The trade-off between the light-harvesting and photoprotective functions of fucoxanthin-chlorophyll proteins dominates light acclimation in Emiliania huxleyi (clone CCMP 1516) Boyd A. McKew 1 , Phillip Davey 1 , Stewart J. Finch 1 , Jason Hopkins 2 , Stephane C. Lefebvre 3 , Metodi V. Metodiev 1 , Kevin Oxborough 4 , Christine A. Raines 1 , Tracy Lawson 1 and Richard J. Geider 1 1 School of Biological Sciences, University of Essex, Colchester, CO4 3SQ, UK; 2 National Oceanography Centre, University of Southampton, European Way, Southampton, SO14 3ZH, UK; 3 J. Craig Venter Institute, San Diego, CA, 92121, USA; 4 CTG Ltd, 55 Central Avenue, West Molesey, KT8 2QZ, UK Authors for correspondence: Richard J. Geider Tel: +44 (0)1206 873312 Email: [email protected] Tracy Lawson Tel: +44 (0)1206 3327 Email: [email protected] Received: 21 March 2013 Accepted: 18 May 2013 New Phytologist (2013) doi: 10.1111/nph.12373 Key words: Emiliania huxleyi, gross photo- synthesis, light acclimation, light harvesting, nonphotochemical quenching, photoinacti- vation, photoinhibition. Summary Mechanistic understanding of the costs and benefits of photoacclimation requires knowl- edge of how photophysiology is affected by changes in the molecular structure of the chloro- plast. We tested the hypothesis that changes in the light dependencies of photosynthesis, non- photochemical quenching and PSII photoinactivation arises from changes in the abundances of chloroplast proteins in Emiliania huxleyi strain CCMP 1516 grown at 30 (Low Light; LL) and 1000 (High Light; HL) lmol photons m 2 s 1 photon flux densities. Carbon-specific light-saturated gross photosynthesis rates were not significantly different between cells acclimated to LL and HL. Acclimation to LL benefited cells by increasing bio- mass-specific light absorption and gross photosynthesis rates under low light, whereas accli- mation to HL benefited cells by reducing the rate of photoinactivation of PSII under high light. Differences in the relative abundances of proteins assigned to light-harvesting (Lhcf), photo- protection (LI818-like), and the photosystem II (PSII) core complex accompanied differences in photophysiology: specifically, Lhcf:PSII was greater under LL, whereas LI818:PSII was greater in HL. Thus, photoacclimation in E. huxleyi involved a trade-off amongst the characteristics of light absorption and photoprotection, which could be attributed to changes in the abundance and composition of proteins in the light-harvesting antenna of PSII. Introduction Photosynthetic organisms must maintain a balance between the interception of light energy, the supply of this excitation energy to the photosynthetic reaction centres, the production of NADPH and ATP within the thylakoid membranes, and the uti- lization of NADPH and ATP for CO 2 fixation and for biosyn- thesis. Changes in photosynthetic photon flux density (PFD) on timescales of seconds to days affect this balance, and as a conse- quence plants and algae must employ a range of mechanisms to adjust their physiologies to maintain this balance (Raven & Geider, 2003; Falkowski & Raven, 2007). Photoacclimation is the term used to designate the process of adjustment of the phenotype of plants, algae and cyanobacteria to changes in the light environment. It operates on timescales of hours to days, which are intermediate between rapid changes involved in the regulation of enzyme activities and long time- scales associated with changes in adaptation of the genotype (Raven & Geider, 2003). Photoacclimation involves changes in ultrastructure, morphology, biochemical and elemental composition, pigment content (Falkowski & La Roche, 1991), and the composition of the proteome (Pandhal et al., 2007; McKew et al., 2013). The contrasting demands of aquatic and terrestrial environ- ments, and the contrasting lifestyles of multicellular vascular plants and unicellular microalgae have led to differences in evolu- tionary adaptations and strategies of physiological acclimation (Geider et al., 2001). For example, many vascular plant leaves photoacclimate to high light by increasing the abundance of Calvin cycle enzymes per unit area whilst maintaining light absorption (Seemann et al., 1987; Givnish, 1988), whereas mic- roalgae reduce light harvesting without increasing Calvin cycle enzymes (Sukenik et al., 1987; McKew et al., 2013). These differ- ences in acclimatory strategy are likely to be a consequence of the fact that vascular plant leaves are optically thick and absorb > 90% of incident solar radiation (Knapp & Carter, 1998), whereas microalgae are typically optically thin and absorb < 50% of the light that falls on their surfaces (Morel & Bricaud, 1981). Additionally, vascular plants can often be characterized into sun and shade ecotypes (Givnish, 1988), whereas most microalgae Ó 2013 The Authors New Phytologist Ó 2013 New Phytologist Trust New Phytologist (2013) 1 www.newphytologist.com Research

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The trade-off between the light-harvesting and photoprotectivefunctions of fucoxanthin-chlorophyll proteins dominates lightacclimation in Emiliania huxleyi (clone CCMP 1516)
Boyd A. McKew1, Phillip Davey1, Stewart J. Finch1, Jason Hopkins2, Stephane C. Lefebvre3, Metodi V. Metodiev1,
Kevin Oxborough4, Christine A. Raines1, Tracy Lawson1 and Richard J. Geider1
1School of Biological Sciences, University of Essex, Colchester, CO4 3SQ, UK; 2National Oceanography Centre, University of Southampton, European Way, Southampton, SO14 3ZH, UK;
3J. Craig Venter Institute, San Diego, CA, 92121, USA; 4CTG Ltd, 55 Central Avenue, West Molesey, KT8 2QZ, UK
Authors for correspondence:Richard J. GeiderTel: +44 (0)1206 873312
Email: [email protected]
Tracy Lawson
Tel: +44 (0)1206 3327
Email: [email protected]
Received: 21 March 2013Accepted: 18 May 2013
New Phytologist (2013)doi: 10.1111/nph.12373
Key words: Emiliania huxleyi, gross photo-synthesis, light acclimation, light harvesting,nonphotochemical quenching, photoinacti-vation, photoinhibition.
Summary
� Mechanistic understanding of the costs and benefits of photoacclimation requires knowl-
edge of how photophysiology is affected by changes in the molecular structure of the chloro-
plast.� We tested the hypothesis that changes in the light dependencies of photosynthesis, non-
photochemical quenching and PSII photoinactivation arises from changes in the abundances
of chloroplast proteins in Emiliania huxleyi strain CCMP 1516 grown at 30 (Low Light; LL)
and 1000 (High Light; HL) lmol photons m�2 s�1 photon flux densities.� Carbon-specific light-saturated gross photosynthesis rates were not significantly different
between cells acclimated to LL and HL. Acclimation to LL benefited cells by increasing bio-
mass-specific light absorption and gross photosynthesis rates under low light, whereas accli-
mation to HL benefited cells by reducing the rate of photoinactivation of PSII under high light.
Differences in the relative abundances of proteins assigned to light-harvesting (Lhcf), photo-
protection (LI818-like), and the photosystem II (PSII) core complex accompanied differences
in photophysiology: specifically, Lhcf:PSII was greater under LL, whereas LI818:PSII was
greater in HL.� Thus, photoacclimation in E. huxleyi involved a trade-off amongst the characteristics of
light absorption and photoprotection, which could be attributed to changes in the abundance
and composition of proteins in the light-harvesting antenna of PSII.
Introduction
Photosynthetic organisms must maintain a balance between theinterception of light energy, the supply of this excitation energyto the photosynthetic reaction centres, the production ofNADPH and ATP within the thylakoid membranes, and the uti-lization of NADPH and ATP for CO2 fixation and for biosyn-thesis. Changes in photosynthetic photon flux density (PFD) ontimescales of seconds to days affect this balance, and as a conse-quence plants and algae must employ a range of mechanismsto adjust their physiologies to maintain this balance (Raven &Geider, 2003; Falkowski & Raven, 2007).
Photoacclimation is the term used to designate the process ofadjustment of the phenotype of plants, algae and cyanobacteriato changes in the light environment. It operates on timescales ofhours to days, which are intermediate between rapid changesinvolved in the regulation of enzyme activities and long time-scales associated with changes in adaptation of the genotype(Raven & Geider, 2003). Photoacclimation involves changesin ultrastructure, morphology, biochemical and elemental
composition, pigment content (Falkowski & La Roche, 1991),and the composition of the proteome (Pandhal et al., 2007;McKew et al., 2013).
The contrasting demands of aquatic and terrestrial environ-ments, and the contrasting lifestyles of multicellular vascularplants and unicellular microalgae have led to differences in evolu-tionary adaptations and strategies of physiological acclimation(Geider et al., 2001). For example, many vascular plant leavesphotoacclimate to high light by increasing the abundance ofCalvin cycle enzymes per unit area whilst maintaining lightabsorption (Seemann et al., 1987; Givnish, 1988), whereas mic-roalgae reduce light harvesting without increasing Calvin cycleenzymes (Sukenik et al., 1987; McKew et al., 2013). These differ-ences in acclimatory strategy are likely to be a consequence of thefact that vascular plant leaves are optically thick and absorb> 90% of incident solar radiation (Knapp & Carter, 1998),whereas microalgae are typically optically thin and absorb < 50%of the light that falls on their surfaces (Morel & Bricaud, 1981).Additionally, vascular plants can often be characterized into sunand shade ecotypes (Givnish, 1988), whereas most microalgae
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 1
www.newphytologist.com
Research

and cyanobacteria are evolutionarily adapted to low light(Richardson et al., 1983), although a few notable exceptions ofhigh and low light ecotypes have been identified (Moore &Chisholm, 1999). Within the microalgae and cyanobacteria, therate of photosynthesis is typically saturated at c. 50–400 lmolphotons m�2 s�1 (Richardson et al., 1983), which is significantlyless than full noon sunlight. In most ocean environments photo-synthesis shows a subsurface maximum at a depth correspondingto c. 10–30% of surface PFD (Behrenfeld & Falkowski, 1997),whereas in many vascular plants photosynthesis of ‘sun’ leavesdoes not saturate even in full sunlight (Bjorkman, 1981).
