REVIEW The role of markers of bone remodeling in multiple myeloma Evangelos Terpos * , Marianna Politou, Amin Rahemtulla Department of Hematology, Faculty of Medicine, Imperial College London, Hammersmith Hospital, Du Cane Road, W12 0NN London, UK Summary Osteolytic bone disease is a frequent complication of multiple mye- loma, resulting in skeletal complications that are a significant cause of morbidity and mortality. A characteristic feature of myeloma bone disease is that the lesions rarely heal and bone scans are often negative in myeloma patients who have exten- sive lytic lesions, offering very little in the follow-up of bone disease. X-rays are also of limited value in monitoring bone destruction during anti-myeloma or anti-resorp- tive treatment. Biochemical markers of bone turnover, such as N- and C-terminal cross-linking telopeptide of type I collagen (NTX, CTX/ICTP, respectively), and newer ones such as the tartrate resistant acid phosphatase isoform 5b, provide information on bone dynamics that in turn may reflect disease activity in bone. Sev- eral studies have shown bone markers to be elevated in myeloma patients and reflect the extent of bone disease, while in some of them bone resorption markers correlate with survival. These markers may also be helpful in identifying those patients likely to respond to bisphosphonate treatment, and monitoring the effec- tiveness of bisphosphonate therapy in the management of myeloma bone disease. This review attempts to summarize the existing data for the role of markers of bone remodeling in assessing the extent of bone destruction in myeloma and monitoring bone turnover during specific anti-myeloma treatment. We also discuss some novel markers that may be of particular interest in the near future. c 2004 Elsevier Ltd. All rights reserved. KEYWORDS Myeloma; Bone markers; N-terminal cross-linking telopeptide of type I collagen (NTX); C-terminal cross-linking telopeptide of type I collagen generated by MMPs (ICTP, CTX-MMP); Pyridinoline; Deoxypyridinoline; Bone-specific alkaline phosphatase; Osteocalcin; Tartrate resistant acid phosphatase isoform 5b; Bone disease Introduction Multiple Myeloma (MM) is a clonal plasma cell disor- der characterized by bone destruction, immunode- ficiency, and renal impairment. Nearly 3500 people in the UK are diagnosed with MM each year. This ac- counts for 10% of blood cancers and 1% of all can- cers. More than 55% of patients who present with MM are aged 60 or older, while less than 3% of MM occurs in patients younger than 40 years; med- ian survival ranges between 3.5 and 5 years. 1,2 A cure in MM appears to be achievable only by alloge- neic transplantation, which is not the treatment of 0268-960X/$ - see front matter c 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.blre.2004.06.001 * Corresponding author. Tel.: +44-208-383-8546; fax: +44- 208-742-9335. E-mail address: [email protected] (E. Terpos). Blood Reviews (2005) 19, 125–142 www.elsevierhealth.com/journals/blre

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Blood Reviews (2005) 19, 125–142
www.elsevierhealth.com/journals/blre
REVIEW
The role of markers of bone remodelingin multiple myeloma
Evangelos Terpos*, Marianna Politou, Amin Rahemtulla
Department of Hematology, Faculty of Medicine, Imperial College London,Hammersmith Hospital, Du Cane Road, W12 0NN London, UK
Summary Osteolytic bone disease is a frequent complication of multiple mye-loma, resulting in skeletal complications that are a significant cause of morbidityand mortality. A characteristic feature of myeloma bone disease is that the lesionsrarely heal and bone scans are often negative in myeloma patients who have exten-sive lytic lesions, offering very little in the follow-up of bone disease. X-rays are alsoof limited value in monitoring bone destruction during anti-myeloma or anti-resorp-tive treatment. Biochemical markers of bone turnover, such as N- and C-terminalcross-linking telopeptide of type I collagen (NTX, CTX/ICTP, respectively), andnewer ones such as the tartrate resistant acid phosphatase isoform 5b, provideinformation on bone dynamics that in turn may reflect disease activity in bone. Sev-eral studies have shown bone markers to be elevated in myeloma patients andreflect the extent of bone disease, while in some of them bone resorption markerscorrelate with survival. These markers may also be helpful in identifying thosepatients likely to respond to bisphosphonate treatment, and monitoring the effec-tiveness of bisphosphonate therapy in the management of myeloma bone disease.This review attempts to summarize the existing data for the role of markers of boneremodeling in assessing the extent of bone destruction in myeloma and monitoringbone turnover during specific anti-myeloma treatment. We also discuss some novelmarkers that may be of particular interest in the near future.
�c 2004 Elsevier Ltd. All rights reserved.
KEYWORDSMyeloma;Bone markers;N-terminal cross-linkingtelopeptide of type Icollagen (NTX);C-terminal cross-linkingtelopeptide of type Icollagen generated byMMPs (ICTP, CTX-MMP);Pyridinoline;Deoxypyridinoline;Bone-specific alkalinephosphatase;Osteocalcin;Tartrate resistantacid phosphataseisoform 5b;Bone disease
0d
2
Introduction
Multiple Myeloma (MM) is a clonal plasma cell disor-der characterized by bone destruction, immunode-
268-960X/$ - see front matter �c 2004 Elsevier Ltd. All rights reseoi:10.1016/j.blre.2004.06.001
* Corresponding author. Tel.: +44-208-383-8546; fax: +44-08-742-9335.
E-mail address: [email protected] (E. Terpos).
ficiency, and renal impairment. Nearly 3500 peoplein the UK are diagnosed with MM each year. This ac-counts for 10% of blood cancers and 1% of all can-cers. More than 55% of patients who present withMM are aged 60 or older, while less than 3% ofMM occurs in patients younger than 40 years; med-ian survival ranges between 3.5 and 5 years.1,2 Acure in MM appears to be achievable only by alloge-neic transplantation, which is not the treatment of
rved.

126 E. Terpos et al.
choice for the majority of patients due to high rateof transplant-related mortality.3 A major clinicalfeature of MM is the presence of osteolytic bonedisease and generalized osteoporosis, which canlead to severe bone pain, pathologic fractures, spi-nal cord compression, and hypercalcemia. Over85% of myeloma patients have bone disease. It thusrepresents a major cause of morbidity and mortal-ity.4 Progression of skeletal disease is often not af-fected by chemotherapy even in respondingpatients.5 The mechanisms of bone destruction ap-pear to be related to increased osteoclastic boneresorption, which is not accompanied by a compa-rable increase in bone formation.6 Thus, a charac-teristic feature of myeloma bone disease is thatthe lesions rarely heal even when the patients arein complete remission.7 This finding is in keepingwith the observation that bone scans are often neg-ative in myeloma patients who have extensive lyticlesions, and offer very little in the follow-up ofbone disease in these patients.8 Furthermore, a re-cent study has shown that sequential measurementof bone mineral density (BMD) using DEXA-scansproduced heterogeneous local BMD changes andthe available data do not support routine use ofsequential DEXA-scans in MM.9 The bone diseasein MM is usually assessed by X-rays of the skeleton.X-rays are useful in the diagnosis of osteolytic le-sions, but do not give any dynamic information onthe ongoing bone resorption. In contrast, biochem-ical markers of bone metabolism specifically re-flect bone resorption or bone formation rates andare strongly affected by the processes active inmyeloma bone disease. Therefore, biochemicalmarkers of bone remodeling are used more oftennowadays in an attempt to improve monitoring ofbone destruction in MM.
In addition, strategies that target osteoclastactivation and proliferation represent an importantapproach to the management of myeloma bone dis-ease. Bisphosphonates consist of a heterogeneousgroup of agents that affect bone metabolism andregulation of calcium homeostasis, mainly throughan inhibitory effect on osteoclasts.10 Several stud-ies have demonstrated their efficacy in myelomabone disease and therefore they are included inall therapeutic regimens for stage II/III myelomapatients.11,12 Biochemical markers of bone resorp-tion and formation have also been used to followup myeloma bone disease during bisphosphonatesadministration. This review attempts to summarizethe existing data for the role of markers of boneremodeling in assessing the extent of bone destruc-tion in myeloma and monitoring bone turnover dur-ing specific anti-myeloma chemotherapy orbisphosphonates administration. We also discuss
novel markers that may be of particular interestin the near future.
Pathogenesis of bone diseasein multiple myeloma
Although the mechanisms responsible for thedevelopment of myeloma bone disease currentlyremain unclear, several studies have begun toshed new light on this process. Histomorphometricstudies have demonstrated that myeloma bonedestruction is related to increased osteoclasticactivity, which is not accompanied by a compara-ble increase in osteoblast formation. This uncou-pling of resorption and formation leads to rapidbone loss.13 A number of cytokines and growthfactors that are produced either by myeloma cellsor by stromal cells, due to interactions betweenthem, have been implicated in the increase inosteoclast formation and activity in MM. Theadherence of myeloma cells to bone marrow stro-mal cells results in enhanced production of cytok-ines, such as interleukin-6 (IL-6), interleukin 1-b(IL-1b), interleukin 11 (IL-11), tumor necrosis fac-tors a and b (TNFa, TNFb), basic fibroblast growthfactor (bFGF), macrophage-colony stimulating fac-tor (M-CSF), which stimulate human osteoclastformation.4 IL-6 also acts as a survival factor formyeloma cells.14 Furthermore, stromal cells alsoproduce receptor activator of nuclear factor j-Bligand (RANKL), a member of the tumor necrosisfactor (TNF) gene family. Following activation ofthe cellular receptor RANK on osteoclasts by itsligand, RANKL, differentiation, proliferation, andsurvival of osteoclasts is enhanced, osteoclastfusion and activation is promoted, and osteoclas-tic apoptosis is suppressed, leading to a dramaticincrease of the number and activity of osteo-clasts.15,16 In addition, production of osteoproteg-erin (OPG), a soluble decoy receptor of RANKLproduced by marrow stromal cells, is suppressedthrough the above interactions and has beenfound to be reduced in patients with MM.17–19
The mechanisms through which OPG levels are de-creased have not been clearly defined yet, but astudy by Standal et al.20 has shown that OPG isbound, internalized, and degraded by the mye-loma cells through CD138. The ratio RANKL/OPGis reversed in myeloma, leading to osteoclast acti-vation and bone destruction.15,19,21–24 Several re-ports have suggested that myeloma cells alsoproduce RANKL and that the expression of RANKLby human myeloma cells mediates osteoclast for-mation in vitro and correlates with bone destruc-

