The role of protein solubilization in antigen removal from xenogeneic tissue for heart valve tissue engineering Maelene L. Wong a, b , J. Kent Leach a , Kyriacos A. Athanasiou a , Leigh G. Griffiths b, * a Department of Biomedical Engineering, University of California, Davis, One Shields Ave., Davis, CA 95616, USA b Department of Veterinary Medicine: Medicine and Epidemiology, University of California, Davis, One Shields Ave., Davis, CA 95616, USA article info Article history: Received 4 May 2011 Accepted 11 July 2011 Available online 31 July 2011 Keywords: Antigen removal Xenogeneic scaffold Decellularization Extracellular matrix Heart valve tissue engineering abstract Decellularization techniques have been developed in an attempt to reduce the antigenicity of xenogeneic biomaterials, a critical barrier in their use as tissue engineering scaffolds. However, numerous studies have demonstrated inadequate removal and subsequent persistence of antigens in the biomaterial following decellularization, resulting in an immune response upon implantation. Thus, methods to enhance antigen removal (AR) are critical for the use of xenogeneic biomaterials in tissue engineering and regenerative medicine. In the present study, AR methods incorporating protein solubilization principles were investigated for their ability to reduce antigenicity of bovine pericardium (BP) for heart valve tissue engineering. Bovine pericardium following AR (BP-AR) was assessed for residual antigenicity, tensile properties, and extracellular matrix composition. Increasing protein solubility during AR signif- icantly decreased the residual antigenicity of BP-ARdby an additional 80% compared to hypotonic solution or 60% compared to 0.1% (w/v) SDS decellularization methods. Moreover, solubilizing agents have a dominant effect on reducing the level of residual antigenicity of BP-AR beyond that achieved by AR additives alone. Tested AR methods did not compromise the tensile properties of BP-AR compared to native BP. Furthermore, residual cell nuclei did not correlate to residual antigenicity, demonstrating that residual nuclei counts may not be an appropriate indicator of successful AR. In conclusion, AR strategies promoting protein solubilization significantly reduced residual antigens compared to decellularization methods without compromising biomaterial functional properties. This study demonstrates the importance of solubilizing protein antigens for their removal in the generation of xenogeneic scaffolds. Published by Elsevier Ltd. 1. Introduction Over 100,000 heart valve replacements are performed annually in the United States [1]. However, residual antigenicity in glutaraldehyde-fixed bioprostheses results in chronic immune- mediated degeneration and subsequent calcification, thus limiting prosthesis lifetime [2,3]. Additionally, fixation prevents biomaterial growth with the patient [4]. Generation of a tissue-engineered heart valve from an appropriate scaffold seeded with autologous cells can potentially overcome drawbacks of glutaraldehyde-fixed bioprostheses. Normal valve function hinges on maintained mobility, pliability, and integrity of the valve leaflets or cusps [5]. As these features are largely dependent on the extracellular matrix (ECM), selection of an appropriate tissue-engineered heart valve scaffold material is crucial. Xenogeneic biomaterials (e.g., bovine pericardium (BP)) are advantageous because they possess the appropriate complex ECM architecture (composition and structure) crucial for physiological heart valve function [6,7]. However, xenoantigens represent a major barrier to the use of xenogeneic scaffolds in tissue engi- neering. Achieving adequately low levels of biomaterial antige- nicity while maintaining appropriate mechanical properties are critical considerations that have yet to be fully addressed. Antigen removal (AR) from xenogeneic biomaterials is required for their use as tissue engineering scaffolds. Initial attempts at scaffold production focused on cell removal (decellularization). The efficacy of osmotic lysis [8e10], chemical [8e10], enzymatic [8,10], or combinatorial [8e10] decellularization techniques for the generation of xenogeneic scaffolds has been investigated previ- ously. These decellularization approaches make two critical assumptions: (1) xenoantigens are only found within the cell and (2) removal of cells as determined under light microscopy is equivalent to biomaterial non-immunogenicity. Previously repor- ted decellularization or AR methods relied predominantly on either osmotic lysis or presence of a detergent to permeabilize the cell * Corresponding author. Tel.: þ1 530 754 0334; fax: þ1 530 752 0414. E-mail address: lggriffi[email protected] (L.G. Griffiths). Contents lists available at ScienceDirect Biomaterials journal homepage: www.elsevier.com/locate/biomaterials 0142-9612/$ e see front matter Published by Elsevier Ltd. doi:10.1016/j.biomaterials.2011.07.030 Biomaterials 32 (2011) 8129e8138

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Biomaterials 32 (2011) 8129e8138
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
The role of protein solubilization in antigen removal from xenogeneic tissuefor heart valve tissue engineering
Maelene L. Wong a,b, J. Kent Leach a, Kyriacos A. Athanasiou a, Leigh G. Griffiths b,*
aDepartment of Biomedical Engineering, University of California, Davis, One Shields Ave., Davis, CA 95616, USAbDepartment of Veterinary Medicine: Medicine and Epidemiology, University of California, Davis, One Shields Ave., Davis, CA 95616, USA
a r t i c l e i n f o
Article history:Received 4 May 2011Accepted 11 July 2011Available online 31 July 2011
Keywords:Antigen removalXenogeneic scaffoldDecellularizationExtracellular matrixHeart valve tissue engineering
* Corresponding author. Tel.: þ1 530 754 0334; faxE-mail address: [email protected] (L.G. Griffi
0142-9612/$ e see front matter Published by Elsevierdoi:10.1016/j.biomaterials.2011.07.030
a b s t r a c t
Decellularization techniques have been developed in an attempt to reduce the antigenicity of xenogeneicbiomaterials, a critical barrier in their use as tissue engineering scaffolds. However, numerous studieshave demonstrated inadequate removal and subsequent persistence of antigens in the biomaterialfollowing decellularization, resulting in an immune response upon implantation. Thus, methods toenhance antigen removal (AR) are critical for the use of xenogeneic biomaterials in tissue engineeringand regenerative medicine. In the present study, AR methods incorporating protein solubilizationprinciples were investigated for their ability to reduce antigenicity of bovine pericardium (BP) for heartvalve tissue engineering. Bovine pericardium following AR (BP-AR) was assessed for residual antigenicity,tensile properties, and extracellular matrix composition. Increasing protein solubility during AR signif-icantly decreased the residual antigenicity of BP-ARdby an additional 80% compared to hypotonicsolution or 60% compared to 0.1% (w/v) SDS decellularization methods. Moreover, solubilizing agentshave a dominant effect on reducing the level of residual antigenicity of BP-AR beyond that achieved byAR additives alone. Tested AR methods did not compromise the tensile properties of BP-AR compared tonative BP. Furthermore, residual cell nuclei did not correlate to residual antigenicity, demonstrating thatresidual nuclei counts may not be an appropriate indicator of successful AR. In conclusion, AR strategiespromoting protein solubilization significantly reduced residual antigens compared to decellularizationmethods without compromising biomaterial functional properties. This study demonstrates theimportance of solubilizing protein antigens for their removal in the generation of xenogeneic scaffolds.
Published by Elsevier Ltd.
1. Introduction
Over 100,000 heart valve replacements are performed annuallyin the United States [1]. However, residual antigenicity inglutaraldehyde-fixed bioprostheses results in chronic immune-mediated degeneration and subsequent calcification, thus limitingprosthesis lifetime [2,3]. Additionally, fixation prevents biomaterialgrowth with the patient [4]. Generation of a tissue-engineeredheart valve from an appropriate scaffold seeded with autologouscells can potentially overcome drawbacks of glutaraldehyde-fixedbioprostheses.