Photoacclimation on timescales of hours to days affects impor-tant physiological processes including the light dependence ofphotosynthesis and the susceptibility of photosystem II (PSII)reaction centres (RCII) to photoinactivation (Richardson et al.,1983; Falkowski & La Roche, 1991; Raven & Geider, 2003).During acclimation to incident light, microalgae modify compo-nents associated with PSII and photosystem I (PSI), (Falkowski& Raven, 2007). These adjustments appear to be driven, in part,through redox and reactive oxygen species (ROS) signalling thatarise from an imbalance between the rate of light absorption andthe rate of energy consumption in photosynthesis and biosynthe-sis (Escoubas et al., 1995; Pfannschmidt et al., 2009), althoughblue-light receptors have also been implicated in the photoaccli-mation of diatoms (Schellenberger Costa et al., 2013).
Knowledge of the cost–benefit trade-offs involved in photo-acclimation is essential for developing a mechanistic understand-ing of how photosynthetic organisms acclimate to the lightenvironment in order to improve fitness (Geider et al., 2009).These costs and benefits can be evaluated within the context ofthree design considerations for structure–function relationshipsin chloroplasts (Raven, 1980). These are the energetic efficiencyof photosynthesis (work output divided by energy input), thecatalytic efficiency of photosynthesis (work output per unit cata-lytic and structural material contained in the chloroplast) andthe provision of mechanisms to ensure safe operation of thephotosynthetic apparatus. Such an analysis is facilitated by thefact that all oxygenic photoautotrophs possess a highly con-served light-driven energy transduction pathway linking O2 evo-lution by PSII to the production of triose phosphates in theCalvin cycle (Blankenship, 2002). By contrast, considerablephylogenetic diversity is evident upstream of PSII in the light-harvesting pigments and proteins amongst the cyanobacteriaand the green and red algal lineages (Blankenship, 2002), aswell as in the downstream metabolism of the products of CO2
fixation (Wilhlem et al., 2006). This includes differences inorganic C storage products, with carbohydrates or neutral lipidsthe major storage products depending on species and growthcondition (Lacour et al., 2012).
In this paper, we examine photoacclimation of the marinehaptophyte Emiliania huxleyi, calcifying strain CCMP 1516,which was the first coccolithophorid to have its genomesequenced by JGI (Read et al., 2013). The haptophytes signifi-cantly affect ocean biogeochemistry through their roles in the car-bon and sulphur cycles. Emiliania huxleyi is a widely distributedspecies that typically blooms in high-latitude seas when and
where the mixed layer is relatively shallow and thus characterizedby high light (Iglesias-Rodriguez et al., 2002). Although its toler-ance of high light is one of the attributes considered to contributeto blooms and the success of this species (Nanninga & Tyrrell,1996), E. huxleyi can grow over a wide range of PFDs (Suggettet al., 2007). As in other photoautotrophs, growth of E. huxleyi inlow light environments requires maximizing both the rate of lightabsorption per unit biomass and the quantum efficiency of pho-tosynthesis, whilst tolerance of high light requires that oxidativestress and consequent cellular damage be minimized by reducingthe production of ROS as well as increasing the scavenging ofunavoidably produced ROS.
We investigated the functional consequences for light absorp-tion, photosynthesis, nonphotochemical excitation energyquenching and photoinactivation that accompany the structuralremodelling of the chloroplast proteome of E. huxleyi in responseto growth at suboptimal and supraoptimal PFDs. We tested thehypotheses that changes in the efficiency of PSII photochemistry,the capacity for nonphotochemical quenching, and the suscepti-bility of RCII to photoinactivation could be explained as conse-quences of changes in the abundances of key chloroplast proteins.To this end, we used selected results from our analysis of theE. huxleyi proteomes under suboptimal and supraoptimal PFDs(McKew et al., 2013) together with new additional measurementsof a suite of photophysiological characteristics. Unlike many pre-viously published investigations, which focused on individualphotophysiological processes in isolation, we have broughttogether observations of photoacclimation of light harvesting,nonphotochemical quenching and photoinactivation in conjunc-tion with changes in the chloroplast proteome to assess the mech-anistic basis of the cost–benefit trade-offs involved inphotoacclimation.
Materials and Methods
Culture conditions and proteomics
Full details of culture conditions and the proteomic analysis aredescribed in McKew et al. (2013). Briefly, cultures of calcifyingEmiliania huxleyi CCMP 1516 were grown to steady-state innutrient-replete turbidostat cultures on a 16 h : 8 h, light : darkcycle at PFDs (k = 400–700 nm) of 30 lmol photons m�2 s�1
(Low Light, LL) or 1000 lmol photons m�2 s�1 (High Light,HL), and nontargeted proteomics of trypsin digested unfraction-ated cell extracts was conducted using LC-electrospray-ionizationtandem mass spectrometry on a hybrid high-resolution LTQ/Or-bitrap Velos instrument.
Pigment content
Samples for pigment analysis were collected on MF300 glassmicrofibre filters (Fisher Scientific, Loughborough, UK). In addi-tion to measuring Chla on methanol extracts (McKew et al.,2013), High Performance Liquid Chromatography (HPLC)analysis was conducted on samples stored under liquid nitrogen.For HPLC analysis, pigments were extracted in 3.0 ml 90%
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist2

acetone using sonication. Extracts were centrifuged to removedebris, and 100-ll aliquots analysed on a 3 lm Hypersil MOS2C8 column using a ThermoFinnigan Spectra HPLC system withThermo Separations AS3000 autosampler, Thermo SeparationsUV6000 diode array absorbance detector, and PC1000 andChromQuest chromotography software (Barlow et al., 1997a,b).Chla (Sigma) and other pigment standards (DHI Institute forWater and Environment, Hørsholm, Denmark), along withretention time and light absorption spectra were used to identifypigments, whilst concentrations were calculated from peak area.
In vivo light absorption
Cells were concentrated by vacuum filtration of 300 ml of cultureonto Whatman 3 lm pore polycarbonate filters (GE Healthcare,Little Chalfont, UK) and resuspended in 5 ml of the filtrate formeasurement of the spectral dependence (380–800 nm) of lightabsorption using a Hitachi U-3000 spectrophotometer fittedwith a 60 mm diameter integrating sphere (Hitachi High-Tech-nologies, Berkshire, UK) correcting for residual scattering as pre-viously described (Suggett et al., 2007). Chla was also measuredon the concentrates to allow Chla-specific light absorption coeffi-cients to be calculated.
Chla-specific light absorption spectra were also reconstructedfrom the measurements of the ratios of accessory pigments toChla and the mass-specific absorption spectra reported for chlo-rophylls, photosynthetic carotenoids and nonphotosynthetic car-otenoids (Bidigare et al., 1990) as described in Suggett et al.(2004) with corrections made for the package effect from mea-surements of equivalent spherical cell diameter and cellular Chlacontent (Morel & Bricaud, 1981).
Fast repetition rate (FRR) fluorometry
Fast repetition rate (FRR) fluorometry (Kolber et al., 1998)was used to obtain fluorescence light curves (FLCs) using theFastTracka II fluorometer and Fast Act laboratory system (bothCTG Ltd, West Molesey, UK). Samples were dark adapted for20 min before each FLC was started. The PFDs used for allFLCs were 5, 19, 34, 48, 121, 237, 444, 853 and1413 lmol m�2 s�1. Each PFD step was run for 5 min. A 20-sdark interval was placed between all adjacent PFD steps, dur-ing which a single dark acquisition was made. FastPro software(CTG Ltd) was used to calculate the dark-adapted values ofminimum and maximum fluorescence (Fo, Fm), the maximalphotochemical efficiency in the dark-adapted state, Fv/Fm andthe PSII effective cross-section (rPSII). At each PFD step thefluorescence under actinic illumination (F 0), the maximum flu-orescence under actinic illumination (Fm0) and the parametersFq0=Fv 0, Fv 0=Fm0 and Fq0=Fm0 were obtained: By conventionFq0=Fm0 is the overall actual operating efficiency of PSII underactinic light, whereas Fv 0=Fm0 provides maximum efficiency ofPSII photochemistry in the light, and Fq0=Fv 0 is the photo-chemical factor, which is nonlinearly related to the proportionof RCII that are in the open state and thus capable ofperforming photochemistry.
Rates of photoinactivation and repair
Rates of PSII photoinactivation were determined at four PFDsfrom 150 to 1200 lmol photons m�2 s�1 by measuring the timedependence of changes in Fv/Fm. Values of Fv/Fm were measuredafter 30 min dark adaptation, following 5, 15, 30 or 60 min lighttreatment at each of the four PFDs. Gross rates of PSII photoin-activation were obtained from measurements on samples incu-bated in the presence of lincomycin (a specific inhibitor ofchloroplast and mitochondrial protein synthesis, Ragni et al.,2008). Rate constants for photoinactivation were obtained bynonlinear curve fitting (SIGMAPLOT�, Systat Software Inc.,London, UK) to the equation:
FvðtÞ=FmðtÞ ¼ Fvð0Þ=Fmð0Þ expð�ki tÞ Eqn 1
(Fv(t)/Fm(t), values measured after 5, 15, 30 and 60 min lightexposure in the presence of lincomycin; ki, inactivation rate con-stant; t, time). The rate constants for net photoinactivation wereobtained from similar measurements made in the absence oflincomycin, and recovery was obtained by difference.
Gross and net oxygen exchange rate The light response curvesfor gross O2 evolution and O2 uptake were determined usingmembrane inlet mass spectrometry (MIMS; model QMG 422;Pfeiffer Vacuum, Asslar, Germany) modified from Kana (1990)as described in Brading et al. (2011). Briefly, samples to which18O2 was added were illuminated for 60–90 min at PFDsbetween 0 and 1200 lmol photons m�2 s�1. The gross O2 evolu-tion rates were calculated from measurements of changes in 36O2
and 32O2 using isotope dilution equations (Kana, 1990). Theserates were then normalized to cell concentration and parametersof the photosynthesis–light (PE) response curve (Jassby & Platt,1976) were determined by nonlinear least squares (SIGMA-PLOT�):
PCell ¼ PCellm � 1� exp �aCell � E
PCellm
! !Eqn 2
(PCell, cell-specific photosynthesis rate; PCellm , light-saturated rate;aCell, initial slope of the PE curve; E, photon flux density). Asexpected, and consistent with Eqn 2, PCell was not significantlydifferent from zero at E = 0.
