APPLIED AND ENVIRONMENTAL MICROBIOLOGY, 0099-2240/00/$04.0010 Feb. 2000, p. 558–565 Vol. 66, No. 2 Copyright © 2000, American Society for Microbiology. All Rights Reserved. The Streptococcus thermophilus Autolytic Phenotype Results from a Leaky Prophage CLARA HUSSON-KAO, 1 JE ´ RO ˆ ME MENGAUD, 2 BE ´ NE ´ DICTE CESSELIN, 3 DOUWE VAN SINDEREN, 4 LAURENT BENBADIS, 2 AND MARIE-PIERRE CHAPOT-CHARTIER 1 * Unite ´ de Biochimie et Structure des Prote ´ines, 1 and Unite ´ de Recherche Laitie `re et Ge ´ne ´tique Applique ´e, 3 INRA, 78352 Jouy-en-Josas, and CIRDC, Danone, 92350 Le Plessis Robinson, 2 France, and Department of Microbiology, University College Cork, Cork, Ireland 4 Received 22 June 1999/Accepted 10 November 1999 Streptococcus thermophilus autolytic strains are characterized by a typical bell-shaped growth curve when grown under appropriate conditions. The cellular mechanisms involved in the triggering of lysis and the bacteriolytic activities of these strains were investigated in this study. Lactose depletion and organic solvents (ethanol, methanol, and chloroform) were shown to trigger a premature and immediate lysis of M17 expo- nentially growing cells. These factors and compounds are suspected to act by altering the cell envelope properties, causing either the permeabilization (organic solvents) or the depolarization (lactose depletion) of the cytoplasmic membrane. The autolytic character was shown to be associated with lysogeny. Phage particles, most of which were defective, were observed in the culture supernatants after both mitomycin C-induced and spontaneous lysis. By renaturing sodium dodecyl sulfate-polyacrylamide gel electrophoresis, a bacteriolytic activity was detected at 31 kDa exclusively in the autolytic strains. This enzyme was detected during both growth and spontaneous lysis with the same intensity. We have shown that it was prophage encoded and homologous to the endolysin Lyt51 of the streptococcal temperate bacteriophage f01205 (M. Sheehan, E. Stanley, G. F. Fitzgerald, and D. van Sinderen, Appl. Environ. Microbiol. 65:569–577, 1999). It appears from our results that the autolytic properties are conferred to the S. thermophilus strains by a leaky prophage but do not result from massive prophage induction. More specifically, we propose that phagic genes are consti- tutively expressed in almost all the cells at a low and nonlethal level and that lysis is controlled and achieved by the prophage-encoded lysis proteins. Autolysis of lactic acid bacteria used as starters appears to be a crucial step in the flavor development of fermented dairy products (12, 15). It indeed causes the opening of cells and the subsequent release into the curd of intracellular enzymes that are involved in the flavor compound formation. The activity of these enzymes can therefore be enhanced by better accessibil- ity to their substrates, proteins, lipids, or carbohydrates from the milk. It has been shown that the main consequence of cell autolysis in cheese is peptidolysis acceleration, leading to an increased rate of free amino acid production and to a decrease in bitter taste (2, 3, 13, 16, 25, 32, 34, 35, 55). Bacterial autolysis results from the degradation of the pep- tidoglycan, which is the major structural component of the bacterial cell wall, by enzymes called peptidoglycan hydrolases. Bacteria synthesize their own peptidoglycan hydrolases, named autolysins (45). These cell wall-associated enzymes are poten- tially lethal for the cell and thus require stringent regulation. It has been proposed that autolysins are involved in different cellular processes including cell wall expansion and turnover, cell division, and transformation (47). Autolysis would result from an uncontrolled action of the bacterial autolysins after inhibition of peptidoglycan synthesis (47). For lysogenic strains, lysis can be caused by induction of the resident prophage. The prophage lysis system is then responsible for the host lysis. It contains in most cases two effectors, a peptidoglycan hydrolase, named endolysin, and a second protein, a so-called holin. Ho- lins are small proteins causing nonspecific lesions in the mem- brane. They thus allow the endolysin, usually devoid of a signal peptide, access to the peptidoglycan to cause subsequent host envelope disruption (56). Hitherto, most of the studies dealing with lactic acid bacteria autolysis concern the genus Lactococcus and to a lesser extent Lactobacillus. Autolysis was investigated by physiological stud- ies and also by analysis of the peptidoglycan hydrolase content. The autolytic character of Lactococcus and Lactobacillus strains was studied in liquid medium (2, 3, 28, 33, 41, 54) and for some of them during cheese ripening (5, 13, 16, 32, 35, 53, 55). It appears to vary from strain to strain. Various factors could account for the different autolytic behaviors: the cellular content in peptidoglycan hydrolases, the regulation of either the expression or the activity of these enzymes, or the cell wall composition. Another alternative is the involvement of a pro- phage in the cell lysis, as demonstrated for the lactococcal strain AM2 (32). The bacteriolytic enzymes of Lactococcus lactis (31, 38, 41) and Lactobacillus spp. (10, 52) have been studied by renaturing sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), which allows detection of peptidoglycan hydrolase activities after renaturation in a sub- strate-containing gel. In L. lactis, an endogenous autolysin (9) and a prophage-encoded enzyme (31) were identified by this technique. Streptococcus thermophilus received little attention regarding autolysis despite its industrial significance as starter used in the manufacture of yogurts and Italian and Swiss type cheeses. For S. thermophilus, spontaneously autolytic strains have been pre- viously identified (43, 50, 58). Lysis occurs at the end of the exponential growth phase, resulting in a typical bell-shaped growth curve. Independent studies of S. thermophilus temper- ate bacteriophages led furthermore to the observation that * Corresponding author. Mailing address: Unite ´ de Biochimie et Structure des Proteines, INRA, 78352 Jouy-en-Josas, France. Phone: 33 (0)134652268. Fax: 33 (0)134652163. E-mail: [email protected] .inra.fr. 558

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

APPLIED AND ENVIRONMENTAL MICROBIOLOGY,0099-2240/00/$04.0010
Feb. 2000, p. 558–565 Vol. 66, No. 2
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
The Streptococcus thermophilus Autolytic Phenotype Resultsfrom a Leaky Prophage
CLARA HUSSON-KAO,1 JEROME MENGAUD,2 BENEDICTE CESSELIN,3 DOUWE VAN SINDEREN,4
LAURENT BENBADIS,2 AND MARIE-PIERRE CHAPOT-CHARTIER1*
Unite de Biochimie et Structure des Proteines,1 and Unite de Recherche Laitiere et Genetique Appliquee,3 INRA,78352 Jouy-en-Josas, and CIRDC, Danone, 92350 Le Plessis Robinson,2 France, and Department of Microbiology,
University College Cork, Cork, Ireland4
Received 22 June 1999/Accepted 10 November 1999
Streptococcus thermophilus autolytic strains are characterized by a typical bell-shaped growth curve whengrown under appropriate conditions. The cellular mechanisms involved in the triggering of lysis and thebacteriolytic activities of these strains were investigated in this study. Lactose depletion and organic solvents(ethanol, methanol, and chloroform) were shown to trigger a premature and immediate lysis of M17 expo-nentially growing cells. These factors and compounds are suspected to act by altering the cell envelopeproperties, causing either the permeabilization (organic solvents) or the depolarization (lactose depletion) ofthe cytoplasmic membrane. The autolytic character was shown to be associated with lysogeny. Phage particles,most of which were defective, were observed in the culture supernatants after both mitomycin C-induced andspontaneous lysis. By renaturing sodium dodecyl sulfate-polyacrylamide gel electrophoresis, a bacteriolyticactivity was detected at 31 kDa exclusively in the autolytic strains. This enzyme was detected during bothgrowth and spontaneous lysis with the same intensity. We have shown that it was prophage encoded andhomologous to the endolysin Lyt51 of the streptococcal temperate bacteriophage f01205 (M. Sheehan, E.Stanley, G. F. Fitzgerald, and D. van Sinderen, Appl. Environ. Microbiol. 65:569–577, 1999). It appears fromour results that the autolytic properties are conferred to the S. thermophilus strains by a leaky prophage butdo not result from massive prophage induction. More specifically, we propose that phagic genes are consti-tutively expressed in almost all the cells at a low and nonlethal level and that lysis is controlled and achievedby the prophage-encoded lysis proteins.
