Robert V Blanden Harald S, Rothenfiuh Paula Zylstra Georg F. Weiller Edward I, Steele The signature of somatic hypermutation appears to be written into the germline IgV segment repertoire Authors' addresses Robert V. BInndeii-, Harold S. Ruihmfliih', Paula Zylslra^ GeoigE ^Veiller'. EJvvardJ. Sicde'', 'Division of Immunology and Cell Biology, Jobn Curtin School of Medical Research. Australian National University, Canberra, Australia, •'Department of Biological Sciences, University of Woliongong, Wollongong. .Australia. ^Bioinformatics Laboratory, Research School of Biological Sciences, Australian National University, Canberra, Australia. Correspondence to: Edward J. S(«le Department of Biological Sciences University of Wollongong Northfields Avenue Wollongong, NSW 2522 Australia Fax: 61 2 42 214135 e-mail: [email protected] Acknowledgements Supported in part hy the Australian Research Council, We are grateful to Rod Langman, Gerry Both and Martin Flajnik for critical discussion. \mrnunologicol Reviews 1998 Vol. 162:117-132 Primed in Denmofk. All rights reserved Copyrighl © Munksgaard 1998 Immunological Reviews 1SSN0J05-2896 Summary: We present here a unifying hypothesis for the molecular mech- anism of somanc hypertnutalion and somatic gene conversion in IgV genes involving rever.se transcription nsing RNA templates from the V-gene loci to produce cDNA which undergoes homologous recombina- tion with chromosomal V(D)J DNA. Experimental evidence produced over the last 20 years is essentially consistent with this hypothesis. We also review evidence suggesting that somatically generated lgV sequences from B lymphocytes have been fed back to gertnline DNA over evolutionary time. Introduction Somatic hypermuCation of rearranged V(D)J genes in B lym- phocytes is a feature of "affinity maturation" of antibody responses. As the immune response matures, B cells with the highest affinity antibodies progressively come to dominate the response. In this review we analyse current data and argue the case for the proposition that somatic mutations of rearranged V(D)j genes have been a part ofthe evolution of unrearranged germline V segments in extant higher vertebrates. This idea was first proposed by Steele (1) in the Somatic Selection Theory some 20 years ago. It was argued (1). on the basis of serological patterns of somatically variable idiotypes in rabbit families, that soma-co-germline flow of somatically mutated IgV genes was the simplest explanation ofthe paternal inheritance data. The soma-to-germline theory invoked a roie for endogenous retro- viral gene transducing vectors (originating in lymphocytes), reverse transcription of Ig mRNA, transfer of the somatically derived IgV cDNA to germ cells and homologous recombina- tion in the shaping ofthe DNA sequence structure of germline V segments. Since the late 1970s, enormous advances in DNA cloning and sequencing together with the use of transgenic mice carry- ing V(D)J genes have advanced our understanding ofthe mech- anism of somatic hypermutation. The accumulated findings strongly suggest a complex mechanism which is unlikely to 117

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Robert V BlandenHarald S, RothenfiuhPaula ZylstraGeorg F. WeillerEdward I, Steele
The signature of somatichypermutation appears to bewritten into the germlineIgV segment repertoire
Authors' addressesRobert V. BInndeii-, Harold S. Ruihmfliih', Paula Zylslra^GeoigE ^Veiller'. EJvvardJ. Sicde'','Division of Immunology and Cell Biology, JobnCurtin School of Medical Research. AustralianNational University, Canberra, Australia,•'Department of Biological Sciences, Universityof Woliongong, Wollongong. .Australia.^Bioinformatics Laboratory, Research School ofBiological Sciences, Australian NationalUniversity, Canberra, Australia.
Correspondence to:Edward J. S(«leDepartment of Biological SciencesUniversity of WollongongNorthfields AvenueWollongong, NSW 2522AustraliaFax: 61 2 42 214135e-mail: [email protected]
AcknowledgementsSupported in part hy the Australian ResearchCouncil, We are grateful to Rod Langman,Gerry Both and Martin Flajnik for criticaldiscussion.
\mrnunologicol Reviews 1998Vol. 162:117-132Primed in Denmofk. All rights reserved
Copyrighl © Munksgaard 1998
Immunological Reviews1SSN0J05-2896
Summary: We present here a unifying hypothesis for the molecular mech-anism of somanc hypertnutalion and somatic gene conversion inIgV genes involving rever.se transcription nsing RNA templates from theV-gene loci to produce cDNA which undergoes homologous recombina-tion with chromosomal V(D)J DNA. Experimental evidence produced overthe last 20 years is essentially consistent with this hypothesis. We alsoreview evidence suggesting that somatically generated lgV sequences fromB lymphocytes have been fed back to gertnline DNA over evolutionarytime.
Introduction
Somatic hypermuCation of rearranged V(D)J genes in B lym-phocytes is a feature of "affinity maturation" of antibodyresponses. As the immune response matures, B cells with thehighest affinity antibodies progressively come to dominate theresponse. In this review we analyse current data and argue thecase for the proposition that somatic mutations of rearrangedV(D)j genes have been a part ofthe evolution of unrearrangedgermline V segments in extant higher vertebrates. This idea wasfirst proposed by Steele (1) in the Somatic Selection Theorysome 20 years ago. It was argued (1). on the basis of serologicalpatterns of somatically variable idiotypes in rabbit families, thatsoma-co-germline flow of somatically mutated IgV genes wasthe simplest explanation ofthe paternal inheritance data. Thesoma-to-germline theory invoked a roie for endogenous retro-viral gene transducing vectors (originating in lymphocytes),reverse transcription of Ig mRNA, transfer of the somaticallyderived IgV cDNA to germ cells and homologous recombina-tion in the shaping ofthe DNA sequence structure of germlineV segments.
Since the late 1970s, enormous advances in DNA cloningand sequencing together with the use of transgenic mice carry-ing V(D)J genes have advanced our understanding ofthe mech-anism of somatic hypermutation. The accumulated findingsstrongly suggest a complex mechanism which is unlikely to
117

Blanden et al • Somatic signature in germline V genes
employ simple error-prone DNA repair processes involvingDNA template-directed DNA synthesis. Just over 10 years ago arole for error-prone reverse transcription in B lymphocytes wasinvoked hy Steele & Pollard (2) for the generation of somaticmutations within rearranged V(D)J genes and their immediate3' and S'-flanking sequences. It appeared at that time to he thesimplest explanation ofthe available DNA sequence data. It wasenvisaged that a mutator process was activated within antigen-stimulated B lymphocytes whereby mutated cDNA retrotrans-cripts ofthe transcriptionally active V(D)] gene were integratedback into the same V(D)J locus by homologous recombination(a form of locus-specific gene conversion). As far as we areaware, most of the current molecular evidence is consistentwith this model (Tobk 1) and this evidence was recently analy-sed and reviewed in Steele et al. (3). This view ofthe mutatorprocess is sitrnmarised in Fig. I.
We have divided this review into two parts. Firstly, we willhriefly analyse the data consistent with the reverse transcripcase(RT) model of somatic hypermutation (2), as embodied in thefunctioning ofthe hypothetical RT-mutatorsome, a telomerase-like particle (Fig. 1). This analysis will also suggest that the pro-cesses of somatic hypermutation in antigen-stimulated mouseand human B lymphocytes and the gene conversion processes(which may not require activation by foreign antigen) fordiversification of V(D)J somatic repertoires in chicken (4-8)and rabbit (9, 10) share common molecular processes viz.error-prone reverse transcription and homologous recombina-tion. Secondly, we will revisit the evidence consistent with theidea that the germline V-gene repertoires in such widely dis-parate vertebrate species as birds and mammals have beendiversified and maintained as functional open reading framesduring evolution by the senna-to-germline flow of cDNA ret-rotranscripts copied from the pre-mRNA of B lymphocytes(1 I). Apparent objections to this idea as outlined in Tomiinsonet al. (I 2) will be addressed.
How can a non-random mutation pattern be generatedin a DNA sequence?
There are several key issues with respect to how a non-randompattern of mutations can be apparent in a DNA base sequence(as occurs during the process of somatic hypermutation).Firstly, given that mutations occur by random errors in the DNAsequence, non-random patterns must be achieved by imposinga selection criterion on either the mutated DNA sequence or itsprotein (or RNA) product, so that those mutated genes thatsuccessfully meet the selection criterion are retained, and thosethat do not are discarded. For mutating B cells in a germinal
Table 1. Features of somatic hypermutation of rearranged V(D)J genes inB lymphocytes of mice'
1. Mainly point mutation, not gene conversion (62, 63).
1. Bias for transitions (59%) over transversions (64).
3, Sli and bias i.e. the mutator mechanism operates on one DNA strand (64).
1, Mutation is roughl/ focused on the V(D)J (65, 66),(a) Upsti-eam boundary is transcription start site (VI.) or L-V intron (y,)(67-70).(b) Downstream boundary is within J-C intron (VH, V,,) or downstream ofC region (V0(fe5. 67,71).
5. Distribution of mutations is asymmetric with a steep slope upstream ofthepeai< and a more gradual slope downstream of the peak into the j-C intron(33. 68).
6. The V(D)J coding sequence and all upstream sequence including the promotercan be exchanged for non-Ig sequence and mutation still occurs (72-74).
7. The Ei/MAR region in the J-C intron is essential for mutation (74). Otherdownstream enhancers of transcription also enhance mutation (74-76). A 300base sequence spanning the A-T rich 3'MAR appears essential (from analysis ofHackettetal. (77) and Azuma etal, (72)); the abundance of poly-A and poly-Tsequences su^ests a capacity for secondary RNA structure in the pre-mRNA,
8. When gene conversion is induced in tandem transgene constructs, there is anobligator/ association between V(D)J-targeted homologous recombinationand somatic point mutation (41).
9. Transgenic constructs with a tRNA coding sequence inserted between theV(D)j and Ei/MAR regions generally do not allow mutation in the V(D)j (39.40),
10. Transgenic constructs with Ei/MAR downstream ofthe C region mutate in theC region (30).
See Steele et al. (3) and Blanden S Steele (21) for more detailed analysis andreferencing.
centre, the selection criterion is antigen-binding affinity oftheir mutant antibody (13-19). Since most mutations reduceaffinity for the original antigen (20), there should logically bea mechanism to ensure that when successful mutation has takenplace, there is no further mntation which may destroy success-ful V(D)J sequences (encoding surface Ig molecules bindingwith equivalent or higher affinity than the antibodies producedat the start of the response). We propose that this is achieved bya feedback "stop mutation" signal resulting from antigen bind-ing by high affinity surface Ig (as indicated by the STOP signalin Fig. 1). But the most important issue with respect to somatichypermutation is the process which ensures that only V(D)Jgenes mutate. Most other genes, encoding housekeeping andtissue-specific functions, must be conserved to ensure properfunction. This can he achieved by invoking a locus-specificdevice in or near intronic enhancer/matrix attachment regions(Fi/MAR) of Ig loci so that the mutatorsome docks (binds)exclusively to the DNA/RNA of rearranged Ig gene loci, thustargeting the transcriptionally active V(D)J for mutation(Fig. 1). Current evidence supports this proposition (3, 21).
118 Immunoloj|i(.iii Rcvims 162/1998