Normal valve function hinges on maintained mobility, pliability,and integrity of the valve leaflets or cusps [5]. As these features arelargely dependent on the extracellular matrix (ECM), selection of anappropriate tissue-engineered heart valve scaffold material iscrucial. Xenogeneic biomaterials (e.g., bovine pericardium (BP)) are
: þ1 530 752 0414.ths).
Ltd.
advantageous because they possess the appropriate complex ECMarchitecture (composition and structure) crucial for physiologicalheart valve function [6,7]. However, xenoantigens representa major barrier to the use of xenogeneic scaffolds in tissue engi-neering. Achieving adequately low levels of biomaterial antige-nicity while maintaining appropriate mechanical properties arecritical considerations that have yet to be fully addressed.
Antigen removal (AR) from xenogeneic biomaterials is requiredfor their use as tissue engineering scaffolds. Initial attempts atscaffold production focused on cell removal (decellularization). Theefficacy of osmotic lysis [8e10], chemical [8e10], enzymatic [8,10],or combinatorial [8e10] decellularization techniques for thegeneration of xenogeneic scaffolds has been investigated previ-ously. These decellularization approaches make two criticalassumptions: (1) xenoantigens are only found within the cell and(2) removal of cells as determined under light microscopy isequivalent to biomaterial non-immunogenicity. Previously repor-ted decellularization or ARmethods relied predominantly on eitherosmotic lysis or presence of a detergent to permeabilize the cell

Fig. 1. Western blot of residual antigens extracted from BP-AR following 2 days of ARwith increasing concentrations of DTT. Densitometry values measured for each ARtreatment after 2 d were normalized to the corresponding 1 m control to determinethe residual antigenicity ratio. Molecular weight ladder in kDa.
M.L. Wong et al. / Biomaterials 32 (2011) 8129e81388130
membrane and thereby expose the intracellular contents of a tissueto the decellularization solution. Such approaches have universallyneglected to consider the need for antigenic components of thebiomaterial to be maintained in a soluble form in order to facilitatetheir removal into aqueous extraction solutions. Indeed, severalpublications have demonstrated the ability of decellularizedxenogeneic biomaterials to elicit an immune response [11e15].These findings have challenged the fundamental assumptions ofdecellularization and consequently prompted a shift in focustowards AR in the development of xenogeneic scaffolds for tissueengineering applications [10,16].
The use of reducing agents [17e20] and salts [21,22] to maintainprotein solubility in proteomic applications is well established.However, the importance of maintaining protein solubility to ach-ieve AR from intact tissues for tissue engineering applications hasnot been investigated. This is a surprising oversight since proteinsolubility is fundamental in permitting protein diffusion intoaqueous solution and from tissue during AR. We hypothesized thatfailure to maintain antigenic proteins in a soluble form during theAR process will result in their precipitation from solution, pre-venting effective removal from the tissue. Furthermore, wehypothesized that AR methods which enhance protein solubilitywill reduce BP antigenicity while maintaining biomaterial func-tional properties. In this study, the importance of promotingprotein solubilization for effective AR was evaluated by assessingresidual antigenicity of BP in the presence and absence ofcompounds (reducing agent and salt) predicted to maintain anti-genic proteins in a soluble form. The effect of each AR protocol onmechanical properties and ECM composition of BP following AR(BP-AR) was assessed by uniaxial tensile testing, biochemicalquantification of ECM components, and histological analysis.
2. Materials and methods
2.1. Tissue harvest
All chemicals were from SigmaeAldrich (St. Louis, MO) unless otherwise stated.Bovine pericardium was harvested as described by Griffiths et al. [23,24] (n ¼ 4).Fresh BP was excised from adult cattle within 8 h of death and placed into PBS (pH7.4) containing 0.1% (w/v) EDTA and 1% (v/v) Antibiotic Antimycotic Solution (AAS).Following removal of pericardial fat and loose connective tissue, BP was stored inDulbecco’s Modified Eagles Medium (DMEM) containing 15% (v/v) dimethyl sulf-oxide (DMSO) at �80 �C.
2.2. Antiserum production
All animal procedures were conducted in accordance with the guidelinesestablished by University of California, Davis IACUC and the Guide for the Care andUse of Laboratory Animals [25]. Anti-BP serumwas generated as previously described[23,24]. Briefly, following subcutaneous injection of BP homogenate and Freund’sadjuvant at a 1:1 ratio into New Zealand white rabbits (n ¼ 4) on days 0, 14, and 28,blood was collected at day 84. Serum was isolated following centrifugation at3000 rpm for 10 min and stored at �80 �C.
2.3. Protein extraction
Protein extraction from minced BP was adapted from the method previouslydescribed by Griffiths et al. [23]. Minced BP (0.2 g) was incubated in standardextraction solution (10 mM TriseHCl (pH 8.0) containing 1 mM dithiothreitol (DTT),2 mM MgCl2, 10mM KCl, and 0.5 mM Pefabloc SC (Roche, Indianapolis, IN)) containing0.1% (w/v) sodium dodecyl sulfate (SDS, Bio-Rad, Hercules, CA) at 1000 rpm, 4 �C for1 h. Following centrifugation at 17,000�g, 4 �C for 25 min, recovered supernatantwas defined as the BP extract. The BP extract was concentrated using Amicon Ultra-0.5 centrifugal filter units with Ultracel-3 membranes (3 kDa cut-off) (Millipore,Billerica, MA) at 14,000�g, 4 �C for 30 min and stored at �80 �C.
2.4. One-dimensional electrophoresis and Western blot (1-DE and WB)
One-dimensional electrophoresis and Western blot (1-DE and WB) was per-formed as previously described by Griffiths et al. [24]. Equal volumes of BP extractwere separated on NuPAGE Novex 4e12% Bis-Tris gels (Invitrogen, Carlsbad, CA)with the XCell SureLock Mini-Cell electrophoresis unit (Invitrogen) and blotted to
nitrocellulose membranes (0.2 mm pores) according to the manufacturer’s protocolusing the XCell II Blot Module. Blots were blocked with Pierce Protein-Free T20Blocking Buffer (Thermo Fisher Scientific, Rockford, IL) containing 0.05% (v/v)Tween-20 (Bio-Rad) (PFBBT) for 1 h and probed with a 1:100 dilution of rabbitserum generated against native BP in PFBBT at 4 �C, overnight. Following three10 minwashes with PFBBT, blots were incubated in a 1:5000 dilution of horseradishperoxidase-conjugated mouse monoclonal anti-rabbit IgG light chain specific anti-body (Jackson ImmunoResearch Laboratories Inc., West Grove, PA) in 10 mM Tris-buffered saline (pH 8.8). Blots underwent five 10 min washes in 10 mM Tris-buffered saline (pH 8.8) with 0.05% (v/v) Tween-20. Protein bands were visualizedwith Immobilon Western Chemiluminescent HRP Substrate (Millipore) and imagedusing the FluorChem Xplor CCD bioimaging system and AlphaView image acquisi-tion and analysis software (Alpha Innotech Corp., Santa Clara, CA) (Fig. 1).
