F.M., Redaelli, A., 2008.Doppler derived quantitative flow estimate in coronary artery bypass graft:a computational multiscale model for the evaluation of the current clinicalprocedure. Med. Eng. Phys. 30, 809e816.
Pop, R., Richardson, G., Waters, S.L., Jensen, O.E., 2007. Shock formation andnonlinear dispersion in a microvascular capillary network. Math. Med. Biol. 24,379e400.
Popel, A.S., Hunter, P.J., 2009. Systems biology and physiome projects. WileyInterdiscip. Rev. Syst. Biol. Med. 1, 153e158.
Popel, A.S., Johnson, P.C., 2005. Microcirculation and hemorheology. Ann. Rev. FluidMech. 27, 43e69.
Popel, A.S., 1989. Theory of oxygen transport to tissue. Crit. Rev. Biomed. Eng. 17,257e321.
Porter, W.T., 1898. The influence of the heart-beat on the flow of blood through thewalls of the heart. Am. J. Physiol. 1, 145e163.
Pries, A.R., Secomb, T.W., 2005. Microvascular blood viscosity in vivo and theendothelial surface layer. Am. J. Physiol. Heart Circ. Physiol. 289, H2657eH2664.
Pries, A.R., Secomb, TW., 2008. Modeling structural adaptation of microcirculation.Microcirculation 15, 753e764.
Pries, A.R., Ley, K., Claassen, M., Gaehtgens, P., 1989. Red cell distribution atmicrovascular bifurcations. Microvasc. Res. 38, 81e101.
Pries, A.R., Secomb, T.W., Gaehtgens, P., Gross, J.F., 1990. Blood flow in microvascularnetworks. Experiments and simulation. Circ. Res. 67, 826e834.
Pries, A.R., Neuhaus, D., Gaehtgens, P., 1992. Blood viscosity in tube flow: depen-dence on diameter and hematocrit. Am. J. Physiol. 263, H1770eH1778.
Pries, A.R., Secomb, T.W., Gessner, T., Sperandio, M.B., Gross, J.F., Gaehtgens, P., 1994.Resistance to blood flow in microvessels in vivo. Circ. Res. 75, 904e915.
Pries, A.R., Secomb, T.W., Gaehtgens, P., 1995. Design principles of vascular beds.Circ. Res. 77 (5), 1017e1023.
Pries, A.R., Secomb, T.W., Gaehtgens, P., 1998. Structural adaptation and stabilityof microvascular networks: theory and simulations. Am. J. Physiol. 275,H349eH360.
Pries, A.R., Secomb, T.W., Gaehtgens, P., 1999. Structural autoregulation of terminalvascular beds: vascular adaptation and development of hypertension. Hyper-tension 33, 153e161.
Pries, A.R., Secomb, T.W., Gaehtgens, P., 2000. The endothelial surface layer. PflugersArch. 440, 653e666.
Pries, A.R., Reglin, B., Secomb, T.W., 2001. Structural adaptation of microvascularnetworks: functional roles of adaptive responses. Am.J. Physiol. Heart Circ.Physiol 281, H1015eH1025.
Pries, A.R., Reglin, B., Secomb, T.W., 2005. Remodeling of blood vessels: responses ofdiameter and wall thickness to hemodynamic and metabolic stimuli. Hyperten46, 725e731.
Quarteroni, A., Formaggia, L., 2004. Mathematical modelling and numerical simu-lation of the cardiovascular system. In: Ayache, N. (Ed.), Computational Modelsfor the Human Body. Elsevier, Amsterdam.
Qutub, A.A., Liu, G., Vempati, P., Popel, A.S., 2009a. Integration of angiogenesismodules at multiple scales: from molecular to tissue. Proc. Pac. Symp. Bio-comput. 14, 316e327.
Qutub, A.A., Mac Gabhann, F., Karagiannis, E.D., Vempati, P., Popel, A.S., 2009b.Multiscale models of angiogenesis. I.E.E.E. Eng. Med. Biol. Mag 28, 14e31.