The role of markers of bone remodeling in multiple myeloma 127
tion in vivo.25–27 However, other investigatorshave not been able to detect RANKL expressionon myeloma cells.21 The amount of RANKL pro-duced by myeloma cells is rather small to stimu-late osteoclast formation by itself and it may besufficient only to prevent osteoclast apoptosis.4,28
Several cytokines produced by both myeloma andstromal cells, such as IL-3, IL-6, IL-11 exerts theireffect through RANKL/RANK/OPG pathway, induc-ing osteoclastogenesis.4,10,29,30
Myeloma cells also produce large amounts ofmacrophage inflammatory protein-1a (MIP-1a),which is a low molecular weight chemokine thatbelongs to the RANTES family, and activates theosteoclasts directly.31–34 MIP-1a levels are ele-vated in both bone marrow plasma and serumof patients with active MM and correlate withthe presence of lytic lesions.35,36 High concentra-tions of MIP-1a were mainly found in patientswith advanced and active disease. Furthermore,MIP-1a has also been shown to increase RANKLexpression on bone marrow stromal cells, thusinducing further bone destruction.33 The majorrole of both RANK/RANKL and MIP-1a pathwaysin myeloma bone disease has been establishedin murine models. The inhibition of RANKL andMIP-1a activity results in markedly decreasedbone destruction and a significant reduction intumor burden.37–40
As mentioned above, in MM the increased boneformation is accompanied by suppression in oste-oblast function. Osteoblasts in active myelomaare functionally exhausted and promptly undergoapoptosis in the presence of myeloma cells frompatients with severe bone disease.41 A recentstudy by Tian et al.,42 reported that myelomacells produce dickkopf 1 (DKK1) protein, an inhib-itor of Wnt signaling pathway, which is crucial forosteoblast differentiation. In this study, marrowplasma from patients with MM that contained>12 ng/ml of DKK1 inhibited osteoblast differenti-ation. Furthermore, gene expression levels ofDKK1 correlated with the extent of bone disease.The presence of a soluble factor produced bymyeloma cells that suppresses osteoblast differ-entiation is a very important finding, which, how-ever, does not entirely explain why myelomabone lesions do not heal when the patients arein complete remission. There may possibly be amore long-lasting change in the marrow microen-vironment that results in an inability of osteo-blast precursors to differentiate, even in theabsence of myeloma cells. However, this hypoth-esis remains to be proven.
Fig. 1 summarizes the currently available dataon osteoclast activation in MM reflecting the role
of myeloma microenvironment in the developmentof myeloma-related bone disease.
Markers of bone remodeling
Throughout life bone undergoes continuousremodeling with removal of old bone andreplacement with new bone. Bone turnover is al-ways initiated by osteoclasts eroding a mineral-ized surface. This process is followed by therecruitment of successive teams of osteoblaststo the outer edge of the erosion cavity that se-crete new bone matrix and gradually fill in theresorption cavity.43 Resorption of old bone andformation of new bone are balanced under nor-mal conditions. In MM, there is a pronouncedimbalance in these processes: increased activa-tion of osteoclasts and suppression of osteoblastfunction. Over the past two decades, the isola-tion and characterization of cellular and extracel-lular components of the skeletal matrix haveresulted in the development of biochemicalmarkers that specifically reflect either bone for-mation or bone resorption. Most of the tradi-tional and new markers of bone resorptionmeasure the collagen degradation products fromosteoclast activity and include urinary hydroxypr-oline, hydroxylysine glycosides, total or freepyridinoline cross-links, and cross-linked N- orC-telopeptides.44 A novel marker, serum tartrateresistant acid phosphatase isoform type 5b(TRACP-5b) which is an enzyme secreted by acti-vated osteoclasts and bone sialoprotein (BSP), anon-collagenous protein have been demonstratedto reflect bone resorptive processes.45,46 The for-mation markers are direct or indirect products ofactive osteoblasts that enter into the circulation(Fig. 2). These include serum bone-specific alka-line phosphatase, osteocalcin and type-I procolla-gen peptides.47 All the above markers aredepicted in Tables 1 and 2. Fasting urinary cal-cium, used by some investigators to measurebone resorption, is not discussed here as it hasbeen found to be of limited value in MM.48
Biochemical markers of bone remodeling arenoninvasive, comparatively inexpensive, and,when applied and interpreted correctly, helpfultools in the assessment of bone diseases. How-ever, factors that affect the markers’ levels,including circadian rhythmicity, diet, age, gen-der, renal function and drugs, should be clearlydefined and appropriately adjusted whenever pos-sible. Another important issue is that biochemicalindices reflect the total bone turnover and give

Figure 1 The adherence of MM cells to bone marrow stromal cells (BMSC)/osteoblasts enhances the production ofRANKL, M-CSF, and other cytokines with OAF activity (IL-6, IL-11, IL-1b, TNFs, bFGF, IGFs), while it suppresses theproduction of OPG (the decoy receptor of RANKL). The above cytokines also modify the bone marrow microenviron-ment, up-regulating the RANKL expression and secretion by both BMSC and osteoblasts. Furthermore, myeloma cellsproduce MIP-1a, HGF, and IL-3 that enhance the proliferation and differentiation of osteoclast precursors, whilemyeloma cells possibly express both transmembrane and soluble RANKL. OPG binds both surface and soluble RANKLinhibiting osteoclast development and bone resorption. Syndecan 1 (CD138) expressed on the surface of, and secretedfrom, the myeloma cells can bind soluble OPG, thus preventing its inhibitory effect on RANKL function. Therefore theratio of RANKL/OPG is increased, leading to osteoclast differentiation, proliferation and activation. Myeloma cells alsoproduce DKK1 protein, which inhibits stromal cell differentiation to osteoblasts, thereby decreasing bone formation.These interactions result in increased bone resorption, as depicted by the increased levels of bone resorption markers(NTX, ICTP, CTX, TRACP-5b).
Figure 2 Osteoblast activation: Through stimulators,the osteoblasts produce enzymes, such as ALP and matrixconstituents, such as collagen type I and osteocalcin.Bone ALP and osteocalcin can be measured in the serumwhile degradation products of collagen type I can bemeasured in both serum and urine.
128 E. Terpos et al.
little information about the function of individualgroups of osteoclasts. All these issues are dis-cussed below.
Biochemistry
Resorption markers
Hydroxyproline and hydroxylysineHydroxyproline (Hyp) is formed in the cell from theposttranslational hydroxylation of proline. Hyp isthe predominant amino acid of collagens, compris-ing about 13% of these proteins. Hydroxylysine(Hyl) is another amino acid essentially unique tocollagenous proteins. Bone is the primary store ofcollagen in the body, but both Hyp and Hyl are pre-sent in essentially all tissues and all genetic typesof collagen. The Hyp released during collagen deg-radation is primarily metabolized in the liver andsubsequently excreted in the urine. However, onlyabout 10% of Hyp-containing products from

Table 1 Markers of bone resorption.
Marker Abbreviationa Tissue of origin Analytical method Analytical specimen
Hydroxyproline Hyp All tissues and all genetic types of collagen Colorimetric, assay, HPLC UrineHydroxylysine Hyl All tissues and all genetic types of collagen Reversed-phase HPLC UrineGalactosyl–hydroxylysine Gal–Hyl Both Gal–Hyl and Glc–Gal–Hyl appears
to be specific for bone collagen degradationReversed-phase HPLC Urine
Glucosyl–galactosyl–hydroxylysine Glc–Gal–HylPyridinoline PYD Bone, cartilage, tendon, blood vessels HPLC, ELISA UrineDeoxypyridinoline DPD Bone, dentin RIA Urine (free DPD can be
also measured in serumor plasma)
N-terminal cross-linkingtelopeptide of type-I collagen
NTX All tissues containing type-I collagen ELISA, RIA Urine, serum
C-terminal cross-linkingtelopeptide of type-I collagen
CTX All tissues containing type-I collagen ELISA, RIA Urine, serum (b-form only)
C-terminal cross-linkingtelopeptide of type-Icollagen generated by MMPs
CTX-MMP or ICTP All tissues containing type-I collagen RIA Serum
Tartrate resistant acidphosphatase isoform 5b
TRACP-5b Bone (osteoclasts) Colorimetric RIA, ELISA Serum, plasma
Bone sialoprotein BSP Bone, dentin, hypertrophic cartilage,cancer cells
RIA, ELISA Serum
a According to the Bone Marker Nomenclature by the Committee of Scientific Advisors of the International Osteoporosis Foundation.115
Therole
ofmarke
rsofboneremodelin
gin
multip
lemye
loma
129

