THE TADPOLE OF PHYLLOMEDUSA ITACOLOMI (ANURA, HYLIDAE), WITH A DESCRIPTION OF THE INTERNAL ORAL MORPHOLOGY TIAGO LEITE PEZZUTI 1,3 , FELIPE SÁ FORTES LEITE 1 , AND FAUSTO NOMURA 2 1 Laboratório de Herpetologia, Departamento de Zoologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, 31270‑901, Belo Horizonte, MG, Brasil. E‑mail: [email protected], [email protected] 2 Laboratório de Ecologia Animal, Departamento de Zoologia e Botânica, Universidade Estadual Paulista, 15054‑000, São José do Rio Preto, SP, Brasil. E‑mail: [email protected] 3 Corresponding author. ABSTRACT. We describe the external morphology and the internal oral features of the tadpole of Phyllomedusa itacolomi (Anura, Hylidae) from Serra de Ouro Branco, Municipality of Ouro Branco, State of Minas Gerais, Brazil. Tadpoles were found at mid water in a temporary pool, in an open montane meadow. Phyllomedusa itacolomi (Gosner’s stage 37) is characterized by a tooth row formula 2(2)/3(1), labial tooth row P3 smaller than P1 and P2, marginal papillae of oral disc with wide dorsal and narrow ventral gaps. External morphological features were compared with other tadpoles of the Phyllomedusa hypochondrialis species group. Additionally, we provide information on egg clutches of P. itacolomi and P. megacephala. KEYWORDS. Anura; Phyllomedusinae; Phyllomedusa itacolomi; Taxonomy; Tadpoles; Southeastern Brazil. INTRODUCTION The neotropical frog genus Phyllomedusa Wagler, comprises 32 valid species (Frost, 2007; Giaretta et al., 2007) arranged in four species groups, with some species not currently assigned to any group (Faivovich, 2005). The Phyllomedusa hypochondria‑ lis species group (sensu Caramaschi, 2006) occurs in Cis-Andean South America in Venezuela, Guyanas, Peru, Ecuador, Brazil, Bolivia, Paraguay, and northern Argentina (Caramaschi, 2006) and is the most diver- sified one. Eleven species are included in the group: P. araguari, P. ayeaye, P. azurea, P. centralis, P. hypo‑ chondrialis, P. itacolomi, P. megacephala, P. nordes‑ tina, P. oreades, P. palliata, and P. rohdei. However, molecular data suggest that P. palliata is not related to any species group (Faivovich et al., 2005). Brandão and Álvares (2009) suggested that P. araguari and P. oreades could be synonymized based on morpho- metrics, coloration pattern, and call similarities. Herein, we describe the external morphology and internal oral features of the tadpole of Phyllomedusa itacolomi, from Serra de Ouro Branco, Municipality of Ouro Branco, State of Minas Gerais, Brazil. Infor- mation on egg clutches of P. itacolomi and P. mega‑ cephala is also provided. MATERIAL AND METHODS Phyllomedusa itacolomi tadpoles at stages 25-37 (Gosner, 1960) were collected from pools formed by temporary streams, surrounded by sparse vegetation and bushes, located in open montane meadows (“cam- pos rupestres”), at the Serra do Ouro Branco, Munici- pality of Ouro Branco, State of Minas Gerais, Brazil, on Feburary 07, 2006. Some tadpoles were reared until the end of metamorphosis to confirm species identity. Tad- poles were anesthetized with 0.1% lidocaine and then fixed in 10% formalin. Specimens used in the descrip- tion are deposited at the DZSJRP Collection, Depart- ment of Zoology and Botany, Universidade Estadual Paulista, São Paulo State, Brazil (Lot DZSJRP 1260.1). External morphology descriptions were based on four- teen tadpoles at stage 37. Measurements and terminol- ogy followed Altig and McDiarmid (1999). Measure- ments were taken with calipers (i.e., total, body and tail lengths) or under a stereomicroscope with an ocular micrometer (other measures) to the nearest 0.1 mm. We present quantitative data as mean (± SD; range; sample size). Three tadpoles at stages 37-38 were dis- sected and the internal oral structures were stained with 1% methylene blue solution for descriptions. All obser- vations were made under a Zeiss stereomicroscope at a magnification of 1.25X. Buccopharyngeal terminol- ogy follows Wassersug (1976). Tadpole coloration is described for life and in preserved specimens. Morpho- logical characteristics of tadpoles of the P. hypochon‑ drialis group (sensu Caramaschi, 2006) were obtained from Lutz and Lutz (1939), Lutz (1966), Pyburn and Glidewell (1971), Cei (1980), Cruz (1982), Lescure et al. (1995), Brandão (2002), Giaretta et al. (2007), and also from comparison with material deposited at the DZSJRP Collection, Department of Zoology and Botany, Universidade Estadual Paulista, São Paulo State, Brazil (P. megacephala Lot DZSJRP 1345.1). South American Journal of Herpetology, 4(2), 2009, 117-124 © 2009 Brazilian Society of Herpetology

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE TADPOLE OF PHYLLOMEDUSA ITACOLOMI (ANURA, HYLIDAE), WITH A DESCRIPTION OF THE INTERNAL ORAL MORPHOLOGY
Tiago LeiTe PezzuTi1,3, FeLiPe Sá ForTeS LeiTe1, and FauSTo nomura2
1 Laboratório de Herpetologia, Departamento de Zoologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, 31270‑901, Belo Horizonte, MG, Brasil. E‑mail: [email protected], [email protected]
2 Laboratório de Ecologia Animal, Departamento de Zoologia e Botânica, Universidade Estadual Paulista, 15054‑000,São José do Rio Preto, SP, Brasil. E‑mail: [email protected]