Raghavan, M.L., Vorp, D.A., 2000. Toward a biomechanical tool to evaluate rupturepotential of abdominal aortic aneurysm: identification of a finite strain consti-tutive model and evaluation of its applicability. J. Biomech. 33 (4), 475e482.
Rammos, K.S., Koullias, G.J., Papou, T.J., Bakas, A.J., Panagopoulos, P.G.,Tsangaris, S.G., 1998. A computer model for the prediction of left epicardialcoronary blood flow in normal, stenotic and bypassed coronary arteries, bysingle or sequential grafting. Cardiov. Surg. 6, 635e648.
Reitsma, S., Slaaf, D., Vink, H., van Zandvoort, M., oude Egbrink, M., 2007. Theendothelial glycocalyx: composition, functions, and visualization. Pflügers Arch.454, 345e359.
Remmelink, M., Sjauw, K.D., Henriques, J.P.S., de Winter, R.J., Koch, K.T., van derSchaaf, R.J., Vis, M.M., Tijssen, J.G.P., Piek, J.J., Baan, J., 2007. Effects of leftventricular unloading by Impella recover LP2.5 on coronary hemodynamics.Catheter Cardiovasc. Interv. 70, 532e537.
Reneman, R.S., Hoeks, A.P., 2008. Wall shear stress as measured in vivo: conse-quences for the design of the arterial system. Med. Biol. Eng. Comput. 46,499e507.
Resnick, N., Yahav, H., Shay-Salit, A., Shushy, M., Schubert, S., Zilberman, L.C.,Wofovitz, E., 2003. Fluid shear stress and the vascular endothelium: for betterand for worse. Prog. Biophys. Mol. Biol. 81, 177e199.
Revellin, R., Rousset, F., Baud, D., Bonjour, J., 2009. Extension of Murray’s law usinga non-Newtonian model of blood flow. Theor. Biol. Med. Model. 6, 7.
Reymond, P., Merenda, F., Perren, F., Rüfenacht, D., Stergiopulos, N., 2009. Validationof a one-dimensional model of the systemic arterial tree. Am. J. Physiol. HeartCirc. Physiol. 297, H208eH222.
Ricotta, J., Pagan, J., Xenos, M., Alemu, Y., Einav, S., Bluestein, D., 2008. Cardiovas-cular disease management: the need for better diagnostics. Med. Biol. Eng.Comput. 46, 1059e1068.
Rodbard, S., 1975. Vascular caliber. Cardiol 60, 4e49.Rubio, R., Berne, R.M., 1975. Regulation of coronary blood flow. Prog. Cardiovasc. Dis.
18, 105e122.Sands, G.B., Gerneke, D.A., Hooks, D.A., Green, C.R., Smaill, B.H., Legrice, I.J., 2005.
Automated imaging of extended tissue volumes using confocal microscopy.Microsc. Res. Tech. 67 (5), 227e239.
Sarkar, A., Jayaraman, G., 1998. Correction to flow rate e pressure drop relation incoronary angioplasty: steady streaming effect. J. Biomech. 31, 781e791.
Sarkar, A., Jayaraman, G., 2001. Nonlinear analysis of oscillatory flow in the annulus ofan elastic tube e application to catheterized artery. Phys. Fluids 13, 2901e2911.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e76 75
Schaper, W., 2009. Collateral circulation: past and present. Bas. Res. Cardiol. 104,5e21.
Scheel, K.W., Mass, H., Williams, S.E., 1989. Pressure-flow characteristics of coronarycollaterals in dogs. Am. J. Physiol. Heart Circ. Physiol. 256, H441eH445.
Schreiner, W., Buxbaum, P.F., 1993. Computer-optimization of vascular trees. IEEE.Trans. Biomed. Eng. 40 (5), 482e491.
Schuijf, J.D., Achenbach, S., Zoghbi, W.A., Boersma, E., Raggi, P., Weber, M., Nagel, E.,Narula, J., Wackers, F.J.T., Poldermans, D., Bax, J.J., 2009. How to identify theasymptomatic high-risk patient? Curr. Probl. Cardiol. 34, 539e577.
