The role of 5-HT 1A and 5-HT 1B receptors in MDMA self-administration By Dane Aronsen A thesis submitted in fulfilment of the requirements for the degree of Doctor of Philosophy Victoria University of Wellington 2016

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The role of 5-HT1A and 5-HT1B receptors in MDMA self-administration
By
Dane Aronsen
A thesis submitted in fulfilment of the requirements for the degree of Doctor of Philosophy
Victoria University of Wellington
2016

2

3
Acknowledgements
To everyone that made this 3 or so years such a positive experience, thank you. I
couldn’t have done it without the family and friends that supported me through thick
and thin.
I want to express sincere gratitude to the team of people I worked with in the Lab, with
a special shout out to Joyce for patiently teaching me whatever I needed to know, and
my partner in all lab shenanigans, Jeremy.
A big thank you to the academic staff that presented me with challenges and
opportunities, and who would happily write me a scholarship reference at the last
minute.
And Sue. You’ve taught me a lot, and some of it was about science. You’ve made sure
I will never forget my experience as a PhD student.
Thanks guys.

4
Contents
Acknowledgements ......................................................................................................... 3
List of abbreviations ........................................................................................................ 6
List of ligands .................................................................................................................. 7
Abstract ........................................................................................................................... 8
General Introduction ..................................................................................................... 10
Brief history of MDMA ............................................................................................ 10
MDMA use ............................................................................................................... 11
Harms associated with MDMA use .......................................................................... 13
Problems associated with studying the harmful effects of MDMA .......................... 13
Animal studies ........................................................................................................... 15
Profile of MDMA self-administration ...................................................................... 18
Pharmacodynamics of MDMA ................................................................................. 23
A focus on 5-HT ....................................................................................................... 24
5-HT1A and 5-HT1B receptors .................................................................................... 26
Summary ................................................................................................................... 35
General Methods ........................................................................................................... 37
Subjects ..................................................................................................................... 37
Drugs ......................................................................................................................... 37
Apparatus and procedures ......................................................................................... 37
Chapter 3: Development of behavioural assays ............................................................ 39
Method ...................................................................................................................... 41
Results ....................................................................................................................... 42
Discussion ................................................................................................................. 43
Chapter 4: Behavioural responses to RU 24969 ........................................................... 45
Method ...................................................................................................................... 46
Results ....................................................................................................................... 48

5
Discussion .................................................................................................................. 51
Chapter 5: Effects of repeated administration of the 5-HT1B/1A receptor agonist, RU
24969, on the acquisition of MDMA self-administration ............................................. 54
Method ....................................................................................................................... 56
Results ....................................................................................................................... 58
Discussion .................................................................................................................. 61
Chapter 6: Predicting the acquisition of MDMA self-administration ........................... 66
Method ....................................................................................................................... 67
Results ....................................................................................................................... 69
Discussion .................................................................................................................. 72
Chapter 7: Response to 5-HT1A and 5-HT1B receptor agonists after
self-administration ........................................................................................................ 75
Method ....................................................................................................................... 78
Results ....................................................................................................................... 79
Discussion .................................................................................................................. 82
General Discussion ........................................................................................................ 84
Summary .................................................................................................................... 84
Synthesis of results .................................................................................................... 85
Limitations ................................................................................................................. 86
Validity of MDMA doses .......................................................................................... 87
Key findings and future directions ............................................................................ 89
Conclusion ................................................................................................................. 93
References ..................................................................................................................... 95
Appendix A: Publication details and permissions ....................................................... 126

6
List of abbreviations
5,7-DHT: 5,7-Dihydroxytryptamine
5CSRTT: 5 choice serial reaction time task
5-HIAA: 5-Hydroxyindoleacetic acid
5-HT: 5-hydroxytryptamine; serotonin
6-OH-DA 6-hydroxydopamine
CSF: Cerebrospinal fluid
DSM: Diagnostic and Statistical Manual of Mental Disorders
EC50: Half maximal effective concentration
EPM: Elevated plus maze
FR: Fixed ratio
GABA: gamma-Aminobutyric acid
GTPγS: guanosine 5'-O-[gamma-thio]triphosphate
Ki: Binding affinity (amount of ligand required to bind 50% of receptors)
MDMA : 3,4-methylenedioxymethamphetamine
NAc: Nucleus accumbens
PFC: Prefrontal cortex
SSRI: Selective serotonin reuptake inhibitor
SUD: Substance use disorder
VTA: Ventral tegmental area

7
List of ligands
5-MeODMT 5-HT2/1A receptor agonist
8-OH-DPAT 5-HT1A/7 receptor agonist
BAY × 3702 5-HT1A receptor agonist
CGS 12066 5-HT1B/2 receptor agonist
CP 93129 5-HT1B receptor agonist
CP 94253 5-HT1B receptor agonist
DOI 5-HT2A/2C receptor agonist
F13640 5-HT1A receptor agonist
F15599 5-HT1A receptor agonist
Fluoxetine Serotonin uptake inhibitor
GBR 12909 Dopamine uptake inhibitor
GR 127935 5-HT1B/1D receptor antagonist
Ketanserin 5-HT2A/2C receptor and H1 receptor antagonist
M100907 5-HT2A receptor antagonist
mCPP 5-HT2/1A receptor agonist
MK 212 5-HT2C receptor agonist
Pindolol β1- and β2-adrenergic receptor antagonist, 5-HT1A antagonist
Propanolol β1- and β2-adrenergic receptor antagonist, 5-HT1A/1B antagonist
Ro 60-175 5-HT2C receptor agonist
RU 24969 5-HT1B/1A receptor agonist
SB 206553 5-HT2C/2B receptor antagonist
SB 224289 5-HT1B receptor inverse agonist
SB 242084 5-HT2C receptor antagonist
SDX 216-525 5-HT1A receptor antagonist
Tianeptine 5-HT uptake facilitator
WAY 101405 5-HT1A receptor antagonist
WAY 100635 5-HT1A receptor antagonist

8
Abstract
Rationale: 3,4-methylenedioxymethamphetamine (MDMA) is a less efficacious
reinforcer than other drugs of abuse. However, following repeated self-administration,
responding increases for some animals and efficacy becomes comparable to other
drugs of abuse. MDMA-stimulated serotonin (5-HT) release was negatively associated
with acquisition of MDMA self-administration, and a neurotoxic 5-HT lesion reduced
the latency to acquire self-administration. These findings suggest that MDMA-
produced 5-HT release is an important component of self-administration. The receptor
mechanisms are not, however, well understood, although it has often been suggested
that the mechanism involves 5-HT-mediated inhibition of dopamine. Both 5-HT1A and
5-HT1B receptors are well localised to regulate dopamine release, and both have been
implicated in modulating the reinforcing effects of many drugs of abuse.
Objectives: The first objective was to establish specific behavioural assays to reflect
5-HT1A and 5-HT1B receptor activation. Then, using the established behavioural
assays, the aim was to determine the role of 5-HT1A and 5-HT1B receptors in the
acquisition of MDMA self-administration. The impact of substantial MDMA self-
administration on 5-HT1A and 5-HT1B receptors was also assessed.
Methods: Firstly, dose-effect relationships for the hyperactive response to the 5-HT1A
receptor agonist, 8-OH-DPAT (0 – 3.0 mg/kg) and the hyperactive and adipsic
response to the 5-HT1B/1A receptor agonist, RU 24969 (0 – 3.0 mg/kg) were
determined. Selectivity of these responses was determined by co-administration of the
5-HT1A receptor antagonist, WAY 100635, or the 5-HT1B/1D receptor antagonist, GR
127935. Secondly, a pretreatment regimen of the RU 24969 (2 × 3.0 mg/kg/day, 3
days), which had been suggested to down-regulate 5-HT1B/1A receptors, was
administered prior to self-administration testing. The effect of this manipulation on
both the acquisition of MDMA self-administration, and the behavioural responses to 5-
HT1A and 5-HT1B receptor activation, was measured. A further study measured
behavioural responses to 5-HT1A or 5-HT1B receptor agonists prior to self-
administration, to determine whether the variability in these responses would predict
the variability in the latency to acquisition of MDMA self-administration. Lastly, the
effect of substantial MDMA self-administration (350 mg/kg) on dose-response curves
for the behavioural effects of 5-HT1A or 5-HT1B receptor activation was assessed.
Results: The hyperactive response to the 5-HT1B/1A receptor agonist, RU 24969, was
blocked by the 5-HT1A receptor antagonist, WAY 100635, but not the 5-HT1B receptor

9
antagonist, GR127935. Similarly, the hyperactive response to the 5-HT1A receptor
agonist, 8-OH-DPAT, was dose-dependently blocked by WAY 100635. GR 127935,
but not WAY 100635, blocked the adipsic response to RU 24969.
Repeated administration of RU 24969 produced rightward shifts in the dose-response
curves for 8-OH-DPAT-produced hyperactivity and RU 24969-produced adipsia, and
also greatly facilitated the acquisition of MDMA self-administration. However, there
was no correlation between latency to acquire MDMA self-administration and the
hyperactive response to 8-OH-DPAT or the adipsic response to RU 24969, and
MDMA self-administration failed to alter these behavioural response to activation of
5-HT1A or 5-HT1B receptors.
Conclusions: The hyperactive response to 8-OH-DPAT and the adipsic response to
RU 24969 reflect activation of 5-HT1A and 5-HT1B receptors, respectively. The
variability in acquisition of MDMA self-administration was reduced by a treatment
that also down-regulated 5-HT1A and 5-HT1B receptors, however there was no further
indication that these receptors play a critical role in the self-administration of MDMA.
Instead, it seems likely that other 5-HT receptors have a greater impact on MDMA
self-administration.

10
General Introduction
Parts of this chapter have been adapted from:
Aronsen & Schenk (2016). MDMA self-administration fails to alter the behavioral
response to 5-HT1A and 5-HT1B agonists. Psychopharmacology, 233 (7), 1323-
1330. DOI 10.1007/s00213-016-4226-9
Aronsen, Bukholt, & Schenk (2016). Repeated administration of the 5-HT1B/1A agonist,
RU 24969, facilitates the acquisition of MDMA self-administration: Role of 5-
HT1A and 5-HT1B receptor mechanisms. Psychopharmacology, 233 (8), 1339-
1347. DOI 10.1007/s00213-016-4225-x
Schenk & Aronsen (2015). Contribution of Impulsivity and Serotonin Receptor
Neuroadaptations to the Development of an MDMA (“ecstasy”) Substance Use
Disorder, in Current Topics in Behavioral Neuroscience: The Misuse of Licit
and Illicit Drugs in Psychopharmacology Susan Schenk, Suzanne Nielson,
Raimondo Bruno (Eds), Springer. DOI 10.1007/7854_2015_421
Aronsen, Webster, & Schenk (2014). RU 24969-produced adipsia and
hyperlocomotion: Differential role of 5HT1A and 5HT1B receptor mechanisms.
Pharmacology, Biochemistry and Behavior, 124, 1-4. DOI
10.1016/j.pbb.2014.05.008
with permission from the publisher (Appendix A).
Brief history of MDMA
3,4-methylenedioxymethamphetamine (MDMA) was initially patented by the
pharmaceutical company, Merck, in 1914 as a precursor for other therapeutically
efficacious compounds (Green, Mechan, Elliott, O'Shea, & Colado, 2003). Some basic
preclinical tests were conducted with MDMA in 1927 (Freudenmann, Öxler, &
Bernschneider-‐Reif, 2006), and in 1953 the US Army sponsored research on the
toxicity of MDMA, concluding that further study in humans should be conducted
(Hardman, Haavik, & Seevers, 1973). In response to the first reports of recreational
MDMA use (Gaston & Rasmussen, 1972) Alexander Shulgin published the first papers
outlining the effects of MDMA in humans (Anderson, Braun, Braun, Nichols, &
Shulgin, 1978; Shulgin, 1978; Shulgin & Nichols, 1978), also encouraging further
human studies. Shulgin was a vocal advocate for the use of MDMA as an adjunct to
psychotherapy, but it has been suggested that his public promotion of MDMA also led
to increased recreational use (Benzenhöfer & Passie, 2010).

11
As recreational use continued to grow, there was pressure on governments to
bring the use of MDMA under legislative control (Beck & Rosenbaum, 1990). MDMA
was scheduled as a Class B Controlled Drug in New Zealand in 1987 (New Zealand
Drug Foundation, 2015), following classification in Schedule I by the USA Drug
Enforcement Agency in 1985 (Beck & Rosenbaum, 1990). The import, manufacture,
supply, or administration of Class B Controlled Drugs carries a jail sentence of up to
14 years in New Zealand (Misuse of Drugs Act 1975). Nonetheless, recreational use of
MDMA, in the form of the street drug, ‘ecstasy’, is popular in New Zealand (Wilkins,
2011; Wilkins & Sweetsur, 2008), and around the world (United Nations Office on
Drugs and Crime, 2015).
Recently, there has been a revival in the push to harness the subjective effects
of MDMA in the treatment of psychiatric disorders. Some therapists claim that
MDMA helps patients talk openly, and fosters an atmosphere of trust (Kupferschmidt,
2014). Clinical trials are currently underway assessing the utility of MDMA as a
therapeutic adjunct in the treatment of post-traumatic stress disorder, and anxiety
associated with a life-threatening illness (National Institutes of Health, 2015).
MDMA use
MDMA is generally consumed as the primary psychoactive component of the
popular street drug, ecstasy (also known as E, Molly, pingers, pills, disco biscuits).
Ecstasy is most commonly available in tablet form, and tablets are usually either
consumed orally or crushed for intranasal administration (De La Garza, Fabrizio, &
Gupta, 2007; Parrott, 2013a; Solowij, Hall, & Lee, 1992). In recent years
recreationally used ecstasy tablets have been shown to contain a wide range of
psychoactive substances, including significant quantities of methamphetamine,
ketamine, caffeine, meta-Chlorophenylpiperazine (mCPP) and mephedrone, and have
sometimes contained no MDMA whatsoever (Brunt, Koeter, Niesink, & van den
Brink, 2012; Morefield, Keane, Felgate, White, & Irvine, 2011; Togni, Lanaro,
Resende, & Costa, 2015; Vogels et al., 2009). Therefore, throughout this thesis, the
term ‘ecstasy’ will be used to refer to the street drug that generally contains MDMA,
while ‘MDMA’ will be used to refer specifically to the psychoactive substance.
Ecstasy became popular in the underground dance party scene of the 1980s, in
part because it increases energy levels, heightens sensual awareness, and facilitates
bonding (McDowell & Kleber, 1994; Schwartz & Miller, 1997). In the 1990s and early

12
2000s ecstasy use became more mainstream, becoming a popular recreational drug
among young adults. A recent study reported worldwide prevalence of ecstasy use to
be the second highest of all illicit drugs (Global Drug Survey, 2014). Recently,
popularity of ecstasy has been facilitated by a ‘re-branding’ of ecstasy as ‘Molly’ in
the mainstream media. ‘Ecstasy’ has associations with the old dance parties of the
1980s, electronic music, and un-masculine displays of affection, misaligning it with
the modern zeitgeist which is heavily influenced by pop and hip-hop culture. On the
other hand, ‘Molly’ has been embraced by the hip-hop and pop communities,
providing a ‘new’ drug that youth can associate with (Carter, 2016).
Although ecstasy use is common, patterns of use differ widely. A recent study
showed that, of 109 subjects who had recently used ecstasy for the first time, 43 did
not take ecstasy again in the following 12 months, while 23 consumed more than 10
ecstasy pills in that time period (Wagner, Becker, Koester, Gouzoulis-‐Mayfrank, &
Daumann, 2013), illustrating that some will use ecstasy very infrequently, while others
will use ecstasy regularly. Furthermore, recent surveys have found a significant
proportion of regular ecstasy users met Diagnostic and Statistical Manual of Mental
Disorders (DSM) -IV-based criteria for dependence (Cottler, Leung, & Abdallah,
2009; Cottler, Womack, Compton, & Ben-Abdallah, 2001; Uosukainen, Tacke, &
Winstock, 2015). The more recent DSM 5 provides diagnostic criteria for ‘substance
use disorders’ (SUDs) rather than ‘dependence’ (American Psychiatric Association,
2013). Although there is no specific ecstasy SUD, some ecstasy users met a number of
SUD criteria, including using more drug than intended (Cottler et al., 2009; Cottler et
al., 2001) unsuccessful efforts to cut down on use (Jansen, 1999), craving (A. K. Davis
& Rosenberg, 2014; Hopper et al., 2006), neglecting activities other than acquiring and
taking drug (Cottler et al., 2009; Cottler et al., 2001; Jansen, 1999; Yen & Hsu, 2007),
use in spite of known negative consequences (Cottler et al., 2009; Cottler et al., 2001;
Jansen, 1999; Schifano & Magni, 1994; Yen & Hsu, 2007), tolerance (Cottler et al.,
2001; Jansen, 1999; Kirkpatrick et al., 2014; Parrott, 2005; Peroutka, Newman, &
Harris, 1988; Yen & Hsu, 2007), and withdrawal (Cottler et al., 2009; Cottler et al.,
2001; Jansen, 1999; Peroutka et al., 1988). Thus, while some ecstasy users take ecstasy
relatively infrequently, a subpopulation of users show regular use, and some show
signs of an SUD.

13
Harms associated with MDMA use
The regular use of ecstasy in some users is of concern, not only because of the
potential to develop an SUD, but also because ecstasy use has been associated with a
number of cognitive, behavioural, and neurochemical deficits. Ecstasy users showed
deficits in learning (Wagner et al., 2013), and in attention and memory (McCann,
Mertl, Eligulashvili, & Ricaurte, 1999) compared to ecstasy-naïve controls or those
with limited ecstasy use. Ecstasy users reported higher levels of depression,
impulsiveness, and sleep disturbances than poly-drug users who did not use ecstasy
(Soar, Turner, & Parrott, 2006; Taurah, Chandler, & Sanders, 2014). These cognitive
and behavioural deficits were persistent, suggesting that regular ecstasy use may cause
long-lasting neuroadaptations (Parrott, 2013a, 2013b; Schifano & Magni, 1994). With
increased experience some heavy ecstasy users report persistent problematic
behaviour, including paranoid delusions (Schifano & Magni, 1994), severe weight loss
(Jansen, 1999; Schifano & Magni, 1994), and suicidal thought (Jansen, 1999; Schifano
& Magni, 1994).
Ecstasy use has also been associated with deficits in the neurotransmitter,
serotonin (5-HT). Ecstasy users had decreased 5-HT transporter binding (Kish et al.,
2010; McCann, Szabo, Scheffel, Dannals, & Ricaurte, 1998), reduced levels of the 5-
HT metabolite, 5-hydroxyindoleacetic acid (5-HIAA), in cerebrospinal fluid (McCann,
Mertl, et al., 1999), reduced 5-HT synthesis in frontal and parietal regions (Booij et al.,
2014), and autopsied striatal tissue from a heavy ecstasy user indicated decreased 5-
HT and 5-HIAA levels (Kish, Furukawa, Ang, Vorce, & Kalasinsky, 2000). These
markers of reduced 5-HT function correlate with lifetime ecstasy use (Kish et al.,
2010; McCann et al., 1998) and levels of behavioural impairment (Kish et al., 2010).
Therefore, it is possible that at least some of these adaptations underlie the long term
behavioural problems seen after repeated ecstasy use. The mechanisms by which
MDMA might produce these effects is not clear.
Problems associated with studying the harmful effects of MDMA
Given the global popularity of ecstasy, and the deficits associated with regular
use, it becomes important to investigate potential treatments to reduce intake, and to
reverse harmful neuroadaptations. However, there are a number of potential confounds
associated with studies that use human subjects to determine the effects of MDMA use
on the brain and/or behaviour. Firstly, results from studies on ecstasy users can be

14
limited by a number of factors. For example, the accuracy of subjects’ reported use and
the range of other drugs the subject also uses may confound results. This concept is
illustrated in the abovementioned report by Jansen (1999) describing the effects of
ecstasy use in three regular users, in which total lifetime ecstasy exposure was
determined by self-report for periods of over two years. This method for determining
drug intake relies on memory for drug taking episodes even though ecstasy use is
associated with memory impairments. Furthermore, the study by Jansen highlights the
poly-drug use typical of regular ecstasy users (Cottler et al., 2009; Cottler et al., 2001)
– the first patient reported regular amphetamine use of 1g/day, the second was
dependent on benzodiazepines, while the third consumed roughly 1 bottle of spirits
every night. Regular use of other drugs makes it more difficult to isolate the effects of
MDMA.
A second potential issue with human studies is that varied individual histories
of ecstasy users can limit the conclusions that can be drawn. For example, while
symptoms of depression and anxiety are widely reported after regular ecstasy use
(Rogers et al., 2009), a causal link cannot be drawn between ecstasy use and
psychological deficits, given that pre-existing problems such as anxiety and depression
might predispose an individual to regular ecstasy consumption as a form of self-
medication (Parrott, 2006, 2013a). Without random allocation and an appropriate
control group, causal links between drug use and its effects cannot be drawn. In an
effort to overcome this limitation a small number of studies have randomly assigned
participants to receive acute administrations of MDMA, but, as the authors of one
study lament, ethical constraints on dosing regimens severely limit the ecological
validity and scope of such studies (Peiró et al., 2013).
Some researchers have gone to great lengths to minimise the impact of such
confounds on the results of their studies. For example, a recent study investigated
current ecstasy users and compared results to a control group of poly-drug users that
have never used ecstasy. Thus, any differences should be attributable to ecstasy use.
The results showed that ecstasy users had higher levels of cognitive and behavioural
disturbances than non-ecstasy poly-drug users (Taurah et al., 2014). Such results
strengthen claims that MDMA use is harmful and help to illustrate the nature of these
harms. However, because of ethical constraints that restrict the doses of MDMA that
can be administered to humans, investigations into the mechanisms behind these
effects of MDMA cannot be readily conducted.

15
Animal studies
For these reasons, animal models are often turned to in order to obtain
information regarding the effects of exposure to MDMA. The real value of animal
laboratory studies is that they allow experimenters some control over the histories of
subjects, the drugs administered, and environmental factors. Furthermore, a wider
range of doses can be administered to animals than is ethically viable with humans.
There is some loss of ecological validity when animal models are employed,
particularly as they necessarily ignore the complex environment in which ecstasy is
consumed, but such studies can be incredibly helpful in evaluating properties of
MDMA that cannot be determined in humans.
A number of studies have replicated the findings of human studies after
administering MDMA to animals. Typically, high doses of MDMA are administered
repeatedly, after which some behavioural or neurochemical measures are made. For
example, exposure to high doses of experimenter-administered MDMA decreased
tissue 5-HT levels (Battaglia, Yeh, & De Souza, 1988; Commins et al., 1987;
McGregor et al., 2003), damaged 5-HT cells (Commins et al., 1987; Jensen et al.,
1993), and reduced 5-HT transporter binding (Battaglia, Yeh, et al., 1988; McGregor
et al., 2003). In behavioural tests, repeated administration of MDMA increased
anxiety-like behaviour in adult (McGregor et al., 2003) and adolescent rats (Bull,
Hutson, & Fone, 2003; Bull, Hutson, & Fone, 2004; Cox et al., 2014), and impaired
novel object discrimination, a measure of recognition memory (Shortall et al., 2013).
Although this method of experimenter-administered, high dose MDMA is useful for
determining the harmful effects of MDMA, these studies have been criticised for
employing a physiologically irrelevant dosing regimen (Baumann & Rothman, 2009;
Cole & Sumnall, 2003; De La Garza et al., 2007; Meyer, Piper, & Vancollie, 2008),
given that this high level of exposure is rarely, if ever, experienced by ecstasy users
(D. Hansen, Maycock, & Lower, 2001; Parrott, 2005; Verheyden, Henry, & Curran,
2003).
One alternative to an experimenter administered drug regimen is to give the
animal control over the delivery of drug, in a manner similar to how humans control
their drug intake. This is the basis of the popular self-administration paradigm, in
which an animal performs some operant (e.g. nose poke, lever press) in order to obtain
a dose of drug. Often, the route of drug administration is intravenous, meaning the
animal requires a surgically implanted indwelling venous catheter. After recovery from

16
this surgery, the animal is placed in an operant chamber and the catheter is connected
via tubing to a syringe encased in a mechanical syringe pump. The operant activates
the syringe pump, resulting in a predetermined intravenous dose of the drug being
investigated. Drug infusions are generally paired with a stimulus (e.g. light, tone).
Usually there is a second manipulandum (e.g. nose poke hole, lever) for which the
operant has no programmed consequence, but responses are recorded as a measure of
non-specific responding.
Human drug taking is a complex behaviour that is influenced by an interaction
of social, economic, and personal factors, and as such it cannot be modelled in a single
animal paradigm. Furthermore, as with all animal models, ecological validity is lost in
order to gain experimental control and practicality. For example, self-administration
studies generally allow an animal to self-administer only the drug of interest (with no
adulterants), in order to draw causal conclusions about this drug. In contrast, human
drug users tend to use a range of drugs, and drugs procured on the street tend not to be
pure. Thus, the self-administration paradigm trades ecological validity for
experimental control (De La Garza et al., 2007). While experimental design can help to
minimise the loss of validity, no self-administration model can perfectly replicate
human drug taking. Nonetheless, as will be explained below, the self-administration
paradigm is an excellent paradigm for MDMA administration, and also allows for
studies in which drug taking is the dependent measure.
A particular strength of the self-administration paradigm is that the animal has
control over their drug intake. Firstly, this reduces concerns over the administration of
irrelevantly large drug doses. Figure 1.1 presents data adapted from Schenk, Gittings,
Johnstone, and Daniela (2003) showing the number of infusions of MDMA that were
self-administered in a session, for different doses of MDMA. It is clear that MDMA
self-administration behaviour adjusts as dose changes, illustrating that the animal
utilises control over responding to regulate total drug intake. Thus, it is less likely that
physiologically irrelevant doses will be administered, as has been suggested for studies
using experimenter-administered MDMA.

17
M D M A (m g /k g /in fu s io n )
Infu
sio
ns
pe
r 2
hr
se
ss
ion
0
2 0
4 0
6 0
8 0
1 0 0
0 .25 0 .5 1 .0 2 .0
Fig. 1.1 Dose response curve for MDMA self-‐administration. Adapted from Schenk et al. (2003).
Secondly, self-administered drug produces neuroadaptations that are not solely
due to the action of the drug. For example, self-administered cocaine produced
significantly greater changes in dopamine transporter binding than the same doses
administered non-contingently, suggesting that the stimulus-response associations
learned in self-administration contribute to the neuroadaptations produced by drugs of
abuse (Miguéns et al., 2008). Because human users also have control over their drug
intake, and because the neuroadaptations produced by drugs may be dependent on this
control, self-administered MDMA is probably a better model of human drug
administration than experimenter-administered MDMA.
A third strength of the self-administration paradigm, and of particular relevance
to MDMA, is that the overall pattern of drug taking is similar in animals and humans.
On their first exposure to MDMA human users generally consume ½ - 1 ecstasy tablet
(D. Hansen et al., 2001) with drug use being intermittent, but with experience some
users may consume upwards of 20 pills in a session (Parrott, 2005; Verheyden et al.,
2003). A similar pattern of low, intermittent initial intake followed by increased intake
in some subjects is seen in MDMA self-administration in rats and monkeys (Banks et
al., 2008; Beardsley, Balster, & Harris, 1986; De La Garza et al., 2007; Schenk,
Colussi-Mas, Do, & Bird, 2012). It is important that, in both animals and humans,
initial exposure to MDMA is low and intermittent, because intermittent or low dose

18
exposure to MDMA was neuroprotective against the neuroadaptations produced by
subsequent high dose administrations (Bhide, Lipton, Cunningham, Yamamoto, &
Gudelsky, 2009; Piper, Ali, Daniels, & Meyer, 2010). Indeed, self-administered
MDMA produced smaller deficits in tissue levels of 5-HT compared to high dose
experimenter-administered MDMA (Do & Schenk, 2011; Scanzello, Hatzidimitriou,
Martello, Katz, & Ricaurte, 1993; Schenk et al., 2007), even though the total amount
self-administered (165-350 mg/kg over 20-30 days of testing) was greater than is
generally administered to produce extensive neurotoxicity (20-80 mg/kg in a single
day). Given that the neuroadaptations produced by MDMA are dependent on the
pattern of prior MDMA exposure, self-administered MDMA likely produces
neuroadaptations more similar to the human condition than those produced by
experimenter-administration.
Furthermore, the self-administration paradigm allows for the behaviour of drug
taking to be studied, which can be useful when investigating how a certain
manipulation might affect drug taking behaviour. In this manner, self-administration
has been a valuable pre-clinical tool in determining the efficacy of purported
treatments for reducing drug consumption. For example, self-administration of a range
of drugs is reduced by vaccines that use the body’s immune system to block drugs
from crossing the blood/brain barrier (Fox et al., 1996; Kantak, 2003; Skolnick, 2015).
Based in part on the results of self-administration studies, a number of these vaccines
have progressed to clinical trials, representing an exciting new potential rehabilitative
tool for reducing drug taking (Heidbreder & Hagan, 2005; Skolnick, 2015).
Overall, the self-administration model allows for direct assessment of drug
taking behaviour, and reduces some of the confounds associated with investigating the
effects of experimenter-administered MDMA on animals (De La Garza et al., 2007;
Fantegrossi, 2007). Furthermore, self-administration of MDMA produces different
neuroadaptations to experimenter administration, and these neuroadaptations are
probably more similar to those produced by regular recreational ecstasy use.
Profile of MDMA self-administration
Just as humans show tremendous variability in their patterns of ecstasy use,
there is considerable variability in the self-administration of MDMA in animals. More
specifically, some individuals are more vulnerable to the reinforcing effects of
MDMA, and the reinforcing efficacy of MDMA appears to increase with repeated

19
exposure. To investigate this variability our laboratory has defined an acquisition
criterion and measured latency to acquisition of MDMA self-administration. To meet
the acquisition criterion a rat must self-administer a total of 90 infusions of MDMA
(1.0 mg/kg/infusion) within 25 self-administration sessions. Our laboratory has shown
that roughly 50% of subjects will acquire MDMA self-administration under these
conditions (Colussi-Mas, Wise, Howard, & Schenk, 2010; Schenk et al., 2012; Schenk
et al., 2003; Schenk et al., 2007). Figure 1.2 presents raw data collected for this thesis
that help to illustrate the variability in acquisition of MDMA self-administration. The
top panel shows the number of MDMA infusions self-administered within the 25
session for a subject (Kererū) that did not meet acquisition criteria. Responding across
the 25 sessions is consistently low, although up to 3 infusions were self-administered
within a session. It should be noted that this rate of self-administration is lower than
that supported by the saline vehicle, which supports roughly 5-7 infusions per session.
The middle panel shows the same data for a subject (Black Stilt) that was slow to
acquire (24 sessions), while the bottom panel shows a subject (Kea) that acquired
relatively quickly (13 sessions). As is typical in these self-administration studies,
initial intake is low in all 3 subjects, but the subjects that did acquire show a sudden
increase in intake. That Kea required less self-administration experience before
increasing intake than Black Stilt shows the underlying variability in the reinforcing
effects of MDMA between these subjects, and this variability is reflected in their
latency to acquisition. The decrease in Kea’s responding in session 7 is typical after
the first exposure to a high dose of MDMA, and can be seen to a lesser extent in the
data from Black Stilt.

20
S e lf-a d m in is tra tio n s e s s io n
Infu
sio
ns
pe
r 2
hr
se
ss
ion
0 5 1 0 1 5 2 0 2 50
5
1 0
1 5
2 0
Subject: Kererū
S e lf-a d m in is tra tio n s e s s io n
Infu
sio
ns
pe
r 2
hr
se
ss
ion
0 5 1 0 1 5 2 0 2 50
5
1 0
1 5
2 0
Subject: Black Stilt
S e lf-a d m in is tra tio n s e s s io n
Infu
sio
ns
pe
r 2
hr
se
ss
ion
0 5 1 0 1 5 2 0 2 50
5
1 0
1 5
2 0
Subject: Kea

21
Fig. 1.2 Number of MDMA infusions (1.0 mg/kg/infusion) self-‐administered over a 25 day acquisition period. Top panel: a subject that did not acquire. Middle panel: A subject that was slow to acquire (24 sessions). Bottom panel: A subject that was relatively quick to acquire (13 sessions). Of the subjects that do acquire MDMA self-administration, some self-
administer more reliably than others. In our laboratory once a subject meets the
acquisition criterion the dose of MDMA is halved, and we expect that responding will
compensate accordingly. Often, we will further increase the FR schedule so that more
responses are required to obtain an infusion of drug. Some subjects will not increase
responding as the FR schedule is increased, while in others responding will
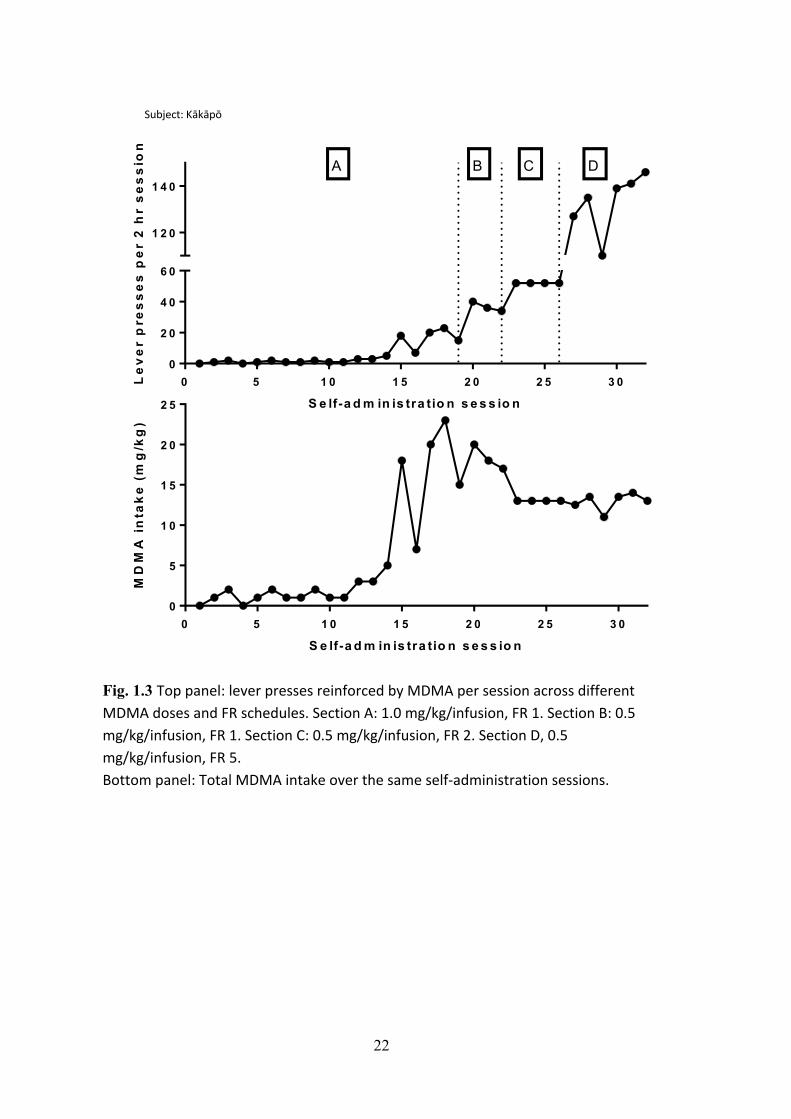
compensate for increases in FR (see Chapter 7). Figure 1.3 illustrates these
compensatory increases in responding with data collected for this thesis from a rat that
shows reliable self-administration. The top panel shows that responding compensated
for the decrease in dose (from section A to section B), and for increases in FR values
(sections C and D). The bottom panel shows that total intake becomes consistent over
time, although initially there is some variability, particularly after the first high dose of
MDMA was self-administered.
These patterns of self-administration behaviour have been a focus of our
laboratory for some time. In particular, we find it interesting that some subjects will
increase responding for MDMA after relatively low MDMA intake, while others will
show similar behaviour after relatively high MDMA intake, and others still will not
increase responding for MDMA within our 25 day cut-off period. We have suggested
that this behavioural profile might reflect the pharmacodynamic profile of MDMA.
22
S e lf-a d m in is tra tio n s e s s io n
Le
ve
r p
res
se
s p
er
2 h
r s
es
sio
n
0 5 1 0 1 5 2 0 2 5 3 00
2 0
4 0
6 0
1 2 0
1 4 0
A B C D
S e lf-a d m in is tra tio n s e s s io n
MD
MA
in
tak
e (
mg
/kg
)
0 5 1 0 1 5 2 0 2 5 3 00
5
1 0
1 5
2 0
2 5
Subject: Kākāpō
Fig. 1.3 Top panel: lever presses reinforced by MDMA per session across different MDMA doses and FR schedules. Section A: 1.0 mg/kg/infusion, FR 1. Section B: 0.5 mg/kg/infusion, FR 1. Section C: 0.5 mg/kg/infusion, FR 2. Section D, 0.5 mg/kg/infusion, FR 5. Bottom panel: Total MDMA intake over the same self-‐administration sessions.

23
Pharmacodynamics of MDMA
MDMA has a diverse pharmacodynamic profile. Battaglia, Brooks,
Kulsakdinun, and De Souza (1988) categorised the 5-HT transporter, 5-HT2 receptors,
α2 adrenergic receptors, and M-1 muscarinic receptors as targets for which MDMA
has high affinity (0-10µM). Moderate affinity (10µM-100µM) targets included the
norepinephrine and dopamine transporters, and 5-HT1 receptors, and low affinity
(>100µM) targets included dopamine-D1 and -D2 receptors and the choline
transporter. A small number of studies have shown that MDMA produces modest
increases in extracellular levels of glutamate (Anneken & Gudelsky, 2012; Nash &
Yamamoto, 1992) and acetylcholine (Acquas et al., 2001; Nair & Gudelsky, 2006a,
2006b), but there is limited evidence for effects on extracellular norepinephrine (Starr,
Page, & Waterhouse, 2012) or GABA (Bankson & Yamamoto, 2004; Yamamoto,
Nash, & Gudelsky, 1995). In contrast, a great deal of research on MDMA has focused
on 5-HT and dopamine mechanisms.
MDMA preferentially releases 5-HT via reverse transport (Gu & Azmitia,
1993; Gudelsky & Nash, 1996; Hekmatpanah & Peroutka, 1990). Although MDMA
has moderate affinity for the norepinephrine and dopamine transporters, MDMA is
more potent at releasing 5-HT (EC50=74.3 nM) than norepinephrine (EC50=136 nM) or
dopamine (EC50=278 nM) (Baumann, Wang, & Rothman, 2007). MDMA also inhibits
the 5-HT transporter (Berger, Gu, & Azmitia, 1992; Rothman & Baumann, 2003),
vesicular monoamine transporter 2 (Bogen, Haug, Myhre, & Fonnum, 2003; Erickson,
Schafer, Bonner, Eiden, & Weihe, 1996; Pifl, Reither, & Hornykiewicz, 2015) and
activity of monoamine oxidase A and B (Leonardi & Azmitia, 1994; Matsumoto et al.,
2014; Scorza et al., 1997). Thus, MDMA enhances extracellular 5-HT levels by
inhibiting the reuptake of 5-HT, directly releasing 5-HT from terminals, inhibiting the
packaging of 5-HT into vesicles, and inhibiting the degradation of 5-HT. Results from
in vivo microdialysis studies reliably show that MDMA preferentially increases
extracellular 5-HT levels (For review see Schenk (2011)). Following acute
administration of MDMA there was an immediate (15 min) and prolonged (2 week)
decrease in tryptophan hydroxylase activity, as measured by a 14CO2-trapping
procedure (Schmidt & Taylor, 1987; Stone, Hanson, & Gibb, 1987; Stone, Johnson,
Hanson, & Gibb, 1988; Stone, Merchant, Hanson, & Gibb, 1987), indicating that
MDMA also inhibits the further production of 5-HT.

