Molecular Cell, Vol. 17, 121–135, January 7, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2004.11.041 The Structure of the Human Adenovirus 2 Penton ton base (polypeptide III) and the externally projecting Chloe Zubieta, 1 Guy Schoehn, 1,2,3 trimeric fiber protein (polypeptide IV), forms the 12 verti- Jadwiga Chroboczek, 2 and Stephen Cusack 1, * ces of the particle. Additionally, a number of proteins 1 European Molecular Biology Laboratory located on the internal or external surface of the capsid, Grenoble Outstation, BP 181 such as IIIa, VI, VIII, and IX, have been implicated in 38042 Grenoble Cedex 9 cementing the hexon-hexon and hexon-penton interac- France tions, (Furcinitti et al., 1989; Stewart et al., 1991; Stewart 2 Institut de Biologie Structurale J.P. Ebel et al., 1993). 38027 Grenoble Cedex 1 The capsid proteins fiber and penton base are the France major players in adenovirus cell entry. Initial attachment 3 Laboratoire de Virologie Mole ´ culaire of the virus occurs through the binding of the distal EA 2939 C-terminal knob domain of the fiber to primary cellular Universite ´ Joseph Fourier receptors. Coxsackie and adenovirus receptor (CAR), Grenoble Cedex 9 an integral membrane protein now known to be part of France the tight junction in epithelial cells (Cohen et al., 2001; Walters et al., 2002), is the primary receptor for adenovi- rus subgroups A and C–F (Bergelson et al., 1997; Tomko Summary et al., 1997; Roelvink et al., 1998). Recently, CD46 has been identified as the receptor for a number of subgroup The adenovirus penton, a noncovalent complex of the B serotypes such as Ad3, Ad11, and Ad33, as well as pentameric penton base and trimeric fiber proteins, Ad37 of subgroup D (Gaggar et al., 2003; Segerman et comprises the vertices of the adenovirus capsid and al., 2003; Sirena et al., 2004; Wu et al., 2004). Addition- contains all necessary components for viral attach- ally, in Ad37, sialic acid may also play a role as a receptor ment and internalization. The 3.3 A ˚ resolution crystal (Burmeister et al., 2004). structure of human adenovirus 2 (hAd2) penton base After initial cell binding, v 3 or v 5 integrins serve as shows that the monomer has a basal jellyroll domain coreceptors by binding to a highly conserved RGD motif and a distal irregular domain formed by two long inser- in the penton base, an interaction that is essential for tions, a similar topology to the adenovirus hexon. The virus internalization by receptor-mediated endocytosis Arg-Gly-Asp (RGD) motif, required for interactions (Wickham et al., 1993). Clustering of integrins induced with cellular integrins, occurs on a flexible surface by this interaction activates signaling pathways that pro- loop. The complex of penton base with bound N-ter- mote rapid internalization into clathrin-coated pits and minal fiber peptide, determined at 3.5 A ˚ resolution, endosomes (Li et al., 1998). Integrin binding is accompa- shows that the universal fiber motif FNPVYPY binds nied by fiber shedding from the capsid (Greber et al., at the interface of adjacent penton base monomers 1993; Nakano et al., 2000). Both penton base and fiber, and results in a localized structural rearrangement in as well as adenovirus protease, have been implicated the insertion domain of the penton base. These results in the escape of the virus from the endosome as the pH give insight into the structure and assembly of the lowers, although the details of this process are unclear adenovirus capsid and will be of use for gene-ther- (Seth et al., 1985; Blumenthal et al., 1986; Nemerow apy applications. and Stewart, 1999; Medina-Kauwe et al., 2001; Medina- Kauwe, 2003). After internalization, partially disassem- Introduction bled capsids are transported along microtubules to the perinuclear envelope where further disintegration of Human adenoviruses are nonenveloped DNA viruses the capsid finally allows import of the viral DNA into the that can cause respiratory, ocular, and enteric disease. nucleus accompanied by protein VI. The critical impor- The approximately fifty known human serotypes are di- tance of fiber and penton base in cell infection is shown vided amongst six subgroups (A–F) based on such fac- by the fact that dodecahedral particles comprising only tors as crossimmunoreactivity, agglutination, nucleic these two viral proteins efficiently enter cells and accu- acid homology, and genetic organization (Bailey and mulate at the periphery of the nucleus, presumably by Mautner, 1994). Adenoviruses have also been isolated using the same pathway as the complete virus (Fender from many other mammals, birds, reptiles, amphibia, et al., 1997). Unlike all other human adenoviruses, sub- and fish (Benko and Harrach, 2003). All adenoviruses group F enteric adenoviruses Ad40 and Ad41 neither exhibit the same overall capsid architecture. The T have an RGD motif nor use integrins for cell infection 25 icosahedral capsid is approximately 1000 A ˚ in diame- (Albinsson and Kidd, 1999). ter and consists primarily of three different polypeptides, Structural studies of intact adenoviruses, individual hexon, penton base, and fiber (van Oostrum and Burnett, capsid proteins, and complexes with receptors by X-ray 1985). Each facet of the capsid contains 12 copies of crystallography and electron microscopy (EM) have pro- the trimeric hexon protein, a major viral antigen. A non- vided a detailed understanding of many aspects of ade- covalent complex of two proteins, the pentameric pen- novirus capsid structure, assembly, and virus-host cell interactions. Cryo-electron microscope (cryo-EM) three- dimensional (3 D) reconstructions of the entire adenovi- *Correspondence: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Cell, Vol. 17, 121–135, January 7, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2004.11.041
The Structure of the Human Adenovirus 2 Penton
ton base (polypeptide III) and the externally projectingChloe Zubieta,1 Guy Schoehn,1,2,3
trimeric fiber protein (polypeptide IV), forms the 12 verti-Jadwiga Chroboczek,2 and Stephen Cusack1,*ces of the particle. Additionally, a number of proteins1European Molecular Biology Laboratorylocated on the internal or external surface of the capsid,Grenoble Outstation, BP 181such as IIIa, VI, VIII, and IX, have been implicated in38042 Grenoble Cedex 9cementing the hexon-hexon and hexon-penton interac-Francetions, (Furcinitti et al., 1989; Stewart et al., 1991; Stewart2 Institut de Biologie Structurale J.P. Ebelet al., 1993).38027 Grenoble Cedex 1
The capsid proteins fiber and penton base are theFrancemajor players in adenovirus cell entry. Initial attachment3 Laboratoire de Virologie Moleculaireof the virus occurs through the binding of the distalEA 2939C-terminal knob domain of the fiber to primary cellularUniversite Joseph Fourierreceptors. Coxsackie and adenovirus receptor (CAR),Grenoble Cedex 9an integral membrane protein now known to be part ofFrancethe tight junction in epithelial cells (Cohen et al., 2001;Walters et al., 2002), is the primary receptor for adenovi-rus subgroups A and C–F (Bergelson et al., 1997; TomkoSummaryet al., 1997; Roelvink et al., 1998). Recently, CD46 hasbeen identified as the receptor for a number of subgroupThe adenovirus penton, a noncovalent complex of theB serotypes such as Ad3, Ad11, and Ad33, as well aspentameric penton base and trimeric fiber proteins,Ad37 of subgroup D (Gaggar et al., 2003; Segerman etcomprises the vertices of the adenovirus capsid andal., 2003; Sirena et al., 2004; Wu et al., 2004). Addition-contains all necessary components for viral attach-ally, in Ad37, sialic acid may also play a role as a receptorment and internalization. The 3.3 A resolution crystal(Burmeister et al., 2004).structure of human adenovirus 2 (hAd2) penton base
After initial cell binding, �v�3 or �v�5 integrins serve asshows that the monomer has a basal jellyroll domaincoreceptors by binding to a highly conserved RGD motifand a distal irregular domain formed by two long inser-in the penton base, an interaction that is essential fortions, a similar topology to the adenovirus hexon. Thevirus internalization by receptor-mediated endocytosisArg-Gly-Asp (RGD) motif, required for interactions(Wickham et al., 1993). Clustering of integrins inducedwith cellular integrins, occurs on a flexible surfaceby this interaction activates signaling pathways that pro-loop. The complex of penton base with bound N-ter-mote rapid internalization into clathrin-coated pits andminal fiber peptide, determined at 3.5 A resolution,endosomes (Li et al., 1998). Integrin binding is accompa-shows that the universal fiber motif FNPVYPY bindsnied by fiber shedding from the capsid (Greber et al.,at the interface of adjacent penton base monomers1993; Nakano et al., 2000). Both penton base and fiber,and results in a localized structural rearrangement inas well as adenovirus protease, have been implicatedthe insertion domain of the penton base. These resultsin the escape of the virus from the endosome as the pHgive insight into the structure and assembly of thelowers, although the details of this process are unclearadenovirus capsid and will be of use for gene-ther-(Seth et al., 1985; Blumenthal et al., 1986; Nemerowapy applications.and Stewart, 1999; Medina-Kauwe et al., 2001; Medina-Kauwe, 2003). After internalization, partially disassem-Introductionbled capsids are transported along microtubules tothe perinuclear envelope where further disintegration of
Human adenoviruses are nonenveloped DNA viruses the capsid finally allows import of the viral DNA into thethat can cause respiratory, ocular, and enteric disease. nucleus accompanied by protein VI. The critical impor-The approximately fifty known human serotypes are di- tance of fiber and penton base in cell infection is shownvided amongst six subgroups (A–F) based on such fac- by the fact that dodecahedral particles comprising onlytors as crossimmunoreactivity, agglutination, nucleic these two viral proteins efficiently enter cells and accu-acid homology, and genetic organization (Bailey and mulate at the periphery of the nucleus, presumably byMautner, 1994). Adenoviruses have also been isolated using the same pathway as the complete virus (Fenderfrom many other mammals, birds, reptiles, amphibia, et al., 1997). Unlike all other human adenoviruses, sub-and fish (Benko and Harrach, 2003). All adenoviruses group F enteric adenoviruses Ad40 and Ad41 neitherexhibit the same overall capsid architecture. The T � have an RGD motif nor use integrins for cell infection25 icosahedral capsid is approximately 1000 A in diame- (Albinsson and Kidd, 1999).ter and consists primarily of three different polypeptides, Structural studies of intact adenoviruses, individualhexon, penton base, and fiber (van Oostrum and Burnett, capsid proteins, and complexes with receptors by X-ray1985). Each facet of the capsid contains 12 copies of crystallography and electron microscopy (EM) have pro-the trimeric hexon protein, a major viral antigen. A non- vided a detailed understanding of many aspects of ade-covalent complex of two proteins, the pentameric pen- novirus capsid structure, assembly, and virus-host cell
interactions. Cryo-electron microscope (cryo-EM) three-dimensional (3 D) reconstructions of the entire adenovi-*Correspondence: [email protected]
Molecular Cell122
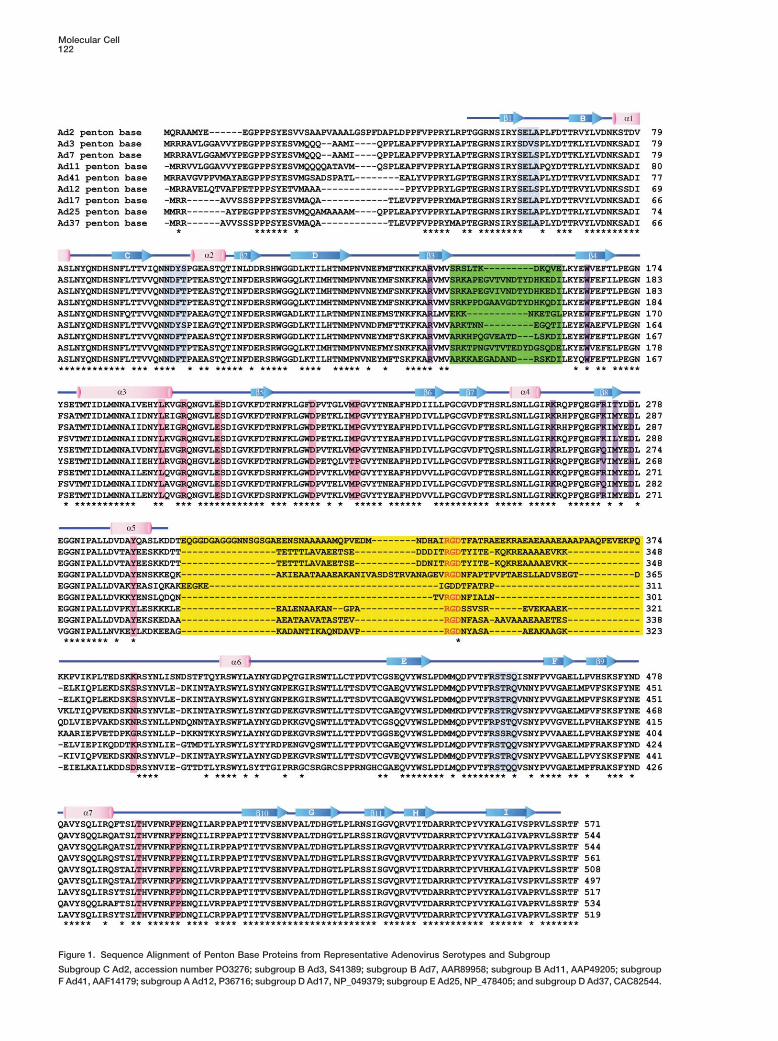
Figure 1. Sequence Alignment of Penton Base Proteins from Representative Adenovirus Serotypes and Subgroup
Subgroup C Ad2, accession number PO3276; subgroup B Ad3, S41389; subgroup B Ad7, AAR89958; subgroup B Ad11, AAP49205; subgroupF Ad41, AAF14179; subgroup A Ad12, P36716; subgroup D Ad17, NP_049379; subgroup E Ad25, NP_478405; and subgroup D Ad37, CAC82544.