Schwartz, L.M., Bukowski, T.R., Revkin, J.H., Bassingthwaighte, J.B., 1999. Cardiacendothelial transport and metabolism of adenosine and inosine. Am. J. Physiol.277, H1241eH1251.
Secomb, T.W., Pries, A.R., 2002. Information transfer in microvascular networks.Microcirc 9, 377e387.
Secomb, T.W., Hsu, R., Pries, A.R., 2001. Motion of red blood cells in a capillary withan endothelial surface layer: effect of flow velocity. Am. J. Physiol. Heart Circ.Physiol. 281, H629eH636.
Secomb, T.W., 1987. Flow-dependent rheological properties of blood in capillaries.Microvasc. Res. 34, 46e58.
Secomb, T.W., 1995. Mechanics of blood flow in the microcirculation. Symp. Soc.Exp.Biol. 49, 305e321.
Secomb, T.W., 2008. Theoretical models for regulation of blood flow. Microcirc 15,765e775.
Semenza, G.L., 2004. Hydroxylation of HIF-1: oxygen sensing at the molecular level.Physiol 19, 176e182.
Sherman, T.F., Popel, A.S., Koller, A., Johnson, P.C., 1989. The cost of departure fromoptimal radii in microvascular networks. J. Theor. Biol. 136 (3), 245e265.
Sherwin, S.J., Franke, V., Peiró, J., Parker, K., 2003. One-dimensional modelling ofa vascular network in space-time variables. J. Eng. Math. 47, 217e250.
Shipley, R.J., Chapman, S.J., 2010. Multiscale modeling of fluid and drug transport invascular tumors. Bull. Math. Biol. 72 (6), 1464e1491.
Shipley, R.J., Chapman, S.J., Roose, T., Multiscale model predictions of fluid andvinblastine perfusion in dorsal skinfold chambers. submitted for publication.
Shiu, Y.T., Weiss, J.A., Hoying, J.B., Iwamoto, M.N., Joung, I.S., Quam, C.T., 2005. Therole of mechanical stresses in angiogenesis. Crit. Rev. Biomed. Eng. 33, 431e510.
Shyy, J.Y., Chien, S., 2002. Role of integrins in endothelial mechanosensing of shearstress. Circ. Res. 91, 769e775.
Siebes, M., Ventikos, Y., 2010. The role of biofluid mechanics in the assessment ofclinical and pathological observations. Ann. Biomed. Eng. 38, 1216e1224.
Siebes, M., Verhoeff, B.-J., Meuwissen, M., de Winter, R.J., Spaan, J.A.E., Piek, J.J.,2004. Single-wire pressure and flow velocity measurement to quantify coro-nary stenosis hemodynamics and effects of percutaneous interventions. Circ109, 756e762.
Siebes, M., Kolyva, C., Verhoeff, B.J., Piek, J.J., Spaan, J.A., 2009. Potential and limi-tations of wave intensity analysis in coronary arteries. Med. Biol. Eng. Comput.47, 233e239.
Sinha Roy, A., Back, L.H., Banerjee, R.K., 2006. Guidewire flow obstruction effect onpressure drop-flow relationship in moderate coronary artery stenosis. J. Bio-mech. 39, 853e864.
Slomka, P.J., Berman, D.S., Germano, G., 2008. Applications and software techniquesfor integrated cardiac multimodality imaging. Expert Rev. Cardiovasc. Ther. 6,27e41.
Smith, N.P., Pullan, A.J., Hunter, P.J., 2000. Generation of an anatomically basedgeometric coronary model. Ann. Biomed. Eng. 28 (1), 14e25.
Smith, N.P., Pullan, A.J., Hunter, P.J., 2002. An anatomically based model of transientcoronary blood flow in the heart. SIAM J. Appl. Math. 62, 990e1018.
Smith, M.L., Long, D.S., Damiano, E.R., Ley, K., 2003. Near-wall m-PIV revealsa hydrodynamically relevant endothelial surface layer in venules in vivo. Bio-phys. J. 85, 637e645.
Smith, N.P., 2004. A computational study of the interaction between coronary bloodflow and myocardial mechanics. Physiol. Meas. 25 (4), 863e877.
