of September 10, 2015. This information is current as Lysosome-Associated Membrane Protein-1 M150, Is a Posttranslational Isoform of The Th1-Specific Costimulatory Molecule, Anil Kumar and Gyan C. Mishra Pinaki P. Banerjee, Pradeep B. Parab, Samit Chattopadhyay, Durbaka V. R. Prasad, Vrajesh V. Parekh, Bimba N. Joshi, http://www.jimmunol.org/content/169/4/1801 doi: 10.4049/jimmunol.169.4.1801 2002; 169:1801-1809; ; J Immunol References http://www.jimmunol.org/content/169/4/1801.full#ref-list-1 , 14 of which you can access for free at: cites 40 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2002 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on September 10, 2015 http://www.jimmunol.org/ Downloaded from by guest on September 10, 2015 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of September 10, 2015.This information is current as
Lysosome-Associated Membrane Protein-1M150, Is a Posttranslational Isoform of The Th1-Specific Costimulatory Molecule,
Anil Kumar and Gyan C. MishraPinaki P. Banerjee, Pradeep B. Parab, Samit Chattopadhyay, Durbaka V. R. Prasad, Vrajesh V. Parekh, Bimba N. Joshi,
http://www.jimmunol.org/content/169/4/1801doi: 10.4049/jimmunol.169.4.1801
2002; 169:1801-1809; ;J Immunol
Referenceshttp://www.jimmunol.org/content/169/4/1801.full#ref-list-1
, 14 of which you can access for free at: cites 40 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

The Th1-Specific Costimulatory Molecule, M150, Is aPosttranslational Isoform of Lysosome-Associated MembraneProtein-11
Durbaka V. R. Prasad,* Vrajesh V. Parekh,* Bimba N. Joshi,* Pinaki P. Banerjee,*Pradeep B. Parab,* Samit Chattopadhyay,* Anil Kumar,† and Gyan C. Mishra2*
In an earlier report, we had shown a 150-kDa protein termed as M150, isolated from the surface of activated macrophages, topossess costimulatory activity for CD4� T cells. Significantly, this protein was found to specifically elicit Th1 responses. In thisstudy, we characterize M150, which belongs to a unique subset of the lysosome-associated membrane protein-1 glycoprotein.Interestingly, the costimulatory activity of M150 depends on its posttranslational modification, which has a distinct glycosylationpattern restricted to macrophages. Furthermore, it has been demonstrated that in addition to stimulating Th1-specific responses,M150 is also capable of driving differentiation of naive CD4� T cells into the Th1 subset. This altered posttranslational modifi-cation of housekeeping protein appears to represent a novel pathway by which APCs can additionally regulate T cellresponses. The Journal of Immunology, 2002, 169: 1801–1809.
T he hallmark of the adaptive immune system is its abilityto induce as well as regulate its responses to a given in-fectious agent. Activation of CD4� T cells minimally re-
quires two distinct signals to be provided by an APC. Although thefirst signal is provided by the MHC class II molecule presentingthe appropriate Ag-derived peptide, the second signal is deliveredby a class of accessory molecules that are also known as costimu-latory molecules (1). A number of such costimulatory moleculeshave now been discovered, along with their counterreceptors on Tcells. The most prominent ones include B7.1 and B7.2 that interactwith CD28 on T cells (2–4), OX40 ligand-OX40 (5, 6), 4-1BBL-4-1BB (7, 8), and B7RP-1 that bind to inducible costimulatory (9,10). On the basis of their ability to secrete specific lymphokines,CD4� T cells have been divided into Th1 and Th2 subsets. Th1lymphocytes contributing to cellular immunity are characterizedby the production of IL-2 and IFN-�, whereas Th2 lymphocytesproduce IL-4, IL-5, IL-10, and IL-13, which are thought to bemainly involved in humoral immunity (11, 12). Th1- and Th2-associated cytokines tend to be reciprocally regulatory. IFN-� in-hibits Th2-associated functions (13), while IL-4 and IL-10 havenegative effect on Th1-associated functions (14). It has been pos-tulated that these two helper cell subsets are not only functionallydifferent, but also show qualitative and quantitative distinctions intheir requirements for costimulation (15).
In addition to ensuring the activation of T cells, accumulatingevidence suggests that costimulatory molecules may also play arole in regulating the qualitative aspects of T cell responses. Forexample, at least when expressed on activated B lymphocytes,B7.1/CD28 has been shown to predominantly activate the Th1subset of CD4� T cells (16–18), whereas B7.2/CD28 appears tobias toward Th2 responses (19–21). Similarly, 4-1BB-4-BBL in-teractions preferentially contribute toward the development of Th2responses. The recently identified costimulatory molecule B7RP-1,which is known to induce IFN-� production, is widely expressedon B cells and macrophages (10). In this context, a new costimu-latory molecule, B7-DC, which is specifically present on dendriticcells, has also been described. This molecule induces Th1-specificpolarization (22).
In an earlier study, we have reported the isolation of a 150-kDaprotein (M150) from the surface of activated macrophages. Thisprotein was shown to possess costimulatory activity, and was ca-pable of stimulating T cell proliferation. In addition, it was alsoable to induce secretion of lymphokines that are typical of Th1responses (23). Furthermore, we showed that macrophages pre-dominantly use M150, relative to B7.1 for costimulating prolifer-ation and IFN-� production from Th cells (24). M150 was alsoshown to restore normal Th1 function upon bystander costimula-tion in diseases like leishmaniasis and tuberculosis, where Th1-likeresponses are supressed (25, 26). However, the biochemical iden-tity of M150 has not been elucidated so far.
In the present study, we identify M150 as a specific posttrans-lational isoform of constitutively produced lysosome-associatedmembrane protein-1 (LAMP-1).3 Interestingly, the costimulatoryactivity of this molecule depends upon its unique pattern of gly-cosylation that is generated only in activated macrophages. Thus,M150 represents a novel example of a housekeeping protein thatcan also function as a costimulatory molecule, and adds to theexpanding list of APC-restricted costimulatory molecules with abiased influence on T cell function.
*National Centre For Cell Science, Pune, India; and †School of Biotechnology, DeviAhilya Vishwavidyalaya, Indore, India
Received for publication October 23, 2001. Accepted for publication June 17, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Department of Biotechnology, Government of India.D.V.R.P. and P.P.B. are the recipients of Senior Research Fellowship from NationalCentre for Cell Science, Department of Biotechnology, Pune, India. V.V.P. is a re-cipient of Senior Research Fellowship, and B.N.J. is a recipient of Research Asso-ciateship from Council for Scientific and Industrial Research, New Delhi, India.2 Address correspondence and reprint requests to Dr. Gyan C. Mishra, National Cen-ter for Cell Science, Ganeshkhind Road, Pune 411007, Maharashtra, India. E-mailaddress: [email protected]
3 Abbreviations used in this paper: LAMP-1, lysosome-associated membrane pro-tein-1; CHO, Chinese hamster ovary.
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists, Inc. 0022-1767/02/$02.00
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Materials and MethodsMacrophage membrane preparation and purification of M150
The peritoneal exudate cells were harvested from BALB/c mice (6–8 wkold) injected 4 days previously with 2 ml of 3% thioglycolate (Difco,Detroit, MI). The peritoneal exudate cells were washed with cold HBSSand macrophages were obtained by adhering for 1 h at 37°C on plastic petridishes and the purity of macrophage was �98%, as analyzed by theirreactivity with anti-mac-1 Ab. The macrophages were washed thrice andpellet was frozen overnight at �70°C. The cells were thawed and homog-enized in the presence of 20 mM Tris-Cl (pH 7.4) and 1 mM EDTA alongwith protease inhibitor mixture (10 �g/ml leupeptin, 10 �g/ml aprotinin,10 �g/ml pepstatin, 10 �g/ml antipain, 10 mM iodoacetamide, 10 �g/mlchymostatin, and 1 mM PMSF). The nuclear fraction was removed bycentrifuging at 700 � g for 10 min at 4°C. The supernatant was subjectedto centrifugation at 110,000 � g for 2 h at 4°C. The pellet was solubilizedovernight in 20 mM Tris-Cl (pH 7.5) containing 1% Triton X-100, 20%glycerol and protease inhibitor mixture, and recentrifuged at 100,000 � gfor 1 h at 4°C to remove the insoluble debris. The membrane proteins (27,28) in the supernatant were separated on 10% SDS-PAGE (29). M150 waseluted from the gel by crushing the gel pieces containing the protein bandfollowed by overnight incubation with 100 mM NH4 HCO3, 50 mM Tris-Cl, 0.1 mM EDTA, and 150 mM NaCl (pH 8.0). SDS from the proteinsolution was removed by passing through Extracti-D gel column (Pierce,Rockford, IL) and the protein content was estimated using the bicincho-ninic acid kit (Pierce). The protein was further purified by fast performanceliquid chromatography on Mono-Q column and the purity was ascertainedby two-dimensional electrophoresis before subjecting it to proteinsequencing.