Structure of the Adenovirus Penton123
rus have enabled location of the major and some of the purified. Because this protein was easily degraded,N-terminal sequencing was used to identify proteolyticminor capsid proteins (Stewart et al., 1991; Stewart et
al., 1993). Atomic-resolution structural data are available products. An N-terminal truncation (49-TGGR…) wasidentified as a stable species and used for all subse-for the Ad5 hexon (Rux and Burnett, 2000); the fiber
head domains of Ad5 (Xia et al., 1994), Ad12 (Bewley et quent structural analysis.All constructs of adenovirus 2 penton base (Ad2pb)al., 1999), Ad3 (Durmort et al., 2001), Ad37 (Burmeister
et al., 2004); and the Ad2 fiber head and part of its shaft formed isolated pentamers in solution as shown by neg-ative stain electron microscopy (Figure 2A). Three crys-(van Raaij et al., 1999). The only major capsid protein
without a high-resolution structure has hitherto been tal forms of Ad2pb were obtained but only those inspacegroup C2, diffracting to 3.3 A resolution (Table 1),the penton base protein. Negative stain electron micro-
scope images of the adenovirus penton show that the could be reproducibly grown. The unit cell dimensions,a � 436.0 A, b � 300.2 A, c � 420.6 A, and � � 104.4�,penton base pentamer forms a bucket-shaped structure
from the center of which the fiber shaft emerges (Ruigrok suggested a large assembly in the unit cell. Indeed, itwas unexpectedly found that under crystallization con-et al., 1990). More detailed information on the penton
base is available from cryo-EM, firstly on dodecahedral ditions, the pentamers had associated into regular do-decahedral particles, as deduced from the characteristicparticles composed of 12 Ad3 penton bases or complete
pentons (Schoehn et al., 1996), and secondly in the con- icosahedral symmetry observed in the self-rotationfunction and EM of dissolved crystals and Ad2pb aftertext of the complete virus, including a complex of Ad12
and a soluble extracellular �v�5 integrin construct (Chiu preincubation in crystallization buffer (Figure 2A). For-mation of dodecahedra of pentons has been observedet al., 1999).
The penton base sequence is exceptionally highly in vivo in infected cells (Norrby, 1964) and with recombi-nant protein (Fender et al., 1997) for other adenovirusconserved even between human and other animal ade-
noviruses (typically 70% homology between any two serotypes, notably Ad3, but not previously for Ad2pb.The Ad2pb/fiber peptide complex was formed by coin-penton bases, Figure 1). It has a peptide chain length
of between 470 and 570 residues with the difference cubation with a 21 amino acid peptide correspondingto the N terminus of the Ad2 fiber protein and crystallizedalmost exclusively being due to a hypervariable loop
region of variable length in the middle of the chain, which in similar conditions with a similar unit cell (Table 1). Acryo-EM model of the Ad3 dodecahedron (Schoehn etin most cases contains the RGD motif (Figure 1). Cryo-
EM structures of the Ad2, Ad3, and Ad12 penton base al., 1996), extended to 15 A resolution, was used tosolve the Ad2pb dodecahedron structure by molecularshow five external protrusions that have been identified
by RGD specific monoclonal antibody binding as the replacement. Subsequent phase extension with 60-foldnoncrystallographic symmetry averaging (one dodeca-flexible, hypervariable RGD loop (Schoehn et al., 1996,
Stewart et al., 1997). Up to five integrins can bind to the hedron per asymmetric unit) yielded a readily interpret-able experimental electron density map at 3.3 A reso-penton base leading to a ring-like structure that may
be important for coreceptor clustering and endocytosis lution.(Chiu et al., 1999).
Given the importance of the penton base in many Overall StructureThe Ad2pb monomer folds into two domains, a loweraspects of adenovirus biology, we set out to determine
the atomic structure of the protein. Here, we report the jellyroll-motif domain facing the capsid interior (40% ofthe residues) and an upper insertion domain facing the3.3 A crystal structure of human Ad2 penton base
(Ad2pb) in a dodecahedral form, which provides the last solvent exposed exterior (60% of the residues) (Figure3A). The monomer is elongated and measures approxi-remaining major piece of the adenovirus capsid puzzle.
We also describe the 3.5 A crystal structure of the penton mately 100 A from the base of the jellyroll domain to thetop of the insertion domain (excluding the disorderedbase in complex with the interacting N-terminal fiber
peptide. This gives insight into the symmetry mismatch RGD loop). The jellyroll motif, common to many viralcapsid proteins, consists of two, four-stranded antipar-implicated in the assembly of the penton from pentam-
eric penton base and trimeric fiber. Finally, this structural allel � sheets, which pack against each other forming a�-barrel (Chelvanayagam et al., 1992). The � strands ofinformation will aid in studies of other less well-under-
stood roles of the penton base, such as in endocytosis, the jellyroll are conventionally labeled CHEF and BIDG(Richardson, 1981), with adjacent strands B and I beingand will potentially allow rational engineering of the pen-
ton base for applications in gene therapy. formed by the opposite extremities of the polypeptide(Figures 3A and 6). Thus, residues 49 and 569 are within�12 A of each other at the base of the protein. TheResults and Discussionlocation of the N-terminal 49 residues absent from thestructure is of considerable importance because thisStructure Determination
A full-length construct of the 571 residue Ad2pb poly- region contains two highly conserved xPPxY motifs, 11-PPPSY and 41-VPPRY, in Ad2pb. The first of thesepeptide was expressed in the baculovirus system and
The hypervariable RGD loop region (yellow), the variable loop (light green), residues involved in dodecahedron formation (blue), fiber peptidebinding residues (pink), and residues implicated in detergent molecule binding (purple) are highlighted. Secondary structure elements aredenoted above the sequence by blue arrows (� strands) and pink cylinders (� helices). Stars below the sequences denote completely conservedresidues. The RGD sequence is highlighted in red.