Sorop, O., Spaan, J.A., Sweeney, T.E., vanBavel, E., 2003. Effect of steady versusoscillatory flow on porcine coronary arterioles: involvement of NO and super-oxide anion. Circ. Res. 92 (12), 1344e1351.
Spaan, J.A., Breuls, N.P., Laird, J.D., 1981. Diastolicesystolic coronary flow differencesare caused by intramyocardial pump action in the anesthetized dog. Circ. Res.49, 584e593.
Spaan, J.A., ter Wee, R., van Teeffelen, J.W., Streekstra, G., Siebes, M., Kolyva, C.,Vink, H., Fokkema, D.S., VanBavel, E., 2005. Visualisation of intramural coronaryvasculature by an imaging cryomicrotome suggests compartmentalisation ofmyocardial perfusion areas. Med. Biol. Eng. Comput. 43 (4), 431e435.
Spaan, J.A., Piek, J.J., Hoffman, J.I., Siebes, M., 2006. Physiological basis of clinicallyused coronary hemodynamic indices. Circ 113 (3), 446e455.
Spaan, J., Kolyva, C., van den Wijngaard, J., ter Wee, R., van Horssen, P., Piek, J.,Siebes, M., 2008. Coronary structure and perfusion in health and disease. Phil.Trans. A Math. Phys. Eng. Sci. 366 (1878), 3137e3153.
Spaan, J.A.E., 1995. Mechanical determinants of myocardial perfusion. Basic Res.Cardiol. 90, 89e102.
Stangeby, D.K., Ethier, C.R., 2002. Computational analysis of coupled blood-wallarterial LDL transport. J. Biomech. Eng. 124, 1e8.
Staughton, T.J., del Rio, J.D., Weinberg, P.D., 2007. Patterns of arterial wall perme-ability at the left coronary bifurcation in rabbits. Athero 196, 207e209.
Steele, B.N., Wan, J., Ku, J.P., Hughes, T.J.R., Taylor, C.A., 2003. In vivo validation ofa one-dimensional finite-element method for predicting blood flow in cardio-vascular bypass grafts. I.E.E.E. Trans. Biomed. Eng. 50, 649e656.
Stefanini, M.O., Wu, F.T., Mac Gabhann, F., Popel, A.S., 2008. A compartment model ofVEGF distribution in blood, healthy and diseased tissues. B.M.C. Syst.Biol. 2, 77.
Steinman, D.A., Taylor, C.A., 2005. Flow imaging and computing: large arteryhemodynamics. Ann. Biomed. Eng. 33, 1704e1709.
Steinman, D.A., 2002. Image-based computational fluid dynamics modeling inrealistic arterial geometries. Ann. Biomed. Engg 30 (4), 483e497.
Stergiopulos, N., Young, D.F., Rogge, T.R., 1992. Computer simulation of arterial flowwith applications to arterial and aortic stenoses. J. Biomech. 25, 1477e1488.
Strahler, A.N., 1957. Quantitative analysis of watershed geomorphology. Trans. Am.Geophys. Union 8 (6), 913e920.
Suga, H., Sagawa, K., Shoukas, A., 1973. Load independance of the instantaneouspressure volume ratio of the canine left ventricle and the effects of epinephrineand heart rate on the ratio. Circ. Res. 32, 314e322.
Sun, Y.H., Anderson, T.J., Parker, K.H., Tyberg, J.V., 2000. Wave-intensity analysis:a new approach to coronary haemodynamics. J. Appl. Physiol. 89, 1636e1644.
Sun, Y.H., Anderson, T.J., Parker, K.H., Tyberg, J.V., 2004. Effects of left ventricularcontractility and coronary vascular resistance on coronary dynamics. Am. J.Physiol. Heart Circ. 286, H1590eH1595.
Suzuki, J., Shimamoto, R., Nishikawa, J., Tomaru, T., Nakajima, T., Nakamura, F.,Shin, W.S., Toyo-oka, T., 1998. Vector analysis of the hemodynamics of athero-genesis in the human thoracic aorta using MR velocity mapping. Am. J. Roent171, 1285e1290.