3 Corresponding author.
abSTracT. We describe the external morphology and the internal oral features of the tadpole of Phyllomedusa itacolomi (Anura, Hylidae) from Serra de Ouro Branco, Municipality of Ouro Branco, State of Minas Gerais, Brazil. Tadpoles were found at mid water in a temporary pool, in an open montane meadow. Phyllomedusa itacolomi (Gosner’s stage 37) is characterized by a tooth row formula 2(2)/3(1), labial tooth row P3 smaller than P1 and P2, marginal papillae of oral disc with wide dorsal and narrow ventral gaps. External morphological features were compared with other tadpoles of the Phyllomedusa hypochondrialis species group. Additionally, we provide information on egg clutches of P. itacolomi and P. megacephala.
KeywordS. Anura; Phyllomedusinae; Phyllomedusa itacolomi; Taxonomy; Tadpoles; Southeastern Brazil.
inTroducTion
The neotropical frog genus Phyllomedusa Wagler, comprises 32 valid species (Frost, 2007; Giaretta et al., 2007) arranged in four species groups, with some species not currently assigned to any group (Faivovich, 2005). The Phyllomedusa hypochondria‑lis species group (sensu Caramaschi, 2006) occurs in Cis-Andean South America in Venezuela, Guyanas, Peru, Ecuador, Brazil, Bolivia, Paraguay, and northern Argentina (Caramaschi, 2006) and is the most diver-sified one. Eleven species are included in the group: P. araguari, P. ayeaye, P. azurea, P. centralis, P. hypo‑chondrialis, P. itacolomi, P. megacephala, P. nordes‑tina, P. oreades, P. palliata, and P. rohdei. However, molecular data suggest that P. palliata is not related to any species group (Faivovich et al., 2005). Brandão and Álvares (2009) suggested that P. araguari and P. oreades could be synonymized based on morpho-metrics, coloration pattern, and call similarities.
Herein, we describe the external morphology and internal oral features of the tadpole of Phyllomedusa itacolomi, from Serra de Ouro Branco, Municipality of Ouro Branco, State of Minas Gerais, Brazil. Infor-mation on egg clutches of P. itacolomi and P. mega‑cephala is also provided.
maTeriaL and meThodS
Phyllomedusa itacolomi tadpoles at stages 25-37 (Gosner, 1960) were collected from pools formed by temporary streams, surrounded by sparse vegetation
and bushes, located in open montane meadows (“cam-pos rupestres”), at the Serra do Ouro Branco, Munici-pality of Ouro Branco, State of Minas Gerais, Brazil, on Feburary 07, 2006. Some tadpoles were reared until the end of metamorphosis to confirm species identity. Tad-poles were anesthetized with 0.1% lidocaine and then fixed in 10% formalin. Specimens used in the descrip-tion are deposited at the DZSJRP Collection, Depart-ment of Zoology and Botany, Universidade Estadual Paulista, São Paulo State, Brazil (Lot DZSJRP 1260.1). External morphology descriptions were based on four-teen tadpoles at stage 37. Measurements and terminol-ogy followed Altig and McDiarmid (1999). Measure-ments were taken with calipers (i.e., total, body and tail lengths) or under a stereomicroscope with an ocular micrometer (other measures) to the nearest 0.1 mm. We present quantitative data as mean (± SD; range; sample size). Three tadpoles at stages 37-38 were dis-sected and the internal oral structures were stained with 1% methylene blue solution for descriptions. All obser-vations were made under a Zeiss stereomicroscope at a magnification of 1.25X. Buccopharyngeal terminol-ogy follows Wassersug (1976). Tadpole coloration is described for life and in preserved specimens. Morpho-logical characteristics of tadpoles of the P. hypochon‑drialis group (sensu Caramaschi, 2006) were obtained from Lutz and Lutz (1939), Lutz (1966), Pyburn and Glidewell (1971), Cei (1980), Cruz (1982), Lescure et al. (1995), Brandão (2002), Giaretta et al. (2007), and also from comparison with material deposited at the DZSJRP Collection, Department of Zoology and Botany, Universidade Estadual Paulista, São Paulo State, Brazil (P. megacephala Lot DZSJRP 1345.1).
South American Journal of Herpetology, 4(2), 2009, 117-124© 2009 Brazilian Society of Herpetology

reSuLTS
Description of the tadpole of Phyllomedusa itacolomi
External morphology – Measurements for all avail-able developmental stages are given in Table 1. Body oval in dorsal and lateral views (Fig. 1A and 1B). To-tal length 2.5-3.0X body length; body 1.5-2.0X lon-ger than wide and 1.5-2.0X longer than high. Snout sloped in lateral view and rounded in dorsal view. Nostrils oval and positioned closer to tip of snout than to eyes. Eyes laterally positioned and directed; inter-orbital distance 0.8-0.9X of maximum body width and 3.6-4.0X of eye diameter. Oral disc anteroventral (Fig. 1C), its width about 0.4X the maximum body width; a single row of alternate marginal papillae on upper lip with a wide dorsal gap; two rows of mar-ginal papillae on lower lip and lateral portions, with a narrow ventral gap. A few scattered submarginal pa-pillae are present on the ventrolateral portions. Tooth
row formula (TRF) 2(2)/3(1); A1 and A2 of the same length; P3 smaller than P1 and P2. Jaw sheaths darkly pigmented and finely serrated on the margins; upper jaw sheath “M” shaped and lower sheath “V” shaped.