Figure 3 Fibrils of collagen showing the N- and C-terminal ends bonding to helical areas of adjacent fibrilsby PYD and DPD.
Table 2 Markers of bone formation.
Marker Abbreviationa Tissue of origin Analytical method Analytical specimen
Osteocalcin (or bone gla-protein) OC Bone, platelets RIA, ELISA, IRMA SerumBone alkaline phosphatase Bone ALP Bone ELISA, IRMA,
colorimetric assaySerum
Procollagen type-IN-propeptide
PINP Bone, soft tissue, skin RIA, ELISA Serum
Procollagen type-IC-propeptide
PICP Bone, soft tissue, skin RIA, ELISA Serum
a According to the Bone Marker Nomenclature by the Committee of Scientific Advisors of the International OsteoporosisFoundation.115
130 E. Terpos et al.
collagen breakdown are excreted in the urine.43
The small pool of urinary Hyp originates from theN-propeptide of type I collagen. Normal ingestionof gelatin or collagen-rich foods such as meat canincrease the level of urinary Hyp, and the urinarypeptides containing Hyp from endogenous collagenbreakdown are indistinguishable from the dietarypeptides.49,50 This is a major disadvantage for theuse of this marker as a specific index of boneresorption.
Hyl is glycosylated and two glycosides areformed, galactosyl–hydroxylysine (Gal–Hyl) andglucosyl–galactosyl–hydroxylysine (Glc–Gal–Hyl),which also appear in the urine.51 While Hyl andits glycosides are less abundant than Hyp in bonecollagen, certain properties make Hyl theoreticallya better marker of bone turnover than urinary Hyp.Gal–Hyl is not metabolized and not influenced bydietary factors.52 In normal urine, 80% of the totalHyl is in the form of Hyl, 10% is free and unglycos-ylated, and the remainder is peptide-bound, whichsuggests that free Hyl is largely metabolized andnot excreted.53 However, in addition to all struc-tural collagens, both Hyp and Hyl are also foundin certain serum proteins, such as the C1q compo-nent of complement.54 This disadvantage, in com-bination with the effect of age and the circadianrhythm (both have their peak excretion after mid-night), make them less specific indices of boneresorption and therefore they have been largelyreplaced by newer markers.
Pyridinoline and deoxypyridinolinecross-links of type I collagenIn the last decade, collagen cross-links haveevolved as promising markers of bone resorption.Pyridinoline (PYD) and deoxypyridinoline (DPD)are formed by the enzymatic action of lysyl oxidaseon lysine and Hyl. Newly deposited collagen fibrilsin the extracellular matrix are stabilized by cross-links formed by the action of lysyl oxidase on lysineand Hyl residues in telopeptide domains of the col-
lagen molecules. The resulting aldehydes condensewith Hyl or lysyl residues on adjacent collagen mol-ecules to form divalent cross-links, which can ma-ture by further condensation with telopeptidealdehydes to the trivalent structures DPD andPYD.43 PYD and DPD act as mature cross-links intype I collagen of all major connective tissues.55
These include type I collagen of bone, dentin, ten-don, vascular walls, muscle, intestine, etc. Fig. 3depicts the basic cross-linking of collagen fibrilsthat construct type I collagen, the protein compris-ing most of the organic matrix of the bone. Each ofthe fibrils contains aminoterminal and carboxyter-minal ends that are termed N-telopeptides (NTX)and C-telopeptide (CTX), respectively. In type Icollagen, these ends are each linked to a helicalportion of a nearby molecule by a PYD or DPDcross-link. The pyridinoline cross-links occur essen-tially at two intermolecular sites in the collagen fi-bril: two amino-telopeptides are linked to a helicalsite at or near residue 930 and two carboxy-telopeptides to helical residue 87.56 DPD is derivedfrom two hydroxylysines and one lysine residue,while PYD is derived from three hydroxylysine res-idues. In all tissues, PYD predominates, while DPDis the minor component. The products of collagendegradation by osteoclasts include NTX and CTXfragments of various sizes, still attached to helicalportions of a nearby molecule by a pyridinium

The role of markers of bone remodeling in multiple myeloma 131
cross-link, that insert into the circulation.57 Withadditional degradation in the liver and kidney thefragments are finally broken down to their constit-uent amino acids, and the pyridiniums, PYD andDPD. Although it is possible that soft tissues con-tribute to the normal excretion of DPD and PYD,bone represents the major reservoir of total colla-gen in the body and turns over faster than most ma-jor connective tissues. In contrast to Hyp and Hyl,the measurement of urine DPD and PYD is not influ-enced by the degradation of newly synthesized col-lagen fibrils or by dietary collagen intake. Thepyridinolines are present in the diet, but unlikeHyp, they are not absorbed.58 Furthermore, unlikeHyp, the pyridinoline amino acids are fully ex-creted with no known pathway of metabolicdegradation.59
Amino- and carboxy-terminal cross-linkingtelopeptide of type I collagenDuring collagen type I degradation by osteoclasts,N- and C-terminal peptide fragments (NTX andCTX, respectively) are released into the circula-tion.57 These fragments represent a spectrum ofproteins having different sizes, as shown in Fig. 4.The majority of these is relatively small and passesthrough the glomerulus into the urine. NTX are re-garded to be specific for bone tissue breakdown asother tissues comprised of type I collagen, e.g.,skin, are not actively metabolized by osteoclasts,and, therefore, different types of fragments areformed during breakdown of non-skeletal tissues.60
An ELISA method has been developed to recognizea discrete pool of NTX isolated from urine.61 Themonoclonal antibody recognizes the a2 chain N-telopeptide fragment. This fragment contains thepyridinium cross-links, but this assay does not rec-ognize the PYD and DPD per se. This implies bonespecificity since the pyridinoline cross-link in boneprimarily involves the a2 chain whereas in othertissues the a1 chain predominates.61 NTX, containsthe cross-linked a2 N-telopeptide sequence,QYDGKGVG, which is a product of osteoclastic pro-teolysis and in which lysine (K) is embodied in a tri-valent cross-linkage.62 Collagen must be broken
15% 38%
40% 7%
Figure 4 Distribution of type I collagen fr
down to small cross-linked peptides that containthis exact sequence before the antibody can bindto the NTX antigen. This suggests that the NTX pep-tide is a direct product of osteoclastic proteolysis,does not require further metabolism in the liver orkidney for generation, and is rapidly cleared by thekidney.63
Other assays have also been developed for themeasurement of epitopes associated with the C-terminal telopeptide of type I collagen (a-CTX, b-CTX, ICTP) in serum and urine.64–66 Due to bonespecificity and their unique characteristics NTX,ICTP, and CTX have almost totally replaced theuse of older resorption indices in the diagnosticassessment of bone diseases.
Tartrate resistant acid phosphataseisoform type 5bTartrate resistant acid phosphatase isoform type5b (TRACP-5b) is a novel marker of bone resorptionwhich has been in use over the last 3–4 years withvery encouraging results. TRACP is produced byboth osteoclasts and activated macrophages andsubsequently is secreted into the circulation.67
Two forms of TRACP circulate in human serum,macrophage-derived TRACP-5a and osteoclast-derived TRACP-5b.68–70 The only structural differ-ence between TRACP-5a and 5b is that the formercontains sialic acid residue(s) that are not foundin TRACP-5b.71 In human serum, TRACP-5b circu-lates in a large complex that contains a2-micro-globulin and calcium.72 Osteoclasts secreteTRACP-5b into the blood circulation as a catalyti-cally active enzyme that is inactivated and de-graded to fragments in the circulation. Thus, allcatalytically active TRACP-5b molecules measuredin the serum are freshly liberated from the osteo-clasts, providing a sensitive resorptive index.45,73
Formation markers
OsteocalcinOsteocalcin is one of the most abundant non-colla-genous bone proteins, produced by osteoblasts,odontoblasts, and hypertrophic chondrocytes. Most
Free (unbound)
Protein-bound forms>3500g/mole
Protein bound forms550 to 1000 g/mole
Protein-bound forms1000 to 3500 g/mole
agments after osteoclastic degradation.