Autolysis of lactic acid bacteria used as starters appears tobe a crucial step in the flavor development of fermented dairyproducts (12, 15). It indeed causes the opening of cells and thesubsequent release into the curd of intracellular enzymes thatare involved in the flavor compound formation. The activity ofthese enzymes can therefore be enhanced by better accessibil-ity to their substrates, proteins, lipids, or carbohydrates fromthe milk. It has been shown that the main consequence of cellautolysis in cheese is peptidolysis acceleration, leading to anincreased rate of free amino acid production and to a decreasein bitter taste (2, 3, 13, 16, 25, 32, 34, 35, 55).
Bacterial autolysis results from the degradation of the pep-tidoglycan, which is the major structural component of thebacterial cell wall, by enzymes called peptidoglycan hydrolases.Bacteria synthesize their own peptidoglycan hydrolases, namedautolysins (45). These cell wall-associated enzymes are poten-tially lethal for the cell and thus require stringent regulation. Ithas been proposed that autolysins are involved in differentcellular processes including cell wall expansion and turnover,cell division, and transformation (47). Autolysis would resultfrom an uncontrolled action of the bacterial autolysins afterinhibition of peptidoglycan synthesis (47). For lysogenic strains,lysis can be caused by induction of the resident prophage. Theprophage lysis system is then responsible for the host lysis. Itcontains in most cases two effectors, a peptidoglycan hydrolase,named endolysin, and a second protein, a so-called holin. Ho-lins are small proteins causing nonspecific lesions in the mem-
brane. They thus allow the endolysin, usually devoid of a signalpeptide, access to the peptidoglycan to cause subsequent hostenvelope disruption (56).
Hitherto, most of the studies dealing with lactic acid bacteriaautolysis concern the genus Lactococcus and to a lesser extentLactobacillus. Autolysis was investigated by physiological stud-ies and also by analysis of the peptidoglycan hydrolase content.The autolytic character of Lactococcus and Lactobacillusstrains was studied in liquid medium (2, 3, 28, 33, 41, 54) andfor some of them during cheese ripening (5, 13, 16, 32, 35, 53,55). It appears to vary from strain to strain. Various factorscould account for the different autolytic behaviors: the cellularcontent in peptidoglycan hydrolases, the regulation of eitherthe expression or the activity of these enzymes, or the cell wallcomposition. Another alternative is the involvement of a pro-phage in the cell lysis, as demonstrated for the lactococcalstrain AM2 (32). The bacteriolytic enzymes of Lactococcuslactis (31, 38, 41) and Lactobacillus spp. (10, 52) have beenstudied by renaturing sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE), which allows detection ofpeptidoglycan hydrolase activities after renaturation in a sub-strate-containing gel. In L. lactis, an endogenous autolysin (9)and a prophage-encoded enzyme (31) were identified by thistechnique.
Streptococcus thermophilus received little attention regardingautolysis despite its industrial significance as starter used in themanufacture of yogurts and Italian and Swiss type cheeses. ForS. thermophilus, spontaneously autolytic strains have been pre-viously identified (43, 50, 58). Lysis occurs at the end of theexponential growth phase, resulting in a typical bell-shapedgrowth curve. Independent studies of S. thermophilus temper-ate bacteriophages led furthermore to the observation that
* Corresponding author. Mailing address: Unite de Biochimie etStructure des Proteines, INRA, 78352 Jouy-en-Josas, France. Phone:33 (0)134652268. Fax: 33 (0)134652163. E-mail: [email protected].
558

lysogens exhibit an identical autolytic phenotype (19). The aimof our study was to investigate the cellular mechanisms in-volved in the triggering of lysis of S. thermophilus and to specifythe link between the autolytic phenotype and lysogeny in thisspecies. For this purpose, 6 different S. thermophilus strainsidentified as autolytic out of 146 S. thermophilus strainsscreened were further characterized in this work. All of themwere found to be lysogenic. Different environmental factors,such as lactose depletion and organic solvents, were identifiedas triggers of premature lysis. A bacteriolytic enzyme of 31 kDawas detected by renaturing SDS-PAGE exclusively in the au-tolytic strains. It was shown to be prophage encoded and ho-mologous to the endolysin Lyt51 of the streptococcal temper-ate bacteriophage f01205 (44). From all these results, wepropose a mechanism of lysis triggering according to which S.thermophilus lysis is triggered and achieved under unfavorableenvironmental conditions via the lysis proteins of a leaky pro-phage.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. The autolytic S. ther-mophilus strains were obtained from two different collections: strains DN-001065and DN-001561 were from the Centre International de Recherche DanielCarasso collection (Danone, Le Plessis-Robinson, France), and strains CNRZ701, CNRZ 1358 (type strain), CNRZ 1205, and CNRZ 1209 were from theCNRZ collection (Institut National de la Recherche Agronomique [INRA],Jouy-en-Josas, France). Nonautolytic S. thermophilus strains CNRZ 302 andCNRZ 1446 were obtained from the INRA collection. Strain CNRZ 1446 is anisogenic representative of the type strain, which was nonautolytic, as opposed tostrain CNRZ 1358. The nonautolytic strains were grown at 42°C in M17 broth(49) (Difco Laboratories, Detroit, Mich.) supplemented with 0.75% (wt/vol)lactose. The autolytic strains were grown in M17 medium containing a highlactose concentration (1.5% [wt/vol]) to prevent lysis or a limited lactose con-centration (1% [wt/vol] for strain DN-001065 and 0.5% [wt/vol] for the otherautolytic strains) to ensure lysis. Growth and lysis were monitored by measuringthe optical density at 580 nm (OD580) in 1-cm cuvettes with a spectrophotometer(model Uvikon 931; Kontron Instruments Inc., Everett, Mass.). Escherichia coliXL1-Blue recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [F9 proAB lacIqZDM15 Tn10 (Tetr)] (Stratagene) was cultured in Luria-Bertani broth (42) con-taining tetracycline (12.5 mg ml21) at 37°C with shaking. The pQE30 plasmidvector (Qiagen) conferring ampicillin resistance was used for high-level expres-sion of six-His-tagged proteins. When needed, ampicillin was used at a concen-tration of 100 mg ml21.
Lysis experiments. The effect of lactose depletion on the triggering of lysis wasstudied as follows. Bacteria grown in M17 medium containing a limited lactoseconcentration were harvested after different culture times by centrifugation at7,500 3 g for 15 min at 15°C and washed once in distilled water at roomtemperature. The cells were then resuspended in the same volume of either freshM17 broth or 50 mM Tris-HCl (pH 7.0) buffer, either supplemented with 0.5%(wt/vol) lactose or devoid of lactose. When specified, the purified recombinantendolysin (six-His-tagged) Lyt51 (see below) was added at various concentra-tions. The cell suspensions were then incubated at 42°C. Cell lysis was monitoredby measuring the OD580 of the bacterial suspensions. The extent of lysis wasexpressed as the percent decrease of OD580 after a given time, and the rate oflysis was expressed as the decrease in OD580 per minute during the first 60 min.
Lactose concentration was determined in cell-free culture supernatants byenzymatic analysis with the lactose/D-galactose determination kit (BoehringerMannheim) according to the supplier’s instructions.
Compounds tested for their ability to trigger a premature lysis were added toexponential-phase cultures. The resulting volume changes were always less than2%, and temperature was held constant.
Mitomycin C induction. S. thermophilus strains were grown in M17 broth at42°C to an OD580 of 0.3. Mitomycin C (Sigma Chemical Co., St. Louis, Mo.) wasadded at a final concentration of 0.2 mg ml21 as described previously (20). Theculture was further incubated at 42°C, and the OD580 was monitored regularly.A culture grown in M17 broth at 42°C was used as the control.
Bacteriophage preparation and electron microscopy observation. The auto-lytic strains were grown in 250 ml of M17 medium. Fifty milliliters of the culturewas induced with mitomycin C as described above, and the remainder of theculture was grown until lysis occurred spontaneously. In both cases, after lysiswas completed, the lysate was incubated for 1 h at 37°C with DNase I (5 mgml21), RNase (12.5 mg ml21), and MgCl2 (1 mM). Lysozyme (1 mg ml21) andmutanolysin (Sigma) (500 U ml21) were also added at this step to ensure anextensive release of phage particles. After addition of NaCl at the final concen-tration of 0.5 M, the lysate was further incubated on ice for 20 min. Cell debriswas subsequently eliminated by a low-speed centrifugation (5,000 3 g, 15 min).Phage particles were collected from the supernatant by centrifugation at
100,000 3 g for 2 h at 15°C in an ultracentrifuge (model Centrikon T-1080;Kontron Instruments Inc.). The phage pellet was resuspended in 240 ml of TMbuffer (10 mM Tris-HCl, 10 mM MgSO4, pH 8.0), and the suspension was filteredthrough a 0.45-mm-pore-size filter (Millex-HA; Millipore S.A., Bedford, Mass.).