Sequencing of M150
The internal and amino-terminal sequencing were conducted independentlyat two different facilities. Typically, the purified M150 was electrophoresedon 10% SDS-PAGE and blotted on to polyvinylidene difluoride membraneusing 10 mM CAPS buffer (US Biochemicals, Cleveland, OH) containing10% methanol at 200 mA for 2 h at 4°C. The blot then was subjected totryptic digestion and the peptides thus obtained were sequenced for internalsequencing at the protein sequencing facility of Worcester Foundation forExperimental Biology. The protein obtained by FPLC was subjected toN-terminal sequencing at the sequencing facility of Purdue University(West Lafayette, IN).
Heteroduplex analysis
Primers were designed from the internal stretches of the M150 protein thatwas sequenced earlier. Sense primer 5�-GAGATCTACACAATGGACTC-3�and antisense primer 5�-GAGTCCATXGTGTAGATCTC-3� were custommade (Life Technologies, Grand Island, NY). The cDNA of LAMP-1 clonedfrom mouse embryo 3T3 cDNA library was a gift from Dr. J. T. August(John Hopkins University, Baltimore, MD). PCR was performed usingthese primers at an annealing temperature of 55°C. RT-PCR was doneusing the same primers from RNA isolated from thioglycolate-elicitedmacrophages using S.N.A.P. total RNA isolation kit (Invitrogen, San Di-ego, CA). A total of 500 ng of total RNA was used to perform RT-PCRusing Titan one tube RT-PCR system (Roche, Indianapolis, IN). The prod-uct was subcloned in PCR cloning vector. After transformation and platingfrom several different colonies, PCR was performed from plasmid prepa-rations using the same primers. A total of 0.5 �g of each PCR product wasmixed with 0.5 �g of PCR product obtained from the cDNA construct ofLAMP-1. Heteroduplex analysis was conducted in TNE buffer, final con-centration Tris:NaCl:EDTA equals 1/10/0.1 mM (30). The PCR productmixtures were made to a final volume of 10 �l and heated at 95°C for 5 minfollowed by rapid cooling in an ice bath for 60 min. The sample was mixedwith loading dye and electrophoresed on an 8% native PAGE using TBEbuffer. DNA pattern was visualized by ethidium bromide staining.
mAb preparation
Lewis rats 8–10 wk old, obtained from National Institute of Immunology(New Delhi, India) were immunized by i.p. injections of thioglycolate-elicited macrophages (107 cells/mice) from BALB/c mice. Three boosterdoses were given after 21 days of primary immunization at an interval of2 wk each. The spleen was removed on the third day after the final boosterdose and the cells were fused to SP2/01-AG14 (American Type CultureCollection, Manassas, VA) using PEG 1500 (Roche). Hybrids were se-lected in hypoxanthine/aminopterin/thymidine medium (Life Technolo-gies). The supernatants were screened for M150 reactivity both by ELISAand Western blot analysis. Positive clones were subcloned by limiting di-lution and the isotype of Abs secreted by individual clones was determined
using mAb-based Rat Ig isotyping kit (BD PharMingen, San Diego, CA).For the present study, anti-M150 Abs were produced as ascites, and puri-fied by sequential chromatography over Sepahcryl S 300, followed by goatanti-rat IgM affinity chromatography.
FACS analysis
The affinity-purified mAb (G1) against M150 and control IgM (normal ratIgM, affinity purified by sheep anti-rat IgM; Pierce) were FITC conjugated.PE-conjugated anti-Mac-1, anti-B220, anti-CD4, anti-CD8, normal IgG2a,normal IgG 2b, and FITC-labeled anti-LAMP-1 Ab, FITC-labeled normalrat IgG2a (isotype control for anti-LAMP-1), and Fc block were purchasedfrom BD PharMingen. Splenocytes and macrophages (106 cells), eitherresting or activated with IFN-� (5 ng/ml for 12 h) from C57 BL/6 mice,were first incubated with Fc block for 20 min at 4°C and the splenocyteswere then dually stained either with FITC-labeled anti-M150 or anti-LAMP-1 along with various PE-conjugated CD markers. Incubations weredone at 4°C in FACS buffer (2% FCS, 0.5% BSA in PBS, pH 7.2) andwashed using the same buffer three times before incubation with the sec-ondary Ab. Finally, cells were washed and fixed in 1% paraformaldehydein PBS and staining was analyzed by flow cytometry (FACSVantage; BDBiosciences, Mountain View, CA). Macrophages were used for the com-petitive binding experiment between the anti-LAMP-1 and anti-M150 Abs.
Western blotting
The samples were electrophoresed using 10% SDS-PAGE (SE 260; Am-ersham Pharmacia Biotech, Uppsala, Sweden) and transferred onto a ni-trocellulose membrane (Schleicher & Schuell, Dassel, Germany) using 20mM Tris containing 125 mM glycine (pH 7.5) and 20% methanol at 200mA for 2 h at 4°C. The membrane was blocked with 2% BSA in TBST (50mM Tris-Cl, 150 mM NaCl, 0.05% Tween 20, pH 8). Incubations andwashings of primary and secondary Abs were done with TBST at roomtemperature. Blots were developed using diaminobenzidine (Sigma-Al-drich, St. Louis, MO).
Generation of LAMP-1-Fc fusion protein
The pCEP4-hFc vector that encodes the Fc portion of human IgG1 was a giftfrom Drs. T. W. Mak and Dr. S. K. Yoshinaga (Amgen, Thousand Oaks, CA).For cloning LAMP-1 in pCEP4-hFc vector, 370 aa from the amino-terminal ofLAMP-1 were fused in frame to the sequence encoding the Fc portion in theamino terminus region. The forward and reverse primers, 5�-CGCAAGCTTATGCGGCCCCCGCGCGCG-3� and 5�-CGCGCGGCCGCGTTGTTACCATCCTGAACACACTC-3� were used for cloning the LAMP-1 truncatedgene into the HindIII and NotI sites within the multiple cloning sites ofpCEP4. The coding sequence of LAMP-1 from N terminus up to the trans-membrane domain, devoid of the region spanning the membrane, was in-corporated into pCEP4-Fc. The highly purified pCEP4-LAMP-1-Fc plas-mid was then transfected into Chinese hamster ovary (CHO) cells and themouse macrophage cell line P388D1 using FuGene 6 transfection reagent(Roche). The transfected cells were grown in serum-free media containinginsulin-transferrin-selenium-A (Life Technologies). Soluble secreted fu-sion proteins were purified from culture supernatants using protein A aga-rose affinity column chromatography (Roche).
Deglycosylation of transfected fusion proteins
A total of 5 �g of fusion proteins, i.e., CHO-LAMP-1-Fc or macrophage-LAMP-1-Fc, were treated with 20 mU endoglycosidase H (Roche) for 16 hat 37°C using 0.5 M sodium citrate buffer (pH 5.5) containing 0.1 M 2-MEand 0.1% SDS.