24
MDMA produces minor and transient reductions in dopamine transporter
function, as measured in ex vivo synaptosomes, but failed to alter dopamine
transporter binding or tyrosine hydroxylase activity in rats (J. P. Hansen et al., 2002;
Stone, Merchant, et al., 1987). Nonetheless, MDMA administration increases
extracellular dopamine levels, as determined by in vivo microdialysis (for review, see
Schenk (2011)). This increase is more modest than the MDMA-produced increase in
extracellular 5-HT levels. For example, there was a 300% increase in extracellular
dopamine concentrations in the nucleus accumbens following 3 mg/kg MDMA, but an
1800% increase in extracellular 5-HT concentrations (Baumann, Clark, & Rothman,
2008).
A focus on 5-HT
A question remains as to which of these effects of MDMA might be related to
its self-administration. A wealth of data indicate that the reinforcing efficacy of a drug
is directly related to its ability to increase synaptic levels of dopamine. For example,
dopamine agonists reduced self-administration in a manner consistent with a leftward
shift in the dose-response curve (Gardner, 2000; Yokel & Wise, 1978), suggesting
enhanced reinforcement. On the other hand, dopamine antagonists produced
responding consistent with a rightward shift in the dose-response curve (de Wit &
Wise, 1977; Ettenberg, Pettit, Bloom, & Koob, 1982; Gardner, 2000; Yokel & Wise,
1975), suggesting a decrease in reinforcement. Similarly, neurotoxic, 6-OH-DA,
lesions also reduced the reinforcing efficacy of drugs of abuse (Gardner, 2000; Lyness,
Friedle, & Moore, 1979; Roberts, Corcoran, & Fibiger, 1977; Roberts & Koob, 1982).
As is true with other drugs of abuse, the reinforcing efficacy of MDMA, and
thus the self-administration of MDMA, results from dopamine release. As indicated
above, however, MDMA preferentially increases 5-HT, an effect that is incompatible
with self-administration. For example, stimulation of 5-HT release inhibited (Rothman
et al., 2005), while neurotoxic 5,7-DHT lesions enhanced (Bradbury et al., 2014; Loh
& Roberts, 1990) self-administration. Self-administration of amphetamine-type drugs
was inversely related to affinity for the 5-HT transporter (Ritz & Kuhar, 1989), or
potency to stimulate 5-HT release (Wee et al., 2005). With specific reference to
MDMA, the (+) isomer that selectively releases dopamine was more readily self-
administered than the (-) isomer that selectively releases 5-HT (Z. Wang &
Woolverton, 2007). That is, higher levels of 5-HT release are inhibitory to self-

25
administration in general, and to MDMA self-administration in particular. Thus,
MDMA-produced 5-HT release would be expected to inhibit MDMA self-
administration, yet, as outlined above, some rats will eventually self-administer
MDMA reliably. It is possible that some rats are less responsive to these 5-HTergic
effects and so self-administer MDMA more readily.
This hypothesis was recently directly tested in our laboratory. Firstly, the 5-
HTergic response to an initial dose of MDMA was determined by in vivo microdialysis
before MDMA self-administration began. 5-HT release produced by this initial
exposure to MDMA was lower in the rats that did acquire MDMA self-administration
than in those that did not, while dopamine release was similar for both groups.
Secondly, the effect of a neurotoxic 5,7-DHT lesion on acquisition of MDMA self-
administration was determined. The lesion reduced 5-HT tissue levels by up to 67%.
Of interest, 100% of the lesion group acquired MDMA self-administration, compared
to approximately 50% of controls, and the latency to acquisition was greatly reduced in
the lesion group (Bradbury et al., 2014). Thus, lower 5-HT release produced by
MDMA, either endogenous or exogenously produced by a lesion, was associated with
enhanced self-administration. These findings support the hypothesis that MDMA-
produced 5-HT release is inhibitory to the acquisition of MDMA self-administration,
but a question remains as to the mechanism for this inhibitory effect.
It has been suggested that the development of MDMA as an efficacious
reinforcer in the self-administration paradigm is due to neuroadaptations that occur in
response to regular MDMA exposure, and that the same neuroadaptations could
underlie the development of ecstasy SUDs (Schenk, 2011; Schenk & Aronsen, 2015).
Microdialysis studies have shown that the 5-HTergic response to MDMA is attenuated
after repeated exposure (Baumann, Clark, Franken, Rutter, & Rothman, 2008;
Reveron, Maier, & Duvauchelle, 2010; Shankaran & Gudelsky, 1999), an effect that
would be expected to facilitate MDMA self-administration. It has been hypothesised
that this reduced 5-HTergic response to MDMA disinhibits the dopaminergic response,
enhancing the reinforcing efficacy of MDMA and making it comparable to other drugs
of abuse (Schenk, 2011). Furthermore, neuroadaptations in 5-HT receptors, as a result
of MDMA exposure, have been suggested to enhance problematic behaviours, like
impulsivity, that are associated with SUDs (Schenk & Aronsen, 2015).
Thus, repeated exposure to MDMA reduces the 5-HTergic response to MDMA,
enhancing its reinforcing effects and producing behaviours that may contribute to

26
problematic drug taking. If the reinforcing effects of MDMA rely on 5-HTergic
deficits, the variability in acquisition of MDMA self-administration might be due to
increased vulnerability to MDMA-produced 5-HTergic neuroadaptations in some rats.
Because 5-HTergic deficits enhance MDMA self-administration via a disinhibition of
dopamine, there are likely specific 5-HT receptors that modulate the dopaminergic
response to, and thus the self-administration of, MDMA.
There are 14 different 5-HT receptor subtypes, arranged into 7 receptor
families, and spread widely throughout the brain (Hoyer et al., 1994). The 5-HT1A and
5-HT1B receptor subtypes have a role in the regulation of dopamine and the
dopaminergic response to drugs of abuse, and as such changes in the activation of
these receptor subtypes might be expected to alter the reinforcing effects of MDMA.
5-HT1A and 5-HT1B receptors
The 5-HT1A receptor is a seven transmembrane receptor that couples to Gi/Go
to inhibit adenylyl cyclase and produce hyperpolarisation (Hamon et al., 1990; Innis,
Nestler, & Aghajanian, 1988; Schoeffter & Hoyer, 1988). In the brain the 5-HT1A
receptor is located both pre- and post-synaptically. Pre-synaptically, the 5-HT1A
receptor is an autoreceptor on 5-HT neurons in the dorsal and median raphe nuclei,
where activation inhibits 5-HT synthesis, and release of 5-HT in terminal regions
(Hamon et al., 1988; Riad et al., 2000; Yoshimoto & McBride, 1992). 5-HT1A
receptors have also been localised to the hippocampus, amygdala, prefrontal cortex
(PFC), and the ventral tegmental area (VTA) where they act as heteroreceptors on
dopamine, glutamate, and GABA cells (Doherty & Pickel, 2001; Hajós, Gartside,
Varga, & Sharp, 2003; Hume et al., 2001; Maeda et al., 2001; Palchaudhuri & Flügge,
2005; Pompeiano, Palacios, & Mengod, 1992; Puig, Artigas, & Celada, 2005; Puig,
Watakabe, Ushimaru, Yamamori, & Kawaguchi, 2010).
The 5-HT1B receptor is also a seven transmembrane protein that couples to
Gi/Go to inhibit adenylyl cyclase and produce hyperpolarisation (Hartig, Branchek, &
Weinshank, 1992; Hoyer & Middlemiss, 1989; Sari, 2004; Seuwen, Magnaldo, &
Pouysségur, 1988; C. Wang et al., 2013). In the brain the 5-HT1B receptor is located
pre-synaptically on the terminals of 5-HTergic or non-5-HTergic cells, as auto- or
heteroreceptors, respectively (Boulenguez et al., 1996; Offord, Ordway, & Frazer,
1988; Sari et al., 1999; Vergé et al., 1986). 5-HT1B receptor binding was high in globus
pallidus, substantia nigra, nucleus accumbens, frontal cortex, striatum, and

27
hippocampus (Bonaventure, Schotte, Cras, & Leysen, 1997; Lindhe et al., 2011). 5-
HT1B mRNA was also abundant in the hypothalamus, thalamus, and amygdala
(Bonaventure et al., 1998). As well as being present on 5-HTergic neurons, 5-HT1B
receptors have been localised to dopaminergic (Sarhan & Fillion, 1999), GABAergic
(Darrow, Strahlendorf, & Strahlendorf, 1990), and glutamatergic (Raiteri, Maura,
Bonanno, & Pittaluga, 1986) terminals.
Changes in 5-HT1A and 5-HT1B receptor protein levels, mRNA levels, or
binding to G proteins, have been shown in response to general interventions such as
exercise (Chennaoui et al., 2001; Fuss et al., 2013), stress (Iyo et al., 2009; S. Wang,
Zhang, Guo, Teng, & Chen, 2009), and steroid administration (Ambar & Chiavegatto,
2009; Kindlundh, Lindblom, Bergström, & Nyberg, 2003). More importantly, changes
have also been shown after repeated exposure to 5-HTergic ligands. 5-HT1A
autoreceptors were down-regulated by chronic exposure to selective 5-HT reuptake
inhibitors (SSRIs) (Castro, Diaz, del Olmo, & Pazos, 2003; Le Poul et al., 2000),
although no changes in 5-HT1A autoreceptors were detected after repeated exposure to
MDMA (Schenk, Abraham, Aronsen, Colussi-Mas, & Do, 2013). Chronic SSRI
treatment also increased post-synaptic 5-HT1A receptor agonist-stimulated binding of
[35S]GTPγS to G proteins (Castro et al., 2003; Moulin-Sallanon et al., 2009). Similarly,
up-regulation of post-synaptic 5-HT1B receptors has been suggested as a result of
repeated SSRI treatment (Le Poul et al., 2000). These findings suggest that the large
increases in synaptic 5-HT produced by MDMA could also produce changes in these
receptor subtypes.
The role of 5-HT1A and 5-HT1B receptors in dopamine modulation
The most commonly used 5-HT1A receptor agonist, 8-hydroxy-2-
dipropylaminotetralin (8-OH-DPAT), has high affinity for 5-HT1A receptors (Peroutka,
1986). Low doses of 8-OH-DPAT preferentially activated 5-HT1A autoreceptors, while
higher doses also activated heteroreceptors (Alex & Pehek, 2007; Hjorth &
Magnusson, 1988). Low doses of 8-OH-DPAT simulated dopamine cell activity in the
VTA (Gronier, 2008) and increased extracellular dopamine concentrations in the PFC
(Arborelius, Nomikos, Hacksell, & Svensson, 1993) and VTA (Chen & Reith, 1995).
Thus, activation of 5-HT1A autoreceptors enhances dopamine cell activity and
extracellular dopamine concentrations.
Higher doses of 8-OH-DPAT inhibited dopamine cell firing in the VTA
(Arborelius, Chergui, et al., 1993) and decreased extracellular dopamine levels in the

28
nucleus accumbens (NAc) (Ichikawa & Meltzer, 2000) and striatum (Rasmusson,
Goldstein, Deutch, Bunney, & Roth, 1994), as measured by in vivo microdialysis. The
relatively new 5-HT1A receptor agonists, F13640, and F15599, both have >1000 fold
selectivity for the 5-HT1A receptor (Colpaert et al., 2002; Newman-Tancredi et al.,
2009), while the slightly older BAY × 3702 has approximately 30 fold selectivity for
the 5-HT1A receptor (De Vry et al., 1998). Each of these agonists, when administered
locally in the PFC, dose-dependently increased extracellular dopamine levels in the
PFC (Díaz-Mataix, Artigas, & Celada, 2006; Díaz-Mataix et al., 2005; Lladó-Pelfort,
Assié, Newman-Tancredi, Artigas, & Celada, 2012; Lladó-‐Pelfort, Assié, Newman-‐
Tancredi, Artigas, & Celada, 2010). It was suggested that this effect was due to
inhibition of PFC GABA and glutamate cells, since endogenous 5-HT release inhibited
electrophysiological recordings from PFC glutamate and GABA cells, and this effect
was attenuated by a 5-HT1A receptor antagonist (Hajós et al., 2003; Puig et al., 2005;
Puig et al., 2010; Sakaue et al., 2000). Thus, the effect of 5-HT1A receptor activation
on dopamine release is region specific – activation of autoreceptors, or heteroreceptors
in the PFC, increased extracellular dopamine concentrations, while global activation of
heteroreceptors decreased extracellular dopamine concentrations in the NAc and
striatum.
There is evidence that activation of 5-HT1A receptors is inhibitory to the
dopaminergic response to drugs of abuse. The 5-HT1A receptor agonist, 8-OH-DPAT,
inhibited amphetamine-induced dopamine release in the PFC (Kuroki, Ichikawa, Dai,
& Meltzer, 1996), striatum and NAc (Ichikawa, Kuroki, Kitchen, & Meltzer, 1995) as
determined by microdialysis. 5-HT1A receptor agonists generally inhibited the
hyperactive response to amphetamine, methamphetamine, and MDMA (Müller, Carey,
Huston, & Silva, 2007), a response that has been associated with enhanced dopamine
neurotransmission (Wise & Bozarth, 1987). Furthermore, the expression and
development of cocaine or amphetamine sensitisation in mice was inhibited by 5-HT1A
receptor agonist administration (Ago et al., 2006; Przegaliñski, Siwanowicz, Baran, &
Filip, 2000). Thus, increased activation of 5-HT1A receptors during MDMA self-
administration might be expected to inhibit the dopaminergic response to MDMA.
In vitro studies showed that activation of 5-HT1B receptors inhibited the release
of dopamine (Sarhan & Fillion, 1999), GABA (Johnson, Mercuri, & North, 1992; Yan
& Yan, 2001b), and glutamate (Muramatsu, Lapiz, Tanaka, & Grenhoff, 1998), but
these studies do not consider interactions between neurotransmitter systems. One of

29
the most widely used 5-HT1B receptor agonists, RU 24969 (5-Methoxy-3-(1,2,5,6-
tetrahydro-4-pyridinyl)-1H-indole), has high affinity for 5-HT1B receptors (Ki = 0.38
nM), but also displays appreciable affinity for the 5-HT1A receptor (Ki = 2.5 nM)
(Peroutka, 1986; Wolf & Kuhn, 1991). Systemic administration of RU 24969
decreased extracellular GABA concentrations in the VTA in vivo, but had no effect on
extracellular dopamine concentrations (Parsons, Koob, & Weiss, 1999). However,
local administration of the 5-HT1B receptor agonist, CP 93129, which has 150 fold
selectivity for 5-HT1B over other 5-HT receptors (Macor et al., 1990), increased
extracellular dopamine concentrations in the PFC (Iyer & Bradberry, 1996), striatum
(Galloway, Suchowski, Keegan, & Hjorth, 1993), and NAc (Hållbus, Magnusson, &
Magnusson, 1997; Yan & Yan, 2001a). Similarly, administration of CP 93129 in the
VTA increased extracellular dopamine levels in the NAc (O'Dell & Parsons, 2004;
Yan & Yan, 2001a; Yan, Zheng, & Yan, 2004) and decreased extracellular GABA
concentrations in the VTA (O'Dell & Parsons, 2004; Yan et al., 2004), without altering
extracellular glutamate concentrations in the VTA (O'Dell & Parsons, 2004). Together
these findings suggest that activation of 5-HT1B receptors enhances dopamine release,
possibly via an inhibition of GABA neurotransmission.
There is evidence that activation of 5-HT1B receptors enhances the
dopaminergic response to drugs of abuse. Cocaine produced significantly greater
increases in extracellular dopamine, and significantly greater reductions in
extracellular GABA, in the NAc after systemic administration of the 5-HT1B/1A
receptor agonist, RU 24969 (Parsons et al., 1999). A similar response to cocaine was
found after infusion of the 5-HT1B receptor agonist, CP 93129, in the VTA (O'Dell &
Parsons, 2004). Systemic administration of the 5-HT1B receptor agonist, CP 94253,
which has approximately 45 fold selectivity for 5-HT1B over other 5-HT receptors
(Koe, Nielsen, Macor, & Heym, 1992), significantly prolonged the increase in
extracellular dopamine in the NAc produced by systemic administration of ethanol
(Yan, Zheng, Feng, & Yan, 2005). 5-HT1B receptor agonists produced a leftward shift
in the cocaine self-administration dose response curve, increased the break points
achieved in cocaine progressive ratio tasks (Parsons, Weiss, & Koob, 1998;
Pentkowski, Acosta, Browning, Hamilton, & Neisewander, 2009; Przegaliñski, Gołda,
Frankowska, Zaniewska, & Filip, 2007), and produced a leftward shift in the self-
administration dose response curve for the dopamine uptake inhibitor, GBR 12909
(Parsons, Weiss, & Koob, 1996). Therefore, activation of 5-HT1B receptors during

30
MDMA self-administration might be expected to enhance the dopaminergic response
to MDMA.
Thus, 5-HT1A and 5-HT1B receptors might be expected to impact the initial
reinforcing effects of MDMA, via dopaminergic modulation. Specifically, activation
of 5-HT1A receptors would be expected to decrease the dopaminergic response to
MDMA, while activation of 5-HT1B receptors would be expected to enhance the
dopaminergic response to MDMA. Furthermore, alterations in the activation of these
receptors might explain the enhancement in the reinforcing efficacy of MDMA after
repeated exposure. As outlined below, individual variability in these receptor
populations, or MDMA-produced changes in these receptor populations, might also
impact the reinforcing effects of MDMA and explain behavioural deficits seen in
regular ecstasy users.
5-HT1A and 5-HT1B receptors and impulsivity
Impulsivity has been broadly defined as action without foresight, referring to
behaviours that are poorly thought out, prematurely executed, or risky (Winstanley,
Eagle, & Robbins, 2006). In drug users impulsivity is a risk factor for initiating drug
taking, escalating drug use, and for developing SUDs (De Wit, 2009; Perry & Carroll,
2008). For example, impulsive traits in youth and young-adulthood positively
predicted future drug use, an earlier onset of drug taking, and the likelihood of
developing an SUD (De Wit, 2009; Kirisci, Tarter, Mezzich, & Vanyukov, 2007;
Sher, Bartholow, & Wood, 2000; Tarter, Kirisci, Feske, & Vanyukov, 2007).
A role of impulsivity in different aspects of drug self-administration in animals
has been determined. Some studies have looked at the acquisition and maintenance of
self-administration, based on the idea that highly impulsive subjects, as is the case with
humans, might be more prone to take drugs (Perry & Carroll, 2008). Typically,
impulsivity is measured by a model of behavioural inhibition, such as the 5 choice
serial reaction time task (5CSRTT), or a model of choice preference for a delayed
reward, such as the delay discounting paradigm. These measures show good validity as
they are variants of those used to assess aspects of impulsive behaviour in humans
(Evenden, 1999b; Robbins, 2002). Delay discounting and reaction time tasks can be
used to determine impulsivity scores across a group of animal subjects, which can then
be divided into ‘low impulsivity’ (LI) groups and ‘high impulsivity’ (HI) groups. HI
subjects are usually defined as those in the upper quartile of impulsivity scores, with
LI subjects being those with impulsivity scores in the bottom quartile. These two

31
groups can then be compared to determine the relationship between impulsivity and
drug self-administration.
When impulsivity was determined using a delay discounting task HI rats
consumed more ethanol (Poulos, Le, & Parker, 1995), or cocaine (Koffarnus &
Woods, 2013; Perry, Larson, German, Madden, & Carroll, 2005; Perry, Nelson, &
Carroll, 2008), and cocaine self-administration was acquired more quickly and in a
higher percentage of HI rats (Perry et al., 2005; Zlebnik & Carroll, 2015). Similarly,
HI rats, as measured by 5CSRTT performance, acquired nicotine self-administration
more readily (Diergaarde et al., 2008), and a strain of mice with high impulsivity
showed enhanced ethanol self-administration (Loos, Staal, Smit, De Vries, & Spijker,
2013). Following acquisition, HI rats, as determined by the 5CSRTT, self-administered
more cocaine per hour than LI rats, and exhibited an upward shift in the cocaine dose
response curve (Dalley et al., 2007). Furthermore, impulsivity as determined by the
5CSRTT predicted the magnitude of the drug-seeking response for MDMA in the
reinstatement paradigm (Bird & Schenk, 2013). Thus, higher levels of impulsivity
would be expected to facilitate self-administration.
Systemic administration of the 5-HT1A receptor agonist, 8-OH-DPAT,
increased premature responding on the 5CSRTT (Carli & Samanin, 2000) while the 5-
HT1A receptor antagonist, WAY 100635 (N-[2-[4-(2-methoxyphenyl)-1-
piperazinyl]ethyl]-N-(2-pyridinyl)cyclohexanecarboxamide), which has >100-fold
selectivity for the 5-HT1A receptor over other receptors (A. Fletcher et al., 1995),
supressed impulsive action in a 3CSRTT (Ohmura et al., 2013). The effects of 5-HT1A
ligands on impulsivity appear to be due to autoreceptor activation, because neither
local administration of 8-OH-DPAT in the PFC, nor systemic administration of the 5-
HT1A post-synaptic preferring receptor agonist F15599, affected premature responding
on the 5CSRTT (Carli, Baviera, Invernizzi, & Balducci, 2006; Lladó-‐Pelfort et al.,
2010; Winstanley et al., 2003).
In humans, 5-HT1B receptor gene polymorphisms are associated with impulsive
aggression (Zouk et al., 2007). Mice that lack the 5-HT1B receptor gene from birth
show increased impulsivity in a behavioural model of response inhibition (Nautiyal et
al., 2015; Pattij et al., 2003). Interestingly, knockdown of 5-HT1B autoreceptors did not
affect impulsivity, suggesting the effect of 5-HT1B receptor activation on impulsivity is
due to heteroreceptor action (Nautiyal et al., 2015). Studies of the effects of 5-HT1B
ligands on impulsivity have been limited due to the fact that agonists have a range of

32
behavioural effects that disrupt operant responding (Evenden, 1999a; van den Bergh,
Bloemarts, Groenink, Olivier, & Oosting, 2006). However, the limited available data
suggest that activation of 5-HT1B receptors reduces impulsive behaviour (Evenden,
1999a). Therefore, activation of 5-HT1B receptors during MDMA self-administration
would be expected to reduce impulsive behaviour, and thus inhibit self-administration.
5-HT1A and 5-HT1B receptors and learning
Before reliable self-administration behaviour can be demonstrated, the subject
must learn the association between performance of the operant response and the
infusion of drug. Enhanced or inhibited ability to learn this association would clearly
also enhance or inhibit the acquisition of self-administration. There is also another
learned association that has been shown to be incredibly important in the self-
administration paradigm - the Pavlovian association between the drug effect and the
contextual stimuli (e.g. the light). These unconditioned contextual stimuli develop
conditioned reinforcement properties over repeated pairings with a drug (Ahrens,
Singer, Fitzpatrick, Morrow, & Robinson, 2016; W. M. Davis & Smith, 1976; P. J.
Fletcher & Korth, 1999b) and these conditioned reinforcers are a powerful driver of
self-administration behaviour. For example, one experiment assessed the acquisition of
nicotine self-administration in two groups of rats – one in which the nicotine infusion
was paired with the illumination of a light, and another in which the infusion was
paired with no specific cues. Rats in the nicotine + cue group took less time to show a
preference for the active self-administration lever and consumed significantly more
nicotine than the nicotine only group, suggesting the Pavlovian association between
drug effect and contextual cues facilitated acquisition of self-administration (Caggiula
et al., 2002). Therefore, enhanced or inhibited learning of either operant or Pavlovian
associations would be expected to enhance or inhibit self-administration, respectively.
The strengthening of stimulus/reward associations is markedly impacted by
pharmacological manipulation of 5-HT1A receptors. Systemic 5-HT1A receptor agonist
administration impaired performance on an appetitive Pavlovian conditioned
responding task (Blair, Bonardi, & Hall, 2004), increased errors in a repeated
acquisition of response sequence task (Winsauer, Rodriguez, Cha, & Moerschbaecher,
1999) and delayed acquisition of operant responding maintained by a food reinforcer
(Frick, Bernardez-Vidal, Hocht, Zanutto, & Rapanelli, 2015). Furthermore, the 5-HT1A
receptor agonist, 8-OH-DPAT, administered after an initial training session, impaired
further operant responding for food (Meneses, 2007). The lack of comprehensive dose-

33
response functions in these studies limits the degree to which the relative roles of 5-
HT1A auto- and heteroreceptors can be disentangled. Importantly, the above results
were noted over a range of 8-OH-DPAT doses that would be expected to activate pre-
and post-synaptic 5-HT1A receptors (up to 1.0 mg/kg). When low doses of 8-OH-
DPAT were used, operant learning was enhanced (Meneses & Hong, 1994b), and this
effect was reversed by the tryptophan hydroxylase inhibitor, pCPA (Meneses & Hong,
1994a). Together, these results suggest that activation of 5-HT1A autoreceptors
enhances, while activation of 5-HT1A heteroreceptors inhibits, learning of
stimulus/reward associations.
The non-selective 5-HT receptor agonist, mCPP, inhibited operant
stimulus/response learning, and this effect was reversed by the non-selective 5-HT1B
receptor antagonist, propranolol (Meneses & Hong, 1997). Moreover, the 5-HT1B
receptor agonist, CGS 12066 impaired (Meneses, 2007), while the 5-HT1B/1D receptor
antagonist, GR 127935, improved (Meneses, Terrón, & Hong, 1997) performance on
the same task. Similarly, the 5-HT reuptake facilitator, tianeptine, enhanced operant
stimulus/response learning, and this effect was reversed by the 5-HT1B receptor inverse
agonist, SB 224289 (Meneses, 2002). These findings suggest that activation of 5-HT1B
receptors inhibits the consolidation of operant learning, and so activation of 5-HT1B
receptors during MDMA self-administration might be expected to inhibit the
development of self-administration.
5-HT1A and 5-HT1B receptors and anxiety
Anxiety disorders are frequently comorbid with SUDs (Ipser, Wilson,
Akindipe, Sager, & Stein, 2015; Merikangas et al., 1998). It has been suggested that
anxiety may underlie the initiation of drug taking, in order to alleviate a negative
emotional state, and negatively reinforce the continuation of drug use to mitigate
withdrawal symptoms (Altman et al., 1996; Belin, Belin-‐Rauscent, Everitt, & Dalley,
2015; Lejuez et al., 2008). In animal models, anxiety is often operationalised in
rodents as an aversion to open or brightly lit spaces (Belin et al., 2015). A popular
method for measuring anxiety is the elevated plus maze (EPM), in which a preference
for the closed (protected) arms of the maze over the open arms is regarded as an
‘anxious’ response (Pellow, Chopin, File, & Briley, 1985). An alternate measure of
rodent anxiety is self-grooming behaviour in response to an environmental change
(Homberg et al., 2002). Rats in the upper quartile for time spent grooming in a novel
environment reached higher break points in progressive ratio cocaine self-

34
administration than the lower quartile group (Homberg et al., 2002). This effect was
not replicated when high anxiety was determined by performance on the EPM,
however in this case high and low anxiety were determined using a median split, thus
possibly masking an effect of anxiety (Bush & Vaccarino, 2007). Higher anxiety on
the EPM was associated with escalation of cocaine self-administration (Dilleen et al.,
2012), and propensity to self-administer alcohol (Spanagel et al., 1995). Thus, higher
levels of anxiety would be expected to facilitate self-administration.
Time spent in the open arms of the EPM was increased by systemic
administration of low doses of the 5-HT1A receptor agonist, 8-OH-DPAT (Kwieciński
& Nowak, 2009; Lalonde & Strazielle, 2010), an effect reversed by the 5-HT1A
receptor antagonist, WAY 100635 (Collinson & Dawson, 1997), suggesting 5-HT1A
autoreceptor activation had an anxiolytic effect. Higher doses of systemically
administered 8-OH-DPAT had an anxiogenic effect in the same task in mice (Miheau
& Van Marrewijk, 1999).When injected into the dorsal or median raphe, 8-OH-DPAT
increased time spent in the open arms of the EPM (De Almeida, Giovenardi, Charchat,
& Lucion, 1998; File & Gonzalez, 1996; File, Gonzalez, & Andrews, 1996), while
injections into the hippocampus (Cheeta, Kenny, & File, 2000a; File et al., 1996; File,
Kenny, & Cheeta, 2000), PFC (Solati, Salari, & Bakhtiari, 2011), or septum (Cheeta,
Kenny, & File, 2000b; De Almeida et al., 1998) increased anxiety-like behaviour in
the EPM. Thus, activation of 5-HT1A autoreceptors had anxiolytic effects in the EPM,
while activation of post-synaptic 5-HT1A receptors was anxiogenic.
Early investigations of the role of 5-HT1B receptors in anxiety states found that
non-selective 5-HT1B receptor agonists decreased time spent in the open arms of an
EPM (Benjamin, Lal, & Meyerson, 1990; Critchley & Handley, 1987; Pellow,
Johnston, & File, 1987), suggesting that activation of 5-HT1B receptors was
anxiogenic. The role of 5-HT1B receptors in modulating anxiety was more recently
confirmed; entries into the open arms of the EPM were dose-dependently reduced by
the 5-HT1B receptor agonist CP 94253, and this effect was reversed by the 5-HT1B/1D
receptor antagonist, GR 127935 (Lin & Parsons, 2002). The relative contribution of 5-
HT1B auto- and heteroreceptors to this effect is not clear. 5-HT1B heteroreceptors on
GABAergic amygdala neurons have been suggested as a possible neuronal mechanism
(Lin & Parsons, 2002; Sari, 2004) because 5-HT1B manipulations of these projections
altered behaviour in the EPM (Audi, De Oliveira, & Graeff, 1991). Furthermore,
activation of 5-HT1B receptors in the PFC produced anxiogenic effects in the EPM

35
(Solati et al., 2011) however, a role of 5-HT1B autoreceptors cannot be ruled out (Sari,
2004).
Summary
MDMA is widely used recreationally in the form of the street drug, ecstasy.
Although the majority of users consume ecstasy intermittently, there is concern that
MDMA produces a range of deficits in regular ecstasy users. Among these deficits,
ecstasy users show increased anxiety and impulsivity, and impaired learning and
memory. Problematically, these behavioural changes might be expected to facilitate
further ecstasy taking.
MDMA is unique among drugs of abuse in that it primarily acts as a 5-HT
releasing agent. 5-HT release has been hypothesised to inhibit the self-administration
of drugs in general, and of MDMA in particular. Nonetheless, MDMA self-
administration is acquired in roughly 50% of animal subjects. It is possible that
MDMA-produced 5-HT release inhibits the reinforcing efficacy of MDMA via
activation of specific 5-HT receptors, but there is likely variability in the 5-HTergic
response to MDMA between individuals. Furthermore, it is possible that
neuroadaptations in 5-HT receptors underlie both the facilitated reinforcement
produced by MDMA after repeated exposure, and the cognitive and behavioural
deficits seen after regular use.
The 5-HT1A and 5-HT1B receptors are good candidates for these effects of
MDMA. Both receptors modulate the reinforcing effects of other drugs of abuse by
regulating dopamine release. Furthermore, these receptors mediate a number of
behaviours associated with self-administration that are impacted by regular ecstasy
use, and receptor up- or down-regulation has been documented in response to a
number of different interventions.
This thesis will explore two ways in which alterations in 5-HT1A and/or 5-HT1B
receptors could influence MDMA self-administration. Firstly, underlying differences
in 5-HT1A and/or 5-HT1B receptors could predispose some subjects to self-administer
more readily. This may explain the variability in acquisition of MDMA self-
administration. If so, it is hypothesised that manipulations that alter 5-HT1A and/or 5-
HT1B receptors will reduce the variability in the acquisition profile for MDMA self-
administration.

36
Secondly, MDMA exposure during self-administration might produce changes
in these receptor populations that might facilitate further drug taking and produce
behavioural deficits. If so, it is hypothesised that these changes will be evident after
substantial MDMA self-administration. Theoretically, if these changes are not the
result of neurotoxicity, they could be partially reversed by repeated administration of
selective agonists or antagonists.

37
General Methods
Subjects
Male Sprague-Dawley rats were bred in the Victoria University of Wellington
vivarium. They were housed in groups of 4 in a temperature- (19-21ºC) and humidity-
(55%) controlled environment until they reached weights of 300-350g, after which
they were housed individually. The housing colony was maintained on a 12 h
light/dark cycle (lights on at 7.00 am) and all tests were conducted during the light
portion of the cycle. Food and water were freely available except during testing.
Drugs
RU 24969 hemisuccinate, WAY 100635 maleate, lithium chloride, ±8-OH-
DPAT hydrobromide, (Tocris, New Zealand), and d-amphetamine sulfate (BDG, New
Zealand) were dissolved in sterilised saline. GR 127935 hydrochloride (Tocris, New
Zealand) was dissolved in distilled water. All injections were a volume of 1.0 ml/kg.
±MDMA hydrochloride (BDG, New Zealand) for self-administration was dissolved in
sterilised saline containing 3IU heparin per ml.
All doses refer to salt weights.
Apparatus and procedures
Water consumption
Water consumption was measured in the home cage. Water bottles were
removed for 24 hours. Drug administration occurred before water bottles were
reintroduced, at times specified in each study. Consumption was measured for a 30
minute period. Fluid consumption was determined by weighing water bottles before
and after the test.
Locomotor Activity
Locomotor activity testing was conducted in clear Plexiglas chambers (Med
Associates Inc., USA; model ENV-515) measuring 42×42×30 cm, set in sound-
attenuating boxes. Forward locomotion was measured with two sets of 16 infrared
beams and sensors spaced evenly along the sides of the chambers producing squares
measuring 25mm × 25mm. The interruption of three adjacent beams (the approximate
size of the body of a rat) was recorded as one activity count. A white noise generator
was used during experiments to mask any outside noise, and chambers were washed
with Virkon ‘S’ disinfectant (Southern Veterinary Supplies, NZ) after testing to control
for olfactory confounds. Experiments were run in a dark room, except for a red light

38
that was used to illuminate the room during drug administrations. Locomotor activity
counts were recorded in 5 minute intervals.
Surgery
For rats that underwent self-administration testing, a silastic catheter was
implanted into the right jugular vein under deep anesthesia produced by i.p. injection
of ketamine (90 mg/kg) and xylazine (9 mg/kg). Areas surrounding skin that was to be
cut were shaved and washed with ethanol and iodine, and eye lubricant (Refresh lacri-
lube) was administered to avoid drying. The catheter was secured in place using
surgical string and a small amount of adhesive (Bostick superglue). The distal end of
the catheter was passed subcutaneously to an exposed part of the skull, attached to a 3
cm piece of 22 gauge stainless steel tubing (BD needles), fixed in place with screws
and a small amount of adhesive, and embedded in dental acrylic (Ostron 100). The
silastic tubing was coated with silicone (Selleys wet area silicone) to protect from the
corrosive nature of the adhesive. Following surgery an analgesic (Carporfen ®, 5.0
mg/kg, s.c.) and electrolyte replacement (Hartman’s solution, 12 ml, s.c.) were
administered. Carprofen was also administered on each of two days following the
surgery. Testing began once pre-surgery weight had been attained, generally within 4-
6 days.
Self-administration
Every day, before self-administration testing, rats were weighed and
administered penicillin dissolved in heparinised saline (0.2 ml, i.v.) to help maintain
general health and catheter patency.
Self-administration was conducted in operant chambers (Med Associates ENV-
001) equipped with two levers. Depression of the active lever resulted in a 12 second
activation of a syringe pump (Razell, Model A, 1 RPM) resulting in a 0.1 ml
intravenous infusion, and the simultaneous illumination of the house light located
above the active lever. Depressions of the inactive lever were recorded, but had no
programmed consequence. Each self-administration session began with an
experimenter-delivered infusion to fill the volume of the catheter. These infusions are
not recorded and do not contribute to calculations of total self-administration intake.

39
Chapter 3: Development of behavioural assays
These first studies were designed to develop behavioural assays for 5-HT1A and
5-HT1B receptor activation, so that further testing of the effects of drug exposure on the
function of these receptor subtypes could be conducted. One assay that seemed
promising was latent inhibition: the impairment of learning that a stimulus predicts an
important event when that stimulus has previously been presented with no
consequence (Cassaday, Hodges, & Gray, 1993).
When a neutral stimulus, for example a tone, is paired with a negative
consequence, for example a footshock, that stimulus will develop conditioned-stimulus
properties and produce freezing behaviour in rodents. A group of rats that had never
been exposed to the tone (control group) would learn this association relatively
quickly. However, if the tone has previously been presented to another group of rats
without consequence (pre-exposure group), learning that the tone now predicts a
footshock will take longer in this group. Therefore, after a small number of pairings,
the tone will produce less freezing behaviour in the pre-exposure group, because the
association between the tone and the footshock is less well learned. Latent inhibition
can be operationalised as this behavioural difference (reduced freezing behaviour)
between groups. Latent inhibition is a robust effect, found across a range of stimulus-
consequence combinations in a wide range of species (Fernández, Giurfa, Devaud, &
Farina, 2012; Ferrari & Chivers, 2011; Lubow, 1989).
A number of studies have implicated 5-HT in latent inhibition. Electrolytic, or
neurotoxic 5,7 DHT, lesions of the median raphe or NAc blocked the latent inhibition
effect (Loskutova, 2001; Loskutova, Luk'yanenko, & Il'yuchenok, 1990; Solomon,
Nichols, Kiernan, Kamer, & Kaplan, 1980). Rats in the pre-exposure group showed
greater 5-HT metabolism in the striatum and amygdala than rats in the control group,
suggesting the latent inhibition effect is associated with increased 5-HTergic activity
(Molodtsova, 2003). Additionally, rats with a genetic deletion of the 5-HT transporter
showed reduced latent inhibition compared to wildtype counterparts (Nonkes et al.,
2012).
The role of 5-HT in latent inhibition is reinforced by studies employing
selective pharmacological ligands. The 5-HT1B/1A receptor agonist, RU 24969 (0.5
mg/kg), administered before each pre-exposure, inhibited the development of latent
inhibition, while the more selective 5-HT1A receptor agonist, 8-OH-DPAT, had no
significant effect (Cassaday et al., 1993), demonstrating a role of 5-HT1B receptors in

40
latent inhibition. On the other hand, the 5-HT1A receptor antagonist, WAY 100635 (0.5
mg/kg), facilitated latent inhibition in the same task (Killcross, Stanhope, Dourish, &
Piras, 1997), suggesting 5-HT1A receptors also impact the expression of latent
inhibition. Thus, RU 24969 inhibited the development of latent inhibition, but the
relative roles of 5-HT1A and 5-HT1B receptor activation in this effect, and the range of
doses over which this effect is produced, have not been determined.
The majority of studies have employed a footshock when investigating latent
inhibition. Given the well-established role of 5-HT, and particularly 5-HT1A receptors,
in the processing of pain (Avila-Rojas et al., 2015; Colpaert, 2006; Colpaert et al.,
2002; Panczyk et al., 2015), I wanted an alternative paradigm to test for latent
inhibition. Latent inhibition can be readily demonstrated using the conditioned taste
aversion paradigm. Conditioned taste aversion refers to the phenomenon whereby an
unfamiliar taste (e.g. a new blend of coffee), paired with a negative internal state (e.g.
feeling sick after drinking), results in future avoidance of that taste (the new coffee).
This association is rapidly learned if the taste stimulus is novel, but this learning is
hampered if the taste stimulus has previously not predicted the illness (e.g. your usual
coffee blend). Thus, prior exposure to the neutral stimulus, without negative
consequence, inhibits the learning of an association, and so latent inhibition can be
demonstrated through an attenuated conditioned taste aversion. In rats this is typically
achieved by pairing flavoured water with a drug (e.g. lithium chloride) that produces
“internal malaise” (Lubow, 1989, p. 5). One pairing of the flavoured water with the
drug is sufficient to ensure the animal avoids the flavoured water in the future, but this
effect is attenuated if, previously, the flavoured water has been consumed without
consequence (Ellenbroek, Knobbout, & Cools, 1997; Mora et al., 1999). This
paradigm has been successfully used to investigate the effects of antipsychotic-type
drugs on latent inhibition (Geyer & Ellenbroek, 2003; Moser, Hitchcock, Lister, &
Moran, 2000).
As a first attempt to assess the roles of 5-HT1A and 5-HT1B receptors in latent
inhibition, the effect of the 5-HT1B/1A receptor agonist, RU 24969, on latent inhibition
in a conditioned taste aversion paradigm was tested.