Taber, L.A., 1998. An optimization principle for vascular radius including the effectsof smooth muscle tone. Biophys. J. 74 (1), 109e114.
Tarbell, J.M., Pahakis, M.Y., 2006. Mechanotransduction and the glycocalyx. J. Intern.Med. 259, 339e350.
Tayebjee, M.H., Lip, G.Y., MacFadyen, R.J., 2004. Collateralization and the response toobstruction of epicardial coronary arteries. Q. J. Med. 97 (5), 259e272.
Thi, M.M., Tarbell, J.M., Weinbaum, S., Spray, D.C., 2004. The role of the glycocalyx inreorganization of the actin cytoskeleton under fluid shear stress: a “bumper-car”0 model. Proc. Natl. Acad. Sci. 101, 16483e16488.
Torii, R., Keegan, J., Wood, N.B., Dowsey, A.W., Hughes, A.D., Yang, G.Z., Firmin, D.N.,Mcg Thom, S.A., Xu, X.Y., 2009. The effect of dynamic vessel motion on hae-modynamic parameters in the right coronary artery: a combined MR and CFDstudy. Br. J. Radiol. 82, S24eS32.
Toyota, E., Ogasawara, Y., Hiramatsu, O., Tachibana, H., Kajiya, F., Yamamori, S.,Chilian, W.M., 2005. Dynamics of flow velocities in endocardial and epicardialcoronary arterioles. Am. J. Physiol. Heart Circ. Physiol. 288, H1598eH1603.
Truesdell, C., Noll, W., 1965. The nonlinear field theories of mechanics. In: Flügge, S.(Ed.), Handbuch der Physik, III/3. Springer, Berlin.
Tschumperlin, D.J., Dai, G., Maly, I.V., Kikuchi, T., Laiho, L.H., McVittie, A.K.,Haley, K.J., Lilly, C.M., So, P.T., Lauffenburger, D.A., Kamm, R.D., Drazen, J.M.,2004. Mechanotransduction through growth-factor shedding into the extra-cellular space. Nature 429, 83e86.
Tsoukias, N.M., Goldman, D., Vadapalli, A., Pittman, R.N., Popel, A.S., 2007. Acomputational model of oxygen delivery by hemoglobin-based oxygen carriersin three-dimensional microvascular networks. J. Theor. Biol. 248 (4), 657e674.
Tsoukias, N.M., 2008. Nitric oxide bioavailability in the microcirculation: insightsfrom mathematical models. Microcirc 15, 813e834.
Tune, J.D., Gorman, M.W., Feigl, E.O., 2004. Matching coronary blood flow tomyocardial oxygen consumption. J. Appl. Physiol. 97, 404e415.
Uhlig, P.N., Baer, R.W., Vlahakes, G.J., Hanley, F.L., Messina, L.M., Hoffman, J.I., 1984.Arterial andvenouscoronarypressure-flowrelations in anesthetizeddogs. Evidencefor a vascular waterfall in epicardial coronary veins. Circ. Res. 55 (2), 238e248.
Urquiza, S.A., Blanco, P.J., Venere, M.J., Feijoo, R.A., 2006. Multidimensionalmodelling for the carotid artery blood flow. Comp. Meth. Appl. Mech. Eng. 195,4002e4017.
Valentin, A., Humphrey, J., 2009a. Modeling effects of axial extension on arterialgrowth and remodeling. Med. Biol. Eng. Comput. 47, 979e987.
Valentin, A., Humphrey, J.D., 2009b. Evaluation of fundamental hypotheses under-lying constrained mixture models of arterial growth and remodelling. Phil.Trans. A Math. Phys. Eng. Sci. 367, 3585e3606.
Valentin, A., Cardamone, L., Baek, S., Humphrey, J.D., 2009. Complementary vaso-activity and matrix remodelling in arterial adaptations to altered flow andpressure. J. R. Soc. Int. 6, 293e306.
