PRIMATES, 19(4): 715-726, October 1978 715 Social Behavior of a Marmoset (Saguinusfuscicollis) Group I" Parental Care and Infant Development JERRY L. VOGT Saint John's University HAROLD CARLSON, and EMIL MENZEL State University of New York ABSTRACT. Development was observed as three infant sets were born into a semi-natural- istic family group. Data on infant carrying through 12 weeks of age revealed that, at almost every week, the father carried more than the mother, and the males carried more than the females. Spatial data for the first two infant sets showed nearest neighbor distances support- ing a two-stage proximity process during development. Behavioral data for 30 weeks were rather variable between the first two infant sets; this suggests that infant behavioral patterns may be more dependent upon the composition of the group than is spatial development or infant carrying. Incidents of "play fathering" and the importance of parental care and infant development in general were discussed. INTRODUCTION Marmosets, which generally include all genera of the New World monkey family Callithricidae (HERSHKOVITZ, 1968), have become for many reasons very popular in primate research (see review by EPPLE, 1975b). These small, arboreal monkeys are interesting in several respects: They are among the smallest in size of the true pri- mates (NAPIER & NAPIER, 1967); they customarily have twin offspring (EPPLE, 1970b; HAMPTON & HAMPTON, 1965); and they seem to live in family group social units--an adult pair and their offspring of several years--in which the parents share infant caretaking (EPPLE, 1972, 1975b; MOYNIHAN, 1970). EPPLE has reported on several aspects of marmoset behavior, including develop- ment and breeding (EPPLE, 1970a), vocal communication patterns (EPPLE, 1968), and the pheromonal communication system (EPPLE, 1970b, 1972). She has recently re- ported quantitative data on infant caretaking for Saguinusfuscieollis ssp. and found that in seven captive groups, dominant males carried most frequently, with mothers and subadults, respectively, carrying next most frequently (EPPLE, 1975a). The present study was also conducted on infant development with this same species (Saguinus fuseieollis) but used a longitudinal rather than a cross-sectional approach. One group of animals, initially numbering four, was observed in a relatively large, semi-naturalistic environment over a 32-month period, during which time data were taken on three sets of infants born into the group. METHOD APPARATUS The monkeys were housed in a 10 • 4 • 3.5 m glass greenhouse which is described in detail in VOGX (1978).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PRIMATES, 19(4): 715-726, October 1978 715
Social Behavior of a Marmoset (Saguinusfuscicollis) Group I" Parental Care and Infant Development
JERRY L. VOGT Saint John's University
HAROLD CARLSON, and EMIL MENZEL State University of New York
ABSTRACT. Development was observed as three infant sets were born into a semi-natural- istic family group. Data on infant carrying through 12 weeks of age revealed that, at almost every week, the father carried more than the mother, and the males carried more than the females. Spatial data for the first two infant sets showed nearest neighbor distances support- ing a two-stage proximity process during development. Behavioral data for 30 weeks were rather variable between the first two infant sets; this suggests that infant behavioral patterns may be more dependent upon the composition of the group than is spatial development or infant carrying. Incidents of "play fathering" and the importance of parental care and infant development in general were discussed.
INTRODUCTION
Marmosets, which generally include all genera of the New World monkey family Callithricidae (HERSHKOVITZ, 1968), have become for many reasons very popular in primate research (see review by EPPLE, 1975b). These small, arboreal monkeys are interesting in several respects: They are among the smallest in size of the true pri- mates (NAPIER & NAPIER, 1967); they customarily have twin offspring (EPPLE, 1970b; HAMPTON & HAMPTON, 1965); and they seem to live in family group social units--an adult pair and their offspring of several years--in which the parents share infant caretaking (EPPLE, 1972, 1975b; MOYNIHAN, 1970).
EPPLE has reported on several aspects of marmoset behavior, including develop- ment and breeding (EPPLE, 1970a), vocal communication patterns (EPPLE, 1968), and the pheromonal communication system (EPPLE, 1970b, 1972). She has recently re- ported quantitative data on infant caretaking for Saguinusfuscieollis ssp. and found that in seven captive groups, dominant males carried most frequently, with mothers and subadults, respectively, carrying next most frequently (EPPLE, 1975a).
The present study was also conducted on infant development with this same species (Saguinus fuseieollis) but used a longitudinal rather than a cross-sectional approach. One group of animals, initially numbering four, was observed in a relatively large, semi-naturalistic environment over a 32-month period, during which time data were taken on three sets of infants born into the group.
METHOD
APPARATUS
The monkeys were housed in a 10 • 4 • 3.5 m glass greenhouse which is described in detail in VOGX (1978).

716 J. L. VOGT, H. CARLSON, & E. MENZEL
SUBJECTS
The original group consisted of four saddle-back tamarins (Saguinus fuscicollis): an adult breeding pair with a 2-month old lab-born female and a 14-month old lab- born male unrelated to and unacquainted with the other three animals. Because of severe fighting the subadult male was removed from the group on September 14, 1973. During the study period four sets of twins were born to the adult female: on March 20, 1973, a male (Spring) and a stillborn; on August 30, 1973, two males (Marc and Rupert); on November 20, 1974, a male (Norman) and a female (Abby); and on August 24, 1975, two females (Daphne and Chloe) (See VOGT, 1978 for further details).