Spiracle ventral, sinistral, short and wide; directed posteriorly and opening on the middle third of the body; inner wall absent. Vent tube dextral, associated with the ventral fin, short and narrow, with opening directed ventrally. Tail length about 0.5-0.7X the to-tal length; tail musculature well developed; dorsal fin emerging on the posterior third of the body, originat-ing anteriorly to the body-tail junction, at a median slope; ventral fin beginning anterior to the vent tube; ventral fin depth approximately 1.2-1.5X the depth of the dorsal fin.
Color in life – Body and tail grayish-yellow with darker gray dots scattered on dorsal and lateral sur-faces of body and on tail musculature; in lateral view
Figure 1. Tadpole of Phyllomedusa itacolomi (DZSJRP 1260.1-A) at stage 37 of Gosner (1960): (A) Dorsal view; (B) lateral view (scale = 10 mm); (C) oral disc (scale = 1 mm).
118 The tadpole of Phyllomedusa itacolomi

the dorsal portion of peritoneum dark bluish; venter silver; dorsal and ventral fins transparent with white and black spots scattered mainly on the ventral fin; iris golden. In metamorphosing specimens, the dorsal surface of body is green and the surfaces of flanks and limbs presents a pattern of yellow markings encircled by deep purple coloration. This pattern of coloration is the same as that of the adults.
Color of fixed specimens – Color pattern in preserva-tive is the same as in life, but faded (e.g., black and white spots of ventral fins less defined). Abdomen becomes more transparent. The coloration of the iris turns black.
Internal oral features – Buccal floor pentagonal (Fig. 2A), as wide as long; oral aperture about 15% of buccal floor width; two large conical infralabial papillae. Two large lingual papillae, which may pres-ent postulated apices. Buccal floor arena pentagonal with seven to fifteen conical papillae on each side; numerous pustulations randomly distributed within buccal floor arena; 20 to 29 pustulations and two to seven conical papillae, smaller than buccal floor are-na papillae, are found anterior to the buccal pocket region; buccal pocket unperforated, wider than long, transversely oriented on the middle region of the buc-cal floor; free velar surface, with distinct glandular zone, with pustulated margins and pustulated on the region anterior to the glottis, which is notched, dis-tinct, open, and fully exposed.
Buccal roof triangular (Fig. 2B), wide posteriorly; prenarial arena with two broad pustulations anteri-orly and two small ones posteriorly. The anterior and posterior pustulations can appear together forming a semicircular ridge. Nares elliptical, parallel oriented in relation to the buccal opening and positioned on the anterior third of the buccal roof; internarial dis-tance smaller than nare opening; narial valve distinct with anterior and posterior wall pustulate and flap-like; post narial arena rectangular with four papillae with pustulate apices, the anteromedial pair is conical and taller than the posterolateral pair and the postero-lateral ones are broader than the anteromedial pair; four pustulations, one pair on each side, located be-tween the anteromedial and posterolateral papillae; one small and broad papillae located anterior to the median ridge, surrounded by three to six pustulations; median ridge triangular, longer than wide, with pus-tulated margins. Buccal roof arena diamond-shaped with seven to eight acute papillae on each side; Ta
bLe
1. M
orph
omet
ric d
ata
[mea
n ±
stan
dard
dev
iatio
n (r
ange
)] in
mm
for P
hyllo
med
usa
itaco
lom
i tad
pole
s fro
m S
erra
do
Our
o B
ranc
o, O
uro
Bra
nco,
Min
as G
erai
s sta
te, s
outh
east
ern
Bra
zil,
for d
evel
opm
enta
l sta
ges 2
5, 2
6, 2
7, 2
8, 2
9, 3
5 an
d 37
(Gos
ner 1
960)
. BL
= bo
dy le
ngth
, TL
= to
tal l
engt
h, T
AL
= ta
il le
ngth
, MTH
= m
axim
um ta
il he
ight
, DF
= m
axim
um h
eigh
t of d
orsa
l tai
l fin
, VF
= m
axim
um h
eigh
t of v
entra
l tai
l fin
, SN
= sn
out-n
aria
l dis
tanc
e, N
P =
naro
pupi
llar d
ista
nce,
IND
= in
tern
aria
l dis
tanc
e, IO
D =
inte
rorb
ital d
ista
nce,
ED
= m
axim
um d
iam
eter
of e
ye,
BW
= m
axim
um b
ody
wid
th, B
H =
max
imum
bod
y he
ight
, OW
= o
ral d
isc
wid
th.
Stag
eN
BL
TLTA
LM
THD
FV
FSN
NP
IND
IOD
EDB
WB
HO
W
254
8.5
± 0.
5 (7
.9-9
.3)
25.1
± 1
.0
(23.
5-26
.1)
16.5
± 0
.5
(15.
6-17
.0)
7.6
± 0.
4 (7
.0-8
.0)
1.7
± 0.
1 (1
.6-1
.9)
2.6
± 0.
1 (2
.5-2
.7)
1.3
± 0.
1 (1
.1-1
.4)
2.7
± 0.
2 (2
.5-3
.1)
2.5
± 0.
