The Semicircular Canal System and Locomotion: The Case of Extinct Lemuroids and Lorisoids ALAN WALKER, TIMOTHY M. RYAN, MARY T. SILCOX, ELWYN L. SIMONS, AND FRED SPOOR Paleontologists reconstruct the locomotor and postural behavior of extinct species by analogy with living forms and biomechanical analyses. In rare cases, behavioral evidence such as footprints can be used to confirm fossil-based reconstructions for predominantly terrestrial orders of mammals. For instance, the chalicothere prints from Laetoli show that these perissodactyls supported their body weight on the metacarpals, as previously reconstructed. 1 Unfortu- nately, primates are mostly arboreal and rarely leave footprints. The cercopithe- cid and hominin prints at Laetoli are a rare exception. We have recently shown that the semicircular canal system can be used to test and augment locomotor reconstructions based on postcranial material or to provide first estimations of locomotor behavior for taxa not known from the postcranium. Using a sample of modern primates, we have been able to demonstrate that the radii of curvature of the semicircular canals are significantly correlated with both body mass and agility of locomotion. 2 This paper reviews those results and examines the rela- tionship between semicircular canal morphology and other evidence in efforts to reconstruct locomotor behavior in subfossil lemurs from the Holocene of Mada- gascar and fossil lorisoids from the Miocene of Africa. The semicircular canal system of the vertebrate inner ear provides neural input for the neck and intrin- sic eye muscle reflexes. These reflexes keep the head steady during locomotion and so stabilize gaze and help integrate the optic flow of reti- nal images. Canals, the bony tubes in the otic capsule, surround three membranous ducts that are part of the endolymph circuit of the inner ear. The term 1 ‘‘semicircular canal system’’ covers the entire functional unit, including both bony and soft- tissue aspects (Box 1). The correlation between semicircu- lar canal dimensions and body mass has been investigated, 3–5 demonstrat- ing that the canals increase in size with strong negative allometry (that is, smaller animals have relatively larger canals). Theoretical functional models of the canal system predicted that double logarithmic plots of canal size against body mass should have slopes between 0.08 and 0.33. 3 Slopes determined for different ver- tebrate groups do fall within this range. 3,4,6 A sample of 174 nonceta- cean mammalian species gave a value of 0.14 for the slope of regres- sion of log mean radius of curvature of the canals on log body mass. 7 Past studies showed that valuable information about the agility or jerki- ness of locomotion is also present in the plots of log canal size against log body mass. It appeared that the size of the canals reflects some quality of an animal’s behavior. A.A. Gray 8 noted, as long ago as 1907, that sloths have very small canals for their body size and suggested that this correlated with their sluggish movements. The semicircular canals of fast flying birds were reported to be relatively larger than those in species with less aero- batic flight. 9–11 Limited comparative quantitative observations on primates showed that, once body size is accounted for, species that were acro- batic or that had very rapid locomo- tion clearly had larger canal arcs than did those that were cautious or slow in their movements. 4,12,13 These pre- liminary findings led us to study the relationship between canal arc size Alan Walker is Evan Pugh Professor of Anthropology and Biology at Penn State University. His 1967 PhD thesis concerned the locomation of the extinct Madagascan lemurs, a sub- ject to which he returns here. E-mail: [email protected] Timothy M. Ryan is a Research Associate in the Department of Anthropology and the Cen- ter for Quantitative Imaging at Penn State University. He is interested in primate locomotor evolution and the functional morphology of the postcranial skeleton. E-mail: [email protected] Mary T. Silcox is an Associate Professor at the University of Winnipeg. Her research focuses on the earliest phases of primate evolution. She is interested in elucidating the ev- olutionary and adaptive context of primate origins, in particular the stem primates referred to as ‘‘plesiadapiforms.’’ E-mail: [email protected] Elwyn L. Simons is James B. Duke Professor Emeritus of Biological Anthropology and Anatomy at Duke University. He is interested in primate evolution and has worked in the field in Egypt, India, Iran, Madagascar, Nepal, and Wyoming. E-mail: [email protected] Fred Spoor is Professor of Evolutionary Anatomy at University College London, United Kingdom, and affiliated with the Koobi Fora Research Project, Kenya. E-mail: [email protected] V V C 2008 Wiley-Liss, Inc. DOI 10.1002/evan.20165 Published online in Wiley InterScience (www.interscience.wiley.com). ARTICLES Evolutionary Anthropology 17:135–145 (2008)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Semicircular Canal System and Locomotion:The Case of Extinct Lemuroids and LorisoidsALAN WALKER, TIMOTHY M. RYAN, MARY T. SILCOX, ELWYN L. SIMONS, AND FRED SPOOR
Paleontologists reconstruct the locomotor and postural behavior of extinctspecies by analogy with living forms and biomechanical analyses. In rare cases,behavioral evidence such as footprints can be used to confirm fossil-basedreconstructions for predominantly terrestrial orders of mammals. For instance,the chalicothere prints from Laetoli show that these perissodactyls supportedtheir body weight on the metacarpals, as previously reconstructed.1 Unfortu-nately, primates are mostly arboreal and rarely leave footprints. The cercopithe-cid and hominin prints at Laetoli are a rare exception. We have recently shownthat the semicircular canal system can be used to test and augment locomotorreconstructions based on postcranial material or to provide first estimations oflocomotor behavior for taxa not known from the postcranium. Using a sample ofmodern primates, we have been able to demonstrate that the radii of curvatureof the semicircular canals are significantly correlated with both body mass andagility of locomotion.2 This paper reviews those results and examines the rela-tionship between semicircular canal morphology and other evidence in efforts toreconstruct locomotor behavior in subfossil lemurs from the Holocene of Mada-gascar and fossil lorisoids from the Miocene of Africa.