2.5. Antigen removal (AR)
All steps of the AR protocol were performed in 2 mL working volume at 4 �C and125 rpm using amethod adapted fromGoncalves et al. [10]. Intact pieces of BP (0.2 g,approximately 1.0 cm � 1.5 cm) were incubated in basic AR buffer (BARB; 0.5 mM
Pefabloc, 1% (v/v) AAS in 10 mM TriseHCl (pH 8.0)) for 2 d; anatomically adjacentpieces of BP subjected to 1 min incubations served as negative AR controls.Following nucleic acid digestion (2.5 Kunitz units/mL deoxyribonuclease I, 7.5 Kunitzunits/mL ribonuclease A, 0.15 M NaCl, 2 mM MgCl2(H2O)6, 1% (v/v) AAS in 10 mM
TriseHCl (pH 7.6)) for 24 h and washout (0.5 mM Pefabloc, 1% (v/v) AAS in 20 mM
Tris-buffered saline (pH 7.5)) for 48 h, BP post-AR (BP-AR) was stored in DMEMwith15% (v/v) DMSO at �80 �C.
Initially, efficiency of AR using no additive, 134 mM 3-(benzyldimethylammonio)propanesulfonate (NDSB-256), or 0.1% (w/v) SDS in either BARB or BARB containing1mMDTT, 2mMMgCl2, and 10mM KCl (solubilizing antigen removing buffer 1, SARB-1) was compared (Table 1 e A1, A2). In phase 1 of DTT optimization, AR efficiency of134 mM NDSB-256 in SARB-1 containing different concentrations of DTT (0, 1, 25, 50,100, 200mM) was assessed (Table 1e B1, B2, B3, B4, B5, B6) (Fig. 1). In phase 2 of DTToptimization, AR efficiency using no additive,134mMNDSB-256, or 0.1% (w/v) SDS ineither SARB-1 or BARB containing 100 mM DTT, 2 mM MgCl2, and 10 mM KCl (solu-bilizing antigen removing buffer 2, SARB-2) was compared (Table 1 e C1, C2). Inphase 1 of KCl optimization, AR efficiency of 134mMNDSB-256 in SARB-2 containingdifferent concentrations of KCl (0, 10, 50, 100, 200, 500 mM) was assessed (Table 1 e
D1, D2, D3, D4, D5, D6). In phase 2 of KCl optimization, AR efficiency using noadditive, 134 mM NDSB-256, or 0.1% (w/v) SDS in either SARB-2 or BARB containing100 mM DTT, 2 mM MgCl2, and 100 mM KCl (optimal solubilizing antigen removingbuffer, opt SARB) was compared (Table 1 e E1, E2). All AR experiments were con-ducted with n ¼ 6 per group (Fig. 2).
For tensile testing, quantitative biochemistry, and histology of BP-AR, AR (usingno additive, 134 mM NDSB-256, or 0.1% (w/v) SDS in either BARB or opt SARB) wasperformed on 0.2 g pieces of BP which were then trimmed into smaller pieces fordownstream analysis (n ¼ 6 per group for all analyses).
2.6. Assessment of residual antigenicity following AR
Proteins extracted from BP-AR (as described in section 2.3 above) were definedas residual antigens. To assess the level of residual antigenicity of BP-AR, extracted

Table 1BARB supplements used in AR. Various concentrations of DTT, MgCl2, and KCl wereused to supplement BARB in a series of AR experiments (treatments not connectedby the same letter were assessed in different experiments) to demonstrate theimportance of protein solubility for AR. For each experiment, the buffer demon-strating a significant reduction in residual antigenicity compared to baseline wasnamed (SARB-1, SARB-2, opt SARB) and used as the negative control in the nextexperiment to determine whether further improvements could be made.
Buffer [DTT] [MgCl2] [KCl]
A1 (BARB) 0 mM 0 mM 0 mM
A2 (SARB-1) 1 mM 2 mM 10 mM
B1 0 mM 2 mM 10 mM
B2 (SARB-1) 1 mM 2 mM 10 mM
B3 25 mM 2 mM 10 mM
B4 50 mM 2 mM 10 mM
B5 (SARB-2) 100 mM 2 mM 10 mM
B6 200 mM 2 mM 10 mM
C1 (SARB-1) 1 mM 2 mM 10 mM
C2 (SARB-2) 100 mM 2 mM 10 mM
D1 100 mM 2 mM 0 mM
D2 (SARB-2) 100 mM 2 mM 10 mM
D3 100 mM 2 mM 50 mM
D4 (opt SARB) 100 mM 2 mM 100 mM
D5 100 mM 2 mM 200 mM
D6 100 mM 2 mM 500 mM
E1 (SARB-2) 100 mM 2 mM 10 mM
E2 (opt SARB) 100 mM 2 mM 100 mM
M.L. Wong et al. / Biomaterials 32 (2011) 8129e8138 8131
proteins were subjected to 1-DE and WB, probed with anti-native BP rabbit serumand assessed for IgG positivity. Densitometry was used to quantify the bandingpattern intensity. Residual antigenicity of BP-AR was defined as the ratio of bandingintensity for extracts following 2 d of AR to the 1 min AR control. Overall residualantigenicity for each AR protocol was calculated with respect to the level of theresidual antigens following AR in BARB by multiplying the intraexperimentalpercentages of residual antigenicity for each step of AR buffer optimization (BARB vs.SARB-1, to SARB-1 vs. SARB-2, to SARB-2 vs. opt SARB).
2.7. Uniaxial tensile testing
Tensile properties of native BP and BP-AR (n ¼ 6 per group) were determinedusing a uniaxial materials testing system (Instron Model 5565, Canton, MA) witha 50 N load cell. Strips of BP (15 � 4 mm) were cut from separate 0.2 g pieces ofnative BP and BP-AR along the circumferential and longitudinal anatomical axeswith samples from adjacent anatomical locations used for each replicate of the ARmethods and control tissue. A 2 mm diameter biopsy punch was used to subtractsemicircles from themidpoint of each 15mm side, creating a dog bone-shaped pieceof BP. BP was mounted under zero strain and subjected to a constant strain rate of 5%gauge length/sec [26]. Initial gauge length, width, and thickness were determinedusing digital calipers. For each sample, a true stressestrain curve was generatedfrom the loadeelongation curve using the instantaneous tissue length and cross-
Fig. 2. AR buffer schematic. Flowchart illustrating evolution of AR buffer components.Residual antigenicity of BP-AR generated using no additive, 134 mM NDSB-256, or 0.1%(w/v) SDS was assessed in each of 4 AR buffers (BARB, SARB-1, SARB-2, opt SARB) eachcontaining a progressionally increasing amount of DTT and/or KCl. All AR experimentswere conducted with n ¼ 6 per group.
sectional area [27] and the Young’s modulus, ultimate tensile stress, and ultimatetensile strain were determined.
2.8. Quantitative biochemistry
To determine whether AR methods change ECM composition, native BP and BP-AR samples (approximately a quarter of the initial 0.2 g piece byarea,n¼ 6 per group)were weighed, frozen at �20 �C overnight, lyophilized for 72 h, and weighed again.Lyophilized BP samples were digested in phosphate buffer containing 125 mg/mLpapain, 5 mM EDTA, and 5 mM N-acetyl-L-cysteine at 65 �C for 18 h. Collagen contentper dry weight (DW) was quantified from papain digests using a modified coloro-metric hydroxyproline assay [28]. Sulfated glycosaminoglycan (GAG) content perDWwas quantified from papain digests using the Blyscan sulfated GAG assay (BiocolorLtd., Carrickfergus, UK). Lyophilized BP samples were incubated in 20 volumes of0.25 M oxalic acid at 100 �C for 60 min. After cooling to room temperature, sampleswere centrifuged at 3000 rpm for 10 min and supernatant containing soluble elastinwas collected. Oxalic acid extractionwas repeated 3 times. Supernatant fromall stepswas pooled and elastin content per DWwas quantified using the Fastin elastin assay(Biocolor Ltd.).