1 (2
.4-2
.6)
5.0
± 0.
1 (4
.8-5
.3)
1.5
± 0.
1 (1
.4-1
.6)
5.4
± 0.
2 (5
.3-5
.8)
5.4
± 0.
4 (5
.0-5
.8)
2.9
± 0.
2 (2
,6-3
.2)
269
9.7
± 0.
6 (8
.9-1
0.6)
28.4
± 1
.4
(26.
3-30
.7)
18.7
± 0
.9
(17.
4-20
.4)
7.6
± 0.
4 (7
.0-8
.0)
2.1
± 0.
4 (1
.7-3
.1)
2.9
± 0.
4 (2
.1-3
.2)
1.8
± 0.
2 (1
.5-2
.1)
3.1
± 0.
2 (2
.8-3
.4)
2.8
± 0.
2 (2
.4-3
.1)
5.6
± 0.
3 (5
.3-6
.1)
1.6
± 0.
1 (1
.5-1
.7)
6.0
± 0.
4 (5
.3-6
.5)
5.7
± 0.
4 (4
.9-6
.2)
2.9
± 0.
2 (2
,6-3
.2)
274
10.6
± 0
.4
(10.
1-11
.0)
31.1
± 1
.2
(29.
8-32
.8)
20.5
± 1
.1
(19.
7-22
.2)
8.7
± 0.
3 (8
.3-9
.3)
2.3
± 0.
2 (2
.1-2
.5)
3.3
± 0.
5 (3
.6-3
.5)
1.6
± 0.
1 (1
.5-1
.6)
3.4
± 0.
1 (3
.3-3
.6)
3.0
± 0.
2 (2
.8-3
.2)
6.1
± 0.
3 (5
.7-6
.4)
1.7
± 0.
1 (1
.6-1
.7)
6.8
± 0.
3 (6
.5-7
.0)
7.2
± 1,
0 (5
.9-8
.5)
2.9
± 0.
2 (2
.6-3
.2)
284
11.3
± 0
.3
(11.
0-11
.7)
32 ±
1.4
(3
0.8-
33.8
)20
.7 ±
1.5
(1
9.3-
22.7
)8.
7 ±
0.2
(8.5
-8.9
)2.
2 ±
0.1
(2.1
-2.2
)3.
4 ±
0.1
(3.2
-3.5
)1.
8 ±
0.3
(1.4
-2.0
)3.
5 ±
0.2
(3.2
-3.7
)3.
2 ±
0.1
(3.3
-3.1
)6.
5 ±
0.2
(6.4
-6.9
)1.
7 ±
0.2
(1.5
-1.9
)7.
0 ±
0.5
(6.6
-7.8
)8.
5 ±
2.0
(6.5
-11.
3)3.
0 ±
0.2
(2.8
-3.2
)
293
12,1
± 0
,4
(11.
7-12
.4)
33.6
± 1
.1
(32.
1-34
.5)
21.5
± 1
.5
(19.
8-22
.8)
9.1
± 0.
6 (8
.7-9
.7)
2.2
± 0.
2 (2
.0-2
.4)
3.5
± 0.
1 (3
.4-3
.6)
1.9
± 0.
1 (1
.8-2
.0)
3.5
± 0.
3 (3
.1-3
.8)
3.3
± 0.
1 (3
.3-3
.2)
7.0
± 0.
3 (6
.7-7
.3)
1.8
± 0.
0 (1
.8-1
.8)
7.8
± 0.
6 (7
.3-8
.4)
6.8
± 0.
7 (6
.0-7
.2)
3.0
± 0.
1 (2
.9-3
.1)
357
14.5
± 0
.7
(13.
2-15
.3)
40.8
± 1
.3
(39.
1-42
.6)
26.6
± 1
.2
(22.
5-31
.2)
9.9
± 0.
7 (8
.7-1
0.6)
2.9
± 0.
2 (2
.6-3
.1)
3.9
± 0.
3 (3
.5-4
.1)
2.1
± 0.
1 (1
.9-2
.3)
4.2
± 0.
4 (3
.6-4
.4)
3.6
± 0.
1 (3
.4-3
.7)
7.6
± 0.
2 (7
.4-7
.8)
2.0
± 0.
1 (2
.0-2
.1)
8.8
± 0.
4 (8
.3-9
.5)
8.4
± 0.
5 (7
.8-9
.4)
3.4
± 0.
2 (3
.2-3
.7)
3714
16.5
± 0
.7
(14.
3-17
.5)
44.3
± 2
.0
(46.
8-39
.4)
28.6
± 2
.1
(22.
5-31
.2)
10.5
± 0
.4
(10.
0-11
.2)
3.1
± 0.
2 (2
.7-3
.4)
4.1
± 0.
2 (3
.6-4
.4)
2.1
± 0.
3 (1
.7-2
.6)
4.5
± 0.
2 (4
.0-4
.7)
3.8
± 0.
2 (3
.5-4
.1)
8.1
± 0.
2 (7
.7-8
.7)
2.1
± 0.
1 (2
.1-2
.2)
9.5
± 0.
8 (8
.4-1
0.9)
9.2
± 0.
6 (8
.2-1
0.2)
3.7
± 0.
1 (3
.5-4
.0)
119Pezzuti, T. L. et al.