T cell proliferation, cytokine, and blocking assays
Spleens from 6- to 8-wk-old female BALB/c obtained from experimentalanimal facility of our institute were used to make single-cell suspensions ofsplenocytes. RBCs were lysed using hemolytic Gey’s solution. Nonadher-ent cells were collected from supernatants after allowing cells to adhere toplastic petri dishes (Corning Glass, Corning, NY) at 37°C in the presenceof 5% CO2 for 2 h. The CD4� T cells were enriched by passing througha nylon wool column (Robbins Scientific, Sunnyvale, CA). Finally, CD4�
T cells were purified to at least 98% purity by negative selection using theCD4� T cell enrichment mixture (StemCell Technologies, Vancouver,British Columbia, Canada). The T cells were cultured in RPMI 1640 (LifeTechnologies) supplemented with penicillin (70 �g/ml), streptomycin (100�g/ml), glutamine (4 mM), 2-ME (50 mM), sodium pyruvate (1 mM),HEPES (20 �M), and heat-inactivated 10% FCS (Life Technologies). Thepurified T cells (105) were cultured along with anti-CD3, either in presenceor absence of varying concentrations of Fc fusion proteins for 72 h in96-well, flat-bottom plate (Costar, Cambridge, MA). These cells were then
1802 GLYCOSYLATION MODULATES THE COSTIMULATORY PROPERTY OF LAMP-1
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

pulsed with [3H]thymidine (1 �Ci/well) during the final 12 h of the cultureperiod, following which the incorporated radioactivity was determined byliquid scintillation counting. For blocking experiments, anti-LAMP-1(ID4B) and IgG2a isotype (BD PharMingen), along with affinity-purifiedanti-M150 and normal rat IgM were used. In parallel experiments, super-natants from such cultures were also collected for determination of IL-2,IFN-�, IL-4, and/or IL-13 by ELISA (R&D Systems, Minneapolis, MN).
RT-PCR for c-Maf and T-bet
Total RNA was prepared from 106 CD4� T cells per well cultured in24-well plate for 24 h either in the presence of CHO-LAMP-1-Fc or mac-rophage-LAMP-1-Fc and 1 �g/ml of soluble anti-CD3 (BD PharMingen).The total RNA was isolated using S.N.A.P. total RNA isolation kit (Invitro-gen). A total of 500 ng of total RNA was used to perform RT-PCR using Titanone tube RT-PCR System (Roche). The T-bet primers used were 5�-ATGGGCATCGTGGAGCCGGGCT-3� and 5�-ACTTGGACCACACAGGTGGTTG-3�, cMaf primers were 5�-ACTGAACCGCAGCTGCGCGGGGTCAG-3�and 5�-CTTCTCGTATTTCTCCTTGTAGGCGTCC-3� (31), which werecustom made (Gemini Biotech, Alachua, FL). The control primers used weredihydrofolatereductase set for RT-PCR (Stratagene, La Jolla, CA).
ResultsThe protein M150 displays amino acid sequence homology withLAMP-1
In our earlier studies, we have isolated a 150-kDa protein from themembrane fraction of activated murine macrophages. This protein,termed as M150, was demonstrated to provide costimulatory ac-tivity to CD4� T cells. Interestingly, expression of M150 on thesurface of the activated macrophages was found to specificallyelicit Th1 responses from CD4� T cells, at least when assessed at
the level of cytokine production in culture supernatants. Further-more, surface expression of M150 could be detected on activatedmacrophages but not on activated B cells. Thus, M150 constitutesa macrophage restricted costimulatory molecule that specificallydrives Th1 responses. Therefore, it was of interest to establish thebiochemical identity of this cell surface protein.
Using a previously established procedure, a homogenous prep-aration of M150 was obtained from the plasma membrane fractionof activated macrophages (Fig. 1, A and B) and the purified M150was subjected to internal and amino-terminal sequencing by twoindependent preparations at two different sequencing facilities.The three partial sequences thus obtained were subsequently usedin a search for homologous sequences within the protein databases.Surprisingly, all of the three partial sequences yielded 100% iden-tity with segments of the murine LAMP-1 protein. As seen in Fig.1C, the amino terminus of M150 was identical with that ofLAMP-1, and the two internal sequences obtained were found tocorrespond to segments between aa 128–139 and 288–293 of theLAMP-1 protein. These results strongly suggested that M150 iseither identical with, or highly homologous to LAMP-1 (32, 33).
To further confirm this, and to check whether there is any al-ternate form of LAMP-1 gene existing in macrophages with inter-nal sequence differences as compared with 3T3 LAMP-1 cDNA, aheteroduplex analysis was performed. For this, corresponding oli-gonucleotide segments from the internal amino acid sequences ofM150 were used as primers to amplify the corresponding segmentfrom cDNA of LAMP-1. In parallel, these primers were also used
FIGURE 1. Electrophoretic analysis of M150 protein and its characterization. A, SDS-PAGE of plasma membrane proteins isolated from thioglycolateexudate macrophages. Macrophage membrane proteins were prepared as described in Materials and Methods. In brief, macrophages (98% mac-1 positive)were homogenized and after removal of the debris by low-speed centrifugation, the supernatant was subjected to ultra centrifugation at 110,000 � g. Thepellet was solubilized and it was recentrifuged at 100,000 � g. The supernatant containing the membrane fraction was separated on a 10% SDS-PAGE andwas stained with Coomassie blue. Lane 1, Molecular mass markers; lane 2, Surface membrane proteins of macrophage; lane 3, Gel-purified M150 underreducing condition. B, Two-dimensional electrophoresis of M150. The purified M150 (as shown in A, lane 3) was further purified by fast performance liquidchromatography on a Mono-Q column and subjected to two-dimensional electrophoresis. The M150 was resolved by isoelectro focusing across a pHgradient of 3.5–9.5 (first dimension) and then electrophoresed on a 10% SDS-PAGE (second dimension). C, Sequence identity of partial amino acidsequences of M150 with LAMP-1. Sequence data of N-terminal and internal amino acid sequences of M150 in comparison with LAMP-1. Peptide sequence1 was derived from the amino-terminal end of M150. Peptide sequences 2 and 3 are two internal sequences derived by tryptic digestion of M150 usinga different preparation as reported by us earlier (23). D, Heteroduplex analysis. Heteroduplex analysis was conducted as described in Materials and Methods.The 500-bp PCR product generated from LAMP-1 cDNA was mixed with PCR product obtained from different clones (representing the RT-PCR productof thioglycolate-elicited macrophages). Duplex DNA thus generated was electrophoresed on 8% native PAGE and visualized by ethidium bromide staining.Lanes 1–3, Results for 3 of the 40 clones tested are shown as representatives.
1803The Journal of Immunology
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
to amplify the cDNA, isolated from thioglycolate-elicited macro-phages. The amplified product was then cloned into an appropriatevector and used for transformation. The plasmid DNA from thesecolonies was then PCR amplified and subjected to a heteroduplexanalysis with the PCR product derived from cDNA of LAMP-1. Itmay be mentioned in this study that mutations or sequence vari-ations are easily detected by gel electrophoresis of heteroduplexes,where minute sequence variations appear as blebs on the gel. Incontrast, identical sequences occur as homoduplexes. All of the40 clones tested in hetroduplex analysis exhibited homoduplexformation. Results of three of these clones are shown in Fig. 1Das a representative example. Therefore, these results stronglysuggest that M150 protein is in fact LAMP-1 protein, which is
an activation-dependent cell surface glycoprotein also known asCD107a (34).