2.9. Histology
Approximately one quarter of the initial 0.2 g piece (by area) of each native BPand BP-AR sample (n ¼ 6 per group) was histologically processed for Verhoeff vanGieson staining (VVG) and hematoxylin and eosin (H&E) staining. For both stains,slides were viewed under light microscopy (Nikon Eclipse E600). Images werecaptured of six random high-powered fields (HPFs) using a Retiga 1300 Q-imagingcamera and processed with Simple PCI software v5.2.1.1906 (Compix Inc., CranberryTownship, PA). Gross ECM structure in BP was assessed with VVG (collagen andelastin) staining. Slides stained with H&E were used to assess general tissuemorphology and quantify residual nuclei on native BP and BP-AR (n ¼ 6 per group, 6HPFs per slide). Correlation between nuclei counts and overall residual antigenicitywas determined by plotting average nuclei counts against overall residual antige-nicity for each AR treatment.
2.10. Statistical analysis
Densitometry values were compared between experimental AR groups andnegative AR control (1 min). Values determined from tensile testing, biochemicalassays, and histology were compared to those for control tissues (native BP). Non-repeated measures ANOVA and TukeyeKramer HSD post hoc analysis were per-formed on samplemeans. Correlationwas determined using bivariate fit analysis. Alldata are presented at mean � standard deviation from the mean. Statistical signif-icance was defined at p < 0.05.
3. Results
3.1. Antigen removal
3.1.1. Basic versus solubilizing antigen removal buffer 1 (BARB vs.SARB-1)
The residual antigenicity of BP-AR using 134 mM NDSB-256 inBARB (NDSB/BARB, 1.08 � 0.23) was not significantly different thanthat of BP-AR using BARB alone (1.36 � 0.22). However, residualantigenicity following use of 0.1% (w/v) SDS in BARB (SDS/BARB,0.64 � 0.20) was significantly reduced compared to both BARB(p < 0.0001) and NDSB/BARB (p < 0.05). The presence of 1 mM DTT,2 mM MgCl2, and 10mM KCl in SARB-1 significantly reduced residualantigenicity of BP-AR following treatment with no additive (SARB-1,0.85 � 0.23, p < 0.005) and 134 mM NDSB-256 (NDSB/SARB-1,0.66 � 0.17, p < 0.05), but not 0.1% (w/v) SDS (SDS/SARB-1,0.55 � 0.18), compared to in BARB (Fig. 3).
3.1.2. DTT optimizationIn phase 1, increasing DTT concentration in solutions containing
134 mM NDSB-256, 2 mM MgCl2, and 10 mM KCl resulted in BP-ARwith decreasing residual antigenicity (p < 0.0001), with a plateauat 100mMDTT (Fig. 4A). Inphase2, AR in thepresence of 100mMDTT(SARB-2) significantly reduced residual antigenicity of BP-ARcompared to 1 mM DTT (SARB-1) for no additive (1.17 � 0.24 to0.75� 0.23, p< 0.005),134mMNDSB-256 (0.96� 0.13 to 0.51�0.11,
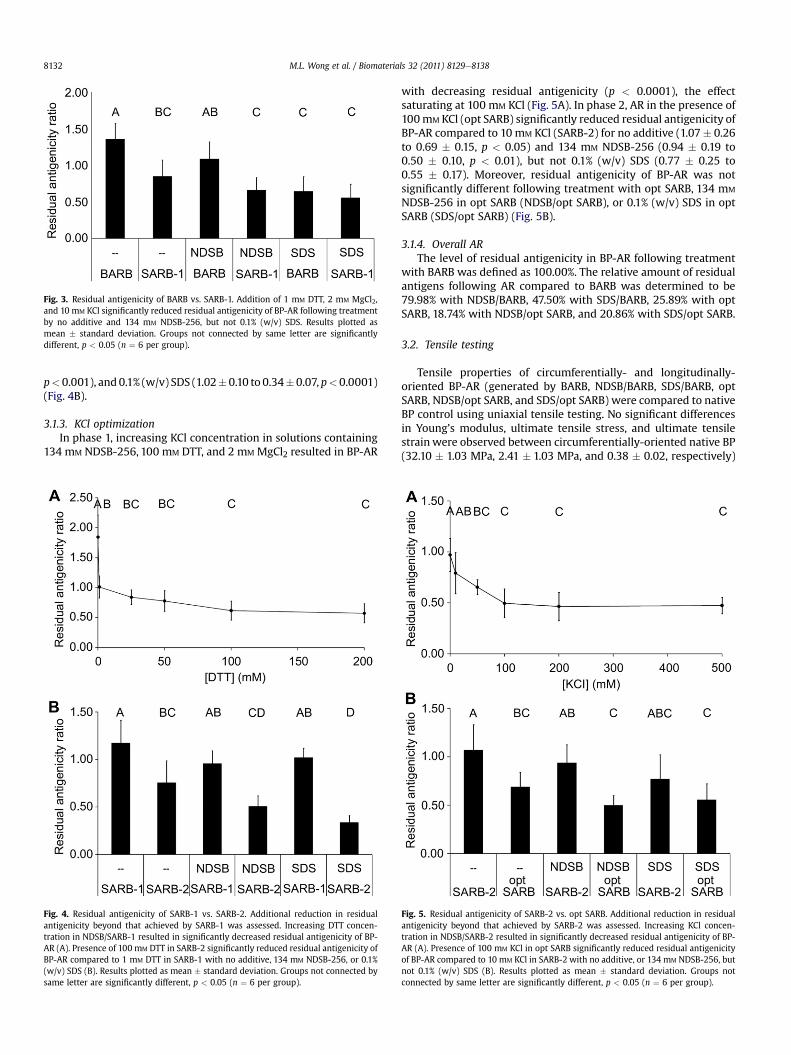
Fig. 3. Residual antigenicity of BARB vs. SARB-1. Addition of 1 mM DTT, 2 mM MgCl2,and 10 mM KCl significantly reduced residual antigenicity of BP-AR following treatmentby no additive and 134 mM NDSB-256, but not 0.1% (w/v) SDS. Results plotted asmean � standard deviation. Groups not connected by same letter are significantlydifferent, p < 0.05 (n ¼ 6 per group).
M.L. Wong et al. / Biomaterials 32 (2011) 8129e81388132
p<0.001), and0.1% (w/v) SDS (1.02�0.10 to 0.34�0.07, p<0.0001)(Fig. 4B).
3.1.3. KCl optimizationIn phase 1, increasing KCl concentration in solutions containing
134 mM NDSB-256, 100 mM DTT, and 2 mM MgCl2 resulted in BP-AR
Fig. 4. Residual antigenicity of SARB-1 vs. SARB-2. Additional reduction in residualantigenicity beyond that achieved by SARB-1 was assessed. Increasing DTT concen-tration in NDSB/SARB-1 resulted in significantly decreased residual antigenicity of BP-AR (A). Presence of 100 mM DTT in SARB-2 significantly reduced residual antigenicity ofBP-AR compared to 1 mM DTT in SARB-1 with no additive, 134 mM NDSB-256, or 0.1%(w/v) SDS (B). Results plotted as mean � standard deviation. Groups not connected bysame letter are significantly different, p < 0.05 (n ¼ 6 per group).
with decreasing residual antigenicity (p < 0.0001), the effectsaturating at 100 mM KCl (Fig. 5A). In phase 2, AR in the presence of100mM KCl (opt SARB) significantly reduced residual antigenicity ofBP-AR compared to 10 mM KCl (SARB-2) for no additive (1.07� 0.26to 0.69 � 0.15, p < 0.05) and 134 mM NDSB-256 (0.94 � 0.19 to0.50 � 0.10, p < 0.01), but not 0.1% (w/v) SDS (0.77 � 0.25 to0.55 � 0.17). Moreover, residual antigenicity of BP-AR was notsignificantly different following treatment with opt SARB, 134 mM
NDSB-256 in opt SARB (NDSB/opt SARB), or 0.1% (w/v) SDS in optSARB (SDS/opt SARB) (Fig. 5B).