Results
Growth rate and pigment composition
Emiliania huxleyi 1516 grew over three times faster under HL(1000 lmol photons m�2 s�1) than LL (30 lmol photons m�2 s�1),and, although HL cells were significantly larger, they containedsignificantly less Chla (Table 1). The b-carotene-to-Chla ratio,although low in absolute values, was c. 30% higher in HL than inLL (Table 1) and the total xanthophyll-to-Chla ratio was c. 20%greater in HL than in LL (Table 1). The major light-harvesting xanthophyll 190-hexanoyloxyfucoanthin accounted for
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 3
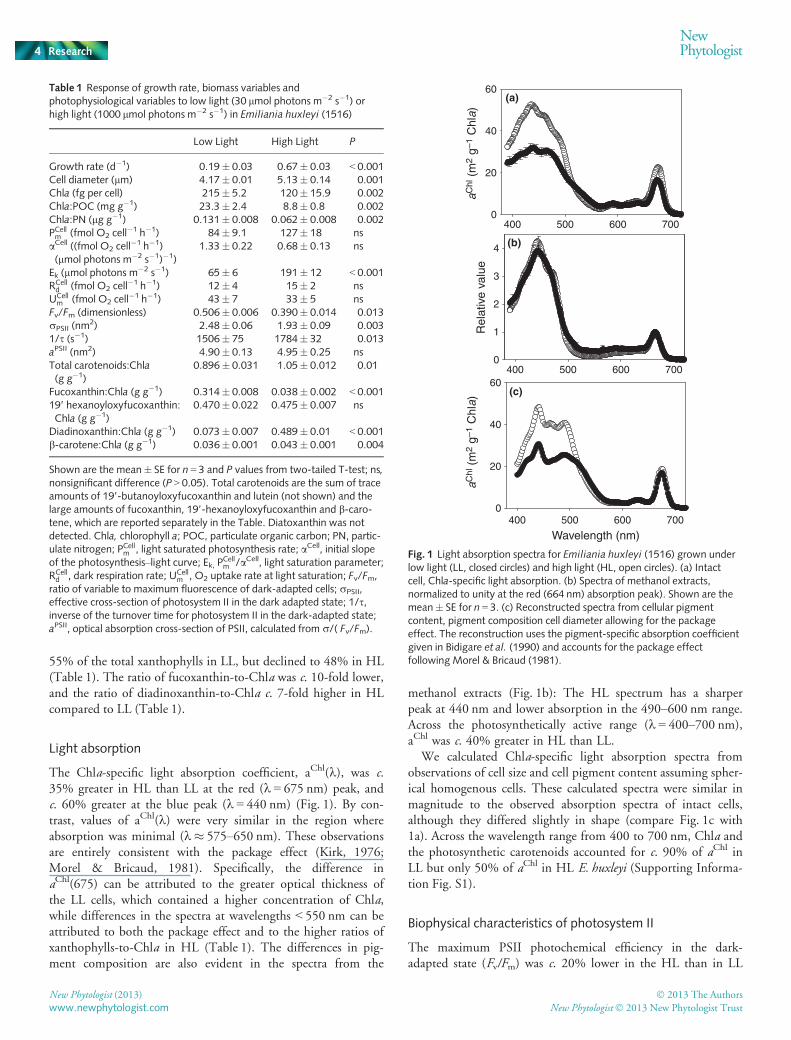
55% of the total xanthophylls in LL, but declined to 48% in HL(Table 1). The ratio of fucoxanthin-to-Chla was c. 10-fold lower,and the ratio of diadinoxanthin-to-Chla c. 7-fold higher in HLcompared to LL (Table 1).
Light absorption
The Chla-specific light absorption coefficient, aChl(k), was c.35% greater in HL than LL at the red (k = 675 nm) peak, andc. 60% greater at the blue peak (k = 440 nm) (Fig. 1). By con-trast, values of aChl(k) were very similar in the region whereabsorption was minimal (k� 575–650 nm). These observationsare entirely consistent with the package effect (Kirk, 1976;Morel & Bricaud, 1981). Specifically, the difference inaChl(675) can be attributed to the greater optical thickness ofthe LL cells, which contained a higher concentration of Chla,while differences in the spectra at wavelengths < 550 nm can beattributed to both the package effect and to the higher ratios ofxanthophylls-to-Chla in HL (Table 1). The differences in pig-ment composition are also evident in the spectra from the
methanol extracts (Fig. 1b): The HL spectrum has a sharperpeak at 440 nm and lower absorption in the 490–600 nm range.Across the photosynthetically active range (k = 400–700 nm),aChl was c. 40% greater in HL than LL.
We calculated Chla-specific light absorption spectra fromobservations of cell size and cell pigment content assuming spher-ical homogenous cells. These calculated spectra were similar inmagnitude to the observed absorption spectra of intact cells,although they differed slightly in shape (compare Fig. 1c with1a). Across the wavelength range from 400 to 700 nm, Chla andthe photosynthetic carotenoids accounted for c. 90% of aChl inLL but only 50% of aChl in HL E. huxleyi (Supporting Informa-tion Fig. S1).
Biophysical characteristics of photosystem II
The maximum PSII photochemical efficiency in the dark-adapted state (Fv/Fm) was c. 20% lower in the HL than in LL
Table 1 Response of growth rate, biomass variables andphotophysiological variables to low light (30 lmol photons m�2 s�1) orhigh light (1000 lmol photons m�2 s�1) in Emiliania huxleyi (1516)
Low Light High Light P
Growth rate (d�1) 0.19� 0.03 0.67� 0.03 < 0.001Cell diameter (lm) 4.17� 0.01 5.13� 0.14 0.001Chla (fg per cell) 215� 5.2 120� 15.9 0.002Chla:POC (mg g�1) 23.3� 2.4 8.8� 0.8 0.002Chla:PN (lg g�1) 0.131� 0.008 0.062� 0.008 0.002PCellm (fmol O2 cell
�1 h�1) 84� 9.1 127� 18 nsaCell ((fmol O2 cell
�1 h�1)(lmol photons m�2 s�1)�1)
1.33� 0.22 0.68� 0.13 ns
Ek (lmol photons m�2 s�1) 65� 6 191� 12 < 0.001RCelld (fmol O2 cell
�1 h�1) 12� 4 15� 2 nsUCell
m (fmol O2 cell�1 h�1) 43� 7 33� 5 ns
Fv/Fm (dimensionless) 0.506� 0.006 0.390� 0.014 0.013rPSII (nm
2) 2.48� 0.06 1.93� 0.09 0.0031/s (s�1) 1506� 75 1784� 32 0.013aPSII (nm2) 4.90� 0.13 4.95� 0.25 nsTotal carotenoids:Chla(g g�1)
0.896� 0.031 1.05� 0.012 0.01
Fucoxanthin:Chla (g g�1) 0.314� 0.008 0.038� 0.002 < 0.00119’ hexanoyloxyfucoxanthin:Chla (g g�1)
0.470� 0.022 0.475� 0.007 ns
Diadinoxanthin:Chla (g g�1) 0.073� 0.007 0.489� 0.01 < 0.001b-carotene:Chla (g g�1) 0.036� 0.001 0.043� 0.001 0.004
Shown are the mean� SE for n = 3 and P values from two-tailed T-test; ns,nonsignificant difference (P > 0.05). Total carotenoids are the sum of traceamounts of 19’-butanoyloxyfucoxanthin and lutein (not shown) and thelarge amounts of fucoxanthin, 19’-hexanoyloxyfucoxanthin and b-caro-tene, which are reported separately in the Table. Diatoxanthin was notdetected. Chla, chlorophyll a; POC, particulate organic carbon; PN, partic-ulate nitrogen; PCellm , light saturated photosynthesis rate; aCell, initial slopeof the photosynthesis–light curve; Ek, P
Cellm /aCell, light saturation parameter;
RCelld , dark respiration rate; UCell
m , O2 uptake rate at light saturation; Fv/Fm,ratio of variable to maximum fluorescence of dark-adapted cells; rPSII,effective cross-section of photosystem II in the dark adapted state; 1/s,inverse of the turnover time for photosystem II in the dark-adapted state;aPSII, optical absorption cross-section of PSII, calculated from r/( Fv/Fm).
0
20
40
60
Rel
ativ
e va
lue
2
3
4
0
1
Wavelength (nm)
400 500 600 700
400 500 600 700
400 500 600 700
aChl
(m
2 g–
1 C
hla)
aC
hl (
m2
g–1
Chl
a)
0
20
40
60
(a)
(b)
(c)
Fig. 1 Light absorption spectra for Emiliania huxleyi (1516) grown underlow light (LL, closed circles) and high light (HL, open circles). (a) Intactcell, Chla-specific light absorption. (b) Spectra of methanol extracts,normalized to unity at the red (664 nm) absorption peak). Shown are themean� SE for n = 3. (c) Reconstructed spectra from cellular pigmentcontent, pigment composition cell diameter allowing for the packageeffect. The reconstruction uses the pigment-specific absorption coefficientgiven in Bidigare et al. (1990) and accounts for the package effectfollowing Morel & Bricaud (1981).