Differential expression of anti-M150 and anti-LAMP-1 reactivityin splenocyte subsets
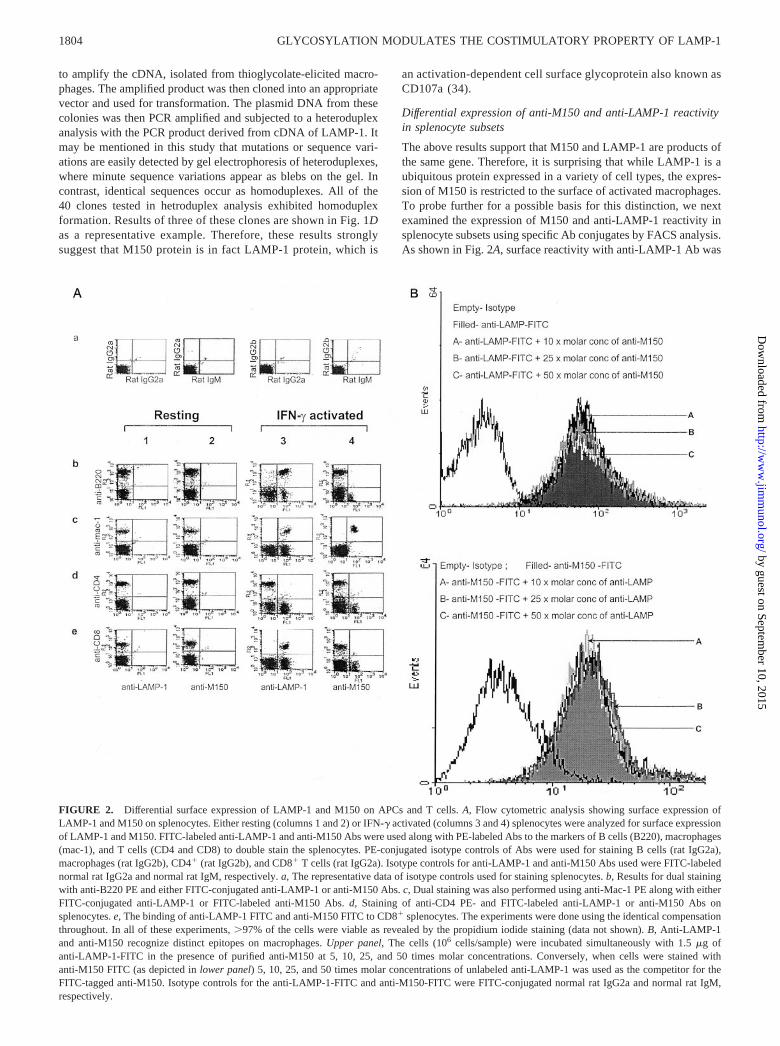
The above results support that M150 and LAMP-1 are products ofthe same gene. Therefore, it is surprising that while LAMP-1 is aubiquitous protein expressed in a variety of cell types, the expres-sion of M150 is restricted to the surface of activated macrophages.To probe further for a possible basis for this distinction, we nextexamined the expression of M150 and anti-LAMP-1 reactivity insplenocyte subsets using specific Ab conjugates by FACS analysis.As shown in Fig. 2A, surface reactivity with anti-LAMP-1 Ab was
FIGURE 2. Differential surface expression of LAMP-1 and M150 on APCs and T cells. A, Flow cytometric analysis showing surface expression ofLAMP-1 and M150 on splenocytes. Either resting (columns 1 and 2) or IFN-� activated (columns 3 and 4) splenocytes were analyzed for surface expressionof LAMP-1 and M150. FITC-labeled anti-LAMP-1 and anti-M150 Abs were used along with PE-labeled Abs to the markers of B cells (B220), macrophages(mac-1), and T cells (CD4 and CD8) to double stain the splenocytes. PE-conjugated isotype controls of Abs were used for staining B cells (rat IgG2a),macrophages (rat IgG2b), CD4� (rat IgG2b), and CD8� T cells (rat IgG2a). Isotype controls for anti-LAMP-1 and anti-M150 Abs used were FITC-labelednormal rat IgG2a and normal rat IgM, respectively. a, The representative data of isotype controls used for staining splenocytes. b, Results for dual stainingwith anti-B220 PE and either FITC-conjugated anti-LAMP-1 or anti-M150 Abs. c, Dual staining was also performed using anti-Mac-1 PE along with eitherFITC-conjugated anti-LAMP-1 or FITC-labeled anti-M150 Abs. d, Staining of anti-CD4 PE- and FITC-labeled anti-LAMP-1 or anti-M150 Abs onsplenocytes. e, The binding of anti-LAMP-1 FITC and anti-M150 FITC to CD8� splenocytes. The experiments were done using the identical compensationthroughout. In all of these experiments, �97% of the cells were viable as revealed by the propidium iodide staining (data not shown). B, Anti-LAMP-1and anti-M150 recognize distinct epitopes on macrophages. Upper panel, The cells (106 cells/sample) were incubated simultaneously with 1.5 �g ofanti-LAMP-1-FITC in the presence of purified anti-M150 at 5, 10, 25, and 50 times molar concentrations. Conversely, when cells were stained withanti-M150 FITC (as depicted in lower panel) 5, 10, 25, and 50 times molar concentrations of unlabeled anti-LAMP-1 was used as the competitor for theFITC-tagged anti-M150. Isotype controls for the anti-LAMP-1-FITC and anti-M150-FITC were FITC-conjugated normal rat IgG2a and normal rat IgM,respectively.
1804 GLYCOSYLATION MODULATES THE COSTIMULATORY PROPERTY OF LAMP-1
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

displayed by activated splenic B cells, macrophages, and CD8� Tcells. In contrast, anti-M150 reactivity was restricted only to theMac-1� subset of activated splenocytes. This suggests that M150is antigenically distinct from LAMP-1. However, as depicted inFig. 2A, the anti-M150 and anti-LAMP-1 Abs have been found tobind only with the IFN-�-activated macrophage populations (i.e.,the Mac-1� populations of the splenocytes). Therefore, we testedthe possibility of these two Abs for competing for the same epitopeof LAMP-1 present on macrophage membrane. To address thisissue, IFN-�-activated peritoneal exudate macrophages werestained with anti-M150 FITC-labeled Ab keeping the unlabeledanti-LAMP-1 Ab as a competitor of its specific site and vice versa.Fig. 2B demonstrates that there is no significant difference in thebinding of either of the labeled Abs in the presence of the other ascompetitor, which conclusively proves that anti-LAMP-1 and anti-M150 Abs do not compete for binding to IFN-�-activated macro-phages. Thus, this result also clearly substantiates that anti-LAMP-1 and anti-M150 bind to different epitopes of LAMP-1.However, NK T cells, dendritic cells, and monocytes were nega-tive for staining with anti-LAMP-1 and anti-M150 Abs (data notshown).