Blanden et al • Somatic signature in germline V genes
.[/"Affinity-based "STOP" signal (Ag-Ab-FDC?)
Ig protein
Mutated L-VDJ-C transcript
Ei/MAR
Mutatorsome particle;-reverse transcriptase (RT)-RT primase (to make E-specific antisense RNA primer)
OR site-specific endonuclease-Various integrases, endonucleases?
Rearranged DNA5'
3'Pre-mRNA
Initiation of reverse transcriptionfrom 3'-OH of a DNA nick or fromantisense RNA primer
Fig. I. The reverse transcriptasemutatorsome. Reverse transcription isinitiated from an antisense RNA primer orfrom a nick or double-strand break in thetarget DNA (see Steeie et al. (3)). Nucleotidemisincorporations (indicated by x) canoriginate during boih transcription andreverse transcription. The cDNA then rt^piacesits germline counterpart via homologousrecombination. Successful competition forfollicuiar dendritic cell-presented epitopes ofantigen-antibody complexes by the mutatedsurface-bound antibody molecule signalstermination of tbe mutator process. Failureto bind antigen can result in cell death(apoptosis) or repeated round (s) of mutation.The relative positions of the Cap site(|^)leader (L), rearranged V region (VDJ),unrearranged J elements (J) and enhancer (E)are indicated.[Figure republished wiih permission of ibepublisher of Iinmunoiogy and Cell Biology,Biackwell Science Asia.l
Wu-Kabat variability patterns amongst related somaticmutants generated during anti-hapten responses
Non-random distributions of point mutations are typical ofcollections of somatically mutated IgV regions derived from thesame germline V segment. Affinity-based selective forceswithin the germinal centre lead to a population of antibodiessuch that amino acid replacement suhstitutions are favoured inthe complementarity-determining regions (CDRs) and conser-vative substitutions (e.g. amino acid-silent codon changes) arefavoured in the framework (FW) regions (13), This is clearlyillustrated for collections of somatic mutant sequences of theheavy chain VHI 86.2 gene following immunization ofC57BL/6J mice with the 4-hydroxy-3-nitrophenyl acetyi (NP)hapten. Wu-Kabat variability plots (22) of such sequence datashow elevations of nucleotide and amino acid variabihty coin-ciding with the CDRl, CDR2 andCDR3 regions (13). CDR3 isa somatic construct in which variability is generated by V(D)Jrearrangement. However, tbe variability peaks in CDRl andCDR2 are the consequence of somatic mutation and antigen-binding selection events acting on Ig heterodimers displayed onthe surface membrane of mutated B lymphocytes.
What has been selected during evolution?
Recent experiments of Zinkernagel's group (23, 24) haveshown that the immune response against certain viral proteinsdoes not display affinity maturation requiring somatic hyper-mutation during a protective antibody response. High affinityantibodies are no better at clearing viral infection than lowaffinity antibodies. These results imply that optimally func-tional antibodies could be selected from the original somaticrepertoire (germline plus combinatorial) without the need formutation during the course of infection. Thus, in an evolution-ary context, somatic hypermutation and affuiity maturationmay now be redundant in higher vertebrates.
At the beginning of the evolution of the adaptive immnnesystem, when the germline V-gene repertoire was muchsmaller than in modern vertebrates (at some point there musthave been a single, primordial gene), somatic hypermutationwould have conferred a great selective advantage by expandingthe limited somatic repertoire during the life of an individualanimal. Moreover, if a soma-to-germline feedback loop wereoperative, then somatic mutation of antibody variable genescould have been a critical evolutionary development to acceler-ate the diversification of germline V-gene repertoires. As far aswe are aware, availahle information is compatible with this sce-nario. Somatic hypermutation is present in all jawed vertehrateswhich possess an adaptive immune system, including the car-tilaginous fish (25-27). We will revisit this issue again shortly.
Immunofogical Reviws 162/1998 119

Blanden et al • Somatic signature in germline V genes
RT-mutatorsome: error-prone copying involvingRNA intermediates
Most DNA polymerase enzyme complexes have several error-correcting and editing functions. Mutations such as hase suh-stituti(nis generated during DNA replication are therefore lowprobability events (28), The maxima! error rate is probably lessthan one base change in 10** bases replicated, although the trueerror rate is prohably much lower at <10"'", This is extraordi-narily high fidelity information copying and it depends onduplex formation allowing the checking of the newly syntbe-sised strand by its base pairing within the double-strandedDNA helix (see (28)), We have lahelled all the alternative mech-anisms to the RT model as "DNA-based" (29) because they alldepend on faulty repair functions of DNA synthesis, and thisincludes the recent transcription-coupled DNA repair model ofPeters &Scorb (30),
In contrast, the producti(ni of single-stranded RNA from aDNA template and the production of cDNA from an RNA tem-plate by reverse transcription are error-prone copying processesgenerating point mutations at error rates of 10 Uo 10+ (28),Repair ofthe sequence cannot be effected because this requiresthe unaltered template strand to engage in douhle-strandedduplex formation with the new strand to allow template-directed correction of the error. Furthermore, the enzymesinvolved in error-prone polynucleotide copying (RNA poly-merase, RT and RNA replicase) do not possess proof-readingand error-correcting repair functions (31), These error ratesare similar to the experimentally determined rates of somaticmutation in vivo (32),
These facts contributed to the development ofthe RT-muta-torsome concept (2, 3). Somatic point mutations are predictedto arise through natural error-prone copying via a low fidelitycopying loop involving an RNA intermediate, Error-pr<DnecDNA synthesis is predicted to he initiated on the pre-mRNA inthe J-C intron at or near the Ei/MAR sequence (Fig. I), thusensuring cDNA synthesis back through the V(D)J region whichmust terminate at or before the 5' end ofthe pre-mRNA. Themutated cDNA species thus generated (see below) are thenintegrated by homologous recombination into the expressedV(D)J region, replacing the original rearranged V(D)J DNAsequence. Somatic mutations are therefore restricted to theV(D)J transcription unit, protecting both upstream promoter(P) elements and downstream constant region (C) elementsfrom the crippling effects of point mutation.
Key features of somatic hypermutation of rearrangedV(D)J genes
In Table 1, we have distilled out the main features of somatichypermutation that we consider require explanation. As wehave analysed elsewhere (3), all the data in Table ( can be acco-modated within the interpretative framework of the RT con-cept. Points 1 through 6 and the main message of point 7 arenon-controversial. However, points 8, 9 and 10 are our inter-pretation ofthe data from the laboratories of E. Seising, P Gear-hart and U, Storb respectively and require complete reading ofthe cited papers. We believe the phenomena summarized inpoints 8 and 9 are incompatible with "DNA-based" models.Point 10 illustrates the crucial role of the Ei/MAR region, addi-tionally documented hy the transgeneic mouse studies of Mil-stein and co-workers (points 6 and 7),
How are the mutations distributed in the target region?
Somatic mutations are localised on a short stretch of DNAsequence in and around the rearranged variable gene (points 4and 5, Table I), They are rarely found furtber upstream beyondthe transcription start site nor are they found to any significantextent downstream beyond what we predict is the "locus-spe-cific device", the Ei/MAR,
In our view, this targeted distrihution illustrates tT,vo keyprinciples of somatic hypermutation. The separation of DNAsequences encoding V and C regions implies tbe evolutionaryadvantage of allowing mutational change in the V genes whilstrigorously conserving the C genes. The focusing of somaticmutations to the V(D)]-region and not the C-region gene isfundamentally important for heavy chains hecause tbey medi-ate antihody functions such as promotion of complement-dependent lysis of target cells, phagocytosis and B-cell signal-ling. This focusing is not needed in the case of light chainsbecause tlieir C regions do not have these functions (point 4b,Table 1) (see Motoyama et al, (71)),
The second principle is that mutations should not occurupstream of the V(D)J gene (points 4a and 5, Table 1) becausethis upstream region contains key regulatory sequences (the Pelements) which allow the binding of the RNA polymerasecomplex initiating transcription and the production of themRNA (at the position indicated by the directional arrow or"Cap" at the S' end ofthe mRNA) (Fig. I),
The positively skewed asymmetrical distribution ofsomatic mutations observed experimentally in populations ofrelated mutant sequences (point 5, Table 1) can be generatedtheoretically by the production of multiple cDNA species of
120 Immunological Revievvi 162/1998

