The role of extracellular vesicles in soma-to-germline communication Author: Jayasooriah, Navind Publication Date: 2018 DOI: https://doi.org/10.26190/unsworks/20944 License: https://creativecommons.org/licenses/by-nc-nd/3.0/au/ Link to license to see what you are allowed to do with this resource. Downloaded from http://hdl.handle.net/1959.4/61243 in https:// unsworks.unsw.edu.au on 2022-08-26

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The role of extracellular vesicles in soma-to-germlinecommunication
Author:Jayasooriah, Navind
Publication Date:2018
DOI:https://doi.org/10.26190/unsworks/20944
License:https://creativecommons.org/licenses/by-nc-nd/3.0/au/Link to license to see what you are allowed to do with this resource.
Downloaded from http://hdl.handle.net/1959.4/61243 in https://unsworks.unsw.edu.au on 2022-08-26

The role of extracellular vesicles in
soma-to-germline communication
Navind Jayasooriah
A thesis in fulfilment for the requirements for the degree of
Doctor of Philosophy
School of Biotechnology and Biomolecular Sciences
Faculty of Science
November 2018


FOR OFFICE USE ONLY Date of completion of requirements for Award:
Thesis/Dissertation Sheet
Abstract 350 words maximum: (PLEASE TYPE) The inheritance of environmentally-induced traits is an established phenomenon, however the underlying molecular mechanism is yet to be elucidated. In some cases, environmental factors may act directly on the germline, however in other cases this appears impossible. While ‘soma-to-germline feedback’ conflicts with the long-held dogma that heritable genetic information flows solely from germline to soma, there is no reason to suppose that germline- associated somatic cells cannot communicate with developing germ cells. A prime candidate for the means of such communication is small RNAs carried within extracellular vesicles (EVs). Small RNAs have been associated with inheritance of acquired phenotypes across phyla, and EVs are known intercellular messengers produced by nearly all cells. In this thesis, I have taken the first steps towards testing this idea by investigating the hypotheses that EVs produced by germline-associated somatic cells transfer small RNA cargo to germ cells, and that the RNA cargo is susceptible to environmental influence. I isolated and characterised EVs from two types of germline-associated somatic cells, Sertoli cells (in vitro) and epididymal epithelial cells (in vitro and in vivo), and characterised their small RNA profiles using high-throughput sequencing. By co-incubating labelled Sertoli EVs with spermatogonial stem cells (SSCs) and labelled epididymal EVs with living sperm, I showed that these EVs interact with germ cells. Furthermore, I also demonstrated the transfer of many Sertoli EV small RNAs to SSCs. I also established that the small RNA profiles of EVs can be influenced by the environment. I used Bisphenol A and dimethyl sulfoxide as environmental stressors of Sertoli cells, varied the concentration of folate available for in vitro epididymal cells, and modelled the response of in vivo epididymal EVs to dietary methyl donor supplementation. The small RNAs affected by these factors included miRNAs associated with transcription and nucleic acid-binding (some of which were transferred from Sertoli EVs to SSCs), and tRNA-derived fragments that have previously been associated with the vertical transmission of diet-induced phenotypes. Taken together, my data show that small RNA cargo from somatic EVs is susceptible to environmental influence, and thus such somatic RNA is capable of being transferred to the germline.
Declaration relating to disposition of project thesis/dissertation
I hereby grant to the University of New South Wales or its agents the right to archive and to make available my thesis or dissertation in whole or in part in the University libraries in all forms of media, now or here after known, subject to the provisions of the Copyright Act 1968. I retain all property rights, such as patent rights. I also retain the right to use in future works (such as articles or books) all or part of this thesis or dissertation.
I also authorise University Microfilms to use the 350 word abstract of my thesis in Dissertation Abstracts International (this is applicable to doctoral theses only).
……………………………………………………………
Signature
……………………………………..………………
Witness Signature
……….……………………...…….…
Date
The University recognises that there may be exceptional circumstances requiring restrictions on copying or conditions on use. Requests for restriction for a period of up to 2 years must be made in writing. Requests for a longer period of restriction may be considered in exceptional circumstances and require the approval of the Dean of Graduate Research.
Surname/Family Name : Jayasooriah Given Name/s : Navind Abbreviation for degree as give in the University calendar : PhD Faculty : Science School : Biotechnology and Biomolecular Sciences Thesis Title : The role of extracellular vesicles in soma-to-germline communication


ORIGINALITY STATEMENT ‘I hereby declare that this submission is my own work and to the best of my knowledge it contains no materials previously published or written by another person, or substantial proportions of material which have been accepted for the award of any other degree or diploma at UNSW or any other educational institution, except where due acknowledgement is made in the thesis. Any contribution made to the research by others, with whom I have worked at UNSW or elsewhere, is explicitly acknowledged in the thesis. I also declare that the intellectual content of this thesis is the product of my own work, except to the extent that assistance from others in the project's design and conception or in style, presentation and linguistic expression is acknowledged.’ Signed …………………………………………….............. Date ……………………………………………..............


COPYRIGHT STATEMENT
‘I hereby grant the University of New South Wales or its agents the right to archive and to make available my thesis or dissertation in whole or part in the University libraries in all forms of media, now or here after known, subject to the provisions of the Copyright Act 1968. I retain all proprietary rights, such as patent rights. I also retain the right to use in future works (such as articles or books) all or part of this thesis or dissertation. I also authorise University Microfilms to use the 350 word abstract of my thesis in Dissertation Abstract International (this is applicable to doctoral theses only). I have either used no substantial portions of copyright material in my thesis or I have obtained permission to use copyright material; where permission has not been granted I have applied/will apply for a partial restriction of the digital copy of my thesis or dissertation.'
Signed ……………………………………………...........................
Date ……………………………………………...........................
AUTHENTICITY STATEMENT
‘I certify that the Library deposit digital copy is a direct equivalent of the final officially approved version of my thesis. No emendation of content has occurred and if there are any minor variations in formatting, they are the result of the conversion to digital format.’
Signed ……………………………………………...........................
Date ……………………………………………...........................


i
ABSTRACT
The inheritance of environmentally-induced traits is an established phenomenon,
however the underlying molecular mechanism is yet to be elucidated. In some cases,
environmental factors may act directly on the germline, however in other cases this
appears impossible. While ‘soma-to-germline feedback’ conflicts with the long-held
dogma that heritable genetic information flows solely from germline to soma, there is
no reason to suppose that germline-associated somatic cells cannot communicate with
developing germ cells. A prime candidate for the means of such communication is
small RNAs carried within extracellular vesicles (EVs). Small RNAs have been
associated with inheritance of acquired phenotypes across phyla, and EVs are known
intercellular messengers produced by nearly all cells. In this thesis, I have taken the
first steps towards testing this idea by investigating the hypotheses that EVs produced
by germline-associated somatic cells transfer small RNA cargo to germ cells, and that
the RNA cargo is susceptible to environmental influence. I isolated and characterised
EVs from two types of germline-associated somatic cells, Sertoli cells (in vitro) and
epididymal epithelial cells (in vitro and in vivo), and characterised their small RNA
profiles using high-throughput sequencing. By co-incubating labelled Sertoli EVs with
spermatogonial stem cells (SSCs) and labelled epididymal EVs with living sperm, I
showed that these EVs interact with germ cells. Furthermore, I also demonstrated the
transfer of many Sertoli EV small RNAs to SSCs. I also established that the small RNA
profiles of EVs can be influenced by the environment. I used Bisphenol A and dimethyl
sulfoxide as environmental stressors of Sertoli cells, varied the concentration of folate
available for in vitro epididymal cells, and modelled the response of in vivo epididymal

ii
EVs to dietary methyl donor supplementation. The small RNAs affected by these
factors included miRNAs associated with transcription and nucleic acid-binding (some
of which were transferred from Sertoli EVs to SSCs), and tRNA-derived fragments that
have previously been associated with the vertical transmission of diet-induced
phenotypes. Taken together, my data show that small RNA cargo from somatic EVs is
susceptible to environmental influence, and thus such somatic RNA is capable of being
transferred to the germline.

iii
ACKNOWLEDGEMENTS
Firstly, I would like to thank my primary supervisor, Catherine Suter. Since joining the
Suter Lab as an Honours student, she has shown patience, and provided guidance and
motivation. She gave me space and independence, allowing me to take control of my
project and my thesis. While the latter stages of my project have been at times
challenging, I admire how you were always able to put on a smile despite the
circumstances. I thank you for your supervision over the past five years and two
degrees.
Second, I would like to thank my co-supervisor Jennifer Cropley. She has shown great
patience and generosity and was always willing to spend time to share her skills from
the bench to the keyboard. I am most grateful for her tutelage on how to communicate
effectively – to convert complicated ideas from my head into a coherent story onto
pages, presentations and posters.
To Matthias, my work-husband, thank you for your companionship over the past 4
years and for sharing this PhD adventure with me. You were the fiery, passionate yin
to my quiet, cautious yang, and thank you for putting up with all my cheeky
comments and silly puns. To Paul, the jack-of-all-trades, thank you for introducing me
to the black-screen terminal (and also reptilia and amphibia!), and for being a great
sounding board for troubleshooting. To Sally, thank you for introducing me to the
world of EVs, and for the mentorship given from my Honours degree to the final time
at the bench. To all other members of the Suter Laboratory (Saeideh, Alastair, Suzy and

iv
many others over the past five years), thank you all for your support and sharing my
trials and tribulations of PhD life.
Next, I would like to thank Christine for her support and patience. I apologise for the
countless times where you waited for me while experiments were running overtime, as
well as for the many weekend visits to the lab.
Finally, I would like to thank my parents for their continued support that in no doubt
impacted on my thesis. To my father, I dedicate this to you.

v
TABLE OF CONTENTS
ABSTRACT .............................................................................................................................................. i
ACKNOWLEDGEMENTS ......................................................................................................................... iii
TABLE OF CONTENTS ............................................................................................................................. v
LIST OF ABBREVIATIONS ....................................................................................................................... ix
LIST OF FIGURES .................................................................................................................................... x
LIST OF TABLES .................................................................................................................................... xiv
PUBLICATIONS AND PRESENTATIONS ARISING FROM WORK IN THIS THESIS ....................................... xvi
1 INTRODUCTION ............................................................................................................................ 1
1.1 THE INHERITANCE OF ACQUIRED TRAITS ................................................................................................... 1
1.1.1 Germ plasm theory and the Weismann Barrier .................................................................... 1
1.1.2 Intergenerational transmission of environmentally-induced traits ...................................... 2
1.1.3 Potential routes of transmission for an environmentally-induced signal in the germline .... 5
1.1.4 Molecular conduits of environmental information between generations ............................ 6
1.2 GERMLINE-ASSOCIATED SOMATIC CELLS ................................................................................................... 9
1.2.1 Sertoli cells .......................................................................................................................... 10
1.2.2 The epididymis .................................................................................................................... 14
1.3 EXTRACELLULAR VESICLES ................................................................................................................... 17
1.3.1 EVs contain selectively packaged cargo ............................................................................. 20
1.3.2 EVs are intercellular messengers ........................................................................................ 21
1.3.3 Functional role of EVs in regular physiology and pathology .............................................. 22
1.3.4 Extracellular vesicles in reproduction ................................................................................. 23
1.4 HYPOTHESIS AND AIMS ....................................................................................................................... 24
2 MATERIALS AND METHODS ........................................................................................................27
2.1 CELL CULTURE AND TISSUE COLLECTION ................................................................................................. 27

vi
2.1.1 Cell lines and culture conditions ..........................................................................................27
2.1.2 Epididymides dissection and epididymal fluid collection ....................................................28
2.1.3 Sperm isolation ...................................................................................................................29
2.2 EXTRACELLULAR VESICLE PURIFICATION AND CHARACTERISATION .................................................................29
2.2.1 Preparation of EV-depleted media ......................................................................................29
2.2.2 Cell line EV production ........................................................................................................30
2.2.3 EV pelleting by ultracentrifugation .....................................................................................31
2.2.4 Nanoparticle tracking analysis ............................................................................................31
2.2.5 Transmission electron microscopy ......................................................................................32
2.2.6 Protein mass spectrometry .................................................................................................32
2.3 IMAGING AND MICROSCOPY .................................................................................................................33
2.3.1 EV labelling .........................................................................................................................33
2.3.2 EV-cell co-incubation ...........................................................................................................33
2.4 RNA ANALYSES .................................................................................................................................35
2.4.1 Extraction ............................................................................................................................35
2.4.2 Gene expression microarray analysis ..................................................................................36
2.4.3 Small RNA libraries ..............................................................................................................37
3 SERTOLI CELL EXTRACELLULAR VESICLES AND THE EFFECTS OF ENVIRONMENTAL STRESS .......... 39
3.1 INTRODUCTION .................................................................................................................................39
3.2 RESULTS ..........................................................................................................................................41
3.2.1 The isolation and characterisation of Sertoli EVs ................................................................41
3.2.2 Sertoli EV small RNA cargo..................................................................................................45
3.2.3 Interactions between Sertoli EVs and germ cells ................................................................56
3.2.4 Environmental effects on Sertoli EVs ..................................................................................65
3.3 DISCUSSION .....................................................................................................................................77
3.3.1 Isolation of Sertoli EVs ........................................................................................................77
3.3.2 Sertoli EVs contain selectively packaged small RNA cargo .................................................79
3.3.3 Sertoli EVs bind to and transfer small RNAs to spermatogonial stem cells ........................81

vii
3.3.4 Environmental effects on Sertoli EV production and cargo ................................................ 84
3.3.5 Conclusion .......................................................................................................................... 87
4 THE EFFECT OF FOLATE AVAILABILITY ON EPIDIDYMAL EXTRACELLULAR VESICLES .....................89
4.1 INTRODUCTION ................................................................................................................................. 89
4.2 RESULTS .......................................................................................................................................... 92
4.2.1 Isolation and characterisation of DC2 epididymal EVs ....................................................... 92
4.2.2 Characterisation of epididymal EV small RNA cargo .......................................................... 96
4.2.3 Effects of folate concentration on epididymal EV production and small RNA cargo ........ 105
4.3 DISCUSSION ................................................................................................................................... 123
4.3.1 The isolation of DC2 epididymal EVs ................................................................................ 123
4.3.2 DC2 epididymal EVs contain selectively packaged cargo ................................................. 125
4.3.3 Effect of folate concentration on epididymal EV production and cargo ........................... 127
4.3.4 Conclusion ........................................................................................................................ 131
5 MOUSE EPIDIDYMAL EXTRACELLULAR VESICLES AND THE EFFECTS OF DIETARY METHYL DONORS
132
5.1 INTRODUCTION ............................................................................................................................... 132
5.2 RESULTS ........................................................................................................................................ 134
5.2.1 Isolation and characterisation of mouse epididymal EVs ................................................. 134
5.2.2 Interactions between epididymal EVs and spermatozoa.................................................. 138
5.2.3 Characterisation of mouse epididymal EV small RNA cargo ............................................ 140
5.2.4 Effects of dietary methyl donor supplementation on epididymal EV production and cargo
144
5.2.5 Mouse epididymal EVs contain a unique processed rRNA transcript ............................... 151
5.3 DISCUSSION ................................................................................................................................... 154
5.3.1 Isolation of mouse epididymal EVs ................................................................................... 155
5.3.2 Characterisation of mouse epididymal EV small RNA cargo ............................................ 156

viii
5.3.3 Dietary methyl donor supplementation has no detectable effect on epididymal EV cargo
157
5.3.4 Mouse epididymal EVs contain a traditionally unmappable ribosomal gene transcript
enriched in mature sperm .............................................................................................................. 159
5.3.5 Conclusion ........................................................................................................................ 160
6 CONCLUDING REMARKS AND FUTURE DIRECTION ................................................................... 161
6.1 LIMITATIONS OF THIS THESIS ............................................................................................................. 166
6.2 CONCLUDING REMARKS ................................................................................................................... 168
7 REFERENCES ............................................................................................................................. 169
8 APPENDIX ................................................................................................................................. 186

ix
LIST OF ABBREVIATIONS
AB apoptotic body BPA Bisphenol A BWW Biggers, Whitten and Whittingham media DAVID Database for Annotation, Visualisation and
Integrated Discovery DMEM Dulbecco’s Modified Eagle Medium DMSO dimethyl sulfoxide ECM extracellular matrix EDC endocrine disrupting chemical EV extracellular vesicle FBP folate-binding protein FBS fetal bovine serum IGS intergenic spacer ILV intraluminal vesicle HFL high folate for long-term HFS high folate for short-term HS horse serum IMDM Iscove’s Modified Dulbecco’s Medium LF low folate MDS methyl donor supplemented MVB multivesicular body miRNA microRNA NTA nanoparticle tracking analysis PBS phosphate buffered saline PC1 principal component one PC2 principal component two PCA principal component analysis RNase ribonuclease rRNA ribosomal RNA RT room temperature SCB Sertoli cell barrier SEM standard error of the mean siRNA short-interfering RNA SSC spermatogonial stem cell TBE tris/borate/EDTA buffer TEM transmission electron microscopy tRF tRNA fragment tRNA transfer RNA

x
LIST OF FIGURES
Figure 1.1 Sertoli and germ cells in the seminiferous tubules ............................................ 11
Figure 1.2 Diagrammatical representation of the epididymis and its segments ............. 15
Figure 1.3 The biogenesis and fate of extracellular vesicles ............................................... 19
Figure 3.1 Physical properties of EVs isolated from Sertoli cells ....................................... 42
Figure 3.2 Proteins identified in Sertoli EVs ......................................................................... 43
Figure 3.3 Size distribution of RNAs in Sertoli cells and EVs ............................................ 46
Figure 3.4 Sertoli cells and EVs small RNA biotype and length distribution .................. 47
Figure 3.5 miRNA read length distribution for Sertoli cells and EVs ............................... 48
Figure 3.6 Sertoli cells and EVs have distinct miRNA profiles .......................................... 49
Figure 3.7 Relative abundance of miRNAs between Sertoli cells and EVs ...................... 50
Figure 3.8 Gene ontologies overrepresented by miRNAs differentially expressed
between Sertoli EVs and cells ................................................................................................. 52
Figure 3.9 tRFs in Sertoli cells and EVs ................................................................................. 54
Figure 3.10 Sertoli cells and EVs can be distinguished by the abundance of the top six
tRFs ............................................................................................................................................. 55
Figure 3.11 tRFs differentially expressed between Sertoli cells and EVs.......................... 56
Figure 3.12 SSCs co-incubated with labelled Sertoli EVs for 15 hours ............................. 58
Figure 3.13 SSCs co-incubated with labelled Sertoli EVs for three hours ........................ 59
Figure 3.14 Changes in SSC miRNA abundance after exposure to Sertoli EVs ............... 61
Figure 3.15 tRFs in SSCs before and after exposure to Sertoli EVs .................................... 63
Figure 3.16 Gene expression changes in SSCs after exposure to Sertoli EVs ................... 64
Figure 3.17 TM4 Sertoli cell viability at varying doses of BPA .......................................... 65

xi
Figure 3.18 Size and yield of EVs produced by Sertoli cells after no, DMSO and BPA
exposure .................................................................................................................................... 67
Figure 3.19 miRNA profiles of Sertoli cells and EVs of different growth environments69
Figure 3.20 BPA effects on Sertoli EV and cellular miRNA cargo .................................... 70
Figure 3.21 Effects of treatment with DMSO on Sertoli EV and cell miRNA cargo ....... 71
Figure 3.22 Gene ontologies overrepresented by the predicted targets of miRNAs
upregulated in Sertoli EVs after DMSO treatment .............................................................. 73
Figure 3.23 tRFs in Sertoli cells and EVs from different growth environments .............. 74
Figure 3.24 tRFs in Sertoli cells and EVs produced under different growth
environments ............................................................................................................................ 75
Figure 3.25 Abundance of top six most abundant tRFs in Sertoli cells and EVs of
different growth environments .............................................................................................. 76
Figure 4.1 Physical properties of EVs isolated from DC2 epididymal cells .................... 93
Figure 4.2 Proteins identified in epididymal EVs ............................................................... 94
Figure 4.3 Size distribution of RNAs in epididymal cells and EVs .................................. 97
Figure 4.4 Epididymal cell and EV biotype and read length distribution ....................... 98
Figure 4.5 Relative abundance of miRNAs between epididymal cells and EVs ........... 100
Figure 4.6 Gene ontologies overrepresented by the predicted targets of miRNAs
selectively packaged into epididymal EVs ......................................................................... 103
Figure 4.7 tRFs in epididymal cells and EVs ...................................................................... 104
Figure 4.8 Schematic of DC2 epididymal cell folate treatments ...................................... 106
Figure 4.9 Size and yield of EVs produced by epididymal cells after different
treatments of folate ................................................................................................................ 107

xii
Figure 4.10 Effect of folate concentration on epididymal cell and EV biotype and read
length distribution .................................................................................................................. 108
Figure 4.11 Effect of folate concentration on epididymal cell and EV miRNA profiles110
Figure 4.12 Effects of folate reduction on epididymal EV miRNA ................................. 111
Figure 4.13 Effects of folate reduction on epididymal cell miRNA ................................. 115
Figure 4.14 Effects of re-exposure to high folate on epididymal EV and cell miRNA . 116
Figure 4.15 Gene ontologies overrepresented by the predicted targets of miRNAs
downregulated in epididymal EVs after short-term exposure to excess folate ............. 118
Figure 4.16 Effect of short- vs long-term exposure on epididymal EV and cell miRNA
profiles ...................................................................................................................................... 119
Figure 4.17 Effect of folate concentration on tRFs in epididymal cells and EVs ........... 120
Figure 4.18 Effect of folate concentration on epididymal cell and EV tRF profiles ....... 121
Figure 5.1 Paternal epigenetic inheritance after dietary methyl donor supplementation
in the Avy mouse ...................................................................................................................... 133
Figure 5.2 Physical properties of EVs isolated from mouse epididymal fluid .............. 135
Figure 5.3 Proteins identified in mouse epididymal EVs ................................................. 136
Figure 5.4 Freshly isolated mouse spermatozoa co-incubated with membrane labelled
mouse epididymal EVs .......................................................................................................... 139
Figure 5.5 Size distribution of RNAs in mouse epididymal EVs ..................................... 140
Figure 5.6 Epididymal EV biotype and read length distribution .................................... 141
Figure 5.7 Gene ontologies overrepresented by the predicted targets of the ten most
abundant miRNAs in mouse epididymal EVs ................................................................... 142
Figure 5.8 tRFs in mouse epididymal EVs .......................................................................... 143

xiii
Figure 5.9 Physical properties of epididymal EVs from control and MDS mice .......... 145
Figure 5.10 Effects of control and MDS diets on mouse epididymal EV small RNA
biotypes and read length distribution ................................................................................. 146
Figure 5.11 Effect of control and MDS diet on mouse epididymal EV miRNA profiles
.................................................................................................................................................. 147
Figure 5.12 Effect of dietary methyl donor supplementation on mouse epididymal EV
miRNA ..................................................................................................................................... 148
Figure 5.13 tRFs in epididymal EVs isolated from mice fed a control MDS diet .......... 149
Figure 5.14 Effect of genotype and diet on mouse epididymal EV tRF profiles ........... 150
Figure 5.15 Abundance of top tRFs in control and MDS epididymal EVs .................... 150
Figure 5.16 Proportion of reads mapped in mouse epididymal EVs, epididymal
epithelial cells and epididymal tissue ................................................................................. 152
Figure 5.17 Abundance of rsRNA-28S in control and MDS epididymal EVs ............... 154
Figure 6.1 Somatic EVs may act as vectors for environmentally-influenced small RNAs
to enter the germline and possibly the next generation ................................................... 168

xiv
LIST OF TABLES
Table 3.1 Common exosome proteins identified in Sertoli EVs ......................................... 44
Table 3.2 Top 10 most abundant miRNAs in Sertoli EVs ................................................... 49
Table 3.3 miRNAs selectively packaged into Sertoli EVs ................................................... 51
Table 3.4 miRNAs selectively excluded from Sertoli EVs .................................................. 51
Table 3.5 miRNAs increased in SSCs after exposure to Sertoli EVs .................................. 62
Table 3.6 EV miRNAs upregulated in response to DMSO treatment ............................... 72
Table 3.7 EV miRNAs downregulated in response to DMSO treatment .......................... 72
Table 4.1 Top 25 ExoCarta proteins and their identification in DC2 epididymal EVs ... 95
Table 4.2 Top 10 most abundant miRNAs in epididymal EVs .......................................... 99
Table 4.3 miRNAs selectively packaged into epididymal EVs ........................................ 101
Table 4.4 miRNAs selectively excluded from epididymal EVs ....................................... 102
Table 4.5 miRNAs upregulated in epididymal EVs after folate reduction .................... 112
Table 4.6 miRNAs downregulated in epididymal EVs after folate reduction ............... 113
Table 4.7 miRNAs upregulated in epididymal EVs after short-term re-exposure to
excess folate ............................................................................................................................. 117
Table 4.8 miRNAs downregulated in epididymal EVs after short-term re-exposure to
excess folate ............................................................................................................................. 117
Table 5.1 Common exosome proteins identified in mouse epididymal EVs ................. 137
Table 5.2 Top 10 most abundant miRNAs in epididymal EVs ........................................ 142
Table 5.3 Top 10 most abundant miRNAs in epididymal EVs from control and MDS
mice ........................................................................................................................................... 148
Table 5.4 Abundant 36 nt unmappable sequence in mouse epididymal EVs ............... 153

xv
Table 8.1 Proteins identified in Sertoli EVs ........................................................................ 186
Table 8.2 miRNAs (normalised to per million miRNA reads and averaged for each
group) detected in Sertoli cells and EVs of different treatments..................................... 195
Table 8.3 tRFs (normalised to per million tRF reads and averaged for each group)
detected in Sertoli cells and EVs of different treatments .................................................. 200
Table 8.4 Proteins identified in DC2 epididymal EVs ...................................................... 202
Table 8.5 miRNAs (normalised to per million miRNA reads) detected in DC2
epididymal cells and EVs of different treatments ............................................................. 207
Table 8.6 tRFs (normalised to per million tRF reads) detected in DC2 epididymal cells
and EVs of different treatments ........................................................................................... 212
Table 8.7 Mouse epididymal fluid pools ............................................................................ 214
Table 8.8 Proteins identified in mouse epididymal EVs ................................................... 215
Table 8.9 miRNAs (normalised to per million miRNA reads and averaged for each
group) detected in mouse epididymal EVs of backgrounds and diets .......................... 235
Table 8.10 tRFs (normalised to per million tRF reads and averaged for each group)
detected in mouse epididymal EVs of different backgrounds and diets ....................... 239
Table 8.11 BLAST hits for unmappable sequence in mouse epididymal EVs ............... 241

xvi
PUBLICATIONS AND PRESENTATIONS ARISING FROM WORK IN
THIS THESIS
Poster: Australasia Extracellular Vesicles Conference 2014 Cairns, Queensland, Australia Poster: EMBL Australia PhD Symposium 2014 University of New South Wales, Sydney, New South Wales, Australia Poster: St Vincent’s Campus Research Symposium 2015 Garvan Institute of Medical Research, Sydney, New South Wales, Australia Poster: Australian Epigenetics Conference 2015 Hobart, Tasmania, Australia Publication: Roll over Weismann: extracellular vesicles in the transgenerational transmission of environmental effects 2015 Eaton, S. A., Jayasooriah, N., Buckland, M. E., Martin, D. I., Cropley, J. E., & Suter, C. M. Epigenomics, 7(7), 1165-1171. doi:10.2217/epi.15.58
Poster: EMBL Australia PhD Symposium 2015 Bio21 Institute, Melbourne, Victoria, Australia
Poster: Abcam Mechanisms and Mysteries in Epigenetics 2016 Garvan Institute of Medical Research, Sydney, New South Wales, Australia Talk: International Society for Extracellular Vesicles Meeting 2016 Rotterdam, Netherlands Talk: Cellular Communications Satellite Meeting – Queenstown Research Week 2016 Nelson, New Zealand Talk: St Vincent’s Campus Research Week Fast Forward Presentation 2016 St Vincent’s Public Hospital, Sydney, New South Wales, Australia

xvii
Talk: International Society for Extracellular Vesicles Workshop on Diet, Environment and Extracellular Vesicles 2017 La Trobe University, Melbourne, Victoria, Australia Talk: Australasia Extracellular Vesicles Conference 2017 Lorne, Victoria, Australia Talk: Australian Society for Medical Research NSW Annual Scientific Meeting 2017 Westmead Hospital, Sydney, New South Wales, Australia


1
1 INTRODUCTION
1.1 The inheritance of acquired traits
1.1.1 Germ plasm theory and the Weismann Barrier
The idea that environmental signals can be passed through the germline was dismissed
by August Weismann in his germ plasm theory of heredity in 1893 (1). He proposed
that the germline alone is responsible for transmitting heritable information to the next
generation, and that such information travels in one direction only, from germline to
soma, and not in reverse. Weismann’s germ plasm theory gained traction as it
harmonised with the discovery and acceptance that DNA (and not protein) was the
agent of heredity (2,3) and Francis Crick’s ‘Sequence hypothesis’ (4), that genetic
information travels from DNA to protein and not in reverse, which formed the basis
for the central dogma of molecular biology (5). Over the ensuing 120 years the concept
of the “Weismann Barrier” became dogmatised without rigorous scientific scrutiny,
especially in animal biology.
Now, evidence suggests that the hypothetical Weismann Barrier may be permeable.
Firstly, the Weismann Barrier is moot in plants – plant somatic cells can undergo
meiosis to form gametes (reviewed in (6)). Additionally, we know that there is no
physical manifestation of the Weismann Barrier. In all multicellular organisms, germ
cells exist directly alongside somatic cells without any physical barrier or separating
structure. For example, in mammals Sertoli cells are intimately connected to both germ
cells and developing sperm in the testis (1.2.1), and maturing sperm are in constant

Introduction
2
contact with epididymal epithelial cells (1.2.2). Thirdly, there are numerous instances
where environmentally-induced traits have been transmitted between generations, and
in some of these, inheritance may have occurred via breach of the Weismann barrier.
1.1.2 Intergenerational transmission of environmentally-induced traits
Environmental factors can influence the phenotype not only of an exposed individual,
but in many cases also that of offspring and even grand-offspring. This vertical
transmission can occur in the absence of continued exposure to the causative
environment, and without inducing a genetic change in the germline. Examples of such
phenomena are rife in the plant kingdom (7), and this may be due to the ability of
plants to derive germ cells from somatic cells (6). However, the transmission of
environmental influences across generations has also been documented in animals,
which segregate a distinct germline early in development (8). In Caenorhabditis elegans,
heritability across three generations has been shown for viral-induced gene silencing (a
host-driven resistance response), and starvation-induced non-fatal developmental
arrest (9,10). In another invertebrate, Drosophila melanogaster, heat and osmotic stress-
induced alterations to heterochromatin are also passed down to the next generation, in
a non-Mendelian manner (11).
Similar examples also exist in mammals. These encompass a wide variety of inducing
environmental factors, ranging from health and metabolic stressors (12,13), toxin and
chemical exposure (14-17), nutrition and diet (18-20), and even psychological stress
(21,22). While in many studies, the alternative mechanisms of transmissions cannot be

Introduction
3
ruled out (such as social inheritance, or via the gestational environment and the
microbiome), there are several well-designed studies in which true-inheritance of
environmentally-induced traits have been demonstrated via the germline. On the other
hand, while less common in the literature, transmission of an acquired paternal state
also occurs, and presents an arguably more tractable system to study the inheritance of
environmentally-influenced traits (23). With paternal transmission, one can exclude
cultural or learned signals via timed matings and separation of mates after copulation
(12,21). Moreover, the paternal contribution can be isolated to sperm itself; with any
signals in seminal fluid excluded through techniques such as in vitro fertilisation
(21,23,24).
In my laboratory, we have reported the paternal transmission of a latent metabolic
phenotype (12). We found that when compared to genetically identical control mice,
the male offspring of obese pre-diabetic sires exhibited hepatic insulin resistance only
after post-weaning exposure to a high-fat diet. Interestingly, this latent metabolic
disorder was also able to be passed from F1 mice not challenged with a high-fat diet
(and thus metabolically normal), to their male offspring, the completely unexposed F2
generation. This F1-F2 inheritance was also associated with altered sperm ncRNA
profiles, with changes in the abundance of several sperm miRNAs and tRNA-
fragments.
We have also reported the paternally-transmissible effects of dietary methyl donor
supplement using the Agouti viable yellow (Avy) mouse model (19,25). The Avy allele
arose from the spontaneous insertion of an intracisternal A particle (IAP)

Introduction
4
retrotransposon upstream of the agouti gene. Unlike most other inserted
retrotransposons which are permanently silenced, the methylation status of the Avy IAP
is epigenetically variable and inversely correlates with expression. This leads to
variation both between isogenic mice as well as between clones of cells in the same
individual. Through its cryptic promoter, the variable Avy IAP can drive ectopic
expression of the downstream agouti gene. Under normal conditions (the original
agouti allele or a silenced Avy IAP), the agouti gene is only Avy expressed at a small
sliver of time in development, giving fur a single yellow band and an agouti colour.
However, after expression from the Avy IAP promoter, the agouti gene leads to a
spectrum of phenotypes ranging from mottled, mosaic yellow banding (indicating Avy
IAP expression in certain clones of cells) to a full coat of yellow fur (Avy IAP expression
in all clones of an individual). In my laboratory we were able to exploit this change in
coat colour as an epigenetic biosensor.
We showed that when the Avy allele is paternally-contributed, gestational dietary
methyl donor supplementation causes a shift in the coat colour spectrum towards a
pseudoagouti phenotype – that the Avy IAP promoter is more likely to be silenced (19).
After both selection for pseuodoagouti sires and continuous dietary methyl donor
supplementation the proportion of pseudoagouti offspring steadily increases from 10%
up to 45% in the 13th generation; this is a progressive and cumulative effect that does
not arise after either selection nor dietary challenge alone (25), suggesting that it is not
due to an induced genetic change. Furthermore, such an effect is reversible in the
short-term – offspring coat colour phenotypes revert to 10% in five generations after
cessation of dietary methyl donor supplementation (25). This heritable,

Introduction
5
environmentally-influenced effect meets all the hallmarks for an epigenetic basis; it
does not occur due to an induced genetic change and is reversible. This effect is not
observed when the Avy allele is maternally-contributed, implying that some sort of
signal about the environment is passed through the male germline.
1.1.3 Potential routes of transmission for an environmentally-induced
signal in the germline
It could be argued that the most parsimonious explanation for the effect of
environment on the germline is that the environmental stressor interacts directly with
the germline. However, in some cases this is unlikely or even extremely unlikely. In the
first instance, the germline is physically protected. In male mammals, the germline
segregates from the soma very early in development, and remains from molecules in
the circulation by structures such as the Sertoli cell barrier (SCB), also known as the
blood-testis-barrier. The SCB acts as a selectively permeable barrier allowing for the
traversal of germ cells as they differentiate while still controlling the biochemical
microenvironment. The SCB is of comparable permeability to the blood-brain barrier,
and is impermeable to all but a select few circulatory molecules (26-29).
Furthermore, in some examples of environmentally-induced inheritance, direct
signalling to the germline does not appear possible, and some sort of transduction of
the signal (by the soma) must be invoked. The prime example is a report by Dias &
Ressler (21), who detailed how a behavioural response to an odour is paternally
transmitted to offspring and grand-offspring. Olfactory receptors are only present in

Introduction
6
the brain and germ cells are innervate and have no way of interpreting or enacting a
behavioural response. For a psychological fear response to be transmitted through the
germline, some brain-derived signal must be passed through the circulation to the
germline in order for a behavioural response to manifest in the next generation. The
notion of a somatic-derived signal being passed through the germline to the next
generation is known as soma-to-germline feedback.
1.1.4 Molecular conduits of environmental information between
generations
The many examples of transgenerational environmental inheritance (TEI) are
demonstrably not due to a genetic or behavioural basis and have been called
“epigenetic”. However, the actual molecular mechanism in the germline has yet to be
elucidated. There are three proposed candidate mechanisms: direct chemical
modifications to DNA bases, chemical modifications to histone proteins, and the action
of small non-coding RNAs. Direct modifications to the DNA bases, most notably the
methylation of cytosine bases, have been implicated in some cases of
environmentally-induced transgenerational epigenetic inheritance, although available
evidence suggests that even diet (a major environmental influence) has little impact on
DNA methylation beyond that of stochastic changes (30). Furthermore, DNA
methylation is generally erased and reset during germ cell development, rendering this
mechanism unlikely to be responsible for cases of penetrant and/or multigenerational
effects. Modifications to histone proteins, eg histone methylation and acetylation, can
alter the chromatin state of DNA and availability of DNA for binding and

Introduction
7
transcription. While histone modifications may be present in the offspring, it is
unlikely that it is the mechanism responsible for heritability, since histones are
replaced with protamines when DNA is compacted in sperm. Small non-coding RNAs
are the most like candidate mechanism as they are known regulators of gene
expression that can be transported throughout the body to exert their effects at distal
locations.
1.1.4.1 Small RNAs
Small RNAs are <200 nt non-coding RNA molecules that regulate gene expression.
They include many types of RNAs which control gene expression in a variety of ways
at both the transcriptional and post-transcriptional level. The most well-known are
microRNAs (miRNAs) and tRNA-fragments (tRFs). miRNAs are 19-22 nt RNA
molecules able to bind to messenger RNA (mRNA) to inhibit translation and trigger
mRNA cleavage and degradation (31). They are able to target specific mRNA
transcripts based on sequence complementarity and recruit the other proteins to form
the RNA-induced silencing complex (32). Another type of small RNA is tRFs, of which
abundance is altered in sperm after environmental stress (24,33,34). tRFs are small 32 nt
sequences deriving from the 5’ or 3’ ends of mature tRNA genes (35). While their full
mechanism of action has not been fully understood, evidence exists that they regulate
translation (36,37) and RNA stability (38,39) and may have a similar mechanism of
action to miRNAs and bind to mRNAs based on complementary sequences to achieve
translational repression and mRNA degradation (40).

Introduction
8
The best evidence for small RNA being involved in inheritance comes from
invertebrates, in which the gene silencing effects of small RNAs can be inherited for
many generations, either with or without continual expression of the inducing small
RNA (9,10,41-43). In these studies, the small RNAs were expressed as a result of
ancestral age (43), in response to viral infection (9), or starvation-induced stress (10), or
acquired from diet (41,42). While C. elegans are unique in that small RNAs can have
systemic effects across the entire organism, there exist examples of vertical
transmission of small RNAs in mammals. One potential method for delivery of small
RNAs to the next generation is via the germline –through the sperm.
1.1.4.2 Sperm small RNAs are susceptible to environmental influence
Mammalian sperm lack the usually dominant ribosomal RNAs and instead are
especially rich in small non-coding RNAs, in particular miRNAs and tRFs (44,45).
While initially thought to be remnants of spermatogenesis, now it is accepted that at
least some RNAs found in sperm are selectively acquired during epididymal transit
(24,34,46). Sperm RNAs have been detected after fertilisation in the embryo (47), and
are likely functional. As sperm are transcriptionally quiescent (48), any acquisition of
sperm small RNA cargo must be dependent on other (somatic) cells (49), thus
providing a means for environmentally-influenced soma-derived RNAs to enter the
germline.
By now, there exist several examples of sperm RNA cargo being affected by
environmental factors. In the study by my laboratory described above in which the
offspring and grand-offspring of obese mice inherited a latent metabolic disorder, the

Introduction
9
sperm of the F1 mice (who were lean but able to transmit the latent metabolic disorder
to the F2 generation) contained altered sperm RNA profiles compared to control mice.
These changes include the increased abundance of the miRNAs miR-10a and miR-10b,
as well as alterations in the levels of many tRFs (12). Altered sperm RNA profiles have
also been induced by other dietary challenges (24,33), as well as chemical (34,50) and
psychological stress (21).
Preliminary evidence suggests that these altered sperm RNA profiles are at least
partially responsible for the observed heritable effects. In one example, small RNAs
that had changed in abundance – specifically the tRF-sized fraction isolated from the
sperm of mice exposed to a high-fat diet were injected into control zygotes (33). These
injected tRFs caused the same metabolic phenotype in offspring as was observed after
fertilisation with whole challenged sperm and were accompanied by changes in the
gene expression of metabolic pathways in the pancreas at the early embryo stage. To
further confirm that tRFs were responsible for the transmitted phenotype, the authors
attempted a similar experiment with artificially synthesised tRFs (as opposed to the
isolated tRF fraction), however this did not yield a similar phenotype. They postulated
that this may due to various RNA modifications that increase RNA stability as
evidenced by the longer half-life of isolated tRFs compared to synthetic ones, however
this remains to be verified.
1.2 Germline-associated somatic cells
If soma-to-germline feedback were possible, the most likely place for it to occur would
be at the somatic-germ cell interface. While signals could be passed from distally

Introduction
10
located somatic tissue through the circulatory system to the germline (as in the
olfactory response described above) (21), such signals might be dilute and difficult to
identify, especially in an initial proof-of-principle investigation. To circumvent these
issues, one can look at the somatic-germ cell interface, where somatic cells lie adjacent
to germ cells without the need for distal transport. Here I will go into detail about two
aspects of the male reproductive system relevant to this thesis.
1.2.1 Sertoli cells
Sertoli cells are somatic cells that line the seminiferous tubules in the testes of male
amniotes (51). They are often referred to as ‘guardians of the germline’ due to their
essential and protective roles in germ cell development. The intimate relationship, both
physiologically and proximally, between Sertoli cells and developing germ cells, as
well as the absence of any separating membrane or structure make them prime
candidates to investigate soma-to-germline feedback.
Sertoli cells are polarised, columnar cells that span the cross-section of seminiferous
tubules from basal membrane to the lumen (52) (Figure 1.1). They comprise the bulk of
the seminiferous epithelium, providing both structural integrity and shape. The
seminiferous epithelium is divided into two compartments by the SCB (Figure 1.1). The
SCB comprises junctional complexes between adjacent Sertoli cells, and physically
divides germ cells into two compartments based on their stage of differentiation (53).
Spermatogonia and early spermatocytes reside in the outer basal compartment of the
seminiferous tubules. As pre-leptotene spermatocytes complete DNA replication, they
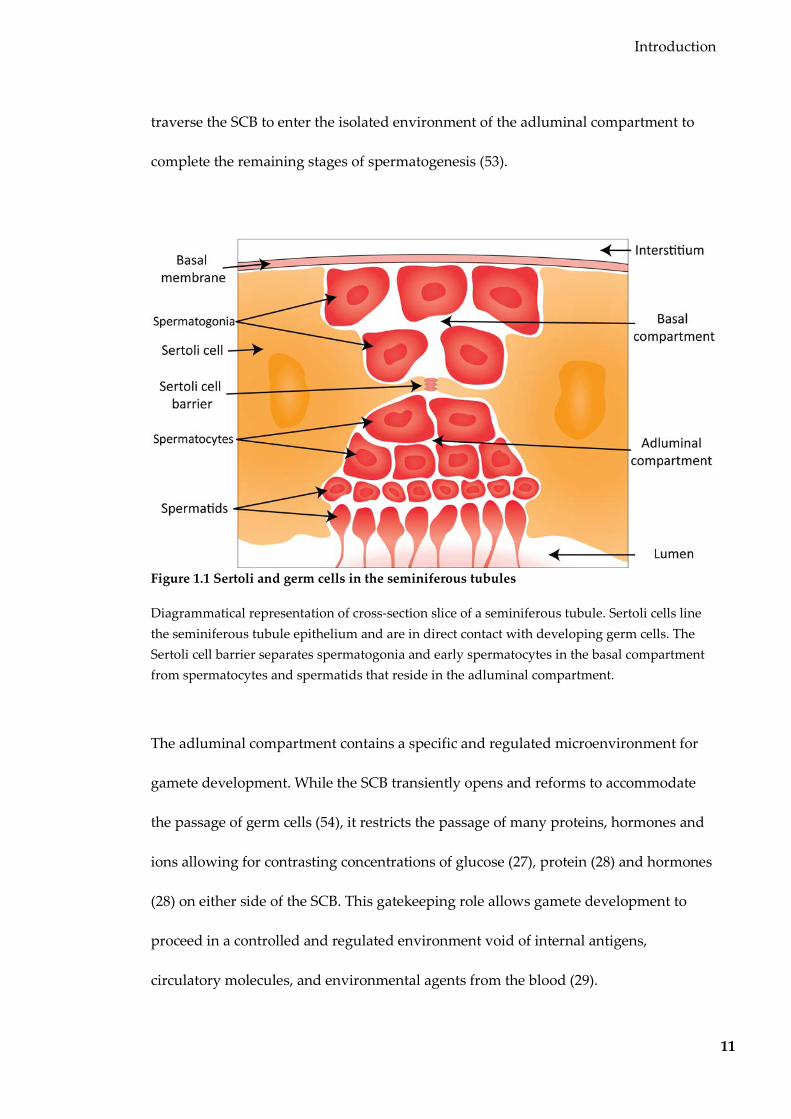
Introduction
11
traverse the SCB to enter the isolated environment of the adluminal compartment to
complete the remaining stages of spermatogenesis (53).
Figure 1.1 Sertoli and germ cells in the seminiferous tubules
Diagrammatical representation of cross-section slice of a seminiferous tubule. Sertoli cells line the seminiferous tubule epithelium and are in direct contact with developing germ cells. The Sertoli cell barrier separates spermatogonia and early spermatocytes in the basal compartment from spermatocytes and spermatids that reside in the adluminal compartment.
The adluminal compartment contains a specific and regulated microenvironment for
gamete development. While the SCB transiently opens and reforms to accommodate
the passage of germ cells (54), it restricts the passage of many proteins, hormones and
ions allowing for contrasting concentrations of glucose (27), protein (28) and hormones
(28) on either side of the SCB. This gatekeeping role allows gamete development to
proceed in a controlled and regulated environment void of internal antigens,
circulatory molecules, and environmental agents from the blood (29).

Introduction
12
Since the SCB is selective to many nutrients and hormones, Sertoli cells also function to
regulate and maintain the specific microenvironment required for spermatogenesis.
Sertoli cells secrete a variety of products into the adluminal compartment including
proteins, proteases, nutrients, hormones, and extracellular matrix components (55).
Many of these secretory products play a role in spermatogenesis, and some have
protective roles and mitigate the effects of germ cell stress, such as the production of
clusterin in response to heat stress (56).
1.2.1.1 Effects of endocrine-disrupting chemicals on Sertoli cell function and
spermatogenesis
Sertoli cells are essential for spermatogenesis, and therefore, any disruption to Sertoli
cell function and the permeability of the SCB can disrupt spermatogenesis. The
interactions between Sertoli cells and developing germ cells can be disrupted by the
effect of various stressors. Androgens are one of the main regulators of the
permeability of the blood-testis-barrier (57), and hence any environmental factors that
influence androgen levels have the ability to influence germ cells. Of particular
relevance is the effect of chemicals that interfere with hormone receptors – endocrine-
disrupting chemicals (EDCs). EDCs include heavy metals (58), fluorocarbons (59), and
pesticides (60). Perhaps the most well-known EDC is Bisphenol A (BPA). BPA is a
xenoestrogen used in the manufacturing process of polycarbonate plastic and epoxy
products. Humans are exposed to BPA both indirectly from the environment and
directly from many plastic products including food and drink containers (61), dental
sealants (62), and thermal printing paper (63). BPA is detectable in human sweat,

Introduction
13
blood, and urine (64-66), and has been detected at increasing concentrations (67),
coinciding with the increased environmental prevalence and consumption of BPA (68).
Although BPA is not mutagenic, exposure has been linked to many deleterious effects
in mammals. BPA exposure has been linked to poor reproductive health, including
meiotic defects during oogenesis (69,70), precocious female puberty (71,72), and
diminished ovarian capacity (73). In Rattus norvegicus, The effects of BPA have also
been reported to extend vertically beyond the exposed generation, resulting in
decreased ovarian, uterine and seminal vesicle weights in up to the F3 generation (74).
BPA has also been linked to male reproductive defects. Several studies in humans have
reported that urinary BPA concentration is inversely correlated with sperm counts,
vitality, and motility (75,76) and sperm DNA damage (77). Such changes in sperm raise
the possibility that effects of BPA can be passed down to offspring via the germline. In
zebrafish, adult BPA exposure resulted in an altered sperm mRNA profile not only in
directly exposed fish, but also in their offspring, along with cardiac abnormalities for
two generations (78). Little is known about the underlying mechanism behind
BPA-induced effects on sperm. A possible mechanism for these alterations in sperm is
through Sertoli cells. BPA is able to interact with Sertoli cells via estrogen receptors, as
well as other alternate endocrine signalling pathways (79), to alter cellular miRNA,
mRNA and protein (80-82). Any disruption of Sertoli cell function and the SCB may
form the basis for the observed sperm defects. It is possible that BPA exposure causes
transgenerational effects via changes to sperm RNA.

Introduction
14
1.2.2 The epididymis
At the completion of spermatogenesis, immature and immotile spermatids are released
from the protective environment of the seminiferous tubule epithelium into the
seminiferous tubule lumen. Following the flow of seminiferous tubule fluid, these
spermatids are transported through the rete testis to the epididymis where they
undergo maturation. The epididymis is a long, convoluted tube with an extended
length of 1-2 m in mice, 6 m in humans and up to 80 m in stallions (83). The tube is
coiled into a crescent-shaped structure that can be divided into three segments (Figure
1.2); the caput (head) where immature sperm enter from the rete testis, the corpus
(body) and the cauda (tail), where motile sperm are stored prior to exit to the vas
deferens (84).

Introduction
15
Figure 1.2 Diagrammatical representation of the epididymis and its segments
After spermatogenesis, immature spermatids flow through the rete testis to the epididymis. Immature sperm acquire their motility and ability to fertilise the ovum during their ~7 day transit in the caput and corpus epididymis. Mature, motile sperm accumulate in the caudal segment before being carried to the ejaculatory duct via the vas deferens.
The epididymis has functions pertinent to sperm development and reproduction.
During sperm transit, epididymal cells absorb much of the fluid from around the
sperm, effectively concentrating sperm and providing the pressure gradient required
for immotile sperm to flow through the epididymis (85). Transit through the
epididymis is also required for spermatozoa to acquire motility and the ability to
fertilise the egg.
Sperm are transcriptionally and translationally inert (48), and so this maturation is
driven almost entirely by interactions with the epididymal microenvironment
(reviewed in (86)). The three segments of the epididymis all have slightly different

Introduction
16
functions, with early and late maturation occurring in the caput and corpus
respectively (86). These segments are then further sub-divided by connective tissue
septa functioning to give each sub-segment a different physiological environment (87).
Sperm maturation involves the gain, loss and modification of proteins (and lipids to an
extent) (88) in sperm in the epididymis (89-92).
1.2.2.1 Environmental influence on epididymal cells
Since the epididymal microenvironment is crucial for sperm maturation, any
environmental-induced changes to the epididymis might impact upon the germline. A
wide range of toxicants affect parameters such as the sperm proteome, transit time,
concentration, motility and fertility (reviewed in (93)), and oxidative stress has been
shown to alter the sperm maturation process to ultimately reduce sperm number,
motility and viability and increase the proportion of abnormal sperm (94).
Of particular interest is the effect of more subtle environment stressors, i.e., those that
do not obviously affect sperm viability and/or motility. A low-protein diet was found
to alter the epididymal cell and sperm small RNA profile in mice (24).This dietary
challenge increased the abundance of tRF5-Gly-GCC, which was then shown to have a
functional effect in embryos. Another study detailed how chronic ethanol exposure
alters not only the abundance of tRFs in mature sperm, but also their modifications
(34). This suggests that changes to sperm RNA profile are associated with epididymal
transit.
Interestingly, similarly altered small RNAs were also detected in the epididymal EVs,
implying that EVs are responsible for the modified sperm RNA profiles. While there

Introduction
17
has been no direct link between the altered epididymal EV and sperm RNAs and the
phenotypic effects caused by a low-protein diet and chronic ethanol exposure, these
studies show that EVs are a possible conduit for the environment to influence the
germline.
1.3 Extracellular vesicles
Extracellular vesicles are lipid-enclosed vesicles that are produced by living cells. They
are released by most, if not all cells, and can be detected in a range of biological fluids
(blood, cerebrospinal fluid, semen, breast milk, saliva, ascites). They contain a subset of
nucleic acid and protein from their cell of origin, and act as intercellular messengers by
delivering their cargo to specific recipient cells. The recent surge in publications on
extracellular vesicles in the last two decades has resulted in many names used to
describe extracellular vesicles, often interchangeably (95), including argosomes,
ectosomes, exosomes, microparticles, microvesicles and oncosomes (96-101). In this
thesis, I use the term extracellular vesicles (EVs) to refer to two classes of vesicles
defined by their method of biogenesis: microparticles and exosomes. Apoptotic bodies
(ABs), larger vesicles that are shed by moribund cells, are not included in the term
extracellular vesicles in this thesis.
Microparticles were first referred to in 1946 as lipoproteins of very high particle weight
that activated blood coagulation (102). In 1967 they were correctly identified as
lipid-enclosed particles released from platelets, and imaged by transmission electron
microscopy (103). It is now known that microparticles are 100-1000 nm EVs that are
formed by membrane budding (Figure 1.3a). Budding is caused by the translocation of

Introduction
18
the lipid phosphatidylserine to the outer leaflet of the phospholipid bilayer caused by
the activation of floppase and inhibition of flippase and scramblase enzymes (104,105).
Microparticles are generally produced at a low basal level, but production can be
increased after cell activation or stress (106). They are not to be confused with ABs,
larger 500-5000 nm vesicles that are produced only from moribund cells (107).
Exosomes were first described in 1981 by Trams et. al. as a smaller sub-population of
EVs isolated from rat C6 glioma cells alongside microparticles (108). Since this time, it
has been found that exosomes are a distinct type of EV produced constitutively by
cells, by an internal process that is distinct from the processes involved in microparticle
production (Figure 1.3a). First, intraluminal vesicles (ILVs) are formed inside
endosomes by invaginations of the endosomal membrane. This process can be
regulated by ‘endosomal sorting complex required for transport’ (ESCRT) protein
complexes (109) or via movement of the lipid ceramide (109,110). Endosomes
containing ILVs are called multi-vesicular bodies (MVBs). Some MVBs are destined for
degradative or recycling fate, while those containing ILVs travel to the cellular surface
for secretion (111). These MVBs then fuse with the cellular plasma membrane,
releasing any contained ILVs as exosomes from the cell.

Introduction
19
Figure 1.3 The biogenesis and fate of extracellular vesicles
(a) Microparticles are shed directly from the cell surface while exosomes are produced inside endosomes and released after endosomal fusion with the cell membrane. (b) EVs can release their cargo to the extracellular space, to recipient cells, trigger cellular signalling cascades, as well as be endocytosed into cells for future fusion or release. Adapted from original by Sally Eaton.
The only definitive way to determine whether EVs are exosomes or microparticles lies
in observing their method of formation, however this is usually difficult, if not
impossible. In general, and in most studies, particle size is often used as a surrogate
criterion, with 30-150 nm sized EVs deemed exosomes and 100-1000 nm sized EVs
deemed microparticles. In some cases, EV cargo can also shed light on their method of
biogenesis and thus type, for example exosomes produced by ESCRT machinery often
contain ESCRT proteins.

Introduction
20
1.3.1 EVs contain selectively packaged cargo
Regardless of their method of production, extracellular vesicles contain molecules from
their producing cell. These include RNA (100,112-115), protein (114,116), and
controversially, DNA (115,117,118). Interestingly, in many cases EV cargo is not
necessarily representative of the contents of the producing cells: some proteins or
RNAs appear to be enriched, or depleted, in EVs relative to their parent cell
(100,112,119-122). Often, this occurs to the extent where the cargo is undetectable in
analysis of the parent cell, but abundant in EVs, and vice versa (100,114,115,119,123).
Selective packaging of proteins in EVs has been observed in glioblastoma EVs, where
many angiogenic proteins were detected at much higher concentrations compared to
cells (114). In this case, the enriched proteins provide a mechanism for how tumour-
derived EVs are able to contribute to angiogenesis. In terms of RNA, this phenomenon
of ‘selective packaging’ is most notable by the absence of intact ribosomal RNA and the
enrichment of certain small RNAs in EVs (98,100,114,124). Work from my laboratory
showed that compared to their parent cells, glioblastoma EVs are enriched with repeat-
derived, intergenic, and tRNA-derived RNA transcripts but depleted in miRNAs (100).
Additionally, within the miRNA population, EVs and cells also exhibited 5p/3p arm
switching; the differential preference for transcripts processed from the two arms of the
mature miRNA molecule. The presence of DNA encapsulated in EVs is controversial,
with several studies reporting the isolation of DNA from EVs without prior DNase
treatment (115), suggesting that that the isolated DNA may likely be extraneous and
carried on the surface of EVs (125). However, a select few studies using unconventional

Introduction
21
reverse transcription techniques report the presence (and sometimes enrichment) of
certain types and sizes of DNA inside EVs (117,118,126). Several studies also report no
detectable DNA inside EVs (98,108), however it remains possible that DNA is present
in EVs produced by certain cell types, and/or not able to be isolated by conventional
methods.
This selective sorting of cargo is believed to be due to part of the biogenesis
mechanism. In exosomes this is believed to be through the action of ESCRT proteins
(109), which select cargo based on marks (such as ubiquitin on proteins) (128), while it
is yet to be elucidated for microparticles.
1.3.2 EVs are intercellular messengers
EVs, with their selectively packaged cargo can interact with (and hence influence)
other cells (Figure 1.3b). If lysed in the extracellular space, proteins and nucleic acids
released from EVs may go on to interact with cells in the proximity or with the
extracellular matrix (ECM) itself (129,130). Intact EVs can interact with select recipient
cells, in multiple ways. Receptor-mediated can result in the triggering of signalling
cascades within the targeted cell (131,132). EVs can also fuse with the recipient cell
membrane, followed by the exchange of cargo. EVs may also be actively internalised
into targeted recipient cells by endocytosis, in a seeming reversal of the exosome
biogenesis process. This results in EVs remaining intact inside endosomal pockets;
internalised EVs may then release their contents into the recipient cell (114,133,134), or
even be re-released from this cell (135). These interactions do not occur between EVs

Introduction
22
and all types of cells – they are restricted to specific cell types based on complementary
surface proteins on EVs and cells (136-138).
EVs can interact with recipient cells that are both close to the site of origin, as well as be
carried to locations all over the body via circulatory fluids such as blood. The
long-range reach of EVs and their ability to target specific tissues are best illustrated in
a proof-of-principle study by Alvarez-Erviti et. al. (137). In this study, exosomes were
engineered to contain RVG, a neuron-specific peptide on their membrane, and loaded
with GAPDH short-interfering RNA (siRNA). When injected into the tail vein of mice,
these exosomes travelled through the circulation where they were able to cross the
blood brain barrier and accumulate in the brain. This was also accompanied by a brain-
specific knockdown of GAPDH, by siRNA delivered by the exosomes.
1.3.3 Functional role of EVs in regular physiology and pathology
In regular physiology (not associated with diseased or pathological states), EVs have a
variety of roles. Such functions were apparent during the first platelet-derived
microparticles discovered that found to carry Tissue Factor and thus have clotting
ability (102,103,139). EVs have also been implicated in a protective/recovery response
to injury. EVs released from human bone marrow mesenchymal cells and liver resident
stem cells aid in the recovery after glycerol-induced acute kidney injury (119,140) and
liver regeneration after hepatectomy (141). The same research group also reported how
EVs from endothelial progenitor cells interacted with endothelial cells to not only
prevent starvation-induced apoptosis but to also promote angiogenesis (120).

Introduction
23
Interestingly, these three functions are believed to be mediated by RNA that was
bound to the surface of EVs and thus is sensitive to RNase treatment, rather than
encapsulated, internalised RNAs (119,120,140).
EVs are perhaps best known for their role in cancer. For example, the number of EVs
released from tumour cell lines is correlated with their metastatic ability (113).
Furthermore, tumour-derived EVs were found to be able to bind and enter monocytes,
where they were able to prevent apoptosis (113). Cancer-derived EVs may also release
their metalloproteinase cargo into the extracellular space to degrade the ECM and
facilitate tumour mobility (130). EVs released by primary glioblastoma cells are
endocytosed by brain microvascular endothelial cells (114). This is followed by the
transfer of many functional RNAs that result in increased the ability for tumour
growth and invasiveness (114). EVs released by prion-infected cells also contain the
misfolded, infectious PrPSc protein, and can spread prion infection (142).
1.3.4 Extracellular vesicles in reproduction
Specifically, EVs have known roles in mammalian reproduction. EVs are present in
ovarian follicular fluid, either from the surrounding granulosa or passing through the
blood-follicular barrier (143). These EVs contain RNAs that can be transferred to the
oocyte, and most likely play a role in the development of the oocyte RNA profile
during maturation (143-145). A similar nurturing role during germ cell development is
also observed in the epididymis. Sperm acquire their motility and ability to fertilise an
egg during their transit through the epididymis. This process is mediated in part by the

Introduction
24
exchange of proteins with epididymal EVs, particularly the acquisition of proteins
associated with acrosome formation, sperm-egg recognition, zona pellucida binding
and fertilisation (89-92).
More recently, evidence has emerged that sperm RNA composition develops during
epididymal transit (46) This is believed to be due to interactions with the epididymal
EVs (24,49), and are likely to have a role in sperm maturation that has not been fully
understood. There exists evidence that epididymal EV cargo can be influenced by
environmental factors. A low protein diet and chronic exposure to ethanol result in
alterations to sperm RNAs (24,34). In these two cases, similar RNAs were identified in
epididymal EVs, which are believed to be the source of the modified sperm RNAs,
suggesting that EVs act as a messenger of environmentally-influenced small RNAs and
the next generation through the sperm.
1.4 Hypothesis and aims
As detailed above, there are a plethora of examples where environmentally-induced
effects are inherited in a pattern that cannot be exampled by classical Mendelian
genetics. While there are many proposed mechanisms, one overlooked possibility is
through small RNAs transferred by EVs. EVs are produced by nearly all cells (and
upregulated in stressed conditions), and carry molecular cargo that can influence the
behaviour of specific recipient cells either locally or via transport in the bloodstream.
Importantly, EVs are rich in small RNAs that have the potential to alter the gene
expression and thus phenotype of recipient cells. While it is known that EVs are
produced by cells of the reproductive system, little is known about their small RNA

Introduction
25
cargo, whether it is susceptible to environment stressors, and the downstream effects of
this cargo in germ cells.
The overarching hypothesis driving this study is that intergenerational communication
of prevailing environmental conditions can occur via soma-to-germline feedback
mediated by small RNAs within germline-associated EVs (146). This hypothesis
requires that (a) EVs are produced from somatic cells that are capable of interaction
with germ cells or gametes, (b) that these EVs carry small RNA cargo, and (c) that the
composition of this EV RNA cargo is susceptible to the environmental influences.
In this thesis I take the first steps towards testing this hypothesis by using
germline-associated somatic cells from the testis and epididymis. I have chosen to
interrogate germline-associated EVs rather than circulatory EVs for reads of
tractability. Germline-associated somatic cells are known to have an intimate
relationship with the germline, and thus any EVs they produce will most likely be in
direct contact with germ cells.
Hence the specific aims of this thesis are to:
1. Isolate and characterise EVs from germline-associated somatic cells.
2. Determine the nature of interaction between soma-derived EVs and germ cells
3. Investigate the effects of a change in environment on EV production, cargo and
germ-cell interactions.

Introduction
26
I begin in Chapter 3 by isolating EVs from a Sertoli cell line and determining if Sertoli
EVs are able to interact with spermatogonial stem cells. Then I investigate the effects of
Bisphenol A exposure on Sertoli cell EV production, cargo and germ-cell interactions.
In Chapter 4, I examine EVs produced by an epididymal cell line, and investigate the
effects of folate availability on epididymal EV production and cargo. In Chapter 5, I
take my investigations into a mouse model, and investigate the effects of dietary
methyl donor supplementation on epididymal EV production and cargo.

