Journal OfNeurochemistry Raven Press, Ltd., New York 0 I993 International Society for Neurochemistry The SV2 Protein of Synaptic Vesicles Is a Keratan Sulfate Proteoglycan Todd W. Scranton, Mineo Iwata, and Steven S. Carlson Department of Physiology and Biophysics, University of Washington, Seattle, Washington, U.S.A Abstract: We have determined that synaptic vesicles con- tain a vesicle-specific keratan sulfate integral membrane proteoglycan. This is a major proteoglycan in electric or- gan synaptic vesicles. It exists in two forms on sodium dodecyl sulfate-polyacrylamide gel electrophoresis, i.e., the L form, which migrates like a protein with an M, of 100,000. and the H form, with a lower mobility that mi- grates with an M, of -250,000. Both forms contain SV2, an epitope located on the cytoplasmic side of the vesicle membrane. In additionto electric organ, we have analyzed the SV2 proteoglycan in vesicle fractions from two other sources, electric fish brain and rat brain. Both the H and L forms of SV2 are present in these vesicles and all are kera- tan sulfate proteoglycans. Unlike previously studied syn- aptic vesicle proteins, this proteoglycancontains a marker specific for a single group of neurons. This marker is an antigenically unique keratan sulfate side chain that is spe- cific for the cells innervating the electric organ; it is not found on the synaptic vesicle keratan sulfate proteoglycan in other neurons of the electric fish brain. Key Words: Pro- teoglycan-Electric organ-Synaptic vesicle-Keratan sulfate-Electric fish brain-SV2 protein. J. Neurochem. 61, 29-44 (1993). Classical neurotransmitters like acetylcholine are secreted from the nerve terminal by exocytosis of small clear synaptic vesicles (Kandel and Schwartz, 1985). In a complete cycle of exocytosis, endocytosis, and reformation of the synaptic vesicle, several activi- ties must occur. The vesicles must fill with neurotrans- mitter, gather at the active zone, and dock with the plasma membrane. Upon an increase in calcium con- centration, the vesicles fuse with the plasma mem- brane, the vesicle membrane proteins are endocy- tosed, and then the vesicles reform from an endo- some. The proteins responsible for many of these functions are unknown. Much progress has been made in characterizingthe components of small clear synaptic vesicles. These ves- icles have been purified from both brain (Huttner et al., 1983;Hell et al., 1989)and electric organ (Carlson et al., 1978;Tashiro and Stadler, 1978). The phospho- lipids of purified electric organ vesicles have been characterized (Deutsch and Kelly, 1981). A number of synaptic vesicle integral membrane proteins have been identified (Carlson, 1989; Sudhof and Jahn, 1991). The cDNAs for several of these proteins have been cloned and sequenced (Buckley et al., 1987; Leube et al., 1987; Sudhof et al., 1987; Trimble et al., 1988; Linial et al., 1989; Archer et al., 1990; Knaus et al., 1990; Perin et al., 1990; Wendland et al., 1991). A proteoglycan has been identified as a specific component of cholinergic synaptic vesicles from elec- tric organ (Stadler and Dowe, 1982; Carlson and Kelly, 1983). This proteoglycan was identified by two different monoclonal antibodies (mAbs) (Tor 70 and anti-SVl) that bind to the same (SV1) epitope (Carl- son and Kelly, 1983; Kushner, 1984; Caroni et al., 1985; Carlson, 1989). This proteoglycan behaves like an integral membrane protein (Carlson and Kelly, 1983;Carlson et al., 1986),with the SVl epitope pres- ent in the lumen of the synaptic vesicle (Carlson and Kelly, 1983; Buckley and Kelly, 1985). The SVl epi- tope is also present on the external nerve terminal surface of the electric organ synapse; it is restricted to the junctional region (Buckley et al., 1983). This pro- teoglycan was thought to be either a keratan sulfate or a heparin/heparan sulfate proteoglycan (Stadler and Dowe, 1982; Carlson and Kelly, 1983). Recent stud- ies have identified this proteoglycan as the receptor Received July 9, 1992; revised manuscript received November 16, 1992; accepted November 18, 1992. Address correspondence and reprint requests to Dr. S. S. Carlson at Department of Physiology and Biophysics, SJ-40, University of Washington, Seattle, WA 98195, U S A . Abbreviations used: BSA, bovine serum albumin; DEAE, dieth- ylaminoethyl; DISA, diazotized iodosulfanilic acid; Ig, irnmuno- globulin; ISA, iodosulfanilic acid mAb, monoclonal antibody; MOPC 2 1, purified mouse IgGl; PBS, phosphate-buffered saline; SDS-PAGE, sodium dodecylsulfate-polyacrylamide gel electrop ho- resis; SVZpg, L and H forms of keratan sulfate proteoglycans; TX- 100, Triton X-100. 29

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal OfNeurochemistry Raven Press, Ltd., New York 0 I993 International Society for Neurochemistry
The SV2 Protein of Synaptic Vesicles Is a Keratan Sulfate Proteoglycan
Todd W. Scranton, Mineo Iwata, and Steven S. Carlson
Department of Physiology and Biophysics, University of Washington, Seattle, Washington, U.S.A
Abstract: We have determined that synaptic vesicles con- tain a vesicle-specific keratan sulfate integral membrane proteoglycan. This is a major proteoglycan in electric or- gan synaptic vesicles. It exists in two forms on sodium dodecyl sulfate-polyacrylamide gel electrophoresis, i.e., the L form, which migrates like a protein with an M, of 100,000. and the H form, with a lower mobility that mi- grates with an M, of -250,000. Both forms contain SV2, an epitope located on the cytoplasmic side of the vesicle membrane. In addition to electric organ, we have analyzed the SV2 proteoglycan in vesicle fractions from two other sources, electric fish brain and rat brain. Both the H and L forms of SV2 are present in these vesicles and all are kera- tan sulfate proteoglycans. Unlike previously studied syn- aptic vesicle proteins, this proteoglycan contains a marker specific for a single group of neurons. This marker is an antigenically unique keratan sulfate side chain that is spe- cific for the cells innervating the electric organ; it is not found on the synaptic vesicle keratan sulfate proteoglycan in other neurons of the electric fish brain. Key Words: Pro- teoglycan-Electric organ-Synaptic vesicle-Keratan sulfate-Electric fish brain-SV2 protein. J. Neurochem. 61, 29-44 (1993).
Classical neurotransmitters like acetylcholine are secreted from the nerve terminal by exocytosis of small clear synaptic vesicles (Kandel and Schwartz, 1985). In a complete cycle of exocytosis, endocytosis, and reformation of the synaptic vesicle, several activi- ties must occur. The vesicles must fill with neurotrans- mitter, gather at the active zone, and dock with the plasma membrane. Upon an increase in calcium con- centration, the vesicles fuse with the plasma mem- brane, the vesicle membrane proteins are endocy- tosed, and then the vesicles reform from an endo- some. The proteins responsible for many of these functions are unknown.
Much progress has been made in characterizing the components of small clear synaptic vesicles. These ves- icles have been purified from both brain (Huttner et al., 1983; Hell et al., 1989) and electric organ (Carlson
et al., 1978; Tashiro and Stadler, 1978). The phospho- lipids of purified electric organ vesicles have been characterized (Deutsch and Kelly, 198 1). A number of synaptic vesicle integral membrane proteins have been identified (Carlson, 1989; Sudhof and Jahn, 199 1). The cDNAs for several of these proteins have been cloned and sequenced (Buckley et al., 1987; Leube et al., 1987; Sudhof et al., 1987; Trimble et al., 1988; Linial et al., 1989; Archer et al., 1990; Knaus et al., 1990; Perin et al., 1990; Wendland et al., 1991).
A proteoglycan has been identified as a specific component of cholinergic synaptic vesicles from elec- tric organ (Stadler and Dowe, 1982; Carlson and Kelly, 1983). This proteoglycan was identified by two different monoclonal antibodies (mAbs) (Tor 70 and anti-SVl) that bind to the same (SV1) epitope (Carl- son and Kelly, 1983; Kushner, 1984; Caroni et al., 1985; Carlson, 1989). This proteoglycan behaves like an integral membrane protein (Carlson and Kelly, 1983; Carlson et al., 1986), with the SVl epitope pres- ent in the lumen of the synaptic vesicle (Carlson and Kelly, 1983; Buckley and Kelly, 1985). The SVl epi- tope is also present on the external nerve terminal surface of the electric organ synapse; it is restricted to the junctional region (Buckley et al., 1983). This pro- teoglycan was thought to be either a keratan sulfate or a heparin/heparan sulfate proteoglycan (Stadler and Dowe, 1982; Carlson and Kelly, 1983). Recent stud- ies have identified this proteoglycan as the receptor
Received July 9, 1992; revised manuscript received November 16, 1992; accepted November 18, 1992.
Address correspondence and reprint requests to Dr. S. S. Carlson at Department of Physiology and Biophysics, SJ-40, University of Washington, Seattle, WA 98195, U S A .
Abbreviations used: BSA, bovine serum albumin; DEAE, dieth- ylaminoethyl; DISA, diazotized iodosulfanilic acid; Ig, irnmuno- globulin; ISA, iodosulfanilic acid mAb, monoclonal antibody; MOPC 2 1 , purified mouse IgGl; PBS, phosphate-buffered saline; SDS-PAGE, sodium dodecylsulfate-polyacrylamide gel electrop ho- resis; SVZpg, L and H forms of keratan sulfate proteoglycans; TX- 100, Triton X-100.
29

30 T. W. SCRANTON ET AL.
for vesamicol, an inhibitor of acetylcholine transport in electric organ vesicles (Bahr et al., 1992).
A glycoprotein of 100 kDa has also been identified as a specific component of purified electric organ syn- aptic vesicles (Buckley and Kelly, 1985). This trans- membrane glycoprotein was found with an mAb to a cytoplasmic epitope, SV2. The SV2 antigen is present in the vesicles of neurons and endocrine cells, but is not detected in exocrine cells (Buckley and Kelly, 1985). This protein has recently been cloned by other investigators (Bajjalieh et al., 1992; Feany et al., 1992). It has homology to a family of bacterial trans- porters (Bajjalieh et al., 1992; Feany et al., 1992) as well as the human glucose transporter and the plasma membrane transporters for neurotransmitters (Feany et al., 1992). The SV2 epitope has also been linked to acetylcholine transport in electric organ synaptic vesi- cles (Bahr and Parsons, 1992).
In this article we show that two forms of the SV2 antigen exist in electric organ synaptic vesicles, i.e., the 100-kDa protein (L form) Buckley and Kelly ( 1985) described previously and a more heavily glyco- sylated protein (H form) of -250 kDa. The H form of the SV2 antigen is the synaptic vesicle proteoglycan identified previously, containing the SV 1 epitope (Carlson and Kelly, 1983). However, both forms are keratan sulfate proteoglycans that we call SV2pg. The SV I epitope is present on a keratan sulfate side chain, and is unique to the H form of electric organ SV2pg. The SV2pg contains the majority of the keratan sul- fate in the synaptic vesicle. We find that the proteogly- can is complexed noncovalently with four other pro- teins in the electric organ synaptic vesicle. In addition to electric organ, we have studied the SV2 antigen in fractions prepared from electric fish brain and rat brain. The SV2 antigen isolated from fish and rat brain also exists in two forms, L and H. In brain both forms contain keratan sulfate.
MATERIALS AND METHODS Elasmobranch electric rays Discopyge ommata were ob-
tained from Mannus Inc. (Long Beach, CA, U.S.A.) and Salt Water City (San Francisco, CA, U.S.A.). Immunobeads [rabbit anti-mouse immunoglobulin (Ig), heavy and light chain specific] and molecular weight standard markers for gel electrophoresis were purchased from Bio-Rad. Sepha- cry1 S-1000, Sephacryl S-500, and Sepharose CL-6B were purchased from Pharmacia LKB Biotechnology Inc. Nitro- cellulose (BA85) and diethylaminoethyl (DEAE)-nitrocel- lulose (NA45) membranes were purchased from Schleicher and Schuell; Triton X- 100 (TX- IOO), endo-P-galactosidase, and n-octylglucoside from Boehringer Mannheim; and x- ray film (X-Omat AR) from Kodak. Na'"I, 1251-labeled sheep anti-mouse Ig, and Rainbow standard protein markers for western blots were obtained from Amersham Corp. Glycosaminoglycan lyases were purchased from Sei- kagaku Kogyo Co., Ltd. (Tokyo, Japan). N-Glycanase was purchased from Genzyme (Boston, MA, U.S.A.); sulfanilic acid from Aldrich; and Iodo-Gen and 0-mercaptoethanol
from Pierce Chemical Co. Ultrapure urea and commercial glycosaminoglycans were purchased from ICN Biomedicals Inc. (Cleveland, OH, U.S.A.); pronase from CalBiochem. Wheat germ agglutinin beads, protease inhibitor peptides, fetuin, and all common chemicals were obtained from Sigma. Anti-keratan sulfate mAbs 1B4 and 2D3 were the kind gift of Dr. Bruce Caterson.