41
Method
Procedure
Firstly, I aimed to establish the conditioned taste aversion effect, based on the
methods of Ellenbroek et al. (1997). All testing was conducted in the home cages.
Water bottles were removed from the home cages and made available for 30 minutes
per day. Rats (see general methods, n=8 per group) were randomly assigned to have
either water (water pre-exposure group) or a 5% sucrose solution (sucrose pre-
exposure group) available for drinking. Water bottles were weighed before and after
each 30 minute drinking period to measure consumption. Once total consumption
during this pre-exposure phase reached 40ml (approximately 3 days; Ellenbroek et al.
(1997)), rats in both groups received the 5% sucrose solution for 30 minutes.
Immediately after this 30 minute drinking period, lithium chloride (75 mg/kg, i.p.;
Ellenbroek et al. (1997)) was administered. The next day both water and the sucrose
solution were made available for 30 minutes. Taste aversion was measured as the
proportion of sucrose consumed on this test day (amount of sucrose solution consumed
divided by total fluid consumption), with lower proportions of sucrose consumption
indicative of greater taste aversion. Thus, latent inhibition was indicated by a lower
taste aversion (i.e. greater proportion of sucrose consumption) in the sucrose pre-
exposure group.
Other groups were tested to determine the effect of RU 24969 pretreatment on
this latent inhibition effect. The same protocol were used, but 15 minutes prior to water
bottles being available during the pre-exposure phase, rats were injected with RU
24969 (0, 0.03, 0.3, 3.0 mg/kg, s.c.). This range of RU 24969 doses has been shown to
be behaviourally effective in different paradigms (Kennett, Dourish, & Curzon, 1987;
Tricklebank, Middlemiss, & Neill, 1986). The 15 minute RU 24969 pretreatment time
is common (Acosta, Boynton, Kirschner, & Neisewander, 2005; P. J. Fletcher &
Korth, 1999b), because maximal effects have been shown between 15 minutes and 4
hours after administration (Tricklebank et al., 1986).
Statistical analyses
Fluid consumption was compared as a function of pre-exposure using one-way
analysis of variance (ANOVA). A 3 (RU 24969 dose) × 3 (Session) mixed model
ANOVA, with session as the within subjects factor, was used to analyse the effect of
RU 24969 on fluid consumption as a function of pre-exposure session. Where
appropriate, post-hoc analyses were conducted using Tukey’s HSD method.

42
Results
Experiment 1: Pilot study on the Conditioned Taste Aversion effect
Rats in both pre-exposure groups (n=8 per group) met the fluid consumption
criterion in 3 daily pre-exposure sessions. There was no difference in total fluid
consumption between the sucrose and water groups (F(1,14)=2.10, p=0.17). Figure 3.1
shows the water pre-exposure group demonstrated conditioned taste aversion, as
indicated by the low proportion of sucrose consumed on the test day. The sucrose pre-
exposure group showed significantly greater sucrose consumption than the water pre-
exposure group (F(1,14)=4.53, p=0.05, ɳp2= 0.25). Figure 3.1 shows that the sucrose
pre-exposure group consumed similar amounts of water and sucrose on the test day,
suggesting the internal malaise produced by lithium chloride was not associated with
the sucrose solution, thus illustrating latent inhibition.
Sucros
eWate
r0.0
0.2
0.4
0.6
0.8
Pre-exposure
Prop
ortio
n su
cros
e
*
Fig. 3.1 Conditioned taste aversion to sucrose after pairing with lithium chloride in rats either pre-‐exposed to sucrose or water. The lack of preference for water over sucrose in the sucrose pre-‐exposure group is indicative of latent inhibition. n = 8 per group, error bars represent SEM. *p=0.05 Experiment 2: The effect of RU 24969 on Conditioned Taste Aversion
Only 11 subjects were available at the beginning of this experiment, so they
were divided into groups that received different doses of RU 24969 (0.0, 0.03, 0.3, 3.0
mg/kg, s.c.) and different pre-exposures (water, sucrose; n=2-3 per group) with the
intention of adding more subjects to each group as they became available. However,

43
the initial groups treated with RU 24969 appeared to consume less fluid in the daily 30
minute sessions. Figure 3.2 shows the fluid consumption over the first 3 sessions
(collapsed across pre-exposure group) as a function of RU 24969 dose. It is clear that
higher RU 24969 dose groups initially consumed less fluid than the lower dose groups.
ANOVA confirmed a significant effect of RU 24969 dose (F(3,54)=25.8, p<0.001,
ɳp2=0.59), and a post-hoc Tukey test showed that the 3.0 and 0.3 mg/kg RU 24969
dose groups both consumed less fluid than the 0.0 and 0.03 mg/kg groups across the
first 3 sessions. This decrease in fluid consumption provided a confound that would
compromise interpretation of a conditioned taste aversion experiment. Therefore, no
further testing was conducted.
P re -e x p o s u r e s e s s io n
Da
ily
flu
id c
on
su
mp
tio
n
(ml)
1 2 30
5
10
15
20
0 .0
0 .0 3
0 .3
3 .0
R U 2 4 9 6 9 (m g /k g )
Fig 3.2 The effect of RU 24969 dose on the amount of fluid consumed by fluid-‐deprived rats during the first 3 drinking sessions. RU 24969 dose-‐dependently decreased fluid consumption, with the 0.3 and 3.0 mg/kg groups consuming significantly less fluid over the 3 sessions. n=4-‐6 per group, error bars represent SEM. Discussion
The latent inhibition effect was successfully produced using the conditioned
taste aversion paradigm. However, the impact of RU 24969 on this effect could not be
assessed because higher doses of RU 24969 reduced fluid consumption. These results
showed that the conditioned taste aversion paradigm was a confounded assay for
measuring behavioural responses to RU 24969 under these conditions.
Serendipitously, the results also suggested a more straightforward measure of
5-HT1B activation, that of reduced drinking, or adipsia. This response to RU 24969 had

44
been alluded to in the literature. For example, RU 24969 non-selectively reduced
intake of both water and sweetened ethanol (Silvestre, Palacios, Fernandez, & O'Neill,
1998), responding maintained by water in water-deprived rats (Carli, Invernizzi,
Cervo, & Samanin, 1988), and the time spent drinking sweetened condensed milk
(Simansky & Vaidya, 1990). To our knowledge there had not been any
pharmacological studies to determine whether this decrease in fluid consumption is
due to effects at 5-HT1A or 5-HT1B receptors. In many ways the adipsic response to RU
24969 would be a preferable behavioural response to measure, because only one drug
exposure is required, and because the effect can be assessed in a relatively short time
period. Thus, the next study aimed to determine the parameters of RU 24969-produced
adipsia, and the relative contribution of 5-HT1A and 5-HT1B receptor subtypes to this
effect.

45
Chapter 4: Behavioural responses to RU 24969
Parts of this chapter appear in:
Aronsen, Webster, & Schenk. (2014). RU 24969-produced adipsia and
hyperlocomotion: Differential role of 5-HT1A and 5-HT1B receptor
mechanisms. Pharmacology, Biochemistry and Behavior, 124, 1-4. DOI
10.1016/j.pbb.2014.05.008
Aronsen, Bukholt, & Schenk (2016). Repeated administration of the 5-HT1B/1A agonist,
RU 24969, facilitates the acquisition of MDMA self-administration: Role of 5-
HT1A and 5-HT1B receptor mechanisms. Psychopharmacology, 233 (8), 1339-
1347. DOI 10.1007/s00213-016-4225-x
The previous chapter showed that latent inhibition, assessed using the
conditioned taste aversion paradigm, is a confounded behavioural assay due to
decreased fluid consumption produced by RU 24969. This decrease in fluid
consumption might, however, be a novel response that could be used to characterise
RU 24969. This effect had been referred to in the literature, but no study had
determined the parameters of this adipsic response to RU 24969, or the contribution of
5-HT1A and 5-HT1B receptors. This was, therefore, one objective of this study.
RU 24969 also produces hyperlocomotion. In contrast to RU 24969-produced
adipsia, this behavioural response to RU 24969 has been well studied. RU 24969-
produced hyperactivity was not attenuated by depletion of brain 5-HT, suggesting a
post synaptic mechanism (Cheetham & Heal, 1993). Studies in mice have generally
attributed RU 24969-induced hyperlocomotion to 5-HT1B mechanisms because it was
selectively attenuated by pretreatment with 5-HT1B, but not 5-HT1A, receptor
antagonists (Cheetham & Heal, 1993; Shanahan et al., 2009). In the rat, however, there
is a lack of full parametric analysis of the roles of 5-HT1A or 5-HT1B activation in this
behavioural response. For example, the 5-HT1B/1D receptor antagonist, GR 127935,
dose-dependently attenuated the hyperactive response to RU 24969 in the Wistar-
Kyoto hyperactive rat, but a control strain was not assessed (Chaouloff, Courvoisier,
Moisan, & Mormede, 1999). Similarly, GR 127935 blocked the hyperactive response
to RU 24969 50-60 minutes after RU 24969 administration (O’Neill & Parameswaran,
1997), while the 5-HT1A receptor antagonists, WAY 100635 and SDX 216-525, but
not GR 127935, blocked the hyperactive response to RU 24969 in the first 15 minutes
after administration (Kalkman, 1995), before maximal effects of RU 24969 are

46
evident. Thus, roles of 5-HT1A and 5-HT1B receptors in RU 24969-produced
hyperlocomotion have been suggested, but the relative contribution of 5-HT1A and 5-
HT1B receptors over the course of RU 24969-produced hyperactivity is not clear.
These studies had 3 aims. Firstly, the adipsic and hyperactive responses to RU
24969 were characterised by administering a range of doses and measuring dose-
dependent behavioural responses. Secondly, the relative contributions of 5-HT1A and
5-HT1B receptors to these effects were determined by pretreating rats with a selective
5-HT1A or 5-HT1B receptor antagonist. Lastly, if a behavioural response to RU 24969
was antagonised by a 5-HT1A receptor antagonist the same response was to be tested
after administration of the selective 5-HT1A receptor agonist, 8-OH-DPAT. A more
selective, well characterised, 5-HT1B receptor agonist was not readily available to us at
the time of these experiments, but 8-OH-DPAT has been widely used as a selective 5-
HT1A receptor agonist. 8-OH-DPAT has approximately 7000 fold preference for 5-
HT1A receptors over 5-HT1B receptors (Hamon, Cossery, Spampinato, & Gozlan,
1986). Therefore, 8-OH-DPAT is a preferable ligand to use when measuring
behavioural responses to 5-HT1A receptor activation.
Method
Water consumption
Standard protocol was used (see General Methods). RU 24969 (0.0 – 3.0
mg/kg, s.c.; n = 10 per group) was administered 15 minutes before water bottles were
reintroduced. These data provided the dose of RU 24969 that was subsequently used in
the antagonist study. Separate groups (n=6-9 per group) were tested in the same
manner to assess the contribution of 5-HT1A or 5-HT1B mechanisms. Either the 5-HT1A
receptor antagonist, WAY 100635 (0.0, 1.0 mg/kg, s.c.), or the 5-HT1B/1D receptor
antagonist, GR 127935 (0.0, 3.0 mg/kg, s.c.) was administered 15 minutes before RU
24969 (1.0 mg/kg, s.c.). These doses were chosen for their documented efficacy in
blocking 5-HT1A and 5-HT1B effects, respectively (Acosta et al., 2005; P. J. Fletcher &
Korth, 1999b).
Locomotor Activity
Rats were placed in the testing chamber for 30 minutes, followed by an
injection of RU 24969 (0.0-3.0 mg/kg, s.c.; n=8 per group), and activity was measured
for 45 minutes post-injection. Separate groups (n=6-12 per group) were placed in the
activity monitoring chambers and 15 minutes later received either WAY 100635 (0.0,

47
1.0 mg/kg, s.c.) or GR 127935 (0.0, 3.0 mg/kg, s.c.), followed 15 minutes later by RU
24969 (3.0 mg/kg, s.c.). In order for the data to be directly comparable to the fluid
consumption protocol, only data collected from 15-45 minutes following the injection
of RU 24969 were analysed.
Separate rats were used to test the hyperactive response to 8-OH-DPAT. Rats
were placed in the testing chamber for 30 minutes, followed by an injection of 8-OH-
DPAT (0.0, 0.03, 0.1, 0.3, 1.0, 3.0 mg/kg, s.c., n=5-7 per group), and activity was
measured for 60 minutes post-injection.
8-OH-DPAT is a selective 5-HT1A receptor agonist but also has appreciable
affinity for 5-HT7 receptors (Bard et al., 1993; Lovenberg et al., 1993). To determine
whether 8-OH-DAT-produced hyperactivity was due to 5-HT1A activation we
determined the effect of the selective 5-HT1A receptor antagonist, WAY 100635, on 8-
OH-DPAT-produced hyperactivity. Rats were placed in the testing chamber and 15
minutes later were injected with WAY 100635 (0, 0.003, 0.3 mg/kg, s.c., n=4-5 per
group). Following a further 15 minutes, 8-OH-DPAT (0.3 mg/kg, s.c.) was injected,
and activity was measured for an additional 60 minutes.
Data analysis
The effect of RU 24969 on water consumption was assessed using a one-way
ANOVA. The effect of RU 24969 on locomotor activity was assessed using a 4 (RU
24969 dose) × 6 (Time after injection) mixed ANOVA with Time as the within
subjects factor. The effects of WAY 100635 and GR 127935 on RU 24969-proudced
adipsia or hyperlocomotion were assessed using separate 2 (antagonist dose) × 2 (RU
24969 dose) ANOVAs. The effect of 8-OH-DPAT on locomotor activity counts was
assessed using a one-way ANOVA. Data for 8-OH-DPAT-produced hyperactivity
after administration of WAY 100635 were analysed using a 3 (Dose) × 12 (Time after
injection) mixed model ANOVA with Time as the within subjects factor. Post-hoc
analyses were conducted using Tukey’s HSD method.

48
Results
0.0 0.03 0.3 1.0 3.0
0
5
10
15
RU 24969 (mg/kg)
Wat
er c
onsu
mpt
ion
(ml)
*
*
*
Fig. 4.1 Effect of RU 24969 on water consumption over 30 minutes in water deprived rats. n= 10 per group, error bars represent SEM. *-‐ p<0.05 compared to 0.0 mg/kg dose.
Figure 4.1 shows the effect of RU 24969 on water consumption. ANOVA
confirmed an effect of dose (F (4, 45) = 24.56, p<0.001, ɳp2= 0.69), and post hoc
Tukey analysis indicated that 0.3, 1.0, and 3.0 mg/kg RU 24969 significantly
decreased water consumption (p<0.05). Effects of the antagonists on RU 24969-
produced adipsia are presented in Figure 4.2
Fig. 4.2 Effect of the 5-‐HT1B/1D receptor antagonist, GR 127935 (left), or the 5-‐HT1A receptor antagonist, WAY 100635 (right), on RU 24969-‐produced adipsia. n=6-‐9 per group, error bars represent SEM. *-‐ p<0.05.
0 .0 1 .00
5
1 0
1 5
R U 2 4 9 6 9 (m g /k g )
Wa
ter
co
ns
um
pti
on
(m
l)
0 .0
1 .0
W A Y 1 0 0 6 3 5 (m g /k g )
0 .0 1 .00
5
1 0
1 5
R U 2 4 9 6 9 (m g /k g )
Wa
ter
co
ns
um
pti
on
(m
l)
0 .0
3 .0
G R 1 2 7 9 3 5 (m g /k g )
*

49
Analysis of the effect of WAY 100635 (dose RU 24969 × dose WAY 100635)
revealed a main effect of RU 24969 (F (1,26) = 26.95, p<.001, ɳp2= 0.51), but no effect
of WAY 100635 (F (1,26) = 0.016, ns) or an interaction (F (1,26) = 0.83, ns). In
contrast, analysis of the effect of GR 127935 (dose RU 24969 × dose GR 127935)
revealed an effect of GR 127935 (F (1,24) = 4.55, p=0.043, ɳp2= 0.16), an effect of RU
24969 (F (1,24) = 29.44, p<0.001 ɳp2= 0.55) and an interaction (F (1,24) = 9.02, p =
0.006 ɳp2= 0.27). Tukey post hoc comparisons confirmed that GR 127935 significantly
reduced RU 24969-produced adipsia (p<0.05).
Figure 4.3 shows that RU 24969 increased locomotor activity (F (3,28) = 8.15,
p<0.001 ɳp2= 0.47). There was no effect of Time (F (5,140) = 0.27, ns) and no
interaction (F (15,140) = 0.45, ns). Post hoc Tukey analysis showed the dose of 3.0
mg/kg was the only dose that significantly increased total forward locomotion.
Fig. 4.3 Effect of RU 24969 on locomotor activity. n=8 per group, error bars represent SEM. *-‐ p<0.05.
Effects of the antagonists on RU 24969-produced hyperlocomotion are
presented in figure 4.4. GR 127935 failed to alter RU 24969-produced hyperactivity;
the effect of GR 127935 (F (1,26) = 0.75, ns) and the interaction (F (1,26) = 0.52, ns)
between the two drugs were not significant. A significant effect of WAY 100635 was
found (F (1,36) = 6.73, p = 0.014, ɳp2= 0.16), and an interaction between WAY
100635 and RU 24969 treatment was significant (F (1,36) = 4.44, p = 0.042, ɳp2=
0.11). Tukey post hoc comparisons confirmed that WAY 100635 significantly reduced
RU 24969-produced hyperactivity (p<0.05).
-3 5 -3 0 -2 5 -2 0 -1 5 -1 0 -5 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0 4 50
2 0 0
4 0 0
6 0 0
8 0 0
1 0 0 0
1 2 0 0
T im e (m in )
Am
bu
lato
ry c
ou
nts
0 .0
0 .3
1 .0
3 .0
R U 2 4 9 6 9 (m g /k g )
R U 2 4 9 6 90 .0 0 .3 1 .0 3 .0
0
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
R U 2 4 9 6 9 (m g /k g )
To
tal a
mb
ula
tory
co
un
ts
*

50
Fig. 4.4 Effect of the 5-‐HT1B/1D receptor antagonist, GR 127935 (left), or the 5-‐HT1A receptor antagonist, WAY 100635 (right), on RU 24969-‐produced hyperactivity. n=6-‐12 per group, error bars represent SEM. *-‐ p<0.05.
8-OH-DPAT dose-dependently increased locomotor activity counts
(F(5,33)=48.63, p<0.001, ɳp2= 0.88). Post hoc analysis revealed that doses of 0.3, 1.0,
and 3.0 mg/kg 8-OH-DPAT significantly increased locomotor activity counts (see fig
4.5).
0 .00 .0
30 .1 0 .3 1 .0 3 .0
0
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
8 -O H -D P A T (m g /k g )
To
tal
am
bu
lato
ry c
ou
nts
**
*
Fig. 4.5 The hyperactive response to 8-‐OH-‐DPAT. n=5-‐7 per group, error bars represent SEM. *-‐ p<0.05 compared to 0.0 mg/kg group.
Figure 4.6 (left panel) shows the time course of the effects of WAY 100635 on
8-OH-DPAT-produced hyperactivity. ANOVA showed a significant interaction
0 .0 3 .00
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
2 5 0 0
3 0 0 0
3 5 0 0
R U 2 4 9 6 9 (m g /k g )
To
tal
am
bu
lato
ry c
ou
nts
0 .0
1 .0
G R 1 2 7 9 3 5 (m g /k g )
0 .0 3 .00
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
2 5 0 0
3 0 0 0
3 5 0 0
R U 2 4 9 6 9 (m g /k g )
To
tal
am
bu
lato
ry c
ou
nts
0 .0
1 .0
W A Y 1 0 0 6 3 5 (m g /k g )*
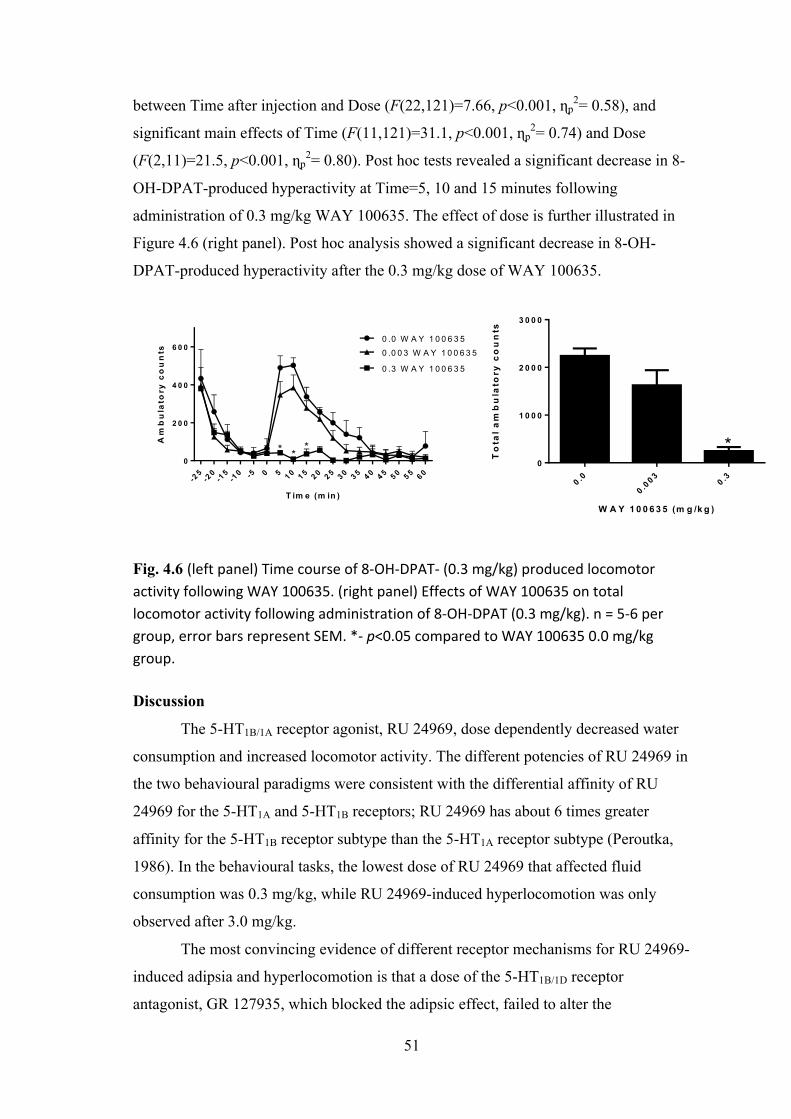
51
between Time after injection and Dose (F(22,121)=7.66, p<0.001, ɳp2= 0.58), and
significant main effects of Time (F(11,121)=31.1, p<0.001, ɳp2= 0.74) and Dose
(F(2,11)=21.5, p<0.001, ɳp2= 0.80). Post hoc tests revealed a significant decrease in 8-
OH-DPAT-produced hyperactivity at Time=5, 10 and 15 minutes following
administration of 0.3 mg/kg WAY 100635. The effect of dose is further illustrated in
Figure 4.6 (right panel). Post hoc analysis showed a significant decrease in 8-OH-
DPAT-produced hyperactivity after the 0.3 mg/kg dose of WAY 100635.
-25
-20
-15
-10 -5 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0 4 5 5 0 5 5 6 0
0
2 0 0
4 0 0
6 0 0
T im e (m in )
Am
bu
lato
ry c
ou
nts
0 .0 W A Y 1 0 0 6 3 5
0 .0 0 3 W A Y 1 0 0 6 3 5
0 .3 W A Y 1 0 0 6 3 5
* **
0 .0
0 .00 3
0 .30
1 0 0 0
2 0 0 0
3 0 0 0
W A Y 1 0 0 6 3 5 (m g /k g )T
ota
l a
mb
ula
tory
co
un
ts
*
Fig. 4.6 (left panel) Time course of 8-‐OH-‐DPAT-‐ (0.3 mg/kg) produced locomotor activity following WAY 100635. (right panel) Effects of WAY 100635 on total locomotor activity following administration of 8-‐OH-‐DPAT (0.3 mg/kg). n = 5-‐6 per group, error bars represent SEM. *-‐ p<0.05 compared to WAY 100635 0.0 mg/kg group. Discussion
The 5-HT1B/1A receptor agonist, RU 24969, dose dependently decreased water
consumption and increased locomotor activity. The different potencies of RU 24969 in
the two behavioural paradigms were consistent with the differential affinity of RU
24969 for the 5-HT1A and 5-HT1B receptors; RU 24969 has about 6 times greater
affinity for the 5-HT1B receptor subtype than the 5-HT1A receptor subtype (Peroutka,
1986). In the behavioural tasks, the lowest dose of RU 24969 that affected fluid
consumption was 0.3 mg/kg, while RU 24969-induced hyperlocomotion was only
observed after 3.0 mg/kg.
The most convincing evidence of different receptor mechanisms for RU 24969-
induced adipsia and hyperlocomotion is that a dose of the 5-HT1B/1D receptor
antagonist, GR 127935, which blocked the adipsic effect, failed to alter the

52
hyperlocomotion effect of RU 24969. Further, a dose of the 5-HT1A receptor
antagonist, WAY 100635, which blocked the locomotor activating effects failed to
alter the adipsic response to RU 24969. The failure of these doses of WAY 100635 and
GR 127935 to alter one behaviour cannot be due to ineffective dosing since the other
behavioural effect of RU 24969 was attenuated by the same dose of the antagonist. It
might be argued that the decrease in drinking reflects the hyperactive response to RU
24969 that might have interfered with the ability to remain at the drinking spout. This
is unlikely since the reduction of fluid consumption was produced by doses of RU
24969 lower than those that increased locomotor activity.
Rather, the data are consistent with the idea that RU 24969-induced adipsia in
rats is mediated by 5-HT1B, but not 5-HT1A, mechanisms, and that RU 24969-induced
hyperactivity in rats is mediated by 5-HT1A, but not 5-HT1B, mechanisms. Another
study (Chaouloff et al., 1999) showed that GR 127935 attenuated RU 24969-induced
hyperactivity in Wistar-Kyoto hyperactive rats. This effect might have been non-
selective since GR 127935 in that study also decreased basal activity levels. We failed
to observe either of these effects in Sprague-Dawley rats, raising the possibility that
there are strain differences in the response to the antagonist. Another study (O’Neill &
Parameswaran, 1997) also showed that RU 24969-induced hyperactivity was
decreased by GR 127935, but this effect was produced 50-60 minutes after RU 24969
administration. This finding raises the possibility that there is an effect of GR 127935
that emerges at time points later than those tested in the present study. In accordance
with our conclusion that RU 24969-induced hyperlocomotion is due to this agonist’s
affinity for the 5-HT1A receptor, 5-HT1A receptor agonists are known to produce
hyperlocomotion (Kalkman & Soar, 1990; Tricklebank, Forler, & Fozard, 1984).
GR 127935 has affinity for a number of serotonin receptors (Centurión et al.,
2000; Price et al., 1997; Watson, Burton, Price, Jones, & Middlemiss, 1996), but it is
noteworthy that the 5-HT1B/1D receptor antagonist is at least 60 times more selective
for the 5-HT1B receptor than any of the other receptors that RU 24969 has notable
affinity for. Therefore, the most likely explanation for the reversal of RU 24969-
induced adipsia by GR 127935 is antagonism of the 5-HT1B receptor.
These results, along with others from the literature, raised the possibility that a
5-HT1A receptor agonist would also produce reliable hyperlocomotion. Given the high
selectivity of 8-OH-DPAT for the 5-HT1A receptor, as well as the low affinity for the
5-HT1B receptor (Hamon et al., 1986; Peroutka, 1986), 8-OH-DPAT-produced

53
hyperactivity would be a preferable behavioural measure of 5-HT1A activation. Thus,
the last study in this chapter aimed to determine the parameters under which 8-OH-
DPAT produces hyperactivity, and the role of 5-HT1A receptor activation in this effect.
8-OH-DPAT dose dependently increased locomotor activity, with a maximal
effect around 1.0 mg/kg. This hyperactive response to the 5-HT1A receptor agonist was
reversed by the 5-HT1A receptor antagonist, WAY 100635. It is unlikely that this
reversal by WAY 100635 was due to a non-specific decrease in locomotor activity
because the higher (1.0 mg/kg) dose of WAY 100635 used in the previous experiment
had no significant effect on locomotor activity. Therefore, these results suggest that 8-
OH-DPAT-produced hyperactivity is due to 5-HT1A receptor activation.
This result is in accordance with other studies that have investigated the
hyperlocomotor response to 8-OH-DPAT. Hyperactivity produced by 8-OH-DPAT
was attenuated by the 5-HT1 receptor antagonist, pindolol (Ahlenius & Salmi, 1995;
Hillegaart, Estival, & Ahlenius, 1996), suggesting a 5-HT1A receptor mechanism. 8-
OH-DPAT produced hyperlocomotion was not attenuated by depletion of monoamines
via reserpine treatment, suggesting this behavioural response to 8-OH-DPAT is not
due to alterations in synthesis and/or release of 5-HT via autoreceptor-mediated
effects, but instead action on post-synaptic 5-HT1A receptors (Ahlenius & Salmi, 1995;
Mignon & Wolf, 2002).
Together, these data show that adipsia and hyperlocomotion provide
dissociable behavioural measures of RU 24969 that are produced by 5-HT1B and 5-
HT1A activation, respectively. Furthermore, 8-OH-DPAT-produced hyperactivity may
be a preferable measure of 5-HT1A activation, because of the selectivity of 8-OH-
DPAT for the 5-HT1A receptor. Because RU 24969-produced adipsia and 8-OH-
DPAT-produced hyperactivity are selective responses to 5-HT1B and 5-HT1A receptor
activation, respectively, these procedures provide straight-forward assays of 5-HT1A
and 5-HT1B receptor function, and so will be used in the following chapters.

54
Chapter 5: Effects of repeated administration of the 5-HT1B/1A receptor agonist,
RU 24969, on the acquisition of MDMA self-administration
Parts of this chapter appear in:
Aronsen, Bukholt, & Schenk (2016). Repeated administration of the 5-HT1B/1A agonist,
RU 24969, facilitates the acquisition of MDMA self-administration: Role of 5-
HT1A and 5-HT1B receptor mechanisms. Psychopharmacology, 233 (8), 1339-
1347. DOI 10.1007/s00213-016-4225-x
As was explained in the General Introduction, self-administration of a range of
substances is inhibited by increased synaptic 5-HT (Loh & Roberts, 1990; Ritz &
Kuhar, 1989; Rothman et al., 2005; Z. Wang & Woolverton, 2007; Wee et al., 2005).
A recent study (Bradbury et al., 2014) tested the idea that MDMA-produced 5-HT
release might be inhibitory to MDMA self-administration and attempted to explain
both the long latency to acquisition, and the small proportion of rats that meet
acquisition criteria. The MDMA-produced increase in synaptic 5-HT was measured by
in vivo microdialysis before MDMA self-administration began. As has been observed
in many studies from the Schenk lab (Colussi-Mas et al., 2010; Schenk et al., 2012;
Schenk et al., 2003; Schenk et al., 2007), about 50% of the rats acquired MDMA self-
administration. Of interest, MDMA-stimulated 5-HT release was lower for the rats that
ultimately met the acquisition criteria, suggesting an inhibitory role of MDMA-
produced 5-HT release on the acquisition of MDMA self-administration. This idea was
experimentally tested by determining the effect of a neurotoxic, 5,7-DHT, lesion on
MDMA self-administration. The lesion reduced 5-HT levels by up to 67%, and greatly
facilitated the acquisition of MDMA self-administration; while approximately 50% of
control rats met acquisition criteria, 100% of the lesion group acquired. Furthermore,
of the control group that acquired, 50% met the criterion within 14 sessions, while only
6 sessions were required for 50% of the lesion group to meet the criterion.
These findings strengthen the idea that variability in the acquisition of MDMA
self-administration is due to variability in sensitivity to MDMA-produced 5-HT
release. Specifically, 5-HT has an inhibitory impact on MDMA self-administration. A
question remains as to the mechanism for this inhibitory effect of 5-HT on the
acquisition of MDMA self-administration. One possibility is that high levels of
synaptic 5-HT produced by MDMA during initial self-administration sessions led to

55
neuroadaptive changes in 5-HT receptor mechanisms that modulate responses
associated with the acquisition of self-administration.
As outlined in the General Introduction, 5-HT1A and 5-HT1B receptors regulate
dopaminergic neurotransmission. Because self-administration is associated with
increased dopamine neurotransmission, activation of 5-HT1A and 5-HT1B receptors
might be expected to impact self-administration. Of particular interest, activation of 5-
HT1A receptors attenuated amphetamine-induced increases in extracellular dopamine
levels (Ichikawa et al., 1995; Kuroki et al., 1996) and, as would therefore be expected,
a range of 5-HT1A receptor agonists have been shown to inhibit self-administration
(Müller et al., 2007). On the other hand, 5-HT1B receptor agonists potentiated the
increase in extracellular dopamine produced by cocaine or ethanol (O'Dell & Parsons,
2004; Parsons et al., 1999; Yan et al., 2005), and generally enhanced self-
administration, producing leftward shifts in the self-administration dose-response
curves for cocaine and GBR 12909 (Parsons et al., 1996, 1998; Pentkowski et al.,
2009; Przegaliñski et al., 2007).
A wealth of data indicate a role of 5-HT1A and 5-HT1B receptor subtypes in the
maintenance of self-administration (P. J. Fletcher, Azampanah, & Korth, 2002;
Neisewander, Cheung, & Pentkowski, 2014; Parsons et al., 1998; Peltier & Schenk,
1993; Przegaliñski et al., 2007) but the role in the acquisition of self-administration has
received far less attention. Given that self-administration is driven by increases in
dopamine neurotransmission, and that the acquisition of MDMA self-administration
was enhanced by a neurotoxic, 5,7-DHT lesion, I wanted to determine whether this
facilitation of MDMA self-administration was due to decreased activation of 5-HT1A
or 5-HT1B receptors. If so, it should be possible to manipulate receptor mechanisms via
repeated agonist or antagonist exposure and to determine the effect on acquisition of
MDMA self-administration.
Tolerance to RU 24969-produced hyperactivity was produced following
repeated exposure to the 5-HT1B/1A receptor agonist (Oberlander, Demassey, Verdu,
Van de Velde, & Bardelay, 1987). As outlined in the previous chapter, we have
recently shown that RU 24969-produced hyperactivity in rats is due to activation of 5-
HT1A, but not 5-HT1B, receptors (Aronsen, Webster, & Schenk, 2014), suggesting that
behavioural tolerance reflects a down-regulation of this receptor subtype. The effect of
RU 24969 pretreatment on 5-HT1B receptor mechanisms has not been specifically
measured, but RU 24969-produced adipsia provides a means of addressing this

56
question (Aronsen et al., 2014). Therefore, in the present study we determined the
effect of repeated exposure to RU 24969 on the acquisition of MDMA self-
administration, and on RU 24969-produced adipsia. In order to assess the effect on 5-
HT1A receptor mechanisms we also measured hyperactivity in response to the selective
5-HT1A receptor agonist, 8-OH-DPAT.
Method
Male Sprague-Dawley rats (see General Methods) were used. For rats that
underwent self-administration testing, an intravenous catheter was implanted, as
outlined in the General Methods.
RU 24969 pretreatment
RU 24969 pretreatment began once pre-surgery weight had been obtained. RU
24969 (3.0 mg/kg, s.c.), or the saline vehicle (1.0 ml/kg), was administered in the
home cage daily at 0900hr and 1600hr, for three consecutive days. This protocol was
adapted from that used in earlier studies (Callaway & Geyer, 1992; Oberlander et al.,
1987) to utilise a dose of RU 24969 that we have previously shown produces both
hyperactivity and adipsia (Aronsen et al., 2014).
Acquisition of MDMA self-administration
Self-administration sessions began the day after the last administration of RU
24969. Self-administration was conducted during 2 hour daily sessions, 6 days per
week. Each self-administration session began with an experimenter-delivered infusion
of drug to clear the line of heparinised saline solution. Thereafter, depression of the
active lever produced an infusion of MDMA (1.0 mg/kg/infusion) according to an FR1
schedule. Responses on the active and inactive levers were recorded. Every seventh
day catheters were infused with sodium pentobarbital (20.0 mg/kg, i.v.). Failure to
demonstrate an immediate loss of the righting reflex suggested a loss of catheter
patency and the rat was excluded from the study. Catheter patency was lost in 4 rats (3
RU 24969 pretreated, 1 saline pretreated), and 3 rats in the RU 24969 pretreatment
group self-administered lethal doses of MDMA, resulting in final sample sizes of 9 and
8 for the RU 24969 and saline pretreated groups, respectively. Self-administration
testing continued for each rat until a total of 90 infusions (90.0 mg/kg) had been self-
administered, or for 25 days, whichever came first. This acquisition criterion is the
same as has been used previously in our laboratory (Bradbury et al., 2014; Oakly,
Brox, Schenk, & Ellenbroek, 2014).