27
2 MATERIALS AND METHODS
2.1 Cell culture and tissue collection
2.1.1 Cell lines and culture conditions
The TM4 Sertoli cell line (147) was purchased from the ATCC (CRL-1715). Cells were
cultured in a 1:1 mixture of Dulbecco’s Modified Eagle’s Medium (DMEM) and Ham’s
F-12 nutrient mixture (Gibco 11320-033) supplemented with 3.151 g/L D-glucose,
2.5 mM L-glutamine, 0.5 mM sodium pyruvate, 5% horse serum (Gibco 16050-122) and
2.5% fetal bovine serum (FBS; Gibco 26140-079). Sertoli Cells were grown at 37oC in 5%
CO2. For Bisphenol A (BPA) sensitivity assays, BPA powder (Sigma Aldrich 133027)
was dissolved in dimethyl sulfoxide (DMSO; Sigma Aldrich D2650), filtered sterilised
through a 0.2 µm polytetrafluoroethylene syringe filter (Merck Millipore), and stored
at 4°C for use within two weeks. Sertoli cells were seeded in a 12-well plate in
increasing concentrations of BPA from 20 to 60 µg/ml (with constant concentrations of
DMSO). At confluence after two days of growth, cells were trypsinised and counted
using a Z2 particle size and count analyser (Beckman Coulter) and viability assessed by
trypan blue exclusion.
The DC2 epididymal epithelial cell line (148) was kindly provided by Prof. Marie-
Claire Orgebin-Crist (Vanderbilt University Medical Centre). They were cultured in
Iscove’s Modified Dulbecco’s Medium (IMDM; Gibco 12440-053) supplemented with
4.5 g/L D-glucose, 4 mM glutamine, 1 mM sodium pyruvate, 0.1 mM non-essential

Materials and Methods
28
amino acids (Gibco, 11140-050), 1 nM 5α-Dihydrotestosterone (Cerilliant, D-073) and
10% FBS (Gibco 26140-079). DC2 cells were grown at 33oC in 5% CO2.
The C18-4 spermatogonial stem cell line (149) was kindly provided by Prof. Marie-
Claude Hofmann (MD Anderson Cancer Center). They were cultured in DMEM (Gibco
11965-092) supplemented with 4.5 g/L D-glucose, 4 mM glutamine and 10% FBS (Gibco
26140-079). C18-4 cells were grown at 33oC in 5% CO2.
For subculturing, all cell lines were trypsinised with 0.05% trypsin/EDTA (Gibco
25300-054, washed once and passaged at a ratio of 1:8 at 3-6 day intervals. Cell
concentrations were obtained using a Z2 particle size and count analyser (Beckman
Coulter) and cell viability was measured at each collection using trypan blue exclusion.
2.1.2 Epididymides dissection and epididymal fluid collection
All animal work was approved by the Garvan/St Vincent’s Animal Ethics Committee
under AEC# 16/31, and performed in accordance with the guidelines of the ‘Animal
Research Act 1985’, the ‘2010 Animal Research Legislation’ and the ‘8th Edition of the
Australia code of practice for the care and use of animals for scientific purposes from
2013’ (NHMRC, Australian Government).
Epididymides were dissected from mice euthanised by CO2 asphyxiation. Dissected
epididymides were opened by slicing with a scalpel and incubated on a heat pad for 30
minutes in 1 ml modified Biggers, Whitten and Whittingham (BWW) media (91.5 mM
NaCl, 4.6 mM KCl, 1.7 mM CaCl2.2H2O, 1.2 mM KH2PO4, 1.2 mM MgSO4.7H2O, 25 mM

Materials and Methods
29
NaHCO3, 5.6 mM D-glucose, 0.27 mM sodium pyruvate, 44 mM sodium lactate, 5
U/mL penicillin, 5 µg/mL streptomycin, 20 mM HEPES buffer and 3 mg/mL bovine
serum albumin) to allow for the diffusion of epididymal fluid. Tissue and sperm were
removed by centrifugations at 50 x g for 3 minutes, 500 x g for 3 minutes, and 1000 x g
for 10 minutes. Epididymal fluid was filtered through a 0.45 µm polyethersulfone
syringe filter (Merck Millipore) and then stored at -20°C for EV purification.
2.1.3 Sperm isolation
Fresh cauda epididymal spermatozoa were isolated via backflushing from the
epididymides. Epididymides with vas deferens attached were dissected from mice
euthanised by CO2 asphyxiation. The cauda segment and vas deferens were
submerged in water-saturated mineral oil heated to 37°C. The vas deferens were
cannulated with a stretched glass tube and secured with silk sutures and a small
incision was made in the tube between the corpus and cauda epididymis. A syringe
attached to the glass pipette was used to backflush sperm, which was collected in a
microcapillary tube from the site of incision. Sperm from each epididymides were
resuspended in 1 mL of modified BWW media and sperm vitality were confirmed by
microscopy.
2.2 Extracellular vesicle purification and characterisation
2.2.1 Preparation of EV-depleted media

Materials and Methods
30
EVs are abundant in HS and FBS, two constituents of culture media. To ensure that
isolated EVs had originated from cell lines and were not endogenous from serum, an
extended ultracentrifugation step was employed to deplete serum of over 95% EVs
(150). HS and FBS was ultracentrifuged at 100,000 x g (average) for 18 hours at 4°C
using a Beckman L-80 floor ultracentrifuge. The EV-depleted supernatant was
aliquoted and stored at -20°C until use.
2.2.2 Cell line EV production
Before seeding flasks for EV harvest, cells in exponential growth phase were washed
twice with PBS to remove any residual EVs. Cell pellets were resuspended in media
made with EV-depleted serum and approximately 2 x 106 cells were seeded into each of
10 x 150 cm2 flasks. For some experiments, Sertoli cells were treated with 20 µg/ml BPA
in DMSO solvent (0.01% final DMSO concentration). BPA treatment started 12 days
prior to EV harvest (9 days prior to seeding cells for EV harvest). For some
experiments, DC2 cells were grown in media with a physiological folate concentration,
prepared by diluting regular IMDM media with custom no-folate IMDM media
(Gibco) at a ratio of 1:127. For all experiments, a corresponding aliquot of cells were
snap-frozen at the time of EV harvest, and stored at -70°C until required for RNA
analyses.
To limit the contamination of EV preparations with ABs produced in response culture-
induced stress, media was collected from flasks with cells that were still in exponential
growth phase with over 95% viability. Detached cells and cell fragments were pelleted

Materials and Methods
31
from the conditioned media by centrifugations at 200 x g for 10 minutes at RT,
followed by a second centrifugation at 1000 x g for 10 min at RT. Any remaining
cellular debris and ABs were removed via filtering conditioned media through a 0.45
µm polyethersulfone syringe filter (Merck Millipore). The resultant pre-cleared
conditioned culture media were stored at -20°C until ultracentrifugation.
2.2.3 EV pelleting by ultracentrifugation
EVs were pelleted from pre-cleared conditioned cell culture media and pooled
epididymal fluid by ultracentrifugation at 100,000 x g (average) for 90 minutes at 4°C
using a Beckman L-80 floor ultracentrifuge. EV pellets were resuspended in 100-200 µl
PBS and stored at -20°C.
2.2.4 Nanoparticle tracking analysis
EV size and concentration was determined by Nanoparticle Tracking Analysis (NTA)
(149) using a NanoSight LM10-HS (Malvern Panalytical) at the Bosch Molecular
Biology Facility (University of Sydney). Samples were diluted with PBS to achieve a
density of 20-80 particles/screen, the optimum density suggested by the manufacturer.
For each EV sample, three 60 second video tracks were recorded at constant 25°C with
the 532 nm laser module and analysed using NTA Software v3.2 with default
parameters.

Materials and Methods
32
2.2.5 Transmission electron microscopy
EV suspensions were adsorbed onto formvar-coated 200 mesh copper grids for one
minute and excess liquid removed by wicking. Grids were negative stained and fixed
with a 2% uranyl acetate solution for 60 seconds and then washed three times in PBS
before being air-dried. Grids were imaged on a JEOL TEM-1400 transmission electron
microscope with an 80 kV electron beam. Grids were imaged at the Electron
Microscope Unit (University of New South Wales).
2.2.6 Protein mass spectrometry
Resuspended EVs were lysed in 50 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100,
and 1% sodium deoxycholate, and protein concentration determined by Direct-detect
infra-red spectroscopy (Merck Millipore). Ten micrograms of EV proteins were
separated by 2D gel electrophoresis on a 4-12% bis-tris polyacrylamide gels for 35
minutes at 200 V in MES SDS running buffer. Gels were stained with SimplyBlue
SafeStain (Life Technologies LC6060). The EV lane was excised and divided into five
segments, with each segment processed separately for LC-MS/MS at the Bioanalytical
Mass Spectrometry Facility (University of New South Wales). Peptide traces were
searched against the SwissProt database using MASCOT (v2.6.0). Proteins were then
subjected to a 0.05 significance threshold and abundance filters of >1 significant
peptide match and >5% protein sequence coverage.
In data analysis, comparisons were made with proteins from two EV cargo compendia:
ExoCarta (v5, downloaded on 2016/11/09) (152) and Vesiclepedia (v3, downloaded on

Materials and Methods
33
2016/06/28) (153). Functional enrichment analysis was performed using the Database
for Annotation, Visualisation and Integrated Discovery (DAVID) v6.8 with all mouse
proteins used as a background set. p-values were adjusted by the Benjamini-Hochberg
correction algorithm to account for multiple hypothesis testing (154).
2.3 Imaging and microscopy
2.3.1 EV labelling
2.3.1.1 Membrane labelling
Purified EV preparations were labelled with PKH26 (Sigma Aldrich MINI26) lipophilic
membrane dye following the manufacturer’s protocol. Briefly, a dye solution
comprising 2 µl dye in 498 µl Diluent C (Sigma Aldrich CGLDIL) was added to a 500
µl mixture of EVs in Diluent C. After three minutes, labelling was halted by an
addition of at least 10 mL EV-depleted media, prior to EV pelleting by
ultracentrifugation as in 2.2.3.
2.3.2 EV-cell co-incubation
2.3.2.1 Live cell imaging
For live-cell imaging, approximately 2 x 104 C18-4 cells were seeded into each well of a
24-well plate. After cells had attached overnight, PKH26-labelled Sertoli EVs were
added to wells at an approximate ratio of 100 EVs per cell. The same volume of dye
control (PKH26-labelled PBS) was added to control wells. Images were captured every
20 minutes for 15 hours in the tissue culture incubator with the JuLi Stage real time cell

Materials and Methods
34
history recorder (NanoEntek), using white light and a 525/50 blue LED and a 580 nm
long pass filter for PKH26 fluorescence. Images were cropped and overlaid using
custom scripts in ImageJ v1.51j.
2.3.2.2 Labelled-EV coincubation for confocal microscopy
2.3.2.2.1 Adherent cell lines
Approximately 1.2 x 106 C18-4 cells were seeded onto 22 mm x 22 mm glass coverslips
placed in a 6-well plate. After two hours, labelled Sertoli EVs in fresh EV-depleted
media were added to C18-4 cells at an approximate ratio of 100 EVs per cell. EVs and
cells were co-incubated for three hours before three PBS washes to remove unattached
EVs.
2.3.2.2.2 Sperm
Labelled EVs were resuspended in BWW media and added to living spermatozoa in
1.5 mL Eppendorf tubes at an approximate ratio of 100 EVs per cell. Sperm and EVs
were left to co-incubate for three hours at 33°C. Co-incubation was ceased by three PBS
washes to remove unattached EVs.
2.3.2.3 Confocal imaging
Immediately following co-incubation, cells on coverslips and live sperm in suspension
were nuclear-stained with Hoechst 33342 nuclear stain (Thermo Fisher H3570) for 30
minutes in the incubator and then washed three times with PBS. Adherent cells and
sperm were fixed with 4% paraformaldehyde for 30 minutes on ice and washed three
times with PBS. Adherent cells/sperm were mounted with coverslips on glass slides in

Materials and Methods
35
7 µl Prolong Gold antifade mountant (Thermo Fisher P10144) and left to cure overnight
in the dark. Coverslips were sealed with nail polish prior to confocal laser imaging on a
Zeiss 720 upright or Zeiss LSM 7 DUO microscope using Zen 2.3 software. Slides were
imaged using a 63x oil-immersion objective lens and 4-fold line averaging. Hoechst
33343 nuclear stain was captured between 410-490 nm after excitation with a 405 nm
laser, CFSE between 496-545 nm after excitation with a 488 nm laser, and PKH26
between 564-621 nm after excitation with a 560 nm laser, with each fluorophore imaged
sequentially.
2.4 RNA analyses
2.4.1 Extraction
Prior to RNA extraction, purified EVs were treated with 50 µg/ml RNase A (Ambion
AM2270) for 30 minutes at RT to remove any unprotected, residual RNA not protected
within EVs. RNA was extracted from EVs and cells using Trizol LS (Invitrogen
10296028) following the manufacturer’s protocol with the exception that 10 mM of
MgCl2 was added to the Trizol LS-sample mixture in order to reduce precipitation bias
(155), and all centrifugation steps were performed at 21,000 x g at 4°C. Phase
separation was conducted in MaXtract High Density gel Tubes (Qiagen 129046) with
an addition of 0.8 volumes chloroform. After centrifugation for 15 minutes, the
RNA-containing aqueous layer was transferred to a new tube. Ten micrograms of
RNase-free glycogen (Invitrogen R0551) was added as a carrier followed by
precipitation with two volumes of isopropanol for 10 minutes at RT. RNA was pelleted
for 10 minutes, washed with 75% ethanol, and then resuspended in RNase free water.

Materials and Methods
36
RNA concentration and integrity were determined using Small RNA, RNA Pico and
RNA Nano chips on an Agilent Bioanalyser (Agilent). RNA was stored at -70°C for
downstream analyses.
2.4.2 Gene expression microarray analysis
Microarrays were performed on RNA from C18-4 cells with and without exposure to
TM4 EVs. C18-4 cells were seeded into a 6-well plate at a density of 1.2 x 105 cells/well
(equivalent to 1:10 split). After four hours when cells had attached, 1.56 x 107 EVs were
added (equivalent to 130 EVs per cell at confluence). After 24 and 48 hours, media was
replaced and an equivalent amount of EVs were added. For the control group,
equivalent volumes of PBS were added. Eighteen hours after the last addition, cells
were washed three times with PBS to remove unattached EVs and Trizol LS was
directly added to cells for RNA extraction. Extracted RNA was quantified and assayed
for purity and integrity using a ND-1000 UV spectrophotometer and an RNA Nano
Chip on an Agilent Bioanalyser Total RNA was used in microarray with a GeneChip
Mouse Gene 2.0 ST whole-transcript arrays (Affymetrix 902119) by the Ramaciotti
Centre for Genomics (University of New South Wales).
Raw microarray data files were processed in Microsoft R Open v3.4.3 using the oligo
v1.42.0 package. Microarray chips were normalised using the RMA (robust-multi-array
average) method with quantile normalisation. A background noise threshold was
calculated by the 95th percentile intensity of all antigenomic probes. Transcripts with
fewer than three samples meeting this background threshold were deemed not

Materials and Methods
37
expressed and removed. Differential expression analysis was performed in Microsoft R
Open v3.4.3 using the limma v3.34.9 package.
2.4.3 Small RNA libraries
2.4.3.1 Small RNA library construction
Small RNA libraries were constructed using the NEBNext Small RNA Library Prep Set
for Illumina (New England Biolabs E7330) essentially following the manufacturer’s
protocol. To increase yields, 3’ SR and 5’ adaptors and the 3’ SR RT primer were
diluted 1:1 in H2O, and an overnight 3’ adaptor ligation at 16°C was used. Adaptor-
ligated RNA was then reverse transcribed to form single-stranded complementary
DNA and then amplified by 15 cycles of PCR. Amplified libraries were electrophoresed
on a vertical 6% polyacrylamide gel in TBE buffer at 150 V for 90 minutes. Gels were
stained with SYBR Gold (Invitrogen S11494) and visualised on a Fujifilm FLA-5100
fluorescent imager (Fuji) through an LPB filter after excitation at 473 nm. Bands
corresponding to libraries with 18-35 nt inserts were excised and library products were
purified using a QiaQuick PCR purification kit (Qiagen 28104). Purified libraries were
verified and quantified on a DNA High sensitivity chip on an Agilent 2100 Bioanalyser
(Agilent).
2.4.3.2 Small RNA sequencing and processing
Purified libraries were pooled to achieve an average read depth of 10-20 million reads
per sample. Pools were sequenced in multiplex on an Illumina NextSeq 500 in rapid
mode at the Ramaciotti Centre for Genomics (University of New South Wales). Small

Materials and Methods
38
RNA sequencing reads were demultiplexed using bcl2fastq2 v2.19 allowing for a one
base mismatch. Adaptors were trimmed using cutadapt v1.8.3. Reads ranging from 18-
35 nt were mapped to the mm10 mouse genome using Bowtie v1.1.2 with an 18 base
seed and a maximum of one mismatch. Mapped reads were then annotated using
custom Perl scripts written by Paul Young, to the following databases in order of
priority: miRbase v21, UCSC genomic tRNA, UCSC RepeatMasker, and UCSC RefSeq.
2.4.3.3 Differential expression analyses
Differential expression analyses were performed in Microsoft R Open v3.4.3 using the
edgeR v3.20.7 package. Reads were normalised within their biotype and subjected to
an abundance filter of >100 normalised counts in at least n samples, where n is the size
of the smallest group. p-values were adjusted by the Benjamini-Hochberg correction
algorithm to account for multiple hypothesis testing (154).
2.4.3.4 miRNA target prediction and functional enrichment analysis
miRNAs were searched against two miRNA target prediction databases:
TargetScanMouse v7.1 (156), and microT-CDS v5 (157) with a stringent prediction
threshold of 0.9. High-confidence predicted gene targets (identified by both
algorithms) were submitted to DAVID v6.8 for functional enrichment analysis, using
all miRNAs identified in the sample as a background set.

39
3 SERTOLI CELL EXTRACELLULAR VESICLES AND THE EFFECTS
OF ENVIRONMENTAL STRESS
3.1 Introduction
Sertoli cells are somatic cells that are essential for spermatogenesis. Any disruption to
the Sertoli cell physiology can alter the Sertoli cell barrier, Sertoli cell secretory
products and ultimately germ cells. One of the main functions of Sertoli cells is the
formation of the SCB, which isolates spermatocytes in a tightly-controlled
microenvironment from less-developed germ cells and circulatory molecules.
Bisphenol A is an EDC with a long-documented history of causing reproductive effects
(70,72,74). BPA can interact with Sertoli cell estrogen receptors (79) to alter the
permeability of the SCB, which ultimately results in decreased sperm quality and
quantity and reproductive defects in offspring (80-82).
While BPA induces many transgenerational effects, little is known about the causative
mechanism. BPA is known to affect the mRNA and miRNA expression profile of
Sertoli cells (80), and it is possible that BPA influences the transfer of small RNAs from
Sertoli cells to germ cells, which are able to act as a conduit between the environment
and the next generation. However, this poses another question, how are these small
RNAs transported from somatic cells to germ cells? One possible vector is EVs, lipid-
enclosed vesicles that contain a selected subset of RNA cargo, that can interact with
specific target cells. Exosome-like secretions from Sertoli cells have been observed in
tissue slices of the Chinese soft-shelled turtle (158), however, there are no known

Sertoli cell extracellular vesicles and the effects of environmental stress
40
studies that have isolated Sertoli EVs or characterised their cargo. Currently, the role of
Sertoli EVs in the complex Sertoli cell-germ cell relationship and the inheritance of
environmentally-induced traits remains unknown.
In this chapter I hypothesise that Sertoli cells release extracellular vesicles that interact
with germ cells through their small RNA cargo, and that this cargo (and any
downstream effects of their cargo on germ cells) is susceptible to environmental
stressors. I test this hypothesis in vitro using the TM4 Sertoli cell line, the C18-4
spermatogonial stem cell line (SSC) to model a recipient germ cell, and BPA as an
environmental stressor. My specific aims are to:
1. Characterise mammalian Sertoli EVs produced in vitro in terms of their size,
shape and concentration
2. Identify the small RNA cargo of Sertoli EVs and their producing cells
3. Determine whether purified EVs interact with SSCs
4. Determine the effects of BPA exposure on Sertoli EV production and cargo

Sertoli cell extracellular vesicles and the effects of environmental stress
41
3.2 Results
3.2.1 The isolation and characterisation of Sertoli EVs
3.2.1.1 Sertoli cells release EVs in vitro
First, I asked whether Sertoli cells produce EVs, by culturing TM4 Sertoli cells for three
days in EV-depleted media before filtration and ultracentrifugation of conditioned
culture media using a differential centrifugation-based protocol optimised to reduce
contamination. The resulting pellet was resuspended in PBS, aliquoted and frozen for
downstream validation and analyses. I then sought to image any particles in the
resuspension using transmission electron microscopy. I was able to detect particles of
approximately 120 nm diameter with a cup-shaped morphology (Figure 3.1a). Both the
size and cup-shaped morphology suggest that these particles are exosomes, one of the
two classes of EVs. I also measured the size and concentration of particles using
nanoparticle tracking analysis (NTA), a particle sizing technique that uses light
scattering to track the movement of particles by Brownian motion and then calculates
particle size and concentration based on individual particles’ velocity using the Stokes-
Einstein equation (151). I detected a primary population of ~120 nm EVs, consistent
with the sizing observed by TEM, as well as secondary peak at ~170 nm, likely to be a
subpopulation of larger EVs or artefacts due to aggregation (Figure 3.1b). Using final
cell counts taken when conditioned culture media was collected and the number of
EVs isolated from NTA, I calculated that 160 EVs were released per cell over the three
days of culture.

Sertoli cell extracellular vesicles and the effects of environmental stress
42
Figure 3.1 Physical properties of EVs isolated from Sertoli cells
(a) Transmission electron micrographs of EVs isolated from Sertoli cells. (b) Sertoli EV Size distribution profile of purified Sertoli EVs as determined by Nanoparticle tracking analysis. n=5 preparations, blue shading indicates SEM.
To further confirm the purification of EVs (and not ABs or organelles), I identified the
constituent proteins by protein mass spectrometry. In 10 µg of Sertoli EV protein, I
identified 330 unique proteins after filtering (>=2 significant matching peptides and
>5% protein coverage) (Table 8.1). The majority (63%) of identified proteins have been
reported in either of the two extracellular vesicles cargo databases, ExoCarta or
Vesiclepedia (Figure 3.2a). Identified Sertoli EV proteins included 24 out of the top 25
most commonly reported exosome protein markers reported on ExoCarta (152); the
remaining (1/25) top exosome protein was detected in my sample but did not meet the
5% total protein coverage cut-off. Functional enrichment analysis revealed that the
‘extracellular exosome’ cellular component was the most overrepresented gene
ontology, associated with >80% of identified Sertoli EV proteins (Figure 3.2b).

Sertoli cell extracellular vesicles and the effects of environmental stress
43
Figure 3.2 Proteins identified in Sertoli EVs
(a) Overlap between proteins identified in Sertoli EVs and those curated in existing EV compendia. (b) The 10 most overrepresented ontologies (retrieved from GO_Direct database using DAVID v6.8) by Sertoli EV proteins. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis. CC=cellular component, MF=molecular function.

Sertoli cell extracellular vesicles and the effects of environmental stress
44
Table 3.1 Common exosome proteins identified in Sertoli EVs
ExoCarta ranka
Protein Gene
symbol
Unique matching peptides
% protein covered
1 CD9 antigen CD9 2 8.4 2 Programmed cell death 6-interacting protein (Alix) PDCD6IP 29 38.2 3 Heat shock cognate 71 kDa protein HSPA8 42 64.2 4 Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) GAPDH 12 36.3 5 Actin, cytoplasmic 1 ACTB 34 77.7 6 Annexin A2 ANXA2 12 32.2 7 CD63 antigen CD63 2 8 8 Syntenin-1 SDCBP 19 49.1 9 Alpha-enolase ENO1 13 32.5
10 Heat shock protein HSP 90-alpha HSP90AA1 12 16.3 11 Tumor susceptibility gene 101 protein TSG101 12 16.4 12 Pyruvate kinase PKM PKM 21 40.6 13 L-lactate dehydrogenase A chain LDHA 8 22 14 Elongation factor 1-alpha 1 EEF1A1 23 53.1 15 14-3-3 protein zeta/delta YWHAZ 14 51.9 16 Phosphoglycerate kinase 1 PGK1 9 28.3 17 Elongation factor 2 EEF2 15 21.9 18 Fructose-bisphosphate aldolase A ALDOA 6 18.4 19 Heat shock protein HSP 90-beta HSP90AB1 24 33.6 20 Annexin A5 ANXA5 14 39.8 21 Fatty acid synthaseb FASN 10 4.6 22 14-3-3 protein epsilon YWHAE 6 20 23 Clathrin heavy chain 1 CLTC 14 8.8 24 CD81 antigen CD81 16 58.4 25 Serum albumin ALB 9 9.3
a From http://exocarta.org/exosome_markers_new (152). b Detected below the protein coverage cut-off of 5%.
Taken together, morphology, size, and protein composition confirm that the particles
purified from conditioned Sertoli culture media are EVs, and suggest that they are
exosomes, one of the two classes of EVs. However, since the mode of biogenesis is
unknown, I am unable to rule out the presence a subpopulation of microparticles, and
thus I will refer to these particles as EVs from hereon in.

Sertoli cell extracellular vesicles and the effects of environmental stress
45
3.2.2 Sertoli EV small RNA cargo
3.2.2.1 Sertoli EVs contain selectively packaged small RNA
There are many reports that EVs contain many types of small RNAs, including EVs. I
asked whether Sertoli EVs similarly carry small RNA cargo. Sertoli EVs were collected
as described previously, and total RNA was extracted. Importantly, purified EVs were
pre-treated with RNase to remove any unencapsulated (and therefore unprotected)
RNAs. For comparison, total RNA was also extracted from the parent cells from which
the EVs were derived.
Electropherograms of the RNA revealed that, as expected, Sertoli cell RNA was
predominantly composed of ribosome RNA (18S and 28S rRNA), along with a smaller
population of <200 nt RNAs (Figure 3.3). Sertoli EVs, however, displayed a unique
electropherogram devoid of rRNA and with a much greater relative abundance of
<200 nt small RNAs.

Sertoli cell extracellular vesicles and the effects of environmental stress
46
Figure 3.3 Size distribution of RNAs in Sertoli cells and EVs
Electropherogram traces of RNA isolated from Sertoli cells and EVs. An Agilent RNA Pico chip used to show the 25-4,000 nt range on the left and an Agilent small RNA Pico chip used to show the 4-150 nt range on the right.
To interrogate the small RNA fraction of EVs, I constructed small RNA libraries from
Sertoli EVs and their parent cells and sequenced them in multiplex on the Illumina
platform. Reads were quality trimmed, mapped to the mm10 genome and annotated
against miRNA, tRNA, repeat and genomic UCSC databases. For an initial parsing of
the data, reads were classed into various types of small RNAs and then conflated by
biotype (Figure 3.4a). On average, 4% and 5% of reads were miRNAs in both Sertoli
cell and EV libraries, fewer than expected especially for the cells. tRNA-derived reads
also made up a lower proportion in Sertoli cells (15%) and EVs (4%). Unexpectedly,
reads predominantly mapped to unannotated intergenic and intronic regions.

Sertoli cell extracellular vesicles and the effects of environmental stress
47
Additionally, biotype proportions varied greatly between samples and thus were not
able to distinguish between Sertoli cell and EV samples.
Further examination of library sequence lengths revealed sample-specific read-length
biases (Figure 3.4b) most likely caused by non-uniform size selection during small
RNA library construction. This size bias explains the unexpected distribution of small
RNA reads and meant that the sequenced small RNA libraries were not representative
of the small RNA in EVs and cells. This bias prevented any valid normalisation based
on total mapped reads, and thus prevented comparisons between the biotype
proportions of each sample. However, I am still able to make comparisons between
reads of similar lengths by normalising reads within a biotype (for example to total
miRNA reads, and thus a specific size range) as opposed to the total mapped reads.
Figure 3.4 Sertoli cells and EVs small RNA biotype and length distribution
(a) Mapping regions (annotated against miRbase v21, and UCSC genomic tRNA, UCSC RepeatMasker, and UCSC RefSeq) for small RNA reads from Sertoli cell and EV libraries. (b) Length of small RNA reads in Sertoli cells and EVs. n=3 and 5 for Sertoli cells and EVs respectively.
3.2.2.2 Sertoli cells and EVs have distinct miRNA profiles
As expected, miRNA reads in both Sertoli cells and EVs had a length distribution
primarily between 20-23 nt, typical of this class of small RNA (Figure 3.5). For

Sertoli cell extracellular vesicles and the effects of environmental stress
48
differential abundance analyses, miRNA counts were normalised within the miRNA
biotype.
Figure 3.5 miRNA read length distribution for Sertoli cells and EVs
Length distribution of miRNA reads from Sertoli cell and EV libraries (n=5 for each group). The majority of reads were ~21-23 nt, as expected for miRNAs.
After processing and filtering, 153 mature miRNA species were considered for further
analyses (Table 8.2). Principal component analysis (PCA; Figure 3.6a) and
unsupervised hierarchical clustering (Figure 3.6b) revealed distinct separation between
Sertoli cell and EV miRNA profiles, suggesting that miRNAs within Sertoli EVs
somehow differs from miRNAs in cells from which they are derived.

Sertoli cell extracellular vesicles and the effects of environmental stress
49
Figure 3.6 Sertoli cells and EVs have distinct miRNA profiles
(a) PCA on miRNA counts in Sertoli cells and EVs. (b) Unsupervised hierarchical clustering based on miRNA counts in Sertoli cells and EVs. n=5 for each group.
Typical for cellular miRNA distributions, the top 10 most abundant miRNAs
accounted for 70% of all miRNA reads in EVs (Table 3.2). miR-21a-5p was the most
abundant miRNA in both Sertoli EVs and cells, accounting for 44% and 41% of all
miRNA reads respectively. The top 10 miRNAs in Sertoli EVs had 443 high-confidence
predicted gene targets (predicted by two prediction algorithms), however functional
annotation enrichment analysis performed on these targets revealed no
overrepresentation of any gene ontology or pathway.
Table 3.2 Top 10 most abundant miRNAs in Sertoli EVs
miRNA Sertoli EVs Sertoli Cells
Normalised counta Rank Normalised counta Rank miR-21a-5p 437,872 1 414,867 1
let-7f-5p 46,074 2 39,835 3 miR-143-3p 34,733 3 38,174 5 miR-10a-5p 33,294 4 38,298 4
let-7i-5p 33,007 5 49,962 2 let-7c-5p 25,618 6 37,065 6 let-7g-5p 23,871 7 28,116 7
miR-10b-5p 18,003 8 15,968 10 let-7a-5p 16,715 9 15,309 11
miR-99b-5p 16,478 10 15,106 12 a Averaged for group and normalised per million miRNA reads
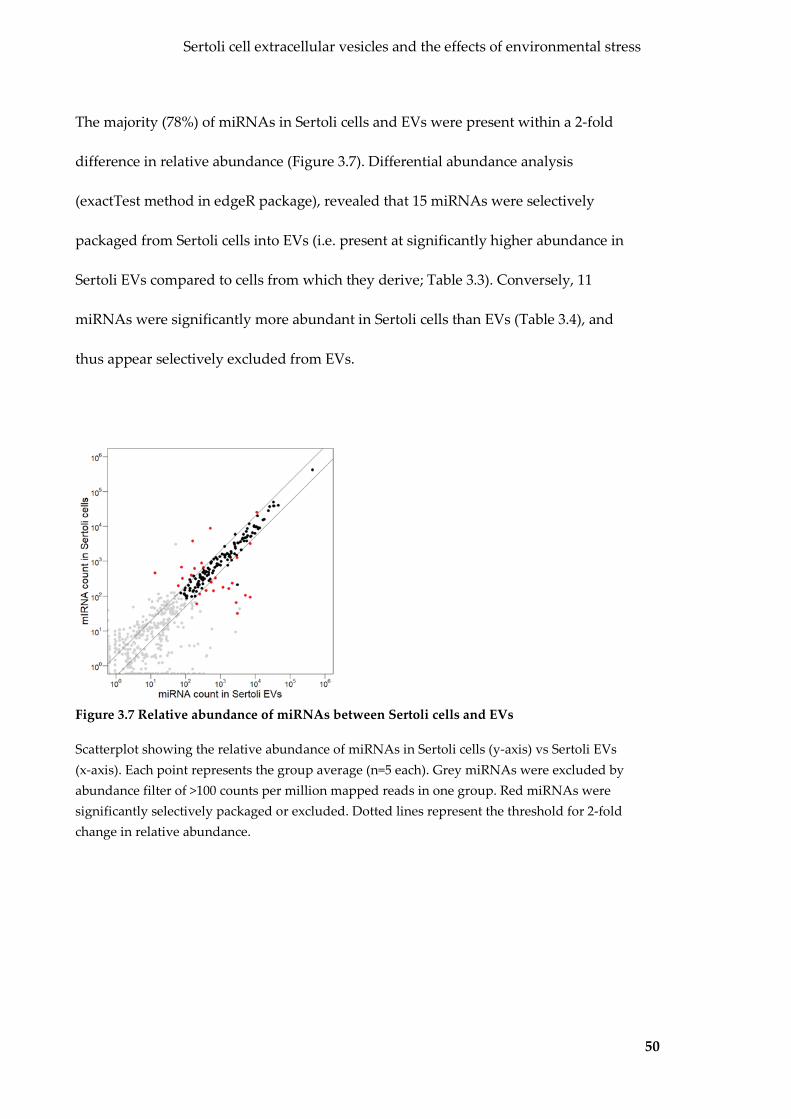
Sertoli cell extracellular vesicles and the effects of environmental stress
50
The majority (78%) of miRNAs in Sertoli cells and EVs were present within a 2-fold
difference in relative abundance (Figure 3.7). Differential abundance analysis
(exactTest method in edgeR package), revealed that 15 miRNAs were selectively
packaged from Sertoli cells into EVs (i.e. present at significantly higher abundance in
Sertoli EVs compared to cells from which they derive; Table 3.3). Conversely, 11
miRNAs were significantly more abundant in Sertoli cells than EVs (Table 3.4), and
thus appear selectively excluded from EVs.
Figure 3.7 Relative abundance of miRNAs between Sertoli cells and EVs
Scatterplot showing the relative abundance of miRNAs in Sertoli cells (y-axis) vs Sertoli EVs (x-axis). Each point represents the group average (n=5 each). Grey miRNAs were excluded by abundance filter of >100 counts per million mapped reads in one group. Red miRNAs were significantly selectively packaged or excluded. Dotted lines represent the threshold for 2-fold change in relative abundance.

Sertoli cell extracellular vesicles and the effects of environmental stress
51
Table 3.3 miRNAs selectively packaged into Sertoli EVs
miRNA Sertoli EV averagea Sertoli cell averagea Fold-enrichment q-value miR-486b-5p 3,074 32 96.12 6.06 x 10-5 miR-101b-3p 7,175 94 76.42 6.83 x 10-5 miR-486a-5p 5,167 106 48.86 1.60 x 10-4 miR-106b-3p 2,823 65 43.11 1.93 x 10-9 miR-106b-5p 1,750 168 10.42 0.04 miR-10a-3p 2,194 240 9.13 0.00 miR-10a-5p 1,178 183 6.44 6.45 x 10-4 miR-10b-5p 630 145 4.34 0.03 miR-122-5p 209 61 3.42 0.00 miR-125a-3p 386 146 2.64 0.00 miR-125a-5p 3,017 1,273 2.37 0.01
miR-125b-1-3p 7,154 3,217 2.22 0.03 miR-125b-5p 252 117 2.15 0.03 miR-126a-3p 715 337 2.12 0.03 miR-128-3p 539 254 2.11 0.03
a Per million miRNA reads
Table 3.4 miRNAs selectively excluded from Sertoli EVs
miRNA Sertoli cell average Sertoli EV average EV fold-enrichment q-value miR-1949 459 13 0.02 1.86 x 10-11 miR-3535 3,821 160 0.04 1.79 x 10-4 miR-5099 8,777 509 0.05 1.36 x 10-6
miR-677-5p 673 76 0.11 1.78 x 10-5 miR-3068-3p 328 81 0.24 0.04 miR-214-5p 618 182 0.29 0.04 miR-145a-5p 877 285 0.32 0.01 miR-130a-3p 203 61 0.30 0.00 miR-16-1-3p 392 146 0.37 3.17 x 10-4
let-7b-5p 25,232 11,007 0.43 0.01 miR-145a-3p 660 324 0.49 0.04 a Per million miRNA reads
Functional annotation enrichment analysis revealed two significantly overrepresented
gene ontologies by the predicted targets of the 15 selective packaged miRNAs: the
‘protein binding’ molecular function and the ‘nucleus’ cellular component (Figure
3.8a). Seven ontologies were overrepresented by the predicted targets of the 11

Sertoli cell extracellular vesicles and the effects of environmental stress
52
miRNAs selectively excluded from EVs, including RNA-binding and processing
related ontologies (Figure 3.8b).
Figure 3.8 Gene ontologies overrepresented by miRNAs differentially expressed between
Sertoli EVs and cells
Overrepresented gene ontologies (retrieved from GO_Direct database using DAVID v6.8) for the predicted gene targets (predicted by both TargetScanMouse v7.1 and microT-CDS v5) of miRNAs selectively packaged into EVs (a) and selectively excluded from EVs (b). q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis.
Taken together, this suggests that Sertoli EV miRNA cargo is not necessarily
representative of their parent cells, with many miRNAs either selectively packaged
into, or selectively excluded from EVs. The predicted targets of these two sets of
miRNAs were found to target different gene ontologies, and thus have contrasting
functions. This lends credence to the idea that Sertoli EVs are intercellular messengers
of specially packaged miRNA cargo.
3.2.2.3 Differential abundance of tRNA-derived reads in Sertoli cells and EVs
miRNAs are not the only small RNA species known to be selectively packaged in EVs
(100,159). Recently, attention has been given to processed tRNA transcripts, as they
appear to be upregulated in response to many stressors, and may regulate gene

Sertoli cell extracellular vesicles and the effects of environmental stress
53
expression by targeting complementary sequences as miRNAs do (35). In Sertoli cells
and EVs, tRNA-derived reads were present as 30-33 nt sequences (Figure 3.9a) that
mapped to the 5’ or 3’ arms of mature tRNAs (Figure 3.9b). The majority of
tRNA-derived reads mapped to the 5’ arm of tRNA molecules (Figure 3.9c), and from
hereon in are referred to as tRNA-fragments (tRFs).
For further analyses, tRFs were conflated based on the anticodon of the gene they
mapped to (for full list see Table 8.3). All anticodons were not evenly represented by
tRFs, with the top six anticodons accounting for 84% and 88% of all tRFs in Sertoli cells
and EVs respectively (Figure 3.10a). This skewed distribution of anticodons does not
mirror their genomic abundance (160), and suggests that these transcripts are not
degradation products, but rather produced by specific processing. Based on the counts
from these top six anticodons, Sertoli cells and EVs could be distinguished by PCA
(Figure 3.10b) and unsupervised hierarchical clustering (Figure 3.10c). In most cases,
tRFs arm preference did not change, however in Sertoli cells tRF5-Val-TAC was most
abundant than tRF3-Val-TAC, while the opposite was observed in Sertoli EVs (Figure
3.9d). This change in arm preference may represent the selective packaging of certain
tRFs.

Sertoli cell extracellular vesicles and the effects of environmental stress
54
Figure 3.9 tRFs in Sertoli cells and EVs
(a) Length distribution of tRFs in Sertoli cells and EVs (b) tRF reads map to the 5’ and 3’ arms of mature tRNA molecules. (c) Wig track of tRF5-Gly-GCC, a representative 5’ tRF. (d) tRF5/3-Val-TAC arm switching between Sertoli cells and EVs. n=5 for each group.
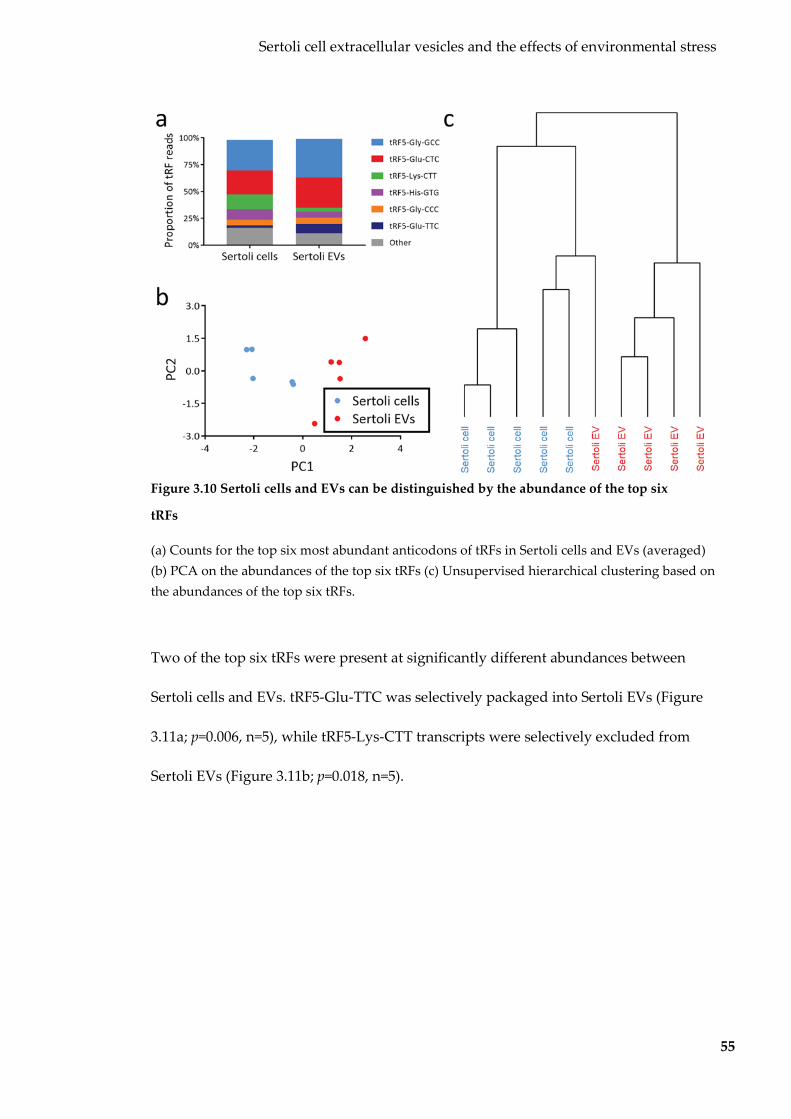
Sertoli cell extracellular vesicles and the effects of environmental stress
55
Figure 3.10 Sertoli cells and EVs can be distinguished by the abundance of the top six
tRFs
(a) Counts for the top six most abundant anticodons of tRFs in Sertoli cells and EVs (averaged) (b) PCA on the abundances of the top six tRFs (c) Unsupervised hierarchical clustering based on the abundances of the top six tRFs.
Two of the top six tRFs were present at significantly different abundances between
Sertoli cells and EVs. tRF5-Glu-TTC was selectively packaged into Sertoli EVs (Figure
3.11a; p=0.006, n=5), while tRF5-Lys-CTT transcripts were selectively excluded from
Sertoli EVs (Figure 3.11b; p=0.018, n=5).

Sertoli cell extracellular vesicles and the effects of environmental stress
56
Figure 3.11 tRFs differentially expressed between Sertoli cells and EVs
(a) tRF5-Glu-TTC is selectively packaged into Sertoli EVs. (b) tRF5-Lys-CTT is selectively excluded from Sertoli EVs. Error bars represent SEM. n=5 for each group.
Taken together, my data suggests that the selective packaging and exclusion of small
RNA cargo between Sertoli cells and EVs is not limited to miRNAs and extends to
tRFs.
3.2.3 Interactions between Sertoli EVs and germ cells
3.2.3.1 Sertoli EVs bind with spermatogonial stem cells
EVs act as intercellular messengers via the transfer of encapsulated small RNA. Before
investigating whether this communication includes environmental signals, I need to
establish that Sertoli EVs can deliver their small RNA cargo to germ cells. First, to
determine if purified Sertoli EVs interact germ cells, I performed co-incubation
experiments with fluorescently-labelled EVs and the C18-4 spermatogonial stem cell
(SSC) line (149). SSCs are representative of germ cells that reside in the basal
compartment of the testes and are in direct contact with Sertoli cells.

Sertoli cell extracellular vesicles and the effects of environmental stress
57
Sertoli EVs were labelled with PKH26 (red membrane dye), washed extensively, added
to a growing culture of SSCs, and then imaged every 20 minutes for 15 hours. A
punctate pattern of fluorescence within cells was observed two hours after the addition
of labelled EVs (Figure 3.12a), perhaps representing hotspots of EV attachment.
Fluorescence intensity increased steadily over the next 13 hours, with fluorescent
hotspots tracking with cells after movement and division. No fluorescence was
observed after the addition of dye-labelled PBS negative controls (Figure 3.12b),
indicating that no detectable dye was transferred by dye micelles.

Sertoli cell extracellular vesicles and the effects of environmental stress
58
Figure 3.12 SSCs co-incubated with labelled Sertoli EVs for 15 hours
Membrane-labelled Sertoli EVs (a) and PBS-labelled negative control (b) were co-incubated with SSCs and imaged for 15 hours.
With the knowledge that EV-binding is detectable after two hours, I sought to
determine the nature of interaction between Sertoli EVs and SSCs. To do this, I
performed a similar co-incubation with PKH67-labelled (green membrane dye) EVs,
however after three hours I washed the cells to remove any unbound EVs before
imaging (Figure 3.13a). After this washing step, a similar punctate fluorescence pattern
indicated that these EVs were not passively bound to SSCs, and were possibly
internalised by SSCs. Dye micelles did not result in any detectable fluorescence (Figure

Sertoli cell extracellular vesicles and the effects of environmental stress
59
3.13b), as previously observed. SSCs labelled directly with the membrane dye exhibited
a distinct uniform, un-speckled fluorescence pattern, suggesting that free dye was not
responsible for the punctate hotspots (Figure 3.13c).
Figure 3.13 SSCs co-incubated with labelled Sertoli EVs for three hours
Membrane-labelled Sertoli EVs (a) and dye-labelled PBS (b) were co-incubated with SSCs for three hours, with unbound EVs washed away prior to imaging. (c) SSCs were directly labelled with membrane dye.

Sertoli cell extracellular vesicles and the effects of environmental stress
60
3.2.3.2 Sertoli EVs transfer small RNA cargo to SSCs
Having established that Sertoli cells produced EVs that contain small RNA cargo and
interacted with SSCs, I next sought to determine whether EV-encapsulated small RNAs
were transferred. To do this I exposed SSCs to Sertoli EVs for three consecutive days.
On day four, I washed the SSCs extensively to remove any unbound EVs, and then
compared the small RNA profiles of SSCs exposed to EVs to those exposed to a PBS
control.
After establishing the miRNA profile of control SSCs (exposed to PBS only), I asked
whether any miRNA present in Sertoli EVs had changed in abundance in the
EV-treated SSCs. Differential expression analysis (exactTest method in edgeR) revealed
that while a group of miRNAs had increased in abundance in SSCs after exposure to
Sertoli EVs, only three reached statistical significance (Figure 3.14). Importantly, no
miRNAs significantly decreased in abundance after exposure to Sertoli EVs.

Sertoli cell extracellular vesicles and the effects of environmental stress
61
Figure 3.14 Changes in SSC miRNA abundance after exposure to Sertoli EVs
Average (n=4) miRNA fold-change (x-axis) for plotted against p-value (y-axis). Red miRNAs are differentially expressed. Dotted line indicates p=0.05.
The miRNAs identified as differentially expressed were not among the most abundant
in SSC control cells. This can be attributed to how the differential expression analysis
was performed. The exact test method used is based on fold-change compared to the
basal abundance of miRNAs in the control SSC sample. This was not ideal, as I sought
miRNAs that had a net-increase in abundance, regardless of their initial abundance in
control SSCs. To incorporate this, I also identified miRNAs that had increased by over
100 normalised (to total mapped reads) reads in SSCs after exposure to Sertoli EVs.
This combined search yielded 21 miRNAs that had increased in SSCs in response to
Sertoli EV exposure (Table 3.5). The majority of these increased miRNAs were either
among the most abundant in Sertoli EVs and/or selectively packaged into EVs by

Sertoli cell extracellular vesicles and the effects of environmental stress
62
Sertoli cells, strongly suggesting that Sertoli EVs were the source of these increased
miRNAs.
Table 3.5 miRNAs increased in SSCs after exposure to Sertoli EVs
miRNA SSC
averagea SSC + EV averagea
Net miRNA increase
Fold-enrichment
EV rank
Sertoli EV vs cell fold-enrichment
miR-122-5p 8 309 300 36.77 32 48.86 miR-486b-5p 27 434 407 15.89 44 96.12 miR-486a-5p 69 1,078 1,010 15.66 24 76.42 miR-21a-5p 29,629 36,331 6,702 1.22 1 1.05 miR-148a-3p 8,482 9,899 1,417 1.16 23 1.43
let-7f-5p 6,639 7,736 1,096 1.16 2 1.15 miR-99b-5p 1,591 2,226 635 1.39 10 1.09
let-7i-5p 4,241 4,856 615 1.14 5 0.66 let-7g-5p 2,339 2,878 539 1.23 7 0.84
miR-10a-5p 3,282 3,679 396 1.12 4 0.86 miR-25-3p 907 1,262 355 1.39 25 2.22
miR-182-5p 1,689 2,018 328 1.19 12 1.43 miR-183-5p 1,450 1,774 324 1.22 27 1.09
let-7c-5p 3,377 3,674 297 1.08 6 0.69 miR-7a-5p 1,296 1,567 270 1.20 121 0.57
miR-30d-5p 1,180 1,451 270 1.22 19 0.89 miR-10b-5p 855 1,064 210 1.24 8 0.89 miR-92a-3p 213 388 175 1.81 17 1.07
let-7b-5p 706 877 171 1.24 15 0.43 miR-151-3p 676 803 127 1.18 52 1.55
miR-148b-3p 364 465 101 1.27 62 1.28 a Normalised per million miRNA reads

63
Five of the top six most abundant tRFs in Sertoli EVs were also increased in SSCs after
Sertoli EV exposure (Figure 3.15), however these increases failed to reach statistical
significance.
Figure 3.15 tRFs in SSCs before and after exposure to Sertoli EVs
Abundances in SSCs before and after exposure to Sertoli EVs shown for top six most abundant tRFs in Sertoli EVs: (a) tRF5-Gly-GCC, (b) tRF5-Gln-CTC, (c) tRF5-Lys-CTT, (d) tRF5-His-GTG, (e) tRF5-Glu-TTC and (f) tRF5-Gly-CCC. Error bars represent SEM. n=4 per group.
To determine whether these transferred small RNAs influence SSC gene expression, I
exposed SSCs to Sertoli EVs, extracted RNA as previously described and run on
Affymetrix microarrays. Surprisingly, out of the 5,506 genes expressed, none were
differentially expressed in response to Sertoli EV exposure (Figure 3.16). This suggests
that while many miRNAs have been transferred to SSCs, they do not have an
immediate, detectable effect on gene expression in the recipient cells. While no changes

Sertoli cell extracellular vesicles and the effects of environmental stress
64
to gene expression were detected in SSCs immediately after EV exposure, this does not
preclude transferred small RNAs from having an effect at a later stage. EVs can be
internalised inside recipient cells via endocytosis for future fusion with the cellular
cytoplasm (114,133,134), thus leaving the EV membrane intact and EV cargo isolated
from cellular gene regulation machinery. Taken together, my data suggests that Sertoli
EVs not only bind and interact with SSCs, but also transfer many of their encapsulated
small RNA cargo, however this does not result in immediate gene expression changes.
Figure 3.16 Gene expression changes in SSCs after exposure to Sertoli EVs
Scatterplot showing the relative expression (average of n=3 per group) of genes in SSCs before (y-axis) and after (x-axis) exposure to Sertoli EVs. Grey genes were expressed below background. Dotted lines represent the threshold for 2-fold change in relative abundance.
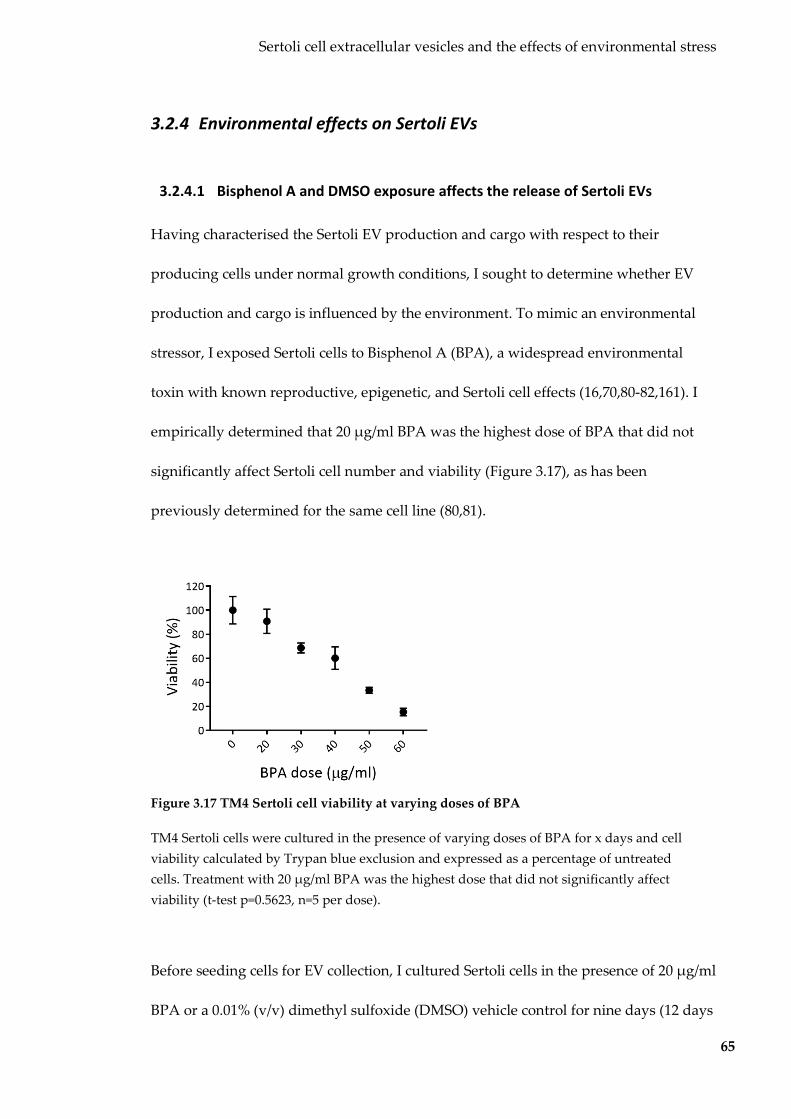
Sertoli cell extracellular vesicles and the effects of environmental stress
65
3.2.4 Environmental effects on Sertoli EVs
3.2.4.1 Bisphenol A and DMSO exposure affects the release of Sertoli EVs
Having characterised the Sertoli EV production and cargo with respect to their
producing cells under normal growth conditions, I sought to determine whether EV
production and cargo is influenced by the environment. To mimic an environmental
stressor, I exposed Sertoli cells to Bisphenol A (BPA), a widespread environmental
toxin with known reproductive, epigenetic, and Sertoli cell effects (16,70,80-82,161). I
empirically determined that 20 µg/ml BPA was the highest dose of BPA that did not
significantly affect Sertoli cell number and viability (Figure 3.17), as has been
previously determined for the same cell line (80,81).
Figure 3.17 TM4 Sertoli cell viability at varying doses of BPA
TM4 Sertoli cells were cultured in the presence of varying doses of BPA for x days and cell viability calculated by Trypan blue exclusion and expressed as a percentage of untreated cells. Treatment with 20 µg/ml BPA was the highest dose that did not significantly affect viability (t-test p=0.5623, n=5 per dose).
Before seeding cells for EV collection, I cultured Sertoli cells in the presence of 20 µg/ml
BPA or a 0.01% (v/v) dimethyl sulfoxide (DMSO) vehicle control for nine days (12 days

Sertoli cell extracellular vesicles and the effects of environmental stress
66
total exposure prior to EV collection), in order to match the length of a cycle of
spermatogenesis in the mouse (162). Then I purified EVs from conditioned culture
media, and determined their concentration, size and cellular yield by NTA as
previously mentioned. To determine the effect of BPA alone on EV production, I
compared EVs produced after treatment with BPA to those produced after treatment
with DMSO, their direct vehicle control. BPA-treated Sertoli cells produced
approximately four times more EVs per cell (Figure 3.18). Surprisingly, treatment with
the DMSO vehicle control alone had an effect in itself, resulting in approximately four-
fold fewer EVs produced per cell when compared to regular media (the direct control
for DMSO treatment). Ultimately, this suggests that treatment with both BPA and
DMSO have opposing effects on EV production, resulting in no apparent change to EV
yield when used together.

Sertoli cell extracellular vesicles and the effects of environmental stress
67
Figure 3.18 Size and yield of EVs produced by Sertoli cells after no, DMSO and BPA
exposure
This graph shows the EV diameter (x-axis) against the EV yield per cell (y-axis). Shading represents SEM. n=6, 4 and 4 for untreated, DMSO and BPA EVs respectively.
3.2.4.2 Growth environment alters miRNA profile in Sertoli EVs but not parent
cells
To determine the effects of treatment with BPA and DMSO on Sertoli EV small RNA
cargo, I extracted RNA from exposed EVs and their producing cells, constructed small
RNA libraries and sequenced them on the Illumina platform. These small libraries
were prepared and sequenced with the earlier untreated batch and were also not size
selected evenly. They were also subject to sample-specific read size bias, as shown by
high inter-group variability in biotype proportions as well as non-uniform read length

Sertoli cell extracellular vesicles and the effects of environmental stress
68
distribution. miRNAs were processed and normalised to per million mapped miRNA
reads as previously described. miRNA reads were subjected to the same abundance
filter, with 204 identified for further analysis (Table 8.2).
For initial analysis, I performed PCA and unsupervised hierarchical clustering on the
filtered miRNA counts. PCA revealed that Sertoli cells and EVs were easily separated
by principal component one (PC1; Figure 3.19a), indicating that sample type accounts
for most of the variation in samples. Both untreated, BPA and DMSO Sertoli cell
samples formed a single cluster, however for EVs, further separation based on growth
environment was achieved by the second principal component. Based on principle
component two (PC2), untreated EVs formed one cluster, while EVs treated with BPA
and DMSO formed a second separate cluster. After unsupervised hierarchical
clustering, a distinct separation was observed between sample types (cells and EVs),
with little separation based on treatment (Figure 3.19b). This suggests that growth
environment affects Sertoli EV miRNA cargo, but not that of their parent cells.
Additionally, it appears that treatment with BPA and DMSO had little effect on Sertoli
EV miRNA profiles when compared to each other, but a distinguishable effect when
compared to untreated EVs, suggesting that DMSO may be the determining factor.
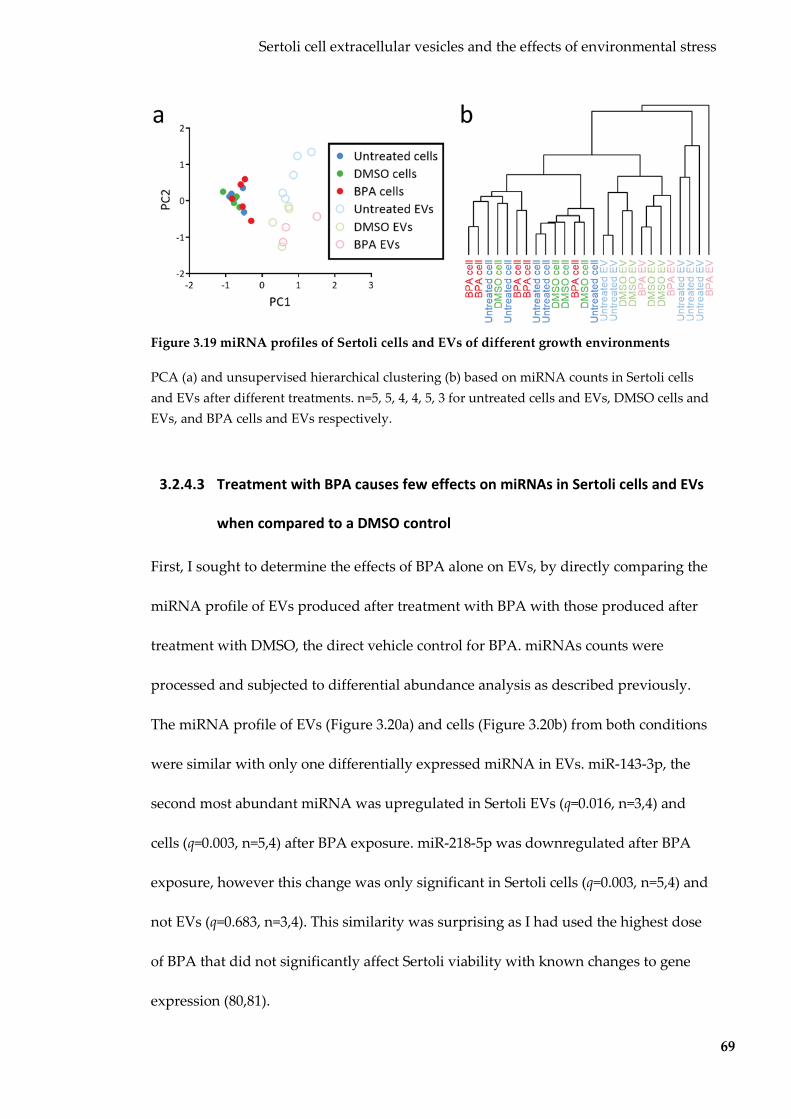
Sertoli cell extracellular vesicles and the effects of environmental stress
69
Figure 3.19 miRNA profiles of Sertoli cells and EVs of different growth environments
PCA (a) and unsupervised hierarchical clustering (b) based on miRNA counts in Sertoli cells and EVs after different treatments. n=5, 5, 4, 4, 5, 3 for untreated cells and EVs, DMSO cells and EVs, and BPA cells and EVs respectively.
3.2.4.3 Treatment with BPA causes few effects on miRNAs in Sertoli cells and EVs
when compared to a DMSO control
First, I sought to determine the effects of BPA alone on EVs, by directly comparing the
miRNA profile of EVs produced after treatment with BPA with those produced after
treatment with DMSO, the direct vehicle control for BPA. miRNAs counts were
processed and subjected to differential abundance analysis as described previously.
The miRNA profile of EVs (Figure 3.20a) and cells (Figure 3.20b) from both conditions
were similar with only one differentially expressed miRNA in EVs. miR-143-3p, the
second most abundant miRNA was upregulated in Sertoli EVs (q=0.016, n=3,4) and
cells (q=0.003, n=5,4) after BPA exposure. miR-218-5p was downregulated after BPA
exposure, however this change was only significant in Sertoli cells (q=0.003, n=5,4) and
not EVs (q=0.683, n=3,4). This similarity was surprising as I had used the highest dose
of BPA that did not significantly affect Sertoli viability with known changes to gene
expression (80,81).

Sertoli cell extracellular vesicles and the effects of environmental stress
70
Figure 3.20 BPA effects on Sertoli EV and cellular miRNA cargo
Scatterplot showing the effect of BPA treatment on Sertoli EV miRNA (a) and Sertoli cell miRNA (b). Grey miRNAs were excluded by abundance filter. Red miRNAs were upregulated and downregulated in response to BPA.
3.2.4.4 Treatment with DMSO alters Sertoli EV miRNA profile but not that of cells
Previous PCA results suggest that treatment with DMSO (compared to untreated
media) has a greater effect on Sertoli EV miRNA cargo than that of treatment with BPA
(compared to the DMSO vehicle control). To further examine this, I compared the
miRNA profiles of Sertoli cells and EVs produced after treatment DMSO to those
produced after no treatment. As suggested in by PCA, miRNA profiles of EVs
produced after treatment with DMSO were dramatically different to miRNA profiles
from untreated EVs (Figure 3.21a). Six miRNAs were significantly upregulated (Table
3.6) while 25 were significantly downregulated after DMSO treatment (a Per million
miRNA reads). EV miRNAs upregulated after treatment with DMSO targeted many
transcription and DNA-binding related gene ontologies (Figure 3.22), while
downregulated miRNAs targeted the ‘nucleus’ cellular component (q-value=2.30 x 10-5,

Sertoli cell extracellular vesicles and the effects of environmental stress
71
45.01% targets). There were no detected changes to DMSO-treated Sertoli cell miRNAs
when compared to untreated cells (Figure 3.21b). This implies that DMSO impacts the
EV cargo sorting process resulting in EVs with different cargo being produced from
virtually indistinguishable sets of cells.
Figure 3.21 Effects of treatment with DMSO on Sertoli EV and cell miRNA cargo
Scatterplot showing the effect of DMSO treatment on Sertoli EV miRNA (a) and Sertoli cell miRNA (b). Grey miRNAs were excluded by abundance filter. Red miRNAs were upregulated and downregulated in response to DMSO.
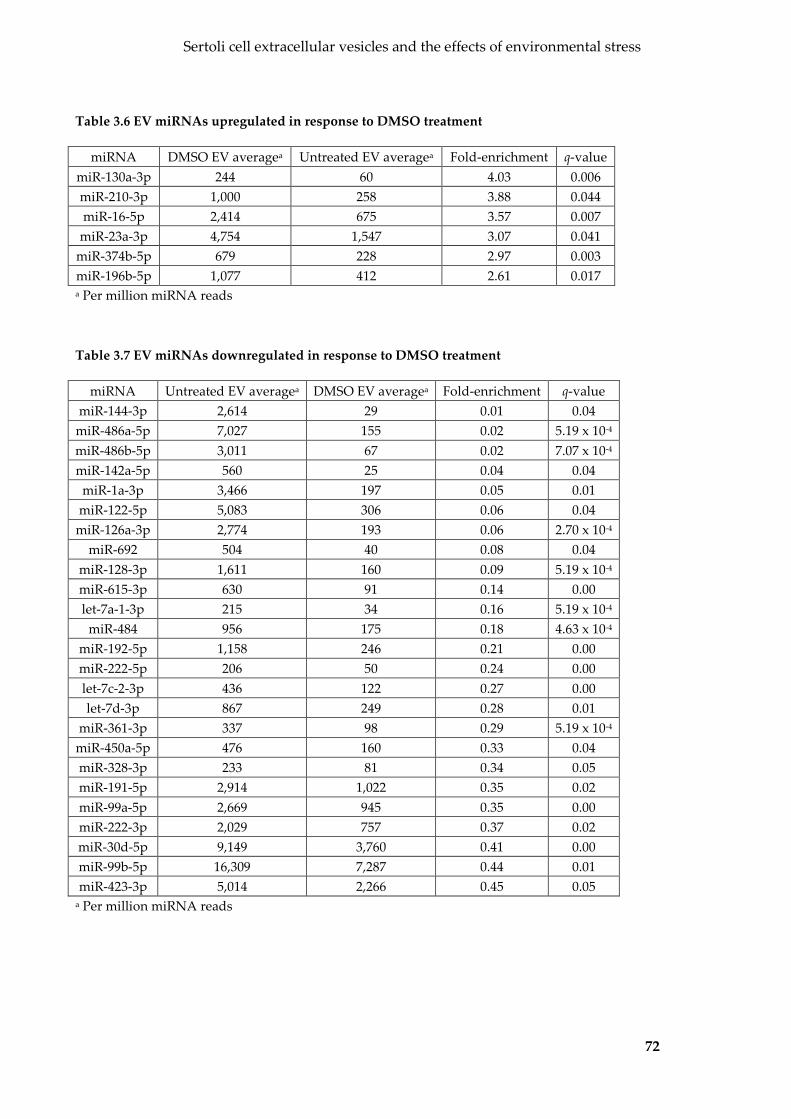
Sertoli cell extracellular vesicles and the effects of environmental stress
72
Table 3.6 EV miRNAs upregulated in response to DMSO treatment
miRNA DMSO EV averagea Untreated EV averagea Fold-enrichment q-value miR-130a-3p 244 60 4.03 0.006 miR-210-3p 1,000 258 3.88 0.044 miR-16-5p 2,414 675 3.57 0.007
miR-23a-3p 4,754 1,547 3.07 0.041 miR-374b-5p 679 228 2.97 0.003 miR-196b-5p 1,077 412 2.61 0.017 a Per million miRNA reads
Table 3.7 EV miRNAs downregulated in response to DMSO treatment
miRNA Untreated EV averagea DMSO EV averagea Fold-enrichment q-value miR-144-3p 2,614 29 0.01 0.04
miR-486a-5p 7,027 155 0.02 5.19 x 10-4 miR-486b-5p 3,011 67 0.02 7.07 x 10-4 miR-142a-5p 560 25 0.04 0.04
miR-1a-3p 3,466 197 0.05 0.01 miR-122-5p 5,083 306 0.06 0.04
miR-126a-3p 2,774 193 0.06 2.70 x 10-4 miR-692 504 40 0.08 0.04
miR-128-3p 1,611 160 0.09 5.19 x 10-4 miR-615-3p 630 91 0.14 0.00 let-7a-1-3p 215 34 0.16 5.19 x 10-4
miR-484 956 175 0.18 4.63 x 10-4 miR-192-5p 1,158 246 0.21 0.00 miR-222-5p 206 50 0.24 0.00 let-7c-2-3p 436 122 0.27 0.00 let-7d-3p 867 249 0.28 0.01
miR-361-3p 337 98 0.29 5.19 x 10-4 miR-450a-5p 476 160 0.33 0.04 miR-328-3p 233 81 0.34 0.05 miR-191-5p 2,914 1,022 0.35 0.02 miR-99a-5p 2,669 945 0.35 0.00 miR-222-3p 2,029 757 0.37 0.02 miR-30d-5p 9,149 3,760 0.41 0.00 miR-99b-5p 16,309 7,287 0.44 0.01 miR-423-3p 5,014 2,266 0.45 0.05
a Per million miRNA reads

Sertoli cell extracellular vesicles and the effects of environmental stress
73
Figure 3.22 Gene ontologies overrepresented by the predicted targets of miRNAs
upregulated in Sertoli EVs after DMSO treatment
Overrepresented gene ontologies (retrieved from GO_Direct database using DAVID v6.8) for the predicted gene targets (predicted by both TargetScanMouse v7.1 and microT-CDS v5) or miRNAs upregulated in Sertoli EVs after DMSO treatment. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis.
3.2.4.5 Effects of treatment with BPA on Sertoli cell and EV tRF cargo
Sertoli cell and EV miRNA data suggests that DMSO itself had a profound effect on
Sertoli EVs (vs untreated), which potentially masks the effect of BPA alone (vs the
DMSO control). To determine if these observations also applied to tRNA-derived
reads, I compared tRNA-derived reads from untreated, BPA treatment and DMSO
treated Sertoli cells and EVs (for full list see Table 8.3). tRNA-derived reads in Sertoli
cells and EVs treated with BPA and DMSO were of a wider size range of 29-33 nts also
mapped to the 5’ and 3’ ends of mature tRNAs. These transcripts were likely to be tRFs
and are referred to as such. Sertoli EVs produced after DMSO treatment (Figure 3.23a)
and BPA treatment (Figure 3.23b) contained tRFs predominantly of 31 nt in length,
while tRFs in parent cells were 32-33 nt long. As seen with untreated EVs and cells,
anticodons were not represented evenly, with the top six anticodons accounting for the
majority of all tRFs in DMSO and BPA treated Sertoli cells and EVs (Figure 3.23c).