Immunocytochemical reagents were purchased from the following companies: biotinylated goat anti-mouse IgG (Fc fragment specific) from Jackson Immunoresearch Labs. (West Grove, PA, U.S.A.); Vectastain ABC kit from Vector Laboratories (Burlingame, CA, U.S.A.); purified mouse IgG1 (MOPC 21) from Sigma Chemical Co. (St. Louis. MO, U.S.A.).
Iodination of synaptic vesicle proteins Synaptic vesicles were purified from D. ommata electric
organ as described previously (Carlson et al., 1978). In brief, perfused electric organ was frozen in liquid nitrogen and ground to a fine powder. This material was then suspended in buffer and homogenized in a blender, and centrifuged at 1 1,000 g. The resulting supernatant was pelleted at 100,000 g, and resuspended in 0.8 M sucrose. The vesicles contained in this resuspended pellet were floated to their equilibrium density on a sucrose gradient. The vesicles were then com- pletely purified by chromatography on controlled pore glass and pelleted at 100,000 g.
[ '251]Iodosulfanilic acid ([ 1251]ISA) was prepared by the Iodo-Gen method as described previously for protein (Carl- son et al., 1986). An aliquot of 300 pCi of ['Z51]ISA was converted to '2SI-diazotized iodosulfanilic acid ([ "'I]DISA) as described previously (Carlson and Kelly. 1980). Synaptic vesicles (28 pg of protein) were solubilized in 0.4 M NaCI, 10 mM HEPES, pH 7.4, and 1% n-octylglucoside, and added to the DISA to a final volume of 110 pl and allowed to react overnight at 4°C. The unbound '''I and [1251]DISA were separated from the bound radioactivity by chromatog- raphy with Sephadex G-25 fine swollen in 0.4 MNaC1,O. I % bovine serum albumin (BSA), 10 mMHEPES, pH 7.4, and 1% TX-100. The final incorporation of 1251 averaged 5.3%, yielding material with a specific activity of 0.57 pCi/pg.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and nitrocellulose transfer procedures
SDS-PAGE was performed by the methods of Laemmli ( 1970). All SDS-PAGE was performed under reducing con- ditions. Gels were poured with a linear gradient of acryl- amide from 2.7 to 14% with a 2.7% stacking gel. Gels for iodinated material were poured on a backing of Gelbond (FMC Corp., Rockland, ME, U.S.A.). The western blotting procedures used were those of Burnette (1 98 1) with the sub- stitution of 5% nonfat milk for 5% BSA. Autoradiography of dried SDS gels and of western blots was performed and the results were quantified as described previously (Carlson et al., 1986). Multiple exposures at different times were taken to establish the linearity of the signals.
Immunoprecipitations All immunoprecipitations were performed with polyacryl-
amide beads to which mAbs were bound. The beads were prepared by binding our monoclonals to rabbit anti-mouse antibodies covalently attached to polyacrylamide (Bio-Rad) as described previously (Iwata and Carlson, 1991). The beads were suspended as a 10% slurry in 0.15 M NaC1, 10
J . Neurochem, Vul 61, No I , 1YY3

SV2 IS A KERATAN SULFATE PROTEOGLYCAN 31
I ~ M HEPES, pH 7.4, 0.1% BSA, and 1.5% TX-100 (im- munoprecipitation buffer), and stored at 4°C before use.
Some protein samples were treated under denaturing and reducing conditions before precipitation. SDS and 0-mer- captoethanol were added to these samples to final concen- trations of 2% each. These samples were boiled for 4 min and diluted 1 :200 with cold (4°C) immunoprecipitation buffer. Immunobeads were then added at a concentration of 15 p1 of beads per 500 p1 of sample. All precipitations of rodinated material were conducted using -0.25 pCi of '251- protein per precipitation. Samples for precipitations of un- labeled synaptic vesicle material contained I pg of purified vesicle protein per microliter of beads; samples for precipita- tion of fish and rat brain material used 12 pg of crude vesicle fraction per microliter of beads. The precipitations were in- cubated overnight at 4"C, then the beads pelleted and washed to remove BSA. Samples for SDS-PAGE were pre- pared by boiling the immunobead pellet in final sample buffer (7.5% glycerol, 2% SDS, 2% 0-mercaptoethanol, 25 mMTris, pH 6.8,O. 125% bromphenol blue; 1 ~1 per micro- liter of bead slurry) for 3 min. The samples were then centri- fuged to remove the polyacrylamide beads and applied to SDS-PAGE.
S-500 gel filtration Protein samples were pretreated by boiling in 2% SDS, 2%
&mercaptoethanol and applied to a 250-ml Sephacryl S-500 column equilibrated with 0.5% SDS, 0.5% P-mercap- toethanol, 25 mM Tris, pH 7.0. Rat brain samples con- tained 2.5 mg of protein from rat synaptosomes, and were centrifuged to remove a small amount of insoluble material. Fish synaptic vesicles contained 600 pg of purified synaptic vesicle protein. The samples were diluted 1:5 with column buffer before application and run at a flow rate of 25 ml per hour. The eluted material was collected in 2-ml fractions. Samples were vacuum dried and resuspended in 10% origi- nal volume 7.5% glycerol, 0.5% 0-mercaptoethanol, and 0.125% bromphenol blue, and then boiled 4 rnin and ap- plied to SDS-PAGE for western blotting. The column void volume was determined from the elution of sheared herring sperm DNA assayed by UV absorbance; the total volume was determined from the elution of phenol red measured by absorbance.
Protein assay
of Schaffer and Weissmann ( 1973).
Glycosaminoglycan lyase digestions Digestions were performed on material that was precipi-
tated from SDS/@-mercaptoethanol-treated synaptic vesi- cles with anti-SV2 as described. The resulting pellets were resuspended in buffer and digested with the glycosaminogly- can lyases keratanase I, keratanase 11, chondroitinase ABC, heparitinase, and endo-b-galactosidase under the condi- tions described in the technical information provided by Seikagaku Kogyo Co. Digestions were conducted for 4 h at 37°C. N-Glycanase digestions were also conducted on pre- cipitated material under the conditions found in the Gen- zyme technical information. The total volume of the diges- tion was 40 p1 in all cases. After digestion samples were prepared for SDS-PAGE as described above for immuno- precipitations.
N-Glycanase control digestions were conducted under identical conditions using 20 pg of fetuin as substrate. The
Protein concentrations were determined by the method
effectiveness of these controls was evaluated by Coomassie Blue staining of SDS-PAGE gels.
Preparation of rat brain material Rat synaptosomes for S-500 gel filtration were prepared
using a Ficoll-sucrose gradient according to methods de- scribed by Cotman (1974). Whole rat brain was homoge- nized in 0.32 M sucrose and centrifuged at 11,000 g to re- move large tissue fragments. The remaining supernatant was then loaded onto a Ficoll gradient with steps of 8.5, 13, and 17% in 0.32 A4 sucrose. These gradients were then cen- trifuged at 68,580 g for 45 rnin and synaptosomes were col- lected from the interface between the 8.5 and 13% Ficoll layers. Synaptosomes were stored at -20°C in the presence of 0.1 mg/ml of iodoacetamide, 0.0 1% diisopropyl fluoro- phosphate, and 0.1 pg/ml of each of the following protease inhibitor peptides: leupeptin, pepstatin, and chymostatin.
The crude vesicle fraction used in immunoprecipitations was a 1 1,000 g supernatant prepared from rat brains frozen and homogenized in liquid nitrogen. Its preparation is taken from the initial stages of the purification of synaptic vesicles from electric organ (Carlson et al., 1978). This mate- rial was then suspended (2 g of tissue/3 ml of buffer) in 0.4 A4 NaCl, 10 mM EGTA, 10 mM HEPES, pH 7.0, with 0.02% azide and protease inhibitors as described above and further homogenized on ice by five passes in a motor-driven Teflon-glass homogenizer. The resulting suspension was centrifuged for 30 rnin at 1 1,000 gin a Beckman SJ-40 rotor at 4°C. The pellet was discarded and the supernatant stored frozen at -20°C.
Preparation of fish brain material The fish brain crude vesicle fraction used in immunopre-
cipitation experiments was a I 1,000 gsupernatant prepared as indicated above for rat brain material.
Irnmunocytochemistry Fish brain was immersion fixed in 4% paraformaldehyde
overnight at 4"C, soaked for 2 days at 4°C in 20% sucrose for cryoprotection, quick-frozen in isopentane cooled with liquid nitrogen, cut into 8-10-pm sections with a cryostat, and mounted on chrome-alum gelatin-coated microscope slides. The slides were incubated for 1 h with 5 % goat serum in phosphate-buffered saline (PBS) (to prevent nonspecific staining of the sections by antibodies), overnight with mAb (monoclonal supernatant diluted 1:5 with 5% horse serum in PBS), 15 rnin with 0.75% hydrogen peroxide to inactivate endogenous peroxidases, 1.5 h with biotinylated secondary antibody (5 pg/ml in 5% horse serum in PBS). The sections were incubated with streptavidin-biotin horseradish peroxi- dase complex using the Vectastain ABC kit of Vector Labo- ratories according to the manufacturer's instructions. A so- lution of 0.3% hydrogen peroxide, 100 pg/ml of 3 3 - diaminobenzidine tetrahydrochloride in 50 mM Tris-HCI, pH 7.6, was used as a substrate for the immunolocalized peroxidase, resulting in a brown reaction product.
Preparation of glycosaminoglycan side chains from synaptic vesicles and Sepharose CL-6B chromatography
Purified synaptic vesicles were subjected to alkaline boro- hydride cleavage and pronase digestion. The released glycos- aminoglycans were analyzed by permeation chromatogra- phy on Sepharose CL-6B. Both methods are from Carlson
J. Neurochern.. Val. 61, No. 1. 1993
32 T. W. SCRANTON ET AL.
and Wight ( I 987). The procedure was as follows: Purified synaptic vesicles ( 1 ml of 0.57 mg of protein/ml) were eth- anol precipitated (Iwata and Carlson, 1991) and the result- ing pellet dissolved in I .7 ml of 0.06 M NaOH and 1.2 M sodium borohydride. After incubation at 45°C for 40 h, the cleavage reaction was stopped by neutralizing the solution with acetic acid. The released glycosaminoglycan side chains were separated from protein by 10% trichloroacetic acid precipitation of the protein from this solution (Iwata and Carlson, 199 1). The free glycosaminoglycan side chains were then ethanol precipitated from this solution and redis- solved in 2 ml of 25 W H E P E S , pH 7.4. The glycosamino- glycan concentration was determined by a dot-blot assay on DEAE-nitrocellulose (Carlson and Wight, 1987). We ob- tained 60 pg of glycosaminoglycan from 600 pg of vesicle protein. To ensure elimination of polypeptides, 15 pg of these glycosaminoglycans in 0.5 ml were digested with pronase at an enzyme concentration of 0.1 pg/pl in 25 mM HEPES, pH 7.4, at 37°C for 18 h, followed by second alka- line borohydrate cleavage. To perform the second cleavage, the glycosaminoglycan solution was made with 0.06 M NaOH and 1.2 Msodium borohydride, incubated as before, the reaction stopped with acetic acid, and the glycosamino- glycans were ethanol precipitated. The glycosaminoglycans were then solubilized in the buffer containing either 25 m M HEPES, pH 7.4,0.2 MNaCI, and 0.02% sodium azide, or 4 A4 guanidine, 50 mM HEPES, pH 7.0. After addition of I00 pg of heparin as a camer, the glycosaminoglycans were ap- plied to a column of Sepharose CL-6B (0.8 X 128 cm) and collected at 0.75 ml/fraction at the speed of 5 ml/h. The antigenicities of SVI, SV2, SV4, and 2D3 were detected by DEAE-nitrocellulose binding assay as described previously (Carlson and Wight, 1987).