Differential glycosylation gives antigenic distinctness to LAMP-1
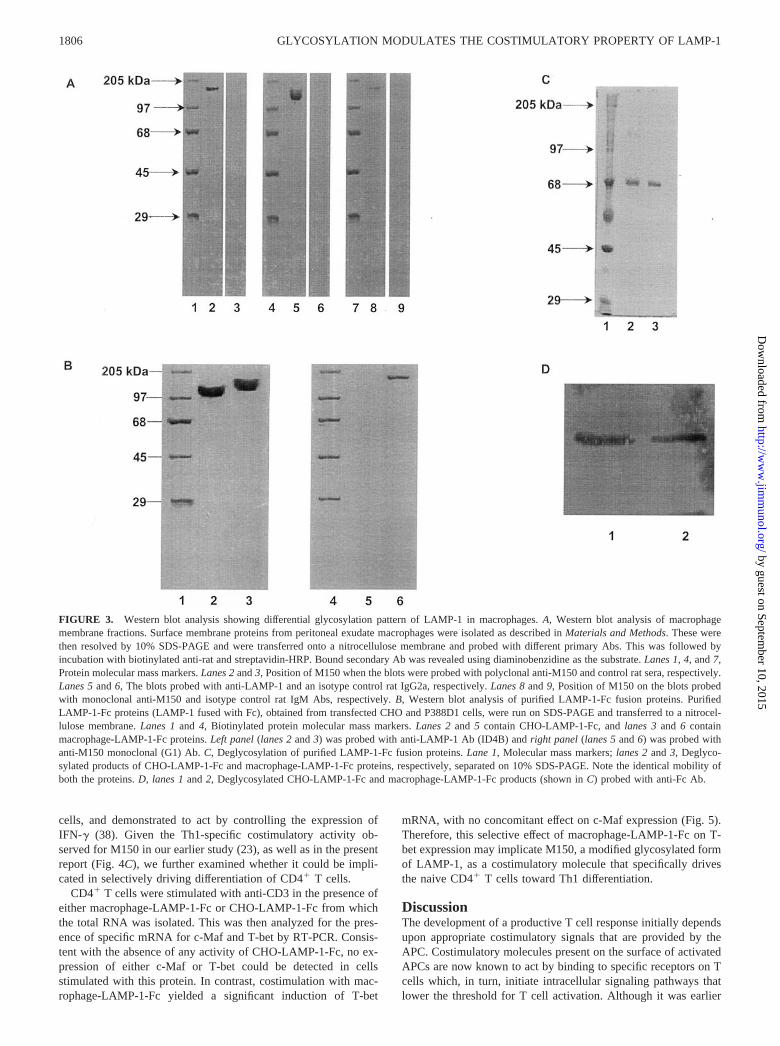
The existence of antigenic variance between M150 and LAMP-1was also confirmed by Western blot analysis of the membranefraction of activated macrophages using either anti-M150 or anti-LAMP-1 Abs as probes. Probing with either polyclonal or mono-clonal anti-M150 Ab (G1) detected a band that centered around amolecular mass of 150 kDa. As opposed to this, anti-LAMP-1 Abidentified a lower band ranging from 120–145 kDa (Fig. 3A). Pre-vious reports have demonstrated that the size heterogeneity ofLAMP-1 is due to heterogeneous glycosylation (32). These studiesalso revealed that the size distribution of LAMP-1 differs signifi-cantly, depending upon the cell type examined. In other words, theglycosylation pattern of LAMP-1 appears to vary depending uponthe cell type in which it is expressed (35).
To further confirm the possible cell type-specific differences inglycosylation of LAMP-1, we expressed a truncated version ofLAMP-1 protein as LAMP-1-Fc chimera either in CHO cells, or inthe mouse macrophage cell line, P388D1. The transfected fusionproteins were then purified and subjected to a Western blot anal-ysis. As shown in Fig. 3B, nonidentical forms of the chimericprotein were produced in CHO and P388D1 cells, at least as de-tected by anti-LAMP-1 and anti-M150 mAbs. The product of CHOcells displayed a molecular mass between 105 and 120 kDa; incontrast, the product from P388D1 cells was significantly of highermolecular mass, ranging from 130–150 kDa. Interestingly, anti-M150 mAb did not show reactivity with LAMP-1 protein ex-pressed in CHO cells while recognizing the LAMP-1 expressed inmacrophages (Fig. 3B, compare lanes 5 and 6). In contrast, anti-LAMP-1 recognized a discrete number of lower molecular massproteins from LAMP-1-transfected CHO and P388D1 cells asshown in Fig. 3B, lanes 2 and 3, respectively. A comparison of theprotein bands (Fig. 3B, lanes 3 and 6) recognized by anti-LAMP-1and anti-M150 Abs suggest that the M150 is a specific form whichconstitutes only a small fraction of the total LAMP-1 produced inmacrophages.
To compare the protein core of these two molecular species, theexpressed products from both CHO and P388D1 cells were firstdeglycosylated using endoglycosidase H. Surprisingly, this proce-dure completely eliminated the reactivity of the anti-LAMP-1 andanti-M150 Abs. Thus, the resultant deglycosylated proteins couldbe detected only with anti-Fc Ab.
Therefore, these results collectively suggest that both Abs prep-arations are directed against the glycosylated portion of the mol-ecules. Thus, from these results it can also be inferred that theexpression of LAMP-1-Fc chimera in CHO and P388D1 cells re-sults from qualitatively distinct glycosylation patterns. Further-more, it is this variation in glycosylation that accounts for theobserved difference in antigenicity, at least with respect to anti-M150 Ab. In other words, the anti-M150 mAb appears to be di-rected against a LAMP-1 subset that is specifically produced onlyin macrophages.
The LAMP-1-Fc chimera displays costimulatory activity whenexpressed in macrophages
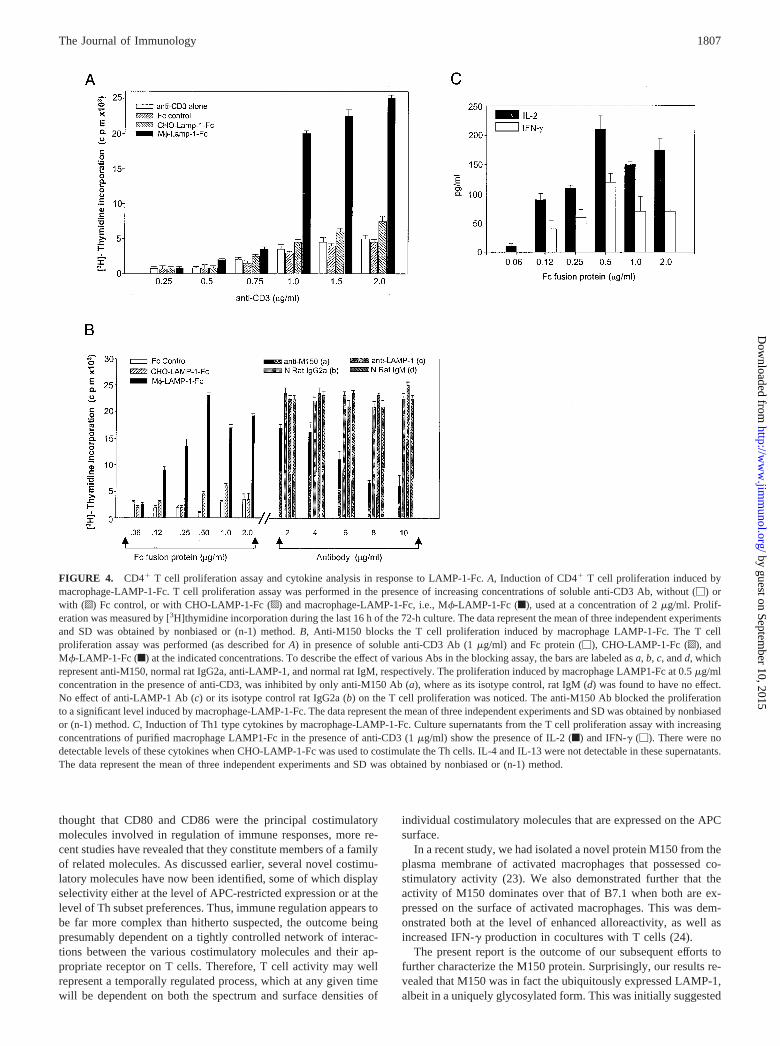
We next examined whether the altered glycosylation pattern of theLAMP-1-Fc chimera also confers altered function for the proteinmolecule when expressed in P388D1 vs CHO cells. Therefore,purified CD4� T cells were stimulated with varying concentrationof anti-CD3 Ab in the presence of a fixed concentration of eitherCHO-LAMP-1-Fc or macrophage-LAMP-1-Fc proteins. A prolif-erative response was observed only in cultures that included mac-rophage-LAMP-1-Fc, but not in those where CHO-LAMP-1-Fcwas added (Fig. 4A).