Sertoli cell extracellular vesicles and the effects of environmental stress
74
Figure 3.23 tRFs in Sertoli cells and EVs from different growth environments
tRF read length distribution in Sertoli cells and EVs after (a) treatment with DMSO and (b) treatment with BPA. (c) Anticodon representation by tRF reads in Sertoli cells and EVs treated with DMSO and BPA. n= 4, 4, 5 and 3 for DMSO cell, DMSO EV, BPA cell and BPA EV respectively.
To determine if treatment with DMSO or BPA influences Sertoli EV and cell tRF
profiles, I performed PCA and unsupervised hierarchical clustering on the counts of
these top six tRFs. PCA showed separation by sample type (cell or EV) by PC1,
however unlike with miRNAs, PC2 did not show any clustering by growth
environment (Figure 3.24a). Unsupervised hierarchical clustering did not separate cells
from EVs as clearly and showed no separation between growth environments (Figure
3.24b).

Sertoli cell extracellular vesicles and the effects of environmental stress
75
Figure 3.24 tRFs in Sertoli cells and EVs produced under different growth environments
(a) PCA and (b) unsupervised hierarchical clustering performed on the top six most abundant tRFs in untreated, DMSO and BPA treated Sertoli cells and EVs. n=5, 5, 4, 4, 5, 3 for untreated cells and EVs, DMSO cells and EVs, and BPA cells and EVs respectively.
To determine if BPA alone had any effect on these abundant tRFs in EVs and cells, I
compared tRF counts for BPA and DMSO treated Sertoli EVs, and then cells (Figure
3.25). As suggested by PCA, none of these anticodons were differentially expressed
between BPA and DMSO EVs nor cells. To determine if any DMSO-specific effects
were masking those of BPA, I compared tRF counts in DMSO-exposed EVs and cells
with untreated EVs and cells respectively. DMSO exposure did not significantly alter
any of the top six most abundant anticodons of tRFs (Figure 3.25). As expected by the
lack of changes, tRF5-Glu-TTC (Figure 3.25e), and TRF5-Lys-CTT (Figure 3.25c)
remained selectively packaged and excluded from Sertoli EVs under all three growth
conditions.
Taken together, this suggests that while Sertoli EV miRNA cargo was affected by
DMSO exposure (BPA alone to a lesser extent), no consistent changes were observed in
tRF cargo.

Sertoli cell extracellular vesicles and the effects of environmental stress
76
Figure 3.25 Abundance of top six most abundant tRFs in Sertoli cells and EVs of different
growth environments
(a) tRF5-Gly-GCC, (b) tRF5-Glu-CTC, (c) tRF5-Lys-CTT, (d) tRF5-His-GTG, (e) tRF5-Glu-TTC, (f) tRF5-Gly-CCC. Error bars represent SEM. n=5, 5, 4, 4, 5, 3 for untreated cells and EVs, DMSO cells and EVs and BPA cells and EVs respectively.

Sertoli cell extracellular vesicles and the effects of environmental stress
77
3.3 Discussion
In this chapter, I investigated whether TM4 Sertoli EVs carry environmentally-
influenced small RNA cargo, and whether this cargo is transferred to SSCs. To do this,
I first isolated Sertoli EVs from conditioned culture media by filtration and differential
centrifugation, and then confirmed their EV identity by examining size, morphology
and protein cargo. Next, I characterised Sertoli EV small RNA cargo with respect to
their parent cells and identified selectively packaged and excluded miRNAs and tRFs.
Then, I established that purified Sertoli EVs bind and interact with spermatogonial
stem cells in vitro, resulting in the transfer of Sertoli EV small RNAs to SSCs. Finally, I
investigated how a change in environment (exposure to BPA and DMSO) alters the
rate at which Sertoli EVs are produced, and also their encapsulated miRNA cargo
uniquely when compared to miRNA in their parent cells.
3.3.1 Isolation of Sertoli EVs
While EVs have been detected in a plethora of biological fluids and cell lines, this is the
first study to purify and characterise Sertoli EVs. Sertoli exosome-like vesicles of
150-200 nm in diameter were found in Pelodiscus sinensis, the Chinese soft-shelled
turtle, however identification was based solely on electron micrographs without any
molecular confirmation (158).
I isolated EVs from Sertoli cell conditioned media using an optimised protocol based
on differential centrifugation. Despite being one of the first techniques used to isolate
EVs, differential centrifugation remains the gold-standard protocol, especially for

Sertoli cell extracellular vesicles and the effects of environmental stress
78
purification from cell culture media (163). To prevent the purification of cells, cellular
fragments and ABs, I ensured that cells were in the exponential growth phase with
>95% viability at the time of harvest. Additionally, I employed a filtration step to
remove any contaminants carried through differential centrifugation. These steps were
evidently successfully, as shown by the lack of organelle proteins and intact rRNA, a
marker of cellular and AB contamination (164,165), in my purified sample. To improve
yield, I opted not to wash my EV pellet after ultracentrifugation as is commonly
performed to improve purity and decrease co-precipitating protein. Despite leaving
out this step, my isolated EV resuspensions did not contain significant protein
contamination or compromised EV purity as shown by protein mass spectrometry.
Sertoli EV size as determined by TEM and NTA was consistent with the 150-200 nm
exosome-like secretions observed in the chinese soft-shelled turtle (158). Sertoli EV size
was within the range expected for exosomes, one of two classes of EVs. While the
distinct cup-shaped morphology observed after TEM is now known to be an artefact
introduced by drying and fixation techniques, it remains a distinguishing feature that
is characteristic of exosomes (166). Proteomic analysis also hinted towards an exosomal
identity, with the ‘extracellular exosome’ ontology being the most overrepresented and
commonly associated for Sertoli EV proteins as well as a significant overlap between
identified Sertoli EV proteins and those curated in ExoCarta (152) and Vesiclepedia
(153), two manually curated EV compendia. Furthermore, the lack of intact rRNA and
enrichment of <200 nt small RNA is characteristic of exosomes isolated from many
other cell types (142,167-169). Based on this data, I am confident that I have purified
Sertoli EVs from Sertoli cell conditioned culture media.

Sertoli cell extracellular vesicles and the effects of environmental stress
79
3.3.2 Sertoli EVs contain selectively packaged small RNA cargo
Initially, I observed that Sertoli EVs and cells had similar small RNA profiles with
surprisingly few miRNA reads. The low proportions of miRNAs in cellular libraries
were unexpected as miRNAs are known to be the most abundant small RNA in the 18-
35 nt size range in Sertoli cells (170). Further examination of the read size distribution
of Sertoli cell and EV libraries (including those exposed to BPA and DMSO) revealed
that the libraries were not size-selected evenly, and thus individual libraries were
variably skewed towards shorter reads. The sample specific bias of sequence length
meant that counts could not be normalised to the total number of mapped reads, and
therefore comparison between small RNA biotypes could not be performed.
Comparisons could still be made between reads of the same biotype by normalising
within the biotype, for example comparing miRNA counts between groups after
normalisation to total miRNA reads, and biotype normalisation was performed
accordingly.
I observed that Sertoli EV miRNA profile was not entirely representative of that of
Sertoli cells, with many miRNAs selectively packaged or excluded. This is consistent
with observation in EVs from other cell types (98,100,114,124). The most abundant
miRNA in Sertoli EVs was miR-21a-5p, accounting for over 40% of all miRNAs in
Sertoli cells and EVs. This miRNA is among the most abundant in EVs released from
glioblastomas (171) and human liver cancer cell lines (172). The human homolog,
miR-21, is well-known for its anti-apoptotic role in cancer (173), and a similar function
has been suggested in germ cells. miR-21a-5p is one of the most abundant miRNAs in

Sertoli cell extracellular vesicles and the effects of environmental stress
80
germ cell cultures enriched with SSCs, and facilitates the maintenance and renewal of
SSCs (174). Also among the most abundant miRNAs are those of the let-7 family,
which also function in SSC maintenance and self-renewal (175). The presence of these
miRNAs in Sertoli EVs imply that they may be transferred to SSCs to support their
growth.
Many 30-32 nt tRNA-derived transcripts that mapped to the 5’ and 3’ arms of mature
tRNAs were identified in both Sertoli cells and EVs. These sequences are likely to be
tRFs, <32 nt small RNAs produced from mature tRNAs by the ribonucleases (35). tRFs
are known components of EVs from a wide variety of sources (176,177) including
semen (159). tRFs have also been identified in increasing concentrations in germline-
associated somatic cells and their EVs, and sperm from the epididymis and beyond,
but not in germ cells of the testes and whole testes tissue (24,33). My data is the first
that identifies tRFs in Sertoli cells and EVs, at the beginning of the male reproductive
system. While unlikely, it is also possible that these tRNA-derived reads may also be
tRNA-halves. Like tRFs, tRNA-halves also map to the 5’ and 3’ ends of mature tRNAs,
however they are usually only produced in response to stress (178). I identified similar
tRFs in the untreated, BPA and DMSO (stressed) EV and cell samples, suggesting that
they are not tRNA-halves.
I observed an uneven representation of anticodons, with the top six tRF anticodons,
accounting for the majority of tRF reads. The overrepresentation of these tRFs did not
reflect their genomic abundance, suggesting that they are not degradation products
and are specifically processed from certain tRNA molecules. Similar

Sertoli cell extracellular vesicles and the effects of environmental stress
81
overrepresentations of these same anticodons have been observed in cells and EVs
(24,34,176), however why and how these anticodons were overrepresented has yet to
be elucidated. The selective packaging of tRF5-Glu-TTC and the exclusion of tRF5-Lys-
CTT from EVs, as well as differential preference of arms for tRF5/3-Val-TAC is
additional evidence suggesting that these tRFs were specifically processed into EVs
and not degradation products of mature tRNAs.
Little is known about the function of tRFs in EVs. tRFs have been linked to the
regulation of translation (36,37), RNA stability (38,39), and cell proliferation and
viability (40,179). tRFs have been proposed as gene expression regulators through a
miRNA-like Argonaute-dependent mechanism (40,180,181), or novel mRNA targeting
pathways (182,183). Several studies report the effect of environment on tRFs in
epididymal EV and sperm small RNA, which result in the regulation of gene targets
after fertilisation (24,33,34). Taken together, this suggests that tRFs in EVs may have a
wide variety of regulatory roles for maturing sperm under normal conditions.
3.3.3 Sertoli EVs bind to and transfer small RNAs to spermatogonial
stem cells
Through labelling and co-incubation experiments, I show that Sertoli EVs bind and
attach to SSCs. EV binding was also observed after the washing of any unbound EVs
from cells, suggesting that EVs have not passively settled or clumped on the cellular
surface and were bound via some specific interaction. EVs bound in a punctate,
spotted pattern, rather than an even pattern as observed with direct cellular staining

Sertoli cell extracellular vesicles and the effects of environmental stress
82
with a dye. This pattern has been observed in other labelled-EV uptake studies
(141,184-186) and may represent how EV attachment and binding may occur in
hotspots at specific locations on the cellular membrane, perhaps where complementary
surface receptors are located. The internalisation of EVs at specific locations on the
cellular surface would also result in the punctate staining pattern I observed.
Additionally, this pattern may also represent the final destination of EV lipids after
fusion with the cell (187), as opposed to any encapsulated cargo which may travel
further from the original site to other regions of the cell.
While lipophilic dyes have been used extensively to determine EV-cell interactions, its
use is not without caveats. Lipophilic dyes can aggregate in aqueous solutions or form
dye micelles in the presence of salts (188), and these aggregates and micelles may be
purified during ultracentrifugation (189). Through the labelling of a PBS control, I
showed that fluorescence due to free-dye aggregates or micelles was undetectable and
thus did not contribute to the fluorescence observed in my EV sample. Recent evidence
emerged indicating that soluble proteins may also account for significant dye transfer
in the absence of EVs (189). While initial analysis suggests that my purified Sertoli EVs
contain little protein contamination, I did not use EV pellets from unconditioned
culture media (not exposed to Sertoli cells) as my negative control and therefore cannot
determine whether residual protein from serum, a constituent of media, contributed to
the transfer of dye.
After exposure to Sertoli EVs, SSCs contained more miRNAs than their unexposed
counterparts. Similar trends were observed in tRFs of Sertoli EV-exposed SSCs,

Sertoli cell extracellular vesicles and the effects of environmental stress
83
however they were not as apparent. This is likely due to the lower abundance of tRFs
in Sertoli EVs compared to miRNAs. Increased individual miRNAs were among the
most abundant in Sertoli EVs and/or selectively packaged from Sertoli cells to EVs,
strongly suggesting that they were of Sertoli EV origin. Taken together, these results
confirm the labelled EV co-incubation observations that Sertoli EVs bind and fuse with
SSCs, show that these interactions result in the transfer of Sertoli EV small RNA cargo.
Many of the miRNAs transferred from Sertoli EVs to SSCs have regulatory functions,
such as miR-21a-5p and those of the let-7 family (174,175). These coincide with how
Sertoli cells and their secretory products aid with the regulation of SSCs (55,56). My
data suggests that this may be mediated in part, by small RNAs transferred via EVs.
Alterations to the abundance of these small RNAs may disrupt this role and thus affect
germ cells and the next generation, however this requires further experimentation
before it can be confirmed.
Despite the transfer of Sertoli EV miRNAs to SSCs, gene expression microarray data
showed no changes to SSC gene expression after exposure to Sertoli EVs. This could be
due to a number of reasons. Due to time and experimental constraints, the gene
expression experiment was performed using a different batch of SSCs and Sertoli EVs
to that of the small RNA sequencing co-incubation experiment. While attempts were
made to add the same amount of EVs, batch effects may account for the lack of
detectable changes in gene expression. Another potential explanation could be that
Sertoli EVs were endocytosed by SSCs. This would mean that the while Sertoli EV
small RNAs were able to be extracted after lysis (as part of the RNA extraction

Sertoli cell extracellular vesicles and the effects of environmental stress
84
procedure), the EV membrane remains intact and EV cargo are not exposed to cellular
gene expression machinery. If this was the case, then these EVs may be stored for
future fusion with the cellular cytoplasm in differentiated germ cells, or potentially
after fertilisation in the zygote. To further confirm the functional nature of transferred
miRNAs, the same set of RNA should be used for both small RNA sequencing and
gene expression microarray experiments.
3.3.4 Environmental effects on Sertoli EV production and cargo
When exposing Sertoli cells to BPA, I chose to use DMSO as a solvent. Surprisingly, the
DMSO vehicle itself had profound effects on Sertoli EV production and miRNA cargo,
while BPA had unexpectedly few. Therefore, instead of considering my samples as
BPA treated and their DMSO vehicle control, I regarded them as BPA and DMSO
treated, and DMSO treated, with untreated EVs as my baseline.
The reduction of EV production after DMSO was unexpected at the low 0.01% (v/v)
concentration of DMSO used. DMSO is commonly used as a polar aprotic solvent, and
it is generally believed (though not scientifically proven for the TM4 Sertoli cell line) to
have few effects at concentrations below 0.1% (v/v). A 20% decrease in in vitro murine
melanoma EV production has previously been reported, however this occurred after
exposure to 5% DMSO, an extremely high concentration of DMSO proven to have
cytotoxic effects (190). A potential mechanism for DMSOs effect on EV production may
be due to alterations to internal membranes. At higher concentrations DMSO increases
the permeability of the cellular membrane (191). Internal plasma membranes such as

Sertoli cell extracellular vesicles and the effects of environmental stress
85
that of the endosome (the site of exosome formation) may be more sensitive to DMSO’s
effects, resulting in the disruption of EV production at low doses.
After exposure to 0.01% DMSO, I identified many dysregulated miRNAs in Sertoli EVs
that were predicted to target many gene ontologies related to DNA-binding and
transcriptional regulation in recipient cells. The effects of DMSO were specific to EVs,
with Sertoli cells themselves being virtually indistinguishable by their small RNA
profile. EV-specific effects were due to DMSO disrupting the EV-cargo sorting process
causing fewer miRNAs to be selective packaged from Sertoli cells to EVs. These types
of effects have never been reported for such a low concentration of DMSO, despite
DMSO being used as a vehicle in many EV publications (128,190,192,193). This is due
in part to how the effects of drugs are directly compared to their solvent. My findings
have many implications for the use of DMSO in EV studies. While it is readily accepted
that a vehicle control should be used when determining the effects of a dissolved drug,
my results suggest that vehicle effects may be extensive and mask changes for the
chemical of interest. I suggest that any effects of drugs be evaluated with respect to an
untreated control as I have performed here, or that other solvents such as ethanol be
used instead when applicable.
Exposure to BPA (compared with the DMSO solvent control) also altered Sertoli EV
production, however in the opposite direction. While DMSO-exposure reduced the
number of EVs released per cell by four-fold compared to untreated conditions, BPA
(in a DMSO solvent) increased EV production by four-fold compared to DMSO alone,
effectively reverting EV/cell yields back to that observed under untreated conditions.

Sertoli cell extracellular vesicles and the effects of environmental stress
86
The increase in EV production caused by BPA alone can be attributed to a BPA-
induced stress response, as EV production is usually upregulated in many stressed and
pathological states (194-196).
By comparing Sertoli EVs exposed to BPA with those exposed to DMSO, I was able to
identify one miRNA upregulated by BPA exposure, miR-143-3p. miR-143-3p is
commonly among the most abundant miRNAs in EVs released by cancer cells and
functions in a protective tumour-suppressive role (197-199). The presence of miR-143-
3p in Sertoli EVs may be part of a similar regulatory role for SSCs, as miR-143-3p was
determined to be transferred from Sertoli EVs to SSCs. While I only identified miRNAs
transferred to SSCs by untreated EVs, these results imply that environmental
perturbations may have effects on SSCs. While not experimentally determined, it can
be assumed that the BPA-induced increase of miR-143-3p in Sertoli EVs results greater
transfer of miR-143-3p to SSCs, thus potentially acting as a link between the
environment and the next generation. miR-143-3p was found to be the most
upregulated miRNA in the sperm of mice fed a high-fat diet (200). My results suggest
that Sertoli EVs may be a potential source for this increase.
More changes were expected, especially considering the dose of BPA I used. I had
chosen a 20µg/ml dose of BPA as I had empirically determined to the be the highest
dose that did not affect Sertoli cell number and viability by >10%. This same dose has
caused alterations to TM4 Sertoli cell miRNAs and proteins, albeit after undisclosed
concentrations of DMSO vehicle and different exposure times (80,81). The lack of
changes may be due to the profound effects of DMSO masking any changes of BPA

Sertoli cell extracellular vesicles and the effects of environmental stress
87
alone. In future, by substituting DMSO with ethanol, an alternate solvent, one would
be able to better discern the effects of BPA alone on Sertoli EV production and small
RNA cargo.
Unlike miRNAs, few changes to Sertoli EV tRFs were observed after both BPA and
DMSO exposure. This was surprising as tRFs have been found to be more susceptible
to environmental changes, albeit in an in vivo setting (33). In mice, dietary protein
restriction resulted in increased levels of tRF5-Gly-GCC in epididymal EVs being
transferred to sperm (24). This same tRF was the most abundant in Sertoli EVs and
appeared to increase in average abundance in Sertoli EVs after BPA and DMSO
exposure (although this did not reach statistical significance). These may be due to tRFs
comprising a smaller proportion of all small RNA reads in Sertoli EVs and cells leading
to changes that are more susceptible to stochastic variation and variability in small
RNA library preparation.
3.3.5 Conclusion
In this chapter, I demonstrated that EVs can be reproducibly isolated from TM4 Sertoli
cell conditioned media. Sertoli EVs contain selectively packaged small RNA cargo,
when compared to the producing cells. Purified Sertoli EVs bound and fused with
spermatogonial stem cells, and delivered their small RNA cargo, thus acting as
intercellular messengers between somatic and germ cells. The presence of BPA and
DMSO in the cellular environment altered the production of Sertoli EVs. DMSO, and to
a lesser extent BPA disrupted the sorting of miRNA cargo from Sertoli cells into EVs,

Sertoli cell extracellular vesicles and the effects of environmental stress
88
resulting in EVs with altered cargo. Taken together, my results indicate that Sertoli EVs
transfer small RNAs to germ cells, and that these transferred small RNAs may be
affected by the environment. My findings suggest that Sertoli EVs may act as a
potential mechanism for the vertical transmission of environmentally-induced traits.

89
4 THE EFFECT OF FOLATE AVAILABILITY ON EPIDIDYMAL
EXTRACELLULAR VESICLES
4.1 Introduction
Despite being transcriptionally inert, sperm contain many types of RNAs (44,45). These
include small RNAs such as miRNAs and tRFs; in fact, sperm are enriched for these
small RNA species and depleted in the normally dominant rRNAs (44,45). The small
RNAs in sperm were initially thought to be remnants from spermatogenesis, however
we now know that the small RNA composition of sperm changes during its transit
through the epididymis (46).
Like most if not all organs, the epididymis produces EVs that can interact with
recipient cells. Epididymal EVs are produced by apocrine secretions from epididymal
epithelial cells into the epididymal lumen (88,201). They have been isolated from the
epididymis of several animals (49,92,202-204), and a cell line called DC2 (205).
Epididymal EVs provide one means by which proteins are transferred to sperm during
maturation (reviewed in (206)); the only other known possibility is lipid carriers (eg
clusterin and apoliporoteins). More recently, epididymal EVs have also been found to
transport small RNAs to sperm (49).
Small non-coding RNAs have been repeatedly implicated in the vertical transmission
of environmentally-induced traits in invertebrates (9,10,43,207,208). There is also some
evidence that suggests a similar role for sperm small RNAs in mammals. For example,
effects on offspring phenotype induced by paternal factors (including psychological

The effect of folate availability on epididymal extracellular vesicles
90
stress (209) and diet (24,33) are known to alter the small RNA composition of sperm.
Furthermore, injection of these altered small RNAs into fertilised oocytes was able to
reproduce the transgenerational phenotypes induced by the stressor (209,210),
implying that small RNAs are somehow responsible. The interaction of sperm with
epididymal components such as epididymal EVs is believed to play a significant part
of sperm ncRNA acquisition (24,49). Thus, if EV ncRNA cargo is affected by the
environment, such interactions may underpin the inheritance of environmentally-
acquired traits.
Extreme dietary concentrations of folate and other methyl donors represent an
environmental stressor with well-studied intergenerational epigenetic effects. Both an
excess, and deficiency, of dietary methyl donors can result in aberrant DNA
methylation in somatic tissues (211-214). These effects are likely caused by both direct
(imbalance in levels of methyl precursors) and indirect means. Effects of folate
concentrations have also been observed in the germline. Low concentrations of folate in
human seminal plasma has been linked to increased sperm DNA fragmentation (215),
and interestingly, folinic acid (a folate derivative) treatment improves many seminal
parameters in infertile men (216). Changes induced by methyl donor availability have
been observed in the maternal and paternal germline and extend to the next generation
(19,214,217). Of note is a finding by my laboratory, which showed that excess dietary
methyl donors during mid-gestation (after the period of epigenetic resetting following
fertilisation) altered DNA methylation at a specific ‘epigenetic reporter’ locus (19). The
altered epigenetic state was accompanied by associated phenotypic changes in both
offspring and grand-offspring.

The effect of folate availability on epididymal extracellular vesicles
91
While there is little available knowledge on the effects of folate concentration on
epididymal EVs or the epididymis itself, there is reason to believe the epididymis is
sensitive to folate. The epididymal lumen exhibits one of the strongest
immunohistochemical signals for folate receptor-α in the human body (218).
Folate-binding protein (FBP) is associated with mature sperm (not testicular sperm but
those that have passed through the epididymis) and their surrounding EVs (91). These
EVs were initially deemed to be prostasomes, however no FBP was detected in the
prostate (91), and thus these EVs must have originated from a different source, most
likely the epididymis.
In this chapter I investigate the hypothesis that extreme folate concentrations affect
epididymal EV production and small RNA cargo. I test this hypothesis by investigating
the production and cargo of epididymal EVs in vitro from DC2 epididymal cells in the
presence of physiological and supraphysiological concentrations of folate. My specific
aims are to:
1. Characterise mammalian epididymal EVs produced in vitro in terms of their
physical properties and concentration
2. Identify the small RNA cargo of epididymal EVs and their producing cells
3. Determine the effects of changes in folate availability on epididymal EV
production and small RNA cargo

The effect of folate availability on epididymal extracellular vesicles
92
4.2 Results
4.2.1 Isolation and characterisation of DC2 epididymal EVs
4.2.1.1 The isolation of EVs from distal caput epididymal cells cultured in vitro
An in vitro system was chosen as the most tractable model system to directly test the
effects of folate availability. The DC2 cell line was established in from epididymal
epithelial cells of the caput epididymis (148). I purified EVs from DC2 epididymal cells
by culturing them for five days to 80% confluence before pelleting EVs from
conditioned culture media using the same previously optimised protocol of filtration
and differential centrifugation used in Chapter 3. Transmission electron microscopy
showed that epididymal EVs were <200 nm in diameter, however clumping and
over-staining prevented any further view of internal morphology (Figure 4.1a). NTA
showed that purified epididymal EVs were a heterogenous population of three sizes:
90 nm, 140 nm and 190 nm (Figure 4.1b). By using the number of producing cells at the
time of media collection, I was able to determine that on average 101 EVs were released
per cell over the five days of culture.

The effect of folate availability on epididymal extracellular vesicles
93
Figure 4.1 Physical properties of EVs isolated from DC2 epididymal cells
(a) Transmission electron micrographs of EVs isolated from epididymal cells. (b) Epididymal EV size distribution profile as determined by NTA. Shading represents SEM. n=2 preparations.
To further confirm the isolation of epididymal EVs, I examined their protein
composition using protein mass spectrometry. In 10 µg of epididymal EV protein, I
identified 162 unique proteins after filtering (>=2 significant matching peptides and
>5% protein coverage) (Table 8.4). The majority (68%) of these proteins have been
identified in other types of EVs and reported in EV cargo databases (Figure 4.2a).
Additionally, identified proteins include 20 out of the top 25 most commonly reported
exosome proteins in ExoCarta (152). Functional enrichment analysis revealed the
‘extracellular exosome’ cellular component was the most overrepresented ontology
and was associated with 80% of identified epididymal EV proteins (Figure 4.2b).

The effect of folate availability on epididymal extracellular vesicles
94
Figure 4.2 Proteins identified in epididymal EVs
(a) Overlap between proteins identified in epididymal EVs and those curated in existing EV compendia. (b) The top 10 most overrepresented ontologies (retrieved from GO_Direct database using DAVID v6.8) by epididymal EV proteins. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis. CC=cellular component, MF=molecular function.
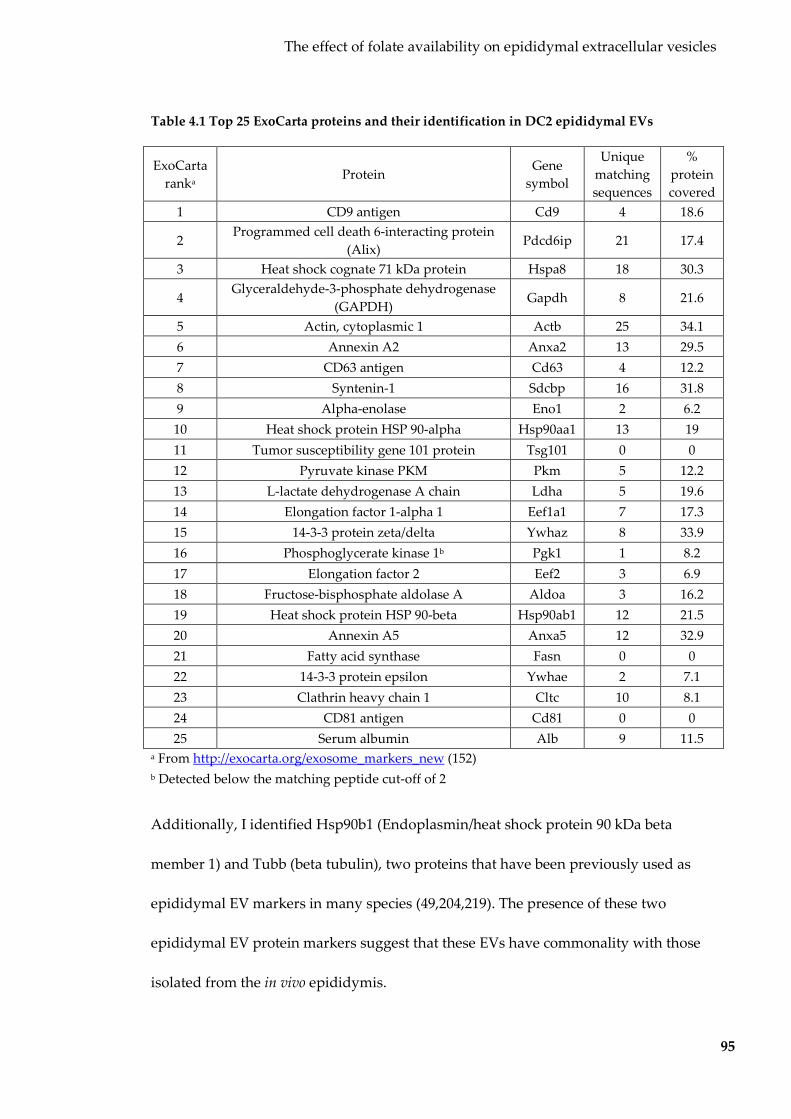
The effect of folate availability on epididymal extracellular vesicles
95
Table 4.1 Top 25 ExoCarta proteins and their identification in DC2 epididymal EVs
ExoCarta ranka
Protein Gene
symbol
Unique matching sequences
% protein covered
1 CD9 antigen Cd9 4 18.6
2 Programmed cell death 6-interacting protein
(Alix) Pdcd6ip 21 17.4
3 Heat shock cognate 71 kDa protein Hspa8 18 30.3
4 Glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) Gapdh 8 21.6
5 Actin, cytoplasmic 1 Actb 25 34.1 6 Annexin A2 Anxa2 13 29.5 7 CD63 antigen Cd63 4 12.2 8 Syntenin-1 Sdcbp 16 31.8 9 Alpha-enolase Eno1 2 6.2 10 Heat shock protein HSP 90-alpha Hsp90aa1 13 19 11 Tumor susceptibility gene 101 protein Tsg101 0 0 12 Pyruvate kinase PKM Pkm 5 12.2 13 L-lactate dehydrogenase A chain Ldha 5 19.6 14 Elongation factor 1-alpha 1 Eef1a1 7 17.3 15 14-3-3 protein zeta/delta Ywhaz 8 33.9 16 Phosphoglycerate kinase 1b Pgk1 1 8.2 17 Elongation factor 2 Eef2 3 6.9 18 Fructose-bisphosphate aldolase A Aldoa 3 16.2 19 Heat shock protein HSP 90-beta Hsp90ab1 12 21.5 20 Annexin A5 Anxa5 12 32.9 21 Fatty acid synthase Fasn 0 0 22 14-3-3 protein epsilon Ywhae 2 7.1 23 Clathrin heavy chain 1 Cltc 10 8.1 24 CD81 antigen Cd81 0 0 25 Serum albumin Alb 9 11.5
a From http://exocarta.org/exosome_markers_new (152) b Detected below the matching peptide cut-off of 2
Additionally, I identified Hsp90b1 (Endoplasmin/heat shock protein 90 kDa beta
member 1) and Tubb (beta tubulin), two proteins that have been previously used as
epididymal EV markers in many species (49,204,219). The presence of these two
epididymal EV protein markers suggest that these EVs have commonality with those
isolated from the in vivo epididymis.

The effect of folate availability on epididymal extracellular vesicles
96
Taken together, the size distribution and protein composition indicate that the particles
purified from epididymal cell conditioned culture medias are EVs; while most likely
exosomes, in this thesis I will continue to refer to them as EVs.
4.2.2 Characterisation of epididymal EV small RNA cargo
4.2.2.1 Epididymal EV small RNA cargo
To determine the profile of RNA carried by epididymal EVs with respect to their
parent cells, I extracted total RNA from epididymal cells and RNase-treated
epididymal EVs. Due to low yields of epididymal EVs and therefore their RNA, six EV
preparations were pooled together prior to RNA extraction. Epididymal cell and EV
RNA was analysed on an Agilent Bioanalyser for initial comparison. Epididymal cells
displayed an electropherogram trace with prominent rRNA peaks, while epididymal
EVs lacked rRNA and were enriched with smaller RNAs less than 200 nt (Figure 4.3).

The effect of folate availability on epididymal extracellular vesicles
97
Figure 4.3 Size distribution of RNAs in epididymal cells and EVs
Electropherogram traces of RNA isolated from DC2 epididymal cells and EVs. An Agilent RNA Pico chip used to show the 25-4,000 nt range on the left and an Agilent small RNA Pico chip used to show the 4-150 nt range on the right.
To interrogate the <40 nt small RNA fraction, I constructed small RNA libraries from
epididymal EVs and cells and sequenced them on the Illumina platform. For initial
analyses, small RNA reads were conflated based on their annotation biotype (Figure
4.4a) and read lengths of all mapped reads were compared (Figure 4.4b). Both
annotation and size indicate that miRNAs and tRNA-derived sequences comprise the
majority of all epididymal cell and EV reads. All further analyses based on counts
normalised within biotypes, as in the previous chapter.

The effect of folate availability on epididymal extracellular vesicles
98
Figure 4.4 Epididymal cell and EV biotype and read length distribution
(a) Mapping regions (annotated against miRbase v21, and UCSC genomic tRNA, UCSC RepeatMasker, and UCSC RefSeq) for small RNA reads of epididymal cells and EVs. (b) Length of small RNA reads in epididymal cells and EVs. Each group represents a pool of 6 matching EV and cell preparations.
4.2.2.2 Selective packaging of miRNA into epididymal EVs
To identify any miRNAs selectively packaged into epididymal EVs, I compared counts
for each of the 171 mature miRNA species that passed the abundance filter (>100
counts per million miRNA reads). The top 10 most abundant miRNAs in each of
epididymal cells and EVs (Table 3.2) accounted for 68% and 70% all miRNA reads in
epididymal cells and EVs respectively. Among the most abundant miRNAs were miR-
10a-5p, miR-10b-5p and miRNAs of the let-7 family – all abundant in sperm (44,220)
and in vivo epididymal EVs (49). No gene ontologies were significantly enriched by the
1086 predicted gene targets of the top 10 most abundant EV miRNAs.

The effect of folate availability on epididymal extracellular vesicles
99
Table 4.2 Top 10 most abundant miRNAs in epididymal EVs
miRNA Epididymal EVs Epididymal cells
Normalised counta Rank Normalised counta Rank let-7i-5p 155,654 1 205,432 1 let-7f-5p 108,896 2 82,442 3
miR-10a-5p 106,137 3 92,659 2 miR-21a-5p 104,972 4 77,832 4
let-7g-5p 47,175 5 38,493 8 let-7c-5p 41,191 6 45,287 5
miR-10b-5p 40,133 7 21,395 12 miR-30a-5p 35,679 8 41,466 6 miR-30d-5p 32,397 9 22,476 11 miR-182-5p 30,159 10 33,547 9
a Per million miRNA reads
To identify any miRNAs that were selectively packaged into or excluded from
epididymal cells EVs, I filtered for miRNAs with a >2-fold change in expression
between epididymal EVs and cells. Forty percent of identified miRNAs were detected
within a 2-fold change in relative abundance (Figure 4.5). Forty-two miRNAs (25%)
were found in EVs at two times the abundance in cells and thus were deemed
selectively packaged (Table 3.3), while 60 miRNAs (35%) were >2-fold more abundant
in cells than EVs and deemed excluded (Table 3.4). To determine whether these
selectively packaged/excluded miRNAs targeted any ontologies, I predicted their gene
targets and submitted them for functional annotation enrichment analysis. The
cytoplasmic cellular component and ‘DNA binding’ and ‘protein binding’ molecular
functions were overrepresented in the predicted gene targets of the 35 selectively
packaged miRNAs (Figure 4.6), while no gene ontologies were overrepresented by the
targets of the selectively excluded miRNAs.

The effect of folate availability on epididymal extracellular vesicles
100
Figure 4.5 Relative abundance of miRNAs between epididymal cells and EVs
Scatterplot showing the relative abundance of individual miRNAs in epididymal cells (y-axis) vs epididymal EVs (x-axis). Grey miRNAs were excluded by abundance filter (<100 counts per million miRNA reads). Red miRNAs were significantly selective packaged into or excluded from epididymal EVs. Dotted lines represent the threshold for 2-fold change in abundance.

The effect of folate availability on epididymal extracellular vesicles
101
Table 4.3 miRNAs selectively packaged into epididymal EVs
miRNA Epididymal EV averagea Epididymal cell averagea EV fold-enrichment miR-382-5p 110 0 ∞
miR-142a-5p 160 0.4 399.82 miR-493-5p 227 1.6 142.19 miR-379-5p 680 7.2 94.48
miR-194-2-3p 162 2.0 81.09 miR-409-3p 205 2.8 73.21
miR-486a-5p 3,145 53 59.15 miR-126a-3p 1,846 38 48.10 miR-486b-5p 1,090 23 47.82 miR-143-3p 2,161 125 17.27 miR-1a-3p 272 24 11.36
miR-199a-3p 131 16 8.17 miR-365-2-5p 133 19 7.07 miR-224-5p 684 98 7.02 miR-122-5p 504 73 6.90
miR-344d-3p 675 102 6.63 miR-20a-5p 2,060 378 5.45
miR-200b-3p 187 39 4.77 miR-1198-5p 236 50 4.77 miR-676-3p 284 65 4.38 miR-340-5p 3,805 905 4.21
miR-181a-2-3p 230 58 3.93 miR-140-5p 160 41 3.88 miR-451a 191 50 3.80
miR-125a-5p 5,156 1,460 3.53 miR-322-3p 534 156 3.41 miR-671-3p 153 46 3.33
let-7d-3p 736 251 2.94 miR-196a-1-3p 110 41 2.71 miR-196a-5p 982 375 2.61 miR-148a-3p 4,097 1,599 2.56 miR-103-3p 5,766 2,351 2.45 miR-100-5p 3,472 1,426 2.43 miR-27b-5p 124 52 2.40 miR-101a-3p 2,598 1,084 2.40 miR-28a-3p 3,114 1,331 2.34 miR-191-5p 3,629 1,672 2.17
miR-3057-5p 164 78 2.12 miR-17-5p 892 424 2.10
miR-672-5p 588 287 2.05 miR-183-5p 24,776 12,155 2.04 miR-671-5p 396 196 2.02
a Per million miRNA reads
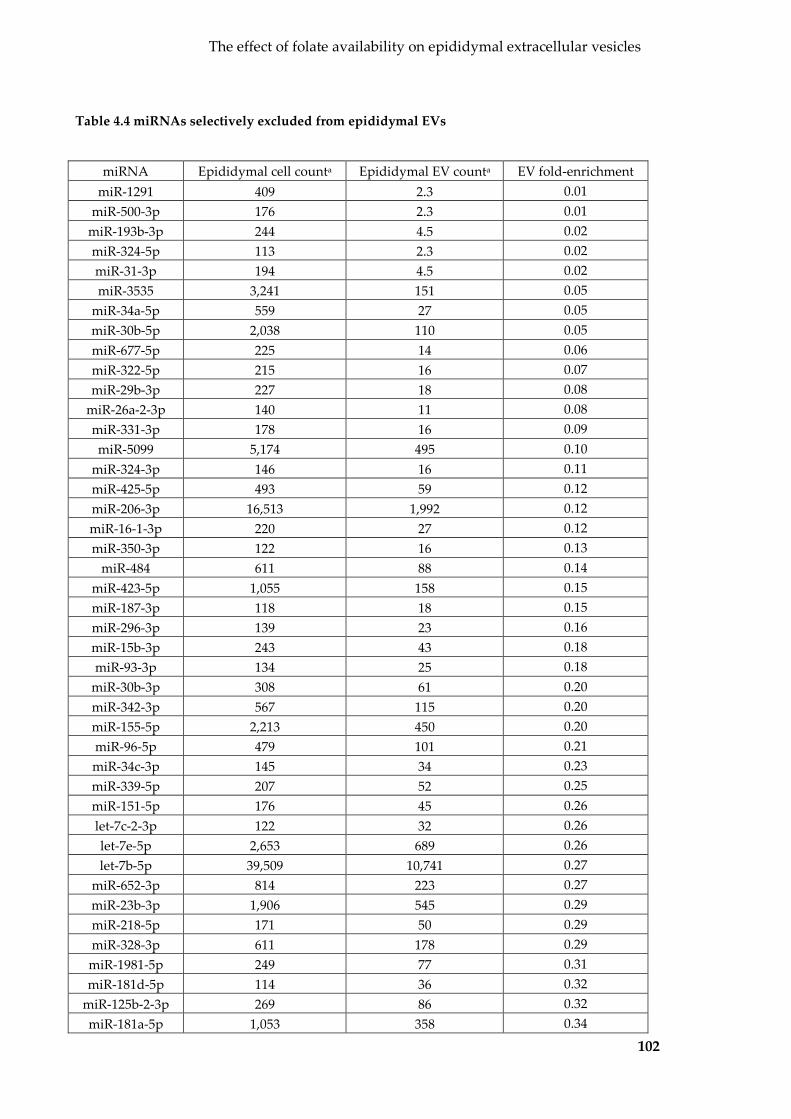
The effect of folate availability on epididymal extracellular vesicles
102
Table 4.4 miRNAs selectively excluded from epididymal EVs
miRNA Epididymal cell counta Epididymal EV counta EV fold-enrichment miR-1291 409 2.3 0.01
miR-500-3p 176 2.3 0.01 miR-193b-3p 244 4.5 0.02 miR-324-5p 113 2.3 0.02 miR-31-3p 194 4.5 0.02 miR-3535 3,241 151 0.05
miR-34a-5p 559 27 0.05 miR-30b-5p 2,038 110 0.05 miR-677-5p 225 14 0.06 miR-322-5p 215 16 0.07 miR-29b-3p 227 18 0.08
miR-26a-2-3p 140 11 0.08 miR-331-3p 178 16 0.09 miR-5099 5,174 495 0.10
miR-324-3p 146 16 0.11 miR-425-5p 493 59 0.12 miR-206-3p 16,513 1,992 0.12 miR-16-1-3p 220 27 0.12 miR-350-3p 122 16 0.13
miR-484 611 88 0.14 miR-423-5p 1,055 158 0.15 miR-187-3p 118 18 0.15 miR-296-3p 139 23 0.16 miR-15b-3p 243 43 0.18 miR-93-3p 134 25 0.18
miR-30b-3p 308 61 0.20 miR-342-3p 567 115 0.20 miR-155-5p 2,213 450 0.20 miR-96-5p 479 101 0.21 miR-34c-3p 145 34 0.23 miR-339-5p 207 52 0.25 miR-151-5p 176 45 0.26 let-7c-2-3p 122 32 0.26 let-7e-5p 2,653 689 0.26 let-7b-5p 39,509 10,741 0.27
miR-652-3p 814 223 0.27 miR-23b-3p 1,906 545 0.29 miR-218-5p 171 50 0.29 miR-328-3p 611 178 0.29 miR-1981-5p 249 77 0.31 miR-181d-5p 114 36 0.32
miR-125b-2-3p 269 86 0.32 miR-181a-5p 1,053 358 0.34

The effect of folate availability on epididymal extracellular vesicles
103
miR-27b-3p 22,806 7,896 0.35 miR-130a-3p 120 43 0.36 miR-181b-5p 2,484 896 0.36
miR-98-5p 3,116 1,135 0.36 miR-421-3p 169 63 0.37 miR-221-3p 4,807 1,844 0.38 miR-320-3p 240 92 0.38 miR-184-3p 169 65 0.39 miR-22-3p 5,258 2,060 0.39
miR-106b-5p 727 295 0.41 miR-23a-3p 2,072 851 0.41 miR-27a-3p 2,387 986 0.41 miR-34b-3p 193 86 0.44
let-7d-5p 4,125 1,828 0.44 miR-374b-5p 383 171 0.45 miR-1306-3p 105 50 0.47 miR-26b-5p 3,798 1,821 0.48
a Per million miRNA reads
Figure 4.6 Gene ontologies overrepresented by the predicted targets of miRNAs
selectively packaged into epididymal EVs
Overrepresented gene ontologies (retrieved from GO_Direct database using DAVID v6.8) by the predicted gene targets (predicted by both TargetScanMouse v7.1 and microT-CDS v5) of miRNAs selectively packaged into epididymal EVs. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis.
4.2.2.3 Selective packaging of tRFs in epididymal cells and EVs
tRNA genes were the 2nd and 3rd most common mapping location for small RNA
reads from epididymal cells and EVs respectively. Most tRNA-derived transcripts in

The effect of folate availability on epididymal extracellular vesicles
104
epididymal cells and EVs were 31-32 nt in length (Figure 4.7a). They mapped almost
exclusively to the 5’ end (and 3’ end in some select cases) of mature tRNA molecules,
as was observed in Sertoli cells and EVs and hence will be hereon in referred to as
tRNA fragments (tRFs).
Figure 4.7 tRFs in epididymal cells and EVs
(a) Size distribution of tRFs in epididymal cells and EVs. (b) Top six most abundant tRFs in epididymal cells and EVs.
Individual tRF sequences were conflated based on the anticodon of the tRNA gene that
they mapped to. Not all anticodons were evenly represented, with the top six
anticodons accounting for 92% and 98% of all tRF reads in cells and EVs respectively
(Figure 4.7b). Interestingly, the epididymal EV tRF profile appeared less diverse than
cells, with the top three tRFs accounting for 95% of all tRF reads. tRF anticodons did
not follow the expected cellular representation of mature tRNAs (160), indicating that
they are unlikely to be remnants of cellular tRNAs but are rather subjected to some
form of processing. Of the six abundant tRFs, tRF5-Glu-CTC (Figure 4.7b) was two
times more abundant in EVs compared to cells, while tRF5-Lys-CTT, tRF5-His-GTG
and tRF5-Glu-TTC were over three times less abundant. Some of the lesser abundant

The effect of folate availability on epididymal extracellular vesicles
105
anticodons (tRF5-Glu-TTC, tRF5-Val-CAC and tRF5-Lys-TTT) were also present in
epididymal cells at over 10,000 counts (per million tRF reads), >3-fold higher than what
was found in EVs. My data shows that tRFs are generally selectively packaged from
epididymal cells to EVs. Importantly, this enrichment matches the increase in sperm
tRF abundance seen after epididymal transit, concurring with the current scientific
understanding that sperm acquire tRFs from epididymal EVs (24,34).
4.2.3 Effects of folate concentration on epididymal EV production and
small RNA cargo
4.2.3.1 Effect of folate concentration on EV production
The extent of small RNA selective packaging and exclusion observed in epididymal
EVs as well as the established transfer of RNAs from epididymal EVs to sperm (24,49)
suggest that any changes due to extreme folate concentrations may have a significant
impact on both epididymal EV and sperm RNA profiles. I sought to determine the
effect of varying folate availability on epididymal EV production and cargo.
Most cell culture media (including that of IMDM media recommended for DC2
epididmyal cells) contain extreme levels of folate acid, two orders of magnitude greater
than concentrations observed in vivo. To determine the effects of folate availability, I
used epididymal cells subjected to three different treatments (Figure 4.8). To determine
a baseline for epididymal EV production and cargo at physiological levels of folate, I
weaned DC2 epididymal cells onto physiological folate levels over seven passages.
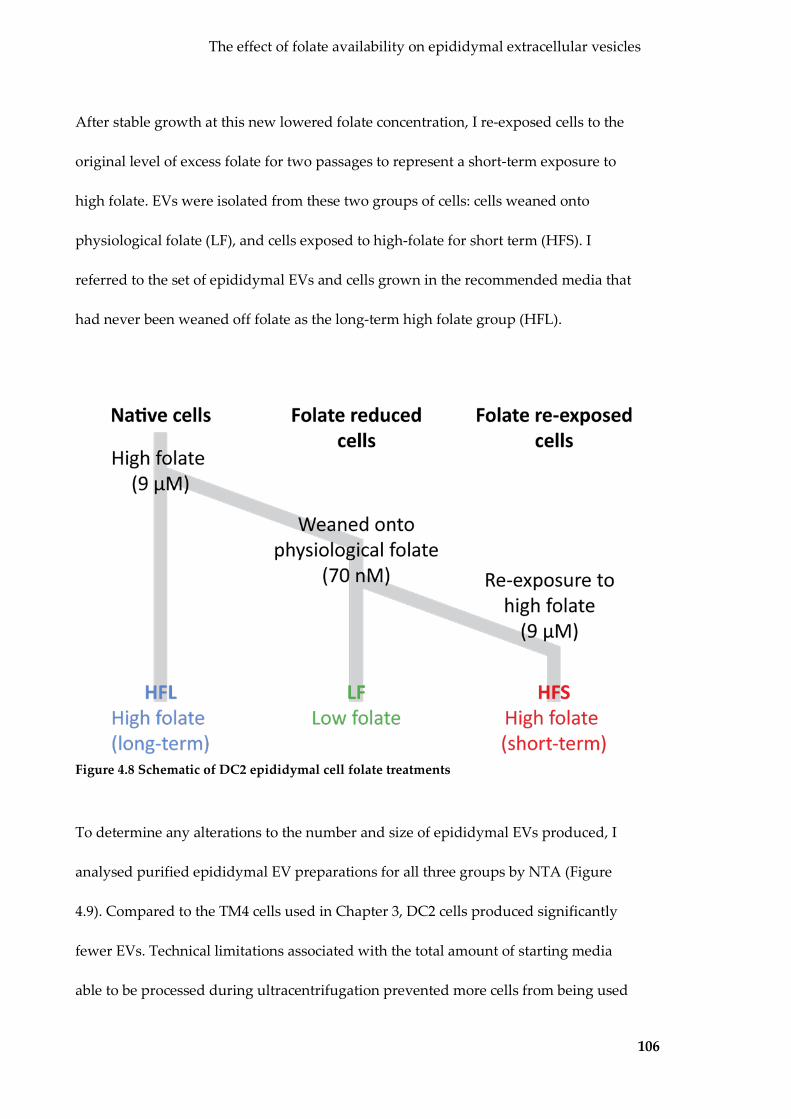
The effect of folate availability on epididymal extracellular vesicles
106
After stable growth at this new lowered folate concentration, I re-exposed cells to the
original level of excess folate for two passages to represent a short-term exposure to
high folate. EVs were isolated from these two groups of cells: cells weaned onto
physiological folate (LF), and cells exposed to high-folate for short term (HFS). I
referred to the set of epididymal EVs and cells grown in the recommended media that
had never been weaned off folate as the long-term high folate group (HFL).
Figure 4.8 Schematic of DC2 epididymal cell folate treatments
To determine any alterations to the number and size of epididymal EVs produced, I
analysed purified epididymal EV preparations for all three groups by NTA (Figure
4.9). Compared to the TM4 cells used in Chapter 3, DC2 cells produced significantly
fewer EVs. Technical limitations associated with the total amount of starting media
able to be processed during ultracentrifugation prevented more cells from being used

The effect of folate availability on epididymal extracellular vesicles
107
during EV harvesting, and time constraints prevented more preparations from being
prepared. As a result, only two replicates were measured by NTA and there was high
variation, however a difference in EV yield was observed. LF cells produced EVs at a
similar rate to HFL cells, however HFS cells produced 40% more EVs over the five days
of culture. There also appeared to be a change in the size of EVs produced. LF and HFS
EVs exhibited two populations of EVs, at ~110 nm and ~180 nm diameter. In addition
to these two groups, HFL EVs contained a third ~140 nm population.
Figure 4.9 Size and yield of EVs produced by epididymal cells after different treatments
of folate
Epididymal EV size distribution profile as determined by NTA. Shading represents SEM. n=2 per group.
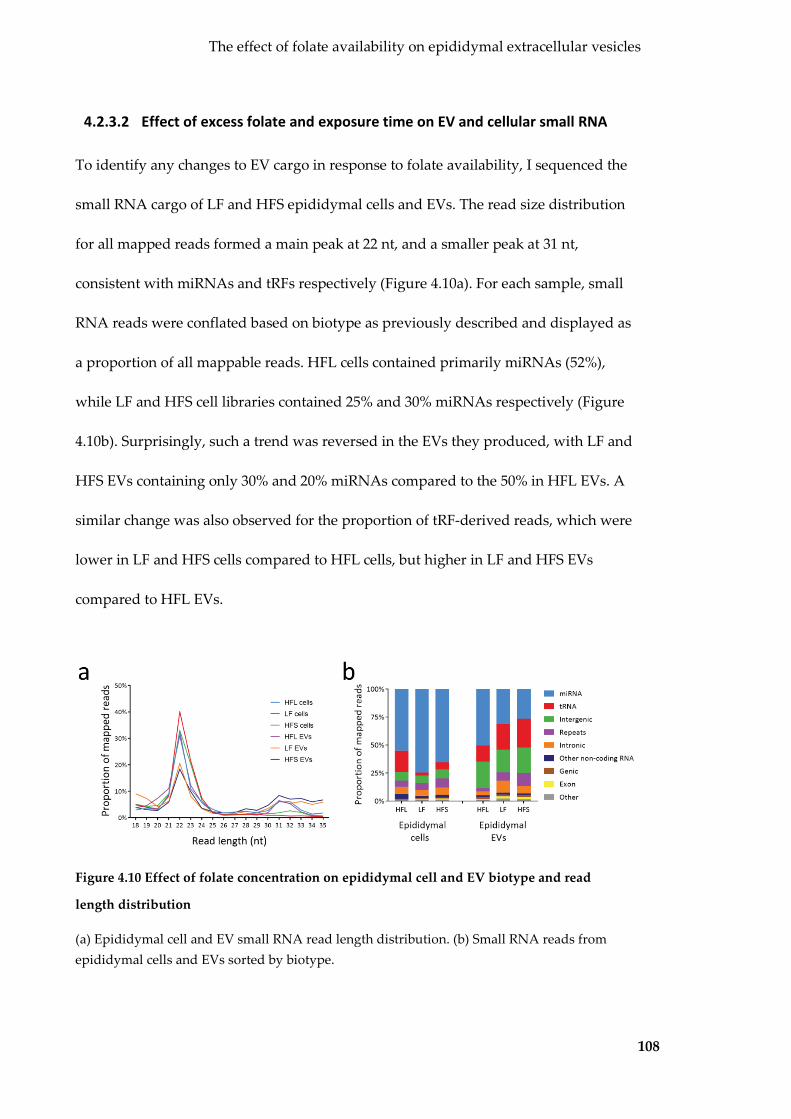
The effect of folate availability on epididymal extracellular vesicles
108
4.2.3.2 Effect of excess folate and exposure time on EV and cellular small RNA
To identify any changes to EV cargo in response to folate availability, I sequenced the
small RNA cargo of LF and HFS epididymal cells and EVs. The read size distribution
for all mapped reads formed a main peak at 22 nt, and a smaller peak at 31 nt,
consistent with miRNAs and tRFs respectively (Figure 4.10a). For each sample, small
RNA reads were conflated based on biotype as previously described and displayed as
a proportion of all mappable reads. HFL cells contained primarily miRNAs (52%),
while LF and HFS cell libraries contained 25% and 30% miRNAs respectively (Figure
4.10b). Surprisingly, such a trend was reversed in the EVs they produced, with LF and
HFS EVs containing only 30% and 20% miRNAs compared to the 50% in HFL EVs. A
similar change was also observed for the proportion of tRF-derived reads, which were
lower in LF and HFS cells compared to HFL cells, but higher in LF and HFS EVs
compared to HFL EVs.
Figure 4.10 Effect of folate concentration on epididymal cell and EV biotype and read
length distribution
(a) Epididymal cell and EV small RNA read length distribution. (b) Small RNA reads from epididymal cells and EVs sorted by biotype.

The effect of folate availability on epididymal extracellular vesicles
109
4.2.3.3 Effects of folate concentration on epididymal cell and EV miRNA profiles
First, I sought to compare the miRNA profiles of epididymal EVs and cells in different
concentrations of folate. To examine whether the expression of individual miRNAs
was affected, I conflated miRNA reads based on their mature miRNA sequence,
processed and normalised to total miRNA reads as previously described (for full list
see Table 8.5). PCA on the counts of all miRNAs showed that all three types of cells
clustered together, separated from EVs by PC1 (Figure 4.11a). Interestingly, HFL EVs
(and cells to a smaller extent) separated with LF and HFS EVs. Unsupervised
hierarchical clustering also showed separation by sample type (EV or cell), with a
similar segregation between LF and HFS EVs, and HFL EVs (Figure 4.11b). These
results suggest that epididymal EVs and cells have distinct miRNA profiles. Reducing
the level of folate to physiological levels altered epididymal cell and EV miRNA cargo
and a short-term re-exposure to original, supraphysiological levels of folate did not
reverse these changes.

The effect of folate availability on epididymal extracellular vesicles
110
Figure 4.11 Effect of folate concentration on epididymal cell and EV miRNA profiles
(a) PCA on epididymal cell and EV miRNA cargo. (b) Unsupervised hierarchical clustering on epididymal cell and EV miRNA cargo.
4.2.3.4 Reducing folate to physiological levels affects epididymal cell and EV
miRNA uniquely
First, I sought to determine any differences in EV cargo with changes in folate levels by
comparing miRNA counts of HFL EVs and LF EVs. Only 46 (30%) of miRNAs were
similarly expressed within a 2-fold change in abundance (Figure 4.12a). Only 37 (24%)
miRNAs were upregulated after folate reduction (Table 4.5), with 71 (46%) miRNAs
downregulated by at least 50% (Table 4.6). The predicted targets for downregulated
and upregulated miRNAs were identified and functional enrichment analysis
performed as previously described. Upregulated miRNAs targeted genes associated
with many nucleic acid-binding-related functions and nuclear-related cellular
components (Figure 4.12b), while the ‘positive regulation of transcription – DNA-
templated’ biological function was overrepresented by downregulated miRNAs
(q=0.028, 6% genes associated).

The effect of folate availability on epididymal extracellular vesicles
111
Figure 4.12 Effects of folate reduction on epididymal EV miRNA
(a) Scatterplot showing the effect of reducing folate on epididymal EV. Grey miRNAs were excluded by abundance filter (<100 counts per million miRNA reads). Red miRNAs were significantly selectively packaged or retained. Dotted lines represent the threshold for 2-fold change in relative abundance. (b) Overrepresented gene ontologies (retrieved from GO_Direct database using DAVID v6.8) by the predicted gene targets (predicted by both TargetScanMouse v7.1 and microT-CDS v5) of miRNAs upregulated in epididymal EVs after reducing folate to physiological levels. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis.
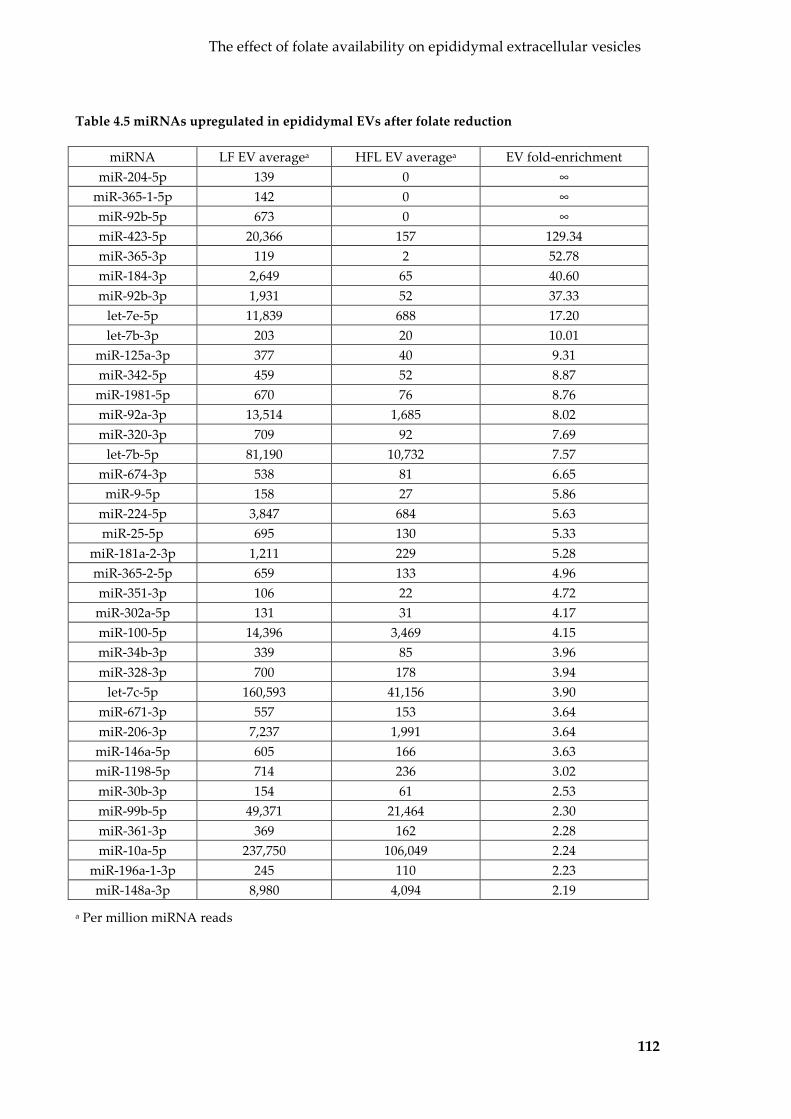
The effect of folate availability on epididymal extracellular vesicles
112
Table 4.5 miRNAs upregulated in epididymal EVs after folate reduction
miRNA LF EV averagea HFL EV averagea EV fold-enrichment miR-204-5p 139 0 ∞
miR-365-1-5p 142 0 ∞ miR-92b-5p 673 0 ∞ miR-423-5p 20,366 157 129.34 miR-365-3p 119 2 52.78 miR-184-3p 2,649 65 40.60 miR-92b-3p 1,931 52 37.33
let-7e-5p 11,839 688 17.20 let-7b-3p 203 20 10.01
miR-125a-3p 377 40 9.31 miR-342-5p 459 52 8.87 miR-1981-5p 670 76 8.76 miR-92a-3p 13,514 1,685 8.02 miR-320-3p 709 92 7.69
let-7b-5p 81,190 10,732 7.57 miR-674-3p 538 81 6.65
miR-9-5p 158 27 5.86 miR-224-5p 3,847 684 5.63 miR-25-5p 695 130 5.33
miR-181a-2-3p 1,211 229 5.28 miR-365-2-5p 659 133 4.96 miR-351-3p 106 22 4.72 miR-302a-5p 131 31 4.17 miR-100-5p 14,396 3,469 4.15 miR-34b-3p 339 85 3.96 miR-328-3p 700 178 3.94
let-7c-5p 160,593 41,156 3.90 miR-671-3p 557 153 3.64 miR-206-3p 7,237 1,991 3.64 miR-146a-5p 605 166 3.63 miR-1198-5p 714 236 3.02 miR-30b-3p 154 61 2.53 miR-99b-5p 49,371 21,464 2.30 miR-361-3p 369 162 2.28 miR-10a-5p 237,750 106,049 2.24
miR-196a-1-3p 245 110 2.23 miR-148a-3p 8,980 4,094 2.19
a Per million miRNA reads

The effect of folate availability on epididymal extracellular vesicles
113
Table 4.6 miRNAs downregulated in epididymal EVs after folate reduction
miRNA miRNA count in HFL EVa miRNA count in LF EVa Fold-enrichment miR-140-5p 160 0 0 miR-671-5p 396 2 0.00 miR-20a-5p 2,058 27 0.01 miR-142a-5p 160 3 0.02 miR-493-5p 227 5 0.02 miR-194-5p 229 6 0.03 miR-362-5p 328 9 0.03 miR-340-5p 3,802 142 0.04 miR-17-5p 891 40 0.04
miR-103-3p 5,761 260 0.05 miR-31-5p 1,451 73 0.05 miR-34c-5p 3,833 198 0.05 miR-652-3p 223 13 0.06
miR-101b-3p 8,766 543 0.06 miR-30a-5p 35,650 2,243 0.06 miR-186-5p 758 49 0.06 miR-93-5p 5,122 369 0.07 miR-16-5p 637 46 0.07
miR-199a-3p 130 9 0.07 miR-542-3p 601 47 0.08 miR-192-5p 1,651 133 0.08
miR-106b-5p 295 24 0.08 miR-143-3p 2,159 187 0.09
let-7g-5p 47,135 4,124 0.09 miR-30e-5p 1,017 90 0.09 miR-361-5p 142 13 0.08937 miR-379-5p 679 66 0.097878 miR-101a-3p 2,596 256 0.098799
miR-451a 191 21 0.107638 miR-185-5p 610 66 0.109074 miR-26b-5p 1,820 204 0.112223
miR-194-2-3p 162 19 0.117298 miR-29a-3p 1,251 150 0.120252 miR-24-3p 2,380 294 0.123728
miR-140-3p 3,903 524 0.134267 miR-152-3p 3,385 469 0.138419 miR-126a-3p 1,845 260 0.140758 miR-1839-5p 877 136 0.155194 miR-345-3p 241 38 0.157859 miR-27b-3p 7,889 1,363 0.172786 miR-15b-5p 169 32 0.187677
miR-374b-5p 171 33 0.194468 miR-27a-3p 985 195 0.197639
The effect of folate availability on epididymal extracellular vesicles
114
miR-7a-5p 7,461 1,488 0.199445 miR-3535 151 33 0.22059
miR-221-3p 1,842 426 0.231159 miR-378a-3p 3,529 866 0.245361
let-7i-5p 155,524 39,430 0.25353 miR-10a-3p 155 40 0.254996 miR-22-3p 2,058 530 0.257671
miR-28a-5p 290 78 0.26733 miR-196a-5p 981 266 0.271184 miR-200b-3p 187 51 0.27134 miR-27b-5p 124 35 0.281515
let-7f-5p 108,805 32,527 0.298952 miR-98-5p 1,134 347 0.305813
miR-322-3p 533 169 0.317744 miR-148b-3p 2,420 785 0.324423 miR-191-5p 3,626 1,303 0.359316 miR-183-5p 24,755 8,983 0.362862 miR-222-5p 130 47 0.364028 miR-24-2-5p 187 68 0.364613 miR-96-5p 101 38 0.375354
miR-532-5p 5,495 2,137 0.388913 miR-382-5p 110 44 0.402164
miR-181b-5p 895 363 0.404943 miR-26a-5p 12,730 5,182 0.40705 miR-30d-5p 32,370 15,266 0.471621 miR-181a-5p 358 171 0.478045 miR-28a-3p 3,111 1,537 0.494127 miR-21a-5p 104,884 51,992 0.495713
a Per million miRNA reads
Next, to determine if miRNAs were similarly dysregulated in the parent cells
themselves, I compared miRNA counts between HFL cells and LF cells. Fewer
miRNAs were upregulated (13%), a similar percentage downregulated (47%), and a
greater proportion (40%) similarly expressed in cells after folate reduction (Figure
4.13). This shows that epididymal cell miRNAs were also affected by folate reduction,
but not to the same extent as EV miRNAs. This implies that folate reduction affects the

The effect of folate availability on epididymal extracellular vesicles
115
EV miRNA cargo sorting process in epididymal cells and suggests that epididymal EV
miRNA is more sensitive to a reduction in folate compared to their parent cells.
Figure 4.13 Effects of folate reduction on epididymal cell miRNA
Scatterplot showing the effect of reducing folate on miRNAs in epididymal cells. Grey miRNAs were excluded by abundance filter (<100 counts per million miRNA reads). Red miRNAs were significantly selectively packaged or excluded. Dotted lines represent the threshold for 2-fold change in abundance.
4.2.3.5 Short-term re-exposure to excess folate does not alter epididymal cell and
EV miRNAs
Having observed changes to epididymal cell and EV miRNA profiles after a reduction
in folate, I asked to see what effect a short-term (10 day) re-exposure to high folate had
on epididymal cells and EVs. Based on initial biotype representation analysis, HFS EVs
contained fewer miRNAs than LF EVs (Figure 4.4a). To determine whether any the
expression of any individual miRNAs had changed, I compared miRNA profiles of LF
EVs with that of HFS EVs. Both sets of EVs were similar with the clear majority (88%)

The effect of folate availability on epididymal extracellular vesicles
116
of all miRNAs expressed within a 2-fold change in abundance (Figure 4.14a). Twelve
miRNAs (9%) were present at over 2-fold greater abundance in HFS EVs vs LF EVs
(Table 4.7), while four miRNAs (3%) had reduced by >50% reduced (Table 4.8). The
‘nucleus’ cellular component gene ontology was enriched by the predicted gene targets
of upregulated miRNAs (q=0.0062, 47% genes associated), while RNA-binding-related
molecular functions were overrepresented in those of downregulated miRNAs (Figure
4.12).
Figure 4.14 Effects of re-exposure to high folate on epididymal EV and cell miRNA
Scatterplot showing the effect of high folate re-exposure on epididymal (a) EV and (b) cell miRNA profiles. Grey miRNAs were excluded by abundance filter (<100 counts per million miRNA reads). Red miRNAs were significantly selectively packaged or excluded. Dotted lines represent the threshold for 2-fold change in abundance.