Molecular Cell124
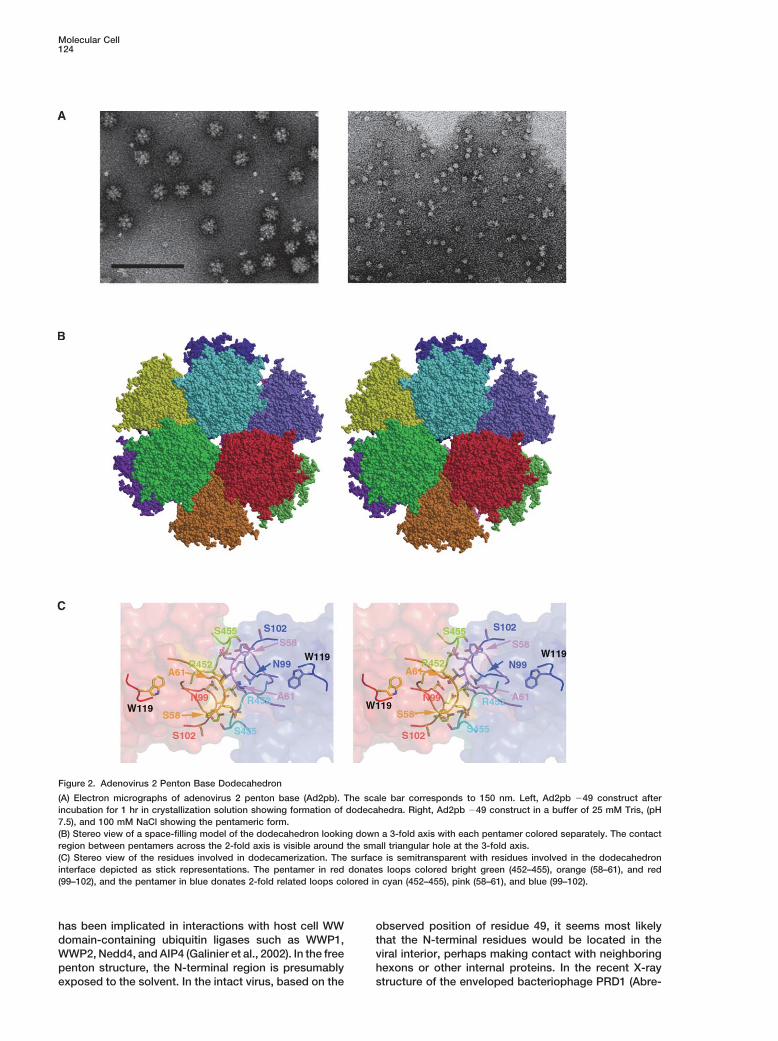
Figure 2. Adenovirus 2 Penton Base Dodecahedron
(A) Electron micrographs of adenovirus 2 penton base (Ad2pb). The scale bar corresponds to 150 nm. Left, Ad2pb �49 construct afterincubation for 1 hr in crystallization solution showing formation of dodecahedra. Right, Ad2pb �49 construct in a buffer of 25 mM Tris, (pH7.5), and 100 mM NaCl showing the pentameric form.(B) Stereo view of a space-filling model of the dodecahedron looking down a 3-fold axis with each pentamer colored separately. The contactregion between pentamers across the 2-fold axis is visible around the small triangular hole at the 3-fold axis.(C) Stereo view of the residues involved in dodecamerization. The surface is semitransparent with residues involved in the dodecahedroninterface depicted as stick representations. The pentamer in red donates loops colored bright green (452–455), orange (58–61), and red(99–102), and the pentamer in blue donates 2-fold related loops colored in cyan (452–455), pink (58–61), and blue (99–102).
has been implicated in interactions with host cell WW observed position of residue 49, it seems most likelythat the N-terminal residues would be located in thedomain-containing ubiquitin ligases such as WWP1,
WWP2, Nedd4, and AIP4 (Galinier et al., 2002). In the free viral interior, perhaps making contact with neighboringhexons or other internal proteins. In the recent X-raypenton structure, the N-terminal region is presumably
exposed to the solvent. In the intact virus, based on the structure of the enveloped bacteriophage PRD1 (Abre-

Structure of the Adenovirus Penton125
Table 1. Data Collection, Averaging, and Refinement Statistics
Ad2 Penton Base Ad2 Penton Base DodecahedronCrystal Contents Dodecahedron Cocrystallized with Fiber Peptide
Space group C2 C2Cell dimensions (A) a � 436.0, b � 300.2, c � 420.6, � � 104.4� a � 438.4, � 299.8, c � 419.6, � � 103.2�
ObservationsTotal measured reflections 657,395 761,440Unique reflections 447,000 354,521
Resolution (A) 20–3.3 30–3.5Completenessa 0.57 (0.22) 0.54 (0.29)Rmerge
a,b 0.21 (0.47) 0.19 (0.45)Average measurement redundancy 1.5 2.1NCS redundancy 60-fold 60-foldInitial NCS averaging statisticsc
Correlationd/Rface 0.87/0.25 0.68/0.37Rigid-body refinement, Rcryst changee 0.32→0.29 0.41→0.32Final NCS averaging statisticsf
Correlationd/Rface 0.90/0.22 0.91/0.20Model refinement Rcryst/Rfree (%)g 31.1/31.1 30.5/30.3Model geometry
RMSD bond lengths (A) 0.012 0.014RMSD bond angles (�) 1.89 1.83
Ramachandran plotFavored � additional regions 95.1% 93.9%
a Values in parentheses refer to the highest resolution shell.b Rmerge � �h�i [Ihi � (Ih)]/�h(Ih), where Ih,i is the ith observation of a symmetry equivalent of reflection h.c Averaging with RAVE to high-resolution limit with strict icosahedral symmetry.d Correlation between the densities of all NCS related points. Rfac � �|Fobs � Fmap|/�Fobs where Fmap is Fourier coefficient of back-transformedaveraged map.e Rigid-body refinement of 60 individual monomers with CNS over resolution range 20–5 A.f Averaging with RAVE to high-resolution limit with relaxed icosahedral symmetry as defined by rigid body refinement e.g Strict NCS was applied with matrices found by rigid-body refinement e. Rcryst � �|Fobs � Fcalc|/�Fobs, where summation is over data used in therefinement. Rfree was calculated by using 2.7% of the observed reflections excluded from refinement. Excluded data were selected from thinshells to minimize correlation with noncrystallographic symmetry related reflections.
scia et al., 2004), which has striking structural similarities first insertion (Figures 1, 3A, and 3C). In Ad2pb, thisloop comprises residues 142–169 and forms a flexibleto adenovirus (Benson et al., 1999), the N-terminal 13
residues of the protein P31, equivalent to the penton � ribbon projecting into the solvent. In other serotypesthe variable loop can be up to ten residues longer (Figurebase, interact with the peripentonal P3 proteins (equiva-
lent to hexon). 1), for instance in the Ad3 penton base where it is promi-nent in the cryo-EM structure (Schoehn et al., 1996).The second, distal domain of Ad2pb has an ordered
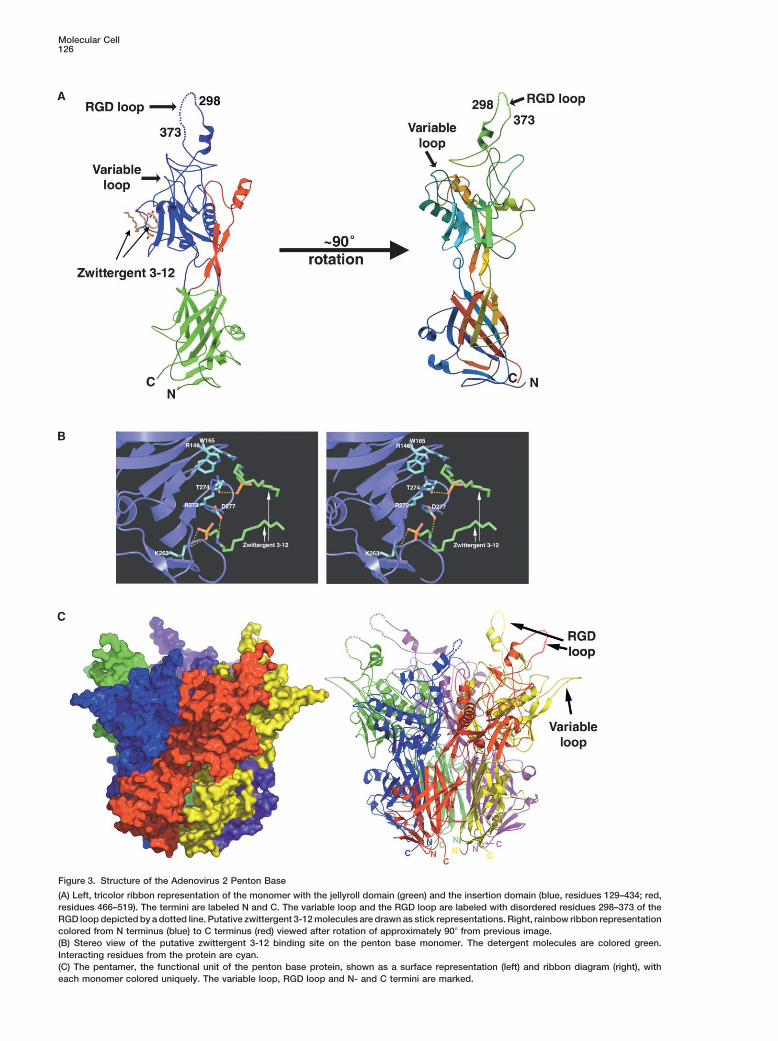
but irregular fold formed by two long insertions, one The function of this variable loop is unknown, but it ispotentially a site where additional sequences could becomprising residues 129–436 between jellyroll strands
D and E and the other formed by residues 468–521 engineered into the structure for gene therapy applica-tions (see Conclusions).between strands F and G (Figure 3A). The first insertion
contains the hypervariable RGD loop region that exhibitsgreat variability in sequence and length amongst adeno- Pentamerization
Ad2pb forms an intimate pentamer that buries approxi-virus serotypes. The RGD loop of Ad2pb consists of�80 residues and is glycine and alanine rich (Figure 1). mately 26% of the total surface area of each monomer
(Figure 2C). The stability gained in pentamer formationIt is clearly flexible because no electron density is seenfor residues 298–375 in NCS averaged maps. At a unique is due mainly to the burying of hydrophobic surface.
Only two salt bridges are formed between adjacentcrystal contact, however, about 15 residues of the RGDloop are visible in one monomer in an unaveraged map monomers upon pentamerization, Arg420-Glu172* and
Arg197-Asp220* (asterisk indicates the second subunit).comprising an � helix followed by a turn and � strand.However, a sequence assignment cannot be made with A ring of jellyroll domains forms the compact base of
the pentamer, and the insertion domains create a wider,certainty, as the density is not continuously connectedto the bulk of the protein. Although this region has rea- � helical-lined top, giving an overall shape resembling a
tulip (but with five “petals” not six). Because the insertionsonably clear electron density, the position relative tothe rest of the penton base is most likely an artifact of domains are tilted relative to the jellyroll domain, the
monomers appear to wrap around each other with athe crystal packing. On the other hand, consistent withEM studies, this observation shows that although the right-handed twist. Interestingly, in contrast to the pen-
ton base, the hexon is wider at the base and narrowerlong RGD loop is flexible (Stewart et al., 1997, Chiu et al.,1999), it may possess at least some structured regions. at the top, suggesting a complementarity of shape be-
tween the two proteins that is most likely important forA second surface loop variable between serotypes,and hence denoted the variable loop, also occurs in the capsid assembly and stability.
Molecular Cell126
Figure 3. Structure of the Adenovirus 2 Penton Base
(A) Left, tricolor ribbon representation of the monomer with the jellyroll domain (green) and the insertion domain (blue, residues 129–434; red,residues 466–519). The termini are labeled N and C. The variable loop and the RGD loop are labeled with disordered residues 298–373 of theRGD loop depicted by a dotted line. Putative zwittergent 3-12 molecules are drawn as stick representations. Right, rainbow ribbon representationcolored from N terminus (blue) to C terminus (red) viewed after rotation of approximately 90� from previous image.(B) Stereo view of the putative zwittergent 3-12 binding site on the penton base monomer. The detergent molecules are colored green.Interacting residues from the protein are cyan.(C) The pentamer, the functional unit of the penton base protein, shown as a surface representation (left) and ribbon diagram (right), witheach monomer colored uniquely. The variable loop, RGD loop and N- and C termini are marked.