RESULTS
Two forms of the SV2 antigen exist in synaptic vesicles, i.e., the 100-kDa protein (L form) and a higher molecular weight form (H form) that contains the SV1 epitope
When '251-labeled, SDS-solubilized, reduced synap- tic vesicle proteins are immunoprecipitated with either the anti-SV1 or anti-SV2 mAbs, the same 250- kDa protein is precipitated. We demonstrated this us- ing successive immunoprecipitations of the I2Tla- beled antigen with the anti-SV 1 and anti-SV2 mAbs. That is, the 1251-labeled antigen was immunoprecipi- tated with one mAb, eluted from the bead pellet with denaturing conditions, and reimmunoprecipitated with the same or different mAb. On SDS-PAGE, the '251-labeled protein twice immunoprecipitated by anti-SV1 (Fig. IA, lane I ) appears as a broad band of -250 kDa (H band). The 1251-labeled protein twice immunoprecipitated by anti-SV2 appears as an iden- tical broad band of -250 kDa (H band), and an addi- tional band is present at 100 kDa (Fig. IA, lane 3). This result suggests that the H band may contain both SVI and SV2 epitopes. To test this idea, we first im- munoprecipitated '251-labeled vesicle proteins with anti-SV1 under the same conditions as before, then reimmunoprecipitated the protein with anti-SV2. On SDS-PAGE, this double-immunoprecipitated protein
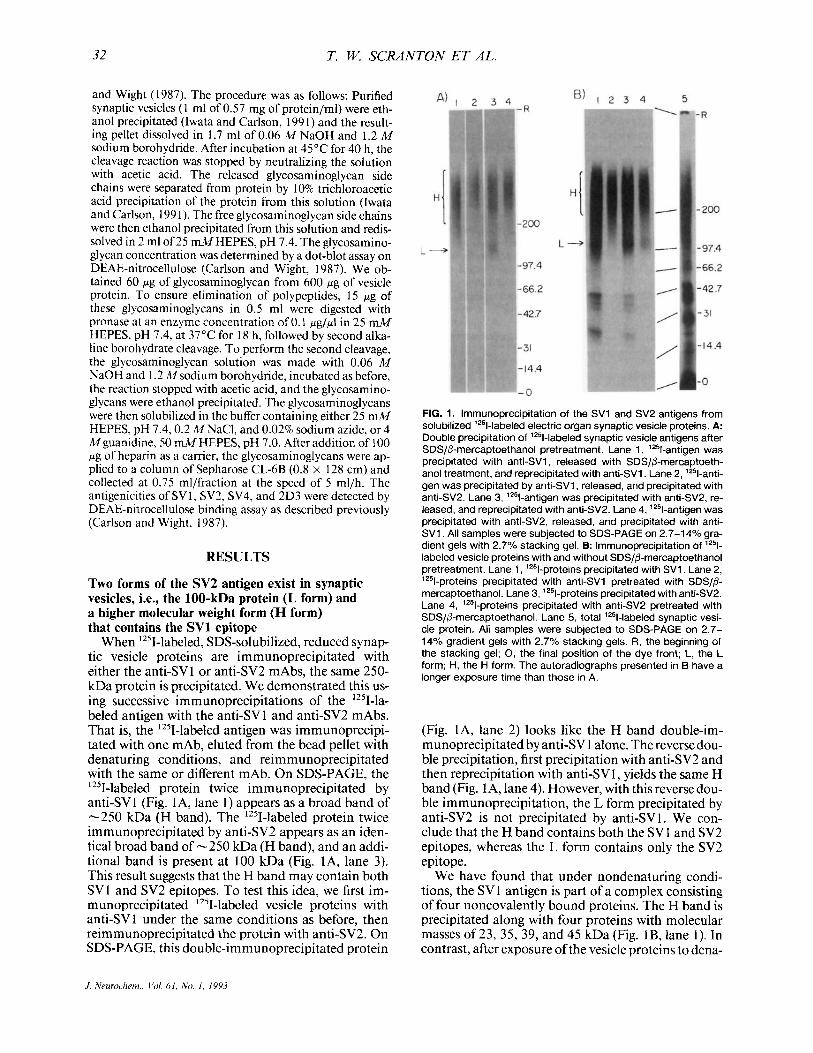
FIG. 1. lmmunoprecipitation of the SV1 and SV2 antigens from solubilized '251-labeled electric organ synaptic vesicle proteins. A: Double precipitation of '251-labeled synaptic vesicle antigens after SDSIP-mercaptoethanol pretreatment. Lane 1, '%antigen was precipitated with anti-SV1, released with SDS/P-mercaptoeth- anol treatment, and reprecipitated with anti-SV1. Lane 2, '%anti- gen was precipitated by anti-SV1, released, and precipitated with anti-SV2. Lane 3, '%antigen was precipitated with anti-SV2, re- leased, and reprecipitated with anti-SV2. Lane 4, '%antigen was precipitated with anti-SV2, released, and precipitated with anti- SV1. All samples were subjected to SDS-PAGE on 2.7-14% gra- dient gels with 2.7% stacking gel. B: lmmunoprecipitation of lZ5l- labeled vesicle proteins with and without SDS/@-mercaptoethanol pretreatment. Lane 1, 'z51-proteins precipitated with SVl . Lane 2, '251-pr~tein~ precipitated with anti-SV1 pretreated with SDS/@- mercaptoethanol. Lane 3, '251-pr~tein~ precipitated with anti-SV2. Lane 4, '251-proteins precipitated with anti-SV2 pretreated with SDS//3-mercaptoethanol. Lane 5, total '251-labeled synaptic vesi- cle protein. All samples were subjected to SDS-PAGE on 2.7- 14% gradient gels with 2.7% stacking gels. R, the beginning of the stacking gel; 0, the final position of the dye front; L, the L form; H, the H form. The autoradiographs presented in B have a longer exposure time than those in A.
(Fig. IA, lane 2) looks like the H band double-im- munoprecipitated by anti-SV 1 alone. The reverse dou- ble precipitation, first precipitation with anti-SV2 and then reprecipitation with anti-SV1, yields the same H band (Fig. 1 A, lane 4). However, with this reverse dou- ble immunoprecipitation, the L form precipitated by anti-SV2 is not precipitated by anti-SVI. We con- clude that the H band contains both the SVl and SV2 epitopes, whereas the L form contains only the SV2 epitope.
We have found that under nondenaturing condi- tions, the SV 1 antigen is part of a complex consisting of four noncovalently bound proteins. The H band is precipitated along with four proteins with molecular masses of 23, 35, 39, and 45 kDa (Fig. lB, lane 1). In contrast, after exposure of the vesicle proteins to dena-
J. Nwrorhem , b'ol 61. No. 1. 1993
SV2 IS A KERA TAN SULFATE PROTEOGLYCAN 33
turing and reducing conditions, only the H band is precipitated by SVl (Fig. lB, lane 2). Similarly, under nondenaturing conditions, immunoprecipitation of vesicle proteins by anti-SV2 reveals the same complex of small proteins as precipitated by anti-SV 1 (Fig. 1 B, compare lanes 3 and 1). Immunoprecipitation by anti-SV2 after denaturing reducing conditions yields the H and L band (Fig. IB, lane 4). We do not im- munoprecipitate all of the iodinated proteins of the purified synaptic vesicles (compare Fig. IB, lane 5) , which indicates a specific interaction between the SV2 protein and the four proteins of 23-45 kDa.
We confirmed the above results by immunoprecipi- tating the unlabeled SV2 antigen from SDS-dena- tured, reduced synaptic vesicles, then subjecting the precipitate to western blot analysis. By these experi- ments we can detect both SVl and SV2 antigenicity (Fig. 2A, lane 1, and Fig. 2B, lane 1). This is exactly as we expected from our hypothesis that SVl and SV2 are on the same molecule of -250 kDa. The pattern of SV2 antigenicity we detect on SDS-PAGE is very similar to that of the immunoprecipitated '251-labeled material, both H and L bands are present (compare Fig. 1, lane 4, with Fig. 2B, lane 1).
The pattern of SVl antigenicity detected on im- munoprecipitated SV2 antigen by western blots (Fig. 2A, lane 1) is different than the SV2 antigenicity de- tected on the same SV2 immunoprecipitates (Fig. 2B, lane 1). No L band is detected by the anti-SV1 anti- body. In addition, the SV 1 antigenicity of the H band distributes into lower and higher molecular weight ranges of SDS-PAGE than the SV2 antigenicity. Like-
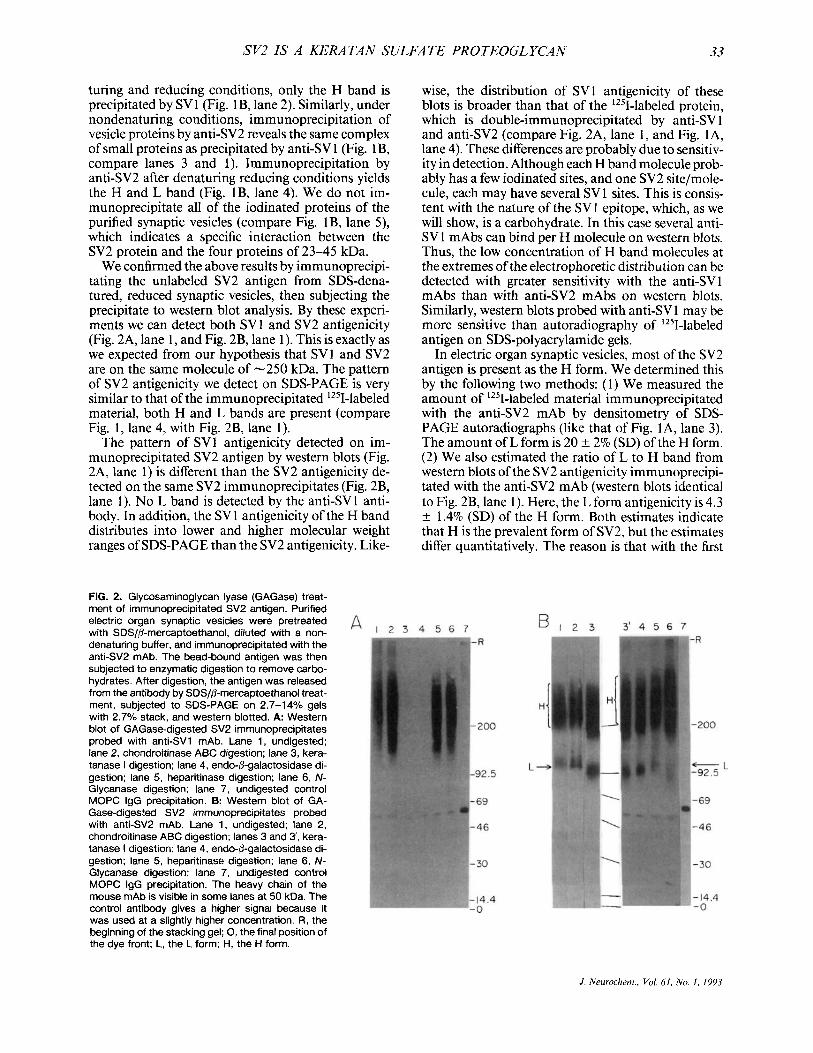
FIG. 2. Glycosaminoglycan lyase (GAGase) treat- ment of immunoprecipitated SV2 antigen. Purified electric organ synaptic vesicles were pretreated with SDS/P-mercaptoethanol, diluted with a non- denaturing buffer, and immunoprecipitated with the anti-SV2 mAb. The bead-bound antigen was then subjected to enzymatic digestion to remove carbo- hydrates. After digestion, the antigen was released from the antibody by SDS/P-mercaptoethanol treat- ment, subjected to SDS-PAGE on 2.7-14% gels with 2.7% stack, and western blotted. A: Western blot of GAGase-digested SV2 immunoprecipitates probed with anti-SV1 mAb. Lane 1, undigested; lane 2, chondroitinase ABC digestion; lane 3, kera- tanase I digestion; lane 4, endo-0-galactosidase di- gestion; lane 5, heparitinase digestion; lane 6, N- Glycanase digestion; lane 7, undigested control MOPC IgG precipitation. B: Western blot of GA- Gase-digested SV2 irnmunoprecipitates probed with anti-SV2 mAb. Lane 1, undigested; lane 2, chondroitinase ABC digestion; lanes 3 and 3', kera- tanase I digestion; lane 4, endo-P-galactosidase di- gestion; lane 5, heparitinase digestion; lane 6, N- Glycanase digestion; lane 7, undigested control MOPC IgG precipitation. The heavy chain of the mouse mAb is visible in some lanes at 50 kDa. The control antibody gives a higher signal because it was used at a slightly higher concentration. R, the beginning of the stacking gel; 0, the final position of the dye front; L, the L form; H, the H form.
wise, the distribution of SVI antigenicity of these blots is broader than that of the '251-labeled protein, which is double-immunoprecipitated by anti-SV 1 and anti-SV2 (compare Fig. 2A, lane 1, and Fig. 1 A, lane 4). These differences are probably due to sensitiv- ity in detection. Although each H band molecule prob- ably has a few iodinated sites, and one SV2 site/mole- cule, each may have several SVl sites. This is consis- tent with the nature of the SVl epitope, which, as we will show, is a carbohydrate. In this case several anti- SV 1 mAbs can bind per H molecule on western blots. Thus, the low concentration of H band molecules at the extremes of the electrophoretic distribution can be detected with greater sensitivity with the anti-SV 1 mAbs than with anti-SV2 mAbs on western blots. Similarly, western blots probed with anti-SV 1 may be more sensitive than autoradiography of 'Z51-labeled antigen on SDS-polyacrylamide gels.
In electric organ synaptic vesicles, most of the SV2 antigen is present as the H form. We determined this by the following two methods: (1) We measured the amount of '251-labeled material immunoprecipitated with the anti-SV2 mAb by densitometry of SDS- PAGE autoradiographs (like that of Fig. lA, lane 3). The amount of L form is 20 f 2% (SD) of the H form. (2) We also estimated the ratio of L to H band from western blots of the SV2 antigenicity immunoprecipi- tated with the anti-SV2 mAb (western blots identical to Fig. 2B, lane 1). Here, the L form antigenicity is 4.3 k 1.4% (SD) of the H form. Both estimates indicate that H is the prevalent form of SV2, but the estimates differ quantitatively. The reason is that with the first
J . Neurochem., Vol. 61, No. I , 1993
34 T. W. SCRANTON ET AL.
estimate, we must assume that the H and the L forms have the same number of iodinated sites per molecule and the same protein core size. With the second esti- mate, we must assume that the H form and the L form behave identically in the immunoblotting process and that the number of SV2 epitopes per protein core is constant. Thus, both estimates rest on different as- sumptions, but both suggest that H is the major form of the SV2 antigen.
