1998;58:1245-1252. Cancer Res Mark L. Gonzalgo, Toshiro Hayashida, Christina M. Bender, et al. Locus in Human Bladder Cancer Cell Lines p19/p16 The Role of DNA Methylation in Expression of the Updated version http://cancerres.aacrjournals.org/content/58/6/1245 Access the most recent version of this article at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications on March 11, 2014. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on March 11, 2014. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1998;58:1245-1252. Cancer Res Mark L. Gonzalgo, Toshiro Hayashida, Christina M. Bender, et al. Locus in Human Bladder Cancer Cell Lines
p19/p16The Role of DNA Methylation in Expression of the
Updated version
http://cancerres.aacrjournals.org/content/58/6/1245
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 58. 1245-1252. March 15. 19981
The Role of DNA Methylation in Expression of the pl9/pl6 Locus in HumanBladder Cancer Cell Lines1
Mark L. Gonzalgo, Toshiro Hayashida, Christina M. Bender, Martha M. Pao, Yvonne C. Tsai, Felicidad A. Gonzales,Hung D. Nguyen, TuDung T. Nguyen, and Peter A. Jones2
Department nf Biochemistry and Molecular Biology, Urologie Cancer Research Laboratory. USC/Norris Comprehensive Cancer Center, University of Southern California. Schoolof Medicine. Los Angeles, California 90033
ABSTRACT
Methylation of CpG sites in the control regions of tumor suppressorgenes may be an important mechanism for their heritable, yet reversible,transcriptional inactivation. These changes in methylation may impair theproper expression and/or function of cell cycle regulatory genes andconfer a selective growth advantage to affected cells. Detailed methylationanalysis using genomic bisulfite sequencing was performed on a series ofsubclones of a bladder cancer cell line in which a hypermethylated pl6gene had been reactivated by transient treatment with 5-aza-2'-deoxycy-
tidine. Methylation of the CpG island in the promoter of the pi6 gene inhuman bladder cancer cells did not stop the formation of a transcriptinitiated 20 kb upstream by the/7/9 promoter but did prevent the expression of a pi6 transcript. Furthermore, we show that reactivant clones thatexpressed pl6 at varying levels contained heterogeneous methylation patterns, suggesting that pió expression can occur even in the presence of arelatively heavily methylated coding region. We also present the tirsifunctional evidence that methylation of only a small number of CpG sitescan significantly down-regulate pJ6 promoter activity, thus providing
support for the model of progressive inactivation of this tumor suppressorgene by DNA methylation.
INTRODUCTION
The overall functions of DNA methylation in mammalian cellsremain enigmatic, despite the fact that it is essential for development(1). Methylation of CpG islands is clearly important for the regulationof imprinting (2-5) and for reinforcing the transcriptional inactivities
of housekeeping genes on the inactive X chromosome (6, 7). Becausemethylation of CpG-rich promoters has been shown to limit theiractivities (8-11 ), it has generally become accepted that methylation ofCpG islands is associated with a lack of transcription (12-14).
Not all genes contain CpG islands, but most islands that have beenidentified are associated with genes (15, 16). The position of CpGislands relative to the transcriptional start sites of genes varies considerably. They are always located in the 5' region of housekeeping
genes but are often confined to the coding regions of tissue-specific
genes. Additionally. CpG islands associated with parasitic repetitiveDNAs are often embedded within the transcriptional units of activegenes, and it has been suggested that the primary function of DNAmethylation may be related to the suppression of parasitic transpos-
able elements (17, 18). Many of these repetitive sequences are observed to be methylated in the genome, presumably as a result of ahost-mediated defense mechanism for their inactivation. Indeed, one-
third of the human genome consists of interspersed repetitive elementssuch as Alus and LINES, many of which fulfill conditions of beingCpG islands (16, 19, 20) and can potentially be transcriptionallyactive (21, 22). Several eukaryotic genes, such as ¡g/2(23, 24) and
Received 6/27/97; accepted 1/16/98.The costs of publication of this article were defrayed in part hy the payment of page
charges. This article must therefore he hereby marked iitÃvertisenient in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by USPHS Grants 5 R35 CA49758 and 5 UdìCA70903
from the National Cancer Institute.2 To whom requests for reprints should be addressed, at USC/Norris Comprehensive
Cancer Center, 1441 Eastlake Avenue. Room 815, Mail Stop #83. Los Angeles. CA90033. Phone: (2131764-0816; Fax: (213)764-0102.
pl6 (25), have CpG-rich promoters downstream of additional promot-
er(s), which can direct transcription through CpG islands. Manytranscriptional units, therefore, contain more than one CpG island, andthese may be associated with promoters, coding regions, or repetitiveelements.
The protein product of the />!6 tumor suppressor gene binds tocyclin-dependent kinases 4 and 6 and inhibits their interaction with
cyclin DI. which normally mediates passage through the G, phase ofthe cell cycle by phosphorylation of pRB (25). The />/6 gene is locatedon chromosome 9p21 and has been reported to be a frequent target ofhomozygous deletion in a variety of human cancer cell lines andtumors (26-28). Intragenic mutation of piò in both cell lines and
primary tumors has been found to be another mechanism of geneinactivation. albeit at a lower frequency than for other tumor suppressor genes such as p53 (13, 29. 30). Methylation of the 5' CpG island
oÃpió has been proposed as yet another mechanism for inactivationof this tumor suppressor gene in —20-40% of certain types of human
cancers and may account for the low frequency of reported pinmutations in primary tumors (31. 32). Numerous studies have documented aberrant methylation of the 5' CpG island of pin in various
human tumors (11, 31, 33, 34). Furthermore, treatment of humancancer cell lines with 5-Aza-CdR' has been correlated with induction
of pl6 expression and demethylation of certain CpG sites as determined by restriction enzyme digestion and Southern analysis (11, 35).It has also been suggested that methylation of the 5' CpG island of
p!6 may play a role in increasing the metastatic potential of tumorcells, although this has not been definitively proven (36).
The novel situation that exists with regard to the pió locus is thepresence of an alternative transcript that is driven by the pl9 promoteroriginating —20kb upstream of the pió exon 1a coding domain. The
pI6 and p 19 transcripts use alternative first exons (la and Iß)joinedthrough the same splice acceptor site to exon 2 coding sequences butin different reading frames (37, 38). The pl9 transcript encodes aprotein that has also been shown to be capable of suppressing growthin mammalian fibroblasts (39). The 5' CpG island of the ¡>lf>gene,
which includes the exon la coding domain, is a highly CpG-rich
sequence through which transcriptional machinery must pass to generate the ¡>I9transcript.
In this report, we show that extensive methylation of the CpG islandin the promoter of the pl6 gene blocks gene expression but noi theexpression of a pl9 transcript. The ability of the />/6 promoter togenerate a transcript was restored by treatment of a human bladdercancer cell line with 5-Aza-CdR, suggesting that promoters located
within other transcriptional units might also be silenced by DNAmethylation. One important function of DNA methylation may be tosilence the activities of CpG-rich promoters present in actively tran
scribed regions of mammalian genes. We have also performed the firstcomprehensive analysis of 67 CpG dinucleotides in the p I ft promoterregion in a human bladder cancer cell line before and after treatmentwith 5-Aza-CdR. Bisulfite genomic sequencing allowed us to char-
'The abbreviations used are: 5-AM-CdR, 5-aza-2'-deoxycytidine; CiAI'DM, glyceral-
dehyde-3-phosphate dchydrogenase; CAT, chloramphenicol acetyltransferasc: RT-PCR.reverse transcription-PCR.