OBSERVATION PROCEDURE
The method of data collection for the first two sets of infants was as follows: the experimenter entered the observation chamber, noted the time of day, temperature, relative humidity, outdoor weather conditions, and then began plotting the location and describing the behavior of the monkeys at 60-second intervals. The click of an electrical timer in the observation booth signaled the instant at which the observer was to note each marmoset's position and behavior. Data maps were used to record position to the nearest 0.5 m unit in each of the three dimensions. During the after- noon 30-minute baseline sessions, averaging 4.5 per week, a cassette tape recorder was used for dictation of the behavioral categories, and this information was subse- quently transcribed to the location data maps. The behavioral categories used for this project are detailed in VOGT (1975, 1978). Those of interest in this study include cling to, groom, play wrestle, play chase, social play (play wrestle and play chase com- bined), sit and look, locomote, eat, drink, and play with object. Data on rate relia- bility were gathered prior to and during the study period, and values typically obtained were greater than +0.90.
Data on the third set of infants (Abby and Norman) were not gathered systematical- ly enough to include in the present analysis, and data collection for the last set of infants (Daphne and Chloe) differed slightly from the above procedure. Data were gathered only on infant carrying; no behavioral or spatial data were recorded. Obser- vation sessions were conducted five times per week, between 1100 and 1400, and were 45 minutes in length. During the first week data were recorded every 60 seconds, and during the following weeks every 30 seconds.
RESULTS AND DISCUSSION
Because of the difficulty in identifying the individual infants early in development, the data on carrying were combined for a set of twin infants. That is, at a particular time period an individual could be scored at, say, 50 ~o by carrying one of the two infants all (100 ~o) of the time, by carrying both of the two infants one-half (50 ~o) of the time, or by some appropriate combination thereof. This method precluded treating the twin infants as individuals but did produce data that were appropriately weighted for the two infants. It should be noted that for the first few weeks of de- velopment twin infants were typically carried by the same individual and as they grew older they were more likely to be on different carriers.
Marmoset Parental Care and Infant Development 717
lot
80'
540- �9 NON
o,o. s
2 4 6 8 10 12 A G E (in weeks)
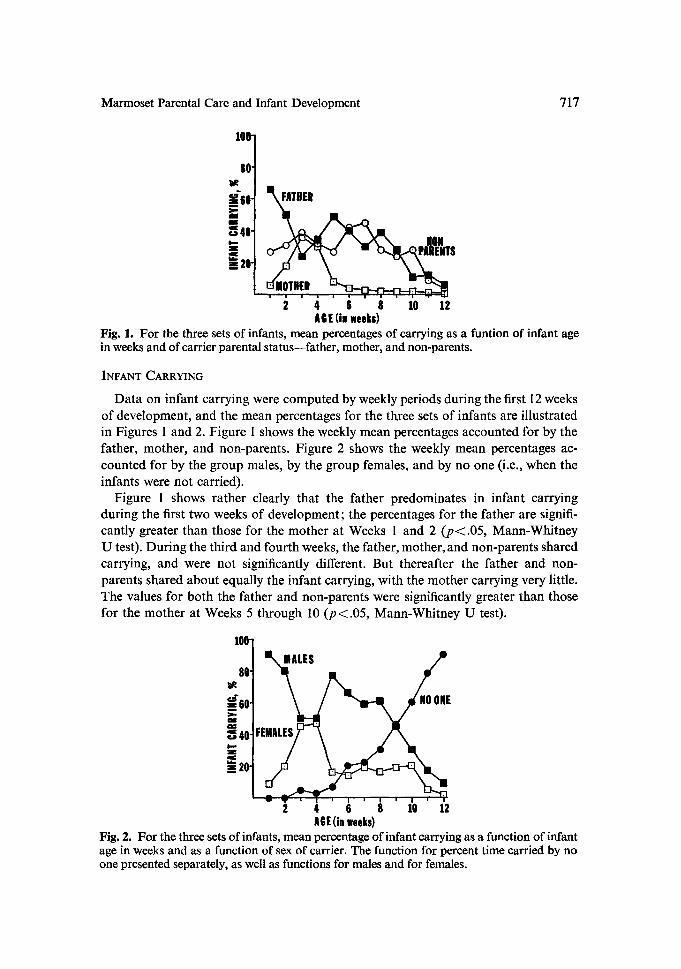
Fig. 1. For the three sets of infants, mean percentages of carrying as a funtion of infant age in weeks and of carrier parental status--father, mother, and non-parents.
Data on infant carrying were computed by weekly periods during the first 12 weeks of development, and the mean percentages for the three sets of infants are illustrated in Figures 1 and 2. Figure 1 shows the weekly mean percentages accounted for by the father, mother, and non-parents. Figure 2 shows the weekly mean percentages ac- counted for by the group males, by the group females, and by no one (i.e., when the infants were not carried).
Figure 1 shows rather clearly that the father predominates in infant carrying during the first two weeks of development; the percentages for the father are signifi- cantly greater than those for the mother at Weeks 1 and 2 ( p ( . 0 5 , Mann-Whitney U test). During the third and fourth weeks, the father, mother, and non-parents shared carrying, and were not significantly different. But thereafter the father and non- parents shared about equally the infant carrying, with the mother carrying very little. The values for both the father and non-parents were significantly greater than those for the mother at Weeks 5 through 10 (p~.05, Mann-Whitney U test).