Phage particles were then negatively stained with 2% (wt/vol) uranyl acetate,as previously described (1), and observed with a Zeiss model EM-10 electronmicroscope at an accelerating voltage of 80 kV.
DNA techniques. Restriction enzymes and T4 DNA ligase were respectivelyobtained from Eurogentec and Boehringer Mannheim and were used as recom-mended by the suppliers. Molecular cloning, purification, and analysis of DNAwere performed by standard procedures (42).
PCR was carried out in a model 2400 Gene Amp PCR system (Perkin-Elmer,Norwalk, Conn.) with Taq DNA polymerase according to the instructions of themanufacturer (Appligene-Oncor, Inc., Gaithersburg, Md.). Total DNA was iso-lated from S. thermophilus strains as reported by Chapot-Chartier et al. (14).
Total DNA was digested with the appropriate restriction enzymes, electro-phoresed in a 0.7% agarose gel, and blotted onto a Hybond-N1 nylon membrane(Amersham International, Amersham, United Kingdom) by the Southernmethod as described by Sambrook et al. (42). A DNA probe corresponding to an825-bp fragment of the S. thermophilus temperate phage f01205 endolysin gene,lyt51 (48) was amplified by PCR from the total DNA of S. thermophilus CNRZ1205. Primers LYS-1 (59-ATGAGCGTAAAACAAAAACTA; position 1 to 21)and LYS-2 (59-GTCGTCCTTATTCCAGCAAGA; position 825 to 805) used forthis purpose were deduced from the previously published lyt51 sequence (48).The probe was labeled with [a-32P]dCTP by using the Nick Translation DNAlabeling kit (Amersham). Hybridization experiments were done under high-stringency conditions (50% formamide, 42°C) according to a standard protocol(42).
SDS-PAGE and renaturing SDS-PAGE. SDS-PAGE was carried out as de-scribed by Laemmli (27) with a Mini Protean II cell unit (Bio-Rad Laboratories,Inc., Hercules, Calif.) and a gel size of 75 by 55 mm. Samples to be analyzed weremixed with sample loading buffer containing (final concentrations) 62.5 mMTris-HCl (pH 6.8), 2.3% (wt/vol) SDS, 50 mM dithiothreitol, 10% (vol/vol)glycerol, and 0.01% (wt/vol) bromophenol blue. They were then heated for 3 minat 100°C.
Renaturing SDS-PAGE was performed as previously described (29, 40). Mi-crococcus lysodeikticus ATCC 4698 (Sigma) autoclaved cells (0.2% [wt/vol]) or S.thermophilus CNRZ 302 autoclaved cells (0.4% [wt/vol]) were included in 12.5%polyacrylamide gels as a substrate for the bacteriolytic enzymes. After electro-phoresis, the gel was washed in distilled water for 30 min at room temperaturewith gentle shaking. It was thereafter transferred into renaturation buffer con-taining 50 mM MES (2-morpholinoethanesulfonic acid) (Sigma), NaOH (pH6.0), and 0.1% (wt/vol) Triton X-100. It was incubated for 16 h at 37°C undergentle shaking. It was rinsed with distilled water, stained with 0.1% (wt/vol)methylene blue in 0.01% (wt/vol) KOH for 2 h at room temperature with gentleshaking, and destained with distilled water. The bacteriolytic activities appear asclear bands on a blue background.
Molecular mass was determined by comparison with prestained molecularmass standards separated by electrophoresis on the same gel. The prestainedstandards were purchased from Bio-Rad Laboratories and contained phosphor-ylase B (107,000 Da), bovine serum albumin (74,000 Da), ovalbumin (49,300Da), carbonic anhydrase (36,400 Da), soybean trypsin inhibitor (28,500 Da), andlysozyme (20,900 Da).
Preparation of cell extracts for renaturing SDS-PAGE. SDS cell extracts wereprepared from exponential-phase cells recovered by centrifugation at 7,500 3 gfor 15 min at 4°C. The cell pellet was resuspended in sample loading buffer,heated for 3 min at 100°C, and centrifuged at 10,000 3 g for 15 min. Thesupernatant (SDS cell extract) was analyzed by renaturing SDS-PAGE.
In order to monitor the expression levels of bacteriolytic enzymes duringgrowth and lysis, cells were harvested by centrifugation at 7,500 3 g for 15 minat 4°C from cultures at different times. The culture supernatant was filteredthrough 0.45-mm-pore-size filters (Milex-HA) and was tested for the presence ofbacteriolytic activities. The cells were resuspended in 50 mM Tris-HCl buffer(pH 7.0) containing 1 mM EDTA and 1 mM AEBSF [4-(2-aminoethyl)benze-nesulfonyl fluoride hydrochloride] (Interchim) and were disrupted with glassbeads 0.1 mm in diameter in a Mini Beadbeater T8M cell disrupter (BiospecProducts, Bartlesville, Ill.) by three 1-min cycles with 1 min cooling on ice aftereach cycle. Glass beads, unbroken cells, and debris were removed by centrifu-gation at 8,000 3 g for 20 min. The protein concentration in the supernatants(glass bead cell extracts) was determined by the Bradford method with theCoomassie protein assay reagent as specified by Pierce Chemical Company(Rockford, Ill.) with bovine serum albumin as the standard. The same proteinamount in each cell extract was analyzed by renaturing SDS-PAGE for thepresence of bacteriolytic activities.
Expression of the Lyt51 endolysin in E. coli, purification of the recombinantprotein, and preparation of antiserum. The Lyt51 endolysin (44) was expressedin E. coli XL1-Blue with an N-terminal six-His tag by using expression vectorpQE30 (Qiagen). The lyt51 endolysin gene was amplified by PCR from S. ther-mophilus CNRZ 1205 total DNA with the primers LYS-3 (59-GGCATGCAGCGTAAAACAAAA [position 4 to 17]) and LYS-4 (59-GGAAGCTTCGTGGTCTATTTG [position 852 to 840]) containing, respectively, SphI and HindIII sitesat their 59 ends and cloned in frame downstream of the six-His box coding
VOL. 66, 2000 AUTOLYTIC S. THERMOPHILUS STRAINS 559

sequence in the pQE30 expression vector. E. coli XL1-Blue competent cells(Stratagene) were transformed with the resulting pTIL72 plasmid. The nucleo-tide sequence of the pTIL72 construction was verified.
The expression of the six-His-tagged Lyt51 was induced by IPTG (isopropyl-b-D-thiogalactopyranoside; 1 mM), according to the Qiagen procedure, in E. coliXL1-Blue harboring the pTIL72 plasmid. After 4 h of induction, the cells werecollected by centrifugation at 4,000 3 g for 15 min and resuspended in 1/25volume of column binding buffer (50 mM NaH2PO4 [pH 8.0], 300 mM NaCl, 10mM imidazole). The cell suspension was then frozen at 220°C. After 2 h it wasthawed at room temperature and replaced in the freezer. This procedure wasrepeated four times, and the cells were finally broken by three ultrasonicationpulses of 20 s with 1-min cooling intervals in an ice bath. Insoluble cell debris wasremoved by centrifugation at 10,000 3 g for 15 min, and the His-tagged Lyt51was purified from the supernatant (cleared lysate) by metal affinity chromatog-raphy on a nickel-nitrilotriacetic acid (Ni-NTA) spin column (Qiagen) accordingto the manufacturer’s instructions. Following elution with imidazole, the purifiedLyt51 protein was dialyzed for 16 h against phosphate-buffered Saline (PBS; 10mM sodium phosphate [pH 7.4], 100 mM NaCl) at 4°C. The protein concentra-tion was determined with the Coomassie protein assay reagent (Pierce).
Purified Lyt51 was injected into a rabbit to raise antibodies (Biological Ser-vices Unit, University College of Cork, Cork, Ireland). Four injections of 50 mgof Lyt51 were performed at 1-week intervals.
The antibodies directed against the His-tagged Lyt51 were purified by affinitychromatography from the rabbit serum obtained 7 days after the third boosterinjection, according to the procedure of Gu et al. (22). Briefly, the His-taggedLyt51 from an E. coli-cleared lysate was bound to a Ni-NTA spin column andthen 600 ml of the crude rabbit antiserum was applied to the column and left tostand for 20 min. The serum was then allowed to filter through, and the columnwas washed five times with 600 ml of 150 mM NaCl–50 mM Tris-HCl, pH 7.4,followed by five washes with 600 ml of 2 M NaCl–50 mM Tris-HCl, pH 7.4. TheLyt51-specific antibodies were thereafter eluted with 600 ml of 4 M MgCl2solution (pH 4.5) and dialyzed against water for 1 h and then against PBS for 16 hat 4°C.