57
Water consumption and locomotor activity
Separate groups of rats were tested to determine the effects of RU 24969
pretreatment on RU 24969-produced adipsia, or 8-OH-DPAT-produced hyperactivity.
The standard water consumption protocol was used (see General Methods), with water
bottles removed the day after the last RU 24969 pretreatment administration. RU
24969 (0.0, 1.0, 3.0 mg/kg, s.c., n= 6-8 per group) was administered 15 minutes before
water bottles were reintroduced. These doses were chosen based on our previous study
(Aronsen et al., 2014) that suggested that adipsia following administration of these
doses of RU 24969 was due to 5-HT1B receptor activation.
The effect of the selective 5-HT1A receptor agonist, 8-OH-DPAT, on locomotor
activity was assessed 2 days after the last administration of RU 24969, in order to
match the delay between pretreatment and the test for RU 24969-induced adipsia. Rats
were placed in the testing chamber (see General Methods) for 30 minutes, followed by
an injection of 8-OH-DPAT (0.0, 0.1, 0.3 mg/kg, s.c., n=4-7 per group), and activity
was measured for 60 minutes post-injection.
To investigate the possibility that RU 24969 pretreatment affected
dopaminergic mechanisms, locomotor activity produced by the dopamine releasing
agent, d-amphetamine, was assessed 2 days after the last administration of RU 24969.
Rats pretreated with either RU 24969, or vehicle, were placed in the testing chamber
(see General Methods) for 30 minutes, followed by an injection of d-amphetamine (0.5
mg/kg, i.p., n=10 per group), and activity was measured for 60 minutes post-injection.
This dose was chosen because it has previously been used in our laboratory to illustrate
dopaminergic sensitisation (Bradbury, Gittings, & Schenk, 2012).
Data analysis
Acquisition of self-administration was compared between pretreatment groups
with a survival analysis, using the log-rank test to compare Kaplan-Meier survival
estimates (Kaplan & Meier, 1958). Right-censoring was applied to data from rats that
did not acquire within the 25 day cut-off period.
RU 24969-produced adipsia was analysed with a 2 (Pretreatment) × 3 (Dose of
RU 24969) ANOVA. Effects of each dose of 8-OH-DPAT on locomotor activity were
analysed by individual 2 (Pretreatment) × 12 (Time after injection) mixed model
ANOVAs with Time as the within subjects factor. Total activity counts as a function
of Dose of 8-OH-DPAT and RU 24969 pretreatment were analysed using a 2
(Pretreatment) × 3 (8-OH-DPAT Dose) ANOVA. The locomotor responses to d-

58
amphetamine was analysed with a 2 (pretreatment) × 12 (Time after injection) mixed
model ANOVA with Time as the within subjects factor.
Results
Figure 5.1 shows survival curves for the acquisition of self-administration for
saline- or RU 24969-treated groups. RU 24969 pretreatment produced a significant
increase in the probability of acquiring MDMA self-administration (χ² (1) = 12.21,
p<0.01). Of the control group that met the acquisition criterion, 50% met the criterion
within 17 sessions, whereas 50% of RU 24969 pretreatment group met the acquisition
criterion within 10 sessions. It is noteworthy that three rats in the RU 24969
pretreatment group self-administered lethal doses of MDMA (>20 mg/kg) during the
first self-administration session and therefore additional data from these rats could not
be obtained. The high intake during the first self-administration session for these 3 rats
supports the other data suggesting RU 24969 pretreatment enhanced the initial
reinforcing effects of MDMA.
0 5 10 15 20 250
25
50
75
100
Self-administration session
Perc
ent a
cqui
red
Saline PretreatmentRU 24969 pretreatment
Fig. 5.1 Cumulative percentage of rats that met the criterion for acquisition of MDMA self-‐administration in the RU 24969 (squares, n=9) and saline (circles, n=8) pretreatment groups.
Figure 5.2 (left panel) shows the effect of RU 24969 pretreatment on RU
24969-produced adipsia. There was a significant interaction between Pretreatment and
Dose (F(2,37)=7.85, p=0.01, ɳp2= 0.30) and a significant effect of Dose
(F(2,37)=53.55, p<0.01, ɳp2= 0.74). Post hoc tests confirmed a significant difference in

59
the adipsic response between the RU 24969 and saline pretreatment groups following
0.0 mg/kg, and 3.0 mg/kg RU 24969. Since there was a decrease in basal water
consumption produced by repeated RU 24969 treatment, the data were further
analysed by expressing drug effects as a percentage of baseline. These data are
presented in Figure 5.2 (right panel). A 2×2 (Pretreatment × Dose) ANOVA revealed a
significant effect of Pretreatment (F(1,27)=20.40, p<0.01, ɳp2= 0.43).
0.0
1.0 3.00
5
10
15
RU 24969 (mg/kg)
Wat
er c
onsu
mpt
ion
(ml)
Saline PretreatmentRU 24969 pretreatment
*
*
1.0 3.00
20
40
60
80
100
RU 24969 (mg/kg)
Wat
er c
onsu
mpt
ion
- P
erce
ntag
e of
bas
elin
e
Fig. 5.2 (Left panel) The adipsic response to RU 24969 after repeated exposure to RU 24969 (grey bars) or saline (black bars). (Right panel) Percentage of baseline water intake as a function of RU 24969 dose for RU 24969 and saline pretreated groups. n=6-‐8 per group. Figures represent the mean + SEM. * -‐ p<0.05.
Locomotor activity produced by the various doses of 8-OH-DPAT as a
function of RU 24969 pretreatment is shown in Figure 5.3. There were no differences
between groups following the 0.0 mg/kg 8-OH-DPAT dose. The data from 0.1 mg/kg
8-OH-DPAT dose produced a Time × Pretreatment interaction (F(11,110)=4.06,
p<0.01, ɳp2= 0.29) and main effects of Time (F(11,110)=19.8, p<0.01, ɳp
2= 0.66) and
Pretreatment (F(1,10)=17.6, p<0.01, ɳp2= 0.64). Post hoc tests revealed significant
decreases in activity during Time=10 and 15 minutes following the injection. There
was a significant Time × Pretreatment interaction (F(11,121)=2.77, p<0.01, ɳp2= 0.20)
and main effects of Time (F(11,121)=62.5, p<0.01, ɳp2= 0.85) and Pretreatment
(F(1,11)=7.45, p<0.05, ɳp2= 0.40) for the 0.3 mg/kg 8-OH-DPAT groups. Post-hoc
tests revealed a significant decrease in activity at Time=25 minutes. Analysis of total
activity counts as a function of Dose and Pretreatment showed a main effect of 8-OH-
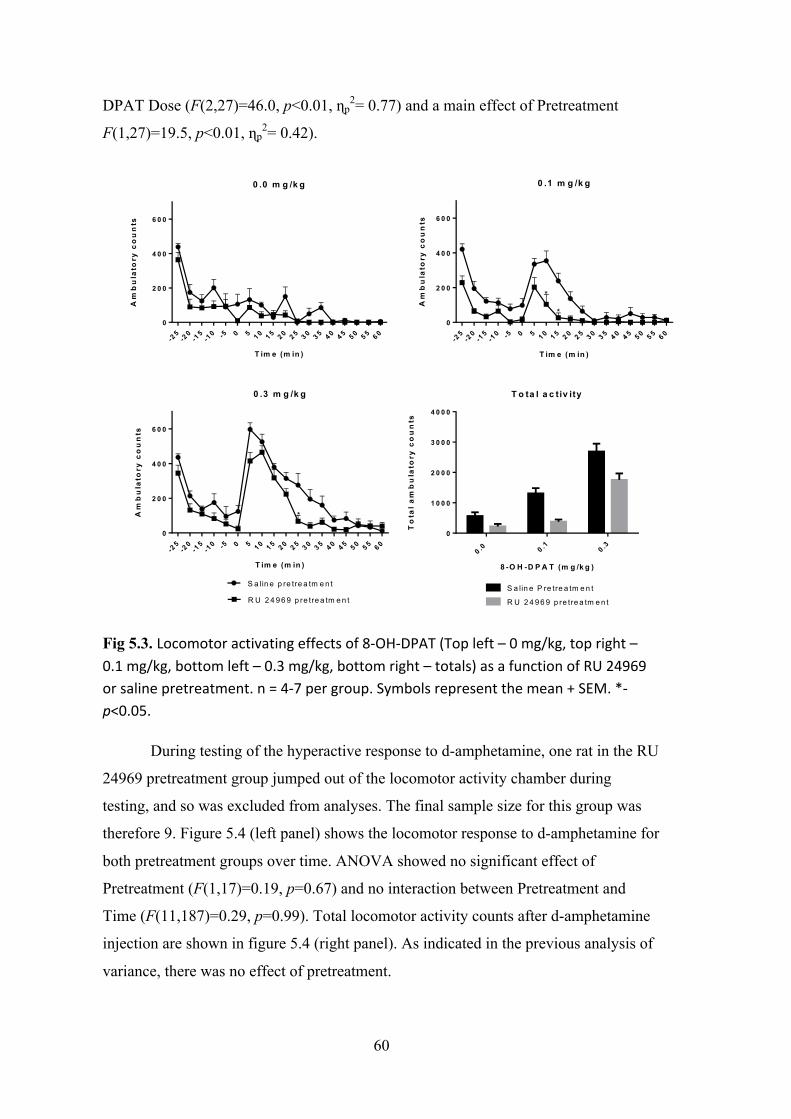
60
DPAT Dose (F(2,27)=46.0, p<0.01, ɳp2= 0.77) and a main effect of Pretreatment
F(1,27)=19.5, p<0.01, ɳp2= 0.42).
-25
-20
-15
-10 -5 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0 4 5 5 0 5 5 6 0
0
2 0 0
4 0 0
6 0 0
0 .0 m g /k g
T im e (m in )
Am
bu
lato
ry c
ou
nts
- 25
-20
-15
-10 -5 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0 4 5 5 0 5 5 6 0
0
2 0 0
4 0 0
6 0 0
0 .1 m g /k g
T im e (m in )
Am
bu
lato
ry c
ou
nts
*
*
-25
-20
-15
-10 -5 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0 4 5 5 0 5 5 6 0
0
2 0 0
4 0 0
6 0 0
0 .3 m g /k g
T im e (m in )
Am
bu
lato
ry c
ou
nts
S a lin e p re tre a tm e n t
R U 2 4 9 6 9 p re tre a tm e n t
*
0 .0
0 .1 0 .30
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
T o ta l a c tiv ity
8 -O H -D P A T (m g /k g )
To
tal
am
bu
lato
ry c
ou
nts
S a lin e P re tre a tm e n t
R U 2 4 9 6 9 p re tre a tm e n t
Fig 5.3. Locomotor activating effects of 8-‐OH-‐DPAT (Top left – 0 mg/kg, top right – 0.1 mg/kg, bottom left – 0.3 mg/kg, bottom right – totals) as a function of RU 24969 or saline pretreatment. n = 4-‐7 per group. Symbols represent the mean + SEM. *-‐ p<0.05.
During testing of the hyperactive response to d-amphetamine, one rat in the RU
24969 pretreatment group jumped out of the locomotor activity chamber during
testing, and so was excluded from analyses. The final sample size for this group was
therefore 9. Figure 5.4 (left panel) shows the locomotor response to d-amphetamine for
both pretreatment groups over time. ANOVA showed no significant effect of
Pretreatment (F(1,17)=0.19, p=0.67) and no interaction between Pretreatment and
Time (F(11,187)=0.29, p=0.99). Total locomotor activity counts after d-amphetamine
injection are shown in figure 5.4 (right panel). As indicated in the previous analysis of
variance, there was no effect of pretreatment.

61
T im e (m in )
Am
bu
lato
ry c
ou
nts
- 25
-20
-15
-10 -5 0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0 4 5 5 0 5 5 6 0
0
2 0 0
4 0 0
6 0 0
S a lin e P re tre a tm e n t
R U 2 4 9 6 9 p re tre a tm e n t
P re tre a tm e n t
To
tal
am
bu
lato
ry c
ou
nts
S a line
R U 24 9 6 9
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
Fig. 5.4 The locomotor response to d-‐amphetamine (0.5 mg/kg) after pretreatment with either RU 24969 or saline. n=9-‐10 per group. Discussion
Pretreatment with RU 24969 decreased the latency to acquisition of MDMA
self-administration, and increased the proportion of rats that acquired MDMA self-
administration. The leftward shift in the acquisition curve for self-administration might
reflect a sensitised reinforcing effect since higher doses of drug have also been shown
to decrease the latency to acquisition of self-administration (Carroll & Lac, 1997;
Schenk & Partridge, 2000).
A remarkable consequence of pretreatment with RU 24969 was the substantial
increase in the proportion of rats that met the criterion for acquisition of MDMA self-
administration. As we have previously reported (Bradbury et al., 2014; Schenk et al.,
2012), 50% of control rats met the criterion within the 25 day cut-off period. Thus,
some rats appear to be inherently more or less sensitive to the reinforcing effects of
MDMA. Following RU 24969 pretreatment, however, all of the rats met the criterion
for acquisition of MDMA self-administration within the limits of the study (25 test
sessions). We have suggested that the initial resistance to self-administration can be
overcome by limiting the impact of 5-HT since a similar increase in the percentage of
subjects that acquired MDMA self-administration was produced following neurotoxic
5,7-DHT lesions in rats (Bradbury et al., 2014) and in 5-HT transporter knock-out rats
(Oakly et al., 2014).
In order to assess the impact of more specific 5-HT mechanisms on the
acquisition of MDMA self-administration, the present study repeatedly administered
the 5-HT1B/1A receptor agonist, RU 24969, as a pretreatment in an attempt to down-

62
regulate 5-HT1A and 5-HT1B receptors. We determined effects of the pretreatment by
measuring behavioural responses that have been attributed to either 5-HT1B (RU
24969-produced adipsia (Aronsen et al., 2014)) or 5-HT1A (8-OH-DPAT-produced
hyperactivity (Hillegaart et al., 1996)) mechanisms.
As previously reported (Aronsen et al., 2014), RU 24969 produced dose-
dependent adipsia. The dose-response curve for this response is relatively narrow;
minimal effects were produced following administration of 0.3 mg/kg and maximal
effects were produced following administration of 3.0 mg/kg (Aronsen et al., 2014).
RU 24969 pretreatment decreased basal water consumption and when this was
accounted for, RU 24969 pretreatment decreased the subsequent RU 24969-produced
adipsic response. These findings are consistent with a rightward shift in the dose-
response curve and suggest a down-regulation of 5-HT1B receptors. 5-HT1B receptor
down-regulation has previously been evidenced by decreased mRNA levels
(Chennaoui et al., 2001; Hiroi & Neumaier, 2009) or decreased binding density
(Kindlundh et al., 2003; Suzuki, Han, & Lucas, 2010), both of which could explain the
present behavioural data.
RU 24969 pretreatment also shifted the dose-response curve for 8-OH-DPAT-
produced hyperactivity to the right; the most pronounced effect of pretreatment was on
hyperactivity produced by the lowest does of 8-OH-DPAT tested. This might explain
why a similar pretreatment with RU 24969 failed to alter hyperactivity produced by a
higher dose of 1.25 mg/kg 8-OH-DPAT (Oberlander et al., 1987).
Although 8-OH-DPAT has appreciable affinity for the 5-HT7 receptor (Bard et
al., 1993; Lovenberg et al., 1993), results from the previous chapter showed that
hyperactivity produced by 8-OH-DPAT was attenuated by the selective receptor
antagonist, WAY 100635, confirming a 5-HT1A receptor mechanism. Of interest, a
similar RU 24969 pretreatment regimen also reduced the locomotor response to RU
24969 (Callaway & Geyer, 1992), a behavioural response that we have attributed to 5-
HT1A receptor activation (Aronsen et al., 2014). Therefore, these findings are
consistent with a down-regulation of 5-HT1A receptors following RU 24969
pretreatment. 5-HT1A down-regulation has been shown via decreased agonist-
stimulated binding of [35S]GTPγS to G proteins (Fuss et al., 2013; Hensler, Vogt, &
Gass, 2010), decreased receptor binding densities or immunoreactivity (Fuss et al.,
2013; Gui et al., 2011), decreased 5-HT1A mRNA (S. Wang et al., 2009), and

63
decreased protein levels (Iyo et al., 2009; S. Wang et al., 2009). It would be of great
interest to determine which, if any, of these mechanisms can explain the present data.
The available literature is consistent with the idea that MDMA self-
administration, like self-administration of other drugs of abuse, progresses as a result
of sensitised dopamine and desensitised 5-HT responses. Thus, repeated exposure to
MDMA increased dopamine (Colussi-Mas et al., 2010; Kalivas, Duffy, & White,
1998) and decreased 5-HT (Baumann, Clark, Franken, et al., 2008; Reveron et al.,
2010; Shankaran & Gudelsky, 1999) synaptic output, as measured by in vivo
microdialysis, dopamine antagonists reduced MDMA self-administration (Brennan,
Carati, Lea, Fitzmaurice, & Schenk, 2009; Daniela, Brennan, Gittings, Hely, &
Schenk, 2004), and dopamine, but not 5-HT, agonists potentiated drug-seeking
following extinction of MDMA self-administration (Schenk, Gittings, & Colussi-Mas,
2011).
MDMA preferentially releases 5-HT and the ensuing activation of post-
synaptic receptors impacts dopamine release, providing potential mechanisms for the
enhanced dopamine response. In this study, both 5-HT1B and 5-HT1A receptor
mechanisms were down regulated, as measured by behavioural assays. Given the
selectivity of RU 24969 for 5-HT1A/1B receptors it is unlikely that alterations in a
different receptor mechanism underlies the facilitated acquisition of self-administration
found in the present study.
Because activation of 5-HT1B receptors enhanced extracellular dopamine
concentrations (Galloway et al., 1993; Hållbus et al., 1997; Iyer & Bradberry, 1996;
O'Dell & Parsons, 2004; Yan & Yan, 2001a; Yan et al., 2004) it is possible that
repeated administration of RU 24969 sensitised dopamine neurons independently of
the effect on 5-HT1A and 5-HT1B receptors. A sensitised dopamine response to MDMA
would be expected to facilitate the acquisition of MDMA self-administration. This
seems unlikely, however, because RU 24969 pretreatment had no effect on
amphetamine-produced hyperactivity. Although the amphetamine dose was chosen
based on other sensitisation studies, it is possible that a sensitised dopamine response
would have been observed if higher doses of amphetamine had been tested.
Activation of 5-HT1B receptors enhanced basal dopamine neurotransmission
(Alex & Pehek, 2007) and the dopaminergic response to drugs of abuse (O'Dell &
Parsons, 2004; Parsons et al., 1999; Yan et al., 2005), so the down-regulation of these
receptor mechanisms, which would be expected to decrease MDMA-produced

64
dopamine, cannot easily explain the facilitated self-administration. On the other hand,
a wealth of data suggest that activation of 5-HT1A receptors is inhibitory to cocaine
self-administration (Müller et al., 2007), possibly via inhibition of dopamine release
(Ichikawa & Meltzer, 2000). Therefore, a down-regulation of this receptor subtype
might be expected to disinhibit MDMA-produced dopamine, leading to more rapid
acquisition of self-administration due to increased reinforcing effects. This might also
explain the facilitated acquisition of MDMA self-administration in serotonin
transporter knockout rats (Oakly et al., 2014), since this manipulation also desensitised
5-HT1A receptor mechanisms (Homberg et al., 2008).
5-HT1A receptors are widely localised in brain and are well-positioned to
modulate activity in a large number of brain systems (Aznar, Qian, Shah, Rahbek, &
Knudsen, 2003). Of importance, these receptors are localised on tyrosine hydroxylase
immunoreactive cells in the VTA (Doherty & Pickel, 2001) and also in dopamine
terminal regions in the NAc (Alex & Pehek, 2007). Systemic administration of 8-OH-
DPAT inhibited amphetamine-produced dopamine release in the NAc (Ichikawa et al.,
1995). The down-regulation produced by RU 24969 pretreatment would, therefore, be
expected to disinhibit stimulated dopamine. Similar studies have not been conducted
using MDMA, but this mechanism could explain the facilitated acquisition of self-
administration.
The acquisition of self-administration is also influenced by factors in addition
to the initial reinforcing effects of the drug and some of these factors are modified by
5-HT1A receptor mechanisms. As explained in the General Introduction, increased
impulsivity, anxiety, or learning, could be expected to facilitate the acquisition of
MDMA self-administration.
5-HT1A activation increased behavioural measures of impulsivity (Carli &
Samanin, 2000). However, individual variability in impulsivity did not predict latency
to acquisition of MDMA self-administration (Bird & Schenk, 2013). This might be
because the impulsive response to 5-HT1A receptor agonists is due to autoreceptor
activation (Carli et al., 2006; Lladó-‐Pelfort et al., 2010; Winstanley et al., 2003). We
have previously shown that repeated exposure to MDMA failed to alter 5-HT1A
autoreceptor mechanisms (Schenk et al., 2013). Therefore, alterations in 5-HT1A-
mediated impulsivity are unlikely to have impacted the present results.
A down-regulation of 5-HT1A receptors would be expected to reduce anxiety
produced by MDMA, because 5-HT1A receptor activation is anxiogenic (Cheeta et al.,

65
2000a, 2000b; File et al., 1996; File et al., 2000; Solati et al., 2011). However, higher
levels of anxiety have been associated with self-administration (Dilleen et al., 2012;
Homberg et al., 2002; Spanagel et al., 1995). Therefore an attenuation of 5-HT1A
receptor-produced anxiety would not explain the facilitated acquisition of MDMA self-
administration.
Reliable self-administration is often facilitated via Pavlovian conditioning
processes by pairing delivery of the drug reinforcer with a discrete, discriminative
stimulus, like a light, as was done in the present study (Di Ciano & Everitt, 2004). As
explained in the General Introduction, strengthening of stimulus/reward associations is
markedly inhibited by administration of 5-HT1A receptor agonists (Blair et al., 2004;
Frick et al., 2015; Winsauer et al., 1999). These findings raise the possibility that
activation of post synaptic 5-HT1A receptors pursuant to MDMA-stimulated 5-HT
release limits the acquisition of MDMA self-administration, in some subjects, by
interfering with associative learning. If so, our data suggest that this effect is mitigated
by exposure to a regimen of RU 24969 pretreatment that down-regulated these
receptor mechanisms, thereby facilitating MDMA self-administration as indicated by
both a leftward and upward shift in the self-administration acquisition curves. This
idea could be tested by administering the same RU 24969 pretreatment as was used in
this study and assessing learning in a stimulus/reward association task. If RU 24969
pretreatment facilitated learning in such a task it would strengthen the claim that the
facilitation of MDMA self-administration seen in the present study was associated
with enhanced learning.

66
Chapter 6: Predicting the acquisition of MDMA self-administration
In the previous chapter, a manipulation that down-regulated 5-HT1A and 5-
HT1B receptors also greatly facilitated the acquisition of MDMA self-administration.
As outlined in the discussion, it is possible that a down-regulation of these receptors
could explain the facilitated acquisition. However, the correlational nature of that
study makes it impossible to ascertain the role of alterations in 5-HT1A and/or 5-HT1B
receptor mechanisms in the acquisition of MDMA self-administration.
There is substantial evidence that the magnitude of MDMA-produced 5-HT
release predicts the latency to acquire MDMA self-administration. A question remains
as to what the mechanism underlying this effect might be. The results from the
previous chapter raise the possibility that this mechanism involves individual
variability in 5-HT1A and/or 5-HT1B receptor-mediated effects.
Some evidence suggests that activation of these receptor subtypes modulates
dopamine neurotransmission, providing a potential mechanism. For example, the 5-
HT1A receptor gonist, 8-OH-DPAT, inhibited amphetamine-produced increases in
extracellular dopamine (Ichikawa et al., 1995; Kuroki et al., 1996). Thus, activation of
5-HT1A receptors by MDMA-stimulated 5-HT would be expected to inhibit dopamine
release. If so, this might explain why MDMA is, at least initially, not a very
efficacious reinforcer. Activation of 5-HT1A receptors also impaired learning in a range
of operant tasks (Blair et al., 2004; Frick et al., 2015; Meneses, 2007; Winsauer et al.,
1999). As a result, 5-HT1A activation during MDMA self-administration could inhibit
learning processes associated with the acquisition of self-administration. Therefore,
subjects with higher sensitivity to 5-HT1A receptor activation may be less likely to
acquire MDMA self-administration, due to inhibition of dopamine release, and/or
impaired ability to learn the operant task. The observation that RU 24969 pretreatment
enhanced MDMA self-administration and down-regulated 5-HT1A receptors is
consistent with this idea.
5-HT1B receptor activation, on the other hand, augmented the increases in
extracellular dopamine produced by cocaine (O'Dell & Parsons, 2004; Parsons et al.,
1999) or ethanol (Yan et al., 2005). Thus, the down-regulation of 5-HT1B receptors
produced by RU 24969 pretreatment might be expected to reduce the reinforcing
efficacy of MDMA. Furthermore, intra-raphe injections of the neurotoxin, 5,7-DHT,
increased 5-HT1B receptor binding in the substantia nigra and NAc (Compan, Segu,

67
Buhot, & Daszuta, 1998), and intraventricular infusion of 5,7-DHT produced an
increase in 5-HT1B binding in the hypothalamus, entorhinal cortex, and substantia
nigra (Manrique et al., 1998; Manrique et al., 1994; Manrique, Segu, Hery, Faudon, &
François-Bellan, 1993; Weissmann, Mach, Oberlander, Demassey, & Pujol, 1986).
This same lesion facilitated MDMA self-administration (Bradbury et al., 2014). In
contrast, intraventricular infusion of 5,7-DHT had no impact on 5-HT1A binding in
substantia nigra, PFC, hippocampus, hypothalamus, or amygdala (Hensler, Kovachich,
& Frazer, 1991; Lawrence, Olverman, Shirakawa, Kelly, & Butcher, 1993; Weissmann
et al., 1986). Thus, the role of 5-HT1B receptor populations in MDMA self-
administration is not clear. Acquisition was facilitated by separate manipulations that
produced both an up- and down-regulation of 5-HT1B receptors, respectively. It is
therefore possible that the acquisition of MDMA self-administration is not related to 5-
HT1B receptor populations.
The purpose of the following studies was to determine whether individual
variability in 5-HT1A and/or 5-HT1B receptors predicted the latency to acquisition of
MDMA self-administration. To this end, the behavioural responses to the 5-HT1A
receptor agonist, 8-OH-DPAT, or the 5-HT1B/1A receptor agonist, RU 24969, were
assessed before MDMA self-administration commenced. Furthermore, to test the idea
that RU 24969 pretreatment facilitated acquisition of MDMA self-administration via a
down-regulation of 5-HT1A receptors, separate groups of rats were administered the 5-
HT1A receptor antagonist, WAY 100635, or vehicle, before each self-administration
session. If 5-HT1A receptor activation does inhibit MDMA self-administration,
pretreatment with WAY 100635 would be expected to facilitate acquisition.
Method
Subjects and procedures
Male Sprague-Dawley rats underwent catheter surgery for self-administration,
as outlined in the General Methods section. Testing began after recovery to pre-
surgery weight.
The hyperlocomotor response to 8-OH-DPAT was assessed using the standard
locomotor activity methods outlined in the General Methods section. Rats were placed
in the activity chambers for 30 minutes, followed by 8-OH-DPAT (0.1, 0.3 mg/kg, s.c.,
n=24 and 30 respectively) administration, and locomotor activity was measured for
another 60 minutes. These doses were chosen because inspection of preliminary data

68
showed both doses produced hyperactivity with considerable between-subject
variability.
Adipsia produced by RU 24969 was assessed using the standard water
consumption methods outlined in the General Methods section. RU 24969 (1.0 mg/kg,
s.c., n=13) was administered 15 minutes before water bottles were reintroduced. This
dose was chosen because in previous studies it produced an adipsic response with
considerable between-subject variability (Aronsen et al., 2014).
MDMA self-administration, as outlined in the General Methods, began the day
after the behavioural response to 8-OH-DPAT or RU 24969 was measured. Self-
administration was conducted during 2 hour daily sessions, 6 days per week. Each self-
administration session began with an experimenter-delivered infusion of drug.
Thereafter, depression of the active lever produced an infusion of MDMA according to
an FR1 schedule. Responses on the active and inactive levers were recorded. Every
seventh day catheters were infused with sodium pentobarbital (20.0 mg/kg, i.v.).
Failure to demonstrate an immediate loss of the righting reflex suggested a loss of
catheter patency and the rat was excluded from the study. Catheter patency was lost in
5 rats (4 after 8-OH-DPAT-produced hyperactivity (3 in the 0.1 mg/kg group, 1 in the
0.3 mg/kg group), 1 after RU 24969-produced adipsia), and one rat self-administered a
lethal dose of MDMA on the first day (0.3 mg/kg 8-OH-DPAT group). The same
acquisition criterion was used as in the last chapter – a total of 90 infusions (90 mg/kg)
self-administered. In order to minimise the number of subjects required, testing
continued for 35 sessions in the groups assigned to 0.3 mg/kg 8-OH-DPAT and 1.0
mg/kg RU 24969. Testing in the 0.1 mg/kg 8-OH-DPAT group continued for 25
sessions, whereupon subjects were used for a different study.
Using the same self-administration procedure, separate groups of rats were
pretreated with either saline vehicle, or the 5-HT1A receptor antagonist, WAY 100635
(1.0 mg/kg, s.c., n=7 per group) 15 minutes before each daily self-administration
session. Of the 14 rats that started self-administration, 4 were removed from the
experiment due to loss of catheter patency (3 in the vehicle group, 1 in the WAY
100635 group).
Statistical analyses
Behavioural responses to either 8-OH-DPAT or RU 24969 were correlated
with latency to acquisition of MDMA self-administration using a Pearson's product-
moment correlation. Data from subjects that did not acquire within the cut off period

69
were not included in these analyses. Analysis of the effect of WAY 100635
pretreatment on self-administration was not possible due to a high attrition rate, but
raw data are presented.
Results
Out of a total of 67 rats that started the locomotor and adipsia studies, 39 met
the acquisition criterion within 25 sessions, and a further 9 met the criterion between
26 and 35 sessions.
Figure 6.1 shows the distribution of days to meet the acquisition criterion and
locomotor response to 0.1 mg/kg 8-OH-DPAT. There was no significant correlation
between these two variables (r(16)=-0.21, p=0.40).
Fig 6.1 Scatterplot of days to acquire MDMA self-‐administration (y-‐axis) and locomotor response to 0.1 mg/kg 8-‐OH-‐DPAT (x-‐axis).
Similarly, there was no correlation between days to acquisition and locomotor
response to 0.3 mg/kg 8-OH-DPAT (r(18)=0.004, p=0.99) (see figure 6.2).
L o c o m o to r a c t iv ity c o u n ts
Da
ys
to
ac
qu
isit
ion
0 5 0 0 1 0 0 0 1 5 0 00
1 0
2 0
3 0
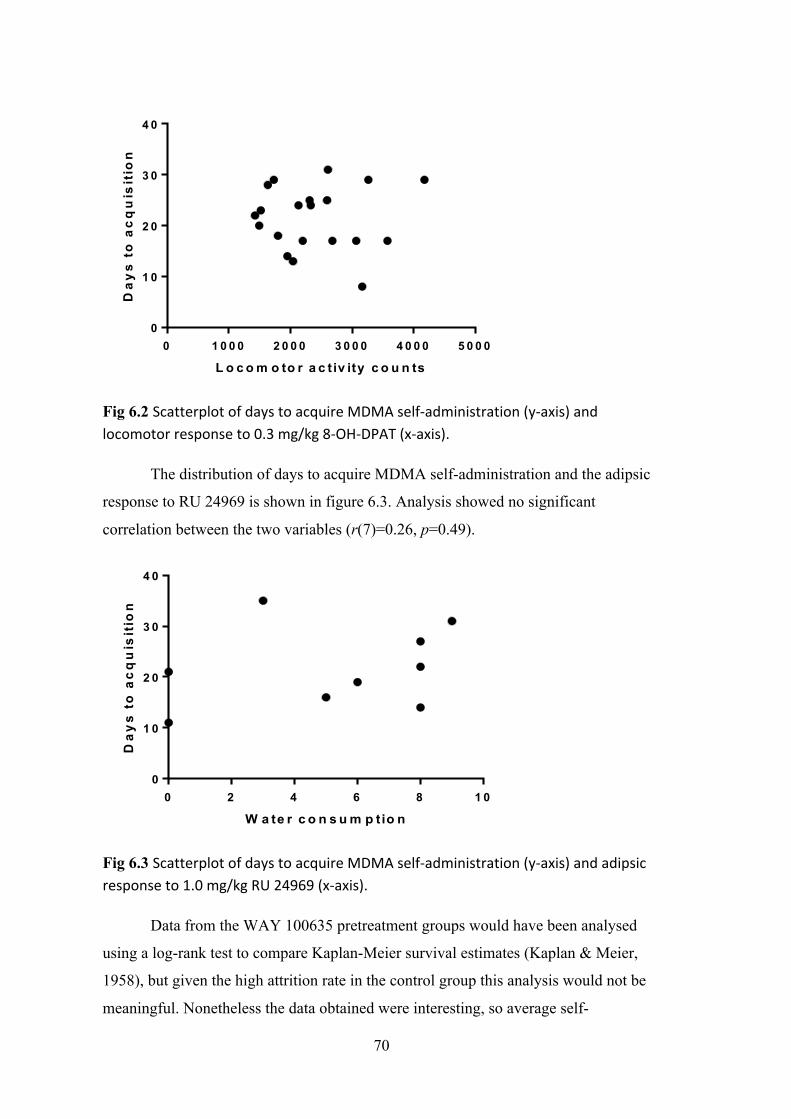
70
L o c o m o to r a c t iv ity c o u n ts
Da
ys
to
ac
qu
isit
ion
0 1 0 0 0 2 0 0 0 3 0 0 0 4 0 0 0 5 0 0 00
1 0
2 0
3 0
4 0
Fig 6.2 Scatterplot of days to acquire MDMA self-‐administration (y-‐axis) and locomotor response to 0.3 mg/kg 8-‐OH-‐DPAT (x-‐axis). The distribution of days to acquire MDMA self-administration and the adipsic
response to RU 24969 is shown in figure 6.3. Analysis showed no significant
correlation between the two variables (r(7)=0.26, p=0.49).
W a te r c o n s u m p tio n
Da
ys
to
ac
qu
isit
ion
0 2 4 6 8 1 00
1 0
2 0
3 0
4 0
Fig 6.3 Scatterplot of days to acquire MDMA self-‐administration (y-‐axis) and adipsic response to 1.0 mg/kg RU 24969 (x-‐axis). Data from the WAY 100635 pretreatment groups would have been analysed
using a log-rank test to compare Kaplan-Meier survival estimates (Kaplan & Meier,
1958), but given the high attrition rate in the control group this analysis would not be
meaningful. Nonetheless the data obtained were interesting, so average self-
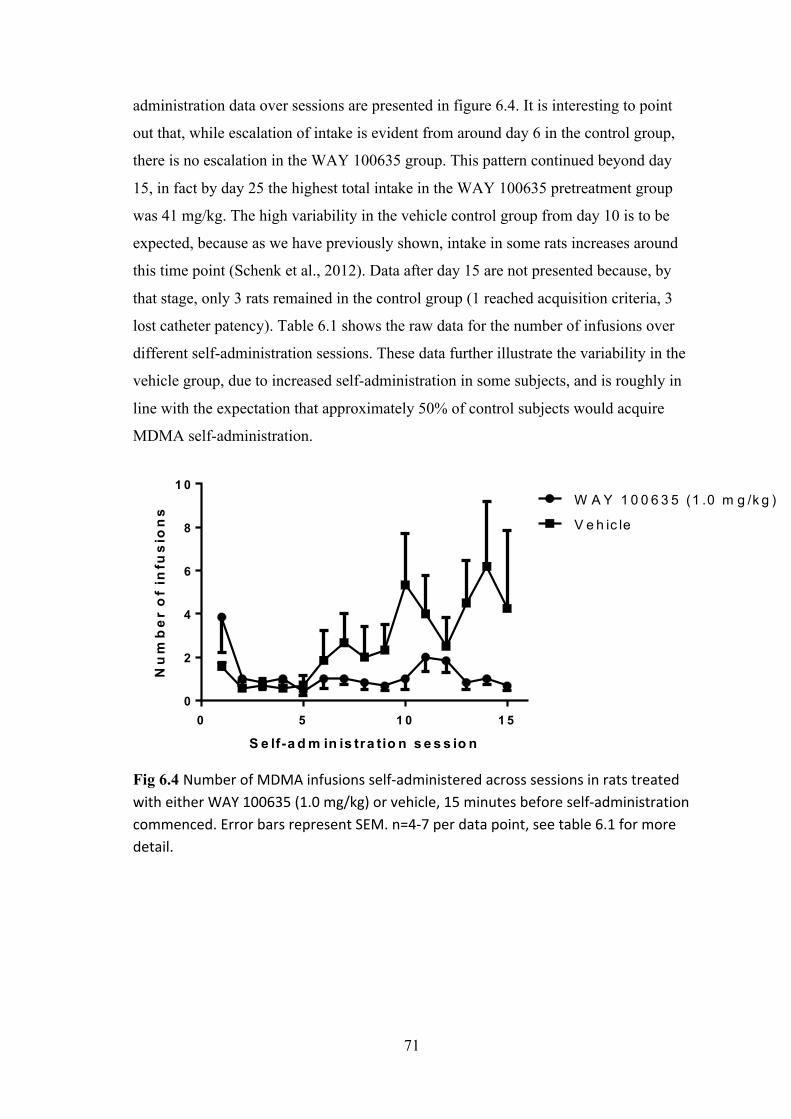
71
administration data over sessions are presented in figure 6.4. It is interesting to point
out that, while escalation of intake is evident from around day 6 in the control group,
there is no escalation in the WAY 100635 group. This pattern continued beyond day
15, in fact by day 25 the highest total intake in the WAY 100635 pretreatment group
was 41 mg/kg. The high variability in the vehicle control group from day 10 is to be
expected, because as we have previously shown, intake in some rats increases around
this time point (Schenk et al., 2012). Data after day 15 are not presented because, by
that stage, only 3 rats remained in the control group (1 reached acquisition criteria, 3
lost catheter patency). Table 6.1 shows the raw data for the number of infusions over
different self-administration sessions. These data further illustrate the variability in the
vehicle group, due to increased self-administration in some subjects, and is roughly in
line with the expectation that approximately 50% of control subjects would acquire
MDMA self-administration.
S e lf-a d m in is tra tio n s e s s io n
Nu
mb
er
of
infu
sio
ns
0 5 1 0 1 50
2
4
6
8
1 0W A Y 1 0 0 6 3 5 (1 .0 m g /k g )
V e h ic le
Fig 6.4 Number of MDMA infusions self-‐administered across sessions in rats treated with either WAY 100635 (1.0 mg/kg) or vehicle, 15 minutes before self-‐administration commenced. Error bars represent SEM. n=4-‐7 per data point, see table 6.1 for more detail.

72
Group Rat
Session 1 Session 5 Session 10 Session 15
WAY 100635
Pūkeko
1 0 removed removed
Rock Wren 4 0 3 1
Saddleback 2 0 0 1
Silvereye
12 0 0 1
Spotless Crake 0 1 0 0
Spotted Shag 7 1 1 0
Stichbird
1 1 2 1
Vehicle
Oystercatcher 1 0 14 removed
Takahē
2 1 2 2
Tomtit
2 1 11 15
Tūī
1 0 4 removed
Weka
1 0 0 0
White Heron 2 3 removed removed
Yellowhead 2 0 1 0
Table 6.1 The number of MDMA infusions (1.0 mg/kg/infusion) self-‐administered by subjects treated with either the 5-‐HT1A receptor antagonist, WAY 100635, or saline vehicle, 15 minutes before each self-‐administration session.
Discussion
These studies failed to show an association between behavioural response to 5-
HT1A or 5-HT1B activation, and latency to acquire MDMA self-administration. These
results were surprising given that, in the previous chapter, a treatment that down-
regulated both receptor subtypes also facilitated acquisition of MDMA self-
administration. Furthermore, lower sensitivity of 5-HT1A, or greater sensitivity of 5-
HT1B, receptors would be expected to enhance the dopaminergic response to MDMA,
which would be expected to enhance self-administration. Thus, it appears that basal
variability in 5-HT1A and 5-HT1B receptors is not associated with the variability in
acquisition of MDMA self-administration.
These results might suggest that facilitated self-administration after RU 24969
pretreatment reported in the previous chapter was not due to the effects of the
pretreatment on 5-HT1A or 5-HT1B receptors. Indeed, a significant correlation between
behavioural response and latency to acquisition in the present studies would have been
evidence for a role of 5-HT1A and/or 5-HT1B receptors in the initial reinforcing effects
of MDMA. However, caution should be exercised before we conclude that the effect

73
of RU 24969 pretreatment on MDMA self-administration was independent of the
effects on 5-HT1A or 5-HT1B receptors. Firstly, RU 24969 is reasonably selective for
these two receptor subtypes, making a non-selective effect less likely. Furthermore, it
is possible that the natural variability in the behaviours measured in this study was not
substantial enough to show an effect. For example, the mean activity count after 0.1
mg/kg 8-OH-DPAT in the present study was 1003 (SD=348), but after RU 24969
pretreatment this mean was 378 (SD=164). Thus, it is possible that lower sensitivity to
8-OH-DPAT is indeed predictive of latency to acquire MDMA self-administration, but
that significantly lower levels of sensitivity are required.
Further study is required to determine the relative roles of 5-HT1A and 5-HT1B
receptors in the facilitation of MDMA self-administration after RU 24969
pretreatment. Pretreatment with RU 24969 and either the 5-HT1A receptor antagonist,
WAY 100635, or the 5-HT1B receptor antagonist, GR 127935, would help to clarify
the roles of each receptor.
There was an unfortunately high attrition rate in the WAY 100635 pretreatment
study. This attrition rate likely reflects a procedural problem during catheter surgery
that has since been identified. Measures have now been put in place to avoid such
levels of attrition in future. The subsequently small size of the saline pretreatment
group precludes meaningful comparisons between the WAY 100635 pretreatment
group and its appropriate control. However, our laboratory, and this thesis, have shown
that approximately 50% of rats acquire MDMA self-administration (Bradbury et al.,
2014; Schenk et al., 2012), and it seems unlikely that saline administration would alter
this acquisition rate. Therefore, it becomes interesting that responding for MDMA was
so low in the WAY 100635 group. If these findings were replicated in a larger sample,
and with an appropriate control, the data would provide evidence for the suggestion
that 5-HT1A receptor activation is required for the development of MDMA self-
administration.
If so, it would be difficult to reconcile these data with the RU 24969
pretreatment data that showed a down-regulation of 5-HT1A receptors was associated
with enhanced MDMA self-administration. Given the limited scope of the present
study, it is not possible to rule out a non-specific effect of WAY 100635. Data from
chapter 4 suggest that this dose of WAY 100635 does not supress locomotor
responding. Furthermore, data from the first self-administration session, and other
research from our lab (Schenk et al., Under Review), show that rats are able to perform

74
an operant response after acute WAY 100635 administration. Thus it seems unlikely
that the low levels of responding after WAY 100635 administration were due to motor
effects. There were no differences between the weights of subjects in the two groups
throughout the experiment (data not shown), suggesting there was no effect of WAY
100635 on eating or drinking.
It might be expected that repeated administration of an antagonist would
upregulate receptor populations. Indeed, administration of a high dose (3 mg/kg) of
WAY 100635 twice per day for 3 days increased 5-HT1A immunoreactivity in the
hippocampus and cortex (Abbas, Nogueira, & Azmitia, 2007). An up-regulation of 5-
HT1A receptors might inhibit self-administration via enhanced inhibition of dopamine.
Alternatively, WAY 100635 may have protected 5-HT1A receptors from important
neuroadaptations in response to high levels of 5-HT. During self-administration
session 1 I noticed that the rats in the WAY 100635 group that self-administered a
large dose of MDMA did not show the characteristic set of symptoms (hyperthermia,
wetness, ‘eagle-fear’, bleeding nose) typically associated with initial self-
administration of high doses. Anecdotally, repeated self-administration of high doses
of MDMA produces tolerance to these effects. Thus, WAY 100635 may have been
preventing neuroadaptations that produce tolerance to some of the aversive effects of
MDMA. Clearly, more research would be required to determine if this is the case.
The results of the present study failed to show an association between basal
responses to 5-HT1A or 5-HT1B receptor activation and latency to acquire MDMA self-
administration. However, the results of the previous chapter, and inferences drawn
from the WAY 100635 study in this chapter, raise the possibility that neuroadaptations
in 5-HT1A and/or 5-HT1B receptors are important for the progression of MDMA self-
administration. As outlined above, basal variability in these receptor subtypes may not
be substantial enough to allow for meaningful analysis. On the other hand, if changes
in these receptor subtypes underlie the development of MDMA as an efficacious
reinforcer, it might be possible to detect differences in these receptor populations after
substantial MDMA self-administration. This possibility will be addressed in the next
chapter.