m•ALES / 8 0
~$0' ONE
_~ 20 i i J i
2 4 6 8 10 12 AGI: (in weeks)
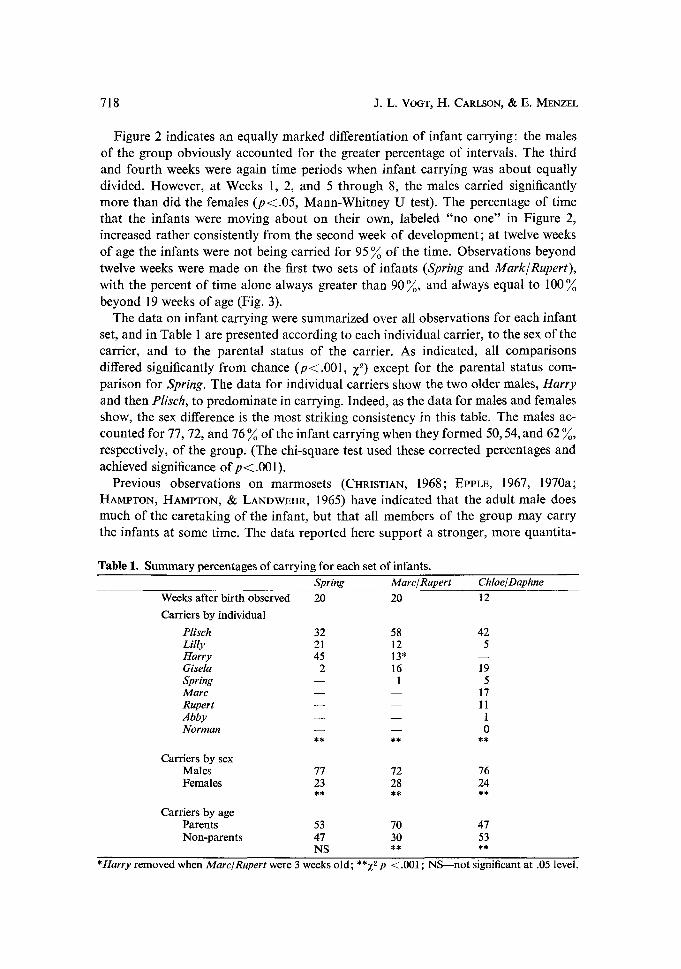
Fig. 2. For the three sets of infants, mean percentage of infant carrying as a function of infant age in weeks and as a function of sex of carrier. The function for percent time carried by no one presented separately, as well as functions for males and for females.
INFANT CARRYING
100
718 J. L. VOGT, H. CARLSON, & E. MENZEL
Figure 2 indicates an equally marked differentiation of infant carrying: the males of the group obviously accounted for the greater percentage of intervals. The third and fourth weeks were again time periods when infant carrying was about equally divided. However, at Weeks 1, 2, and 5 through 8, the males carried significantly more than did the females (p< .05 , Mann-Whitney U test). The percentage of time that the infants were moving about on their own, labeled "no one" in Figure 2, increased rather consistently f rom the second week of development; at twelve weeks of age the infants were not being carried for 95 % of the time. Observations beyond twelve weeks were made on the first two sets of infants ( S p r i n g and M a r k ~ R u p e r t ) ,
with the percent of time alone always greater than 90 %, and always equal to 100 % beyond 19 weeks of age (Fig. 3).
The data on infant carrying were summarized over all observations for each infant set, and in Table 1 are presented according to each individual carrier, to the sex of the carrier, and to the parental status of the carrier. As indicated, all comparisons differed significantly f rom chance (p<.001, Z 2) except for the parental status com- parison for S p r i n g . The data for individual carriers show the two older males, H a r r y
and then P l i s c h , to predominate in carrying. Indeed, as the data for males and females show, the sex difference is the most striking consistency in this table. The males ac- counted for 77, 72, and 76 % of the infant carrying when they formed 50, 54, and 62 %, respectively, of the group. (The chi-square test used these corrected percentages and achieved significance of p<.001) .
Previous observations on marmosets (CHRISTIAN, 1968; EPPLE, 1967, 1970a; HAMPTON, HAMPTON, & LANDWEHR, 1965) have indicated that the adult male does much of the caretaking of the infant, but that all members of the group may carry the infants at some time. The data reported here support a stronger, more quantita-
Table 1. Summary percentages of carrying for each set of infants. Spr ing Marc~Ruper t Chloe~Daphne
Weeks after birth observed 20 20 12 Carriers by individual
Pl i sch 32 58 42 Li l l y 21 12 5 H a r r y 45 13 * - - Gisela 2 16 19 Spr ing - - 1 5 M a r c - - - - 17 R u p e r t - - - - 11 A b b y - - - - 1 N o r m a n - - - - 0
Carriers by sex Males 77 72 76 Females 23 28 24
Carriers by age Parents 53 70 47 Non-parents 47 30 53
NS ** ** *Harry removed when Marc~Ruper t were 3 weeks old; **ZZp <.001 ; NS--not significant at .05 level.
Marmoset Parental Care and Infant Development 719
tive statement, for the males in this group seem to take the primary responsibility of carrying the infants. And the strong interest that the subadult male, Harry, showed in young infants is most striking. During Spring's development he accounted for the greatest amount (45 ~ ) of the carrying (Table 1), and in his last week in the group Harry accounted for 50 ~ of the infant carrying of Marc and Rupert. Both the high level of infant carrying by Harry and the mounting tension between Harry and Plisch were probably reflections of Harry's emerging adulthood. The paternal moti- vation of the adult and near-adult males in this group seems comparable in form and function to the great interest in infants (play-mothering) by females of all ages in many Old and New World primate species. It certainly had the result of allowing practice in caring for the infants and increasing parental investment (TRIVERS, 1972) by the male.
BEHAVIORAL AND SPATIAL DEVELOPMENT
In addition to parental care, data on spatial and behavioral development were also gathered. Except for percent time not carried (Fig. 3), these data were exclusively from the first two sets of infants, Spring and Marc/Rupert.