132 E. Terpos et al.
of the circulating osteocalcin is a product of oste-oblast activity and thus considered a marker ofbone formation. It is a small protein of 49 aminoacids and in most species contains three residues(at 17, 21, and 24) of c-carboxy glutamic acid, acalcium-binding amino acid.74 This vitamin K-de-pendent post-translational modification of newlysynthesized proteins results in c-carboxylation ofspecific glutamate residues. The reaction is compa-rable to the activation of vitamin K-dependentblood coagulation factors and is inhibited by warfa-rin.75 The human osteoblast produces an 11-kDamolecule consisting of a 23-residue hydrophobicsignal peptide, a 26-residue propeptide, and the49-residue mature protein. The pro-region containsa c-carboxylation recognition site homologous tocorresponding regions in the vitamin K-dependentclotting factors. After the hydrophobic regionscleaved by a signal peptidase, pro-osteocalcin isc-carboxylated. Subsequently, the propeptide isremoved and the mature protein is secreted.43 Inserum, osteocalcin is degraded so that both the in-tact peptide and fragments coexist in the circula-tion.76 Therefore, assays that evaluate bothintact osteocalcin and fragments are more accu-rate for the measurement of serum osteocalcin.
A fraction of newly synthesized osteocalcin is se-creted into the circulation, while during boneresorption osteocalcin is also degraded. Thus thereis some question whether osteocalcin should beconsidered an indicator of osteoblast activity or amarker of bone matrix metabolism or turnover.60
The human osteocalcin gene is located at thedistal long arm of chromosome 1.77 Various pro-moter elements contribute to basal expressionand osteoblast specificity. The gene is further mod-ulated by vitamin D and glucocorticoid responseelements.78,79 Although osteocalcin is present insignificant amounts in bone, dentin, and calcifiedcartilage, it has recently also been found in osteo-sarcomas, prostate, ovarian, lung and brain can-cer.80 Osteocalcin function has not clearlydefined yet. However, it is assumed that much ofthe newly synthesized protein is incorporated intothe bone matrix binding calcium. Serum levels ofosteocalcin are significantly influenced by gender,age, and renal function.76
Bone-specific alkaline phosphataseAlkaline phosphatase (ALP) is a ubiquitously ex-pressed, cell-membrane enzyme. ALP belongs tothe category of molecules that are localized to cellmembranes through a COOH-terminal glycan–phosphatidylinositol anchor, providing a basis forunderstanding the generation of different isoforms
observed in plasma. Isoforms produced by differen-tial cleavage or preservation of the glycan–phos-phatidylinositol anchor originate from differenttissues, such as liver, bone, intestine, spleen, kid-ney and placenta.81 Liver and bone (bALP) isoformsaccount for almost 95% of the total ALP activity inthe serum. Bone ALP is produced by osteoblastsand has been demonstrated in matrix vesiclesdeposited as ‘‘buds’’ derived from the cells mem-brane. These deposits seem to play an importantrole in bone formation.60 Bone ALP is produced inextremely high amounts during bone formationphase of bone turnover, and is, therefore, anexcellent indicator of total bone formationactivity.81
Type I procollagen propeptidesCollagen type I, a 300-kDa protein, makes up 90% ofthe organic bone matrix and is synthesized by oste-oblasts in the form of procollagen, having a molec-ular weight (MW) of 450-kDa. Extracellularprocessing of procollagen before fiber assembly in-cludes cleavage of the N- and C-terminal extensionpropeptides, that are termed procollagen type I N-and C-propeptide (PINP, and PICP, respectively),having a MW of 35 and 100 kDa, respectively.60
PINP is cleared via the scavenger endothelial sys-tem in the liver, and PICP via the mannose recep-tors on liver endothelial cells.82 Because thesepeptides are generated in a stoichiometric 1:1 ratiowith newly formed collagen molecules, their levelsin serum are considered an index of collagen syn-thesis and thus of bone formation.83 Serum levelsof the PICP have been demonstrated to correlatewith histomorphometric measures of bone forma-tion, and hormone replacement or bisphosphonatetherapy leads to a reduction in the circulating con-centration of this marker. Most recent studies,however, suggest that the PINP has a greater diag-nostic validity than PICP.84
Markers of bone remodelingin myeloma bone disease
Comparison with normal individuals andcorrelations with the extent of myelomabone destruction
Markers of both resorption and formation havebeen used in an attempt to better evaluate the ex-tent of bone disease in multiple myeloma and as-sess clinical correlations. Table 3 summarizes theavailable data for the use of markers of bone

Table 3 Studies describing the levels of markers of bone remodeling in MM patients and correlation with clinicaldata.
Authors–year Number ofpatients
Parameterstudied
Comparisonwith controls(symbolrefers to MMpatients)
Correlationwith extent ofbone disease
Correlationwith survival
Nawawi et al., 199685 17 DPD ›a NA NATRACP NSOC NSbALP NSPICP NS
Abildgaard et al., 199786 109 ICTP ›a NA YesOC NS YesbALP ›a NoPICP NS NoPIIINP ›a No
Withold et al., 199887 15 DPD, PYD ›a NA NAbALP, PICP fla
Carlson et al., 199988 73 DPD ›a Yes NoICTP ›a Yes YesOC NS No NoPICP NS No No
Woitge et al., 199966 18 PYD ›a NA NADPD ›a
NTX ›a
CTX ›a
BSP ›a
Fonseca et al., 200089 313 ICTP ›a Yes YesTRACP ›a No NoOC fla No NobALP NS Yes NoPICP NS No No
Terpos et al., 200090 62 NTX ›a NA NAOC fla
bALP fla
Woitge et al., 2001103 43 OC NS NA NAbALP fla
Corso et al., 200191 52 DPD ›a Yes NAOC fla NobALP NS No
Jakob et al., 200292b 57 DPD ›a Yes NoNTX NS No p=0.05ICTP ›a Yes Yes
Alexandrakis et al., 200293 38 PYD ›a Yes NADPD ›a YesNTX ›a Yes
(continued on next page)
The role of markers of bone remodeling in multiple myeloma 133
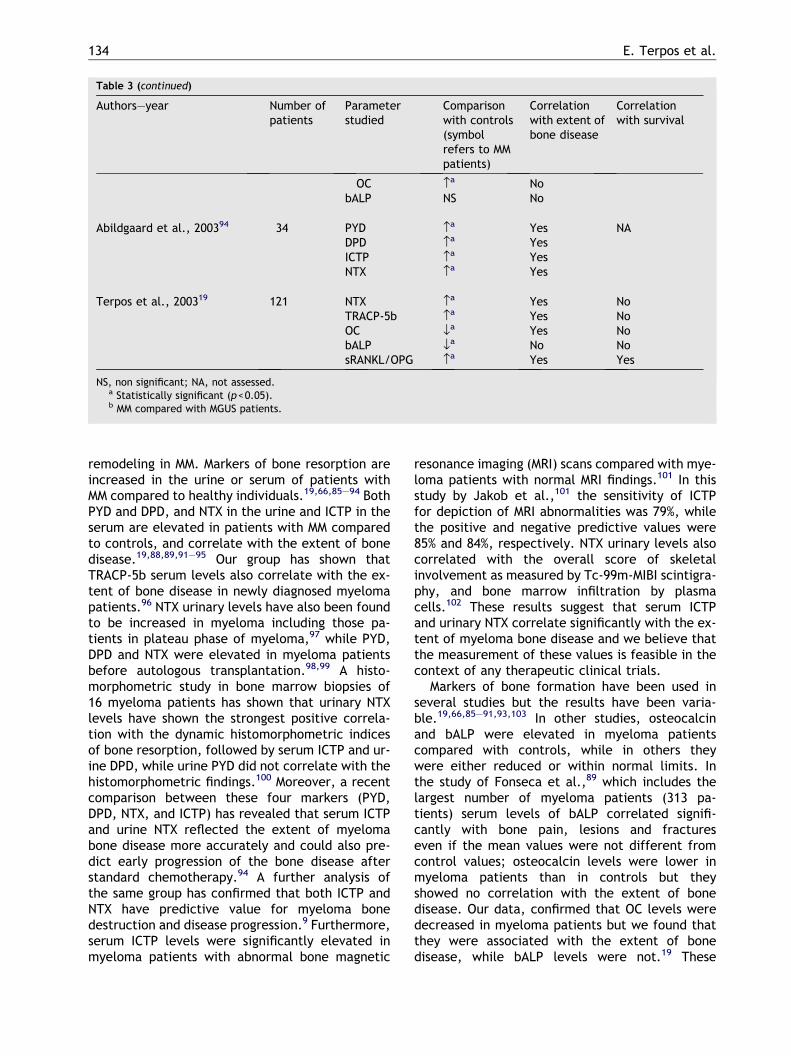
Table 3 (continued)
Authors–year Number ofpatients
Parameterstudied
Comparisonwith controls(symbolrefers to MMpatients)
Correlationwith extent ofbone disease
Correlationwith survival
OC ›a NobALP NS No
Abildgaard et al., 200394 34 PYD ›a Yes NADPD ›a YesICTP ›a YesNTX ›a Yes
Terpos et al., 200319 121 NTX ›a Yes NoTRACP-5b ›a Yes NoOC fla Yes NobALP fla No NosRANKL/OPG ›a Yes Yes
NS, non significant; NA, not assessed.a Statistically significant (p<0.05).b MM compared with MGUS patients.
134 E. Terpos et al.
remodeling in MM. Markers of bone resorption areincreased in the urine or serum of patients withMM compared to healthy individuals.19,66,85–94 BothPYD and DPD, and NTX in the urine and ICTP in theserum are elevated in patients with MM comparedto controls, and correlate with the extent of bonedisease.19,88,89,91–95 Our group has shown thatTRACP-5b serum levels also correlate with the ex-tent of bone disease in newly diagnosed myelomapatients.96 NTX urinary levels have also been foundto be increased in myeloma including those pa-tients in plateau phase of myeloma,97 while PYD,DPD and NTX were elevated in myeloma patientsbefore autologous transplantation.98,99 A histo-morphometric study in bone marrow biopsies of16 myeloma patients has shown that urinary NTXlevels have shown the strongest positive correla-tion with the dynamic histomorphometric indicesof bone resorption, followed by serum ICTP and ur-ine DPD, while urine PYD did not correlate with thehistomorphometric findings.100 Moreover, a recentcomparison between these four markers (PYD,DPD, NTX, and ICTP) has revealed that serum ICTPand urine NTX reflected the extent of myelomabone disease more accurately and could also pre-dict early progression of the bone disease afterstandard chemotherapy.94 A further analysis ofthe same group has confirmed that both ICTP andNTX have predictive value for myeloma bonedestruction and disease progression.9 Furthermore,serum ICTP levels were significantly elevated inmyeloma patients with abnormal bone magnetic
resonance imaging (MRI) scans compared with mye-loma patients with normal MRI findings.101 In thisstudy by Jakob et al.,101 the sensitivity of ICTPfor depiction of MRI abnormalities was 79%, whilethe positive and negative predictive values were85% and 84%, respectively. NTX urinary levels alsocorrelated with the overall score of skeletalinvolvement as measured by Tc-99m-MIBI scintigra-phy, and bone marrow infiltration by plasmacells.102 These results suggest that serum ICTPand urinary NTX correlate significantly with the ex-tent of myeloma bone disease and we believe thatthe measurement of these values is feasible in thecontext of any therapeutic clinical trials.
Markers of bone formation have been used inseveral studies but the results have been varia-ble.19,66,85–91,93,103 In other studies, osteocalcinand bALP were elevated in myeloma patientscompared with controls, while in others theywere either reduced or within normal limits. Inthe study of Fonseca et al.,89 which includes thelargest number of myeloma patients (313 pa-tients) serum levels of bALP correlated signifi-cantly with bone pain, lesions and fractureseven if the mean values were not different fromcontrol values; osteocalcin levels were lower inmyeloma patients than in controls but theyshowed no correlation with the extent of bonedisease. Our data, confirmed that OC levels weredecreased in myeloma patients but we found thatthey were associated with the extent of bonedisease, while bALP levels were not.19 These