3.1.4. Overall ARThe level of residual antigenicity in BP-AR following treatment
with BARB was defined as 100.00%. The relative amount of residualantigens following AR compared to BARB was determined to be79.98% with NDSB/BARB, 47.50% with SDS/BARB, 25.89% with optSARB, 18.74% with NDSB/opt SARB, and 20.86% with SDS/opt SARB.
3.2. Tensile testing
Tensile properties of circumferentially- and longitudinally-oriented BP-AR (generated by BARB, NDSB/BARB, SDS/BARB, optSARB, NDSB/opt SARB, and SDS/opt SARB) were compared to nativeBP control using uniaxial tensile testing. No significant differencesin Young’s modulus, ultimate tensile stress, and ultimate tensilestrain were observed between circumferentially-oriented native BP(32.10 � 1.03 MPa, 2.41 � 1.03 MPa, and 0.38 � 0.02, respectively)
Fig. 5. Residual antigenicity of SARB-2 vs. opt SARB. Additional reduction in residualantigenicity beyond that achieved by SARB-2 was assessed. Increasing KCl concen-tration in NDSB/SARB-2 resulted in significantly decreased residual antigenicity of BP-AR (A). Presence of 100 mM KCl in opt SARB significantly reduced residual antigenicityof BP-AR compared to 10 mM KCl in SARB-2 with no additive, or 134 mM NDSB-256, butnot 0.1% (w/v) SDS (B). Results plotted as mean � standard deviation. Groups notconnected by same letter are significantly different, p < 0.05 (n ¼ 6 per group).

M.L. Wong et al. / Biomaterials 32 (2011) 8129e8138 8133
and BP-AR generated by any AR buffer (Fig. 6). Similarly, nosignificant differences in Young’s modulus, ultimate tensile stress,and ultimate tensile strain were observed between longitudinally-oriented native BP (28.51 � 10.15 MPa, 2.12 � 0.60 MPa, and0.36 � 0.02, respectively) and BP-AR generated by any AR buffer(Fig. 6). No significant differences were observed between cir-cumferentially- and longitudinally-oriented native BP.
3.3. Quantitative biochemistry
Thewater contentof BP-ARgeneratedusingBARB (74.83�1.38%),NDSB/BARB (75.24 � 0.87%), SDS/BARB (71.50 � 4.01%), opt SARB
Fig. 6. Tensile properties of BP. Native BP and BP-AR were tested along anatomicallycircumferential and longitudinal axes. Young’s modulus (A), ultimate tensile strength(B), and ultimate tensile strain (C) of BP-AR were not found to significantly differ fromnative BP in the circumferential or longitudinal directions. Results plotted asmean � standard deviation. Groups not connected by same letter are significantlydifferent, p < 0.05 (n ¼ 6 per group).
(74.41 � 0.66%), NDSB/opt SARB (76.17 � 1.16%), or SDS/opt SARB(76.22� 1.66%) was not significantly different from that of native BP(73.79 � 1.97%) (Fig. 7A).
The collagen content per DW of BP-AR was not significantlydifferent than that of native BP (33.94 � 7.71% per DW) followingtreatment with BARB (45.15 � 18.21% per DW), NDSB/BARB(41.69 � 11.03% per DW), SDS/BARB (33.68 � 6.62% per DW), optSARB (34.05 � 6.52% per DW), NDSB/opt SARB (42.48 � 18.94% perDW), or SDS/opt SARB (38.46 � 9.33% per DW) (Fig. 7B).
The elastin content per DW of BP-AR following treatment withBARB (10.08 � 3.36% per DW), NDSB/BARB (8.04 � 1.89% per DW),opt SARB (7.54 � 3.25% per DW), or NDSB/opt SARB (7.40 � 1.20%per DW) was not significantly different from that in native BP(7.59 � 1.41% per DW). However, elastin in BP-AR was significantlyreduced compared to that in native BP when SDS/BARB(3.14 � 2.23% per DW, p < 0.05) or SDS/opt SARB (3.70 � 1.77% perDW, p < 0.05) was used for AR (Fig. 7C).
The GAG content per DWof BP-ARwas not significantly differentfrom that of native BP (0.55 � 0.04% per DW) when treated withBARB (0.52 � 0.05% per DW). However, AR using opt SARB(0.39 � 0.05% per DW), NDSB/BARB (0.47 � 0.04% per DW), orNDSB/opt SARB (0.34 � 0.05% per DW) significantly decreased GAGcontent per DW compared to that in native BP (p< 0.0001, p< 0.05,p < 0.0001, respectively) (Fig. 7D). The presence of residual SDS inBP-AR subjected to SDS/BARB and SDS/opt SARB interferedwith theBlyscan assay (data not shown).
3.4. Histology
Presence of nuclei was assessed in H&E-stained native BP andBP-AR sections (generated by BARB, NDSB/BARB, SDS/BARB, optSARB, NDSB/opt SARB, and SDS/opt SARB) (Fig. 8). All AR treatmentssignificantly reduced the number of nuclei per HPF compared tonative BP (85.72 � 20.35) (p < 0.005) except for BARB(83.28 � 19.66). In both BARB and opt SARB, the number ofpersistent nuclei following treatment in 0.1% (w/v) SDS(18.83 � 9.78 with BARB and 29.72 � 14.12 with opt SARB) wassignificantly less than that from 134 mM NDSB-256 (64.61 � 16.07with BARB, p < 0.0001; 58.00 � 16.44 with opt SARB, p < 0.0001),which was significantly less than that with no additive(83.28� 19.66 with BARB, p< 0.0001; 69.94�16.44with opt SARB,p < 0.0001). Although opt SARB resulted in significantly fewernuclei than BARB (p< 0.05), therewas not a significant difference innuclear count between BARB and opt SARB in the presence of134 mM NDSB-256 or 0.1% (w/v) SDS (Fig. 9). No statisticallysignificant correlation was observed between nuclei counts andoverall residual antigenicity of BP-AR (p ¼ 0.239, R2 ¼ 0.3234)(Fig. 10). Qualitatively, differences in collagen and elastin contentper DWand organizationwere not observed between native BP andBP-AR in VVG-stained sections (Fig. 11).
4. Discussion
The objectives of this study were (1) to determine whetherstrategies promoting protein solubility could reduce the antige-nicity of xenogeneic tissue beyond that achieved by decellulariza-tion methods, (2) to identify under which conditions proteinsolubilization-mediated AR was most effectively achieved, and (3)to assess whether these AR methods adversely affect the xenoge-neic biomaterial. We have demonstrated that (1) promotion ofprotein solubility enhances AR without compromising biomaterialproperties, (2) solubilizing factors are critical determinants of ARefficiency, overwhelming the effect of AR additives (hypotonicsolution, 134 mM NDSB-256, or 0.1% (w/v) SDS) in the level of

Fig. 7. Quantitative biochemical analysis of BP composition. Water, collagen, elastin, and GAG content per DW in BP. Water (A) and collagen (B) content per DW of BP-AR was notsignificantly different following AR. Elastin content per DWwas significantly decreased in BP-AR with 0.1% (w/v) SDS treatment (C). GAG content per DWwas significantly decreasedin BP-AR following treatment with opt SARB and/or 134 mM NDSB-256 (D). Results plotted as mean � standard deviation. Groups not connected by same letter are significantlydifferent, p < 0.05 (n ¼ 6 per group).