It is possible that the difference in activity observed for macro-phage-LAMP-1-Fc and CHO-LAMP-1-Fc in Fig. 4A simply re-flects differences in dose requirements for the two chimeric prod-ucts. To verify this, T cells were stimulated with a constant dose of1 �g/ml of anti-CD3 Ab in the presence of increasing concentra-tions of either macrophage-LAMP-1-Fc or CHO-LAMP-1-Fc. Forthese experiments, a dose-dependent proliferation of CD4� T cellswas obtained with macrophage-LAMP-1-Fc (Fig. 4B). In contrastto this, CHO-LAMP-1-Fc was unable to induce T cell proliferationat any of its tested concentrations (Fig. 4B). Furthermore, the spec-ificity of the macrophage-LAMP-1-Fc-dependent T cell responseis also evident from the fact that this effect could be inhibited byaddition of anti-M150 Ab, but not by anti-LAMP-1 Ab (Fig. 4B).
We have previously demonstrated that M150 serves as a co-stimulatory molecule that specifically elicits cytokines typical ofthe Th1 subset of CD4� T cells. As shown in Fig. 4C, the mac-rophage-LAMP-1-Fc product also reproduced this activity whenused in conjunction with anti-CD3 for stimulation of CD4� Tcells. Significant levels of both IL-2 and IFN-� were detected inthe supernatants from these cultures (Fig. 4C). In contrast, stimu-lation in presence of CHO-LAMP-1-Fc failed to induce detectablelevels of Th1 representative cytokines (Fig. 4C). It is pertinent tomention that no detectable levels of Th2 representative cytokineswere obtained from the culture supernatants of Th cells activatedeither by CHO-LAMP-1-Fc or macrophage LAMP-1-Fc. Collec-tively, these results categorically identify that LAMP-1 can indeeddisplay Th1-specific costimulatory activity, but this activity is con-tingent upon its expression in macrophages.
Macrophage-LAMP-1-Fc induces expression of T-bet in naiveCD4� T cells
It is now becoming evident that differentiation of CD4� T cellsinto either the Th1 or Th2 commitment pathways is regulated bythe activation of independent transcription factors (36). Subset-specific transcription factors have been identified and their role indistinct T cell differentiation is being actively explored. For ex-ample, STAT-6 has been shown to synergize with an antigenicstimulus leading to the up-regulation of GATA3, a potent inducerof Th2 differentiation (31). The transcription factor c-Maf, whichnegatively regulates Th1 differentiation, has now been shown to beresponsible for the tissue-specific expression of IL-4 (37). Re-cently, T-bet was identified as a transcription factor specific to Th1
1805The Journal of Immunology
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
cells, and demonstrated to act by controlling the expression ofIFN-� (38). Given the Th1-specific costimulatory activity ob-served for M150 in our earlier study (23), as well as in the presentreport (Fig. 4C), we further examined whether it could be impli-cated in selectively driving differentiation of CD4� T cells.
CD4� T cells were stimulated with anti-CD3 in the presence ofeither macrophage-LAMP-1-Fc or CHO-LAMP-1-Fc from whichthe total RNA was isolated. This was then analyzed for the pres-ence of specific mRNA for c-Maf and T-bet by RT-PCR. Consis-tent with the absence of any activity of CHO-LAMP-1-Fc, no ex-pression of either c-Maf or T-bet could be detected in cellsstimulated with this protein. In contrast, costimulation with mac-rophage-LAMP-1-Fc yielded a significant induction of T-bet
mRNA, with no concomitant effect on c-Maf expression (Fig. 5).Therefore, this selective effect of macrophage-LAMP-1-Fc on T-bet expression may implicate M150, a modified glycosylated formof LAMP-1, as a costimulatory molecule that specifically drivesthe naive CD4� T cells toward Th1 differentiation.
DiscussionThe development of a productive T cell response initially dependsupon appropriate costimulatory signals that are provided by theAPC. Costimulatory molecules present on the surface of activatedAPCs are now known to act by binding to specific receptors on Tcells which, in turn, initiate intracellular signaling pathways thatlower the threshold for T cell activation. Although it was earlier
FIGURE 3. Western blot analysis showing differential glycosylation pattern of LAMP-1 in macrophages. A, Western blot analysis of macrophagemembrane fractions. Surface membrane proteins from peritoneal exudate macrophages were isolated as described in Materials and Methods. These werethen resolved by 10% SDS-PAGE and were transferred onto a nitrocellulose membrane and probed with different primary Abs. This was followed byincubation with biotinylated anti-rat and streptavidin-HRP. Bound secondary Ab was revealed using diaminobenzidine as the substrate. Lanes 1, 4, and 7,Protein molecular mass markers. Lanes 2 and 3, Position of M150 when the blots were probed with polyclonal anti-M150 and control rat sera, respectively.Lanes 5 and 6, The blots probed with anti-LAMP-1 and an isotype control rat IgG2a, respectively. Lanes 8 and 9, Position of M150 on the blots probedwith monoclonal anti-M150 and isotype control rat IgM Abs, respectively. B, Western blot analysis of purified LAMP-1-Fc fusion proteins. PurifiedLAMP-1-Fc proteins (LAMP-1 fused with Fc), obtained from transfected CHO and P388D1 cells, were run on SDS-PAGE and transferred to a nitrocel-lulose membrane. Lanes 1 and 4, Biotinylated protein molecular mass markers. Lanes 2 and 5 contain CHO-LAMP-1-Fc, and lanes 3 and 6 containmacrophage-LAMP-1-Fc proteins. Left panel (lanes 2 and 3) was probed with anti-LAMP-1 Ab (ID4B) and right panel (lanes 5 and 6) was probed withanti-M150 monoclonal (G1) Ab. C, Deglycosylation of purified LAMP-1-Fc fusion proteins. Lane 1, Molecular mass markers; lanes 2 and 3, Deglyco-sylated products of CHO-LAMP-1-Fc and macrophage-LAMP-1-Fc proteins, respectively, separated on 10% SDS-PAGE. Note the identical mobility ofboth the proteins. D, lanes 1 and 2, Deglycosylated CHO-LAMP-1-Fc and macrophage-LAMP-1-Fc products (shown in C) probed with anti-Fc Ab.
1806 GLYCOSYLATION MODULATES THE COSTIMULATORY PROPERTY OF LAMP-1
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
thought that CD80 and CD86 were the principal costimulatorymolecules involved in regulation of immune responses, more re-cent studies have revealed that they constitute members of a familyof related molecules. As discussed earlier, several novel costimu-latory molecules have now been identified, some of which displayselectivity either at the level of APC-restricted expression or at thelevel of Th subset preferences. Thus, immune regulation appears tobe far more complex than hitherto suspected, the outcome beingpresumably dependent on a tightly controlled network of interac-tions between the various costimulatory molecules and their ap-propriate receptor on T cells. Therefore, T cell activity may wellrepresent a temporally regulated process, which at any given timewill be dependent on both the spectrum and surface densities of
individual costimulatory molecules that are expressed on the APCsurface.
In a recent study, we had isolated a novel protein M150 from theplasma membrane of activated macrophages that possessed co-stimulatory activity (23). We also demonstrated further that theactivity of M150 dominates over that of B7.1 when both are ex-pressed on the surface of activated macrophages. This was dem-onstrated both at the level of enhanced alloreactivity, as well asincreased IFN-� production in cocultures with T cells (24).