Blanden et al • Somatic signature in germline V sene
varying lengths (3, 33) by reverse transcription of the pre-mRNA beginning at various points upstream of Ei/MAR(downstream boundary) and proceeding close to the upstreamb(mndary defined by the 5' end ofthe pre-mRNA. This distri-bution emphasises that in the analysis of somatic hypermuta-tion data it is important to determine the entire 5'-to-3' distri-bution. If this is not done, then unwarranted interpretativeemphasis may he placed on a portion of the mutational targetregion (34, 35),
Interim summary: mechanism of somatic hypermutationof V(D)J genes
The distribution of mutations plus the kn<jw-n error rates ofnucleic acid copying involving RNA intermediates were twokey facts which led to the development of the prototype RTmodel by Steele & Pollard (2). Its conception by one of us(F, J. Steele) was inspired by, and thns derived from, theSomatic Selection soma-to-germline idea (36), However, criti-cal re-evaluation by R. V Blanden has suggested that in evolu-tionary time, RT-based somatic bypermutation should precedethe hypothesised soma-to-germline transmission of V-genecDNA as discussed ahove. This idea was the stimulus fordetailed analysis of the genetic recombination signature ofgermline V genes (below) (37),
The RT model is consistent with the vast majority of exper-imental results concerning somatic hypermutation as exempli-fied by the list in Table i, It can be extended to explain molecularmechanisms which operate to somatically diversify chickenrearranged variahle region V(D)J genes, in a process hithertoknown as "gene conversion" (below) (21).
We have proposed that the "molecular machine" whichmutates rearranged V(D)J DNA at a high rate he termed the RT-mutatorsome (Fig, 1), which uses the unspliced pre-mRNA as atemplate for synthesising cDNA. The telomerase enzyme com-plex (38) provides a precedent for RT activity in a cellularmolecular machine. We suggest that reverse transcription is ini-tiated at any one ofa number of potential reverse transcriptionprimer sites (3) downstream of the V(D)J near the Ei/MARregion and proceeds upstream co or near to the Cap site (the 5'end of the pre-mRNA template). We propose that Ei/MARallows "docking" of the RT-mntacorsome onto the rearrangedV(D)J gene with respect to hoth chromosomal DNA and pre-mRNA hecause of specific RNA and DNA structures bound hymutatorsome protein subunits. Experimental testing of thesepredictions is in progress. We also propose that tbe mutatedcDNA copy of the coding strand of the V(D)J region is inte-grated back into the chromosome and displaces the original,
unmutated sequence (indicated by the looping arrow in Fig, 1),thus explaining the observed strand bias of hypermutation (sat-isfying point 3, Table 1), This theoretical scenario ensures thatupstream P elements and downstream C regions are hcDth pro-tected, but the non-coding DNA in the immediate environs ofthe V(D)J is mutated at a very high rate (the same rate of errorgeneration known for transcription and reverse transcription ofabout 10" to 10 " per base per copying cycle (31)). The molec-ular directional rules of copying from DNA or RNA templatescoupled with intrinsically error-prone RNA and cDNA synthe-sis are therefore entirely consistent with the requirements forsomatic mutaticni.
In the mouse, the observed upstream boundary of hyper-mutation in data available at this time is near the transcriptionstart site for H chains and somewhere in the L-V intron for lightchains (point 4, Table 1), These are consistent with the siteswhere the proposed cDNA synthesis could terminate, eitherwhen the RT comes to the 5' end of the pre-mRNA template(the Cap site), or at the boundaries of the L-V intron, if chesplicing ofthe pre-mRNA template intervenes; 5'-to-3' exonu-clease activity acting on che pre-mRNA could also move the 5'boundary downstream.
Data outside tbe ambit of conventional DNA-based modelsyet predicted by the RT model
Firsdy, as far as we are aware, tbere is no experimental evidenceof a mechanism which selectively stops DNA synthesis. Oncebegun, synthesis proceeds until the end of the template isreached. Thus, DNA-based hypermutation models require theinventi(Dn of special rules of DNA synthesis. In contrast, the RTconcept is based entirely on documented molecular mecha-nisms.
Secondly, in experiments reported by Gearhart's group, a"tRNA reporter" sequence was located just downstream oftherearranged V(D)J region in the J-C intron hut upstream of theEi/MAR (39, 40). The majority of these data (3, 33) shows thatthe tRNA reporter sequence suppresses mutation in the V(D)Jtarget area (point 9, Table 1), a result incompatible with DNA-based theories, but consistent with the RT model. Our interpre-tation is that the tRNA reporter prevented cDNA synthesis pro-ceeding into the V(D)J target area in one or more of three ways.Eirstly, when DNA sequence encoding the tRNA reporter is trans-cribed, the RNA should fold into the clover leaf tRNA structure.In our model, reverse transcription may not proceed throughthe tRNA secondary structure and cDNA synthesis would termi-nate before reaching the V(D)J, Secondly, the tRNA sequencewithin the pre-mRNA may be rapidly cleaved by endonucleases
Immunoiogira] Review's 162/1998 121

Blanden et al • Somatic signature in germline V genes
specific for the 5' and 3' ends ofthe tRNA, releasing it from thepre-mRNA and prematurely truncating the pre-mRNA templatefrom which cDNA is reverse transcribed. Thirdly, known chem-ical modification of the ribonucleotide bases in the tRNA couldalso inhibit cDNA synthesis by the RT-mutatorsome.
Finally, we recall that onr model involves an ohligatoryrequirement for homologous recombination following error-prone reverse transcription (Fig. 1). Xu & Seising (41) haveshown that in a transgenic mouse, nsing two shghtly differentrearranged V(D)J genes linked close together, somatic pointmutation of the target V(D)J always involved recombination(gene conversion) with the other closely related V(D)Jsequence (point 8, Table 1). The ohligatory association ofsomatic point mutation with recombination is the strikingfinding. This result is inconsistent with DNA-based hypermuta-tion theories hecause they shonid allow hypermutation to takeplace without obligatory recombination. However, as can beseen in Fig. 1, the RT model demands "homologous recombi-nation"; it is integral to the mutator process. Point mutationsdo not accrue in the chromosomal V(D)J target DNA sequenceunless the mntated cDNA homologonsly recombines so as todisplace the original, unmntated V(D)J sequence.
What is the signal for somatic mutation to stop?
We argue (3) that when a cell surface Ig receptor on aB lymphocyte (centrocyte) has mutated to a point where it pos-sesses high affmity for the antigen (displayed as antigen-anti-body complexes on the foUicular dendritic cells (FDCs) in thegerminal centre), a signal shonid he given to the B cell to stopit mutating further. There is theoretical selective advantage inavoiding the risk of losing binding affinity for the antigen byfurther mutation. Most mutations that change amino acidseqnence adversely affect affinity illustrated by the recent site-directed mutagenesis work from Rittenberg's laboratory (20,42). We propose that when a mutant B cell strongly binds toepitopes presented on a FDC, a "STOP" signal is delivered toturn off the somatic mutation process (Fig. 1), for example byturning off production ofthe protein subunits ofthe RT-muta-torsome. Evidence that memory B cells preferentially remain inthe outer periarteriolar lymphoid sheath and differentiate intoplasma cells rather than initiate germinal centre reactions(reviewed in (43)) is consistent with this idea.
Is cell division necessary for mutation?
All DNA-based models of mntation are cell-division dependentbecause they assume DNA replication (and thus cell division)
must take place for mutations to be generated. The mainstreamassumption seems to be that mutations are generated duringthe rapid cell division of centroblas:s which provides the largeB-cell population (centrocytes) in the germinal centre (e.g. see(16, 18). Flowever, the hypothesised RT-based process isdependent on neither DNA replication nor cell division. Celldivision would be a selective advantage for the animal in B cellswith high affinity antihody. but wasteful in mutant B cellswhich have lost affinity for the antigen. Therefore, we suggestthat the RT-mutatorsome is most likely assembled at the end ofthe rapid prohferative phase of centroblasts during differentia-tion of centrocytes (3).
The paradigm that "cell division is necessary f(5r mutation"has become so deeply entrenched that the rate of somatic muta-tion is expressed as " 10" to 1 0"'* per base per cell generation".Thus, the conventional description of mutation rate tacitlybiases thinking about the mechanism.
Because the RT m(3del is cell-division independent, wefavour a cychc process based on "mutate -^ pause (for expres-sion of Ig) -> affmity test" occurring within a non-dividing cellwhich is transcriptionally active i.e. producing mRNA and pro-tein molecules of H and L antihody chains and the componentsof the RT-mutatorsome. This would be a more advantageousway for evolution to have organised the mutation/selectionprocess so as to maximise the production of high affmity anti-bodies than a process hased on random mutation during celldivision.
Unification of the molecular mechanism of somatic mutationand gene conversion
The B-cell repertoire in birds and rabbits is generated in gut-associated lymphoid tissues by a process described as gene con-version (4—10). Hitherto this one-way homologous recombi-nation process has been thought of as heing different from theprocess of somatic hypermutation. However, theoretically theycan be linked at the molecular level by the processes of error-prone reverse transcription and homokjgous recomhination(21).
We will illustrate this by reference to the chicken lightchain (IgL) locus as a prototype for discussion of the possibleshared molecular mechanisms. The chicken IgL locus containsonly one fully functional Vj.-gene segment capable of rearrange-ment with a Jx segment, and subsequent expression as L-chainprotein. A 1.8 kb stretch of DNA is excised (released as a circu-lar episonne) on rearrangement hetween V and J. Upstream ofthe V gene there is an array of 25 V pseudogenes tightly packedinto 19 kb of DNA (5). During the generation ofthe antigen-
122 Immunolo}iicni 162/199S