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist4
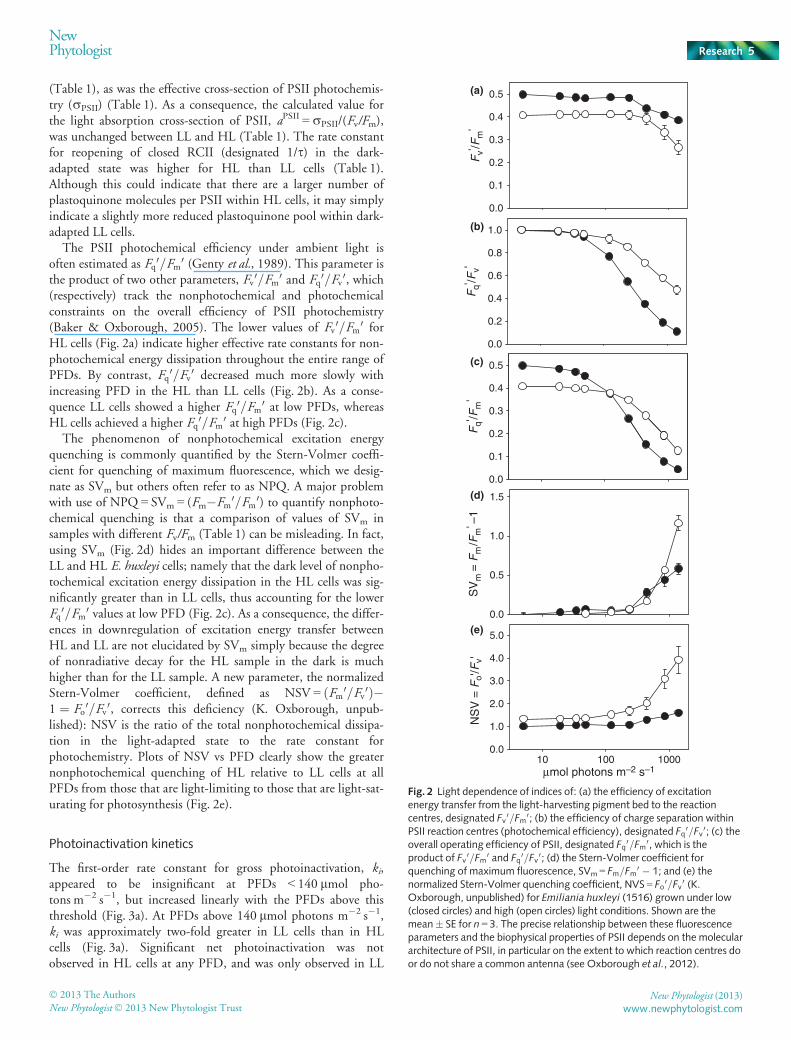
(Table 1), as was the effective cross-section of PSII photochemis-try (rPSII) (Table 1). As a consequence, the calculated value forthe light absorption cross-section of PSII, aPSII = rPSII/(Fv/Fm),was unchanged between LL and HL (Table 1). The rate constantfor reopening of closed RCII (designated 1/s) in the dark-adapted state was higher for HL than LL cells (Table 1).Although this could indicate that there are a larger number ofplastoquinone molecules per PSII within HL cells, it may simplyindicate a slightly more reduced plastoquinone pool within dark-adapted LL cells.
The PSII photochemical efficiency under ambient light isoften estimated as Fq0=Fm0 (Genty et al., 1989). This parameter isthe product of two other parameters, Fv 0=Fm0 and Fq0=Fv 0, which(respectively) track the nonphotochemical and photochemicalconstraints on the overall efficiency of PSII photochemistry(Baker & Oxborough, 2005). The lower values of Fv 0=Fm0 forHL cells (Fig. 2a) indicate higher effective rate constants for non-photochemical energy dissipation throughout the entire range ofPFDs. By contrast, Fq0=Fv 0 decreased much more slowly withincreasing PFD in the HL than LL cells (Fig. 2b). As a conse-quence LL cells showed a higher Fq0=Fm0 at low PFDs, whereasHL cells achieved a higher Fq0=Fm0 at high PFDs (Fig. 2c).
The phenomenon of nonphotochemical excitation energyquenching is commonly quantified by the Stern-Volmer coeffi-cient for quenching of maximum fluorescence, which we desig-nate as SVm but others often refer to as NPQ. A major problemwith use of NPQ = SVm = (Fm�Fm0=Fm0) to quantify nonphoto-chemical quenching is that a comparison of values of SVm insamples with different Fv/Fm (Table 1) can be misleading. In fact,using SVm (Fig. 2d) hides an important difference between theLL and HL E. huxleyi cells; namely that the dark level of nonpho-tochemical excitation energy dissipation in the HL cells was sig-nificantly greater than in LL cells, thus accounting for the lowerFq0=Fm0 values at low PFD (Fig. 2c). As a consequence, the differ-ences in downregulation of excitation energy transfer betweenHL and LL are not elucidated by SVm simply because the degreeof nonradiative decay for the HL sample in the dark is muchhigher than for the LL sample. A new parameter, the normalizedStern-Volmer coefficient, defined as NSV = ðFm0=Fv 0Þ�1 ¼ Fo0=Fv 0, corrects this deficiency (K. Oxborough, unpub-lished): NSV is the ratio of the total nonphotochemical dissipa-tion in the light-adapted state to the rate constant forphotochemistry. Plots of NSV vs PFD clearly show the greaternonphotochemical quenching of HL relative to LL cells at allPFDs from those that are light-limiting to those that are light-sat-urating for photosynthesis (Fig. 2e).
Photoinactivation kinetics
The first-order rate constant for gross photoinactivation, ki,appeared to be insignificant at PFDs < 140 lmol pho-tons m�2 s�1, but increased linearly with the PFDs above thisthreshold (Fig. 3a). At PFDs above 140 lmol photons m�2 s�1,ki was approximately two-fold greater in LL cells than in HLcells (Fig. 3a). Significant net photoinactivation was notobserved in HL cells at any PFD, and was only observed in LL
Fv' /F
m'
Fq' /F
v'F
q' /Fm
'S
Vm
= F
m/F
m' –
1N
SV
= F
o'/F
v'
0.0
0.1
0.2
0.3
0.4
0.5
0.0
0.2
0.4
0.6
0.8
1.0
0.3
0.4
0.5
0.0
0.1
0.2
0.0
0.5
1.0
1.5
µmol photons m–2 s–110 100 1000
0.0
1.0
2.0
3.0
4.0
5.0
(a)
(b)
(c)
(d)
(e)
Fig. 2 Light dependence of indices of: (a) the efficiency of excitationenergy transfer from the light-harvesting pigment bed to the reactioncentres, designated Fv
0=Fm 0; (b) the efficiency of charge separation withinPSII reaction centres (photochemical efficiency), designated Fq
0=Fv 0; (c) theoverall operating efficiency of PSII, designated Fq
0=Fm 0, which is theproduct of Fv 0=Fm 0 and Fq
0=Fv 0; (d) the Stern-Volmer coefficient forquenching of maximum fluorescence, SVm = Fm=Fm
0 � 1; and (e) thenormalized Stern-Volmer quenching coefficient, NVS = Fo
0=Fv 0 (K.Oxborough, unpublished) for Emiliania huxleyi (1516) grown under low(closed circles) and high (open circles) light conditions. Shown are themean� SE for n = 3. The precise relationship between these fluorescenceparameters and the biophysical properties of PSII depends on the moleculararchitecture of PSII, in particular on the extent to which reaction centres door do not share a common antenna (see Oxborough et al., 2012).
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 5

cells when they were exposed to a PFD of 1200 lmol photonsm�2 s�1 (Table S1). Even in this extreme case, the net photoin-activation rate was < 20% of the gross photoinactivation rate(Table S1).
A strict photon-dose response would result in ki being directlyproportional to the product of the absorption cross-section fortarget, the probability that an absorbed photon will inactivate thereaction centre, and the photon flux density:
ki ¼ atargetWE Eqn 3
(ki, rate constant for photoinactivation (s�1); atarget, absorptioncross-section of the putative target; Ψ, probability of a PSII beingphotoinactivated by an incident photon (photon�1); E, photonflux density (mol photons m�2 s�1)). Although we observed alinear relationship between ki and PFD, this applied only above athreshold of c. 140 lmol photons m�2 s�1 (Fig. 3). Conse-quently, our data are not consistent with this model if both atargetand Ψ are assumed to be constant (see Fig. S2a). This conclusionprompted us to examine an alternative model in which the triggerfor inactivation is double reduction of QA, as proposed by Vasset al. (1992). This would require that the observed rate of pho-toinactivation is proportional to the absorption cross-section ofPSII light harvesting at closed RCIIs, corrected for downregula-tion, as is given by Eqn 4:
ki ¼ aPSIIWðFm0=FmÞð1� ðFq0=Fv 0ÞÞE Eqn 4
(aPSII, absorption cross-section of PSII light harvesting; Fm0/Fm,correction for downregulation; (1 � (Fq0=Fv 0)), ratio of the sumof aPSII for light harvesting systems serving only closed RCII tothe sum of aPSII for light harvesting systems serving all RCII (Ox-borough et al., 2012; Notes S1)). When we calculated Ψ fromrearrangement of Eqn 4 we found a systematic decline withincreasing PFD (Fig. S2b), indicating that our observations areinconsistent with this model as well. Finally, we considered a sce-nario where we assumed that the photoinactivation target is inde-pendent of PSII light harvesting, but is only active within a PSIIcomplex with a closed RCII:
ki ¼ atargetWð1� ðFq0=Fv 0ÞÞE Eqn 5
In this scenario, (1 �Fq0=Fv 0) provides an estimate of the frac-tion of RCII in the closed state, and although this estimation isconnectivity-dependent, our fluorescence data are entirely consis-tent with the use of Fq0=Fv 0 in this instance. Our observations arein very good agreement with this model (Figs 3b, S2c), which is,to some extent, a hybrid of Eqns 3 and 4.