Our precipitation experiments demonstrate that the SV 1 epitope is present on the H form of SV2. Is all the SVl epitope present on the SV2 antigen or is it also present on other vesicle proteins? It is clear from Fig. 1 (lanes A I and B4) that the anti-SV 1 mAb essen- tially precipitates only 1251-labeled protein that mi- grates with the H band of the SV2 antigen. No other vesicle proteins are precipitated. Does any of the SVI- containing protein that migrates as the H form of SV2 antigen not contain the SV2 epitope? Most SVl epi- tope is probably associated with SV2 epitope. We at- tempted to deplete the SV2 antigen from reduced and SDS-denatured synaptic vesicles by several successive rounds of immunoprecipitations with anti-SV2 mAb. The amount of SV2 and SV 1 antigenicity remaining in the supernatant after these immunoprecipitations was assayed by western blot as in Fig. 2 (data not shown). We could not produce a solution containing only SVl without SV2 by this method.
The SVl and SV2 antigenicities coelute from Sephacryl S-500 like a protein of 250 kDa
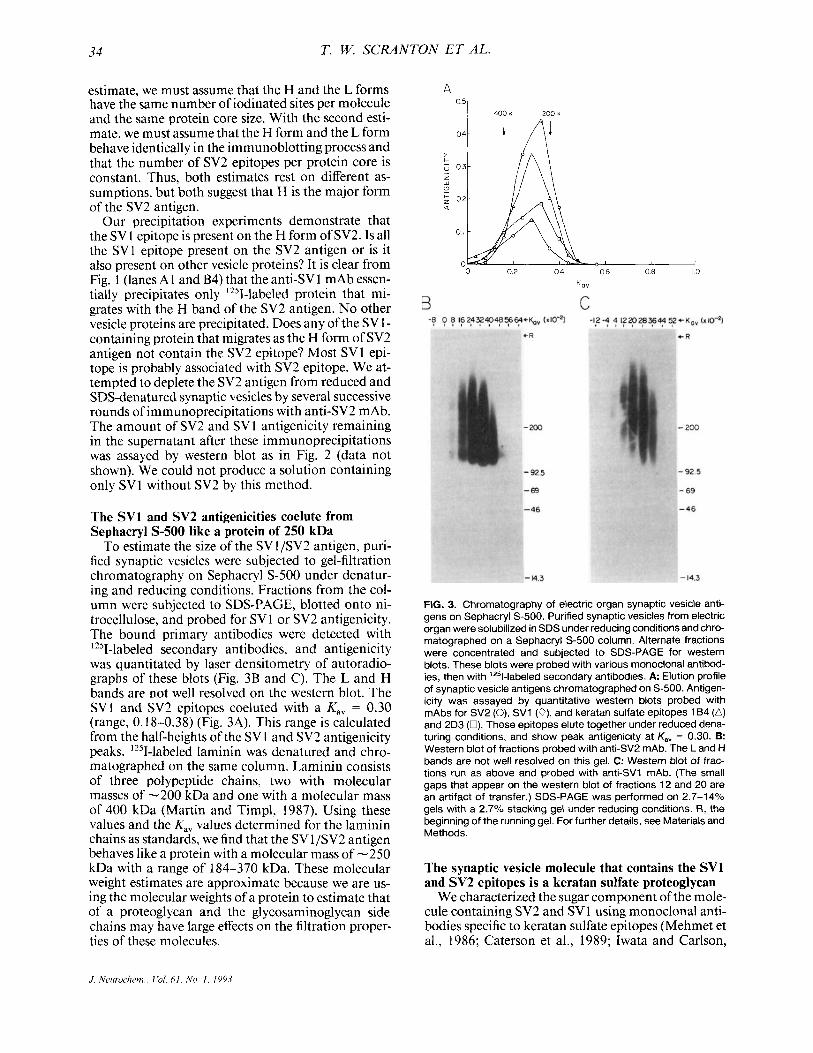
To estimate the size of the SV 1 /SV2 antigen, pun- fied synaptic vesicles were subjected to gel-filtration chromatography on Sephacryl S-500 under denatur- ing and reducing conditions. Fractions from the col- umn were subjected to SDS-PAGE, blotted onto ni- trocellulose, and probed for SV l or SV2 antigenicity. The bound primary antibodies were detected with '251-labeled secondary antibodies, and antigenicity was quantitated by laser densitometry of autoradio- graphs of these blots (Fig. 3B and C). The L and H bands are not well resolved on the western blot. The SVl and SV2 epitopes coeluted with a K,, = 0.30 (range, 0.18-0.38) (Fig. 3A). This range is calculated from the half-heights of the SV 1 and SV2 antigenicity peaks. '2sI-labeled laminin was denatured and chro- matographed on the same column. Laminin consists of three polypeptide chains, two with molecular masses of -200 kDa and one with a molecular mass of 400 kDa (Martin and Timpl, 1987). Using these values and the K,, values determined for the laminin chains as standards, we find that the SV 1 jSV2 antigen behaves like a protein with a molecular mass of -250 kDa with a range of 184-370 kDa. These molecular weight estimates are approximate because we are us- ing the molecular weights of a protein to estimate that of a proteoglycan and the gl ycosaminoglycan side chains may have large effects on the filtration proper- ties of these molecules.
A
05[ 400h 200 k
KO"
B C
FIG. 3. Chromatography of electric organ synaptic vesicle anti- gens on Sephacryl s-500. Purified synaptic vesicles from electric organ were solubilized in SDS under reducing conditions and chro- matographed on a Sephacryl 5-500 column. Alternate fractions were concentrated and subjected to SDS-PAGE for western blots. These blots were probed with various monoclonal antibod- ies, then with "51-labeled secondary antibodies. A: Elution profile of synaptic vesicle antigens chromatographed on S-500. Antigen- icity was assayed by quantitative western blots probed with mAbs for SV2 (0), SV1 (o), and keratan sulfate epitopes 164 (A) and 2D3 (0). These epitopes elute together under reduced dena- turing conditions, and show peak antigenicity at K,, = 0.30. B: Western blot of fractions probed with anti-SV2 mAb. The Land H bands are not well resolved on this gel. C: Western blot of frac- tions run as above and probed with anti-SV1 mAb. (The small gaps that appear on the western blot of fractions 12 and 20 are an artifact of transfer.) SDS-PAGE was performed on 2.7-14% gels with a 2.7% stacking gel under reducing conditions. R, the beginning of the running gel. For further details, see Materials and Methods.
The synaptic vesicle molecule that contains the SV1 and SV2 epitopes is a keratan sulfate proteoglycan
We characterized the sugar component of the mole- cule containing SV2 and SVl using monoclonal anti- bodies specific to keratan sulfate epitopes (Mehmet et al., 1986; Caterson et al., 1989; Iwata and Carlson,
J . Neriroch~~rn.. L ' d . 61. No I , 1993
SV2 IS A KERATAN SULFATE PROTEOGLYCAN 35
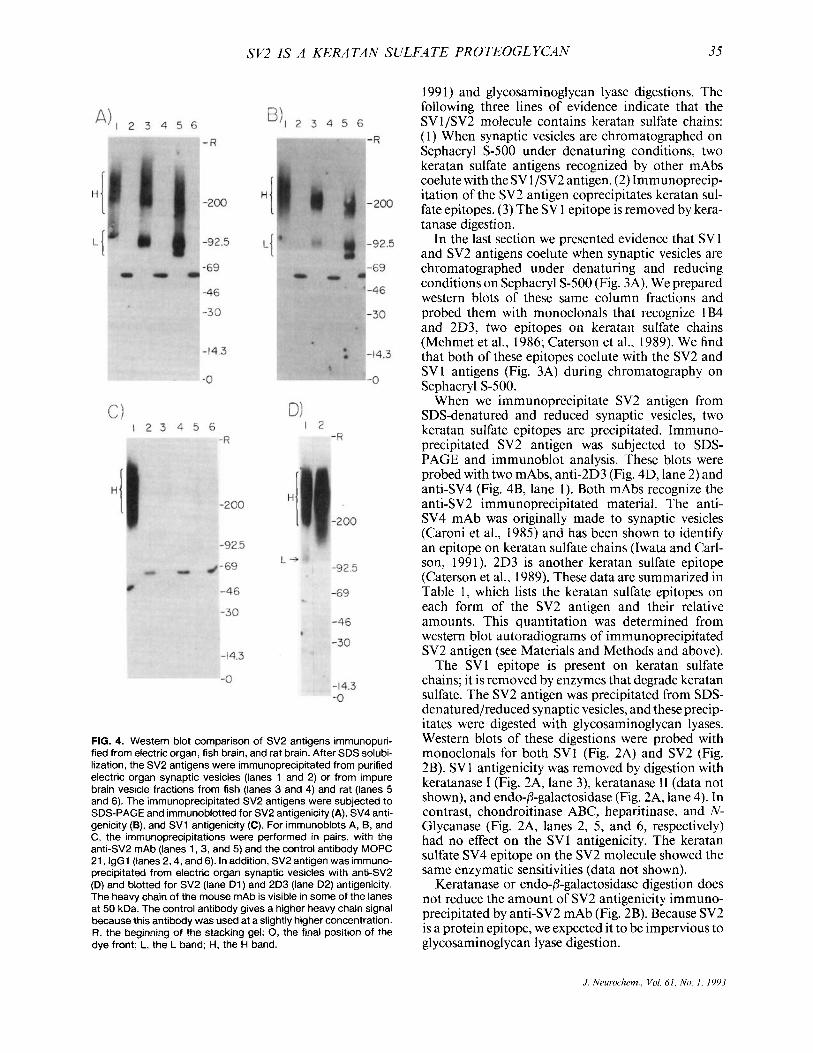
FIG. 4. Western blot comparison of SV2 antigens immunopuri- fied from electric organ, fish brain, and rat brain. After SDS solubi- lization, the SV2 antigens were immunoprecipitated from purified electric organ synaptic vesicles (lanes 1 and 2) or from impure brain vesicle fractions from fish (lanes 3 and 4) and rat (lanes 5 and 6). The immunoprecipitated SV2 antigens were subjected to SDS-PAGE and immunoblotted for SV2 antigenicity (A), SV4 anti- genicity (B), and SV1 antigenicity (C). For immunoblots A, B, and C, the immunoprecipitations were performed in pairs, with the anti-SV2 mAb (lanes 1,3, and 5) and the control antibody MOPC 21, lgGl (lanes 2,4, and 6). In addition, SV2 antigen was immuno- precipitated from electric organ synaptic vesicles with anti-SV2 (D) and blotted for SV2 (lane D1) and 2D3 (lane D2) antigenicity. The heavy chain of the mouse mAb is visible in some of the lanes at 50 kDa. The control antibody gives a higher heavy chain signal because this antibody was used at a slightly higher concentration. R, the beginning of the stacking gel; 0, the final position of the dye front; L, the L band; H, the H band.
199 1) and glycosaminoglycan lyase digestions. The following three lines of evidence indicate that the SV 1/SV2 molecule contains keratan sulfate chains: (1) When synaptic vesicles are chromatographed on Sephacryl S-500 under denaturing conditions, two keratan sulfate antigens recognized by other mAbs coelute with the SVl/SV2 antigen. (2) Immunoprecip- itation of the SV2 antigen coprecipitates keratan sul- fate epitopes. (3) The SV1 epitope is removed by kera- tanase digestion.
In the last section we presented evidence that SV 1 and SV2 antigens coelute when synaptic vesicles are chromatographed under denaturing and reducing conditions on Sephacryl S-500 (Fig. 3A). We prepared western blots of these same column fractions and probed them with monoclonals that recognize 1B4 and 2D3, two epitopes on keratan sulfate chains (Mehmet et al., 1986; Caterson et al., 1989). We find that both of these epitopes coelute with the SV2 and SVl antigens (Fig. 3A) during chromatography on Sephacryl S-500.
When we immunoprecipitate SV2 antigen from SDS-denatured and reduced synaptic vesicles, two keratan sulfate epitopes are precipitated. Immuno- precipitated SV2 antigen was subjected to SDS- PAGE and immunoblot analysis. These blots were probed with two mAbs, anti-2D3 (Fig. 4D, lane 2) and anti-SV4 (Fig. 4B, lane 1). Both mAbs recognize the anti-SV2 immunoprecipitated material. The anti- SV4 mAb was originally made to synaptic vesicles (Caroni et al., 1985) and has been shown to identify an epitope on keratan sulfate chains (Iwata and Carl- son, 1991). 2D3 is another keratan sulfate epitope (Caterson et al., 1989). These data are summarized in Table 1, which lists the keratan sulfate epitopes on each form of the SV2 antigen and their relative amounts. This quantitation was determined from western blot autoradiograms of immunoprecipitated SV2 antigen (see Materials and Methods and above).