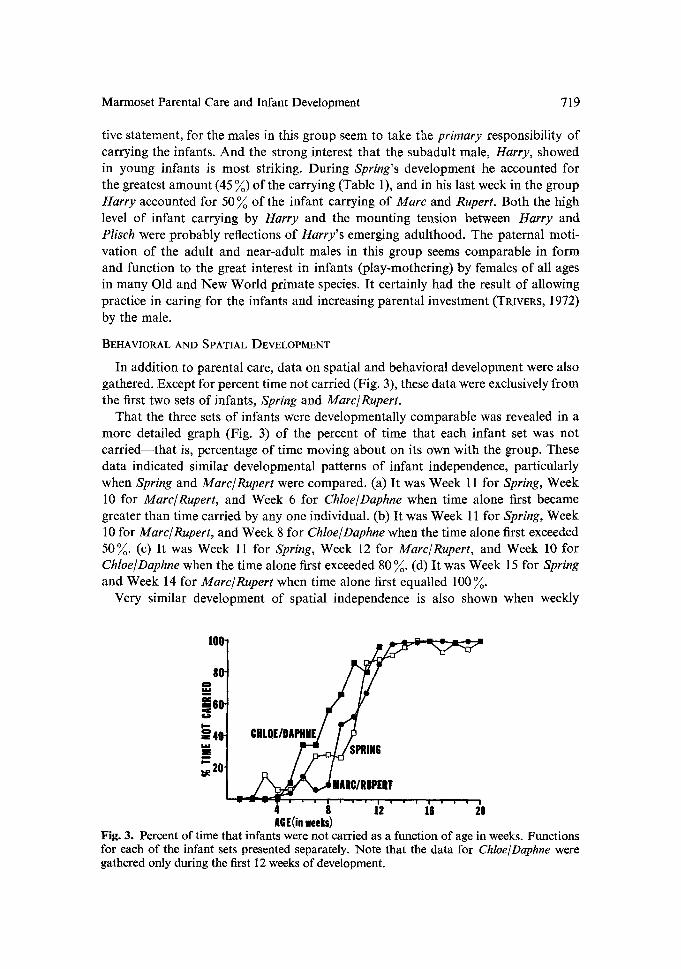
That the three sets of infants were developmentally comparable was revealed in a more detailed graph (Fig. 3) of the percent of time that each infant set was not car r ied~that is, percentage of time moving about on its own with the group. These data indicated similar developmental patterns of infant independence, particularly when Spring and Marc~Rupert were compared. (a) It was Week 11 for Spring, Week 10 for Marc/Rupert, and Week 6 for Chloe/Daphne when time alone first became greater than time carried by any one individual. (b) It was Week 11 for Spring, Week 10 for Marc/Rupert, and Week 8 for Chloe~Daphne when the time alone first exceeded 50~. (c) It was Week 11 for Spring, Week 12 for Marc~Rupert, and Week 10 for Chloe/Daphne when the time alone first exceeded 80 ~ . (d) It was Week 15 for Spring and Week 14 for Marc/Rupert when time alone first equalled 100~.
Very similar development of spatial independence is also shown when weekly
'~176 SO 1
is01
~201 MARC/RUPtRT r . / . 7 . ~ 'AR!/R.UPE R, . . . . . . . . 4 8 12 16 20 AGE(in weeks)
Fig. 3. Percent of time that infants were not carried as a function of age in weeks. Functions for each of the infant sets presented separately. Note that the data for Chloe/Daphne were gathered only during the first 12 weeks of development.
i i ~Q 0 g
0 0
C~
0 0 gE
5'
~.
~-.~
"
~.~
~~
~
~ ~
~ NE
ARES
T NE
IGHB
OR
~" ~
.. ~
0 ~8~
~"
0
0 N
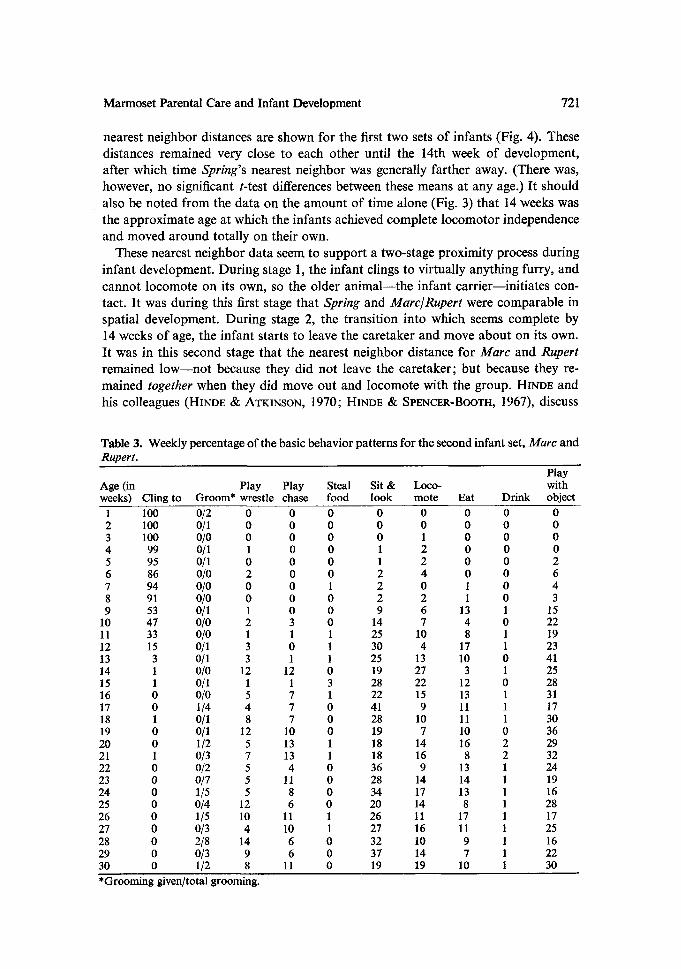
Marmoset Parental Care and Infant Development 721
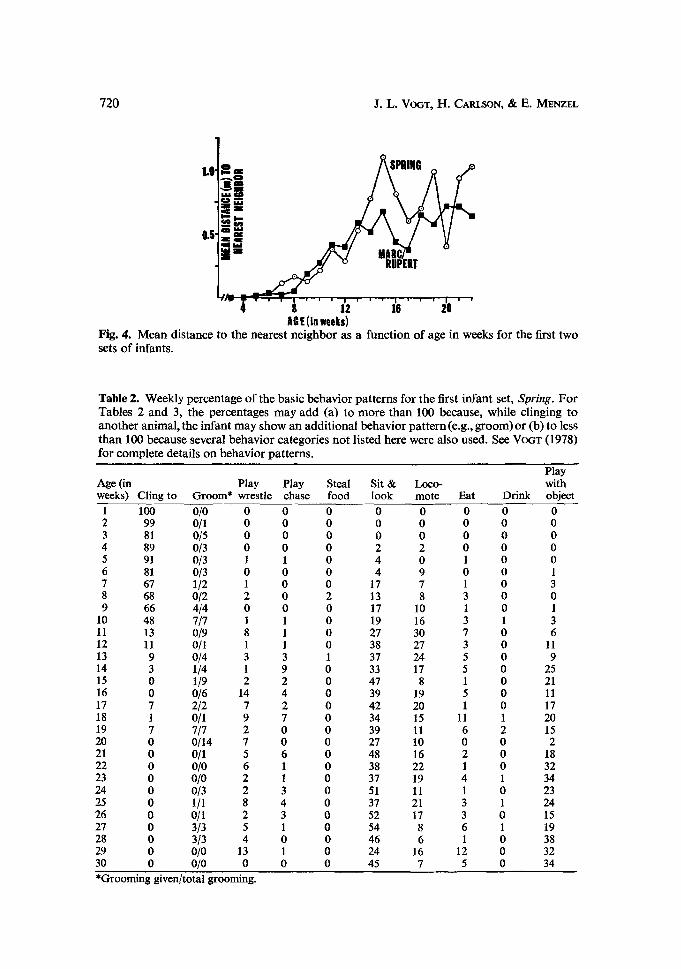
nearest ne ighbor distances are shown for the first two sets o f infants (Fig. 4). These
distances remained very close to each other unt i l the 14th week of development,
after which time Spring's nearest neighbor was generally farther away. (There was,
however, no significant t-test differences between these means at any age.) I t should
also be noted from the data on the a m o u n t of t ime alone (Fig. 3) that 14 weeks was
the approximate age at which the infants achieved complete locomotor independence
and moved a round totally on their own.
These nearest ne ighbor data seem to support a two-stage proximity process dur ing