Photosynthesis–light response curve
Cellular gross light-saturated photosynthesis (PCellm ) and dark res-piration rates (RCell
d ) were not significantly different in LL andHL cells (Table 1), nor was the rate of oxygen uptake (UCell
m ) at asaturating PFD of 1000 lmol photons m�2 s�1 (Table 1). Bycontrast, the Chla-specific light-saturated rate of gross photosyn-thesis (PChlm ) was three-fold higher in HL, relative to LL (Fig. 4;Table 2), as was the PFD required to saturate gross
µmol photons m–2 s–1
k i (
h–1 )
0
2
4
6
E (1 - Fq'/Fv')
µmol photons m–2 s–1
0 300 600 900 1200
0 300 600 900 1200
k i (
h–1 )
0
2
4
6
(a)
(b)
Fig. 3 (a) Dependence of the rate constant for gross photoinhibition ofFv/Fm (ki) on the PFD for Emiliania huxleyi (1516) grown under low(closed circles) and high (open circles) light. Linear regression of ki on PFDyielded x-intercepts of 136� 18 and 146� 31 lmol photons m�2 s�1
(intercept� SD) for HL and LL, respectively. (b) Dependence of ki on theproduct of PFD and the proportion of RCII that are reduced. The linearrelationship which extrapolates to the origin provides a test of Eqn 5.
Gro
ss O
2 ev
olut
ion
(mol
O2
(g C
hla)
–1 h
–1)
0.5
1.0
0.0
µmol photons m–2 s–1
0 300 600 900 1200
0 300 600 900 1200
O2
upta
ke(m
ol O
2 (g
Chl
a)–1
h–1
)
–0.5
0.0
(a)
(b)
Fig. 4 Typical light responses of (a) gross photosynthetic O2 evolution and(b) O2 consumption for Emiliania huxleyi (1516) grown under low (closedcircles) and high (open circles) light. Summary statistics for cell-specificphotosynthetic parameters and dark respiration rates are given in Table 1,and summary statistics for Chla-specific and organic carbon-specificparameters are given in Table 2.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist6

photosynthesis, EK (Table 2). The Chla-specific initial slopes(aChl) of the photosynthesis–light (PE) response curves were notsignificantly different between LL and HL (Fig. 4; Table 2), butthe maximum quantum efficiency (/m = aChl/aChl) of photosyn-thesis was lower in HL than LL (Table 2).
Impact of light on photosynthetic proteins
Our recent findings (McKew et al., 2013) indicated that the pho-tosynthetic proteins accounted for a greater proportion of thetotal proteome in LL, and in addition that the relative abun-dances of specific photosynthetic proteins also changed signifi-cantly between HL and LL. We reanalysed these data by firstnormalizing to the abundance of PSII core complex proteins(sum of spectral counts for detected proteins PsbA, B, C, D, E,H and V) as an internal photosynthetic unit reference before tak-ing the ratio of these PSII-normalized relative abundancesbetween LL and HL (Fig. 5) to facilitate interpretation of thephotophysiological data. The ratios PSI:PSII and cyt b6/f:PSIIwere identical in LL and HL treatments, and Lhcf(Red):PSII didnot change significantly between LL and HL treatments (Fig 5).Relative to PSII, there were significant decreases in diatom/ha-ptophyte groups I and II Lhcfs in HL, countered by significantincreases in the LI818-like group of fucoxanthin chlorophyllbinding proteins. Similarly Lhcz:PSII was greater in HL althoughthe difference was not significant. The ratios Calvin cycle:PSIIand ATPsynthase:PSII were also significantly lower in HL.
Discussion
We evaluated changes in the photophysiological characteristicsof Emiliana huxleyi within the context of three design consider-ations for the structure–function relationship in chloroplastsidentified by Raven (1980). These considerations are: the
energetic efficiency of photosynthesis; the catalytic efficiency ofphotosynthesis; and the provision of mechanisms to ensure safeoperation of the photosynthetic apparatus. Protein typicallyaccounts for c. 50% of the mass of a microalga under nutrient-replete conditions, and as such the proteome is expected toadjust and respond to selective pressures associated with changesin the environment. The greatest response of the E. huxleyi pro-teome to HL and LL conditions was observed in the antennaeproteins of PSII: these proteins are involved in both light har-vesting and excitation energy dissipation (McKew et al., 2013).Here, we discuss the implications of these changes in the prote-ome for the photophysiology of E. huxleyi, focusing on charac-teristics that map onto Raven’s (1980) three designconsiderations (Table 3).
The Chla-specific initial slope, aChl, of the photosynthesis–light response curve (PE curve) was unaffected by acclima-tion to low or high light
The PE curve (Fig. 4a) has a central role in elucidating andunderstanding different strategies of photoacclimation in micro-algae (Richardson et al., 1983). The initial slope (aChl) of thiscurve quantifies the linear increase of the rate of photosynthesiswith PFD under light-limited conditions (Jassby & Platt, 1976;MacIntyre et al., 2002), and is the product of the light absorption
Lhcf grp I
Lhcf grp II
Lhcz
Lhcf(red)
LI818
LL :
HL
of s
pect
ral c
ount
sno
rmal
ized
to P
SII
0
1
2
3
4
2
**
*
PS I
Calvin Cycle
ATP synthase
Cyt b6/f c
omplexPS II
LL :
HL
of s
pect
ral c
ount
sno
rmal
ized
to P
SII
0
1
* *
(a)
(b)
Fig. 5 (a) Changes in the ratios of Lhcfs and LI818-like proteins to PSIIbetween low light (LL) and high light (HL) Emiliania huxleyi (1516)expressed as the LL-to-HL ratio. (b) Changes in the ratios of thylakoidmembrane complexes and Calvin cycle enzymes to PSII between LL andHL. (Shown are the mean� SE of three technical replicates across threereplicate cultures of each light treatment; n = 3). *, Significant difference atP < 0.05. Calculated from data reported for spectral counts in McKewet al. (2013).
Table 2 Response of photophysiological variables to growth under lowlight (30 lmol photons m�2 s�1) or high light (1000 lmol photons m�2 s�1)in Emiliania huxleyi (1516)
Variable Dimensions Low Light High Light
PChlm mol O2 (g Chla)�1 h�1 0.39� 0.04 1.18� 0.24aChl mol O2 (mol photons)�1
m2 (g Chla)�11.7� 0.3 1.7� 0.4
aChl m2 (g Chla)�1 14.6� 1.2 20.3� 0.5/m mol O2 (mol photons)�1 0.12� 0.02 0.08� 0.02PCm mol O2 (mol C)�1 h�1 0.11� 0.02 0.11� 0.02aC mol O2 (mol photons)�1
m2 (g C)�10.14� 0.03 0.050� 0.011
aC m2 (mg C)�1 0.34� 0.05 0.18� 0.02
Chla specific, carbon-specific photosynthesis rates were calculated fromcell-specific data (Table 1) with error propagated assuming uncorrelatederrors.PChlm , the Chla-specific light saturated gross oxygen evolution rate; aChl, theinitial slope of the PChl vs light curve; aChl, the Chla-specific light absorptioncoefficient; /m, the maximum quantum efficiency of gross oxygen evolu-tion; PCm, light saturated gross oxygen evolution rate; aC, the initial slope ofthe PC vs light curve; aC, the organic C-specific light absorption coefficient.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 7

coefficient (aChl) and the maximum quantum efficiency of photo-synthesis (/m):
aChl ¼ aChl/m Eqn 6
In the context of Raven’s (1980) design considerations, aChl isa catalytic efficiency, which when multiplied by the PFD givesthe rate of photon absorption, whereas /m is an energetic effi-ciency that relates the output of O2 to the input of photons. Wecalculated that /m was 25% lower in HL than LL from our mea-surements of aChl and aChl. This decrease is consistent with the22% lower value of Fv/Fm for the HL cells (Table 1), which alongwith the lower values of Fv 0=Fm0 in the light (Fig. 2) can beascribed to the increase in the contribution of the photoprotectivepigments to light absorption in the blue region of the spectrum(Fig. S1).
The independence of aChl from growth PFD in E. huxleyiresults from compensatory changes between the catalytic effi-ciency of light absorption and the energetic efficiency of excita-tion energy transfer from the light harvesting pigments to thePSII reaction centres. Similar reciprocal changes of aChl and /m
in response to growth at different PFDs have been observed inother microalgae (Geider et al., 1998; Anning et al., 2000). Thisinvariance of aChl within a taxon facilitates the formulation ofsimple models of algal growth and acclimation (e.g. Geider et al.,1998); however, it is not immediately obvious why this compen-sation occurs. To begin to address this issue, we now turn to thebio-optical and biophysical mechanisms that underpin changesof cellular pigment content, aChl and /m.
Acclimation of E. huxleyi to low light increased both thesize and number of photosynthetic units
Changes in the protein composition of the thylakoid membranesthat accompany increased cellular pigment content in low lightcan involve: an increase in the number of photosynthetic units ofconstant ‘size’; or an increase in the sizes of a constant numberphotosynthetic units; or some combination of the two (Falkowski& La Roche, 1991). The number of RCII per cell is typically
quantified by measuring the abundances of PSII and PSIIreaction centre proteins using either biophysical techniques(Falkowski et al., 1981) or Western blotting (Six et al., 2008),whereas photosynthetic unit size is typically quantified by theratio Chla:RCII. In E. huxleyi the proportion of the proteomeattributable to PSII was greater in LL than HL (McKew et al.,2013), and the ratio Lhcf:PSII was also significantly greater in LLcompared to HL (Fig. 5), consistent with previous observationsthat photoacclimation in this species involves changes of bothphotosynthetic unit size and number (Suggett et al., 2007).
The higher photosynthetic unit number and photosyntheticunit size in LL E. huxleyi accounted for c. 55% of the 2.8-foldgreater C-specific initial slope of the PE curves in LL treatments,with the remaining 45% of the increase attributable to the greater/m in LL than HL. Thus, by increasing pigment content underLL E. huxleyi achieved a higher growth rate under LL than wouldhave been possible in the absence of photoacclimation, therebycontributing to increased fitness.