The effect of folate availability on epididymal extracellular vesicles
117
Table 4.7 miRNAs upregulated in epididymal EVs after short-term re-exposure to excess
folate
miRNA HFS EV counta LF EV counta Fold-enrichment miR-351-3p 371 106 3.50 miR-10a-3p 130 40 3.28 miR-342-5p 1,430 459 3.11 miR-185-5p 203 66 3.05 miR-16-5p 135 46 2.94
miR-23b-5p 171 63 2.69 miR-31-5p 189 73 2.60
miR-365-1-5p 351 142 2.47 miR-1843b-3p 161 70 2.31 miR-200b-3p 109 51 2.15 miR-298-5p 146 68 2.14 miR-1249-3p 145 70 2.07
a Per million miRNA reads
Table 4.8 miRNAs downregulated in epididymal EVs after short-term re-exposure to
excess folate
miRNA HFS EV counta LF EV cell counta Fold-enrichment miR-184-3p 2,649 119 0.04 miR-1a-3p 283 18 0.06 miR-9-5p 158 58 0.37
miR-148a-3p 8,980 4,425 0.49
a Per million miRNA reads

The effect of folate availability on epididymal extracellular vesicles
118
Figure 4.15 Gene ontologies overrepresented by the predicted targets of miRNAs
downregulated in epididymal EVs after short-term exposure to excess folate
Overrepresented gene ontologies (retrieved from GO_Direct database using DAVID v6.8) by the predicted gene targets (predicted by both TargetScanMouse v7.1 and microT-CDS v5) of miRNAs downregulated in epididymal EVs after short-term exposure to excess folate. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis.
Cells were comparably similar with the clear majority (94%) of miRNAs expressed
within a 2-fold difference in abundance (Figure 4.14b), suggesting that miRNA
shuttling into EVs was not affected by the short-term exposure to excess folate. Taken
together, these data suggest that a short-term exposure to excess folate does not reverse
the alterations caused by a stable reduction in folate availability.
4.2.3.6 Differences between short- and long-term high folate exposure on
epididymal cell and EV miRNA cargo
Previous data (PCA, unsupervised hierarchical clustering, and miRNA comparisons)
suggest that the length of folate exposure impacts on epididymal cell and EV miRNA
cargo. To confirm this, I compared miRNA counts for epididymal cells and EVs
produced after short- and long-term exposure to folate. Fifty (32%) miRNAs were
present within a 2-fold change in relative abundance, 40 (26%) miRNAs were present

The effect of folate availability on epididymal extracellular vesicles
119
at a higher relative abundance in HFS EVs, while 66 (42%) were more abundant in HFL
EVs (Figure 4.16a). Similar differences were also observed between HFL and HFS cells
(50%, 12% and 38% respectively; Figure 4.16b). EVs appeared to be more affected by
length of folate exposure, consistent with previous observations that EVs are more
sensitive to environmental stressors than cells in Chapter 3. These proportions were
similar to that seen between HFL and LF EVs and cells, which was expected
considering that LF and HFS EVs and cells were very similar.
Figure 4.16 Effect of short- vs long-term exposure on epididymal EV and cell miRNA
profiles
Scatterplot showing the effect of short- vs long-term folate exposure on epididymal EV miRNA (a) and cell miRNA (b). Grey miRNAs were excluded by abundance filter. Red miRNAs were >2-fold differentially expressed.
Taken together, this confirms that a short-term re-exposure to excess folate does not
reverse the effects of weaning epididymal cells from high to physiological folate.
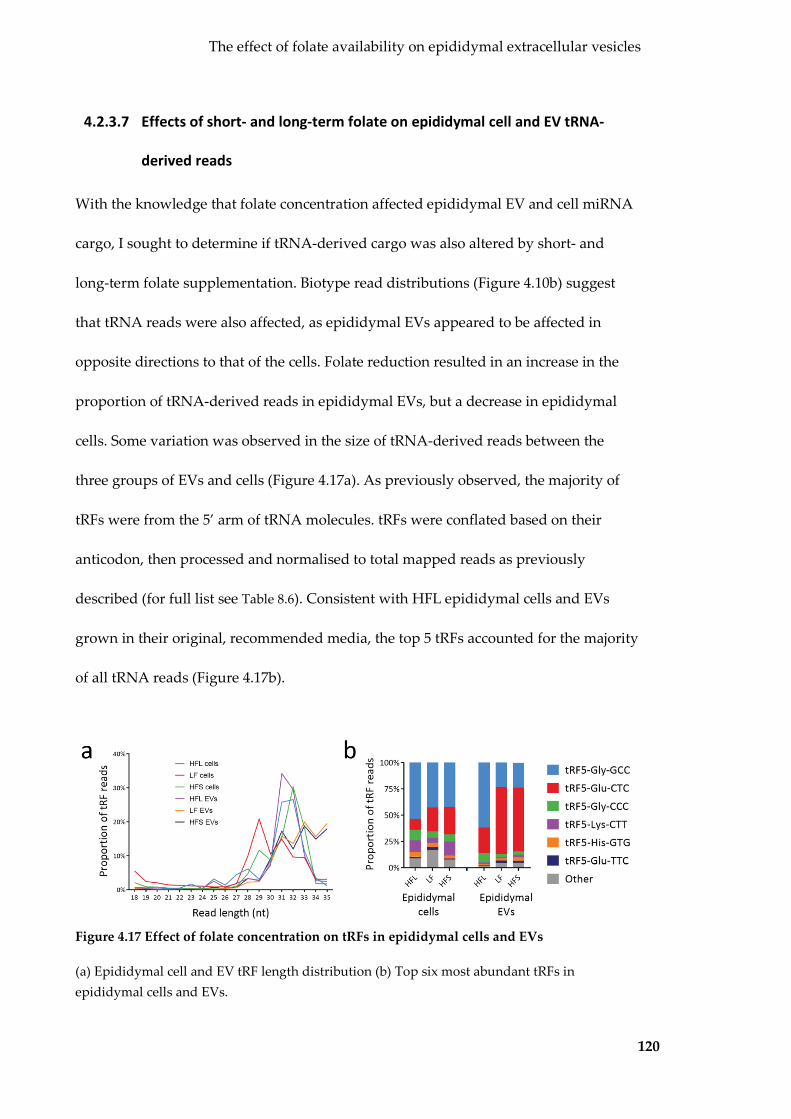
The effect of folate availability on epididymal extracellular vesicles
120
4.2.3.7 Effects of short- and long-term folate on epididymal cell and EV tRNA-
derived reads
With the knowledge that folate concentration affected epididymal EV and cell miRNA
cargo, I sought to determine if tRNA-derived cargo was also altered by short- and
long-term folate supplementation. Biotype read distributions (Figure 4.10b) suggest
that tRNA reads were also affected, as epididymal EVs appeared to be affected in
opposite directions to that of the cells. Folate reduction resulted in an increase in the
proportion of tRNA-derived reads in epididymal EVs, but a decrease in epididymal
cells. Some variation was observed in the size of tRNA-derived reads between the
three groups of EVs and cells (Figure 4.17a). As previously observed, the majority of
tRFs were from the 5’ arm of tRNA molecules. tRFs were conflated based on their
anticodon, then processed and normalised to total mapped reads as previously
described (for full list see Table 8.6). Consistent with HFL epididymal cells and EVs
grown in their original, recommended media, the top 5 tRFs accounted for the majority
of all tRNA reads (Figure 4.17b).
Figure 4.17 Effect of folate concentration on tRFs in epididymal cells and EVs
(a) Epididymal cell and EV tRF length distribution (b) Top six most abundant tRFs in epididymal cells and EVs.

The effect of folate availability on epididymal extracellular vesicles
121
After PCA using the counts of the top six tRFs, EVs appeared to separate based on PC1
by treatment and EVs and cells separated by PC2 (Figure 4.18a). Unsupervised
hierarchical clustering was able to distinguish cell samples from EV samples (Figure
4.18b). While RNA from different preparations were pooled prior to sequencing
leading to no replicates being available, these results indicate that epididymal EVs can
be separated from cells based on counts for the top six tRFs. Additionally, the top six
tRFs may also be able distinguish epididymal EVs produced under physiological
concentrations of folate from those produced under short- and long-term excess folate.
Figure 4.18 Effect of folate concentration on epididymal cell and EV tRF profiles
(a) PCA on top six tRFs in epididymal cells and EVs (b) Unsupervised hierarchical clustering on top six tRFs in epididymal cells and EVs
4.2.3.8 Folate reduction alters the abundance of tRFs in epididymal cells and EVs
Folate reduction resulted in more tRFs in epididymal EVs, and contrastingly, fewer in
their parent cells (Figure 4.10b). While the most parsimonious explanation would be
that tRFs are being shuttled into EVs, more work is required to confirm this. To further
investigate the effects of a reduction in folate on epididymal EV tRFs, I compared the

The effect of folate availability on epididymal extracellular vesicles
122
abundance of individual tRFs in HFL EVs and LF EVs. The general increase in the
proportion of tRFs in EVs could be attributed to the >2-fold increase of certain
anticodons: tRF5-Glu-CTC, tRF5-Glu-TTC, and tRF5-His-GTG. Despite the 50%
increase in tRFs in LF EVs compared to HFL EVs, several individual tRFs, tRF5-
Gly-GCC and tRF5-Gly-CCC, had decreased by >50%. This suggests that not all
epididymal EV tRFs were affected evenly and in the same direction by a reduction in
folate.
Taken together, this shows that a reduction in folate affects tRFs in epididymal EVs
and cells in opposing directions. This disparity has been seen before with miRNAs (see
section 4.2.3.4), and together implies that that folate availability influences EV cargo
directly by dysregulating the sorting of tRFs (in addition to miRNAs) from epididymal
cells into EVs.
4.2.3.9 Effect of short-term exposure to excess folate on tRFs in epididymal EVs
and cells
Next, I compared the tRF profiles of LF and HFS EVs. Short-term exposure to excess
folate caused a slight increase in the proportion of tRFs when compared to all other
small RNA types (Figure 4.10b). The top six anticodons were similarly expressed, being
present within a 2-fold change in abundance, except for tRF5-Lys-CTT which was two
times higher in HF EVs than LF EVs (Figure 4.17b). Short-term folate cells also had two
times tRF reads (Figure 4.10b), that was spread over the top six most abundant
anticodons (Figure 4.17b).

The effect of folate availability on epididymal extracellular vesicles
123
These changes were not as large as those caused by a reduction in folate and indicate
that a short-term exposure to excess folate does not reverse the previous effects of
folate reduction. This results in epididymal EVs and cells produced after short- and
long-term exposures to excess folate having a distinctly different tRF profiles, as was
the case with miRNAs. Taken together, a short-term re-exposure to excess folate does
not reverse the effects of folate reduction on epididymal EV tRFs.
4.3 Discussion
In this chapter, I established the small RNA cargo of epididymal EVs and asked
whether this cargo could be influenced by environmental signals. To do this, I purified
epididymal EVs from an established epididymal cell line, DC2, and validated their EV
and epididymal identity by TEM, NTA and protein mass spectrometry. I characterised
epididymal EV small RNA cargo with respect to their parent cells and identified
miRNAs and tRFs that were selective packaged inside EVs. Finally, I documented the
effects of reducing folate concentration to physiological levels followed by a short-term
re-exposure to excess folate on epididymal EV production, and the miRNA and tRF
cargo of epididymal EVs and their parent cells.
4.3.1 The isolation of DC2 epididymal EVs
Epididymal EVs, have been previous isolated and their function in sperm maturation
investigated (89-92). To date, the vast majority of epididymal EV studies involve the
isolation of EVs from biological fluids, with little focus on epididymal EVs produced
and isolated in vitro. Despite the lack of in vitro knowledge on epididymal EVs, I

The effect of folate availability on epididymal extracellular vesicles
124
decided to start with an in vitro model as it is a simpler, tractable system that allows for
the detection of any changes directly stemming from a change in folate concentration.
In this chapter I use the DC2 epididymal cell line as my epididymal cell model. The
DC2 epididymal cell line was established in 1980 from epididymal epithelial cells taken
from the distal caput cells taken of 10-12 week-old mice harbouring the SV40-LTA gene
(148). These cells are regarded as principal epithelial cells – the type that release
epididymal EVs – and have been used as a model epididymal principal cell line
(205,221).
I isolated DC2 epididymal EVs using my previously optimised method of EV isolated
as described in the Chapter 3. This method allows for the reproducible isolation of pure
EVs from conditioned culture media, while limiting the production of typical
contaminants such as cell fragments, ABs and intracellular organelles. While my TEM
images were slightly over-exposed and internal morphology was not clear, I was still
able to determine the size range of these EVs, which matched the size range of in vivo
epididymal EVs (34,49,92). Proteomic analyses showed great overlap with the most
common exosome protein markers, as well as curated EV proteins from two
compendia. While fewer exosome protein markers were identified than with Sertoli
EVs, this can be attributed to the lower total number of proteins identified in
epididymal EVs. No enrichment of ribosomal or intracellular organelle-derived
proteins was detected, and along with the absence of intact rRNA, suggests that I have
captured a population of EVs that is devoid of significant contamination (164,165).

The effect of folate availability on epididymal extracellular vesicles
125
Despite having similar EV/cell yields to TM4 Sertoli cells, DC2 epididymal cells grew
slower, and to a lower density at confluence, leading to lower absolute EV yields. To
collect substantial EVs for measurement and sequencing, I had to pool together many
EV preparations. Due to these low yields and time constraints, NTA results were
obtained from only two replicates. Nevertheless, EV size determined by NTA was
consistent with that determined by TEM and was within the range expected for
epididymal EVs (34,49,92). Proteomic analyses and RNA electropherogram traces
strongly suggest the successful purification of epididymal EVs without contamination.
One study has reported the isolation of epididymal EVs from the same DC2
epididymal cell line that I have used (205). While these isolated EVs were of the
expected size range of epididymal EVs (and similar to what I observed in my samples),
the authors did not perform extensive protein validation as the study was a targeted
analysis a select few factors. Additionally, these EVs were produced in response to
calcium ionophore induction and thus may not be representative of regular epididymal
EVs. My data demonstrates the reproducible isolation of epididymal EVs from cells of
the DC2 cell line, with validation by TEM, NTA and protein mass spectrometry.
4.3.2 DC2 epididymal EVs contain selectively packaged cargo
Due to low EV and RNA yields, I had pooled together six individual EV preparations
from which to prepare a single small RNA library. While I did not have any replicates
for statistical analyses, each sequenced library represents the rough average of six
replicates, and thus the effect of any outliers would be minimal. Epididymal cells and

The effect of folate availability on epididymal extracellular vesicles
126
EVs both contained small RNA profiles primarily comprising miRNAs and tRNA-
derived reads.
As seen before for EVs, miRNAs in epididymal EVs are found at different
concentrations to that in their producing cells (100,114). Among the most abundant EV
miRNAs were those of the let-7 family, miR-21a-5p, and others previously associated
with epididymal EVs in vivo (49). Little is known about the function of these miRNAs
in the epididymis and sperm maturation. One common role for miR-21a-5p and those
of the let-7 family is that of tumour suppression (173,222), which may suggest some
sort of regulatory role for maturating sperm. A group of miRNAs were identified at >2-
fold higher abundance in epididymal EVs than their parent cells, which were predicted
to target DNA- and protein-binding molecular functions. Compared to Sertoli EVs,
fewer miRNAs were selectively packaged inside epididymal EVs. Despite this, similar
gene ontologies were enriched by selectively packaged miRNAs, suggesting a similar
role for EVs produced from germline-associated somatic cells.
tRNA-derived reads were the 2nd and 3rd most abundant small RNA type in
epididymal cells and EVs respectively. These transcripts bear high similarity with
those that I found in Sertoli EVs, as well as many other EVs from different cell types
(100,142,159,176), and are likely to be tRFs. tRFs have been identified in epididymal
EVs and sperm (24,33,34), and appear to increase in abundance from the caput to the
cauda epididymis. My findings coincide with the current understanding that
epididymal EVs traffic tRFs to sperm (24,34). A notable difference is that tRFs make up
a smaller proportion of small RNA reads compared to those purified from the mouse

The effect of folate availability on epididymal extracellular vesicles
127
(24,34), the reasons for which could be manifold. They could be due to differences in
mouse strain, the segmental origin of the EVs (DC2 cells were established from
epithelial cells of the caput epididymis), in vitro vs in vivo source, and differences in
RNA extraction and library construction.
Not all tRF anticodons were present at similar abundances, with the top three
anticodons in epididymal EVs accounting for 95% of all tRF reads. These same tRFs are
known constituents of EVs from the epididymis (24,34) and other cell types (176). The
tRFs that were selectively packaged into or excluded from epididymal EVs were not
necessarily of the same anticodons as those in Sertoli EVs. Similarities were observed
for tRF5-Lys-CTT, which was excluded from both epididymal and Sertoli EVs.
However, tRF5-Glu-TTC, was excluded from epididymal EVs (four-fold depleted
compared to cells), but enriched in Sertoli EVs (three-fold compared to cells). This
suggests that the selective packaging of tRFs is not consistent between cell types, even
those within the male reproductive system. It remains to be elucidated whether tRFs
share a similar role or vary based on cell type.
4.3.3 Effect of folate concentration on epididymal EV production and
cargo
Since epididymal cells were grown in media that contained excess high folate (HFL;
long-term), I first weaned the cells onto a low physiological dose of folate (LF). This
process could be regarded as a stress in itself as a sub-population of cells may have
been selected for leading to LF cells to contain a genetically diverged population of

The effect of folate availability on epididymal extracellular vesicles
128
cells from the original HFL cells. To circumvent this potential confounder, I re-exposed
the LF epididymal cells to the original high folate for two passages (HFS) to act as a
direct control for this weaning process.
While both HFL and LF epididymal cells produced similar amounts of EVs, EV
production was increased by 50% from HFS cells. A 10-fold increase in folate has been
shown to increase growth and metabolic activity in the HT29 colon cancer cell line
(223). If a similar increase in metabolic activity occurred in the DC2 epididymal cell
line, then it may be responsible for the observed increase in EV production. Another
notable difference was in the sizes of EVs produced, where LF and HFS EV samples
lacked the 140 nm population observed in HFL samples. This extra population may
represent aggregation, as seen in TEM, or may be a sub-population of EVs only
released after long-term folate supplementation. Another reason could be the selection
and stress caused by the folate weaning process. Further evidence suggesting that the
process of weaning epididymal cells off folate had a significant impact on their EVs lie
in small RNA sequencing data. PCA and unsupervised hierarchical clustering of
miRNAs suggest that HFS and LF EVs separate from HFL EVs, with the same also
observed for their producing cells.
The abundance of many miRNAs had changed in LF vs HFL EVs, with the majority
having decreased. Among the decreased miRNAs included those of the let-7 family,
shown to be abundance in other epididymal EVs (49). Epididymal EVs are known
interactors with sperm and transfer many RNA (24,34) and proteins (89-92,206) to
sperm essential for maturation. While the effects of the altered miRNAs have not been

The effect of folate availability on epididymal extracellular vesicles
129
explicitly tested, any change in cargo may influence the RNAs transferred to sperm,
and thus have downstream effects on fertilisation, development of offspring.
The changes between HFL and LF EVs were triggered by a change in environment,
however it is not known whether they result from a folate reduction or stress during
the folate weaning process. It is possible that the cell population had been altered by
the folate reduction process. While care was taken to slowly wean the cells off the
supraphysiological folate levels (folate levels halved at each passage) and to allow cells
to recover after (cells were grown at physiological folate level for several passages
before EV harvest), it is possible that I had selected for a sub-population of cells and
thus my LF cells were no longer representative of DC2 epididymal cells. Additionally,
it is possible that the DC2 cell had adapted to growing in such high folate levels since
their establishment in 2002 (148). If this is the case, then the DC2 epididymal cells
would have been starved and stressed after being weaned onto what is normally a
regular physiological concentration of folate.
Re-exposing epididymal cells to excess folate for two passages did not reverse the
changes to cellular and EV miRNA cargo. HFS EVs and cells were most similar to LF
EVs and cells respectively, than their HFL counterparts. The lack of detectable changes
after folate re-exposure may be due to the short length of re-exposure. The process of
weaning cells off folate took over seven passages (28 days), and the short-term re-
exposure to excess folate may not have been long enough to affect the cells. There may
be a lag time for intracellular folate reserves to increase or replenish, and there may be
a lag time for folate receptors to increase on the cell surface in response to this increase

The effect of folate availability on epididymal extracellular vesicles
130
in folate. It is also possible that the cells had adapted to the low folate levels over the
weaning and were not able to make use of the excess folate available to them in such a
relatively short timeframe. Further experiments such as the measuring folate in the
media to see whether it has been taken up by cells, as well as measuring intracellular
folate concentrations may be able to determine this.
As discussed above, it is also possible that the epididymal cell population had been
stressed and altered during the folate weaning process, leading to no reversal after
folate re-exposure. Since DC2 cell line were grown in a supraphysiological folate
environment since their establishment, they may have adapted to this concentration.
During the folate weaning process I may have selected for a small population of cells,
thus resulting in a different population of cells used in the LF experiment. This can be
confirmed by comparing the cargo of LF EVs and cells that have been exposed to
excess folate for at least seven passages – the time taken for cells to be weaned off
supraphysiological folate.
The effects of folate reduction and re-exposure on tRF cargo appear more complicated.
While basic biotype representations suggest that folate reduction causes an increase in
tRFs in EVs but a decrease in their producing cells, such changes were not as obvious
after PCA and unsupervised hierarchical clustering. While many tRFs were
dysregulated, more work is required to fully understand their functional significance.
Greater differences were observed in epididymal EV miRNA profiles than those of
their parent cells, which suggests that EVs are more sensitive to environmental stress.
This observation concurs with findings from my previous chapter where BPA and

The effect of folate availability on epididymal extracellular vesicles
131
DMSO exposure have greater effects on Sertoli EV cargo than their parent cells. Unlike
the previous chapter, only one sequencing library (albeit comprising a pool of 6 EV
preparations) was prepared for each sample due to low yields. Since no replicates were
available, it would have been ideal to validate and confirm the presence of the miRNAs
and tRFs mentioned above by qPCR and confirm any differential expression.
Unfortunately, this was not possible as the low RNA yields which resulted in
insufficient leftover RNA after library prep for RT-qPCR.
4.3.4 Conclusion
In this chapter, I characterised EVs isolated from the conditioned culture media of DC2
epididymal cells. These epididymal EVs are much like their in vivo counterparts and
contain selectively packaged small RNA cargo. Reducing the concentration of folate to
physiological levels increases miRNAs and decreases tRFs in epididymal EVs, while
affecting miRNAs and tRFs in opposite directions in their producing cells. A short-
term re-exposure to excess folate was not able to reverse these effects. Taken together
my findings show that changes in folate availability influence the production of
epididymal EVs, as well as their small RNA cargo.

132
5 MOUSE EPIDIDYMAL EXTRACELLULAR VESICLES AND THE
EFFECTS OF DIETARY METHYL DONORS
5.1 Introduction
In the previous chapter, I established that folate availability can have profound effects
on epididymal EV production and cargo in vitro. In this chapter, I take my
investigations further and ask whether similar changes may occur in vivo, using
epididymal fluid derived from a mouse model of dietary methyl donor
supplementation.
Methyl donors have been associated with both maternal and paternal effects on
offspring (15,224-226). In my laboratory, we have studied the effects of dietary methyl
donor supplementation in the Avy mouse model (19,25). We showed that exposure to
excess dietary methyl donors during gestation allows for increased stability of the
epigenetic state of the Avy allele when inherited through the paternal germline (Figure
5.1a). Furthermore, epigenetic state can be passed down again to the next generation,
the grand-offspring of the initially exposed mouse (Figure 5.1b). In another
experiment, we reported a cumulative epigenetic effect in the paternal germline, after
both dietary methyl donor supplementation and selection of coat colour, for five
generations (25). This experiment has continued and the sub-colony of Avy mice has
now been exposed to dietary methyl donor supplementation for over 14 generations.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
133
Figure 5.1 Paternal epigenetic inheritance after dietary methyl donor supplementation in
the Avy mouse
Offspring coat colour phenotypes in the (a) F1 and (b) F2 generation after dietary methyl donor supplementation. In the Avy mouse, coat colour is an epigenetic biosensor for the epigenetic state of the Avy locus. Adapted from Cropley et. al. (19).
The two examples mentioned above demonstrate how a dietary methyl donor
supplementation results in a paternally-inherited phenotype. Thus, this Avy model
provides an ideal tool for me to use to investigate whether dietary methyl donors alter
epididymal EV production and small RNA cargo.
In this chapter I hypothesise that dietary methyl donors influence epididymal EV
concentration and small RNA cargo. I will test this hypothesis by comparing the
epididymal EVs produced from Avy/a mice fed a normal chow diet with those fed a
methyl donor supplemented (MDS) diet for over 14 generations. My specific aims are
to:
1. Optimise the isolation and characterisation of epididymal EVs from mouse
epididymal fluid.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
134
2. Determine the effects of dietary methyl donors on mouse epididymal EV
concentration and small RNA cargo
5.2 Results
5.2.1 Isolation and characterisation of mouse epididymal EVs
5.2.1.1 Isolation of EVs from mouse epididymal fluid
For the optimisation of techniques and initial characterisation, C57BL/6 mice were used
instead of Avy/a experimental mice as they were in abundance. Epididymal fluid was
collected from dissected mouse epididymides and pre-cleared of any cells and cellular
debris. Due to low yields epididymal fluid from five mice was pooled and used for
each EV preparation. Three pools were organised such that the average ages were
matched (Table 8.1)
Transmission electron microscopy (TEM) on EV pellet resuspensions revealed a
heterogenous population of EVs comprising <150 nm electron-dense particles with a
cup-shaped morphology (Figure 5.2a) and larger 150-200 nm electron-lucent particles
(Figure 5.2b). Both these populations of EVs have been previously observed in in vivo
epididymal EV preparations (49,92,219,227,228), however only the former was
observed in DC2 epididymal EVs. Nanoparticle tracking analysis (NTA) showed that
the main population was approximately 125 nm with a less abundant populations of
~170 nm and ~260 nm EVs (Figure 5.2c), coordinating with the two types shown by
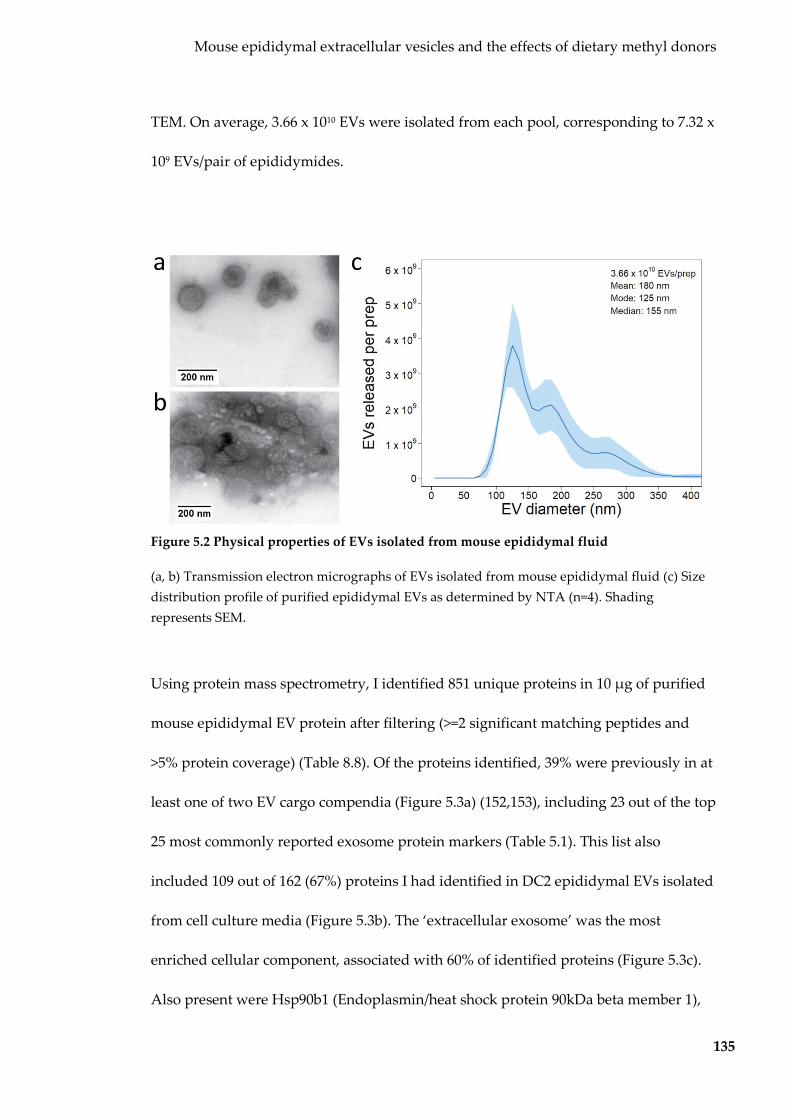
Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
135
TEM. On average, 3.66 x 1010 EVs were isolated from each pool, corresponding to 7.32 x
109 EVs/pair of epididymides.
Figure 5.2 Physical properties of EVs isolated from mouse epididymal fluid
(a, b) Transmission electron micrographs of EVs isolated from mouse epididymal fluid (c) Size distribution profile of purified epididymal EVs as determined by NTA (n=4). Shading represents SEM.
Using protein mass spectrometry, I identified 851 unique proteins in 10 µg of purified
mouse epididymal EV protein after filtering (>=2 significant matching peptides and
>5% protein coverage) (Table 8.8). Of the proteins identified, 39% were previously in at
least one of two EV cargo compendia (Figure 5.3a) (152,153), including 23 out of the top
25 most commonly reported exosome protein markers (Table 5.1). This list also
included 109 out of 162 (67%) proteins I had identified in DC2 epididymal EVs isolated
from cell culture media (Figure 5.3b). The ‘extracellular exosome’ was the most
enriched cellular component, associated with 60% of identified proteins (Figure 5.3c).
Also present were Hsp90b1 (Endoplasmin/heat shock protein 90kDa beta member 1),

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
136
Tubb (beta tubulin), and PSMD7 (26S proteasome non-ATPase regulatory subunit 7),
three cross-species epididymal EV markers (49,204,219). ALOX15 (arachidonate 15-
lipoxygenase), a marker for sperm cytoplasmic droplets, a similarly-sized contaminant
found in the epididymis (229), was not detected.
Figure 5.3 Proteins identified in mouse epididymal EVs
(a) Overlap between proteins identified in epididymal EVs and those curated in existing EV compendia. (b) Overlap between mouse epididymal EV proteins and in vitro produced epididymal EVs. (c) Top 10 most overrepresented ontologies (retrieved from GO_Direct database using DAVID v6.8) by epididymal EV proteins. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis. CC=cellular component, MF=molecular function.
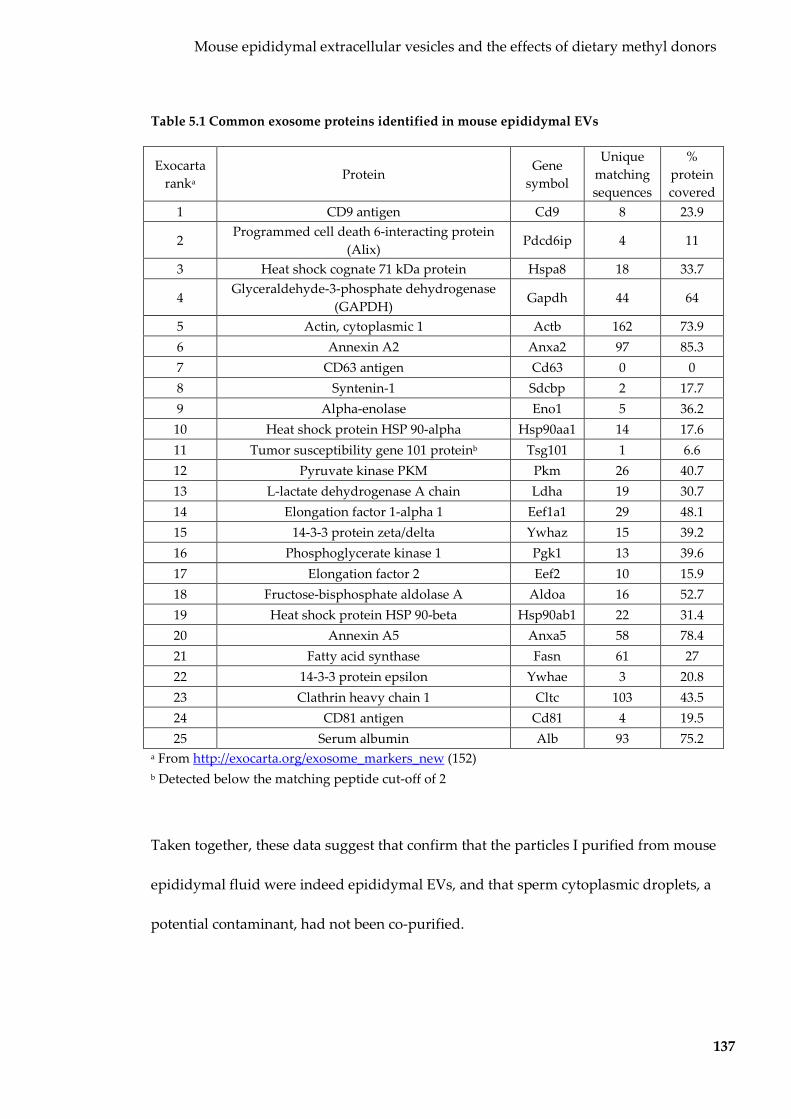
Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
137
Table 5.1 Common exosome proteins identified in mouse epididymal EVs
Exocarta ranka
Protein Gene
symbol
Unique matching sequences
% protein covered
1 CD9 antigen Cd9 8 23.9
2 Programmed cell death 6-interacting protein
(Alix) Pdcd6ip 4 11
3 Heat shock cognate 71 kDa protein Hspa8 18 33.7
4 Glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) Gapdh 44 64
5 Actin, cytoplasmic 1 Actb 162 73.9 6 Annexin A2 Anxa2 97 85.3 7 CD63 antigen Cd63 0 0 8 Syntenin-1 Sdcbp 2 17.7 9 Alpha-enolase Eno1 5 36.2 10 Heat shock protein HSP 90-alpha Hsp90aa1 14 17.6 11 Tumor susceptibility gene 101 proteinb Tsg101 1 6.6 12 Pyruvate kinase PKM Pkm 26 40.7 13 L-lactate dehydrogenase A chain Ldha 19 30.7 14 Elongation factor 1-alpha 1 Eef1a1 29 48.1 15 14-3-3 protein zeta/delta Ywhaz 15 39.2 16 Phosphoglycerate kinase 1 Pgk1 13 39.6 17 Elongation factor 2 Eef2 10 15.9 18 Fructose-bisphosphate aldolase A Aldoa 16 52.7 19 Heat shock protein HSP 90-beta Hsp90ab1 22 31.4 20 Annexin A5 Anxa5 58 78.4 21 Fatty acid synthase Fasn 61 27 22 14-3-3 protein epsilon Ywhae 3 20.8 23 Clathrin heavy chain 1 Cltc 103 43.5 24 CD81 antigen Cd81 4 19.5 25 Serum albumin Alb 93 75.2
a From http://exocarta.org/exosome_markers_new (152) b Detected below the matching peptide cut-off of 2
Taken together, these data suggest that confirm that the particles I purified from mouse
epididymal fluid were indeed epididymal EVs, and that sperm cytoplasmic droplets, a
potential contaminant, had not been co-purified.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
138
5.2.2 Interactions between epididymal EVs and spermatozoa
I next determined whether my purified epididymal EVs were able to interact with
spermatozoa. To do this I labelled purified mouse epididymal EVs with PKH26, a red
membrane dye and co-incubated them with live spermatozoa for three hours.
Spermatozoa were then washed, nuclear-stained, fixed and mounted on slides for
imaging via confocal microscopy. Mouse epididymal EVs bound to the sperm head,
and to some extent the midpiece (Figure 5.4a). No fluorescence was detected in the
PBS-labelled negative control (Figure 5.4b), indicating that no detectable fluorescence
was caused by dye micelles. The pattern of fluorescence observed with labelled EVs
was also distinct different to that of directly-labelled spermatozoa, which was evenly
distributed across the entire sperm structure (head, midpiece and tail) (Figure 5.4c),
suggesting that epididymal EVs specifically targeted the sperm head and midpiece.
This staining pattern of sperm is consistent with that previously observed by with both
murine and bovine epididymal EVs (92,227).
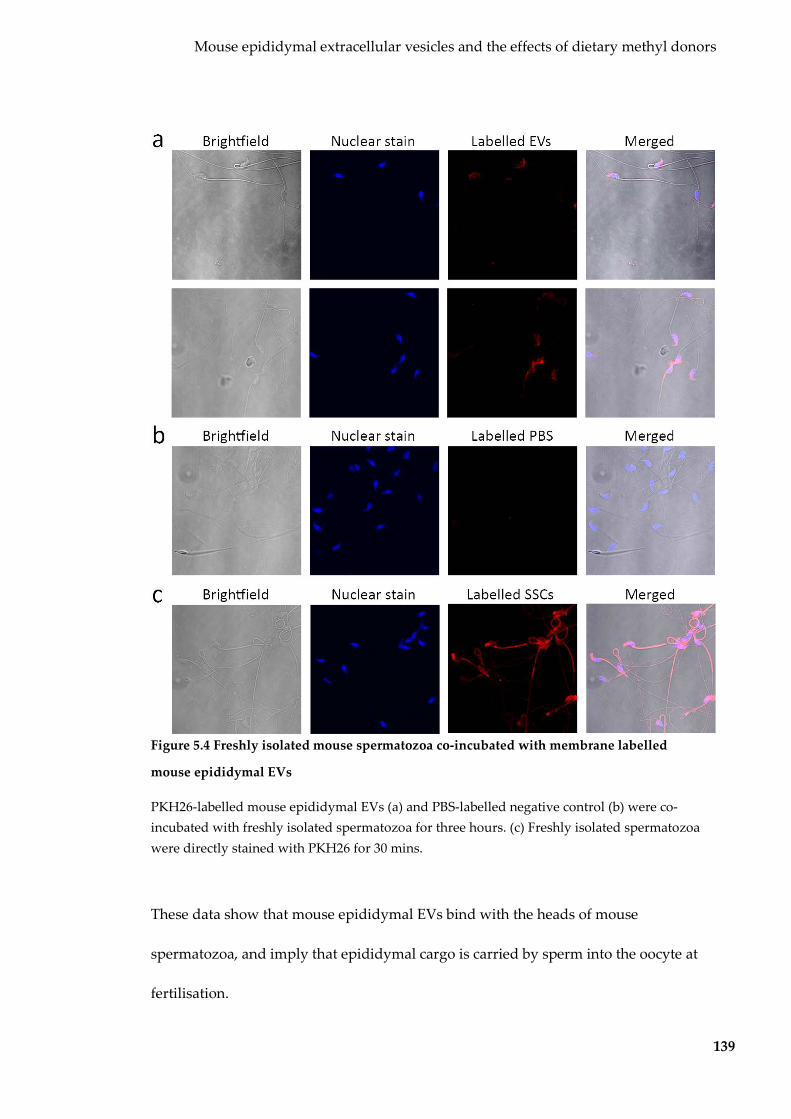
Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
139
Figure 5.4 Freshly isolated mouse spermatozoa co-incubated with membrane labelled
mouse epididymal EVs
PKH26-labelled mouse epididymal EVs (a) and PBS-labelled negative control (b) were co-incubated with freshly isolated spermatozoa for three hours. (c) Freshly isolated spermatozoa were directly stained with PKH26 for 30 mins.
These data show that mouse epididymal EVs bind with the heads of mouse
spermatozoa, and imply that epididymal cargo is carried by sperm into the oocyte at
fertilisation.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
140
5.2.3 Characterisation of mouse epididymal EV small RNA cargo
5.2.3.1 Small RNA cargo
To identify the small RNA cargo of mouse epididymal EVs, I extracted RNA from
RNase-treated epididymal EVs. Initial profiling on an Agilent Bioanalyser showed that
mouse epididymal EVs contained very little rRNA and are enriched <200 nt small
RNAs (Figure 5.2), a profile characteristic of EVs (164,165).
Figure 5.5 Size distribution of RNAs in mouse epididymal EVs
Electropherogram traces of RNA isolated from mouse epididymal EVs. An Agilent RNA Pico chip used to show the 25-4,000 nt range on the left and an Agilent small RNA Pico chip used to show the 4-150 nt range on the right.
Small RNA libraries were created from each pool, sequenced and processed as
previously described. On average, most small RNA reads in mouse epididymal EVs
mapped to tRNA genes, followed by unannotated intergenic regions and miRNAs
(Figure 5.6a). Read size distributions reflected the biotype representation with peaks
observed for miRNAs and tRFs (Figure 5.6b).

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
141
Figure 5.6 Epididymal EV biotype and read length distribution
(a) Mapping regions for small RNA reads of mouse epididymal EVs. (b) Length of small RNA reads in mouse epididymal EVs. n=3 pools.
5.2.3.2 Mouse epididymal EV miRNA profiles
Mouse epididymal miRNA reads were conflated and processed as previously
described. One hundred and twelve miRNAs were identified in mouse epididymal
EVs (Table 8.9). The top 10 most abundant miRNAs accounted for 75% of all miRNA
reads (Table 5.2). Interestingly, many of these miRNAs are primary constituents
miRNAs of sperm, such as miR-148-3p, miR-10a-5p, miR-10b-5p and the let-7 family
(44,46,220). Most of the top 10 most abundant miRNAs were also amongst the most
abundant in DC2 epididymal EVs (DC2 cells were established from the same strain of
mice, C57BL/6), except for miR-148a-3p, miR-200a-3p, and miR-143-3p. These top 10
miRNAs were predicted to have 711 high-confidence gene targets, from which DNA-
binding and transcription-related gene ontologies were overrepresented (Figure 5.7).
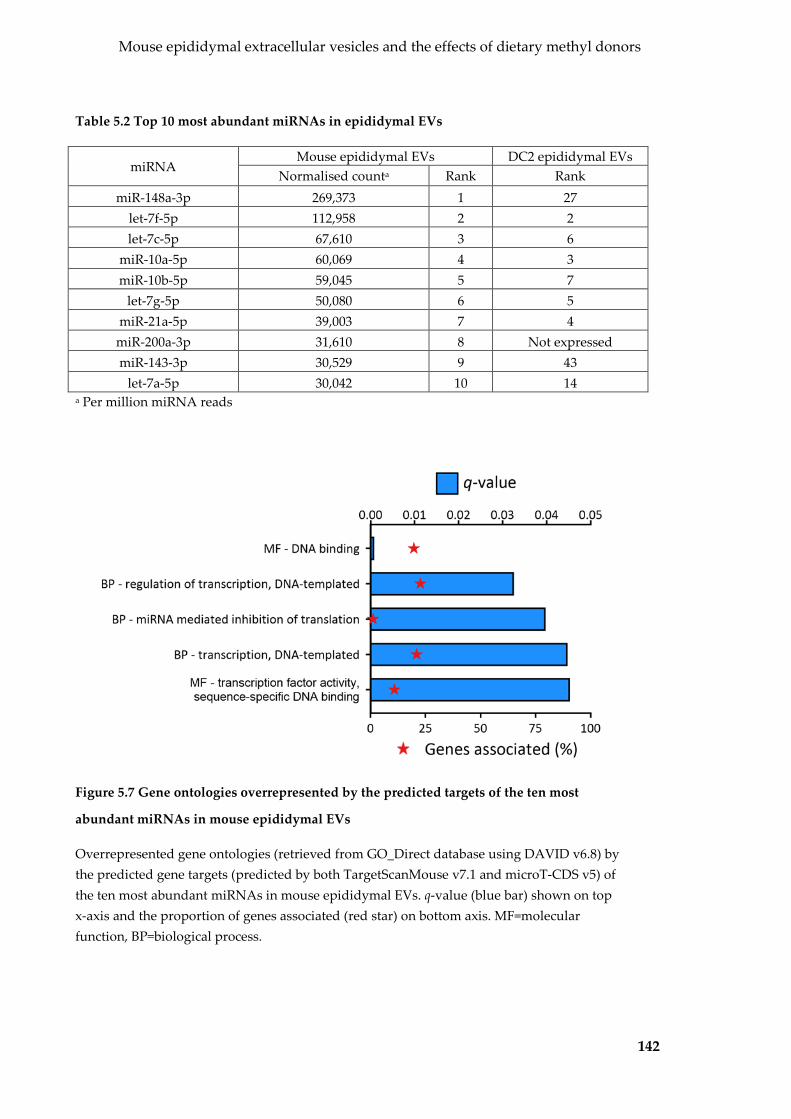
Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
142
Table 5.2 Top 10 most abundant miRNAs in epididymal EVs
miRNA Mouse epididymal EVs DC2 epididymal EVs
Normalised counta Rank Rank miR-148a-3p 269,373 1 27
let-7f-5p 112,958 2 2 let-7c-5p 67,610 3 6
miR-10a-5p 60,069 4 3 miR-10b-5p 59,045 5 7
let-7g-5p 50,080 6 5 miR-21a-5p 39,003 7 4
miR-200a-3p 31,610 8 Not expressed miR-143-3p 30,529 9 43
let-7a-5p 30,042 10 14 a Per million miRNA reads
Figure 5.7 Gene ontologies overrepresented by the predicted targets of the ten most
abundant miRNAs in mouse epididymal EVs
Overrepresented gene ontologies (retrieved from GO_Direct database using DAVID v6.8) by the predicted gene targets (predicted by both TargetScanMouse v7.1 and microT-CDS v5) of the ten most abundant miRNAs in mouse epididymal EVs. q-value (blue bar) shown on top x-axis and the proportion of genes associated (red star) on bottom axis. MF=molecular function, BP=biological process.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
143
Taken together, my purified mouse epididymal EVs have a miRNA profile similar to
mouse epididymal EVs isolated by others, sperm, to which they transfer RNA to, as
well as epididymal EVs that I isolated in vitro in the previous chapter. The most
abundant miRNAs are predicted to have roles with DNA binding and transcription,
suggesting that they may be able to alter gene expression after fertilisation.
5.2.3.3 Mouse epididymal EV tRF profiles
tRNA-derived reads comprised the majority of 18-32 nt small RNA reads in mouse
epididymal EVs (for full list see Table 8.10). As seen previously in both Sertoli and DC2
epididymal EVs, tRNA-derived reads were 31-32 nt in length (Figure 5.8a) and
mapped to the 5’ and 3’ end of tRNA genes, consistent with tRFs. The top six most
abundant tRF anticodons represented on average 98% of all tRF reads (Figure 5.8b).
The top three tRFs accounted for 95% of all tRF reads in mouse epididymal EVs.
Figure 5.8 tRFs in mouse epididymal EVs
(a) Top six most abundant tRFs in mouse epididymal EVs. (b) Read length distribution of tRFs in mouse epididymal EVs. n=3 (pools).

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
144
5.2.4 Effects of dietary methyl donor supplementation on epididymal EV
production and cargo
5.2.4.1 Dietary methyl donor supplementation on epididymal EV production
To investigate the impact of excess dietary methyl donors on epididymal EV
production, I used Avy/a mice from an existing mouse study. This study showed that
methyl donor diet supplementation results in the vertical transmission of paternal
epigenetic state (19). I used two groups of Avy/a mice, those fed a normal chow diet and
those fed an MDS diet for over 14 generations. Epididymides were dissected from
mice, epididymal fluid was pooled into age-matched pools of five mice (Table 8.1) and
epididymal EVs were purified as previously described. I performed NTA to detect any
effect of diet on EV size or concentration. EVs from control and MDS mice were of
similar size by NTA, with a main population of ~150 nm EVs and a secondary
population of 200-300 nm EVs (Figure 5.9). More EVs were part of the secondary
population in control compared to MDS mice.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
145
Figure 5.9 Physical properties of epididymal EVs from control and MDS mice
This graph shows the concentration and size of EVs (averaged for each pool). Shading represents SEM. n=3 and 4 for control and MDS EVs respectively.
5.2.4.2 Effect of dietary methyl donor supplementation on epididymal EV small
RNA profiles
RNA was extracted from pools of epididymal EVs from control and MDS mice. Small
RNA libraries were constructed, sequenced and processed as previously described. In
epididymal EVs from both control and MDS mice, tRFs were the most common type of
small RNA read, followed by unannotated and then miRNA reads as was observed
with C57BL/6 control mice (Figure 5.10a). Both groups had very similar small RNA
profiles, indicating that diet had little effect on biotype proportions. The length
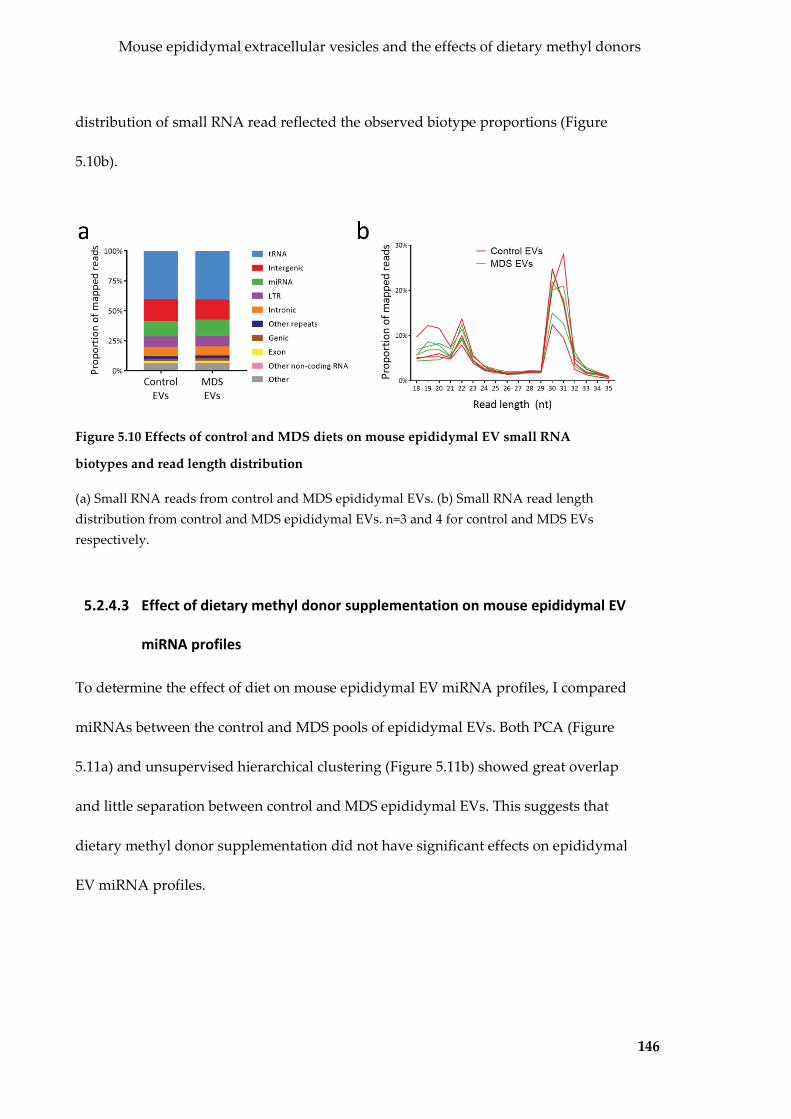
Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
146
distribution of small RNA read reflected the observed biotype proportions (Figure
5.10b).
Figure 5.10 Effects of control and MDS diets on mouse epididymal EV small RNA
biotypes and read length distribution
(a) Small RNA reads from control and MDS epididymal EVs. (b) Small RNA read length distribution from control and MDS epididymal EVs. n=3 and 4 for control and MDS EVs respectively.
5.2.4.3 Effect of dietary methyl donor supplementation on mouse epididymal EV
miRNA profiles
To determine the effect of diet on mouse epididymal EV miRNA profiles, I compared
miRNAs between the control and MDS pools of epididymal EVs. Both PCA (Figure
5.11a) and unsupervised hierarchical clustering (Figure 5.11b) showed great overlap
and little separation between control and MDS epididymal EVs. This suggests that
dietary methyl donor supplementation did not have significant effects on epididymal
EV miRNA profiles.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
147
Figure 5.11 Effect of control and MDS diet on mouse epididymal EV miRNA profiles
(a) PCA on epididymal EV miRNA cargo from control and MDS mice. (b) Unsupervised hierarchical clustering on epididymal EV miRNAs from control and MDS mice. n=3 and 4 for control and MDS EVs respectively.
5.2.4.4 Diet does not affect epididymal EV miRNA profiles
To further investigate the effects of diet on mouse epididymal EV miRNA cargo, I
compared individual epididymal EV miRNAs from control and MDS mice (for full
list see Table 8.9). The top 10 epididymal EV miRNAs from control and MDS mice
both accounted for 73% of all miRNA reads (Table 5.3). Many of the top 10 miRNAs
were shared between the two groups. All miRNAs but one were detected within a
2-fold change in relative abundance (Figure 5.12a). miR-1a-3p was two times more
abundant in MDS epididymal EVs than control EVs, however this did not reach
statistical significance.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
148
Table 5.3 Top 10 most abundant miRNAs in epididymal EVs from control and MDS mice
miRNA Control EVs MDS EVs
Normalised counta Rank Normalised counta Rank miR-148a-3p 209,431 1 160,525 2
let-7f-5p 156,258 2 200,788 1 miR-10a-5p 59,453 3 54,127 5
let-7g-5p 59,002 4 63,376 3 miR-21a-5p 53,476 5 55,761 4 miR-10b-5p 51,330 6 51,294 6
let-7c-5p 50,684 7 49,207 7 miR-143-3p 31,009 8 38,054 8 miR-200a-3p 30,266 9 25,837 10
let-7a-5p 25,479 10 20,722 11 let-7i-5p 23,298 11 27,944 9
a Per million miRNA reads
Figure 5.12 Effect of dietary methyl donor supplementation on mouse epididymal EV
miRNA
Scatterplot showing the effect dietary methyl donor supplementation on mouse epididymal EV miRNAs. Grey miRNAs were excluded by abundance filter (<100 counts per million miRNA reads). Dotted lines represent the threshold for 2-fold change in abundance.
Taken together, these data suggest that dietary methyl donor supplementation has no
detectable effect on epididymal EV miRNA.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
149
5.2.4.5 Effect of dietary methyl donor supplementation on epididymal EV tRF
profiles
tRNA-derived reads were the most abundant read biotype in control and MDS
epididymal EV small RNA libraries (Figure 5.10a). These tRNA-derived reads were 32
nt tRNA in length (Figure 5.13a) and mapped to the 5’ and 3’ ends of tRNA genes;
consistent with tRFs and referred to as such. Like C57BL/6 mice, the same six tRFs
comprised 97% and 98% of all tRF reads (and 39% of all mapped reads) in control and
methyl mice respectively (Figure 5.13b).
Figure 5.13 tRFs in epididymal EVs isolated from mice fed a control MDS diet
(a) tRF read length distribution from control and MDS epididymal EVs. (b) The six most abundant tRFs in control and MDS epididymal EVs. n=3 and 4 for control and MDS EVs respectively.
To determine whether dietary methyl donor supplementation affected epididymal EV
tRF profiles, I compared tRF counts in control and MDS epididymal EVs (for full list
see Table 8.10). First, I performed PCA and unsupervised hierarchical clustering on the
top six most abundant tRFs. PCA (Figure 5.14a) and unsupervised hierarchical
clustering (Figure 5.14b) failed to separate samples based on diet. No significant
differences in abundance were detected in the most abundant tRFs (Figure 5.15).

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
150
Figure 5.14 Effect of genotype and diet on mouse epididymal EV tRF profiles
(a) PCA on the top six most tRFs in control and MDS epididymal EVs. (b) Unsupervised hierarchical clustering on the top six tRFs in control and MDS epididymal EVs. n=3 and 4 for control and MDS EVs respectively.
Figure 5.15 Abundance of top tRFs in control and MDS epididymal EVs
(a) tRF5-Gly-GCC, (b) tRF5-Gly-GCC, (c) tRF5-Glu-CTC, (d) tRF5-Val-CAC, (e) tRF5-His-GTG, (f) tRF5-Glu-TTC. Error bars represent SEM. n=3 and 4 for control and MDS EVs respectively.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
151
Taken together, both PCA, unsupervised hierarchical clustering and individual tRF
counts suggest that dietary methyl donor supplementation has no detectable effect on
mouse epididymal EV tRF profiles.
5.2.5 Mouse epididymal EVs contain a unique processed rRNA transcript
5.2.5.1 Small RNA libraries contain a higher than expected proportion of
unmappable reads
I observed an unusually low percentage of mappable reads in my three groups of
mouse epididymal EV small RNA libraries (C57BL/6 optimisation set, and control and
MDS groups). While unmappable reads were discarded during processing prior
analysis (as usually occurs during small RNA sequencing processing workflows), I
discovered that only 45-65% of my sequences mapped to the mouse genome (Figure
5.16a), compared to the usual >80%. To determine whether this was a biological or
technical, I downloaded two published publicly available small RNA sequencing
datasets for epididymal epithelial cells (the cells that produce the EVs), whole
epididymal tissue (both GSE70197) (230), and mouse epididymal EVs (GSE79500) (49)
and processed them through my pipeline. In this publicly available data, I observed
similarly low mapping percentages in mouse epididymal EVs (Figure 5.16b) and the
epididymal epithelial cells that produce them (Figure 5.16c), but high mapping
proportions in whole epididymis tissue (Figure 5.16d). This suggests that these
unmappable sequences are characteristic of epididymal EVs and their epididymal
epithelial parent cells and most likely have a biological basis.
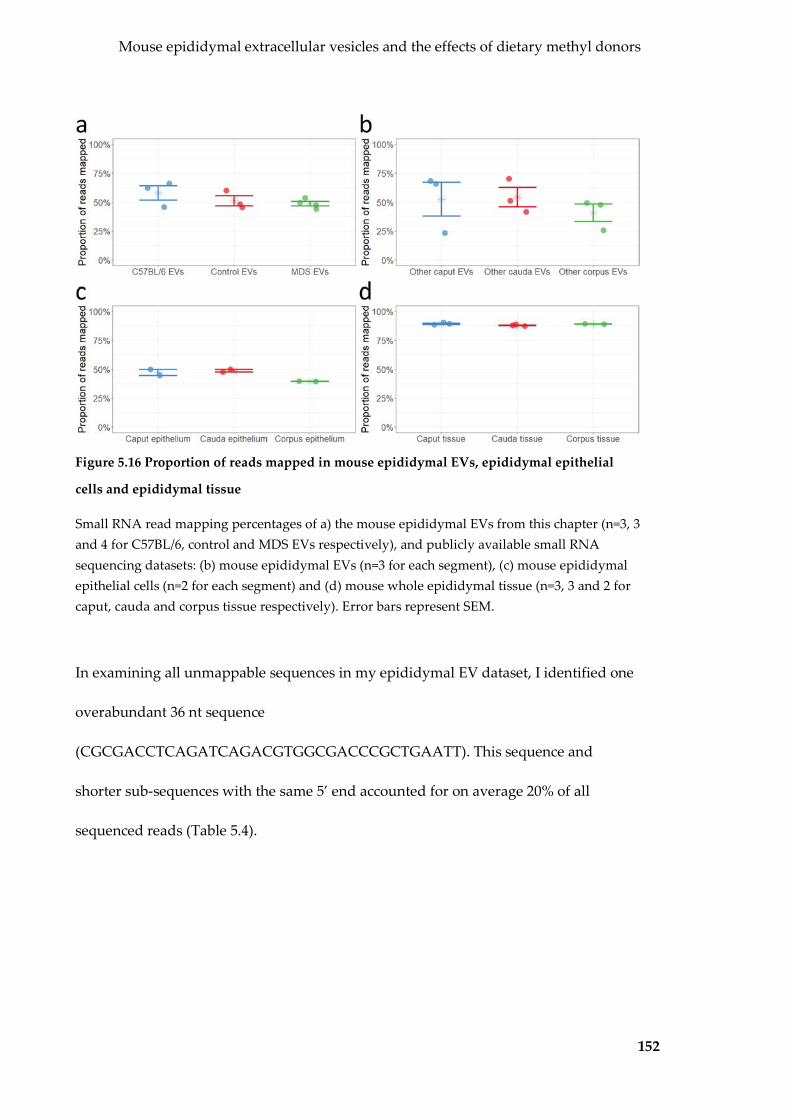
Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
152
Figure 5.16 Proportion of reads mapped in mouse epididymal EVs, epididymal epithelial
cells and epididymal tissue
Small RNA read mapping percentages of a) the mouse epididymal EVs from this chapter (n=3, 3 and 4 for C57BL/6, control and MDS EVs respectively), and publicly available small RNA sequencing datasets: (b) mouse epididymal EVs (n=3 for each segment), (c) mouse epididymal epithelial cells (n=2 for each segment) and (d) mouse whole epididymal tissue (n=3, 3 and 2 for caput, cauda and corpus tissue respectively). Error bars represent SEM.
In examining all unmappable sequences in my epididymal EV dataset, I identified one
overabundant 36 nt sequence
(CGCGACCTCAGATCAGACGTGGCGACCCGCTGAATT). This sequence and
shorter sub-sequences with the same 5’ end accounted for on average 20% of all
sequenced reads (Table 5.4).