The semicircular canal system ofthe vertebrate inner ear providesneural input for the neck and intrin-sic eye muscle reflexes. Thesereflexes keep the head steady duringlocomotion and so stabilize gaze and
help integrate the optic flow of reti-nal images. Canals, the bony tubes inthe otic capsule, surround threemembranous ducts that are part ofthe endolymph circuit of the innerear. The term1 ‘‘semicircular canal
system’’ covers the entire functionalunit, including both bony and soft-tissue aspects (Box 1).The correlation between semicircu-
lar canal dimensions and body mass
has been investigated,3–5 demonstrat-
ing that the canals increase in size
with strong negative allometry (that
is, smaller animals have relatively
larger canals). Theoretical functional
models of the canal system predicted
that double logarithmic plots of
canal size against body mass should
have slopes between 0.08 and 0.33.3
Slopes determined for different ver-
tebrate groups do fall within this
range.3,4,6 A sample of 174 nonceta-
cean mammalian species gave a
value of 0.14 for the slope of regres-
sion of log mean radius of curvature
of the canals on log body mass.7
Past studies showed that valuableinformation about the agility or jerki-ness of locomotion is also present inthe plots of log canal size against logbody mass. It appeared that the size ofthe canals reflects some quality of ananimal’s behavior. A.A. Gray8 noted,as long ago as 1907, that sloths havevery small canals for their body sizeand suggested that this correlatedwith their sluggish movements. Thesemicircular canals of fast flying birdswere reported to be relatively largerthan those in species with less aero-batic flight.9–11 Limited comparativequantitative observations on primatesshowed that, once body size isaccounted for, species that were acro-batic or that had very rapid locomo-tion clearly had larger canal arcs thandid those that were cautious or slowin their movements.4,12,13 These pre-liminary findings led us to study therelationship between canal arc size
Alan Walker is Evan Pugh Professor of Anthropology and Biology at Penn State University.His 1967 PhD thesis concerned the locomation of the extinct Madagascan lemurs, a sub-ject to which he returns here. E-mail: [email protected] M. Ryan is a Research Associate in the Department of Anthropology and the Cen-ter for Quantitative Imaging at Penn State University. He is interested in primate locomotorevolution and the functional morphology of the postcranial skeleton.E-mail: [email protected] T. Silcox is an Associate Professor at the University of Winnipeg. Her researchfocuses on the earliest phases of primate evolution. She is interested in elucidating the ev-olutionary and adaptive context of primate origins, in particular the stem primates referredto as ‘‘plesiadapiforms.’’ E-mail: [email protected] L. Simons is James B. Duke Professor Emeritus of Biological Anthropology andAnatomy at Duke University. He is interested in primate evolution and has worked in thefield in Egypt, India, Iran, Madagascar, Nepal, and Wyoming. E-mail: [email protected] Spoor is Professor of Evolutionary Anatomy at University College London, UnitedKingdom, and affiliated with the Koobi Fora Research Project, Kenya.E-mail: [email protected]
VVC 2008 Wiley-Liss, Inc.DOI 10.1002/evan.20165Published online in Wiley InterScience (www.interscience.wiley.com).
ARTICLES
Evolutionary Anthropology 17:135–145 (2008)

and locomotion in a large compara-tive database of 210 mammal species,including 91 primates ranging in aver-age body mass from Microcebus rufus(43 g) to Gorilla gorilla (121 kg). Bodymasses were taken from the litera-ture.40 Canal dimensions were meas-ured using computed tomographydata following the method of Spoor
and Zonneveld.4 Six basic categoriesof the degree of agility of motion,from extremely slow to very jerky,were assessed through nonquantita-tive field observations.
The study by Spoor and co-workers,2 showed a strong relation-ship between semicircular canal sizeand jerkiness of locomotion in a vari-
ety of primate groups. As can be seenin Box 2 Figure 1, semicircular canalradius of curvature is positively cor-related with agility of locomotion inprimates. The leaping tarsiers andgalagos have large canals relative totheir body size, while the slow quad-rupedal lorises, although of similarbody size, lie on the lower end of the
Box 1. The Semicircular Canal System
The bony labyrinth inside the pet-rous part of the mammalian temporalbone houses the inner ear, includingthe organ of hearing and the senseorgans for the perception of move-ment and spatial orientation. The lat-ter are phylogenetically older. Alljawed vertebrates have inner earswith three membranous semicircularducts, anterior, posterior and lateral,which sense angular head motion.Each duct is connected with themembranous utricle inside the vesti-bule of the bony labyrinth, and thusforms a fluid circuit filled with endo-lymph (Fig. 1). The ampulla, a swollensection at one end of each duct, con-tains a gelatinous mass, the cupula,which seals the duct. Head rotationcauses the endolymph to lag behindbecause of inertia, and the resultingfluid pressure slightly deforms thecupula. This deformation is detectedby the cilia of hair cells embedded inthe cupula. Resulting changes in thefiring rate of cilia provides the outputthat encodes the head rotation. Thesignal that stimulates the semicircularcanal system is angular acceleration.Mechanically, however, the systeminstantly integrates this input and theneural output is directly proportionalto the angular velocity of head move-ments. The three semicircular ductsare oriented in approximately orthog-onal planes; any head rotation willthus be sensed by at least one duct.Moreover, the output from both earsis integrated to provide comprehen-sive representation of self-rotation.This information, combined with oto-lithic, visual, and proprioceptive input,helps coordinate posture and bodymovements during locomotion. Thebest understood function of the semi-circular canal system is its contribu-
tion to the stabilization of gaze duringlocomotion. This is accomplished viathe vestibulocollic and vestibuloocularreflexes that trigger compensatorymovements of the head and eyes bythe extraocular and neck muscles,respectively. Stabilization of vision isespecially important in birds and ar-boreal and/or gliding mammals, suchas most primates, dermopterans,scandentians, some marsupials, andmany rodents, that have to rely oneyesight when moving quicklythrough the air or trees.The basis for a functional relation-
ship between the semicircular canalsystem and locomotor repertoire of aspecies is that physical dimensions ofthe ducts influence the sensitivity andother mechanical response character-istics of the sensory system.36 The
latter are fine-tuned by morphologicalchanges to match the requirementsof particular locomotor behav-iors.2,7,14,37 Empirical studies investi-gating this functional relationshiphave mainly considered the bonysemicircular canals of extant and fos-sil species.2,7,14,37 Valid interpretationof these osteological data is possiblebecause each canal reliably reflectsthe functionally important length ofthe enclosed membranous duct(along its arc). However, among pri-mates in particular, a secondfunctionally important aspect of thecanal system, the lumen size (cross-section) of each duct, is more difficultto assess on the basis of the bonycanal because, to variabledegree,8,38,39 the duct occupies thecanal lumen (Fig. 1).
Figure 1. Lateral view of a left baboon inner ear after removal of the surrounding bonyotic capsule (modified after Grey8). On the left, the anterior (A), posterior (P), and lat-eral (L) semicircular canals and their enclosed membranous ducts are shown. Thecochlea, with the organ of hearing, is located at bottom left. On the right, the poste-rior canal has been isolated and stylized to demonstrate the principle morphology ofthe canal system. The duct, utricle (u), and ampulla with cupula (c), forming an endo-lymph circuit, is shown in black. The radius of curvature (R) quantifies the length of theendolymph circuit. Scale of 5 mm pertains to the left image.
136 Walker et al. ARTICLES

Box 2. Semicircular canals and Locomotor agility
The graphical results from analysisof the comparative database of pri-mates. The log transformed radiusof curvature of the three semicircularcanals are plotted against log trans-formed species mean body masses(data from Smith and Jungers40).
Different symbols represent each agil-ity category: red triangles, fast; graytriangles, medium fast; white boxes,medium; dark gray circles, mediumslow; blue circles, slow. Abbreviations:ASC-R, radius of anterior semicircularcanal; PSC-R, radius of posterior
semicircular canal; LSC-R, radius oflateral semicircular canal; SC-R, ra-dius of mean of the three semicircu-lar canals. See Table 1 for regressioncoefficients. Note agility categoriesare separated best by the lateralcanal plot.