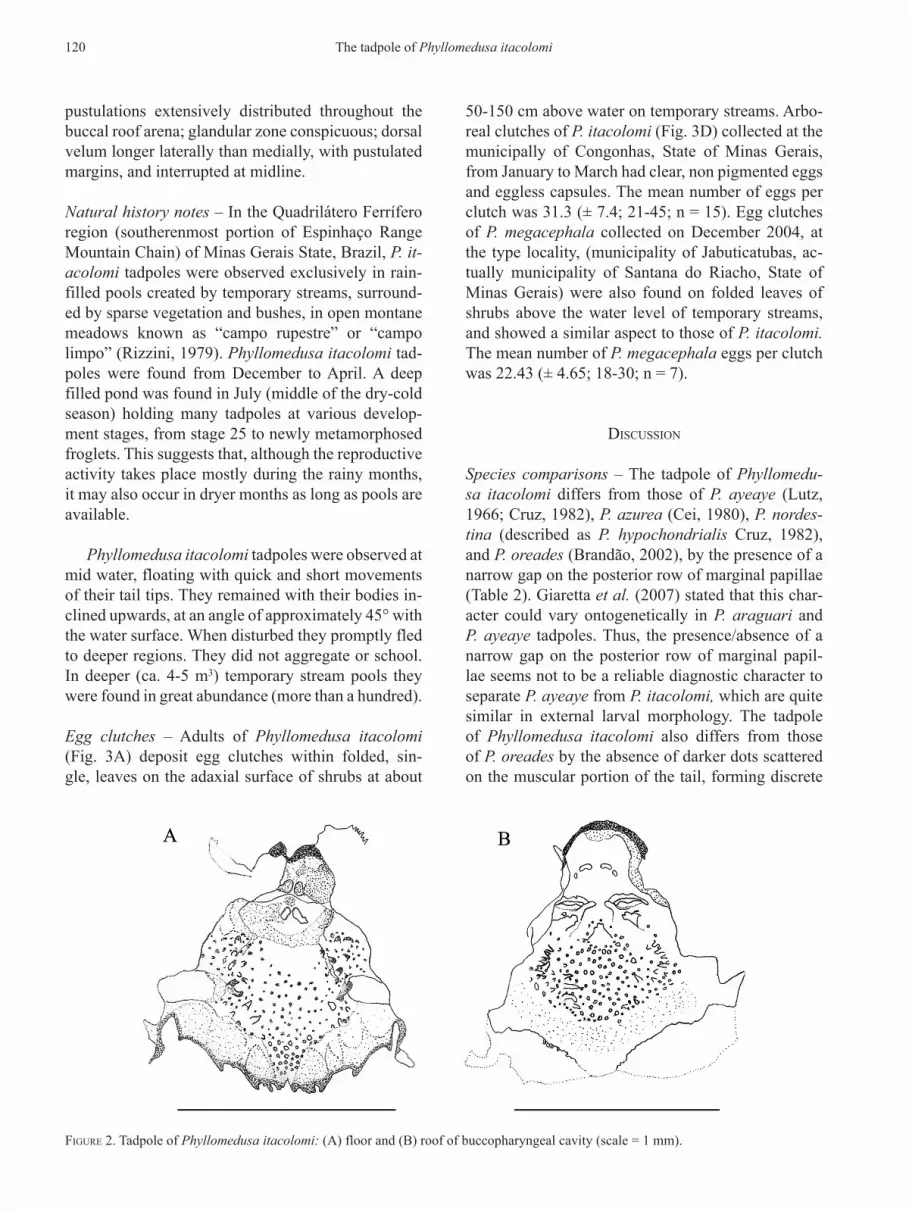
pustulations extensively distributed throughout the buccal roof arena; glandular zone conspicuous; dorsal velum longer laterally than medially, with pustulated margins, and interrupted at midline.
Natural history notes – In the Quadrilátero Ferrífero region (southerenmost portion of Espinhaço Range Mountain Chain) of Minas Gerais State, Brazil, P. it‑acolomi tadpoles were observed exclusively in rain-filled pools created by temporary streams, surround-ed by sparse vegetation and bushes, in open montane meadows known as “campo rupestre” or “campo limpo” (Rizzini, 1979). Phyllomedusa itacolomi tad-poles were found from December to April. A deep filled pond was found in July (middle of the dry-cold season) holding many tadpoles at various develop-ment stages, from stage 25 to newly metamorphosed froglets. This suggests that, although the reproductive activity takes place mostly during the rainy months, it may also occur in dryer months as long as pools are available.
Phyllomedusa itacolomi tadpoles were observed at mid water, floating with quick and short movements of their tail tips. They remained with their bodies in-clined upwards, at an angle of approximately 45° with the water surface. When disturbed they promptly fled to deeper regions. They did not aggregate or school. In deeper (ca. 4-5 m3) temporary stream pools they were found in great abundance (more than a hundred).