M.L. Wong et al. / Biomaterials 32 (2011) 8129e81388134
residual antigenicity, and (3) the number of residual nuclei may notbe an appropriate assessment of residual antigenicity.
Dithiothreitol and KCl have previously been utilized to facilitateprotein extraction from homogenized BP into aqueous solution[23,24], however solubilizing agents have yet to be incorporatedinto AR protocols. In this study, AR with hypotonic solution, 134 mM
NDSB-256, and 0.1% (w/v) SDS were assessed for their ability toreduce BP antigenicity in the presence and absence of compoundspredicted to maintain antigenic proteins in a soluble form. Hypo-tonic solution [8e10] and 0.1% (w/v) SDS [29] correspond toosmotic lysis and detergent-based decellularization, respectively.Use of 134 mM NDSB-256 was motivated by previous studies whichdemonstrated the effectiveness of the compound in proteinextraction protocols [23,24]. The addition of low levels of reducingagent and salt (1 mM DTT, 2 MgCl2, and 10 mM KCl) to BARB (SARB-1) significantly reduced the residual antigenicity of BP-AR treatedwith no additive or 134 mM NDSB-256. In contrast, insignificantimprovement in AR observed with 0.1% (w/v) SDS in SARB-1 can beexplained by the relatively high efficiency of SDS/BARB to removeantigens, owing to the ability of the anionic detergent to readilydenature and solubilize proteins. Low levels of DTT and KCl are ableto achieve a reduction in residual antigenicity that is not signifi-cantly different from that of 0.1% (w/v) SDS. These findings supportthe hypothesis that maintaining proteins in solution facilitates theirremoval from intact BP.
The effect of DTT and KCl concentration on AR enhancementwas investigated in a stepwise fashion as a series of two-phasestudies. Phase 1 aimed to determine the effect of increasing DTTor KCl concentration on BP-AR residual antigenicity. The optimal
concentrationwas then tested in phase 2 for its ability to influenceAR by BARB, NDSB/BARB, and SDS/BARB. For each study, theconcentration determined to be most effective for AR in theprevious study served as the new negative control to be improvedupon, such that the results reflect a progressive improvement inAR beyond that achieved in the previous step (Fig. 2). Solutionscontaining 100 mM DTT (SARB-2) were found to significantlyreduce residual antigenicity of BP-AR compared to 1 mM DTT(SARB-1). These data indicate that not only can the efficiency of ARbe modulated based on dose of reducing agent, but that a reducingagent can improve the degree of AR accomplished by the goldstandard for decellularization agents (SDS). The intracellularenvironment is maintained in a reduced state by thioredoxin andglutathione systems [30]. Water-soluble cytoplasmic proteins areconsequentially maintained in a reduced state within the cell.Following cell lysis in the decellularization process, the cellularreducing environment can no longer be maintained. Oxidation ofcytoplasmic proteins increases the tendency for disulfide bondformation to occur between proteins, resulting in protein aggre-gation and precipitation from solution [31]. By discouraging theformation of intermolecular disulfide bridges [17,20,31], presenceof a reducing agent prevents disulfide bridge-mediated aggrega-tion of proteins and their subsequent precipitation from solution[19]. The beneficial effect of DTT on AR suggests that maintenanceof a reducing environment during cell lysis is critical in facilitatingwater-soluble antigenic protein solubility and removal intoaqueous AR solutions.
The effect of altering KCl concentration on AR enhancement wasassessed for solutions containing the previously identified optimal

Fig. 8. Hematoxylin and eosin staining of BP. Representative images of H&E-stained BP sections showing preserved gross tissue morphology and presence of residual nuclei with allAR treatments. Scale bar represents 20 mm.
M.L. Wong et al. / Biomaterials 32 (2011) 8129e8138 8135
DTTconcentration (100mM). The addition of 100mM KCl (opt SARB)significantly reduced residual antigenicity of BP-AR compared to10 mM KCl (SARB-2) with no additive or 134 mM NDSB-256. Saltconcentration is known to affect protein solubility in aqueoussolutions, with protein solubility initially increasing as salt is addedto a solution (salting in) and decreasing as additional salt is added
Fig. 9. Residual nuclei per high-powered field in BP. Following AR, SDS/BARB and SDS/opt SARB most significantly reduced nuclei compared to native BP. Results plotted asmean � standard deviation. Groups not connected by same letter are significantlydifferent, p < 0.05 (n ¼ 6 per group).
(salting out) [21]. By incorporating a physiologically relevantconcentration of salt [32] in opt SARB (100 mM KCl), most biologicalproteins aremaintained between the points of salting in and saltingout, mitigating the aggregation of protein molecules via pro-teineprotein interactions [21], permitting maintenance of protein
Fig. 10. Residual nuclei per high-powered field vs. overall residual antigenicity.Residual nuclei counts per high powered field do not correlate to the overall residualantigenicity of BP-AR (p ¼ 0.239, R2 ¼ 0.3234). Nuclei counts represent the averagevalue from 6 replicates per AR treatment group (6 HPFs per replicate). Residual anti-genicity was calculated from n � 6 replicates per AR treatment group.

Fig. 11. Verhoeff van Gieson staining of BP. Representative images of VVG-stained BP sections showing gross collagen and elastin structure was preserved following AR. Scale barrepresents 20 mm.
M.L. Wong et al. / Biomaterials 32 (2011) 8129e81388136
molecules in solution for subsequent removal from BP. Takentogether, the results for the addition of both DTT and KCl to the ARbuffer support the hypothesis that maintaining solubility of water-soluble antigens is critical to achieving efficient AR from intact BP.Remarkably, differences in AR efficiency by BARB, NDSB/BARB, andSDS/BARB were mitigated by addition of 100 mM DTT, 2 mM MgCl2,and 100 mM KCl, suggesting that factors promoting solubility weremore influential than AR additives in reducing the level of residualantigenicity of BP-AR.
A successful AR process must maintain biomaterial mechanicalproperties in a range appropriate for physiological function.Closed heart valves experience planar tension as a result of bloodflow reversal [33]. The organization of ECM components confersdirectionality to heart valves important for their function.Circumferentially-oriented collagen fibers provide strength to theheart valve and the ability to accommodate the load imparted byblood flow during the cardiac cycle [34]. Radially-aligned elastinmolecules modulate the radial strain during valve opening andclosing [35]. In the present study, Young’s modulus, ultimatetensile stress, and ultimate tensile strain of BP-AR were notsignificantly different from native BP control (for both circumfer-ential and longitudinal axes), suggesting that AR using opt SARBdoes not compromise BP tensile properties. Moreover, the valuesmeasured for BP-AR generated using opt SARB are comparable toor exceed those of cadaveric human aortic and pulmonary valvesalong the circumferential and radial axes determined by Stradins
et al. [36]. Since tensile properties of BP-AR approximate those ofnative human heart valve, we reason that BP-AR generated usingopt SARB will be capable of withstanding the tensile forces expe-rienced by heart valves. Tension represents one important type ofloading experienced by heart valves. Future studies using cyclicalloading, stress relaxation, and biaxial tensile testing will betterinform how BP-AR will perform under in vivo conditions.