1245
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ROLE OF DNA METHYLATION IN EXPRESSION OF PIWPI6
acterize methylation patterns of the pió 5' CpG island in reactivant
clones as a function of gene expression. Furthermore, we have investigated the effects of site-specific methylation of the p!6 promoter asa potential initiating event in the progressive down-regulation of this
tumor suppressor gene.
MATERIALS AND METHODS
Cell Lines and 5-Aza-CdR Treatment. T24 and J82 human bladdercarcinoma-derived cell lines were obtained from the American Type CultureCollection (Rockville. MD). DNA and total RNA were isolated from noncon-fluent, exponentially growing cells. Cell lines were plated at I X IO5 per75-mnr tlask and treated 24 h later with a 3 X 1CT7 M dose of 5-Aza-CdR.
Cell medium was changed 24 h after addition of 5-Aza-CdR. and single cell
subclones were isolated between 48 and 72 h after removal of the drug.Conditioned media was used for the initial culturing of single-cell subclones.
DNA and RNA were isolated after at least 20 population doublings from eachof the subclones.
RT-PCR Analysis of pl6, pl9, and GAPDH Expression. Total RNA(~2.5 j^g) was isolated from cells and reverse transcribed using random
hexamers, deoxynucleotide triphosphates (Boehringer Mannheim, Indianapolis. IN), and Superscript II reverse transcriptase (Life Technologies, Inc.,Gaithersburg, MD) in a 25-ß\reaction. cDNA was amplified using primers
specific for pI6, pl9, or GAPDH. Briefly, PCR reactions were performed in25-f¿lvolumes for 94°Cfor 3 min, 24 cycles at 94°Cfor 1 min. 56°Cfor 30 s,72°Cfor 40 s (pió and pI9 amplification), and 94°Cfor 1 min, 20 cycles at94°Cfor I min, 58°Cfor 30 s. 72°Cfor 45 s (GAPDH amplification). Primers
sequences have been described previously (33) and are as follows: pl6 sense,5'-AGC CTT COG CTG ACT GGC TGG-3'; pió antisense. 5'-CTG CCCATC ATC ATG ACC TOG A-3': pl9 sense. 5'-TGG CGC TOC TCA CCTCTG GT-3': pI9 antisense. 5'-GCG CTG CCC ATC ATC ATG AC-3';GAPDH sense. 5'-CAG CCG AGC CAC ATC G-3': and GAPDH antisense,5'-TGA GGC TGT TGT CAT ACT TCT C-3'. PCR products were resolved
on 2% agarose gels, transferred to Zetaprobe membrane (Bio-Rad, Richmond,CA), and probed with a digoxigenin-labeled internal oligonucleotide.
PCR-based Methylation Assay. A PCR-based methylation assay de
scribed previously (33) was used to analyze various regions of the pI9/plogene locus. Briefly. 2 /j.g of genomic DNA was incubated with 2 units of eitherHpall, Cfi>\, or Msp\ restriction enzymes prior to PCR amplification. Primersused for methylation analysis of each region are as follows: exon 1/3 sense.5'-GCT CAC CTC TGG TGC CAA AGG GC-3'; exon Ißantisense, 5'-AGGAGC CAG CGT CTA GGG CAG-3'; 5' Alu sense, 5'-TCA GGG ACT AAGTTC TTC TTC GTC-3': 5' Alu antisense, 5'-TGC TTT GGA TAG AAT TATCAC TAT TAC-3'; 3' Alu sense, 5'-GTA ATA GTG ATA ATT CTA TCCAAA GCA-3'; 3' Alu antisense, 5'-GCA ATC TTC TTT TOA GGC ATATA A-3'; exon la sense, 5'-AGC CTT CGC CTG ACT GGC TGG-3', exonla antisense, 5'-CTG GAT CGG CCT CCG ACC GTA-3': exon 2 sense,5'-CTG CTT GGC GGT GAG GGG G-3': and exon 2 antisense, 5'-CCT CACCTG AGG G AC CTT C-3'. PCR conditions for each region were: exon Iß.22cycles at 94°Cfor 1 min. 57°Cfor 30 s, 72°Cfor 1 min; 5' and 3' Alus, 30cycles at 94°Cfor 1 min, 58°Cfor 1 min, 72°Cfor 1 min: and exons la and2. 28 cycles at 94°C for 1 min, 56°Cfor 45 s, 72°Cfor 45 s. Sequence
information for the region upstream of the piò gene has been deposited in theGenBank database (accession no. AF022809).
Bisulfite Genomic Sequencing Protocol. The method described by From-mer's group (40. 41 ) was used to react —2.0/^g of DNA isolated from parent
bladder cancer cell lines and subclones with sodium bisulfite. Primary andsecondary PCR reactions were performed on bisulfite-treated DNA for 30cycles each at 94°Cfor 1 min. 58°Cfor 1 min, and 72°Cfor 1 min. PrimersAB 1a (5'-AAT AAT GTG TAA TTT TGT TTT TAG AAT ATT GAG-3') andABlb [5'-CTC (A/G)CC AAA ACC AAC (A/G)TT AAC-3'] were used togenerate the primary PCR template for regions A and B. Primers Cla (5'-GAGGGG GTA GGG GGA TAT-3') and Clb [5'-ACC AAT CAA CC(A/G) AAAACT CCA TAC TA-3'] were used for primary PCR amplification for regionC. Primers Dia (5'-GTA GGT GGG GAG GAG TTT ACT T-3') and Dlb(5'-CTA CAÕ ACC CTC TAC CCA CCT AA-3') were used for primary PCR
amplification for region D. Primers ABla and A2b (5'-AAA ACT ATT CTTTCC TAA TTA TAA AAA CC-3') were used for secondary PCR amplifica
tion of region A. Primers B2a (5'-TTT GGT GGG GTT TTT ATA ATTAGG-3') and B2b (5'-AAA CTA AAC TCC TCC CCA CCT A-3') were usedfor secondary PCR amplification of region B. Primers Dia and D2b (5'-CCC
ACC CTC TAA TAA CCA ACC AA-3') were used for secondary PCRamplification of region C. Primers D2a (5'-AGA GGA GGG GTT GGT TGGTTA TT-3') and Dlb were used for secondary PCR amplification of region D.
PCR products were gel purified and ligated into the pCRII cloning vector(Invitrogen. San Diego, CA). Individual plasmid molecules were then se-
quenced using the Sequenase version 2.0 kit (Amersham Corp., Cleveland,
OH).Construction, Methylation, and Transfection of pl6PRO-CAT Vector.
A 2.1-kb DNA fragment isolated from a genomic DNA library (Stratagene, La
Jolla. CA) was used as a template for PCR amplification of the region upstreamof the pió exon la ATG initiation codon. The 5' primer included the froRIsite at position —869and the 3' primer replaced the ATG with a Hindttl site.