Egg clutches – Adults of Phyllomedusa itacolomi (Fig. 3A) deposit egg clutches within folded, sin-gle, leaves on the adaxial surface of shrubs at about
50-150 cm above water on temporary streams. Arbo-real clutches of P. itacolomi (Fig. 3D) collected at the municipally of Congonhas, State of Minas Gerais, from January to March had clear, non pigmented eggs and eggless capsules. The mean number of eggs per clutch was 31.3 (± 7.4; 21-45; n = 15). Egg clutches of P. megacephala collected on December 2004, at the type locality, (municipality of Jabuticatubas, ac-tually municipality of Santana do Riacho, State of Minas Gerais) were also found on folded leaves of shrubs above the water level of temporary streams, and showed a similar aspect to those of P. itacolomi. The mean number of P. megacephala eggs per clutch was 22.43 (± 4.65; 18-30; n = 7).
diScuSSion
Species comparisons – The tadpole of Phyllomedu‑sa itacolomi differs from those of P. ayeaye (Lutz, 1966; Cruz, 1982), P. azurea (Cei, 1980), P. nordes‑tina (described as P. hypochondrialis Cruz, 1982), and P. oreades (Brandão, 2002), by the presence of a narrow gap on the posterior row of marginal papillae (Table 2). Giaretta et al. (2007) stated that this char-acter could vary ontogenetically in P. araguari and P. ayeaye tadpoles. Thus, the presence/absence of a narrow gap on the posterior row of marginal papil-lae seems not to be a reliable diagnostic character to separate P. ayeaye from P. itacolomi, which are quite similar in external larval morphology. The tadpole of Phyllomedusa itacolomi also differs from those of P. oreades by the absence of darker dots scattered on the muscular portion of the tail, forming discrete
Figure 2. Tadpole of Phyllomedusa itacolomi: (A) floor and (B) roof of buccopharyngeal cavity (scale = 1 mm).
120 The tadpole of Phyllomedusa itacolomi

transverse bars. It differs from those of P. azurea, P. nordestina, and P. rodhei by the shape of the upper jaw sheath (“arc-shaped” in P. azurea, P. nordestina, and P. rodhei and “M-shaped” in P. itacolomi) and
from those of P. azurea by the TRF (Table 2). The tadpole of P. itacolomi differs from those of P. rohdei and from those of P. araguari by the origin of the dorsal fin, which originates at the body-tail junction
Figure 3. (A) Phyllomedusa itacolomi in life, male adult specimen photographed in calling activity (unvouchered photo); (B) froglet of P. itacolomi (unvouchered photo); (C) reproductive habitat of P. itacolomi; (D) open clutch of P. itacolomi (unvouchered photo). All photos are from the Municipality of Congonhas, State of Minas Gerais, Brazil.
TabLe 2. Morphological characters and habitat occurrence of the tadpoles of Phyllomedusa hypochondrialis species group.
TL mm (gosner’s
stage)Tooth row formula
Gap on the posterior row of marginal
papillae
Shape of the upper jaw sheath Occurrence (Temporary stream or pound)
P. araguari 45,7(33) 2(2)/3(1) Absent/presentc Md Temporary stream/PoundP. ayeaye 33,0(31) 2(2)/3a; 2(2)/3(1)b Absent/presentc M Temporary stream/PoundP. azurea — 2(2)/2(1) Absent Arch PoundP. hypochondrialis — 2(2)/3(1) Present M PoundP. itacolomi 44,3(37) 2(2)/3(1) Present M Temporary streamP. megacephala 36,0(30) 2(2)/3(1) Present M Temporary streamP. nordestina 46,0(37) 2(2)/3(1) Absent Arch PoundP. oreades 46,3(37) 2(2)/3e; 2(2)/3(1)f Absent M Temporary streamP. rohdei 43,0(36) 2(2)/3(1) Present Arch Pound
a = TRF presented by Lutz (1966); b = TRF presented by Cruz (1982); c = Ontogenetic variation (Giaretta et al., 2007); d = Giaretta et al. (2007) presented the upper jaw sheath as arc-shaped with a medial point; e = Brandão (2002); f = R. A. Brandão and G. Álvares (pers. comm.).
121Pezzuti, T. L. et al.

in P. rohdei and in P. araguari and anteriorly to the body-tail junction in P. itacolomi (Lutz and Lutz, 1939; Cruz, 1982; Giaretta et al., 2007). It also dif-fers from those of P. azurea, P. nordestina, and P. ro‑hdei by the dorsal fin height (lower in P. azurea, P. nordestina, and P. rohdei and higher in P. itaco‑lomi). Phyllomedusa itacolomi and P. megacephala (as P. centralis, Cruz, 1982), are quite similar in larval external morphology characters, but differ by the origin of the dorsal fin, (at the body-tail junction in P. itacolomi, and posterior to the body-tail junc-tion in P. megacephala), and by the life coloration of the dorsal peritoneum (dark-blue in P. itacolomi and black in P. megacephala). The tadpole of P. itacolomi could not be distinguished from those of P. hypocon‑drialis. Pyburn and Glidewell (1971), and Lescure et al. (1995) described the oral disc and dorsal pattern of coloration of P. hypochondrialis tadpoles. These characteristics per se are not enough to distinguish the larvae of these two species.