TABLE 1. Results of PGLS analyses with log semicircular canal radius as dependent variable for extant strepsirhine
primates. Note that the association between canal radius and the log10 of agility is highly significant for all three
canals, and for their average radius (SCR)
Canal Variable Coefficient SE F df p r2 ln ML Likelihood
ASCR logBM 0.1905 0.0118 259.2092 1,25 <0.0001 0.9224 62.0743logAGILITY 0.1045 0.0387 7.2837 1,25 <0.05Y intercept 20.3208 0.0375
PSCR logBM 0.1734 0.0145 143.0360 1,25 <0.0001 0.8766 56.3145logAGILITY 0.1565 0.0474 10.8866 1,25 <0.01Y intercept 20.3532 0.0460
LSCR logBM 0.1337 0.0112 142.7350 1,25 <0.0001 0.8946 65.1933logAGILITY 0.2425 0.0388 39.0771 1,25 <0.0001Y intercept 20.3308 0.0380
SCR logBM 0.1669 0.0107 244.2745 1,25 <0.0001 0.9240 66.1217logAGILITY 0.1691 0.0357 22.3790 1,25 <0.0001Y intercept 20.3352 0.0347
Figure 1. See text above.
ARTICLES The Semicircular Canal System and Locomotion 137
distribution, with relatively smallcanals. At larger body masses, thisrelationship also holds. The acro-batic brachiating gibbons have rela-tively large canals for their body sizeas compared with the great apes.Within Old World monkeys, Colobuspolykomos has larger canals thandoes the similarly sized Procolobusbadius. The former frequently leapsbetween trees but the latter appearsto be a less skilled leaper, especiallyin landing (S. McGraw, personalcommunication), which is possiblywhy P. badius is more heavily pre-dated upon by chimpanzees acrossAfrica. Ateles geoffroyi, an acrobaticprehensile-tailed monkey, has largeanterior and posterior canals for itsbody size whereas the less agileAlouatta seniculus has smaller
canals. The radius of each individualcanal acts as part of an integratedfunctional system and does not nec-essarily impart the same informationabout head jerkiness. This maydepend on the planes of head motioninvolved and, perhaps, spatial con-traints on the otic capsule. For exam-ple, during hominin evolution onlythe anterior and posterior canalsenlarge with the emergence of mod-ern-human-like bipedal locomo-tion.14 In contrast, tarsiers and gala-gos on one hand and lorises on theother are most distinct in lateralcanal size. Likewise, the small lateralcanal of Alouatta seniculus is consist-ent with its less agile behavior. How-ever, its anterior canal appears unex-pectedly large, possibly as a conse-quence of spatial constraints on the
subarcuate fossa,15 which opens intothe endocranial cavity through thearc of the anterior canal and housesa lobule of the cerebellum.To allow for the most relevant
comparison to fossil and subfossilstrepsirhines, we have, for the cur-rent study, limited our statisticalanalysis to the living members ofthat group. The same pattern ofresults holds for this more restrictedsample, with more agile strepsirhinessuch as galagids having relativelylarger canals than do forms with lessjerky locomotion, such as lorisids(Box 2). Phylogeny can have a con-founding effect in regression analy-ses that include different species16;forms may appear similar becausethey are closely related, not becausethey share underlying functional
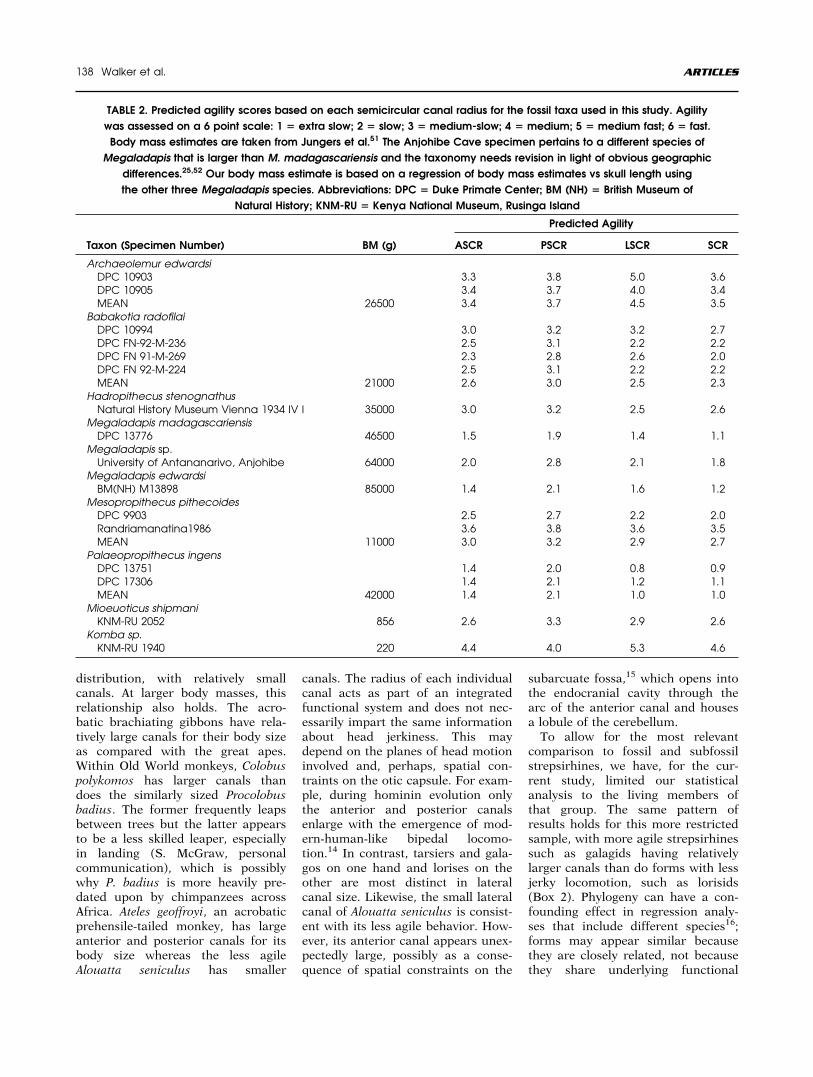
TABLE 2. Predicted agility scores based on each semicircular canal radius for the fossil taxa used in this study. Agility
was assessed on a 6 point scale: 1 5 extra slow; 2 5 slow; 3 5 medium-slow; 4 5 medium; 5 5 medium fast; 6 5 fast.