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
153
Table 5.4 Abundant 36 nt unmappable sequence in mouse epididymal EVs
Sequence Length Normalised counta
CGCGACCTCAGATCAGAC 18 5,962 CGCGACCTCAGATCAGACG 19 184,225 CGCGACCTCAGATCAGACGT 20 49,932 CGCGACCTCAGATCAGACGTG 21 21,042 CGCGACCTCAGATCAGACGTGG 22 36,848 CGCGACCTCAGATCAGACGTGGC 23 16,479 CGCGACCTCAGATCAGACGTGGCG 24 533 CGCGACCTCAGATCAGACGTGGCGA 25 190 CGCGACCTCAGATCAGACGTGGCGAC 26 410 CGCGACCTCAGATCAGACGTGGCGACC 27 1,532 CGCGACCTCAGATCAGACGTGGCGACCC 28 5,585 CGCGACCTCAGATCAGACGTGGCGACCCG 29 10,141 CGCGACCTCAGATCAGACGTGGCGACCCGC 30 6,195 CGCGACCTCAGATCAGACGTGGCGACCCGCT 31 4,440 CGCGACCTCAGATCAGACGTGGCGACCCGCTG 32 1,796 CGCGACCTCAGATCAGACGTGGCGACCCGCTGA 33 436 CGCGACCTCAGATCAGACGTGGCGACCCGCTGAA 34 608 CGCGACCTCAGATCAGACGTGGCGACCCGCTGAAT 35 2,845 CGCGACCTCAGATCAGACGTGGCGACCCGCTGAATT 36 3,458 Total 18-36 352,657 a Per million mapped reads, averaged for all C57BL/6, control and MDS epididymal EV pools.
I then used BLAST to identify 11 perfect full-length matches for the 36 nt contig to
mouse ribosomal RNA genes (Table 8.2). This same sequence had been previously
identified in mature sperm and named rsRNA-28S (231). rsRNA-28S is enriched in
mature sperm and the cauda epididymis, compared to testicular sperm and earlier
segments of the epididymis (231). I did not detect any differences in abundance
between control and MDS epididymal EVs (Figure 5.17).

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
154
Figure 5.17 Abundance of rsRNA-28S in control and MDS epididymal EVs
Counts were normalised to per million unmapped reads. Error bars represent SEM. n=3 and 4 for control and MDS EVs.
Through searching for differences in small RNA between control and MDS epididymal
EVs, I found that rsRNA-28S is at relatively high abundance in epididymal EVs. My
findings implicate epididymal EVs as a potential source for rsRNA-28S in mature
sperm.
5.3 Discussion
In this chapter, I investigated whether dietary methyl donor supplementation
influences mouse epididymal RV small RNA profiles. First, I optimised the isolation of
EVs from the epididymal fluid of C57BL/6 mice. Purified EVs were validated by TEM,

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
155
NTA and protein mass spectrometry. I identified the miRNA and tRF cargo of mouse
epididymal EVs, and compared them to their in vitro counterparts, DC2 EVs. Next, I
ensured that the purified mouse epididymal EVs were able to bind with freshly
isolated spermatozoa – that the transfer of cargo was possible. Then, I identified that
dietary methyl donor supplementation have no detectable effect on epididymal EV
miRNA and tRF profiles. Inadvertently, with the help of several publicly available
datasets, I discovered that epididymal EVs contain rsRNA-28S, a rRNA gene transcript
that is unmappable by traditional methods. rsRNA-28S was previously found in
mature sperm, and my data implicates epididymal EVs as the likely source.
5.3.1 Isolation of mouse epididymal EVs
All seminal work on epididymal EVs was performed with EVs isolated from
epididymal fluid. In comparison to conditioned culture media, biological fluids such as
epididymal fluid are usually more complex and thus require more extensive isolation
protocols to purify EVs. My own protocols of filtration and differential centrifugation
was sufficient to isolate epididymal EVs, as evidenced by the expected size and shape
(49), protein composition (152,153), and interactions with spermatozoa (92,227,228).
Contamination by cells, cellular debris or sperm cytoplasmic droplets was able to be
excluded.
Mouse epididymal EVs shared many similarities with DC2 epididymal EVs. A large
overlap of 109 proteins were observed between the two types of epididymal EVs,
consistent with their similar producing cell type. These 109 commonly detected

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
156
proteins represented the majority of proteins identified in DC2 epididymal EVs but
only 13% of the proteins detected in mouse epididymal EVs – this may be due to the
five-fold difference in the number of proteins identified. The difference in the number
of proteins identified could be due to an increased complexity in mouse epididymal EV
proteins, however this seems unlikely given the great difference in identified proteins
between DC2 and mouse epididymal EVs. While care was taken to use the same
amount of EV protein for mass spectrometry for both experiments, I am unable rule
out any batch-specific effects.
5.3.2 Characterisation of mouse epididymal EV small RNA cargo
Mouse epididymal EVs contained many miRNAs previously identified in epididymal
EVs (49,232). Many are of these miRNAs are also abundant in sperm (46), consistent
with the transfer of small RNAs to developing sperm in the epididymis (49). Many of
the most abundant miRNAs were also amongst the most abundant in DC2 epididymal
EVs, which should be expected considering they are produced from the same parent
cell type. However, there were three miRNAs that were not expressed to the same
relative abundance in in vitro EVs: miR-148a-3p, miR-200a-3p and miR-143-3p. These
three miRNAs are known to be amongst the most abundant in in vivo epididymal EVs
and sperm (46,49,232), and thus their absence from DC2 epididymal EVs is surprising.
While the literature on in vitro epididymal EVs is very limited (there is one paper that
details targeted, qualitative analyses on specific RNAs in DC2 epididymal EVs (205)),
these differences in abundance may be due to physiological differences between the in
situ and cell line production. These three miRNAs all have similar anti-proliferative

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
157
roles. miR-148a-3p and miR-143-3p are oncomiRs implicated with tumour-suppression
(199,233). miR-200a-3p on the other hand functions to regulate epididymal cell
proliferation in epididymal tissue in adulthood (234). DC2 epididymal cells were
immortalised from transgenic mice over-expressing the SV-40 large T antigen, a viral
oncogene (148). Additionally, tissue culture cells are always kept in a proliferative state
of exponential growth, while more regulation is required for cells in situ.
Consequently, one might expect miRNAs with anti-proliferative function to be
downregulated, possibly to below the limit of detection as seen with miR-200a-3p.
Compared to miRNAs, the epididymal EV tRF population was less diverse, with tRFs
from three anticodons accounting for 95% of all tRF reads. These same tRFs have been
previously detected in epididymal EVs (DC2 epididymal EVs and others (24,34).
Anticodon representation does not correlate with the number of genomic loci (160),
and the basis for their high abundance is presently unknown.
5.3.3 Dietary methyl donor supplementation has no detectable effect on
epididymal EV cargo
Given the results of the previous chapter on the effect of methyl donors on EVs in vitro,
it was surprisingly to find that dietary methyl donor supplementation had no
detectable effect on mouse epididymal EV small RNA cargo. Small RNA biotype
proportions were as expected (24,34), and were virtually identical between control and
MDS epididymal EVs. Within miRNA and tRF populations, pools of control and MDS

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
158
EV were indistinguishable by PCA and unsupervised hierarchical clustering. This was
reflected by the extremely similar abundances of individual miRNAs and tRFs.
The observed phenotypic differences between control and MDS mice suggests that
some signal caused be dietary methyl donor supplementation is passed through the
germline (19), leading to a cumulative, yet reversible effect (25). This notion has
precedence – both dietary protein restriction (24) and chronic ethanol exposure (34)
both changes to epididymal EV small RNA cargo, which go on to have an effect on
expression after fertilisation in the developing embryo.
While I did not detect any effect of diet on small RNA, it is possible that the model
system was not sensitive enough to detect changes. Due to time and animal constraints,
I had pooled together the whole epididymal fluid from five mice to form one pool,
from which RNA was extracted and small RNA libraries were made. It has been shown
however, that the small RNA profiles of epididymal EVs differ based on the segment
from which they are derived (e.g. caput is different to cauda) (49). The small RNA
changes in DC2 EVs caused by folate availability in vitro may be due to the caput origin
of DC2 cells (148). In other words, by pooling together EVs from the whole epididymis,
I may have obscured any differences present only in one segment. If so, this may
explain why I observed slight differences (though not statistically significant) in the
abundance of specific tRFs. It is worth nothing that the two studies detailing the
influences of diet on epididymal EV small RNA cargo specifically looked at cauda
epididymal EVs (24,34). In future experiments, pools should be made based on
epididymal segment.

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
159
Diet may be able to influence small RNAs in more ways than just abundance. Chen et.
al. (33) reported that a high fat diet altered tRFs post-translational modifications.
Artificially synthesised tRFs lacking the modifications did not have the same
phenotypic effects as isolated sperm tRFs, suggesting that these modifications play a
crucial role. These modifications are not distinguishable by the small RNA sequencing
approach I used. While tRF abundance may be similar, diet could influence the type
and extent of modification and thus remain undetected.
5.3.4 Mouse epididymal EVs contain a traditionally unmappable
ribosomal gene transcript enriched in mature sperm
The effect of dietary methyl donor supplementation aside, the data I generated led to
an unexpected discovery with epididymal EVs: the overabundance of a specific
sequence called rsRNA-28S (231). Despite being identified as rsRNA-28S, a rRNA-
derived protein transcript, this transcript was not able to be mapped to the mouse
genome. This is believed to be due to the presence of intergenic spacers (IGS) in
ribosomal genes (231). A large portion of IGS have not been sequenced or assembled,
thus resulting in ribosomal gene transcripts escaping genome mapping.
rsRNA-28S had been identified before in the epididymis and mature sperm, but not
testicular sperm, and thus was proposed that it was enriched in mature sperm (231).
This thesis is the first to show that rsRNA-28S is also present in epididymal EVs and
their producing cells. Considering that epididymal EVs transfer proteins and RNAs to
sperm (49,89-92,206), these findings imply that epididymal EVs also transfer rsRNA-

Mouse epididymal extracellular vesicles and the effects of dietary methyl donors
160
28S to sperm. The function of rsRNA-28S remains unknown. rsRNA-28S abundance
has been linked to inflammation and stress (231). I did not observe any differences
between control and MDS epididymal EVs, but again this may be confounded by the
study of the whole epididymis as opposed to individual segments. Interestingly, I did
not detect an abnormally high abundance of rsRNA-28S or unusual mapping
percentages in DC2 epididymal EVs and cells, however these differences may be in
vivo vs in vitro in nature, as observed and discussed with miR-148a-3p, miR-200a-3p,
and miR-143-3p.
This overabundant transcript accounts for a significant proportion of epididymal EV
small RNA libraries, however they have never been reported in many epididymal EV
sequencing studies (24,34,49,232), possibly discarded after failing to map to the
genome. I propose that rsRNA-28S should be examined in all epididymal EV studies,
especially in the context of effects of an environmental stress on small RNA cargo.
5.3.5 Conclusion
In this chapter, I characterised EVs isolated from the epididymal fluid of mice. These
EVs exhibited the expected physical characteristics of epididymal EVs. I showed that
dietary methyl donor supplementation had little detectable effect on epididymal EV
production and small RNA cargo overall. During my investigations, I also identified
rsRNA-28S in epididymal EVs, an abundant ribosomal gene transcript that is not
mappable to the genome. This finding implies that epididymal EVs may be the source
for rsRNA-28S in mature sperm.

161
6 CONCLUDING REMARKS AND FUTURE DIRECTION
In this thesis, I sought evidence to support the idea that soma-derived EVs could be
involved in the inheritance of environmentally-influenced traits. Small RNAs have
emerged as key molecules in such inheritance. Using three model systems – Sertoli
cells, epididymal cells, and epididymal fluid – I sought to determine whether EVs from
these somatic sources carried small RNA, and could interact with germ cells.
Importantly, I asked whether EV small RNA cargo could be influenced by
environmental factors. I was able to show that not only do soma-derived EVs contain
small RNA cargo that can be influenced by the environment, but these soma-derived
EVs can interact with germ cells.
In Chapter 3, I used the TM4 Sertoli cell line as a model system. Sertoli cells have an
essential role in spermatogenesis (29,55), and preliminary evidence exists that Sertoli
cells produce EVs – at least in the Chinese soft-shelled turtle (158). I present the first
report of isolated Sertoli EVs and have characterised their production and small RNA
cargo with respect to their parent cells. In Sertoli EVs I identified many miRNAs
known to regulate and help maintain spermatogonial stem cells (SSCs). I also showed
that Sertoli EVs bind and fuse with SSCs resulting in the transfer small RNA cargo.
Interestingly, I was not able to detect any changes in gene expression after such
interaction, however this is likely due time limitations and possible batch-specific
effects. Further experiments such as small RNA sequencing and gene expression
analysis on the same recipient cell sample would be able to confirm whether

Concluding remarks and future direction
162
transferred small RNAs regulate gene expression in SSCs. Regardless, my findings
suggest a new facet for how Sertoli cells facilitate spermatogenesis.
BPA is a known endocrine toxin that causes many deleterious reproductive effects (74),
some possibly due to its action on Sertoli cells (80-82). After exposure to BPA and the
vehicle DMSO, I detailed changes to Sertoli EV production and small RNA cargo.
Interestingly the abundance of many small RNAs were altered in EVs, while remaining
unchanged in the Sertoli cells themselves. The EV-specific effects of DMSO exposure
are extremely pertinent, especially since DMSO is widely used as a solvent for many
chemicals since it does not have significant effects on cellular small RNAs and
physiology. My findings suggest that while cells may be unaffected, the EVs they
produce may harbour many undetected but significant changes. While I was able
detect some BPA-specific effects to Sertoli EVs and their cargo by comparing them with
the direct DMSO control, these effects were dwarfed by that of DMSO alone (compared
to untreated media). Therefore, it is possible that DMSO masked some effects of BPA.
This is extremely important to recognise, as DMSO is commonly used as a solvent in
many EV studies (128,190,192,193). To further determine the effects of BPA alone on
Sertoli EVs and their small RNA cargo, a different vehicle solvent such as ethanol may
be better suited. Providing that the vehicle solvent does not have drastic EV-specific
effects like DMSO, then it may provide a better baseline for detecting the effects of BPA
alone.
Due to time limitations I was only able to perform any co-incubations experiments
between untreated Sertoli EVs and SSCs. I have shown that it is possible and likely that

Concluding remarks and future direction
163
environmentally-influenced small RNAs enter the germline, as several small RNAs
shown to be transferred from Sertoli EVs to SSCs are altered by BPA and DMSO
exposure. However, I have not been able to explicitly demonstrate this. Similar
co-incubation experiments but between SSCs and BPA- and DMSO-treated EVs would
be sufficient to rectify this. Regardless, my findings from this chapter show that Sertoli
EVs carry small RNAs influenced by the parent cell environment and can pass these
environmental signals onto SSCs and possibly the next generation.
In Chapter 4, I switched models to the DC2 epididymal cell line. Epididymal EVs have
a well-documented role in sperm maturation. They are responsible for transferring
many proteins to sperm to aid with motility and fertilisation capability (89-92). There is
also limited evidence that they transfer RNA (24,34,49). I purified and isolated
epididymal EVs from the DC2 epididymal cell line, and characterised their physical
properties, and cargo with respect to the cells themselves. I used folate concentration as
my model environmental stressor. I detected many miRNAs with an altered
abundance after a reduction of folate from original supraphysiological concentrations
in default to physiological concentrations.
Additionally, my findings suggest that a short-term exposure to excess folate is not
equivalent to a long-term exposure to excess folate. However, there remains a
possibility these short- vs long-term changes may represent selection caused by the
folate-weaning process. Further experiments would be able to separate the direct
effects of a change in folate concentration from those stemming from the folate-
weaning process. These include using the same low-folate population as baseline from

Concluding remarks and future direction
164
which sub-lines were prepared and exposed to excess folate for a range of timepoints.
By using the epididymal cells grown in physiological folate as a baseline, I would be
able to isolate the effects of the folate-weaning process, and using varying timepoints,
would better document how exposure to excess folate alters epididymal EV production
and small RNA cargo over time.
Due to low EV yields and time limitations, I had to pool together RNA from multiple
preparations of EVs. While this pooling provided adequate starting material for small
RNA library construction and meaningful analysis, it would be desirable to perform
this experiment with replicates. Nevertheless, my findings show that, like Sertoli EVs,
epididymal EVs carry small RNA influenced by the parent cell environment.
In Chapter 5, I ventured in vivo, into the mouse epididymis – a more complicated but
arguably more realistic model system. I purified mouse epididymal EVs from the
epididymal fluid of mice and characterised the physical properties and small RNA
cargo of these EVs. Through labelling experiments, I was able to confirm that
epididymal EVs interacted with live spermatozoa, setting the foundations for any
possible transfer of small RNAs from epididymal EVs to living sperm.
I used excess dietary methyl donor supplementation as my environmental stressor. I
purified epididymal EVs from two sub-strains of mice fed a methyl donor
supplemented and normal chow diet for over 14 generations. However, I could not
detect any differences in epididymal EV production and cargo as a result of dietary
methyl donor supplementation. This may be due to many reasons. While it is possible
that dietary methyl donor supplementation has transgenerational effects that are

Concluding remarks and future direction
165
independent on small RNAs transferred from epididymal EVs to sperm, the findings
from in vitro epididymal EVs suggest otherwise. A more realistic possibility could lie
within experimental design. Other studies have shown that diet influences the small
RNA profile of epididymal EVs (24,34). Instead of using EVs purified from the whole
epididymis, these studies split the epididymis up into different segments. Different
segments of the epididymis contain sperm and EVs with significantly different small
RNA cargo (46). Thus, it is possible that by collecting EVs from the whole epididymis,
changes within only a specific segment could not be detected. This could be easily
addressed by preparing pools of EVs from each segment, however this would also
require a greater number of experimental animals, which were not available for this
thesis.
Additionally, I discovered an interesting potential function of epididymal EVs. I
noticed that my epididymal EV small RNA libraries had a higher than expected level
of unmappable reads, primarily due to a particular rRNA-derived transcript called
rsRNA-28S. Using publicly available small RNA sequencing datasets, I was able to
show that this rRNA transcript was a property of epididymal EVs and epididymal
epithelial cells, but not whole epididymal tissue. This same rRNA-28S transcript was
previously found in mature sperm after epididymal transit (231). My findings suggest
that epididymal EVs are the likely source.
This thesis shows that soma-derived EV production and small RNA cargo is influenced
by the environment, along with some strong, but preliminary evidence suggesting that
these altered small RNAs can be transferred to germ cells. The next immediate steps

Concluding remarks and future direction
166
include preliminary experiments to confirm this, such as co-incubation experiments
between treated EVs and germ cells. Then the next steps lie in investigating what effect
any transferred environmentally-influenced small RNAs have in the germ cells,
following the overarching hypothesis (146). This would involve detecting any
transferred small RNAs at later stages of male germ cells (e.g. if small RNAs
transferred from Sertoli EVs to SSCs are retained after sperm maturation). I can also
look at cells after fertilisation, from the zygote, to the blastocyst to see if the transferred
small RNAs brought any changes to gene expression. Furthermore, I can look in the
grown individual itself to determine any effects on phenotype. Any effect can also be
isolated down to the transferred small RNA cargo as well, by isolating altered small
RNAs from germ cells exposed to EVs, and then injecting them directly into control
embryos.
6.1 Limitations of this thesis
The work presented here in this thesis forms a strong foundation for future studies on
the role of extracellular vesicles in soma-to-germline feedback and the inheritance of
environmentally-induced traits. In Chapters 3 and 4, my work involved somatic EVs
produced in tissue culture. While the use of an in vitro model suited the introductory
and exploratory analysis (the first reports of characterisation of Sertoli and DC2 EVs
and their encapsulated cargo), there remains the question whether these results can be
extrapolated to an in vivo or in situ setting. In the case of Sertoli cells, a more complex
model incorporating the tubular structure of the seminiferous tubules (eg that
described by Gassei et al (235)), the presence of germ cells during EV production as

Concluding remarks and future direction
167
well as the presence of Leydig cells (somatic cells that support spermatogenesis albeit
with a much lesser impact than Sertoli cells) may be more representative of in vivo
biology. It also may be possible to isolate Sertoli EVs in vivo (236), however this
technique remains untested and is plagued with many technical issues. Few studies
have been able to collect seminiferous tubule fluid via cannulation, however technical
issues remain such as a suitable EV isolation method as well as the number of
experimental animals required for substantial EV yield.
My results from Chapter 4 suggested that epididymal EV cargo is influenced by the
environment, specifically the concentration of folate and the level of exposure. In
Chapter 5, I attempted to take my investigations of the same cell type (epididymal
epithelial cells) into the mouse model by investigating the effects of dietary methyl
donor supplementation on epididymal EVs isolated from epididymal fluid. The results
from this in vivo experiment did not complement that of the previous chapter and the
reasons for this are not clear. While it is possible that the results achieved from the in
vitro model do not represent in vivo biology, further experimentation is required to
address some experimental and technical limitations. For example, due to the limited
number of experimental mice available, I isolated EVs from epididymal fluid from the
whole epididymis (as opposed to the three epididymal segments being treated
separation as performed in several other studies (24,33,34,49)), and as a result, RNA
changes in individual segments may have been masked and undetected.

Concluding remarks and future direction
168
6.2 Concluding remarks
In conclusion, the work in this thesis demonstrates that germline-associated somatic
cells release EVs that can interact and transfer RNAs to germ cells. EV production and
small RNA cargo are susceptible to the environment, and thus these EVs may act as a
vector for soma-to-germline feedback. The work performed in this thesis suggests that
environmentally-influenced small RNAs in somatic cells could be transported by EVs
to germ cells (Figure 6.1). This thesis sets the foundations for further studies into the
potential role of EVs in the inheritance of environmentally acquired traits.
Figure 6.1 Somatic EVs may act as vectors for environmentally-influenced small
RNAs to enter the germline and possibly the next generation
In this thesis, I have isolated EVs and their small RNA cargo from somatic cells of the male reproductive system. These EVs are in close contact with developing germ cells and maturing sperm, and this thesis shows that germ cells are able to take up these EVs and their small RNA cargo. Additionally, I show that EV cargo is influenced by the environment. Taken together, somatic EVs may be able to act as vectors for transgenerational inheritance via the transfer of small RNAs to male germ cells and the next generation.

169
7 REFERENCES
1. Weismann, A. (1893) The Germ-plasm: a theory of heredity. Translated by W. Newton Parker and Harriet Rönnfeldt, Scribner, New York
2. Avery, O. T., MacLeod, C. M., and McCarty, M. (1944) Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types : Induction of Transformation by a Desoxyribonucleic Acid Fraction Isolated from Pneumococcus Type III. The Journal of Experimental Medicine 79, 137-158
3. Hershey, A. D., and Chase, M. (1952) Independent functions of viral protein and nucleic acid in growth of bacteriophage. The Journal of General Physiology 36, 39-56
4. Crick, F. H. (1958) On protein synthesis. Symposia of the Society for Experimental Biology 12, 138-163
5. Crick, F. (1970) Central dogma of molecular biology. Nature 227, 561-563 6. Schmidt, A., Schmid, M. W., and Grossniklaus, U. (2015) Plant germline
formation: common concepts and developmental flexibility in sexual and asexual reproduction. Development 142, 229-241
7. Jablonka, E., and Raz, G. (2009) Transgenerational epigenetic inheritance: prevalence, mechanisms, and implications for the study of heredity and evolution. The Quarterly Review of Biology 84, 131-176
8. Extavour, C. G., and Akam, M. (2003) Mechanisms of germ cell specification across the metazoans: epigenesis and preformation. Development 130, 5869-5884
9. Rechavi, O., Minevich, G., and Hobert, O. (2011) Transgenerational inheritance of an acquired small RNA-based antiviral response in C. elegans. Cell 147, 1248-1256
10. Rechavi, O., Houri-Ze'evi, L., Anava, S., Goh, W. S. S., Kerk, S. Y., Hannon, G. J., and Hobert, O. (2014) Starvation-induced transgenerational inheritance of small RNAs in C. elegans. Cell 158, 277-287
11. Seong, K. H., Li, D., Shimizu, H., Nakamura, R., and Ishii, S. (2011) Inheritance of stress-induced, ATF-2-dependent epigenetic change. Cell 145, 1049-1061
12. Cropley, J. E., Eaton, S. A., Aiken, A., Young, P. E., Giannoulatou, E., Ho, J. W. K., Buckland, M. E., Keam, S. P., Hutvagner, G., Humphreys, D. T., Langley, K. G., Henstridge, D. C., Martin, D. I. K., Febbraio, M. A., and Suter, C. M. (2016) Male-lineage transmission of an acquired metabolic phenotype induced by grand-paternal obesity. Molecular Metabolism 5, 699-708
13. Li, C. C., Young, P. E., Maloney, C. A., Eaton, S. A., Cowley, M. J., Buckland, M. E., Preiss, T., Henstridge, D. C., Cooney, G. J., Febbraio, M. A., Martin, D. I., Cropley, J. E., and Suter, C. M. (2013) Maternal obesity and diabetes induces latent metabolic defects and widespread epigenetic changes in isogenic mice. Epigenetics 8, 602-611
14. Anway, M. D., Memon, M. A., Uzumcu, M., and Skinner, M. K. (2006) Transgenerational effect of the endocrine disruptor vinclozolin on male spermatogenesis. Journal of Andrology 27, 868-879
15. Dolinoy, D. C., Huang, D., and Jirtle, R. L. (2007) Maternal nutrient supplementation counteracts bisphenol A-induced DNA hypomethylation in

References
170
early development. Proceedings of the National Academy of Sciences of the United States of America 104, 13056-13061
16. Wolstenholme, J. T., Edwards, M., Shetty, S. R. J., Gatewood, J. D., Taylor, J. A., Rissman, E. F., and Connelly, J. J. (2012) Gestational Exposure to Bisphenol A Produces Transgenerational Changes in Behaviors and Gene Expression. Endocrinology 153, 3828-3838
17. Salian, S., Doshi, T., and Vanage, G. (2009) Impairment in protein expression profile of testicular steroid receptor coregulators in male rat offspring perinatally exposed to Bisphenol A. Life sciences 85, 11-18
18. Dolinoy, D. C., Weidman, J. R., Waterland, R. A., and Jirtle, R. L. (2006) Maternal genistein alters coat color and protects Avy mouse offspring from obesity by modifying the fetal epigenome. Environmental Health Perspectives 114, 567-572
19. Cropley, J. E., Suter, C. M., Beckman, K. B., and Martin, D. I. (2006) Germ-line epigenetic modification of the murine A vy allele by nutritional supplementation. Proceedings of the National Academy of Sciences of the United States of America 103, 17308-17312
20. Bygren, L. O., Kaati, G., and Edvinsson, S. (2001) Longevity determined by paternal ancestors' nutrition during their slow growth period. Acta Biotheoretica 49, 53-59
21. Dias, B. G., and Ressler, K. J. (2014) Parental olfactory experience influences behavior and neural structure in subsequent generations. Nature Neuroscience 17, 89-96
22. Rodgers, A. B., Morgan, C. P., Bronson, S. L., Revello, S., and Bale, T. L. (2013) Paternal stress exposure alters sperm microRNA content and reprograms offspring HPA stress axis regulation. The Journal of Neuroscience 33, 9003-9012
23. Crean, A. J., and Bonduriansky, R. (2014) What is a paternal effect? Trends in Ecology & Evolution 29, 554-559
24. Sharma, U., Conine, C. C., Shea, J. M., Boskovic, A., Derr, A. G., Bing, X. Y., Belleannee, C., Kucukural, A., Serra, R. W., Sun, F., Song, L., Carone, B. R., Ricci, E. P., Li, X. Z., Fauquier, L., Moore, M. J., Sullivan, R., Mello, C. C., Garber, M., and Rando, O. J. (2016) Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science 351, 391-396
25. Cropley, J. E., Dang, T. H., Martin, D. I., and Suter, C. M. (2012) The penetrance of an epigenetic trait in mice is progressively yet reversibly increased by selection and environment. Proceedings of the Royal Society B: Biological Sciences 279, 2347-2353
26. Okumura, K., Lee, I. P., and Dixon, R. L. (1975) Permeability of selected drugs and chemicals across the blood-testis barrier of the rat. Journal of Pharmacology and Experimental Therapeutics 194, 89-95
27. Turner, T. T., D'Addario, D. A., and Howards, S. S. (1980) [3H]3-O-methyl-D-glucose transport from blood into the lumina of the seminiferous and epididymal tubules in intact and vasectomized hamsters. Journal of Reproduction and Fertility 60, 285-289
28. Turner, T. T., Jones, C. E., Howards, S. S., Ewing, L. L., Zegeye, B., and Gunsalus, G. L. (1984) On the androgen microenvironment of maturing spermatozoa. Endocrinology 115, 1925-1932

References
171
29. Johnson, L., Thompson, D. L., Jr., and Varner, D. D. (2008) Role of Sertoli cell number and function on regulation of spermatogenesis. Animal Reproduction Science 105, 23-51
30. Shea, J. M., Serra, R. W., Carone, B. R., Shulha, H. P., Kucukural, A., Ziller, M. J., Vallaster, M. P., Gu, H., Tapper, A. R., Gardner, P. D., Meissner, A., Garber, M., and Rando, O. J. (2015) Genetic and Epigenetic Variation, but Not Diet, Shape the Sperm Methylome. Developmental cell 35, 750-758
31. Bartel, D. P. (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281-297
32. Bartel, D. P. (2009) MicroRNAs: target recognition and regulatory functions. Cell 136, 215-233
33. Chen, Q., Yan, M., Cao, Z., Li, X., Zhang, Y., Shi, J., Feng, G. H., Peng, H., Zhang, X., Zhang, Y., Qian, J., Duan, E., Zhai, Q., and Zhou, Q. (2016) Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science 351, 397-400
34. Rompala, G. R., Mounier, A., Wolfe, C. M., Lin, Q., Lefterov, I., and Homanics, G. E. (2018) Heavy Chronic Intermittent Ethanol Exposure Alters Small Noncoding RNAs in Mouse Sperm and Epididymosomes. Frontiers in Genetics 9, 32
35. Keam, S. P., and Hutvagner, G. (2015) tRNA-Derived Fragments (tRFs): Emerging New Roles for an Ancient RNA in the Regulation of Gene Expression. Life 5, 1638-1651
36. Sobala, A., and Hutvagner, G. (2013) Small RNAs derived from the 5' end of tRNA can inhibit protein translation in human cells. RNA Biology 10, 553-563
37. Gebetsberger, J., Zywicki, M., Kunzi, A., and Polacek, N. (2012) tRNA-derived fragments target the ribosome and function as regulatory non-coding RNA in Haloferax volcanii. Archaea 2012, 260909
38. Couvillion, M. T., Bounova, G., Purdom, E., Speed, T. P., and Collins, K. (2012) A Tetrahymena Piwi bound to mature tRNA 3' fragments activates the exonuclease Xrn2 for RNA processing in the nucleus. Molecular Cell 48, 509-520
39. Goodarzi, H., Liu, X., Nguyen, H. C., Zhang, S., Fish, L., and Tavazoie, S. F. (2015) Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement. Cell 161, 790-802
40. Maute, R. L., Schneider, C., Sumazin, P., Holmes, A., Califano, A., Basso, K., and Dalla-Favera, R. (2013) tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proceedings of the National Academy of Sciences of the United States of America 110, 1404-1409
41. Ashe, A., Sapetschnig, A., Weick, E. M., Mitchell, J., Bagijn, M. P., Cording, A. C., Doebley, A. L., Goldstein, L. D., Lehrbach, N. J., Le Pen, J., Pintacuda, G., Sakaguchi, A., Sarkies, P., Ahmed, S., and Miska, E. A. (2012) piRNAs can trigger a multigenerational epigenetic memory in the germline of C. elegans. Cell 150, 88-99
42. Buckley, B. A., Burkhart, K. B., Gu, S. G., Spracklin, G., Kershner, A., Fritz, H., Kimble, J., Fire, A., and Kennedy, S. (2012) A nuclear Argonaute promotes multigenerational epigenetic inheritance and germline immortality. Nature 489, 447-451

References
172
43. Grentzinger, T., Armenise, C., Brun, C., Mugat, B., Serrano, V., Pelisson, A., and Chambeyron, S. (2012) piRNA-mediated transgenerational inheritance of an acquired trait. Genome Research 22, 1877-1888
44. Krawetz, S. A., Kruger, A., Lalancette, C., Tagett, R., Anton, E., Draghici, S., and Diamond, M. P. (2011) A survey of small RNAs in human sperm. Human Reproduction 26, 3401-3412
45. Jodar, M., Selvaraju, S., Sendler, E., Diamond, M. P., Krawetz, S. A., and Reproductive Medicine, N. (2013) The presence, role and clinical use of spermatozoal RNAs. Human Reproduction Update 19, 604-624
46. Nixon, B., Stanger, S. J., Mihalas, B. P., Reilly, J. N., Anderson, A. L., Tyagi, S., Holt, J. E., and McLaughlin, E. A. (2015) The microRNA signature of mouse spermatozoa is substantially modified during epididymal maturation. Biology of Reproduction 93, 91
47. Ostermeier, G. C., Miller, D., Huntriss, J. D., Diamond, M. P., and Krawetz, S. A. (2004) Reproductive biology: delivering spermatozoan RNA to the oocyte. Nature 429, 154
48. Grunewald, S., Paasch, U., Glander, H. J., and Anderegg, U. (2005) Mature human spermatozoa do not transcribe novel RNA. Andrologia 37, 69-71
49. Reilly, J. N., McLaughlin, E. A., Stanger, S. J., Anderson, A. L., Hutcheon, K., Church, K., Mihalas, B. P., Tyagi, S., Holt, J. E., Eamens, A. L., and Nixon, B. (2016) Characterisation of mouse epididymosomes reveals a complex profile of microRNAs and a potential mechanism for modification of the sperm epigenome. Scientific Reports 6, 31794
50. Schuster, A., Skinner, M. K., and Yan, W. (2016) Ancestral vinclozolin exposure alters the epigenetic transgenerational inheritance of sperm small noncoding RNAs. Environmental Epigenetics 2
51. Yoshida, S. (2016) From cyst to tubule: innovations in vertebrate spermatogenesis. Wiley interdisciplinary reviews. Developmental biology 5, 119-131
52. Hess, R. A., and França, L. R. (2005) Chapter 3 - Structure of the Sertoli Cell. in Sertoli Cell Biology, Academic Press, San Diego. pp 19-40
53. Mruk, D. D., and Cheng, C. Y. (2015) The Mammalian Blood-Testis Barrier: Its Biology and Regulation. Endocrine Reviews 36, 564-591
54. Smith, B. E., and Braun, R. E. (2012) Germ cell migration across Sertoli cell tight junctions. Science 338, 798-802
55. Mruk, D. D., and Cheng, C. Y. (2004) Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis. Endocrine Reviews 25, 747-806
56. Matsushita, K., Miyake, H., Chiba, K., and Fujisawa, M. (2016) Clusterin produced by Sertoli cells inhibits heat stress-induced apoptosis in the rat testis. Andrologia 48, 11-19
57. Meng, J., Holdcraft, R. W., Shima, J. E., Griswold, M. D., and Braun, R. E. (2005) Androgens regulate the permeability of the blood-testis barrier. Proceedings of the National Academy of Sciences of the United States of America 102, 16696-16700
58. Ramos-Trevino, J., Bassol-Mayagoitia, S., Ruiz-Flores, P., Espino-Silva, P. K., Saucedo-Cardenas, O., Villa-Cedillo, S. A., and Nava-Hernandez, M. P. (2017) In Vitro Evaluation of Damage by Heavy Metals in Tight and Gap Junctions of Sertoli Cells. DNA and Cell Biology 36, 829-836

References
173
59. Qiu, L., Zhang, X., Zhang, X., Zhang, Y., Gu, J., Chen, M., Zhang, Z., Wang, X., and Wang, S. L. (2013) Sertoli cell is a potential target for perfluorooctane sulfonate-induced reproductive dysfunction in male mice. Toxicological Sciences 135, 229-240
60. Dalsenter, P. R., Faqi, A. S., Webb, J., Merker, H. J., and Chahoud, I. (1996) Reproductive toxicity and tissue concentrations of lindane in adult male rats. Human & Experimental Toxicology 15, 406-410
61. Carwile, J. L., Luu, H. T., Bassett, L. S., Driscoll, D. A., Yuan, C., Chang, J. Y., Ye, X., Calafat, A. M., and Michels, K. B. (2009) Polycarbonate bottle use and urinary bisphenol A concentrations. Environmental Health Perspectives 117, 1368-1372
62. Olea, N., Pulgar, R., Perez, P., OleaSerrano, F., Rivas, A., NovilloFertrell, A., Pedraza, V., Soto, A. M., and Sonnenschein, C. (1996) Estrogenicity of resin-based composites and sealants used in dentistry. Environmental Health Perspectives 104, 298-305
63. Hormann, A. M., vom Saal, F. S., Nagel, S. C., Stahlhut, R. W., Moyer, C. L., Ellersieck, M. R., Welshons, W. V., Toutain, P. L., and Taylor, J. A. (2014) Holding Thermal Receipt Paper and Eating Food after Using Hand Sanitizer Results in High Serum Bioactive and Urine Total Levels of Bisphenol A (BPA). PLOS One 9, 12
64. Calafat, A. M., Ye, X., Wong, L. Y., Reidy, J. A., and Needham, L. L. (2008) Exposure of the U.S. population to bisphenol A and 4-tertiary-octylphenol: 2003-2004. Environmental Health Perspectives 116, 39-44
65. Genuis, S. J., Beesoon, S., Birkholz, D., and Lobo, R. A. (2012) Human excretion of bisphenol A: blood, urine, and sweat (BUS) study. Journal of Environmental and Public Health 2012, 185731
66. Lakind, J. S., and Naiman, D. Q. (2011) Daily intake of bisphenol A and potential sources of exposure: 2005-2006 National Health and Nutrition Examination Survey. Journal of Exposure Science and Environmental Epidemiology 21, 272-279
67. Matsumoto, A., Kunugita, N., Kitagawa, K., Isse, T., Oyama, T., Foureman, G. L., Morita, M., and Kawamoto, T. (2003) Bisphenol A levels in human urine. Environmental Health Perspectives 111, 101-104
68. Corrales, J., Kristofco, L. A., Steele, W. B., Yates, B. S., Breed, C. S., Williams, E. S., and Brooks, B. W. (2015) Global Assessment of Bisphenol A in the Environment: Review and Analysis of Its Occurrence and Bioaccumulation. Dose Response 13, 1559325815598308
69. Machtinger, R., Combelles, C. M., Missmer, S. A., Correia, K. F., Williams, P., Hauser, R., and Racowsky, C. (2013) Bisphenol-A and human oocyte maturation in vitro. Human Reproduction 28, 2735-2745
70. Hunt, P. A., Koehler, K. E., Susiarjo, M., Hodges, C. A., Ilagan, A., Voigt, R. C., Thomas, S., Thomas, B. F., and Hassold, T. J. (2003) Bisphenol a exposure causes meiotic aneuploidy in the female mouse. Current Biology 13, 546-553
71. Howdeshell, K. L., Hotchkiss, A. K., Thayer, K. A., Vandenbergh, J. G., and vom Saal, F. S. (1999) Exposure to bisphenol A advances puberty. Nature 401, 763-764
72. Honma, S., Suzuki, A., Buchanan, D. L., Katsu, Y., Watanabe, H., and Iguchi, T. (2002) Low dose effect of in utero exposure to bisphenol A and diethylstilbestrol on female mouse reproduction. Reproductive Toxicology 16, 117-122

References
174
73. Souter, I., Smith, K. W., Dimitriadis, I., Ehrlich, S., Williams, P. L., Calafat, A. M., and Hauser, R. (2013) The association of bisphenol-A urinary concentrations with antral follicle counts and other measures of ovarian reserve in women undergoing infertility treatments. Reproductive Toxicology 42, 224-231
74. Manikkam, M., Tracey, R., Guerrero-Bosagna, C., and Skinner, M. K. (2013) Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLOS One 8, e55387
75. Li, D. K., Zhou, Z., Miao, M., He, Y., Wang, J., Ferber, J., Herrinton, L. J., Gao, E., and Yuan, W. (2011) Urine bisphenol-A (BPA) level in relation to semen quality. Fertility and Sterility 95, 625-630 e621-624
76. Lassen, T. H., Frederiksen, H., Jensen, T. K., Petersen, J. H., Joensen, U. N., Main, K. M., Skakkebaek, N. E., Juul, A., Jorgensen, N., and Andersson, A. M. (2014) Urinary bisphenol A levels in young men: association with reproductive hormones and semen quality. Environmental Health Perspectives 122, 478-484
77. Meeker, J. D., Yang, T., Ye, X., Calafat, A. M., and Hauser, R. (2011) Urinary concentrations of parabens and serum hormone levels, semen quality parameters, and sperm DNA damage. Environmental Health Perspectives 119, 252-257
78. Lombo, M., Fernandez-Diez, C., Gonzalez-Rojo, S., Navarro, C., Robles, V., and Herraez, M. P. (2015) Transgenerational inheritance of heart disorders caused by paternal bisphenol A exposure. Environmental Pollution 206, 667-678
79. Nadal, A., Ropero, A. B., Laribi, O., Maillet, M., Fuentes, E., and Soria, B. (2000) Nongenomic actions of estrogens and xenoestrogens by binding at a plasma membrane receptor unrelated to estrogen receptor alpha and estrogen receptor beta. Proceedings of the National Academy of Sciences of the United States of America 97, 11603-11608
80. Cho, H., Kim, S., Park, H.-W., Oh, M.-J., Yu, S., Lee, S., Park, C., Han, J., Oh, J.-H., Hwang, S., and Yoon, S.-J. (2010) A relationship between miRNA and gene expression in the mouse Sertoli cell line after exposure to bisphenol A. BioChip Journal 4, 75-81
81. Lee, D. Y., Lee, S. S., Joo, W. A., Lee, E. J., and Kim, C. W. (2004) Analysis of differentially regulated proteins in TM4 cells treated with bisphenol A. Bioscience, Biotechnology, and Biochemistry 68, 1201-1208
82. Tabuchi, Y., Takasaki, I., and Kondo, T. (2006) Identification of genetic networks involved in the cell injury accompanying endoplasmic reticulum stress induced by bisphenol A in testicular Sertoli cells. Biochemical and Biophysical Research Communications 345, 1044-1050
83. Simerly, C., Castro, C., Hartnett, C., Lin, C. C., Sukhwani, M., Orwig, K., and Schatten, G. (2016) Post-Testicular Sperm Maturation: Centriole Pairs, Found in Upper Epididymis, are Destroyed Prior to Sperm's Release at Ejaculation. Scientific Reports 6, 31816
84. De Grava Kempinas, W., and Klinefelter, G. R. (2014) Interpreting histopathology in the epididymis. Spermatogenesis 4, e979114
85. Turner, T. T. (1984) Resorption versus secretion in the rat epididymis. Journal of Reproduction and Fertility 72, 509-514
86. Cornwall, G. A., and von Horsten, H. H. (2007) Sperm Maturation in the Epididymis. in The Genetics of Male Infertility (Carrell, D. T. ed.), Humana Press, Totowa, NJ. pp 211-231

References
175
87. Turner, T. T., Bomgardner, D., Jacobs, J. P., and Nguyen, Q. A. (2003) Association of segmentation of the epididymal interstitium with segmented tubule function in rats and mice. Reproduction 125, 871-878
88. Rejraji, H., Sion, B., Prensier, G., Carreras, M., Motta, C., Frenoux, J. M., Vericel, E., Grizard, G., Vernet, P., and Drevet, J. R. (2006) Lipid remodeling of murine epididymosomes and spermatozoa during epididymal maturation. Biology of Reproduction 74, 1104-1113
89. Skerget, S., Rosenow, M. A., Petritis, K., and Karr, T. L. (2015) Sperm Proteome Maturation in the Mouse Epididymis. PLOS One 10, e0140650
90. Frenette, G., and Sullivan, R. (2001) Prostasome-like particles are involved in the transfer of P25b from the bovine epididymal fluid to the sperm surface. Molecular Reproduction and Development 59, 115-121
91. Malm, J., Birn, H., Frohm, B., Hansen, S. I., Hoier-Madsen, M., and Holm, J. (2005) A minor fraction of a high-affinity folate binding protein from the epididymis is associated with membranous vesicles and spermatozoa in human semen. Int J Androl 28, 267-274
92. Griffiths, G. S., Galileo, D. S., Reese, K., and Martin-Deleon, P. A. (2008) Investigating the role of murine epididymosomes and uterosomes in GPI-linked protein transfer to sperm using SPAM1 as a model. Molecular Reproduction and Development 75, 1627-1636
93. Hess, R. A. (1998) Effects of environmental toxicants on the efferent ducts, epididymis and fertility. Journal of reproduction and fertility. Supplement 53, 247-259
94. Garcia-Diaz, E. C., Gomez-Quiroz, L. E., Arenas-Rios, E., Aragon-Martinez, A., Ibarra-Arias, J. A., and del Socorro, I. R.-M. M. (2015) Oxidative status in testis and epididymal sperm parameters after acute and chronic stress by cold-water immersion in the adult rat. Systems biology in reproductive medicine 61, 150-160
95. Simpson, R. J., and Mathivanan, S. (2012) Extracellular Microvesicles: the need for internationally recognised nomenclature and stringer purification criteria. J Proteomics Bioinform 5, ii
96. Greco, V., Hannus, M., and Eaton, S. (2001) Argosomes: a potential vehicle for the spread of morphogens through epithelia. Cell 106, 633-645
97. Hess, C., Sadallah, S., Hefti, A., Landmann, R., and Schifferli, J. A. (1999) Ectosomes released by human neutrophils are specialized functional units. Journal of immunology 163, 4564-4573
98. Valadi, H., Ekström, K., Bossios, A., Sjöstrand, M., Lee, J. J., and Lötvall, J. O. (2007) Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature cell biology 9, 654–659
99. Yuana, Y., Bertina, R. M., and Osanto, S. (2011) Pre-analytical and analytical issues in the analysis of blood microparticles. Thrombosis and haemostasis 105, 396-408
100. Li, C. C., Eaton, S. A., Young, P. E., Lee, M., Shuttleworth, R., Humphreys, D. T., Grau, G. E., Combes, V., Bebawy, M., Gong, J., Brammah, S., Buckland, M. E., and Suter, C. M. (2013) Glioma microvesicles carry selectively packaged coding and noncoding RNAs which alter gene expression in recipient cells. RNA Biology 10, 44–55
101. Al-Nedawi, K., Meehan, B., Micallef, J., Lhotak, V., May, L., Guha, A., and Rak, J. (2008) Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nature cell biology 10, 619-624

References
176
102. Chargaff, E., and West, R. (1946) The biological significance of the thromboplastic protein of blood. The Journal of biological chemistry 166, 189-197
103. Wolf, P. (1967) The nature and significance of platelet products in human plasma. British journal of haematology 13, 269-288
104. Bevers, E. M., Comfurius, P., Dekkers, D. W., and Zwaal, R. F. (1999) Lipid translocation across the plasma membrane of mammalian cells. Biochimica et biophysica acta 1439, 317-330
105. Zwaal, R. F. A., Comfurius, P., and Bevers, E. M. (2005) Surface exposure of phosphatidylserine in pathological cells. Cellular and Molecular Life Sciences 62, 971-988
106. Gyorgy, B., Szabo, T. G., Pasztoi, M., Pal, Z., Misjak, P., Aradi, B., Laszlo, V., Pallinger, E., Pap, E., Kittel, A., Nagy, G., Falus, A., and Buzas, E. I. (2011) Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cellular and Molecular Life Sciences 68, 2667-2688
107. Hristov, M., Erl, W., Linder, S., and Weber, P. C. (2004) Apoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood 104, 2761-2766
108. Trams, E. G., Lauter, C. J., Salem, N., Jr., and Heine, U. (1981) Exfoliation of membrane ecto-enzymes in the form of micro-vesicles. Biochimica et biophysica acta 645, 63-70
109. Hurley, J. H. (2008) ESCRT complexes and the biogenesis of multivesicular bodies. Current opinion in cell biology 20, 4-11
110. Trajkovic, K., Hsu, C., Chiantia, S., Rajendran, L., Wenzel, D., Wieland, F., Schwille, P., Brugger, B., and Simons, M. (2008) Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 319, 1244-1247
111. Gruenberg, J., and Stenmark, H. (2004) The biogenesis of multivesicular endosomes. Nature Reviews Molecular Cell biology 5, 317-323
112. Skinner, A. M., O'Neill, S. L., and Kurre, P. (2009) Cellular microvesicle pathways can be targeted to transfer genetic information between non-immune cells. PLOS One 4, e6219
113. Baj-Krzyworzeka, M., Szatanek, R., Weglarczyk, K., Baran, J., Urbanowicz, B., Branski, P., Ratajczak, M. Z., and Zembala, M. (2006) Tumour-derived microvesicles carry several surface determinants and mRNA of tumour cells and transfer some of these determinants to monocytes. Cancer Immunology, Immunotherapy 55, 808-818
114. Skog, J., Würdinger, T., van Rijn, S., Meijer, D. H., Gainche, L., Sena-Esteves, M., Curry, W. T., Jr., Carter, B. S., Krichevsky, A. M., and Breakefield, X. O. (2008) Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nature cell biology 10, 1470–1476
115. Waldenström, A., Genneback, N., Hellman, U., and Ronquist, G. (2012) Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLOS One 7, e34653
116. Turiak, L., Misjak, P., Szabo, T. G., Aradi, B., Paloczi, K., Ozohanics, O., Drahos, L., Kittel, A., Falus, A., Buzas, E. I., and Vekey, K. (2011) Proteomic characterization of thymocyte-derived microvesicles and apoptotic bodies in BALB/c mice. Journal of Proteomics 74, 2025-2033

References
177
117. Guescini, M., Genedani, S., Stocchi, V., and Agnati, L. F. (2010) Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. Journal of Neural Transmission 117, 1-4
118. Ronquist, K. G., Ronquist, G., Carlsson, L., and Larsson, A. (2009) Human prostasomes contain chromosomal DNA. The Prostate 69, 737-743
119. Collino, F., Deregibus, M. C., Bruno, S., Sterpone, L., Aghemo, G., Viltono, L., Tetta, C., and Camussi, G. (2010) Microvesicles derived from adult human bone marrow and tissue specific mesenchymal stem cells shuttle selected pattern of miRNAs. PLOS One 5, e11803
120. Deregibus, M. C., Cantaluppi, V., Calogero, R., Lo Iacono, M., Tetta, C., Biancone, L., Bruno, S., Bussolati, B., and Camussi, G. (2007) Endothelial progenitor cell derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood 110, 2440–2448
121. Ratajczak, J., Miekus, K., Kucia, M., Zhang, J., Reca, R., Dvorak, P., and Ratajczak, M. Z. (2006) Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia 20, 847-856
122. Renzulli, J. F., 2nd, Del Tatto, M., Dooner, G., Aliotta, J., Goldstein, L., Dooner, M., Colvin, G., Chatterjee, D., and Quesenberry, P. (2010) Microvesicle induction of prostate specific gene expression in normal human bone marrow cells. The Journal of urology 184, 2165-2171
123. Valadi, H., Ekstrom, K., Bossios, A., Sjostrand, M., Lee, J. J., and Lotvall, J. O. (2007) Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature cell biology 9, 654-659
124. Chen, C., Skog, J., Hsu, C. H., Lessard, R. T., Balaj, L., Wurdinger, T., Carter, B. S., Breakefield, X. O., Toner, M., and Irimia, D. (2010) Microfluidic isolation and transcriptome analysis of serum microvesicles. Lab Chip 10, 505–511
125. Miranda, K. C., Bond, D. T., McKee, M., Skog, J., Paunescu, T. G., Da Silva, N., Brown, D., and Russo, L. M. (2010) Nucleic acids within urinary exosomes/microvesicles are potential biomarkers for renal disease. Kidney international 78, 191-199
126. Balaj, L., Lessard, R., Dai, L., Cho, Y. J., Pomeroy, S. L., Breakefield, X. O., and Skog, J. (2011) Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nature communications 2, 180
127. Liu, Y., Shah, S. V., Xiang, X., Wang, J., Deng, Z. B., Liu, C., Zhang, L., Wu, J., Edmonds, T., Jambor, C., Kappes, J. C., and Zhang, H. G. (2009) COP9-associated CSN5 regulates exosomal protein deubiquitination and sorting. The American journal of pathology 174, 1415-1425
128. Liu, S., Hossinger, A., Hofmann, J. P., Denner, P., and Vorberg, I. M. (2016) Horizontal Transmission of Cytosolic Sup35 Prions by Extracellular Vesicles. mBio 7
129. Kubikova, I., Konecna, H., Sedo, O., Zdrahal, Z., Rehulka, P., Hribkova, H., Rehulkova, H., Hampl, A., Chmelik, J., and Dvorak, P. (2009) Proteomic profiling of human embryonic stem cell-derived microvesicles reveals a risk of transfer of proteins of bovine and mouse origin. Cytotherapy 11, 330-340, 331 p following 340
130. Millimaggi, D., Festuccia, C., Angelucci, A., D'Ascenzo, S., Rucci, N., Flati, S., Bologna, M., Teti, A., Pavan, A., and Dolo, V. (2006) Osteoblast-conditioned media stimulate membrane vesicle shedding in prostate cancer cells. International journal of oncology 28, 909-914

References
178
131. Gasser, O., and Schifferli, J. A. (2004) Activated polymorphonuclear neutrophils disseminate anti-inflammatory microparticles by ectocytosis. Blood 104, 2543-2548
132. Raposo, G., Nijman, H. W., Stoorvogel, W., Liejendekker, R., Harding, C. V., Melief, C. J., and Geuze, H. J. (1996) B lymphocytes secrete antigen-presenting vesicles. The Journal of Experimental Medicine 183, 1161-1172
133. Escrevente, C., Keller, S., Altevogt, P., and Costa, J. (2011) Interaction and uptake of exosomes by ovarian cancer cells. BMC Cancer 11, 108
134. Montecalvo, A., Larregina, A. T., Shufesky, W. J., Stolz, D. B., Sullivan, M. L., Karlsson, J. M., Baty, C. J., Gibson, G. A., Erdos, G., Wang, Z., Milosevic, J., Tkacheva, O. A., Divito, S. J., Jordan, R., Lyons-Weiler, J., Watkins, S. C., and Morelli, A. E. (2012) Mechanism of transfer of functional microRNAs between mouse dendritic cells via exosomes. Blood 119, 756-766
135. Grapp, M., Wrede, A., Schweizer, M., Huwel, S., Galla, H. J., Snaidero, N., Simons, M., Buckers, J., Low, P. S., Urlaub, H., Gartner, J., and Steinfeld, R. (2013) Choroid plexus transcytosis and exosome shuttling deliver folate into brain parenchyma. Nature communications 4, 2123
136. Koppler, B., Cohen, C., Schlondorff, D., and Mack, M. (2006) Differential mechanisms of microparticle transfer toB cells and monocytes: anti-inflammatory propertiesof microparticles. European journal of immunology 36, 648-660
137. Alvarez-Erviti, L., Seow, Y., Yin, H., Betts, C., Lakhal, S., and Wood, M. J. A. (2011) Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nature biotechnology 29, 341-345
138. Smyth, T. J., Redzic, J. S., Graner, M. W., and Anchordoquy, T. J. (2014) Examination of the specificity of tumor cell derived exosomes with tumor cells in vitro. Biochimica et biophysica acta 1838, 2954-2965
139. Spek, C. A. (2004) Tissue factor: from 'just one of the coagulation factors' to a major player in physiology. Blood Coagulation & Fibrinolysis 15, S3-S10
140. Bruno, S., Grange, C., Deregibus, M. C., Calogero, R. A., Saviozzi, S., Collino, F., Morando, L., Busca, A., Falda, M., Bussolati, B., Tetta, C., and Camussi, G. (2009) Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. Journal of the American Society of Nephrology 20, 1053-1067
141. Herrera, M. B., Fonsato, V., Gatti, S., Deregibus, M. C., Sordi, A., Cantarella, D., Calogero, R., Bussolati, B., Tetta, C., and Camussi, G. (2010) Human liver stem cell-derived microvesicles accelerate hepatic regeneration in hepatectomized rats. Journal of cellular and molecular medicine 14, 1605-1618
142. Bellingham, S. A., Coleman, B. M., and Hill, A. F. (2012) Small RNA deep sequencing reveals a distinct miRNA signature released in exosomes from prion-infected neuronal cells. Nucleic acids research 40, 10937-10949
143. Sang, Q., Yao, Z., Wang, H., Feng, R., Wang, H., Zhao, X., Xing, Q., Jin, L., He, L., Wu, L., and Wang, L. (2013) Identification of microRNAs in human follicular fluid: characterization of microRNAs that govern steroidogenesis in vitro and are associated with polycystic ovary syndrome in vivo. The Journal of Clinical Endocrinology & Metabolism 98, 3068-3079
144. Santonocito, M., Vento, M., Guglielmino, M. R., Battaglia, R., Wahlgren, J., Ragusa, M., Barbagallo, D., Borzi, P., Rizzari, S., Maugeri, M., Scollo, P., Tatone, C., Valadi, H., Purrello, M., and Di Pietro, C. (2014) Molecular characterization of exosomes and their microRNA cargo in human follicular fluid: bioinformatic analysis reveals that exosomal microRNAs control

References
179
pathways involved in follicular maturation. Fertility and Sterility 102, 1751-1761 e1751
145. Sohel, M. M., Hoelker, M., Noferesti, S. S., Salilew-Wondim, D., Tholen, E., Looft, C., Rings, F., Uddin, M. J., Spencer, T. E., Schellander, K., and Tesfaye, D. (2013) Exosomal and Non-Exosomal Transport of Extra-Cellular microRNAs in Follicular Fluid: Implications for Bovine Oocyte Developmental Competence. PLOS One 8, e78505
146. Eaton, S. A., Jayasooriah, N., Buckland, M. E., Martin, D. I., Cropley, J. E., and Suter, C. M. (2015) Roll over Weismann: extracellular vesicles in the transgenerational transmission of environmental effects. Epigenomics 7, 1165-1171
147. Mather, J. P. (1980) Establishment and characterization of two distinct mouse testicular epithelial cell lines. Biology of Reproduction 23, 243-252
148. Araki, Y., Suzuki, K., Matusik, R. J., Obinata, M., and Orgebin-Crist, M. C. (2002) Immortalized epididymal cell lines from transgenic mice overexpressing temperature-sensitive simian virus 40 large T-antigen gene. Journal of Andrology 23, 854-869
149. Hofmann, M. C., Braydich-Stolle, L., Dettin, L., Johnson, E., and Dym, M. (2005) Immortalization of mouse germ line stem cells. Stem Cells 23, 200-210
150. Shelke, G. V., Lasser, C., Gho, Y. S., and Lotvall, J. (2014) Importance of exosome depletion protocols to eliminate functional and RNA-containing extracellular vesicles from fetal bovine serum. Journal of extracellular vesicles 3
151. Dragovic, R. A., Gardiner, C., Brooks, A. S., Tannetta, D. S., Ferguson, D. J. P., Hole, P., Carr, B., Redman, C. W. G., Harris, A. L., Dobson, P. J., Harrison, P., and Sargent, I. L. (2011) Sizing and phenotyping of cellular vesicles using Nanoparticle Tracking Analysis. Nanomedicine: Nanotechnology, Biology and Medicine 7, 780-788
152. Keerthikumar, S., Chisanga, D., Ariyaratne, D., Al Saffar, H., Anand, S., Zhao, K., Samuel, M., Pathan, M., Jois, M., Chilamkurti, N., Gangoda, L., and Mathivanan, S. (2016) ExoCarta: A Web-Based Compendium of Exosomal Cargo. Journal of Molecular Biology 428, 688-692
153. Kalra, H., Simpson, R. J., Ji, H., Aikawa, E., Altevogt, P., Askenase, P., Bond, V. C., Borras, F. E., Breakefield, X., Budnik, V., Buzas, E., Camussi, G., Clayton, A., Cocucci, E., Falcon-Perez, J. M., Gabrielsson, S., Gho, Y. S., Gupta, D., Harsha, H. C., Hendrix, A., Hill, A. F., Inal, J. M., Jenster, G., Kramer-Albers, E. M., Lim, S. K., Llorente, A., Lotvall, J., Marcilla, A., Mincheva-Nilsson, L., Nazarenko, I., Nieuwland, R., Nolte-'t Hoen, E. N., Pandey, A., Patel, T., Piper, M. G., Pluchino, S., Prasad, T. S., Rajendran, L., Raposo, G., Record, M., Reid, G. E., Sanchez-Madrid, F., Schiffelers, R. M., Siljander, P., Stensballe, A., Stoorvogel, W., Taylor, D., Thery, C., Valadi, H., van Balkom, B. W., Vazquez, J., Vidal, M., Wauben, M. H., Yanez-Mo, M., Zoeller, M., and Mathivanan, S. (2012) Vesiclepedia: a compendium for extracellular vesicles with continuous community annotation. PLOS Biology 10, e1001450
154. Benjamini, Y., and Hochberg, Y. (1995) Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. Journal of the Royal Statistical Society. Series B (Methodological) 57, 289-300

References
180
155. Kim, Y. K., Yeo, J., Kim, B., Ha, M., and Kim, V. N. (2012) Short structured RNAs with low GC content are selectively lost during extraction from a small number of cells. Molecular Cell 46, 893–895
156. Agarwal, V., Bell, G. W., Nam, J. W., and Bartel, D. P. (2015) Predicting effective microRNA target sites in mammalian mRNAs. eLife 4
157. Paraskevopoulou, M. D., Georgakilas, G., Kostoulas, N., Vlachos, I. S., Vergoulis, T., Reczko, M., Filippidis, C., Dalamagas, T., and Hatzigeorgiou, A. G. (2013) DIANA-microT web server v5.0: service integration into miRNA functional analysis workflows. Nucleic acids research 41, W169-173
158. Ahmed, N., Yufei, H., Yang, P., Muhammad Yasir, W., Zhang, Q., Liu, T., Hong, C., Lisi, H., Xiaoya, C., and Chen, Q. (2016) Cytological study on Sertoli cells and their interactions with germ cells during annual reproductive cycle in turtle. Ecology and Evolution 6, 4050-4064
159. Vojtech, L., Woo, S., Hughes, S., Levy, C., Ballweber, L., Sauteraud, R. P., Strobl, J., Westerberg, K., Gottardo, R., Tewari, M., and Hladik, F. (2014) Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic acids research 42, 7290-7304
160. Coughlin, D. J., Babak, T., Nihranz, C., Hughes, T. R., and Engelke, D. R. (2009) Prediction and verification of mouse tRNA gene families. RNA Biology 6, 195-202
161. Drobna, Z., Henriksen, A. D., Wolstenholme, J. T., Montiel, C., Lambeth, P. S., Shang, S., Harris, E. P., Zhou, C., Flaws, J. A., Adli, M., and Rissman, E. F. (2017) Transgenerational effects of Bisphenol A on gene expression and DNA methylation of imprinted genes in brain. Endocrinology
162. Oakberg, E. F. (1956) Duration of spermatogenesis in the mouse and timing of stages of the cycle of the seminiferous epithelium. Am J Anat 99, 507-516
163. Thery, C., Clayton, A., Amigorena, S., and Raposo, G. (2006) Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Current Protocols in Cell Biology Chapter 3, Unit 3 22
164. Crescitelli, R., Lasser, C., Szabo, T. G., Kittel, A., Eldh, M., Dianzani, I., Buzas, E. I., and Lotvall, J. (2013) Distinct RNA profiles in subpopulations of extracellular vesicles: apoptotic bodies, microvesicles and exosomes. Journal of extracellular vesicles 2
165. Lunavat, T. R., Cheng, L., Kim, D. K., Bhadury, J., Jang, S. C., Lasser, C., Sharples, R. A., Lopez, M. D., Nilsson, J., Gho, Y. S., Hill, A. F., and Lotvall, J. (2015) Small RNA deep sequencing discriminates subsets of extracellular vesicles released by melanoma cells--Evidence of unique microRNA cargos. RNA Biology 12, 810-823
166. Chernyshev, V. S., Rachamadugu, R., Tseng, Y. H., Belnap, D. M., Jia, Y., Branch, K. J., Butterfield, A. E., Pease, L. F., 3rd, Bernard, P. S., and Skliar, M. (2015) Size and shape characterization of hydrated and desiccated exosomes. Analytical and Bioanalytical Chemistry 407, 3285-3301
167. Taylor, D. D., and Gercel-Taylor, C. (2008) MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecologic oncology 110, 13-21
168. Michael, A., Bajracharya, S. D., Yuen, P. S. T., Zhou, H., Star, R. A., Illei, G. G., and Alevizos, I. (2010) Exosomes from human saliva as a source of microRNA biomarkers. Oral Diseases 16, 34-38
169. Ekstrom, K., Valadi, H., Sjostrand, M., Malmhall, C., Bossios, A., Eldh, M., and Lotvall, J. (2012) Characterization of mRNA and microRNA in human mast