Blanden et al • 5omatic signature in germline V genes
"Close-packed"V pseudogenes
Triplet base insertionsand deletionsPieces of DJ sequenceand N nucleotides
long read-through transcript of RNA from the un rearranged locus
5'-to-3' exonuclease
cDNAdonated to VJ by homologous recombination
Fig. 2. Gene conversion in chickens. Aschematic outline of the mechanism ofdonation of pseudogene sequence torearranged VJ. The number of light chainpseudogenes has been reduced from 25 to 4 fordiagrammatic clarity. The top line shows thechromosomal DNA in a rearranged locus. Thesecond hne shows a RNA transcript from theunrearranged locus which in the thkd line hasformed a loop by base pairing as described in tbetext. We postulate tbat the RNA binds to tbechromosomal DNA via locus-specific structuresassociated with the Ei/MAR region. Thus,pseudogene sequences are brought physicallycloser to the rearranged VJ locus. cDNA producedby reverse transcription of any of the RNAsequences witbin or outside the loop arepotential donors by boinologous recombinationinto tbe chromosomal DNA of the recipient VJsequence, FTom Blanden & Steele (21).[Figure republi.shed with the permission ofthepublisher of Immunology and Cell Biology,Blackwell Science Asia,]
independent pre-immune B-cell repertoire in the bursa of Fab-ricius (after rearrangement of the single functional V to theJ-gene segment), variahle length blocks of V-pseudogenesequence are donated by homologous recombination to therearranged VJ. The precise recombination mechanism isunknown, although it apparently involves unidirectionaJ dona-tion of nucleotide seqnence from pseudogene to functionalgene via a homologous recomhination process. It is widelyassumed that the mechanism involves direct DNA-to-DNAdonation and has been termed gene conversion.
Firstly, we assume that such a mechanism must be limitedto Ig loci because, if it occurred in other multigene familiesdttring development ofthe animal, severe biological problemswould result. Therefore, we have explored mechanisms analo-gous to the RT-mutatorsome concept outline above (Fig. 1).
Because it is necessary to protect the C region from muta-tional change, we again postulated a role for an Ei/MAR ana-logue in the J-C intron. We then formulated RT models hasedupon long "sterile" transcripts of RNA originating upstream ofthe close-packed V-pseudogene array and continuing throughthe functional V and J genes, the J-C intron and the C region.As before, secondary structure in Ei/MAR KNA and DNA wouldlocalise a molecular complex analogous to the RT-mutatorsomepostulated above. We first envisaged the long RNA transcriptbehaving like a loose hall of string with protruding randomloops which provide RNA templates for cDNA synthesis ofpseudogene sequences. These cDNA sequences could nndergohomologous recombination with the rearranged VJ chromo-somal DNA,
As we applied this model to tbe chicken IgL locus, itbecame clear that available data suggests non-random processesin the sequence donation process. Thus, 8 out of the 25pseudogenes are orientated in the opposite direction to the sin-gle functional VL element and the remaining 17 pseudogenes.These 8 pseudogenes donate sequence much more frequentlythan the other 17 pseudogenes; additionally, more distalpseudogenes generally donate less frequently than more prox-imal pseudogenes (see McCormack & Thompson (7)). Howcan non-random loops be formed in the long RNA transcript?The simplest mechanism would involve substantial stretches ofG-C or A-U base pairing. Examination ofthe DNA sequences ofthe 25 VL pseudogenes reveals that two of them (VIO and VI 3)have poly-A sequences of 1 2 and 15 hases, respectively, imme-diately adjoining the downstream end ofthe coding sequence(5). A search for a poly-T(U) sequence, which could base pairwith either one of these poly-As and produce a loop in an RNAtranscript, revealed a 14T stretch about 1 kb downstream ofthefunctional V gene and about 500 bases upstream of the func-tional J gene. In our view, such highly non-random sequencestretches are very likely to have functional significance. How-ever, the intervening sequence between tbe funcdonal V and Jgenes is excised during DNA rearrangement. This means that,in a iTiodel invoking loops formed by the 14U stretch, the RNAtemplates must be provided by sterile transcripts from theunrearranged locus. Invoking an essential functional role forsuch a transcript would demand that in mature B lymphocytesof the chicken which have undergone gene conversion, thereshould be one nnrearranged V locus. This seems to be the case.
Immimoioj|iLai Rtvifws 162/1998 123

Blanden et al • Somatic signature in germline V genes
which is in marked contrast to B lympbocytes in mice, inwhich both loci are often rearranged (review hy McCormack &Thompson (44)).
The mechanism that we propose for the so-called geneconversion process is shown in Fig. 2. We postulate that a loopforms in the long transcript of RNA from the unrearrangedlocus by base pairing of the 14U sequence in the V-J intronwith the 12A sequence ahutting VIO, or with the ] 5A sequenceabutting V13. Because V13 is orientated in the same directionas the functional V and VJ sequences, the diagram showing theloop in the figure represents the case where the 15A sequenceabutting VI3 is used to base pair with the 14U sequence, sothat V13 is immediately outside the loop and V10 is inside theloop. Since VIO is orientated in the opposite direction to thefunctional V and VJ sequences, if the 12A sequence abuttingV10 is used to base pair and form the loop. V10 will be imme-diately within the loop. (Of course, in this antisense case, the1 2As adjacent to VI0 will actually be 12Us and the intron willcontain a 14A stretch, but to avoid confusion we have nsed thesense terminology.)
With the loop formed and "anchored" in the rearranged VJlocus via postulated secondary structure of the RNA and DNAofthe Ei/MAR analogue and binding of proteins of a molecularcomplex analogous to the murine RT-mutatorsome, cDNAwould be produced using RNA templates either inside or out-side the loop. The first function of the loop is to bring thepseudogene donor sequences in the RNA transcript physicallycloser to the target rearranged VJ chromosome DNA to opti-mise homologous recombination of cDNA and chromosomalDNA, The second function is to protect RNA sequences withinthe loop from degradation by 5'-to-3' exonuclease activity
The model fits the published data with one major apparentcontradiction. Firstly, it provides an explanation for the pres-ence of one unrearranged IgV locus in mature chicken B cells.Secondly, it accounts for the presence of point mutations inaddition to apparent gene conversion in rearranged chickenand rabbit IgV sequences (5, 10, 45), since the production ofRNA and cDNA are error-prone copying processes. Thirdly, thelocation of the postulated loops in the long RNA transcriptfrom the unrearranged locus also correlates strikingly witbpublished data in Figure 4 of McCormack & Thompson (/).This figure shows the frequency of V pseudogenes as gene con-version donors. As a group, the 8 pseudogenes with oppositeorientation (antisense) to the functional V gene donatesequence more frequently than their counterparts orientated intbe same direction (sense) as the functional V gene. Our inter-pretation of this result is that the production of long RNA trans-cripts in the anti-sense orientation is quantitatively greater than
that in the sense orientation (because they will be indepen-dendy regulated). Tbis wciuld provide a quantitatively greateropportunity for donation from the antisense transcript. Theother interesting result is that as a group, pseudogenes locatedwithin tbe putative loops of RNA (V12 to V1) donate sequencemore frequently than those outside the loops (VI 3 to V25);this is particularly striking for genes in the sense orientation.The model presented here accounts for this by the expectedvulnerahihty of single-stranded RNA templates outside the loopto degradation by 5'-to-3' exonuclease activity (see Fig. 2).
The apparent contradiction concerns the interpretation ofdonations of Carlson et al. (46) that their data obtained from F,hybrid chickens indicates that pseudogene donations alwaysoccur in cis. However, in the RT model presented here, any genelocated within a postulated RNA loop could serve as a high fre-quency trans donor. This includes the functional V sequencefrom the unrearranged locus. Because ofthe high homology inthe 5' L and L-V intron regions between this sequence and therearranged VJ, and tbe 5'-to-3' direction of integratioiKjf cDNA(see McCormack & Thompson (7)) we postulate that it almostinvariably dominates donation. Additional smaller donationsfrom the V pseudogenes lacking L regions would be superim-posed on the large donation from the functional V sequence.
In summary, this RT model unifies, at the level ofthe basicmolecular mechanisms, somatic hypermutation and somaticgene conversion in B lymphocytes of higher vertebrates. Sincethe chicken (5, 45) and rabbit (10) employ both somatic muta-tion and gene conversion to generate a B-cel! repertoire, itseems more probable that common molecular mechanisms areinvolved than otherwise. The model can be tested by manipu-lation ofthe Ig loci with the goal of interfering with the postu-lated single-stranded nucleic acid intermediates.
Inheritance of acquired somatic mutations?
The weight of current data is consistent with a RT-based, error-prone, V(D)J-gene feedback process operating within a B lym-phocyte, whether this be of avian or mammalian origin. Giventhe fact of extensive somatic mutation during immuneresponses, does any of this somatic genetic variabihty feed backto germ cells and play a role in the evolution of multigene Vfamihes? Can tbe homologous recombination event that we haveproposed to take place within an individual B lymphocyte beextended to the transfer of cDNAs of V-coding sequence from aB lymphocyte to the germline DNA? We addressed these issues5 years ago in Imniunological Reviews (13). Here we will extend thisanalysis to current data and outline our explanation of ho\v thegermline DNA sequences of antihody variable genes have
124 Immunologicd Rwims 162/1998