Body mass estimates are taken from Jungers et al.51 The Anjohibe Cave specimen pertains to a different species of
Megaladapis that is larger than M. madagascariensis and the taxonomy needs revision in light of obvious geographic
differences.25,52 Our body mass estimate is based on a regression of body mass estimates vs skull length using
the other three Megaladapis species. Abbreviations: DPC 5 Duke Primate Center; BM (NH) 5 British Museum of
Natural History; KNM-RU 5 Kenya National Museum, Rusinga Island
Taxon (Specimen Number) BM (g)
Predicted Agility
ASCR PSCR LSCR SCR
Archaeolemur edwardsiDPC 10903 3.3 3.8 5.0 3.6DPC 10905 3.4 3.7 4.0 3.4MEAN 26500 3.4 3.7 4.5 3.5
Babakotia radofilaiDPC 10994 3.0 3.2 3.2 2.7DPC FN-92-M-236 2.5 3.1 2.2 2.2DPC FN 91-M-269 2.3 2.8 2.6 2.0DPC FN 92-M-224 2.5 3.1 2.2 2.2MEAN 21000 2.6 3.0 2.5 2.3
Hadropithecus stenognathusNatural History Museum Vienna 1934 IV I 35000 3.0 3.2 2.5 2.6
Megaladapis madagascariensisDPC 13776 46500 1.5 1.9 1.4 1.1
Megaladapis sp.University of Antananarivo, Anjohibe 64000 2.0 2.8 2.1 1.8
Megaladapis edwardsiBM(NH) M13898 85000 1.4 2.1 1.6 1.2
Mesopropithecus pithecoidesDPC 9903 2.5 2.7 2.2 2.0Randriamanatina1986 3.6 3.8 3.6 3.5MEAN 11000 3.0 3.2 2.9 2.7
Palaeopropithecus ingensDPC 13751 1.4 2.0 0.8 0.9DPC 17306 1.4 2.1 1.2 1.1MEAN 42000 1.4 2.1 1.0 1.0
Mioeuoticus shipmaniKNM-RU 2052 856 2.6 3.3 2.9 2.6
Komba sp.KNM-RU 1940 220 4.4 4.0 5.3 4.6
138 Walker et al. ARTICLES

similarities. In this study, attemptshave been made to control for theseeffects by using Phylogenetic Gener-alized Least Squares (PGLS) (Box 3).The results of PGLS analyses assess-ing the correlation between semicir-cular canal size and both body massand agility confirm, in general, theresults of the uncorrected regressionanalysis, but with much better likeli-hoods.2 Box 1 Table 1 gives theresults from the PGLS analyses foreach semicircular canal for extantstrepsirhine primates. The same dataused in these analyses can be used toproduce predictive equations for agil-ity in extinct species; these are givenin Table 2 for the subfossil lemurs ofMadagascar and the fossil lorisoidsfrom the Miocene of Africa.
THE RECENTLY EXTINCTMADAGASCAN LEMUROIDS
Several genera of large lemuroidshave become extinct in the past twothousand years.17 Because these spe-cies have only become extinct veryrecently, they can be regarded aspart of the same ecological commu-nity as living lemurs. These subfos-sils have been collected for nearly a
century and a half. Many of the spe-cies are known from complete, ornearly complete, skeletons. The goodquality of preservation of the post-cranium in many of these formsmeans that much can be said abouttheir locomotion based on traditionalbiomechanical and comparativeanalyses; the most recent accounts oftheir postcranial adaptations arethose of Jungers, Godfrey, and col-leagues.18–25 Study of the dimensionsof the semicircular canals allows fora test of the conclusions from suchanalyses. These dimensions also pro-vide information of a somewhat dif-ferent type, in that they relate tojerkiness, and thus agility of locomo-tion. This is a variable that is at bestindirectly reflected in the postcra-nium, but that is critical to ananimal’s ability to escape quicklyfrom predators and to monopolizeresources.
Semicircular canals were studiedin eight species of subfossil lemurs(Table 2) belonging to three subfami-lies: Palaeopropithecinae, Megalada-pinae, and Archaeolemurinae. Somereconstructed bony labyrinths ofextant and extinct species are shownin Figure 1.
THE SLOTH LEMURS(PALAEOPROPITHECINAE)
The sloth lemurswere a remarkableradiation that showsconvergent adapta-tions on edentate
sloths.19,22,26,27 Weexamined the bony labyrinths ofthree genera, Mesopropithecus, Baba-kotia, and Palaeopropithecus, thepostcranial remains of which showprogressively more extreme adapta-tions to below-branch quadrupedalsuspensory behavior. The first isreconstructed as a slow-climbing lor-isid-like quadruped and the last as afully sloth-like suspensory form.Babakotia is reconstructed as inter-mediate in its suspensory behavior.These behaviors are reconstructedby, among other things, the increas-ingly high intermembral indices,increasingly curved phalanges, anddecreasing height of the greater tu-bercle of the humerus and thegreater trochanter of the femur.Many details of the postcranial skele-ton of Palaeopropithecus show re-markable convergence on the condi-
Figure 1. Reconstructed left bony labyrinths for selected extant and subfossil strepsirhines. Scale bar for each is 1 mm. Reconstructionsare shown in lateral view and are scaled for body mass based on the strepsirhine regression for the mean canal radius.
ARTICLES The Semicircular Canal System and Locomotion 139

tion in Bradypus and Choeloepus, thetwo genera of modern sloths.19,22
Animals more specialized for sloth-like quadrupedal suspensory locomo-tion would be predicted to have rela-tively smaller semicircular canals.The relative sizes of the semicircu-
lar canals of these species follow thepattern of jerkiness of locomotionpredicted from the postcranial recon-structions. Figure 2 shows the rela-tionship between body mass and thelateral semicircular canal diameter.The three sloth lemurs are situatedin order of the degree of their post-cranial suspensory adaptations, withMesopropithecus and Babakotia hav-ing quite small canals, so that theyare positioned like lorisids relative tothe general strepsirhine trend. Palae-opropithecus falls well below as anoutlier from the strepsirhine trend.Living sloths also fall well below thegeneral mammalian trend in thesame way as does Palaeopropithecus.2
An animal with semicircular canalsas small as those observed for Palae-opropithecus would be estimated toweigh only 302 gm, three-quarterssmaller than Cheirogaleus major. Theactual body mass for this animal isestimated from limb bones to bemore than two orders of magnitudelarger (42 kg). The predicted agility
scores for the sloth lemurs based onthe mean canal radius are in orderof reconstructed locomotion. M. pith-ecoides, 2.7 (medium-slow to slow);B. radofilai, 2.3 (medium-slow toslow); and P. ingens, 1.0 (extra slow;
Table 2). For comparison, living lori-sids were assigned agility scores of 2(slow) and living sloths scores of 1(extra slow).
THE KOALA LEMURS(MEGALADAPINAE)
Megaladapis specieshave body propor-tions that are some-what like those ofPhascolarctos ciner-eus, the marsupialkoala.25 They have
large skulls relative to the rest of thebody, very long forelimbs relative totheir short hindlimbs, and extremelylarge feet. We studied two species ofMegaladapis (Table 2). The consensusfor postural and locomotor recon-structions for this genus suggests itwas a large-bodied, deliberate pri-mate that relied on strong graspinghands and feet for stability in thetrees. This would lead us to predictthat they would have relatively smallsemicircular canals, similar to theliving koalas, which were assignedan agility score of 2 (slow).2
Megaladapis is larger than all livingstrepsirrhines, necessitating an
Figure 3. Bivariate log-log plot of lateral semicircular canal radius (LSC-R) against bodymass for strepsirhine primates including the koala lemurs. Different symbols representeach agility category: red triangles, fast; white boxes, medium; blue circles, slow; Mm,Megaladapis madagascariensis; Mcf, Megaladapis sp. (northern variety); Me, Megalada-pis edwardsi.