Van Bavel, E., Spaan, J.A., 1992. Branching patterns in the porcine coronary arterialtree. Estimation of flow heterogeneity. Circ. Res. 71 (5), 1200e1212.
van den Akker, J., Schoorl, M.J., Bakker, E.N., Vanbavel, E., 2010. Small arteryremodeling: current concepts and questions. J. Vasc. Res. 47 (3), 183e202.
van den Berg, B.M., Spaan, J.A.E., Rolf, T.M., Vink, H., 2006. Atherogenic region anddiet diminish glycocalyx dimension and increase intima-to-media ratios atmurine carotid artery bifurcation. Am. J. Physiol. 290, H915eH920.
van den Broek, C.N., van der Horst, A., Rutten, M.C.M., van de Vosse, F.N., A genericconstitutive model for the passive porcine coronary artery. Biomech. Mod.Mechanobiol., in press.
van den Wijngaard, J., Kolyva, C., Siebes, M., Dankelman, J., van Gemert, M., Piek, J.,Spaan, J., 2008. Model prediction of subendocardial perfusion of the coronarycirculation in the presence of an epicardial coronary artery stenosis. Med. Biol.Eng. Comput. 46, 421e432.
van den Wijngaard, J., Siebes, M., Westerhof, B., 2009. Comparison of arterial wavesderived by classical wave separation and wave intensity analysis in a model ofaortic coarctation. Med. Biol. Eng. Comput. 47, 211e220.
van der Heiden, K., Groenendijk, B.C.W., Hierck, B.P., Hogers, B., Koerten, H.K.,Mommaas, A.M., 2006. Monocilia on chicken embryonic endocardium in lowshear stress areas. Devel. Dyn. 235, 19e28.

S.L. Waters et al. / Progress in Biophysics and Molecular Biology 104 (2011) 49e7676
van der Heiden, K., Hierck, B.P., Krams, R., de Crom, R., Cheng, C., Baiker, M., 2008.Endothelial primary cilia in areas of disturbed flow are at the base of athero-sclerosis. Athero 196, 542e550.
Van Doormaal, M.A., Zhang, J., Wada, S., Shaw, J.E., Won, D., Cybulsky, M.I., Yip, C.M.,Ethier, C.R., 2009. Variations in mass transfer to single endothelial cells. Bio-mech. Mod. Mechanobiol. 8 (3), 183e193.
van Horssen, P., van den Wijngaard, J., Nolte, F., Hoefer, I., Haverslag, R., Spaan, J.,Siebes, M., 2009. Extraction of coronary vascular tree and myocardial perfusiondata from stacks of cryomicrotome images. In: Ayache, N., Delingette, H.,Sermesant, M. (Eds.), Functional Imaging and Modeling of the Heart (LNCS vol.5528). Springer, Berlin-Heidelberg, pp. 486e494.
van Horssen, P., Siebes, M., Hoefer, I., Spaan, J.A.E., van denWijngaard, J.P.H.M., 2010.Improved detection of fluorescently labeled microspheres and vessel architec-ture with an imaging cryomicrotome. Med. Biol. Eng. Comput. 4, 735e744.
Van Meerveld, J., Waters, S.L., 2001. Numerical computation of the steady secondaryflow in a tube with timeedependent curvature. Q.J.Mech. Appl. Maths. 54,631e640.
Vankan, W.J., Huyghe, J.M.R.J., Janssen, J.D., Huson, A., Hacking, W., Schreiner, W.,1997. Finite element analysis of blood perfusion through biological tissue. Int. J.Engng Sci. 35 (4), 375e385.
VanTeeffelen, J.W., Merkus, D., Bos, L.J., Vergroesen, I., Spaan, J.A., 1998. Impairmentof contraction increases sensitivity of epicardial lymph pressure for leftventricular pressure. Am. J. Physiol. Heart Circ. Physiol. 274, H187eH192.
Verberne, H.J., Meuwissen, M., Chamuleau, S.A., Verhoeff, B.J., van Eck-Smit, B.L.,Spaan, J.A., Piek, J.J., Siebes, M., 2007. Effect of simultaneous intracoronaryguidewires on the predictive accuracy of functional parameters of coronarylesion severity. Am. J. Physiol. Heart Circ. Physiol. 292 (5), H2349eH2355.