To ensure the structureefunction relationship was preserved,ECM composition of BP-AR was assessed. Following AR with optSARB, collagen content per DW of BP-AR was not observed to besignificantly different from that in native BP. This is in agreementwith the lack of significant difference in ultimate tensile stressbetween native BP and opt SARB-treated BP-AR, and furthersupports the promise of generating BP-AR using opt SARB withoutcompromising the functional properties of native BP. Additionally,AR using opt SARB did not significantly alter the amount of elastinin BP. However, the elastin content per DW of BP-AR was signifi-cantly decreased in the presence of 0.1% (w/v) SDS during AR. Aselastin is necessary for the large deformations heart valves undergoduring the cardiac cycle, this result represents a potential detri-mental effect of protocols utilizing 0.1% (w/v) SDS for AR. It shouldbe noted, however, that differences in elastin content per DW didnot appear to translate to differences in ultimate tensile strain ofBP-AR following 0.1% (w/v) SDS treatment compared to native BP.Antigen removal with opt SARB resulted in a significant decrease inGAG content per DW compared to that in native BP. Despite this

M.L. Wong et al. / Biomaterials 32 (2011) 8129e8138 8137
observed reduction in GAGs, significant differences in watercontent were not observed between native BP and BP-AR, likely dueto the fact that GAGs are not amajor contributor to the DWof nativeBP (0.55 � 0.04% per DW). The GAG content per DWof BP-AR couldnot be determined for 0.1% (w/v) SDS treated samples due to assayincompatibility with residual SDS in BP-AR extracts (data notshown). Overall, AR methods involving opt SARB do not drasticallycompromise the collagen, elastin, or GAG content per DWof BP-AR.
Previous studies have utilized residual nuclei counts as theprimary outcome measure for success of decellularization[8,9,29,37e39]. In the current study, cell nuclei were uniformlydistributed throughout sections of native BP. No significantdecrease in number of nuclei was noted following AR using BARB.Although treatment with opt SARB and/or 134 mM NDSB-256significantly reduced nuclei on BP-AR compared to native BP,treatment with 0.1% (w/v) SDS significantly reduced residual nucleifurther. Following the reasoning used in decellularization studies,these results suggest that AR buffers containing 0.1% (w/v) SDS arethe most effective at producing a non-immunogenic biologicalmaterial, regardless of the presence or absence of a reducing agentand salt. However, this conclusion is in stark contrast to the findingsfrom the assessment of residual antigenicity for BP-AR whichutilized a more specific, antibody-based assay. The IgG-basedassessment employed in this study provides a rigorous determi-nant of biomaterial antigenicity as it probes for residual antigensusing specific antibodyeantigen interactions that elicit the immuneresponse in vivo. Moreover, since B-cell isotype switching to IgGproduction requires T-helper cell costimulation [40], limitingassessment to IgG positivity indicates the residual antigensassessed in this study are capable of inducing both a humoral andcell-mediated immune response. The absence of a significantcorrelation between residual nuclei counts and the overall residualantigenicity suggests that the presence or absence of residual cellnuclei may not be an appropriate indicator of biomaterial antige-nicity. Following decellularization of rat aortic valve leaflets byhypotonic lysis or Triton X-100, Meyer et al. reported the presenceof residual major histocompatibility complex molecules inspite ofapparent tissue acellularity on H&E-stained sections [41]. Kasimiret al. observed persistent galactosyl a-1,3-galactose (a-gal) xen-oantigens and cellular debris in commercially-available SynerGraftdecellularized porcine grafts [16] despite being branded as anacellular graft [37]. The present study confirms that assessment ofcellularity under light microscopy as an indication of reduction inbiomaterial antigenicity is prone to both underestimation (e.g., optSARB and NDSB/opt SARB) and overestimation (e.g., SDS/BARB) ofAR efficiency. It is becoming increasingly clear that a nuclei-basedassessment of a xenogeneic scaffold may be an insufficientmeasure of its residual antigenicity. Overestimation of AR efficiencyby assessment of residual nuclei counts is concerning as high levelsof residual antigenicity in an apparently acellular scaffold may becapable of eliciting an immune response following in vivoimplantation. Underestimation of AR efficiency by assessment ofresidual nuclei counts also raises important questions regarding theeffect of residual nuclei on the immunological potential of thescaffold following in vivo implantation. Future in vivo implantationstudies are therefore required to elucidate how xenogeneic scaffoldresidual antigenicity and residual nuclei counts translate to theimmunological potential of the biomaterial.
5. Conclusions
This present study indicates that AR buffers promoting proteinsolubility (opt SARB) significantly reduce residual antigens in BP-ARcompared to previously reported decellularization and/or ARbuffers that do not maintain protein solubility. By accounting for
the solubility of antigenic proteins to be removed, the total residualantigenicity of BP-AR can be reduced beyond that attained bycommon decellularization protocols. Factors promoting solubility(i.e., reducing agent and salt) have a dominant effect on AR effi-ciency beyond that achieved by the presence of a particular additivealone. Antigen removal using opt SARB permitted maintenance ofBP-AR tensile properties similar to those of native BP. Moreover, thecollagen and elastin content per DW are unchanged in BP-ARgenerated using opt SARB. However, AR with 0.1% (w/v) SDSsignificantly decreased elastin content per DW. Furthermore, theprimary endpoint of decellularization methods (absence of cellsunder light microscopy) may not be an appropriate measure for theremoval of antigenic proteins from xenogeneic materials. Takentogether, these results demonstrate that by promoting proteinsolubilization, AR methods utilizing opt SARB (1) yield BP-AR withsignificantly reduced residual antigenicity compared to currentmethods of decellularization and (2) do not compromise thefunctional properties of the biomaterial. Adaptation of the pre-sented AR strategy to other tissues of the body has potential for thegeneration of xenogeneic scaffolds with an immunogenic potentialsignificantly lower than that achieved by current decellularizationmethods.
Acknowledgments
The authors would like to thank Janelle L. Wong for her tech-nical assistance with the quantitative biochemical assays andhistology. The authors would also like to thank Dr. Jerry Hu, SriramV. Eleswarapu, Donald Responte, and Vincent P. Willard from theAthanasiou lab for their advice regarding tensile testing andbiochemical assays and Dr. Clare E. Yellowley for use of hermicroscope. This work was funded by the American Heart Asso-ciation (11SDG4980023, LGG) and HHMI Med into Grad Initiative(56005706, MLW).
References
[1] Rosamond W, Flegal K, Friday G, Furie K, Go A, Greenlund K, et al. Heartdisease and stroke statisticse2007 update: a report from the American HeartAssociation Statistics Committee and Stroke Statistics Subcommittee. Circu-lation 2007;115:e69e171.
[2] Human P, Zilla P. Characterization of the immune response to valve bio-prostheses and its role in primary tissue failure. Ann Thorac Surg 2001;71:S385e8.
[3] Manji RA, Zhu LF, Nijjar NK, Rayner DC, Korbutt GS, Churchill TA, et al.Glutaraldehyde-fixed bioprosthetic heart valve conduits calcify and fail fromxenograft rejection. Circulation 2006;114:318e27.
[4] Shinoka T, Breuer C, Tanel R, Zund G, Miura T, Ma P, et al. Tissue engineeringheart valves: valve leaflet replacement study in a lamb model. Ann ThoracSurg 1995;60:S513e6.
[5] Sacks MS, Schoen FJ, Mayer JE. Bioengineering challenges for heart valve tissueengineering. Annu Rev Biomed Eng 2009;11:289e313.
[6] Mendelson K, Schoen F. Heart valve tissue engineering: concepts, approaches,progress, and challenges. Ann Biomed Eng 2006;34:1799e819.