infant development. Dur ing stage 1, the infant clings to virtually anything furry, and
canno t locomote on its own, so the older a n i m a l - - t h e infant carr ier- - ini t ia tes con-
tact. I t was dur ing this first stage that Spring and Marc/Rupert were comparable in
spatial development. Dur ing stage 2, the t ransi t ion into which seems complete by
14 weeks of age, the in fan t starts to leave the caretaker and move abou t on its own.
It was in this second stage that the nearest ne ighbor distance for Marc and Rupert remained l o w - - n o t because they did no t leave the caretaker; bu t because they re-
mained together when they did move out and locomote with the group. H~No~ and
his colleagues (HINDE & ATKINSON, 1970; HINDE & SPENCER-BOOTH, 1967), discuss
Table 3. Weekly percentage of the basic behavior patterns for the second infant set, Marc and Rupert.
Play Age (in Play Play Steal Sit & Loco- with weeks) Cling to Groom* wrestle chase food look mote Eat Drink object
1 100 0/2 0 0 0 0 0 0 0 0 2 100 0/1 0 0 0 0 0 0 0 0 3 100 0/0 0 0 0 0 1 0 0 0 4 99 0/1 1 0 0 1 2 0 0 0 5 95 0/1 0 0 0 1 2 0 0 2 6 86 0/0 2 0 0 2 4 0 0 6 7 94 0/0 0 0 1 2 0 1 0 4 8 91 0/0 0 0 0 2 2 1 0 3 9 53 0/1 1 0 0 9 6 13 1 15
10 47 0/0 2 3 0 14 7 4 0 22 11 33 0/0 1 1 1 25 10 8 1 19 12 15 0/1 3 0 1 30 4 17 1 23 13 3 0/1 3 1 1 25 13 10 0 41 14 1 0/0 12 12 0 19 27 3 1 25 15 1 0/1 1 1 3 28 22 12 0 28 16 0 0/0 5 7 1 22 15 13 1 31 17 0 I/4 4 7 0 41 9 11 1 17 18 1 0/1 8 7 0 28 10 11 1 30 19 0 0/1 12 10 0 19 7 10 0 36 20 0 I/2 5 13 1 18 14 16 2 29 21 1 0/3 7 13 1 18 16 8 2 32 22 0 0/2 5 4 0 36 9 13 1 24 23 0 0/7 5 11 0 28 14 14 1 19 24 0 1/5 5 8 0 34 17 13 1 16 25 0 0/4 12 6 0 20 14 8 1 28 26 0 1/5 10 11 1 26 11 17 1 17 27 0 0/3 4 10 1 27 16 11 1 25 28 0 2/8 14 6 0 32 10 9 1 16 29 0 0/3 9 6 0 37 14 7 1 22 30 0 1/2 8 11 0 19 19 10 1 30 *Grooming given/total grooming.

722 J. L. VOGT, H. CARLSON, & E. MENZEL
a quite similar process that takes place in captive rhesus (Macaca mulatta) mother- infant relations. The mother is initially responsible for maintaining proximity, but later on, as the infant starts locomoting more on its own, contact and proximity maintenance are primarily determined by the infants.