References
181
cell-derived exosomes and their transfer to other mast cells and blood CD34 progenitor cells. Journal of extracellular vesicles 1
170. Ortogero, N., Hennig, G. W., Langille, C., Ro, S., McCarrey, J. R., and Yan, W. (2013) Computer-assisted annotation of murine Sertoli cell small RNA transcriptome. Biology of Reproduction 88, 3
171. Akers, J. C., Ramakrishnan, V., Kim, R., Skog, J., Nakano, I., Pingle, S., Kalinina, J., Hua, W., Kesari, S., Mao, Y., Breakefield, X. O., Hochberg, F. H., Van Meir, E. G., Carter, B. S., and Chen, C. C. (2013) MiR-21 in the extracellular vesicles (EVs) of cerebrospinal fluid (CSF): a platform for glioblastoma biomarker development. PLOS One 8, e78115
172. Berardocco, M., Radeghieri, A., Busatto, S., Gallorini, M., Raggi, C., Gissi, C., D'Agnano, I., Bergese, P., Felsani, A., and Berardi, A. C. (2017) RNA-seq reveals distinctive RNA profiles of small extracellular vesicles from different human liver cancer cell lines. Oncotarget 8, 82920-82939
173. Kumarswamy, R., Volkmann, I., and Thum, T. (2011) Regulation and function of miRNA-21 in health and disease. RNA Biology 8, 706-713
174. Niu, Z., Goodyear, S. M., Rao, S., Wu, X., Tobias, J. W., Avarbock, M. R., and Brinster, R. L. (2011) MicroRNA-21 regulates the self-renewal of mouse spermatogonial stem cells. Proceedings of the National Academy of Sciences of the United States of America 108, 12740-12745
175. Tong, M. H., Mitchell, D., Evanoff, R., and Griswold, M. D. (2011) Expression of Mirlet7 family microRNAs in response to retinoic acid-induced spermatogonial differentiation in mice. Biology of Reproduction 85, 189-197
176. Nolte-'t Hoen, E. N., Buermans, H. P., Waasdorp, M., Stoorvogel, W., Wauben, M. H., and t Hoen, P. A. (2012) Deep sequencing of RNA from immune cell-derived vesicles uncovers the selective incorporation of small non-coding RNA biotypes with potential regulatory functions. Nucleic acids research 40, 9272-9285
177. Guzman, N., Agarwal, K., Asthagiri, D., Yu, L., Saji, M., Ringel, M. D., and Paulaitis, M. E. (2015) Breast Cancer-Specific miR Signature Unique to Extracellular Vesicles Includes "microRNA-like" tRNA Fragments. Molecular Cancer Research 13, 891-901
178. Fu, H., Feng, J., Liu, Q., Sun, F., Tie, Y., Zhu, J., Xing, R., Sun, Z., and Zheng, X. (2009) Stress induces tRNA cleavage by angiogenin in mammalian cells. FEBS letters 583, 437-442
179. Lee, Y. S., Shibata, Y., Malhotra, A., and Dutta, A. (2009) A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes & development 23, 2639-2649
180. Cole, C., Sobala, A., Lu, C., Thatcher, S. R., Bowman, A., Brown, J. W., Green, P. J., Barton, G. J., and Hutvagner, G. (2009) Filtering of deep sequencing data reveals the existence of abundant Dicer-dependent small RNAs derived from tRNAs. RNA 15, 2147-2160
181. Haussecker, D., Huang, Y., Lau, A., Parameswaran, P., Fire, A. Z., and Kay, M. A. (2010) Human tRNA-derived small RNAs in the global regulation of RNA silencing. RNA 16, 673-695
182. Wang, Q., Lee, I., Ren, J., Ajay, S. S., Lee, Y. S., and Bao, X. (2013) Identification and functional characterization of tRNA-derived RNA fragments (tRFs) in respiratory syncytial virus infection. Molecular Therapy 21, 368-379
183. Deng, J., Ptashkin, R. N., Chen, Y., Cheng, Z., Liu, G., Phan, T., Deng, X., Zhou, J., Lee, I., Lee, Y. S., and Bao, X. (2015) Respiratory Syncytial Virus

References
182
Utilizes a tRNA Fragment to Suppress Antiviral Responses Through a Novel Targeting Mechanism. Molecular Therapy 23, 1622-1629
184. Pap, E., Pallinger, E., Falus, A., Kiss, A. A., Kittel, A., Kovacs, P., and Buzas, E. I. (2008) T lymphocytes are targets for platelet- and trophoblast-derived microvesicles during pregnancy. Placenta 29, 826-832
185. Chen, L., Wang, Y., Pan, Y., Zhang, L., Shen, C., Qin, G., Ashraf, M., Weintraub, N., Ma, G., and Tang, Y. (2013) Cardiac progenitor-derived exosomes protect ischemic myocardium from acute ischemia/reperfusion injury. Biochemical and Biophysical Research Communications 431, 566-571
186. Madison, R. D., McGee, C., Rawson, R., and Robinson, G. A. (2014) Extracellular vesicles from a muscle cell line (C2C12) enhance cell survival and neurite outgrowth of a motor neuron cell line (NSC-34). Journal of extracellular vesicles 3
187. Mulcahy, L. A., Pink, R. C., and Carter, D. R. (2014) Routes and mechanisms of extracellular vesicle uptake. Journal of extracellular vesicles 3
188. (2017) PKH26 [package insert], Sigma Aldrich, St Louis, MO. 189. Takov, K., Yellon, D. M., and Davidson, S. M. (2017) Confounding factors in
vesicle uptake studies using fluorescent lipophilic membrane dyes. Journal of extracellular vesicles 6, 1388731
190. Matsumoto, A., Takahashi, Y., Nishikawa, M., Sano, K., Morishita, M., Charoenviriyakul, C., Saji, H., and Takakura, Y. (2017) Accelerated growth of B16BL6 tumor in mice through efficient uptake of their own exosomes by B16BL6 cells. Cancer Science 108, 1803-1810
191. de Menorval, M. A., Mir, L. M., Fernandez, M. L., and Reigada, R. (2012) Effects of dimethyl sulfoxide in cholesterol-containing lipid membranes: a comparative study of experiments in silico and with cells. PLOS One 7, e41733
192. Takasugi, M., Okada, R., Takahashi, A., Virya Chen, D., Watanabe, S., and Hara, E. (2017) Small extracellular vesicles secreted from senescent cells promote cancer cell proliferation through EphA2. Nature communications 8, 15729
193. Majumdar, R., Tavakoli Tameh, A., and Parent, C. A. (2016) Exosomes Mediate LTB4 Release during Neutrophil Chemotaxis. PLOS Biology 14, e1002336
194. King, H. W., Michael, M. Z., and Gleadle, J. M. (2012) Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer 12, 421
195. Kanemoto, S., Nitani, R., Murakami, T., Kaneko, M., Asada, R., Matsuhisa, K., Saito, A., and Imaizumi, K. (2016) Multivesicular body formation enhancement and exosome release during endoplasmic reticulum stress. Biochemical and Biophysical Research Communications 480, 166-172
196. Aubertin, K., Silva, A. K., Luciani, N., Espinosa, A., Djemat, A., Charue, D., Gallet, F., Blanc-Brude, O., and Wilhelm, C. (2016) Massive release of extracellular vesicles from cancer cells after photodynamic treatment or chemotherapy. Scientific Reports 6, 35376
197. Lin, T., Dong, W., Huang, J., Pan, Q., Fan, X., Zhang, C., and Huang, L. (2009) MicroRNA-143 as a tumor suppressor for bladder cancer. The Journal of urology 181, 1372-1380
198. Liu, L., Yu, X., Guo, X., Tian, Z., Su, M., Long, Y., Huang, C., Zhou, F., Liu, M., Wu, X., and Wang, X. (2012) miR-143 is downregulated in cervical cancer and promotes apoptosis and inhibits tumor formation by targeting Bcl-2. Molecular medicine reports 5, 753-760

References
183
199. He, Z., Yi, J., Liu, X., Chen, J., Han, S., Jin, L., Chen, L., and Song, H. (2016) MiR-143-3p functions as a tumor suppressor by regulating cell proliferation, invasion and epithelial–mesenchymal transition by targeting QKI-5 in esophageal squamous cell carcinoma. Molecular Cancer 15, 51
200. Fullston, T., Ohlsson-Teague, E. M., Print, C. G., Sandeman, L. Y., and Lane, M. (2016) Sperm microRNA Content Is Altered in a Mouse Model of Male Obesity, but the Same Suite of microRNAs Are Not Altered in Offspring's Sperm. PLOS One 11, e0166076
201. Hermo, L., and Jacks, D. (2002) Nature's ingenuity: bypassing the classical secretory route via apocrine secretion. Molecular Reproduction and Development 63, 394-410
202. Frenette, G., Girouard, J., and Sullivan, R. (2006) Comparison between epididymosomes collected in the intraluminal compartment of the bovine caput and cauda epididymidis. Biology of Reproduction 75, 885-890
203. Fornes, M. W., Barbieri, A., and Cavicchia, J. C. (1995) Morphological and enzymatic study of membrane-bound vesicles from the lumen of the rat epididymis. Andrologia 27, 1-5
204. Thimon, V., Frenette, G., Saez, F., Thabet, M., and Sullivan, R. (2008) Protein composition of human epididymosomes collected during surgical vasectomy reversal: a proteomic and genomic approach. Human Reproduction 23, 1698-1707
205. Jerczynski, O., Lacroix-Pepin, N., Boilard, E., Calvo, E., Bernet, A., Fortier, M. A., Bjorkgren, I., Sipila, P., and Belleannee, C. (2016) Role of Dicer1-Dependent Factors in the Paracrine Regulation of Epididymal Gene Expression. PLOS One 11, e0163876
206. Martin-DeLeon, P. A. (2015) Epididymosomes: transfer of fertility-modulating proteins to the sperm surface. Asian Journal of Andrology 17, 720-725
207. Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E., and Mello, C. C. (1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806-811
208. Devanapally, S., Ravikumar, S., and Jose, A. M. (2015) Double-stranded RNA made in C. elegans neurons can enter the germline and cause transgenerational gene silencing. Proceedings of the National Academy of Sciences of the United States of America 112, 2133-2138
209. Gapp, K., Jawaid, A., Sarkies, P., Bohacek, J., Pelczar, P., Prados, J., Farinelli, L., Miska, E., and Mansuy, I. M. (2014) Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nature Neuroscience
210. Chen, Z., Zuo, X., He, D., Ding, S., Xu, F., Yang, H., Jin, X., Fan, Y., Ying, L., Tian, C., and Ying, C. (2017) Long-term exposure to a 'safe' dose of bisphenol A reduced protein acetylation in adult rat testes. Scientific Reports 7, 40337
211. Kim, Y. I., Pogribny, I. P., Basnakian, A. G., Miller, J. W., Selhub, J., James, S. J., and Mason, J. B. (1997) Folate deficiency in rats induces DNA strand breaks and hypomethylation within the p53 tumor suppressor gene. The American Journal of Clinical Nutrition 65, 46-52
212. Varela-Moreiras, G., Ragel, C., and Perez de Miguelsanz, J. (1995) Choline deficiency and methotrexate treatment induces marked but reversible changes in hepatic folate concentrations, serum homocysteine and DNA methylation rates in rats. Journal of the American College of Nutrition 14, 480-485

References
184
213. Kim, H. W., Kim, K. N., Choi, Y. J., and Chang, N. (2013) Effects of paternal folate deficiency on the expression of insulin-like growth factor-2 and global DNA methylation in the fetal brain. Molecular nutrition & food research 57, 671-676
214. Lambrot, R., Xu, C., Saint-Phar, S., Chountalos, G., Cohen, T., Paquet, M., Suderman, M., Hallett, M., and Kimmins, S. (2013) Low paternal dietary folate alters the mouse sperm epigenome and is associated with negative pregnancy outcomes. Nature communications 4, 2889
215. Boxmeer, J. C., Smit, M., Utomo, E., Romijn, J. C., Eijkemans, M. J. C., Lindemans, J., Laven, J. S. E., Macklon, N. S., Steegers, E. A. P., and Steegers-Theunissen, R. P. M. (2009) Low folate in seminal plasma is associated with increased sperm DNA damage. Fertility and Sterility 92, 548-556
216. Bentivoglio, G., Melica, F., and Cristoforoni, P. (1993) Folinic acid in the treatment of human male infertility. Fertility and Sterility 60, 698-701
217. Aarabi, M., San Gabriel, M. C., Chan, D., Behan, N. A., Caron, M., Pastinen, T., Bourque, G., MacFarlane, A. J., Zini, A., and Trasler, J. (2015) High-dose folic acid supplementation alters the human sperm methylome and is influenced by the MTHFR C677T polymorphism. Human Molecular Genetics 24, 6301-6313
218. Weitman, S. D., Weinberg, A. G., Coney, L. R., Zurawski, V. R., Jennings, D. S., and Kamen, B. A. (1992) Cellular localization of the folate receptor: potential role in drug toxicity and folate homeostasis. Cancer research 52, 6708-6711
219. Girouard, J., Frenette, G., and Sullivan, R. (2011) Comparative proteome and lipid profiles of bovine epididymosomes collected in the intraluminal compartment of the caput and cauda epididymidis. International Journal of Andrology 34, e475-486
220. Kotaja, N. (2014) MicroRNAs and spermatogenesis. Fertility and Sterility 101, 1552-1562
221. Ruan, Y. C., Shum, W. W., Belleannee, C., Da Silva, N., and Breton, S. (2012) ATP secretion in the male reproductive tract: essential role of CFTR. The Journal of physiology 590, 4209-4222
222. Roush, S., and Slack, F. J. (2008) The let-7 family of microRNAs. Trends in cell biology 18, 505-516
223. Pellis, L., Dommels, Y., Venema, D., Polanen, A., Lips, E., Baykus, H., Kok, F., Kampman, E., and Keijer, J. (2008) High folic acid increases cell turnover and lowers differentiation and iron content in human HT29 colon cancer cells. British Journal of Nutrition 99, 703-708
224. Wolff, G. L., Kodell, R. L., Moore, S. R., and Cooney, C. A. (1998) Maternal epigenetics and methyl supplements affect agouti gene expression in Avy/a mice. The FASEB Journal 12, 949-957
225. Pauwels, S., Truijen, I., Ghosh, M., Duca, R. C., Langie, S. A. S., Bekaert, B., Freson, K., Huybrechts, I., Koppen, G., Devlieger, R., and Godderis, L. (2017) The effect of paternal methyl-group donor intake on offspring DNA methylation and birth weight. Journal of Developmental Origins of Health and Disease 8, 311-321
226. Ryan, D. P., Henzel, K. S., Pearson, B. L., Siwek, M. E., Papazoglou, A., Guo, L., Paesler, K., Yu, M., Muller, R., Xie, K., Schroder, S., Becker, L., Garrett, L., Holter, S. M., Neff, F., Racz, I., Rathkolb, B., Rozman, J., Ehninger, G., Klingenspor, M., Klopstock, T., Wolf, E., Wurst, W., Zimmer, A., Fuchs, H., Gailus-Durner, V., Hrabe de Angelis, M., Sidiropoulou, K., Weiergraber, M.,

References
185
Zhou, Y., and Ehninger, D. (2018) A paternal methyl donor-rich diet altered cognitive and neural functions in offspring mice. Molecular Psychiatry 23, 1345-1355
227. Caballero, J. N., Frenette, G., Belleannee, C., and Sullivan, R. (2013) CD9-positive microvesicles mediate the transfer of molecules to Bovine Spermatozoa during epididymal maturation. PLOS One 8, e65364
228. Schwarz, A., Wennemuth, G., Post, H., Brandenburger, T., Aumuller, G., and Wilhelm, B. (2013) Vesicular transfer of membrane components to bovine epididymal spermatozoa. Cell and tissue research 353, 549-561
229. Fischer, K. A., Van Leyen, K., Lovercamp, K. W., Manandhar, G., Sutovsky, M., Feng, D., Safranski, T., and Sutovsky, P. (2005) 15-Lipoxygenase is a component of the mammalian sperm cytoplasmic droplet. Reproduction 130, 213-222
230. Nixon, B., Stanger, S. J., Mihalas, B. P., Reilly, J. N., Anderson, A. L., Dun, M. D., Tyagi, S., Holt, J. E., and McLaughlin, E. A. (2015) Next Generation Sequencing Analysis Reveals Segmental Patterns of microRNA Expression in Mouse Epididymal Epithelial Cells. PLOS One 10, e0135605
231. Chu, C., Yu, L., Wu, B., Ma, L., Gou, L. T., He, M., Guo, Y., Li, Z. T., Gao, W., Shi, H., Liu, M. F., Wang, H., Chen, C. D., Drevet, J. R., Zhou, Y., and Zhang, Y. (2017) A sequence of 28S rRNA-derived small RNAs is enriched in mature sperm and various somatic tissues and possibly associates with inflammation. Journal of Molecular Cell Biology 9, 256-259
232. Belleannee, C., Calvo, E., Caballero, J., and Sullivan, R. (2013) Epididymosomes convey different repertoires of microRNAs throughout the bovine epididymis. Biology of Reproduction 89, 30
233. Wang, X., Liang, Z., Xu, X., Li, J., Zhu, Y., Meng, S., Li, S., Wang, S., Xie, B., Ji, A., Liu, B., Zheng, X., and Xie, L. (2016) miR-148a-3p represses proliferation and EMT by establishing regulatory circuits between ERBB3/AKT2/c-myc and DNMT1 in bladder cancer. Cell Death & Disease 7, e2503
234. Wu, X., Zhao, B., Li, W., Chen, Y., Liang, R., Li, L., Jin, Y., and Ruan, K. (2012) MiR-200a is involved in rat epididymal development by targeting β-catenin mRNA. Acta biochimica et biophysica Sinica 44, 233-240
235. Gassei, K., Ehmcke, J., Wood, M. A., Walker, W. H., and Schlatt, S. (2010) Immature rat seminiferous tubules reconstructed in vitro express markers of Sertoli cell maturation after xenografting into nude mouse hosts. Mol Hum Reprod 16, 97-110
236. Clulow, J., and Jones, R. C. (2004) Composition of luminal fluid secreted by the seminiferous tubules and after reabsorption by the extratesticular ducts of the Japanese quail, Coturnix coturnix japonica. Biology of Reproduction 71, 1508-1516

186
8 APPENDIX
Table 8.1 Proteins identified in Sertoli EVs
Protein Gene symbol Unique
matching peptides
% protein covered
Alpha-2-macroglobulin-P A2mp 11 5.7 Actin, alpha skeletal muscle Acta1 20 28.9 Actin, aortic smooth muscle Acta2 32 68.4
Actin, cytoplasmic 1 Actb 34 77.7 Beta-actin-like protein 2 Actbl2 20 49.7
Disintegrin and metalloproteinase domain-containing protein 10 Adam10 5 9.2 Adiponectin Adipoq 4 14.6
Adipocyte enhancer-binding protein 1 Aebp1 7 8.6 Adenosylhomocysteinase Ahcy 7 18.3
Axin interactor, dorsalization-associated protein Aida 2 8.5 Aldose reductase Akr1b1 5 13 Serum albumin Alb 9 9.3
Fructose-bisphosphate aldolase A Aldoa 6 18.4 Protein AMBP Ambp 2 5.8
Annexin A1 Anxa1 11 39 Annexin A2 Anxa2 12 32.2 Annexin A3 Anxa3 4 19.5 Annexin A4 Anxa4 10 33.9 Annexin A5 Anxa5 14 39.8 Annexin A6 Anxa6 23 34.5
Rho GDP-dissociation inhibitor 1 Arhgdia 2 15.2 Rho GDP-dissociation inhibitor 2 Arhgdib 2 10.5
Actin-related protein 2/3 complex subunit 4 Arpc4 3 19 Sodium/potassium-transporting ATPase subunit alpha-1 Atp1a1 18 22.5 Sodium/potassium-transporting ATPase subunit alpha-2 Atp1a2 7 11.2 Sodium/potassium-transporting ATPase subunit alpha-4 Atp1a4 3 5.1
Brain acid soluble protein 1 Basp1 3 15.5 Protein BRICK1 Brk1 2 28
Basigin Bsg 7 18.8 Complement C1q tumor necrosis factor-related protein 3 C1qtnf3 6 14.2
Complement C3 C3 31 17.6 Complement C4-B C4b 9 5.4
Macrophage-capping protein Capg 2 7.1 F-actin-capping protein subunit beta Capzb 3 10.5
T-complex protein 1 subunit beta Cct2 3 9 T-complex protein 1 subunit gamma Cct3 4 7.7 T-complex protein 1 subunit epsilon Cct5 3 6.7

Appendix
187
T-complex protein 1 subunit eta Cct7 2 5 CD151 antigen Cd151 2 6.7 CD63 antigen Cd63 2 8 CD81 antigen Cd81 16 58.4 CD9 antigen Cd9 2 8.4
Cell division control protein 42 homolog Cdc42 3 23.6 CDC42 small effector protein 2 Cdc42se2 2 25
Cyclin-dependent kinase 1 Cdk1 4 14.1 Cofilin-1 Cfl1 5 20.5 Cofilin-2 Cfl2 3 12
Charged multivesicular body protein 1b-1 Chmp1b1 3 14.1 Charged multivesicular body protein 4b Chmp4b 3 11.2
Claudin domain-containing protein 1 Cldnd1 3 9.5 Chloride intracellular channel protein 1 Clic1 3 19.5 Chloride intracellular channel protein 4 Clic4 4 9.5
Clathrin heavy chain 1 Cltc 14 8.8 Collagen alpha-1(I) chain Col1a1 24 15.3 Collagen alpha-2(I) chain Col1a2 45 34.8
Collagen alpha-1(III) chain Col3a1 14 12.7 Collagen alpha-1(V) chain Col5a1 10 5 Collagen alpha-1(VI) chain Col6a1 6 7.6
Carboxypeptidase B2 Cpb2 4 7.4 Carboxypeptidase N catalytic chain Cpn1 5 10.5
Catenin alpha-1 Ctnna1 5 8.9 Catenin beta-1 Ctnnb1 8 11.4 Catenin delta-1 Ctnnd1 6 6.7
NADH-cytochrome b5 reductase 3 Cyb5r3 2 9 Cytoplasmic FMR1-interacting protein 1 Cyfip1 20 16.8 Cytoplasmic FMR1-interacting protein 2 Cyfip2 10 7.5
N(G),N(G)-dimethylarginine dimethylaminohydrolase 1 Ddah1 4 11.9 N(G),N(G)-dimethylarginine dimethylaminohydrolase 2 Ddah2 5 27.4
DnaJ homolog subfamily A member 1 Dnaja1 14 28.7 DnaJ homolog subfamily A member 2 Dnaja2 7 15
Destrin Dstn 4 15.8 Elongation factor 1-alpha 1 Eef1a1 23 53.1 Elongation factor 1-alpha 2 Eef1a2 10 19
Elongation factor 1-delta Eef1d 4 21.4 Elongation factor 1-gamma Eef1g 6 12.9
Elongation factor 2 Eef2 15 21.9 EH domain-containing protein 1 Ehd1 7 19 EH domain-containing protein 2 Ehd2 4 7.6 EH domain-containing protein 3 Ehd3 4 8.5
Eukaryotic translation initiation factor 3 subunit H Eif3h 2 8.8 Eukaryotic translation initiation factor 3 subunit I Eif3i 2 6.2
Eukaryotic initiation factor 4A-I Eif4a1 5 13.3 Eukaryotic initiation factor 4A-II Eif4a2 4 9.6

Appendix
188
Eukaryotic translation initiation factor 5A-1 Eif5a 2 9.7 EMILIN-1 Emilin1 7 13.5
Homeobox protein engrailed-1 En1 5 9.2 Homeobox protein engrailed-2 En2 10 31.1
Alpha-enolase Eno1 13 32.5 Ephrin type-A receptor 2 Epha2 21 21.4 Ephrin type-B receptor 4 Ephb4 6 5.7
S-formylglutathione hydrolase Esd 2 7.1 Protein eva-1 homolog B Eva1b 2 17.7
Ezrin Ezr 8 10 Prothrombin F2 14 11.9
Coagulation factor V F5 14 5.7 FERM, RhoGEF and pleckstrin domain-containing protein 1 Farp1 16 16.7
Peptidyl-prolyl cis-trans isomerase FKBP4 Fkbp4 2 5.2 Formin-like protein 3 Fmnl3 4 6
Fibronectin Fn1 103 38.2 Ferritin heavy chain Fth1 2 12 Ferritin light chain 1 Ftl1 5 38.3 Ferritin light chain 2 Ftl2 3 21.3
Glyceraldehyde-3-phosphate dehydrogenase Gapdh 12 36.3 Rab GDP dissociation inhibitor alpha Gdi1 4 11.4 Rab GDP dissociation inhibitor beta Gdi2 8 23.2
Glial fibrillary acidic protein Gfap 3 5.8 PDZ domain-containing protein GIPC1 Gipc1 3 9.6
Gap junction alpha-1 protein Gja1 14 35.1 Guanine nucleotide-binding protein subunit alpha-12 Gna12 3 5 Guanine nucleotide-binding protein subunit alpha-13 Gna13 5 10.9
Guanine nucleotide-binding protein G(i) subunit alpha-1 Gnai1 7 15.5 Guanine nucleotide-binding protein G(i) subunit alpha-2 Gnai2 15 34.9 Guanine nucleotide-binding protein G(k) subunit alpha Gnai3 10 23.4
Guanine nucleotide-binding protein G(olf) subunit alpha Gnal 3 5 Guanine nucleotide-binding protein G(o) subunit alpha Gnao1 3 5.4
Guanine nucleotide-binding protein G(s) subunit alpha isoforms XLas
Gnas 11 8.6
Guanine nucleotide-binding protein G(t) subunit alpha-1 Gnat1 4 7.7 Guanine nucleotide-binding protein G(t) subunit alpha-2 Gnat2 5 9.6
Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-1
Gnb1 5 17.9
Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2
Gnb2 4 14.1
Guanine nucleotide-binding protein subunit beta-2-like 1 Gnb2l1 4 13.2 Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit
beta-3 Gnb3 2
5.3 Guanine nucleotide-binding protein subunit beta-4 Gnb4 3 8.8
Guanine nucleotide-binding protein G(I)/G(S)/G(O) subunit gamma-12
Gng12 3 34.7
Guanine nucleotide-binding protein G(I)/G(S)/G(O) subunit Gng5 2 22.1

Appendix
189
gamma-5 Aspartate aminotransferase, cytoplasmic Got1 2 6.1
Glypican-1 Gpc1 14 28.2 Glypican-4 Gpc4 7 12.7
Glucose-6-phosphate isomerase Gpi 2 5.7 Retinoic acid-induced protein 3 Gprc5a 2 7.6
Gelsolin Gsn 32 34.6 Histone H2A.V H2afv 5 30.5 Histone H2AX H2afx 7 32.9
H-2 class I histocompatibility antigen, Q10 alpha chain H2-Q10 2 7.4 Histone H3.3 H3f3a 11 30.9
Histone H3.3C H3f3c 16 39.6 Hemoglobin subunit alpha Hba 7 44.4 Hemoglobin subunit beta-1 Hbb-b1 4 25.8
Histone H1.1 Hist1h1a 5 27.7 Histone H1.5 Hist1h1b 3 13.9 Histone H1.2 Hist1h1c 5 26.4 Histone H1.3 Hist1h1d 5 25.8 Histone H1.4 Hist1h1e 6 26
Histone H2A type 1-F Hist1h2af 17 66.85 Histone H2A type 1-H Hist1h2ah 7 43 Histone H2B type 1-A Hist1h2ba 29 59.85 Histone H2B type 1-B Hist1h2bb 51 73
Histone H2B type 1-C/E/G Hist1h2bc 48 93.65 Histone H2B type 1-K Hist1h2bk 22 73
Histone H3.1 Hist1h3a 12 40.4 Histone H4 Hist1h4a 33 85.95
Histone H2A type 2-A Hist2h2aa1 10 71.5 Histone H2A type 2-B Hist2h2ab 7 39.9 Histone H2A type 2-C Hist2h2ac 4 31.8 Histone H2B type 2-E Hist2h2be 24 60.35 Histone H2B type 3-A Hist3h2ba 17 59.5
GTPase HRas Hras 2 12.2 Heat shock protein HSP 90-alpha Hsp90aa1 12 16.3 Heat shock protein HSP 90-beta Hsp90ab1 24 33.6 Heat shock 70 kDa protein 1A Hspa1a 8 14.8
Heat shock-related 70 kDa protein 2 Hspa2 10 14.8 78 kDa glucose-regulated protein Hspa5 4 8.3
Heat shock cognate 71 kDa protein Hspa8 42 64.2 Basement membrane-specific heparan sulfate proteoglycan core
protein Hspg2 83
29.4 Isocitrate dehydrogenase [NADP] cytoplasmic Idh1 3 7 Interferon-induced transmembrane protein 2 Ifitm2 4 11.2 Interferon-induced transmembrane protein 3 Ifitm3 6 20.4
Immunoglobulin superfamily member 8 Igsf8 11 18.8 Interleukin-1 receptor accessory protein Il1rap 3 5.1

Appendix
190
Immunity-related GTPase family M protein 1 Irgm1 2 6.6 IST1 homolog Ist1 3 8.8
Integrin alpha-3 Itga3 13 14.6 Integrin beta-1 Itgb1 9 13.7
Inter-alpha-trypsin inhibitor heavy chain H1 Itih1 9 7.2 Inter-alpha-trypsin inhibitor heavy chain H2 Itih2 19 21.7 Inter-alpha-trypsin inhibitor heavy chain H3 Itih3 12 10.5
GTPase KRas Kras 3 18.5 L-lactate dehydrogenase A chain Ldha 8 22 L-lactate dehydrogenase B chain Ldhb 2 5.1
Galectin-1 Lgals1 2 17.8 Galectin-3-binding protein Lgals3bp 18 36.2
Protein lin-7 homolog C Lin7c 2 10.7 Lumican Lum 2 5.3
Myristoylated alanine-rich C-kinase substrate Marcks 2 11 Malate dehydrogenase, cytoplasmic Mdh1 4 13.8
Lactadherin Mfge8 22 49.1 72 kDa type IV collagenase Mmp2 2 5.7 MOB kinase activator 1A Mob1a 2 8.3
Mas-related G-protein coupled receptor member F Mrgprf 4 15.1 Moesin Msn 16 18.9
Multivesicular body subunit 12A Mvb12a 3 12.5 Major vault protein Mvp 6 6
Myeloid-associated differentiation marker Myadm 6 13.8 UPF0160 protein MYG1, mitochondrial Myg1 3 10.5
Myosin-9 Myh9 14 6.8 Unconventional myosin-Ic Myo1c 9 9.2
Alpha-soluble NSF attachment protein Napa 2 8.8 Nck-associated protein 1 Nckap1 10 10.3
Nidogen-1 Nid1 5 7.1 Nucleoside diphosphate kinase A Nme1 6 32.9 Nucleoside diphosphate kinase B Nme2 5 26.3
GTPase NRas Nras 3 18.5 Polyadenylate-binding protein 1 Pabpc1 3 6.5
Multifunctional protein ADE2 Paics 4 12.2 Pantothenate kinase 3 Pank3 2 5.4 Protein deglycase DJ-1 Park7 2 11.6
Poly(rC)-binding protein 1 Pcbp1 3 7.9 Poly(rC)-binding protein 2 Pcbp2 3 9.1 Poly(rC)-binding protein 3 Pcbp3 2 5.4
Proliferating cell nuclear antigen Pcna 3 18 Procollagen C-endopeptidase enhancer 1 Pcolce 28 52.6
Programmed cell death protein 6 Pdcd6 3 14.1 Programmed cell death 6-interacting protein Pdcd6ip 29 38.2
Xaa-Pro dipeptidase Pepd 3 5.1 Profilin-1 Pfn1 4 30.7
Appendix
191
Phosphoglycerate mutase 1 Pgam1 6 26.8 Phosphoglycerate mutase 2 Pgam2 2 8.3 Phosphoglycerate kinase 1 Pgk1 9 28.3 Phosphoglycerate kinase 2 Pgk2 3 6
Pyruvate kinase PKM Pkm 21 40.6 Pleckstrin homology domain-containing family B member 2 Plekhb2 2 6.8
Plasminogen Plg 7 5.1 Phospholipid scramblase 1 Plscr1 4 7 Phospholipid scramblase 3 Plscr3 3 13.2
Serum paraoxonase/arylesterase 1 Pon1 3 6.7 Peptidyl-prolyl cis-trans isomerase A Ppia 12 53.7 Peptidyl-prolyl cis-trans isomerase B Ppib 5 26.4 Peptidyl-prolyl cis-trans isomerase C Ppic 2 11.8
Peroxiredoxin-1 Prdx1 4 20.1 Peroxiredoxin-2 Prdx2 2 14.6 Peroxiredoxin-6 Prdx6 2 15.2
cAMP-dependent protein kinase catalytic subunit alpha Prkaca 2 5.1 cAMP-dependent protein kinase type I-alpha regulatory subunit Prkar1a 2 5
Protein arginine N-methyltransferase 1 Prmt1 3 11.3 Phosphoserine aminotransferase Psat1 2 5.9 Proteasome subunit alpha type-1 Psma1 2 11.4 Proteasome subunit alpha type-3 Psma3 2 9 Proteasome subunit alpha type-7 Psma7 2 8.9 26S protease regulatory subunit 8 Psmc5 2 6.4
26S protease regulatory subunit 10B Psmc6 2 6.4 26S proteasome non-ATPase regulatory subunit 13 Psmd13 2 6.1 26S proteasome non-ATPase regulatory subunit 14 Psmd14 3 13.9 26S proteasome non-ATPase regulatory subunit 7 Psmd7 3 10.6
Prostaglandin F2 receptor negative regulator Ptgfrn 22 24.9 Pituitary tumor-transforming gene 1 protein-interacting protein Pttg1ip 3 12.1
Transcriptional activator protein Pur-alpha Pura 5 17.4 Transcriptional activator protein Pur-beta Purb 7 24.7
Ras-related protein Rab-10 Rab10 4 21.5 Ras-related protein Rab-11A Rab11a 3 14.4 Ras-related protein Rab-14 Rab14 3 16.7 Ras-related protein Rab-1A Rab1A 8 33.2 Ras-related protein Rab-1B Rab1b 7 35.8
Ras-related protein Rab-22A Rab22a 2 12.9 Ras-related protein Rab-2A Rab2a 3 16 Ras-related protein Rab-31 Rab31 2 12.9 Ras-related protein Rab-35 Rab35 2 11.4 Ras-related protein Rab-5A Rab5a 7 23.3 Ras-related protein Rab-5C Rab5c 5 19.9 Ras-related protein Rab-7a Rab7a 6 30 Ras-related protein Rab-8A Rab8a 5 22.7 Ras-related protein Rab-8B Rab8b 2 11.6
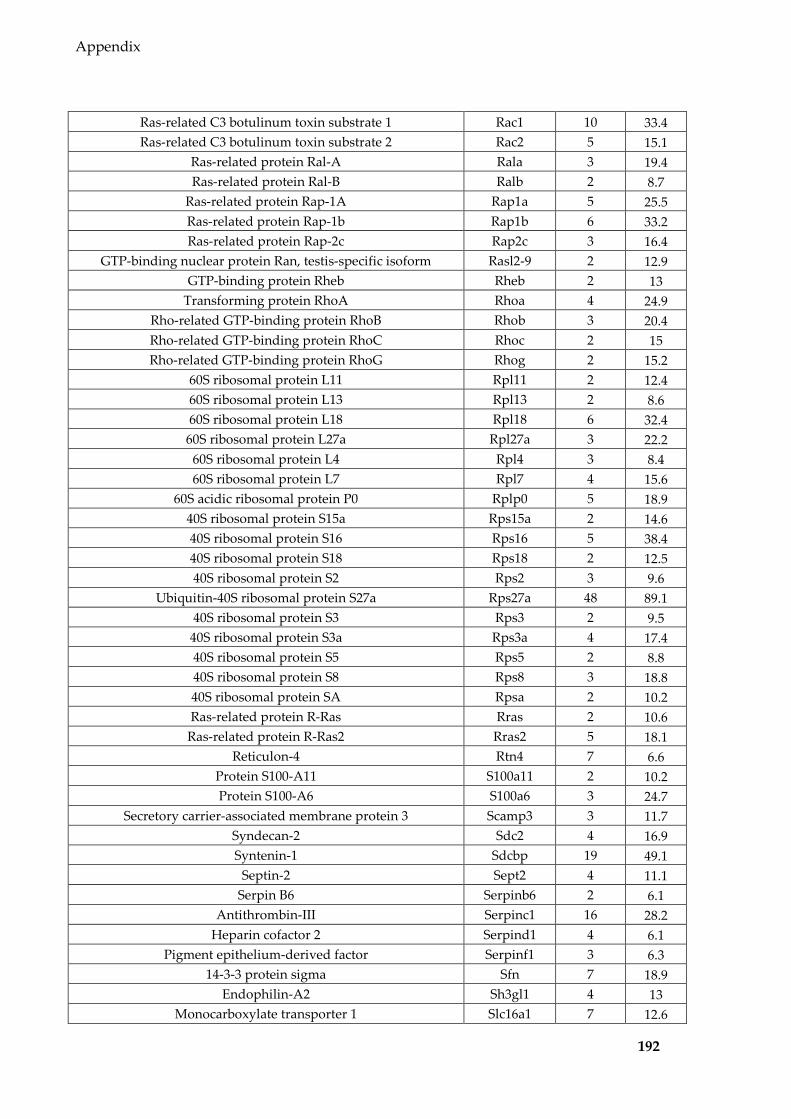
Appendix
192
Ras-related C3 botulinum toxin substrate 1 Rac1 10 33.4 Ras-related C3 botulinum toxin substrate 2 Rac2 5 15.1
Ras-related protein Ral-A Rala 3 19.4 Ras-related protein Ral-B Ralb 2 8.7
Ras-related protein Rap-1A Rap1a 5 25.5 Ras-related protein Rap-1b Rap1b 6 33.2 Ras-related protein Rap-2c Rap2c 3 16.4
GTP-binding nuclear protein Ran, testis-specific isoform Rasl2-9 2 12.9 GTP-binding protein Rheb Rheb 2 13
Transforming protein RhoA Rhoa 4 24.9 Rho-related GTP-binding protein RhoB Rhob 3 20.4 Rho-related GTP-binding protein RhoC Rhoc 2 15 Rho-related GTP-binding protein RhoG Rhog 2 15.2
60S ribosomal protein L11 Rpl11 2 12.4 60S ribosomal protein L13 Rpl13 2 8.6 60S ribosomal protein L18 Rpl18 6 32.4
60S ribosomal protein L27a Rpl27a 3 22.2 60S ribosomal protein L4 Rpl4 3 8.4 60S ribosomal protein L7 Rpl7 4 15.6
60S acidic ribosomal protein P0 Rplp0 5 18.9 40S ribosomal protein S15a Rps15a 2 14.6 40S ribosomal protein S16 Rps16 5 38.4 40S ribosomal protein S18 Rps18 2 12.5 40S ribosomal protein S2 Rps2 3 9.6
Ubiquitin-40S ribosomal protein S27a Rps27a 48 89.1 40S ribosomal protein S3 Rps3 2 9.5
40S ribosomal protein S3a Rps3a 4 17.4 40S ribosomal protein S5 Rps5 2 8.8 40S ribosomal protein S8 Rps8 3 18.8 40S ribosomal protein SA Rpsa 2 10.2 Ras-related protein R-Ras Rras 2 10.6
Ras-related protein R-Ras2 Rras2 5 18.1 Reticulon-4 Rtn4 7 6.6
Protein S100-A11 S100a11 2 10.2 Protein S100-A6 S100a6 3 24.7
Secretory carrier-associated membrane protein 3 Scamp3 3 11.7 Syndecan-2 Sdc2 4 16.9 Syntenin-1 Sdcbp 19 49.1
Septin-2 Sept2 4 11.1 Serpin B6 Serpinb6 2 6.1
Antithrombin-III Serpinc1 16 28.2 Heparin cofactor 2 Serpind1 4 6.1
Pigment epithelium-derived factor Serpinf1 3 6.3 14-3-3 protein sigma Sfn 7 18.9
Endophilin-A2 Sh3gl1 4 13 Monocarboxylate transporter 1 Slc16a1 7 12.6

Appendix
193
Neutral amino acid transporter A Slc1a4 3 6.8 Neutral amino acid transporter B(0) Slc1a5 5 15.2
Equilibrative nucleoside transporter 1 Slc29a1 5 11.3 Solute carrier family 2, facilitated glucose transporter member 1 Slc2a1 10 13.8
Sodium-coupled neutral amino acid transporter 2 Slc38a2 7 18.8 4F2 cell-surface antigen heavy chain Slc3a2 9 20.9
Large neutral amino acids transporter small subunit 1 Slc7a5 5 8 Acid sphingomyelinase-like phosphodiesterase 3b Smpdl3b 2 6.4
Sepiapterin reductase Spr 2 10.3 Spermidine synthase Srm 2 7
Erythrocyte band 7 integral membrane protein Stom 8 45 Serine-threonine kinase receptor-associated protein Strap 2 7.1
Syntaxin-4 Stx4 2 10.1 T-complex protein 1 subunit alpha Tcp1 5 10.6
Serotransferrin Tf 7 7.6 Transferrin receptor protein 1 Tfrc 4 7.1
Transforming growth factor-beta-induced protein ig-h3 Tgfbi 3 5.3 Thrombospondin-1 Thbs1 10 8.5 Thrombospondin-2 Thbs2 16 14.1
Metalloproteinase inhibitor 2 Timp2 2 13.6 Talin-1 Tln1 18 11.6
Tenascin Tnc 24 13 Tumor necrosis factor receptor superfamily member 12A Tnfrsf12a 3 32.6
Triosephosphate isomerase Tpi1 4 16.4 Tumor susceptibility gene 101 protein Tsg101 5 16.4
Tetraspanin-14 Tspan14 3 11.5 Tetraspanin-9 Tspan9 4 15.1
Protein tweety homolog 3 Ttyh3 2 5.2 Tubulin alpha-1A chain Tuba1a 14 43.2 Tubulin alpha-1B chain Tuba1b 6 17.7 Tubulin alpha-3 chain Tuba3a 10 31.4
Tubulin alpha-4A chain Tuba4a 8 25.4 Tubulin alpha-8 chain Tuba8 4 15.6 Tubulin beta-1 chain Tubb1 4 7.7
Tubulin beta-2A chain Tubb2a 8 21.6 Tubulin beta-3 chain Tubb3 4 8.4
Tubulin beta-4A chain Tubb4a 3 6.3 Tubulin beta-4B chain Tubb4b 7 18.2 Tubulin beta-5 chain Tubb5 11 26.5
Twinfilin-1 Twf1 2 5.2 Ubiquitin-conjugating enzyme E2 N Ube2n 2 12.5
Vasodilator-stimulated phosphoprotein Vasp 2 7.8 Synaptic vesicle membrane protein VAT-1 homolog Vat1 2 5.7
Vinculin Vcl 8 8.1 Transitional endoplasmic reticulum ATPase Vcp 31 37.1
Vacuolar protein sorting-associated protein 28 homolog Vps28 2 5.9

Appendix
194
Vacuolar protein-sorting-associated protein 36 Vps36 3 10.1 Vacuolar protein sorting-associated protein 37B Vps37b 2 8.4
Vitronectin Vtn 7 12.2 14-3-3 protein beta/alpha Ywhab 12 40.3
14-3-3 protein epsilon Ywhae 6 20 14-3-3 protein gamma Ywhag 13 45.4
14-3-3 protein eta Ywhah 11 38.2 14-3-3 protein theta Ywhaq 8 22.4
14-3-3 protein zeta/delta Ywhaz 14 51.9 Palmitoyltransferase ZDHHC5 Zdhhc5 2 5.6

Appendix
195
Table 8.2 miRNAs (normalised to per million miRNA reads and averaged for each group)
detected in Sertoli cells and EVs of different treatments
miRNA Untreated
cells Untreated
EVs DMSO
cells DMSO
EVs BPA cells BPA EVs
mmu-let-7a-1-3p 210 215 202 34 101 26 mmu-let-7a-5p 15,228 16,520 14,092 13,557 18,034 18,346 mmu-let-7b-3p 168 132 119 31 71 9 mmu-let-7b-5p 25,076 10,879 25,376 12,649 27,299 13,518
mmu-let-7c-2-3p 623 436 553 122 304 103 mmu-let-7c-5p 36,863 25,346 36,308 27,095 39,217 26,645 mmu-let-7d-3p 794 867 722 249 502 530 mmu-let-7d-5p 5,876 4,524 6,102 4,001 5,265 6,306 mmu-let-7e-5p 3,290 2,428 3,097 3,239 3,611 3,008 mmu-let-7f-5p 39,601 45,670 32,253 53,992 57,577 57,240 mmu-let-7g-5p 27,949 23,650 22,394 27,158 30,568 32,438 mmu-let-7i-5p 49,664 32,694 35,843 32,937 40,419 39,808
mmu-miR-100-5p 9,248 11,447 8,052 7,887 9,288 4,462 mmu-miR-101a-3p 239 2,156 447 1,009 483 2,105 mmu-miR-101b-3p 3,792 4,774 4,512 7,858 5,342 7,323 mmu-miR-103-3p 1,266 2,991 2,006 3,224 1,831 3,350
mmu-miR-106b-3p 1,409 1,845 938 1,979 1,022 2,014 mmu-miR-106b-5p 282 461 493 569 384 349 mmu-miR-107-3p 47 58 65 99 77 133 mmu-miR-10a-3p 137 160 141 161 165 154 mmu-miR-10a-5p 38,089 32,977 38,820 37,129 41,167 27,264 mmu-miR-10b-5p 15,881 17,824 16,955 16,634 19,928 13,211 mmu-miR-1198-5p 79 101 103 55 83 34 mmu-miR-122-5p 105 5,087 69 306 298 5,861
mmu-miR-125a-3p 137 202 147 87 141 39 mmu-miR-125a-5p 3,632 3,741 5,462 1,431 2,712 1,105
mmu-miR-125b-1-3p 1,583 1,092 926 552 785 507 mmu-miR-125b-5p 9,821 10,552 15,807 4,823 5,586 3,122 mmu-miR-126a-3p 65 2,776 107 193 180 1,633 mmu-miR-127-3p 0 324 0 6 9 60 mmu-miR-128-3p 754 1,613 675 160 488 637 mmu-miR-1291 3,049 51 2,780 17 3,417 15
mmu-miR-130a-3p 202 60 346 244 243 190 mmu-miR-133a-3p 2 394 0 29 1 330 mmu-miR-140-3p 3,726 5,567 4,937 8,375 5,541 6,812 mmu-miR-140-5p 725 769 1,302 1,275 898 906
mmu-miR-142a-5p 11 560 4 26 49 1,281 mmu-miR-143-3p 37,951 34,426 29,631 39,505 87,218 103,998 mmu-miR-143-5p 100 188 109 114 168 193 mmu-miR-144-3p 9 2,617 2 29 27 342
mmu-miR-145a-3p 657 320 586 210 976 304

Appendix
196
mmu-miR-145a-5p 871 283 894 478 1,982 861 mmu-miR-146a-5p 944 551 1,051 932 430 1,516 mmu-miR-146b-5p 309 282 271 339 269 380 mmu-miR-148a-3p 5,024 7,144 5,401 10,139 14,632 14,384 mmu-miR-148a-5p 72 16 74 26 204 78 mmu-miR-148b-3p 1,371 1,755 1,163 2,147 1,728 2,744 mmu-miR-149-5p 318 191 228 94 120 104 mmu-miR-151-3p 1,621 2,502 1,764 1,339 1,349 1,858 mmu-miR-151-5p 85 62 107 145 135 52 mmu-miR-152-3p 1,884 1,995 2,109 3,720 3,151 3,066 mmu-miR-155-5p 311 302 309 354 231 362 mmu-miR-15a-5p 52 28 83 51 71 83 mmu-miR-15b-3p 98 147 95 65 118 51 mmu-miR-15b-5p 116 250 256 509 193 532 mmu-miR-16-1-3p 390 144 299 144 242 74 mmu-miR-16-5p 649 676 1,106 2,415 896 1,883 mmu-miR-17-5p 1,057 2,053 1,699 2,719 1,252 2,197
mmu-miR-181a-2-3p 170 288 161 153 95 56 mmu-miR-181a-5p 1,474 1,368 2,251 1,241 1,435 1,073 mmu-miR-181b-5p 1,516 1,239 2,100 1,378 1,359 1,095 mmu-miR-182-5p 8,368 11,992 10,659 17,356 8,146 12,533 mmu-miR-183-5p 5,441 5,907 5,158 7,751 5,002 6,753
mmu-miR-1839-5p 527 624 470 766 594 797 mmu-miR-185-5p 144 619 234 265 255 974 mmu-miR-186-5p 1,370 1,821 1,367 1,363 1,450 1,896 mmu-miR-18a-5p 119 91 212 114 143 72 mmu-miR-191-5p 1,457 2,917 1,534 1,023 1,663 3,698 mmu-miR-192-5p 182 1,159 152 246 167 579
mmu-miR-193a-3p 86 17 76 91 329 108 mmu-miR-193a-5p 67 44 111 24 86 28 mmu-miR-193b-3p 106 56 83 100 344 399
mmu-miR-1949 456 13 495 13 413 104 mmu-miR-195a-5p 44 44 95 131 46 32 mmu-miR-196a-5p 414 455 657 569 651 435 mmu-miR-196b-5p 387 412 921 1,078 586 902 mmu-miR-199a-3p 9,002 13,409 12,002 19,495 12,894 14,057 mmu-miR-199a-5p 8,600 5,824 9,329 7,032 9,358 6,084 mmu-miR-199b-3p 6,282 9,621 8,380 13,391 8,839 8,707 mmu-miR-199b-5p 2,624 1,288 2,684 2,330 2,324 1,728 mmu-miR-19a-3p 104 98 169 104 117 238 mmu-miR-19b-3p 372 262 693 520 478 565 mmu-miR-1a-3p 43 3,469 5 197 19 1,048
mmu-miR-206-3p 204 121 152 244 73 69 mmu-miR-20a-5p 5,129 7,578 7,182 9,062 5,944 8,435 mmu-miR-210-3p 396 258 2,291 1,001 949 463 mmu-miR-2137 167 1,717 597 462 374 1,490
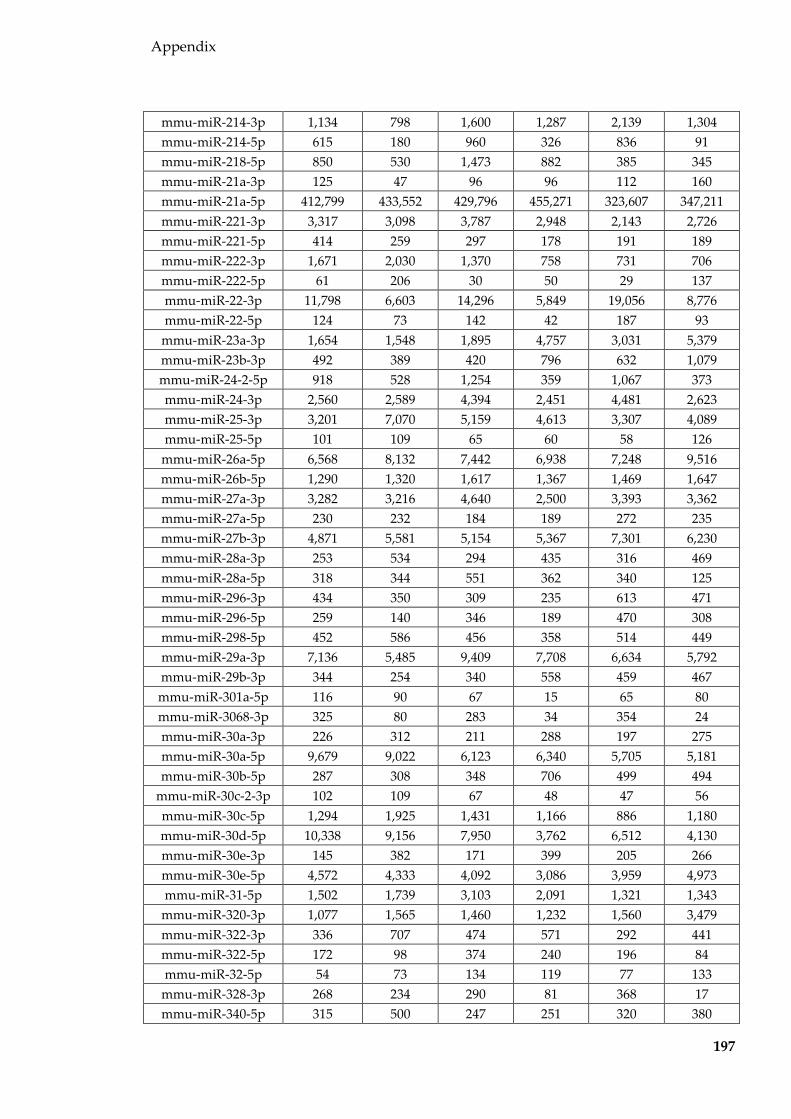
Appendix
197
mmu-miR-214-3p 1,134 798 1,600 1,287 2,139 1,304 mmu-miR-214-5p 615 180 960 326 836 91 mmu-miR-218-5p 850 530 1,473 882 385 345 mmu-miR-21a-3p 125 47 96 96 112 160 mmu-miR-21a-5p 412,799 433,552 429,796 455,271 323,607 347,211 mmu-miR-221-3p 3,317 3,098 3,787 2,948 2,143 2,726 mmu-miR-221-5p 414 259 297 178 191 189 mmu-miR-222-3p 1,671 2,030 1,370 758 731 706 mmu-miR-222-5p 61 206 30 50 29 137 mmu-miR-22-3p 11,798 6,603 14,296 5,849 19,056 8,776 mmu-miR-22-5p 124 73 142 42 187 93
mmu-miR-23a-3p 1,654 1,548 1,895 4,757 3,031 5,379 mmu-miR-23b-3p 492 389 420 796 632 1,079 mmu-miR-24-2-5p 918 528 1,254 359 1,067 373 mmu-miR-24-3p 2,560 2,589 4,394 2,451 4,481 2,623 mmu-miR-25-3p 3,201 7,070 5,159 4,613 3,307 4,089 mmu-miR-25-5p 101 109 65 60 58 126
mmu-miR-26a-5p 6,568 8,132 7,442 6,938 7,248 9,516 mmu-miR-26b-5p 1,290 1,320 1,617 1,367 1,469 1,647 mmu-miR-27a-3p 3,282 3,216 4,640 2,500 3,393 3,362 mmu-miR-27a-5p 230 232 184 189 272 235 mmu-miR-27b-3p 4,871 5,581 5,154 5,367 7,301 6,230 mmu-miR-28a-3p 253 534 294 435 316 469 mmu-miR-28a-5p 318 344 551 362 340 125 mmu-miR-296-3p 434 350 309 235 613 471 mmu-miR-296-5p 259 140 346 189 470 308 mmu-miR-298-5p 452 586 456 358 514 449 mmu-miR-29a-3p 7,136 5,485 9,409 7,708 6,634 5,792 mmu-miR-29b-3p 344 254 340 558 459 467 mmu-miR-301a-5p 116 90 67 15 65 80 mmu-miR-3068-3p 325 80 283 34 354 24 mmu-miR-30a-3p 226 312 211 288 197 275 mmu-miR-30a-5p 9,679 9,022 6,123 6,340 5,705 5,181 mmu-miR-30b-5p 287 308 348 706 499 494
mmu-miR-30c-2-3p 102 109 67 48 47 56 mmu-miR-30c-5p 1,294 1,925 1,431 1,166 886 1,180 mmu-miR-30d-5p 10,338 9,156 7,950 3,762 6,512 4,130 mmu-miR-30e-3p 145 382 171 399 205 266 mmu-miR-30e-5p 4,572 4,333 4,092 3,086 3,959 4,973 mmu-miR-31-5p 1,502 1,739 3,103 2,091 1,321 1,343
mmu-miR-320-3p 1,077 1,565 1,460 1,232 1,560 3,479 mmu-miR-322-3p 336 707 474 571 292 441 mmu-miR-322-5p 172 98 374 240 196 84 mmu-miR-32-5p 54 73 134 119 77 133
mmu-miR-328-3p 268 234 290 81 368 17 mmu-miR-340-5p 315 500 247 251 320 380

Appendix
198
mmu-miR-342-3p 111 92 93 113 82 99 mmu-miR-345-3p 52 157 53 117 84 138 mmu-miR-34c-5p 5,798 2,619 4,861 2,741 5,334 2,485 mmu-miR-350-3p 68 55 83 74 82 50 mmu-miR-351-5p 844 778 705 269 560 368 mmu-miR-3535 3,786 158 2,039 44 2,682 148
mmu-miR-361-3p 529 337 444 98 403 97 mmu-miR-361-5p 67 187 103 123 109 108 mmu-miR-362-3p 61 96 72 213 95 145 mmu-miR-362-5p 472 310 584 370 471 481
mmu-miR-365-2-5p 34 36 32 87 29 92 mmu-miR-365-3p 296 132 232 91 316 96
mmu-miR-374b-5p 184 229 250 679 263 532 mmu-miR-378a-3p 3,022 2,487 3,276 3,345 5,527 5,099
mmu-miR-378c 144 166 180 158 222 208 mmu-miR-423-3p 4,695 5,018 4,052 2,267 4,348 2,457 mmu-miR-423-5p 983 1,020 770 547 1,020 1,770 mmu-miR-425-5p 122 177 117 65 157 46
mmu-miR-450a-5p 570 476 584 161 253 139 mmu-miR-450b-5p 82 76 72 58 52 189
mmu-miR-451a 214 3,016 7 77 1,245 18,107 mmu-miR-467a-5p 101 50 136 154 214 133 mmu-miR-467c-5p 31 44 77 53 92 46 mmu-miR-467e-5p 48 11 69 46 108 96
mmu-miR-484 1,343 957 1,336 176 831 408 mmu-miR-486a-5p 93 7,034 65 155 221 1,967 mmu-miR-486b-5p 32 3,014 26 67 92 1,363 mmu-miR-497a-5p 56 89 126 46 62 15 mmu-miR-501-3p 230 221 241 172 242 251 mmu-miR-503-5p 340 313 442 588 324 290 mmu-miR-5099 8,713 503 6,941 168 5,692 130 mmu-miR-5112 0 13 0 119 1 930 mmu-miR-5126 17 76 90 9 158 263
mmu-miR-532-5p 4,264 4,152 2,972 2,414 3,715 3,003 mmu-miR-542-3p 324 278 394 431 299 151 mmu-miR-574-3p 51 92 91 104 117 57 mmu-miR-598-3p 87 105 91 69 68 95 mmu-miR-615-3p 1,139 631 919 91 897 291 mmu-miR-6238 36 259 60 133 81 208
mmu-miR-6516-3p 114 17 155 24 122 0 mmu-miR-652-3p 377 169 419 463 648 625 mmu-miR-6538 144 129 143 22 81 140
mmu-miR-669a-5p 102 30 97 35 88 34 mmu-miR-669c-5p 150 93 194 167 343 273 mmu-miR-671-5p 39 37 102 74 92 100 mmu-miR-674-3p 215 245 327 164 168 156
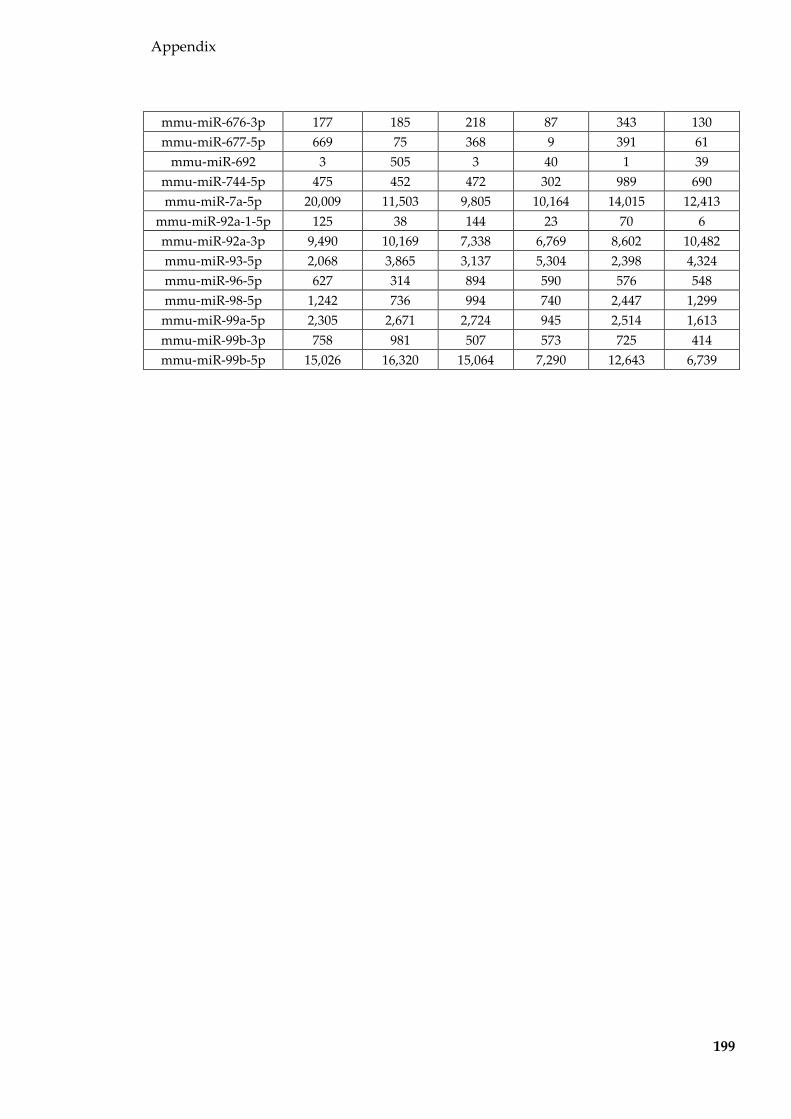
Appendix
199
mmu-miR-676-3p 177 185 218 87 343 130 mmu-miR-677-5p 669 75 368 9 391 61
mmu-miR-692 3 505 3 40 1 39 mmu-miR-744-5p 475 452 472 302 989 690 mmu-miR-7a-5p 20,009 11,503 9,805 10,164 14,015 12,413
mmu-miR-92a-1-5p 125 38 144 23 70 6 mmu-miR-92a-3p 9,490 10,169 7,338 6,769 8,602 10,482 mmu-miR-93-5p 2,068 3,865 3,137 5,304 2,398 4,324 mmu-miR-96-5p 627 314 894 590 576 548 mmu-miR-98-5p 1,242 736 994 740 2,447 1,299
mmu-miR-99a-5p 2,305 2,671 2,724 945 2,514 1,613 mmu-miR-99b-3p 758 981 507 573 725 414 mmu-miR-99b-5p 15,026 16,320 15,064 7,290 12,643 6,739

Appendix
200
Table 8.3 tRFs (normalised to per million tRF reads and averaged for each group) detected
in Sertoli cells and EVs of different treatments
tRF Untreated cells Untreated EVs DMSO cells DMSO EVs BPA cells BPA EVs tRF-Ala-AGC 547 2,678 713 1,556 429 1,652 tRF-Ala-CGC 868 2,579 766 1,342 580 1,663 tRF-Ala-TGC 1,797 2,163 1,311 1,678 447 2,400 tRF-Arg-ACG 4,097 1,596 3,850 573 1,077 430 tRF-Arg-CCG 1,819 1,928 2,061 870 1,148 611 tRF-Arg-CCT 3,854 2,310 4,518 1,400 1,663 1,693 tRF-Arg-TCG 692 516 490 201 301 138 tRF-Arg-TCT 1,618 785 2,494 489 931 294 tRF-Asn-GTT 1,030 1,271 987 863 464 499 tRF-Asp-GTC 8,683 14,879 10,466 14,749 6,654 11,109 tRF-Cys-GCA 5,396 4,844 4,048 4,453 3,322 6,560 tRF-Gln-CTG 5,133 8,173 7,709 4,308 4,020 3,018 tRF-Gln-TTG 2,005 2,507 3,043 1,715 2,073 1,205 tRF-Glu-CTC 223,412 280,888 219,763 159,593 212,231 149,087 tRF-Glu-TTC 24,943 87,904 27,078 115,008 26,811 87,111 tRF-Gly-ACC 194 261 244 221 131 624 tRF-Gly-CCC 50,339 57,406 43,972 71,186 58,006 85,093 tRF-Gly-GCC 283,523 359,889 240,385 455,237 329,616 523,987 tRF-Gly-TCC 1,160 1,147 1,635 1,665 786 1,217 tRF-His-ATG 148 69 121 28 32 12 tRF-His-GTG 98,797 59,947 75,178 80,537 100,754 46,893 tRF-Ile-AAT 7,107 1,659 9,556 833 2,889 1,266 tRF-Ile-TAT 289 403 330 150 163 173
tRF-Leu-AAG 742 852 681 665 453 813 tRF-Leu-CAA 20,952 1,755 18,698 603 6,916 627 tRF-Leu-CAG 19,662 2,440 14,783 1,065 6,281 846 tRF-Leu-TAA 5,054 2,318 4,891 1,104 1,371 1,284 tRF-Leu-TAG 581 505 631 338 335 474 tRF-Lys-CTT 138,086 33,563 185,816 18,649 161,705 17,355 tRF-Lys-TTT 20,217 10,321 27,989 12,442 24,434 14,827 tRF-Met-CAT 17,355 3,785 19,686 2,404 9,695 1,959 tRF-Phe-GAA 454 306 301 248 178 316 tRF-Pro-AGG 2,160 2,652 2,226 1,742 823 1,445 tRF-Pro-CGG 1,499 1,593 1,287 1,124 512 835 tRF-Pro-TGG 2,413 3,305 2,324 2,244 976 1,953 tRF-Ser-AGA 3,018 4,685 3,850 2,941 2,596 1,450 tRF-Ser-CGA 210 210 281 111 152 150 tRF-Ser-GCT 1,682 1,371 1,794 732 888 546 tRF-Ser-GGA 2 87 2 67 8 125 tRF-Ser-TGA 639 1,288 859 842 692 468
tRF-STOP-TTA 26 21 28 11 10 15 tRF-STOP-TCA 713 1,260 935 728 816 841
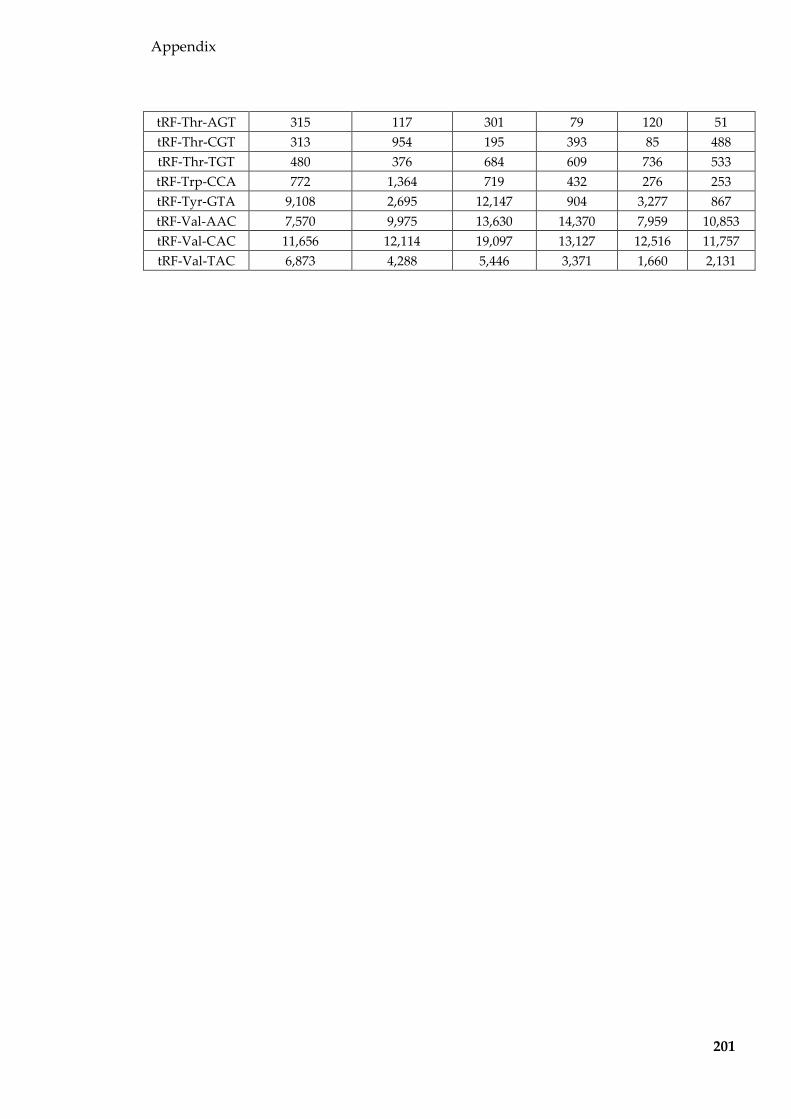
Appendix
201
tRF-Thr-AGT 315 117 301 79 120 51 tRF-Thr-CGT 313 954 195 393 85 488 tRF-Thr-TGT 480 376 684 609 736 533 tRF-Trp-CCA 772 1,364 719 432 276 253 tRF-Tyr-GTA 9,108 2,695 12,147 904 3,277 867 tRF-Val-AAC 7,570 9,975 13,630 14,370 7,959 10,853 tRF-Val-CAC 11,656 12,114 19,097 13,127 12,516 11,757 tRF-Val-TAC 6,873 4,288 5,446 3,371 1,660 2,131