Verhoeff, B.-J., Siebes, M., Meuwissen, M., Atasever, B., Voskuil, M., de Winter, R.J.,Koch, K.T., Tijssen, J.G.P., Spaan, J.A.E., Piek, J.J., 2005. Influence of percutaneouscoronary intervention on coronary microvascular resistance index. Circulation111, 76e82.
Vincent, P.E., Sherwin, S.J., Weinberg, P.D., 2008. Viscous flow over outflow slitscovered by an anisotropic Brinkman medium: a model of flow above inter-endothelial cell clefts. Phys. Fluids 20, 063106.
Vink, H., Duling, B.R., 1996. Identification of distinct luminal domains for macro-molecules, erythrocytes, and leukocytes within mammalian capillaries. Circ.Res. 79, 581e589.
Vink, H., Duling, B.R., 2000. Capillary endothelial surface layer selectively reducesplasma solute distribution volume. Am. J. Physiol. Heart Circ. Physiol. 278,H285eH289.
Vink, H., Constantinescu, A.A., Spaan, J.A., 2000. Oxidized lipoproteins degrade theendothelial surface layer: implications for plateleteendothelial cell adhesion.Circulation 101, 1500e1502.
Vis, M.A., Bovendeerd, P.H., Sipkema, P., Westerhof, N., 1997. Effect of ventricularcontraction, pressure, and wall stretch on vessels at different locations in thewall. Am. J. Physiol. Heart Circ. Physiol. 272, H2963eH2975.
Voitl, P., Vollkron, M., Bergmeister, H., Wieselthaler, G., Schima, H., 2009. Coronaryhemodynamics and myocardial oxygen consumption during support withrotary blood pumps. Artif. Organs 33, 77e80.
von Restorff, W., Holtz, J., Bassenge, E., 1977. Exercise induced augmentation ofmyocardial oxygen extraction in spite of normal coronary dilatory capacity indogs. Pflugers Arch. 372, 181e185.
Walley, K.R., 1996. Heterogeneity of oxygen delivery impairs oxygen extraction byperipheral tissues: theory. J. Appl. Physiol. 81, 885e894.
Wang, J.J., O’Brien, A.B., Shrive, N.G., Parker, K.H., Tyberg, J.V., 2003. Time-domainrepresentation of ventricularearterial coupling as a windkessel and wavesystem. Am. J. Physiol. Heart Circ. Physiol. 284, H1358eH1368.
Wang, J.J., Flewitt, J.A., Shrive, N.G., Parker, K.H., Tyberg, J.V., 2006. Systemic venouscirculation. Waves propagating on a windkessel: relation of arterial and venouswindkessels to systemic vascular resistance. Am. J. Physiol. Heart Circ. Physiol.290, H154eH162.
Waters, S.L., Pedley, T.J., 1999. Oscillatory flow in a tube of time-dependent curva-ture: part I. Perturbation to flow in a stationary curved tube. J. Fluid Mech. 282,327e352.
Weinbaum, S., Zhang, X., Han, Y., Vink, H., Cowin, S.C., 2003. Mechanotransductionand flow across the endothelial glycocalyx. Proc. Natl. Acad. Sci. U S A 100 (13),7988e7995.
Weinbaum, S., Tarbell, J.M., Damiano, E.R., 2007. The structure and function of theendothelial glycocalyx layer. Annu. Rev. Biomed. Eng. 9, 121e167.
Weinberg, E., Shahmirzadi, D., Mofrad, M., On the multiscale modeling of heartvalve biomechanics in health and disease. Biomech. Model Mechanobiol., inpress.
Westerhof, N., Boer, C., Lamberts, R.R., Sipkema, P., 2006. Cross-talk between cardiacmuscle and coronary vasculature. Physiol. Rev. 86 (4), 1263e1308.
Weydahl, E.S., Moore, J.E., 2001. Dynamic curvature strongly affects wall shear ratesin a coronary artery bifurcation model. J. Biomech. 34, 1189e1196.
Wieringa, P.A., Stassen, H.G., Van Kan, J.J., Spaan, J.A., 1993. Oxygen diffusion ina network model of the myocardial microcirculation. Int. J. Microcirc. Clin. Exp.13, 137e169.