Internal oral features – The description of the inter-nal oral morphology was available for only three oth-ers Phyllomedusa species: Phyllomedusa aff. rohdei (Dias, 2007), from Mogi das Cruzes, Brazil, Phyllo‑medusa azurea (treated as P. hypochondrialis), and Phyllomedusa sauvagii (Candioti, 2007), from For-mosa and Salta, Argentina, respectively. The oral in-ternal features of these tadpoles differ mainly in the distribution and quantity of papillae. For example, the buccal roof arena is delimited by papillae only in P. itacolomi and P. sauvagii, whereas in P. azurea and P. aff. rohdei the papillae are distributed only on the central region of the buccal roof arena (Candioti, 2007; Dias, 2007; this work). In P. sauvagii six papil-lae delimited the buccal roof arena (Candioti, 2007), whereas we found up to eight papillae in P. itacolomi. The pre-narial arena of the buccal roof in Phyllom‑edusa species has a medially interrupted ridge, which could be low (as in P. rohdei Dias, 2007) or well de-veloped (as in P. azurea and P. sauvagii Candioti, 2007). In P. itacolomi, this character is variable as an uninterrupted low semicircular ridge or consisting of four pustulations rather than a ridge. Also the median ridge of P. itacolomi is distinct from those of other phyllomedusines: in P. itacolomi, it is triangular-shaped, whereas for P. azurea and P. aff. rohdei it is semicircular (Candioti, 2007; Dias, 2007). It is also triangular in P. sauvagii (Candioti, 2007), however it is lower than that of P. itacolomi. On the buccal floor, only P. itacolomi has infralabial papillae with no pro-jections, whereas they are bifurcated in P. azurea and
are more segmented in P. aff. rohdei and P. sauvagii (Candioti, 2007; Dias, 2007). Two lingual papillae are present in phyllomedusine tadpoles, but they can be conical and tall (as in P. aff. rohdei), conical and short (P. azurea), bifurcated and tall (P. sauvagii), or pustulate and tall (P. itacolomi Candioti, 2007; Dias, 2007; this work). Phyllomedusa itacolomi also has the highest number of papillae on the buccal floor arena, up to 15 papillae on each side, when compared with other phyllomedusine species: two pairs of pa-pillae in P. azurea, six in P. sauvagii, and six to seven in P. aff. rohdei (Candioti, 2007; Dias, 2007).
Natural history – The tadpoles of P. itacolomi also differ from those of P. azurea, P. hypochondrialis, P. nordestina, and P. rohdei by their occurrence in temporary streams (tadpoles of P. azurea, P. hypo‑chondrialis, P. nordestina, and P. rohdei develop in lenthic ponds) (Table 2). Phyllomedusa itacolomi egg clutches differ from those of P. araguari and P. mega‑cephala by the larger number of eggs [31.3 (± 7.4; 21-45; n = 15) in P. itacolomi; 24 in P. araguari (Gi-aretta et al., 2007); 22.43 (± 4.65; 18-30; n = 7) in P. megacephala]. Otherwise, P. itacolomi egg clutch-es have less eggs than those of P. hypochondrialis and P. rohdei that have been reported to range between 44-110 and 44-80 eggs, respectively (Pyburn and Glidewell, 1971; Lutz and Lutz, 1939). Despite the limited available information, it is possible that the species in the P. hypochondrialis group which are characterized by a reticulated pattern on the hidden parts of the flanks and limbs, a characteristic distri-bution on mountain ranges, and reproduction occur-ring in streams and rivulets (i.e., P. ayeaye, P. ara‑guari, P. centralis, P. itacolomi, P. megacephala, and P. oreades) lay smaller clutches of eggs than lowland species with striped pattern on the hidden parts of the flanks and limbs (i.e., P. azurea, P. hypochondrialis, P. nordestina, P. palliata, and P. rohdei).
Caramaschi (2006) redefined the P. hypochon‑drialis species group based on osteology, adult and larval external characters. The larval characters com-mon for this group were: anterior non-umbelliform oral disc, surrounded by a row of marginal papil-, surrounded by a row of marginal papil-lae interrupted by a wide dorsal gap and sometimes by a narrow ventral gap; vent tube short, and fused with the ventral fin; TRF 2(2)/3(1), with P3 smaller than P1 and P2. However, P. azurea presents TRF 2(2)/2(1) (Cei 1980), which increases the variation of TRF in this species group. Lutz (1966) described the tadpoles of P. ayeaye with a TRF 2(2)/3, but Cruz
122 The tadpole of Phyllomedusa itacolomi

(1982), based on the same specimens, found a TRF 2(2)/3(1) (Table 2). Also, the tadpoles of P. oreades were described as having a TRF 2(2)/3 (Brandão, 2002), but topotypes have a distinct interruption on P1 (R. A. Brandão and G. Álvares, pers. comm.). These differences could represent ontogenic or inter-populational variation.
Adults of P. itacolomi and P. ayeaye are not easy to diagnose (Caramaschi et al., 2007) and their tad-poles show quite similar external larval morphology. Brandão and Álvares (2009) noted the importance of assessing population variation of described species to avoid describing such variation as new species. These authors recommend an integrative taxonomic study, including statistical comparisons of morphometry, bioacoustics, and molecular differentiation, to clarify the taxonomic status of similar species in the P. hypo‑chondrialis species group.
reSumo
No presente artigo são descritos a morfologia externa e as características orais internas do girino de Phyllomedusa itacolomi (Anura, Hylidae) da Serra de Ouro Branco, Município de Ouro Branco, Estado de Minas Gerais, Brasil. Os girinos foram encontrados à meia-água de uma poça de riacho temporário, em área de campo rupestre. Phyllomedusa itacolomi (Estágio 37 de Gosner) é caracterizada por apresentar fórmula oral 2(2)/3(1), fileira de dentículos P3 menor que P1 e P2, papilas marginais do disco oral com ampla interrupção dorsal e pequena interrupção ventral. As características morfológicas externas de P. itacolomi foram comparadas às dos girinos pertencentes ao grupo de espécies de Phyllomedusa hypochondrialis. Adicionalmente, são disponibilizadas informações sobre as desovas de P. itacolomi e P. megacephala.
acKnowLedgmenTS
We are grateful to field course “Instrumentação em Herpetologia” of the Programa de Pós Graduação em Zoologia de Vertebrados – PUC Minas and K. Kopp, M. Wachlevski, L. G. Afonso and P. C. Eterovick for field work; C. A. G. Cruz, and D. Baêta, for kindly providing us relevant bibliographic material; P. C. Eterovick, G. Álvares, R. C. Santos, R. O. de Sá, J. A. Langone, and one anonymous reviewer for the suggestions on the manuscript; IBAMA for collection permit (license 282/05-IBAMA/RAN, process 0215.011808/05-06); R. Ludicanti, from the Junkie Dogs Holding Company for oral disc tadpole drawing; J. R. Thompson for tadpole drawings; Centro de Pesquisas Del-Rey for field support. Internal oral morphology drawings by F. Nomura.