Figure 2. Bivariate log-log plot of lateral semicircular canal radius (LSC-R) against bodymass for strepsirhine primates including the three sloth lemurs. Different symbols representeach agility category: red triangles, fast; white boxes, medium; blue circles, slow; Mp,Mesopropithecus pithecoides; Br, Babakotia radofilai; Pi, Palaeopropithecus ingens.
140 Walker et al. ARTICLES

extrapolation of the regression linebeyond the measured range (Fig. 3).If such extrapolations are reasona-ble, it seems that Megaladapis specieshave canals the same relative size asthose of lorisids. The only living pri-mates that are comparable to theMegaladapis in body mass are thegreat apes and humans, which havelarger semicircular canals than thekoala lemurs.2 Notably, orangutans,which are the primates most similarto the koala lemurs in intermembralindices, have larger semicircularcanals than do M. edwardsi, eventhough that taxon is reconstructed ashaving been somewhat larger inbody mass. This suggests an evenslower form of locomotion than thatof living Pongo, which was assignedan agility score of 2 (slow). The pre-dicted agility scores for the koalalemurs based on the mean canal ra-dius are in order of reconstructedlocomotion: Megaladapis sp. (thenorthern variety), 1.8 (extra slow toslow); M. edwardsi, 1.2 (extra slow toslow); M. madagascariensis, 1.1(extra slow to slow; Table 2).
THE ‘‘MONKEY LEMURS’’(ARCHAEOLEMURINAE)
The archaeolemur-ines are relativelylarge-brained, havelost their toothcombs in favor ofthick nipping inci-
sors, and have bilophodont molarteeth. These features have engen-dered comparisons to Old WorldMonkeys, leading to the informalappellation of ‘‘monkey lemurs’’ forthe group. But as well as havingcharacters reminiscent of Old Worldmonkeys, they also have features likethose of African apes and lemurs.They are stockier in body build thanare modern Old and New Worldmonkeys, and have strange handsand feet with broad terminal pha-langes on all digits.20,21,25 Althoughboth Archaeolemur and Hadropithe-cus show clear terrestrial adapta-tions, like most terrestrial primatesthey would have been capable ofclimbing in trees, but probably theydid so cautiously. Hadropithecus may
have climbed in trees or cliffs, but ithad no suspensory or leaping adap-tations.
Old World monkeys were assignedagility scores of 4-6 (medium to fast),with baboons and geladas, the formsmost often compared to the archaeo-lemurines, receiving scores of 4 (me-dium). For A. edwardsi, the predictedagility score based on the meancanal radius is 3.5 (medium-slow tomedium), slightly lower than theassigned scores for living Old Worldmonkeys. However, its agility score,predicted from the lateral semicircu-lar canal, which provides the bestseparation of agility categories (Box2), is 4.5, putting it within the OldWorld monkey range and suggestingthat a model based on a baboon or agelada might be appropriate for thisform. In contrast, the predicted agil-ity score for H. stenognathus is only2.6 (slow to medium slow; Table 2,Fig. 4) based on the average canal ra-dius, and even lower (2.5) from thelateral canal. The radii of curvatureof the semicircular canals in H. sten-ognathus are similar to those ofPapio, but the subfossil taxon is esti-mated to be about 10 kg heavier.These values suggest that locomo-tion in H. stenognathus was less ag-
ile than that of living Old Worldmonkeys, and that living cercopithe-cids are imperfect models for thisspecies.
MIOCENE LORISOIDS
With the possible exception of theadapoids, the fossil record for strep-sirhines that predate the Malagasysubfossil forms is relatively limited.Prior to the Miocene, the crownstrepsirrhine record is limited tothree lorisoid species from theEocene of the Fayum (Karanisiaclarki, Saharagalago misrensis, andWadilemur elegans).28,29 The Fayummaterial demonstrates that the twolorisoid families, Lorisidae and Gala-gidae, had already split by at leastthe Late Eocene. The fossil evidenceof the split between the two familiesis still between 15 and 20 Ma afterthe Early Eocene date (50–55 Ma)for the separation of these familiesestimated from molecular-basedphylogenies and molecular clocks.30
There are no cranial remains for anyof the pre-Miocene crown strepsir-rhines, although one postcranial ele-ment for the galagid Wadilemur sug-gests that it was already practicing a
Figure 4. Bivariate log-log plot of lateral semicircular canal radius (LSC-R) against bodymass for strepsirhine primates including the monkey lemurs. Different symbols representeach agility category: red triangles, fast; white boxes, medium; blue circles, slow; Ae,Archaeolemur edwardsi; Hs, Hadropithecus stenognathus.
ARTICLES The Semicircular Canal System and Locomotion 141

locomotor mode similar to that ofliving galagids, including someamount of leaping.29
The fossil record for lorisoidsimproves somewhat in the Miocene,
with multiple species being recog-nized in the genera Progalago,Komba, and Mioeuoticus. In particu-lar, two early lorisoid specimens thatpreserve the petrous bones have been
collected from the Early Miocene ofKenya. The first of these, a partialnatural endocast and the inferiorparts of the cranium, was originallyassigned to Progalago31 but on the
Box 3. Phylogenetic Generalized Least Squares Method
The phylogenetic relationshipsamong taxa in an analysis can havea confounding effect on the regres-sion results because measurementstaken on closely related species maynot be independent data points dueto shared ancestry. Several phyloge-netic regression methods have beendeveloped to account for this poten-tial problem.53 The most commonlyused method is Felsenstein’s phylo-genetic independent contrast (PIC)method. In this study, we used thephylogenetic generalized leastsquares (PGLS) method, which isfunctionally equivalent to the PICmethod.41,42 Given the same set ofspecies and phylogenetic tree, bothmethods produce the same resultsand therefore are considered variantsof one another.1 In PIC analyses,contrasts are calculated as the differ-ence between trait values betweensister taxa normalized by the squareroot of the sum of their branchlengths. Once calculated, the con-trasts can be used in any number ofstatistical analyses. In a PGLS analy-sis, the phylogenetic tree is con-verted to a variance-covariance ma-trix in which the diagonals representthe branch length from the root toeach tip species, while the off-diago-nals represent the branch lengthshared by pairs of tips from the rootof the tree to their last commonancestor.41,42 This phylogeneticallydetermined matrix is then used in aweighted linear regression.1 Becausethe off-diagonal elements in this ma-trix represent shared commonancestry, the data points for eachspecies have different weight in theanalysis. In both PIC and PGLS anal-yses, a variety of branch lengthtransformations can be applied tothe phylogenetic tree either to opti-mize the fit of the data or to test
specific models of character evolu-tion. In the current analysis, the Orn-stein-Uhlenbeck (OU) transform,41,43
Grafen’s q,44,45 and Pagel’s k46,47
branch length transformations wereused.