A Sad site was created in the pió promoter region by changing the nucleotideat position -341 relative to ATG (GAGCÇC to GAGCTC) by site-directed
mutagenesis. This nucleotide change did not reduce basal promoter activity inthe CAT assay system. The DNA fragment was ligated into the pSVO-CATvector to create pl6PRO-CAT. In vitro methylation was performed by incubating the pl6PRO-CAT plasmid with either 3 units of M.Hpall or M.Hhal. A
methylation cassette assay was also used to generate additional constructs byincubating l ¡¿gof plasmid DNA with either 3 units of M.Hpall (New EnglandBiolabs, Beverly. MA) followed by digestion with Sad and BslEll or 3 unitsof M.S.s.vIfollowed by digestion with EcoRl and Sad. Restricted DNA fragments were separated by agarose gel electrophoresis, purified, and ligated toeither unmethylated or methylated plasmid DNA to produce the desiredconstruct. Equal amounts of plasmid were introduced into C-33A cells byliposome-mediated transfection with pRSV-LacZ. CAT activity was measured48 h after transfection by '4C radioactive quantitation. The relative promoter
activity was calculated and normalized via ß-galactosidase activity.
RESULTS
Analysis of pI9 and pl6 Expression in Subclone Populations.The correlation between methylation and transcriptional silencing ofthe p 16 gene has been well established (11, 31, 33) as well as themodulation of gene expression in culture by the use of demethylatingagents such as 5-Aza-CdR (11. 42. 43). We examined the relationshipbetween CpG island methylation and expression in the p 19/16 transcriptional unit in the J82 and T24 bladder cancer cell lines that do ordo not express a p!6 transcript, respectively (Fig. 1). Both cell typeswere found to express equivalent levels of the p 19 gene. Clones ofT24 cells expressing substantial quantities of pió mRNA could beobtained following brief treatment with 5-Aza-CdR. and these showedno changes in the levels of pl9 transcript (Fig. 1). The J82 humanbladder carcinoma-derived cell line was used as a positive control forpió expression because this cell line normally expresses pl6 at highlevels. A T24 subclone derived from a parent culture that was nottreated with 5-Aza-CdR (clone 1) showed no detectable p 16 expression. Clone 2 was similar to many of the clones derived from parentcultures treated with 5-Aza-CdR and also showed no detectable pl6expression. Reactivant clone 7 had the least expression of piómRNAfollowed in increasing order by clone 3, then clone 4. The levels ofpl9 mRNA expression were virtually identical for all samples analyzed, indicating that transcription of this gene was not affected bydifferential expression of p!6.
Long Range Methylation Analysis of the pi9 and pi6 Loci. Fig.2 shows results from PCR-based methylation analysis oipl9 exon Iß,Alu elements located upstream of exon la. piò exon la codingregion, and exon 2 in the bladder cancer cell lines described previously. This assay is qualitative rather than quantitative, and thepresence of bands in the uncut (U) DNA lanes served as a positivecontrol for PCR amplification, whereas absence of bands in the Cfo\(C)- and Hpall (H)-digested DNAs indicated that methylation was notpresent in at least one of the enzyme sites examined. The methylation
1246
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ROLE OF DNA METHYLATION [N EXPRESSION OF PlWPItt
Z T- oà n "t r*.Q O 4) 4) Q) Q)U C C C C C
m O .2 .2 .2 .2 £" Z O O O O O
ptomRNA
pÃSmRNA
GAPDHmRNA
Fig. 1. Induction of pit) expression assayed by RT-PCR in T24 subclones. Single-cellsubclones were isolated after treatment of parent cultures with 3 X 10"7 M 5-A/.a-CdR.
J82 cells were used as a positive control for/>/6 expression. T24 clone 1 was derived fromcells that were not treated with 5-Aza-CdR and showed no detectable levels of pl6expression. Clone 2 originated from a parent culture treated with 5-Aza-CdR and showedno pió expression. Varying levels of pió expression were observed in subclones derivedfrom T24 parent cultures treated with 3 X 10~7 M 5-Aza-CdR. In increasing order of pió
expression were: clone 7 < clone 3 < clone 4. Parent T24 cultures were treated for 24 hwith 5-Aza-CdR. followed by subcloning —48h after drug removal. Levels of ;>/9 and
GAPDH expression were constant for all cell lines examined.
during S phase of the first cell division after drug treatment. We havedefined 5-Aza-CdR-affected molecules as those which contained atleast two unmethylated CpGs in tandem or a total of three unmethy-lated CpG sites within a region amplified by PCR of bisulfite-treated
DNA, because these methylation patterns were never observed inmolecules sequenced from the untreated parent T24 culture. 5-Aza-CdR-unaffected molecules were defined as having less than three
unmethylated CpG sites for each region amplified by PCR and werepresumably derived from parent DNA strands that did not incorporate5-Aza-CdR during S phase of the first division after drug treatment.
Interestingly, 105 of 245 (43%) of the total number of individualmolecules examined for all />/6-expressing subclones were classifiedas being 5-Aza-CdR unaffected, and 140 of 245 (57%) were definedas being 5-Aza-CdR-affected molecules. Our hypothesis was thatDNA molecules that were methylated (5-Aza-CdR-unaffected) did
not express p¡6, whereas molecules that were demethylated by theeffects of 5-Aza-CdR (5-Aza-CdR-affected) were competent for pió
expression. This is also supported by the finding that clone 2, whichwas derived from a parent culture treated with 5-Aza-CdR but showedno detectable pl6 expression, contained an almost fully methylated 5'
CpG island similar to the untreated T24 cultures (data not shown).Consensus methylation patterns at each CpG site were, therefore,generated for each of the T24 subclones that expressed pI6, considering the data from 5-Aza-CdR-affected molecules to characterize
status of seven Cfol sites and one Hpall site in exon Ißof the pl9gene was monitored using the PCR-based assay. Analysis of both 5'and 3' Alii elements located approximately 1.3 kb upstream of the pl6
exon la translation start codon revealed methylation at all sites (twoCfol sites and one Hpa\\ site in each/4/«) as indicated by the presenceof bands in the Cfol- and //pall-digested lanes for all samples ana
lyzed. Analysis of two Cfol sites and one Hpall site in the exon 1acoding region showed methylation in T24 cell line DNA. absence ofmethylation in J82 cell line DNA, and partial methylation in clone 4DNA. Exon 2 was found to be methylated at seven Cfol sites and fourHpall sites in all of the samples. These findings demonstrated that apl9 transcript can be formed, even in the presence of heavily methylated CpG-rich sequences located downstream of its promoter.
Methylation Status of T24 and J82 Cell Lines. Bisulfite genomicsequencing (40. 41) was used to determine the methylation patterns inbladder cancer cell lines in greater detail by dividing a 1-kb region
that included p 16 exon la and upstream sequences into 4 regions (Ato D) defined by the PCR primers used to amplify bisulfite-treatedDNA. DNA sequences extending to position —869relative to the pió
ATG initiation codon were included in this study because this regionhas been reported previously to be sufficient for basal pI6 promoteractivity (44). PCR products were cloned into the TA vector (Invitro-
gen. San Diego, CA) after amplification of each region, and individualplasmid molecules were sequenced. Sequencing of 80 plasmid clonesrevealed a complete absence of methylation at all 67 CpG sites in theJ82 cell line (0%) and a close to fully methylated pattern at all CpGsin the T24 cell line (97%; data not shown).