Appendix
202
Table 8.4 Proteins identified in DC2 epididymal EVs
Protein Gene
symbol
Unique matching peptides
% protein covered
14-3-3 protein beta/alpha Ywhab 6 18.7 14-3-3 protein epsilon Ywhae 2 7.1 14-3-3 protein gamma Ywhag 6 31.6
14-3-3 protein zeta/delta Ywhaz 8 33.9 4F2 cell-surface antigen heavy chain Slc3a2 2 7.2
60S ribosomal protein L18 Rpl18 3 11.2 78 kDa glucose-regulated protein Hspa5 2 5
Acid sphingomyelinase-like phosphodiesterase 3b Smpdl3b 2 10.1 Actin, alpha cardiac muscle 1 Actc1 24 28.6 Actin, alpha skeletal muscle Acta1 24 28.6 Actin, aortic smooth muscle Acta2 24 28.6
Actin, cytoplasmic 1 Actb 25 34.1 Actin, cytoplasmic 2 Actg1 25 34.1
Actin, gamma-enteric smooth muscle Actg2 24 28.7 Alpha-enolase Eno1 2 6.2
Annexin A1 Anxa1 13 35 Annexin A2 Anxa2 13 29.5 Annexin A3 Anxa3 5 15.2 Annexin A4 Anxa4 9 32 Annexin A5 Anxa5 12 32.9 Annexin A6 Anxa6 11 26.2
Antithrombin-III Serpinc1 2 8.2 Apolipoprotein A-I Apoa1 2 6.4 Apolipoprotein M Apom 5 15.3
Basement membrane-specific heparan sulfate proteoglycan core protein
Hspg2 79 20.3
CD151 antigen Cd151 3 18.2 CD63 antigen Cd63 4 12.2 CD82 antigen Cd82 2 7.9 CD9 antigen Cd9 4 18.6
Charged multivesicular body protein 4b Chmp4b 4 14.7 Clathrin heavy chain 1 Cltc 10 8.1 Clathrin light chain A Clta 2 10.6 Coagulation factor X F10 6 6.4
Cofilin-1 Cfl1 3 17.5 Complement C1q tumor necrosis factor-related protein
3 C1qtnf3 4 5.3
Complement C3 C3 7 6.9 CUB and zona pellucida-like domain-containing
protein 1 Cuzd1 18 24.4
Disco-interacting protein 2 homolog B Dip2b 6 6.4 Disintegrin and metalloproteinase domain-containing Adam10 2 5.7

Appendix
203
protein 10 Elongation factor 1-alpha 1 Eef1a1 7 17.3
Elongation factor 2 Eef2 3 6.9 EMILIN-2 Emilin2 2 6.1
Endoplasmin Hsp90b1 7 9.7 Erythrocyte band 7 integral membrane protein Stom 3 11.3
Ferritin light chain 1 Ftl1 4 39.3 Fibronectin Fn1 35 16.6
Fibulin-2 Fbln2 13 10.5 Fructose-bisphosphate aldolase A Aldoa 3 16.2
Glutamate dehydrogenase 1, mitochondrial Glud1 6 8.6 Glutathione S-transferase Mu 1 Gstm1 3 12.4
Glyceraldehyde-3-phosphate dehydrogenase Gapdh 8 21.6 Golgi-associated plant pathogenesis-related protein 1 Glipr2 4 20.8
GTP-binding nuclear protein Ran Ran 4 21.3 Guanine nucleotide-binding protein G(k) subunit
alpha Gnai3 3 12.1
Heat shock 70 kDa protein 1A Hspa1a 5 6.9 Heat shock 70 kDa protein 1B Hspa1b 5 6.9
Heat shock 70 kDa protein 1-like Hspa1l 5 12.8 Heat shock cognate 71 kDa protein Hspa8 18 30.3 Heat shock protein HSP 90-alpha Hsp90aa1 13 19 Heat shock protein HSP 90-beta Hsp90ab1 12 21.5
Hemoglobin subunit alpha Hba 9 16.9 Hemoglobin subunit beta-1 Hbb-b1 3 6.8 Hemoglobin subunit beta-2 Hbb-b2 3 6.8
Hemoglobin subunit epsilon-Y2 Hbb-y 3 6.8 High mobility group protein B1 Hmgb1 5 26
Histone H2A type 1 Hist1h2ab 6 16.2 Histone H2A type 1-F Hist1h2af 6 16.2 Histone H2A type 1-H Hist1h2ah 6 16.4 Histone H2A type 1-K Hist1h2ak 6 16.2 Histone H2A type 2-A Hist2h2aa1 6 16.9 Histone H2A type 2-B Hist2h2ab 6 23.1 Histone H2A type 2-C Hist2h2ac 6 17.1
Histone H2A type 3 Hist3h2a 6 16.2 Histone H2A.J H2afj 6 17.1 Histone H2AX H2afx 6 31.5
Histone H2B type 1-B Hist1h2bb 20 61.1 Histone H2B type 1-C/E/G Hist1h2bc 21 61.1 Histone H2B type 1-F/J/L Hist1h2bf 20 61.1
Histone H2B type 1-H Hist1h2bh 21 61.1 Histone H2B type 1-K Hist1h2bk 20 61.1 Histone H2B type 1-M Hist1h2bm 21 61.1 Histone H2B type 1-P Hist1h2bp 20 61.1 Histone H2B type 2-B Hist2h2bb 21 61.1

Appendix
204
Histone H3.1 Hist1h3a 15 38.2 Histone H3.1 H3110 15 38.2 Histone H3.2 Hist1h3b 15 38.2 Histone H3.2 H312 15 38.2 Histone H3.3 H3f3a 15 38.2
Histone H3.3C H3f3c 15 38.2 Histone H4 Hist1h4a 9 46.6
Integrin alpha-3 Itga3 4 6.6 Integrin beta-1 Itgb1 9 13.5
Inter-alpha-trypsin inhibitor heavy chain H2 Itih2 8 7.4 Junction plakoglobin Jup 3 9.4
Keratin, type I cuticular Ha4 Krt34 3 10.7 Keratin, type I cuticular Ha5 Krt35 7 7.7 Keratin, type I cytoskeletal 10 Krt10 13 10.5 Keratin, type I cytoskeletal 14 Krt14 12 12.8 Keratin, type I cytoskeletal 42 Krt42 11 14.2 Keratin, type II cuticular Hb4 Krt84 2 7.6 Keratin, type II cuticular Hb5 Krt85 10 19.5 Keratin, type II cytoskeletal 1 Krt1 5 6
Keratin, type II cytoskeletal 2 epidermal Krt2 3 5.9 Keratin, type II cytoskeletal 2 oral Krt76 5 7.6
Keratin, type II cytoskeletal 5 Krt5 5 14.7 Keratin, type II cytoskeletal 6A Krt6a 7 17.9 Keratin, type II cytoskeletal 73 Krt73 2 5 Keratin, type II cytoskeletal 75 Krt75 5 13.2
Lactadherin Mfge8 31 37.1 L-lactate dehydrogenase A chain Ldha 5 19.6 L-lactate dehydrogenase B chain Ldhb 4 11.7
Major vault protein Mvp 5 10.8 MARCKS-related protein Marcksl1 2 14
Moesin Msn 4 12.3 Monocarboxylate transporter 1 Slc16a1 4 10.5
Myosin-8 Myh8 4 5.8 Nidogen-1 Nid1 7 8.6
Nucleoside diphosphate kinase B Nme2 2 19.7 Peptidyl-prolyl cis-trans isomerase A Ppia 2 7.9
Peroxiredoxin-1 Prdx1 4 14.1 Phosphoglycerate mutase 1 Pgam1 3 11.4 Phosphoglycerate mutase 2 Pgam2 3 12.3
Pigment epithelium-derived factor Serpinf1 3 5.8 Pituitary tumor-transforming gene 1 protein-
interacting protein Pttg1ip 4 34.5
Plakophilin-1 Pkp1 2 7.3 Polyubiquitin-B Ubb 28 17 Polyubiquitin-C Ubc 28 7.1 Prelamin-A/C Lmna 2 5.1
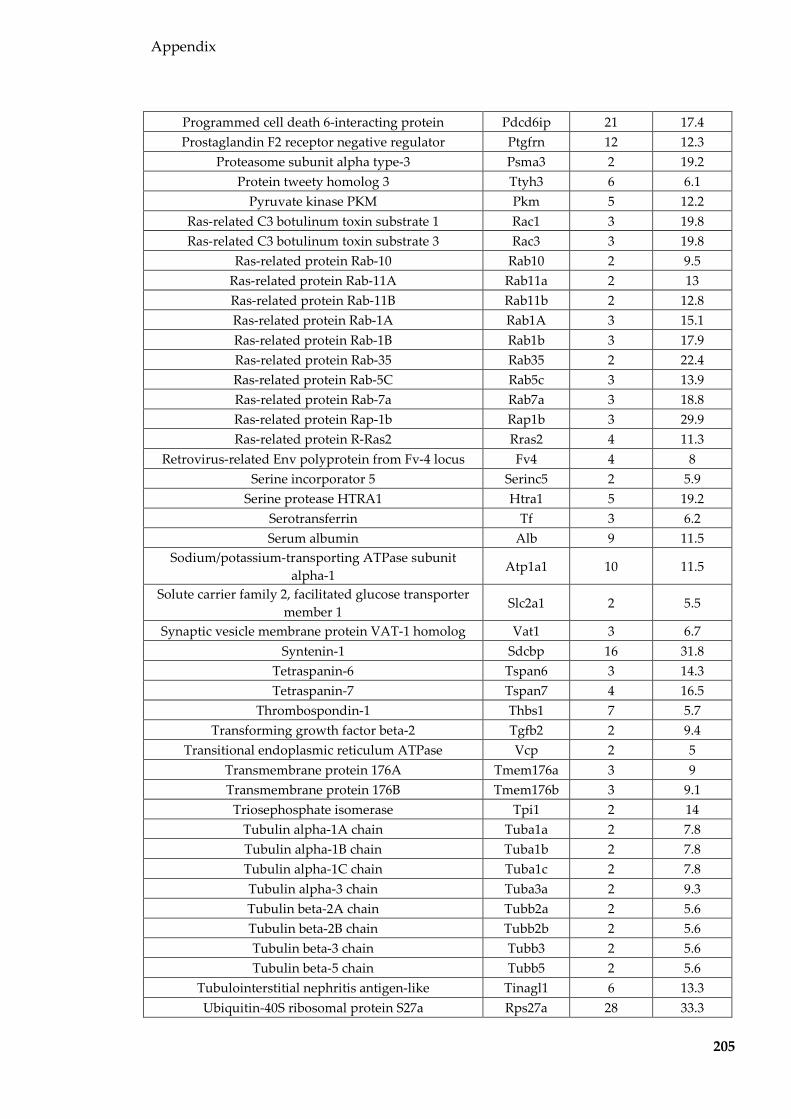
Appendix
205
Programmed cell death 6-interacting protein Pdcd6ip 21 17.4 Prostaglandin F2 receptor negative regulator Ptgfrn 12 12.3
Proteasome subunit alpha type-3 Psma3 2 19.2 Protein tweety homolog 3 Ttyh3 6 6.1
Pyruvate kinase PKM Pkm 5 12.2 Ras-related C3 botulinum toxin substrate 1 Rac1 3 19.8 Ras-related C3 botulinum toxin substrate 3 Rac3 3 19.8
Ras-related protein Rab-10 Rab10 2 9.5 Ras-related protein Rab-11A Rab11a 2 13 Ras-related protein Rab-11B Rab11b 2 12.8 Ras-related protein Rab-1A Rab1A 3 15.1 Ras-related protein Rab-1B Rab1b 3 17.9 Ras-related protein Rab-35 Rab35 2 22.4 Ras-related protein Rab-5C Rab5c 3 13.9 Ras-related protein Rab-7a Rab7a 3 18.8 Ras-related protein Rap-1b Rap1b 3 29.9 Ras-related protein R-Ras2 Rras2 4 11.3
Retrovirus-related Env polyprotein from Fv-4 locus Fv4 4 8 Serine incorporator 5 Serinc5 2 5.9
Serine protease HTRA1 Htra1 5 19.2 Serotransferrin Tf 3 6.2 Serum albumin Alb 9 11.5
Sodium/potassium-transporting ATPase subunit alpha-1
Atp1a1 10 11.5
Solute carrier family 2, facilitated glucose transporter member 1
Slc2a1 2 5.5
Synaptic vesicle membrane protein VAT-1 homolog Vat1 3 6.7 Syntenin-1 Sdcbp 16 31.8
Tetraspanin-6 Tspan6 3 14.3 Tetraspanin-7 Tspan7 4 16.5
Thrombospondin-1 Thbs1 7 5.7 Transforming growth factor beta-2 Tgfb2 2 9.4
Transitional endoplasmic reticulum ATPase Vcp 2 5 Transmembrane protein 176A Tmem176a 3 9 Transmembrane protein 176B Tmem176b 3 9.1
Triosephosphate isomerase Tpi1 2 14 Tubulin alpha-1A chain Tuba1a 2 7.8 Tubulin alpha-1B chain Tuba1b 2 7.8 Tubulin alpha-1C chain Tuba1c 2 7.8 Tubulin alpha-3 chain Tuba3a 2 9.3 Tubulin beta-2A chain Tubb2a 2 5.6 Tubulin beta-2B chain Tubb2b 2 5.6 Tubulin beta-3 chain Tubb3 2 5.6 Tubulin beta-5 chain Tubb5 2 5.6
Tubulointerstitial nephritis antigen-like Tinagl1 6 13.3 Ubiquitin-40S ribosomal protein S27a Rps27a 28 33.3
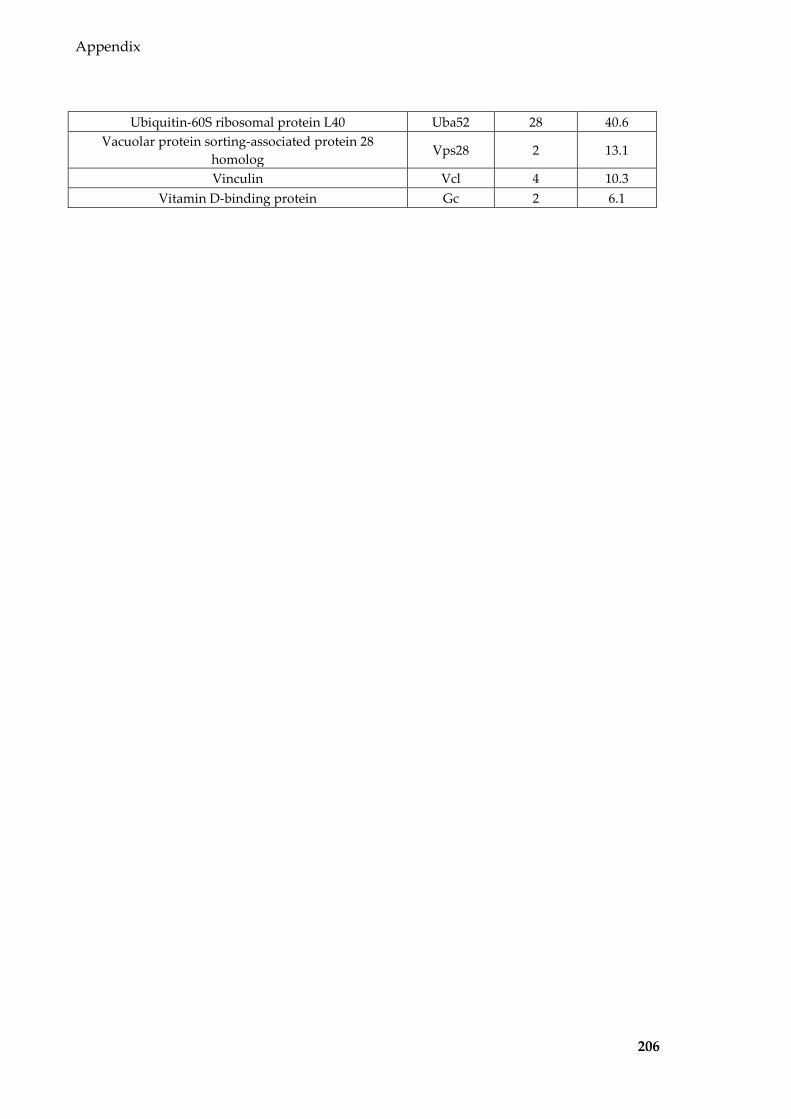
Appendix
206
Ubiquitin-60S ribosomal protein L40 Uba52 28 40.6 Vacuolar protein sorting-associated protein 28
homolog Vps28 2 13.1
Vinculin Vcl 4 10.3 Vitamin D-binding protein Gc 2 6.1

Appendix
207
Table 8.5 miRNAs (normalised to per million miRNA reads) detected in DC2 epididymal
cells and EVs of different treatments
miRNA HFL cells HFL EVs LF cells LF EVs HFS cells HFS EVs mmu-let-7a-5p 12,043 18,970 9,598 32,839 12,669 37,271 mmu-let-7b-3p 60 20 130 203 105 215 mmu-let-7b-5p 39,472 10,732 22,708 81,190 18,722 90,329
mmu-let-7c-2-3p 122 31 80 17 98 23 mmu-let-7c-5p 45,245 41,156 72,452 160,593 63,348 166,059 mmu-let-7d-3p 250 736 606 989 392 1,448 mmu-let-7d-5p 4,121 1,827 2,341 2,638 2,608 4,216 mmu-let-7e-5p 2,650 688 1,777 11,839 1,692 11,506 mmu-let-7f-5p 82,366 108,805 54,586 32,527 63,438 34,433 mmu-let-7g-5p 38,458 47,135 17,535 4,124 26,828 6,096 mmu-let-7i-5p 205,243 155,524 107,098 39,430 121,334 42,668
mmu-let-7j 124 88 70 35 83 36 mmu-miR-100-5p 1,425 3,469 6,891 14,396 6,943 12,070
mmu-miR-101a-3p 1,083 2,596 2,255 256 1,914 225 mmu-miR-101b-3p 6,384 8,766 6,217 543 4,860 441 mmu-miR-103-3p 2,349 5,761 1,563 260 2,163 405
mmu-miR-106b-3p 7,904 7,653 7,609 4,732 6,218 3,977 mmu-miR-106b-5p 727 295 346 24 546 23 mmu-miR-10a-3p 206 155 117 40 223 130 mmu-miR-10a-5p 92,573 106,049 203,908 237,750 150,085 218,380 mmu-miR-10b-5p 21,375 40,099 44,436 45,395 31,848 41,230 mmu-miR-1198-5p 50 236 92 714 111 867 mmu-miR-122-5p 73 504 1 562 1 416
mmu-miR-1249-3p 82 0 33 70 32 145 mmu-miR-125a-3p 67 40 30 377 28 530 mmu-miR-125a-5p 1,458 5,151 1,205 7,423 1,964 10,290
mmu-miR-125b-1-3p 426 394 116 454 127 483 mmu-miR-125b-2-3p 269 85 61 13 87 12 mmu-miR-125b-5p 1,575 2,580 1,526 3,024 1,912 3,960 mmu-miR-126a-3p 38 1,845 39 260 79 339 mmu-miR-128-3p 772 794 316 1,056 462 1,384 mmu-miR-1291 408 2 8 5 20 2
mmu-miR-1306-3p 105 49 12 85 19 84 mmu-miR-130a-3p 120 43 59 19 207 8 mmu-miR-140-3p 3,551 3,903 2,416 524 1,973 564 mmu-miR-140-5p 41 160 24 0 46 9
mmu-miR-142a-5p 0 160 0 3 0 4 mmu-miR-143-3p 125 2,159 413 187 276 179
mmu-miR-146a-5p 255 166 2,333 605 4,607 1,046 mmu-miR-146b-5p 735 835 797 705 756 545 mmu-miR-148a-3p 1,598 4,094 14,491 8,980 8,449 4,425 mmu-miR-148b-3p 3,568 2,420 5,322 785 4,482 718

Appendix
208
mmu-miR-151-3p 8,828 12,433 9,936 19,443 7,320 15,929 mmu-miR-151-5p 175 45 56 0 82 28 mmu-miR-152-3p 2,351 3,385 2,404 469 2,992 588 mmu-miR-155-5p 2,211 450 1,224 242 1,051 373 mmu-miR-15b-3p 242 43 76 51 97 34 mmu-miR-15b-5p 159 169 535 32 611 86 mmu-miR-16-1-3p 220 27 179 30 205 12 mmu-miR-16-5p 919 637 936 46 1,263 135 mmu-miR-17-5p 423 891 121 40 234 31
mmu-miR-181a-2-3p 58 229 236 1,211 224 1,192 mmu-miR-181a-5p 1,052 358 270 171 962 325 mmu-miR-181b-5p 2,482 895 448 363 980 434 mmu-miR-181d-5p 113 36 43 5 84 20 mmu-miR-182-5p 33,516 30,134 23,369 17,698 13,994 14,524 mmu-miR-183-5p 12,144 24,755 16,747 8,983 11,072 7,939
mmu-miR-1839-5p 1,128 877 526 136 413 104 mmu-miR-1843b-3p 4 38 10 70 13 161
mmu-miR-184-3p 169 65 28 2,649 42 119 mmu-miR-185-5p 719 610 347 66 572 203 mmu-miR-186-5p 1,150 758 265 49 1,092 81 mmu-miR-187-3p 118 18 40 14 67 26 mmu-miR-191-5p 1,670 3,626 1,401 1,303 1,998 1,952 mmu-miR-192-5p 1,072 1,651 391 133 449 167
mmu-miR-193b-3p 244 4 12 8 9 0 mmu-miR-194-2-3p 2 162 5 19 3 63 mmu-miR-194-5p 211 229 33 6 63 16
mmu-miR-1964-3p 14 81 16 135 29 133 mmu-miR-196a-1-3p 41 110 55 245 66 178 mmu-miR-196a-5p 375 981 221 266 228 377 mmu-miR-1981-5p 249 76 85 670 99 840 mmu-miR-199a-3p 16 130 7 9 11 8 mmu-miR-19b-3p 105 58 52 3 75 4 mmu-miR-1a-3p 24 272 9 283 8 18
mmu-miR-200b-3p 39 187 65 51 81 109 mmu-miR-204-5p 1 0 23 139 35 185 mmu-miR-206-3p 16,498 1,991 1,611 7,237 1,745 6,833 mmu-miR-20a-5p 377 2,058 196 27 510 43 mmu-miR-210-3p 52 27 60 51 118 27 mmu-miR-218-5p 171 49 95 11 96 32 mmu-miR-21a-5p 77,760 104,884 181,576 51,992 226,434 49,922 mmu-miR-221-3p 4,802 1,842 907 426 1,716 588 mmu-miR-221-5p 401 306 502 214 328 230 mmu-miR-222-3p 1,670 1,145 344 942 640 1,495 mmu-miR-222-5p 73 130 46 47 56 66 mmu-miR-22-3p 5,253 2,058 658 530 1,278 713
mmu-miR-224-5p 97 684 728 3,847 1,185 3,692

Appendix
209
mmu-miR-23a-3p 2,070 850 1,666 673 1,708 849 mmu-miR-23b-3p 1,904 544 1,778 692 1,877 1,004 mmu-miR-23b-5p 9 45 6 63 7 171 mmu-miR-24-2-5p 360 187 77 68 144 78 mmu-miR-24-3p 3,539 2,380 1,006 294 1,472 485 mmu-miR-25-3p 15,373 24,022 21,383 25,324 24,029 24,912 mmu-miR-25-5p 81 130 49 695 65 1,160
mmu-miR-26a-2-3p 139 11 43 2 33 3 mmu-miR-26a-5p 17,551 12,730 10,308 5,182 13,804 8,921 mmu-miR-26b-5p 3,795 1,820 1,541 204 1,870 226 mmu-miR-27a-3p 2,385 985 825 195 1,369 237 mmu-miR-27a-5p 304 288 287 161 303 216 mmu-miR-27b-3p 22,785 7,889 13,617 1,363 18,324 1,371 mmu-miR-27b-5p 52 124 56 35 42 56 mmu-miR-28a-3p 1,330 3,111 899 1,537 718 1,704 mmu-miR-28a-5p 389 290 57 78 105 85 mmu-miR-296-3p 139 22 28 62 30 120 mmu-miR-298-5p 136 97 48 68 95 146 mmu-miR-29a-3p 2,087 1,251 545 150 1,045 267 mmu-miR-29b-3p 227 18 60 5 75 0 mmu-miR-302a-5p 18 31 4 131 86 80 mmu-miR-3057-5p 77 164 80 173 55 208 mmu-miR-30a-3p 5,232 3,660 3,459 2,083 2,838 2,117 mmu-miR-30a-5p 41,428 35,650 20,780 2,243 20,715 2,176 mmu-miR-30b-3p 308 61 148 154 108 120 mmu-miR-30b-5p 2,036 110 697 100 701 48
mmu-miR-30c-2-3p 255 409 626 313 361 352 mmu-miR-30c-5p 5,156 2,819 3,102 3,336 4,295 3,663 mmu-miR-30d-5p 22,455 32,370 22,953 15,266 23,502 15,675 mmu-miR-30e-3p 1,080 963 1,937 747 1,450 776 mmu-miR-30e-5p 1,021 1,017 363 90 723 96 mmu-miR-31-3p 194 4 38 3 23 3 mmu-miR-31-5p 2,794 1,451 971 73 1,373 189
mmu-miR-320-3p 240 92 143 709 179 1,082 mmu-miR-322-3p 156 533 516 169 548 209 mmu-miR-322-5p 214 16 52 0 80 0 mmu-miR-324-3p 146 16 24 3 35 4 mmu-miR-324-5p 113 2 12 0 21 0 mmu-miR-328-3p 611 178 748 700 493 1,233 mmu-miR-331-3p 178 16 44 13 44 1 mmu-miR-339-5p 207 52 93 74 123 91 mmu-miR-340-5p 904 3,802 901 142 1,867 206 mmu-miR-342-3p 566 115 244 195 230 306 mmu-miR-342-5p 47 52 7 459 11 1,430
mmu-miR-344d-3p 102 675 74 678 71 647 mmu-miR-345-3p 234 241 75 38 82 82
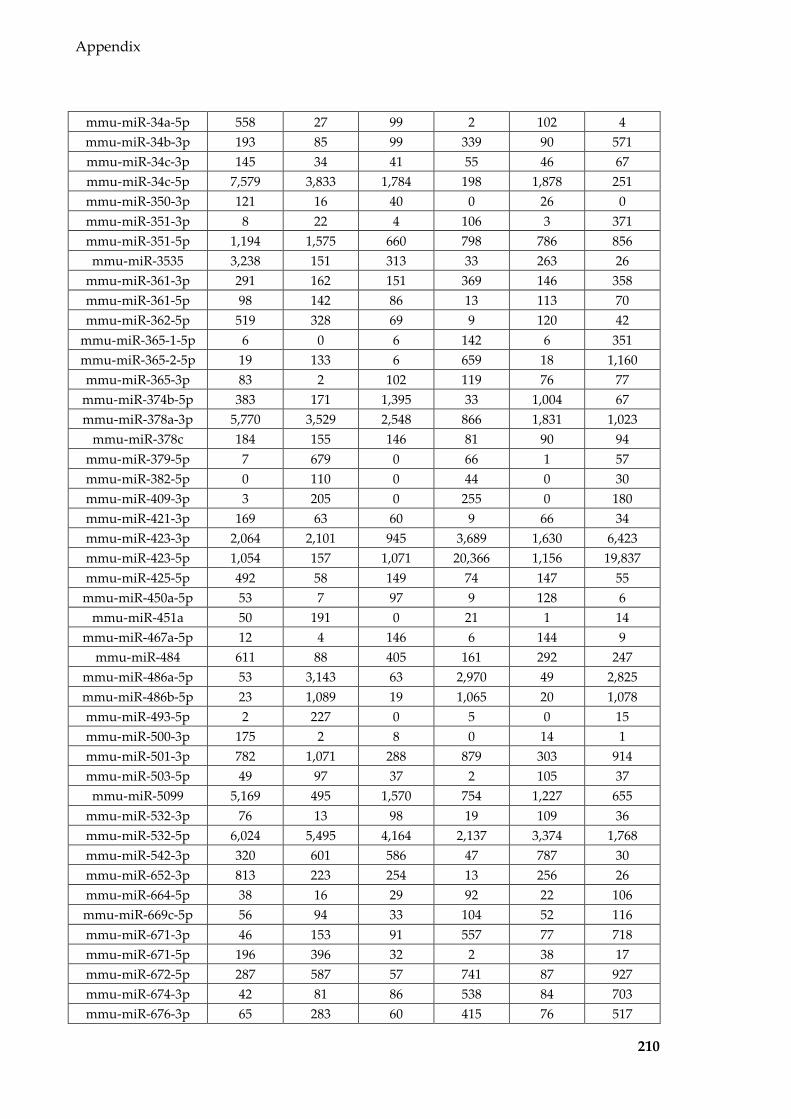
Appendix
210
mmu-miR-34a-5p 558 27 99 2 102 4 mmu-miR-34b-3p 193 85 99 339 90 571 mmu-miR-34c-3p 145 34 41 55 46 67 mmu-miR-34c-5p 7,579 3,833 1,784 198 1,878 251 mmu-miR-350-3p 121 16 40 0 26 0 mmu-miR-351-3p 8 22 4 106 3 371 mmu-miR-351-5p 1,194 1,575 660 798 786 856 mmu-miR-3535 3,238 151 313 33 263 26
mmu-miR-361-3p 291 162 151 369 146 358 mmu-miR-361-5p 98 142 86 13 113 70 mmu-miR-362-5p 519 328 69 9 120 42
mmu-miR-365-1-5p 6 0 6 142 6 351 mmu-miR-365-2-5p 19 133 6 659 18 1,160 mmu-miR-365-3p 83 2 102 119 76 77
mmu-miR-374b-5p 383 171 1,395 33 1,004 67 mmu-miR-378a-3p 5,770 3,529 2,548 866 1,831 1,023
mmu-miR-378c 184 155 146 81 90 94 mmu-miR-379-5p 7 679 0 66 1 57 mmu-miR-382-5p 0 110 0 44 0 30 mmu-miR-409-3p 3 205 0 255 0 180 mmu-miR-421-3p 169 63 60 9 66 34 mmu-miR-423-3p 2,064 2,101 945 3,689 1,630 6,423 mmu-miR-423-5p 1,054 157 1,071 20,366 1,156 19,837 mmu-miR-425-5p 492 58 149 74 147 55
mmu-miR-450a-5p 53 7 97 9 128 6 mmu-miR-451a 50 191 0 21 1 14
mmu-miR-467a-5p 12 4 146 6 144 9 mmu-miR-484 611 88 405 161 292 247
mmu-miR-486a-5p 53 3,143 63 2,970 49 2,825 mmu-miR-486b-5p 23 1,089 19 1,065 20 1,078 mmu-miR-493-5p 2 227 0 5 0 15 mmu-miR-500-3p 175 2 8 0 14 1 mmu-miR-501-3p 782 1,071 288 879 303 914 mmu-miR-503-5p 49 97 37 2 105 37 mmu-miR-5099 5,169 495 1,570 754 1,227 655
mmu-miR-532-3p 76 13 98 19 109 36 mmu-miR-532-5p 6,024 5,495 4,164 2,137 3,374 1,768 mmu-miR-542-3p 320 601 586 47 787 30 mmu-miR-652-3p 813 223 254 13 256 26 mmu-miR-664-5p 38 16 29 92 22 106 mmu-miR-669c-5p 56 94 33 104 52 116 mmu-miR-671-3p 46 153 91 557 77 718 mmu-miR-671-5p 196 396 32 2 38 17 mmu-miR-672-5p 287 587 57 741 87 927 mmu-miR-674-3p 42 81 86 538 84 703 mmu-miR-676-3p 65 283 60 415 76 517

Appendix
211
mmu-miR-677-5p 225 13 51 71 96 42 mmu-miR-744-5p 473 742 386 1,311 488 1,957
mmu-miR-7688-5p 49 85 42 169 42 305 mmu-miR-7a-5p 14,005 7,461 4,730 1,488 5,508 1,073
mmu-miR-92a-3p 1,914 1,685 2,041 13,514 2,312 12,843 mmu-miR-92b-3p 50 52 146 1,931 134 2,588 mmu-miR-92b-5p 16 0 8 673 13 945 mmu-miR-93-3p 134 25 70 28 38 27 mmu-miR-93-5p 3,662 5,122 1,714 369 3,514 371 mmu-miR-9-5p 32 27 304 158 343 58
mmu-miR-96-5p 479 101 114 38 209 38 mmu-miR-98-5p 3,113 1,134 561 347 675 480
mmu-miR-99a-5p 5,448 5,750 4,647 10,267 6,711 14,716 mmu-miR-99b-3p 869 1,176 891 627 819 692 mmu-miR-99b-5p 11,085 21,464 15,515 49,371 12,748 41,063

Appendix
212
Table 8.6 tRFs (normalised to per million tRF reads) detected in DC2 epididymal cells and
EVs of different treatments
tRF HFS cells HFS EVs LF cells LF EVs HFL cells HFL EVs tRF-Ala-AGC 215 408 476 265 170 438 tRF-Ala-CGC 1,062 1,178 1,949 1,825 618 230 tRF-Ala-TGC 7,148 1,538 18,552 1,495 1,224 407 tRF-Arg-ACG 598 405 1,054 299 317 100 tRF-Arg-CCG 618 891 1,498 978 618 261 tRF-Arg-CCT 5,608 768 12,527 798 2,136 1,175 tRF-Arg-TCG 247 32 786 34 295 23 tRF-Arg-TCT 368 133 490 100 1,998 207 tRF-Asn-GTT 1,279 99 2,421 135 1,342 261 tRF-Asp-GTC 8,007 9,525 8,555 5,830 7,200 1,351 tRF-Cys-GCA 2,968 360 9,560 277 1,086 1,897 tRF-Gln-CTG 9,420 1,216 31,794 2,079 571 676 tRF-Gln-TTG 4,290 462 11,755 781 646 161 tRF-Glu-CTC 261,889 608,623 229,853 636,268 101,938 240,646 tRF-Glu-TTC 10,323 18,201 28,933 24,918 18,448 5,767 tRF-Gly-ACC 17 18 28 30 68 15 tRF-Gly-CCC 65,153 32,941 62,485 32,688 101,385 90,177 tRF-Gly-GCC 420,423 236,121 423,513 230,463 536,517 617,777 tRF-Gly-TCC 2,735 988 4,144 756 3,576 276 tRF-His-ATG 19 2 35 0 3 0 tRF-His-GTG 27,507 34,137 38,743 22,208 43,494 8,684 tRF-Ile-AAT 337 135 624 330 314 61 tRF-Ile-TAT 97 44 102 42 72 131
tRF-Leu-AAG 532 501 1,653 326 1,119 146 tRF-Leu-CAA 2,089 1,203 3,791 1,143 663 1,067 tRF-Leu-CAG 1,503 454 3,312 328 1,664 192 tRF-Leu-TAA 2,692 318 5,113 318 1,638 514 tRF-Leu-TAG 525 474 1,434 265 1,391 100 tRF-Lys-CTT 137,292 23,076 48,761 9,766 110,153 16,778 tRF-Lys-TTT 3,375 1,018 3,323 593 12,256 1,943 tRF-Met-CAT 2,879 883 6,632 599 2,650 514 tRF-Phe-GAA 134 34 236 30 60 15 tRF-Pro-AGG 1,796 2,824 4,729 3,173 301 77 tRF-Pro-CGG 927 1,616 2,551 1,791 186 38 tRF-Pro-TGG 1,920 3,157 5,476 3,802 418 177 tRF-Ser-AGA 478 1,411 1,043 1,025 1,290 2,012 tRF-Ser-CGA 217 60 641 49 249 8 tRF-Ser-GCT 393 418 853 191 464 31 tRF-Ser-GGA 5 0 25 0 5 0 tRF-Ser-TGA 273 430 567 320 881 461
tRF-STOP-TCA 297 326 669 258 806 376 tRF-STOP-TTA 2 1 14 0 0 0
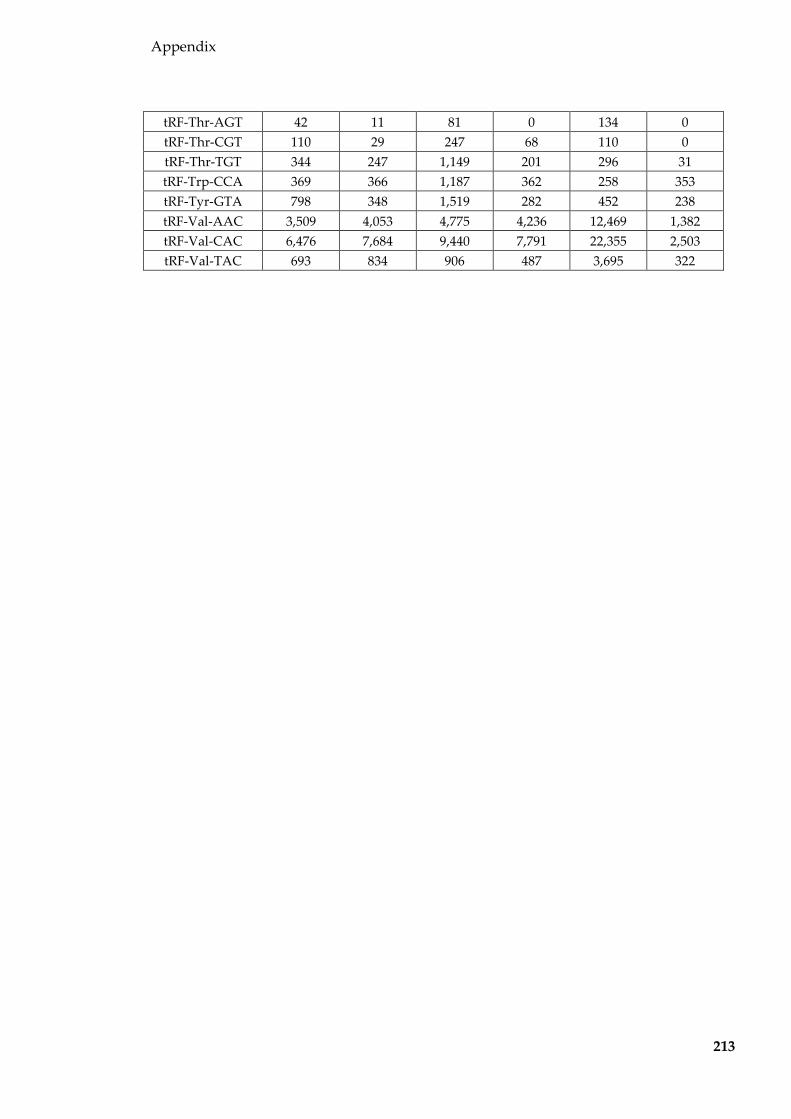
Appendix
213
tRF-Thr-AGT 42 11 81 0 134 0 tRF-Thr-CGT 110 29 247 68 110 0 tRF-Thr-TGT 344 247 1,149 201 296 31 tRF-Trp-CCA 369 366 1,187 362 258 353 tRF-Tyr-GTA 798 348 1,519 282 452 238 tRF-Val-AAC 3,509 4,053 4,775 4,236 12,469 1,382 tRF-Val-CAC 6,476 7,684 9,440 7,791 22,355 2,503 tRF-Val-TAC 693 834 906 487 3,695 322

Appendix
214
Table 8.7 Mouse epididymal fluid pools
Group C57BL/6 Control MDS
Pool number
Number of mice in
pool Age
Number of mice in
pool Age
Number of mice in
pool Age
1 5
206
5
68
5
61 208 329 324 225 331 352 266 391 369 282 444 431
2 5
207
5
86
5
101 227 307 351 239 329 356 269 385 362 300 439 367
3 5
208
5
262
5
123 227 285 326 227 313 329 248 318 347 265 380 380
4
5
293 313 322 322 324
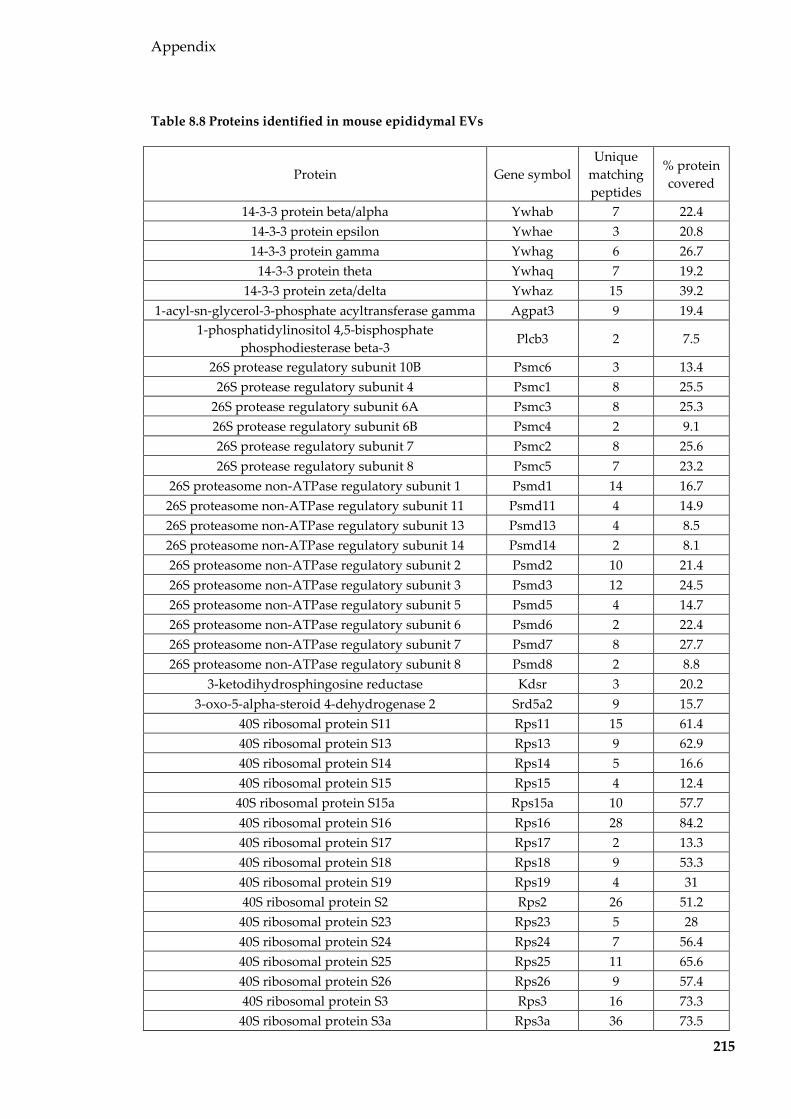
Appendix
215
Table 8.8 Proteins identified in mouse epididymal EVs
Protein Gene symbol Unique
matching peptides
% protein covered
14-3-3 protein beta/alpha Ywhab 7 22.4 14-3-3 protein epsilon Ywhae 3 20.8 14-3-3 protein gamma Ywhag 6 26.7
14-3-3 protein theta Ywhaq 7 19.2 14-3-3 protein zeta/delta Ywhaz 15 39.2
1-acyl-sn-glycerol-3-phosphate acyltransferase gamma Agpat3 9 19.4 1-phosphatidylinositol 4,5-bisphosphate
phosphodiesterase beta-3 Plcb3 2 7.5
26S protease regulatory subunit 10B Psmc6 3 13.4 26S protease regulatory subunit 4 Psmc1 8 25.5
26S protease regulatory subunit 6A Psmc3 8 25.3 26S protease regulatory subunit 6B Psmc4 2 9.1 26S protease regulatory subunit 7 Psmc2 8 25.6 26S protease regulatory subunit 8 Psmc5 7 23.2
26S proteasome non-ATPase regulatory subunit 1 Psmd1 14 16.7 26S proteasome non-ATPase regulatory subunit 11 Psmd11 4 14.9 26S proteasome non-ATPase regulatory subunit 13 Psmd13 4 8.5 26S proteasome non-ATPase regulatory subunit 14 Psmd14 2 8.1 26S proteasome non-ATPase regulatory subunit 2 Psmd2 10 21.4 26S proteasome non-ATPase regulatory subunit 3 Psmd3 12 24.5 26S proteasome non-ATPase regulatory subunit 5 Psmd5 4 14.7 26S proteasome non-ATPase regulatory subunit 6 Psmd6 2 22.4 26S proteasome non-ATPase regulatory subunit 7 Psmd7 8 27.7 26S proteasome non-ATPase regulatory subunit 8 Psmd8 2 8.8
3-ketodihydrosphingosine reductase Kdsr 3 20.2 3-oxo-5-alpha-steroid 4-dehydrogenase 2 Srd5a2 9 15.7
40S ribosomal protein S11 Rps11 15 61.4 40S ribosomal protein S13 Rps13 9 62.9 40S ribosomal protein S14 Rps14 5 16.6 40S ribosomal protein S15 Rps15 4 12.4
40S ribosomal protein S15a Rps15a 10 57.7 40S ribosomal protein S16 Rps16 28 84.2 40S ribosomal protein S17 Rps17 2 13.3 40S ribosomal protein S18 Rps18 9 53.3 40S ribosomal protein S19 Rps19 4 31 40S ribosomal protein S2 Rps2 26 51.2
40S ribosomal protein S23 Rps23 5 28 40S ribosomal protein S24 Rps24 7 56.4 40S ribosomal protein S25 Rps25 11 65.6 40S ribosomal protein S26 Rps26 9 57.4 40S ribosomal protein S3 Rps3 16 73.3
40S ribosomal protein S3a Rps3a 36 73.5
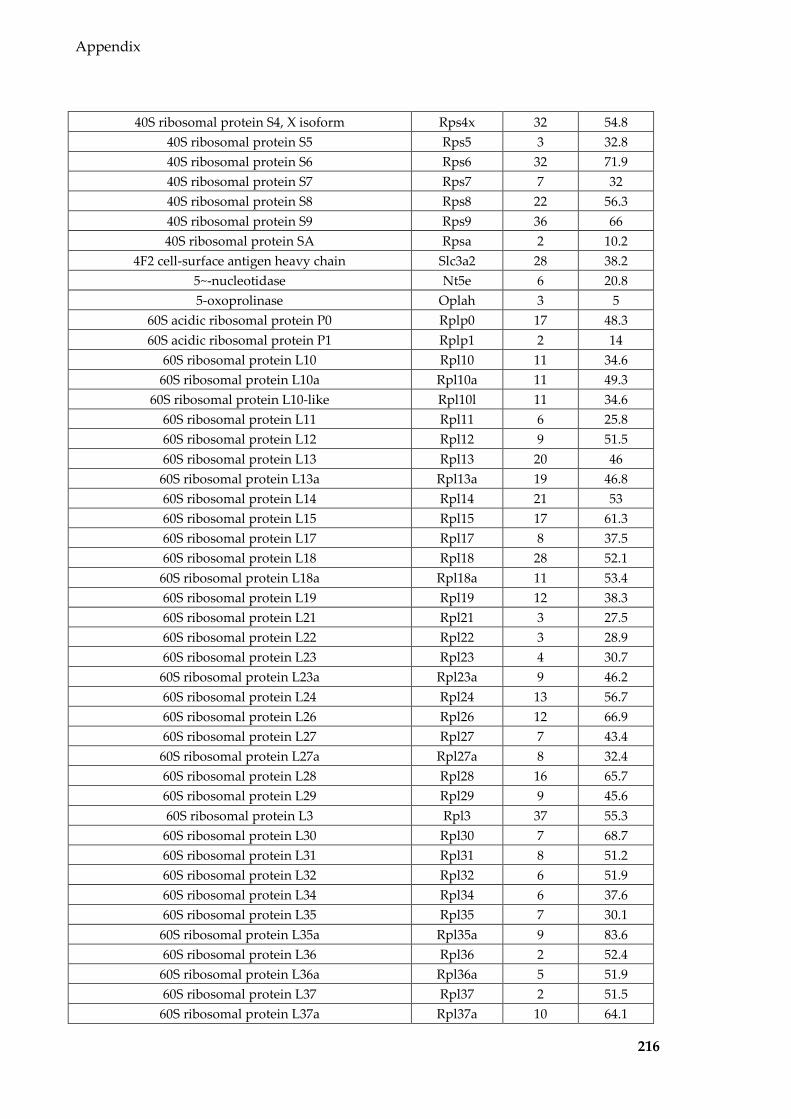
Appendix
216
40S ribosomal protein S4, X isoform Rps4x 32 54.8 40S ribosomal protein S5 Rps5 3 32.8 40S ribosomal protein S6 Rps6 32 71.9 40S ribosomal protein S7 Rps7 7 32 40S ribosomal protein S8 Rps8 22 56.3 40S ribosomal protein S9 Rps9 36 66 40S ribosomal protein SA Rpsa 2 10.2
4F2 cell-surface antigen heavy chain Slc3a2 28 38.2 5~-nucleotidase Nt5e 6 20.8 5-oxoprolinase Oplah 3 5
60S acidic ribosomal protein P0 Rplp0 17 48.3 60S acidic ribosomal protein P1 Rplp1 2 14
60S ribosomal protein L10 Rpl10 11 34.6 60S ribosomal protein L10a Rpl10a 11 49.3
60S ribosomal protein L10-like Rpl10l 11 34.6 60S ribosomal protein L11 Rpl11 6 25.8 60S ribosomal protein L12 Rpl12 9 51.5 60S ribosomal protein L13 Rpl13 20 46
60S ribosomal protein L13a Rpl13a 19 46.8 60S ribosomal protein L14 Rpl14 21 53 60S ribosomal protein L15 Rpl15 17 61.3 60S ribosomal protein L17 Rpl17 8 37.5 60S ribosomal protein L18 Rpl18 28 52.1
60S ribosomal protein L18a Rpl18a 11 53.4 60S ribosomal protein L19 Rpl19 12 38.3 60S ribosomal protein L21 Rpl21 3 27.5 60S ribosomal protein L22 Rpl22 3 28.9 60S ribosomal protein L23 Rpl23 4 30.7
60S ribosomal protein L23a Rpl23a 9 46.2 60S ribosomal protein L24 Rpl24 13 56.7 60S ribosomal protein L26 Rpl26 12 66.9 60S ribosomal protein L27 Rpl27 7 43.4
60S ribosomal protein L27a Rpl27a 8 32.4 60S ribosomal protein L28 Rpl28 16 65.7 60S ribosomal protein L29 Rpl29 9 45.6 60S ribosomal protein L3 Rpl3 37 55.3
60S ribosomal protein L30 Rpl30 7 68.7 60S ribosomal protein L31 Rpl31 8 51.2 60S ribosomal protein L32 Rpl32 6 51.9 60S ribosomal protein L34 Rpl34 6 37.6 60S ribosomal protein L35 Rpl35 7 30.1
60S ribosomal protein L35a Rpl35a 9 83.6 60S ribosomal protein L36 Rpl36 2 52.4
60S ribosomal protein L36a Rpl36a 5 51.9 60S ribosomal protein L37 Rpl37 2 51.5
60S ribosomal protein L37a Rpl37a 10 64.1

Appendix
217
60S ribosomal protein L4 Rpl4 54 71.8 60S ribosomal protein L5 Rpl5 3 28.3 60S ribosomal protein L6 Rpl6 48 77 60S ribosomal protein L7 Rpl7 32 65.2
60S ribosomal protein L7a Rpl7a 30 73.7 60S ribosomal protein L8 Rpl8 22 43.2 60S ribosomal protein L9 Rpl9 3 25
6-phosphogluconate dehydrogenase, decarboxylating Pgd 6 14.9 6-phosphogluconolactonase Pgls 2 6.2
78 kDa glucose-regulated protein Hspa5 45 60.5 Acetoacetyl-CoA synthetase Aacs 2 6.4
Acetolactate synthase-like protein Ilvbl 15 25.5 Acetyl-CoA acetyltransferase, cytosolic Acat2 3 23.7
Acetyl-CoA carboxylase 1 Acaca 3 5.3 Acid ceramidase Asah1 7 10.7
Acid sphingomyelinase-like phosphodiesterase 3b Smpdl3b 3 7 Actin, alpha cardiac muscle 1 Actc1 133 63.1 Actin, aortic smooth muscle Acta2 133 63.1
Actin, cytoplasmic 1 Actb 162 73.9 Actin, cytoplasmic 2 Actg1 162 73.9
Actin-related protein 2/3 complex subunit 4 Arpc4 4 33.3 Actin-related protein 3 Actr3 2 10.5
Adenosine 3~-phospho 5~-phosphosulfate transporter 1 Slc35b2 5 9 Adenosylhomocysteinase Ahcy 3 25.2
Adenylate kinase isoenzyme 1 Ak1 6 35.6 Adenylyl cyclase-associated protein 1 Cap1 3 18.6
Adhesion G-protein coupled receptor G2 Adgrg2 13 9.7 ADP/ATP translocase 1 Slc25a4 7 19.1 ADP/ATP translocase 2 Slc25a5 12 38.6
ADP-ribosylation factor 1 Arf1 7 53.6 ADP-ribosylation factor 3 Arf3 7 53.6 ADP-ribosylation factor 4 Arf4 6 33.3 ADP-ribosylation factor 5 Arf5 4 27.8 ADP-ribosylation factor 6 Arf6 4 17.7
ADP-ribosylation factor-like protein 1 Arl1 4 12.7 ADP-ribosylation factor-like protein 8B Arl8b 2 17.2
Aflatoxin B1 aldehyde reductase member 2 Akr7a2 4 17.7 Alcohol dehydrogenase [NADP(+)] Akr1a1 4 14.2
Alcohol dehydrogenase 1 Adh1 3 20.3 Aldehyde dehydrogenase, cytosolic 1 Aldh1a7 12 32.9
Aldehyde dehydrogenase, mitochondrial Aldh2 5 15.2 Aldose reductase Akr1b1 8 23.4
Aldose reductase-related protein 1 Akr1b7 60 80.1 Alkaline phosphatase, tissue-nonspecific isozyme Alpl 17 36.1
Alpha-1,3/1,6-mannosyltransferase ALG2 Alg2 3 7.2 Alpha-1-antitrypsin 1-1 Serpina1a 17 24.2

Appendix
218
Alpha-1-antitrypsin 1-2 Serpina1b 10 21.5 Alpha-1-antitrypsin 1-4 Serpina1d 12 29.8 Alpha-1-antitrypsin 1-5 Serpina1e 9 22.8 Alpha-1-antitrypsin 1-6 Serpina1f 6 25.1 Alpha-2-macroglobulin A2m 74 36.1
Alpha-actinin-1 Actn1 4 6.2 Alpha-actinin-4 Actn4 4 9.3
Alpha-aminoadipic semialdehyde dehydrogenase Aldh7a1 3 9.1 Alpha-centractin Actr1a 6 20.7
Alpha-enolase Eno1 5 36.2 Alpha-mannosidase 2C1 Man2c1 8 10.6
Alpha-soluble NSF attachment protein Napa 2 7.8 Aminoacyl tRNA synthase complex-interacting
multifunctional protein 2 Aimp2 3 10
Aminopeptidase N Anpep 15 17.8 Anaphase-promoting complex subunit 2 Anapc2 2 8.8
Angiotensin-converting enzyme Ace 59 34.6 Ankyrin repeat domain-containing protein 6 Ankrd6 2 11.1
Annexin A1 Anxa1 62 85 Annexin A11 Anxa11 7 21.3 Annexin A13 Anxa13 38 77.6 Annexin A2 Anxa2 97 85.3 Annexin A3 Anxa3 20 42.4 Annexin A4 Anxa4 23 50.5 Annexin A5 Anxa5 58 78.4 Annexin A6 Anxa6 60 64.9 Annexin A7 Anxa7 17 39.5 Anoctamin-1 Ano1 7 15
Anterior gradient protein 2 homolog Agr2 3 20 AP-1 complex subunit beta-1 Ap1b1 7 18.6
AP-1 complex subunit gamma-1 Ap1g1 5 12.8 AP-1 complex subunit mu-2 Ap1m2 3 13.9
AP-2 complex subunit alpha-1 Ap2a1 13 22.2 AP-2 complex subunit alpha-2 Ap2a2 21 29.3
AP-2 complex subunit beta Ap2b1 12 20.6 AP-2 complex subunit mu Ap2m1 11 20.2
AP-2 complex subunit sigma Ap2s1 4 27.5 Apolipoprotein A-I Apoa1 24 58.3
Apolipoprotein A-IV Apoa4 3 11.4 Apolipoprotein E Apoe 12 46.6
Aquaporin-1 Aqp1 2 9.7 Aquaporin-9 Aqp9 2 5.8
Arginine--tRNA ligase, cytoplasmic Rars 5 18.6 Argininosuccinate lyase Asl 2 12.5
Argininosuccinate synthase Ass1 4 14.1 Aspartate aminotransferase, cytoplasmic Got1 3 12.3
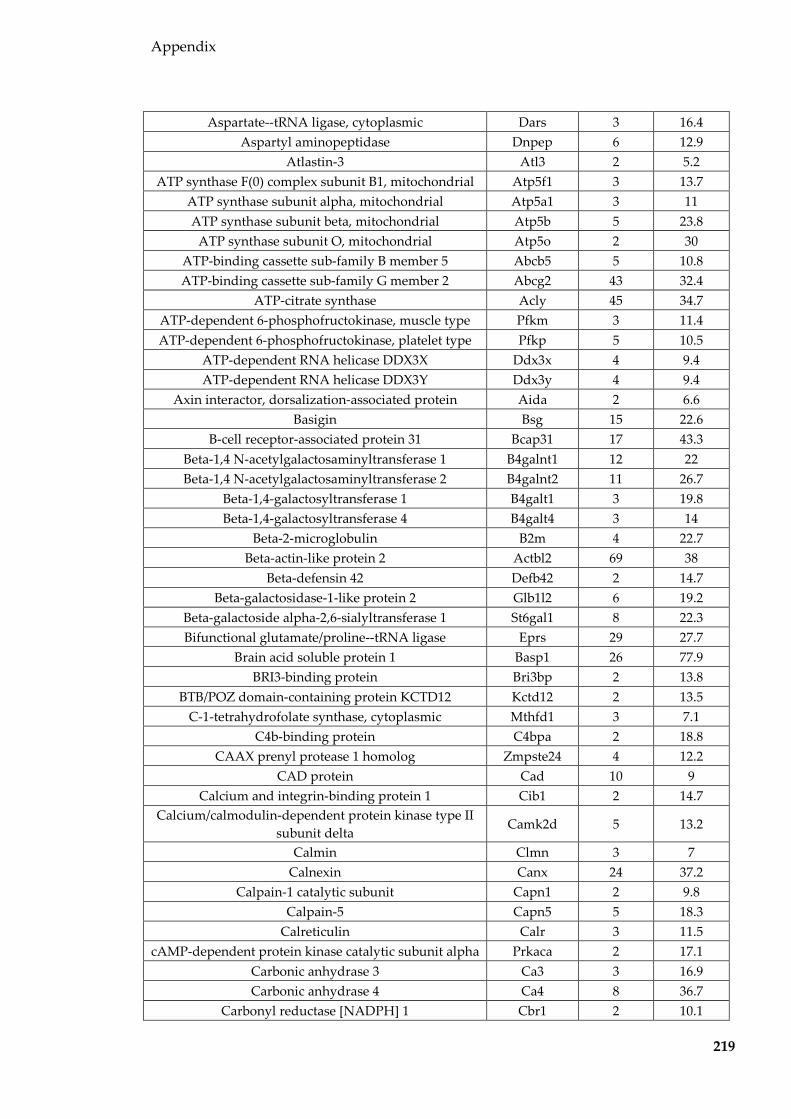
Appendix
219
Aspartate--tRNA ligase, cytoplasmic Dars 3 16.4 Aspartyl aminopeptidase Dnpep 6 12.9
Atlastin-3 Atl3 2 5.2 ATP synthase F(0) complex subunit B1, mitochondrial Atp5f1 3 13.7
ATP synthase subunit alpha, mitochondrial Atp5a1 3 11 ATP synthase subunit beta, mitochondrial Atp5b 5 23.8
ATP synthase subunit O, mitochondrial Atp5o 2 30 ATP-binding cassette sub-family B member 5 Abcb5 5 10.8 ATP-binding cassette sub-family G member 2 Abcg2 43 32.4
ATP-citrate synthase Acly 45 34.7 ATP-dependent 6-phosphofructokinase, muscle type Pfkm 3 11.4 ATP-dependent 6-phosphofructokinase, platelet type Pfkp 5 10.5
ATP-dependent RNA helicase DDX3X Ddx3x 4 9.4 ATP-dependent RNA helicase DDX3Y Ddx3y 4 9.4
Axin interactor, dorsalization-associated protein Aida 2 6.6 Basigin Bsg 15 22.6
B-cell receptor-associated protein 31 Bcap31 17 43.3 Beta-1,4 N-acetylgalactosaminyltransferase 1 B4galnt1 12 22 Beta-1,4 N-acetylgalactosaminyltransferase 2 B4galnt2 11 26.7
Beta-1,4-galactosyltransferase 1 B4galt1 3 19.8 Beta-1,4-galactosyltransferase 4 B4galt4 3 14
Beta-2-microglobulin B2m 4 22.7 Beta-actin-like protein 2 Actbl2 69 38
Beta-defensin 42 Defb42 2 14.7 Beta-galactosidase-1-like protein 2 Glb1l2 6 19.2
Beta-galactoside alpha-2,6-sialyltransferase 1 St6gal1 8 22.3 Bifunctional glutamate/proline--tRNA ligase Eprs 29 27.7
Brain acid soluble protein 1 Basp1 26 77.9 BRI3-binding protein Bri3bp 2 13.8
BTB/POZ domain-containing protein KCTD12 Kctd12 2 13.5 C-1-tetrahydrofolate synthase, cytoplasmic Mthfd1 3 7.1
C4b-binding protein C4bpa 2 18.8 CAAX prenyl protease 1 homolog Zmpste24 4 12.2
CAD protein Cad 10 9 Calcium and integrin-binding protein 1 Cib1 2 14.7
Calcium/calmodulin-dependent protein kinase type II subunit delta
Camk2d 5 13.2
Calmin Clmn 3 7 Calnexin Canx 24 37.2
Calpain-1 catalytic subunit Capn1 2 9.8 Calpain-5 Capn5 5 18.3
Calreticulin Calr 3 11.5 cAMP-dependent protein kinase catalytic subunit alpha Prkaca 2 17.1
Carbonic anhydrase 3 Ca3 3 16.9 Carbonic anhydrase 4 Ca4 8 36.7
Carbonyl reductase [NADPH] 1 Cbr1 2 10.1

Appendix
220
Carboxylesterase 5A Ces5a 19 29 Casein kinase I isoform gamma-2 Csnk1g2 2 6.5
Casein kinase II subunit alpha Csnk2a1 3 9 Casein kinase II subunit beta Csnk2b 2 10.2 Catechol O-methyltransferase Comt 3 17.7
Catenin beta-1 Ctnnb1 5 6.8 Cation-dependent mannose-6-phosphate receptor M6pr 4 26.3
Cation-independent mannose-6-phosphate receptor Igf2r 4 6.3 Caveolin-1 Cav1 6 51.7
CD151 antigen Cd151 3 24.9 CD81 antigen Cd81 4 19.5 CD82 antigen Cd82 3 7.9 CD9 antigen Cd9 8 23.9
CDGSH iron-sulfur domain-containing protein 2 Cisd2 2 20.7 CDGSH iron-sulfur domain-containing protein 3,
mitochondrial Cisd3 2 5.1
CDK5 regulatory subunit-associated protein 2 Cdk5rap2 2 5.8 CDP-diacylglycerol--inositol 3-phosphatidyltransferase Cdipt 4 29.6
Ceruloplasmin Cp 20 20.7 Chloride intracellular channel protein 1 Clic1 18 55.2 Chloride intracellular channel protein 3 Clic3 3 18.6 Chloride intracellular channel protein 4 Clic4 9 32
Choline transporter-like protein 1 Slc44a1 5 9.2 Choline transporter-like protein 2 Slc44a2 11 17.1 Choline transporter-like protein 4 Slc44a4 18 12.7
Clathrin heavy chain 1 Cltc 103 43.5 Clathrin light chain A Clta 4 21.3
Claudin-3 Cldn3 2 14.2 Cleft lip and palate transmembrane protein 1 homolog Clptm1 2 5.7
Clusterin Clu 34 44.6 CMP-sialic acid transporter Slc35a1 5 14.3
Coactosin-like protein Cotl1 6 33.1 Coatomer subunit alpha Copa 85 46.6 Coatomer subunit beta Copb1 23 26.1
Coatomer subunit beta~ Copb2 36 38.2 Coatomer subunit epsilon Cope 11 39.9
Coatomer subunit gamma-1 Copg1 8 15 Coatomer subunit gamma-2 Copg2 11 13.4
Cofilin-1 Cfl1 5 29.5 Cofilin-2 Cfl2 5 16.9
Complement C1q subcomponent subunit B C1qb 5 13.8 Complement C1q subcomponent subunit C C1qc 4 21.5
Complement C3 C3 53 34.2 Complement C4-B C4b 6 6.8
Complement factor H Cfh 9 8.4 Conserved oligomeric Golgi complex subunit 1 Cog1 4 6