Acclimation of E. huxleyi to low light increased the effectivecross-section of PSII photochemistry (rPSII) but not theabsorption cross-section of PSII light harvesting (aPSII)
Our proteomic and biophysical data yielded markedly differentconclusions regarding the response of PSU ‘size’ to the light treat-ments. The 2.3-fold greater Lhcf:PSII in LL than HL (Fig. 5)indicates that the number of pigment molecules, and mass ofprotein, associated with each PSII was markedly greater in LLthan HL. By contrast, there was no significant change in theabsorption cross-section of PSII light harvesting between LL andHL (aPSII = rPSII/[Fv/Fm] = 4.9 in both LL and HL) because the1.3-fold higher rPSII under LL was offset by an equivalentincrease in Fv/Fm (Table 1).
To summarize, despite the markedly higher resource allocationto Lhcfs in LL cells, the average light absorption cross-section ofeach individual PSII light-harvesting system was the same forboth LL and HL cells. The likely explanation for this result is thatthe extent of intracellular self-shading due to the pigment packag-ing effect, which quantifies the decrease in the efficiency of light
Table 3 Summary of responses of differences in photophysiological characteristics of Emiliania huxleyi (1516) between low light (LL) and high light (HL)treatments categorized according to Raven’s (1980) design considerations for structure–function relationships in eukaryotic chloroplasts
Design consideration Photophysiological characteristic assessed SymbolTreatmenteffect
Change in chloroplastproteins
Energetic efficiency of the processdefined as the ratio of useful workoutput to energy input
Maximum quantum efficiency of photosynthesis /m LL >HL Increase of LI818:Lhcf in HLreduces energetic efficiencyMaximum efficiency of PSII in the dark-adapted state Fv/Fm LL >HL
Catalytic efficiency of the processdefined as work output per unitcatalytic and structural materialused
Chla-specific light absorption coefficient aChl LL <HL Increase of Lhcf:RCII in LLincreases rate of lightabsorption
Organic carbon-specific light absorption coefficient aC LL >HLLight harvesting (light absorption) cross section of PSII aPSII LL =HLOrganic carbon-specific light-saturated photosynthesis rate PCm LL =HL
Provision of mechanisms to ensuresafe operation of the photosyntheticapparatus
Rate constant for photoinactivation of RCII at a given PFD ki LL >HL Increase of LI818:RCII in HLmaintains more RCII in theopen state
Susceptibility of reduced RCII to photoinactivation atarget Ψ LL =HLProportion of PSII reaction centres in the ‘open’ state athigh PFD
Fq0=Fv0 LL <HL
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist8

absorption by pigments when present at extremely high concen-trations in cells compared with the same mass of pigment whenmore diffusely distributed in solution (Morel & Bricaud, 1981),was much greater in highly pigmented and slightly smaller LLcells than in less pigmented, larger HL cells. Thus, aPSII wasunchanged between LL and HL because the decline in aChl wasoffset by the increase in Lhcf:PSII. Our data indicate that tomaintain the same PSII light absorption cross-section (when PSIIabundance was 1.7-fold greater) in LL relative to HL required,on average, that each PSII of a LL E. huxleyi cell contained abouta 2.3-fold greater amount of Lhcfs. In conclusion, there was amarked decline in the marginal rate of increase in light intercep-tion per unit investment of the proteome into Lhcfs and PSII ofLL E. huxleyi due to the package effect, with each pigment mole-cule only c. 50% as effective in LL as in HL cells.
Our observations differ from those reported for Ostreococcuswhere changes in rPSII were correlated with changes in Lhcp:PSII(Six et al., 2008). Cells of Ostreococcus are much smaller thanthose of E. huxleyi, and the package effect plays a less significantrole in such small cells (Morel & Bricaud, 1981). Thus, in largerorganisms like E. huxleyi, it is likely that the diminishing returnon investment in Lhcfs is greater than in picoplankton likeOstreococcus species. This has implications for integrating opti-mality considerations (Smith et al., 2011) into trait-based modelsof phytoplankton ecophysiology (Litchman & Klausmeier,2008).
Different costs and benefits are associated with acclimationof PSU number and size
In their study of photoacclimation of two Ostreococcus ecotypes,Six et al. (2008) contrasted n-type photoacclimation, whichinvolved changes in PSU number per cell, with r-type photoacc-limation, which involved changes in the effective cross-section ofPSII photochemistry (rPSII). Six et al. (2008) suggested that thephotoacclimation strategy employed by a microalga can havesignificant implications for the capital costs of investment inthylakoid proteins, as well as the operating and/or opportunitycosts associated with the D1 damage and repair cycle. Increasingthe number of photosynthetic units in response to low light ismore nitrogen-expensive than adding peripheral light harvestingcomplexes to the PSII and PSI cores (Dubinsky et al., 1986;Moore et al., 2006; Six et al., 2008). Similarly, in E. huxleyiinvestment in Lhcfs with a ratio of c. 0.37 mol Chla (kgprotein)�1 will be more efficient in light absorption per unitnitrogen than investment in core PSII complex which have a ratioof c. 0.10 mol Chla (g protein)�1 (see Table S2). Exacerbatingthe difference in Chla:protein between antennae Lhcfs and thePSII core is the fact that much of light absorption by E. huxleyi isdue to xanthophylls (Fig. S1), which are bound to Lhcfs.
There are, however, potential costs of increasing the ratio ofLhcfs to PSII in a r-type strategy. For example, increasing aPSII
may increase the susceptibility to photoinhibition by increasingthe rate of excitation delivery to the reaction centres (Mooreet al., 2006; Six et al., 2008). Although this may be the case inOstreococcus (Six et al., 2008), aPSII was not significantly
different between LL and HL E. huxleyi. Six et al. (2008) alsosuggested that a large antenna may impede the rate of RCIIrepair following photoinactivation, which would have conse-quences for intermittent exposure to supraoptimal light oftenexperienced in nature. By contrast, a large cellular pool ofRCIIs with smaller light harvesting absorption cross-sectionsmay provide a buffer against the accumulation of inactive PSIIunits (Behrenfeld et al., 1998). Taking all of these consider-ations into account, Six et al. (2008) suggested that r-typephotoacclimation is expected in organisms evolutionallyadapted to stable low-light environments with n-type strategyexpected in organisms adapted to environment characterized byhigh variability in light.
The carbon-specific light-saturated photosynthesis rate,PCm, was unaffected by acclimation to low or high light
Although the Chla-specific light-saturated photosynthesis rate(PChlm ) was about three-fold greater in HL than LL E. huxleyi(Fig. 4a), the organic C-specific light-saturated photosynthesisrate (PCm) did not differ between LL and HL cells (Table 2). Thelimited variability of PCm in response to growth at different PFDsis also observed in other microalgae (MacIntyre et al., 2002), andis consistent with observations that the abundance of the Calvincycle enzyme Rubisco is largely unaffected by growth at differentPFDs in a number of species (Sukenik et al., 1987; Fisher et al.,1989; Six et al., 2008; Harris et al., 2009; Lefebvre et al., 2010).In fact, our own data suggest that Rubisco is slightly more abun-dant in LL than in HL acclimated cells of E. huxleyi 1516(McKew et al., 2013).
The invariance of PCm in E. huxleyi was accompanied by a two-fold reduction in the cell-specific initial slope of the PE curve(Table 1), and a three-fold reduction in the carbon-specific initialslope, in HL relative to LL (Table 2). Thus, photoacclimation toHL does not appear to benefit E. huxleyi, or most other microal-gae that have been examined (Anning et al., 2000; MacIntyreet al., 2002), by increasing the biomass-specific photosynthesisrate at the growth PFD. Thus, we must seek an alternative expla-nation to account for the benefit of acclimation to HL inE. huxleyi. This benefit is likely to be a reduction in the suscepti-bility of RCII to photoinactivation, although it may also includemechanisms that reduce the production of ROS and associatedoxidative stress.
Blooms of E. huxleyi tend to occur in water columns whereincident solar radiation exceeds a mean of c. 500 lmol photonsm�2 s�1 over the day (Nanninga & Tyrrell, 1996). Thus, condi-tions under which E. huxleyi or other microalgae would experi-ence prolonged exposure to PFDs as high as 1000 lmol photonsm�2 s�1 will be limited to water columns where the surface layerbecomes diurnally stratified under clear skies. Under these condi-tions, PFD can exceed 1000 lmol photons m�2 s�1 for severalhours at a time. However, nocturnal destratification will redis-tribute cells vertically, making it unlikely that the same cells (ortheir daughter cells) will experience such high PFDs for severaldays running. Thus, there would appear to be little selectionpressure in nature to develop an acclimation strategy to
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 9

prolonged exposure to such high PFDs. Instead, the acclimationstrategy is likely to involve development of tolerance to episodicexposure to high PFDs whilst maintaining the capacity to photo-synthesize efficiently in low light.
Acclimation of E. huxleyi to high light reduced thesusceptibility of RCII to photoinactivation by increasingenergy dissipation in the antenna complexes
It is inevitable that some excitation energy is dissipated by uncon-trolled reactions which generate free radicals, which in turn cancause photo-oxidative damage (Salin, 1987). The D1 reactioncentre protein of PSII is one of most rapidly turned over proteinsin the photosynthetic electron transfer chain because it is particu-larly susceptible to photo-oxidative damage (Raven, 2011).When the rate of D1 photoinactivation exceeds the rate of repair,photoinhibition occurs. We observed that the rate of D1 damagewas about two-fold greater in E. huxleyi growing under LL thanin E. huxleyi grown in HL (Fig. 3).
As discussed by Anderson et al. (1998) and Oxborough &Baker (2000), one mechanism to reduce the susceptibility to pho-toinactivation during prolonged exposure to supra-optimal lightis to increase the capacity for nonphotochemical quenching, aprocess that is referred to as downregulation (Krause & Behrend,1986; Oxborough & Horton, 1988; Li et al., 2002). Downregu-lation operates on timescales of seconds to minutes, to adjust thelevel of nonphotochemical de-excitation in response to changesin both PFD and the requirement for NADPH and ATP. Thisprocess increases the effective rate constant for nonradiativedecay, which competes directly with both PSII photochemistryand Chla fluorescence for excitons within the PSII pigmentmatrix. The result is an increase in the level of nonphotochemicalde-excitation, which shortens exciton lifetime within the PSIIpigment matrix. Invariably, down regulation results in a nonpho-tochemical quenching of the fluorescence signal (Schreiber et al.,1986), which decreases maximum fluorescence (Fm in the dark-adapted state, Fm0 in the light-adapted state). Despite the fact thatdownregulation competes directly with PSII photochemistry, theprocess may have minimal impact on the rate of PSII photo-chemistry, particularly under high light conditions where the ratebecomes limited on the reducing side of PSII.