Figure 1. Phylogenetic tree. The base of strepsirhines was placed at 69 mya fromYoder and Yang,48 the base of Lorisiformes at 55 from Yoder,30 and the base ofLemuriformes at 62.7 mya from Yoder and Yang.48 Internal branching patterns anddivergence dates for strepsirrhines are based on several molecular studies.30,48–50 Thesplit between African and Asian lorises was set at 42 mya; the splits between the re-spective loris genera were set at 36 mya.50 The divergence between Galagoides andthe Galago-Otolemur clade was set to 30 mya, with the splits between G. elegantulusand the other bushbabies arbitrarily set to 15 mya. Daubentonia is set as the initialbranch from the other Lemuriformes at 62.7 mya. The split between cheirogaleids andthe rest of the Malagasy taxa is set at approximately 43 mya.48,50 Internal branchingdates within cheirogaleids are after Yoder and Yang.48 Phylogenetic positions of thesubfossil lemurs taken from Karanth and coworkers.49 Branch lengths for the subfossilsare slightly shorter than contemporary ones to reflect their status as recently extinct.Estimates of last occurrence are from Burney Robinson, and Burney.17
142 Walker et al. ARTICLES

basis of size, geological age, and ge-ography, we think is likely to be fromthe galagid Komba. The braincase, asoriginally figured, had the posteriorpart of the skull preserved. However,large pieces of this were broken offafterwards, by a preparator, presum-ably to expose the posterior part ofthe endocast. Fortunately, both tym-panic regions are still preserved. Theother specimen, a nearly completecranium, was also originally assignedto Progalago,31 but later placed in thelorisid genus Mioeuoticus based onresemblances to living lorisids.32
Although there are no securely asso-ciated postcranial remains of theEarly Miocene lorisoids, galagid-likelimb bones are relatively well repre-sented in collections. Some lorisid-like bones are also known, such asthe partial humerus reported to be ofthe right size to belong to Mioeuoti-cus.33 From the limb bones, itappears that leaping adaptations hadalready been established in theseMiocene galagids, while at the sametime slow-climbing lorisid-like adap-tations had appeared in the lorisids.The semicircular canal data offer theopportunity to further test this ideaand to evaluate the taxonomic
assignments of the postcranial mate-rial. Living galagids have muchlarger canals than do living lorisids,so if these assignments are correct,then Komba should have notablylarger canals than Mioeuoticus. Inthe absence of associated skeletonsfor these lorisids, we had to assumethat the crania had the same relativesize to body mass as occurs in extantspecies.
The canals of Komba are similar insize to those of modern galagids ofsimilar braincase dimensions, whilethe canals of Mioeuticus match thosefound in modern Nycticebus, whichhas the same cranial size. Agilityscores for living galagids range from4–6 (medium to fast). The predictedagility score for Komba is in thesame range (4.6, based on meancanal radius). Modern lorisids wereall assigned agility scores of 2 (slow),while the predicted score based onthe mean canal radius for Mioeuoti-cus is 2.6 (medium slow to slow)(Fig. 5; Table 2). These results con-firm that both galagid and lorisidlocomotor patterns were alreadypresent in the Miocene and supportthe taxonomic assignments of theknown postcranial bones. This result
also demonstrates that semicircularcanal measurements can be used topredict locomotion in fossil primatesusing a modern comparative sampleat least as far back as the Miocene.Other studies in progress will con-sider the utility of these measures forreconstructing locomotion in olderfossils.34,35
FUTURE DIRECTIONS
As discussed, the size of the semi-circular canals relates, in part, to thejerkiness of locomotion. Studyingthese canals therefore offers a win-dow into an element of locomotorbehavior that is only indirectlyreflected in other sorts of data.Unfortunately, few data directlyrelated to this variable are availablefor living primates, which compli-cates attempts to form analogies.Locomotor behavior should ideallybe characterized in terms of spectraof angular head velocities and accel-erations rather than in six basic cate-gories of the degree of agility, as wedid in our analysis. Miniaturizationof accelerometers and telemetricequipment or the development of 3-D video and analytical software thatcould be used during the day ornight might give us this informationfor captive specimens in controlledenvironments. Although it mighteventually be possible to measurehead velocities and accelerations infreely moving species in the wild, itprobably will not be possible to do itfor more than a few species because ofcost, conservation measures, and poli-tics. So for now our current approachis the only realistic option for analyz-ing many species. It is likely, however,that the imprecision in defining theseagility categories is contributing to theoverlap between categories in someanalyses; an even stronger signal mightbe detectable if quantitative data onjerkiness were available.In spite of these limitations, we
found that results of both the con-ventional and the phylogeneticallycorrected regression models showthat the semicircular canal systemholds a very strong functional signalrelated to locomotor agility. Such anapparently robust functional rela-tionship across primates (and other
Figure 5. Bivariate log-log plot of lateral semicircular canal radius (LSC-R) against bodymass for strepsirhine primates including the two Miocene lorisoids. Different symbols repre-sent each agility category: red triangles, fast; white boxes, medium; blue circles, slow; M,Mioeuoticus; K, Komba.
ARTICLES The Semicircular Canal System and Locomotion 143

mammals) reveals that the canal sys-tem responds adaptively to differentpostural and locomotor repertoires.The fact that the reconstructed canalsizes for both subfossil lemuroidsand fossil lorisoids generally matchpredications based on the knownpostcranial material confirms theutility of the semicircular canals inreconstructing behavior in extinctspecies.
ACKNOWLEDGMENTS
We thank G. Krovitz and T. Gar-land Jr. for their contributions tothis project and A. Grader, P. Hal-leck, and O. Karacan of the Centerfor Quantitative Imaging, Penn StateUniversity, for scanning facilities andadvice. We thank the following forgiving us access to specimens forscanning or access to CT scans: P.Chatrath, H. Chatterjee, J. Dines, K.Doyle, L. Godfrey, U. Golich, L. Gor-don, G. Hock, T. Holmes, W.Jungers, H. Kafka, D. Lieberman, R.Martin, E. Mbua, P. Morris, S.McLaren, J. Mead, T. Rasmussen, J.Rossie, D. Rothrock, J. Thewissen,and J. Wible. M. Nakatsukasa and G.Weber kindly scanned some fossilspecimens. The following institutionsprovided access to specimens: TheCarnegie Museum of Natural His-tory, Duke University Division ofFossil Primates; Field Museum ofNatural History; Grant Museum ofZoology and Napier Collection; Uni-versity College London; NationalMuseums of Kenya; National Mu-seum of Natural History; SmithsonianInstitution; Natural History Museumof Los Angeles County; Pratt Museum;Amherst College; Royal College of Sur-geons, London; University of KansasNatural History Museum; and the Nat-ural History Museum of Vienna. Wealso thank J. Fleagle and S. McGrawfor assistance with locomotor behav-ioral classifications and J. Cheverud, A.Ives, N. Jeffrey, R. Smith, and N. Vaseyfor advice. The comments of threeanonymous reviewers and John Fleaglegreatly improved this manuscript. Thisresearch was supported by theNational Science Foundation BCS-0003920 to A.W. and F.S.
REFERENCES
1 Guerin C, Demathieu G. 1993. Rhinocerotidae(Mammalia, Perissodactyla) tracks and tracesfrom the Upper Pliocene site of Laetoli (Tanza-nia). Geobios 26:497–513.
2 Spoor F, Garland T, Krovitz G, Ryan TM, Sil-cox MT, Walker A. 2007. The primate semicir-cular canal system and locomotion. Proc NatlAcad Sci USA 104:10808–10812.