Effects of 5-Aza-CdR on Methylation Patterns in T24 Sub-
clones. Typical results from sequencing individual plasmid molecules for determination of CpG methylation status in a/j/6-expressing
subclone (clone 4) isolated after treatment of the parent T24 cells with5-Aza-CdR are shown in Fig. 3. Because 5-Aza-CdR only acts as an
inhibitor of methylation of newly synthesized DNA (45), we were notsurprised to find heterogeneous mixtures of molecules in which someDNA molecules were completely unaffected by the drug, whereasother DNA molecules contained areas of demethylation consistentwith the incorporation of 5-Aza-CdR into parental DNA strands
JLJLI.. . .U...3 .III.... . .A.1II 1 ' 1 «. Il «.1ZOObpT24U
C H1
w K
J82MU C M MJL
..nati._sJ1—î—1"BClone
4UC HMExon
1ß—̂ ••T24U
C HJ82M U C H MClone4U
C HM5'Alu
—* --- -••---T24U
C HJ82M U C H MClone4U
C HMy
Alu —* --.---T24U
C HJ82M U C H MClone4U
C HMExonla—
999 ••••T24U
C HJ82M U C H MClone4U
C HMExon2
^— ... — -
Fig. 2. PCR-based melhylation analysis of exon 1ß,Alu elements, exon l a, and exon2 in human bladder cancer cell lines. T<>/>.relative positions of exon coding regions, Aluelements, CpG sites (tick marks), and restriction enzymes sites used for PCR-basedmethylation analysis. Lanes marked U (undigested), C (Cf<>\).H (Hpa\\), and M (Afv/ïl)indicate that DNAs were digested with the corresponding restriction enzyme before PCRamplification using primers flanking the region of interest. The presence of bandsindicates melhylation at all monitored CpG sites within the region of PCR amplification.The absence of bands indicates lack of methylalion of at least one site within the regionamplified by PCR. Uncut ((/) DNA served as a control for PCR amplification; digestionof DNA with A/.v/il(M) before PCR served as a control for restriction en/yme digestion.
1247
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
ROLE OF DNA METHYLATION IN EXPRESSION OF PIWP16
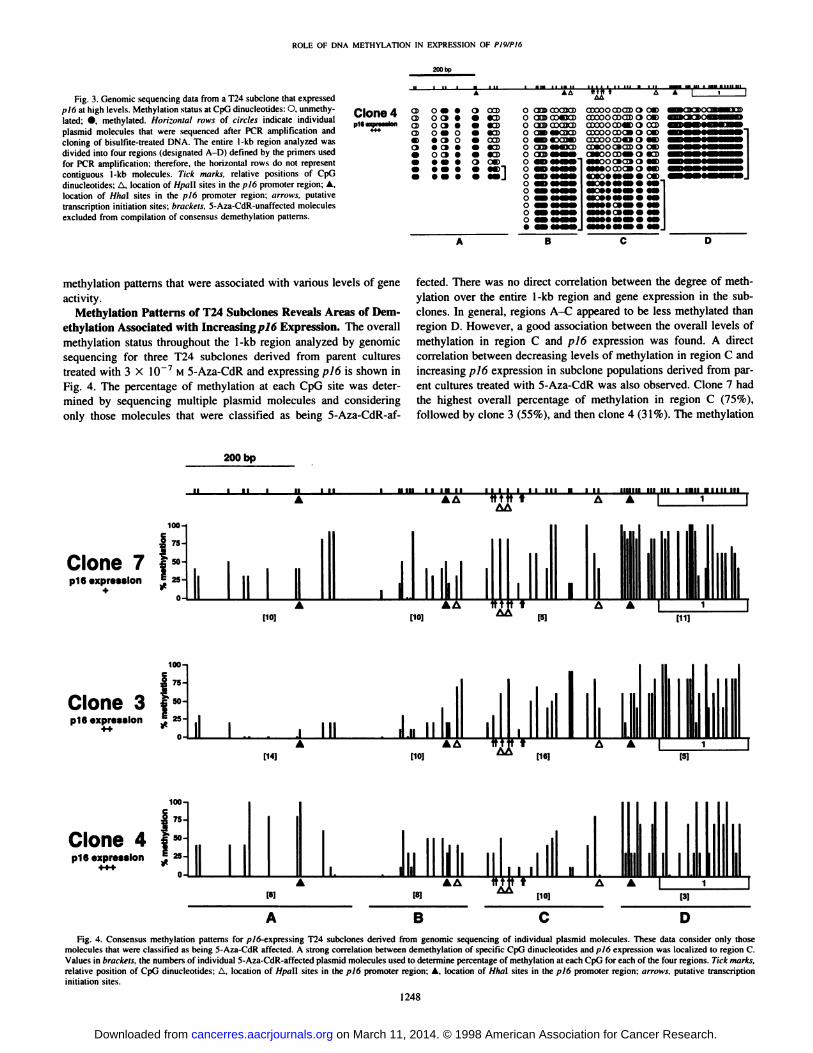
Fig. 3. Cienomic sequencing data from a T24 subclone thai expressedl¡/6at high levels. Methylation status at CpG dinucleoiides: O. unmethy-
lated; •.methylated. Horizontal rows of circles indicate individualplasmid molecules that were sequenced after PCR amplification andcloning of bisulfite-treated DNA. The entire 1-kb region analyzed wasdivided into four regions (designated A-D) defined by the primers used
for PCR amplification: therefore, the horizontal rows do not representcontiguous l-kb molecules. Tick murks, relative positions of CpGdinucleotides: A. location of HfitiU sites in the piò promoter region; A.location of Hha\ sites in the ¡>16promoter region; arrows, putativetranscription initiation sites; brackets, 5-Aza-CdR-unaffected moleculesexcluded from compilation of consensus demethylation patterns.
Clone 4p16expression
d>CBCD0>
o •O C»O C»o ••o•oo a
'IMV '"
CO)•CD«D•CDCCD•CD•CD»
o anp ŒXUCDO nrm cOeEJCDO CESCOCOJCDo a»*»caa>O «MHDOMDo aa>«Moa>O CBSOoo
AA
COOOOCOODO C*>ccooocoaso c*>CDOOOœ«DOCCDCDOOOCDOiO CM)axoocotaoccaooooDO o«CMOOCOiVai «CD
OC»CEtOo coa* o
*:»:;•«••»•;c*>
methylution patterns that were associated with various levels of gene
activity.Methylation Patterns of T24 Subclones Reveals Areas of Dem
ethylation Associated with Increasing pi6 Expression. The overallmethylation status throughout the l-kb region analyzed by genomic
sequencing tor three T24 subclones derived from parent culturestreated with 3 X 10 7 M 5-Aza-CdR and expressing p!6 is shown in
Fig. 4. The percentage of methylation at each CpG site was determined by sequencing multiple plasmid molecules and consideringonly those molecules that were classified as being 5-Aza-CdR-af-
fected. There was no direct correlation between the degree of methylation over the entire l-kb region and gene expression in the sub-clones. In general, regions A-C appeared to be less methylated than
region D. However, a good association between the overall levels ofmethylation in region C and pió expression was found. A directcorrelation between decreasing levels of methylation in region C andincreasing pI6 expression in subclone populations derived from parent cultures treated with 5-Aza-CdR was also observed. Clone 7 had
the highest overall percentage of methylation in region C (75%),followed by clone 3 (55%), and then clone 4 (31%). The methylation
200 bp
Clone 7
i n in i i i il 11 i i i i i i i 111 • in niiiii in in i inn nini inAA ttttt ItA A| 1 |
AA
100-
I 75-
= sopì6 expression * 25-
+ *O-
AA A A[10] [10] [5] [11]
100-I-Clone
3I50p16expression J 25~
++0-ll1 1 III1A[14]J| III!llAA
ItIilI
IIIfiFi
A A | 1|[10]"" [16] [5]
100-
iW-Clone
4t50pi6 expression J25~+++0-[1lini iII
liIIIL .. ¡JillHINIill 11111iAAA Ittttt[8]
[8]""A
B,|_liy[10]CA
A |1I[3]D
Fig. 4. Consensus methylution patterns for /?/6-expressing T24 subclones derived from genomic sequencing of individual plasmid molecules. These data consider only thosemolecules that were classified as being 5-Aza-CdR affected. A strong correlation between demethylation of specific CpG dinucleotides and ¡y}f>expression was localized to region C.Values in brackets, the numbers of individual 5-Aza-CdR-affected plasmid molecules used to determine percentage of methylation at each CpG for each of the four regions. Tick marks,relative position of CpG dinucleotides: A. location of Hpa\\ sites in the pió promoter region; A. location of Hha\ sites in the piò promoter region: arrows, putative transcriptioninitiation sites.