Wischgoll, T., Choy, J.S., Kassab, G.S., 2009. Extraction of morphometry andbranching angles of porcine coronary arterial tree from CT images. Am. J.Physiol. Heart Circ. Physiol. 297, H1949eH1955.
Wu, F., Zhang, E.Y., Zhang, J., Bache, R.J., Beard, D.A., 2008. Phosphate metaboliteconcentrations and ATP hydrolysis potential in normal and ischaemic hearts. J.Physiol. 586, 4193e4208.
Wu, F.T., Stefanini, M.O., Gabhann, F.M., Kontos, C.D., Annex, B.H., Popel, A.S., 2009a.Computational kinetic model of VEGF trapping by soluble VEGF receptor-1:effects of transendothelial and lymphatic macromolecular transport. Physiol.Genomics 38, 29e41.
Wu, F.T., Stefanini, M.O., Mac Gabhann, F., Popel, A.S., 2009b. A compartment modelof VEGF distribution in humans in the presence of soluble VEGF receptor-1acting as a ligand trap. PLoS One 4, e5108.
Wu, F., Zhang, J., Beard, D.A., 2009c. Experimentally observed phenomena oncardiac energetics in heart failure emerge from simulations of cardiac metab-olism. Proc. Natl. Acad. Sci. USA 106, 7143e7148.
Yang, J., Clark Jr., J.W., Bryan, R.M., Robertson, C., 2003a. The myogenic response inisolated rat cerebrovascular arteries: smooth muscle cell model. Med. Eng. Phys.25, 691e709.
Yang, J., Clark Jr., J.W., Bryan, R.M., Robertson, C.S., 2003b. The myogenic response inisolated rat cerebrovascular arteries: vessel model. Med. Eng. Phys. 25, 711e717.
Yang, J., Clark, J.W., Bryan, R.M., Robertson, C.S., 2005. Mathematical modeling of thenitric oxide/cGMP pathway in the vascular smooth muscle cell. Am. J. Physiol.Heart Circ. Physiol. 289, H886eH897.
Yao, Y., Rabodzey, A., Dewey Jr., C.F., 2007. Glycocalyx modulates the motility andproliferative response of vascular endothelium to fluid shear stress. Am. J.Physiol. 293, H1023eH1030.
Young, D.F., 1979. Fluid mechanics of arterial stenoses. J. Biomech. Eng. 101,157e175.
Zamir, M., Bigelow, D.C., 1984. Cost of departure from optimality in arterialbranching. J. Theor. Biol. 109 (3), 401e409.
Zamir, M., 1976. Optimality principles in arterial branching. J. Theor. Biol. 62 (1),227e251.
Zamir, M., 1998. Mechanics of blood supply to the heart: wave reflection effects ina right coronary artery. Proc. Roy. Soc. B. 265, 439e444.
Zhang, W., Herrera, C., Atluri, S.N., Kassab, G.S., 2004. Effect of surrounding tissue onvessel fluid and solid mechanics. J. Biomech. Eng. 126, 760e769.
Zhang, C., Rogers, P.A., Merkus, D., Muller-Delp, J.M., Tiefenbacher, C.P., Potter, B.,Knudson, J.D., Rocic, P., Chilian, W.M., 2008. Regulation of Coronary Microvas-cular Resistance, in Health and Disease in Handbook of Physiology: Microcir-culation. Academic Press.
Zhang, J., Johnson, P.C., Popel, A.S., 2009. Effects of erythrocyte deformability andaggregation on the cell free layer and apparent viscosity of microscopic bloodflows. Microvasc. Res. 77, 265e272.
Zinemanas, D., Beyar, R., Sideman, S., 1994. Effects of myocardial contraction oncoronary blood flow: an integrated model. Ann. Biomed. Eng. 22, 638e652.
Zulliger, M.A., Rachev, A., Stergiopulos, N., 2004. A constitutive formulation ofarterial mechanics including vascular smooth muscle tone. Am. J. Physiol. HeartCirc. Physiol. 287, 1335e1343.
Related Documents