3 Jones GM, Spells KE. 1963. A theoretical andcomparative study of the functional dependenceof the semicircular canal upon its physicaldimensions. Proc R Soc Lond B Biol Sci157:403–19.
4 Spoor F, Zonneveld F. 1998. Comparativereview of the human bony labyrinth. YearbookPhys Anthropol 41:211–251.
5 Watt HJ. 1924. Dimensions of the labyrinth cor-related. Proc R Soc Lond B Biol Sci 96:334–338.
6 Wilson VJ, Melvill-Jones G. 1979. Mamma-lian vestibular physiology. New York: PlenumPress.
7 Spoor F, Thewissen JGM. 2008. Balance: com-parative and functional anatomy in aquatic mam-mals. In: Thewissen JGM, Nummela S, editors.Sensory evolution on the threshold. Berkeley: Uni-versity of California Press. p 257–284.
8 Gray AA. 1907. The labyrinth of animals, vol.1. London: Churchill.
9 Hadziselimovic H, Savkovic LJ. 1964. Appear-ance of semicircular canals in birds in relationto mode of life. Acta Anat (Basel) 57:306–315.
10 Tanturri V. 1933. Zur Anatomie und Physio-logie des Labyrinthes der Voegel. Mschr.Ohrenheilk. Laryngo-Rhinol 67:1–27.
11 Turkewitsch BG. 1934. Zur Anatomie desGehororgans der Vogel (Canales semicircu-lares). Ztschr Anat Entwicklungsgesch 103:551–608.
12 Matano S, Kubo T, Matsunaga T, Niemitz C,Gunther M. 1986. On the size of the semicircu-lar canals organ in Tarsius bancanus. In: TaubDM, King FA, editors. Current perspectives inprimate biology. New York: Van Nostrand Rein-hold. pp. 122–129.
13 Matano S, Kubo T, Niemitz C, Guenther M.1985. Semicircular canal organ in 3 primatespecies and behavioral correlations. FortschrZool 30:677–680.
14 Spoor F. 2003. The semicircular canal sys-tem and locomotor behaviour, with special ref-erence to hominin evolution. Cour Forsch Inst.Senckenberg 243:93–104.
15 Jeffery N, Spoor F. 2006. The primate sub-arcuate fossa and its relationship to the semi-circular canals part I: prenatal growth. J HumEvol 51:537–549.
16 Felsenstein J. 1985. Phylogeny and the com-parative method. Am Nat 125:1–15.
17 Burney DA, Robinson GS, Burney LP. 2003.Sporormiella and the late Holocene extinctionsin Madagascar. Proc Natl Acad Sci USA100:10800–10805.
18 Godfrey LR, Jungers WL, Reed KE, SimonsEL, and Chatrath PS. 1997. Subfossil lemurs:inferences about past and present primate com-munities. In: Goodman SM, Patterson B, edi-tors. Natural change and human impact inMadagascar. Washington, D.C.: SmithsonianInstitution Press. p 218–256.
19 Godfrey LR, Jungers WL. 2003. The extinctsloth lemurs of Madagascar. Evol Anthropol12:252–263.
20 Godfrey LR, Jungers WL, Burney DA, Vasey N,Ramilisonina, Wheeler W, Lemelin P, Shapiro LJ,Schwartz GT, King SJ, Ramarolahy MF, Rahariv-
ony LL, Randria GFN. 2006. New discoveries ofskeletal elements of Hadropithecus stenognathusfrom Andrahomana Cave, southeastern Madagas-car. J HumEvol 51:395–410.
21 Godfrey LR, Jungers WL, Wunderlich RE,Richmond BG. 1997. Reappraisal of the post-cranium of Hadropithecus (Primates, Indroi-dea). Am J Phys Anthropol 103:529–556.
22 Jungers WL, Godfrey LR, Simons EL, Cha-trath PS. 1997. Phalangeal curvature and posi-tional behavior in extinct sloth lemurs (Prima-tes, Palaeopropithecidae). Proc Natl Acad SciUSA 94:11998–12001.
23 Jungers WL, Godfrey LR, Simons EL, Cha-trath PS, Rakotosamimanana B. 1991. Phyloge-netic and functional affinities of Babakotia (pri-mates), a fossil lemur from northern Madagas-car. Proc Natl Acad Sci USA 88:9082–9086.
24 Jungers WL, Lemelin P, Godfrey LR, Wunder-lich RE, Burney DA, Simons EL, Chatrath PS,James HF, Randria GF. 2005. The hands and feetof Archaeolemur: metrical affinities and theirfunctional significance. J HumEvol 49:36–55.
25 Godfrey LR, Jungers WL. 2002. Quaternaryfossil lemurs. In: Hartwig WC, editor. The pri-mate fossil record. Cambridge: Cambridge Uni-versity Press. p 97–121.
26 Carleton A. 1936. The limb bones and verte-brae of the extinct lemurs of Madagascar. ProcZool Soc Lond 106:281–307.
27 Simons EL, Godfrey LR, Jungers WL, Cha-trath PS, Rakotosamimanana B. 1992. A newgiant subfossil lemur, Babakotia, and the evolu-tion of the sloth lemurs. Folia Primatol (Basel)58:197–203.
28 Seiffert ER, Simons EL, Attia Y. 2003. Fossilevidence for an ancient divergence of lorisesand galagos. Nature 422:421–424.
29 Seiffert ER, Simons EL, Ryan TM, Attia Y.2005. Additional remains of Wadilemur elegans,a primitive stem galagid from the late Eoceneof Egypt. Proc Natl Acad Sci USA 102:11396–11401.
30 Yoder AD. 1997. Back to the future: a syn-thesis of Strepsirrhine systematics. Evol Anthro-pol 6: 11–22.
31 Le Gros Clark WE, Thomas DP. 1952. TheMiocene lemuroids of East Africa. Br Mus NatHist Fossil Mamm Afr 5:1–20.
32 Phillips E, Walker A. 2002. Fossil lorisoids.In: Hartwig WC, editor. The primate fossil re-cord. Cambridge, England: Cambridge Univer-sity Press. p 83–95.
33 Gebo DL, MacLatchy L, Kityo R. 1997. Anew lorisid humerus from the early Miocene ofUganda. Primates 38:423–427.
34 Ryan TM, Silcox MT, Simons E, Spoor F,Walker A. n. d. The semicircular canal systemin anthropoids. J Hum Evol. In Preparation.
35 Silcox MT, Bloch JI, Boyer DM, Godinot M,Ryan TM, Spoor F, Walker A. n. d. The semicir-cular canal system in early primates and eupri-mates. J Hum Evol. In Preparation.
36 Rabbitt RD, Damiano ER, Grant JW. 2004.Biomechanics of the vestibular semicircularcanals and otolith organs. In: Highstein SM,Popper A, Fay R, editors. The vestibular system.New York: Springer-Verlag. p 153–201.
37 Georgi JA, Sipla JS. 2008. Balance: comparativeanatomy and physiology in secondarily aquaticreptiles and birds. In: Thewissen JGM, NummelaS, editors. Sensory evolution on the threshold.Berkeley: University of California Press. p 233–256.