In E. huxleyi, as in other microalgae, different Chla bindingproteins contribute to light-harvesting and nonphotochemicalquenching. Fucoxanthin-Chl binding proteins in the groups Iand II Lhcf clades and the Lhcf(red) clade contribute to light-harvesting and efficient excitation energy transfer to the reactioncentres, and the abundances of these complexes were up-regu-lated in LL (Fig. 5a). By contrast, LI818 and LI818-like genes ofvarious microalgal species are upregulated under high light(Lefebvre et al., 2010; Park et al., 2010; Zhu & Green, 2010).Consistent with the photoprotective role of downregulation, thecontribution of LI818 proteins to spectral counts of all of thefucoxanthin binding proteins increased from c. 3% in LL to 25%in HL (see Table S2 in McKew et al., 2013). This difference wasassociated with an increase in diadinoxanthin:Chla (Table 1),decrease in Fv/Fm (Table 1), decrease of Fv 0=Fm0 (Fig. 2a), and a
decrease in the rate constant for gross photoinactivation (Fig. 3a).Thus, in E. huxleyi, as previously shown for Chlamydomonas(Peers et al., 2009), there is a direct functional correlationbetween accumulation of LI818 proteins and photoprotection.Our data also suggest a photoprotective role for the Lhcz-likeproteins, which were more abundant (relative to PSII) in HLthan LL (Fig. 5), although the difference was not as pronouncedas for the LI818-like proteins.
Significantly, our data indicate that photoinactivation of Fv/Fmin E. huxleyi can be explained by a process in which some compo-nent of a closed RCII is the target (Fig. 3c). Thus, increased pro-tection of RCII from photoinactivation in HL cells appears to beconferred by higher capacity of these cells to downregulate excita-tion energy transfer from the light-harvesting antenna to RCII,thus maintaining a greater proportion of RCII in the open state.
Conclusion
Differences in photophysiology of E. huxleyi between low-lightand high-light treatments were associated primarily with changesin the abundances of different groups of fucoxanthin-Chl bind-ing proteins. The acclimation of photophysiology reported in thispaper supports our conclusion based on whole-cell proteomicdata (McKew et al., 2013) that the benefit to E. huxleyi of accli-mation to light energy-limited conditions is an increase in therate of gross photosynthesis at the growth irradiance, whereas thebenefit of acclimation to supra-optimal, light-saturating condi-tions is a reduction in the rate of photoinactivation, and possiblyother sources of photodamage, at the growth PFD. Significantly,the decreased carbon-specific initial slope of the PE curve thataccompanied reduced pigment:biomass in E. huxleyi acclimatedto high light was not accompanied by an increase in the carbon-specific light-saturated photosynthesis rate.
Photoacclimation was most evident in the protein and pig-ment composition of the antenna of PSII and in the ratio ofperipheral antenna to the PSII core. These changes reflect differ-ences in resource allocation between the light-harvesting andphotoprotective functions within the PSII antenna. The func-tional consequences of these changes in the molecular structureof PSII included reducing the susceptibility of RCII to photoin-activation in cells acclimated to high-light and increasing thedelivery of excitation energy to RCII in cells acclimated to lowlight. The specific changes are as follows: an increase of LI818-like proteins and xanthophyll cycle pigments in HL reduced theenergetic efficiency of photosynthesis; an increase of Lhcf pro-teins and photosynthetic pigments in LL increased light harvest-ing and the light-limited rate of photosynthesis; an increase inthe ratio LI818:Lhcf in HL reduced susceptibility of RCII tophotoinactivation by maintaining a greater proportion of RCIIin the open state (Table 3).
Our results support Raven’s (1980) analysis that structure–function relationships in chloroplasts of eukaryotic cells areconstrained by trade-offs to maximize the energetic and catalyticefficiencies of photosynthesis, subject to the provision of mecha-nisms to ensure safety from photooxidative stress. Quantifyingthe cost–benefit trade-offs associated with different acclimation
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist10

and adaptation strategies is still in its infancy. This is despite theneed for better mechanistic descriptions of algal growth based onoptimal allocation principles (Smith et al., 2011) for inclusion inlarger scale models of open ocean ecology and biogeochemistry(Follows & Dutkiewicz, 2011). Our results confirm thatRaven’s (1980) design considerations of energetic efficiency, cata-lytic efficiency and safety provide a framework that can be usedto inform optimality-based models of chloroplast function andalgal growth.
Acknowledgements
This research was supported by NERC grant NE/G003688/1awarded to R.J.G., C.A.R. and M.V.M. J.H. and S.J.F. were sup-ported by NERC studentships.
References
Anderson JM, Park Y-I, ChowWS. 1998. Unifying model for the
photoinactivation of photosystem II in vivo under steady-state conditions.Photosynthesis Research 56: 1–13.
Anning T, Pratt SM, Sammes PJ, MacIntyre HL, Gibb S, Geider RJ. 2000.
Photoacclimation in the marine diatom Skeletonema costatum. Limnology andOceanography 45: 1807–1817.
Baker NR, Oxborough K. 2005. Chlorophyll fluorescence as a probe of
photosynthetic productivity. In: Papageorgiou GC, Govindjee, eds. Chlorophylla fluorescence – a signature of photosynthesis. Dordrecht, the Netherlands:
Springer, 65–82.Barlow RG, Cummings DG, Gibb SW. 1997a. Improved resolution of mono-
and divinyl chlorophylls a and b and zeaxanthln and lutein in phytoplankton
extracts using reverse phase C-8 HPLC.Marine Ecology Progress Series 161:303–307.
Barlow RG, Mantoura RFC, Cummings DG, Fileman TW. 1997b. Pigment
chemotaxonomic distributions of phytoplankton during summer in the western
Mediterranean. Deep-Sea Research II 44: 833–850.BehrenfeldMJ, Falkowski PG. 1997. Photosynthetic rates derived from satellite-
based chlorophyll concentration. Limnology and Oceanography 42: 1–20.Behrenfeld MJ, Pr€asil O, Kolber ZS, Babin M, Falkowski PG. 1998.
Compensatory changes in photosystem II electron turnover rates protect
photosynthesis from photoinhibition. Photosynthesis Research 58: 259–268.Bidigare RR, Ondrusek ME, Morrow JH, Kiefer DA. 1990. In vivo absorptionproperties of algal pigments. Proceedings of the Society of Photo-opticalInstrumentation Engineers 1302: 290–302.
Bjorkman O 1981. Responses to different quantum flux densities. In: Lange OL,
Nobel PS, Osmond CB, Zeiger H, eds. Encyclopaedia of plant physiology, Newseries, Vol 12A. Berlin, Germany: Springer, 57–107.
Blankenship RE. 2002Molecular mechanisms of photosynthesis. Oxford, UK:
Blackwell Science Ltd.
Brading P, Warner ME, Davey P, Smith DJ, Achterberg EP, Suggett DJ. 2011.
Differential effects of ocean acidification on growth and photosynthesis among
phylotypes of Symbiodinium (Dinophyceae). Limnology and Oceanography 56:927–938.
Dubinsky Z, Falkowski PG, Wyman K. 1986. Light harvesting and utilization by
phytoplankton. Plant Cell Physiology 27: 1335–1349.Escoubas J-M, Lomas M, La Roche J, Falkowski PG. 1995. Light intensity
regulation of cab gene transcription is signalled by the redox state of the
plastoquinone pool. Proceedings of the National Academy of Sciences, USA 92:
10 237–10 241.Falkowski PG, La Roche J. 1991. Acclimation to spectral irradiance in algae.
Journal of Phycology 27: 8–14.Falkowski PG, Owens TG, Ley AC, Mauzerall DC. 1981. Effects of growth
irradiance levels on the ratio of reaction centers in two species of marine
phytoplankton. Plant Physiology 69: 969–973.
Falkowski PG, Raven JA. 2007. Aquatic photosynthesis, 2nd edn. Princeton, NJ,
USA: Princeton University Press.
Fisher T, Shurtz-Swirski R, Gepstein S, Dubinsky Z. 1989. Changes in levels of
ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) in Tetraedronminimum (Chlorophyta) during light and shade adaptation. Plant and CellPhysiology 30: 221–228.
Follows MJ, Dutkiewicz S. 2011.Modeling diverse communities of marine
microbes. Annual Review of Marine Science 3: 427–451.Geider RJ, DeLucia EH, Falkowski PG, Finze A, Grime JP, Grace J, Kana TM,
LaRoche J, Long SP, Osborne BA, et al. 2001. Forum. Primary productivity
of planet earth: biological determinants and physical constraints in terrestrial
and aquatic habitats. Global Change Biology 7: 849–882.Geider RJ, MacIntyre HL, Kana TM. 1998. A dynamic regulatory model of
phytoplankton acclimation to light, nutrients and temperature. Limnology andOceanography 43: 679–694.
Geider RJ, Moore CM, Ross ON. 2009. The role of cost–benefit analysis inmodels of phytoplankton growth and acclimation. Plant Ecology and Diversity2: 165–178.
Genty B, Briantais J-M, Baker NR. 1989. The relationship between the quantum
yield of photosynthetic electron transport and quenching of chlorophyll
fluorescence. Biochimica et Biophysica Acta 990: 87–92.Givnish TJ. 1988. Adaptation to sun and shade: a whole plant perspective.
Australian Journal of Plant Physiology 15: 63–92.Harris GN, Scanlan D, Geider RJ. 2009. Responses of Emiliania huxleyi(Prymnesiophyceae) to step changes in photon flux density. European Journal ofPhycology 44: 31–48.