Structure of the Adenovirus Penton127
A pore occurs along the 5-fold axis of the pentamer the second molecule forms a salt bridge with Lys263,and the ammonio group is positioned within hydrogenwith diameter 15–20 A at the base and tapering to 5–10 A
at the top of the insertion domain. The top, narrow part bonding distance of Asp277 (Figure 3B).It has recently been found that the Ad3 penton baseof the pore is predominantly hydrophobic, apart from
Asp478, and lined by bulky residues Phe489, Leu485, dodecahedra, but not Ad3 virions, selectively bind hepa-rin and heparan sulfate with high affinity and that thisTyr482, Tyr476, and Phe475 coming from helix �7 and
the preceding strand �9 of each monomer of the inser- interaction at the cell surface can promote internaliza-tion (Vives et al., 2004). The binding of detergent mole-tion domain. Notably, fiber binding modifies this region
(see below). The five side chains of Arg-473 point into the cules in the Ad2pb structures could mimic this interac-tion due to the resemblance of the sulphonate-containingcentral pore forming a tight constriction at the bottom
of the insertion domain. Beneath this, the lower, wider moiety of zwittergent 3-12 to the sulfate groups in hepa-rin and heparan. Obstruction of these sites due to thesection of the pore formed by His471 from helix �8 and
residues from � strands I, G, and D from the jellyroll presence of neighboring hexons might explain the in-ability of the Ad3 virus itself to use heparan sulphatesdomain is much more hydrophilic with residues Asp65,
Asp122, Asp253, Lys124, Arg564, and Glu518 and two as receptors (Vives et al., 2004).These observations are interesting in the light of thesets of salt bridges formed by Asp122-Arg564 and
Lys124-Glu525 exposed to the solvent-filled pore. The little understood but critical activity of the penton basein pH-dependent lipid binding and endocytosis (Medina-possibilities of inserting the fiber protein into the pore
or exposing to the surface the N terminus of the penton Kauwe, 2003). Mutating the conserved Trp165, to whichwe find one detergent molecule binding, to histidinebase through the pore are excluded, as it is too narrow
for a polypeptide chain to be threaded all the way along. resulted in penton bases that were severely deficient incell attachment and somewhat in internalization, consis-tent with the idea that Trp165 may be involved in lipidDodecahedron Formationbinding (Karayan et al., 1997). Clearly, further studies ofThe Ad2 dodecahedron is a highly regular particle ofthis putative lipid binding site, including the effect ofdiameter 280 A with an internal spherical cavity of diame-pH, are required.ter about 80 A (Figures 2A and 2B). The particle is formed
by interactions between three loop regions in the jellyrolldomain, which, however, do not form an extensive inter- Fiber Bindingface. Residues 99–102 (NDYS) and 58–61 (SELA) from The penton complex is formed by the noncovalent bind-one monomer and 452–455 (RSTS) from an adjacent ing of the N-terminal extremity of the fiber polypeptidemonomer in the same pentamer (bold type indicates with the penton base top surface (Devaux et al., 1987).residue conservation, Figure 1) interact with the corre- The penton is very stable and can be dissociated onlysponding loops from a neighboring pentamer in the do- by treatment with chaotropic agents at elevated temper-decahedron about the icosahedral 2-fold axis (Figure 2C). ature (Boudin and Boulanger, 1982). There has been
Under normal conditions, Ad2pb does not spontane- much speculation about the nature and significance ofously form dodecahedra, unlike the penton base of Ad3. the symmetry mismatch between the trimeric fiber andAs monitored by negative stain EM, Ad2pb dodecahedra the pentameric base, which we can now largely resolve.formed only upon addition of the crystallization buffer, A peptide corresponding to the 21 N-terminal residueswhich contains both 1.5 M ammonium sulfate and 10% of adenovirus 2 fiber protein (1-MKRARPSEDTFNPVYdioxane (Figure 2A). Thus, changes in the solvent envi- PYDTEC) was cocrystallized with Ad2pb. The 3.5 A aver-ronment are sufficient to favor association of pentamers aged electron density map gave unambiguous densityinto dodecahedra. Because there are no residues in for a central 11 residue portion of the fiber tail peptide,the three loop areas that are exclusive to those penton but not the preceding basic nuclear location signal (Fig-bases that spontaneously form dodecahedra (Figure 1), ure 4). The observed residues, 10-TFNPVYPYDTE, in-it is difficult to identify the key residues except by further clude the FNPVYPY motif extremely highly conservedmutational analysis. amongst fiber proteins in diverse adenovirus serotypes
(Figure 4D). The fiber peptide lies in a groove on the topsurface of the protein formed between two adjacentDetergent Binding
Ad2pb and the Ad2pb/fiber peptide complex were crys- monomers and resulting from a conformational changein helix �7 of the insertion domain. Fiber peptide bindingtallized in the presence of zwittergent 3-12 (3-dodecyl-
dimethyl ammonio-propane-1-sulphonate), a zwitter- requires this helix to kink by almost 45� between resi-dues Gln482 and Ile484, and additionally, the loop regionionic detergent. This detergent additive was required
for reproducible growth of the high resolution C2 crystal C-terminal to helix �7 is rearranged (Figure 5). Structuralchanges in the penton base are restricted to a 23-resi-form. Two unexplained strong peaks in the averaged
map for Ad2pb were assigned to the sulphonate and due stretch from residues 482–505, indicating that pep-tide binding does not have global repercussions.ammonio head group of two putative detergent mole-
cules with electron density extending to two or three It has been previously proposed that the peptide 253-SRLSNLLG is the fiber binding site (Hong and Bou-carbons of the aliphatic tail (Figures 3A and 3B). The
detergent molecules bind to solvent exposed basic langer, 1995). The crystal structure shows that this pep-tide forms a turn and short, largely buried helix �4 (Figurepatches on the insertion domain. The sulphonate head
group of one putative detergent molecule is positioned 1) that is at least 25 A from the peptide and is thereforenot part of the fiber binding site. In reality, the peptidein a hydrophilic pocket formed by Arg272, Arg146,
Thr274, and the nitrogen of Trp165. The sulphonate of binding site is multipartite and distributed over adjacent
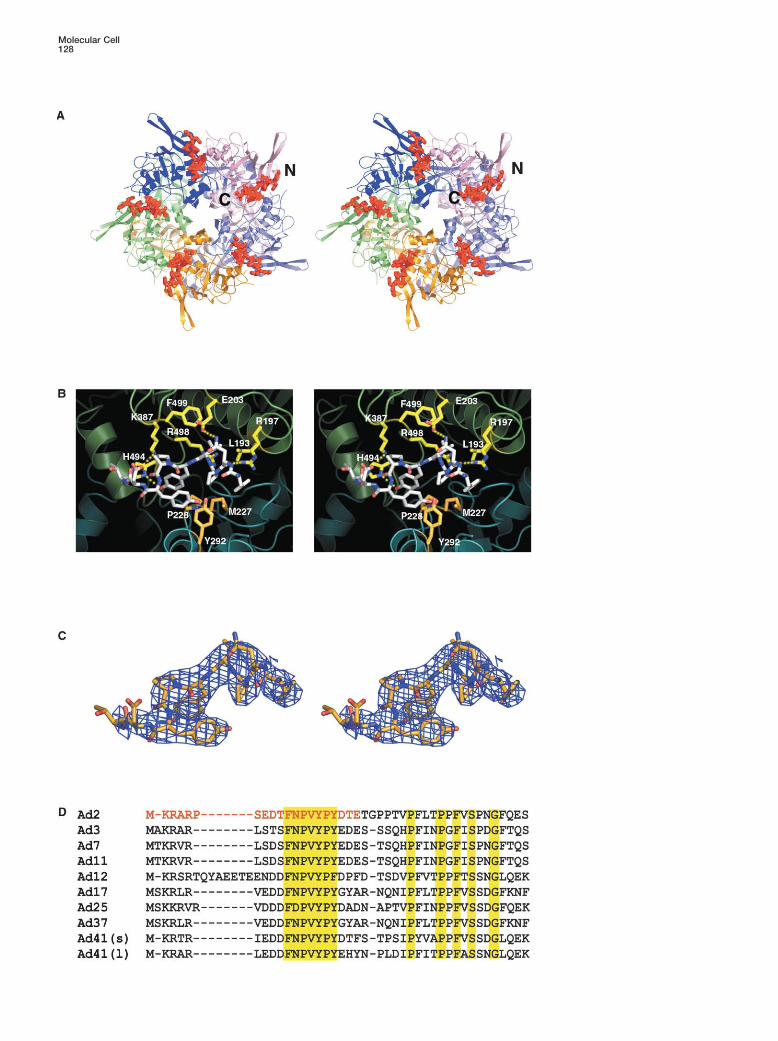
Molecular Cell128

Structure of the Adenovirus Penton129
monomers. It is formed by helices �7 and �3 and strand is consistent with observations from EM showing thefiber shaft emerging rigidly from the center of the penton�5 at the interface between adjacent monomers of the
pentamer (Figure 4B). The monomers in turn are con- base (Chroboczek et al., 1995; Wu et al., 2003). Thereare two possible arrangements; either the three fibernected in this region by a salt bridge between Asp220
(monomer B) and Arg197 (monomer A), which is also tails occupy three successive sites 72� apart, or two arein neighboring sites and one is separated by 144� as inpresent in the native structure. Hydrophobic residues
Leu193, Phe499, and the aliphatic portion of Arg498 Figure 5C. The latter solution is most likely becauseit approximates more closely to 3-fold symmetry. Thecontributed by monomer A and residues Met227,