Immunoblotting analysis. Immunoblotting was carried out as described byTowbin and Gordon (51) using the purified anti-Lyt51 antibodies. The sameprotein amount from each SDS cell extract and the purified His-tagged Lyt51 (2ng) were electrophoresed on a 12.5% SDS-polyacrylamide gel and transferred toa nitrocellulose membrane (Schleicher & Schuell). After incubation with theprimary anti-Lyt51 antibodies (1/500), the membrane was incubated with proteinG-horseradish peroxidase conjugate (Bio-Rad) (1/3,000). Antigen-antibody com-plexes were subsequently detected using chemiluminescence with the ECL PlusWestern blotting detection kit (Amersham) according to the manufacturer’sinstructions.
RESULTS
Lactose depletion triggers the lysis of the autolytic S. ther-mophilus strains. S. thermophilus DN-001065 (Fig. 1A) as wellas the other five S. thermophilus autolytic strains (data notshown) exhibited a typical bell-shaped growth curve whengrown at 42°C in M17 medium supplemented with lactose inlimited concentrations (1% for strain DN-001065 and 0.5% forthe other strains). After reaching maximal growth, cells lysedrapidly, resulting in a sharp decrease of OD580. The onset oflysis was coincident with lactose exhaustion from the medium(data not shown).
These results suggested that lactose depletion was the trig-gering factor for cell lysis. To confirm this hypothesis, the lyticbehavior of exponential-phase cells transferred in fresh M17medium devoid of lactose was examined. Upon their transfer,the cells lysed immediately with a lysis rate (1.4 3 1022 OD580unit min21 for strain DN-001065) similar to that of spontane-ous lysis occurring at the end of the exponential-growth phase(Fig. 1A). When transferred in M17 medium containing lac-tose, the exponential-phase cells did not lyse and resumedgrowth after a short lag phase (Fig. 1A). The lysis phenomenonobserved in the absence of lactose was specific to the autolyticstrains. Indeed, when nonautolytic strain CNRZ 1446 cellswere used in the same experiment, they stopped growing butdid not lyse (data not shown).
In addition, we observed that lactose can arrest the lysis ofthe autolytic strains. When a lysing culture was transferred intofresh M17 medium containing lactose, lysis stopped immedi-
ately and a renewed growth phase was observed. In absence oflactose, lysis continued (Fig. 1A).
The protective role of lactose against lysis was also investi-gated with buffer solutions. For this purpose, S. thermophilusautolytic strains at the beginning of the lysing phase in M17medium were transferred in 50 mM Tris-HCl buffer, pH 7.0,supplemented with 0.5% lactose or not supplemented, andincubated at 42°C. In the absence of lactose, the cell suspen-sion turbidity decreased linearly during 2 h (Fig. 1B). Forstrain DN-001065 the extent of lysis reached 70% after 19 h ofincubation. By contrast, when lactose was present in the buffer,the OD580 remained stable over the same period of time.These results thus show that lactose also protects resting cellsagainst lysis.
Thus, the depletion of lactose, which is the unique carbonsource in the growth medium, appears to be the trigger of celllysis. The fact that even exponential-phase cells lyse underlactose depletion suggests that the triggering of lysis does notrequire the accumulation in the cells of lysis effectors at the
FIG. 1. Effect of lactose depletion on S. thermophilus DN-001065 lysis. (A) S.thermophilus DN-001065 was grown in M17 medium with 1% lactose at 42°C (E).Cells were harvested during the exponential-growth phase (h, ■) or during thelysing phase (‚, Œ). They were resuspended in the same volume of fresh M17medium with 0.5% lactose (h, ‚) or devoid of lactose (■, Œ) and furtherincubated at 42°C. Arrows indicate the times at which cells were harvested. (B)Cells were harvested at the beginning of the lysing phase, washed in distilledwater at room temperature, and resuspended in the same volume of 50 mMTris-HCl buffer, pH 7.0, with 0.5% lactose (‚) or devoid of lactose (Œ). The cellsuspensions were further incubated at 42°C.
560 HUSSON-KAO ET AL. APPL. ENVIRON. MICROBIOL.
end of the exponential-growth phase but rather that these lysiseffectors are present in the cells at any stage of growth.
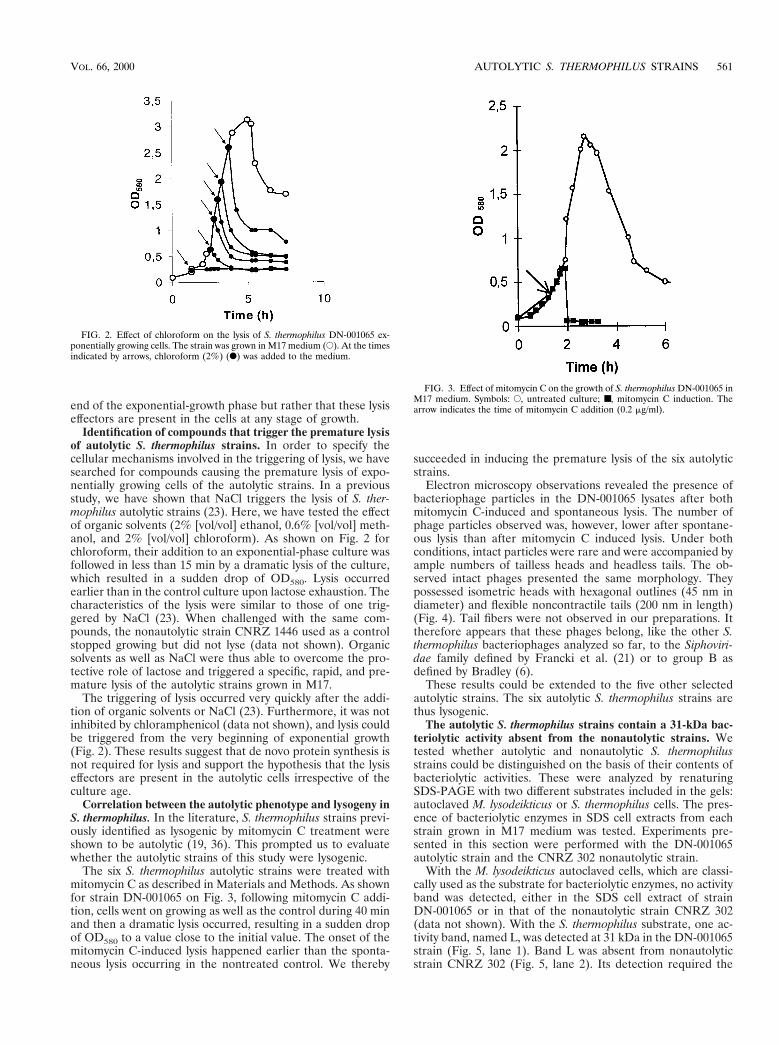
Identification of compounds that trigger the premature lysisof autolytic S. thermophilus strains. In order to specify thecellular mechanisms involved in the triggering of lysis, we havesearched for compounds causing the premature lysis of expo-nentially growing cells of the autolytic strains. In a previousstudy, we have shown that NaCl triggers the lysis of S. ther-mophilus autolytic strains (23). Here, we have tested the effectof organic solvents (2% [vol/vol] ethanol, 0.6% [vol/vol] meth-anol, and 2% [vol/vol] chloroform). As shown on Fig. 2 forchloroform, their addition to an exponential-phase culture wasfollowed in less than 15 min by a dramatic lysis of the culture,which resulted in a sudden drop of OD580. Lysis occurredearlier than in the control culture upon lactose exhaustion. Thecharacteristics of the lysis were similar to those of one trig-gered by NaCl (23). When challenged with the same com-pounds, the nonautolytic strain CNRZ 1446 used as a controlstopped growing but did not lyse (data not shown). Organicsolvents as well as NaCl were thus able to overcome the pro-tective role of lactose and triggered a specific, rapid, and pre-mature lysis of the autolytic strains grown in M17.
The triggering of lysis occurred very quickly after the addi-tion of organic solvents or NaCl (23). Furthermore, it was notinhibited by chloramphenicol (data not shown), and lysis couldbe triggered from the very beginning of exponential growth(Fig. 2). These results suggest that de novo protein synthesis isnot required for lysis and support the hypothesis that the lysiseffectors are present in the autolytic cells irrespective of theculture age.