The SVl epitope is present on keratan sulfate chains; it is removed by enzymes that degrade keratan sulfate. The SV2 antigen was precipitated from SDS- denatured/reduced synaptic vesicles, and these precip- itates were digested with glycosaminoglycan lyases. Western blots of these digestions were probed with monoclonals for both SVl (Fig. 2A) and SV2 (Fig. 2B). SVl antigenicity was removed by digestion with keratanase I (Fig. 2A, lane 3), keratanase I1 (data not shown), and endo-b-galactosidase (Fig. 2A, lane 4). In contrast, chondroitinase ABC, heparitinase, and N- Glycanase (Fig. 2A, lanes 2, 5, and 6, respectively) had no effect on the SVl antigenicity. The keratan sulfate SV4 epitope on the SV2 molecule showed the same enzymatic sensitivities (data not shown).
Keratanase or endo-/I-galactosidase digestion does not reduce the amount of SV2 antigenicity immuno- precipitated by anti-SV2 mAb (Fig. 2B). Because SV2 is a protein epitope, we expected it to be impervious to glycosaminoglycan lyase digestion.
J . Neurochem., Vol. 61, No. I , I993
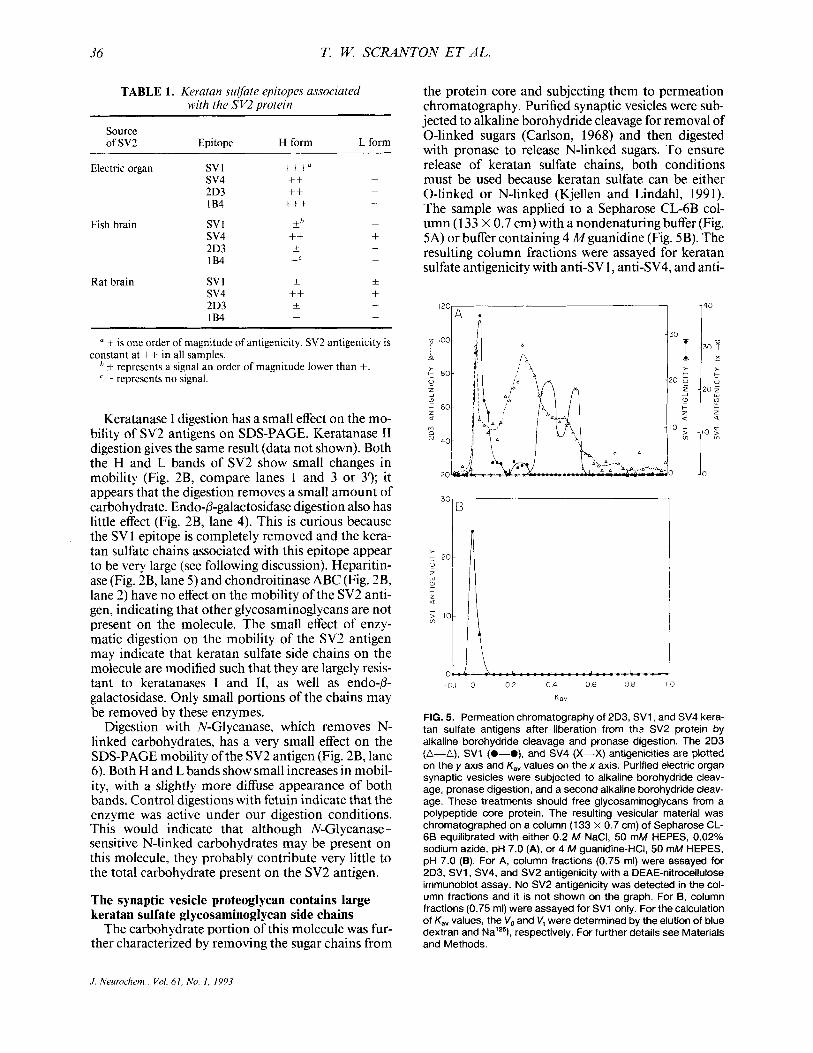
36 T. W. SCRANTON E T AL.
-
TABLE 1. Kerutun sulfate epitopes ussociuted with the SV2 urotein
740
-30 f I
t
U c
20 ;
-10 2
L? z 4
0
Source of sv2 Epitope H form L form
Electric organ sv 1 + + + a ~
- sv4 ++ 2D3 ++ 184 +++
- -
Fish brain sv 1 f h ~
sv4 ++ + 2D3 -
1 B4 - +
- ~
Rat brain sv I f * sv4 ++ + 2D3 f I B4
-
- -
a + is one order of magnitude of antigenicity. SV2 antigenicity is constant at ++ in all samples. ’ k represents a signal an order of magnitude lower than f.
c. - represents no signal.
Keratanase I digestion has a small effect on the mo- bility of SV2 antigens on SDS-PAGE. Keratanase I1 digestion gives the same result (data not shown). Both the H and L bands of SV2 show small changes in mobility (Fig. 2B, compare lanes I and 3 or 3’); it appears that the digestion removes a small amount of carbohydrate. Endo-P-galactosidase digestion also has little effect (Fig. 2B, lane 4). This is curious because the SV1 epitope is completely removed and the kera- tan sulfate chains associated with this epitope appear to be very large (see following discussion). Heparitin- ase (Fig. 2B, lane 5) and chondroitinase ABC (Fig. 2B, lane 2) have no effect on the mobility of the SV2 anti- gen, indicating that other glycosaminoglycans are not present on the molecule. The small effect of enzy- matic digestion on the mobility of the SV2 antigen may indicate that keratan sulfate side chains on the molecule are modified such that they are largely resis- tant to keratanases I and 11, as well as endo-0- galactosidase. Only small portions of the chains may be removed by these enzymes.
Digestion with N-Glycanase, which removes N- linked carbohydrates, has a very small effect on the SDS-PAGE mobility of the SV2 antigen (Fig. 2B, lane 6). Both H and L bands show small increases in mobil- ity, with a slightly more diffuse appearance of both bands. Control digestions with fetuin indicate that the enzyme was active under our digestion conditions. This would indicate that although N-Glycanase- sensitive N-linked carbohydrates may be present on this molecule, they probably contribute very little to the total carbohydrate present on the SV2 antigen.
The synaptic vesicle proteoglycan contains large keratan sulfate glycosaminoglycan side chains
The carbohydrate portion of this molecule was fur- ther characterized by removing the sugar chains from
the protein core and subjecting them to permeation chromatography. Purified synaptic vesicles were sub- jected to alkaline borohydride cleavage for removal of 0-linked sugars (Carlson, 1968) and then digested with pronase to release N-linked sugars. To ensure release of keratan sulfate chains, both conditions must be used because keratan sulfate can be either 0-linked or N-linked (Kjellen and Lindahl, 1991). The sample was applied to a Sepharose CL-6B col- umn (1 33 X 0.7 cm) with a nondenaturing buffer (Fig. 5A) or buffer containing 4 Mguanidine (Fig. 5B). The resulting column fractions were assayed for keratan sulfate antigenicity with anti-SV 1, anti-SV4, and anti-
I20
.;a 100
: 80
4
0
a F 60 w u
ro
40
20
-01 0 0 2 0 4 0 6 0 8 10
K a v
FIG. 5. Permeation chromatography of 2D3, SV1, and SV4 kera- tan sulfate antigens after liberation from thz SV2 protein by alkaline borohydride cleavage and pronase digestion. The 2D3 (A-A), SV1 (0-O), and SV4 (X-X) antigenicities are plotted on the y axis and K,, values on the x axis. Purified electric organ synaptic vesicles were subjected to alkaline borohydride cleav- age, pronase digestion, and a second alkaline borohydride cleav- age. These treatments should free glycosaminoglycans from a polypeptide core protein. The resulting vesicular material was chromatographed on a column (133 x 0.7 cm) of Sepharose CL- 66 equilibrated with either 0.2 M NaCI, 50 mM HEPES, 0.02% sodium azide, pH 7.0 (A), or 4 M guanidine-HCI, 50 mM HEPES, pH 7.0 (B). For A, column fractions (0.75 mi) were assayed for 2D3, SVI , SV4, and SV2 antigenicity with a DEAE-nitrocellulose immunoblot assay. No SV2 antigenicity was detected in the col- umn fractions and it is not shown on the graph. For B, column fractions (0.75 ml) were assayed for SV1 only. For the calculation of K,, values, the V, and V, were determined by the elution of blue dextran and Nalz51, respectively. For further details see Materials and Methods.
.I. Neurochern , Vol. 6 I , No. I , 1993

SV2 IS A KERATAN SULFATE PROTEOGLYCAN 37
2D3, as well as for SV2 antigenicity, by DEAE-nitro- cellulose dot-blot.
Under nondenaturing conditions, keratan sulfate antigenicity, as detected by mAb 2D3, eluted from Sepharose CL-6B in three main peaks (Fig. 5A), in the void volume and at K,, values of 0.26 and 0.45. We can obtain rough molecular weight estimates corre- sponding to these K,, values by using the published data of Wasteson (197 l), who determined the rela- tionship between K,, values and molecular weight for a series of linear chondroitin sulfate polymers. Based on these data, our Kay values correspond to molecular weights of 80,000 and 27,000. The half-heights of the leading edge of the second peak (K,, of 0.26) and the trailing edge of the third peak (Kav of 0.45) elute with Kay values of 0.16 and 0.54, respectively. These K,, values indicate the range of keratan sulfate chains and corresponding molecular weights of 140,000- 17,000. We cannot make a molecular weight estimate for the keratan sulfate-containing material in the void vol- ume, but it must be > 140,000. Heterogeneous sizes of the keratan sulfate chains may be responsible for the extremely broad bands that we see on SDS-PAGE of the intact SV 1 /SV2 antigen.
Another keratan sulfate epitope on the SVljSV2 antigen, SV4, elutes as two peaks with Kay values of 0.38 and 0.52 (Fig. 5A). The half-heights of the lead- ing edge of the first peak and the trailing edge of the second peak elute with K,, values of 0.55 and 0.34, respectively. Thus, we estimate the molecular weight range for the SVCcontaining keratan sulfate chains to be 16,000-50,000. The SV4 chains represent the smaller keratan sulfate chains of the proteoglycan.
SV 1 antigenicity elutes as two peaks, a large peak in the void volume and a small peak with a K,, of 0.26, which coelute with the major keratan sulfate (2D3) peaks (Fig. 5A). The eluted SVl antigenicity is thus distinct from the SV4 antigen, which elutes after the second SVl peak. These epitopes must therefore be located on different keratan sulfate side chains.
The SV 1 -containing keratan sulfate chains on the SV2 molecule are largest, most > 140,000 daltons, some -80,000 daltons. Initially, we found that the freed SV1 chains chromatographed as large chains. We were concerned that our cleavage of the oligosac- charide chains from the protein might be incomplete. It was possible that some of the polypeptide chain might still link these sugar chains and cause them to behave as a larger species during permeation chroma- tography on Sepharose CL-6B. So we performed two cycles of alkaline borohydride cleavage plus a pronase digestion, the experiments shown in Fig. 5. Given the harsh conditions used to free these chains and the ab- sence of the SV2 antigenicity eluting from the column (data not shown), we consider it unlikely that the chains could be linked by protein.
We were concerned that the aggregation of isolated carbohydrate chains could cause them to appear larger during chromatography. To test this possibility,
we conducted chromatography of the freed oligosac- charide chains with a denaturing buffer containing 4 A4 guanidine-HC1. Under these conditions, the peak of the SVl antigenicity elutes in the column void (Fig. 5B), as it does under nondenaturing conditions (Fig. 5A), indicating that chain aggregation is probably not a factor.
Fish and rat brain SV2 antigens are keratan sulfate proteoglycans
When the SV2 antigen is isolated from fish or rat brain, both the L and the H forms are present. In brain, both forms of SV2 antigen contain keratan sul- fate chains. We immunoprecipitated SV2 antigen from purified electric organ synaptic vesicles, from an impure vesicle fraction of fish brain without its elec- tromotor nucleus, and from a rat brain impure vesicle fraction. Before precipitation, the proteins in these samples were solubilized with SDS under reducing conditions. The immunoprecipitates were subjected to SDS-PAGE, immunoblotted, and probed with anti-SV2 (Fig. 4A) and anti-SV4 (Fig. 4B). Like elec- tric organ (Fig. 4A, lane l), fish brain contains a 100- kDa (L) SV2 antigen as well as the large H band (Fig. 4A, lane 3). The H and L bands are also apparent in rat brain (Fig. 4A, lane 5). Here, the L form appears as a doublet with proteins at 83 and 98 kDa. In fish and rat brain, all forms of the SV2 antigen contain the SV4 epitope (Fig. 4B, lanes 3 and 5) . The keratan sulfate epitope 2D3 is found in small amounts on the H form of these molecules as well, though they lack the 1B4 epitope (data not shown). The SV 1 epitope is not present in significant amounts in brain tissue from either species (see below). Thus, the SV2 anti- gens in fish and rat brain are keratan sulfate proteogly- cans, though some chemical differences are indicated by the observed differences in antigenicity. The data are summarized in Table 1.