The role of markers of bone remodeling in multiple myeloma 135
differences among the different studies may bedue to different study population and the differ-ent phase of bone turnover in each population.PICP values do not predict the extent of myelomabone disease.85–89 As shown in Table 3, we be-lieve that markers of bone formation may be ofsome value but may not necessarily reflect theextent of myeloma bone destruction.
Correlations with disease activity andsurvival
There are several studies in which markers of boneremodeling have shown strong correlation with thestage of myeloma. Serum levels of ICTP and urinaryvalues of NTX were higher in myeloma stage II/IIthan stage I disease.86,89,93 Abnormal high DPD uri-nary levels also correlated with advanced myelomastage.91,92 Our group has also reported in 121 newlydiagnosed myeloma patients, that stage of the dis-ease correlated with both TRACP-5b and NTX(p<0.0001, and p=0.014, respectively), showing aborderline correlation with OC (p=0.046) but nocorrelation with bALP (p=0.73).19 Fig. 5 depictsthe differences in NTX urinary levels among mye-loma patients at different disease stages, patientswith MGUS and normal individuals reported by ourgroup.104 The association of osteocalcin with stage
N =
GROUP
LGN
TX
3.0
2.5
2.0
1.5
1.0
.5
144
188
181
4045
MMMGUSControls
Parameter MM (n=42)
MGUS (n=40)
NTX (nm BCE/mM creatinine)
132.0 (25-571)
81.2 (19.1-418.6)
Figure 5 NTX urinary levels in MGUS patients
III disease has also been shown in the study ofWoitge et al.,103 while two other studies showedno correlation of either osteocalcin or bALP withstage or other clinical parameters.89,91 Markers ofboth formation and resorption also correlated withmarkers of disease activity, such as b2-microglobu-lin and IL-6, in several studies.19,89,91,100
However, it is clear that markers of bone resorp-tion and in particular, ICTP correlates with survivalin myeloma.86,88,89,92,105 Fonseca et al.89 haveshown that the median survival was 4.1 and 3.5years for patients with low and high ICTP levels,respectively (p=0.02). Furthermore, Jakobet al.92 have shown that ICTP is a prognostic factorfor overall survival (p<0.03), while urinary NTXshowed borderline significance (p=0.05). Turessonet al.105 has shown in an unselected series of 394myeloma patients that ICTP correlated with sur-vival in the univariate analysis; something thatwas not confirmed in the multivariate analysis.We had the same results in terms of associationof urinary NTX levels with survival; univariate anal-ysis showed a predictive value of NTX, but this wasnot confirmed by the multivariate analysis.19 Re-cently, Abildgaard et al.,9 using sequential meas-urements of both ICTP and NTX showed that bothmarkers predict progression of myeloma. No mar-ker of bone formation has been shown to correlatewith survival.
2
111
3111
MM Stage III Stage I/II
Controls (n=45)
Significance MM vs MGUS
Significance MGUS vs controls
37.9 (11-56.2)
P<0.001 P<0.0001
, myeloma patients and normal controls.104

Figure 6 Pamidronate (90 mg per month) in combination with chemotherapy (group I) results in a dramatic decreaseof urinary NTX levels compared to controls (group II in (a)) and patients receiving a combination of ibandronate (4 mgper month) and chemotherapy (group II in (b)). Pamidronate (group I) was also more effective than ibandronate (groupII) in reducing serum TRACP-5b levels in MM patients (c).90,108
136 E. Terpos et al.
Bone markers for monitoringbisphosphonates or anti-myeloma treatment
Biochemical markers of bone turnover have beenused in patients with MM in order to monitor bis-phosphonate treatment and identify who willbenefit the most from bisphosphonate therapy.Both ICTP and NTX have shown a dramatic de-crease after clodronate, pamidronate or zoled-ronic acid administration, in myeloma patients,confirming the strong anti-resorptive activity of
these agents that consist part of anti-myelomatherapy in patients with bone lesions.87,90,97,106–108 Elomaa et al.106 have shown that clodronateadministration resulted in a significant decreaseof PINP and ICTP in 244 myeloma patients com-pared to the control group; furthermore highbaseline ICTP, PINP and ALP levels indicated apoor prognosis. Our group has shown that pamidr-onate in combination with chemotherapy reducesurine NTX levels significantly, compared withmyeloma patients who receive chemotherapy

Median values of NTX post-ASCT
-50
0
50
100
150
200
Months post-ASCT
NT
X (
nM
BC
E/m
M c
reat
inin
e)
Median values of TRACP-5b post-ASCT
0
1
2
3
4
5
6
7
8
1 2 3 4 5 6 7 8 9 10 11 12 13
Months post-ASCT
TR
AC
P-5
b (
U/L
)
(a)
(b)
Figure 7 (a) Median values (±SD) of NTX post ASCT. (b) Median values (±SD) of TRACP-5b post ASCT.99
The role of markers of bone remodeling in multiple myeloma 137
alone. Fig. 6(a) depicts this difference betweenthe two groups of patients.90 Furthermore, inaddition to the inhibition of osteoclastic activity,pamidronate in combination with interferon-awas shown to induce bone formation in patientswith myeloma in the plateau phase.97 In thispoint it is reasonable to note that bALP and/orOC rises after successful treatment of MM inthose patients with bone disease as a sign of re-pair. Zoledronic acid, which is a potent inhibitorof osteoclastic activity has also shown a reduc-tion in NTX levels at a dose of 4 mg per monthin myeloma patients,107 while we reported thatpamidronate at a dose of 90 mg could reduceboth NTX and TRACP-5b levels more effectivelythan 4 mg of ibandronate (Fig. 6(b) and (c)).108
In addition, bone remodeling markers seem tobe normalized after high dose chemotherapy withautologous stem cell support (Fig. 7).98,99
However, we believe that bisphosphonatetreatment has to be continued post ASCT evenif all markers of bone remodeling have beennormalized due to the possible antimyeloma ef-fect of bisphosphonates.109
A recent study has shown that high levels of ICTPand NTX correlated with an increased risk for earlyprogression of bone lesions during standard melph-alan–prednisolone treatment in myeloma. This
study suggests that ICTP and NTX are clinically use-ful for identifying patients with increased risk ofearly progression of bone disease and thereforeof disease progression.9
Future developments
RANKL/Osteoprotegerin
RANKL and OPG play a crucial role in the develop-ment of myeloma bone disease (Fig. 1). It has beendemonstrated that patients with advanced pros-tate cancer have significantly higher serum OPGlevels than patients with early stage disease.110 Inanother study, significant differences in OPG serumlevels were found between prostate cancer pa-tients with bone metastases compared withhealthy controls, patients with non-metastasizedcancer, and patients with benign hyperplasia. Evenwhen compared with NTX serum levels, OPG wasfound to be of higher sensitivity in detecting bonemetastases than NTX.111 However, serum levels ofsoluble RANKL have no correlation with bone dis-ease in prostate cancer.112
OPG levels have been found to be reduced inmyeloma patients, while the ratio of sRANKL/OPG

83281045N =
Bone Disease
CBAControl
Log
sRA
NK
L/O
PG
2.0
1.5
1.0
.5
0.0
-.5
-1.0
-1.5
68
3612130
52
159149134158
LGRATIO2.01.51.0.50.0-.5-1.0-1.5
LGT
RA
CP
-5b
1.8
1.6
1.4
1.2
1.0
.8
.6
.4
.2
0.0
LGRATIO2.01.51.0.50.0-.5-1.0-1.5
LGN
TX
2.8
2.6
2.4
2.2
2.0
1.8
1.6
1.4
1.2
(a)
(b) (c)
Figure 8 (a) Association between log sRANKL/OPG ratio and extent of bone disease as assessed by radiographicevaluation. Median values (range) of the sRANKL/OPG ratio for controls and patients were as follows: controls, 0.33(0.10–0.59); group A (no lytic lesions and/or osteoporosis), 0.57 (0.10–1.01); group B (1–3 lytic lesions), 0.84 (0.26–4.16); group C (>3 lytic lesions and/or fractures), 2.25 (0.43–44.04). Circled numbers referred to patient outliers.(b) Correlation between sRANKL/OPG ratio and TRACP-5b serum levels (r=0.87, p<0.0001). (c) Correlation betweensRANKL/OPG ratio and NTX urinary levels (r=0.69, p<0.0001).19
138 E. Terpos et al.
is increased.17–19 RANKL/OPG ratio has also re-ported to be increased in patients with cancerand severe osteolysis.24 Our group has shown thatthe ratio of sRANKL/OPG correlates with the extentof bone disease in myeloma, and with markersof bone resorption, such as NTX and TRACP-5b(Fig. 8).19 Recently, the results of a phase I studyusing a recombinant osteoprotegerin construct inpatients with multiple myeloma or breast cancer-related bone metastases were published. A singledose of AMGN-0007, a recombinant osteoproteg-erin construct, suppressed bone resorption as indi-cated by a rapid, sustained, and profound decreaseof urinary NTX levels, and the role of OPG as a
therapeutic tool in this area will be clarified in fu-ture clinical trials.113
Other potential markers reflecting bonedestruction in MMBone sialoprotein (BSP) is a phosphorylated 70–80kDA glycoprotein that accounts for 5–10% of thenon-collagenous bonematrix. The protein is amajorsynthetic product of active osteoclasts and odonto-blasts. In bone remodeling BSP is involved in theadhesion of bone resorbing cells to the extracellularbone matrix.114 BSP levels were associated withskeletal involvement and tumor cell burden and sur-vival in MM patients.46 The quantification of serum