Appendix
221
Conserved oligomeric Golgi complex subunit 4 Cog4 2 5.7 Copine-1 Cpne1 4 9.1 Copine-3 Cpne3 2 9
CUB and zona pellucida-like domain-containing protein 1
Cuzd1 192 43.7
Cullin-associated NEDD8-dissociated protein 1 Cand1 7 12.7 Cystathionine gamma-lyase Cth 4 17.3
Cysteine and glycine-rich protein 1 Csrp1 2 15.5 Cysteine-rich protein 1 Crip1 6 36.4
Cysteine-rich secretory protein 1 Crisp1 8 36.1 Cytochrome b5 type B Cyb5b 2 13
Cytochrome b561 Cyb561 2 6.8 Cytochrome b-c1 complex subunit 2, mitochondrial Uqcrc2 4 14.3
Cytochrome c oxidase subunit 4 isoform 1, mitochondrial
Cox4i1 3 29
Cytochrome c oxidase subunit 6C Cox6c 2 44.7 Cytoplasmic aconitate hydratase Aco1 4 15.1
Cytoplasmic dynein 1 heavy chain 1 Dync1h1 53 17.6 Cytoplasmic dynein 1 light intermediate chain 1 Dync1li1 3 13.4
Cytoplasmic FMR1-interacting protein 1 Cyfip1 16 17.8 Cytosol aminopeptidase Lap3 25 61.7
Cytosolic non-specific dipeptidase Cndp2 5 16 D-3-phosphoglycerate dehydrogenase Phgdh 7 18.6
Dedicator of cytokinesis protein 1 Dock1 2 6.9 Dehydrogenase/reductase SDR family member 7B Dhrs7b 2 20.4
Delta(14)-sterol reductase Tm7sf2 5 16.3 Delta-aminolevulinic acid dehydratase Alad 7 39.4
Destrin Dstn 3 15.8 Dihydrolipoyllysine-residue acetyltransferase
component of pyruvate dehydrogenase complex, mitochondrial
Dlat 2 7.8
Dihydropyrimidinase-related protein 2 Dpysl2 2 9.1 Dimethylaniline monooxygenase [N-oxide-forming] 2 Fmo2 21 35.9
Dipeptidase 1 Dpep1 8 33.4 Dipeptidase 3 Dpep3 11 18.5
Dipeptidyl peptidase 4 Dpp4 40 34.6 Disintegrin and metalloproteinase domain-containing
protein 1b Adam1b 4 8.6
Disintegrin and metalloproteinase domain-containing protein 28
Adam28 2 7.4
Disintegrin and metalloproteinase domain-containing protein 7
Adam7 27 32.1
DNA (cytosine-5)-methyltransferase 1 Dnmt1 3 9.8 DNA-directed RNA polymerases I, II, and III subunit
RPABC1 Polr2e 2 15.7
DnaJ homolog subfamily C member 10 Dnajc10 3 8.7 Dolichyl-diphosphooligosaccharide--protein Ddost 13 22

Appendix
222
glycosyltransferase 48 kDa subunit Dolichyl-diphosphooligosaccharide--protein
glycosyltransferase subunit 1 Rpn1 27 36.5
Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit 2
Rpn2 13 33.9
Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit STT3A
Stt3a 11 13.3
Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit STT3B
Stt3b 7 11.4
Dolichyl-phosphate beta-glucosyltransferase Alg5 3 13.9 Dynactin subunit 1 Dctn1 8 12.7
Dysferlin Dysf 2 5.6 E3 ubiquitin-protein ligase MYCBP2 Mycbp2 9 5.9 E3 ubiquitin-protein ligase NEDD4 Nedd4 2 6 E3 ubiquitin-protein ligase UBR4 Ubr4 16 6.4
Ectonucleoside triphosphate diphosphohydrolase 2 Entpd2 25 37 EH domain-containing protein 1 Ehd1 6 17.6 EH domain-containing protein 2 Ehd2 3 19.9
Elongation factor 1-alpha 1 Eef1a1 29 48.1 Elongation factor 1-gamma Eef1g 7 14
Elongation factor 2 Eef2 10 15.9 Elongator complex protein 1 Ikbkap 5 8.1 Elongator complex protein 3 Elp3 2 11.7
Embigin Emb 7 14.5 Endoplasmic reticulum resident protein 44 Erp44 3 10.8 Endoplasmic reticulum-Golgi intermediate
compartment protein 1 Ergic1 16 31.4
Endoplasmin Hsp90b1 25 35.3 Endothelial lipase Lipg 2 11
Epididymal secretory glutathione peroxidase Gpx5 14 35.3 Epididymal-specific lipocalin-5 Lcn5 11 41.1 Epididymal-specific lipocalin-8 Lcn8 2 18.9
Epithelial cell adhesion molecule Epcam 12 38.7 Epoxide hydrolase 1 Ephx1 7 21.8
ER membrane protein complex subunit 1 Emc1 3 5.5 ER membrane protein complex subunit 2 Emc2 3 16.8 ER membrane protein complex subunit 3 Emc3 3 22.2 ER membrane protein complex subunit 8 Emc8 2 10.1
Erlin-2 Erlin2 7 46.5 Erythrocyte band 7 integral membrane protein Stom 7 21.5
Estradiol 17-beta-dehydrogenase 11 Hsd17b11 5 10.1 Eukaryotic translation initiation factor 2 subunit 3, X-
linked Eif2s3x 6 16.3
Eukaryotic translation initiation factor 3 subunit A Eif3a 18 22.8 Eukaryotic translation initiation factor 3 subunit C Eif3c 6 12.6 Eukaryotic translation initiation factor 3 subunit H Eif3h 3 10.5
Eukaryotic translation initiation factor 5B Eif5b 2 7

Appendix
223
Excitatory amino acid transporter 3 Slc1a1 7 12 Extended synaptotagmin-1 Esyt1 6 10.1
Ezrin Ezr 120 75.9 F-actin-capping protein subunit alpha-2 Capza2 4 23.8
F-actin-capping protein subunit beta Capzb 5 19.5 Far upstream element-binding protein 2 Khsrp 3 8.3
Fatty acid synthase Fasn 61 27 Fatty aldehyde dehydrogenase Aldh3a2 3 15.1 Fatty-acid amide hydrolase 1 Faah 2 13.3
Ferritin heavy chain Fth1 15 50.5 Ferritin light chain 1 Ftl1 31 77
Fibrinogen alpha chain Fga 20 23.8 Fibrinogen beta chain Fgb 11 29.9
Fibrinogen gamma chain Fgg 9 30.7 Fibronectin Fn1 6 5.7 Filamin-A Flna 17 11.9
Fructose-1,6-bisphosphatase 1 Fbp1 6 27.2 Fructose-bisphosphate aldolase A Aldoa 16 52.7
Galactokinase Galk1 3 14.5 Galectin-3-binding protein Lgals3bp 29 49.9
Gamma-glutamyltranspeptidase 1 Ggt1 27 40.3 Gamma-soluble NSF attachment protein Napg 2 12.2
GDP-fucose transporter 1 Slc35c1 2 7.7 Gelsolin Gsn 13 26.2
General vesicular transport factor p115 Uso1 3 5.7 GLIPR1-like protein 2 Glipr1l2 3 16.6
Glucose-6-phosphate isomerase Gpi 20 46.8 Glutamate carboxypeptidase 2 Folh1 9 17.6
Glutamate dehydrogenase 1, mitochondrial Glud1 7 6.6 Glutamine synthetase Glul 52 62.7
Glutamine--fructose-6-phosphate aminotransferase [isomerizing] 1
Gfpt1 3 6.2
Glutathione peroxidase 3 Gpx3 6 29.2 Glutathione S-transferase A3 Gsta3 3 23.1
Glutathione S-transferase Mu 1 Gstm1 20 63.8 Glutathione S-transferase Mu 2 Gstm2 22 78.9 Glutathione S-transferase Mu 5 Gstm5 16 50 Glutathione S-transferase Mu 6 Gstm6 5 16.1 Glutathione S-transferase Mu 7 Gstm7 18 62.4 Glutathione S-transferase P 1 Gstp1 5 24.8
Glyceraldehyde-3-phosphate dehydrogenase Gapdh 44 64 Glycogen phosphorylase, brain form Pygb 13 24.1 Glycogen phosphorylase, liver form Pygl 7 8.9
Glycogen phosphorylase, muscle form Pygm 10 17.8 Glycosaminoglycan xylosylkinase Fam20b 2 13.9
Glycosyltransferase 8 domain-containing protein 1 Glt8d1 2 5.7
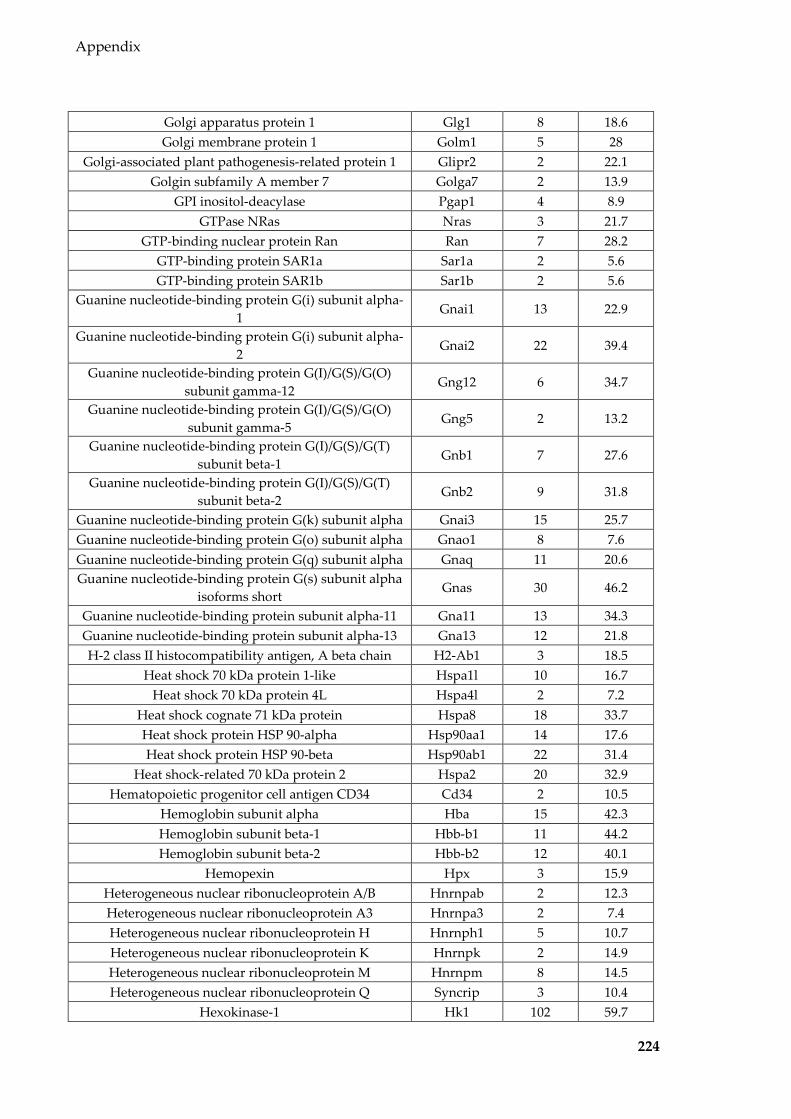
Appendix
224
Golgi apparatus protein 1 Glg1 8 18.6 Golgi membrane protein 1 Golm1 5 28
Golgi-associated plant pathogenesis-related protein 1 Glipr2 2 22.1 Golgin subfamily A member 7 Golga7 2 13.9
GPI inositol-deacylase Pgap1 4 8.9 GTPase NRas Nras 3 21.7
GTP-binding nuclear protein Ran Ran 7 28.2 GTP-binding protein SAR1a Sar1a 2 5.6 GTP-binding protein SAR1b Sar1b 2 5.6
Guanine nucleotide-binding protein G(i) subunit alpha-1
Gnai1 13 22.9
Guanine nucleotide-binding protein G(i) subunit alpha-2
Gnai2 22 39.4
Guanine nucleotide-binding protein G(I)/G(S)/G(O) subunit gamma-12
Gng12 6 34.7
Guanine nucleotide-binding protein G(I)/G(S)/G(O) subunit gamma-5
Gng5 2 13.2
Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-1
Gnb1 7 27.6
Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2
Gnb2 9 31.8
Guanine nucleotide-binding protein G(k) subunit alpha Gnai3 15 25.7 Guanine nucleotide-binding protein G(o) subunit alpha Gnao1 8 7.6 Guanine nucleotide-binding protein G(q) subunit alpha Gnaq 11 20.6 Guanine nucleotide-binding protein G(s) subunit alpha
isoforms short Gnas 30 46.2
Guanine nucleotide-binding protein subunit alpha-11 Gna11 13 34.3 Guanine nucleotide-binding protein subunit alpha-13 Gna13 12 21.8 H-2 class II histocompatibility antigen, A beta chain H2-Ab1 3 18.5
Heat shock 70 kDa protein 1-like Hspa1l 10 16.7 Heat shock 70 kDa protein 4L Hspa4l 2 7.2
Heat shock cognate 71 kDa protein Hspa8 18 33.7 Heat shock protein HSP 90-alpha Hsp90aa1 14 17.6 Heat shock protein HSP 90-beta Hsp90ab1 22 31.4
Heat shock-related 70 kDa protein 2 Hspa2 20 32.9 Hematopoietic progenitor cell antigen CD34 Cd34 2 10.5
Hemoglobin subunit alpha Hba 15 42.3 Hemoglobin subunit beta-1 Hbb-b1 11 44.2 Hemoglobin subunit beta-2 Hbb-b2 12 40.1
Hemopexin Hpx 3 15.9 Heterogeneous nuclear ribonucleoprotein A/B Hnrnpab 2 12.3 Heterogeneous nuclear ribonucleoprotein A3 Hnrnpa3 2 7.4 Heterogeneous nuclear ribonucleoprotein H Hnrnph1 5 10.7 Heterogeneous nuclear ribonucleoprotein K Hnrnpk 2 14.9 Heterogeneous nuclear ribonucleoprotein M Hnrnpm 8 14.5 Heterogeneous nuclear ribonucleoprotein Q Syncrip 3 10.4
Hexokinase-1 Hk1 102 59.7

Appendix
225
Hexokinase-2 Hk2 9 14.6 Hippocalcin-like protein 1 Hpcal1 2 15.5 Histidine-rich glycoprotein Hrg 3 5.7
Histone deacetylase 6 Hdac6 4 7.3 Histone H1.2 Hist1h1c 5 43.4 Histone H1.4 Hist1h1e 8 39.7
Histone H2A type 1 Hist1h2ab 2 16.9 Histone H2A type 1-F Hist1h2af 2 16.9 Histone H2A type 1-H Hist1h2ah 2 17.2 Histone H2A type 1-K Hist1h2ak 2 16.9 Histone H2A type 2-A Hist2h2aa1 2 16.9 Histone H2A type 2-C Hist2h2ac 2 17.1
Histone H2A type 3 Hist3h2a 2 16.9 Histone H2A.J H2afj 2 17.1 Histone H2A.V H2afv 2 30.5 Histone H2A.Z H2afz 2 30.5 Histone H2AX H2afx 2 21.7
Histone H2B type 1-C/E/G Hist1h2bc 13 61.9 Histone H2B type 1-H Hist1h2bh 13 61.9 Histone H2B type 1-M Hist1h2bm 13 61.9 Histone H2B type 2-B Hist2h2bb 13 71.4
Histone H3.1 Hist1h3a 4 40.4 Histone H3.1 H3110 4 40.4 Histone H3.2 Hist1h3b 4 40.4 Histone H3.2 H312 4 40.4 Histone H3.3 H3f3a 4 40.4
Histone H3.3C H3f3c 4 40.4 Histone H4 Hist1h4a 5 62.1
Histone lysine demethylase PHF8 Phf8 2 5.6 Hormone-sensitive lipase Lipe 3 5
Huntingtin-interacting protein 1-related protein Hip1r 2 5.9 Hyaluronidase PH-20 Spam1 2 6.1
Hypoxanthine-guanine phosphoribosyltransferase (Fragment)
Hprt1 4 20.1
Hypoxanthine-guanine phosphoribosyltransferase Hprt1 4 19.7 Hypoxia up-regulated protein 1 Hyou1 15 19
Ig alpha chain C region #VALUE! 2 7.3 Ig gamma-1 chain C region secreted form Ighg1 2 8.3
Ig gamma-2A chain C region secreted form #VALUE! 3 17.3 Ig gamma-2B chain C region Igh-3 7 20 Ig gamma-3 chain C region #VALUE! 2 7
Ig heavy chain V region AC38 205.12 #VALUE! 4 42.4 Ig kappa chain C region #VALUE! 5 24.5
Ig mu chain C region Ighm 12 30.6 Importin subunit beta-1 Kpnb1 3 5.7
Inactive serine protease 39 Prss39 2 9.8
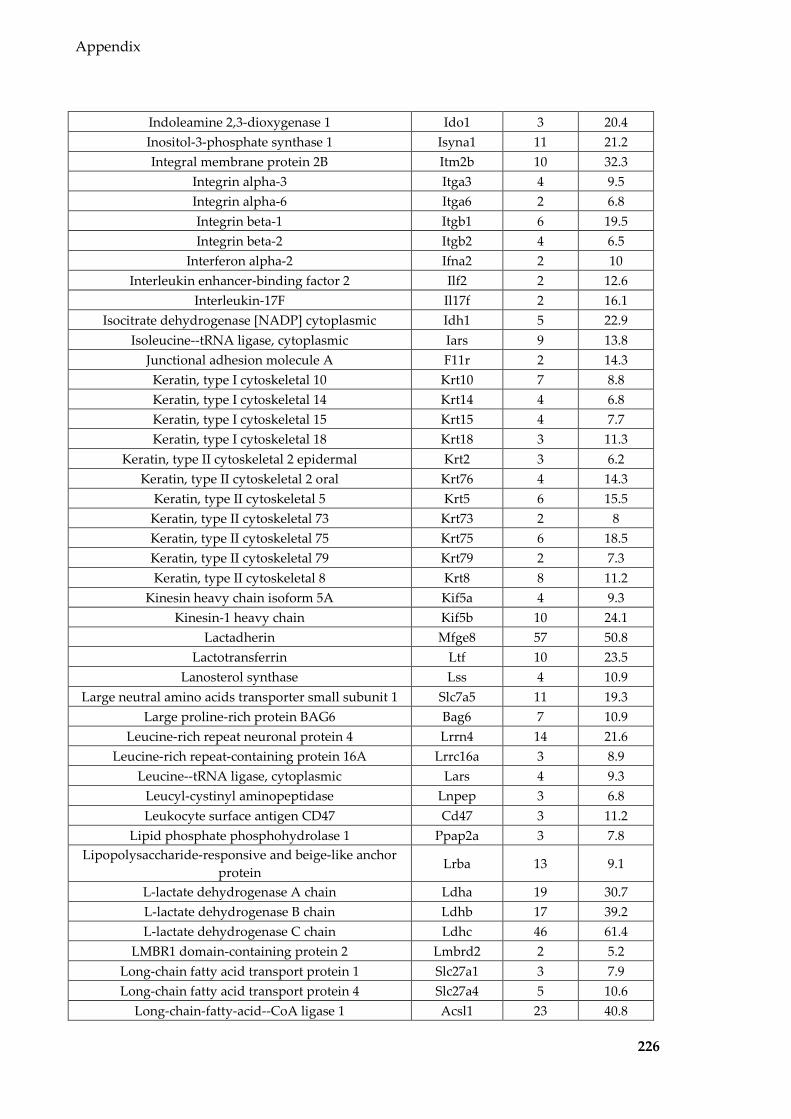
Appendix
226
Indoleamine 2,3-dioxygenase 1 Ido1 3 20.4 Inositol-3-phosphate synthase 1 Isyna1 11 21.2 Integral membrane protein 2B Itm2b 10 32.3
Integrin alpha-3 Itga3 4 9.5 Integrin alpha-6 Itga6 2 6.8 Integrin beta-1 Itgb1 6 19.5 Integrin beta-2 Itgb2 4 6.5
Interferon alpha-2 Ifna2 2 10 Interleukin enhancer-binding factor 2 Ilf2 2 12.6
Interleukin-17F Il17f 2 16.1 Isocitrate dehydrogenase [NADP] cytoplasmic Idh1 5 22.9
Isoleucine--tRNA ligase, cytoplasmic Iars 9 13.8 Junctional adhesion molecule A F11r 2 14.3 Keratin, type I cytoskeletal 10 Krt10 7 8.8 Keratin, type I cytoskeletal 14 Krt14 4 6.8 Keratin, type I cytoskeletal 15 Krt15 4 7.7 Keratin, type I cytoskeletal 18 Krt18 3 11.3
Keratin, type II cytoskeletal 2 epidermal Krt2 3 6.2 Keratin, type II cytoskeletal 2 oral Krt76 4 14.3
Keratin, type II cytoskeletal 5 Krt5 6 15.5 Keratin, type II cytoskeletal 73 Krt73 2 8 Keratin, type II cytoskeletal 75 Krt75 6 18.5 Keratin, type II cytoskeletal 79 Krt79 2 7.3 Keratin, type II cytoskeletal 8 Krt8 8 11.2
Kinesin heavy chain isoform 5A Kif5a 4 9.3 Kinesin-1 heavy chain Kif5b 10 24.1
Lactadherin Mfge8 57 50.8 Lactotransferrin Ltf 10 23.5
Lanosterol synthase Lss 4 10.9 Large neutral amino acids transporter small subunit 1 Slc7a5 11 19.3
Large proline-rich protein BAG6 Bag6 7 10.9 Leucine-rich repeat neuronal protein 4 Lrrn4 14 21.6
Leucine-rich repeat-containing protein 16A Lrrc16a 3 8.9 Leucine--tRNA ligase, cytoplasmic Lars 4 9.3
Leucyl-cystinyl aminopeptidase Lnpep 3 6.8 Leukocyte surface antigen CD47 Cd47 3 11.2
Lipid phosphate phosphohydrolase 1 Ppap2a 3 7.8 Lipopolysaccharide-responsive and beige-like anchor
protein Lrba 13 9.1
L-lactate dehydrogenase A chain Ldha 19 30.7 L-lactate dehydrogenase B chain Ldhb 17 39.2 L-lactate dehydrogenase C chain Ldhc 46 61.4
LMBR1 domain-containing protein 2 Lmbrd2 2 5.2 Long-chain fatty acid transport protein 1 Slc27a1 3 7.9 Long-chain fatty acid transport protein 4 Slc27a4 5 10.6
Long-chain-fatty-acid--CoA ligase 1 Acsl1 23 40.8
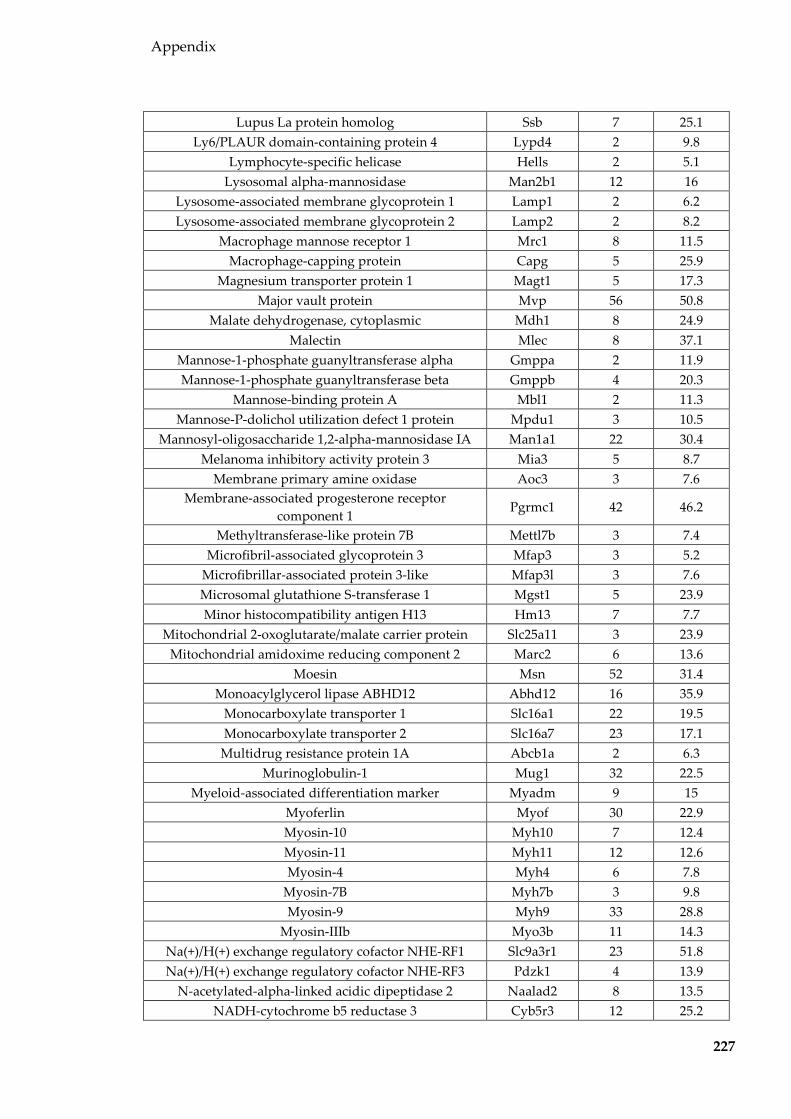
Appendix
227
Lupus La protein homolog Ssb 7 25.1 Ly6/PLAUR domain-containing protein 4 Lypd4 2 9.8
Lymphocyte-specific helicase Hells 2 5.1 Lysosomal alpha-mannosidase Man2b1 12 16
Lysosome-associated membrane glycoprotein 1 Lamp1 2 6.2 Lysosome-associated membrane glycoprotein 2 Lamp2 2 8.2
Macrophage mannose receptor 1 Mrc1 8 11.5 Macrophage-capping protein Capg 5 25.9
Magnesium transporter protein 1 Magt1 5 17.3 Major vault protein Mvp 56 50.8
Malate dehydrogenase, cytoplasmic Mdh1 8 24.9 Malectin Mlec 8 37.1
Mannose-1-phosphate guanyltransferase alpha Gmppa 2 11.9 Mannose-1-phosphate guanyltransferase beta Gmppb 4 20.3
Mannose-binding protein A Mbl1 2 11.3 Mannose-P-dolichol utilization defect 1 protein Mpdu1 3 10.5
Mannosyl-oligosaccharide 1,2-alpha-mannosidase IA Man1a1 22 30.4 Melanoma inhibitory activity protein 3 Mia3 5 8.7
Membrane primary amine oxidase Aoc3 3 7.6 Membrane-associated progesterone receptor
component 1 Pgrmc1 42 46.2
Methyltransferase-like protein 7B Mettl7b 3 7.4 Microfibril-associated glycoprotein 3 Mfap3 3 5.2
Microfibrillar-associated protein 3-like Mfap3l 3 7.6 Microsomal glutathione S-transferase 1 Mgst1 5 23.9 Minor histocompatibility antigen H13 Hm13 7 7.7
Mitochondrial 2-oxoglutarate/malate carrier protein Slc25a11 3 23.9 Mitochondrial amidoxime reducing component 2 Marc2 6 13.6
Moesin Msn 52 31.4 Monoacylglycerol lipase ABHD12 Abhd12 16 35.9
Monocarboxylate transporter 1 Slc16a1 22 19.5 Monocarboxylate transporter 2 Slc16a7 23 17.1
Multidrug resistance protein 1A Abcb1a 2 6.3 Murinoglobulin-1 Mug1 32 22.5
Myeloid-associated differentiation marker Myadm 9 15 Myoferlin Myof 30 22.9 Myosin-10 Myh10 7 12.4 Myosin-11 Myh11 12 12.6 Myosin-4 Myh4 6 7.8
Myosin-7B Myh7b 3 9.8 Myosin-9 Myh9 33 28.8
Myosin-IIIb Myo3b 11 14.3 Na(+)/H(+) exchange regulatory cofactor NHE-RF1 Slc9a3r1 23 51.8 Na(+)/H(+) exchange regulatory cofactor NHE-RF3 Pdzk1 4 13.9
N-acetylated-alpha-linked acidic dipeptidase 2 Naalad2 8 13.5 NADH-cytochrome b5 reductase 3 Cyb5r3 12 25.2

Appendix
228
NADP-dependent malic enzyme Me1 5 12.4 NADPH--cytochrome P450 reductase Por 3 13.3
Nck-associated protein 1 Nckap1 5 6 Neprilysin Mme 24 35.7
Neuronal membrane glycoprotein M6-a Gpm6a 13 16.2 Neuron-specific calcium-binding protein hippocalcin Hpca 2 18.7
Neuroplastin Nptn 2 8.1 Neutral amino acid transporter B(0) Slc1a5 2 5.8
Neutrophil gelatinase-associated lipocalin Lcn2 4 22 Nicastrin Ncstn 3 5.9
Nodal modulator 1 Nomo1 3 7.9 Nucleolin Ncl 17 26.4
Nucleoside diphosphate kinase A Nme1 5 34.9 Nucleoside diphosphate kinase B Nme2 5 44.1
Nucleoside diphosphate-linked moiety X motif 13 Nudt13 2 6.8 Oligosaccharyltransferase complex subunit OSTC Ostc 2 14.1
Ovostatin homolog Ovos 10 8.9 OX-2 membrane glycoprotein Cd200 2 10.8
Peptidyl-prolyl cis-trans isomerase A Ppia 6 35.4 Peptidyl-prolyl cis-trans isomerase B Ppib 5 27.3
Peptidyl-prolyl cis-trans isomerase FKBP11 Fkbp11 4 17.9 Perilipin-4 Plin4 7 5.8
Peroxiredoxin-1 Prdx1 15 49.2 Peroxiredoxin-2 Prdx2 4 18.2
Peroxiredoxin-5, mitochondrial Prdx5 2 25.2 Peroxiredoxin-6 Prdx6 5 12.9
PGAP2-interacting protein Cwh43 3 6.2 Phosphate carrier protein, mitochondrial Slc25a3 4 21.3
Phosphatidylethanolamine-binding protein 1 Pebp1 6 36.4 Phosphatidylinositide phosphatase SAC1 Sacm1l 12 23.9
Phosphoglycerate kinase 1 Pgk1 13 39.6 Phosphoglycerate kinase 2 Pgk2 37 50.4 Phosphoglycerate mutase 2 Pgam2 13 34
Phospholipase B1, membrane-associated Plb1 3 8.1 Phospholipid-transporting ATPase IK Atp8b3 4 5.4
Pituitary tumor-transforming gene 1 protein-interacting protein
Pttg1ip 4 27
Placenta-specific gene 8 protein Plac8 2 29.5 Plasma membrane calcium-transporting ATPase 1 Atp2b1 29 24.8
Plastin-2 Lcp1 25 36.7 Plastin-3 Pls3 39 51.3 Plexin-B2 Plxnb2 7 7.3
Poly(rC)-binding protein 1 Pcbp1 6 20.8 Polyadenylate-binding protein 1 Pabpc1 8 22.2
Polymerase I and transcript release factor Ptrf 7 12.2 Polypeptide N-acetylgalactosaminyltransferase 1 Galnt1 2 9.5
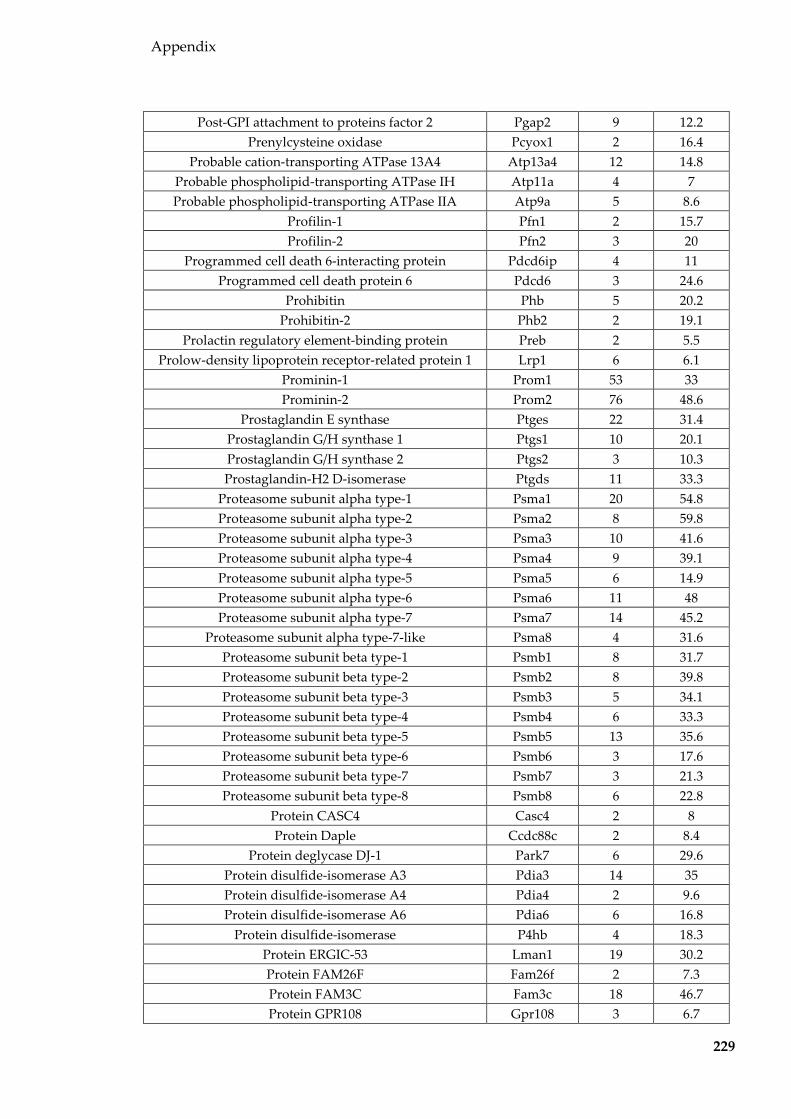
Appendix
229
Post-GPI attachment to proteins factor 2 Pgap2 9 12.2 Prenylcysteine oxidase Pcyox1 2 16.4
Probable cation-transporting ATPase 13A4 Atp13a4 12 14.8 Probable phospholipid-transporting ATPase IH Atp11a 4 7 Probable phospholipid-transporting ATPase IIA Atp9a 5 8.6
Profilin-1 Pfn1 2 15.7 Profilin-2 Pfn2 3 20
Programmed cell death 6-interacting protein Pdcd6ip 4 11 Programmed cell death protein 6 Pdcd6 3 24.6
Prohibitin Phb 5 20.2 Prohibitin-2 Phb2 2 19.1
Prolactin regulatory element-binding protein Preb 2 5.5 Prolow-density lipoprotein receptor-related protein 1 Lrp1 6 6.1
Prominin-1 Prom1 53 33 Prominin-2 Prom2 76 48.6
Prostaglandin E synthase Ptges 22 31.4 Prostaglandin G/H synthase 1 Ptgs1 10 20.1 Prostaglandin G/H synthase 2 Ptgs2 3 10.3 Prostaglandin-H2 D-isomerase Ptgds 11 33.3
Proteasome subunit alpha type-1 Psma1 20 54.8 Proteasome subunit alpha type-2 Psma2 8 59.8 Proteasome subunit alpha type-3 Psma3 10 41.6 Proteasome subunit alpha type-4 Psma4 9 39.1 Proteasome subunit alpha type-5 Psma5 6 14.9 Proteasome subunit alpha type-6 Psma6 11 48 Proteasome subunit alpha type-7 Psma7 14 45.2
Proteasome subunit alpha type-7-like Psma8 4 31.6 Proteasome subunit beta type-1 Psmb1 8 31.7 Proteasome subunit beta type-2 Psmb2 8 39.8 Proteasome subunit beta type-3 Psmb3 5 34.1 Proteasome subunit beta type-4 Psmb4 6 33.3 Proteasome subunit beta type-5 Psmb5 13 35.6 Proteasome subunit beta type-6 Psmb6 3 17.6 Proteasome subunit beta type-7 Psmb7 3 21.3 Proteasome subunit beta type-8 Psmb8 6 22.8
Protein CASC4 Casc4 2 8 Protein Daple Ccdc88c 2 8.4
Protein deglycase DJ-1 Park7 6 29.6 Protein disulfide-isomerase A3 Pdia3 14 35 Protein disulfide-isomerase A4 Pdia4 2 9.6 Protein disulfide-isomerase A6 Pdia6 6 16.8
Protein disulfide-isomerase P4hb 4 18.3 Protein ERGIC-53 Lman1 19 30.2 Protein FAM26F Fam26f 2 7.3 Protein FAM3C Fam3c 18 46.7 Protein GPR108 Gpr108 3 6.7
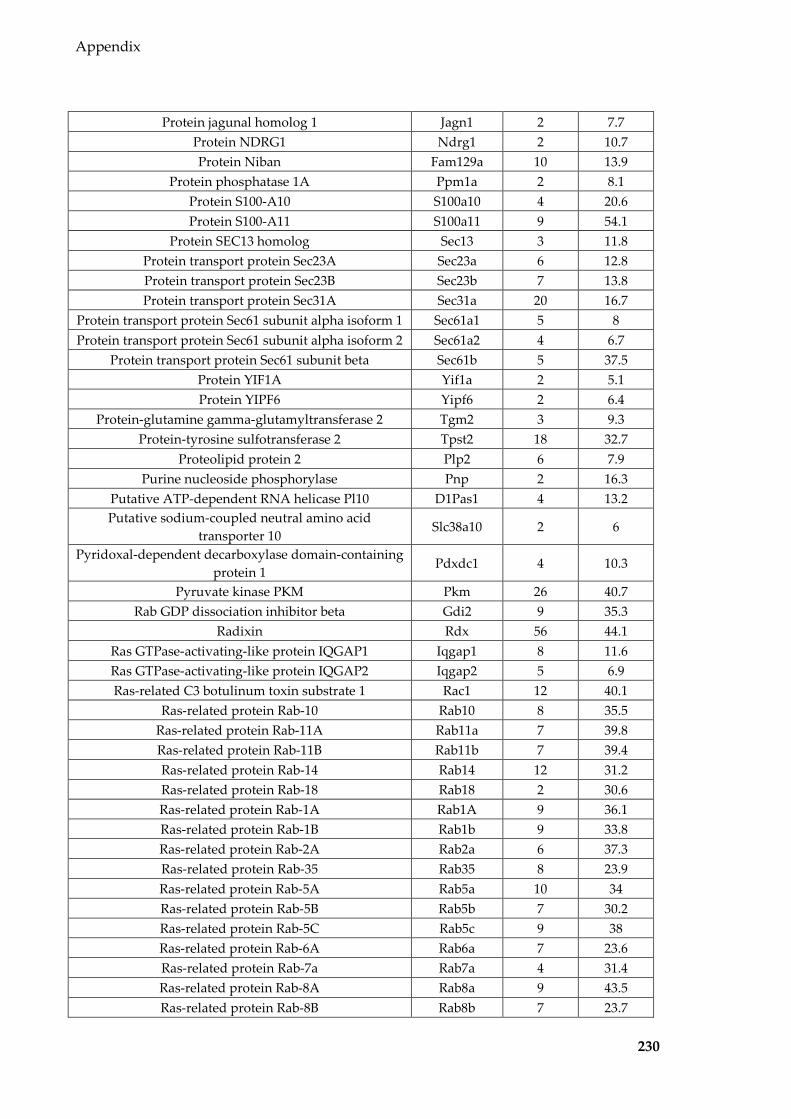
Appendix
230
Protein jagunal homolog 1 Jagn1 2 7.7 Protein NDRG1 Ndrg1 2 10.7 Protein Niban Fam129a 10 13.9
Protein phosphatase 1A Ppm1a 2 8.1 Protein S100-A10 S100a10 4 20.6 Protein S100-A11 S100a11 9 54.1
Protein SEC13 homolog Sec13 3 11.8 Protein transport protein Sec23A Sec23a 6 12.8 Protein transport protein Sec23B Sec23b 7 13.8 Protein transport protein Sec31A Sec31a 20 16.7
Protein transport protein Sec61 subunit alpha isoform 1 Sec61a1 5 8 Protein transport protein Sec61 subunit alpha isoform 2 Sec61a2 4 6.7
Protein transport protein Sec61 subunit beta Sec61b 5 37.5 Protein YIF1A Yif1a 2 5.1 Protein YIPF6 Yipf6 2 6.4
Protein-glutamine gamma-glutamyltransferase 2 Tgm2 3 9.3 Protein-tyrosine sulfotransferase 2 Tpst2 18 32.7
Proteolipid protein 2 Plp2 6 7.9 Purine nucleoside phosphorylase Pnp 2 16.3
Putative ATP-dependent RNA helicase Pl10 D1Pas1 4 13.2 Putative sodium-coupled neutral amino acid
transporter 10 Slc38a10 2 6
Pyridoxal-dependent decarboxylase domain-containing protein 1
Pdxdc1 4 10.3
Pyruvate kinase PKM Pkm 26 40.7 Rab GDP dissociation inhibitor beta Gdi2 9 35.3
Radixin Rdx 56 44.1 Ras GTPase-activating-like protein IQGAP1 Iqgap1 8 11.6 Ras GTPase-activating-like protein IQGAP2 Iqgap2 5 6.9 Ras-related C3 botulinum toxin substrate 1 Rac1 12 40.1
Ras-related protein Rab-10 Rab10 8 35.5 Ras-related protein Rab-11A Rab11a 7 39.8 Ras-related protein Rab-11B Rab11b 7 39.4 Ras-related protein Rab-14 Rab14 12 31.2 Ras-related protein Rab-18 Rab18 2 30.6 Ras-related protein Rab-1A Rab1A 9 36.1 Ras-related protein Rab-1B Rab1b 9 33.8 Ras-related protein Rab-2A Rab2a 6 37.3 Ras-related protein Rab-35 Rab35 8 23.9 Ras-related protein Rab-5A Rab5a 10 34 Ras-related protein Rab-5B Rab5b 7 30.2 Ras-related protein Rab-5C Rab5c 9 38 Ras-related protein Rab-6A Rab6a 7 23.6 Ras-related protein Rab-7a Rab7a 4 31.4 Ras-related protein Rab-8A Rab8a 9 43.5 Ras-related protein Rab-8B Rab8b 7 23.7

Appendix
231
Ras-related protein Ral-A Rala 4 34.5 Ras-related protein Ral-B Ralb 5 23.3
Ras-related protein Rap-1A Rap1a 8 40.2 Ras-related protein Rap-1b Rap1b 9 39.7 Ras-related protein R-Ras2 Rras2 6 41.2
Redox-regulatory protein FAM213A Fam213a 2 18.3 Reticulon-4 Rtn4 3 7.1
Retinal dehydrogenase 1 Aldh1a1 29 60.9 Retina-specific copper amine oxidase Aoc2 3 5.5
Retinol dehydrogenase 11 Rdh11 4 32.9 Retinol-binding protein 1 Rbp1 2 28.1
Rho GDP-dissociation inhibitor 1 Arhgdia 7 35.8 Rho guanine nucleotide exchange factor 18 Arhgef18 6 5.1
Rho-related GTP-binding protein RhoC Rhoc 4 36.8 Rho-related GTP-binding protein RhoG Rhog 2 22.5
Ribosome-binding protein 1 Rrbp1 31 24.7 RuvB-like 1 Ruvbl1 3 10.3 RuvB-like 2 Ruvbl2 2 5.2
Sarcoplasmic/endoplasmic reticulum calcium ATPase 1 Atp2a1 9 12.1 Sarcoplasmic/endoplasmic reticulum calcium ATPase 2 Atp2a2 31 32.3
SEC23-interacting protein Sec23ip 6 7 Secretory carrier-associated membrane protein 1 Scamp1 4 15.4 Secretory carrier-associated membrane protein 2 Scamp2 3 15.8 Secretory carrier-associated membrane protein 3 Scamp3 3 9.7
Septin-7 Sept7 3 11.7 Septin-8 Sept8 2 5.4
Serine incorporator 3 Serinc3 3 7.4 Serine protease 46 Prss46 4 6.7
Serine protease inhibitor A3K Serpina3k 9 22.7 Serine protease inhibitor Kazal-type 8 Spink8 2 18.1 Serine/threonine-protein kinase A-Raf Araf 2 8.6 Serine/threonine-protein kinase LATS1 Lats1 2 5.1 Serine/threonine-protein kinase mTOR Mtor 3 5.4
Serotransferrin Tf 32 66.4 Serum albumin Alb 93 75.2
Signal peptidase complex catalytic subunit SEC11A Sec11a 5 27.9 Signal peptidase complex subunit 2 Spcs2 6 35.8 Signal peptidase complex subunit 3 Spcs3 2 10.6
Signal recognition particle subunit SRP68 Srp68 6 17.9 Sodium/potassium-transporting ATPase subunit alpha-
1 Atp1a1 54 39.3
Sodium/potassium-transporting ATPase subunit alpha-2
Atp1a2 29 27
Sodium/potassium-transporting ATPase subunit alpha-4
Atp1a4 44 37
Sodium/potassium-transporting ATPase subunit beta-1 Atp1b1 4 19.7 Sodium/potassium-transporting ATPase subunit beta-3 Atp1b3 7 35.6
Appendix
232
Sodium-coupled neutral amino acid transporter 5 Slc38a5 14 10.6 Solute carrier family 2, facilitated glucose transporter
member 3 Slc2a3 66 20.9
Solute carrier family 2, facilitated glucose transporter member 5
Slc2a5 6 10.8
Solute carrier family 22 member 21 Slc22a21 4 5.3 Solute carrier family 28 member 3 Slc28a3 2 7.1
Solute carrier family 35 member F2 Slc35f2 4 6.1 Solute carrier family 52, riboflavin transporter, member
3 Slc52a3 4 8
Solute carrier organic anion transporter family member 2A1
Slco2a1 3 9.5
Sorbitol dehydrogenase Sord 10 32.8 Sorcin Sri 2 8.1 Sortilin Sort1 17 19.5
Sorting nexin-1 Snx1 2 7.1 Spectrin alpha chain, non-erythrocytic 1 Sptan1 12 12.7 Spectrin beta chain, non-erythrocytic 1 Sptbn1 21 12.9
Sphingosine-1-phosphate lyase 1 Sgpl1 3 8.1 Sphingosine-1-phosphate phosphatase 1 Sgpp1 2 5.3
Splicing factor, proline- and glutamine-rich Sfpq 3 7.3 Sterol-4-alpha-carboxylate 3-dehydrogenase,
decarboxylating Nsdhl 10 41.4
Structural maintenance of chromosomes protein 1A Smc1a 2 9.8 Sulfhydryl oxidase 2 Qsox2 2 11.3
Superoxide dismutase [Cu-Zn] Sod1 4 49.4 Surfeit locus protein 4 Surf4 12 26
Sushi domain-containing protein 2 Susd2 5 6.2 Synaptic vesicle membrane protein VAT-1 homolog Vat1 20 36.7
Synaptogyrin-4 Syngr4 4 10.7 Synaptophysin-like protein 1 Sypl1 2 17.6
Synaptosomal-associated protein 23 Snap23 2 17.6 Syntaxin-12 Stx12 3 20.1 Syntaxin-3 Stx3 2 14.9
Syntaxin-binding protein 2 Stxbp2 2 10.1 Syntenin-1 Sdcbp 2 17.7
Talin-1 Tln1 21 13.9 TAR DNA-binding protein 43 Tardbp 3 8
T-complex protein 1 subunit alpha Tcp1 12 36.7 T-complex protein 1 subunit beta Cct2 24 52.5 T-complex protein 1 subunit delta Cct4 25 44
T-complex protein 1 subunit epsilon Cct5 9 33.6 T-complex protein 1 subunit eta Cct7 16 29.8
T-complex protein 1 subunit gamma Cct3 20 45.1 T-complex protein 1 subunit theta Cct8 13 26.3 T-complex protein 1 subunit zeta Cct6a 14 33.1
Tetraspanin-3 Tspan3 2 18.2

Appendix
233
Tetraspanin-8 Tspan8 2 9.4 THAP domain-containing protein 4 Thap4 2 7.4
Thy-1 membrane glycoprotein Thy1 2 7.4 Trafficking protein particle complex subunit 5 Trappc5 2 17 Trafficking protein particle complex subunit 9 Trappc9 3 6.4
Transcriptional activator protein Pur-alpha Pura 12 44.2 Transcriptional activator protein Pur-beta Purb 4 17.9
Transforming protein RhoA Rhoa 4 36.8 Transgelin Tagln 2 10.4
Trans-Golgi network integral membrane protein 1 Tgoln1 2 5.1 Transitional endoplasmic reticulum ATPase Vcp 50 54.3
Transketolase Tkt 11 24.7 Translocon-associated protein subunit delta Ssr4 3 23.3
Translocon-associated protein subunit gamma Ssr3 2 12.4 Transmembrane 9 superfamily member 2 Tm9sf2 22 19.9 Transmembrane 9 superfamily member 3 Tm9sf3 3 10.1 Transmembrane 9 superfamily member 4 Tm9sf4 12 15.1
Transmembrane channel-like protein 4 Tmc4 6 10.5 Transmembrane channel-like protein 5 Tmc5 4 6.9
Transmembrane emp24 domain-containing protein 10 Tmed10 11 37.4 Transmembrane emp24 domain-containing protein 2 Tmed2 4 32.8 Transmembrane emp24 domain-containing protein 3 Tmed3 5 21.7 Transmembrane emp24 domain-containing protein 4 Tmed4 5 20.3 Transmembrane emp24 domain-containing protein 9 Tmed9 5 18.7
Transmembrane epididymal protein 1A Teddm1a 3 11.8 Transmembrane protein 109 Tmem109 9 21.8 Transmembrane protein 205 Tmem205 4 21.7 Transmembrane protein 33 Tmem33 2 8.5
Transthyretin Ttr 4 16.3 Triokinase/FMN cyclase Tkfc 9 20.6
Triosephosphate isomerase Tpi1 6 21.1 Tripeptidyl-peptidase 2 Tpp2 2 5.5
tRNA-splicing ligase RtcB homolog Rtcb 6 20.4 Tryptophan--tRNA ligase, cytoplasmic Wars 14 40.7
Tubulin alpha-1A chain Tuba1a 44 46.1 Tubulin alpha-1B chain Tuba1b 43 46.1 Tubulin alpha-4A chain Tuba4a 32 36.2 Tubulin beta-2A chain Tubb2a 45 50.3 Tubulin beta-2B chain Tubb2b 45 51.2 Tubulin beta-4B chain Tubb4b 47 51.2 Tubulin beta-5 chain Tubb5 42 50.5
Tyrosine-protein kinase FRK Frk 3 7 Ubiquitin carboxyl-terminal hydrolase isozyme L3 Uchl3 2 15.7
Ubiquitin-40S ribosomal protein S27a Rps27a 47 85.9 Ubiquitin-conjugating enzyme E2 N Ube2n 2 23.7
Ubiquitin-like modifier-activating enzyme 1 Uba1 5 12.2

Appendix
234
UDP-glucose:glycoprotein glucosyltransferase 1 Uggt1 12 7.7 UDP-glucuronosyltransferase 1-6 Ugt1a6 4 13.6
UDP-glucuronosyltransferase 1-7C Ugt1a7c 5 22.2 Unconventional myosin-Ib Myo1b 43 39.9 Unconventional myosin-Ic Myo1c 26 24.8 Unconventional myosin-Id Myo1d 16 23 Unconventional myosin-VI Myo6 21 19.2
Uroplakin-3b Upk3b 6 21.8 UTP--glucose-1-phosphate uridylyltransferase Ugp2 4 16.7 Vacuolar protein sorting-associated protein 35 Vps35 4 7.7
Valine--tRNA ligase Vars 20 22.2 Very-long-chain (3R)-3-hydroxyacyl-CoA dehydratase
2 Hacd2 7 13.4
Very-long-chain (3R)-3-hydroxyacyl-CoA dehydratase 3
Hacd3 4 8.6
Very-long-chain 3-oxoacyl-CoA reductase Hsd17b12 6 22.1 Very-long-chain enoyl-CoA reductase Tecr 17 31.2
Vesicle transport protein GOT1B Golt1b 7 26.8 Vesicle-associated membrane protein 2 Vamp2 3 28.4 Vesicle-associated membrane protein 3 Vamp3 3 32
Vesicle-trafficking protein SEC22b Sec22b 4 20 Vesicular integral-membrane protein VIP36 Lman2 13 20.1
Vigilin Hdlbp 3 7.2 Vimentin Vim 2 7.7 Vinculin Vcl 14 24.3
Vitronectin Vtn 10 28.7 Voltage-dependent anion-selective channel protein 1 Vdac1 3 10.5 Voltage-dependent anion-selective channel protein 2 Vdac2 7 11.2 Voltage-dependent calcium channel subunit alpha-
2/delta-1 Cacna2d1 6 9.7
V-type proton ATPase 116 kDa subunit a isoform 1 Atp6v0a1 7 13.2 V-type proton ATPase subunit B, brain isoform Atp6v1b2 4 11.2
V-type proton ATPase subunit d 1 Atp6v0d1 3 19.4 WAP four-disulfide core domain protein 15B Wfdc15b 11 62.5 WAP four-disulfide core domain protein 6B Wfdc6b 5 20.3
Xanthine dehydrogenase/oxidase Xdh 9 12.7

Appendix
235
Table 8.9 miRNAs (normalised to per million miRNA reads and averaged for each group)
detected in mouse epididymal EVs of backgrounds and diets
miRNA C57BL/6 EVs Control EVs MDS EVs mmu-let-7a-5p 29,859 25,465 20,713 mmu-let-7b-5p 17,546 13,050 12,170 mmu-let-7c-5p 67,213 50,657 49,185 mmu-let-7d-5p 11,786 11,609 9,911 mmu-let-7e-5p 2,355 2,043 1,844 mmu-let-7f-5p 112,255 156,183 200,699 mmu-let-7g-5p 49,749 58,971 63,347 mmu-let-7i-5p 21,617 23,286 27,932
mmu-miR-100-5p 9,810 8,934 8,085 mmu-miR-101a-3p 3,784 5,306 4,904 mmu-miR-101b-3p 1,174 1,375 1,693 mmu-miR-103-3p 687 996 1,035
mmu-miR-106b-3p 450 216 185 mmu-miR-10a-3p 208 270 254 mmu-miR-10a-5p 59,712 59,422 54,103 mmu-miR-10b-3p 92 112 98 mmu-miR-10b-5p 58,717 51,303 51,271 mmu-miR-122-5p 77 121 27
mmu-miR-1249-3p 49 171 196 mmu-miR-125a-5p 294 367 284
mmu-miR-125b-2-3p 170 284 231 mmu-miR-125b-5p 745 1,011 1,003 mmu-miR-126a-3p 627 1,007 1,098 mmu-miR-126a-5p 70 97 98 mmu-miR-127-3p 78 105 121 mmu-miR-127-5p 55 90 141
mmu-miR-133a-3p 394 340 292 mmu-miR-134-5p 78 116 140
mmu-miR-135a-5p 191 318 336 mmu-miR-136-3p 135 147 209 mmu-miR-136-5p 84 260 392 mmu-miR-140-3p 1,577 1,430 1,505 mmu-miR-141-3p 1,236 1,850 1,661 mmu-miR-141-5p 91 134 115
mmu-miR-142a-5p 122 160 113 mmu-miR-143-3p 30,337 30,992 38,037
mmu-miR-145a-3p 101 94 91 mmu-miR-145a-5p 356 473 638 mmu-miR-146a-5p 2,064 2,783 3,103 mmu-miR-146b-5p 2,111 1,916 2,066 mmu-miR-148a-3p 267,782 209,314 160,451 mmu-miR-148a-5p 145 148 111

Appendix
236
mmu-miR-148b-3p 1,456 1,139 1,146 mmu-miR-150-5p 77 107 138 mmu-miR-151-3p 406 386 300 mmu-miR-152-3p 4,140 4,390 5,347 mmu-miR-16-5p 352 472 606 mmu-miR-17-5p 144 213 328
mmu-miR-181a-5p 910 1,218 1,429 mmu-miR-181b-5p 180 203 222 mmu-miR-181c-5p 42 87 122 mmu-miR-181d-5p 71 103 105 mmu-miR-182-5p 10,599 9,922 8,479 mmu-miR-183-5p 3,389 3,454 3,456
mmu-miR-1839-5p 5,421 4,741 5,699 mmu-miR-1843a-5p 78 103 99 mmu-miR-1843b-5p 131 141 153
mmu-miR-184-3p 914 415 549 mmu-miR-185-5p 205 275 269 mmu-miR-186-5p 339 404 479 mmu-miR-187-3p 62 108 116 mmu-miR-191-5p 2,094 2,515 2,995 mmu-miR-192-5p 741 606 602 mmu-miR-194-5p 121 111 96
mmu-miR-195a-5p 112 203 200 mmu-miR-196a-5p 382 367 350 mmu-miR-196b-5p 349 557 602 mmu-miR-1981-5p 141 175 243 mmu-miR-199a-3p 4,241 4,764 5,334 mmu-miR-199a-5p 546 824 1,087 mmu-miR-199b-3p 2,973 3,397 3,693 mmu-miR-199b-5p 460 730 903 mmu-miR-19b-3p 80 82 114 mmu-miR-1a-3p 1,725 1,565 3,149
mmu-miR-200a-3p 31,397 30,249 25,825 mmu-miR-200a-5p 238 307 206 mmu-miR-200b-3p 15,326 17,882 18,582 mmu-miR-200b-5p 91 168 216 mmu-miR-200c-3p 7,478 9,432 9,985 mmu-miR-203-3p 724 803 918 mmu-miR-204-5p 779 985 992 mmu-miR-205-5p 369 640 677 mmu-miR-206-3p 1,533 1,554 1,600 mmu-miR-20a-5p 284 497 661 mmu-miR-210-3p 169 197 229 mmu-miR-2137 71 96 97
mmu-miR-214-3p 67 132 186 mmu-miR-218-5p 223 281 364

Appendix
237
mmu-miR-21a-5p 38,736 53,450 55,737 mmu-miR-221-3p 548 588 705 mmu-miR-222-3p 506 673 842 mmu-miR-22-3p 638 721 714
mmu-miR-224-5p 117 70 64 mmu-miR-23a-3p 2,831 3,312 3,191 mmu-miR-23b-3p 1,454 1,859 1,900 mmu-miR-24-2-5p 169 252 234 mmu-miR-24-3p 765 1,143 1,158 mmu-miR-25-3p 502 536 405
mmu-miR-26a-5p 14,163 15,239 16,052 mmu-miR-26b-5p 3,207 3,464 3,845 mmu-miR-27a-3p 1,222 1,525 1,444 mmu-miR-27a-5p 122 169 122 mmu-miR-27b-3p 3,965 4,771 4,248 mmu-miR-28a-3p 149 116 108 mmu-miR-29a-3p 4,907 5,727 5,896 mmu-miR-29b-3p 306 457 592 mmu-miR-29c-3p 131 272 380 mmu-miR-30a-3p 887 900 818 mmu-miR-30a-5p 15,142 14,981 13,497 mmu-miR-30b-5p 2,755 3,225 3,134
mmu-miR-30c-2-3p 217 170 124 mmu-miR-30c-5p 5,134 5,998 5,385 mmu-miR-30d-5p 2,100 2,208 1,617 mmu-miR-30e-3p 509 416 329 mmu-miR-30e-5p 887 1,319 1,222 mmu-miR-320-3p 1,492 1,147 955 mmu-miR-324-3p 82 85 98 mmu-miR-328-3p 79 164 167 mmu-miR-340-5p 1,114 1,585 1,800 mmu-miR-342-3p 69 136 145 mmu-miR-345-5p 128 140 188 mmu-miR-34a-5p 120 248 389 mmu-miR-34b-3p 121 116 201 mmu-miR-34c-5p 1,265 1,040 1,514 mmu-miR-3535 175 122 138
mmu-miR-374b-5p 275 379 429 mmu-miR-375-3p 1,137 1,225 1,191
mmu-miR-378a-3p 1,440 1,894 1,806 mmu-miR-378c 105 136 109
mmu-miR-379-5p 1,984 2,433 3,464 mmu-miR-381-3p 90 121 123 mmu-miR-382-5p 154 130 170 mmu-miR-411-5p 430 634 798 mmu-miR-423-3p 122 199 165

Appendix
238
mmu-miR-423-5p 350 278 261 mmu-miR-425-5p 199 245 246 mmu-miR-429-3p 1,729 2,542 3,016 mmu-miR-434-3p 75 119 141 mmu-miR-451a 160 259 249
mmu-miR-465a-5p 178 160 156 mmu-miR-465b-5p 1,295 629 703 mmu-miR-465c-5p 763 428 514 mmu-miR-467a-5p 41 77 102 mmu-miR-470-5p 270 277 279
mmu-miR-486a-5p 129 363 207 mmu-miR-486b-5p 52 144 94 mmu-miR-532-5p 336 252 235 mmu-miR-652-3p 1,175 1,364 1,416 mmu-miR-669c-5p 187 253 462 mmu-miR-672-5p 886 866 751 mmu-miR-676-3p 121 118 95
mmu-miR-743a-3p 85 59 91 mmu-miR-743b-3p 116 91 80 mmu-miR-744-5p 104 156 150 mmu-miR-7a-5p 1,339 1,460 2,203
mmu-miR-878-5p 313 215 281 mmu-miR-881-3p 3,123 1,210 1,097 mmu-miR-92a-3p 447 758 614 mmu-miR-93-5p 251 364 471 mmu-miR-9-3p 189 343 453 mmu-miR-9-5p 1,451 2,964 3,010
mmu-miR-96-5p 133 179 279 mmu-miR-98-5p 781 795 857
mmu-miR-99a-5p 12,380 16,407 14,054 mmu-miR-99b-5p 1,276 1,207 983

Appendix
239
Table 8.10 tRFs (normalised to per million tRF reads and averaged for each group)
detected in mouse epididymal EVs of different backgrounds and diets
tRF C57BL/6 EVs Control EVs MDS EVs tRF-Ala-AGC 94 108 81 tRF-Ala-CGC 316 490 625 tRF-Ala-TGC 89 124 99 tRF-Arg-ACG 107 70 51 tRF-Arg-CCG 265 229 244 tRF-Arg-CCT 447 724 546 tRF-Arg-TCG 52 42 35 tRF-Arg-TCT 46 46 46 tRF-Asn-GTT 179 191 144 tRF-Asp-GTC 1,275 3,233 3,303 tRF-Cys-GCA 428 267 213 tRF-Gln-CTG 930 895 845 tRF-Gln-TTG 338 332 300 tRF-Glu-CTC 85,519 105,080 87,793 tRF-Glu-TTC 19,161 18,342 15,702 tRF-Gly-ACC 84 154 134 tRF-Gly-CCC 114,415 105,594 105,372 tRF-Gly-GCC 744,940 688,874 684,439 tRF-Gly-TCC 307 359 446 tRF-His-ATG 3 1 2 tRF-His-GTG 9,691 22,656 26,949 tRF-Ile-AAT 87 116 110 tRF-Ile-TAT 61 95 82
tRF-Leu-AAG 178 267 209 tRF-Leu-CAA 109 132 114 tRF-Leu-CAG 513 800 597 tRF-Leu-TAA 55 83 108 tRF-Leu-TAG 174 223 206 tRF-Lys-CTT 1,950 2,440 2,438 tRF-Lys-TTT 1,105 2,281 2,284 tRF-Met-CAT 855 1,089 1,523 tRF-Phe-GAA 35 31 26 tRF-Pro-AGG 137 147 150 tRF-Pro-CGG 89 105 101 tRF-Pro-TGG 182 188 177 tRF-Ser-AGA 151 172 202 tRF-Ser-CGA 19 27 29 tRF-Ser-GCT 209 478 425 tRF-Ser-GGA 1 0 1 tRF-Ser-TGA 68 81 82
tRF-STOP-TCA 118 127 121 tRF-STOP-TTA 0 0 0

Appendix
240
tRF-Thr-AGT 52 40 42 tRF-Thr-CGT 29 29 28 tRF-Thr-TGT 67 66 60 tRF-Trp-CCA 218 130 99 tRF-Tyr-GTA 114 157 100 tRF-Val-AAC 4,548 13,236 19,851 tRF-Val-CAC 9,774 28,602 42,275 tRF-Val-TAC 417 1,046 1,190

Appendix
241
Table 8.11 BLAST hits for unmappable sequence in mouse epididymal EVs
Sequence definition Accession Match Match length
Start End
Mus musculus 18S ribosomal RNA genes, partial sequence; 5.8S ribosomal RNA gene, complete sequence; and 28S ribosomal RNA
gene, partial sequence
AH002077.2 100% 36 3,279 3,314
Mus musculus clone contig 3 chromocenter region genomic sequence
KX121612.1 100% 36 622 587
Mus musculus 45S pre-ribosomal RNA (Rn45s), ribosomal RNA
NR_046233.2 100% 36 8,123 8,158
TPA_exp: Mus musculus ribosomal DNA, complete repeating unit
BK000964.3 100% 36 8,123 8,158
Mus musculus strain BALB/c 45S ribosomal RNA region genomic sequence
GU372691.1 100% 36 11,134 11,169
Mus musculus 28S ribosomal RNA (Rn28s1), ribosomal RNA
NR_003279.1 100% 36 1 36
Mus musculus CNR gene for cadherin-related neuronal receptor, complete cds
AB114630.1 100% 36 2,305 2,340
Mus musculus 18S ribosomal RNA gene, partial sequence; internal transcribed spacer
1, 5.8S ribosomal RNA gene and internal transcribed spacer 1, complete sequence; and
28S ribosomal RNA gene, partial sequence
J00623.1 100% 36 2,477 2,512
M.musculus 45S pre rRNA gene X82564.1 100% 36 13,758 13,793 Mouse 28S large subunit rRNA, 5' end M19226.1 100% 36 1 36
Mouse 28S ribosomal RNA X00525.1 100% 36 1 36
Related Documents