Pro228, and Tyr292 from monomer B form part of the tightness of the trimeric fiber binding is probably dueto the avidity effect of the three binding sites and the factpeptide binding surface. Although the binding groove
is relatively hydrophobic, a number of putative hydrogen that the conformational switch leads to a more stableconformation of the penton base, notably of helix �7bonds are also formed between the peptide and the
penton base. The side chain amine of Asn12(fiber) forms and the following residues, which are poorly ordered inthe unliganded state.a hydrogen bond with the carboxylate of Glu203. Addi-
tionally, the �1-nitrogen of Arg197 is within 3 A of the There are possibly additional fiber interactions withthe penton base not seen in our structure due to themain chain carbonyl of Asn12(fiber). The main chain
carbonyl of Val14(fiber) forms a hydrogen bond with the shortness of the tail peptide used. The presence of theintact fiber in the penton base could alter and/or stabilize�1-nitrogen Arg496, and a salt bridge occurs between
the carboxylate of Asp18(fiber) and the side chain amino the conformation of the RGD loop possibly through di-rect interactions as suggested by comparative EM re-group of Lys387. The -1 nitrogen of His494 hydrogen
bonds to the side chain hydroxyl of Tyr15(fiber). Tyr17- constructions of Ad3 penton base and penton dodeca-hedra, where a conformational change in the penton(fiber) is stacked between Tyr15(fiber) and Tyr292, do-
nated from the adjacent monomer B (Figure 4B). All cited base upon fiber binding was reported (Schoehn et al.,1996).interactions are to the universally conserved FNPVYPY
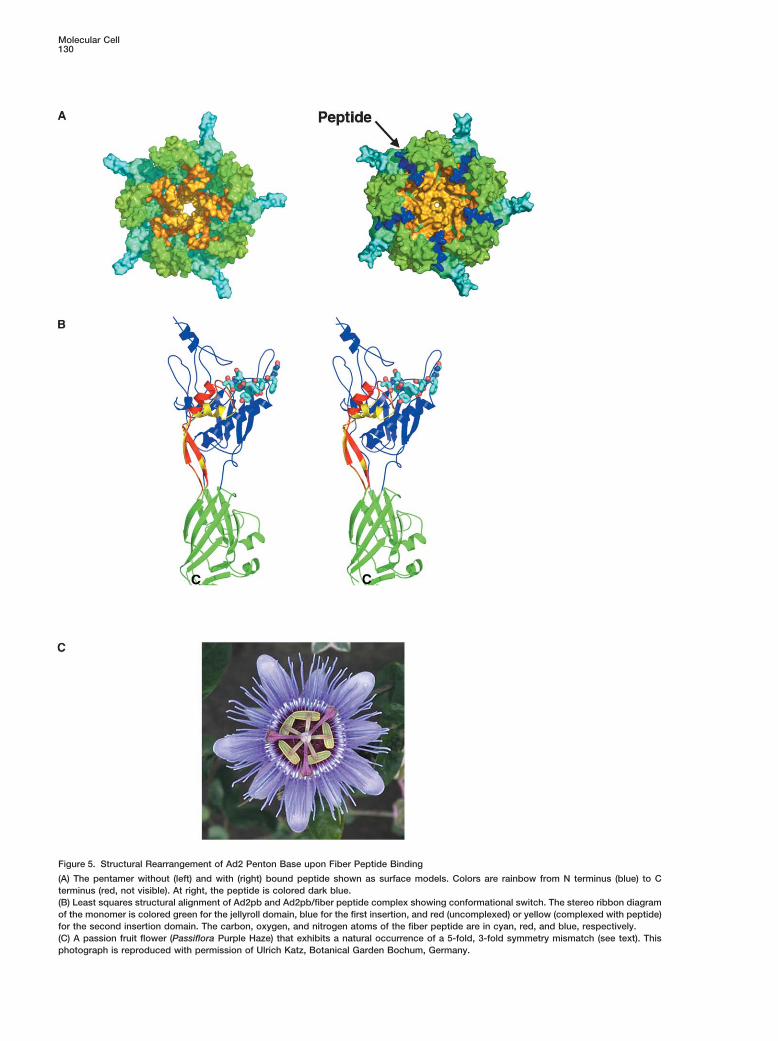
motif of the fiber tail. In addition, all cited residues on In the spirit of poetic license, we note that natureexhibits a trimeric/pentameric symmetry mismatch phe-the penton base with which the peptide interacts are
also very highly conserved with the exception of Lys387 nomenon in the beautiful flowers of the Passiflora family,which have five stamens and three stigma. An example(Figure 1). Thus, this is likely to be a universal mode of
association for adenovirus penton base and fiber. is shown in Figure 5C that appears to us to be a plausiblerepresentation of how the three fiber tails might bind atHow does the trimeric fiber fit into a pentameric pen-
ton base? The clear electron density for the fiber peptide the interface of the five penton base subunits, leavingtwo sites vacant.after 60-fold NCS averaging implies a conserved and
stable binding conformation for the fiber peptide in the It remains unclear how some avian adenoviruses arecapable of binding two fiber proteins, one short andpenton base monomer, with each fiber binding site
equivalent and one penton being capable of binding one long, per penton base. Sequence analysis and EMsuggest that the short avian fiber is structurally similarfive peptides. Comparison of adjacent monomers in the
bound and unbound conformations shows that they are to mammalian adenovirus fibers and interacts normallywith the penton base, whereas the long fiber is clearlysterically incompatible because Thr488 of one monomer
with bound peptide clashes with Gln482 and Val479 of abnormal both in sequence (with a long polyglycine se-quence immediately following the penton base bindingan adjacent monomer without peptide. Thus, the confor-
mational switch upon fiber binding is likely to be cooper- motif) and in disposition (it appears to come out horizon-tally from the penton base) (Hess et al., 1995). Assumingative around the penton base. This would allow the three
independently flexible fiber tails to be accommodated in the short fiber binds in three binding sites, these obser-vations could be explained by two of the long fiber tailsthree binding sites, leaving two sites unsatisfied (Figure
5C). However, all five sites would likely adopt a confor- interacting with the remaining two binding sites (whichmust both be on the same half of the penton base) andmation more closely resembling a fiber peptide bound
state. The peptide lies almost horizontal and radial on the polyglycine stretch, providing sufficient flexibility toavoid steric clashes between the fibers.the top surface of the penton base with its N terminus
external and the C-terminal part central (Figure 4A). Thisarrangement allows the three flexible tails to come to- Structural Comparisons with Other Proteins
Based on the DALI algorithm (Holm and Sander, 1993),gether in the center to form a trimeric structure probablyinvolving the following conserved proline-rich regions the Ad2pb monomer is structurally most closely related
to proteins such as Drosophila nucleoplasmin-like pro-(27-PFLTPPF) prior to the first repeating shaft motif. This
Figure 4. Fiber Peptide Interactions with the Penton Base
(A) Stereo diagram of the fiber peptide (red) bound to the penton base subunit interface viewed looking down the 5-fold axis.(B) Stereo view of the peptide binding site. The ribbon diagrams of the monomers are colored blue and green and the bound peptide isdepicted as sticks with carbons colored white. Residues forming the peptide binding pocket are colored yellow for monomer A and orangefor monomer B. Putative hydrogen bonding interactions are denoted by dashed yellow lines.(C) Stereo view of the experimental averaged electron density of the bound peptide.(D) Sequence alignment of the N-terminal portion of representative fiber proteins from serotypes Ad2, accession number P03275; Ad3, P04501;Ad7, AAR89975; Ad11, NP_852715; Ad12, DAA00575; Ad17, DAA00599; Ad25, NP_478420; Ad37, AAB71734; Ad41 short fiber, P16883; andAd41 long fiber, P14267. Conserved residues (yellow) and the peptide sequence used for structural analysis (red) are highlighted.
Molecular Cell130
Figure 5. Structural Rearrangement of Ad2 Penton Base upon Fiber Peptide Binding
(A) The pentamer without (left) and with (right) bound peptide shown as surface models. Colors are rainbow from N terminus (blue) to Cterminus (red, not visible). At right, the peptide is colored dark blue.(B) Least squares structural alignment of Ad2pb and Ad2pb/fiber peptide complex showing conformational switch. The stereo ribbon diagramof the monomer is colored green for the jellyroll domain, blue for the first insertion, and red (uncomplexed) or yellow (complexed with peptide)for the second insertion domain. The carbon, oxygen, and nitrogen atoms of the fiber peptide are in cyan, red, and blue, respectively.(C) A passion fruit flower (Passiflora Purple Haze) that exhibits a natural occurrence of a 5-fold, 3-fold symmetry mismatch (see text). Thisphotograph is reproduced with permission of Ulrich Katz, Botanical Garden Bochum, Germany.
Structure of the Adenovirus Penton131
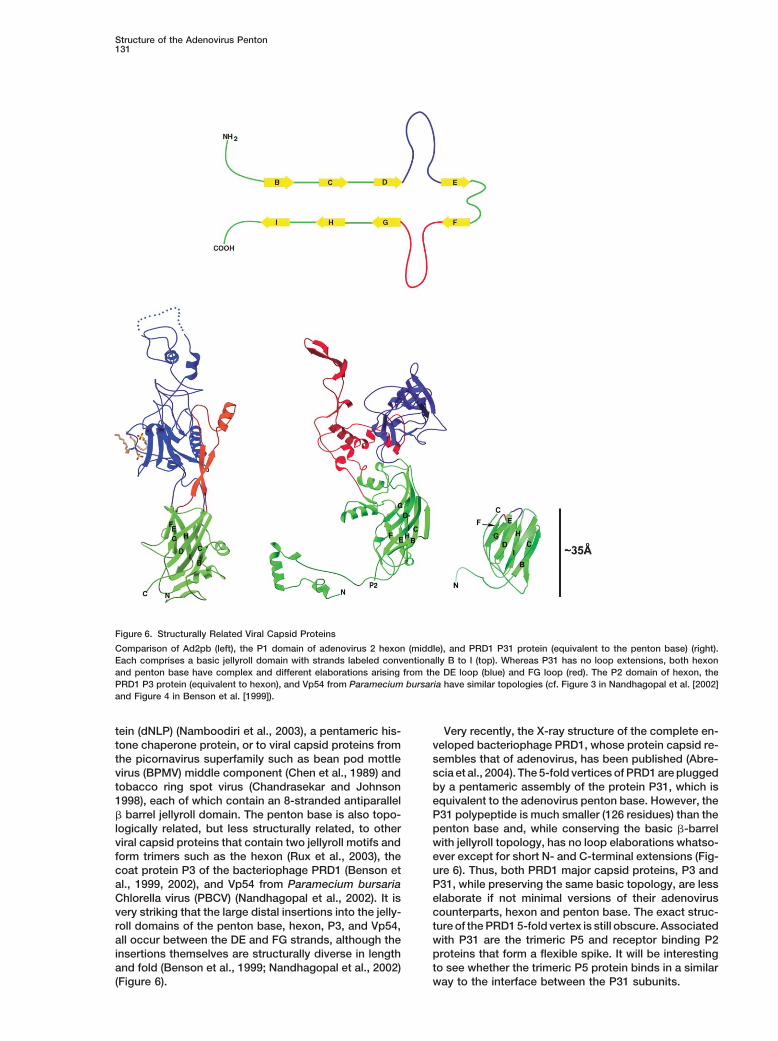
Figure 6. Structurally Related Viral Capsid Proteins
Comparison of Ad2pb (left), the P1 domain of adenovirus 2 hexon (middle), and PRD1 P31 protein (equivalent to the penton base) (right).Each comprises a basic jellyroll domain with strands labeled conventionally B to I (top). Whereas P31 has no loop extensions, both hexonand penton base have complex and different elaborations arising from the DE loop (blue) and FG loop (red). The P2 domain of hexon, thePRD1 P3 protein (equivalent to hexon), and Vp54 from Paramecium bursaria have similar topologies (cf. Figure 3 in Nandhagopal et al. [2002]and Figure 4 in Benson et al. [1999]).