The role of markers of bone remodeling in multiple myeloma 139
BSP may be a non-invasive method for the diagnosisand follow-up, and may improve the prognostic va-lue of conventional staging in myeloma.
Finally, the novel protein DKK1, a WNT path-way inhibitor, has been linked to the function ofosteoblasts in myeloma. Immunohistochemicalanalysis of bone marrow-biopsy specimens showedthat myeloma cells contained detectable DKK1.Elevated DKK1 levels in bone marrow plasmaand peripheral blood from patients with multiplemyeloma correlated with the gene-expressionpatterns of DKK1 and were associated with thepresence of focal bone lesions.42 Measurementof DKK1 protein in more patients and correlationwith markers of bone remodeling will show ifthe measurement of this protein will provide anaccurate reflection of the extent of bone diseasein myeloma.
Conclusions
Biochemical markers of bone remodeling arepotentially useful tools for the evaluation of theextent of bone disease and the follow-up of anti-resorptive treatment in patients with multiplemyeloma. In patients with established bone lyticlesions, most bone markers are abnormal, indicat-ing that these parameters faithfully reflect changesin bone metabolism associated with the malignantprocess. However, ICTP and NTX seem to be bettermarkers in reflecting both, the severity of bonedestruction and the efficacy of bisphosphonatetreatment in myeloma. TRACP-5b also seems tobe a useful marker but needs further evaluation.Although it is unlikely that a single marker of boneremodeling has sufficient diagnostic or prognosticvalue in myeloma bone disease, the combinationof these markers with other laboratory tests andimaging techniques is likely to improve the clinicalassessment of patients with myeloma.
There is a correlation between serum ICTP andurinary NTX and an increased risk of progressivebone disease, and consequently to myeloma pro-gression. Patients with smoldering myeloma andmyeloma patients with normal radiographs are sub-groups in which the assays might be particularlyuseful. Elevated levels of these resorption markersmay support a decision to initiate anti-myelomatherapy in patients with smoldering myeloma orof starting bisphosphonate treatment in patientswithout lytic bone disease. In the present erawhere new anti resorptive agents are in develop-ment (more potent bisphosphonates, osteoproteg-erin, Rank-Fc, etc.) these assays might be of
particular value. However, further trials areneeded to establish the predictive value of thesemarkers before introducing them into routine use.
References
1. Bataille R, Harousseau JL. New Engl J Med1997;336:1657–64.
2. Kyle RA, Gertz MA, Witzig TE, et al. Review of 1027patients with newly diagnosed multiple myeloma. MayoClin Proc 2003;78:21–33.
3. Barlogie B, Shaughnessy J, Tricot G, et al. Treatment ofmultiple myeloma. Blood 2004;103:20–32.
4. Roodman GD. Pathogenesis of myeloma bone disease.Blood Cell Mol Dis 2004;32:290–2.
5. Raje N, Anderson KC. Multiple myeloma. Curr Treat OptionOncol 2000;1:73–82.
6. Terpos E, Politou M, Rahemtulla A. New insights intothe pathophysiology and management of bone diseasein multiple myeloma. Br J Haematol 2003;123:758–69.
7. Callander NS, Roodman GD. Myeloma bone disease. SeminHematol 2001;38:276–85.
8. Mileshkin L, Blum R, Seymour JF, Patrikeos A, Hicks RJ,Prince HM. A comparison of fluorine-18 fluoro-deoxyglu-cose PET and technetium-99m sestamibi in assessingpatients with multiple myeloma. Eur J Haematol2004;72:32–7.
9. Abildgaard N, Brixen K, Eriksen EF, Kristensen JE, NielsenL, Heickendorff L. Sequential analysis of biochemicalmarkers of bone resorption and bone densitometry inmultiple myeloma. Haematologica 2004;89:567–77.
10. Ashcroft AJ, Davies FE, Morgan GJ. Aetiology of bonedisease and the role of bisphosphonates in multiplemyeloma. Lancet Oncol 2003;4:284–92.
11. Terpos E, Rahemtulla A. Bisphosphonate treatment formultiple myeloma. Drugs Today 2004;40:29–40.
12. Berenson JR, Hillner BE, Kyle RA, et al. American Societyof Clinical Oncology clinical practice guidelines: the role ofbisphosphonates in multiple myeloma. J Clin Oncol2002;20:3719–36.
13. Roodman GD. Biology of osteoclast activation in cancer. JClin Oncol 2001;19:3562–71.
14. Klein B, Zhang XG, Lu ZY, Bataille R. Interleukin-6 in humanmultiple myeloma. Blood 1995;85:863–72.
15. Sezer O, Heider U, Zavrski I, Kuhne CA, Hofbauer LC. RANKligand and osteoprotegerin in myeloma bone disease. Blood2003;101:2094–8.
16. Vanderkerken K, Asosingh K, Croucher P, Van Camp B.Multiple myeloma biology: lessons from the 5TMM models.Immunol Rev 2003;194:196–206.
17. Seidel C, Hjertner O, Abildgaard N, et al. Serum osteopro-tegerin levels are reduced in patients with multiplemyeloma with lytic bone disease. Blood 2001;98:2269–71.
18. Lipton A, Ali SM, Leitzel K, et al. Serum osteoprotegerinlevels in healthy controls and cancer patients. Clin CancerRes 2002;8:2306–10.
19. Terpos E, Szydlo R, Apperley JF, et al. Soluble receptoractivator of nuclear factor jB ligand–osteoprotegerin ratiopredicts survival in multiple myeloma: proposal for a novelprognostic index. Blood 2003;102:1064–9.
20. Standal T, Seidel C, Hjertner O, et al. Osteoprotegerin isbound, internalized, adegraded degraded by multiplemyeloma cells. Blood 2002;100:3002–7.