Correlation between the autolytic phenotype and lysogeny inS. thermophilus. In the literature, S. thermophilus strains previ-ously identified as lysogenic by mitomycin C treatment wereshown to be autolytic (19, 36). This prompted us to evaluatewhether the autolytic strains of this study were lysogenic.
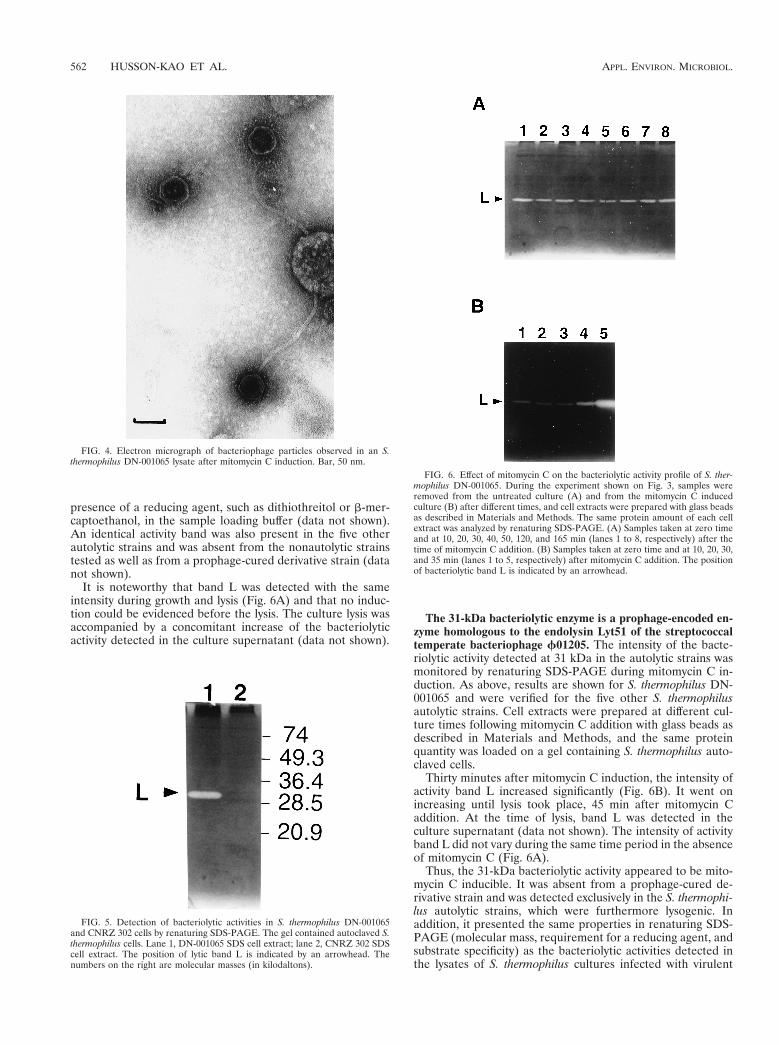
The six S. thermophilus autolytic strains were treated withmitomycin C as described in Materials and Methods. As shownfor strain DN-001065 on Fig. 3, following mitomycin C addi-tion, cells went on growing as well as the control during 40 minand then a dramatic lysis occurred, resulting in a sudden dropof OD580 to a value close to the initial value. The onset of themitomycin C-induced lysis happened earlier than the sponta-neous lysis occurring in the nontreated control. We thereby
succeeded in inducing the premature lysis of the six autolyticstrains.
Electron microscopy observations revealed the presence ofbacteriophage particles in the DN-001065 lysates after bothmitomycin C-induced and spontaneous lysis. The number ofphage particles observed was, however, lower after spontane-ous lysis than after mitomycin C induced lysis. Under bothconditions, intact particles were rare and were accompanied byample numbers of tailless heads and headless tails. The ob-served intact phages presented the same morphology. Theypossessed isometric heads with hexagonal outlines (45 nm indiameter) and flexible noncontractile tails (200 nm in length)(Fig. 4). Tail fibers were not observed in our preparations. Ittherefore appears that these phages belong, like the other S.thermophilus bacteriophages analyzed so far, to the Siphoviri-dae family defined by Francki et al. (21) or to group B asdefined by Bradley (6).
These results could be extended to the five other selectedautolytic strains. The six autolytic S. thermophilus strains arethus lysogenic.
The autolytic S. thermophilus strains contain a 31-kDa bac-teriolytic activity absent from the nonautolytic strains. Wetested whether autolytic and nonautolytic S. thermophilusstrains could be distinguished on the basis of their contents ofbacteriolytic activities. These were analyzed by renaturingSDS-PAGE with two different substrates included in the gels:autoclaved M. lysodeikticus or S. thermophilus cells. The pres-ence of bacteriolytic enzymes in SDS cell extracts from eachstrain grown in M17 medium was tested. Experiments pre-sented in this section were performed with the DN-001065autolytic strain and the CNRZ 302 nonautolytic strain.
With the M. lysodeikticus autoclaved cells, which are classi-cally used as the substrate for bacteriolytic enzymes, no activityband was detected, either in the SDS cell extract of strainDN-001065 or in that of the nonautolytic strain CNRZ 302(data not shown). With the S. thermophilus substrate, one ac-tivity band, named L, was detected at 31 kDa in the DN-001065strain (Fig. 5, lane 1). Band L was absent from nonautolyticstrain CNRZ 302 (Fig. 5, lane 2). Its detection required the
FIG. 2. Effect of chloroform on the lysis of S. thermophilus DN-001065 ex-ponentially growing cells. The strain was grown in M17 medium (E). At the timesindicated by arrows, chloroform (2%) (F) was added to the medium.
FIG. 3. Effect of mitomycin C on the growth of S. thermophilus DN-001065 inM17 medium. Symbols: E, untreated culture; ■, mitomycin C induction. Thearrow indicates the time of mitomycin C addition (0.2 mg/ml).
VOL. 66, 2000 AUTOLYTIC S. THERMOPHILUS STRAINS 561
presence of a reducing agent, such as dithiothreitol or b-mer-captoethanol, in the sample loading buffer (data not shown).An identical activity band was also present in the five otherautolytic strains and was absent from the nonautolytic strainstested as well as from a prophage-cured derivative strain (datanot shown).
It is noteworthy that band L was detected with the sameintensity during growth and lysis (Fig. 6A) and that no induc-tion could be evidenced before the lysis. The culture lysis wasaccompanied by a concomitant increase of the bacteriolyticactivity detected in the culture supernatant (data not shown).
The 31-kDa bacteriolytic enzyme is a prophage-encoded en-zyme homologous to the endolysin Lyt51 of the streptococcaltemperate bacteriophage f01205. The intensity of the bacte-riolytic activity detected at 31 kDa in the autolytic strains wasmonitored by renaturing SDS-PAGE during mitomycin C in-duction. As above, results are shown for S. thermophilus DN-001065 and were verified for the five other S. thermophilusautolytic strains. Cell extracts were prepared at different cul-ture times following mitomycin C addition with glass beads asdescribed in Materials and Methods, and the same proteinquantity was loaded on a gel containing S. thermophilus auto-claved cells.
Thirty minutes after mitomycin C induction, the intensity ofactivity band L increased significantly (Fig. 6B). It went onincreasing until lysis took place, 45 min after mitomycin Caddition. At the time of lysis, band L was detected in theculture supernatant (data not shown). The intensity of activityband L did not vary during the same time period in the absenceof mitomycin C (Fig. 6A).
Thus, the 31-kDa bacteriolytic activity appeared to be mito-mycin C inducible. It was absent from a prophage-cured de-rivative strain and was detected exclusively in the S. thermophi-lus autolytic strains, which were furthermore lysogenic. Inaddition, it presented the same properties in renaturing SDS-PAGE (molecular mass, requirement for a reducing agent, andsubstrate specificity) as the bacteriolytic activities detected inthe lysates of S. thermophilus cultures infected with virulent
FIG. 4. Electron micrograph of bacteriophage particles observed in an S.thermophilus DN-001065 lysate after mitomycin C induction. Bar, 50 nm.
FIG. 5. Detection of bacteriolytic activities in S. thermophilus DN-001065and CNRZ 302 cells by renaturing SDS-PAGE. The gel contained autoclaved S.thermophilus cells. Lane 1, DN-001065 SDS cell extract; lane 2, CNRZ 302 SDScell extract. The position of lytic band L is indicated by an arrowhead. Thenumbers on the right are molecular masses (in kilodaltons).