tein (dNLP) (Namboodiri et al., 2003), a pentameric his- Very recently, the X-ray structure of the complete en-veloped bacteriophage PRD1, whose protein capsid re-tone chaperone protein, or to viral capsid proteins from
the picornavirus superfamily such as bean pod mottle sembles that of adenovirus, has been published (Abre-scia et al., 2004). The 5-fold vertices of PRD1 are pluggedvirus (BPMV) middle component (Chen et al., 1989) and
tobacco ring spot virus (Chandrasekar and Johnson by a pentameric assembly of the protein P31, which isequivalent to the adenovirus penton base. However, the1998), each of which contain an 8-stranded antiparallel
� barrel jellyroll domain. The penton base is also topo- P31 polypeptide is much smaller (126 residues) than thepenton base and, while conserving the basic �-barrellogically related, but less structurally related, to other
viral capsid proteins that contain two jellyroll motifs and with jellyroll topology, has no loop elaborations whatso-ever except for short N- and C-terminal extensions (Fig-form trimers such as the hexon (Rux et al., 2003), the
coat protein P3 of the bacteriophage PRD1 (Benson et ure 6). Thus, both PRD1 major capsid proteins, P3 andP31, while preserving the same basic topology, are lessal., 1999, 2002), and Vp54 from Paramecium bursaria
Chlorella virus (PBCV) (Nandhagopal et al., 2002). It is elaborate if not minimal versions of their adenoviruscounterparts, hexon and penton base. The exact struc-very striking that the large distal insertions into the jelly-
roll domains of the penton base, hexon, P3, and Vp54, ture of the PRD1 5-fold vertex is still obscure. Associatedwith P31 are the trimeric P5 and receptor binding P2all occur between the DE and FG strands, although the
insertions themselves are structurally diverse in length proteins that form a flexible spike. It will be interestingto see whether the trimeric P5 protein binds in a similarand fold (Benson et al., 1999; Nandhagopal et al., 2002)
(Figure 6). way to the interface between the P31 subunits.

Molecular Cell132
Bac-to-Bac system and Htb expression vector were from Invitrogen,Conclusionsand Sf21 and HighFive insect cells were from GIBCO. ZwittergentRecombinant adenoviruses are potentially an important3-12 was obtained from Hampton. The hAd2 fiber N-terminal peptidevehicle for gene delivery for therapeutic applications.(MKRARPSEDTFNPVYPYDTEC) was purchased from DiverDrugs.
Despite extensive development, their utility has been The C-terminal cysteine was added to the native sequence for possi-restricted by promiscuous tropism and host inflamma- ble heavy atom derivativisation.tory immune response (see for example, Nemerow Purification of the full-length construct revealed closely spaced
bands that by N-terminal sequencing corresponded to truncations[2000]; Young and Mautner [2001]; Amalfitano and Parksof �49 and �30 residues. New �30 and �49 constructs for expres-[2002]; and Einfeld and Roelvink [2002]). Structural stud-sion in the baculovirus system were obtained with forward oligomersies of adenoviral capsid proteins are critical in being5-TATTTTCAGGGCGCCATGGGATCCCCCTTCGATGCTCCC-3
able to overcome these limitations. Most frequently, re- (�30 construct) and 5-TATTTTCAGGGCGCCATGGGAACCGGGGtargeting of adenovirus vectors has been achieved GGAGAAACAGC-3 (�49 construct) and a reverse oligomer, 5-through alteration of the knob domain of the fiber pro- GTAGGCCTTTGAATTCCTCAAAAAGTGCGGCTCGAT-3 with Nco1
(forward) and EcoR1 (reverse) cleavage sites for cloning. Virus ampli-tein, the site of primary receptor binding. Insertion offication was performed in a monolayer of Sf21 cells and proteintargeting sequences into the penton base has also beenexpression was done in HighFive cells in shaker flasks (135 rpm,successfully used in the design of adenoviral vectors27�C, 3 days) with a multiplicity of infection between 1 and 5. Cellsexhibiting nonnative tropism (Einfeld et al., 1999). Tro-were collected at 1000 rpm for 5 min and stored at �80�C. Cells
pism can be successfully modulated in this manner, were resuspended in 25 mM Tris, (pH 7.5) containing 100 mM NaClwhereas the immune response to the vector is not elimi- (buffer A) plus protease inhibitors (Roche) and lysed by sonication.nated. The immune system responds to the adenovirus The cell debris was pelleted at 100,000 � g for 45 min and the
supernatant collected. Ammonium sulfate (30%) was added to thevector by first producing antifiber antibodies recogniz-supernatant and the precipitant collected. The precipitant was re-ing the trimeric form of the C-terminal knob domainsuspended in buffer A and the ammonium sulfate removed by dial-(Gahery-Segard et al., 1998). One potential method ofysis against the same buffer. The solution was then applied to a
improving adenovirus vectors for gene therapy applica- MONO Q column (Pharmacia) by using a linear gradient from 100tion is therefore to use fiberless adenoviruses (Von Seg- mM to 1 M NaCl, in 25 mM Tris (pH 7.5). The Ad2pb was eluted atgern et al., 1999; Li et al., 2000). However, a serious approximately 220 mM NaCl. The fractions of interest were concen-limitation of the fiberless vectors is the reduced stability trated to 6–9 mg/ml and stored at �80�C. Due to higher expression
levels and greater proteolytic stability, only the �49 construct wasof the capsid (Von Seggern et al., 1999) perhaps due toused in extensive crystallization trials.a nonoptimal conformation of the penton base in the
absence of the fiber. Based on the results presentedEMhere, the conformational change in the penton basePurified samples of Ad2pb (full-length and �49 construct) at 6 mg/mlupon fiber binding can be to a large extent duplicatedwere diluted to �0.1 mg/ml in crystallization buffer (1.6 M ammonium
with short peptides and possibly small molecule mimics sulfate, 10% dioxane, 0.1 M MES, and [pH 6.5]) and incubated atof the fiber protein N-terminal region. Thus, the stability room temperature for at least 1 hr to allow formation of dodecahedralof the fiberless vector could be improved for instance particles. Protein samples were similarly diluted in a 1.6 M ammo-
nium sulfate solution buffered with 0.1 M MES (pH 6.5) or a 10%by engineering into the RGD loop the fiber tail motif.dioxane solution buffered with 0.1 M MES (pH 6.5) and incubatedFurthermore, the extensive structural information nowat room temperature. Samples were then applied to the clean sideavailable on the three major capsid proteins allows theof carbon on mica (carbon/mica interface) and negatively staineddesign and addition of amino acid sequences shown towith 1% sodium silico-tungstate (pH 7.0). Micrographs were taken
interact with cellular receptors into surface loop regions under low-dose conditions with a JEOL 1200 EX II microscope atof these proteins. Such a region in the penton base 100 kV and a nominal magnification of 40,000�.could be the second variable loop (residues 150–160),which forms a surface accessible protuberance (Figures Crystallization
Crystals were grown in hanging drops against 1.5–1.6 M ammonium2C and 5A). Vectors based on such constructs wouldsulfate, 10% dioxane, and 0.1 M MES (pH 6.5). 1 �l of the crystalliza-have the benefit of nonnative tropism and potentiallytion solution was added to 1 �l of the protein solution at concentra-less antigenic response.tion 6–8 mg/ml (approximately 0.1 mM) in buffer A. ApproximatelyFinally, due to the very high conservation of the inter-0.3 �l of a 40 mM zwittergent 3-12 stock solution was added directly
action, a compound that would block the fiber tail bind- to the hanging drop as an additive. Crystals of Ad2pb grew overing to the penton base could be a general inhibitor of 4–8 weeks at 15�C, reaching dimensions of 100 � 75 � 30 �m3.adenoviral assembly and multiplication. Such a com- Prior to flash freezing, crystals were soaked for �5 min in 20%
sucrose, 1.8 M ammonium sulfate, 10% dioxane, and 0.1 M MESpound might be useful as an agent for treating diverse(pH 6.5) as a stabilizing and cryoprotecting solution. For the crystalli-adenoviral infections, which can still be the cause ofzation of the Ad2 penton base/fiber peptide complex, a 3-fold molarserious epidemics especially for children and immuno-excess of peptide was added to the Ad2pb protein and incubatedcompromised patients (Kolavic-Gray et al., 2002; Kimon ice for 30 min prior to addition of crystallization buffer and zwitter-
et al., 2003; Kojaoghlanian et al., 2003). By using the gent 3-12. The average crystal size was �75 � 75 � 10 �m3 for thecrystal structure of the peptide-penton base complex, Ad2pb fiber peptide complex.lead compounds of this kind could be evolved into pen-tavalent ligands with very tight avidity based binding. Structure DeterminationSuch an approach has been explored for inhibitors of Crystallographic data for Ad2pb were collected on European Syn-
chrotron Radiation Facility (ESRF) beamlines ID14-EH1, EH2, andpentameric cholera and other toxins (Zhang et al., 2002).EH4 in 0.5� frames. Due to radiation sensitivity, data from ten crystalswere merged to create a more complete dataset. For the Ad2pb/Experimental Proceduresfiber peptide complex, data were collected on the ESRF microfocusbeamline ID13 with the EMBL microdiffractometer (Perrakis et al.,Protein Expression and Purification1999). Radiation damage was readily apparent after a few expo-A full-length construct of hAd2 penton base was a gift from Pierre
Boulanger (Karayan et al., 1994). For baculovirus expression, the sures, which limited the amount of usable data per exposed position

Structure of the Adenovirus Penton133
to 2� (four 0.5� frames). One large crystal (100 � 75 � 20 �m3) Bailey, A., and Mautner, V. (1994). Phylogenetic relationships amongadenovirus serotypes. Virology 205, 438–452.provided 104 frames (52�) of usable data to a maximum resolution
of 3.5 A, the combination of a 10 �m beam and the precision of Benko, M., and Harrach, B. (2003). Molecular evolution of adenovi-the microdiffractometer permitting exposure of multiple volumes. A ruses. Curr. Top. Microbiol. Immunol. 272, 3–35.smaller crystal (75 � 75 � 10 �m3) gave an additional 12 frames (6�) Benson, S.D., Bamford, J.K., Bamford, D.H., and Burnett, R.M.of data to 3.5 A. All data were integrated with MOSFLM (Leslie 1992) (1999). Viral evolution revealed by bacteriophage PRD1 and humanand scaled with SCALA (CCP4, 1994). adenovirus coat protein structures. Cell 98, 825–833.