Behavioral development during the first 30 weeks of the first two infant sets is shown in Tables 2 and 3. These illustrate the percentage of selected behaviors at weekly intervals for Spring and for Marc and Rupert averaged. First it should be mentioned that qualitative observations during times other than the afternoon sessions indicated that the eating activity (Steal Food, Eat, and Drink) shown in Tables 2 and 3 was much higher at other times, especially during morning feeding. The data for several of the behaviors in Tables 2 and 3 are condensed into 2-week blocks and presented graphically in Figure 5. For the social behavior of Groom both sets of infants showed grooming very early in development, within the first two weeks. However, Spring showed relatively high groom levels in the weeks following, while Marc and Rupert showed elevated Groom interactions only some 18 weeks later. It should be noted from Table 3 that active grooming of another animal (as opposed to the receiving of grooming from another animal) for Marc and Rupert was not present in 24 of the first 30 weeks of their development and active grooming never amounted to over one- half of their groom interactions. Spring (Table 2), however, was showing a relatively high frequency of active grooming within his first ten weeks of development, and his enhanced level of groom interactions (Fig. 5) was probably attributable to this active grooming process. The other social behaviors considered in this study were the activities of Play Chase and Play Wrestle which were combined into one category, Social Play, in Figure 5. The plotted data for Social Play in Figure 5 revealed rather similar frequencies for both sets of infants until about 20 weeks of age. Beyond that point, Marc and Rupert clearly showed more play, which suggested a strong social facilitation effect in Social Play among similar-age siblings.
The other behavior patterns graphed in Figure 5, Sit and Look, Eat, Play with Object, and Locomote, were nonsocial behaviors, as none of them involved social interaction with another animal. The percentages for these behaviors were generally comparable, though Spring showed more Locomote and Sit and Look, and Marc and Rupert more Eat and Play with Object. Except for Locomote, which Spring showed quite frequently at 8 to 12 weeks of age, and for Groom, which accounted for a relatively small percentage of the infant's behavior, Marc and Rupert were the more active infants, more involved in eating and in social and nonsocial play. This in- creased activity on the part of the twin infants, Marc and Rupert, again strongly suggests a social facilitation process important in development.
The data on behavioral development indicate that the age at which each set of infants shows significant activity in several of the behaviors differs greatly. In groom- ing, for instance, Spring showed a high grooming level, including active grooming comparable to adult interactions, by ten weeks of age. Rather than engaging in groom interactions as did Spring, the twin infants, Marc and Rupert, showed in- creased Social Play activity--Play Chase and Play Wrestle--which would strongly suggest a social facilitation effect probably as a result of having same and similar age companions in the group.
In general, then, the behavior of the twin infants was much more action oriented,
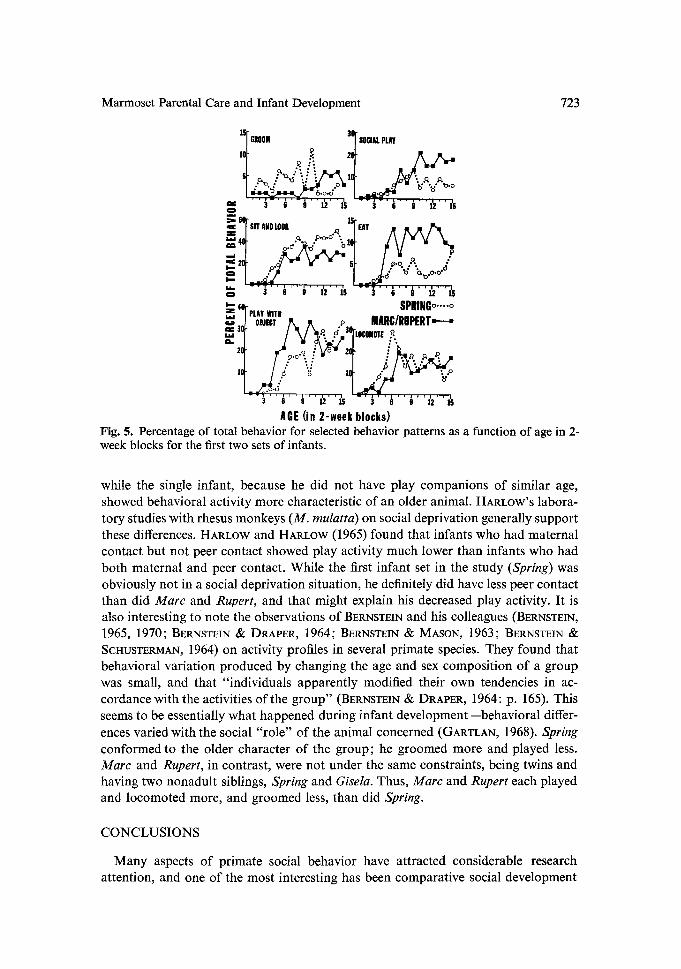
Marmoset Parental Care and Infant Development 723
15 3 [GROOM O[ SOCIAL PLAY
g .:- : : " "
_ [S,YA..,OA .~. i "r ~ r , , ~ / N _ ~ . . % .? .... o'~- 1 \ / " ~ x, .
O" ~ 9 , e l �9 O ;
/ ..or / d.d" ~ ~.o.o'" ~- l - ~ . ~ / _ _.o_~ "v
- ' ~ " 6 " ~ " i 2 " i s " ' t " ~ " ~ " h " ~ ~or SPRING', . . . . .