140 E. Terpos et al.
21. Pearse RN, Sordillo EM, Yaccoby S, et al. Multiple myelomadisrupts the TRANCE/osteoprotegerin cytokine axis totrigger bone destruction and promote tumor progression.Proc Natl Acad Sci USA 2001;98:11581–6.
22. Giuliani N, Bataille R, Mancini C, Lazzaretti M, Barille S.Myeloma cells induce imbalance in the osteoprotegerin/osteoprotegerin ligand system in the human bone marrowenvironment. Blood 2001;98:3527–33.
23. Hofbauer LC, Neubauer A, Heufelder AE. Receptoractivator of nuclear factor-jB ligand and osteoproteg-erin: potential implications for the pathogenesis andtreatment of malignant bone diseases. Cancer2001;92:460–70.
24. Grimaud E, Soubigou L, Couillaud S, et al. Receptoractivator of nuclear factor jB ligand (RANKL)/osteopro-tegerin (OPG) ratio is increased in severe osteolysis. Am JPathol 2003;163:2021–31.
25. Sezer O, Heider U, Jakob C, et al. Immunocytochemistryreveals RANKL expression of myeloma cells. Blood2002;99:4646–7. letter.
26. Heider U, Langelotz C, Jakob C, et al. Expression ofreceptor activator of nuclear factor jB ligand on bonemarrow plasma cells correlates with osteolytic bonedisease in patients with multiple myeloma. Clin CancerRes 2003;9:1436–40.
27. Farrugia AN, Atkins GJ, To LB, et al. Receptor activator ofnuclear factor-jB ligand expression by human myelomacells mediates osteoclast formation in vitro and correlateswith bone destruction in vivo. Cancer Res2003;63:5438–45.
28. Giuliani N, Colla S, Rizzoli V, Barille-Nion S, Bataille R. Dohuman myeloma cells directly produce the receptor acti-vator of nuclear factor jB ligand (RANKL) or induce RANKLin the bone marrow microenvironment?. Cancer Res2004;64:772–3. [letter].
29. Zhang XG, Gu JJ, Lu ZY, et al. Ciliary neurotropic factor,interleukin 11, leukemia inhibitory factor, and oncostatinM are growth factors for human myeloma cell lines usingthe interleukin 6 signal transducer gp130. J Exp Med1994;179:1337–42.
30. Lee JW, Chung HY, Ehrlich LA, Jelinek DF, Callander NS,Roodman GD, et al. IL-3 expression by myeloma cellsincreases both osteoclast formation and growth of mye-loma cells. Blood 2004;103:2308–15.
31. Choi SJ, Cruz JC, Craig F, Chung H, Devlin RD, Roodman GD,et al. Macrophage inflammatory protein 1-a is a potentialosteoclast stimulatory factor in multiple myeloma. Blood2000;96:671–5.
32. Han JH, Choi SJ, Kurihara N, Koide M, Oba Y, Roodman GD.Macrophage inflammatory protein-1a is an osteoclasto-genic factor in myeloma that is independent of receptoractivator of nuclear factor jB ligand. Blood2001;97:3349–53.
33. Abe M, Hiura K, Wilde J, et al. Role for macrophageinflammatory protein (MIP)-1a and MIP-1b in the develop-ment of osteolytic lesions in multiple myeloma. Blood2002;100:2195–202.
34. Hashimoto T, Abe M, Oshima T, Shibata H, Ozaki S, Inoue D,et al. Ability of myeloma cells to secrete macrophageinflammatory protein (MIP)-1a and MIP-1b correlates withlytic bone lesions in patients with multiple myeloma. Br JHaematol 2004;125:38–41.
35. Uneda S, Hata H, Matsuno F, Harada N, Mitsuya Y, KawanoF, et al. Macrophage inflammatory protein-1a is producedby human multiple myeloma (MM) cells and its expressioncorrelates with bone lesions in patients with MM. Br JHaematol 2003;120:53–5.
36. Terpos E, Politou M, Szydlo R, Goldman JM, Apperley JF,Rahemtulla A. Serum levels of macrophage inflammatoryprotein-1a (MIP-1a) correlate with the extent of bonedisease and survival in patients with multiple myeloma. BrJ Haematol 2003;123:106–9.
37. Vanderkerken K, De Leenheer E, Shipman C, Asosingh K,Willems A, Van Camp B, et al. Recombinant osteoproteg-erin decreases tumor burden and increases survival in amurine model of multiple myeloma. Cancer Res2003;63:287–9.
38. Sordillo EM, Pearse RN. RANK-Fc: a therapeutic antagonistfor RANK-L in myeloma. Cancer 2003;97(Suppl. 3):802–12.
39. Oyajobi BO, Franchin G, Williams PJ, et al. Dual effects ofmacrophage inflammatory protein-1a on osteolysis andtumor burden in the murine 5TGM1 model of myeloma bonedisease. Blood 2003;102:311–9.
40. Choi SJ, Oba Y, Gazitt Y, Alsina M, Cruz J, AndersonJ, et al. Antisense inhibition of macrophage inflam-matory protein 1-a blocks bone destruction in amodel of myeloma bone disease. J Clin Invest2001;108:1833–41.
41. Silvestris F, Cafforio P, Tucci M, Grinello D, Dammacco F.Upregulation of osteoblast apoptosis by malignant plasmacells: a role in myeloma bone disease. Br J Haematol2003;122:39–52.
42. Tian E, Zhan F, Walker R, Rasmussen E, Ma Y, Barlogie B,et al. The role of the Wnt-signaling antagonist DKK1 in thedevelopment of osteolytic lesions in multiple myeloma.New Engl J Med 2003;349:2483–94.
43. Calvo MS, Eyre DR, Gundberg CM. Molecular basis andclinical application of biological markers of bone turnover.Endocrine Rev 1996;17:333–68.
44. Hannon RA, Eastell R. Biochemical markers of boneturnover and fracture prediction. J Br Menopause Soc2003;9:10–5.
45. Hannon RA, Clowes JA, Eagleton AC, Al Hadari A, Eastell R,Blumsohn A. Clinical performance of immunoreactivetartrate-resistant acid phosphatase isoform 5b as a markerof bone resorption. Bone 2004;34:187–94.
46. Woitge HW, Pecherstorfer M, Horn E, et al. Serum bonesialoprotein as a marker of tumour burden and neoplasticbone involvement and as a prognostic factor in multiplemyeloma. Br J Cancer 2001;84:344–51.
47. Seibel MJ. Biochemical markers of bone metabolism in theassessment of osteoporosis: useful or not?. J EndocrinolInvest 2003;26:464–71.
48. Taube T, Beneton MN, McCloskey EV, Rogers S, Greaves M,Kanis JA. Abnormal bone remodeling in patients withmyelomatosis and normal biochemical indices of boneresorption. Eur J Haematol 1992;49:192–8.
49. Meilman E, Urivetzky MM, Rapoport CM. Urinary hydrox-yproline peptides. J Clin Invest 1963;42:40–50.
50. Prockop DJ, Keiser HR, Sjoerdsma A. Gastrointestinalabsorption and renal excretion of hydroxyproline peptides.Lancet 1962;2:527–8.
51. Segrest JP. Urinary metabolites of collagen. Meth Enzymol1982;82:398–410.
52. Prockop DJ, Kivirikko KI. Collagens: molecular biology,diseases, and potentials for therapy. Annu Rev Biochem1995;64:403–34.
53. Askenasi R, Rao VH, Devos A. Peptide-bound hydroxylysineand large polypeptides related to collagen synthesis. Eur JClin Invest 1976;6:361–3.
54. Arlaud GJ, Gaboriaud C, Thielens NM, Rossi V. Structuralbiology of C1. Biochem Soc Trans 2002;30(pt 6):1001–6.
55. Eyre DR. Bone biomarkers as tools in osteoporosis manage-ment. Spine 1997;22(Suppl. 24):17S–24S.

The role of markers of bone remodeling in multiple myeloma 141
56. Eyre DR, Paz MA, Gallop PM. Cross-linking in collagen andelastin. Annu Rev Biochem 1984;53:717–48.
57. Knott L, Bailey AJ. Collagen cross-links in mineralizingtissues: a review of their chemistry, function, and clinicalrelevance. Bone 1998;22:181–7.
58. Robins SP. Collagen crosslinks in metabolic bone disease.Acta Orthop Scand Suppl 1995;266:171–5.
59. Garnero P, Delmas PD. New developments in biochemicalmarkers for osteoporosis. Calcif Tissue Int 1996;59(Suppl.):S2–9.
60. Christenson RH. Biochemical markers of bone metabolism:an overview. Clin Biochem 1997;30:573–93.
61. Hanson DA, Weis MA, Bollen AM, Maslan SL, Singer FR, EyreDR. A specific immunoassay for monitoring human boneresorption: quantitation of type I collagen cross-linked N-telopeptides in urine. J Bone Miner Res 1992;7:1251–8.
62. Eyre DR. The specificity of collagen cross-links as markersof bone and connective tissue degradation. Acta OrthopScand Suppl 1995;266:166–70.
63. Apone S, Lee MY, Eyre DR. Osteoclasts generate cross-linked collagen N-telopeptides (NTx) but not free pyridin-olines when cultured on human bone. Bone1997;21:129–36.
64. Bonde M, Garnero P, Fledelius C, Qvist P, Delmas PD,Christiansen C. Measurement of bone degradation productsin serum using antibodies reactive with an isomerized formof an 8 amino acid sequence of the C-telopeptide of type Icollagen. J Bone Miner Res 1997;12:1028–34.
65. Rosenquist C, Fledelius C, Christgau S, Pedersen BJ, BondeP, Qvist P, et al. Serum CrossLaps One Step ELISA. Firstapplication of monoclonal antibodies for measurement inserum of bone-related degradation products from C-terminal telopeptides of type I collagen. Clin Chem1998;44:2281–9.
66. Woitge HW, Pecherstorfer M, Li Y, Keck AV, Horn E, ZieglerR, et al. Novel serum markers of bone resorption: clinicalassessment and comparison with established urinary indi-ces. J Bone Miner Res 1999;14:792–801.
67. Janckila AJ, Takahashi K, Sun SZ, Yam LT. Tartrate-resistant acid phosphatase isoform 5b as serum markerfor osteoclastic activity. Clin Chem 2001;47:74–80.
68. Janckila AJ, Neustadt DH, Nakasato YR, Halleen JM,Hentunen T, Yam LT. Serum tartrate-resistant acid phos-phatase isoforms in rheumatoid arthritis. Clin Chim Acta2002;320:49–58.
69. Halleen JM, Alatalo SL, Suominen H, Cheng S, Janckila AJ,Vaananen HK. Tartrate-resistant acid phosphatase 5b: anovel serum marker of bone resorption. J Bone Miner Res2000;15:1337–45.
70. Alatalo SL, Halleen JM, Hentunen TA, Monkkonen J,Vaananen HK. Rapid screening method for osteoclastdifferentiation in vitro that measures tartrate-resistantacid phosphatase 5b activity secreted into the culturemedium. Clin Chem 2000;46:1751–4.
71. Lam WK, Eastlund DT, Li CY, Yam LT. Biochemicalproperties of tartrate-resistant acid phosphatase in serumof adults and children. Clin Chem 1978;24:1105–8.
72. Ylipahkala H, Halleen JM, Kaija H, Vihko P, Vaananen HK.Tartrate-resistant acid phosphatase 5B circulates in humanserum in complex with a2-macroglobulin and calcium.Biochem Biophys Res Commun 2003;308:320–4.
73. Halleen JM. Tartrate-resistant acid phosphatase 5B is aspecific and sensitive marker of bone resorption. Antican-cer Res 2003;23:1027–9.
74. Gallop PM, Lian JB, Hauschka PV. Carboxylated calcium-binding proteins and vitamin K. New Engl J Med1980;302:1460–6.
75. Nelsestuen GL, Shah AM, Harvey SB. Vitamin K-dependentproteins. Vitam Horm 2000;58:355–89.
76. Young MF, Kerr JM, Ibaraki K, Heegaard AM, Robey PG.Structure, expression, regulation of the major noncolla-genous matrix proteins of bone. Clin Orthop1992;281:275–94.
77. Puchacz E, Lian JB, Stein GS, Wozney J, Huebner K, CroceC. Chromosomal localization of the human osteocalcingene. Endocrinology 1989;124:2648–50.
78. Javed A, Gutierrez S, Montecino M, van Wijnen AJ, SteinGS, Stein GS, et al. Multiple Cbfa/AML sites in the ratosteocalcin promoter are required for basal and vitamin D-responsive transcription and contribute to chromatinorganization. Mol Cell Biol 1999;19:7491–500.
79. Sierra J, Villagra A, Paredes R, et al. Regulation of thebone-specific osteocalcin gene by p300 requires Runx2/Cbfa1 and the vitamin D3 receptor but not p300 intrinsichistone acetyltransferase activity. Mol Cell Biol2003;23:3339–51.
80. Koeneman KS, Kao C, Ko SC, et al. Osteocalcin-directedgene therapy for prostate-cancer bone metastasis. World JUrol 2000;18:102–10.
81. Moss DW. Perspectives in alkaline phosphatase research.Clin Chem 1992;38:2486–92.
82. Smedsrod B, Melkko J, Risteli L, Risteli J. Circulating C-terminal propeptide of type I procollagen is cleared mainlyvia the mannose receptor in liver endothelial cells.Biochem J 1990;271:345–50.
83. Risteli J, Risteli L. Assays of type I procollagen domains andcollagen fragments: problems to be solved and futuretrends. Scand J Clin Lab Invest Suppl 1997;227:105–13.
84. Fohr B, Dunstan CR, Seibel MJ. Clinical review 165: markersof bone remodeling in metastatic bone disease. J ClinEndocrinol Metab 2003;88:5059–75.
85. Nawawi H, Samson D, Apperley J, Girgis S. Biochemicalbone markers in patients with multiple myeloma. Clin ChimActa 1996;253:61–77.
86. Abildgaard N, Bentzen SM, Nielsen JL, Heickendorff L.Serum markers of bone metabolism in multiple myeloma:prognostic value of the carboxy-terminal telopeptide oftype I collagen (ICTP). Br J Haematol 1997;96:103–10.
87. Withold W, Arning M, Schwarz M, Wolf HH, Schneider W.Monitoring of multiple myeloma patients by simultaneouslymeasuring marker substances of bone resorption andformation. Clin Chim Acta 1998;269:21–30.
88. Carlson K, Larsson A, Simonsson B, Turesson I, Westin J,Ljunghall S. Evaluation of bone disease in multiple mye-loma: a comparison between the resorption markersurinary deoxypyridinoline/creatinine (DPD) and serumICTP, and an evaluation of the DPD/osteocalcin andICTP/osteocalcin ratios. Eur J Haematol 1999;62:300–6.
89. Fonseca R, Trendle MC, Leong T, et al. Prognostic value ofserum markers of bone metabolism in untreated multiplemyeloma patients. Br J Haematol 2000;109:24–9.
90. Terpos E, Palermos J, Tsionos K, et al. Effect of pamidr-onate administration on markers of bone turnover anddisease activity in multiple myeloma. Eur J Haematol2000;65:331–6.
91. Corso A, Arcaini L, Mangiacavalli S, et al. Biochemicalmarkers of bone disease in asymptomatic early stagemultiple myeloma. A study on their role in identifying highrisk patients. Haematologica 2001;86:394–8.
92. Jakob C, Zavrski I, Heider U, et al. Bone resorptionparameters [carboxy-terminal telopeptide of type-I colla-gen (ICTP), amino-terminal collagen type-I telopeptide(NTx), and deoxypyridinoline (Dpd)] in MGUS and multiplemyeloma. Eur J Haematol 2002;69:37–42.