75
Chapter 7: Response to 5-HT1A and 5-HT1B receptor agonists after self-
administration
Parts of this chapter have been adapted from:
Aronsen & Schenk (2016). MDMA self-administration fails to alter the response to 5-
HT1A and 5-HT1B agonists. Psychopharmacology, 233 (7), 1323-1330. DOI
10.1007/s00213-016-4226-9
with permission from the publisher (Appendix A).
Some users regularly consume large quantities of ecstasy (Cottler et al., 2001;
Degenhardt, Barker, & Topp, 2004; Topp, Hall, & Hando, 1997), and repeated ecstasy
use produces a range of negative consequences, including cognitive and emotional
deficits. While these deficits are worrisome in and of themselves, it has been suggested
that they could also facilitate further ecstasy taking, and thus contribute to the
development of an SUD (Schenk, 2009; Schenk & Aronsen, 2015). The mechanisms
underlying these deficits are not, however, well understood.
Ecstasy users showed deficits in learning (Wagner et al., 2013), and in attention
and memory (McCann, Mertl, et al., 1999) compared to ecstasy-naïve controls or those
with limited ecstasy use. Ecstasy users reported higher levels of depression,
impulsiveness, and sleep disturbances than poly-drug users who did not use ecstasy
(Taurah et al., 2014). These cognitive and behavioural deficits were persistent,
suggesting that regular ecstasy use may cause long-lasting neuroadaptations (Parrott,
2013b).
Animal studies have shown that a number of these adverse effects associated
with ecstasy use are modulated by pharmacological manipulation of 5-HT receptors.
For example, the 5-HT1A receptor agonist, 8-OH-DPAT, impaired learning and
memory in water maze (Carli & Samanin, 1992), passive avoidance (Carli, Tranchina,
& Samanin, 1992), and conditioned reinforcement (Meneses, 2007) tasks, while the 5-
HT1A receptor antagonist, WAY 101405, improved learning in the Morris water maze
(Hirst et al., 2008). 5-HT1A receptor agonists and antagonists also altered performance
in the forced swim test and conditioned stress-induced ultrasonic vocalisations (Assié
et al., 2010; Lucki, Singh, & Kreiss, 1994) and altered sleep and wakefulness, as
measured by EEG and EMG (Monti & Jantos, 1992; Monti et al., 1990). Activation of
5-HT1A receptors increased impulsive responding on the five-choice serial reaction

76
time task (Carli & Samanin, 2000), while the 5-HT1A receptor antagonist, WAY
100635, supressed impulsive action (Ohmura et al., 2013).
Pharmacological manipulation of the 5-HT1B receptor subtype also affected
learning and memory as measured by a conditioned reinforcement task (Meneses,
2001, 2007), altered EEG and EMG recordings of sleep and wakefulness (Bjorvatn &
Ursin, 1994; Monti, Monti, Jantos, & Ponzoni, 1995), and affected immobility time in
the forced swim test (Dawson et al., 2006; Tatarczynska, Klodzinska, Stachowicz, &
Chojnacka-Wojcik, 2004). Therefore, it is possible that some of the cognitive and
behavioural deficits that accompany ecstasy use might be due to MDMA-produced
neuroadaptations in these receptor mechanisms.
A small number of studies have assessed the effects of repeated exposure to
MDMA on 5-HT1A and 5-HT1B receptor mechanisms. Repeated experimenter-
administered MDMA reduced 5-HT1A binding in the dorsal raphe, suggesting a down-
regulation of 5-HT1A autoreceptors, and increased 5-HT1A binding in the frontal cortex,
suggesting an up-regulation of 5-HT1A heteroreceptors (Aguirre, Ballaz, Lasheras, &
Del Rio, 1998; Aguirre, Frechilla, García-‐Osta, Lasheras, & Del RIo, 1997; Aguirre,
Galbete, Lasheras, & Del Río, 1995). These effects were only produced following
exposure to high doses (2x20-30 mg/kg/day, 4 consecutive days); exposure to lower
doses (4x5 mg/kg/day, 2 consecutive days (McGregor et al., 2003)), or intermittent
doses (2x10mg/kg/day, every 5th day (Piper, Vu, Safain, Oliver, & Meyer, 2006)) of
MDMA failed to alter cortical or subcortical 5-HT1A densities. Repeated
administration of racemic MDMA increased 5-HT1B receptor mRNA (Kindlundh-
Högberg, Svenningsson, & Schiöth, 2006), and receptor binding densities were
increased in some brain regions, but decreased in others, after repeated MDMA
administration (McGregor et al., 2003). Repeated administration of (+) MDMA,
however, failed to produce persistent changes in 5-HT1B mRNA or 5-HT1B receptor
binding (Sexton, McEvoy, & Neumaier, 1999).
Functional evidence for these receptor changes is equivocal. Repeated
administration of MDMA attenuated the autoreceptor-mediated decrease in 5-HT
release produced by the 5-HT1A receptor agonist, F13640, in mice (Lanteri et al.,
2014). Repeated administration of MDMA did not, however, alter 8-OH-DPAT-
produced lower lip retraction or hypolocomotion, behaviours associated with 5-HT1A
autoreceptor activation (Schenk et al., 2013). On the other hand, 8-OH-DPAT-
produced hypothermia was increased after repeated MDMA administration in one

77
study (Aguirre et al., 1998) but unchanged in others (McNamara, Kelly, & Leonard,
1995; Mechan, O'Shea, Elliott, Colado, & Green, 2001; Piper et al., 2006). MDMA
pretreatment also attenuated the 8-OH-DPAT-produced 5-HT syndrome (Piper et al.,
2006) and forepaw treading (Granoff & Ashby, 2001), but had no effect on the
prosocial response (Thompson, Callaghan, Hunt, & McGregor, 2008), or the
hyperactive response (Granoff & Ashby, 2001) to 8-OH-DPAT. Differences might be
due to a number of paradigmatic variables including dosing regimen and subject
sample.
The hyperactive response to the 5-HT1B/1A receptor agonist, RU 24969, was
decreased after repeated administration of racemic MDMA (Callaway & Geyer, 1992),
but enhanced after repeated administration of the (+) MDMA isomer (McCreary,
Bankson, & Cunningham, 1999). It was suggested that this behavioural response to
RU 24969 reflected 5-HT1B receptor activation (Callaway & Geyer, 1992), but some
studies have suggested that RU 24969-produced hyperactivity is due to 5-HT1A
receptor activation (Aronsen et al., 2014; Kalkman, 1995). Repeated MDMA
administration (2x20 mg/kg/day, 4 consecutive days) failed, however, to alter
hyperactivity produced by the 5-HT1A receptor agonist, 8-OH-DPAT (Granoff &
Ashby, 2001). Therefore, the effect of MDMA exposure on the function of 5-HT1B
receptors is equivocal.
Studies on the effects of repeated exposure to MDMA have generally
administered a regimen that produces extensive, and persistent, neurotoxic effects. For
example, alterations in 5-HT1A binding, decreased tissue levels of 5-HT (Aguirre et al.,
1998) and decreased 5-HT transporter binding (Aguirre et al., 1995) were produced by
exposure to high doses (2x30mg/kg/day, 4 consecutive days) of MDMA. This high
level of exposure is rarely, if ever, experienced by ecstasy users (D. Hansen et al.,
2001; Parrott, 2005; Verheyden et al., 2003), which questions the external validity of
findings derived from these experiments (Baumann & Rothman, 2009; Cole &
Sumnall, 2003; De La Garza et al., 2007; Meyer et al., 2008).
MDMA exposure during self-administration is quite different from most
studies that employ experimenter-administered MDMA. In rats, MDMA self-
administration is initially limited, but with repeated testing intake gradually increases
for some subjects (Schenk et al., 2012). Given the differences in exposure as well as
the well documented differences between effects of contingent and non-contingent
drug administrations (Dworkin, Mirkis, & Smith, 1995; Miguéns et al., 2008), self-

78
administered MDMA might be expected to produce different effects than those seen
after experimenter-administration. Indeed, self-administered MDMA produced smaller
deficits in tissue levels of 5-HT compared to high dose experimenter-administered
MDMA (Do & Schenk, 2011; Scanzello et al., 1993; Schenk et al., 2007) even though
the total amount self-administered (165-350 mg/kg over 20-30 days of testing) was
greater than is generally administered to produce extensive neurotoxicity (20-80 mg/kg
in a single day). Additionally, intermittent or low dose exposure to MDMA was
neuroprotective against the toxic effects of subsequent high dose administrations
(Bhide et al., 2009; Piper et al., 2010).
Because of the limited amount of information concerning effects of self-
administered MDMA on brain and/or behaviour and the potential role of specific
neuroadaptations in some of the adverse effects of MDMA, this study determined the
effect of extensive MDMA self-administration on behavioural responses to 5-HT1A
and 5-HT1B receptor agonists.
Method
Subjects and procedures
Male Sprague-Dawley rats underwent catheter surgery as outlined in the
General Methods section.
MDMA self-administration
Rats were randomly assigned to self-administer either MDMA, or vehicle,
using the standard self-administration equipment outlined in the General Methods
section. Self-administration was conducted during 2 hour daily sessions, 6 days per
week. Initially, active lever responses were reinforced with MDMA (1.0 mg/kg), or
vehicle (0.1 ml) infusions according to an FR1 schedule. The vehicle control group
continued on this contingency for the remainder of the experiment. The MDMA self-
administration group continued with this contingency until a total of 90 infusions had
been self-administered, or 25 test sessions had been completed, whichever came first.
Rats that failed to self-administer 90 infusions within this 25 day cut-off period
(approximately 50%, as we have previously reported (Schenk et al., 2012)) were not
tested further. For those that met this criterion, the dose of MDMA was decreased to
0.5 mg/kg. The reinforcement schedule was then increased to FR2 for a minimum of 5
days and then FR5. Testing continued until a total intake of 350 mg/kg MDMA was
self-administered. Between 20 and 58 self-administration sessions were required to

79
reach a total intake of 350 mg/kg. Where possible, each rat in the vehicle self-
administration group was matched to a rat in the MDMA self-administration group to
ensure a comparable number of test sessions. A total of 73 rats met the initial criterion
of 90 infusions of MDMA (1.0 mg/kg/infusion) within the 25 day cut-off period. Of
these, some did not progress further due to loss of catheter patency (n=1), failure to
increase responding when the FR schedule was increased (n=12), or MDMA toxicity
(n=3). The remaining rats (n=57) completed testing and self-administered 350 mg/kg
MDMA. A total of 62 rats initiated vehicle self-administration, but 1 was removed
from the study due to an inner ear infection, leaving a total of 61 that self-administered
vehicle. Separate groups of rats that completed self-administration testing were then
randomly assigned to groups to measure the effects of either 8-OH-DPAT-produced
hyperactivity or RU 24969-produced adipsia.
Locomotor activity
Locomotor activity was assessed 2 days after the last self-administration
session. Rats were placed in the testing chamber for 30 minutes, followed by an
injection of 8-OH-DPAT (0.0, 0.03, 0.1, 0.3, 1.0 mg/kg, s.c., n=5-7 per group).
Horizontal activity counts were recorded in 5 minute intervals during the 30 minutes
prior to, and 60 minutes following, the 8-OH-DPAT injection.
Water consumption
The day following the last self-administration session, water bottles were
removed from the home cages for 24 hours. Fifteen minutes before water bottles were
reintroduced, RU 24969 (0, 0.3, 1.0, 3.0 mg/kg, s.c., n= 6-9 per group) was
administered, as previously reported (Aronsen et al., 2014). Water bottles were
weighed before, and after 30 minutes of access, to measure water consumption.
Data analysis
Effects of 8-OH-DPAT on locomotor activity were analysed by a 2 (self-
administration group) × 5 (Dose of 8-OH-DPAT) ANOVA. RU 24969-produced
adipsia was analysed with a 2 (self-administration group) × 4 (Dose of RU 24969)
ANOVA.
Results
Self-administration
The average amount of MDMA that was self-administered during the last 5
days of testing was 13.2 mg/kg/day (SEM=0.55). Figure 7.1 shows the distribution of

80
the number of rats that self-administered 350 mg/kg of MDMA as a function of test
session. Most of the rats met the criterion within 25-44 test sessions. The mean number
of test sessions required to complete testing was 35.7 (SEM=1.3). The average number
of days to complete testing reported in this study is similar to data that we have
previously reported. For example, an average of 37 +/- 2.3 days was required to self-
administer a slightly lesser total of 315 mg/kg that resulted in decreased tissue levels of
5-HT (Do & Schenk, 2011). The vehicle self-administration group was tested for an
average of 36 sessions (SEM= 1.4). These rats were matched to the MDMA self-
administration rats to minimise any confounds associated with the self-administration
procedure.
N u m b e r o f s e s s io n s
Nu
mb
er
of
rats
(n
=5
7)
2 0 -24
2 5 -29
3 0 -34
3 5 -39
4 0 -44
4 5 -49
5 0 -54
5 5 -59
0
5
1 0
1 5
Fig. 7.1 Frequency distribution of the number of rats that self-‐administered 350 mg/kg MDMA as a function of test session. 8-OH-DPAT-produced hyperactivity
Figure 7.2 shows the hyperactive response to 8-OH-DPAT after self-administration of MDMA or vehicle. ANOVA showed an effect of 8-OH-DPAT dose (F(4,47) = 27.27, p<0.01, ɳp
2= 0.70), but no effect of self-administration (F(1,47) = 0.79, p=0.38), and no interaction (F(4,47) = 0.50, p=0.50).

81
8-OH-DPAT (mg/kg)
Tota
l am
bula
tory
cou
nts
0.10
1000
2000
3000
4000VehicleMDMA
0.0 0.03 0.3 1.0
Fig. 7.2 Effect of MDMA self-‐administration (350 mg/kg total) on 8-‐OH-‐DPAT-‐produced hyperactivity. Rats in these groups met the criterion of 350 mg/kg MDMA after 25-‐58 test sessions. Symbols represent mean ± SEM. n = 5-‐7 per group. RU 24969-produced adipsia
As we have previously shown (Aronsen et al., 2014), RU 24969 produced a
dose-dependent adipsic response (F(3,51) = 65.68, p<0.01, ɳp2= 0.79; Fig 7.3). There
was no statistically significant effect of self-administration (F(1,51) = 2.86, p=0.10)
and no statistically significant interaction (F(3,51) = 1.60, p=0.20).
RU 24969 (mg/kg)
Wat
er c
onsu
mpt
ion
(ml)
0
5
10
15 VehicleMDMA
0.0 0.3 3.01.0
Fig. 7.3 Effect of MDMA self-‐administration (350 mg/kg total) on RU 24969-‐produced adipsia. Rats in these groups met the criterion of 350 mg/kg MDMA after 20-‐58 test sessions. Symbols represent mean ± SEM. n = 6-‐9 per group.

82
Discussion
MDMA self-administration failed to alter 8-OH-DPAT-produced hyperactivity,
or RU 24969-produced adipsia. It is unlikely that the MDMA exposure was
insufficient because similar or lower doses of self-administered MDMA produced
decreases in 5-HT transporter binding (Schenk et al., 2007), decreases in tissue levels
of 5-HT (Do & Schenk, 2011; Schenk et al., 2011), and behavioural deficits (Do &
Schenk, 2011). Instead, the present data suggest that 5-HT1A and 5-HT1B receptor
mechanisms are not altered by MDMA self-administration.
These findings were surprising because prolonged activation by MDMA-
produced 5-HT release might have been expected to down-regulate these receptor
subtypes. Alternatively, the decrease in MDMA-produced 5-HT release that has been
reported following MDMA self-administration (Reveron et al., 2010) might have been
expected to result in a compensatory up-regulation of these receptors. A neurotoxic
5,7-DHT lesion increased 5-HT1B receptor binding (Compan et al., 1998; Crino, Vogt,
Volicer, & Wiley, 1990; Frankfurt, Mendelson, McKittrick, & McEwen, 1993;
Manrique et al., 1998; Manrique et al., 1994; Manrique et al., 1993; Offord et al.,
1988; Weissmann et al., 1986). Furthermore, repeated agonist treatment decreased 5-
HT1B receptor binding (Pranzatelli & Razi, 1994), and behavioural responses to 5-
HT1A (De Souza, Goodwin, Green, & Heal, 1986; Hensler, 2003) and 5-HT1B (Frances
& Monier, 1991) receptor agonists.
Repeated exposure to other drugs that increase synaptic 5-HT levels altered 5-
HT1A and 5-HT1B receptors. For example, chronic treatment with the selective 5-HT
reuptake inhibitor (SSRI), fluoxetine, decreased 5-HT1B receptor binding (Duncan,
Hester, Hopper, & Franklin, 2010). It is important to note, however, that many of the
effects of SSRI treatment reflect alterations that are most likely attributed to
autoreceptor, rather than post-synaptic receptor, desensitisation. For example, repeated
treatment with fluoxetine (8 mg/kg/day, 2-3 weeks) reduced 5-HT1A mRNA in the
raphe nuclei (Le Poul et al., 2000). Higher doses also produced a decrease in 5-HT1A
receptor binding (Welner, De Montigny, Desroches, Desjardins, & Suranyi-‐Cadotte,
1989) and 8-OH-DPAT stimulated [35S]GTPγS binding (Castro et al., 2003) in the
dorsal raphe. Repeated exposure to MDMA failed to alter a number of 5-HT1A
autoreceptor mediated behavioural or neurochemical responses (Schenk et al., 2013),
suggesting differences between effects of these two classes of drugs. Repeated
administrations of cocaine increased 5-HT1B receptor binding (Przegaliński, Czepiel,

83
Nowak, Dlaboga, & Filip, 2003) and 5-HT1B mRNA (Hoplight, Vincow, & Neumaier,
2007). Cocaine self-administration also increased the behavioural and physiological
responses to 5-HT1A and 5-HT1B receptor agonists (O'Dell, Manzardo, Polis, Stouffer,
& Parsons, 2006).
The present data do not rule out the possibility that repeated ecstasy use leads
to cognitive and behavioural deficits via dysregulation of these receptor subtypes, but
our results suggest that other 5-HT receptors are more likely to make important
contributions. One potential candidate is the 5-HT2A receptor, because it has also been
implicated in impulsivity (Cunningham & Anastasio, 2014) , sleep (Sharpley, Elliott,
Attenburrow, & Cowen, 1994) and memory (Dhonnchadha & Cunningham, 2008;
Howell & Cunningham, 2015), behaviours that are impacted by regular ecstasy use.
MDMA exposure increased 5-HT2A receptor binding (Benningfield & Cowan, 2013;
Urban et al., 2012) and behavioural responses to the 5-HT2A/2C receptor agonist, DOI
(Biezonski, Courtemanche, Hong, Piper, & Meyer, 2009). Additional studies assessing
the impact of MDMA self-administration on this receptor mechanism is warranted.

84
General Discussion
Summary
MDMA is widely used in the form of the street drug, ecstasy. Regular use of
ecstasy has been associated with a number of behavioural and neurochemical deficits,
and some of these deficits likely contribute to further, problematic drug taking. While
most drugs of abuse primarily enhance dopamine neurotransmission, MDMA
preferentially releases 5-HT. This 5-HT release has been hypothesised to inhibit the
dopaminergic response to MDMA, thus inhibiting the reinforcing efficacy of MDMA.
However, with repeated exposure to MDMA, the 5-HTergic response is attenuated,
disinhibiting the dopaminergic response and making MDMA similar to other drugs of
abuse. The mechanism for this 5-HTergic inhibition of dopamine is not known, but
one possibility is that activation of specific 5-HTergic receptors, via MDMA-produced
5-HT release, alters the dopaminergic response to MDMA. Of the 14 different 5-HT
receptors, the 5-HT1A and 5-HT1B receptors were investigated because of a
documented role in regulating basal and drug-produced dopamine release, as well as
behaviours associated with ecstasy use. The purpose of this thesis was to test the role
of these receptors in the self-administration of MDMA in rats, and to document any
changes in these receptor populations produced by MDMA.
Firstly, appropriate behavioural assays for 5-HT1A and 5-HT1B receptor
activation needed to be identified. Latent inhibition, measured using the conditioned
taste aversion paradigm, was chosen as a behavioural response to 5-HT1B receptor
activation, but this response was confounded by the adipsic response to the 5-HT1B/1A
receptor agonist, RU 24969. After further testing, I found that this adipsic response to
RU 24969 was dose-dependent, and blocked by a 5-HT1B, but not a 5-HT1A, receptor
antagonist. Thus, the adipsic response to RU 24969 was chosen as a behavioural
measure of 5-HT1B receptor activation. In contrast, the hyperactive response to RU
24969 was blocked by a 5-HT1A, but not a 5-HT1B, receptor antagonist. A similar result
was obtained with the more selective and well characterised 5-HT1A receptor agonist,
8-OH-DPAT, thus the locomotor response to 8-OH-DPAT was chosen as a
behavioural response to 5-HT1A receptor activation.
To test whether 5-HT1A and/or 5-HT1B receptors regulated MDMA self-
administration I attempted to alter the activity of these receptors and measure the

85
impact on the acquisition of MDMA self-administration. To this end, rats were
repeatedly administered a high dose of the 5-HT1B/1A receptor agonist, RU 24969,
before commencing MDMA self-administration. The pretreatment down-regulated 5-
HT1A and 5-HT1B receptors, and greatly facilitated the acquisition of MDMA self-
administration. Because drug produced dopamine release is inhibited by activation of
5-HT1A receptors, but enhanced by 5-HT1B receptors, the impact of RU 24969
pretreatment on acquisition of MDMA self-administration was hypothesised to be
associated with the down-regulation of 5-HT1A receptors.
The role of 5-HT1A and 5-HT1B receptors in the acquisition of MDMA self-
administration was further tested by investigating the relationship between basal
responses to receptor activation and latency to acquire MDMA self-administration.
Based on the role of these receptors in regulating the dopaminergic response to other
drugs of abuse, and the facilitated acquisition of MDMA self-administration after
repeated exposure to RU 24969, it was expected that behavioural responses to
activation of these receptors would predict the latency to acquire MDMA self-
administration. This hypothesis was not supported in any of the studies. Furthermore,
because an inhibitory role of 5-HT1A receptor activation in the acquisition of MDMA
self-administration was hypothesised, I investigated the effect of 5-HT1A receptor
antagonist treatment during the acquisition phase. Again, results did not support the
hypothesis, in fact, the results suggested that 5-HT1A receptor blockade inhibited
MDMA self-administration.
Acquisition studies had returned mainly negative results, but there was still
reason to believe that 5-HT1A and/or 5-HT1B receptors regulated the self-administration
of MDMA. Therefore, behavioural responses to 5-HT1A or 5-HT1B receptor activation
were measured after the self-administration of a high dose of MDMA. It was expected
that prolonged exposure to MDMA would alter behavioural responses to agonist
administration, but again this hypothesis was not supported. Although these studies do
not rule out the possibility of 5-HT1A or 5-HT1B receptor neuroadaptations in response
to MDMA self-administration, they do suggest that other 5-HT receptors are more
likely to make important contributions.
Synthesis of results
Overall, the data presented in this thesis are difficult to reconcile. On the one
hand, there is a sound theoretical basis to expect that 5-HT1A and 5-HT1B receptors

86
would modulate the reinforcing efficacy of MDMA, and that these receptor
mechanisms would be altered by prolonged exposure to MDMA. Furthermore, the RU
24969 pretreatment, that down-regulated 5-HT1A and 5-HT1B receptors, also facilitated
the acquisition of MDMA self-administration. On the other hand, behavioural
responses to 5-HT1A and 5-HT1B receptor activation did not predict the acquisition of
MDMA self-administration, acquisition was blocked by the 5-HT1A receptor
antagonist, WAY 100635, and there were no changes in dose response curves for 5-
HT1A or 5-HT1B mediated responses after the self-administration of a high dose of
MDMA.
Together, the most likely explanation for the results of the studies comprising
this thesis is that 5-HT1A and 5-HT1B receptors have a limited role in the self-
administration of MDMA. If true, this conclusion would suggest that the facilitated
self-administration produced by RU 29496 pretreatment was due to some non-specific
effect. To test this possibility, it would be important to co-administer a 5-HT1A or 5-
HT1B receptor antagonist with RU 24969 during pretreatment and test for latency to
acquire MDMA self-administration.
Limitations
It is possible that the conclusions made in this thesis were skewed by the
behavioural measures used. Although it was demonstrated that RU 24969-produced
adipsia and 8-OH-DPAT-produced locomotor activity are measures of 5-HT1B and 5-
HT1A receptor activation, respectively, there is no consensus on what population of 5-
HT1B or 5-HT1A receptors produce these effects. Systemic administration of 8-OH-
DPAT produced dose-dependent hyperactivity, but this locomotor response is the net
result of global 5-HT1A activation. Specific 5-HT1A populations alter locomotor
activity in different ways, for example local injections of 8-OH-DPAT in the PFC did
not alter locomotor activity (Solati et al., 2011), while administration in the NAc
decreased locomotor activity (Hillegaart, Ahlenius, & Larsson, 1991; Plaznik et al.,
1994). Similarly, the population of 5-HT1B receptors responsible for the adipsic
response to RU 24969 is not known. One study showed that local infusion of RU
24969, or the more selective 5-HT1B receptor agonist, CP 93129, in the NAc reduced
responding for water (P. J. Fletcher & Korth, 1999a), but it is not clear what other
populations of 5-HT1B receptors might also influence this behavioural response. Thus,
care needs to be taken when interpreting these behavioural data. It is possible, for

87
example, that MDMA self-administration did alter some 5-HT1A and/or 5-HT1B
receptor populations, but not those that impact the locomotor response to 8-OH-DPAT
or the adipsic response to RU 24969.
Extensive study would be required to address the possibility that the negative
results found in this thesis were due to the choice of behavioural responses. Because of
the time, rats, and drugs required, it was not possible to investigate further for this
thesis, but our lab has started to probe this possibility in further detail. The logical first
step is to directly investigate the effect of MDMA self-administration on 5-HT1A and
5-HT1B receptor binding. There is a widely used and well characterised 5-HT1A
antibody (Abbas et al., 2007; Kia et al., 1996; Say, Machaalani, & Waters, 2007;
Tachibana, Endoh, Fujiwara, & Nawa, 2005), allowing for an immunohistochemistry
investigation, but 5-HT1B receptors are best mapped using a radioactively labelled
ligand (Domenech, Beleta, & Palacios, 1997; Lindhe et al., 2011). Our lab is currently
conducting 5-HT1A immunohistochemistry on tissue from rats that have extensive
MDMA self-administration history. Such an approach allows for a detailed, region
specific, analysis of the effect of MDMA self-administration. Similar data could be
obtained to determine the effect of RU 24969 pretreatment. Receptor populations that
are similarly affected by both manipulations might underlie the development of
MDMA as an efficacious reinforcer. Thus, local drug administrations in these areas
could be used, first to obtain a behavioural response to predict the acquisition of
MDMA self-administration, then for pharmacological treatments to reduce MDMA
self-administration.
Validity of MDMA doses
The self-administration paradigm was used in this thesis because it likely
produces neuroadaptations similar to those produced by ecstasy use in humans. It is
important to note that, when compared across species, the MDMA doses self-
administered in this thesis were of relevance to human users. The issue of interspecies
scaling is based on the fact that, in general, smaller animals have relatively larger
organs and a shorter blood circulation time, and so will metabolise drug faster
(Mordenti & Chappell, 1989). Therefore, as long as there are no species-specific
mechanisms of drug metabolism, smaller animals require larger doses in order for
effects to be comparable to those produced in larger animals. Most recreational users
consume 1-2 tablets per ecstasy-taking session (Parrott, 2005; Verheyden et al., 2003),

88
and although the contents of ecstasy tablets procured ‘on the street’ vary widely,
median MDMA content per tablet has been shown to be around 70-80 mg (Vogels et
al., 2009). Therefore, a 70kg user is consuming approximately 1-2 mg/kg MDMA in
recreational settings. Interspecies scaling can help to determine the doses that should
be administered in animal studies to best mimic the effects of such doses in
recreational users.
As a starting point for investigating drugs across species the USA Food and
Drug Administration (FDA) suggest that drug doses should be scaled across species
based on the body weight and surface area of these species (Food and Drug
Administration, 2005). The FDA recommendation is that the effects of a 1.0 mg/kg
dose in a human are roughly comparable with the effects of a 6.2 mg/kg dose in a rat.
The FDA scaling suggestions are not drug-specific and are meant merely as a
guideline for determining safe initial doses in clinical trials.
With specific reference to MDMA, some researchers have used the following
algorithm to scale doses between species:
DHuman = DAnimal (WHuman / WAnimal) k (Equation 1)
where D is the drug dose in mg, W is weight in kg, and k is an estimated value that
reflects the logarithmic relationship between bodyweight and metabolic rate (that is,
the slope of the curve fitted to the log transforms of empirical values for weight and
drug clearance times of different species). This process, in which doses are determined
by transforming bodyweight to a different physiological variable through a power
function, is called ‘allometric scaling’ (Mordenti & Chappell, 1989). The precise value
of the scaling factor, k, has been seriously contended in the literature, with suggestions
ranging between 0.67 and 0.77 (Food and Drug Administration, 2005; Mordenti &
Chappell, 1989; Travis & White, 1988; Watanabe, Bois, & Zeise, 1992). In studies
using MDMA, a k value of 0.7 has been adopted (McCann & Ricaurte, 2001; Ricaurte,
Yuan, & McCann, 2000). Based on Equation 1, with k set at 0.7, a 1.0 mg/kg dose in a
70kg human would be equivalent to a 5.0 mg/kg dose in a 330g rat. It should be noted
that this suggested dose could vary from 3.4 mg/kg to 5.9 mg/kg if the highest or
lowest suggested k value is used, respectively.
Vollenweider, Jones, and Baggott (2001) have suggested that allometric scaling
is not relevant to MDMA because there is evidence for species differences in MDMA
pharmacokinetics, and because MDMA has active metabolites that may contribute to
the drug effect. Instead, they suggest that pharmacokinetic data (e.g. area under the

89
curve (AUC) of MDMA plasma levels) should be compared between species to
determine similar doses. Based on equation 1, McCann and Ricaurte (2001) claim that
a 20 mg/kg dose of MDMA in a 220g rat is comparable to a 1.4 mg/kg dose in a
human, however AUC of MDMA plasma concentrations in humans after
approximately 1.8 mg/kg was 70% lower than the AUC in rats after 20 mg/kg
(Vollenweider et al., 2001). Although comparing pharmacokinetic data can account for
some potential flaws with allometric scaling, comparisons can only be made with
empirical data, so finding similar doses across species becomes a ‘trial and error’ type
task.
Both allometric scaling and comparisons of pharmacokinetic data (AUC)
suggest that a human dose of approximately 1 mg/kg is comparable to a rat dose of
roughly 5 mg/kg (De La Torre et al., 2000; Fitzgerald, Blanke, & Poklis, 1990). Thus,
during initial self-administration sessions, in which rats self-administer less than 5
infusions per session, rats are consuming less MDMA than a human user might be
expected to use recreationally. As intake increases, rats will self-administer 5-10
mg/kg per session, which roughly scales to the human recreational dose. The total
intake of 350 mg/kg MDMA used in this thesis is comparable to that of a heavy
ecstasy user, after roughly 70 recreational doses. Most studies have shown cognitive or
behavioural deficits in ecstasy users to be present at levels of total intake around or
below 70 doses (Booij et al., 2014; A. K. Davis & Rosenberg, 2014; McCann, Mertl, et
al., 1999; Wagner et al., 2013). Therefore, the doses of MDMA self-administered by
rats in this thesis are relevant to human ecstasy users, particularly heavy users.
Key findings and future directions
This thesis made a number of novel and important findings. Firstly,
characterising the adipsic and hyperactive responses to RU 24969 as 5-HT1B and 5-
HT1A receptor mediated, respectively, was important for clarifying previous findings
and facilitating further research. Earlier studies had not clearly shown the mechanism
by which RU 24969 produced hyperactivity, and some had interpreted the hyperactive
response as a behavioural measure of 5-HT1B receptor activation (Callaway & Geyer,
1992). The results from this thesis clearly show a role of 5-HT1A, but not 5-HT1B
receptors in RU 24969-produced hyperactivity. Furthermore, this thesis provides a
straightforward behavioural assay for 5-HT1B receptor activation, and this behavioural
measure might be useful in preclinical tests. As outlined above, further study of the

90
population of 5-HT1B receptors that produce the the adipsic response to RU 24969
would make this behavioural assay more useful.
As one reviewer pointed out, the facilitation of MDMA self-administration
produced by RU 24969 pretreatment is a ‘novel and important’ finding for the
addiction field, although more work needs to be done to understand this effect. I would
strongly encourage further investigation of the effects of RU 24969 pretreatment on 5-
HTergic systems so that the mechanism by which this pretreatment facilitated MDMA
self-administration can be elucidated. Another novel finding made in this thesis was
that MDMA self-administration had no effect on behavioural responses to 5-HT1A or
5-HT1B activation. Again, a reviewer commented that these results are interesting and
important, even though the results were negative.
The studies in this thesis were based on the theory that a decreased 5-HTergic
response to MDMA after repeated exposure could enhance the reinforcing effects of
MDMA via altered activation of 5-HT receptors that regulate dopamine
neurotransmission. Although this thesis suggests that 5-HT1A and 5-HT1B receptors
likely play a limited role in the enhanced reinforcing efficacy of MDMA after repeated
exposure, the theoretical basis for these studies is still sound. Thus, it is possible that
there are other 5-HT receptors that regulate the reinforcing efficacy of MDMA and
that also underlie cognitive and behavioural deficits following repeated exposure. Two
5-HT receptors that have been shown to regulate dopaminergic neurotransmission are
the 5-HT2A and 5-HT2C receptors.
5-HT2C receptors are well localised to mediate the dopaminergic responses to
drugs of abuse, with high levels of 5-HT2C receptors reported in dopamine terminal
areas of the PFC, striatum, and NAcc, and in the VTA (Bubar & Cunningham, 2006;
Clemett, Punhani, Duxon, Blackburn, & Fone, 2000; Di Matteo, De Blasi, Di Giulio,
& Esposito, 2001; Eberle-‐Wang, Mikeladze, Uryu, & Chesselet, 1997; Ji et al., 2006).
5-HT2C receptor agonists inhibited, while 5-HT2C receptor antagonists enhanced, the
firing rate of VTA dopamine neurons, and extracellular dopamine levels in the nucleus
accumbens and PFC (Alex, Yavanian, McFarlane, Pluto, & Pehek, 2005; Di Matteo et
al., 2001).
The 5-HT2C/2B receptor antagonist, SB 206553, and the more selective 5-HT2C
receptor antagonist, SB 242084, both potentiated the cocaine-produced increase in
extracellular dopamine in the nucleus accumbens and striatum (Navailles, De
Deurwaerdere, Porras, & Spampinato, 2004).The 5-HT2C receptor agonist, Ro 60-175,

91
inhibited the self-administration of cocaine, ethanol, and nicotine, an effect that was
reversed by the 5-HT2C receptor antagonist, SB 242084 (P. J. Fletcher, Chintoh,
Sinyard, & Higgins, 2004; P. J. Fletcher, Rizos, Sinyard, Tampakeras, & Higgins,
2007; Grottick, Corrigall, & Higgins, 2001; Grottick, Fletcher, & Higgins, 2000;
Tomkins et al., 2002). Mice that lack the 5-HT2C gene reached higher breakpoints in a
progressive ratio paradigm reinforced by cocaine, and also showed enhanced levels of
cocaine-induced dopamine release in the NAcc (Rocha et al., 2002).
These findings are consistent with the idea that activation of 5-HT2C receptors
is inhibitory to, while blockade of 5-HT2C receptors facilitates, the dopaminergic
response to drugs of abuse. As such, it is possible that a down-regulation of 5-HT2C
receptors, in response to repeated exposure to MDMA, underlies the development of
MDMA as an efficacious reinforcer
Unfortunately, there is no clear evidence for 5-HT2C receptor down-regulation
in response to MDMA exposure. On the one hand, male ecstasy users showed blunted
neuroendocrine responses to the 5-HT2/1A receptor agonist, m-CPP, compared to
MDMA-naïve controls (McCann, Eligulashvili, Mertl, Murphy, & Ricaurte, 1999),
and repeated administration of MDMA decreased 5-HT2C receptor protein levels in the
hippocampus of young-adult rats (García-Cabrerizo & García-Fuster, 2015),
suggesting a possible down-regulation of 5-HT2C receptors after MDMA exposure. On
the other hand, repeated exposure to MDMA enhanced the inhibition of MDMA-
produced hyperlocomotion by the 5-HT2C receptor agonist, MK 212 (Ramos, Goni-
Allo, & Aguirre, 2005) and increased sensitivity to the 5-HT2/1A receptor agonist, m-
CPP (Taffe et al., 2002). Furthermore, repeated exposure to MDMA increased 5-HT2C
mRNA in cortex and hypothalamus (Kindlundh-Högberg et al., 2006). Further still,
some animal studies have failed to show any effect of MDMA exposure on
neuroendocrine or behavioural responses to m-CPP (Bull et al., 2003; Jones, Brennan,
Colussi-Mas, & Schenk, 2010).
It is entirely possible that repeated exposure to MDMA in the self-
administration paradigm would down-regulate 5-HT2C receptors, but so far there is
limited evidence to suggest this would be the case. Significantly more research is
required to determine the effects of MDMA self-administration on 5-HT2C receptor
mechanisms. On the other hand, there is substantial evidence to suggest that
neuroadaptations in 5-HT2A receptor mechanisms might underlie the development of
MDMA as an efficacious reinforcer in the self-administration paradigm.