1248
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ROLE OF DNA METHYt.ATION IN F.XPRI-ISSION Öl I'lv/Pln
' '•"'¿A''MW" ' "i T "Cat'. : ]-»-
Fig. 5. Relative CAT expression following invitra methylalion and Iransfection of plasmid DNAcontaining the piò promoter region upstream of theCM7 reporter gene. A methylation cassette assaywas used to make Ihe appropriate constructs asindicated. Tick murks, relative positions of CpGdinucleotides; A. location of Hpa\\ sites in the ¡tlopromoter region; A. location of Hha\ sites in thep/6 promoter region: arrows, putative transcriptioninitiation sites; horizontal shaded bars, regionsmethylated by incubation of plasmid DNA witheither M.Sj.sl. M.HpaU, or M.Hhal. horizontalwhite bars, unmethylated regions of DNA.
Enzyme
none
M.SssI
M.SssI
M.SssI
M.Hpall
M.Hpall
M.Hpall
M.H/731
relative CAT activity
levels of CpGs located at the Hpall and Sma\ sites in the region ofputative transcription start in T24 subclones was also confirmed bymethylation-specific PCR (Ref. 46; data not shown).
The methylation patterns generated by bisulfite genomic sequencing of reactivant clones also suggested that pió expression couldoccur in the presence of a relatively heavily methylated coding domain (region D). In general, region D contained a higher proportion of5-Aza-CdR-unaffected molecules than other regions, but even the5-Aza-CdR-affected molecules used to generate the composite meth
ylation profiles for this region still contained a large number ofmethylated CpGs. No other correlation between the number of 5-Aza-CdR-affected alÃelesand gene expression was found among the reactivant clones, suggesting that the proportion of 5-Aza-CdR-affected
alÃelesin each clone is not responsible for the observed differences inthe levels of pió expression.
We have also generated a total of four additional subclones of clone4 to further investigate the relationship between methylation patternsand p/6 transcriptional activity. Three of the four subclones derivedfrom the clone 4 culture did not express p!6 by RT-PCR analysis,
whereas one subclone was found to express pió at a level that wassimilar to the original clone 4 culture (data not shown). The percentage of methylation of the pl6 5' CpG island was monitored at three
CpG sites in each of these subclones using the quantitative Ms-SNuPE
method as described previously (47,48). Methylation analysis showedthat the single subclone that expressed pió had an average of 52%methylation, whereas the three nonexpressing subclones had an average of between 96 and 99% methylation at the monitored CpG sites.These results showed that the clone 4 culture, which was originallygenerated from a single cell, had diverged into a more heterogeneouspopulation of cells after drug treatment that contained either lowlevels of methylation and expressed pI6 or contained virtually fullymethylated promoter regions and did not express the pió gene.
Methylation of Hpall Sites Is Capable of Down-Regulating pi6Promoter Activity. A plasmid containing the upstream region of ¡>16exon la was used in a CAT reporter assay system to determine theeffect of CpG methylation on pió promoter activity. The p ^PRO-
CAT construct was made by inserting a fragment that containedsequences between the EcoKl site at position -869 and a Hindlll sitethat replaced the normal ATG translational start site by PCR-directedmutagenesis into the pSVO-CAT plasmid vector. This fragment has
been characterized as being sufficient for basal p Id promoter activityin pRB-negative cell lines (44) and contained 49 CpG dinucleotides.
A total of 13 Hpa\\ sites and 11 Hhal sites are located in the vectorsequences flanking the EcoRl-Hiini\l\ pió promoter fragment. Methylation of specific regions of the pl6PRO-CAT construct was accom
plished using a cassette assay as described previously (49).Data obtained from CAT reporter gene expression after in vitro
methylation and transfection of plasmid DNA into the pRB-negativehuman cervical carcinoma-derived cell line C-33A is shown in Fig. 5.The unmethylated pl6PRO-CAT plasmid was defined as having
100% CAT activity. Methylation of the entire construct with M.S.s.vIsignificantly reduced CAT expression by 96%, whereas methylationof all 24 CpG sites located within the EcoR\-Sac\ fragment of the p 16
promoter reduced CAT activity by 74%. Promoter activity was furtherreduced by 97% when all CpG sites located outside of the EcoRl-Sacl
fragment were methylated. For technical reasons, we were unable togenerate a cassette with all CpGs methylated by S.s.vlin region C:however, methylation of the pl6PRO-CAT plasmid with M.Hpall
reduced promoter activity by 75%. Interestingly, methylation of onlythree Hpall sites located within the S«rI-fl.vfEIIfragment (which
corresponds to sequences located within region C) resulted in approximately a 67% decrease in CAT reporter gene expression. On the otherhand, methylation of 1 Hpctll site located outside of region C and 13Hpall sites within the plasmid resulted in a 48% decrease in promoteractivity. Methylation of all Hhal sites in the plasmid, which includedthree CpG sites located within the pl6 promoter, reduced CAT activity by only 4%.
The in vitro methylation experiments indicate that methylation ofonly a few CpG sites in the pió promoter can significantly reducetranscriptional activity. In fact, methylation of a single Hpall sitelocated in the pió promoter reduced CAT activity by 48%. Presumably, this repression is not attributable to methylation of the ///»illsites in the vector backbone because the number and location of Hpallsites in the vector is almost identical to that of the Hhal sites (13Hpall sites compared to 11 ///ml sites), which did not reduce geneactivity. Furthermore, previous studies have demonstrated no effect ofmethylation at Hpall or Hhal sites on C47gene expression in plas-
mids containing the SV40 promoter (50. 51). Methylation of threeHpall sites in the pI6 promoter reduced transcriptional activity by67%, whereas methylation of all Hpall sites, which included four
1249
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Fig. 6. Summary of the relationship betweenmelhylation and transcriptional activilies of pl9 andpl6. The J82 human bladder carcinoma cell linewas methylated in only the Alii elements and exon2. This cell line also showed detectable expressionof both pl9 and p/6. The T24 cell line was shownto be extensively methylated in all regions examined except for exon 1/3. This cell line expressedonly the ¡>I9transcript. Clone 4 was a subclonederived from treatment of a parent T24 culture with5-Aza-CdR. This clone was found to be extensivelymethylated in the Alu elements, exon la codingdomain, and exon 2. No methylation was detectedin exon Iß.and lower levels of methylation weredetected in the ¡tlopromoter region. Clone 4 wascapable of expressing both pl9 and pl6 transcripts.