Iglesias-Rodriguez MD, Brown CW, Doney SC, Kleypas JA, Kolber D, Kolber
Z, Hayes PK, Falkowski PG. 2002. Representing key phytoplankton
functional groups in ocean carbon cycle models: Coccolithophorids. GlobalBiogeochemical Cycles 16: 47-1–47-20.
Jassby AD, Platt T. 1976.Mathematical formulation of the relationship between
photosynthesis and light for phytoplankton. Limnology and Oceanography 21:540–547.
Kana T. 1990. Light-dependent oxygen cycling measured by an oxygen-18
isotope dilution technique.Marine Ecology Progress Series 64: 293–300.Kirk JTO. 1976. A theoretical analysis of the contribution of algal cells to the
attenuation of light within natural waters. III. Cylindrical and spheroidal cells.
New Phytologist 77: 341–358.Knapp AK, Carter GA. 1998. Variability in leaf optical properties among 26
species from a broad range of habitats. American Journal of Botany 85: 940–946.
Kolber ZS, Pr�asil O, Falkowski PG. 1998.Measurements of variable chlorophyll
fluorescence using fast repetition rate techniques: defining methodology and
experimental protocols. Biochimica et Biophysica Acta 1367: 88–106.Krause GH, Behrend U. 1986. pH-dependent chlorophyll fluorescence
quenching indicating a mechanism of protection against photoinhibition of
chloroplasts. FEBS Letters 200: 298–302.Lacour T, Sciandra A, Talec A, Mayzaud P. 2012. Diel variations of
carbohydrates and neutral lipids in nitrogen-sufficient and nitrogen-starved
cyclostat cultures of Isochrysis sp. Journal of Phycology 48: 966–975.Lefebvre SC, Harris G, Webster R, Leonardos N, Geider RJ, Raines CA, Read
BA, Garrido JL. 2010. Characterization and expression analysis of the LHCF
gene family in Emiliania huxleyi (Haptophyta) reveals differential responses to
light and CO2. Journal of Phycology 46: 123–134.Li XP, Muller-Moule P, Gilmore AM, Niyogi KK. 2002. PsbS-dependent
enhancement of feedback de-excitation protects photosystem II from
photoinhibition. Proceedings of the National Academy of Sciences, USA 99:
15 222–15 227.Litchman E, Klausmeier CA. 2008. Trait-based community ecology of
phytoplankton. Annual Reviews of Ecology and Systematics 39: 615–639.MacIntyre HL, Anning T, Kana TM, Geider RJ. 2002. Photoacclimation of
photosynthesis-irradiance response curves and photosynthetic pigments in
microalgae and cyanobacteria. Journal of Phycology 38: 17–38.McKew B, Lefebvre SC, Achterberg EP, Metodieva G, Raines CA, Metodiev
MV, Geider RJ. 2013. Plasticity in the proteome of Emiliania huxleyi toextremes of light is highly targeted. New Phytologist. doi: 10.1111/nph.12352.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 11

Moore CM, Suggett DJ, Hickman AE, Kim Y-N, Tweddle JF, Sharples J,
Geider RJ, Holligan PM. 2006. Phytoplankton photoacclimation and
photoadaptation in response to environmental gradients in a shelf sea.
Limnology and Oceanography 51: 936–949.Moore LR, Chisholm SW. 1999. Photophysiology of the marine cyanobacterium
Prochlorococcus: ecotypic differences among cultured isolates. Limnology andOceanography 44: 628–638.
Morel A, Bricaud A. 1981. Theoretical results concerning light absorption in a
discrete medium, and application to specific absorption of phytoplankton.
Deep-Sea Research 28A: 1375–1393.Nanninga HJ, Tyrrell T. 1996. Importance of light for the formation of algal
blooms by Emiliania huxleyi.Marine Ecology Progress Series 136: 195–203.Oxborough K, Baker NR. 2000. An evaluation of the potential triggers of
photoinactivation of photosystem II in the context of a Stern-Volmer model for
down-regulation and the reversible radical pair equilibrium model.
Philosophical Transactions of the Royal Society B 355: 1489–1498.Oxborough K, Horton P. 1988. A study of the regulation and function of
energy-dependent quenching in pea chloroplasts. Biochimica et Biophysica Acta934: 135–143.
Oxborough K, Moore CM, Suggett DJ, Lawson T, Chan HG, Geider RJ. 2012.
Direct estimation of functional PSII reaction centre concentration and PSII
electron flux on a volume basis: a new approach to the analysis of Fast
Repetition Rate fluorometry (FRRf) data. Limnology and Oceanography Methods10: 142–154.
Pandhal J, Wright PC, Biggs CA. 2007. A quantitative proteomic analysis of
light adaptation in a globally significant marine cyanobacterium ProchlorococcusmarinusMED4. Journal of Proteome Research 6: 996–1005.
Park S, Jung G, Hwang YS, Jin E. 2010. Dynamic response of the transcriptome
of a psychrophilic diatom, Chaetoceros neogracile, to high irradiance. Planta231: 349–360.
Peers G, Truong TB, Ostendorf E, Busch A, Elrad D, Grossman AR, Hippler
M, Niyogi KK. 2009. An ancient light-harvesting protein is critical for the
regulation of algal photosynthesis. Nature 462: 518–521.Pfannschmidt T, Br€autigam K, Wagner R, Dietzel L, Schr€oter Y, Steiner S,
Nykytenko A. 2009. Potential regulation of gene expression in photosynthetic
cells by redox and energy state: approaches towards better understanding.
Annals of Botany 103: 599–607.Ragni M, Airs R, Leonardos N, Geider R. 2008. Photoinhibition of PSII in
Emiliania huxleyi (Haptophyta) under high light stress: the roles of
photoacclimation, photoprotection, and photorepair. Journal of Phycology 44:670–683.
Raven JA. 1980. Chloroplasts of eukaryotic micro-organisms. In: Gooday GW,
Lloyd D, Trinci APJ, eds. The eukaryotic microbial cell – 13th Symposium of theSociety for General Microbiology. Cambridge, UK: Cambridge University Press,
181–205.Raven JA. 2011. The cost of photoinhibition. Physiologia Plantarum 142: 87–104.
Raven JA, Geider RJ. 2003. Adaptation, acclimation and regulation in algal
photosynthesis. In: Larkum AWD, Douglas S, Raven JA, eds. Photosynthesis ofalgae. Dordrecht, the Netherlands: Kluwer Academic, 385–412.
Read BA, Kegel J, Klute MJ, Kuo A, Lefebvre SC, Maumus F, Mayer C, Miller
J, Monier A, Salamov A et al. 2013. Emiliania’s pan genome drives the
phytoplankton’s global distribution. Nature doi: 10.1038/nature12221.Richardson K, Beardall J, Raven JA. 1983. Adaptation of unicellular algae to
irradiance: an analysis of strategies. New Phytologist 93: 157–191.Salin ML. 1987. Toxic oxygen species and protective systems of the chloroplast.
Physiologia Plantarum 72: 681–689.Schellenberger Costa B, Jungandreas A, Jakob T, Weisheit W, Mittag M,
Wilhelm C. 2013. Blue light is essential for high light acclimation and
photoprotection in the diatom Phaeodactylum tricornutum. Journal ofExperimental Botany 64: 483–493.
Schreiber U, Schliwa U, Bilger W. 1986. Continuous recording of
photochemical and non-photochemical chlorophyll fluorescence
quenching with a new type of modulation fluorometer. Photosynthesis Research10: 51–62.
Seemann JR, Sharkey TD, Wang JL, Osmond BC. 1987. Environmental effects
on photosynthesis, nitrogen-use efficiency, and metabolite pools in leaves of
sun and shade plants. Plant Physiology 84: 796–802.Six C, Finkel Z, Rodriguez F, Marie D, Partensky F, Campbell DA. 2008.
Contrasting photoacclimatation costs in ecotypes of the marine picoplankter
Ostreococcus. Limnology and Oceanography 53: 255–265.Smith SL, Pahlow M, Merico A, Wirtz KW. 2011.Optimality-based modelling
of planktonic organisms. Limnology and Oceanography 56: 2080–2094.Suggett DJ, Le Floc’H E, Harris G, Leonardos N, Geider RJ. 2007. Different
strategies of photoacclimation by two strains of Emiliania huxleyi(Haptophyta). Journal of Phycology 43: 1209–1222.
Suggett DJ, MacIntyre HL, Geider RJ. 2004. Evaluation of biophysical and
optical determinations of light absorption by photosystem II in phytoplankton.
Limnology and Oceanography Methods 2: 316–332.Sukenik A, Bennett J, Falkowski P. 1987. Light-saturated photosynthesis –limitation by electron transport or carbon fixation? Biochimica et BiophysicaActa 891: 205–215.
Vass I, Styring S, Hundal T, Koivuniemi A, Aro EM, Andersson B. 1992.
Reversible and irreversible intermediates during photoinhibition of
photosystem II. 2. Stable reduced QA species promote chlorophyll triplet
formation. Proceedings of the National Academy of Sciences, USA 89:
1408–1412.Wilhelm C, Buchel C, Fisahn J, Goss R, Jakob T, La Roche J, Lavaud J, Lohr
M, Riebesell U, Stehfest K et al. 2006. The regulation of carbon and nutrient
assimilation in diatoms is significantly different from green algae. Protist 157:91–124.
Zhu SH, Green BR. 2010. Photoprotection in the diatom Thalassiosirapseudonana: role of LI818-like proteins in response to high light stress.
Biochimica et Biophysica Acta 1797: 1449–1457.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1Contributions of Chla and accessory pigments to lightabsorption.
Fig. S2 PFD dependence of the probability of photoinactivationcalculated from Eqns 3–5.
Table S1 Rate constants for gross and net photoinactivation
Table S2Nitrogen and carbon contents of Lhcfs and PSII
Notes S1Derivation of Eqn 4 in the main paper.
Please note: Wiley-Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist12
Related Documents