anden et al • Somatic signature in germline V genes
changed in response Lo anugenic stimulation over evolutionarytime. The evidence is consistent with the operation of a genefeedback loop involving the flow of genetic information fromsomatic cells (lymphocytes) into the germline genes (11).
Why should non-random Wu-Kabat variabilitybe found in germline DNA?
Given that non-random variability in V-gene DNA in the germ-line can only be fashioned during evolution by selection viaantigen-binding by functional protein heterodimers in whichthe complementary folding of the variable portions {encodedby rearranged V(D)J genes) of H and L protein chains forms anantigen-ccDnibining site, how is it possible for unrearrangedgermhne V-gene segments to show highly detailed evidence ofantigen-binding selection? The DNA sequence data consistentwith strong selection in moiise, human, amphibian and avianspecies has been documented (11, 47, 48).
The Somatic Selection Theory predicts the soma-to-germ-line transmission of acquired somatic mutations of antibodyV-region genes (i). It could be effected via the agency of RTsof either cellular or viral origin plus the movement of lympho-cytes to germ tissue (49) or the ubiquitous, naturally occur-ring, endogenous RNA retroviruses (produced by lympho-cytes) acting as "gene shuttles" ferrying mutated V-regiongene sequences into germ cells. Physical integration of somati-cally derived DNA sequences into the germline DNA by homol-ogous recombination (so as to replace a pre-existing genesequence) would also be required.
Is the somatic mutation/selection "signature" writteninto germline V genes?
ha contrast to other genes, it is the clear distinction between thegermline (unrearranged) configuration versus the somatic(rearranged) configuration of V genes which allows a rationalapproach to this problem. Functional, rearranged V(D)J genesoccur and are expressed only in mature B lymphocytes, andonly the products of such clonally expressed genes are thedirect targets of the rigorous antigen-driven somatic mutationand selection process in the germinal centre. Why is it then thatclear features of DNA sequences derived from the somatic con-figuration can also be present in the V segments in the germlineconfiguration?
Let us amplify the key points (Toble 2). Firstly germline Vsegments can never be the direct targets of antigen-bindingselection. Only a fully assembled Ig H-l-L heterodimer can beselected or tested for antigen-binding function. Germline
Tflbie 2. Some key features ot unrearranged germline V segments'
1, Germline V segments, in general, display a significant deficit of stop codonsand point mutation-gene rated pseudogenes below numbers expected fromrandom point mLitation.
2, Codon-by-codon analysis of the V-coding segments shows an excess ofnon-synonymous changes in CDRs and synonymous changes in FW regions,
3, Insertion-deletion events (assumed to be diagnostic of previousrecombination errors) display two patterns. Within the coding regionsthere are insertions or deletions of triplet codons or multiples thereof. Innon-coding regions there are variable length insertions and deletionsclustered around the transcription start site, within the leader (L-V) intronand immediately downstream of the 3' edge of the V-coding segment.
4, Heavy chain V chicken pseudogenes contain 3' termini with pieces ofD sequence, all in the preferred reading frame (R. Langman, personalcommunication) and the 3' ends of both the H and L-chain V pseudogenesshow evidence of nucleotide addition and trimming diagnostic of somaticV(D)) rearrangement.
5, Phylogenetic analysis of 5'-flanking sequences compared with thetranscription unit or V-coding segments indicates that the flanking and codingregions are evolving independently
6,'- Recombination analysis using the phylogenetic profile algorithm withinfamilies of mouse (VHI 86,2-related) and human (V ) germline V segmentsshows that major sites of recombination coincide with the borders of theL-V intron, the transcription start srte and the 3' end of the V segment.
-Summarised from Rothenfluh et al. (11) for V, ,186,2-related genes and V^Oxi -related genes from mice, related subfamilies of Vn gene segments from human,rabbit and Xenopus, V pseudogenes from chicken L and H-chain germline loci.
' for phylogenetic profile analysis see Weiller et al. (37).
V-gene segments are never translated; they are only expressedin mature B lymphocytes following V(D)J rearrangement.Functional studies thus far have found only about half of thehuman germline V-gene repertoire in V(D)J sequences (50).Many may never be expressed as V(D)J rearrangements andthus cannot be subjected to selection. This is a conundrum forconventional evolutionary theory.
Secondly, there are numerous germline V genes in themouse and human genome (about one hundred each for the Hchain and the L chain) and, whilst they are very homologousin sequence within their related subgroups (>75% sequencesimilarity), they are also diverse and distinctly very different inDNA sequence (again, mainly in their CDR regions). The germ-line V genes of a homologous subset generate a non-randomWu-Kabat variability plot (11) which sh<Duld be produced onlyby direct antigen-binding selection acdng on an Ig protein het-erodimer.
Thirdly, germline V segments in mice and humans exhibita relatively low incidence of translation stop codons, with manyopen reading frames. There is a statistically significant deficit inthe observed frequency of stop codons within the V-codingregion from the frequency expected from a process of randompoint mntation (11, 48). This observation also applies to the
Immunoloj|k(il Raiewi 1 6 2 / 1 9 9 8 125
Blanden et al • Somatic sienature in eermline V eenes
300
20 40 60 80 100Amino acid position
100 200 300Nucleotide position
CDR1 CDR2
400
CDR1 CDR2 CDR3
20 40 60 80 100 120Amino acid position• ^
CDR1 CDR2
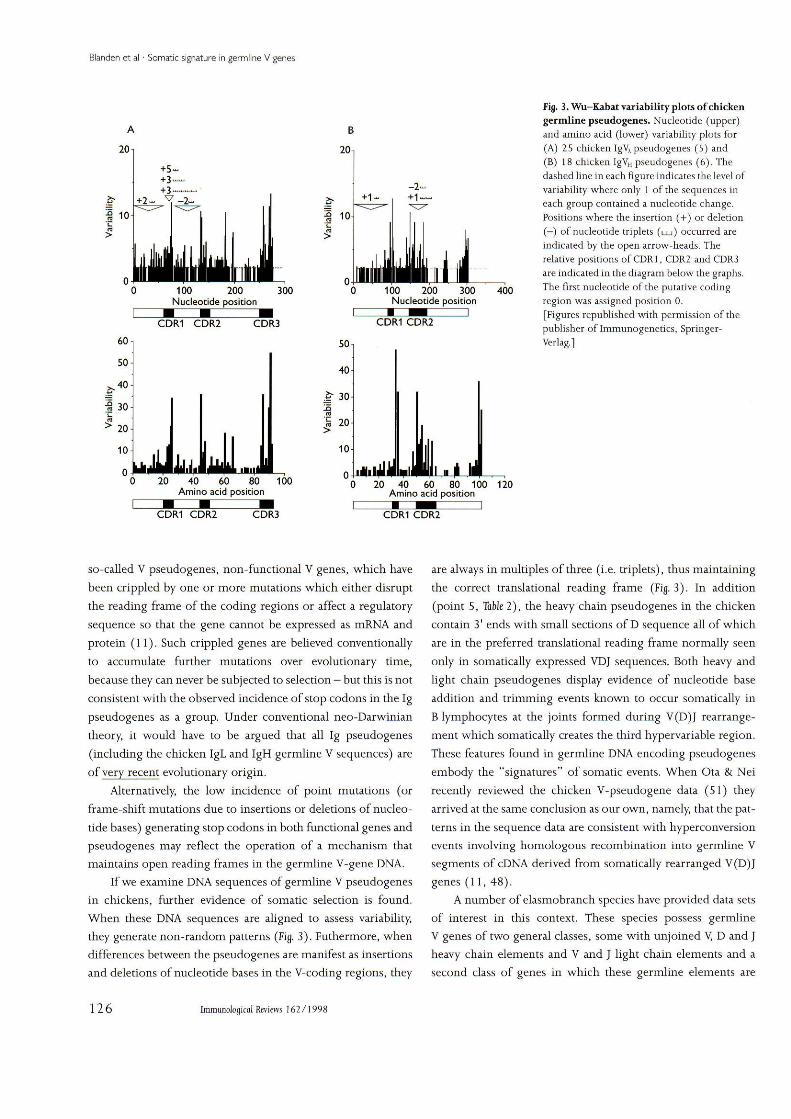
Fig. 3. Wu-Kabat variability plots of chickengermline psendogenes. Nueleotide (upper)and amino acid (lower) variability plots for(A) 25 chicken IgV^pseudogenes (5) and(B) 18 chicken IgVH pseudogenes (6). Thedashed line in each figure indicates the level ofvariability where only I of the seqnences ineach group contained a nucleotide change.Positions where the insertion (-I-) or deletion(—) of nucleotide triplets (u_j) occurred areindicated hy the open arrow-heads. Therelative positions of CDR 1. CDR2 and CDR3are indicated in the diagram below the graphs.The first nucleotide of the putative codingregion was assigned position 0.
[Figures republished widi permission of thepublisher of Ininiunogenetics, Springer-Verlag.]
so-called V psendogenes, non-functional V genes, \vhich havebeen crippled by one or more mutations which either disruptthe reading frame of the coding regions or affect a regnlatorysequence so that the gene cannot be expressed as rtiRNA andprotein (11). Such crippled genes are believed conventionallyto accumulate further mutations over evolutionary time,because they can never be subj ected to selection — but this is notconsistent with the ohserved incidence of stop codons in the Igpseudogenes as a group. Under conventional neo-Darwiniantheory, it would have to be argued that all Ig pseudogenes(including the chicken IgL and igH germline V sequences) areof very recent evolutionary origin.
Alternatively, the low incidence of point mutations (orframe-shift mutations due to insertions or deletions of nucleo-tide bases) generating stop codons in both functional genes andpseudogenes may reflect the operation of a mechanism thatmaintains open reading frames in the germline V-gene DNA.
If we examine DNA sequences of germline V pseudogenesin chickens, further evidence of somatic selection is found.When these DNA sequences are aligned to assess variability,they generate non-random patterns (Fig. 3). Futhermore, whendifferences hetv/een the pseudogenes are manifest as insertionsand deletions of nucleotide bases in the V-coding regions, they
are always in mnltiples of three (i.e. triplets), thus maintainingthe correct translational reading frame (Fig. 3). In addition(point 5, Table 2), the heavy chain psendogenes in the chickencontain 3' ends with small sections of D sequence all of whichare in the preferred translational reading frame normally seenonly in somatically expressed VDJ sequences. Both heavy andlight chain psendogenes display evidence of nncleotide baseaddition and trimming events known to occur somatically inB lymphocytes at the joints formed during V(D)J rearrange-ment which somatically creates the third hypervariable region.These features found in germline DNA encoding pseudogenesemhody the "signatures" of somatic events. When Ota & Neirecently review ed the chicken V-pseudogene data (51) theyarrived at the same conclusion as our own, namely, that the pat-terns in the sequence data are consistent with hyperconversionevents involving homologous recombination into germline Vsegments of cDNA derived from somatically rearranged V(D)Jgenes (11, 48).
A numher of elasmobranch species have provided data setsof interest in this context. These species possess germlineV genes of two general classes, some with unjoined V, D and Jheavy chain elements and V and J light chain elements and asecond class of genes in which these germline elements are
126 Ifflmunoiogicai Reviews 162/1998

Blanden et a! • Somatic signature in germline V genes
joined (52-56). There are two alternaiive explanations for theexistence of the joined genes. Either they represent a primordialform of V gene which predated separation of V, D and J ele-ments, or they have arisen by joining of previously separate ele-ments. In the latter case, a RT-based process producing cDNAcopies of pre-mRNA in B lymphocytes, followed by integrationof the entire V(D)J cDNA into germhne DNA, is a possiblemechanism for the origin of such a gene family.
The appearance of somatic patterns amongst a wide rangeof vertebrate germline V genes and V pseudogenes, particularlythe striking features of chicken pseudogenes described above,strongly suggests the operation of a somatic cell-to-germ cellgene feedback process active during the known 400 to 500million years of V-gene evolution. We have no way of measur-ing the frequency of such events over evolutionary time,although the strength of the somatic signattire apparent in thegermline DNA, differences in patterns in different species anddifferences in V-gene sequences among inbred mouse strains(II, 48) suggest that it is likely to he a frequent evolutionary
occurrence.
A response to Tomtinson and co-workers
In a receni communication, Tomlinson and colleagues com-pared patterns of somatic hypermutation established in anti-bodies raised in mice against small chemical haptens with pat-terns of variation in human germhne V genes (12). Since thesepatterns are different but complementary with respect to thelocation of amino acid changes in the antigen-combining siteof the Ig, they concluded that these data are incompatible withhomologous recombination between somatically derived DNAand germline V genes proposed by us. We disagree with theiranalysis. Firstly, experimental somatic hypermutation hasinvolved synthetic hapten molecules that could not have beeninvolved in the natural evolution of germline V genes. Untilmuch more data is accumulated on patterns of somatic hyper-mutation using protein epitopes that were involved in the evo-lution of adaptive immunity, their conclusion is premature.Indeed, as we have mentioned above, the work of Zinkernageland colleagties on mouse antibody responses to viral proteinantigens demonstrates that litde affinity maturation andsomatic hypermutatioti appears to occur in the generation ofsuch antibodies (23, 24). These results imply that the structuralrequirements (5 7) for antibodies with optimum affmity forprotein epitopes can be met by germline V genes plus ccjmbi-natorial diversity without the need for somatic mntation. Sec-ondly, Tomlinson et al. confmed their discussion to only one(possibly invalid) criterion in comparing somatic and germline
IgV sequences; they have not explained the ratige of featurespreviously published in Rothenfluh et al. (11) and expandedhere in Table 2.
Can we define a soma-to-germline "integration signature"for IgV genes?
The Somatic Selection model to explain the evolution of anti-body V genes proposes that new variant V sequences are"born" in the soma by hypermutation of an antigen-selectedV(D)J gene expressed in a B lymphocyte. Somatic hypermuta-tion and antigen-mediated selection in a germinal centre leadto the emergence of new, successful V(D)J DNA sequences withamino acid replacement mutations accumulating in the CDRregions and predominantly silent changes accumulating in FWregions. These new DNA sequences are then transported toreproductive tissues. Homologous recombination then takesplace to integrate the new gene into the chromosome, replac-ing a pre-existing germline V gene with a very similarsequetice.
If a somatic gene sequence recombined into germlineDNA, evidence of diis recombination should be detectable insome cases (point 4, Table 2). For example, imperfect joiningshould result in base insertions or deletions of variable lengthbordering the germline genes. In the same way that a non-ran-dom Wu-Kabat variabihty plot is indicative of direct antigen-binding selection, a non-random distribution of insertion-deletion events would be consistent with the operation of asoma-to-germline feedback loop. Such evidence exists in allrelevant vertebrate germline V-gene families where sufTicientdata are available to analyse (11, 48). This non-random distri-bution of putative recombination errors fits precisely with thepredictions of the soma-to-germline model.
The Weiller algorithm and the precise definition of the"recombination signature" of germline V segments
Recendy we employed a more precise method to define recom-bination sites in germline V genes.
The presence of the leader intron in germline V segmentsleads to clear predictions based on the RT model of somatichypermutation of rearranged V(D)J seqnences in B lympho-cytes producing various cDNA species, depending upon thetiming of intervention of splicing of the pre-mRNA template.Four species of cDNA molecules could be produced, three ofwdiich are relevant in the present contexi (see Fig. 4). Firstly,cDNA production may proceed unchecked to the 5' end of thepre-mRNA template (the Cap site). Secondly, cDNA production
Reviews 162/1998 127