The present report is the outcome of our subsequent efforts tofurther characterize the M150 protein. Surprisingly, our results re-vealed that M150 was in fact the ubiquitously expressed LAMP-1,albeit in a uniquely glycosylated form. This was initially suggested
FIGURE 4. CD4� T cell proliferation assay and cytokine analysis in response to LAMP-1-Fc. A, Induction of CD4� T cell proliferation induced bymacrophage-LAMP-1-Fc. T cell proliferation assay was performed in the presence of increasing concentrations of soluble anti-CD3 Ab, without (�) orwith (o) Fc control, or with CHO-LAMP-1-Fc (p) and macrophage-LAMP-1-Fc, i.e., M�-LAMP-1-Fc (f), used at a concentration of 2 �g/ml. Prolif-eration was measured by [3H]thymidine incorporation during the last 16 h of the 72-h culture. The data represent the mean of three independent experimentsand SD was obtained by nonbiased or (n-1) method. B, Anti-M150 blocks the T cell proliferation induced by macrophage LAMP-1-Fc. The T cellproliferation assay was performed (as described for A) in presence of soluble anti-CD3 Ab (1 �g/ml) and Fc protein (�), CHO-LAMP-1-Fc (o), andM�-LAMP-1-Fc (f) at the indicated concentrations. To describe the effect of various Abs in the blocking assay, the bars are labeled as a, b, c, and d, whichrepresent anti-M150, normal rat IgG2a, anti-LAMP-1, and normal rat IgM, respectively. The proliferation induced by macrophage LAMP1-Fc at 0.5 �g/mlconcentration in the presence of anti-CD3, was inhibited by only anti-M150 Ab (a), where as its isotype control, rat IgM (d) was found to have no effect.No effect of anti-LAMP-1 Ab (c) or its isotype control rat IgG2a (b) on the T cell proliferation was noticed. The anti-M150 Ab blocked the proliferationto a significant level induced by macrophage-LAMP-1-Fc. The data represent the mean of three independent experiments and SD was obtained by nonbiasedor (n-1) method. C, Induction of Th1 type cytokines by macrophage-LAMP-1-Fc. Culture supernatants from the T cell proliferation assay with increasingconcentrations of purified macrophage LAMP1-Fc in the presence of anti-CD3 (1 �g/ml) show the presence of IL-2 (f) and IFN-� (�). There were nodetectable levels of these cytokines when CHO-LAMP-1-Fc was used to costimulate the Th cells. IL-4 and IL-13 were not detectable in these supernatants.The data represent the mean of three independent experiments and SD was obtained by nonbiased or (n-1) method.
1807The Journal of Immunology
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

from a partial amino acid sequence analysis of the intact M150protein. Our inference was further supported by heteroduplex anal-ysis, which unambiguously demonstrated the identity betweenM150 and LAMP-1. Finally, expression of the extracellular do-main part of LAMP-1 as a fusion protein in macrophages, but notCHO cells, also yielded a product with M150-like activity.
The functional properties of LAMP-1 continue to remain elu-sive. Observations that LAMP-1 expression on the surface of sev-eral tumors increases their metastatic potential suggest a role forthis protein in mediating cell adhesion to vascular endothelium(39). However, direct evidence that supports either an in vivo func-tion, or a physiological role for LAMP-1 is currently lacking. Aparticularly striking aspect of our results is the finding that auniquely glycosylated form of LAMP-1 possesses costimulatoryactivity. To our knowledge, this is the first report, which indicatesthat T cell costimulating potential can also derive from posttrans-lational modification of an otherwise inactive molecule. However,the nature of glycosylation that confers costimulatory activity toLAMP-1 remains to be characterized. Our present results particu-larly underscore the versatility that is built into immune regulatoryprocesses. Thus, in addition to using “professional” costimulatorymolecules, recruitment of housekeeping proteins by selective post-translational modifications may constitute an additional pathwayby which T cell responses may be modulated. A possible signifi-cance of the latter pathway could be that it facilitates APC-specificeffects on T cell costimulation. Significantly, our results demon-strate that costimulation of T cells with macrophage-LAMP-1-Fcinduces expression of T-bet may also implicate a role of M150 indriving differentiation of naive CD4� T cells into the Th1pathway.
In summary, M150 is a uniquely glycosylated form of LAMP-1that is produced only in activated macrophages. However, it re-mains to be ascertained if LAMP-1 represents an isolated example,or if there are other such housekeeping molecules that can alsofunction as costimulatory molecules under specific circumstances.As the catalogue of APC-surface molecules that provide costimu-lation to T cells increases, the question of their cumulative in-volvement in the regulation of immune responses also assumesincreasing importance. It would also be to determine whether thisapparent redundancy of costimulatory molecules simply provides asafeguard against evasive strategies evolved by pathogens, or ifthey constitute a cooperative network that appropriately regulatesthe response depending on the nature of “danger” perceived (40).
AcknowledgmentsWe thank Drs. Tak W. Mak and S. K. Yoshinaga SK for providingpCEP4-Fc vector, and Dr. J. Thomas August for providing LAMP-1cDNA. We are extremely grateful to Dr. K. V. S. Rao (International Centerfor Genetic Engineering and Biotechnology, New Delhi, India) for criticalevaluation and suggestions. A part of the manuscript was presented toNational Academy of Sciences (Allahabad, India) as presidential address(Biological Section). We also thank Drs. A. Sahu, K. Shastry, and S. Bapatfor critical reading of the manuscript, and Atul Suple for excellent technicalassistance.
References1. Schwartz, R. H. 1985. T-lymphocyte recognition of antigen in association with
gene products of the major histocompatibility complex. Annu. Rev. Immunol.3:237.
2. Lenschow, D. J., T. L. Walunas, and J. A. Bluestone. 1996. CD28/B7 system ofT cell costimulation. Annu. Rev. Immunol. 14:233.
3. Linsley, P. S., W. Brady, L. Grosmaire, A. Aruffo, N. K. Damle, andJ. A. Ledbetter. 1991. Binding of the B cell activation antigen B7 to CD28costimulates T cell proliferation and interleukin 2 mRNA accumulation. J. Exp.Med. 173:721.
4. Croft, M., and C. Dubey. 1997. Accessory molecule and costimulation require-ments for CD4 T cell response. Crit. Rev. Immunol. 17:89.
5. Flynn, S., K. M. Toellner, C. Raykundalia, M. Goodall, and P. Lane. 1998. CD4T cell cytokine differentiation: the B cell activation molecule, OX40 ligand, in-structs CD4 T cells to express interleukin 4 and upregulates expression of thechemokine receptor, Blr-1. J. Exp. Med. 188:297.
6. Gramaglia, I., A. D. Weinberg, M. Lemon, and M. Croft. 1998. Ox-40 ligand: apotent costimulatory molecule for sustaining primary CD4 T cell responses.J. Immunol. 161:6510.
7. Saoulli, K., S. Y. Lee, J. L. Cannons, W. C. Yeh, A. Santana, M. D. Goldstein,N. Bangia, M. A. DeBenedette, T. W. Mak, Y. Choi, and T. H. Watts. 1998.CD28-independent, TRAF2-dependent costimulation of resting T cells by 4-1BBligand. J. Exp. Med. 187:1849.
8. Shuford, W. W., K. Klussman, D. D. Tritchler, D. T. Loo, J. Chalupny,A. W. Siadak, T. J. Brown, J. Emswiler, H. Raecho, C. P. Larsen, et al. 1997.4-1BB costimulatory signals preferentially induce CD8� T cell proliferation andlead to the amplification in vivo of cytotoxic T cell responses. J. Exp. Med.186:47.
9. Hutloff, A., A. M. Dittrich, K. C. Beier, B. Eljaschewitsch, R. Kraft,I. Anagnostopoulos, and R. A. Kroczek. 1999. ICOS is an inducible T-cell co-stimulator structurally and functionally related to CD28. Nature 397:263.
10. Yoshinaga, S. K., J. S. Whoriskey, S. D. Khare, U. Sarmiento, J. Guo, T. Horan,G. Shih, M. Zhang, M. A. Coccia, T. Kohno, et al. 1999. T-cell co-stimulationthrough B7RP-1 and ICOS. Nature 402:827.
11. Mosmann, T. R., and R. L. Coffman. 1989. TH1 and TH2 cells: different patternsof lymphokine secretion lead to different functional properties. Annu. Rev. Im-munol. 7:145.
12. Romagnani, S. 1994. Lymphokine production by human T cells in disease states.Annu. Rev. Immunol. 12:227.
13. Mosmann, T. R. 1991. Role of a new cytokine, interleukin-10, in the cross-regulation of T helper cells. Ann. NY Acad. Sci. 628:337.