PLAY WITH ~ OHJm ~ . ~ MARC/RUPs =
�9 . : ' .
i _ ~ . / ~ . o . g i ~ . / " ' ~ " i " ~"r "' ~ " ~ " ~ " h " i s
AGE (in 2-week blocks) Fig. 5. Percentage o f total behavior for selected behavior patterns as a function of age in 2- week blocks for the first two sets o f infants.
while the single infant, because he did not have play companions of similar age, showed behavioral activity more characteristic of an older animal. HARLOW'S labora- tory studies with rhesus monkeys (M. mulatta) on social deprivation generally support these differences. HARLOW and HARLOW (1965) found that infants who had maternal contact but not peer contact showed play activity much lower than infants who had both maternal and peer contact. While the first infant set in the study (Spring) was obviously not in a social deprivation situation, he definitely did have less peer contact than did Marc and Rupert, and that might explain his decreased play activity. It is also interesting tO note the observations of BERNSTEIN and his colleagues (BERNSTEIN, 1965, 1970; BERNSTEIN & DRAPER, 1964; BERNSTEIN & MASON, 1963; BERNSTEIN & SCt-ItJSTERMAN, 1964) on activity profiles in several primate species. They found that behavioral variation produced by changing the age and sex composition of a group was small, and that "individuals apparently modified their own tendencies in ac- cordance with the activities of the group" (BERNSTEIN & DRAPER, 1964: p. 165). This seems to be essentially what happened during infant development--behavioral differ- ences varied with the social "role" of the animal concerned (GARTLAN, 1968). Spring conformed to the older character of the group; he groomed more and played less. Marc and Rupert, in contrast, were not under the same constraints, being twins and having two nonadult siblings, Spring and Gisela. Thus, Marc and Rupert each played and locomoted more, and groomed less, than did Spring.
CONCLUSIONS
Many aspects of primate social behavior have attracted considerable research attention, and one of the most interesting has been comparative social development

724 J. L. "VOGT, H. CARLSON, & E. MENZEL
(e.g., HINDE, 1974; POIRIER, 1972). The species of the marmoset family Callithricidae are particularly noteworthy in this regard, as they typically have twin offspring, and as the father does much of the infant caretaking. Two recent studies have reported observations on different species of marmosets: EPPLE (1975a) on Saguinusfuscicollis, the same species as in this study; and Box (1975) on Callithrix jacchus. EPPLE re- corded data from seven laboratory groups during the first 40 days of development and found that the dominant male (presumably the father) did most of the infant carrying, although all group members did carry the infants at some time. EPPLE commented, however, on the variability among these seven groups, and on the fact that in some groups the mother carried more than the father. Box (1975) reported on this last type of group; she observed during the first 28 days of development that the mother carried the infants more frequently than did the father, and the females more frequent- ly than the males.
The longer term data of the present study generally agree with EPPLE (1975a): the father accounts for more of the infant carrying than the mother. An even more clear cut conclusion came out of these data when the group males-group females compari- son was made. The males of the group almost always accounted for more carrying than did the females. During the three developmental periods when observations were made in this study, over 70 ~ of the time the males were carrying the infants. This paternal care was most conspicuous in the behavior of the subadult male Harry. As he was approaching adulthood at two years of age, intense fighting suddenly broke out one day between Harry and the adult male Plisch. Harry was removed from the group the same day, which was a few weeks after the birth of the second set of infants, Marc and Rupert. Up until the time he was removed, Harry was the most frequent carrier of the infants. During the development of the first infant (Spring) Harry accounted for the greatest amount (45 ~o) of the carrying and his last week in the group Harry accounted for 50 7oo of the infant carrying of Marc and Rupert. There were also several incidents of infant stealing by Harry. Once the infant was clinging to him, Harry would often rush to the opposite end of the greenhouse--a behavior that was not observed among the other animals. Moreover, this paternal motivation shown by Harry and the other males in the group seems quite comparable in form and function to the preoccupation with infants (play-mothering) by females of all ages observed in many Old and New World species: e.g., red howler monkeys, Alouatta seniculus (NEVILLE, 1972); squirrel monkeys, Saimiri sciureus (BALDWIN, 1971); and vervet monkeys, Cercopithecus aethiops (LANCASTER, 1971). Although caution must be used because of Harry's nonfamily status, one might refer to Harry's behavior as "play- fathering." Indeed it may have been Harry's emerging, adult paternal motivation that helped precipitate the fights and chases, and Harry's ouster from the group.
Development of spatial development of the first two sets of infants, Spring and Marc/Rupert, was quite comparable for both measures of spatial independence, per- centage of time alone (i.e., not being carried) and distance to the nearest neighbor. When taken together, the measures support a two-stage process of spatial develop- ment quite similar to that described by HINDE and ATKINSON 0970) and HINDE and SPENCER-BOOTH (1967) in captive rhesus mother-infant relations. The infant caretaker is initially responsible for spatial proximity, but the young animal gradually acquires

Marmoset Parental Care and Infant Development 725
the ability to locomote on its own so that, by 14 weeks of age for marmosets, the infant determines its spatial closeness to others. I t should be emphasized that com- plete spatial independence is not achieved at 14 weeks of age, or at any age, for that matter. Rather, the relative spatial dependence, in terms of distance to the other group members, becomes less obvious to the casual observer, though to be sure this factor is always present and important for the group living animal.
Comparison of selected behaviors for the first two infant sets showed that greater differences were present here than in either spatial development or infant carrying. I t may be that infant behavioral patterns are more dependent upon the composition of the group (cf., BER~STEIN & DRAPER, 1964) than are infant carrying and spatial development. Both of these latter activities seem to reflect a more basic and funda- mental process that occurs independently of the particular members of the group. While not exclusive of the context o f the social group, a basic process like infant care will take place regardless of group structure, so long as the rudiments of socialization are present. In contrast, it seems that behavioral development is much more influenced by the activity of the other group members.