92
5-HT2A receptors are strongly expressed as excitatory 5-HTergic receptors on
non-5-HTergic cells in the PFC (Eison & Mullins, 1995), where their activation has
been shown to increase dopamine activity in the VTA (Bortolozzi, Díaz-Mataix,
Scorza, Celada, & Artigas, 2005). This increased mesocorticolimbic dopamine release
is a product of increased glutamatergic activity in projections from the PFC to the
VTA (Aghajanian & Marek, 1999; Pehek, Nocjar, Roth, Byrd, & Mabrouk, 2005). The
5-HT2A/2C receptor antagonist, ketanserin, attenuated the dopaminergic response to
MDMA in the striatum (Nash, 1990), and a similar effect was produced by local
administration of the selective 5-HT2A receptor antagonist, M100907, in the striatum
(Schmidt, Sullivan, & Fedayal, 1994). On the other hand the non-selective 5-HT2
receptor agonists, DOI and 5-MeODMT, both enhanced the dopaminergic response to
MDMA in the striatum (Gudelsky, Yamamoto, & Nash, 1994). These data suggest that
activation of the 5-HT2A receptor via MDMA-induced 5-HT release would enhance the
reinforcing efficacy of MDMA.
Compellingly, repeated MDMA was associated with increased 5-HT2A receptor
binding (Benningfield & Cowan, 2013; Urban et al., 2012)(but see McGregor et al.
(2003)), suggesting that MDMA self-administration might also up-regulate 5-HT2A
receptors. Thus, with repeated exposure to MDMA, enhanced activation of 5-HT2A
receptors could underlie the development of MDMA as an efficacious reinforcer. This
hypothesis is in agreement with the finding that a neurotoxic 5,7-DHT lesion, which
also facilitated the acquisition of MDMA self-administration (Bradbury et al., 2014),
produced an increase in 5-HT2A receptor binding density in mice (Heal, Philpot,
Molyneux, & Metz, 1985).
An up-regulation of 5-HT2A receptors might also underlie the increased
impulsivity produced by repeated exposure to MDMA. The 5-HT2A/2C receptor agonist,
DOI, increased premature responding on the 5CSRTT (Koskinen, Haapalinna, & Sirvi,
2003; Koskinen & Sirviö, 2001), and this effect was blocked by the 5-HT2A/2C receptor
antagonist, ketanserin (Koskinen, Ruotsalainen, Puumala, et al., 2000; Koskinen,
Ruotsalainen, & Sirviö, 2000), while ketanserin (P. J. Fletcher, Tampakeras, Sinyard,
& Higgins, 2007; Passetti, Dalley, & Robbins, 2003; Ruotsalainen et al., 1997; Talpos,
Wilkinson, & Robbins, 2006) and the more selective 5-HT2A receptor antagonist,
M100907 (P. J. Fletcher, Tampakeras, et al., 2007; Winstanley, Theobald, Dalley,
Glennon, & Robbins, 2004), decreased premature responses on the 5CSRTT, or the

93
similar 1CSRTT (Anastasio et al., 2011). Increased impulsivity, due to an up-
regulation of 5-HT2A receptors after repeated exposure to MDMA, would be expected
to facilitate drug taking, and as such could underlie the development of an MDMA
SUD (Schenk & Aronsen, 2015).
The role of the 5-HT2A receptor in MDMA self-administration has not been
studied. Some studies have shown no effect of regular, repeated MDMA
administration on the head-twitch or locomotor responses to the 5-HT2A/2C receptor
agonist, DOI (Granoff & Ashby Jr, 1998), or the behavioural response to the non-
selective 5-HT2 receptor agonist, mCPP (Jones et al., 2010). On the other hand, an
intermittent dosing regimen of MDMA increased the head-twitch responses to DOI
(Biezonski et al., 2009), suggesting that adaptations in 5-HT2A receptors may be
dependent on dosing regimen. Ecstasy use is typically intermittent, and 5-HT2A
binding was increased in human ecstasy users, compared to naïve controls, with
increased exposure to MDMA associated with increased 5-HT2A binding density (Di
Iorio et al., 2012; Urban et al., 2012). Thus, it is possible that an up-regulation of 5-
HT2A receptors would be evident after MDMA self-administration.
It would be interesting to selectively up-regulate 5-HT2A receptors and test for
latency to acquire MDMA self-administration. An up-regulation may be achieved by
repeatedly administering the selective 5-HT2A receptor antagonist, M100907 (Minabe,
Hashimoto, Watanabe, & Ashby, 2001). Behavioural assessment is difficult, however,
because of the lack of selective 5-HT2A receptor agonists. Ideally, a behavioural
response to the selective antagonist, M100907, would be determined. There are some
reports that M100907 enhanced the inhibition of a startle response in the pre-pulse
inhibition paradigm (Padich, McCloskey, & Kehne, 1996; Zhang, Engel, Jackson,
Johansson, & Svensson, 1997), although more parametric work for this behavioural
response is required (Geyer, Krebs-Thomson, & Varty, 1999; Varty, Bakshi, & Geyer,
1999)
Conclusion
Repeated exposure to MDMA enhances the reinforcing efficacy of MDMA. It
is possible that this increased reinforcement is due to adaptations in 5-HT receptors
that regulate dopaminergic responses to MDMA. This thesis showed that 5-HT1A and
5-HT1B receptors likely play a limited role in the self-administration of MDMA, and
thus likely do not explain the enhanced reinforcing efficacy of MDMA after repeated

94
exposure. Future research should consider the role of the 5-HT2A receptor in
neuroadaptations that might underlie the self-administration of MDMA.

95
References Abbas, S. Y., Nogueira, M. I., & Azmitia, E. C. (2007). Antagonist-‐induced increase in 5-‐HT1A-‐
receptor expression in adult rat hippocampus and cortex. Synapse, 61(7), 531-‐539. Acosta, J. I., Boynton, F. A., Kirschner, K. F., & Neisewander, J. L. (2005). Stimulation of 5-‐HT1B
receptors decreases cocaine-‐and sucrose-‐seeking behavior. Pharmacology Biochemistry and Behavior, 80(2), 297-‐307.
Acquas, E., Marrocu, P., Pisanu, A., Cadoni, C., Zernig, G., Saria, A., & Di Chiara, G. (2001). Intravenous administration of ecstasy (3, 4-‐methylendioxymethamphetamine) enhances cortical and striatal acetylcholine release in vivo. European journal of pharmacology, 418(3), 207-‐211.
Aghajanian, G. K., & Marek, G. J. (1999). Serotonin, via 5-‐HT2A receptors, increases EPSCs in layer V pyramidal cells of prefrontal cortex by an asynchronous mode of glutamate release. Brain research, 825(1–2), 161-‐171. doi:http://dx.doi.org/10.1016/S0006-‐8993(99)01224-‐X
Ago, Y., Nakamura, S., Hayashi, A., Itoh, S., Baba, A., & Matsuda, T. (2006). Effects of osemozotan, ritanserin and azasetron on cocaine-‐induced behavioral sensitization in mice. Pharmacology Biochemistry and Behavior, 85(1), 198-‐205.
Aguirre, N., Ballaz, S., Lasheras, B., & Del Rio, J. (1998). MDMA (Ecstasy') enhances 5-‐HT1A receptor density and 8-‐OH-‐DPAT-‐induced hypothermia: blockade by drugs preventing 5-‐hydroxytryptamine depletion. European journal of pharmacology, 346(2), 181-‐188.
Aguirre, N., Frechilla, D., García-‐Osta, A., Lasheras, B., & Del RIo, J. (1997). Differential Regulation by Methylenedioxymethamphetamine of 5-‐Hydroxytryptamine1A Receptor Density and mRNA Expression in Rat Hippocampus, Frontal Cortex, and Brainstem: The Role of Corticosteroids. Journal of Neurochemistry, 68(3), 1099-‐1105.
Aguirre, N., Galbete, J., Lasheras, B., & Del Río, J. (1995). Methylenedioxymethamphetamine induces opposite changes in central pre-‐and postsynaptic 5-‐HT1A receptors in rats. European journal of pharmacology, 281(1), 101-‐105.
Ahlenius, S., & Salmi, P. (1995). Antagonism of Reserpine-‐Induced Suppression of Spontaneous Motor Activity by Stimulation of 5-‐HT1A Receptors in Rats. Pharmacology & toxicology, 76(2), 149-‐156.
Ahrens, A. M., Singer, B. F., Fitzpatrick, C. J., Morrow, J. D., & Robinson, T. E. (2016). Rats that sign-‐track are resistant to Pavlovian but not instrumental extinction. Behavioural brain research, 296, 418-‐430.
Alex, K. D., & Pehek, E. (2007). Pharmacologic mechanisms of serotonergic regulation of dopamine neurotransmission. Pharmacology & therapeutics, 113(2), 296-‐320.
Alex, K. D., Yavanian, G. J., McFarlane, H. G., Pluto, C. P., & Pehek, E. A. (2005). Modulation of dopamine release by striatal 5-‐HT2C receptors. Synapse, 55(4), 242-‐251. doi:10.1002/syn.20109
Altman, J., Everitt, B., Robbins, T., Glautier, S., Markou, A., Nutt, D., . . . Phillips, G. (1996). The biological, social and clinical bases of drug addiction: commentary and debate. Psychopharmacology, 125(4), 285-‐345.
Ambar, G., & Chiavegatto, S. (2009). Anabolic-‐androgenic steroid treatment induces behavioral disinhibition and downregulation of serotonin receptor messenger RNA in

96
the prefrontal cortex and amygdala of male mice. Genes, Brain and Behavior, 8(2), 161-‐173.
American Psychiatric Association. (2013). Diagnostic and statistical manual of mental disorders (5 ed.). Arlington, VA: American Psychiatric Publishing.
Anastasio, N. C., Stoffel, E. C., Fox, R. G., Bubar, M. J., Rice, K. C., Moeller, F. G., & Cunningham, K. A. (2011). The serotonin (5-‐HT) 5-‐HT2A receptor: Association with inherent and cocaine-‐evoked behavioral disinhibition in rats. Behavioural pharmacology, 22(3), 248.
Anderson, G., Braun, G., Braun, U., Nichols, D. E., & Shulgin, A. T. (1978). Absolute configuration and psychotomimetic activity. NIDA Research Monograph, 22, 8-‐15.
Anneken, J. H., & Gudelsky, G. A. (2012). MDMA produces a delayed and sustained increase in the extracellular concentration of glutamate in the rat hippocampus. Neuropharmacology, 63(6), 1022-‐1027.
Arborelius, L., Chergui, K., Murase, S., Nomikos, G. G., Höök, B. B., Chouvet, G., . . . Svensson, T. H. (1993). The 5-‐HT1A receptor selective ligands,(R)-‐8-‐OH-‐DPAT and (S)-‐UH-‐301, differentially affect the activity of midbrain dopamine neurons. Naunyn-‐Schmiedeberg's archives of pharmacology, 347(4), 353-‐362.
Arborelius, L., Nomikos, G., Hacksell, U., & Svensson, T. (1993). (R)-‐8-‐OH-‐DPAT preferentially increases dopamine release in rat medial prefrontal cortex. Acta physiologica scandinavica, 148(4), 465-‐466.
Aronsen, D., Webster, J., & Schenk, S. (2014). RU 24969-‐produced adipsia and hyperlocomotion: Differential role of 5HT1A and 5HT1B receptor mechanisms. Pharmacology Biochemistry and Behavior, 124, 1-‐4.
Assié, M.-‐B., Bardin, L., Auclair, A. L., Carilla-‐Durand, E., Depoortère, R., Koek, W., . . . Newman-‐Tancredi, A. (2010). F15599, a highly selective post-‐synaptic 5-‐HT1A receptor agonist: in-‐vivo profile in behavioural models of antidepressant and serotonergic activity. International Journal of Neuropsychopharmacology, 13(10), 1285-‐1298.
Audi, E., De Oliveira, R., & Graeff, F. (1991). Microinjection of propranolol into the dorsal periaqueductal gray causes an anxiolytic effect in the elevated plus-‐maze antagonized by ritanserin. Psychopharmacology, 105(4), 553-‐557.
Avila-‐Rojas, S. H., Velazquez-‐Lagunas, I., Salinas-‐Abarca, A. B., Barragan-‐Iglesias, P., Pineda-‐Farias, J. B., & Granados-‐Soto, V. (2015). Role of spinal 5-‐HT5A, and 5-‐HT1A/1B/1D, receptors in neuropathic pain induced by spinal nerve ligation in rats. Brain research, 1622, 377-‐385.
Aznar, S., Qian, Z., Shah, R., Rahbek, B., & Knudsen, G. M. (2003). The 5-‐HT1A serotonin receptor is located on calbindin-‐and parvalbumin-‐containing neurons in the rat brain. Brain research, 959(1), 58-‐67.
Banks, M. L., Czoty, P. W., Gage, H. D., Bounds, M. C., Garg, P. K., Garg, S., & Nader, M. A. (2008). Effects of cocaine and MDMA self-‐administration on serotonin transporter availability in monkeys. Neuropsychopharmacology, 33(2), 219-‐225.
Bankson, M. G., & Yamamoto, B. K. (2004). Serotonin–GABA interactions modulate MDMA-‐induced mesolimbic dopamine release. Journal of Neurochemistry, 91(4), 852-‐859.
Bard, J. A., Zgombick, J., Adham, N., Vaysse, P., Branchek, T. A., & Weinshank, R. L. (1993). Cloning of a novel human serotonin receptor (5-‐HT7) positively linked to adenylate cyclase. Journal of Biological Chemistry, 268(31), 23422-‐23426.

97
Battaglia, G., Brooks, B. P., Kulsakdinun, C., & De Souza, E. B. (1988). Pharmacologic profile of MDMA (3, 4-‐methylenedioxymethamphetamine) at various brain recognition sites. European journal of pharmacology, 149(1), 159-‐163.
Battaglia, G., Yeh, S., & De Souza, E. B. (1988). MDMA-‐induced neurotoxicity: parameters of degeneration and recovery of brain serotonin neurons. Pharmacology Biochemistry and Behavior, 29(2), 269-‐274.
Baumann, M. H., Clark, R. D., Franken, F. H., Rutter, J. J., & Rothman, R. B. (2008). Tolerance to 3, 4-‐methylenedioxymethamphetamine in rats exposed to single high-‐dose binges. Neuroscience, 152(3), 773-‐784.
Baumann, M. H., Clark, R. D., & Rothman, R. B. (2008). Locomotor stimulation produced by 3, 4-‐methylenedioxymethamphetamine (MDMA) is correlated with dialysate levels of serotonin and dopamine in rat brain. Pharmacology Biochemistry and Behavior, 90(2), 208-‐217.
Baumann, M. H., & Rothman, R. B. (2009). Neural and cardiac toxicities associated with 3, 4-‐methylenedioxymethamphetamine (MDMA). International review of neurobiology, 88, 257-‐296.
Baumann, M. H., Wang, X., & Rothman, R. B. (2007). 3, 4-‐Methylenedioxymethamphetamine (MDMA) neurotoxicity in rats: a reappraisal of past and present findings. Psychopharmacology, 189(4), 407-‐424.
Beardsley, P. M., Balster, R. L., & Harris, L. S. (1986). Self-‐administration of methylenedioxymethamphetamine (MDMA) by rhesus monkeys. Drug and alcohol dependence, 18(2), 149-‐157.
Beck, J., & Rosenbaum, M. (1990). The scheduling of MDMA ('ecstasy'). In J. A. Inciardi (Ed.), Handbook of Drug Control in the United States (pp. 303-‐316). New York: Greenwood.
Belin, D., Belin-‐Rauscent, A., Everitt, B. J., & Dalley, J. W. (2015). In search of predictive endophenotypes in addiction: insights from preclinical research. Genes, Brain and Behavior.
Benjamin, D., Lal, H., & Meyerson, L. R. (1990). The effects of 5-‐HT1B characterizing agents in the mouse elevated plus-‐maze. Life sciences, 47(3), 195-‐203.
Benningfield, M. M., & Cowan, R. L. (2013). Brain serotonin function in MDMA (ecstasy) users: evidence for persisting neurotoxicity. Neuropsychopharmacology, 38(1), 253-‐255. doi:10.1038/npp.2012.178
Benzenhöfer, U., & Passie, T. (2010). Rediscovering MDMA (ecstasy): the role of the American chemist Alexander T. Shulgin. Addiction, 105(8), 1355-‐1361.
Berger, U. V., Gu, X. F., & Azmitia, E. C. (1992). The substituted amphetamines 3, 4-‐methylenedioxymethamphetamine, methamphetamine, p-‐chloroamphetamine and fenfluramine induce 5-‐hydroxytryptamine release via a common mechanism blocked by fluoxetine and cocaine. European journal of pharmacology, 215(2), 153-‐160.
Bhide, N. S., Lipton, J. W., Cunningham, J. I., Yamamoto, B. K., & Gudelsky, G. A. (2009). Repeated exposure to MDMA provides neuroprotection against subsequent MDMA-‐induced serotonin depletion in brain. Brain research, 1286, 32-‐41.
Biezonski, D. K., Courtemanche, A. B., Hong, S. B., Piper, B. J., & Meyer, J. S. (2009). Repeated adolescent MDMA (“Ecstasy”) exposure in rats increases behavioral and neuroendocrine responses to a 5-‐HT2A/2C agonist. Brain research, 1252, 87-‐93.

98
Bird, J., & Schenk, S. (2013). Contribution of impulsivity and novelty-‐seeking to the acquisition and maintenance of MDMA self-‐administration. Addiction Biology, 18(4), 654-‐664.
Bjorvatn, B., & Ursin, R. (1994). Effects of the selective 5-‐HT1B agonist, CGS 12066B, on sleep/waking stages and EEG power spectrum in rats. Journal of sleep research, 3(2), 97-‐105.
Blair, C., Bonardi, C., & Hall, G. (2004). Differential effects of 8-‐OH-‐DPAT on two forms of appetitive pavlovian conditioning in the rat. Behavioral neuroscience, 118(6), 1439.
Bogen, I. L., Haug, K. H., Myhre, O., & Fonnum, F. (2003). Short-‐and long-‐term effects of MDMA (“ecstasy”) on synaptosomal and vesicular uptake of neurotransmitters in vitro and ex vivo. Neurochemistry international, 43(4), 393-‐400.
Bonaventure, P., Schotte, A., Cras, P., & Leysen, J. (1997). Autoradiographic mapping of 5-‐HT1B-‐and 5-‐HT1D receptors in human brain using [3H] alniditan, a new radioligand. Receptors & channels, 5(3-‐4), 225-‐230.
Bonaventure, P., Voorn, P., Luyten, W., Jurzak, M., Schotte, A., & Leysen, J. (1998). Detailed mapping of serotonin 5-‐HT1B and 5-‐HT1D receptor messenger RNA and ligand binding sites in guinea-‐pig brain and trigeminal ganglion: clues for function. Neuroscience, 82(2), 469-‐484.
Booij, L., Soucy, J. P., Young, S. N., Regoli, M., Gravel, P., Diksic, M., . . . Benkelfat, C. (2014). Brain serotonin synthesis in MDMA (ecstasy) polydrug users: an alpha-‐[11C] methyl-‐l-‐tryptophan study. Journal of Neurochemistry, 131(5), 634-‐644.
Bortolozzi, A., Díaz-‐Mataix, L., Scorza, M. C., Celada, P., & Artigas, F. (2005). The activation of 5-‐HT2A receptors in prefrontal cortex enhances dopaminergic activity. Journal of Neurochemistry, 95(6), 1597-‐1607. doi:10.1111/j.1471-‐4159.2005.03485.x
Boulenguez, P., Rawlins, J., Chauveau, J., Joseph, M., Mitchell, S., & Gray, J. (1996). Modulation of dopamine release in the nucleus accumbens by 5-‐HT1B agonists: involvement of the hippocampo-‐accumbens pathway. Neuropharmacology, 35(11), 1521-‐1529.
Bradbury, S., Bird, J., Colussi-‐Mas, J., Mueller, M., Ricaurte, G., & Schenk, S. (2014). Acquisition of MDMA self-‐administration: pharmacokinetic factors and MDMA-‐induced serotonin release. Addiction Biology, 19(5), 874-‐884.
Bradbury, S., Gittings, D., & Schenk, S. (2012). Repeated exposure to MDMA and amphetamine: sensitization, cross-‐sensitization, and response to dopamine D1-‐and D2-‐like agonists. Psychopharmacology, 223(4), 389-‐399.
Brennan, K. A., Carati, C., Lea, R. A., Fitzmaurice, P. S., & Schenk, S. (2009). Effect of D1-‐like and D2-‐like receptor antagonists on methamphetamine and 3, 4-‐methylenedioxymethamphetamine self-‐administration in rats. Behavioural pharmacology, 20(8), 688-‐694.
Brunt, T. M., Koeter, M. W., Niesink, R. J., & van den Brink, W. (2012). Linking the pharmacological content of ecstasy tablets to the subjective experiences of drug users. Psychopharmacology, 220(4), 751-‐762.
Bubar, M. J., & Cunningham, K. A. (2006). Serotonin 5-‐HT2A and 5-‐HT2C receptors as potential targets for modulation of psychostimulant use and dependence. Current topics in medicinal chemistry, 6(18), 1971-‐1985.

99
Bull, E. J., Hutson, P., & Fone, K. (2003). Reduced social interaction following 3, 4-‐methylenedioxymethamphetamine is not associated with enhanced 5-‐HT 2C receptor responsivity. Neuropharmacology, 44(4), 439-‐448.
Bull, E. J., Hutson, P. H., & Fone, K. C. (2004). Decreased social behaviour following 3, 4-‐methylenedioxymethamphetamine (MDMA) is accompanied by changes in 5-‐HT2A receptor responsivity. Neuropharmacology, 46(2), 202-‐210.
Bush, D. E., & Vaccarino, F. J. (2007). Individual differences in elevated plus-‐maze exploration predicted progressive-‐ratio cocaine self-‐administration break points in Wistar rats. Psychopharmacology, 194(2), 211-‐219.
Caggiula, A. R., Donny, E. C., White, A. R., Chaudhri, N., Booth, S., Gharib, M. A., . . . Sved, A. F. (2002). Environmental stimuli promote the acquisition of nicotine self-‐administration in rats. Psychopharmacology, 163(2), 230-‐237.
Callaway, C. W., & Geyer, M. A. (1992). Tolerance and cross-‐tolerance to the activating effects of 3, 4-‐methylenedioxymethamphetamine and a 5-‐hydroxytryptamine1B agonist. Journal of Pharmacology and Experimental Therapeutics, 263(1), 318-‐326.
Carli, M., Baviera, M., Invernizzi, R. W., & Balducci, C. (2006). Dissociable contribution of 5-‐HT1A and 5-‐HT2A receptors in the medial prefrontal cortex to different aspects of executive control such as impulsivity and compulsive perseveration in rats. Neuropsychopharmacology, 31(4), 757-‐767.
Carli, M., Invernizzi, R., Cervo, L., & Samanin, R. (1988). Neurochemical and behavioural studies with RU-‐24969 in the rat. Psychopharmacology, 94(3), 359-‐364.
Carli, M., & Samanin, R. (1992). 8-‐Hydroxy-‐2-‐(di-‐n-‐propylamino) tetralin impairs spatial learning in a water maze: role of postsynaptic 5-‐HT1Areceptors. British journal of pharmacology, 105(3), 720.
Carli, M., & Samanin, R. (2000). The 5-‐HT1A receptor agonist 8-‐OH-‐DPAT reduces rats’ accuracy of attentional performance and enhances impulsive responding in a five-‐choice serial reaction time task: role of presynaptic 5-‐HT1A receptors. Psychopharmacology, 149(3), 259-‐268.
Carli, M., Tranchina, S., & Samanin, R. (1992). 8-‐Hydroxy-‐2-‐(di-‐n-‐propylamino) tetralin, a 5-‐HT1A receptor agonist, impairs performance in a passive avoidance task. European journal of pharmacology, 211(2), 227-‐234.
Carroll, M. E., & Lac, S. T. (1997). Acquisition of iv amphetamine and cocaine self-‐administration in rats as a function of dose. Psychopharmacology, 129(3), 206-‐214.
Carter, L. (2016). Molly overtakes Marijuana in Rap and Hip Hop Music. Recovery Happens Counseling Services. Retrieved from http://www.recoveryhappens.com/Molly_overtakes_weed_in_music_culture.pdf
Cassaday, H. J., Hodges, H., & Gray, J. A. (1993). The effects of ritanserin, RU 24969 and 8-‐OH-‐DPAT on latent inhibition in the rat. Journal of Psychopharmacology, 7(1 suppl), 63-‐71.
Castro, M. E., Diaz, A., del Olmo, E., & Pazos, A. (2003). Chronic fluoxetine induces opposite changes in G protein coupling at pre and postsynaptic 5-‐HT1A receptors in rat brain. Neuropharmacology, 44(1), 93-‐101.
Centurión, D., Sánchez-‐López, A., Ortiz, M. I., De Vries, P., Saxena, P. R., & Villalón, C. M. (2000). Mediation of 5-‐HT-‐induced internal carotid vasodilatation in GR127935-‐and

100
ritanserin-‐pretreated dogs by 5-‐HT7 receptors. Naunyn-‐Schmiedeberg's archives of pharmacology, 362(2), 169-‐176.
Chaouloff, F., Courvoisier, H., Moisan, M., & Mormede, P. (1999). GR 127935 reduces basal locomotor activity and prevents RU 24969-‐, but not D-‐amphetamine-‐induced hyperlocomotion, in the Wistar-‐Kyoto hyperactive (WKHA) rat. Psychopharmacology, 141(3), 326-‐331.
Cheeta, S., Kenny, P. J., & File, S. E. (2000a). Hippocampal and septal injections of nicotine and 8-‐OH-‐DPAT distinguish among different animal tests of anxiety. Progress in Neuro-‐Psychopharmacology and Biological Psychiatry, 24(7), 1053-‐1067.
Cheeta, S., Kenny, P. J., & File, S. E. (2000b). The role of 5-‐HT1A receptors in mediating the anxiogenic effects of nicotine following lateral septal administration. European Journal of Neuroscience, 12(10), 3797-‐3802.
Cheetham, S., & Heal, D. (1993). Evidence that RU 24969-‐induced locomotor activity in C57/B1/6 mice is specifically mediated by the 5-‐HT1B receptor. British journal of pharmacology, 110(4), 1621-‐1629.
Chen, N. H., & Reith, M. E. (1995). Monoamine Interactions Measured by Microdialysis in the Ventral Tegmental Area of Rats Treated Systemically with (±)-‐8-‐Hydroxy-‐2-‐(Di-‐n-‐Propylamino) tetralin. Journal of Neurochemistry, 64(4), 1585-‐1597.
Chennaoui, M., Drogou, C., Gomez-‐Merino, D., Grimaldi, B., Fillion, G., & Guezennec, C. (2001). Endurance training effects on 5-‐HT1B receptors mRNA expression in cerebellum, striatum, frontal cortex and hippocampus of rats. Neuroscience letters, 307(1), 33-‐36.
Clemett, D. A., Punhani, T., Duxon, S., Blackburn, T. P., & Fone, K. C. (2000). Immunohistochemical localisation of the 5-‐HT2C receptor protein in the rat CNS. Neuropharmacology, 39(1), 123-‐132.
Cole, J. C., & Sumnall, H. R. (2003). The pre-‐clinical behavioural pharmacology of 3, 4-‐methylenedioxymethamphetamine (MDMA). Neuroscience & Biobehavioral Reviews, 27(3), 199-‐217.
Collinson, N., & Dawson, G. (1997). On the elevated plus-‐maze the anxiolytic-‐like effects of the 5-‐HT1A agonist, 8-‐OH-‐DPAT, but not the anxiogenic-‐like effects of the 5-‐HT1A partial agonist, buspirone, are blocked by the 5-‐HT1A antagonist, WAY 100635. Psychopharmacology, 132(1), 35-‐43.
Colpaert, F. C. (2006). 5-‐HT (1A) receptor activation: new molecular and neuroadaptive mechanisms of pain relief. Current opinion in investigational drugs (London, England: 2000), 7(1), 40-‐47.
Colpaert, F. C., Tarayre, J., Koek, W., Pauwels, P., Bardin, L., Xu, X.-‐J., . . . Assie, M. (2002). Large-‐amplitude 5-‐HT1A receptor activation: a new mechanism of profound, central analgesia. Neuropharmacology, 43(6), 945-‐958.
Colussi-‐Mas, J., Wise, R. J., Howard, A., & Schenk, S. (2010). Drug seeking in response to a priming injection of MDMA in rats: relationship to initial sensitivity to self-‐administered MDMA and dorsal striatal dopamine. The International Journal of Neuropsychopharmacology, 13(10), 1315-‐1327.
Commins, D., Vosmer, G., Virus, R., Woolverton, W., Schuster, C., & Seiden, L. (1987). Biochemical and histological evidence that methylenedioxymethylamphetamine

101
(MDMA) is toxic to neurons in the rat brain. Journal of Pharmacology and Experimental Therapeutics, 241(1), 338-‐345.
Compan, V., Segu, L., Buhot, M., & Daszuta, A. (1998). Selective increases in serotonin 5-‐HT1B/1D and 5-‐HT2A/2C binding sites in adult rat basal ganglia following lesions of serotonergic neurons. Brain research, 793(1), 103-‐111.
Cottler, L. B., Leung, K. S., & Abdallah, A. B. (2009). Test–re-‐test reliability of DSM-‐IV adopted criteria for 3, 4-‐methylenedioxymethamphetamine (MDMA) abuse and dependence: a cross-‐national study. Addiction, 104(10), 1679-‐1690.
Cottler, L. B., Womack, S. B., Compton, W. M., & Ben-‐Abdallah, A. (2001). Ecstasy abuse and dependence among adolescents and young adults: applicability and reliability of DSM-‐IV criteria. Human Psychopharmacology: Clinical and Experimental, 16(8), 599-‐606.
Cox, B. M., Shah, M. M., Cichon, T., Tancer, M. E., Galloway, M. P., Thomas, D. M., & Perrine, S. A. (2014). Behavioral and neurochemical effects of repeated MDMA administration during late adolescence in the rat. Progress in Neuro-‐Psychopharmacology and Biological Psychiatry, 48, 229-‐235.
Crino, P. B., Vogt, B. A., Volicer, L., & Wiley, R. G. (1990). Cellular localization of serotonin 1A, 1B and uptake sites in cingulate cortex of the rat. Journal of Pharmacology and Experimental Therapeutics, 252(2), 651-‐656.
Critchley, M., & Handley, S. (1987). Effects in the X-‐maze anxiety model of agents acting at 5-‐HT1 and 5-‐HT2 receptors. Psychopharmacology, 93(4), 502-‐506.
Cunningham, K. A., & Anastasio, N. C. (2014). Serotonin at the nexus of impulsivity and cue reactivity in cocaine addiction. Neuropharmacology, 76, 460-‐478.
Dalley, J. W., Fryer, T. D., Brichard, L., Robinson, E. S., Theobald, D. E., Lääne, K., . . . Probst, K. (2007). Nucleus accumbens D2/3 receptors predict trait impulsivity and cocaine reinforcement. Science, 315(5816), 1267-‐1270.
Daniela, E., Brennan, K., Gittings, D., Hely, L., & Schenk, S. (2004). Effect of SCH 23390 on (±)-‐3,4-‐methylenedioxymethamphetamine hyperactivity and self-‐administration in rats. Pharmacology Biochemistry and Behavior, 77(4), 745-‐750. doi:http://dx.doi.org/10.1016/j.pbb.2004.01.008
Darrow, E. J., Strahlendorf, H. K., & Strahlendorf, J. C. (1990). Response of cerebellar Purkinje cells to serotonin and the 5-‐HT1A agonists 8-‐OH-‐DPAT and ipsapirone in vitro. European journal of pharmacology, 175(2), 145-‐153.
Davis, A. K., & Rosenberg, H. (2014). The prevalence, intensity, and assessment of craving for MDMA/Ecstasy in recreational users. Journal of psychoactive drugs, 46(2), 154-‐161.
Davis, W. M., & Smith, S. G. (1976). Role of conditioned reinforcers in the initiation, maintenance and extinction of drug-‐seeking behavior. The Pavlovian journal of biological science: official journal of the Pavlovian, 11(4), 222-‐236.
Dawson, L. A., Hughes, Z. A., Starr, K. R., Storey, J. D., Bettelini, L., Bacchi, F., . . . Hagan, J. J. (2006). Characterisation of the selective 5-‐HT1B receptor antagonist SB-‐616234-‐A (1-‐[6-‐(cis-‐3, 5-‐dimethylpiperazin-‐1-‐yl)-‐2, 3-‐dihydro-‐5-‐methoxyindol-‐1-‐yl]-‐1-‐[2ʹ′-‐methyl-‐4ʹ′-‐(5-‐methyl-‐1, 2, 4-‐oxadiazol-‐3-‐yl) biphenyl-‐4-‐yl] methanone hydrochloride): in vivo neurochemical and behavioural evidence of anxiolytic/antidepressant activity. Neuropharmacology, 50(8), 975-‐983.

102
De Almeida, R. M. M., Giovenardi, M., Charchat, H., & Lucion, A. B. (1998). 8-‐OH-‐DPAT in the median raphe nucleus decreases while in the medial septal area it may increase anxiety in female rats. Neuroscience & Biobehavioral Reviews, 23(2), 259-‐264.
De La Garza, R., 2nd, Fabrizio, K., & Gupta, A. (2007). Relevance of rodent models of intravenous MDMA self-‐administration to human MDMA consumption patterns. Psychopharmacology, 189(4), 425-‐434.
De La Torre, R., Farre, M., Ortuno, J., Mas, M., Brenneisen, R., Roset, P., . . . Cami, J. (2000). Non-‐linear pharmacokinetics of MDMA (‘ecstasy’) in humans. British journal of clinical pharmacology, 49(2), 104-‐109.
De Souza, R., Goodwin, G., Green, A., & Heal, D. (1986). Effect of chronic treatment with 5-‐HT1 agonist (8-‐OH-‐DPAT and RU 24969) and antagonist (isapirone) drugs on the behavioural responses of mice to 5-‐HT1 and 5-‐HT2 agonists. British journal of pharmacology, 89(2), 377.
De Vry, J., Schohe-‐Loop, R., Heine, H.-‐G., Greuel, J. M., Mauler, F., Schmidt, B., . . . Glaser, T. (1998). Characterization of the aminomethylchroman derivative BAY× 3702 as a highly potent 5-‐hydroxytryptamine1A receptor agonist. Journal of Pharmacology and Experimental Therapeutics, 284(3), 1082-‐1094.
De Wit, H. (2009). Impulsivity as a determinant and consequence of drug use: a review of underlying processes. Addiction Biology, 14(1), 22-‐31.
de Wit, H., & Wise, R. A. (1977). Blockade of cocaine reinforcement in rats with the dopamine receptor blocker pimozide, but not with the noradrenergic blockers phentolamine or phenoxybenzamine. Canadian Journal of Psychology/Revue canadienne de psychologie, 31(4), 195.
Degenhardt, L., Barker, B., & Topp, L. (2004). Patterns of ecstasy use in Australia: findings from a national household survey. Addiction, 99(2), 187-‐195.
Dhonnchadha, B. Á. N., & Cunningham, K. A. (2008). Serotonergic mechanisms in addiction-‐related memories. Behavioural brain research, 195(1), 39-‐53.
Di Ciano, P., & Everitt, B. J. (2004). Conditioned reinforcing properties of stimuli paired with self-‐administered cocaine, heroin or sucrose: implications for the persistence of addictive behaviour. Neuropharmacology, 47 Suppl 1, 202-‐213. doi:10.1016/j.neuropharm.2004.06.005
Di Iorio, C. R., Watkins, T. J., Dietrich, M. S., Cao, A., Blackford, J. U., Rogers, B., . . . Kessler, R. M. (2012). Evidence for chronically altered serotonin function in the cerebral cortex of female 3, 4-‐methylenedioxymethamphetamine polydrug users. Archives of general psychiatry, 69(4), 399-‐409.
Di Matteo, V., De Blasi, A., Di Giulio, C., & Esposito, E. (2001). Role of 5-‐HT2C receptors in the control of central dopamine function. Trends in pharmacological sciences, 22(5), 229-‐232.
Díaz-‐Mataix, L., Artigas, F., & Celada, P. (2006). Activation of pyramidal cells in rat medial prefrontal cortex projecting to ventral tegmental area by a 5-‐HT 1A receptor agonist. European Neuropsychopharmacology, 16(4), 288-‐296.
Díaz-‐Mataix, L., Scorza, M. C., Bortolozzi, A., Toth, M., Celada, P., & Artigas, F. (2005). Involvement of 5-‐HT1A receptors in prefrontal cortex in the modulation of dopaminergic activity: role in atypical antipsychotic action. The Journal of neuroscience, 25(47), 10831-‐10843.

103
Diergaarde, L., Pattij, T., Poortvliet, I., Hogenboom, F., de Vries, W., Schoffelmeer, A. N., & De Vries, T. J. (2008). Impulsive choice and impulsive action predict vulnerability to distinct stages of nicotine seeking in rats. Biological Psychiatry, 63(3), 301-‐308.
Dilleen, R., Pelloux, Y., Mar, A. C., Molander, A., Robbins, T. W., Everitt, B. J., . . . Belin, D. (2012). High anxiety is a predisposing endophenotype for loss of control over cocaine, but not heroin, self-‐administration in rats. Psychopharmacology, 222(1), 89-‐97.
Do, J., & Schenk, S. (2011). Self-‐administered MDMA produces dose-‐and time-‐dependent serotonin deficits in the rat brain. Addiction Biology.
Doherty, M. D., & Pickel, V. M. (2001). Targeting of serotonin 1A receptors to dopaminergic neurons within the parabrachial subdivision of the ventral tegmental area in rat brain. Journal of Comparative Neurology, 433(3), 390-‐400.
Domenech, T., Beleta, J., & Palacios, J. (1997). Characterization of human serotonin 1D and 1B receptors using [3H]-‐GR-‐125743, a novel radiolabelled serotonin 5HT1D/1B receptor antagonist. Naunyn-‐Schmiedeberg's archives of pharmacology, 356(3), 328-‐334.
Duncan, M. J., Hester, J. M., Hopper, J. A., & Franklin, K. M. (2010). The effects of aging and chronic fluoxetine treatment on circadian rhythms and suprachiasmatic nucleus expression of neuropeptide genes and 5-‐HT1B receptors. European Journal of Neuroscience, 31(9), 1646-‐1654.
Dworkin, S. I., Mirkis, S., & Smith, J. E. (1995). Response-‐dependent versus response-‐independent presentation of cocaine: differences in the lethal effects of the drug. Psychopharmacology, 117(3), 262-‐266.
Eberle-‐Wang, K., Mikeladze, Z., Uryu, K., & Chesselet, M. F. (1997). Pattern of expression of the serotonin2C receptor messenger RNA in the basal ganglia of adult rats. Journal of Comparative Neurology, 384(2), 233-‐247.
Eison, A. S., & Mullins, U. L. (1995). Regulation of central 5-‐HT2A receptors: a review of in vivo studies. Behavioural brain research, 73(1), 177-‐181.
Ellenbroek, B., Knobbout, D., & Cools, A. (1997). The role of mesolimbic and nigrostriatal dopamine in latent inhibition as measured with the conditioned taste aversion paradigm. Psychopharmacology, 129(2), 112-‐120.
Erickson, J. D., Schafer, M., Bonner, T. I., Eiden, L. E., & Weihe, E. (1996). Distinct pharmacological properties and distribution in neurons and endocrine cells of two isoforms of the human vesicular monoamine transporter. Proceedings of the National Academy of Sciences, 93(10), 5166-‐5171.
Ettenberg, A., Pettit, H. O., Bloom, F. E., & Koob, G. F. (1982). Heroin and cocaine intravenous self-‐administration in rats: mediation by separate neural systems. Psychopharmacology, 78(3), 204-‐209.
Evenden, J. L. (1999a). The pharmacology of impulsive behaviour in rats VII: the effects of serotonergic agonists and antagonists on responding under a discrimination task using unreliable visual stimuli. Psychopharmacology, 146(4), 422-‐431.
Evenden, J. L. (1999b). Varieties of impulsivity. Psychopharmacology, 146(4), 348-‐361. Fantegrossi, W. E. (2007). Reinforcing effects of methylenedioxy amphetamine congeners in
rhesus monkeys: are intravenous self-‐administration experiments relevant to MDMA neurotoxicity? Psychopharmacology, 189(4), 471-‐482.
Fernández, V. M., Giurfa, M., Devaud, J.-‐M., & Farina, W. M. (2012). Latent inhibition in an insect: The role of aminergic signaling. Learning & memory, 19(12), 593-‐597.