38 Gray AA. 1908. The labyrinth of animals,vol. 2. London: Churchill.
39 Ramprashad F, Landolt JP, Money KE, Lau-fer J. 1984. Dimensional analysis and dynamic
144 Walker et al. ARTICLES

response characterization of mammalian pe-ripheral vestibular structures. Am J Anat 169:295–313.
40 Smith RJ, Jungers WL. 1997. Body mass incomparative primatology. J HumEvol 32:523–559.
41 Garland T, Bennett AF, Rezende EL. 2005.Phylogenetic approaches in comparative physi-ology. J Exp Biol 208:3015–3035.
42 Garland T, Ives AR. 2000. Using the past topredict the present: confidence intervals forregression equations in phylogenetic compara-tive methods. Am Nat 155:346–364.
43 Blomberg SP, Garland T, Ives AR. 2003.Testing for phylogenetic signal in comparativedata: behavioral traits are more labile. Evolu-tion 57:717–745.
44 Grafen A. 1989. The phylogenetic regression.Philos Trans R Soc Lond B Biol Sci 326:119–157.
45 Grafen A. 1992. The uniqueness of the phy-logenetic regression. J Theor Biol 156:405–423.
46 Pagel M. 1997. Inferring evolutionary proc-esses from phylogenies. Zool Scripta 26:331–348.
47 Pagel M. 1999. Inferring the historical patternsof biological evolution. Nature 401:877–884.
48 Yoder AD, Yang ZH. 2004. Divergence datesfor Malagasy lemurs estimated from multiplegene loci: geological and evolutionary context.Mol Ecol 13:757–773.
49 Karanth KP, Delefosse T, Rakotosamima-nana B, Parsons TJ, Yoder AD. 2005. AncientDNA from giant extinct lemurs confirms singleorigin of Malagasy primates. Proc Natl AcadSci USA 102:5090–5095.
50 Roos C, Schmitz J, Zischler H. 2004. Pri-mate jumping genes elucidate strepsirrhine
phylogeny. Proc Natl Acad Sci USA 101:10650–10654.
51 Jungers WL, Demes B, Godfrey LR. 2008.How big were the ‘‘giant’’ extinct lemurs ofMadagascar? In: Fleagle JG, Gilbert CC, editors.Elwyn Simons: A Search for Origins. New York:Springer.
52 Vuillaumerandriamanantena M, GodfreyLR, Jungers WL, Simons EL. 1992. Morphol-ogy, taxonomy and distribution of Megaladapis,giant subfossil lemur from Madagascar. C RAcad Sci III 315:1835–1842.
53 Nunn CL, Barton RA. 2001. Comparativemethods for studying primate adaptation andallometry. Evol Anthropol 10:81–98.
VVC 2008 Wiley-Liss, Inc.
Books Received
� Pinhasi, R. and Mays, S., Eds.(2008). Advances in HumanPalaeopathology. 389 pp. Chi-chester: Wiley. ISBN 978-0-470-03602-0. $150.00 (cloth).
� Radick, G. (2008). The SimianTongue: The Long Debate AboutAnimal Language. 544 pp. Chi-cago: University of ChicagoPress. ISBN 978-0-226-70224-7.$45.00 (cloth).
� Tattersall, I. (2008). The WorldFrom Beginnings to 4000 BCE.143 pp. Oxford: Oxford Univer-sity Press. ISBN 978-0-19-533315-2. $19.95 (paper).
� Jones, A. (2007). Memory andMaterial Culture. 258 pp. Cam-bridge, Cambridge UniversityPress. ISBN 978-0-521-5455108.$25.99 (paper).
� Perry, S. with Manson, J.H.(2008). Manipulative Monkeys:The Capuchins of Lomas Bar-budal. 358 pp. Cambridge: Har-vard University Press. ISBN 978-0-674-02664-3. $45.00 (cloth).
� Shubin, N. (2008). Your InnerFish: A Journey into the 3.5-Bil-lion-Year History of the HumanBody. 229 pp. New York: Pan-theon Books. ISBN 978-0-375-42447-2. $24.00 (cloth).
� Wilson, D. S. (2007). Evolutionfor Everyone: How Darwin’sTheory Can Change the Way
We Think About Our Lives. 400pp. New York: Bantam Dell.ISBN 978-0-385-34092-2. $15.00(paper).
� Sommer, M. (2008). Bones andOchre: The Curious Afterlife ofthe Red Lady of Paviland. 398pp. Cambridge: Harvard Uni-versity Press. ISBN 978-0-674-02499-1. $39.95 (cloth).
� Owen, R. (2008). On the Natureof Limbs: A Discourse. 119 pp.Chicago: University of ChicagoPress. ISBN 0-226-09280-1.$20.00 (paper).
� Furuichi, T. and Thompson, J.,Eds. (2008). The Bonobos: Behav-ior, Ecology, and Conservation.xiii þ 327 pp. New York:Springer. ISBN 978-0-387-74785-9. $149.00 (cloth).
� Krebs, C. (2008). The Ecologi-cal World View. xviii 1 574 pp.Berkeley: University of Califor-nia Press. ISBN 978-0-520-25479. $59.95 (paper).
� Nudds, J. R. and Selden, P. A.(2008). Fossil Ecosystems ofNorth America: A Guide to theSites and Their ExtraordinaryBiotas. 288 pp. Chicago: Uni-versity of Chicago Press. ISBN0-226-60722-4. $39.00 (paper).
� Adams, B. J. and Byrd, J. E.(2008). Recovery, Analysis, andIdentification of Commingled
Human Remains. 374 pp.Totowa, New Jersey: HumanaPress. ISBN 978-1-58829-769-3.$129.00 (cloth).
� Livingstone, D. N. (2008). Adam’sAncestors: Race, Religion, andthe Politics of Human Origins.301 pp. Baltimore: The JohnsHopkins University Press. ISBN978-0-8018-8813-7. $35.00 (cloth).
� Bearzi, M. and Stanford, C. B.(2008). Beautiful Minds: TheParallel Lives of Great Apesand Dophins. 351 pp. Cambri-dge: Harvard University Press.ISBN 978-0-674-02781-7. $24.95(cloth).
� Rick, T. C. and Erlandson, J. M.,Eds. (2008). Human Impacts onAncient Marine Ecosystems: AGlobal Perspective. 319 pp.Berkeley: University of CaliforniaPress. ISBN 978-0-520-25343-8.$60.00 (cloth).
� Chapais, B. (2008). PrimevalKinship: How Pair-BondingGave Birth to Human Society.349 pp. Cambridge: HarvardUniversity Press. ISBN 978-0-674-02782-4. $39.95 (cloth).
� Gottschall, J. (2008). The Rapeof Troy: Evolution, Violence,and the World of Homer. 223pp. Cambridge: Cambridge Uni-versity Press. ISBN 978-0-521-69047-8. $34.99 (paper).
ARTICLES The Semicircular Canal System and Locomotion 145
Related Documents