14. Sher, A., and R. L. Coffman. 1992. Regulation of immunity to parasites by T cellsand T cell derived cytokines. Annu. Rev. Immunol. 10:385.
15. Lichtman, A. H., J. Chin, J. A. Schmidt, and A. K. Abbas. 1988. Role of inter-leukin 1 in the activation of T lymphocytes. Proc. Natl. Acad. Sci. USA 85:9699.
16. Kuchroo, V. K., M. P. Das, J. A. Brown, A. M. Ranger, S. S. Zamvil, R. A. Sobel,H. L. Weiner, N. Nabavi, and L. H. Glimcher. 1995. B7-1 and B7-2 costimulatorymolecules activate differentially the Th1/Th2 developmental pathways: applica-tion to autoimmune disease therapy. Cell 10:707.
17. Bluestone, J. A. 1995. New perspectives of CD28–B7-mediated T cell costimu-lation. Immunity 2:555.
18. Abbas, A. K., K. M. Murphy, and A. Sher. 1996. Functional diversity of helperT lymphocytes. Nature 383:787.
19. Freeman, G. J., V. A. Boussiotis, A. Anumanthan, G. M. Bernstein, X. Y. Ke,P. D. Rennert, G. S. Gray, J. G. Gribben, and L. M. Nadler. 1995. B7-1 and B7-2do not deliver identical costimulatory signals, since B7-2 but not B7-1 preferen-tially costimulates the initial production of IL-4. Immunity 2:523.
20. Ranger, A. M., M. P. Das, V. K. Kuchroo, and L. H. Glimcher. 1996. B7-2(CD86) is essential for the development of IL-4-producing T cells. Int. Immunol.8:1549.
21. Vijayakrishnan, L., K. Natarajan, V. Manivel, S. Raisuddin, and K. V. Rao. 2000.B cell responses to a peptide epitope. IX. The kinetics of antigen binding differ-entially regulates costimulatory capacity of activated B cells. J. Immunol. 164:5605.
22. Tseng, S. Y., M. Otsuji, K. Gorski, X. Huang, J. E. Slansky, S. I. Pai, A. Shalabi,T. Shin, D. M.Pardoll, and H. Tsuchiya. 2001. B7-DC, a new dendritic cellmolecule with potent costimulatory properties for T cells. J. Exp. Med. 193:839.
23. Agrewala, J. N., D. S. Vinay, A. Joshi, and G. C. Mishra. 1994. A 150-kDamolecule of macrophage membrane stimulates interleukin-2 and interferon-� pro-duction and proliferation of ovalbumin-specific CD4� T cells. Eur. J. Immunol.24:2092.
FIGURE 5. Induction of T-bet, a Th1-specific transcription factor. TotalRNA was prepared from CD4� T cells activated in presence of anti-CD3(1 �g/ml) and with CHO-LAMP-1-Fc (2 �g/ml), or with macrophage-LAMP-1-Fc for 24 h and analyzed for the messages for c-Maf and T-bet byRT-PCR. Lanes 1 and 2, Equal loading controls which are the RT-PCRproduct for dihydrofolatereductase (DHFR) from the RNA prepared fromT cells activated with CHO-LAMP-1-Fc protein and M�-LAMP-1-Fc fu-sion proteins, respectively. Lanes 3 and 4, The RT-PCR products of c-Mafand T-bet mRNA, respectively, from T cells activated with CHO-LAMP-1-Fc. Lane 5 and 6, The RT-PCR product of c-Maf and T-bet mRNAderived from M�-LAMP-1-Fc-activated T cells, respectively.
1808 GLYCOSYLATION MODULATES THE COSTIMULATORY PROPERTY OF LAMP-1
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

24. Agrewala, J. N., S. Suvas, R. K. Verma, and G. C. Mishra.1998. Differentialeffect of anti-B7-1 and anti-M150 antibodies in restricting the delivery of co-stimulatory signals from B cells and macrophages. J. Immunol. 160:1067.
25. Das, G., H. Vohra, B. Saha, J. N. Agrewala, and G. C. Mishra. 1998. Leishmaniadonovani infection of a susceptible host results in apoptosis of Th1-like cells:rescue of anti-leishmanial CMI by providing Th1-specific bystander costimula-tion. Microbiol. Immunol. 42:795.
26. Das, G., H. Vohra, B. Saha, and G. C. Mishra. 2000. Th1-specific bystandercostimulation imparts resistance against Mycobacterium tuberculosis infection.Scand. J. Immunol. 52:515.
27. Stoppelli, M. P., C. Tacchetti, M. V. Cubellis, A. Corti, V. J. Hearing, G. Cassani,E. Appella, and F. Blasi. 1986. Autocrine saturation of pro-urokinase receptors onhuman A431 cells. Cell 45:675.
28. Vinay, D. S., M. Raje, R. K. Verma, and G. C. Mishra. 1995. Characterization ofnovel costimulatory molecules: a protein of 38–42 kDa from B cell surface isconcerned with T cell activation and differentiation. J. Biol. Chem. 270:23429.
29. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227:680.
30. White, M. B., M. Carvalho, D. Derse, S. J. O’Brien, and M. Dean. 1992. De-tecting single base substitutions as heteroduplex polymorphisms. Genomics 12:301.
31. Ouyang, W., M. Lohning, Z. Gao, M. Assenmacher, S. Ranganath, A. Radbruch,and K. M. Murphy. 2000. Stat6-independent GATA-3 autoactivation directs IL-4-independent Th2 development and commitment. Immunity 12:27.
32. Chen, J. W., Y. Cha, K. U. Yuksel, R. W. Gracy, and J. T. August. 1988. Isolationand sequencing of a cDNA clone encoding lysosomal membrane glycoprotein
mouse LAMP-1: sequence similarity to proteins bearing onco-differentiation an-tigens. J. Biol. Chem. 263:8754.
33. Heffernan, M., S. Yousefi, and J. W. Dennis. 1989. Molecular characterization ofP2B/LAMP-1, a major protein target of a metastasis-associated oligosaccharidestructure. Cancer Res. 49:6077.
34. Kannan, K., R. M. Stewart, W. Bounds, S. R. Carlsson, M. Fukuda,K. W. Betzing, and R. F. Holcombe. 1996. Lysosome-associated membrane pro-teins h-LAMP1 (CD107a) and h-LAMP2 (CD107b) are activation-dependent cellsurface glycoproteins in human peripheral blood mononuclear cells which me-diate cell adhesion to vascular endothelium. Cell Immunol. 171:10.
35. Chen, J. W., W. Pan, M. P. D’Souza, and J. T. August. 1985. Lysosome-asso-ciated membrane proteins: characterization of LAMP-1 of macrophage P388 andmouse embryo 3T3 cultured cells. Arch. Biochem. Biophys. 239:574.
36. Murphy, K. M., W. Ouyang, J. D. Farrar, J. Yang, S. Ranganath, H. Asnagli,M. Afkarian, and T. L.Murphy. 2000. Signaling and transcription in T helperdevelopment. Annu. Rev. Immunol. 18:451.
37. Ho, I. C., M. R. Hodge, J. W. Rooney, and L. H. Glimcher. 1996. The proto-oncogene c-Maf is responsible for tissue-specific expression of interleukin-4. Cell85:973.
38. Szabo, S. J., S. T. Kim, G. L. Costa, X. Zhang, C. G. Fathman, andL. H. Glimcher. 2000. A novel transcription factor, T-bet, directs Th1 lineagecommitment. Cell 100:655.
39. Sawada, R., S. Tsuboi, and M. Fukuda. 1994. Differential E-selectin-dependentadhesion efficiency in sublines of a human colon cancer exhibiting distinct met-astatic potentials. J. Biol. Chem. 269:1425.
40. Matzinger, P., and E. J. Fuchs. 1996. Beyond self and non-self: immunity is aconversation, not a war. J. NIH Res. 8:35.
1809The Journal of Immunology
by guest on September 10, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents











