Cell Tiss. Res. 191, 27-37 (1978) Cell and Tissue Research © by Springer-Verlag 1978 The Spermatozoon of Oikopleura dioica Fol (Larvacea, Tunicata) Per R. Flood* and Bj6rn A. Afzelius** * Institute of Anatomy, University of Bergen, Bergen, Norway and ** Wenner-Gren Institute, University of Stockholm, Stockholm, Sweden Summary. The spermatozoon of Oikopleuradioica is about 30 Ixm long, with a spherical head, about 1 Ixm wide, a 3~tm long and 1 Ixm wide midpiece, and a 25lam long tail with a tapered end piece. The head contains a nucleus with the chromatin volume limited to about 0.1 ~tm3. A small acrosome is found in an anterior inpocketing, and a flagellar basal body in a posterior inpocketing ofthe nucleus. The midpiece contains a single mitochondrion with the flagellar axoneme embedded in a groove along its medial surface. The flagellar axoneme has the typical 9+2 substructure, and the basal body the typical 9+0 substructure. A second centriole and special anchoring fibres are absent. Key words: Spermatozoon - Oikopleura dioica - Electron microscopy. Introduction During an extensive comparative study of animal spermatozoa Gustaf Retzius (1905) found that the spermatozoon of Oikopleura dioica is smaller than any other sperm type he had studied. In other respects it was claimed to have a rather conventional appearance. With its small, round head plus midpiece and its thin tail, it was regarded as a "primitive spermatozoon". According to Retzius it resembled the teleost spermatozoon. The diminutive size makes the Oikopleura spermatozoon less suitable for light microscopical investigations, but has prompted us to examine it with the electron microscope. It has also been considered of interest to compare its fine structure with that of spermatozoa from other deuterostome metazoa. Send offprintrequests to." Dr. Bj6rn Afzelius, Wenner-Grens Institut, Norrtullsgatan 16, S-113 45 Stockholm, Sverige, Sveden 0302-766X/78/0191/0027/$02.20

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cell Tiss. Res. 191, 27-37(1978) Cell and TissueResearch© by Springer-Verlag 1978
The Spermatozoon of Oikopleura dioica Fol(Larvacea, Tunicata)
Per R. Flood* and Bj6rn A. Afzelius**
* Institute of Anatomy, University of Bergen, Bergen, Norway and** Wenner-Gren Institute, University of Stockholm, Stockholm, Sweden
Summary. The spermatozoon of Oikopleura dioica is about 30 Ixm long, with aspherical head, about 1 Ixm wide, a 3~tm long and 1 Ixm wide midpiece, and a25 lam long tail with a tapered end piece. The head contains a nucleus with thechromatin volume limited to about 0.1 ~tm3. A small acrosome is found in ananterior inpocketing, and a flagellar basal body in a posterior inpocketing o f thenucleus. The midpiece contains a single mitochondrion with the flagellaraxoneme embedded in a groove along its medial surface. The flagellar axonemehas the typical 9+2 substructure, and the basal body the typical 9+0substructure. A second centriole and special anchoring fibres are absent.
Key words: Spermatozoon - Oikopleura dioica - Electron microscopy.
Introduction
During an extensive comparative study of animal spermatozoa Gustaf Retzius(1905) found that the spermatozoon of Oikopleura dioica is smaller than any othersperm type he had studied. In other respects it was claimed to have a ratherconventional appearance. With its small, round head plus midpiece and its thin tail,it was regarded as a "primitive spermatozoon". According to Retzius it resembledthe teleost spermatozoon.
The diminutive size makes the Oikopleura spermatozoon less suitable for lightmicroscopical investigations, but has prompted us to examine it with the electronmicroscope. It has also been considered of interest to compare its fine structure withthat of spermatozoa from other deuterostome metazoa.
Send offprint requests to." Dr. Bj6rnAfzelius, Wenner-Grens Institut, Norrtullsgatan 16, S-113 45Stockholm, Sverige, Sveden
0302-766X/78/0191/0027/$02.20

28 P.R. Flood and B.A. Afzelius
Materials and Methods
Oikopleura dioica i (Fol, 1872) has alength of afew millimeters and inhabits the superficial layers ofmostmarine waters. During certain seasons of the year it may be present in great numbers in ordinaryplankton samples. However, using conventional plankton nets wehavebeen unable to obtain anythingbut animals withvery immature or emptied gonads. It was not until weisolated the animals, withintactjelly houses (cf. Fenaux, 1976) directly out of undisturbed sea water insmall beakers that wewere able tofind males with mature gonads. Such males could be detected by the naked eye because of an opaquewhite to yellowish colour of the gonads.
Immediately upon addition of fixative the animals escaped from their houses and emptied theirgonads. The semen was left undisturbed as coagulated threads in the fixative for some hours andprocessed for microscopyas tissue blocks. Somesemen was left inthe fixative for months before furtherprocessing. The fixative contained 1% formaldehyde freshly prepared from paraformaldehyde, 2.5glutaraldehyde, 30mM HEPES buffer pH 7.4, and60 %sea water.After two rinsesinbuffered seawater,the blocks werepostfixed in 1% osmium tetroxide in 40mM HEPES buffer and 75 % sea water. Surplusosmium was removedby a bath of distilled waterand dehydration followed inincreasing concentrationsof ethanol. Epon was used for embedding. Ultrathin sections were stained in uranyl acetate and leadcitrate, and examined in a Philips EM 300 or a JEOL 100c transmission electron microscope.
For lightmicroscopya fewsamples, still in the aldehyde fixative, were mountedandexaminedbyoilimmersion phase contrast in a Zeiss photomicroscope.
For scanning electron microscopy the material was fixed and rinsed as for transmission microscopy,spread on aluminium foils, air dried, and coated by argon sputtered gold-palladium. Somespermatozoawere also examined on the feeding filters of critical point dried jelly houses (Flood, 1978).
Results
Light Microscopic Observations
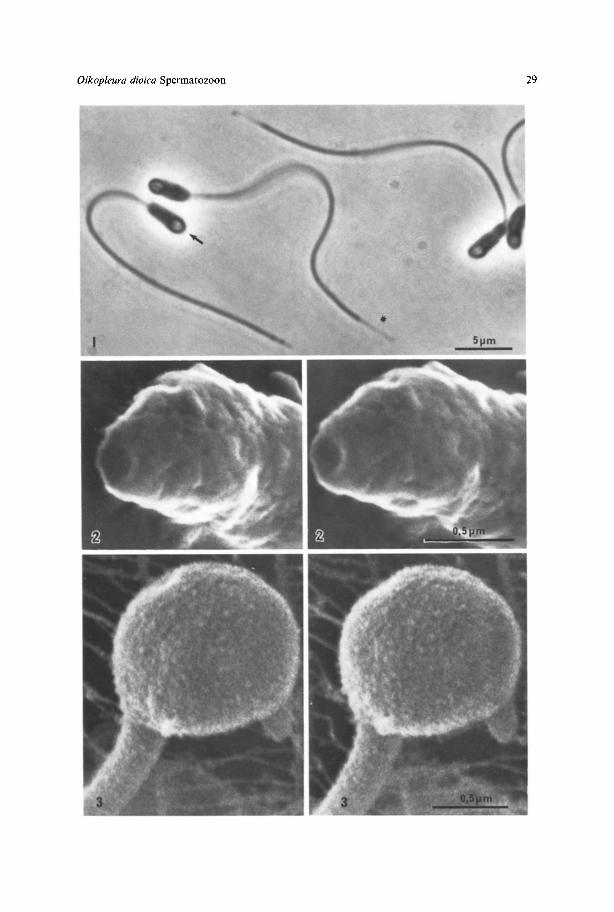
Spermatozoa f r o m di f fe ren t animals are in d i f fe ren t stages o f m a t u r a t i o n : I n some
animals t h e cells conta in head and midpiece in a roundish o r k idney-shapedst ructure less t h a n 2 g m long and d o n o t have t h e t a p e r e d end piece . I n o t h e r
animals t h e cells have a roundish h e a d , a b o u t 1 g m w i d e , with a f a in t axial str ing,but n o visible acrosome. Fur ther , t h e r e is a midpiece, 2 t o 3 g m long and slightly
narrower t h a n t h e h e a d . T h e tail is a b o u t 30 g m long and ends in a 2 - 3 g m longt a p e r e d end piece (Fig. 1). T h e e longa ted cells probably represen t t h e m a t u r e
1 We thank Mr. Terje Flood and Mr. Gunnar A. Flood for collecting the animals with intact jelly-houses, Mrs. Gro Bernstrom Tveit, Mrs. Kari Weltzin, Mr. Egil Erichsen, Mr. Harald Knutsen,Mr. Rune Haakonsen, Mr. Ragnar Jensen, and Mr. Jacob Roli for technical assistance, and theNorwegian Research Council for Science and the Humanities for financial support (grant no. C 21.30-8)
Fig. 1. Phase contrast light micrograph of aldehyde fixed spermatozoa. Note the faint axialstring in thehead (arrow), the elongated midpiece and tapered tail tip (asterisk) of these presumably maturespermatozoa, x 3000
Fig. 2. Stereopair of scanning electronmicrographs of the head region of an air dried spermatozoon.Note the circular depression of the plasma membrane at the anterior end. x 47,000
Fig. 3. Stereopair of scanning electron micrographs of the globular head-midpiece of a critical pointdried spermatid trapped in the feedingfilterof the animal's jelly house. Note the irregular dots coveringmost of the plasma membrane on the head and tail. x 40,000
Oikopleura dioica Spermatozoon 29

30 P.R. Flood and B.A.Afzelius
spermatozoa whereas those with a more roundish and asymmetrical head plusmidpiece are believed are believed to be spermatids.
Scanning Electron Microscopic Observations
Elongated spermatozoa have been examined only after air drying and reveal anirregularly bulged surface on most of the head and midpiece. However, a t theiranterior end a 0.21xm wide circular cavity is constantly found (Fig. 2). Theimmature spermatids which have also been examined after critical point drying,reveal a relatively smooth surface covered by tiny (10-50 gm) irregular dots (Fig. 3).
Transmission Electron Microscopic Observations
Both spermatozoa and late spermatids have been examined. Their general form is ingood agreement with the light and scanning electron microscopic observationsreported above (Figs. 4, 5).
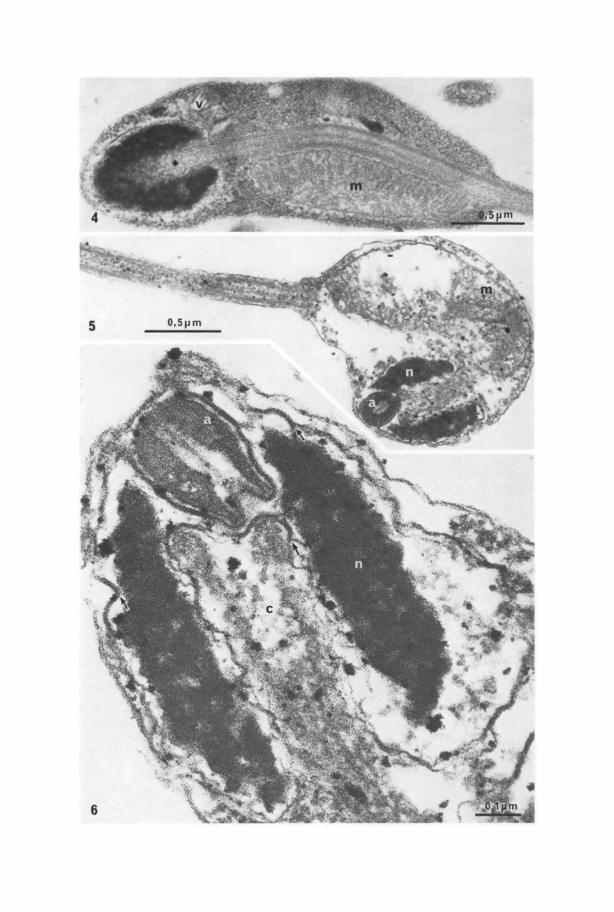
The cells contain an acrosome closely associated with a slightly depressed areao f the plasma membrane, a nucleus and the basal body of the flagellum. The nucleusforms a ring around the acrosome and basal body. However, the nuclearmembranes are continuous across the gap between the acrosome and basal body sothat the nuclear envelope constitutes a sphere with two deep inpocketings whereasthe chromatin is an annulus (Fig. 6). The midpiece is made up of a variable amountof granular cytoplasm and a single mitochondrion partly embracing the proximalflagellar shaft (Figs. 4, 11). The basal body and flagellum are fairly straight inspermatozoa (Fig. 4), whereas in spermatids the proximal flagellar shaft is sharplybent (Fig. 5). In the spermatozoa the midpiece, accordingly, constitutes a 2-3 lxmlong segment in continuation of the head, whereas in spermatids, the midpiececonstitutes an irregular cytoplasmic mass which forms a kidney-shaped or globularstructure together with the head.
Acrosome. The acrosome region contains an acrosomal vesicle and a diminutivesubacrosomal space. The acrosomal vesicle is oval, 0.3 Ixm long and up to 0.2 lamwide. Its long axis coincides roughly, but not exactly, with the cell axis. The vesicle
Fig.4. Transmission electron micrograph of an elongated spermatozoon longitudinally sectionedthrough the head and midpiece. The flagellar axoneme courses from a nuclear inpocketing (asterisk)along onesideofthe mitochondrion (m)towardsthe slender tail.Vesicles(v)and ribosome-like granulesare present in the cytoplasm, x 39,000
Fig.5. Transmission electron micrograph of a roundish spermatid head and midpiece. Note theacrosome (a), the nucleus (n), andthe kink of the flagellar shaft (asterisk) near the mitochondrion (m).x 41,000
Fig.6. Transmission electron micrograph asin Figure 5. Note theacrosomal (a) substructure and thepentalaminar nuclear membrane (arrows) bordering the entire anterior end ofthe nucleus (n)and thedeep portion of the centriolar (c) inpocketing, x 120,000

32 P.R. Flood and B.A. Afzelius
has a rather indistinct limiting membrane and a content which isfairly uniform anddense, apart from a central fibrous strand surrounded by a less dense zone(Figs. 6, 7). The plasma membrane is closely apposed to the anterior end of theacrosome and slightly invaginated relative to the surrounding membrane (Fig. 6).Laterally and posteriorly the acrosome borders on the nuclear envelope.
There is a small posterior notch in the acrosomal vesicle, and although thenuclear envelope shows a corresponding evagination that partly fills the notch,there is a small gap between the two membranes. This gap has a maximal width of0.05 lxm and may be regarded as a vestigial subacrosomal space (Fig. 6).
Nucleus. The nucleus is 0.8 ~tm long and 0.6 ~tm wide. It has two inpocketings: ananterior one where the acrosome is inserted and a posterior one housing thecentriole and a portion o f the axoneme. The two inpocketings traverse the entirenucleus, yet without piercing the nuclear envelope. The two inpocketings areusually aligned. When they are not, the posterior end of the acrosome and the frontend of the centriole overlap (Fig. 6).
The volume of the chromatin can be estimated to about 0.13 lain3 which is byone order of magnitude less than that o f other primitive spermatozoa, such as thatof sea urchins (Afzelius, 1955). The chromatin is the most electron dense componento f the cell. No substructure can be resolved in it.
The nuclear envelope has two membranous components. Along the posteriorand lateral surfaces o f the nucleus there is a well defined perinuclear cisterna. Alongthe anterior periphery and along the entire anterior nuclear inpocketing and in thebottom of the posterior one, the cisterna is collapsed, and the two membranouscomponents form a five-layered compound membrane (dense-lucid-dense-lucid-dense).
Midpiece. As previously mentioned, the form of the midpiece varies in sperm cellsfrom different gonads. Cells with the head and midpiece organelles gathered in aglobular or kidney-shaped structure (Fig. 5) are considered to be less mature thanthose with the midpiece organelles gathered in an elongated swelling o f the tailbehind the head organelles (Fig. 4).
The midpiece contains one mitochondrion about 2 ~tm long and 0.5 to 1 pmwide. The flagellar axoneme courses through a furrow on one side of themitochondrion and seems always to have an axial orientation with themitochondrion on one side of a plane drawn through the two central singletfilaments (Fig. 11). Most o f the mitochondrial cristae are tubular or slightlylamellar and are oriented perpendicularly to the axis of the axoneme (Fig. 4). Theremainder o f the midpiece cytoplasm contains a few vesicles and tightly packedgranules, probably both ribosomes and glycogen (Figs. 4, 10). In the spermatid, theflagellar shaft usually exhibits a sharp kink or bend, corresponding to the anteriorpart of the mitochondrion. This is probably an artifact due to imperfect fixation(Fig. 5).
Tail. The specimens that remained in the aldehyde fixative for 3 months beforeembedding proved to be very useful for the examination of the flagellar
Oikopleura dioica Spermatozoon 33
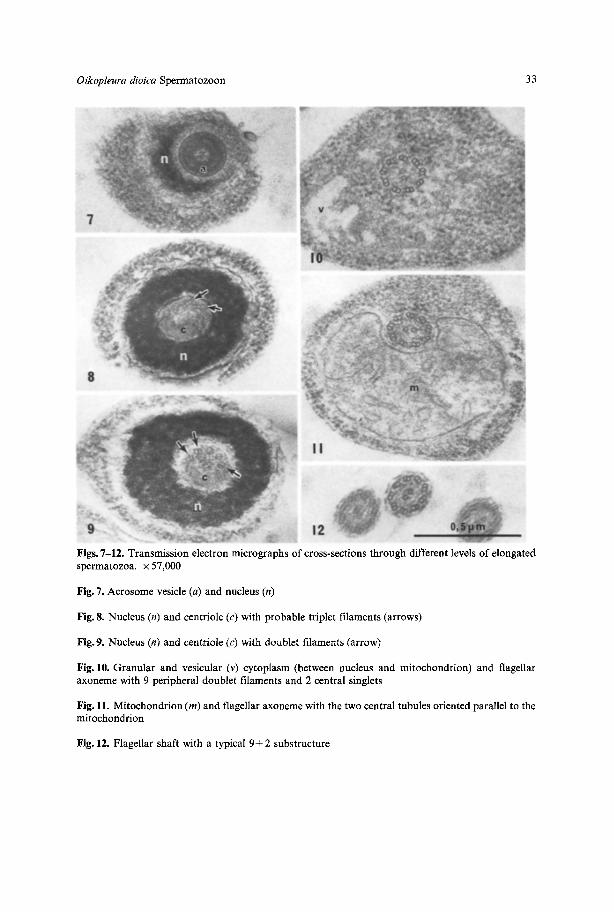
Figs. 7-12. Transmission electronmicrographs of cross-sections through different levels of elongatedspermatozoa, x 57,000
Fig. 7. Acrosome vesicle(a) and nucleus (n)
Fig. 8. Nucleus (n) and centriole (c) with probable triplet filaments (arrows)
Fig. 9. Nucleus (n) and centriole (c) with doublet filaments (arrow)
Fig. 10. Granular and vesicular (v) cytoplasm (between nucleus and mitochondrion) and flagellaraxoneme with 9 peripheral doublet filaments and 2 central singlets
Fig. 11. Mitochondrion (m) and flagellar axonemewith the two central tubules orientedparallel to themitochondrion
Fig. 12. Flagellar shaft with a typical 9+ 2 substructure

34 P.R. Flood and B.A.Afzelius
Fig. 13. Diagram ofan elongated O. dioica spermatozoon
substructure (Figs. 8-12), although the plasma membrane and some cytoplasmicmembranes appeared fragmented (Figs. 10-12). The tail has an axoneme with thecommon 9 + 2 microtubule pattern. Its basal body is the centriole, which occupiesthe anterior half o f the intranuclear canal. It differs from the remainder o f theflagellum by having a 9+ 0 structure. A short segment just behind the acrosomeprobably has triplets in each of the nine units (Fig. 8). The main part of the centriole,however, contains nine doublets (Fig. 9). There is no proximal centriole, nor is thereany anchoring fibre apparatus, which suspends the basal body in the anterior partof the midpiece, as iscommon in primitive spermatozoa (Afzelius et al., 1973, 1978).
The "free" part of the flagellum is about 25 ~tmlong. An end piece region, 2 to3 pm long, is also found on most cells (Fig. 1). The main features of the O. dioicaspermatozoon are given in a schematic drawing (Fig. 13).
Discuss ion
Our classification of some cells as spermatids and others as spermatozoa may bequestioned as we have not followed the differentiation of the spermatids ofOikopleura dioica in a temporal sense and not checked the fertility of the sperm cellswe have examined. Our definitions are mainly based on a superficial comparison tothe situation in other animals where an elongated slender cell usually represents amore mature stage than a roundish cell. Whether our elongated "spermatozoa"really are mature cells remains to be proved. The presence of relatively abundantcytoplasm with ribosome-like granules may indicate that some differentiation isstill in progress. In this connection it should be mentioned that the total generationtime of O. dioicahas recently been established to be 9.5 days at 13° C (Paffenh6fer,1973), 3 to 5 days at 22° C, and 10 to 12 days at 14° C (Fenaux, 1976) and that thelate gonadal maturation is particularly rapid (Fenaux, 1976).
In spite of our uncertainty as to the exact stage of maturation of the cellsexamined this study provides important information on the spermatozoon ofO. dioica, which is an exception among the appendicularia as it is the only specieswith separate sexes (Drach, 1966). As long as the spermatozoa of other appendicu-laria have not been studied, it is impossible to know how representative the O. dioicaspermatozoon is for the appendicularian group.
When compared to other animal groups, the O. dioica spermatozoon posessestwo striking features. It is very small and its structure deviates somewhat from thato f the typical primitive spermatozoon. Spermatozoa from most animalspecies have

Oikopleura dioica Spermatozoon 35
undergone a marked reduction in size during spermiogenesis and are small cells. Itis believed that one reason for the diminution is the necessity to store a greatnumber o fcells (in a limited space) to ensure that a sufficient proportion o feggs arefertilized. When fertilization is internal and few eggs are fertilized, the spermatozoatend to be larger than in species with external fertilization, although exceptions tothis rule exist. Termite spermatozoa, for instance, may be very reduced in size (andcomplexity), probably because of the need of fertilizing a great number of eggs inone mating (Baccetti et al., 1973).
In O. dioica spermatozoa we have estimated the chromatin volume to be in theorder o f 0.13~tm3. From nuclear dimensions given by, or derived from theillustrations of, Schabtach and Ursprung (1965) we have estimated the chromatinvolume of Ascidia nigra spermatozoa to be about 0.25 ~tm3. Comparable valuesderived from the work of Franz6n (1976) on Corella parallelogramma and ofWollacott (1977) on Ciona intestinalis (both belonging to the Ascidiacea) are0.3 lam3 and 0.6 Izma. All these values are lower than those found in spermatozoa o fother species characterized by external fertilization.
Whereas the gametes o f most multicellular eukaryotes have a DNA contentseveral hundred fold greater than that present in bacteria, Ciona intestinalis (andDrosophila) are an exception in having only about 35 times as much DNA in itsspermatozoa as is present in the E. eoli haploid genome (for reference see Atkins andOhno, 1967 and Laird, 1971). Using the nuclear volume values reported above andassuming the chromatin to be condensed to the same degree in O. dioica as inC. intestinalis spermatozoa, the DNA content o f an O. dioica spermatozoon wouldbe only 9 times that o f E. coli. This, of course, makes one believe that thespermatozoon contains only a fraction of the haploid genome or that the chromatinis unusually densely packed. It should be added that tunicate cells in general aresmall (Szarski, 1968) and that the nuclei o fmost internal epithelial cells o fO. dioicahave a volume of about 8 ~tm3 (Flood, unpublished observation). Other somaticcells of adult O. dioica especially in the oikoplasts (Fenaux, 1971) and tail musclecells (Seeliger, 1900), have large, probably polyploid nuclei.
It has been shown by Franz6n (1956) that spermatozoa involved in externalfertilization usually have a typical construction which is called the primitive type ofspermatozoon. Such primitive spermatozoa have been characterized at the ultra-structural level by a.o. Afzelius et al. (1973, 1978). Typically, the nucleus is roundedand surmounted by a twopartite acrosome; the midpiece contains four mitochon-dria and two centrioles, the distal one being suspended by an anchoring fibreapparatus.
Within the deuterostome line of animals, the lowest group, namely the crinoidechinoderms, have spermatozoa which come closest to this description, the onlydeviation from the above characterization being the presence o f only one ring-shaped mitochondrion rather than four separate round ones (Afzelius, 1977).
The O. dioiea spermatozoon has a single U-shaped mitochondrion, but deviatesin further respects: The subacrosomal space is almost completely reduced. There isonly one centriole and there is no anchoring apparatus for it. This structure may becompared to that of the ascidian type of spermatozoon, as described by Schabtachand Ursprung (1965), Franz6n (1976) and Woollacott (1977). The ascidians areclosely related to the appendicularians and have a sperm structure which deviates

36 P.R. Flood and B.A. Afzelius
even more strongly from the primitive sperm type. The entire acrosome is reducedin size, or absent . The only mitochondrion lies laterally to the elongated nucleus,and there is no anchoring fibre apparatus . The spermatozoon of the third group oftunicates, the Thaliacea, has been examined by light microscopy (Franz6n, 1958)but not yet by electron microscopy (Berrill, 1975). Although the ascidian andappendicularian sperm types have certain features in common, neither can bederived from the other, nor does the appendicularian (O. dioica) spermatozoonshow any g r e a t resemblance to spermatozoa from other primitive chordate groupsthat have been examined with the electron microscope: the cephalochordateBranch ios toma (Baccetti et al., 1973; Wickstead, 1975), and the cyclostomesMyxinidae (Jespersen, 1975).
The n o r m a l mode of sperm release in Oikopleurae is poorly understood. Fol(1872, PL I, Figs. 1 and 2), D r a c h (1966, Fig. 355) and Fenaux (1973, PL 4, Fig. 1)describe a sperm duct leading from the testis to the exterior of various speciesincluding O. dioica. However, in the case of the latter species, the size of the testissuggests a very immature stage of gonadal development, a stage in which we havebeen unable to find any spermatid-like cells by transmission electron microscopy.Gradual sperm release over prolonged periods seems unlikely for another reasontoo: most Appendicularia spend only a few minutes of their life outside theirsecreted jelly houses (Gai t , 1972; Alldredge, 1976a) which are very efficient foodt r a p s that would also withhold spermatozoa (cf. Fig. 3 and Flood, 1978).
In our material the entire sperm mass was released through a rupture in theectoderm and gonadal wall when the animals forced their way out of their jellyhouses upon chemical irritation. Similar jett ison movements normally take placeevery 4 or 5 h when the feeding filters become clogged and the animals buildthemselves new houses (Gai t , 1972; Alldredge, 1976b). Massive sperm release byrupture in the jettisoned state (probably followed by the death of the animal) wouldbe a very efficient way of spreading the gametes.
References
Afzelius, B.A.: Thefine structure ofthe seaurchin spermatozoa as revealedby the electron microscope.Z. Zellforsch. 42, 134-148 (1955)
Afzelius, B.A.: Spermatozoa and spermatids of the crinoid Antedonpetasus, with a note on primitivespermatozoa from deuterostome animals. J. Ultrastruct. Res. 59, 272-281 (1977)
Afzelius, B.A., Ferraguti, M.: The spermatozoon ofPriapulus caudatus Lamarck.J. submicrosc. Cytol.10, 71 79 (1978)
Afzelius, B.A., Franz6n,/~.: The spermatozoon ofthe jellyfish Nausithoe. J. Ultrastruct. Res. 37,186-199(1971)
Alldredge,A.: Field behavior and adaptive strategies of appendicularians (Chordata, Tunicata). Mar.Biol. 38, 29-39 (1976a)
Alldredge, A.: Appendicularians. Sci. Amer. 235, 95-102 (1976b)Atkins, N.B.,Ohno, S.: DNAvalues offour primitive chordates. Chromosoma(Berl.) 23, 10 13(1967)Baccetti, B.,Burrini,A.G., Dallai, R.: The spermatozoon of Branchiostoma lanceolatum L. J. Morph.
136, 211-226 (1972)Baccetti, B., Dallai, R., Rosati, F., Giusti, F., Bernini, F., Selmi, G.: The spermatozoon of Isoptera,
Embioptera and Dermaptera. J. Microsc. (Paris) 21, 159-172 (1974)Drach,P.: Les appendiculaires. D6veloppement embryonaire, organes g6nitaux. In: Trait~ de zoologie,
anatomie, syst6matique, biologie (P.-P. Grass6, ed.),Vol.11, pp. 891-892. Paris: Masson &Cie.1966

Oikopleura dioicaSpermatozoon 37
Fenaux, R.: Cycle vital d'un appendiculaire Oikopleura dioica Fol, 1872. Description et chronologie.Ann. Inst. Oceanogr. Paris 52, 89-101 (1976)
Flood, P.R.: Filter characteristics of appendicularian food catching nets. Experientia (in press, 1978)Fol, M.H. : Etudes surles Appendiculaires du detroit deMessine.M6m.Soc.Phys. Hist. Nat. Gen6ve21,
446-499 (1872)Franzbn, A.: On spermiogenesis, morphology of the spermatozoon, and biology of fertilization among
invertebrates. Zool. Bidrag Uppsala 31, 355482 (1956)Franzbn, A.: On sperm morphology and acrosome filament formation in some annelida, echiuroidea,
and tunicata. Zool. Bidrag Uppsala 33, 1-28 (1958)Franzbn, A.: The fine structure of spermatid differentiation in a tunicate, Corella parallelogramma
(Mfiller). Zoon 4, 115-120 (1976)Gait, C.P.: Development of Oikopleuradioica (Urochordata, Larvacea). Ontogeny of behavior and of
organ systems related to constructionand use ofthe house.PH.D.ThesisUniversity ofWashington85 pp. (1972)
Jesperson, A.: Fine structure of spermiogenesis in EasternPacificspecies of a hagfish(Myxinidae).Actazool. (Stockh.) 56, 189-198 (1975)
Laird, C.D.: Chromatin structure: Relationship between DNA content and nucleotide sequencediversity. Chromosoma (Berl.) 32,378406 (1971)
Paffenh6fer, G.-A.: The cultivation of an appendicularian through numerous generations. Mar. Biol.22, 183-185 (1973)
Retzius, G.: Zur Kenntnis der Spermien der Evertebraten II. Biol. Untersuch. (Stockh.) 12, 79-102(1905)
Schabtach, E., Ursprung, H. : The fine structure of the sperm ofa tunicate, Ascidia nigra. J. exp. Zool.159, 357-366 (1965)
Seeliger, O.: Einige Bemerkungenfiber den Bau des Ruderschwanzes derAppendicularien. Z. wiss. Zool.67, 63-70 (1900)
Szarski, H.: Evolution of cell size in lower vertebrate phylogeny. In: Problems of lower vertebratephylogeny (T. Orvig, ed.), Nobel Symposium 4,pp. 445453. Stockholm: Almqvist &Wiksell 1968
Wickstead, J.H.: Chordata: Acrania (Cephalochordata). In: Reproduction in marine invertebrates(A. Giese and J.S. Pearse, eds.), Vol. 2, pp. 283-319. New York-London: Academic Press 1975
Woollacott, R.M.: Spermatozoa of Ciona intestinalis and analysis of ascidian fertilization. J. Morph.152, 77-88 (1977)
Accepted January 30, 1978
Related Documents