MOLREP (Vagin and Teplyakov, 2000) was used to place the cryo-Benson, S.D., Bamford, J.K., Bamford, D.H., and Burnett, R.M.EM electron density of the Ad3 dodecahedron (Schoehn et al., 1996)(2002). The X-ray crystal structure of P3, the major coat protein ofin the unit cell. Subsequent 60-fold NCS-averaging and gradualthe lipid-containing bacteriophage PRD1, at 1.65 A resolution. Actaphase extension from 20 A with RAVE (Kleywegt and Jones, 1999)Crystallogr. D Biol. Crystallogr. 58, 39–59.allowed the generation of high quality electron density maps (TableBergelson, J.M., Cunningham, J.A., Droguett, G., Kurt-Jones, E.A.,1). Model building was performed in O (Jones et al., 1991) andKrithivas, A., Hong, J.S., Horwitz, M.S., Crowell, R.L., and Finberg,refinements carried out with CNS (Brunger et al., 1998). Only oneR.W. (1997). Isolation of a common receptor for Coxsackie B virusesmonomer was built, and all refinements used strict 60-fold icosahe-and adenoviruses 2 and 5. Science 275, 1320–1323.dral NCS constraints. Unaveraged maps calculated with CNS from
the model were used to locate deviations from perfect icosahedral Bewley, M.C., Springer, K., Zhang, Y.B., Freimuth, P., and Flanagan,symmetry at crystal contacts. In the case of the Ad2pb-fiber peptide J.M. (1999). Structural analysis of the mechanism of adenoviruscomplex, the native Ad2pb dodecahedron structure was used for binding to its human cellular receptor, CAR. Science 286, 1579–molecular replacement and phase extension recommenced from 1583.low resolution. However, maintaining strict 60-fold icosahedral NCS Blumenthal, R., Seth, P., Willingham, M.C., and Pastan, I. (1986).restraints gave poor final averaging statistics and poor density for pH-dependent lysis of liposomes by adenovirus. Biochemistry 25,the peptide. By using rigid-body refinement in CNS to relax the 2231–2237.icosahedral symmetry for each of the 60 monomers and then re-
Boudin, M.L., and Boulanger, P. (1982). Assembly of adenoviruspeating the averaging with newly defined NCS matrices vastly im-penton base and fiber. Virology 116, 589–604.proved the map and allowed the peptide and interacting region ofBrunger, A.T., Adams, P.D., Clore, G.M., DeLano, W.L., Gros, P.,the penton base to be built unambiguously.Grosse-Kunstleve, R.W., Jiang, J.S., Kuszewski, J., Nilges, M.,According to PROCHECK (Laskowski et al., 1993) (Table 1), onlyPannu, N.S., et al. (1998). Crystallography & NMR system: a newsix (Tyr84, Phe137, Asp156, Glu167, Glu172, and Ser176) and foursoftware suite for macromolecular structure determination. Acta(Ser58, Asp65, Asp156, and Ser176) residues for the Ad2pb andCrystallogr. D Biol. Crystallogr. 54, 905–921.Ad2pb/fiber peptide complex structures, respectively, were in disal-
lowed regions of the Ramachandran plot. These generally lie in Burmeister, W.P., Guilligay, D., Cusack, S., Wadell, G., and Arnberg,N. (2004). Crystal structure of species D adenovirus fiber knobs andpoorly ordered regions. Residues 568–571 (native C terminus) and
residues 298–335 were disordered and not modeled in the Ad2pb their sialic acid binding sites. J. Virol. 78, 7727–7736.structure. Similarly, residues 49–51, 297–373, and 568–571 were CCP4 (Collaborative Computational Project, Number 4) (1994). Thenot modeled in the Ad2pb/fiber peptide structure. Figures were CCP4 suite: programs for protein crystallography. Acta Crystallogr.generated with PyMOL (DeLano 2002). D Biol. Crystallogr. 50, 760–763.
Chandrasekar, V., and Johnson, J.E. (1998). The structure of tobaccoAcknowledgments ringspot virus: a link in the evolution of icosahedral capsids in the
picornavirus superfamily. Structure 6, 157–171.The authors wish to thank the European Molecular Biology Labora-
Chelvanayagam, G., Heringa, J., and Argos, P. (1992). Anatomy andtory and the European Synchrotron Radiation Facility (ESRF) staff,evolution of proteins displaying the viral capsid jellyroll topology. J.in particular Christian Riekel, for access to ESRF beamlines; ManfredMol. Biol. 228, 220–242.Burghammer and David Flot for assistance in data collection onChen, Z.G., Stauffacher, C., Li, Y., Schmidt, T., Bomu, W., Kamer,ID13; John Johnson and colleagues (Scripps Research Institute) forG., Shanks, M., Lomonossoff, G., and Johnson, J.E. (1989). Protein-discussions about icosahedral symmetry and self-rotation func-RNA interactions in an icosahedral virus at 3.0 A resolution. Sciencetions; Fasseli Coulibaly and Felix Rey (Gif-sur-Yvette) for help with245, 154–159.NCS averaging in RAVE; Pierre Boulanger (Lyon) for the Ad2 penton
base construct; James Conway and Elisabeth Hewat (IBS) for elec- Chiu, C.Y., Mathias, P., Nemerow, G.R., and Stewart, P.L. (1999).tron microscopy; Catherine Mazza and Evelyne Gout for help with Structure of adenovirus complexed with its internalization receptor,baculovirus expression; Wim Burmeister, Pascal Fender, Ulrike alphavbeta5 integrin. J. Virol. 73, 6759–6768.Kapp, and Catherine Stehlin for initiating the penton base crystallog- Chroboczek, J., Ruigrok, R.W., and Cusack, S. (1995). Adenovirusraphy project; Max Nanao for help with figures; and Rob Ruigrok fiber. Curr. Top. Microbiol. Immunol. 199, 163–200.for frequent discussions. We thank Ulrich Katz (Botanical Garden
Cohen, C.J., Shieh, J.T., Pickles, R.J., Okegawa, T., Hsieh, J.T., andBochum, Germany) for Figure 5C.
Bergelson, J.M. (2001). The coxsackievirus and adenovirus receptoris a transmembrane component of the tight junction. Proc. Natl.
Received: August 25, 2004 Acad. Sci. USA 98, 15191–15196.Revised: November 10, 2004
DeLano, W.L. (2002). The PyMOL Molecular Graphics System. (SanAccepted: November 24, 2004Carlos, CA: DeLano Scientific).Published online: December 16, 2004Devaux, C., Caillet-Boudin, M.L., Jacrot, B., and Boulanger, P.(1987). Crystallization, enzymatic cleavage, and the polarity of theReferencesadenovirus type 2 fiber. Virology 161, 121–128.
Abrescia, N.G., Cockburn, J.J., Grimes, J.M., Sutton, G.C., Diprose, Durmort, C., Stehlin, C., Schoehn, G., Mitraki, A., Drouet, E., Cusack,J.M., Butcher, S.J., Fuller, S.D., San Martin, C., Burnett, R.M., Stuart, S., and Burmeister, W.P. (2001). Structure of the fiber head of Ad3,D.I., et al. (2004). Insights into assembly from structural analysis of a non-CAR-binding serotype of adenovirus. Virology 285, 302–312.bacteriophage PRD1. Nature 432, 68–74. Einfeld, D.A., and Roelvink, P.W. (2002). Advances towards tar-Albinsson, B., and Kidd, A.H. (1999). Adenovirus type 41 lacks an getable adenovirus vectors for gene therapy. Curr. Opin. Mol. Ther.RGD alpha(v)-integrin binding motif on the penton base and under- 4, 444–451.goes delayed uptake in A549 cells. Virus Res. 64, 125–136. Einfeld, D.A., Brough, D.E., Roelvink, P.W., Kovesdi, I., and Wick-
ham, T.J. (1999). Construction of a pseudoreceptor that mediatesAmalfitano, A., and Parks, R.J. (2002). Separating fact from fiction:assessing the potential of modified adenovirus vectors for use in transduction by adenoviruses expressing a ligand in fiber or penton
base. J. Virol. 73, 9130–9136.human gene therapy. Curr. Gene Ther. 2, 111–133.

Molecular Cell134
Fender, P., Ruigrok, R.W., Gout, E., Buffet, S., and Chroboczek, J. G.R. (2000). Signaling antibodies complexed with adenovirus cir-cumvent CAR and integrin interactions and improve gene delivery.(1997). Adenovirus dodecahedron, a new vector for human gene
transfer. Nat. Biotechnol. 15, 52–56. Gene Ther. 7, 1593–1599.
Furcinitti, P.S., van Oostrum, J., and Burnett, R.M. (1989). Adenovi- Medina-Kauwe, L.K. (2003). Endocytosis of adenovirus and adenovi-rus polypeptide IX revealed as capsid cement by difference images rus capsid proteins. Adv. Drug Deliv. Rev. 55, 1485–1496.from electron microscopy and crystallography. EMBO J. 8, 3563– Medina-Kauwe, L.K., Maguire, M., Kasahara, N., and Kedes, L.3570. (2001). Nonviral gene delivery to human breast cancer cells by tar-Gaggar, A., Shayakhmetov, D.M., and Lieber, A. (2003). CD46 is a geted Ad5 penton proteins. Gene Ther. 8, 1753–1761.cellular receptor for group B adenoviruses. Nat. Med. 9, 1408–1412. Nakano, M.Y., Boucke, K., Suomalainen, M., Stidwill, R.P., andPublished online October 19, 2003. 10.1038/nm952. Greber, U.F. (2000). The first step of adenovirus type 2 disassemblyGahery-Segard, H., Farace, F., Godfrin, D., Gaston, J., Lengagne, R., occurs at the cell surface, independently of endocytosis and escapeTursz, T., Boulanger, P., and Guillet, J.G. (1998). Immune response to to the cytosol. J. Virol. 74, 7085–7095.recombinant capsid proteins of adenovirus in humans: antifiber and Namboodiri, V.M., Dutta, S., Akey, I.V., Head, J.F., and Akey, C.W.anti-penton base antibodies have a synergistic effect on neutralizing (2003). The crystal structure of Drosophila NLP-core provides insightactivity. J. Virol. 72, 2388–2397. into pentamer formation and histone binding. Structure (Camb.)Galinier, R., Gout, E., Lortat-Jacob, H., Wood, J., and Chroboczek, 11, 175–186.J. (2002). Adenovirus protein involved in virus internalization recruits Nandhagopal, N., Simpson, A.A., Gurnon, J.R., Yan, X., Baker, T.S.,ubiquitin-protein ligases. Biochemistry 41, 14299–14305. Graves, M.V., Van Etten, J.L., and Rossmann, M.G. (2002). The struc-Greber, U.F., Willetts, M., Webster, P., and Helenius, A. (1993). Step- ture and evolution of the major capsid protein of a large, lipid-wise dismantling of adenovirus 2 during entry into cells. Cell 75, containing DNA virus. Proc. Natl. Acad. Sci. USA 99, 14758–14763.477–486. Published online October 31, 2002. 10.1073/pnas.232580699Hess, M., Cuzange, A., Ruigrok, R.W., Chroboczek, J., and Jacrot, Nemerow, G.R. (2000). Adenoviral vectors–new insights. Trends Mi-B. (1995). The avian adenovirus penton: two fibres and one base. crobiol. 8, 391–394.J. Mol. Biol. 252, 379–385.