Rat brain SV2 antigen behaves like a smaller mole- cule than the SV2 antigen from electric organ. As a source of rat brain SV2 antigen, we prepared synapto- somes from rat brain and applied the SDS-solubilized membranes under reducing conditions to the same Sephacryl S-500 column described above (for Fig. 3). The SV2 antigenicity elutes as a single peak with a K,, of 0.36, and half-heights of the leading and trailing edge are 0.3 and 0.43 (data not shown). Using laminin chains of 200 and 400 kDa chromatographed on the same column, we calculate that the SV2 antigen from rat brain behaves like a protein of 200 kDa with a molecular mass range of 160-250 kDa.
The SV1 epitope is highly enriched on the SV2 antigen in electromotor neurons of the electric fish; it is present in very low amounts elsewhere in the brain
The SV1 epitope is present on the molecule con- taining the SV2 antigen only in the neurons of the electromotor nucleus in the electric fish brain. These are the neurons that innervate the electric organ. We
J. Neurochem., Vol. 61, No. I , 1993

38 T. W. SCRANTON ET AL.
demonstrate this both immunocytochemically and biochemically.
Consecutive cross sections of the electric fish brain, which contain the electromotor nucleus, are stained with either the anti-SV2 or anti-SV1 mAbs (Fig. 6). As expected, the SV2 antigenicity is present throughout the brain, including the cells of the electromotor nu- cleus and their process projecting to the electric organ (Fig. 6A). In contrast, SV1 antigenicity is located ex- clusively in the neurons of the electromotor nucleus and their processes (Fig. 6B). SVl antigenicity is not seen in any other part of the fish brain (Fig. 6B), nor is it present at the neuromuscularjunction in this organ- ism (data not shown). This indicates that the carbohy- drate modification that is identified by anti-SV1 is specific to electromotor neurons, though it is cova- lently attached to the SV2 protein core found throughout the electric fish brain.
When the SV2 antigen is isolated from fish brain tissue without the electromotor nucleus, very little SV1 is present on the molecule. Electric fish brain was dissected so as not to include the electromotor nu- cleus, and a crude synaptic vesicle fraction was pre- pared. The SV2 antigen was immunoprecipitated after exposure to SDS and reducing conditions, and subjected to SDS-PAGE and immunoblot analysis. When these blots were probed with anti-SV1, we de- tected very little antigenicity (Fig. 4C, lane 3). How- ever, anti-SV2 shows a strong antigenic signal (Fig. 4B, lane 3). Similar experiments demonstrate that lit- tle SV 1 antigen is present on the rat brain SV2 antigen (Fig. 4B, lane 5) .
Although the SVl epitope is found on keratan sul- fate chains, it is not a common keratan sulfate epi- tope. When we assayed commercially available prepa- rations of glycosaminoglycans, the SV 1 antigen was completely absent (Table 2). This is quite unlike the SV4 epitope that is present on the synaptic vesicle proteoglycan (Caroni et al., 1985; Carlson, 1989); a large chondroitin/keratan sulfate proteoglycan, PG- 1000 from electric organ (Carlson and Wight, 1987; Iwata and Carlson, 1991), in commercial prepara-
4
FIG. 6. lmmunocytochemical localization of SV1 and SV2 anti- genicities in the electromotor nucleus of electric fish brain. Ten- micrometer horizontal cross-sections of fish brain containing electromotor nucleus were exposed to anti-SV2 mAb (A), anti- SV1 mAb (B), or a control antibody (MOPC 21, IgG1) (C). A la- beled cartoon outline of the area sectioned is also shown (D). The bound antibodies were visualized with a biotinylated goat anti- mouse IgG, streptavidin, and biotinylated horseradish peroxi- dase. The SVl antigenicity (6) is present only in electromotor neuronal cell bodies and their processes; it is not present in other neurons. The SV2 antigenicity (A) is present throughout the sec- tion, including the electromotor neuronal cell bodies and their pro- cesses. For most neurons, the SV2 staining in the cell body and process is low (Buckley and Kelly, 1985). Presumably, electromo- tor neurons synthesize large amounts of synaptic vesicle proteins because they have very large nerve terminal arbors to maintain. Bars, 418 pm.
-
J. Neurochem.. Vol. 61, No. I , 1993

SV2 IS A KERATAN SULFATE PROTEOGLYCAN 39
TABLE 2. The absence of SVI antigenicity in commercial preuarat ions of nlvcosam inonlvcans
Sample
Whale cartilage chondroitin sulfate Porcine skin dermatan sulfate Shark cartilage chondroitin sulfate Bovine cornea keratan sulfate Heparin Bovine kidney heparan sulfate Synaptic vesicle proteoglycan
sv 1 antigenicitf
0 0 0 0 0 0
+++
sv4 antigenicity"
+ 0
+++ ++ 0 0
NT
NT, not determined in the experiment shown here. + is one order of magnitude antigenicity per microgram of gly-
cosaminoglycan.
tions of keratan sulfate, and on the keratan sulfate that heavily contaminates commercial preparations of chondroitin sulfate (Table 2; Iwata and Carlson, 1991). The sensitivity of the SVI epitope to keratan- ase and its absence in commercial preparations sug- gest that it is a unique keratan sulfate structure.
DISCUSSION Previous investigators demonstrated that SV2 is an
epitope of a synaptic vesicle protein on the cytoplas- mic side of the vesicle membrane (Buckley and Kelly, 1985). The recent cloning of the protein core contain- ing this epitope has revealed many interesting charac- teristics (Bajjalieh et al., 1992; Feany et al., 1992). The cDNA, cloned from rat brain and PC 12 cells, encodes a protein core of 82.7 kDa. The amino acid sequence predicts a cytoplasmic N-terminal domain, 12 mem- brane spanning domains, and two large loops, one cytoplasmic and one luminal (Bajjalieh et al., 1992; Feany et al., 1992). The N-terminal cytoplasmic do- main contains the SV2 epitope (Bajjalieh et al., 1992). The large luminal domain contains three potential N- glycosylation sites (Bajjalieh et al., 1992; Feany et al., 1992).
In this study, we have definitively shown that the SV2 protein is a keratan sulfate proteoglycan. Two SDS-PAGE forms of the SV2 proteoglycan (SV2pg) exist in fish electric organ, fish brain, and rat brain, i.e., the L form that migrates like a protein of 100-kDa and the more slowly migrating H form. The keratan sulfate epitope SV4 is present on the H and L forms in rat and fish brain; the L form in electric organ lacks this epitope. Although the electric organ L form lacks the SV4 epitope, we see an increase in its mobility on SDS-PAGE after keratanase digestion (Fig. 2B, lanes 3 and 3'). Thus, the electric organ L form may also contain keratan sulfate chains.
In electric organ synaptic vesicles, the H form con- tains the SVl epitope that was the original marker of the synaptic vesicle proteoglycan. The investigators (Carlson and Kelly, 1983; Buckley and Kelly, 1985; Caroni et al., 1985) who initially described the SVI
and SV2 epitopes did not recognize that they were present on the same molecule. This misunderstand- ing was probably due to the SDS-polyacrylamide gel systems previously used. In these systems, the L form would enter the resolving gel, whereas the H form would remain in the stacking gel and not enter the resolving phase. The stacking gel is usually discarded and not used for immunoblot analysis. Thus, only the L form with a mobility of a 1 00-kDa protein would be observed. The 2.7-14% gradient gel system we use does not suffer from this problem; essentially no pro- teins remain in the 2.7% stacking gel at the end of electrophoresis.
The H form from electric organ contains keratan sulfate side chains that vary greatly in size. SV I, SV4, and 2D3 epitopes are antigenic sites on keratan sul- fate chains of the H form. SV4 and 2D3 are known epitopes on keratan sulfate chains (Caterson et al., 1989; Iwata and Carlson, 199 1). The removal of SV 1 from the SV2pg by keratanase digestion (Fig. 2) dem- onstrates that an SVI antigen site is also present on a keratan sulfate chain. When glycosaminoglycan side chains are cleaved from SV2pg and chromatographed on Sepharose CLdB, the glycosaminoglycans bearing the SV 1, SV4, and 2D3 epitopes chromatograph like oligosaccharides of 17,000 to > 140,000 daltons.
The H and L forms differ in their mobility on SDS-
sv4 H Form
sv2 f7
sv4 H Form
KSase .L
FIG. 7 . Hypothetical model for SV2pg in the electric organ synap- tic vesicle. Both forms of the SV2 antigen are shown, i e , the H and the L forms. The model for the SV2 polypeptide chain is based on that inferred by Feany et al. (1992) and Bajjalieh et al. (1992) from the rat SV2 cDNA sequence. Here we assume that all three potential N-linked glycosylation sites contain N-linked kera- tan sulfate. The H form contains both small and large keratan sulfate chains that have the keratanase sensitive epitopes SVI , SV4, and 2D3. The data suggest that these epitopes may be localized toward the nonreducing ends of the sugar chains. The L form contains small chains with small keratanase (KSase)-sensi- tive regions. The SV4 epitope is present on the fish and rat brain molecule L forms, but not on the L form of the molecule from purified electric organ vesicles.
J . Neurochem., Vol. 61, No. 1, 1993

40 T. W. SCRANTON ET AL.
PAGE (Figs. 1, 2, and 4). The H form migrates as a very broad band with the average mobility of a 250- kDa protein, and the L form like a 100-kDa protein. However, when the SDS-denatured/reduced H and L forms are chromatographed on Sephacryl S-500 under the same denaturing conditions as SDS-PAGE, the two forms elute at very similar volumes. Both forms elute as would an SDS-denatured protein of 250 kDa. This discrepancy probably reflects the dif- ferent physical properties each method uses to sepa- rate proteins and the peculiar physical properties of proteoglycans. Proteins are thought to migrate on SDS-PAGE as a result of the electrical driving force acting on the protein due to the net charge of the pro- tein/SDS micelle. Opposing the forward migration of the protein, the sieving effect of the gel matrix creates a retarding drag on the micelle (van Holde, 1985). The elution of proteins during gel filtration is thought to be dependent on their diffusional properties (Siege1 and Monty, 1966; Scopes, 1987). The empirical rela- tionship between log(molecu1ar weight) and electro- phoretic mobility (or elution volume) for nonglycosy- lated proteins breaks down in the case of heavily gly- cosylated proteins (Blackshear, 1984). For this relationship to hold during SDS-PAGE, the SDS/pro- tein micelles must be rods with the same charge den- sity (van Holde, 1985); for this relationship to hold for gel filtration in denaturing conditions, proteins must be randomly coiled linear homopolymers (Fish et al., 1969; Mann and Fish, 1972). It is likely that neither of these conditions is met by proteoglycans, which are branched heteropolymers containing variable amounts of charge. Thus, it is not surprising that the H and L forms of SV2 demonstrate anomalous behav- ior when fractionated because they may differ radi- cally in those physical properties important for their separation by these methods.
Although a quantitative relationship between SDS- PAGE mobilities and molecular weight has not been established for proteoglycans, it has been found that enzymatic removal of glycosaminoglycan side chains from a proteoglycan changes its SDS-PAGE mobility from a broad heterogeneous band to a faster migrat- ing discrete band (for example, Jalkanen et al., 1985; Vogel and Evanko, 1987; Funderburgh and Conrad, 1990). Thus, we expected the removal of large hetero- geneous glycosaminoglycan chains containing SV 1 and SV4 ( I 7,000 to > 140,000 daltons) to have a signif- icant impact on the heterogeneous mobility of the H form. However, this does not occur. Digestion of the SV2pg with keratanases (I and 11) and endo-p-galacto- sidase results in removal of the SV 1 and SV4 epitopes, but no decrease in the heterogeneous mobility of the H form and only a small increase in the SDS-PAGE mobility of the H and L forms (Fig. 2). Because the heterogeneous mobility of the H form changes little, presumably little carbohydrate is removed from the SV2pg by keratanase or endo-P-galactosidase diges- tion. Perhaps the keratan sulfate side chains represent
only a minor amount of the total carbohydrate on the SV2 molecule. Alternatively, most of the keratan sul- fate side chain may be resistant to keratanase diges- tion and only the SVl and SV4 epitopes, localized at the nonreducing end of the keratan sulfate side chains away from the polypeptide backbone, are removed. The first explanation is possible, but less likely. If these large keratan sulfate chains make only a minor contribution to the mass of the molecule, large amounts of additional carbohydrate must be present. This hypothetical carbohydrate could not be other known glycosaminoglycans, heparin, heparan sulfate, dermatan sulfate, or chondroitin sulfate, because the enzymes that degrade these molecules have little or no effect on the SDS mobility of SV2 antigenicity. Also, previous experiments indicated that this molecule was either a keratan sulfate or a heparan sulfate pro- teoglycan (Stadler and Dowe, 1982; Carlson and Kelly, 1983), based on the mobility of the isolated chains on thin layer electrophoresis.