FIG. 6. Effect of mitomycin C on the bacteriolytic activity profile of S. ther-mophilus DN-001065. During the experiment shown on Fig. 3, samples wereremoved from the untreated culture (A) and from the mitomycin C inducedculture (B) after different times, and cell extracts were prepared with glass beadsas described in Materials and Methods. The same protein amount of each cellextract was analyzed by renaturing SDS-PAGE. (A) Samples taken at zero timeand at 10, 20, 30, 40, 50, 120, and 165 min (lanes 1 to 8, respectively) after thetime of mitomycin C addition. (B) Samples taken at zero time and at 10, 20, 30,and 35 min (lanes 1 to 5, respectively) after mitomycin C addition. The positionof bacteriolytic band L is indicated by an arrowhead.
562 HUSSON-KAO ET AL. APPL. ENVIRON. MICROBIOL.

phages (data not shown). All together, these findings suggestedthat the 31-kDa enzyme was encoded by a prophage.
As it was shown that genes involved in the lysis module ofS. thermophilus bacteriophages were highly conserved at theDNA sequence level (17, 44), we examined whether activityband L could be a bacteriolytic enzyme homologous to therecently identified endolysin Lyt51 of S. thermophilus temper-ate bacteriophage f01205 (44).
We first investigated the presence of a gene homologous tolyt51 in the six autolytic strains by probing strain DNA digestswith the PCR-generated fragment consisting of nucleotides 1to 825 of lyt51 (data not shown). The results showed that the sixautolytic strains contain a DNA fragment which is not presentin the nonautolytic strain CNRZ 302 and which hybridizes withthe lyt51 probe.
We then examined whether a protein similar to Lyt51 couldbe detected in the autolytic strains. For this purpose, antibod-ies were raised against Lyt51, which was overexpressed as asix-His-tagged fusion protein in E. coli and purified by Ni-NTAaffinity chromatography. The antibodies were used in immu-noblotting experiments on SDS cell extracts of the autolyticstrains.
In the S. thermophilus DN-001065 SDS cell extract, a proteinof 31 kDa was recognized by the purified antibodies (Fig. 7,lane 2) and the band intensity was higher in the mitomycinC-induced cells (Fig. 7, lane 3). One band was observed at thesame molecular mass in the cell extracts of the five otherautolytic strains as well (data not shown). The purified His-tagged Lyt51 used for antibody production was used as thecontrol (Fig. 7, lane 1). The observed difference in electro-phoretical mobility between the His-tagged endolysin and theprotein detected in the DN-001065 cell extracts can be attrib-uted to the fact that the molecular mass of the recombinantprotein (32.2 kDa) is higher than that of the native protein(31.1 kDa) due to the six-His tag fusion. As expected, no bandwas detected at 31 kDa in the SDS cell extract of S. thermophi-lus CNRZ 302 (Fig. 7, lane 4). Protein bands detected at about74 kDa, as well as the one at 29 kDa, were equally present instrain CNRZ 302 and thus appear nonspecific.
Thus, the 31-kDa bacteriolytic activity detected in the S.thermophilus autolytic strains is encoded by a prophage and is
homologous to the Lyt51 endolysin identified in the f01025temperate bacteriophage of S. thermophilus.
Effect of exogenous endolysin on the growth and lysis of S.thermophilus autolytic strains. The effect of exogenous endo-lysin on the growth and lysis of the autolytic DN-001065 strainwas tested. For this purpose, exponential-phase cells were col-lected and resuspended in fresh M17 medium supplementedwith lactose and several amounts (0, 10, 25, and 100 ng ml21)of purified six-His-tagged Lyt51. The concentration of 100 ngml21 corresponds to the maximum amount that is released inthe culture supernatant by spontaneous lysis, as estimated byrenaturing SDS-PAGE.
At concentrations of 25 ng ml21 and below, the endolysinLyt51 has no effect on growth and lysis triggering; the culturesbehave like the control without endolysin (data not shown). Ata concentration of 100 ng ml21, the growth rate was reducedbut lysis was triggered at the same time as for the control (datanot shown).
These results thus rule out the hypothesis that the cells lysedfrom the outside, that is, from the action of endolysin releasedby a fraction of the cell population. They rather indicate thatmost of the lysis observed in response to the identified trigger-ing factors is from within.
DISCUSSION
S. thermophilus autolytic strains are characterized by a typ-ical bell-shaped growth curve, reflecting their propensity todramatically lyse after a normal growth phase. The cellularmechanisms involved in triggering the lysis of these strainswere investigated in the present study. Lysis was rapidly trig-gered under unfavorable environmental conditions such as lac-tose depletion and organic solvent addition. This autolytic phe-notype appears to be linked to the lysogenic character of thestrains. It is also associated with the presence of a 31-kDaprophage-encoded endolysin, homologous to Lyt51 of thestreptococcal temperate bacteriophage f01205 (44) and de-tected in the cells during both growth and lysis.
Lysis of the autolytic strains appears to be triggered by thedepletion of lactose, the unique carbon source in the growthmedium. In addition, our results indicate that lactose preventsthe lysis of starved or resting cells. As the triggering of lysis ofS. thermophilus autolytic strains grown on lactose and also onsaccharose was previously reported to be concomitant withsugar depletion from the medium (50, 58), we assumed thatlysis is caused by the carbon source depletion irrespective ofits nature. The cellular integrity of S. thermophilus autolyticstrains thus appears to depend on the presence of metabolicenergy in the medium as observed for some strains in otherbacterial genus such as Enterococcus hirae (46), L. lactis (41),Propionibacterium spp. (30), and Bacillus subtilis (24).
Since S. thermophilus does not contain storage polymers,lactose starvation may immediately result in energy starvationand in the subsequent rapid dissipation of the proton motiveforce (PMF), as described previously for L. lactis (39). Organicsolvents as well as NaCl (23) were shown to remove the pro-tective effect of lactose against lysis. Following their addition,a sharp and immediate lysis, which does not seem to require denovo protein synthesis, occurred. It is worth noting that or-ganic solvents are known to permeabilize the cytoplasmicmembrane and that NaCl was also shown to lower the PMF atconcentrations above 50 mM (26). The environmental factorstriggering the cell lysis thus present the common feature ofaffecting the cell envelope properties by causing either thedepolarization or the permeabilization of the cytoplasmicmembrane.
FIG. 7. Immunoblotting with anti-Lyt51 antibodies. Lane 1, purified six-His-tagged Lyt51; lane 2, DN-001065; lane 3, mitomycin C-induced DN-001065; lane4, CNRZ 302. The SDS cell extracts were prepared 40 min after mitomycin Cinduction of strain DN-001065. For each cell extract, the same protein amountwas transferred on a nitrocellulose membrane. The blot was incubated with thepurified antibodies directed against the six-His-tagged Lyt51 and then with pro-tein G coupled to horseradish peroxidase. Positions of the six-His-tagged Lyt51and the 31-kDa protein reacting with the anti-Lyt51 antibodies are indicated byarrowheads.
VOL. 66, 2000 AUTOLYTIC S. THERMOPHILUS STRAINS 563

In the present survey all the strains selected for their auto-lytic character are lysogenic. Reciprocally, according to theliterature (19, 36), all the S. thermophilus lysogenic strainsidentified by mitomycin C induction are autolytic. The uniqueexception is the lysogen St18 described by Carminati and Gi-raffa (11), which did not exhibit the autolytic phenotype; thiswas most probably due to inappropriate growth conditions thatled to lysis inhibition. The relation between the autolytic phe-notype and the presence of a mitomycin C-inducible prophagein the strain genomes thus appears to be symmetrical in S.thermophilus. The involvement of a prophage in the autolyticphenotype is moreover supported by the nonautolytic charac-ter of strains cured for their prophage and the restoration ofthe autolytic phenotype by relysogenization (19, 36). The uni-formity of the S. thermophilus lysogen phenotype might belinked to the recently evidenced genetic similarity among thestreptococcal temperate bacteriophages (8, 17, 18, 20, 48) andtheir lysogeny modules (37).
A unique bacteriolytic enzyme of 31 kDa was detected byrenaturing SDS-PAGE exclusively in the autolytic strains. Weshowed that this enzyme is prophage encoded and homologousto the endolysin Lyt51 of the streptococcal temperate bacte-riophage f01205 (44). The identification of the 31-kDa en-zyme as an endolysin homologous to Lyt51 was based first onthe similar biochemical properties of both enzymes observedby renaturing SDS-PAGE, i.e., their molecular masses andtheir requirement for a reducing agent to be detected as well astheir apparently narrow substrate specificities. The 31-kDa en-zyme identification was further substantiated by the detectionof DNA homologous to lyt51 in the autolytic strains, and finallyconfirmed by the immunodetection of a 31-kDa protein react-ing with anti-Lyt51 antibodies.