ROLE OF DNA METHYLATION IN EXPRESSION OK
iLiliL.i, . f .In I. i ||—i nmi insinu
200 bp
J82
T24 999 ff ff ffffff f ff( Alu )[ Alu )
Expression
T?ta?j '/ TT' ( Alut?999999.H
Alu ) |1a|,ff|
2_TJTp19
o16
CI 4 999 ff ff f f f f ff[ Alu )[ Alu] Mal
f Tf
CpGs in the promoter, caused a 75% reduction in CAT reporteractivity. These results suggest that the degree of transcriptional down-regulation may be dependent upon the density of CpG methylation.However, the location of methylated CpG sites may also be animportant factor because methylation of 24 CpG sites in regions A-B
reduced promoter activity by 74%, which was similar to the degree oftranscriptional repression caused by methylation of only three to fourHpall sites in the promoter (67-75%). Although it is difficult todistinguish whether down-regulation of pió promoter activity is dependent upon the location or density of methylated CpG sites, thelarge reduction in promoter activity by methylation of Hpall site(s)located within the p 16 promoter provides the first functional evidencethat site-specific methylation may potentially act as an initiating eventin the progressive inactivation of this tumor suppressor gene.
DISCUSSION
A summary of the relationship between the methylation status ofregions examined in this report and transcriptional activities of p/9and pióis shown in Fig. 6. The J82 cell line was unmethylated at sitesin exon l ß,methylated in each of the Alu elements, unmethylated inthe 5' CpG island oÃp 16 including exon la. and methylated in exon
2. The expression of both pl9 and pl6 mRNA was detected byRT-PCR. The T24 cell line was methylated in all regions examinedexcept for exon Iß.and only the pl9 transcript was detected. Thesituation observed in the T24 cell line demonstrated that extensivemethylation of repetitive sequences and CpG islands downstream ofthe pi9 promoter did not block expression of the pI9 transcript butprevented expression of a pl6 transcript. Finally, the data for clone 4is summarized showing absence of methylation in exon l ß,presenceof methylation in each of the Alu sequences, variable (partial) methylation of the pió promoter, more extensive methylation of the exonla coding region, and methylation of exon 2. Interestingly, clone 4showed expression of both pI9 and pI6 transcripts. Similar methylation patterns were also observed for the other subclones (clones 2 and7), which expressed pl9 at the same level as clone 4, but showed
higher levels of pJ6 promoter methylation and lower levels of pióexpression.
Methylation of repetitive sequences such as Alus and Lis may bethe result of a host defense system that is aimed at abrogating thepotential transcriptional activities of such elements. Alu sequenceshave been found previously to be located in close association withgrowth-regulatory genes such as p53 (52, 53), and the presence ofrepetitive sequences near gene coding regions or promoters may affectthe methylation patterns of these adjacent regions of DNA (18, 52,54). De novo methylation of sequences associated with repetitiveelements may also result in increased mutation rates such as thoseobserved for certain regions of the p53 gene (52).
Prior investigation of apo-E and /g/2r have shown that downstreammethylation of CpG-rich sequences is permissive for gene expression(55, 56). In the case of ¡gßr,the parental alÃelethat expressed thegene was shown to be methylated in downstream sequences. Theeffect of regional DNA methylation has also been studied in thehamster apri gene, demonstrating that methylation at the 3' end did
not interfere with transcription (57). The hamster aprt gene, however,has a high density of CpGs at its 5' end but is relatively CpG depletedat the 3' end. On the other hand, methylation of both the 5' and 3'
ends of the hamster tk gene, which contains a high number of CpGsover its entire length, inhibited transcriptional activity (57). We haveshown that methylation of the p!6 5' CpG islands blocks expression
of the pl6 transcript but not p!9 expression, as demonstrated by ouranalysis of methylation patterns and gene activity in human bladdercancer cell lines. Methylation of Alu repetitive elements and genecoding sequences downstream of the p 19 promoter also did not inhibitpI9 expression. Therefore, methylation of CpG islands, including thepl6 promoter region and pió exon 2 which are located within thetranscriptional unit of the pl9 gene, do not block pl9 expression.
A number of previous studies correlating CpG methylation withtranscriptional silencing of p!6 in cell lines and tumors have examined sites in the exon la coding domain (31-33). These reports haveused Southern analysis or PCR-based methylation assays and werelimited to the possible CpG sites analyzed. Although methylation of
1250
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ROLE OF DNA METHYLATION IN EXPRESSION OK PIV/PI6
the p 16 exon la coding region has been correlated previously with alack of gene expression (11, 31-34, 36), we have shown the presenceof extensive methylation of the exon la coding domain in reactivantclones expressing pió. Our data also suggest that localized areas ofdemethylation in the pió promoter, including CpGs associated withputative transcriptional start sites, may be permissive for pi6 expression and may also serve as a better marker for methylation-associatedtranscriptional silencing of pió.
Although it is not surprising that in vitro methylation of thepl6PRO-CAT construct reduced transcriptional activity, it is remarkable that methylation of only a few Hpa\\ sites was sufficientto confer such a high degree of transcriptional down-regulation.Interestingly, methylation of three Hhai sites in the pl6 promoterdid not have any significant effect on promoter activity. Althoughit is difficult to determine whether density or location of CpGmethylation is responsible for the observed reduction in CATexpression, it appears that each of these components may playsome role in decreasing promoter activity. The presence of methylation at a small number of CpG sites may, therefore, be aninitiating event resulting in progressive inactivation of the piógene. The partial methylation of previously unmethylated CpGislands located in the promoters of tumor suppressor genes mayresult in transcriptional down-regulation and thus lead to an increased rate of cellular growth. The aberrant methylation of CpGislands associated with such growth-regulatory genes may alsoresult in a clonal selection of cells that possess increased growthpotential because of decreased tumor suppressor gene activity. Theuse of 5-Aza-CdR for possible treatment of certain cancers may befeasible because demethylation of only a few CpG sites could becapable of restoring tumor suppressor gene function. Indeed, it hasbeen shown previously that demethylation at one critical CpG siteis sufficient for transcriptional activation of the EBV latency Cpromoter (49).
The genomic sequencing data from single-cell subclones in conjunction with the methylation cassette experiments using the ploPRO-CAT plasmid illustrate several important points that can be madeabout the role of DNA methylation in regulating pin promoter activity: (a) methylation of CpG islands within the/>/9 transcriptional unitdoes not block expression of the pI9 gene. Previous reports that haveinvestigated the effects of downstream methylation on transcriptionalactivity have not considered the effects of methylation of an internalCpG island-containing promoter. Additionally, methylation of otherCpG-rich sequences, including Alii elements and pl6 exon 2, did notblock pl9 expression; (b) methylation of certain regions upstream ofthe p 16 exon la coding domain, particularly region C, may be criticalfor transcriptional activity. The consensus methylation patterns of thereactivant clones that expressed pl6 showed a direct correlationbetween decreased levels of methylation in region C and increasedgene expression and also suggested that pióexpression can occur inthe presence of a relatively heavily methylated coding domain: and (c)methylation of Hpall site(s) within the piópromoter can significantlydown-regulate transcriptional activity. These results suggest that arelatively low number of methylation errors may be sufficient toinitiate a reduction of gene expression and thus provide support for themodel of progressive inactivation of this tumor suppressor gene byDNA methylation.
ACKNOWLEDGMENTS
REFERENCES
10.
29.
30.
We thank Dr. Amy Lee for supplying the pSVO-CAT vector used for the in
vitro methylation and transfection assays.