Blanden et al • Somatic signature in germline V genes
Fig, 4. Predicted species of cDNA donors for soma-to-germline geneconversion. Esseiiiial features of the hypothetical nieLlianisra forhypermutaiion of rearranged VDI sequences in B lymphocytes. Thechromosomal DNA is shown with non-essential details omitted for clarity(e,g. split leader exons, possible additional J coding sequences in the J-Cintron and details of C-coding regions). Relevant coding regions arcindicated in hoxes: L - leader, VDI - variable-diversity-]oining region;;.The promoter (P) and the intronic enhancer/matrix attachment region(Ei/MAR) are also shown. The transcription start site (TSS) is indicated.The pre-mRNA is proposed to be a template for the production of cDNAby a reverse transcripcase (RT) complex designated the "mutatorsome"whieh binds to locus-specific secondary structure within the Ei/MARregion. Priming for initiation of reverse transcription is postulated looccnr at a number of different sites within the I-C intron as indicated bythe dotted line at the beginning of the cDNA strand. Depending upon thetiming of cDNA synthe.sis relative Co the intervention of splicing, differentspecies of cDNA molecules with different termination points will beproduced as described in the text.[Figure republished with the perinission of the publisher of Immunologyand Cell Biology, Blaekwell Science Asia,]
may extend to a point near the 5'-splice border of the leaderintron hnt be terminated there because of binding of (or cleav-age by) the spliceosome to the pre-mRNA template. Thirdly,cDNA synthesis may proceed through the V(D)J coding regionbut then terminate at the npstreani honndary of the V segment(3' border of the leader intron) because of the intervention oflariat formation and splicing at that point. Fourthly, cDNA mayterminate at the upstream boundary of the J-C intron if spliceo-some binding intervenes at that point. The first three of thesehypothetical cDNA species will provide sequences with highhomology to parts of the tandem array of germline V segmentsand with the potential for homologons recomhinaticni in thesegermline sites. The fourth is not relevant to the generation ofvariation (jr the maintenance of open reading frames in germ-line V coding elements. Because of possible degradation of the5' ends of tbe cut pre-mRNA by 5'-to-3' exonuclease activity,the cDNAs may not terminate precisely at the points describedabove,
Weiiler (58) has developed a novel algorithm to detectrecombination sites in gene sequences. Testing of the algorithmusing knowai reconihination points in related sets of bacterialDNA sequences (58) shows that it can precisely locate the sitesof recombination in collections of homologous gene
o o
seqnences. This "phylogenetic profile" method graphicallyrepresents the coherence of the phykDgenetic signal over theentire length of a mnltiple homologous sequence alignment.The PhylPro computer program (59) has been devised whichimplements the algorithm. We have used the phylogenetic pro-file method (the Weiller algorithm) to detect genetic recombi-nation in germhne V element DNA sequences (37).
The method introduces the "phylogenetic correlation"measure, which quantifies the coherence of the sequence inter-relationships in two different regions of an alignment involvingmnltiple homologous seqnences. The phylogenetic correlati<niof any given position is determined by evaluating sequencerelationships of regions immediately upstream and down-stream of this positi(ni. Positions where the sequence relation-ships of the upstream region differ significandy frotn theirdownstream connterparts are likely recombination sites; suchpositions will exhihit low phylogenetic correlations (see Fig. 5).The phylogenetic correlations are computed at every positionusing a sliding window technique for each individual sequencein the alignment. The phylogenetic correlation index can beplotted against the sequence position giving a phylogeneticprofile for that sequence in that collection. The profiles of allindividual sequences are typically superimposed in a single dia-gram which may also include a mean profile representing theaverage phylogenetic correlations of all sequences (the dark-ened line in Fig. 5). Such profiles indicate the major sites ofgenetic recomhination in that collection of homologoussequences. The values of the phylogenetic correlation index arelimited to values between -1-1.0 and -1,0, with high positivevalues representing a high phylogenetic correlation (and there-fore indicative of little or no recombination) and values dip-ping to around 0 representing uncorrelated relationships thatindicate sites of genetic recombination.
Analysis is presented here of DNA seqnences of humangermline IgV,, gene segments isolated and sequeiiced by Zachauand colleagues (60) using conventional phage and plasmidcloning techniques. Fig. 5 shows the analysis of 49 homologousgenes of V suhgroup I. The three pred<Dniinant recombinationsites in these genes approximate the 5' boundary of the leaderintron, the 5' boundary of the V-coding region extending intothe first framework region (Fl) and the Cap site. These resultsare in very good agreement with predictions hased on thesoma-to-germline model with the qualification that some exo-nuclease-mediated trimming of pre-mRNA templates may haveoccnrred after the intervention of the spliceosome. We are notaware of alternative explanations as to why recombinationshould occur predominantly at the sites shown in Fig. 5 (seeWeiller et al. (37) for further analysis and discussion). There-
128 162/1998
Blanden et al • Somatic signature in germline V genes
300 400
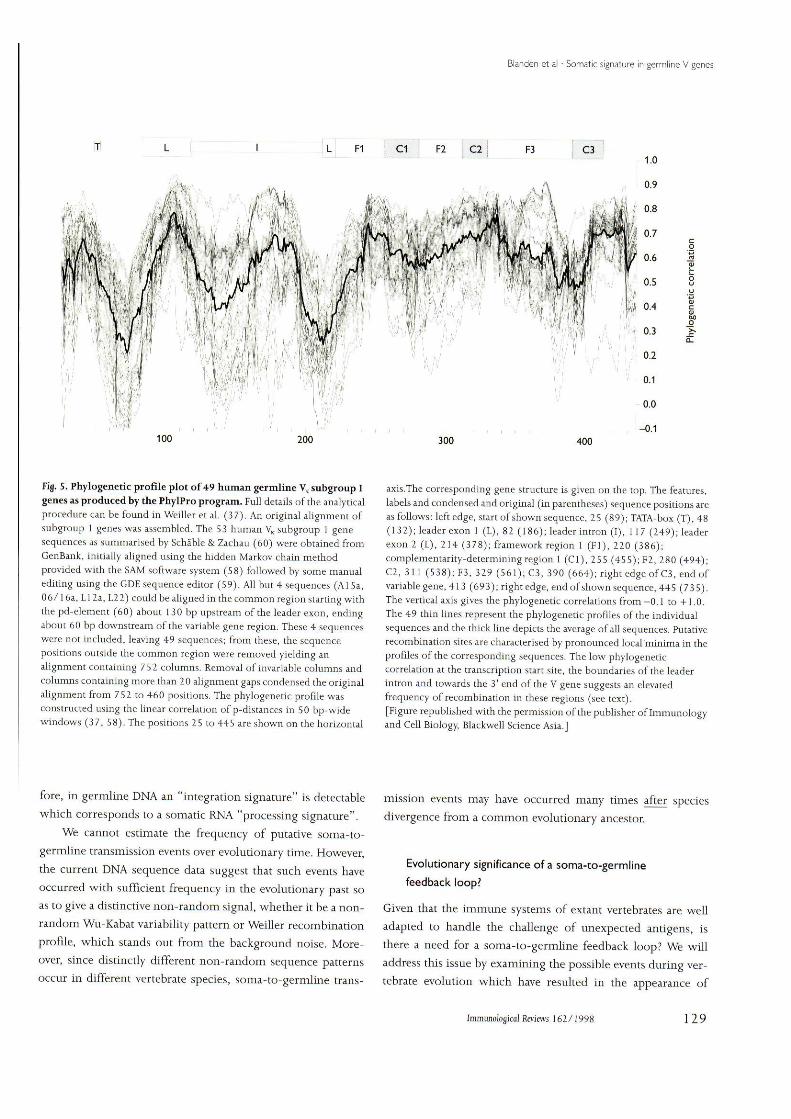
Fig. 5. Phylogenetic profile plot of 49 hnman germline V subgronp Igenes as produced by the PhylPro program. Full details of the analyticalprocedure can be found in Weiller et al. (37). An original alignment ofsubgroup 1 genes was assembled. The 53 human V subgroup 1 genesequences as summarised by Schable & Zachau (60) were obtained fromGenBank, initially aligned using tbe hidden Markov chain methodprovided with the SAM software system (58) followed by some manualediting using the GDE sequence editor (59), All but 4 sequences (Al 5a,06/16a, LI 2a, L22) could be aligned in the common region starting withthe pd-element (60) about 1 30 bp upstream of the leader exon, endingabout 60 bp downstream of the variable gene region. These 4 sequenceswere not included, leaving 49 sequences; from these, the sequencepositions outside the common region were removed yielding analignment containing 752 columns. Removal of invariable columns andcolumns containing more than 20 alignment gaps condensed the originalalignment from 752 to 460 positions. The phylogenetic profile wasconstructed using the linear correlation of p-distances in 50 bp-widewindows (37, 58). The positions 25 to 445 are shown on the horizontal
axis.The corresponding gene structure is given on the top. The features,labels and condensed and original (in parentheses) sequence positions areas follows: left edge, start of shown sequence, 25 (89); TATA-box (T), 48(132); leader exon 1 (L), 82 (186); leader intron (I), 117 (249); leaderexon 2 (L), 214 (378); framework region 1 (Fl), 220 (386);completnentarity-determiningregion 1 (Cl), 255 (455); F2, 280 (494);C2, 311 (538); F3, 329 (561); C3, 390 (664); right edge of C3, end ofvariable gene, 413 (693); right edge, end of shown sequence, 445 (735).The vertical axis gives the phylogenetic correlations from -0.1 to +1.0.The 49 thin lines represent the phylogenetic profiles of the individualsequences and the thick line depicts the average of all sequences. Putativerecombination sites are characterised by pronounced local minima in theprofiles of tbe corresponding sequences. The low phylogeneticcorrelation at the transcription start site, the boundaries of the leaderintron and towards the 3' end of the V gene suggests an elevatedfrequency of recombination in these regions (see text).[Figure republished with tbe permission of the pnbli.sher of Immunologyand Cell Biology, Blackweli Science Asia.]
fore, in germline DNA an "integration signature" is detectablewhich corresponds to a somatic RNA "processing signature".
We cannot estimate the frequency of putative soma-to-germline transmission events over evohidonary time. However,the current DNA sequence data snggest that snch events haveoccurred with sufficient frequency in the evolutionary past soas to give a distinctive non-random signal, whether it be a non-random Wu-Kabat variability pattern or Weiller recombinationprofile, which stands out from the background noise. More-over, since distinctly different noti-random sequence patternsoccur in different vertebrate species, soma-to-germline trans-
mission events may have occnrred tnany times after speciesdivergence from a common evolutionary ancestor.
Evolutionary significance of a soma-to-germlinefeedback loop?
Given that the immune systems of extant vertebrates are welladapted to handle the challenge of nnexpected antigens, isthere a need for a soma-to-germline feedback loop? We willaddress this issue by examining the possible events during ver-tebrate evolution which have resulted in the appearance of
ImmunoiogicQi Revims 162/1998 129