Acknowledgements. The collection of data for this project was supported by National Science Foundation Grant GB-3850 to the Psychobiology Program at the State University of New York at Stony Brook. The writing of this paper was partly supported by the Faculty Research Fund at Saint John's University. The authors wish to thank Dr. GISELA EPPLE for supplying the animals in this study.
Much of the data reported here formed part of a Ph. D. dissertation submitted by the first author to the State University of New York at Stony Brook.
REFERENCES
BALDWIN, J. D., 1971. The social organization of a semifree-ranging troop of squirrel mon- keys ( Saimiri sciureus). Folia primat., 14: 23-50.
BERNSXEIN, I. S., 1965. Activity patterns in a cebus monkey group. Foliaprimat., 3 �9 211-214. - - , 1970. Activity patterns in pigtail monkey groups. Folia primat., 12: 187-198. - - & W. A. DRAPER, 1964. The behavior of juvenile rhesus monkeys in groups. Anita.
Behav., 12: 84-91. - - & W. A. MASON, 1963. Activity patterns of rhesus monkeys in a social group. Anim.
Behav., 11: 455--460. - - & R. S. SCHUSTERMAN, 1964. The activity of gibbons in a social group. Foliaprimat.,
2: 161-170. Box, H. O., 1975. A social developmental study of young monkeys (Callithrixjacchus) within
a captive family group. Primates, 16: 419-435. ChRiSTIAN, A., 1968. Haltung und Brutbiologie von Cebulla. Folia primat., 8: 41-49. EPPLE, G., 1967. Vergleichende Untersuchen uber Sexual und Sozialverhalten der Krallenaf-
fen (Hapalidae). Folia primat., 7: 37-65. - - , 1968. Comparative studies on vocalizations in marmoset monkeys (Hapalidae).
Folia primat., 8: 1-40. - - - - , 1970a. Maintenance, breeding, and development of marmoset monkeys (Callithri-
cidae) in captivity. Folia primat., 12: 56-76. - - , 1970b. Quantitative studies on scent marking in the marmoset (Callithrixjacchus)
in captivity. Foliaprimat., 13 : 48-62. ,1972. Social communication by olfactory signals in marmosets. Inter. Zoo Yrbk.,
12: 36--42.

726 J. L. VOGT, H. CARLSON, ~r E. MENZEL
- - , 1975a. Parental behavior in Saguinus fuscicollis ssp. (Callithricidae). Folia primat., 24: 221-238.
- - , 1975b. The behavior of marmoset monkeys (Callithricidae). In: Primate Behavior: Developments in FieM and Laboratory Research, Vol. 4, L. ROSENBLUM (ed.), Academic Press, New York, pp. 195-239.
GARTLAN, J. S., 1968. Structure and function in primate society. Foliaprimat., 8: 89-120. HAMPTON, J. K. & S. H. HAMPTON, 1965. Marmosets (Hapalidae): Breeding seasons, twin-
ning and sex of offspring. Science, 150: 915-917. - - , & B. T. LANDWEnR, 1965. Observations on a successful breeding colony
of the marmoset Oedipomidas oedipus. Foliaprimat., 4: 265-287. HARLOW, H. F. & M. K. HARLOW, 1965. The affectional systems. In: Behavior of Nonhuman
Primates, Vol. 2, A. SCHRIER, H. HARLOW, & F. STOLLNITZ (eds.), Academic Press, New York, pp. 287-334.
HERSnKOVITZ, P., 1968. Metachromism or the principle of evolutionary change in mamma- lian tegumentary colors. Evolution, 22: 556-575.
HINDE, R. A., 1974. Biological Bases of Human Social Behaviour. McGraw-Hill , New York. - - & S. ATICINSON, 1970. Assessing the roles of social partners in maintaining mutual
proximity as exemplified by mother-infant relations in monkeys. Anim. Behav., 18: 169- 176.
& Y. SPENCER-BOOTr~, 1967. The behaviour of socially living rhesus monkeys in their first two and a half years. Anita. Behav., 15: 169-196.
LANCASTER, S. B., 1971. Play mothering: The relations between juvenile females and young infants among free-ranging vervet monkeys (Cercopithecus aethiops). Folia primat., 15: 161-182.
MOYNmAN, M., 1970. Some behavior patterns of platyrrhine monkeys. I I : Saguinus geoffroyi and some other tamarins. Smithson. Contrib. Zool., 28: 1-77.
NAPIER, J. R. & P. H. NAPIER, 1967. A Handbook oJ Living Primates. Academic Press, New York.
NEVILLE, M. K., 1972. Social relations within troops of red howler monkeys (Alouatta senicu- lus). Folia primat., 18: 47-77.
POIRIER, F. E., 1972. Primate Socialization. Random House, New York. TRIVERS, R. L., 1972. Parental investment and sexual selection. In: Sexual Selection and the
Descent of Man, B. CAMPBELL (ed.), Aldine, Chicago, pp. 136-179. VOGT, J. L., 1975. The social structure of a marmoset monkey group. Ph.D. diss., State Univ.
of New York at Stony Brook. - - , 1978. The social behavior of a marmoset (Saguinus fuscicollis) Group I I : Behavior
patterns and social interaction. Primates, 19: 287-300.
- - Received January 13, 1977; Accepted August 29, 1977
Authors' Present Addresses: JERRY L. VOGT, Department of Psychiatry and Behavioral Sciences, Stanford University School of Medicine, Stanford, California 94305, U.S.A.; HAROLD CARLSON and EMIL MENZEL, Department of Psychology, State University of New York, Stony Brook, New York 11794, U.S.A.
Related Documents