[7] Schmidt CE, Baier JM. Acellular vascular tissues: natural biomaterials for tissuerepair and tissue engineering. Biomaterials 2000;21:2215e31.
[8] Yang M, Chen C-Z, Wang X-N, Zhu Y-B, Gu YJ. Favorable effects of thedetergent and enzyme extraction method for preparing decellularized bovinepericardium scaffold for tissue engineered heart valves. J Biomed Mater Res BAppl Biomater 2009;91B:354e61.
[9] Mirsadraee S, Wilcox HE, Korossis SA, Kearney JN, Watterson KG, Fisher J, et al.Development and characterization of an acellular human pericardial matrixfor tissue engineering. Tissue Eng 2006;12:763e73.
[10] Goncalves AC, Griffiths LG, Anthony RV, Orton EC. Decellularization of bovinepericardium for tissue-engineering by targeted removal of xenoantigens.J Heart Valve Dis 2005;14:212e7.
[11] Simon P, Kasimir MT, Seebacher G, Weigel G, Ullrich R, Salzer-Muhar U, et al.Early failure of the tissue engineered porcine heart valve SYNERGRAFT(TM) inpediatric patients. Eur J Cardiothorac Surg 2003;23:1002e6.
[12] Rieder E, Seebacher G, Kasimir M-T, Eichmair E, Winter B, Dekan B, et al.Tissue engineering of heart valves: decellularized porcine and human valvescaffolds differ importantly in residual potential to attract monocytic cells.Circulation 2005;111:2792e7.

M.L. Wong et al. / Biomaterials 32 (2011) 8129e81388138
[13] Kasimir M, Rieder E, Seebacher G, Nigisch A, Dekan B, Wolner E, et al.Decellularization does not eliminate thrombogenicity and inflammatorystimulation in tissue-engineered porcine heart valves. J Heart Valve Dis 2006;15:278e86.
[14] Wilhelmi M, Rebe P, Leyh R, Wilhelmi M, Haverich A, Mertsching H. Role ofinflammation and ischemia after implantation of xenogeneic pulmonary valveconduits: histological evaluation after 6 to 12 months in sheep. Int J ArtifOrgans 2003;26:411e20.
[15] Bastian F, Stelzmüller M-E, Kratochwill K, Kasimir M-T, Simon P, Weigel G. IgGdeposition and activation of the classical complement pathway involvementin the activation of human granulocytes by decellularized porcine heart valvetissue. Biomaterials 2008;29:1824e32.
[16] Kasimir MT, Rieder E, Seebacher G, Wolner E, Weigel G, Simon P. Presence andelimination of the xenoantigen gal (alpha1, 3) gal in tissue-engineered heartvalves. Tissue Eng 2005;11:1274e80.
[17] Shaw MM, Riederer BM. Sample preparation for two-dimensional gel elec-trophoresis. Proteomics 2003;3:1408e17.
[18] Herbert BR, Molloy MP, Gooley AA, Walsh BJ, Bryson WG, Williams KL.Improved protein solubility in two-dimensional electrophoresis using tributylphosphine as reducing agent. Electrophoresis 1998;19:845e51.
[19] Weiss W, Görg A. Protein solubilization. In: Hagen Jv, editor. Proteomicssample preparation. Weinheim: Wiley-VCH; 2008. p. 135e7.
[20] Rabilloud T. Protein solubility in two-dimensional electrophoresis. In:Walker JM, editor. The protein protocols handbook. Totowa: Humana Press;2002. p. 131e40.
[21] Koolman J, Roehm K-H. Isolation and analysis of proteins. Color atlas ofbiochemistry. 2nd ed. Stuttgart and New York: Thieme; 2005. p. 78e79.
[22] Upreti GC, Davis C, Oliver J. Preparation of representative homogenates ofbiological tissues: effect of salt on protein extraction. Anal Biochem 1991;198:298e301.
[23] Griffiths LG, Choe L, Lee KH, Reardon KF, Orton EC. Protein extraction and2-DE of water- and lipid-soluble proteins from bovine pericardium, a low-cellularity tissue. Electrophoresis 2008;29:4508e15.
[24] Griffiths LG, Choe LH, Reardon KF, Dow SW, Christopher Orton E. Immuno-proteomic identification of bovine pericardium xenoantigens. Biomaterials2008;29:3514e20.
[25] National Research Council. Institute of Laboratory Animal ResourcesCommission on Life Sciences. Guide for the care and use of laboratory animals.Washington DC: National Academy Press; 1996.
[26] Sanchez DM, Gaitan DM, Leon AF, Mugnier J, Briceno JC. Fixation of vasculargrafts with increased glutaraldehyde concentration enhances mechanicalproperties without increasing calcification. ASAIO J 2007;53:257e62.
[27] Ling Y. Uniaxial true stress-strain after necking. AMP J Tech 1996;5:37e48.[28] Woessner JF. The determination of hydroxyproline in tissue and protein
samples containing small proportions of this imino acid. Arch Biochem Bio-phys 1961;93:440e7.
[29] Gilbert TW, Sellaro TL, Badylak SF. Decellularization of tissues and organs.Biomaterials 2006;27:3675e83.
[30] Masutani H, Ueda S, Yodoi J. The thioredoxin system in retroviral infectionand apoptosis. Cell Death Differ 2005;12:991e8.
[31] De Bernardez Clark E. Refolding of recombinant proteins. Curr Opin Bio-technol 1998;9:157e63.
[32] Roussel A, Cohen N, Holland P, Taliaoferro L, Green R, Benson P, et al. Alter-ations in acid-base balance and serum electrolyte concentrations in cattle:632 cases (1984e1994). J Am Vet Med Assoc 1998;212:1769e75.
[33] Merryman WD, Engelmayr JGC, Liao J, Sacks MS. Defining biomechanicalendpoints for tissue engineered heart valve leaflets from native leafletproperties. Prog Ped Card 2006;21:153e60.
[34] Thubrikar M. The aortic valve. Boca Raton: CRC Press, Inc;; 1990.[35] Vesely I. Theroleof elastin inaorticvalvemechanics. J Biomech1997;31:115e23.[36] Stradins P, Lacis R, Ozolanta I, Purina B, Ose V, Feldmane L, et al. Comparison
of biomechanical and structural properties between human aortic andpulmonary valve. Eur J Cardiothorac Surg 2004;26:634e9.
[37] O’Brien M, Goldstein S, Walsh S, Black K, Elkins R, Clarke D. The SynerGraftvalve: a new acellular (nonglutaraldehyde-fixed) tissue heart valve forautologous recellularization first experimental studies before clinicalimplantation. Semin Thorac Cardiovasc Surg 1999;11:194e200.
[38] Kasimir MT, Rieder E, Seebacher G, Silberhumer G, Wolner E, Weigel G, et al.Comparison of different decellularization procedures of procine heart valves.Int J Artif Organs 2003;26:421e7.
[39] Schenke-Layland K, Vasilevski O, Opitz F, König K, Riemann I, Halbhuber KJ,et al. Impact of decellularization of xenogeneic tissue on extracellular matrixintegrity for tissue engineering of heart valves. J Struct Biol 2003;143:201e8.
[40] Stavnezer J, Guikema JEJ, Schrader CE. Mechanism and regulation of classswitch recombination. Annu Rev Immunol 2008;26:261e92.
[41] Meyer SR, Chiu B, Churchill TA, Zhu L, Lakey JRT, Ross DB. Comparison of aorticvalve allograft decellularization techniques in the rat. J Biomed Mater Res A2006;79A:254e62.
Related Documents