LiTeraTure ciTed
aLTig, r. and r. w. mcdiarmid. 1999. Body plan: development and morphology; pp. 24-51. In: R. W. McDiarmid and R. Altig (Eds.), Tadpoles: the Biology of Anuran Larvae. The University of Chicago Press, Chicago.
brandão, r. a. 2002. A New Species of Phyllomedusa Wagler, 1830 (Anura: Hylidae) from Central Brazil. Journal of Herpetology, 36:571-578.
brandão, r. a. and g. F. r. áLvareS. 2009. Remarks on “A new Phyllomedusa Wagler (Anura, Hylidae) with reticulated pattern on flanks from Southeastern Brazil”. Zootaxa, 2044:61-64.
candioTi, m. F. v. 2007. Anatomy of anuran tadpoles from lentic water bodies: systematic relevance and correlation with feeding habits. Zootaxa, 1600:1-175.
caramaSchi, u. 2006. Redefinição do grupo de Phyllomedusa hypochondrialis, com redescrição de P. megacephala (Miranda-Ribeiro, 1926), revalidação de P. azurea Cope, 1862 e descrição de uma nova espécie (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional, 64:159-179.
caramaSchi, u., c. a. g. cruz, and r. n. Feio. 2006. A new species of Phyllomedusa Wagler, 1830 from the State of Minas Gerais, Brazil (Amphibia, Anura, Hylidae). Boletim do Museu Nacional, 524:1-8.
cei, J. m. 1980. Amphibians of Argentina. Monitore Zoologico Italiano, 2:1-609.
cruz, c. a. g. 1982. Conceituação de grupos de espécies de Phyllomedusinae brasileiras com base em caracteres larvários (Amphibia, Anura, Hylidae). Arquivos da Universidade Federal Rural do Rio de Janeiro, 5:147-171.
diaS, n. y. n. 2008. Estrutura de taxocenoses de girinos de Mata Atlântica. MSc. Dissertation, Universidade Estadual Paulista, São José do rio Preto, SP, 130 pp.
Faivovich, J., c. F. b. haddad, P. c. a. garcia, d. r. FroST, J. a. camPbeLL, and w. c. wheeLer. 2005. Systematic review of the frog Family Hylidae, with special reference to Hylinae: phylogenetic analysis and taxonomic revision. Bulletin of the American Museum of Natural History, 294:1-240.
FroST, d. 2007. Amphibian Species of the World: an Online Reference. Version 5.0. American Museum of Natural History, New York, USA. Available at: http://research.amnh.org/herpetology/amphibia/index.php.
giareTTa, a. a., J. c. oLiveira-FiLho, and m. n. c. KoKubum. 2007. A new Phyllomedusa Wagler (Anura, Hylidae) with reticulated pattern on flanks from Southeastern Brazil. Zootaxa, 1614:31-41.
goSner, K. L. 1960. A simplified table for staging anuran embryo and larvae with notes on identification. Herpetologica, 16:183-190.
LeScure, J., v. marTy, c. marTy, F. STarace, m. auber-Thomay, and F. LeTeLLier. 1995. Contribution a l’etude des amphibiens de Guyane Francaise X. Les Phyllomedusa (Anura, Hylidae). Revue Française d’ Aquariologie, Herpétologie 22(1-2):35-50.
LuTz, b. 1966. Pithecopus ayeaye, a new Brazilian hylid with vertical pupils and grasping feet. Copeia, 2:236-240.
LuTz, a. and b. LuTz. 1939. Notes on the genus Phyllomedusa Wagler. Observations on small Phyllomedusae without vomerine teeth or conspicuous paratoids found in the region of Rio de Janeiro. Annais da Academia Brasileira de Sciencias, 11:219-250.
123Pezzuti, T. L. et al.

nomura, F., d. c. roSSa-FereS, and v. h. m. Prado. 2003. The tadpole of Physalaemus fuscomaculatus (Anura: Leptodactylidae), with a description of internal oral morphology. Zootaxa, 370:1-8.
Pyburn, w. F. and J. r. gLideweLL. 1971. Nests and breeding behavior of Phyllomedusa hypochondrialis in Colombia. Journal of Herpetology, 5(1-2):49-52.
rizzini, c. T. 1979. Tratado de Fitogeografia do Brasil. Hucitec and Editora da Universidade de São Paulo, São Paulo, 374 pp.
waSSerSug, r. J. 1976. Oral morphology of anuran larvae: terminology and general description. Occasional Papers of the Museum of Natural History University of Kansas, 48:1-23.
Submitted 23 October 2008 Accepted 28 July 2009
124 The tadpole of Phyllomedusa itacolomi
Related Documents