104
Ferrari, M. C., & Chivers, D. P. (2011). Learning about non-‐predators and safe places: the forgotten elements of risk assessment. Animal cognition, 14(3), 309-‐316.
File, S. E., & Gonzalez, L. E. (1996). Anxiolytic effects in the plus-‐maze of 5-‐HT1A-‐receptor ligands in dorsal raphe and ventral hippocampus. Pharmacology Biochemistry and Behavior, 54(1), 123-‐128.
File, S. E., Gonzalez, L. E., & Andrews, N. (1996). Comparative Study of Pre-‐and Postsynaptic 5-‐HT1A Receptor Modulation of Anxiety in Two Ethological Animal Tests. The Journal of neuroscience, 16(15), 4810-‐4815.
File, S. E., Kenny, P. J., & Cheeta, S. (2000). The role of the dorsal hippocampal serotonergic and cholinergic systems in the modulation of anxiety. Pharmacology Biochemistry and Behavior, 66(1), 65-‐72.
Fitzgerald, R. L., Blanke, R. V., & Poklis, A. (1990). Stereoselective pharmacokinetics of 3, 4-‐methylenedioxymethamphetamine in the rat. Chirality, 2(4), 241-‐248.
Fletcher, A., Forster, E. A., Bill, D. J., Brown, G., Cliffe, I. A., Hartley, J. E., . . . Critchley, D. J. (1995). Electrophysiological, biochemical, neurohormonal and behavioural studies with WAY-‐100635, a potent, selective and silent 5-‐HT1A receptor antagonist. Behavioural brain research, 73(1), 337-‐353.
Fletcher, P. J., Azampanah, A., & Korth, K. M. (2002). Activation of 5-‐HT1B receptors in the nucleus accumbens reduces self-‐administration of amphetamine on a progressive ratio schedule. Pharmacology Biochemistry and Behavior, 71(4), 717-‐725.
Fletcher, P. J., Chintoh, A. F., Sinyard, J., & Higgins, G. A. (2004). Injection of the 5-‐HT2C receptor agonist Ro60-‐0175 into the ventral tegmental area reduces cocaine-‐induced locomotor activity and cocaine self-‐administration. Neuropsychopharmacology.
Fletcher, P. J., & Korth, K. (1999a). Activation of 5-‐HT1B receptors in the nucleus accumbens reduces amphetamine-‐induced enhancement of responding for conditioned reward. Psychopharmacology, 142(2), 165-‐174.
Fletcher, P. J., & Korth, K. (1999b). RU-‐24969 disrupts d-‐amphetamine self-‐administration and responding for conditioned reward via stimulation of 5-‐HT1B receptors. Behavioural pharmacology, 10(2), 183-‐193.
Fletcher, P. J., Rizos, Z., Sinyard, J., Tampakeras, M., & Higgins, G. A. (2007). The 5-‐HT2C receptor agonist Ro60-‐0175 reduces cocaine self-‐administration and reinstatement induced by the stressor yohimbine, and contextual cues. Neuropsychopharmacology, 33(6), 1402-‐1412.
Fletcher, P. J., Tampakeras, M., Sinyard, J., & Higgins, G. A. (2007). Opposing effects of 5-‐HT2A and 5-‐HT2C receptor antagonists in the rat and mouse on premature responding in the five-‐choice serial reaction time test. Psychopharmacology, 195(2), 223-‐234.
Food and Drug Administration. (2005). Guidance for Industry Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers. Retrieved from Rockville, MD:
Fox, B. S., Kantak, K. M., Edwards, M. A., Black, K. M., Bollinger, B. K., Botka, A. J., . . . Greenstein, J. L. (1996). Efficacy of a therapeutic cocaine vaccine in rodent models. Nature medicine, 2(10), 1129-‐1132.
Frances, H., & Monier, C. (1991). Tolerance to the behavioural effect of serotonergic (5-‐HT1B) agonists in the isolationinduced social behavioural deficit test. Neuropharmacology, 30(6), 623-‐627.

105
Frankfurt, M., Mendelson, S. D., McKittrick, C. R., & McEwen, B. S. (1993). Alterations of serotonin receptor binding in the hypothalamus following acute denervation. Brain research, 601(1), 349-‐352.
Freudenmann, R. W., Öxler, F., & Bernschneider-‐Reif, S. (2006). The origin of MDMA (ecstasy) revisited: the true story reconstructed from the original documents*. Addiction, 101(9), 1241-‐1245.
Frick, L. R., Bernardez-‐Vidal, M., Hocht, C., Zanutto, B. S., & Rapanelli, M. (2015). Dual role of serotonin in the acquisition and extinction of reward-‐driven learning: Involvement of 5-‐HT1A, 5-‐HT2A and 5-‐HT3 receptors. Behavioural brain research, 277, 193-‐203.
Fuss, J., Vogt, M. A., Weber, K. J., Burke, T. F., Gass, P., & Hensler, J. G. (2013). Hippocampal serotonin-‐1A receptor function in a mouse model of anxiety induced by long-‐term voluntary wheel running. Synapse, 67(10), 648-‐655.
Galloway, M. P., Suchowski, C. S., Keegan, M. J., & Hjorth, S. (1993). Local infusion of the selective 5HT-‐1B agonist CP-‐93,129 facilitates striatal dopamine release in vivo. Synapse, 15(1), 90-‐92.
García-‐Cabrerizo, R., & García-‐Fuster, M. J. (2015). Chronic MDMA induces neurochemical changes in the hippocampus of adolescent and young adult rats: Down-‐regulation of apoptotic markers. Neurotoxicology, 49, 104-‐113.
Gardner, E. L. (2000). What We Have Learned about Addiction from Animal Models of Drug Self-‐Administration. The American Journal on Addictions, 9(4), 285-‐313.
Gaston, T. R., & Rasmussen, G. T. (1972). Identification of 3, 4-‐methylenedioxymethamphetamine. Microgram, 5, 60.
Geyer, M. A., & Ellenbroek, B. (2003). Animal behavior models of the mechanisms underlying antipsychotic atypicality. Progress in Neuro-‐Psychopharmacology and Biological Psychiatry, 27(7), 1071-‐1079.
Geyer, M. A., Krebs-‐Thomson, K., & Varty, G. B. (1999). The effects of M100907 in pharmacological and developmental animal models of prepulse inhibition deficits in schizophrenia. Neuropsychopharmacology, 21, S134-‐S142.
Global Drug Survey. (2014). Last 12 month prevalence of top 20 drugs. Retrieved from London:
Granoff, M. I., & Ashby, C. R. J. (2001). Effect of the repeated administration of (±)-‐3, 4-‐methylenedioxymethamphetamine on the behavioral response of rats to the 5-‐HT1A receptor agonist (±)-‐8-‐hydroxy-‐(di-‐n-‐propylamino) tetralin. Neuropsychobiology, 43(1), 42-‐48.
Granoff, M. I., & Ashby Jr, C. R. (1998). The Effect of the Repeated Administration of the Compound 3, 4-‐Methylenedioxymethamphetamineon the Response of Rats to the5-‐HT2A/C Receptor Agonist (±)-‐1-‐(2, 5-‐dimethoxy-‐4-‐iodophenyl)-‐2-‐aminopropane (DOI). Neuropsychobiology, 37(1), 36-‐40.
Green, A. R., Mechan, A. O., Elliott, J. M., O'Shea, E., & Colado, M. I. (2003). The pharmacology and clinical pharmacology of 3, 4-‐methylenedioxymethamphetamine (MDMA,“ecstasy”). Pharmacological reviews, 55(3), 463-‐508.
Gronier, B. (2008). Involvement of glutamate neurotransmission and N-‐methyl-‐d-‐aspartate receptor in the activation of midbrain dopamine neurons by 5-‐HT1A receptor agonists: an electrophysiological study in the rat. Neuroscience, 156(4), 995-‐1004.

106
Grottick, A. J., Corrigall, W., & Higgins, G. (2001). Activation of 5-‐HT2C receptors reduces the locomotor and rewarding effects of nicotine. Psychopharmacology, 157(3), 292-‐298.
Grottick, A. J., Fletcher, P. J., & Higgins, G. A. (2000). Studies to investigate the role of 5-‐HT2C receptors on cocaine-‐and food-‐maintained behavior. Journal of Pharmacology and Experimental Therapeutics, 295(3), 1183-‐1191.
Gu, X. F., & Azmitia, E. C. (1993). Integrative transporter-‐mediated release from cytoplasmic and vesicular 5-‐hydroxytryptamine stores in cultured neurons. European journal of pharmacology, 235(1), 51-‐57.
Gudelsky, G. A., & Nash, J. F. (1996). Carrier-‐Mediated Release of Serotonin by 3, 4-‐Methylenedioxymethamphetamine: Implications for Serotonin-‐Dopamine Interactions. Journal of Neurochemistry, 66(1), 243-‐249.
Gudelsky, G. A., Yamamoto, B. K., & Nash, J. F. (1994). Potentiation of 3, 4-‐methylenedioxymethamphetamine-‐induced dopamine release and serotonin neurotoxicity by 5-‐HT2 receptor agonists. European journal of pharmacology, 264(3), 325-‐330.
Gui, Z., Zhang, Q., Liu, J., Zhang, L., Ali, U., Hou, C., . . . Hui, Y. (2011). Unilateral lesion of the nigrostriatal pathway decreases the response of fast-‐spiking interneurons in the medial prefrontal cortex to 5-‐HT1A receptor agonist and expression of the receptor in parvalbumin-‐positive neurons in the rat. Neurochemistry international, 59(5), 618-‐627.
Hajós, M., Gartside, S. E., Varga, V., & Sharp, T. (2003). In vivo inhibition of neuronal activity in the rat ventromedial prefrontal cortex by midbrain-‐raphe nuclei: role of 5-‐HT 1A receptors. Neuropharmacology, 45(1), 72-‐81.
Hållbus, M., Magnusson, T., & Magnusson, O. (1997). Influence of 5-‐HT1B/1D receptors on dopamine release in the guinea pig nucleus accumbens: a microdialysis study. Neuroscience letters, 225(1), 57-‐60.
Hamon, M., Cossery, J., Spampinato, U., & Gozlan, H. (1986). Are there selective ligands for 5-‐HT1A and 5-‐HT1B receptor binding sites in brain? Trends in pharmacological sciences, 7, 336-‐338.
Hamon, M., Fattaccini, C., Adrien, J., Gallissot, M., Martin, P., & Gozlan, H. (1988). Alterations of central serotonin and dopamine turnover in rats treated with ipsapirone and other 5-‐hydroxytryptamine1A agonists with potential anxiolytic properties. Journal of Pharmacology and Experimental Therapeutics, 246(2), 745-‐752.
Hamon, M., Gozlan, H., Mestikawy, S., Emerit, M., Bolanos, F., & Schechter, L. (1990). The Central 5-‐HT1A Receptors: Pharmacological, Biochemical, Functional, and Regulatory Properties. Annals of the New York Academy of Sciences, 600(1), 114-‐129.
Hansen, D., Maycock, B., & Lower, T. (2001). ‘Weddings, parties, anything…’, a qualitative analysis of ecstasy use in Perth, Western Australia. International Journal of Drug Policy, 12(2), 181-‐199.
Hansen, J. P., Riddle, E. L., Sandoval, V., Brown, J. M., Gibb, J. W., Hanson, G. R., & Fleckenstein, A. E. (2002). Methylenedioxymethamphetamine decreases plasmalemmal and vesicular dopamine transport: mechanisms and implications for neurotoxicity. Journal of Pharmacology and Experimental Therapeutics, 300(3), 1093-‐1100.

107
Hardman, H. F., Haavik, C. O., & Seevers, M. H. (1973). Relationship of the structure of mescaline and seven analogs to toxicity and behavior in five species of laboratory animals. Toxicology and applied Pharmacology, 25(2), 299-‐309.
Hartig, P. R., Branchek, T. A., & Weinshank, R. L. (1992). A subfamily of 5-‐HT1D receptor genes. Trends in pharmacological sciences, 13, 152-‐159.
Heal, D. J., Philpot, J., Molyneux, S. G., & Metz, A. (1985). Intracerebroventricular administration of 5,7-‐dihydroxytryptamine to mice increases both head-‐twitch response and the number of cortical 5-‐HT2 receptors. Neuropharmacology, 24(12), 1201-‐1205.
Heidbreder, C. A., & Hagan, J. J. (2005). Novel pharmacotherapeutic approaches for the treatment of drug addiction and craving. Current opinion in pharmacology, 5(1), 107-‐118.
Hekmatpanah, C. R., & Peroutka, S. J. (1990). 5-‐Hydroxytryptamine uptake blockers attenuate the 5-‐hydroxytryptamine-‐releasing effect of 3, 4-‐methylenedioxymethamphetamine and related agents. European journal of pharmacology, 177(1), 95-‐98.
Hensler, J. G. (2003). Regulation of 5-‐HT1A receptor function in brain following agonist or antidepressant administration. Life sciences, 72(15), 1665-‐1682.
Hensler, J. G., Kovachich, G. B., & Frazer, A. (1991). A quantitative autoradiographic study of serotonin1A receptor regulation: effect of 5, 7-‐dihydroxytryptamine and antidepressant treatments. Neuropsychopharmacology.
Hensler, J. G., Vogt, M. A., & Gass, P. (2010). Regulation of cortical and hippocampal 5-‐HT 1A receptor function by corticosterone in GR+/− mice. Psychoneuroendocrinology, 35(3), 469-‐474.
Hillegaart, V., Ahlenius, S., & Larsson, K. (1991). Region-‐selective inhibition of male rat sexual behavior and motor performance by localized forebrain 5-‐HT injections: a comparison with effects produced by 8-‐OH-‐DPAT. Behavioural brain research, 42(2), 169-‐180.
Hillegaart, V., Estival, A., & Ahlenius, S. (1996). Evidence for specific involvement of 5-‐HT1A and 5-‐HT2A/C receptors in the expression of patterns of spontaneous motor activity of the rat. European journal of pharmacology, 295(2), 155-‐161.
Hiroi, R., & Neumaier, J. F. (2009). Estrogen decreases 5-‐HT1B autoreceptor mRNA in selective subregion of rat dorsal raphe nucleus: inverse association between gene expression and anxiety behavior in the open field. Neuroscience, 158(2), 456-‐464.
Hirst, W. D., Andree, T. H., Aschmies, S., Childers, W. E., Comery, T. A., Dawson, L. A., . . . Harrison, B. L. (2008). Correlating efficacy in rodent cognition models with in vivo 5-‐hydroxytryptamine1A receptor occupancy by a novel antagonist,(R)-‐N-‐(2-‐methyl-‐(4-‐indolyl-‐1-‐piperazinyl) ethyl)-‐N-‐(2-‐pyridinyl)-‐cyclohexane carboxamide (WAY-‐101405). Journal of Pharmacology and Experimental Therapeutics, 325(1), 134-‐145.
Hjorth, S., & Magnusson, T. (1988). The 5-‐HT1A receptor agonist, 8-‐OH-‐DPAT, preferentially activates cell body 5-‐HT autoreceptors in rat brain in vivo. Naunyn-‐Schmiedeberg's archives of pharmacology, 338(5), 463-‐471.
Homberg, J. R., De Boer, S. F., Raasø, H. S., Olivier, J. D., Verheul, M., Ronken, E., . . . Vanderschuren, L. J. (2008). Adaptations in pre-‐and postsynaptic 5-‐HT1A receptor function and cocaine supersensitivity in serotonin transporter knockout rats. Psychopharmacology, 200(3), 367-‐380.

108
Homberg, J. R., Van Den Akker, M., Raasø, H. S., Wardeh, G., Binnekade, R., Schoffelmeer, A. N., & De Vries, T. J. (2002). Enhanced motivation to self-‐administer cocaine is predicted by self-‐grooming behaviour and relates to dopamine release in the rat medial prefrontal cortex and amygdala. European Journal of Neuroscience, 15(9), 1542-‐1550.
Hoplight, B., Vincow, E., & Neumaier, J. (2007). Cocaine increases 5-‐HT1B mRNA in rat nucleus accumbens shell neurons. Neuropharmacology, 52(2), 444-‐449.
Hopper, J. W., Su, Z., Looby, A. R., Ryan, E. T., Penetar, D. M., Palmer, C. M., & Lukas, S. E. (2006). Incidence and patterns of polydrug use and craving for ecstasy in regular ecstasy users: An ecological momentary assessment study. Drug and alcohol dependence, 85(3), 221-‐235.
Howell, L. L., & Cunningham, K. A. (2015). Serotonin 5-‐HT2 receptor interactions with dopamine function: implications for therapeutics in cocaine use disorder. Pharmacological reviews, 67(1), 176-‐197.
Hoyer, D., Clarke, D. E., Fozard, J. R., Hartig, P., Martin, G. R., Mylecharane, E. J., . . . Humphrey, P. (1994). International Union of Pharmacology classification of receptors for 5-‐hydroxytryptamine (Serotonin). Pharmacological reviews, 46(2), 157-‐203.
Hoyer, D., & Middlemiss, D. N. (1989). Species differences in the pharmacology of terminal 5-‐HT autoreceptors in mammalian brain. Trends in pharmacological sciences, 10(4), 130-‐132.
Hume, S., Hirani, E., Opacka-‐Juffry, J., Myers, R., Townsend, C., Pike, V., & Grasby, P. (2001). Effect of 5-‐HT on binding of [11C] WAY 100635 to 5-‐HT1A receptors in rat brain, assessed using in vivo microdialysis and PET after fenfluramine. Synapse, 41(2), 150-‐159.
Ichikawa, J., Kuroki, T., Kitchen, M. T., & Meltzer, H. Y. (1995). R (+)-‐8-‐OH-‐DPAT, a 5-‐HT1A receptor agonist, inhibits amphetamine-‐induced dopamine release in rat striatum and nucleus accumbens. European journal of pharmacology, 287(2), 179-‐184.
Ichikawa, J., & Meltzer, H. Y. (2000). The effect of serotonin1A receptor agonism on antipsychotic drug-‐induced dopamine release in rat striatum and nucleus accumbens. Brain research, 858(2), 252-‐263.
Innis, R. B., Nestler, E. J., & Aghajanian, G. K. (1988). Evidence for G protein mediation of serotonin-‐and GABA B-‐induced hyperpolarization of rat dorsal raphe neurons. Brain research, 459(1), 27-‐36.
Ipser, J. C., Wilson, D., Akindipe, T. O., Sager, C., & Stein, D. J. (2015). Pharmacotherapy for anxiety and comorbid alcohol use disorders. status and date: Edited (no change to conclusions), published in(1).
Iyer, R. N., & Bradberry, C. W. (1996). Serotonin-‐mediated increase in prefrontal cortex dopamine release: pharmacological characterization. Journal of Pharmacology and Experimental Therapeutics, 277(1), 40-‐47.
Iyo, A. H., Kieran, N., Chandran, A., Albert, P. R., Wicks, I., Bissette, G., & Austin, M. C. (2009). Differential regulation of the serotonin 1 A transcriptional modulators five prime repressor element under dual repression-‐1 and nuclear-‐deformed epidermal autoregulatory factor by chronic stress. Neuroscience, 163(4), 1119-‐1127.
Jansen, K. L. (1999). Ecstasy (MDMA) dependence. Drug and alcohol dependence, 53(2), 121-‐124.

109
Jensen, K. F., Olin, J., Haykal-‐Coates, N., O’Callaghan, J., Miller, D. B., & de Olmos, J. S. (1993). Mapping toxicant-‐induced nervous system damage with a cupric silver stain: a quantitative analysis of neural degeneration induced by 3, 4-‐methylenedioxymethamphetamine. NIDA Res Monogr, 136, 133-‐149.
Ji, S.-‐P., Zhang, Y., Van Cleemput, J., Jiang, W., Liao, M., Li, L., . . . Zhang, X. (2006). Disruption of PTEN coupling with 5-‐HT2C receptors suppresses behavioral responses induced by drugs of abuse. Nature medicine, 12(3), 324-‐329.
Johnson, S. W., Mercuri, N. B., & North, R. (1992). 5-‐hydroxytryptamine1B receptors block the GABAB synaptic potential in rat dopamine neurons. The Journal of neuroscience, 12(5), 2000-‐2006.
Jones, K., Brennan, K., Colussi-‐Mas, J., & Schenk, S. (2010). Tolerance to 3, 4-‐methylenedioxymethamphetamine is associated with impaired serotonin release. Addiction Biology, 15(3), 289.
Kalivas, P. W., Duffy, P., & White, S. R. (1998). MDMA elicits behavioral and neurochemical sensitization in rats. Neuropsychopharmacology, 18(6), 469-‐479.
Kalkman, H. O. (1995). RU 24969-‐induced locomotion in rats is mediated by 5-‐HT1A receptors. Naunyn-‐Schmiedeberg's archives of pharmacology, 352(5), 583-‐584.
Kalkman, H. O., & Soar, J. (1990). Determination of the 5-‐HT receptor subtype involved in 8-‐OH-‐DPAT-‐induced hyperlocomotion: potential difficulties arising from inadequate pharmacological tools. European journal of pharmacology, 191(3), 383-‐390.
Kantak, K. M. (2003). Vaccines against drugs of abuse. Drugs, 63(4), 341-‐352. Kaplan, E. L., & Meier, P. (1958). Nonparametric estimation from incomplete observations.
Journal of the American statistical association, 53(282), 457-‐481. Kennett, G., Dourish, C., & Curzon, G. (1987). 5-‐HT1B agonists induce anorexia at a
postsynaptic site. European journal of pharmacology, 141(3), 429-‐435. Kia, H. K., Miquel, M. C., Brisorgueil, M. J., Daval, G., Riad, M., Mestikawy, S. E., . . . Verge, D.
(1996). Immunocytochemical localization of serotonin1A receptors in the rat central nervous system. Journal of Comparative Neurology, 365(2), 289-‐305.
Killcross, A., Stanhope, K., Dourish, C., & Piras, G. (1997). WAY100635 and latent inhibition in the rat: selective effects at preexposure. Behavioural brain research, 88(1), 51-‐57.
Kindlundh-‐Högberg, A. M., Svenningsson, P., & Schiöth, H. B. (2006). Quantitative mapping shows that serotonin rather than dopamine receptor mRNA expressions are affected after repeated intermittent administration of MDMA in rat brain. Neuropharmacology, 51(4), 838-‐847.
Kindlundh, A., Lindblom, J., Bergström, L., & Nyberg, F. (2003). The anabolic–androgenic steroid nandrolone induces alterations in the density of serotonergic 5HT1B and 5HT2 receptors in the male rat brain. Neuroscience, 119(1), 113-‐120.
Kirisci, L., Tarter, R., Mezzich, A., & Vanyukov, M. (2007). Developmental trajectory classes in substance use disorder etiology. Psychology of Addictive Behaviors, 21(3), 287.
Kirkpatrick, M. G., Baggott, M. J., Mendelson, J. E., Galloway, G. P., Liechti, M. E., Hysek, C. M., & de Wit, H. (2014). MDMA effects consistent across laboratories. Psychopharmacology, 231(19), 3899-‐3905.
Kish, S. J., Furukawa, Y., Ang, L., Vorce, S., & Kalasinsky, K. (2000). Striatal serotonin is depleted in brain of a human MDMA (Ecstasy) user. Neurology, 55(2), 294-‐296.

110
Kish, S. J., Lerch, J., Furukawa, Y., Tong, J., McCluskey, T., Wilkins, D., . . . Wilson, A. A. (2010). Decreased cerebral cortical serotonin transporter binding in ecstasy users: a positron emission tomography/[11C] DASB and structural brain imaging study. Brain, 133(6), 1779-‐1797.
Koe, B. K., Nielsen, J. A., Macor, J. E., & Heym, J. (1992). Biochemical and behavioral studies of the 5-‐HT1B receptor agonist, CP-‐94,253. Drug development research, 26(3), 241-‐250.
Koffarnus, M. N., & Woods, J. H. (2013). Individual differences in discount rate are associated with demand for self-‐administered cocaine, but not sucrose. Addiction Biology, 18(1), 8-‐18.
Koskinen, T., Haapalinna, A., & Sirvi, J. (2003). α-‐Adrenoceptor-‐Mediated Modulation of 5-‐HT2 Receptor Agonist Induced Impulsive Responding in a 5-‐Choice Serial Reaction Time Task. Pharmacology & toxicology, 92(5), 214-‐225.
Koskinen, T., Ruotsalainen, S., Puumala, T., Lappalainen, R., Koivisto, E., Männistö, P. T., & Sirviö, J. (2000). Activation of 5-‐HT 2A receptors impairs response control of rats in a five-‐choice serial reaction time task. Neuropharmacology, 39(3), 471-‐481.
Koskinen, T., Ruotsalainen, S., & Sirviö, J. (2000). The 5-‐HT 2 receptor activation enhances impulsive responding without increasing motor activity in rats. Pharmacology Biochemistry and Behavior, 66(4), 729-‐738.
Koskinen, T., & Sirviö, J. (2001). Studies on the involvement of the dopaminergic system in the 5-‐HT 2 agonist (DOI)-‐induced premature responding in a five-‐choice serial reaction time task. Brain research bulletin, 54(1), 65-‐75.
Kupferschmidt, K. (2014). Can ecstasy treat the agony of PTSD? Science, 345(6192), 22-‐23. doi:10.1126/science.345.6192.22
Kuroki, T., Ichikawa, J., Dai, J., & Meltzer, H. Y. (1996). R (+)-‐8-‐OH-‐DPAT, a 5-‐HT1A receptor agonist, inhibits amphetamine-‐induced serotonin and dopamine release in rat medial prefrontal cortex. Brain research, 743(1), 357-‐361.
Kwieciński, A., & Nowak, P. (2009). Gestational manganese intoxication and anxiolytic-‐like effects of diazepam and the 5-‐HT1A receptor agonist 8-‐OH-‐DPAT in male Wistar rats. Pharmacological Reports, 61(6), 1061-‐1068.
Lalonde, R., & Strazielle, C. (2010). Relations between open-‐field, elevated plus-‐maze, and emergence tests in C57BL/6J and BALB/c mice injected with GABA-‐and 5HT-‐anxiolytic agents. Fundamental & clinical pharmacology, 24(3), 365-‐376.
Lanteri, C., Doucet, E., Vallejo, S. H., Godeheu, G., Bobadilla, A., Salomon, L., . . . Tassin, J. (2014). Repeated exposure to MDMA triggers long-‐term plasticity of noradrenergic and serotonergic neurons. Molecular psychiatry, 19(7), 823-‐833.
Lawrence, J. A., Olverman, H. J., Shirakawa, K., Kelly, J. S., & Butcher, S. P. (1993). Binding of 5-‐HT1A receptor and 5-‐HT transporter ligands in rat cortex and hippocampus following cholinergic and serotonergic lesions. Brain research, 612(1-‐2), 326-‐329.
Le Poul, E., Boni, C., Hanoun, N. m., Laporte, A.-‐M., Laaris, N., Chauveau, J., . . . Lanfumey, L. (2000). Differential adaptation of brain 5-‐HT1A and 5-‐HT1B receptors and 5-‐HT transporter in rats treated chronically with fluoxetine. Neuropharmacology, 39(1), 110-‐122.
Lejuez, C., Zvolensky, M. J., Daughters, S. B., Bornovalova, M. A., Paulson, A., Tull, M. T., . . . Otto, M. W. (2008). Anxiety sensitivity: A unique predictor of dropout among inner-‐

111
city heroin and crack/cocaine users in residential substance use treatment. Behaviour research and therapy, 46(7), 811-‐818.
Leonardi, E. T. K., & Azmitia, E. C. (1994). MDMA (ecstasy) inhibition of MAO type A and type B: comparisons with fenfluramine and fluoxetine (Prozac). Neuropsychopharmacology, 10(4), 231-‐238.
Lin, D., & Parsons, L. H. (2002). Anxiogenic-‐like effect of serotonin1B receptor stimulation in the rat elevated plus-‐maze. Pharmacology Biochemistry and Behavior, 71(4), 581-‐587.
Lindhe, Ö., Almqvist, P., Kågedal, M., Gustafsson, S.-‐Å., Bergström, M., Nilsson, D., & Antoni, G. (2011). Autoradiographic Mapping of 5-‐H T 1 B/1 D Binding Sites in the Rhesus Monkey Brain Using [carbonyl-‐11 C] zolmitriptan. International journal of molecular imaging, 2011.
Lladó-‐Pelfort, L., Assié, M.-‐B., Newman-‐Tancredi, A., Artigas, F., & Celada, P. (2012). In vivo electrophysiological and neurochemical effects of the selective 5-‐HT1A receptor agonist, F13640, at pre-‐and postsynaptic 5-‐HT1A receptors in the rat. Psychopharmacology, 221(2), 261-‐272.
Lladó-‐Pelfort, L., Assié, M. B., Newman-‐Tancredi, A., Artigas, F., & Celada, P. (2010). Preferential in vivo action of F15599, a novel 5-‐HT1A receptor agonist, at postsynaptic 5-‐HT1A receptors. British journal of pharmacology, 160(8), 1929-‐1940.
Loh, E. A., & Roberts, D. C. (1990). Break-‐points on a progressive ratio schedule reinforced by intravenous cocaine increase following depletion of forebrain serotonin. Psychopharmacology, 101(2), 262-‐266.
Loos, M., Staal, J., Smit, A. B., De Vries, T. J., & Spijker, S. (2013). Enhanced alcohol self-‐administration and reinstatement in a highly impulsive, inattentive recombinant inbred mouse strain. Frontiers in Behavioral Neuroscience, 7. doi:10.3389/fnbeh.2013.00151
Loskutova, L. (2001). The effects of a serotoninergic substrate of the nucleus accumbens on latent inhibition. Neuroscience and Behavioral Physiology, 31(1), 15-‐20.
Loskutova, L., Luk'yanenko, F. Y., & Il'yuchenok, R. Y. (1990). Interaction of serotonin-‐and dopaminergic systems of the brain in mechanisms of latent inhibition in rats. Neuroscience and Behavioral Physiology, 20(6), 500-‐505.
Lovenberg, T. W., Baron, B. M., de Lecea, L., Miller, J. D., Prosser, R. A., Rea, M. A., . . . Siegel, B. W. (1993). A novel adenylyl cyclase-‐activating serotonin receptor (5-‐HT7) implicated in the regulation of mammalian circadian rhythms. Neuron, 11(3), 449-‐458.
Lubow, R. E. (1989). Latent inhibition and conditioned attention theory (Vol. 9): Cambridge University Press.
Lucki, I., Singh, A., & Kreiss, D. S. (1994). Antidepressant-‐like behavioral effects of serotonin receptor agonists. Neuroscience & Biobehavioral Reviews, 18(1), 85-‐95.
Lyness, W., Friedle, N., & Moore, K. (1979). Destruction of dopaminergic nerve terminals in nucleus accumbens: effect on d-‐amphetamine self-‐administration. Pharmacology Biochemistry and Behavior, 11(5), 553-‐556.
Macor, J. E., Burkhart, C. A., Heym, J. H., Ives, J. L., Lebel, L. A., Newman, M. E., . . . Schulz, D. W. (1990). 3-‐(1, 2, 5, 6-‐Tetrahydropyrid-‐4-‐yl) pyrrolo [3, 2-‐b] pyrid-‐5-‐one: a potent and selective serotonin (5-‐HT1B) agonist and rotationally restricted phenolic analog of

112
5-‐methoxy-‐3-‐(1, 2, 5, 6-‐tetrahydropyrid-‐4-‐yl) indole. Journal of medicinal chemistry, 33(8), 2087-‐2093.
Maeda, J., Suhara, T., Ogawa, M., Okauchi, T., Kawabe, K., Zhang, M. R., . . . Suzuki, K. (2001). In vivo binding properties of [carbonyl-‐11C] WAY-‐100635: Effect of endogenous serotonin. Synapse, 40(2), 122-‐129.
Manrique, C., Bosler, O., Becquet, D., Héry, F., Faudon, M., & François-‐Bellan, A. (1998). Post-‐lesion up-‐regulation of 5-‐HT1B binding sites in the suprachiasmatic nucleus may be reversed after spontaneous or graft-‐induced serotonin reinnervation. Brain research, 788(1), 332-‐336.
Manrique, C., Francois-‐Bellan, A., Segu, L., Becquet, D., Hery, M., Faudon, M., & Hery, F. (1994). Impairment of serotoninergic transmission is followed by adaptive changes in 5HT1B binding sites in the rat suprachiasmatic nucleus. Brain research, 663(1), 93-‐100.
Manrique, C., Segu, L., Hery, M., Faudon, M., & François-‐Bellan, M. (1993). Increase of central 5-‐HT1B binding sites following 5, 7-‐dihydroxytryptamine axotomy in the adult rat. Brain research, 623(2), 345-‐348.
Matsumoto, T., Maeno, Y., Kato, H., Seko-‐Nakamura, Y., Monma-‐Ohtaki, J., Ishiba, A., . . . Aoki, Y. (2014). 5-‐hydroxytryptamine-‐and dopamine-‐releasing effects of ring-‐substituted amphetamines on rat brain: A comparative study using in vivo microdialysis. European Neuropsychopharmacology, 24(8), 1362-‐1370.
McCann, U. D., Eligulashvili, V., Mertl, M., Murphy, D. L., & Ricaurte, G. A. (1999). Altered neuroendocrine and behavioral responses to m-‐chlorophenylpiperazine in 3, 4-‐methylenedioxymethamphetamine (MDMA) users. Psychopharmacology, 147(1), 56-‐65.
McCann, U. D., Mertl, M., Eligulashvili, V., & Ricaurte, G. A. (1999). Cognitive performance in (±) 3, 4-‐methylenedioxymethamphetamine (MDMA,“ecstasy”) users: a controlled study. Psychopharmacology, 143(4), 417-‐425.
McCann, U. D., & Ricaurte, G. A. (2001). Caveat emptor: editors beware. Neuropsychopharmacology, 24(3), 333-‐334.
McCann, U. D., Szabo, Z., Scheffel, U., Dannals, R., & Ricaurte, G. (1998). Positron emission tomographic evidence of toxic effect of MDMA (“Ecstasy”) on brain serotonin neurons in human beings. The Lancet, 352(9138), 1433-‐1437.
McCreary, A. C., Bankson, M. G., & Cunningham, K. A. (1999). Pharmacological studies of the acute and chronic effects of (+)-‐3, 4-‐methylenedioxymethamphetamine on locomotor activity: role of 5-‐hydroxytryptamine1A and 5-‐hydroxytryptamine1B/1D receptors. Journal of Pharmacology and Experimental Therapeutics, 290(3), 965-‐973.
McDowell, D. M., & Kleber, H. D. (1994). MDMA: its history and pharmacology. Psychiatric Annals, 24(3), 127-‐130.
McGregor, I. S., Clemens, K. J., Van der Plasse, G., Li, K. M., Hunt, G. E., Chen, F., & Lawrence, A. J. (2003). Increased anxiety 3 months after brief exposure to MDMA (" Ecstasy") in rats: association with altered 5-‐HT transporter and receptor density. Neuropsychopharmacology.
McNamara, M. G., Kelly, J. P., & Leonard, B. E. (1995). Some behavioural and neurochemical aspects of subacute (±) 3, 4-‐methylenedioxymethamphetamine administration in rats. Pharmacology Biochemistry and Behavior, 52(3), 479-‐484.

113
Mechan, A. O., O'Shea, E., Elliott, M. J., Colado, M., & Green, R. A. (2001). A neurotoxic dose of 3, 4-‐methylenedioxymethamphetamine (MDMA; ecstasy) to rats results in a long term defect in thermoregulation. Psychopharmacology, 155(4), 413-‐418.
Meneses, A. (2001). Could the 5-‐HT1B receptor inverse agonism affect learning consolidation? Neuroscience & Biobehavioral Reviews, 25(2), 193-‐201.
Meneses, A. (2002). Tianeptine: 5-‐HT uptake sites and 5-‐HT1-‐7 receptors modulate memory formation in an autoshaping Pavlovian/instrumental task. Neuroscience & Biobehavioral Reviews, 26(3), 309-‐319.
Meneses, A. (2007). Stimulation of 5-‐HT1A, 5-‐HT1B, 5-‐HT2A/2C, 5-‐HT3 and 5-‐HT4 receptors or 5-‐HT uptake inhibition: short-‐and long-‐term memory. Behavioural brain research, 184(1), 81-‐90.
Meneses, A., & Hong, E. (1994a). Mechanism of action of 8-‐OH-‐DPAT on learning and memory. Pharmacology Biochemistry and Behavior, 49(4), 1083-‐1086.
Meneses, A., & Hong, E. (1994b). Modification of 8-‐OH-‐DPAT effects on learning by manipulation of the assay conditions. Behavioral and neural biology, 61(1), 29-‐35.
Meneses, A., & Hong, E. (1997). Role of 5-‐HT1B, 5-‐HT2A and 5-‐HT2C receptors in learning. Behavioural brain research, 87(1), 105-‐110.
Meneses, A., Terrón, J. A., & Hong, E. (1997). Effects of the 5-‐HT receptor antagonists GR127935 (5-‐HT1B/1D) and MDL100907 (5-‐HT2A) in the consolidation of learning. Behavioural brain research, 89(1), 217-‐223.
Merikangas, K. R., Mehta, R. L., Molnar, B. E., Walters, E. E., Swendsen, J. D., Aguilar-‐Gaziola, S., . . . Dewit, D. J. (1998). Comorbidity of substance use disorders with mood and anxiety disorders: results of the International Consortium in Psychiatric Epidemiology. Addictive behaviors, 23(6), 893-‐907.
Meyer, J. S., Piper, B. J., & Vancollie, V. E. (2008). Development and characterization of a novel animal model of intermittent MDMA (“Ecstasy”) exposure during adolescence. Annals of the New York Academy of Sciences, 1139(1), 151-‐163.
Mignon, L., & Wolf, W. A. (2002). Postsynaptic 5-‐HT1A receptors mediate an increase in locomotor activity in the monoamine-‐depleted rat. Psychopharmacology, 163(1), 85-‐94.
Miguéns, M., Crespo, J. A., Del Olmo, N., Higuera-‐Matas, A., Montoya, G. L., García-‐Lecumberri, C., & Ambrosio, E. (2008). Differential cocaine-‐induced modulation of glutamate and dopamine transporters after contingent and non-‐contingent administration. Neuropharmacology, 55(5), 771-‐779.
Miheau, J., & Van Marrewijk, B. (1999). Stimulation of 5-‐HT1A receptors by systemic or medial septum injection induces anxiogenic-‐like effects and facilitates acquisition of a spatial discrimination task in mice. Progress in Neuro-‐Psychopharmacology and Biological Psychiatry, 23(6), 1113-‐1133.
Minabe, Y., Hashimoto, K., Watanabe, K. I., & Ashby, C. R. (2001). Acute and repeated administration of the selective 5-‐HT2A receptor antagonist M100907 significantly alters the activity of midbrain dopamine neurons: An in vivo electrophysiological study. Synapse, 40(2), 102-‐112.
Misuse of Drugs Act, http://www.legislation.govt.nz/ (1975). Misuse of Drugs Act 1975. New Zealand Statutes (1975).