Blanden et al • Somatic signature in germline V genes
immunoglobulin genes. A minimal model for evolutionarydevelopment of a tandem array of germline IgV genes is givenbelow.1) The first step would possibly have been the emergence of a
very small set of germline antigen-recognition variablegenes in a vertebrate during ihe Cambrian explosiciii about550-500 million years ago. Selection pressure would favoursuccessive duplications of a single primordial gene (bytinequal crossing over) followed by mutational change inthe DNA sequences of the duplicates, resnlting in a tandemarray of closely related, thongh different, genes.
2) There would also be intense selection pressure on such avertebrate to somatically hypermutale its V genes to rapidlygenerate an antigen-recognition repertoire (to cope withinfections diseases) larger than that provided by the small,slowly mutating germline set.
3) Tbere would be continued selection pressure lo furtherexpand the size of the germline V-gene repertoire. Randomgermline mntation followed by Darwinian whole-animal"natural selection" would be a very slow way of building arepertoire. Furthermore, as we have already discussed (11),the evolution of randomly formed heterodimeric Ig bind-ing sites together with segmentation of the germline genes(into \ D, J segments) necessitating somatic rearrangementof the DNA sequence to produce cDNA and allow antibodybe expressed, markedly blxints whole-animal selection ofindividual germhne V segments. Moreover, each change ingermhne V-gene structure would require the generation ofa new repertoire of germline genes. In these circumstances,scmia-to-germline feedback of successful, functional,mutated V-gene sequences would confer great selectiveadvantage. Finally, if random mutation followed by whole-animal selection of germline genes for function were themechanism, it would favour single chain antihodies and no
DNA rearrangement. Thus, soma-to-germline feedbackmay have been an integral part of the evolutionary emer-gence of both H+L heterodimers and rearrangement ofvariable V(D)J genes.
However, is somatic mutation per se necessary in contemporaryvertebrates? Extensive somatic hypermutation can certainly bedemonstrated experimentally in antihapten responses. But theexperiments reported by Zinkernagel and co-workers usingviruses suggest that somatic hypermutation either does notoccnr in antiviral responses or does not confer a functionaladvantage on the antiviral immune response of the mouse (23,24). Therefore, somatic hypermutation may be redundant inmouse and human immune responses; there is sufficient germ-line diversity and combinatorial diversity of genetic elementsencoding heavy and light chains of antibodies to provide therapid somatic generation of a large repertoire sufficient to pro-tect against novel infectious agents.
Why, then, has such a sophisticated cellular and molecnlarprocess resulting in somatic hypermutation and selection ofV(D)J genes been retained in extant vertebrates? It may stillccmfer selective advantage because of its generative role as asource of tested, successful, new IgV open reading frames to befed back to the germiine. Such a RT-based soma-to-germlinefeedback loop would act to reduce the potentially cripplingeffects on the germline V-gene repertoire of random geneticdrift which may introduce translational stop codons inio cod-ing regions via point mutations or insertion/deletions ofnucleotides.
The soma-to-germline feedback loop seems to us toremain consistent with experimental evidence in immunologywhich has emerged in the 20-year period since the originalproposal (1). Extension of the analysis outside the immune sys-tem is heyond the scope of this review and will be publishedelsewhere (61).
ReferencesSteele EJ. Somatic seleciion and adaptiveevolution; on the inheritance of acquiredcharacters. Toronto: Williams and Wallace;1979; 2uded. Chicago: University of ChicagoPress; 1981.Steele EJ, Pollard JW Hypothesis: somatichypermutatioii by gene conversion via theerror prone DNA—>RNA—»DNA informationloop.Mol Immunol 1987;24:667-673.Steele FJ, Rolhenflnh HS, Blandeu RVMechanism of auiigeu-driven somatichypermutation of rearrangedimmunc)globulin V(D)J genes in the mouse.hiimuuol Cell Biol l997;7S:82-95.
4. Thompson CB, Neiman PE. Somaticdiversification of the chickenimmunoglobulin light chain gene is limitedto the rearranged variable gene segment.Cdl l987;48:369-378.
5. Reyiiaud C-A, Anquez V, Grimal H. Weill J-C,A hyperconversion mechanism generates thecliicken light chain preimmune repertoire.Cell 1987;48;379~388.
6. Reynaud C-A. Dahan A, Anquez V, Weill J-C.Somatic hyperconversion diversifies thesingle VH gene of che chicken with a highincidence in the D region.Cell 1989;59:171-183.
7. McCormack WT, Thompson CB. Chicken IgLvariable gene conversion display pseudogenedonor preference and 5' to 3' polarity.Genes Dev 1990;4:548-SS8.
B. McCormack WT, Hurley EA, Thompson CB.Germ line maintenance of the pseudogenedonor pool for somatic immunoglobulingene conversion in ciiickens.MolCellBiol 1993;13:8Zl-830.
9. Knight KL. Restricted VH gene usage andgeneration of antibody diversity in rabbit.Amiu Rev Immunol 1992; 10:593-616.
130 Inimunologicai Reiiews 162/1998