Li. E.. Besior. T.. and Jaenisch. R. Targeted mutation of the DNA methyltransferasegene results in embryonic lethality. Cell. 6V: 915-926, 1992.Feinberg. A. I". Genomic imprinting and gene activation in cancer. Nat Genet., 4:110-113, 1993.
Rainier. S.. and Feinberg, A. P. Genomic imprinting. DNA methylation. and cancer.J. Nati. Cancer Inst.. 86: 753-759, 1994.Ferguson-Smith. A. C.. Sasaki. H.. Cattanach. B. M.. and Surani. M. A. Parentalorigin-specific epigenetic nunlification of the mouse HIV gene. Nature (I.ond.). .ÃŽ62:751-755. 1993.
Li. E.. Beard. C.. and Jaenisch. R. Role for DNA methylalion in genomic imprinting.Nature (Lond.). 366: 362-365. 1993.Pfeifer. G. P.. Sleigerwald. S. D.. Hansen. R. S.. Gartler. S. M.. and Riggs. A. D.Polymerase chain reaction-aided genomic sequencing of an X chromosome-linked Cp(i
island: methylation patterns suggest clonal inheritance. CpG site autonomy, and anexplanation of activity state stability. Proc. Nail. Acad. Sci. USA. 87: 8252-8256. I99().Hansen. R. S., Ellis. N. A., and Gartler. S. M. Demethylalion of specific sites in (he5' region of the inactive X-linked human phosphoglycerate kinase gene correlates
with the appearance of nuclease sensitivity and gene expression. Mol. Cell. Biol.. fi:4692-4699, 1988.
Ontani Fujita. N., Fujita. T.. Aoike. A.. Osifchin. N. E., Robbins, P. D., and Sakai. T.CpG methylation inactivates the promoter activity of the human relinohlastomatumor-suppressor gene. Oncogene. X: 11)63-1(167. 1993.
Zhang. Y.. Shields, T.. Crenshaw. T., Hao. Y.. Moulton. T., and Tyeko. B. Imprintingof human H19: allele-specific CpG methylation. loss of the active alÃelein Wilmstumor, and potential for somatic alÃeleswitching. Am. J. Hum. (ienet.. 5.?.- I 13-124,
1993.Sakai. T.. Toguchida. J.. Ohlani. N.. Yandell. D. W.. Rapaport. J. M.. and Dryja, T. P.Allele-specific hypermethvlation of the retinoblastoma tumor-suppressor gene. Am. J.Hum. Genet.. 4X: 880-888. 1991.
Merlo. A.. Herman. J. G.. Mao, L., Lee. D. J., Gabrielson. E., Burger, P. C., Baylin.S. B.. and Sidransky. D. 5' CpG island methylation is associated with transcriplional
silencing of the tumour suppressor pl6/CDKN2/MTSl in human cancers. Nat. Med../.- 686-692. 1995.
Jones. P. A. DNA methylation errors and cancer. Cancer Res.. 56: 2463-2467. 1996.Gon/algo. M. L.. and Jones, P. A. Mutagenic and epigenetic effects iti DNAmethylation. Mutât Res.. .186: 107-118. 1997.
Baylin. S. B.. Makos. M.. Wu. J. J.. Yen. R. W., de Buslros. A.. Venino. P.. andNelkin. B. D. Abnormal patterns of DNA methylation in human neoplasia: potentialconsequences for tumor progression. Cancer Cells, 3: 383-390. 1991.
Larsen. F.. Ciundersen. G.. Lope/. R.. and Pryd¿.H. CpG islands as gene markers inthe human genome. Genomics. 13: 1095-1107. 1992.Bird. A. P. CpG-rich islands and the function of DNA methylation. Nature (Lond.).321: 209-213. 1986.
Bestor. T. H., and Tycko. B. Creation of genomic methylation patterns. Nat. (ienet..12: 363-367. 1996.
Yoder. J. A.. Walsh. C. P., and Bestor, T. H. Cytosine methylation and the ecologyof intragenomic parasites. Trends Genet., 13: 335-340. 1997.Bird. A. P. The essentials of DNA methylation. Cell, 70: 5-8. 1992.Gardiner-Garden. M.. and Frommer. M. CpG islands in vertebrate genomes. J. Mol.Biol.. 196: 261-282. 1987.
Smit. A. F. The origin of interspersed repeats in the human genome. Curr. Opin.Genet. Dev.. 6: 743-748. 1996.Schmid. C. W Alu: structure, origin, evolution, significance and function of one-tenthof human DNA. Prog. Nucleic Acid Res. Mol. Biol.. 5.?: 283-319, 1996.
Steenman, M. J.. Rainier. S.. Dobry. C. J.. Grundy. P.. Horon. I. L.. and Fcinberg.A. P. Loss of imprinting of IGF2 is linked to reduced expression and abnormalmethylation of H19 in Wilms' tumour. Nat. Genet.. 7: 433-439, 1994.
Sasaki. M.. Jones. P. A.. Chaillel. J. R.. Ferguson-Smith. A. C.. Barton. S. C.. Reik.W.. and Surani, M. A. Parental imprinting: potentially active chromatin of therepressed maternal alÃeleof the mouse insulin-like growth factor II (Igf2) gene. GenesDev.. 6: 1843-1856. 1992.Serrano. M.. Hannon. G. J.. and Beach. D. A new regulatory motif in cell-cyclecontrol causing specific inhibition of cyclin D/CDK4. Nature (Lond.), 366: 704-707.
1993.Cairns. P., Polascik. T. J.. Eby, Y., Tokino. K., Califano, J.. Merlo, A.. Mao, 1...Herath. J.. Jenkins. R.. Westra, W., Rutter. J. L.. Buckler. A., Gahrielson. E..Tockman, M., Cho. K. R.. Hedrick. L., Bova. G. S.. Isaacs, W.. Koch. W.. Schwab.D.. and Sidransky. D. Frequency of homozygous deletion at p]6/CDKN2 in primaryhuman tumours. Nat. Genet.. //: 210-212. 1995.Kamb. A., Gruis. N. A., Weaver Feldhaus. J., Liu, Q., Harshman. K.. Tavtigian. S. V..Stocken. E.. Day, R. S., Johnson. B. E.. and Skolnick. M. H. A cell cycle regulatorpotentially involved in genesis of many tumor types. Science (Washington DC), 264:436-440. 1994.Nobori. T.. Miura. K.. Wu. D. J.. Lois. A.. Takabayashi. K.. and Carson. D A.Deletions of the cyclin-dependent kinase-4 inhibitor gene in multiple human cancers.Nature (Lond.). 368: 753-756. 1994.Pollock. P. M.. Pearson. J. V.. and Hayward. N. K. Compilation of somatic mutationsof the CDKN2 gene in human cancers: non-random distribution of base substitutions.Genes Chromosomes Cancer. 15: 77-88. 1996.Greenblalt. M. S., Bennett. W. P.. Hollstein. M., and Harris. C. C. Mutations in thep53 tumor suppressor gene: clues to cancer etiology and molecular pathogenesis.Cancer Res., 54: 4855-4878, 1994.
Herman. J. G.. Merlo. A.. Mao. L., Lapidus. R. G.. Issa, J. P.. Davidson. N. E..Sidransky. D.. and Baylin. S. B. Inactivation of the Cl)KN2/iil6/MTSI gene is
1251
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ROLF. OF DNA METHYI.ATION IN EXPRF.SSION OF P/WPI6
frequently associated with aberrant DNA melhylation in all common human cancers.Cancer Res., 55: 4525-4530, 1995.