We favor the explanation that the keratan sulfate chains are resistant to keratanase digestion, and thus the enzyme has no great effect on the mobility of the SV2pg on SDS-PAGE. Presumably, the disaccharides in most of these chains are modified so that the galac- tose-N-acetylglucosamine backbone is no longer a substrate for the glycosaminoglycan lyase enzymes. A small keratanase-sensitive region of the chain (con- taining the SVl and SV4 epitopes) could be at the nonreducing end of the side chain. This modification is probably not simply a matter of over- or undersulfa- tion of keratan sulfate. In addition to keratanase I, we used endo-0-galactosidase, which degrades unsulfated keratan sulfate (lactosaminoglycans) as well as kera- tan sulfate (Fukuda and Matsumura, 1976), and kera- tanase I1 (data not shown), which degrades oversul- fated keratan sulfate (Seikagaku Kogyo Co. technical data). All these enzymes had the same effect on the SV2pg, producing no reduction in the heterogeneity of the H form’s mobility on SDS-PAGE and only a small increase in electrophoretic mobility ofthe L and H forms. It has been proposed that branched sugars on the polysaccharide backbone may explain the resis- tance of some keratan sulfate chains to digestion (Fu- kuda and Matsumura, 1976; Melrose and Ghosh, 1988). Disaccharide units containing branched sugars may be poor substrates for keratanases or P-galactosi- dase. Fucose (Tai et al., 199 1) and perhaps sialic acid residues (Nieduszynski et al., 1990) have been found attached to the repeating disaccharide unit in some keratan sulfates. Keratan sulfate chains containing N- sulfated glucosamine residues have also been de- scribed (Tang et al., 1986) and shown to be resistant to keratanase digestion (Block et al., 1992). This type of modification is one possible explanation of the resis- tance to digestion that we have observed. We have observed a similar resistance to enzymatic digestion in keratan sulfate chains immunopurified from shark cartilage fractions (CS-C) with the anti-SV4 mAb.
J. Neurochrm., Val. 61, No. 1. 1993

SV2 IS A KERATAN SULFATE PROTEOGLYCAN 41
Large (-50,000 kDa) sections of these chains remain after digestion with mixtures of keratanase I and I1 (T. W. Scranton, unpublished data). Such modifications have not been reported for other glycosaminoglycans; for example, the known glycosaminoglycan lyases are able to completely digest the heparin/heparan sulfate and chondroitin sulfate families.
We have observed that N-Glycanase digestion causes only a small change in the mobility of the SV2pg on SDS-PAGE. This result is surprising, given that the amino acid sequence of this protein shows the presence of three potential N-glycosylation sites (Baj- jalieh et al., 1992; Feany et al., 1992). Keratan sulfate chains can be either 0-linked or N-linked (Kjellen and Lindahl, 1991), and it is possible that 0-linked keratan sulfate chains are present on the SV2pg. How- ever, the consensus sequence identified in bovine ag- grecan for the attachment of 0-linked keratan sulfate (Antonsson et al., 1989) is not present in this mole- cule, making this less likely. For the L form, our re- sults appear to contrast with data reported by Buckley and Kelly (1985). They demonstrated that treatment with tunicamycin, which prevents the addition of N- linked sugars, causes a greater size shift in the L form of the SV2 antigen produced by PC12 cells, from a molecular weight of 100,000 to 62,000. Volknandt and Zimmermann (1 990) achieved similar results us- ing a mixture of deglycosylating enzymes to digest the L form of SV2pg isolated from Torpedo vesicles and from bovine brain. N-Glycanase digestion may not, therefore, provide an accurate assessment of the total molecular weight contribution of N-linked sugars on the L form of SV2. This enzyme can have trouble digesting N-linked sugars from proteins.
The model in Fig. 7 reflects protein structural pre- dictions based on the amino acid sequence of the SV2pg (Bajjalieh et al., 1992; Feany et al., 1992) and our hypothesis that the L and H forms in electric or- gan synaptic vesicles are composed of the same core protein, but differ in glycosylation. The keratan sul- fate chains are shown attached to the three potential N-linked glycosylation sites in the luminal loop of the SV2 proteoglycan. The H form contains keratan sul- fate chains that are large, heterogeneous, and contain the SVl , SV4, and 2D3 epitopes. The L form contains keratan sulfate chains without the SV1, SV4, and 2D3 epitopes. (Of course, in fish and rat brain, the L form contains the SV4 epitope.) Alternatively, it is possible that the core protein for the H and L forms could represent alternative splicing patterns of the same gene, with the H form having attachment regions for oligosaccharide chains containing SV1, SV4, and 2D3.
Electric organ synaptic vesicles probably contain one, or at most two, major types of proteoglycans. In past experiments, the glycosaminoglycan content of purified electric organ synaptic vesicles was analyzed by thin layer electrophoresis. The glycosaminoglycan side chains were freed from protein by protease diges- tion of delipidated vesicles (Carlson and Kelly, 1983).
On this thin layer system, only one band ofglycosami- noglycans copurified with synaptic vesicles. These gly- cosaminoglycans, as well as heparin and keratan sul- fate, all had the same electrophoretic mobility on this thin layer system. No other glycosaminoglycans co- purified with synaptic vesicles. In addition (Carlson and Kelly, 1983), the electrophoretic band of glycos- aminoglycans appeared to be associated with the pro- tein bearing the SV 1 epitope (identified by the Tor 70 mAb). These data indicated that either heparin or ker- atan sulfate, or both of these glycosaminoglycans, might be present on the proteoglycan. In the experi- ments we present here, we again prepared glycosami- noglycans from purified electric organ synaptic vesi- cles and found that they contain keratan sulfate epi- topes (Fig. 5). The SV2 protein that bears these antigenic chains shows no sensitivity to heparinase or heparitinase (Fig. 2, and data not shown), indicating that heparin chains are probably not present on the proteoglycan. Thus, electric organ synaptic vesicles contain a keratan sulfate proteoglycan, although it is possible that they could contain a heparin proteogly- can as well.
The SV2pg probably contains the majority of the keratan sulfate epitopes of electric organ synaptic vesi- cles. It is presumably the major keratan sulfate proteo- glycan of the vesicle. This is suggested by these epi- topes appearing inseparable from intact SV2. (1) Under denaturing conditions, the keratan sulfate epi- topes SV1,2D3, and lB4 coelute with SV2 from Se- phacryl S-500 (Fig. 3). None of these keratan sulfate antigens elute without SV2. (2) The immunoprecipi- tation experiments (Fig. 1) suggest that the majority of the SVI epitope is present on the SV2pg. (3) The keratan sulfate epitopes recognized by antibodies 2D3 and anti-SV4 can be precipitated from denatured vesi- cles by the anti-SV2 mAb (Fig. 4B and D). We have attempted to deplete the SV2pg from a solution of SDS-solubilized synaptic vesicles by exhaustive im- munoprecipitation with the anti-SV2 mAb. We are unable to create a solution containing keratan sulfate epitopes in the absence of SV2 (data not shown).
We have also shown that the molecule in I2’I-la- beled electric organ vesicles that contains SV1 and SV2 is part of a complex in the membrane of these vesicles (Fig. IB). This complex consists of four smaller labeled molecules in the molecular weight range of 23,000-45,000. Anti-SVl and anti-SV2 both specifically precipitate this complex. Western blots of these precipitations probed with an antisynaptic vesi- cle serum (Carlson and Kelly, 1980, 1983) demon- strate that many vesicle components are not precipi- tated in this complex. This antiserum was previously shown to contain antibodies to several specific compo- nents of the synaptic vesicle, including the synaptic vesicle proteoglycan (Carlson and Kelly, 1983), p65 or synaptotagmin (Matthew, 198 I), VAT-1 (Linial et al., 1989), and VAMP-1 or synaptobrevin (Trimble et al., 1988). Only the synaptic vesicle proteoglycan is
J . Neuroehem.. Val. 61, No. 1. 1993

417 T. W. SCRANTON ET AL.
stained by this antiserum and not any of the smaller molecular weight proteins of 23,000-45,000. These results point to the existence of a specific membrane complex containing SV2pg and new additional syn- aptic vesicle-specific proteins. The complex is disrupted by denaturing detergents without p-mer- captoethanol, indicating that it is held together by purely noncovalent interactions (Fig. 1 B). Our results contrast with those of Bennett et al. (1992), who ob- served the coprecipitation of a small amount of syn- aptotagmin with SV2 precipitated from rat brain vesi- cles under similar solubilization conditions. This could indicate a species difference in the strength of the interaction between these proteins.
The amino acid sequence of SV2 shows homology with that of several other molecules that have been identified as transporters. The N-terminal sequence surrounding the first six transmembrane regions of the molecule are homologous to several bacterial su- gar transporters (Ballajieh et al., 1992; Feany et al., 1992); the C-terminal region containing the other six membrane-spanning domains is homologous to sev- eral members of the family of plasma membrane neu- rotransmitter transporters (Feany et al., 1992). Using vesicles isolated from electric organ, Parsons and col- leagues have shown that both the SVl and SV2 epi- topes are associated with the receptor for vesamicol, a blocker of acetylcholine transport in these vesicles (Bahr and Parsons, 1992; Bahr et a]., 1992). This strongly suggests that the SV2pg has a transport func- tion in synaptic vesicles. Because SV2pg appears to be present in most, if not all, synapses (Buckley and Kelly, 1985), it may be a common subunit of neuro- transmitter transporters. One of the 23-45-kDa pro- teins associated with the SV2pg may differ between synapse types and determine which neurotransmitter is transported.
SV 1 is a unique epitope on keratan sulfate that ap- pears to be created in the nerve terminal in electric organ. It is not a common antigenic site because it is not present in commercial preparations of keratan sulfate. SVl is also pathway specific, i.e., it is present only on the PI form of the SV2pg in electromotor neurons and not in any significant amounts on the SV2pg elsewhere in the fish brain. Axonal transport studies suggest that the SV 1 epitope originates at the nerve terminal (Caroni et al., 1985). When the elec- tromotor nerve is crushed, the SV 1 epitope accumu- lates sixfold in the region of the nerve distal to the nerve crush compared with the proximal region. Under the same conditions, the SV2 protein shows a distal-to-proximal ratio of only 1.5: 1 .O. Thus, SV 1 ret- rograde transport appears to be greater than the an- terograde transport, whereas the SV2 antigen shows about the same transport in both directions. Consis- tent with this result is the finding that denervation of the electromotor nucleus decreases the amount of SVI in the nucleus about sixfold, whereas the SV2 increases - 1.5-fold. These data suggest the hypothe-
sis that a modification of the keratan sulfate chains on the SV2pg occurs in the nerve terminal and this modi- fication creates the SV 1 epitope. Such a modification could be the covalent attachment of some small mole- cule by a transferase, or simply cleavage of the keratan sulfate chain. The cleavage of a glycosaminoglycan, which creates new epitopes, is known to occur on chondroitin sulfate proteoglycans. When a chondroi- tin sulfate side chain is cleaved with chondroitin ABC lyase, a carbon-carbon double bond is created in the terminal uronic acid residue of the short oligosaccha- ride, which remains attached to the protein core. This oligosaccharide is very immunogenic (Caterson et al., 1987).
Our findings regarding the SV 1 epitope raise some interesting questions. Why should a synaptic vesicle proteoglycan contain a modified keratan sulfate chain that is specific to one group of neurons? Why does this modification occur in the nerve terminal? One possi- bility is that this carbohydrate modification is neces- sary for the transport function of the SV2pg. Removal of keratan sulfate chains from electric organ vesicle SV2pg has been shown to destroy the vesamicol bind- ing activity of these molecules, perhaps through con- formational changes in the SV2 molecule (Bahr el al., 1992). The chain modification could therefore pro- vide a mechanism of modulating neurotransmitter storage.
This modified chain is also known to be present on the nerve terminal surface in the synaptic cleft region (Buckley et al., I983), presumably as a result of synap- tic vesicle exocytosis. What is the function of a neu- ron-specific glycosaminoglycan modification at the nerve terminal? One suggestion might be that this pro- teoglycan is involved in nerve terminal-extracellular matrix adhesive interactions (Schubert, 199 l), much like the integral membrane heparan sulfate proteogly- can syndecan found on epithelial cells (Jalkanen et al., 1985; Gallagher, 1989; Saunderset al., 1989). The SVI keratan sulfate derivative could be a nerve termi- nal-specific ligand for an electric organ lectin. This would be similar to the fucosylated polylactosamine (unsulfated keratan sulfate) chains present on neutro- phils, which are ligands for a specific lectin (selectin) of endothelial cells (Springer and Lasky, 1991). Ifthis speculation is correct, we should find a receptor for this keratan sulfate derivative at the electric organ synapse.
Acknowledgment: We thank Rebecca Cruz for biochemi- cal and immunocytochemical technical assistance and Dr. Bruce Caterson for the monoclonal antibodies to keratan sulfate epitopes (1B4, 2D3). We also thank Regis Kelly, Kathleen Buckley, and Tom Wight for their helpful com- ments. We thank Connie Missimer for editorial help. This work was supported by the W. M. Keck Foundation and a grant from the National Institutes of Health (NS22367) to S. S. Carlson. T. Scranton was supported by NIH Training Grant GM07 1 OX.

SV2 IS A KERATAN SULFATE PROTEOGLYCAN 43
REFERENCES Antonsson P., Heinegard D., and Oldberg A. (1 989) The keratan
sulfate-enriched region of bovine cartilage proteoglycan con- sists of a consecutively repeated hexapeptide motif. J. Biol. Chem 264, 16170-16173.