Although the autolytic phenotype of the S. thermophilusstrains is linked to their lysogenic character and is observedunder unfavorable physiological conditions, cell lysis does notappear to result from massive prophage induction in responseto a stress. Indeed, lysis resulting from lysogenic induction,such as that caused by mitomycin C treatment, occurs afterabout 45 min, and, over this period of time, expression ofphage genes is massively induced, as shown for the 31-kDaendolysin. By contrast, following lactose depletion and organicsolvent or NaCl addition, lysis occurs more quickly and doesnot appear to require de novo protein synthesis. In addition,the 31-kDa endolysin is detected with the same intensity up tothe onset of lysis. It thus appears that the two lysis events donot result from the same triggering mechanism and thereforethat the autolytic phenotype is not caused by massive prophageinduction.
It rather seems that the autolytic phenotype results from anincomplete prophage repression. Several observations suggestthat the S. thermophilus lysogens failed to efficiently control theprophage state; linearized phage DNA (7, 19) and the 31-kDaendolysin were detected in cells that were not treated withmitomycin C, and phage particles were liberated by the spon-taneous lysis of the lysogens. As an extensive culture lysis wastriggered under appropriate conditions and as most of the celllysis appears to come from within, we assumed that the pro-phage induction takes place in almost all the cells and not onlyin a fraction of the population. According to this hypothesis,the prophage lysis proteins are also expressed in the cells, asshown for the endolysin. In S. thermophilus phages, as in mostbacteriophages, the lysis system contains, in addition to theendolysin, holin proteins which are required by the endolysinto pass through the cytoplasmic membrane and reach the pep-tidoglycan (17, 44, 48). The high cell toxicity of these mem-brane-embedded proteins implies that the prophage induction
occurs at a low and therefore nonlethal level. We propose thatthe prophage lysis proteins control and achieve cell lysis andare thus responsible for the autolytic phenotype. It has beensuggested that the culture lysis resulting from the endolysinaction can be viewed as a reporter event for holin function (4).The involvement of holin proteins in lysis triggering was sug-gested by the fact that the environmental triggers of S. ther-mophilus lysis, which are factors leading to the membranedepolarization, are also known to trigger premature lysis ofinduced E. coli lambda lysogens by causing premature holeformation by holins (4, 56, 57). These data suggest that non-induced S. thermophilus lysogens express holin proteins at anonlethal level and that, according to the well-documentedmodel of lysis triggering in lambda lysogens, collapse of analready unstable membrane potential, which is weakened be-cause of the expression of holins, would cause the conversionof holins from a dormant to an active state. Our hypothesis isthus that the prophage lysis proteins, which are the 31-kDaendolysin and the holins, would accomplish lysis and that thescheduling of lysis would be controlled by mechanisms govern-ing the activity of the holins present in small amounts in themembranes during growth.
Because they can lyse in response to several specific envi-ronmental signals such as NaCl (23), lactose depletion, andorganic solvents, S. thermophilus lysogens appear to be a veryuseful tool to evaluate the impact of cell lysis on the flavordevelopment of fermented dairy products. In respect to thesignificant problem of phage infection in the dairy industry, therestriction to their wide industrial use is their lysogenic char-acter, although we did not detect virulent phages in the lysissupernatants. Further studies will thus be required in order tospecify the role of S. thermophilus lysogeny in phage infection.
ACKNOWLEDGMENTS
This work was supported by the Centre International de RechercheDaniel Carasso of the Danone Group. C.H.-K. was subsidized by aCIFRE convention (no. 95762) from the French Association de laRecherche Technique and the T3Net program from the Commissionof the European Communities.
We thank Patrick Taillez for the access to the randomly amplifiedpolymorphic DNA classification of the S. thermophilus strains from theCNRZ collection. We acknowledge Michelle Sheehan for helpful ad-vice on Lyt51 purification and Kate Pollock for antiserum production.We are very grateful to Jean-Claude Gripon and Gerard Denariaz fortheir interest in this work. We warmly thank Bertrand Nicolas forphotographic work.
REFERENCES
1. Accolas, J.-P., and H. Spillman. 1979. The morphology of six bacteriophagesof Streptococcus thermophilus. J. Appl. Bacteriol. 47:135–144.
2. Bie, R., and G. Sjostrom. 1975. Autolytic properties of some lactic acidbacteria used in cheese production. Part I: Material and methods. Milchwis-senschaft 30:653–657.
3. Bie, R., and G. Sjostrom. 1975. Autolytic properties of some lactic acidbacteria used in cheese production. Part II. Experiments with fluid substratesand cheese. Milchwiss. 30:739–747.
4. Blasi, U., and R. Young. 1996. Two beginnings for a single purpose: thedual-start holins in the regulation of phage lysis. Mol. Microbiol. 21:675–682.
5. Botazzi, V., B. Battistotti, M. Vescovo, A. Rebecchi, and F. Bianchi. 1992.Development and lysis of homofermentative thermophilic lactobacilli micro-colonies in grana cheese. Ann. Microbiol. Enzimol. 42:227–247.
6. Bradley, D. E. 1967. Ultrastructure of bacteriophages and bacteriocins. Bac-teriol. Rev. 31:230–314.
7. Brussow, H., and A. Bruttin. 1995. Characterization of a temperate Strepto-coccus thermophilus bacteriophage and its genetic relationship with lyticphages. Virology 212:632–640.
8. Brussow, H., A. Probst, M. Fremont, and J. Sidoti. 1994. Distinct Strepto-coccus thermophilus bacteriophages share an extremely conserved DNA frag-ment. Virology 200:854–857.
9. Buist, G., J. Kok, K. J. Leenhouts, M. Dabrowska, G. Venema, and A. J.Haandrikman. 1995. Molecular cloning and nucleotide sequence of the gene
564 HUSSON-KAO ET AL. APPL. ENVIRON. MICROBIOL.

encoding the major peptidoglycan hydrolase of Lactococcus lactis, a mur-amidase needed for cell separation. J. Bacteriol. 177:1554–1563.
10. Cappa, F., and V. Botazzi. 1996. Characterization of autolytic enzymes inLactobacillus casei subsp. casei 1St261. Ann. Microbiol. Enzimol. 46:299–310.
11. Carminati, D., and G. Giraffa. 1992. Evidence and characterization of tem-perate bacteriophage in Streptococcus thermophilus subsp. thermophilus St18.J. Dairy Res. 59:71–79.
12. Chapot-Chartier, M.-P. 1996. Les autolysines des bacteries lactiques. Lait76:91–109.
13. Chapot-Chartier, M.-P., C. Deniel, M. Rousseau, L. Vassal, and J. C. Gri-pon. 1994. Autolysis of two different strains of Lactococcus lactis duringcheese ripening. Int. Dairy J. 4:251–269.
14. Chapot-Chartier, M.-P., F. Rul, M. Nardi, and J.-C. Gripon. 1994. Genecloning and characterization of PepC, a cysteine aminopeptidase from Strep-tococcus thermophilus, with sequence similarity to the eukaryotic bleomycinhydrolase. Eur. J. Biochem. 224:497–506.
15. Crow, V. L., T. Coolbear, P. K. Gopal, F. G. Martley, L. L. McKay, and H.Riepe. 1995. The role of autolysis of lactic acid bacteria in the ripening ofcheese. Int. Dairy J. 5:855–875.
16. Crow, V. L., F. G. Martley, T. Coolbear, and S. J. Roundhill. 1995. Theinfluence of phage-assisted lysis of Lactococcus lactis subsp. lactis ML8 onCheddar cheese ripening. Int. Dairy J. 5:454–472.
17. Desiere, F., S. Lucchini, and H. Brussow. 1998. Evolution of Streptococcusthermophilus bacteriophage genomes by modular exchanges followed bypoint mutations and small deletions and insertions. Virology 241:345–356.
18. Desiere, F., S. Lucchini, A. Bruttin, M. C. Zwahlen, and H. Brussow. 1997.A highly conserved DNA replication module from Streptococcus thermophi-lus phages is similar in sequence and topology to a module from Lactococcuslactis phages. Virology 234:372–382.