142 E. Terpos et al.
93. Alexandrakis MG, Passam FH, Malliaraki N, Katachanakis C,Kyriakou DS, Margioris AN. Evaluation of bone disease inmultiple myeloma: a correlation between biochemicalmarkers of bone metabolism and other clinical parametersin untreated multiple myeloma patients. Clin Chim Acta2002;325:51–7.
94. Abildgaard N, Brixen K, Kristensen JE, Eriksen EF, NielsenL, Heickendorff L. Comparison of five biochemical markersof bone resorption in multiple myeloma: elevated pre-treatment levels of S-ICTP and U-Ntx are predictive forearly progression of the bone disease during standardchemotherapy. Br J Haematol 2003;120:235–42.
95. Xue Y, Jia W, Zhang H, Dong J, Clemens JD, Tian X. Urinarycross-linked N-telopeptides of type I collagen and bonemetabolic diseases. Chin Med J 1999;112:149–52.
96. Terpos E, de Terpos la Fuente J, Szydlo R, et al. Tartrate-resistant acid phosphatase isoform 5b: a novel serummarker for monitoring bone disease in multiple myeloma.Int J Cancer 2003;106:455–7.
97. Terpos E, Palermos J, Viniou N, Vaiopoulos G, Meletis J,Yataganas X. Pamidronate increases markers of boneformation in patients with multiple myeloma in plateauphase under interferon-a treatment. Calcif Tissue Int2001;68:285–90.
98. Clark RE, Flory AJ, Ion EM, Woodcock BE, Durham BH,Fraser WD. Biochemical markers of bone turnover followinghigh-dose chemotherapy and autografting in multiplemyeloma. Blood 2000;96:2697–702.
99. Terpos E, Politou M, Szydlo R, et al. Autologous stem celltransplantation normalizes abnormal bone remodeling andsRANKL/osteoprotegerin ratio in patients with multiplemyeloma. Leukemia [in press] doi:10.1038/sj.leu.2403423.
100. Abildgaard N, Glerup H, Rungby J, et al. Biochemicalmarkers of bone metabolism reflect osteoclastic andosteoblastic activity in multiple myeloma. Eur J Haematol2000;64:121–9.
101. Jakob C, Zavrski I, Heider U, et al. Serum levels ofcarboxy-terminal telopeptide of type-I collagen are ele-vated in patients with multiple myeloma showing skeletalmanifestations in magnetic resonance imaging but lackinglytic bone lesions in conventional radiography. Clin CancerRes 2003;9:3047–51.
102. Alexandrakis MG, Kyriakou DS, Passam FH, Malliaraki N,Vlachonikolis IG, Karkavitsas N. Urinary N-telopeptidelevels in multiple myeloma patients, correlation with Tc-99m-sestaMIBI scintigraphy and other biochemical markersof disease activity. Hematol Oncol 2003;21:17–24.
103. Woitge HW, Horn E, Keck AV, Auler B, Seibel MJ, Pecher-storfer M. Biochemical markers of bone formation in
patients with plasma cell dyscrasias and benign osteopo-rosis. Clin Chem 2001;47:686–93.
104. Politou M, Terpos E, Anagnostopoulos A, et al. Role ofreceptor activator of nuclear factor-jB ligand (RANKL),osteoprotegerin and macrophage inflammatory protein-1a(MIP-1a) in monoclonal gammopathy of undeterminedsignificance (MGUS). Br. J. Haematol. [in press].
105. Turesson I, Abildgaard N, Ahlgren T, et al. Prognosticevaluation in multiple myeloma: an analysis of the impactof new prognostic factors. Br J Haematol1999;106:1005–12.
106. Elomaa I, Risteli L, Laakso M, Lahtinen R, Virkkunen P,Risteli J. Monitoring the action of clodronate with type Icollagen metabolites in multiple myeloma. Eur J Cancer1996;32A:1166–70.
107. Berenson JR, Rosen LS, Howell A, et al. Zoledronic acidreduces skeletal-related events in patients with osteolyticmetastases. Cancer 2001;91:1191–200.
108. Terpos E, Viniou N, de la Fuente J, et al. Pamidronate issuperior to ibandronate in decreasing bone resorption,interleukand and b2-microglobulin in multiple myeloma.Eur J Haematol 2003;70:34–42.
109. Gordon S, Helfrich MH, Sati HI, et al. Pamidronate causesapoptosis of plasma cells in vivo in patients with multiplemyeloma. Br J Haematol 2002;119:475–83.
110. Brown JM, Vessella RL, Kostenuik PJ, Dunstan CR, LangeE, Corey E. Serum osteoprotegerin levels are increased inpatients with advanced prostate cancer. Clin Cancer Res2001;7:2977–83.
111. Jung K, Lein M, von Hosslin K, Brux B, Schnorr D, LoeningSA, et al. Osteoprotegerin in serum as a novel marker ofbone metastatic spread in prostate cancer. Clin Chem2001;47:2061–3.
112. Jung K, Stephan C, Semjonow A, Lein M, Schnorr D, LoeningSA. Serum osteoprotegerin and receptor activator ofnuclear factor-jB ligand as indicators of disturbed osteo-clastogenesis in patients with prostate cancer. J Urol2003;170:2302–5.
113. Body JJ, Greipp P, Coleman RE, et al. A phase I studyof AMGN-0007, a recombinant osteoprotegerin con-struct, in patients with multiple myeloma or breastcarcinoma related bone metastases. Cancer 2003;97:887–92.
114. Nanci A. Content and distribution of noncollagenous matrixproteins in bone and cementum: relationship to speed offormation and collagen packing density. J Struct Biol1999;126:256–69.
115. Delmas PD. Bone marker nomenclature. Bone2001;28:575–6.
Related Documents