Blanden et al • Somatic signature in germline V genes
10. Laimiiig DK, Knight KL, Sonidtkhypernuiration, Muiations 3' of'rahhit VDJH-chain ^ciies.] Immunol 1 997;159:4403-4407.
I I. Roihcnfluh HS, Blaiiden RV, Steelc EJ,Evolution ot V genes: DNA sequencestrticturc of functional germ-line gems andpseudogenes.Immunogenetics 1995;42:1 59-1 7 1,
12. Tomlinson IM, Walter G, Jones PT, Dear PH,Sonnhammer ELL, Winicr G. The imprint ofsomatic; hypermutatioii on the repertoire ofhuman germline V genes.JMolBiol 1996:256:813-817.
1 3. Steele FJ, Rothcnfluh HS, Ada GL, Blanden RV,Affinity maturation of lymphocyte receptorsand positive selection of T cells in the ihymu;Immunol Rev 1 993;13S:S-49.
14. MacLcnnan ICM, Gray D. Antigen-drivenselection of virgin and memory B cells.Immunol Rev 1986;91:6 1-85.
I 5, Bcrck C. Milstein C. MutatioTi drifl andrepertoire shift in the mattiraiion of theimmune response.Immunol Rev 1987:96:23-41.
I 6, Jacob J, Kelsoe G, Rajewsky K, Weiss U.Intraclona! generation of antibody mutants ingerminal centres.Nature 1991:354:389-392.
17. Berek C, Berger A, Apel M, Maturation of theimmune response in germinal centers.Cell 199I;67:1121-1129,
1 8. MacLennan I. The centre of liypermuiaiion.Nature 1991;354:3S2-3.S3.
19. MacLennan ICM, Lui Y-J, Johnson GD.Maturation and dispersal of B-cell clonesduring T cell-dependent antibody responsesImmtinolRev 1992:126:143-161.
20. Chen C, Roberts VA, Rittenberg MB,Generation and analysis of random pointmutations in an antibody CDR2 sequence:many mutaced antibodies lose their ability tobind antigen.JExp Med 1992:176:8 5 5-866.
21. Blanden RV, Steele EJ, A Linifying hypothesisfor the molecular mechanism of somaticmutation and gene conversion in rearrangedimmunoglobulin variable genes.Immunol Cell Biol 1998;76 (In press).
22. Wii T, Kabat EA. An analysis of the sequencesof the variable regions of Bence Jones proteinsand myelome light chains and theirimplications for antibody complementarity,JExp Mod 1970; 132:21 1-250.
23. Roost H-R et al. Early high-affmityneutralizing ajiti-viral IgG responses withoutfurther improvements of affmityProc Nad Acad Sci USA 199S:92:1 2S7-1 261.
24. Bachniann MF, et al. The role of antibodyconcentration and avidity in antiviralprotection.Science 1997:276:2024-2027.
IS. Parham P A boost to immunity from nursesharks.CurrBiol 1995:5:696-699.
26. Greenbcrg AS, Avila D, Hughes M, Hughes A,McKJnney EC, Hajnik MF. A nev^ antigenreceptor gene family that undergoesrearrangement and extensive somaticdiversification in sharks.Nature 1995:374:168-173.
27. Hinds-Frey KR, Nishikata H, Litman RT,Litman GW SoT7iatic variation precedesextensive diversification of germlinesequences and combinatorial joining in theevolution of iTnmunoglobulin heavy chaindiversity.JExp Med 1993:178:825-834.
28. Reanney DC. Genetic error and genomedesign.Trends Genet 1986;2:4l-46.
29. Steele EJ, PollardJ\y Taylor L, Both GWEvaluation of possible mutator mechanismsactive on mammalian variable region genes.In: Steele EJ, ed. Somatic hypermutation inV-regions. Boca Raton (EL): CRC Press: 1991.p. 137-148,
30. Peters A, Storb U. Somatic hypermutation ofimmunoglobulin genes is linked totranscription initiation.Immunity 1996;4:l-9.
3 I. Kunkcl TA. Hypermutation during DNAsythesis in vitro. In: Steele EJ, ed. Somatichypertnutation in V-regions. Boca Raton(FL): CRC Press: 1991. p. 159-1 78,
32. McKean D, Huppi K, Bell M, Staudt L,Weigert M, Generation of antibody diversityin the immune response of BALB/c mice toinfluenza virus hemagglutinin.Proc Nad Acad Sci USA 1984:81:3 1 80-3 184.
33. Steeie EJ, Rothenfluh HS, Both GW Definingthe nucleic acid substrate for somatichypermutation.Immunol Cell Biol 1992:70:129-144,
34. Tumas-Brundage KM, Manser T. Thetranscription a I promoter regulateshypermntation on the antibody heavy chainlocus.JExp Med 1997:185:239-250.
3 5. Rada C, Yelamos J, Dean V , Milstein C. The 5'hypermutation boundary of K chains isindependent of local and neighbouringsequences and related to the distance from theinitiation of transcription.Eur J Immunol 1997;27:3 11 5-3 1 20.
36. Steele EJ. Mechanism of directionalmutations?MolReprodDev 1989:25:231-232.
3 7. Weillor GF, et al. Recombination signature ofgermline immunoglobulin variable genes.Immunol Cell Biol 1998:76 (In press).
3 8. Blackburn EH. Telomerascs,Annu Rev Biochem 1 992:61:11 3-1 29.
39. Umar A, Schweitzer PA, Levy NS. Gearhart ID,Gearhart PJ. Muiatlon in a reporter genedepends on proximity to and transcription ofimmunoglobulin variable transgenes.Proc Natl Acad Sci USA 1 99 I ;88:4902-4906.
40. Umar A, Gearhart PJ. Reciprocal homologousrecombinion ni or near antibody VDJ.Eur J Immunol 1995:25:2392-2400.
41. Xu B, Seising E. Analysis of sequence transfersresembhng gene conversion In a mouseantibody trans gene.Science 1994:265:1590-1593.
42. Chen C, Roherts VA, Stevens S, Brown M,Stenzel-Poore MP, Rittenberg MB.Enhancement and destruction of antibodyfunction by somatic mutation: unequaloccurrence is controlled by V genecombinatorial associations.EMBOJ 1995:14:2784-2794.
43. Liu Y-J. Sites of B lymphocyte selection,activation, tolerance in spleen.JExp Med 1997:186:625-629,
44. McCormack WT, Thompson CB, Somaticdiversification of the chickenimmunoglobiJin light-chain gene,Advlmmiuiol 1990;48:4i-67,
45. Arakavi'a H, Furusawa S, Ekino S,Yamagishi H. Immunoglobulin genehyper conversion ongoing in the chickensplenic germinal centers.EMBOJ 1996:15:2540-2546.
46. Carlson LM, McCormack WT, Postenia CE,Humphries EH, Thompson CB. Templatedinsertions in the rearranged chicken IgLV gene segment arise hy intrachromosomalgene conversion.Genes Dev 1990;4:536-547,
47. Rothenfluh HS, Steele EJ, Origin andmaintenance of germline V genes.Immunol Cell Biol 1993:71:227-232.[Pubhshed corrigendum appears in ImmunolCell Biol 1994;72:102J.
48. Rothenlluh HS. Gibbs AJ, Blanden R, Steele EJ,Analysis of patterns of DNA sequencevariation in flanking and coding regions ofmurine germline immunoglobulin variablegenes - evolutionary imphcations.Proc Nad Acad Sci USA1994;91:l2163-i2167.
49. Rothenfluh HS. Hypothesis: a memoryl)iiiphocyte-specificsoma-to-germlinegenetic feedback loop,Immunol Cell Biol 1 99 5;73:1 74-180.
50. Cox JPL, Tomlinson IM, Winter GA,A directory of human germline Vic segmentsreveals a strong bias in their usage,Eur J Immunol 1994:24:827-836.
5 1. Ota T, Nei M, Evolution of immunoglobulinVH pseudogenes in chickens,MolBiol Evol 1995;12:94-1 02.
162/1998 131

Blatiden et al • Somatic signature in getmline V genes
5 2. KokubuE, Litman R, Shamblott MJ, Hinds K,Litman GW Diverse organization ofimmtuioglobulin VH gene loci in a primitivevertebrate.EMBOJ 1988;7:3413-3422,
53. Harding FA, Cohen N, Utman GW,Immunoglobulin heavy chain geneorganization and complexity in the skate, Rajaerinacea.Nucleic Acids Res 1990;18:1015-1020.
54. Litman GW; Haire RN. Hhids KR,Amemiya CT, Rast JP, Hulst MA. Evolutionarydevelopment ofthe B ceU repertoire,AnnNYAcadSci 1992:651:360-368.
55. Rast JP, et al. Immunoglobulin light chainclass multiplicity and alternativeorganizational forms in early vertebrateph>'logeiiy.Immimogenetics 1994:40:83-99.
56. Hohman VS. Schuchman, DB, Schluter SE,Marchalonis jj. Genomic clone for thesandbar shark lambda light chain: generationof diversity in tJie absence of generearrangement.Proc Natl Acad Sci USA 1993:90:9882-9886.
57. Wedemayer GJ, Pattern PA, Wang LH,Schultze PG, Stevens RC. Structural insightsinto the evoJution of an antibody combiiiijigsite.Science i 997:276: J 665-1 669.
58. Weiiler GE. PJiylogenetic profiles: a graphicalmethod for detecting genetic recombinationsin homologous sequences.MolBiol Evol 199S;15i326-335.
59. Weiiler GF. Australian National University,Canberra, ACT; 1997 (Computer program),
60. Schable KE, Zachau H-G, The variable genes ofthe human immunoglobulin K locus.Biol Chem Hoppe Seyler1993:374:1001-1022.
6 1. Steele EJ, Lfndley RA, Blanden RV Lamarck'ssignature: how retrogenes are changingDarwin's natural selection paradigm. Sydney:Allen & Unwin: 1998. Reading (MT):Addison Wesley Longman; 1998.
62. Chien NC. Pollock RR, Desaymard C.Scharff MD. Point mutations cause ihesomatic diversification of IgM and IgG2aanti-phosphorylcholine antibodies.JExp Med 1988:167:954-973.
6 3. Gonzalez-Eernandez A, Milstein C. Analysis ofsotiiatic hypermutation in motise Peyer'spatches using immunoglobuliu K light-chaintransgenes,Proc Natl Acad Sci USA 1993:90:9862-9866,
64, Betz AG, Rada C, Pannell R. Milstein C,Neuberger MS. Passenger transgenes revealintrinsic specificity of the antibodyhypermutaiion mechanism: clustering,polarity, specific hotspots.Proc Natl Acad Sci USA l993;90:2385-2388.
65. Lebecque SG, Gearhart PJ. Boundaries ofsomatic mutation in rearrangedimmunoglobtihn genes: 5' boundary is nearthe promoter, 3' boundary is approximately Ikb from V-D-J gene.JExp Med 1990:172:1717-1727.
66. Weber JS, Berry J, Manser T, ClallinJL,Position ofthe rearranged VK and its 5'flanking sequences determines the location ofsomatic mtitations in the JK locus.JImmunol 1991:146:3652-3655.
67, Boih GW, Taylor L, Pollard JW, Steeie EJ.Distribution of mutations around rearrangedheavy-chain antibody variable-region genes.Mol Cell Biol 1990:10:5187-5196.
68. Rothenfluh HS, Taylor L, Bothwell ALM,Both GV Steele EJ. Somatic hypermutation in5' flanking regions of heavy chain antibodyvariable genes.EurJImmunol 1993;23:2152-2159.
69, Rada C, Gonzalez-Fernandez A, Jarvis JM,Milstein C. The 5' boutidary of somatichypermutation in a VK gene is in the leaderintron.EurJImmunol 1994:24:14-53-1457.
70. Rogerson BJ. Mapping the upstreamboundary of somatic mutations in rearrangedimmunoglobulin traiiigenes and endogenousgenes.Mol Immtmol 1994:31:83-98,
71. Motoyama N, Okada H, Azuma T. Somaticmutation in constant regions of mouse X]liglit cliains.Proc Natl Acad Sci USA 1991:88:7933-7937.
72. Azuma T, Moioyama N, Eields LH, Loll DY.Mutations ofthe chloramphenicol acetyltransferase transgene driven by theimmunoglobulin promoter and intronenhancerhit Immtmol 1993:5:121-130.
73. Yelamos), ec al. Targeting of lion-Igsequences in place of the V segment bysotnatic hypermutation.Namre 1995:376:225-229,
74. Beiz AG, Milstein C, Gonzalez-Fernandez A,Pannell R, Larson T, Neuberger MS. Elementsregulating somatic hypermutatioii of animmunoglobulin K gene: critical role of theinlron eiihancer7inatrix attachment region.Cell 1994:77:239-248,
75. Sohii ], Gerstein RM, Hsieh C-L, Lemer M,Seising E, Somatic hypermutation of animmunoglobulin u heavy chain transgene.j Exp Med 1993:177:493-504.
76. Giusti .AM, Manser T. Hypermutation isobserved only in antibody H chain V regiontransgenes that have recombined vi'ithendogenous immunoglobulin H DNA:implications for the location of ds-aciingelements required for somatic mutation.JExp Med 1993,177:797-809.
77. Hackett J, Stebbins C, Rogerson B, Davis MM,Storb U. Analysis of a T cell receptor gene as atarget of the somatic hypermutationalmechanism.JExp Med 1992:176:225-23!.
132 immunologicQl Reviews 162/1998

Related Documents