19. Fayard, B. 1993. PhD thesis. Nancy I, Nancy, France.20. Fayard, B., M. Haefliger, and J.-P. Accolas. 1993. Interactions of temperate
bacteriophages of Streptococcus salivarius subsp. thermophilus with lysogenicindicators affect phage DNA restriction patterns and host ranges. J. DairyRes. 60:385–399.
21. Francki, R. I. B., C. M. Fauquet, D. L. Knudson, and D. L. B. 1991. Clas-sification and nomenclature of viruses. Fifth report of the InternationalCommittee on Taxonomy of Viruses. Arch. Virol. Suppl. 2:1–450.
22. Gu, J., C. G. Stephenson, and M. J. Iadarola. 1994. Recombinant proteinsattached to a nickel-NTA column: use in affinity purification of antibodies.BioTechniques 17:257–262.
23. Husson-Kao, C., J. Mengaud, J.-C. Gripon, L. Benbadis, and M.-P. Chapot-Chartier. 2000. The autolysis of S. thermophilus DN-001065 is triggered byseveral food-grade environmental signals. Int. Dairy. J. 9:715–723.
24. Joliffe, L. K., R. J. Doyle, and U. N. Streips. 1981. The energized membraneand cellular autolysis in Bacillus subtilis. Cell 25:753–763.
25. Kawabata, S., L. Vassal, D. LeBars, B. Cesselin, M. Nardi, J.-C. Gripon, andM.-P. Chapot-Chartier. 1997. Phage-induced lysis of Lactococcus lactis dur-ing Saint Paulin cheese ripening and its impact on proteolysis. Lait 77:229–239.
26. Konings, W. N., B. Poolman, and A. J. M. Driessen. 1989. Bioenergetics andsolute transport in lactococci. Crit. Rev. Microbiol. 16:419–476.
27. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 277:680–685.
28. Langsrud, T., A. Landaas, and H. B. Castberg. 1987. Autolytic properties ofdifferent group N streptococci. Milchwissenschaft 42:556–560.
29. Leclerc, D., and A. Asselin. 1989. Detection of bacterial cell wall hydrolasesafter denaturing polyacrylamide gel electrophoresis. Can. J. Microbiol. 35:749–753.
30. Lemee, R., S. Lortal, B. Cesselin, and J. van Heijenoort. 1994. Involvementof an N-acetylglucosaminidase in autolysis of Propionobacterium freuden-reichii CNRZ 725. Appl. Environ. Microbiol. 60:4351–4358.
31. Lepeuple, A. S., E. van Gemert, and M.-P. Chapot-Chartier. 1998. Analysisof the bacteriolytic enzymes of the autolytic Lactococcus lactis subsp. cremo-ris strain AM2 by renaturing polyacrylamide gel electrophoresis: identifica-tion of a prophage-encoded enzyme. Appl. Environ. Microbiol. 64:4142–4148.
32. Lepeuple, A.-S., L. Vassal, B. Cesselin, A. Delacroix-Buchet, J.-C. Gripon,and M.-P. Chapot-Chartier. 1998. Involvement of a prophage in the lysis ofLactococcus lactis subsp. cremoris AM2 during cheese ripening. Int. Dairy J.8:667–674.
33. Lortal, S., P. Boyaval, and J. van Heijenoort. 1989. Influence de plusieurs
facteurs sur l’autolyse de Lactobacillus helveticus CNRZ 414. Lait 69:223–231.
34. Lowrie, R. J., R. C. Lawrence, and M. F. Peberdy. 1974. Cheddar cheeseflavour. V. Influence of bacteriophage and cooking temperatures on cheesemade under controlled bacteriological conditions. New Zealand J. Dairy Sci.Technol. 9:116–121.
35. Meijer, W., C. Dobbelaar, and J. Hugenholtz. 1998. Thermoinducible lysis inLactococcus lactis subsp. cremoris SK 110: implications for cheese ripening.Int. Dairy J. 8:275–280.
36. Mercenier, A. 1990. Molecular genetics of Streptococcus thermophilus. FEMSMicrobiol. Rev. 87:61–78.
37. Neve, H., K. I. Zenz, F. Desiere, A. Koch, K. J. Heller, and H. Brussow. 1998.Comparison of the lysogeny module from the temperate Streptococcus ther-mophilus bacteriophages TP-J34 and Sfi 21: implications for the modulartheory of phage evolution. Virology 241:61–72.
38. Ostlie, H., G. Vegarud, and T. Langsrud. 1995. Autolysis of lactococci:detection of lytic enzymes by polyacrylamide gel electrophoresis and char-acterization in buffer systems. Appl. Environ. Microbiol. 61:3598–3603.
39. Otto, R., J. Vije, B. ten Brink, B. Klont, and W. N. Konings. 1985. Energymetabolism in Streptococcus cremoris during lactose starvation. Arch. Micro-biol. 141:348–352.
40. Potvin, C., D. Leclerc, G. Tremblay, A. Asselin, and G. Bellemare. 1988.Cloning, sequencing and expression of a Bacillus bacteriolytic enzyme inEscherichia coli. Mol. Gen. Genet. 214:241–248.
41. Riepe, H. R., C. J. Pillidge, P. K. Gopal, and L. L. McKay. 1997. Charac-terization of the highly autolytic Lactococcus lactis subsp. cremoris strainsCO and 2250. Appl. Environ. Microbiol. 63:3757–3763.
42. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
43. Sandholm, E., and S. S. Sarimo. 1981. Autolysis of Streptococcus thermophi-lus. FEMS Microbiol. Lett. 11:125–129.
44. Sheehan, M. M., E. Stanley, G. F. Fitzgerald, and D. van Sinderen. 1999.Identification and characterization of a lysis module present in a large pro-portion of bacteriophages infecting Streptococcus thermophilus. Appl. Envi-ron. Microbiol. 65:569–577.
45. Shockman, G. D., and J. F. Barrett. 1983. Structure, function, and assemblyof cell walls of gram-positive bacteria. Annu. Rev. Microbiol. 37:501–527.
46. Shockman, G. D., M. J. Conover, J. J. Kolb, P. M. Philipps, L. S. Riley, andG. Toennies. 1961. Lysis of Streptococcus faecalis. J. Bacteriol. 81:36–43.
47. Shockman, G. D., and J.-V. Holtje. 1994. Microbial peptidoglycan (murein)hydrolases. New comprehensive biochemistry, p. 131–166. In J.-M. Ghuysenand R. Hakenbeck (ed.), Bacterial cell wall. Elsevier Science, London,United Kingdom.
48. Stanley, E., G. Fitzgerald, C. Le Marrec, B. Fayard, and D. van Sinderen.1997. Sequence analysis and characterization of f01205, a temperate bacte-riophage infecting Streptococcus thermophilus CNRZ1205. Microbiology 143:3417–3429.
49. Terzaghi, B. E., and W. E. Sandine. 1975. Improved medium for lacticstreptococci and their bacteriophages. Appl. Microbiol. 29:807–813.
50. Thomas, T. D., and V. L. Crow. 1983. Lactose and sucrose utilization byStreptococcus thermophilus. FEMS Microbiol. Lett. 17:13–17.
51. Towbin, H., and J. Gordon. 1984. Immunoblotting and dot immunobind-ing—current status and outlook. J. Immunol. Methods 72:313–340.
52. Valence, F., and S. Lortal. 1995. Zymogram and preliminary characterizationof Lactobacillus helveticus autolysins. Appl. Environ. Microbiol. 61:3391–3399.
53. Valence, F., R. Richoux, A. Thierry, A. Palva, and S. Lortal. 1998. Autolysisof Lactobacillus helveticus and Propionobacterium freudenreichii in Swisscheeses: first evidence by using species-specific lysis markers. J. Dairy Res.65:609–620.
54. Vegarud, G., H. B. Castberg, and T. Langsrud. 1983. Autolysis of group Nstreptococci. Effect of media composition modifications and temperature. J.Dairy Sci. 66:2294–2303.
55. Wilkinson, M. G., T. P. Guinee, D. M. O’Callaghan, and P. F. Fox. 1994.Autolysis and proteolysis in different strains of starter bacteria during Ched-dar cheese ripening. J. Dairy Res. 61:249–262.
56. Young, R. 1992. Bacteriophage lysis: mechanism and regulation. Microbiol.Rev. 56:430–481.
57. Young, R., and U. Blasi. 1995. Holins: form and function in bacteriophagelysis. FEMS Microbiol. Rev. 17:191–205.
58. Zourari, A. 1991. PhD thesis. INA-PG, Paris, France.
VOL. 66, 2000 AUTOLYTIC S. THERMOPHILUS STRAINS 565
Related Documents