32. Reed. A. U Califano, J., Cairns. P.. Westra. W. H.. Jones, R. M., Koch, W., Ahrendt.S., Eby, Y., Sewell, D.. Nawroz, H.. Bartek. J., and Sidransky, D. High frequency ofpl6 (CDKN2/MTS-1/1NK4AI inactivation in head and neck squamous cell carcinoma. Cancer Res., 56: 3630-3633. 1996.
33. Gonzalez-Zulueta, M. Bender. C. M., Yang, A. S., Nguyen. T.. Bean. R. W.. VanTomout. J. M.. and Jones. P. A. Methylation of the 5' CpG island of the pl6/CDKN2
tumor suppressor gene in normal and transformed human tissues correlates with genesilencing. Cancer Res., 55: 4531-4535, 1995.
34. Lo, K. W.. Cheung, S. T., Leung, S. F., van Hasselt, A., Tsang. Y. S., Mak, K. F.,Chung. Y. F., Woo, J. K.. Lee, J. C., and Huang. D. P. Hypermethylation of the ;>/6gene in nasopharyngeal carcinoma. Cancer Res.. 50: 2721-2725. 1996.
35. Costello. J. F., Berger. M. S.. Huang. H. S., and Cavenee. W. K. Silencing ofpl6/CDKN2 expression in human gliomas by methylation and chromatin condensation. Cancer Res.. 56: 2405-2410. 1996.
36. Maesawa. C.. Tamura. G.. Nishi/uka, S.. Ogasawara. S.. Ishida. K.. Terashima. M-,Sakata. K., Sato, N., Sailo, K.. and Satodate. R. Inactivation of the CDKN2 gene byhomo/ygous deletion and tic noro methylation is associated with advanced stageesophageal squamous cell carcinoma. Cancer Res.. 56: 3875-3878. 1996.
37. Mao. L., Merlo. A.. Bedi. G.. Shapiro. G. I., Edwards, C. D.. Rollins. B. J., andSidransky. D. A novel pl6/INK4a transcript. Cancer Res.. 55: 2995-2997. 1995.
38. Stone. S.. Jiang, P., Dayananth. P., Tavtigian, S. V.. Katcher. H.. Parry. D.. Peters, G.,and Kamb. A. Complex structure and regulation of the pió (A/757) locus. CancerRes.. 55: 2988-2994. 1995.
39. Quelle, D. E., Zindy. F.. Ashmun. R. A., and Sherr. C. J. Alternative reading framesof the INK4u tumor suppressor gene encode two unrelated proteins capable ofinducing cell cycle arrest. Cell. Hj: 993-1000. 1995.
40. Clark. S. J.. Harrison. J.. Paul. C. L.. and Frommer, M. High sensitivity mapping ofmethylated cytosines. Nucleic Acids Res.. 22: 2990-2997. 1994.
41. Frommer. M.. McDonald. L. E., Millar. D. S.. Collis. C. M., Watt. F.. Grigg, G. W..Molloy, P. L., and Paul. C. L. A genomic sequencing protocol that yields a positivedisplay of 5-methylcytosine residues in individual DNA strands. Proc. Nail. Acad.Sci. USA, 89: 1827-1831. 1992.
42. Jones. P. A. Altering gene expression with 5-azacytidine. Cell. 40: 485-486. 1985.43. Bender. C. M.. Pao. M. M., and Jones. P. A. Inhibition of DNA melhylation by
5-aza-2'-deoxycytidine suppresses the growth of human tumor cell lines. Cancer Res..
58: 95-101. 1998.
44. Hará.E.. Smith, R.. Parry, D.. Tahara, H., Stone. S.. and Peters, G. Regulation ofpl6CDKN2 expression and its implications for cell immortalization and senescence.Mol. Cell. Biol.. 16: 859-867. 1996.
45. Jones, P. A. Gene activation by 5-azacytidine. In: A. Razin, H. Cedar, and A. D.Riggs (eds.). DNA Methylation: Biochemistry and Biological Significance, pp. 165-187. New York: Springer-Verlag, 1984.
46. Herman. J. G.. Graff. J. R.. Myohanen. S.. Nelkin. B. D., and Baylin. S. B. Melhy-lation-specillc PCR: a novel PCR assay for methylation status of CpG islands. Proc.Nati. Acad. Sci. USA. 93: 9821-9826. 1996.
47. Gonzalgo. M. L.. and Jones. P. A. Rapid quantitation of methylation differences atspecific sites using methylation-sensitive single nucleotide primer extension (Ms-SNuPE). Nucleic Acids Res.. 25: 2529-2531, 1997.
48. Gonzalgo. M. L., Bender. C. M.. You. E. H.. Glendening, J. M.. Flores. J. F.. Walker.G. J., Hayward, N. K.. Jones, P. A., and Fountain. J. W. Low frequency of pl6/CDKN2A methylation in sporadic melanoma: comparative approaches for methylation analysis of primary tumors. Cancer Res., 57: 5336-5347. 1997.
49. Robertson. K. D.. Hayward. S. D.. Ling, P. D.. Samid. D., and Ambinder. R. F.Transcriptional activation of the Epstein-Barr virus latency C promoter after 5-aza
cytidine treatment: evidence that demethylation at a single CpG site is crucial. Mol.Cell. Biol., 15: 6150-6159, 1995.
50. Bryans. M.. Kass. S., Seivwright. C., and Adams. R. L. Vector methylation inhibitstranscription from the SV40 early promoter. FEBS Lett., .109: 97-102, 1992.
51. Graessmann. M.. Graessmann. A.. Wagner. H.. Werner. E., and Simon, D. CompleteDNA methylation does not prevent polyoma and simian virus 40 virus early geneexpression. Proc. Nati. Acad. Sci. USA. 80: 6470-6474. 1983.
52. Magewu, A. N., and Jones. P. A. Ubiquitous and tenacious methylation of the CpGsite in codon 248 of the ¡t53gene may explain its frequent appearance as a mutationalhotspot in human cancer. Mol. Cell. Biol.. 14: 4225-4232, 1994.
53. Yang, A. S.. Gonzalgo. M. L., Zingg, J. M.. Millar. R. P.. Buckley. J. D., and Jones,P. A. The rate of CpG mutation in Alu repetitive elements within the /)5.i tumorsuppressor gene in the primate germline. J. Mol. Biol., 258: 240-250. 1996.
54. Hasse. A., and Schulz. W. A. Enhancement of reporter gene tit' nitro methylation byDNA fragments from the a-fetoprotein control region. J. Biol. Chem.. 269: 1821-
1826, 1994.55. Larsen. F.. Solheim, J.. and Prydz. H. A methylated CpG island 3' in the
iipoliliopt'oti'in-E gene does not repress its transcription. Hum. Mol. Genet.. 2:
775-780. 1993.
56. Stoger. R.. Kubicka. P.. Liu. C. G.. Kafri. T.. Razin, A.. Cedar. H.. and Barlow. D. P.Maternal-specific methylation of the imprinted mouse Ixf2r locus identifies theexpressed locus as carrying the imprinting signal. Cell. 73: 61-71. 1993.
57. Keshet, I.. Yisraeli, J.. and Cedar, H. Effect of regional DNA methylation on geneexpression. Proc. Nati. Acad. Sci. USA. 82: 2560-2564. 1985.
1252
on March 11, 2014. © 1998 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
Related Documents