Nemerow, G.R., and Stewart, P.L. (1999). Role of alpha(v) integrinsHolm, L., and Sander, C. (1993). Protein structure comparison by in adenovirus cell entry and gene delivery. Microbiol. Mol. Biol. Rev.alignment of distance matrices. J. Mol. Biol. 233, 123–138. 63, 725–734.Hong, S.S., and Boulanger, P. (1995). Protein ligands of the human Norrby, E. (1964). The relationship between the soluble antigensadenovirus type 2 outer capsid identified by biopanning of a phage- and the virion of adenovirus type 3. I. Morphological characteristics.displayed peptide library on separate domains of wild-type and Virology 1, 236–248.mutant penton capsomers. EMBO J. 14, 4714–4727.
Perrakis, A., Cipriani, F., Castagna, J.C., Claustre, L., Burghammer,Jones, T.A., Zou, J.Y., Cowan, S.W., and Kjeldgaard (1991). Im- M., Riekel, C., and Cusack, S. (1999). Protein microcrystals and theproved methods for building protein models in electron density design of a microdiffractometer: current experience and plans atmaps and the location of errors in these models. Acta Crystallogr. EMBL and ESRF/ID13. Acta Crystallogr. D Biol. Crystallogr. 55,A 47, 110–119. 1765–1770.Karayan, L., Gay, B., Gerfaux, J., and Boulanger, P.A. (1994). Oligo- Richardson, J.S. (1981). The anatomy and taxonomy of protein struc-merization of recombinant penton base of adenovirus type 2 and ture. Adv. Protein Chem. 34, 167–339.its assembly with fiber in baculovirus-infected cells. Virology 202,
Roelvink, P.W., Lizonova, A., Lee, J.G., Li, Y., Bergelson, J.M., Fin-782–795.berg, R.W., Brough, D.E., Kovesdi, I., and Wickham, T.J. (1998).
Karayan, L., Hong, S.S., Gay, B., Tournier, J., d’Angeac, A.D., and The coxsackievirus-adenovirus receptor protein can function as aBoulanger, P. (1997). Structural and functional determinants in ade- cellular attachment protein for adenovirus serotypes from sub-novirus type 2 penton base recombinant protein. J. Virol. 71, 8678– groups A, C, D, E, and F. J. Virol. 72, 7909–7915.8689.
Ruigrok, R.W., Barge, A., Albiges-Rizo, C., and Dayan, S. (1990).Kim, Y.J., Hong, J.Y., Lee, H.J., Shin, S.H., Kim, Y.K., Inada, T., Structure of adenovirus fibre. II. Morphology of single fibres. J. Mol.Hashido, M., and Piedra, P.A. (2003). Genome type analysis of ade- Biol. 215, 589–596.novirus types 3 and 7 isolated during successive outbreaks of lower
Rux, J.J., and Burnett, R.M. (2000). Type-specific epitope locationsrespiratory tract infections in children. J. Clin. Microbiol. 41, 4594–revealed by X-ray crystallographic study of adenovirus type 5 hexon.4599.Mol. Ther. 1, 18–30.
Kleywegt, G.J., and Jones, T.A. (1999). Software for handling macro-Rux, J.J., Kuser, P.R., and Burnett, R.M. (2003). Structural and phylo-molecular envelopes. Acta Crystallogr. D Biol. Crystallogr. 55,genetic analysis of adenovirus hexons by use of high-resolution941–944.x-ray crystallographic, molecular modeling, and sequence-based
Kojaoghlanian, T., Flomenberg, P., and Horwitz, M.S. (2003). Themethods. J. Virol. 77, 9553–9566.
impact of adenovirus infection on the immunocompromised host.Schoehn, G., Fender, P., Chroboczek, J., and Hewat, E.A. (1996).Rev. Med. Virol. 13, 155–171.Adenovirus 3 penton dodecahedron exhibits structural changes ofKolavic-Gray, S.A., Binn, L.N., Sanchez, J.L., Cersovsky, S.B., Pol-the base on fibre binding. EMBO J. 15, 6841–6846.yak, C.S., Mitchell-Raymundo, F., Asher, L.V., Vaughn, D.W., Feigh-Segerman, A., Atkinson, J.P., Marttila, M., Dennerquist, V., Wadell,ner, B.H., and Innis, B.L. (2002). Large epidemic of adenovirus typeG., and Arnberg, N. (2003). Adenovirus type 11 uses CD46 as a4 infection among military trainees: epidemiological, clinical, andcellular receptor. J. Virol. 77, 9183–9191.laboratory studies. Clin Infect Dis 35, 808–818. Published online
September 10, 2002. Seth, P., Willingham, M.C., and Pastan, I. (1985). Binding of adenovi-rus and its external proteins to Triton X-114. Dependence on pH. J.Laskowski, R.A., MacArthur, M.W., Moss, D.S., and Thornton, J.M.Biol. Chem. 260, 14431–14434.(1993). PROCHECK: a program to check the stereochemical quality
of protein structures. J. Appl. Crystallogr. 26, 283–291. Sirena, D., Lilienfeld, B., Eisenhut, M., Kalin, S., Boucke, K., Beerli,R.R., Vogt, L., Ruedl, C., Bachmann, M.F., Greber, U.F., and Hemmi,Leslie, A.G.W. (1992). Recent changes to the MOSFLM package forS. (2004). The human membrane cofactor CD46 is a receptor forprocessing film and image plate data. Joint CCP4 � ESF-EAMCBspecies B adenovirus serotype 3. J. Virol. 78, 4454–4462.Newsletter on Protein Crystallography 26.
Stewart, P.L., Burnett, R.M., Cyrklaff, M., and Fuller, S.D. (1991).Li, E., Stupack, D., Bokoch, G.M., and Nemerow, G.R. (1998). Adeno-Image reconstruction reveals the complex molecular organizationvirus endocytosis requires actin cytoskeleton reorganization medi-of adenovirus. Cell 67, 145–154.ated by Rho family GTPases. J. Virol. 72, 8806–8812.
Li, E., Brown, S.L., Von Seggern, D.J., Brown, G.B., and Nemerow, Stewart, P.L., Fuller, S.D., and Burnett, R.M. (1993). Difference im-

Structure of the Adenovirus Penton135
aging of adenovirus: bridging the resolution gap between X-raycrystallography and electron microscopy. EMBO J. 12, 2589–2599.
Stewart, P.L., Chiu, C.Y., Huang, S., Muir, T., Zhao, Y., Chait, B.,Mathias, P., and Nemerow, G.R. (1997). Cryo-EM visualization of anexposed RGD epitope on adenovirus that escapes antibody neutral-ization. EMBO J. 16, 1189–1198.
Tomko, R.P., Xu, R., and Philipson, L. (1997). HCAR and MCAR: thehuman and mouse cellular receptors for subgroup C adenovirusesand group B coxsackieviruses. Proc. Natl. Acad. Sci. USA 94, 3352–3356.
Vagin, A., and Teplyakov, A. (2000). An approach to multi-copysearch in molecular replacement. Acta Crystallogr. D Biol. Crys-tallogr. 56, 1622–1624.
van Oostrum, J., and Burnett, R.M. (1985). Molecular compositionof the adenovirus type 2 virion. J. Virol. 56, 439–448.
van Raaij, M.J., Mitraki, A., Lavigne, G., and Cusack, S. (1999). Atriple beta-spiral in the adenovirus fibre shaft reveals a new struc-tural motif for a fibrous protein. Nature 401, 935–938.
Vives, R.R., Lortat-Jacob, H., Chroboczek, J., and Fender, P. (2004).Heparan sulfate proteoglycan mediates the selective attachmentand internalization of serotype 3 human adenovirus dodecahedron.Virology 321, 332–340.
Von Seggern, D.J., Chiu, C.Y., Fleck, S.K., Stewart, P.L., and Nem-erow, G.R. (1999). A helper-independent adenovirus vector with E1,E3, and fiber deleted: structure and infectivity of fiberless particles.J. Virol. 73, 1601–1608.
Walters, R.W., Freimuth, P., Moninger, T.O., Ganske, I., Zabner, J.,and Welsh, M.J. (2002). Adenovirus fiber disrupts CAR-mediatedintercellular adhesion allowing virus escape. Cell 110, 789–799.
Wickham, T.J., Mathias, P., Cheresh, D.A., and Nemerow, G.R.(1993). Integrins alpha v beta 3 and alpha v beta 5 promote adenovi-rus internalization but not virus attachment. Cell 73, 309–319.
Wu, E., Pache, L., Von Seggern, D.J., Mullen, T.M., Mikyas, Y., Stew-art, P.L., and Nemerow, G.R. (2003). Flexibility of the adenovirus fiberis required for efficient receptor interaction. J. Virol. 77, 7225–7235.
Wu, E., Trauger, S.A., Pache, L., Mullen, T.M., von Seggern, D.J.,Siuzdak, G., and Nemerow, G.R. (2004). Membrane cofactor proteinis a receptor for adenoviruses associated with epidemic keratocon-junctivitis. J. Virol. 78, 3897–3905.
Xia, D., Henry, L.J., Gerard, R.D., and Deisenhofer, J. (1994). Crystalstructure of the receptor-binding domain of adenovirus type 5 fiberprotein at 1.7 A resolution. Structure 2, 1259–1270.
Young, L.S., and Mautner, V. (2001). The promise and potentialhazards of adenovirus gene therapy. Gut 48, 733–736.
Zhang, Z., Merritt, E.A., Ahn, M., Roach, C., Hou, Z., Verlinde, C.L.,Hol, W.G., and Fan, E. (2002). Solution and crystallographic studiesof branched multivalent ligands that inhibit the receptor-binding ofcholera toxin. J. Am. Chem. Soc. 124, 12991–12998.
Accession Numbers
The native penton base structure and complex with fiber peptidehave PDB codes 1X9P and 1X9T, respectively.
Related Documents