Archer B. T. Ill, Ozcelik T., Jahn R., Francke U., and Siidhof T. C. (1990) Structures and chromosomal localizations of two hu- man genes encoding synaptobrevins 1 and 2. J. Biol. Chem.
Bahr B. A. and Parsons S. M. ( I 992) Purification of the vesamicol receptor. Biochemistry 31, 5763-5769.
Bahr B. A,, Noremberg K., Rogers G. A., Hicks B. W., and Parsons S. M. (1 992) Linkage of the acetylcholine transporter-vesami- col receptor to proteonlvcan in synaptic vesicles. Biochemistrv
265, 17267-17273.
_ . - . 31,5778-5784:
Baiialieh S. M.. Peterson K.. Shinehal R.. and Scheller R. H. (1992) L , I ~I
SV2, a brain synaptic vesicle protein homologous to bacterial transporters. Science 257, 1271-1273.
Bennett M. K., Calakos N., Kreiner T., and Scheller R. H. (1992) Synaptic vesicle membrane proteins interact to form a multi- meric complex. J. Cell Biol. 116, 761-775.
Blackshear P. J. ( 1984) Systems for polyacrylamide gel electrophore- sis. Methods Enzymol. 104, 237-254.
Block J. A,, Inerot S. E., and Kimura J. H. (1992) Heterogeneity of keratan sulfate substituted on human chondrocytic large pro- teoglycans. J. Biol. Chem. 267, 1245-7252.
Buckley K. and Kelly R. B. (1985) ldentification of a transmem- brane glycoprotein specific for secretory vesicles of neural and endocrine cells. J. Cell Biol. 100, 1284-1294.
Buckley K., Schweitzer E. S., Miljanich G. P., Clift-OGrady L., Kushner P. D., Reichardt L. F., and Kelly R. B. (1983) A synaptic vesicle antigen is restricted to the junctional region of the presynaptic plasma membrane. Proc. Nail. Acad. Sci. USA
Buckley K., Floor E., and Kelly R. B. (1987) Cloning and sequence analysis of cDNA encoding p38, a major synaptic vesicle pro- tein. J. Cell Biol. 105, 2447-2456.
Burnette W. N. (198 I ) “Western blotting”: Electrophoretic transfer of proteins from sodium dodecyl sulphate-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal. Biochem. 112,
Carlson D. M. (1968) Structures and immunological properties of oligosaccharides isolated from pig submaxillary mucins. J. Biol. Chem. 243,616-626.
Carlson S. S. (1989) Synaptic vesicle glycoproteins and proteogly- cans, in Neurohiology of’Glycoconjugates (Margolis R. U. and Margolis R. K., eds), pp. 309-336. Plenum Publishing, New York.
Carlson S. S. and Kelly R. B. (1980) An antiserum specific for cholinergic synaptic vesicles from electric organ. J. Cell Biol.
Carlson S. S. and Kelly R. B. (1983) A highly antigenic proteogly- can-like component of cholinergic synaptic vesicles. J. Biol. Chem. 258, 11082-11091.
Carlson S. S. and Wight T. (1987) Nerve terminal anchorage pro- tein 1 (TAP-I) is a chondroitin sulfate proteoglycan: biochemi- cal and electron microscopic characterization. J. Cell Biol. 105,3075-3086.
Carlson S. S., Wagner J. A., and Kelly R. B. (1978) Purification of synaptic vesicles from elasmobranch electric organ and the use of biophysical criteria to demonstrate purity. Biochemistry 17, 1188-1199.
Carlson S. S., Caroni P., and Kelly R. B. (1986) A nerve terminal anchorage protein from electric organ. J. Cell Biol. 103, 509- 520.
Caroni P., Carlson S. S., Schweitzer E., and Kelly R. B. (1985) Presynaptic neurones may contribute a unique glycoprotein to the extracellular matrix at the synapse. Nature 314, 441-443.
Caterson B., Calabro T., and Hampton A. ( I 987) Monoclonal anti- bodies as probes for elucidating proteoglycan structure and
80,7342-7346.
195-203.
87,98-103.
function, in Biology of Proteoglycans (Wight T. N. and Me- cham R. P., eds), pp. 1-26. Academic Press, Orlando, Florida.
Caterson B., Brooks K., Sattsangi S., Ratcliffe A.. Hardingham T., and Muir H. (1989) Factors affecting the determination of ker- atan sulphate using monoclonal antibodies in immunoassay procedures, in Keratan Sulfate-Chemistuy, Biology, and Chemical Pathology(Grei1ing H. and Scott J. E., eds), pp. 199- 2 14. British Biochemical Society, London.
Cotman C. W. (1974) Isolation of synaptosomal and synaptic plasma membrane fractions. Methods Enzymol. 28,445-452.
Deutsch J. W. and Kelly R. B. (1981) Lipids of synaptic vesicles: relevance to the mechanism of membrane fusion. Biochernis-
Feany M. B., Lee S., Edwards R. H., and Buckley K. M. ( 1 992) The synaptic vesicle protein SV2 is a novel type of transmembrane transporter. Cell 70, 86 1-867.
Fish W. W., Mann K. G., and Tanford C. (1969) The estimation of polypeptide chain molecular weights by gel filtration in 6 M guanidine hydrochloride. J. Biol. Chem. 244,4989-4994.
Fukuda M. N. and Matsumura G. (1976) Endo-/3-galactosidase of Escherichia freundii. J . Biol. Chem 251, 62 18-6225.
Funderburgh W. W. and Conrad G. W. (1990) Isoforms of corneal keratan sulfate proteoglycan. J. Bid. Chem. 265, 8297-8303.
Gallagher J. T. (1 989) The extended family of proteoglycans: social residents of the pericellular zone. Curr. Opin. Cell Biol. 1,
Hell J. W., Maycox P. R., Stadler H., and Jahn R. (1989) Uptake of rat brain synaptic vesicles isolated by a new procedure. EMBO
Huttner W. B., Schiebler W., Greengard P., and De Camilli P. (1983) Synapsin 1 (protein I), a nerve terminal-specific phos- phoprotein. 111. Its association with synaptic vesicles studied in a highly purified synaptic vesicle preparation. J. Cell Bid. 96,
Iwata M. and Carlson S. S. ( 1 99 1) A large chondroitin sulfate base- ment membrane-associated proteoglycan exists as a disulfide- stabilized complex of several proteins. J. Biol. Chem. 266,
Jalkanen M., Nguyen H., Rapraeger A., Kurn N., and Bernfield M. (1985) Heparan sulfate proteoglycans from mouse mammary epithelial cells: localization on the cell surface with a monoclo- nal antibody. J. Cell Biol. 101, 976-984.
Kandel E. and Schwartz J. (1985) Principles ofNetiral Science. El- sevier, New York.
Kjellen L. and Lindahl U. (1991) Proteoglycans: structures and interactions. Annu. Rev. Biochem. 60, 443-475.
Knaus P., Marqueze-Pouey B., Scherer H., and Betz H. (1990) Syn- aptoporin, a novel putative channel protein of synaptic vesi- cles. Neuron 5, 453-462.
Kushner P. D. (1984) A library of monoclonal antibodies to Tor- pedo cholinergic synaptosomes. J. Neurochem. 43, 775-786.
Laemmli U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nafure 227, 680- 685.
Leube R. E., Kaiser P., Seiter A,, Zimbelmann R., Franke W. W., Rehm H., Knaus P., Prior P., Betz H., Reinke H., Beyreuther K., and Wiedenmann B. ( 1 987) Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. EMBO J.
Linial M., Miller K., and Scheller R. H. (1989) VAT-I: an abun- dant membrane protein from Torpedocholinergic synapticves- icles. Neiiron 2, 1265-1273.
Mann K. G. and Fish W. W. (1972) Protein polypeptide chain molecular weights by gel chromatography in guanidinium chlo- ride. Melhods Enzymol. 26, 28-42.
Martin G. R. and Timpl R. ( I 987) Laminin and other basement membrane components. Annu. Rev. Cell Biol. 3, 57-85.
Matthew W. D. (1981) Biochemical studies using monoclonal anti- bodies to neural antigens. Ph.D. thesis, University of Califor- nia, San Francisco, California.
Mehmet H., Scudder P., Tang P. W., Hounsell E. F., Caterson B., and Feizi T. (1986) The antigenic determinants recognized by
try 20, 378-385.
120 1-1 2 18.
J . 7,3023-3029.
1374-1388.
323-333.
6,3261-3268.
J . Neurochem., Vol. 61, No. 1. 1993

44 T. W. SCRANTON ET AL.
three monoclonal antibodies to keratan sulfate involve sul- fated hepta- or larger oligosaccharides of the poly(N-acetyllac- tosamine) series. Eur. J. Biochem. 157, 385-391.
Melrose J. and Ghosh P. ( I 988) The quantitative discrimination of corneal type I, but not skeletal type 11, keratan sulphate in glycosaminoglycan mixtures by using a combination of di- methylene blue and endo-beta-D-galactosidase digestion. Anal. Biochem. 170,293-300.
Nieduszynski I. A., Huckerby T. M., Dickerson J. M., Brown G. M., Tai G. H., and Bayliss M. T. (1990) Structural aspects of skeletal keratan sulphates. Bzochem. Soc. Trans. 18, 792- 793.
Perin M. S., Fried V. A,, Slaughter C. A., and Siidhof T. C. ( I 990) Phospholipid binding by a synaptic vesicle protein homolo- gous to the regulatory region of protein kinase C. Nature 345,
Perin M. S., Johnston P. A., Ozcelik T., Jahn R., Francke U., and Siidhof T. C. ( I 99 1 ) Structural and functional conservation of synaptotagmin (p65). J. Biol. Chem. 266,623-629.
Saunders S., Jalkanen M., O’Farrell S., and Bernfield M. (1989) Molecular cloning of syndecan, an integral membrane proteo- glycan. J. Cell Biol. 108, 1547-1556.
Schaffer W. and Weissmann C. (1973) A rapid, sensitive, and spe- cific method for the determination of protein in dilute solu- tions. Anal. Biochem. 56, 502-5 14.
Schubert D. ( 199 1) The possible role ofadhesion in synaptic modifi- cation. Trends Neurosci. 14, 127-130.
Scopes R. K. (1 987) Protein Purification. Springer-Verlag Inc., New York.
Siege1 L. M. and Monty K. L. (1966) Determination of molecular weights and frictional ratios of proteins in impure systems by use of gel filtration and density gradient centrifugation. Bio- chim. Biophys. Acta 112, 346-362.
Springer T. A. and Lasky L. A. (1991) Sticky sugar for selectins. Nature349, 196-197.
Stadler H. and Dowe G. H. C. (1982) Identification of a heparan
260-263.
sulfate-containing proteoglycan as a specific core component of cholinergic synaptic vesicles from Torpedo marmorata. EMBO J. 1, 1381-1384.
Siidhof T. C. and Jahn R. (199 I ) A synaptic vesicle protein with a novel cytoplasmic domain and four transmembrane regions. Neuron 6,665-677.
Siidhof T. C., Lottspeich F., Greengard P., Mehl E., and Jahn R. (1987) Proteins of synaptic vesicles involved in exocytosis and membrane recycling. Science 238, I 142-1 144.
Tai G. H., Brown G. M., Morris H. G., Huckerby T. M., and Nie- duszynski I. A. (1991) Fucose content in keratan sulphates from bovine articular cartilage. Biochem. J. 273, 307-310.
Tang P. W., Scudder P., Mehmet H., Hounsell E. F., and Fiezi T. (1986) Sulphate groups are involved in the antigenicity of kera- tan sulphate and mask i antigen expression on their po1y-N- acetyllactosamine backbones. Eur. J. Biochem. 160,537-545.
Tashiro T. and Stadler H. (1978) Chemical composition of cholin- ergic synaptic vesicles from Torpedo marmorata based on im- proved purification. Eur. J . Biochem. 90,479-487.
Trimble W. S., Cowan D. M., and Scheller R. H. (1988) VAMP-1: a synaptic vesicle-associated integral membrane protein. Proc. Nail. Acad. Sci. USA 85, 4538-4542.
van Holde K. E. (1985) Physical Biochemistry. Prentice-Hall Inc., Englewood Cliffs, New Jersey.
Vogel K. C. and Evanko S. P. ( 1987) Proteoglycans of fetal bovine tendon. J. Biol. Chem. 262, 13607-13613.
Volknandt W. and Zimmermann H. ( I 990) Identical properties of transmembrane synaptic vesicle protein M, 100,000 in Tor- pedo and M, 86,000 in bovine brain. Neurochem. Int. 16,539- 541.
Wasteson A. (197 1) A method for the determination ofthe molecu- lar weight and molecular weight distribution of chondroitin sulfate. J . Chromatogr. 59, 87-97.
Wendland B., Miller K. C., Schilling J., and Scheller R. H. (1991) Differential expression of the p65 gene family. Neuron 6,993- 1007.
J Neurochem., Vol. 61, No. I , 1993
Related Documents