114
Molodtsova, G. (2003). Differences in serotonin and dopamine metabolism in the rat brain in latent inhibition. Neuroscience and Behavioral Physiology, 33(3), 217-‐222.
Monti, J. M., & Jantos, H. (1992). Dose-‐dependent effects of the 5-‐HT1A receptor agonist 8-‐OH-‐DPAT on sleep and wakefulness in the rat. Journal of sleep research, 1(3), 169-‐175.
Monti, J. M., Monti, D., Jantos, H., & Ponzoni, A. (1995). Effects of selective activation of the 5-‐HT1B receptor with CP-‐94,253 on sleep and wakefulness in the rat. Neuropharmacology, 34(12), 1647-‐1651.
Monti, J. M., Pineyro, G., Orellana, C., Boussard, M., Jantos, H., Labraga, P., . . . Alvarino, F. (1990). 5-‐HT receptor agonists 1-‐(2, 5-‐dimethoxy-‐4-‐iodophenyl)-‐2-‐aminopropane (DOI) and 8-‐OH-‐DPAT increase wakefulness in the rat. Biogenic Amines, 7(2), 145-‐151.
Mora, P. D. O., Fouquet, N., Oberling, P., Gobaille, S., Graeff, F. G., & Sandner, G. (1999). A neurotoxic lesion of serotonergic neurones using 5, 7-‐dihydroxytryptamine does not disrupt latent inhibition in paradigms sensitive to low doses of amphetamine. Behavioural brain research, 100(1), 167-‐175.
Mordenti, J., & Chappell, W. (1989). The Use of Interspecies Scaling in Toxicokinetics. In A. Yacobi, J. P. Skelly, & V. K. Batra (Eds.), Toxicokinetics and new drug development: Pergamon Press.
Morefield, K. M., Keane, M., Felgate, P., White, J. M., & Irvine, R. J. (2011). Pill content, dose and resulting plasma concentrations of 3, 4-‐methylendioxymethamphetamine (MDMA) in recreational ‘ecstasy’users. Addiction, 106(7), 1293-‐1300.
Moser, P. C., Hitchcock, J. M., Lister, S., & Moran, P. M. (2000). The pharmacology of latent inhibition as an animal model of schizophrenia. Brain Research Reviews, 33(2), 275-‐307.
Moulin-‐Sallanon, M., Charnay, Y., Ginovart, N., Perret, P., Lanfumey, L., Hamon, M., . . . Millet, P. (2009). Acute and chronic effects of citalopram on 5-‐HT1A receptor—Labeling by [18F] MPPF and—Coupling to receptors-‐G proteins. Synapse, 63(2), 106-‐116.
Müller, C. P., Carey, R. J., Huston, J. P., & Silva, M. A. D. S. (2007). Serotonin and psychostimulant addiction: focus on 5-‐HT1A-‐receptors. Progress in neurobiology, 81(3), 133-‐178.
Muramatsu, M., Lapiz, M. D. S., Tanaka, E., & Grenhoff, J. (1998). Serotonin inhibits synaptic glutamate currents in rat nucleus accumbens neurons via presynaptic 5-‐HT1B receptors. European Journal of Neuroscience, 10(7), 2371-‐2379.
Nair, S. G., & Gudelsky, G. A. (2006a). 3, 4-‐Methylenedioxymethamphetamine enhances the release of acetylcholine in the prefrontal cortex and dorsal hippocampus of the rat. Psychopharmacology, 184(2), 182-‐189.
Nair, S. G., & Gudelsky, G. A. (2006b). Effect of a serotonin depleting regimen of 3, 4-‐methylenedioxymethamphetamine (MDMA) on the subsequent stimulation of acetylcholine release in the rat prefrontal cortex. Brain research bulletin, 69(4), 382-‐387.
Nash, J. F. (1990). Ketanserin pretreatment attenuates MDMA-‐induced dopamine release in the striatum as measured by in vivo microdialysis. Life sciences, 47(26), 2401-‐2408.
Nash, J. F., & Yamamoto, B. K. (1992). Methamphetamine neurotoxicity and striatal glutamate release: comparison to 3, 4-‐methylenedioxymethamphetamine. Brain research, 581(2), 237-‐243.

115
National Institutes of Health. (2015). Clinical Trials. Retrieved from https://clinicaltrials.gov/ Nautiyal, K. M., Tanaka, K. F., Barr, M. M., Tritschler, L., Le Dantec, Y., David, D. J., . . . Ahmari,
S. E. (2015). Distinct circuits underlie the effects of 5-‐HT1b receptors on aggression and impulsivity. Neuron, 86(3), 813-‐826.
Navailles, S., De Deurwaerdere, P., Porras, G., & Spampinato, U. (2004). In vivo evidence that 5-‐HT2C receptor antagonist but not agonist modulates cocaine-‐induced dopamine outflow in the rat nucleus accumbens and striatum. Neuropsychopharmacology, 29(2), 319-‐326.
Neisewander, J. L., Cheung, T. H., & Pentkowski, N. S. (2014). Dopamine D3 and 5-‐HT1B receptor dysregulation as a result of psychostimulant intake and forced abstinence: Implications for medications development. Neuropharmacology, 76, 301-‐319.
New Zealand Drug Foundation. (2015). About a Drug: MDMA. Retrieved from https://www.drugfoundation.org.nz/content/about-‐drug-‐mdma
Newman-‐Tancredi, A., Martel, J.-‐C., Assié, M.-‐B., Buritova, J., Lauressergues, E., Cosi, C., . . . Vacher, B. (2009). Signal transduction and functional selectivity of F15599, a preferential post-‐synaptic 5-‐HT1A receptor agonist. British journal of pharmacology, 156(2), 338-‐353.
Nonkes, L. J., van de Vondervoort, I. I., de Leeuw, M. J., Wijlaars, L. P., Maes, J. H., & Homberg, J. R. (2012). Serotonin transporter knockout rats show improved strategy set-‐shifting and reduced latent inhibition. Learning & memory, 19(5), 190-‐193.
O'Dell, L. E., Manzardo, A. M., Polis, I., Stouffer, D. G., & Parsons, L. H. (2006). Biphasic alterations in Serotonin-‐1B (5-‐HT1B) receptor function during abstinence from extended cocaine self-‐administration. Journal of Neurochemistry, 99(5), 1363-‐1376.
O'Dell, L. E., & Parsons, L. H. (2004). Serotonin1B receptors in the ventral tegmental area modulate cocaine-‐induced increases in nucleus accumbens dopamine levels. Journal of Pharmacology and Experimental Therapeutics, 311(2), 711-‐719.
O’Neill, M., & Parameswaran, T. (1997). RU24969-‐induced behavioural syndrome requires activation of both 5HT1A and 5HT1B receptors. Psychopharmacology, 132(3), 255-‐260.
Oakly, A., Brox, B., Schenk, S., & Ellenbroek, B. (2014). A genetic deletion of the serotonin transporter greatly enhances the reinforcing properties of MDMA in rats. Molecular psychiatry, 19(5), 534-‐535.
Oberlander, C., Demassey, Y., Verdu, A., Van de Velde, D., & Bardelay, C. (1987). Tolerance to the serotonin 5-‐HT1 agonist RU 24969 and effects on dopaminergic behaviour. European journal of pharmacology, 139(2), 205-‐214.
Offord, S. J., Ordway, G. A., & Frazer, A. (1988). Application of [125I] iodocyanopindolol to measure 5-‐hydroxytryptamine1B receptors in the brain of the rat. Journal of Pharmacology and Experimental Therapeutics, 244(1), 144-‐153.
Ohmura, Y., Kumamoto, H., Tsutsui-‐Kimura, I., Minami, M., Izumi, T., Yoshida, T., & Yoshioka, M. (2013). Tandospirone Suppresses Impulsive Action by Possible Blockade of the 5-‐HT1A Receptor. Journal of pharmacological sciences, 122(2), 84-‐92.
Padich, R. A., McCloskey, T. C., & Kehne, J. H. (1996). 5-‐HT modulation of auditory and visual sensorimotor gating: II. Effects of the 5-‐HT2A antagonist MDL 100,907 on disruption of sound and light prepulse inhibition produced by 5-‐HT agonists in Wistar rats. Psychopharmacology, 124(1-‐2), 107-‐116.

116
Palchaudhuri, M., & Flügge, G. (2005). 5-‐HT1A receptor expression in pyramidal neurons of cortical and limbic brain regions. Cell and tissue research, 321(2), 159-‐172.
Panczyk, K., Golda, S., Waszkielewicz, A., Zelaszczyk, D., Gunia-‐Krzyzak, A., & Marona, H. (2015). Serotonergic System and Its Role in Epilepsy and Neuropathic Pain Treatment: A Review Based on Receptor Ligands. Current pharmaceutical design, 21(13), 1723-‐1740.
Parrott, A. C. (2005). Chronic tolerance to recreational MDMA (3,4-‐methylenedioxymethamphetamine) or Ecstasy. Journal of Psychopharmacology, 19(1), 71-‐83. doi:10.1177/0269881105048900
Parrott, A. C. (2006). MDMA in humans: factors which affect the neuropsychobiological profiles of recreational ecstasy users, the integrative role of bioenergetic stress. Journal of Psychopharmacology, 20(2), 147-‐163.
Parrott, A. C. (2013a). Human psychobiology of MDMA or ‘Ecstasy’: an overview of 25 years of empirical research. Human Psychopharmacology: Clinical and Experimental, 28(4), 289-‐307.
Parrott, A. C. (2013b). MDMA, serotonergic neurotoxicity, and the diverse functional deficits of recreational ‘Ecstasy’users. Neuroscience & Biobehavioral Reviews, 37(8), 1466-‐1484.
Parsons, L. H., Koob, G. F., & Weiss, F. (1999). RU 24969, a 5-‐HT1B/1A receptor agonist, potentiates cocaine-‐induced increases in nucleus accumbens dopamine. Synapse, 32(2), 132-‐135.
Parsons, L. H., Weiss, F., & Koob, G. F. (1996). Serotonin1B receptor stimulation enhances dopamine-‐mediated reinforcement. Psychopharmacology, 128(2), 150-‐160.
Parsons, L. H., Weiss, F., & Koob, G. F. (1998). Serotonin1B receptor stimulation enhances cocaine reinforcement. The Journal of neuroscience, 18(23), 10078-‐10089.
Passetti, F., Dalley, J. W., & Robbins, T. W. (2003). Double dissociation of serotonergic and dopaminergic mechanisms on attentional performance using a rodent five-‐choice reaction time task. Psychopharmacology, 165(2), 136-‐145.
Pattij, T., Broersen, L. M., van der Linde, J., Groenink, L., van der Gugten, J., Maes, R. A., & Olivier, B. (2003). Operant learning and differential-‐reinforcement-‐of-‐low-‐rate 36-‐s responding in 5-‐HT1A and 5-‐HT1B receptor knockout mice. Behavioural brain research, 141(2), 137-‐145.
Pehek, E. A., Nocjar, C., Roth, B. L., Byrd, T. A., & Mabrouk, O. S. (2005). Evidence for the preferential involvement of 5-‐HT2A serotonin receptors in stress-‐and drug-‐induced dopamine release in the rat medial prefrontal cortex. Neuropsychopharmacology, 31(2), 265-‐277.
Peiró, A., Farré, M., Roset, P., Carbó, M., Pujadas, M., Torrens, M., . . . De la Torre, R. (2013). Human pharmacology of 3, 4-‐methylenedioxymethamphetamine (MDMA, ecstasy) after repeated doses taken 2 h apart. Psychopharmacology, 225(4), 883-‐893.
Pellow, S., Chopin, P., File, S. E., & Briley, M. (1985). Validation of open: closed arm entries in an elevated plus-‐maze as a measure of anxiety in the rat. Journal of neuroscience methods, 14(3), 149-‐167.
Pellow, S., Johnston, A. L., & File, S. E. (1987). Selective agonists and antagonists for 5-‐hydroxytryptamine receptor subtypes, and interactions with yohimbine and FG 7142

117
using the elevated plus-‐maze test in the rat. Journal of pharmacy and pharmacology, 39(11), 917-‐928.
Peltier, R., & Schenk, S. (1993). Effects of serotonergic manipulations on cocaine self-‐administration in rats. Psychopharmacology, 110(4), 390-‐394.
Pentkowski, N. S., Acosta, J. I., Browning, J. R., Hamilton, E. C., & Neisewander, J. L. (2009). Stimulation of 5-‐HT1B receptors enhances cocaine reinforcement yet reduces cocaine-‐seeking behavior. Addiction Biology, 14(4), 419-‐430.
Peroutka, S. J. (1986). Pharmacological Differentiation and Characterization of 5-‐HT1A, 5-‐HT1B, and 5-‐HT1C Binding Sites in Rat Frontal Cortex. Journal of Neurochemistry, 47(2), 529-‐540.
Peroutka, S. J., Newman, H., & Harris, H. (1988). Subjective effects of 3, 4-‐methylenedioxymethamphetamine in recreational users. Neuropsychopharmacology, 1(4), 273-‐277.
Perry, J. L., & Carroll, M. E. (2008). The role of impulsive behavior in drug abuse. Psychopharmacology, 200(1), 1-‐26.
Perry, J. L., Larson, E. B., German, J. P., Madden, G. J., & Carroll, M. E. (2005). Impulsivity (delay discounting) as a predictor of acquisition of IV cocaine self-‐administration in female rats. Psychopharmacology, 178(2-‐3), 193-‐201.
Perry, J. L., Nelson, S. E., & Carroll, M. E. (2008). Impulsive choice as a predictor of acquisition of IV cocaine self-‐administration and reinstatement of cocaine-‐seeking behavior in male and female rats. Experimental and clinical psychopharmacology, 16(2), 165.
Pifl, C., Reither, H., & Hornykiewicz, O. (2015). The profile of mephedrone on human monoamine transporters differs from 3, 4-‐methylenedioxymethamphetamine primarily by lower potency at the vesicular monoamine transporter. European journal of pharmacology, 755, 119-‐126.
Piper, B. J., Ali, S. F., Daniels, L. G., & Meyer, J. S. (2010). Repeated intermittent methylenedioxymethamphetamine exposure protects against the behavioral and neurotoxic, but not hyperthermic, effects of an MDMA binge in adult rats. Synapse, 64(6), 421-‐431.
Piper, B. J., Vu, H. L., Safain, M. G., Oliver, A. J., & Meyer, J. S. (2006). Repeated adolescent 3, 4-‐methylenedioxymethamphetamine (MDMA) exposure in rats attenuates the effects of a subsequent challenge with MDMA or a 5-‐hydroxytryptamine1A receptor agonist. Journal of Pharmacology and Experimental Therapeutics, 317(2), 838-‐849.
Plaznik, A., Stefanski, R., Palejko, W., Bidzinski, A., Kostowski, W., Jessa, M., & Nazar, M. (1994). Antidepressant treatment and limbic serotonergic mechanisms regulating rat locomotor activity. Pharmacology Biochemistry and Behavior, 48(2), 315-‐325.
Pompeiano, M., Palacios, J. M., & Mengod, G. (1992). Distribution and cellular localization of mRNA coding for 5-‐HT1A receptor in the rat brain: correlation with receptor binding. The Journal of neuroscience, 12(2), 440-‐453.
Poulos, C. X., Le, A., & Parker, J. (1995). Impulsivity predicts individual susceptibility to high levels of alcohol self-‐administration. Behavioural pharmacology.
Pranzatelli, M. R., & Razi, P. (1994). Drug-‐induced regulation of [125I] iodocyanopindolol-‐labeled 5-‐hydroxytryptamine1B receptor binding sites in the central nervous system. Neuropsychopharmacology, 10(4), 259-‐264.

118
Price, G., Burton, M., Collin, L., Duckworth, M., Gaster, L., Göthert, M., . . . Middlemiss, D. (1997). SB-‐216641 and BRL-‐15572–compounds to pharmacologically discriminate h5-‐HT1B and h5-‐HT1D receptors. Naunyn-‐Schmiedeberg's archives of pharmacology, 356(3), 312-‐320.
Przegaliński, E., Czepiel, K., Nowak, E., Dlaboga, D., & Filip, M. (2003). Withdrawal from chronic cocaine up-‐regulates 5-‐HT1B receptors in the rat brain. Neuroscience letters, 351(3), 169-‐172.
Przegaliñski, E., Gołda, A., Frankowska, M., Zaniewska, M., & Filip, M. (2007). Effects of serotonin 5-‐HT1B receptor ligands on the cocaine-‐and food-‐maintained self-‐administration in rats. European journal of pharmacology, 559(2), 165-‐172.
Przegaliñski, E., Siwanowicz, J., Baran, L., & Filip, M. (2000). Activation of serotonin (5-‐HT) 1A receptors inhibits amphetamine sensitization in mice. Life sciences, 66(11), 1011-‐1019.
Puig, M. V., Artigas, F., & Celada, P. (2005). Modulation of the activity of pyramidal neurons in rat prefrontal cortex by raphe stimulation in vivo: involvement of serotonin and GABA. Cerebral Cortex, 15(1), 1-‐14.
Puig, M. V., Watakabe, A., Ushimaru, M., Yamamori, T., & Kawaguchi, Y. (2010). Serotonin modulates fast-‐spiking interneuron and synchronous activity in the rat prefrontal cortex through 5-‐HT1A and 5-‐HT2A receptors. The Journal of neuroscience, 30(6), 2211-‐2222.
Raiteri, M., Maura, G., Bonanno, G., & Pittaluga, A. (1986). Differential pharmacology and function of two 5-‐HT1 receptors modulating transmitter release in rat cerebellum. Journal of Pharmacology and Experimental Therapeutics, 237(2), 644-‐648.
Ramos, M., Goni-‐Allo, B., & Aguirre, N. (2005). Administration of SCH 23390 into the medial prefrontal cortex blocks the expression of MDMA-‐induced behavioral sensitization in rats: an effect mediated by 5-‐HT2C receptor stimulation and not by D1 receptor blockade. Neuropsychopharmacology, 30(12), 2180-‐2191.
Rasmusson, A. M., Goldstein, L. E., Deutch, A. Y., Bunney, B. S., & Roth, R. H. (1994). 5-‐HT1A agonist±-‐8-‐OH-‐DPAT modulates basal and stress-‐induced changes in medial prefrontal cortical dopamine. Synapse, 18(3), 218-‐224.
Reveron, M. E., Maier, E. Y., & Duvauchelle, C. L. (2010). Behavioral, thermal and neurochemical effects of acute and chronic 3, 4-‐methylenedioxymethamphetamine (“Ecstasy”) self-‐administration. Behavioural brain research, 207(2), 500-‐507.
Riad, M., Garcia, S., Watkins, K. C., Jodoin, N., Doucet, É., Langlois, X., . . . Descarries, L. (2000). Somatodendritic localization of 5-‐HT1A and preterminal axonal localization of 5-‐HT1B serotonin receptors in adult rat brain. The Journal of Comparative Neurology, 417(2), 181-‐194. doi:10.1002/(sici)1096-‐9861(20000207)417:2<181::aid-‐cne4>3.0.co;2-‐a
Ricaurte, G. A., Yuan, J., & McCann, U. D. (2000). (±) 3, 4-‐Methylenedioxymethamphetamine (‘Ecstasy’)-‐induced serotonin neurotoxicity: studies in animals. Neuropsychobiology, 42(1), 5-‐10.
Ritz, M. C., & Kuhar, M. J. (1989). Relationship between self-‐administration of amphetamine and monoamine receptors in brain: comparison with cocaine. Journal of Pharmacology and Experimental Therapeutics, 248(3), 1010-‐1017.

119
Robbins, T. (2002). The 5-‐choice serial reaction time task: behavioural pharmacology and functional neurochemistry. Psychopharmacology, 163(3-‐4), 362-‐380.
Roberts, D. C., Corcoran, M. E., & Fibiger, H. C. (1977). On the role of ascending catecholaminergic systems in intravenous self-‐administration of cocaine. Pharmacology Biochemistry and Behavior, 6(6), 615-‐620.
Roberts, D. C., & Koob, G. F. (1982). Disruption of cocaine self-‐administration following 6-‐hydroxydopamine lesions of the ventral tegmental area in rats. Pharmacology Biochemistry and Behavior, 17(5), 901-‐904.
Rocha, B. A., Goulding, E. H., O'Dell, L. E., Mead, A. N., Coufal, N. G., Parsons, L. H., & Tecott, L. H. (2002). Enhanced locomotor, reinforcing, and neurochemical effects of cocaine in serotonin 5-‐hydroxytryptamine 2C receptor mutant mice. The Journal of neuroscience, 22(22), 10039-‐10045.
Rogers, G., Elston, J., Garside, R., Roome, C., Taylor, R., Younger, P., . . . Somerville, M. (2009). The harmful health effects of recreational ecstasy: a systematic review of observational evidence. Health Technology Assessment, 13(6), xii-‐338.
Rothman, R. B., & Baumann, M. H. (2003). Monoamine transporters and psychostimulant drugs. European journal of pharmacology, 479(1), 23-‐40.
Rothman, R. B., Blough, B. E., Woolverton, W. L., Anderson, K. G., Negus, S. S., Mello, N. K., . . . Baumann, M. H. (2005). Development of a rationally designed, low abuse potential, biogenic amine releaser that suppresses cocaine self-‐administration. Journal of Pharmacology and Experimental Therapeutics, 313(3), 1361-‐1369.
Ruotsalainen, S., Sirviö, J., Jäkälä, P., Puumala, T., MacDonald, E., & Riekkinen Sr, P. (1997). Differential effects of three 5-‐HT receptor antagonists on the performance of rats in attentional and working memory tasks. European Neuropsychopharmacology, 7(2), 99-‐108.
Sakaue, M., Somboonthum, P., Nishihara, B., Koyama, Y., Hashimoto, H., Baba, A., & Matsuda, T. (2000). Postsynaptic 5-‐hydroxytryptamine1A receptor activation increases in vivo dopamine release in rat prefrontal cortex. British journal of pharmacology, 129(5), 1028-‐1034.
Sarhan, H., & Fillion, G. (1999). Differential sensitivity of 5-‐HT1B auto and heteroreceptors. Naunyn-‐Schmiedeberg's archives of pharmacology, 360(4), 382-‐390.
Sari, Y. (2004). Serotonin1B receptors: from protein to physiological function and behavior. Neuroscience & Biobehavioral Reviews, 28(6), 565-‐582.
Sari, Y., Miquel, M.-‐C., Brisorgueil, M.-‐J., Ruiz, G., Doucet, E., Hamon, M., & Verge, D. (1999). Cellular and subcellular localization of 5-‐hydroxytryptamine1B receptors in the rat central nervous system: immunocytochemical, autoradiographic and lesion studies. Neuroscience, 88(3), 899-‐915.
Say, M., Machaalani, R., & Waters, K. A. (2007). Changes in serotoninergic receptors 1A and 2A in the piglet brainstem after intermittent hypercapnic hypoxia (IHH) and nicotine. Brain research, 1152, 17-‐26.
Scanzello, C. R., Hatzidimitriou, G., Martello, A. L., Katz, J. L., & Ricaurte, G. (1993). Serotonergic recovery after (+/-‐) 3, 4-‐(methylenedioxy) methamphetamine injury: observations in rats. Journal of Pharmacology and Experimental Therapeutics, 264(3), 1484-‐1491.

120
Schenk, S. (2009). MDMA self-‐administration in laboratory animals: a summary of the literature and proposal for future research. Neuropsychobiology, 60(3-‐4), 130-‐136.
Schenk, S. (2011). MDMA (“ecstasy”) abuse as an example of dopamine neuroplasticity. Neuroscience & Biobehavioral Reviews, 35(5), 1203-‐1218.
Schenk, S., Abraham, B., Aronsen, D., Colussi-‐Mas, J., & Do, J. (2013). Effects of repeated exposure to MDMA on 5HT1a autoreceptor function: behavioral and neurochemical responses to 8-‐OHDPAT. Psychopharmacology, 227(2), 355-‐361.
Schenk, S., & Aronsen, D. (2015). Contribution of Impulsivity and Serotonin Receptor Neuroadaptations to the development of an MDMA (“ecstasy”) Substance Use Disorder. In S. Schenk, S. Nielson, & R. Bruno (Eds.), Current Topics in Behavioral Neuroscience: The Misuse of Licit and Illicit Drugs in Psychopharmacology (pp. 1-‐16): Springer.
Schenk, S., Colussi-‐Mas, J., Do, J., & Bird, J. (2012). Profile of MDMA self-‐administration from a large cohort of rats: MDMA develops a profile of dependence with extended testing. Journal of Drug and Alcohol Research, 1, 1-‐6.
Schenk, S., Foote, J., Aronsen, D., Bukholt, N., Highgate, Q., Van de Wetering, R., & Webster, J. (Under Review). Comparison of the role of serotonin in MDMA and cocaine self-‐administration and drug-‐seeking in rats. British journal of pharmacology.
Schenk, S., Gittings, D., & Colussi-‐Mas, J. (2011). Dopaminergic mechanisms of reinstatement of MDMA-‐seeking behaviour in rats. British journal of pharmacology, 162(8), 1770-‐1780.
Schenk, S., Gittings, D., Johnstone, M., & Daniela, E. (2003). Development, maintenance and temporal pattern of self-‐administration maintained by ecstasy (MDMA) in rats. Psychopharmacology, 169(1), 21-‐27.
Schenk, S., Hely, L., Lake, B., Daniela, E., Gittings, D., & Mash, D. C. (2007). MDMA self-‐administration in rats: acquisition, progressive ratio responding and serotonin transporter binding. European Journal of Neuroscience, 26(11), 3229-‐3236.
Schenk, S., & Partridge, B. (2000). Sensitization to cocaine's reinforcing effects produced by various cocaine pretreatment regimens in rats. Pharmacology Biochemistry and Behavior, 66(4), 765-‐770.
Schifano, F., & Magni, G. (1994). MDMA (“Ecstasy”) abuse: psychopathological features and craving for chocolate: a case series. Biological Psychiatry, 36(11), 763-‐767.
Schmidt, C. J., Sullivan, C. K., & Fedayal, G. M. (1994). Blockade of Striatal 5-‐Hydroxytryptmine2 Receptors Reduces the Increase in Extracellullar Concentrations of Dopamine Produced by the Amphhetamine analogue 3,4-‐Methylenedioxymethamphetamine. Journal of Neurochemistry, 62(4), 1382-‐1389. doi:10.1046/j.1471-‐4159.1994.62041382.x
Schmidt, C. J., & Taylor, V. L. (1987). Depression of rat brain tryptophan hydroxylase activity following the acute administration of methylenedioxymethamphetamine. Biochemical pharmacology, 36(23), 4095-‐4102.
Schoeffter, P., & Hoyer, D. (1988). Centrally acting hypotensive agents with affinity for 5-‐HT1A binding sites inhibit forskolin-‐stimulated adenylate cyclase activity in calf hippocampus. British journal of pharmacology, 95(3), 975-‐985.
Schwartz, R. H., & Miller, N. S. (1997). MDMA (ecstasy) and the rave: a review. Pediatrics, 100(4), 705-‐708.

121
Scorza, M. C., Carrau, C., Silveira, R., Zapata-‐Torres, G., Cassels, B. K., & Reyes-‐Parada, M. (1997). Monoamine oxidase inhibitory properties of some methoxylated and alkylthio amphetamine derivatives: structure–activity relationships. Biochemical pharmacology, 54(12), 1361-‐1369.
Seuwen, K., Magnaldo, I., & Pouysségur, J. (1988). Serotonin stimulates DNA synthesis in fibroblasts acting through 5–HT1B receptors coupled to a Gi-‐protein. Nature, 335(6187), 254-‐256.
Sexton, T., McEvoy, C., & Neumaier, J. (1999). (+) 3, 4-‐methylenedioxymethamphetamine ('ecstasy') transiently increases striatal 5-‐HT1B binding sites without altering 5-‐HT1B mRNA in rat brain. Molecular psychiatry, 4(6), 572-‐579.
Shanahan, N. A., Holick Pierz, K. A., Masten, V. L., Waeber, C., Ansorge, M., Gingrich, J. A., . . . Dulawa, S. C. (2009). Chronic reductions in serotonin transporter function prevent 5-‐HT1B-‐induced behavioral effects in mice. Biological Psychiatry, 65(5), 401-‐408.
Shankaran, M., & Gudelsky, G. A. (1999). A neurotoxic regimen of MDMA suppresses behavioral, thermal and neurochemical responses to subsequent MDMA administration. Psychopharmacology, 147(1), 66-‐72.
Sharpley, A., Elliott, J., Attenburrow, M.-‐J., & Cowen, P. (1994). Slow wave sleep in humans: role of 5-‐HT2A and 5-‐HT2C receptors. Neuropharmacology, 33(3), 467-‐471.
Sher, K. J., Bartholow, B. D., & Wood, M. D. (2000). Personality and substance use disorders: a prospective study. Journal of consulting and clinical psychology, 68(5), 818.
Shortall, S. E., Macerola, A. E., Swaby, R. T., Jayson, R., Korsah, C., Pillidge, K. E., . . . Fone, K. C. (2013). Behavioural and neurochemical comparison of chronic intermittent cathinone, mephedrone and MDMA administration to the rat. European Neuropsychopharmacology, 23(9), 1085-‐1095.
Shulgin, A. T. (1978). Psychotomimetic drugs: Structure-‐activity relationships Stimulants (pp. 243-‐333): Springer.
Shulgin, A. T., & Nichols, D. E. (1978). Characterization of three new psychotomimetics. The Pharmacology of Hallucinogens. Pergamon, New York.
Silvestre, J. S., Palacios, J. M., Fernandez, A. G., & O'Neill, M. F. (1998). Comparison of effects of a range of 5-‐HT receptor modulators on consumption and preference for a sweetened ethanol solution in rats. Journal of Psychopharmacology, 12(2), 168-‐176.
Simansky, K. J., & Vaidya, A. (1990). Behavioral mechanisms for the anorectic action of the serotonin (5-‐HT) uptake inhibitor sertraline in rats: comparison with directly acting 5-‐HT agonists. Brain research bulletin, 25(6), 953-‐960.
Skolnick, P. (2015). Biologic Approaches to Treat Substance-‐Use Disorders. Trends in pharmacological sciences, 36(10), 628-‐635.
Soar, K., Turner, J., & Parrott, A. C. (2006). Problematic versus non-‐problematic ecstasy/MDMA use: the influence of drug usage patterns and pre-‐existing psychiatric factors. Journal of Psychopharmacology, 20(3), 417-‐424.
Solati, J., Salari, A.-‐A., & Bakhtiari, A. (2011). 5HT1A and 5HT1B receptors of medial prefrontal cortex modulate anxiogenic-‐like behaviors in rats. Neuroscience letters, 504(3), 325-‐329.
Solomon, P. R., Nichols, G. L., Kiernan, J. M., Kamer, R. S., & Kaplan, L. J. (1980). Differential effects of lesions in medial and dorsal raphe of the rat: latent inhibition and

122
septohippocampal serotonin levels. Journal of comparative and physiological psychology, 94(1), 145.
Solowij, N., Hall, W., & Lee, N. (1992). Recreational MDMA use in Sydney: a profile of ‘ecstasy’users and their experiences with the drug. British journal of addiction, 87(8), 1161-‐1172.
Spanagel, R., Montkowski, A., Allingham, K., Shoaib, M., Holsboer, F., & Landgraf, R. (1995). Anxiety: a potential predictor of vulnerability to the initiation of ethanol self-‐administration in rats. Psychopharmacology, 122(4), 369-‐373.
Starr, M. A., Page, M. E., & Waterhouse, B. D. (2012). Effects of repeated 3, 4-‐methylenedioxymethamphetamine administration on neurotransmitter efflux and sensory-‐evoked discharge in the ventral posterior medial thalamus. Journal of Pharmacology and Experimental Therapeutics, 340(1), 73-‐82.
Stone, D. M., Hanson, G., & Gibb, J. (1987). Differences in the central serotonergic effects of methylenedioxymethamphetamine (MDMA) in mice and rats. Neuropharmacology, 26(11), 1657-‐1661.
Stone, D. M., Johnson, M., Hanson, G. R., & Gibb, J. W. (1988). Role of endogenous dopamine in the central serotonergic deficits induced by 3, 4-‐methylenedioxymethamphetamine. Journal of Pharmacology and Experimental Therapeutics, 247(1), 79-‐87.
Stone, D. M., Merchant, K., Hanson, G., & Gibb, J. (1987). Immediate and long-‐term effects of 3, 4-‐methylenedioxymethamphetamine on serotonin pathways in brain of rat. Neuropharmacology, 26(12), 1677-‐1683.
Suzuki, H., Han, S. D., & Lucas, L. R. (2010). Chronic passive exposure to aggression decreases D2 and 5-‐HT1B receptor densities. Physiology & behavior, 99(5), 562-‐570.
Tachibana, T., Endoh, M., Fujiwara, N., & Nawa, T. (2005). Receptors and transporter for serotonin in Merkel cell-‐nerve endings in the rat sinus hair follicle. An immunohistochemical study. Archives of histology and cytology, 68(1), 19-‐28.
Taffe, M. A., Davis, S. A., Yuan, J., Schroeder, R., Hatzidimitriou, G., Parsons, L. H., . . . Gold, L. H. (2002). Cognitive performance of MDMA-‐treated rhesus monkeys: sensitivity to serotonergic challenge. Neuropsychopharmacology, 27(6), 993-‐1005.
Talpos, J. C., Wilkinson, L. S., & Robbins, T. W. (2006). A comparison of multiple 5-‐HT receptors in two tasks measuring impulsivity. Journal of Psychopharmacology, 20(1), 47-‐58.
Tarter, R. E., Kirisci, L., Feske, U., & Vanyukov, M. (2007). Modeling the pathways linking childhood hyperactivity and substance use disorder in young adulthood. Psychology of Addictive Behaviors, 21(2), 266.
Tatarczynska, E., Klodzinska, A., Stachowicz, K., & Chojnacka-‐Wojcik, E. (2004). Effects of a selective 5-‐HT1B receptor agonist and antagonists in animal models of anxiety and depression. Behavioural pharmacology, 15(8), 523-‐534.
Taurah, L., Chandler, C., & Sanders, G. (2014). Depression, impulsiveness, sleep, and memory in past and present polydrug users of 3, 4-‐methylenedioxymethamphetamine (MDMA, ecstasy). Psychopharmacology, 231(4), 737-‐751.
Thompson, M. R., Callaghan, P. D., Hunt, G. E., & McGregor, I. S. (2008). Reduced sensitivity to MDMA-‐induced facilitation of social behaviour in MDMA pre-‐exposed rats. Progress in Neuro-‐Psychopharmacology and Biological Psychiatry, 32(4), 1013-‐1021.

123
Togni, L. R., Lanaro, R., Resende, R. R., & Costa, J. L. (2015). The Variability of Ecstasy Tablets Composition in Brazil. Journal of forensic sciences, 60(1), 147-‐151.
Tomkins, D. M., Joharchi, N., Tampakeras, M., Martin, J., Wichmann, J., & Higgins, G. (2002). An investigation of the role of 5-‐HT2C receptors in modifying ethanol self-‐administration behaviour. Pharmacology Biochemistry and Behavior, 71(4), 735-‐744.
Topp, L., Hall, W., & Hando, J. (1997). Is there a dependence syndrome for ecstasy? Travis, C. C., & White, R. K. (1988). Interspecific scaling of toxicity data. Risk Analysis, 8(1),
119-‐125. Tricklebank, M. D., Forler, C., & Fozard, J. R. (1984). The involvement of subtypes of the 5-‐HT1
receptor and of catecholaminergic systems in the behavioural response to 8-‐hydroxy-‐2-‐(di-‐n-‐Propylamino) tetralin in the rat. European journal of pharmacology, 106(2), 271-‐282.
Tricklebank, M. D., Middlemiss, D., & Neill, J. (1986). Pharmacological analysis of the behavioural and thermoregulatory effects of the putative 5-‐HT1 receptor agonist, RU 24969, in the rat. Neuropharmacology, 25(8), 877-‐886.
United Nations Office on Drugs and Crime. (2015). World Drug Report, 2015 (United Nations publication, Sales No. E.15.XI.6). Retrieved from http://www.unodc.org/documents/wdr2015/World_Drug_Report_2015.pdf
Uosukainen, H., Tacke, U., & Winstock, A. R. (2015). Self-‐reported prevalence of dependence of MDMA compared to cocaine, mephedrone and ketamine among a sample of recreational poly-‐drug users. International Journal of Drug Policy, 26(1), 78-‐83.
Urban, N. B., Girgis, R. R., Talbot, P. S., Kegeles, L. S., Xu, X., Frankle, W. G., . . . Laruelle, M. (2012). Sustained Recreational Use of Ecstasy Is Associated with Altered Pre and Postsynaptic Markers of Serotonin Transmission in Neocortical Areas: A PET Study with [11C] DASB and [11C] MDL 100907. Neuropsychopharmacology, 37(6), 1465-‐1473.
van den Bergh, F. S., Bloemarts, E., Groenink, L., Olivier, B., & Oosting, R. S. (2006). Delay aversion: Effects of 7-‐OH-‐DPAT, 5-‐HT1A/1B-‐receptor stimulation and d-‐cycloserine. Pharmacology Biochemistry and Behavior, 85(4), 736-‐743.
Varty, G. B., Bakshi, V. P., & Geyer, M. A. (1999). M100907, a serotonin 5-‐HT2A receptor antagonist and putative antipsychotic, blocks dizocilpine-‐induced prepulse inhibition deficits in Sprague–Dawley and Wistar rats. Neuropsychopharmacology, 20(4), 311-‐321.
Vergé, D., Daval, G., Marcinkiewicz, M., Patey, A., El Mestikawy, S., Gozlan, H., & Hamon, M. (1986). Quantitative autoradiography of multiple 5-‐HT1 receptor subtypes in the brain of control or 5, 7-‐dihydroxytryptamine-‐treated rats. The Journal of neuroscience, 6(12), 3474-‐3482.
Verheyden, S. L., Henry, J. A., & Curran, H. V. (2003). Acute, sub-‐acute and long-‐term subjective consequences of ‘ecstasy’(MDMA) consumption in 430 regular users. Human Psychopharmacology: Clinical and Experimental, 18(7), 507-‐517.
Vogels, N., Brunt, T. M., Rigter, S., Van Dijk, P., Vervaeke, H., & Niesink, R. J. (2009). Content of ecstasy in the Netherlands: 1993–2008. Addiction, 104(12), 2057-‐2066.
Vollenweider, F. X., Jones, R. T., & Baggott, M. J. (2001). Caveat emptor: editors beware. Neuropsychopharmacology, 24(4), 461-‐463.

124
Wagner, D., Becker, B., Koester, P., Gouzoulis-‐Mayfrank, E., & Daumann, J. (2013). A prospective study of learning, memory, and executive function in new MDMA users. Addiction, 108(1), 136-‐145.
Wang, C., Jiang, Y., Ma, J., Wu, H., Wacker, D., Katritch, V., . . . Vardy, E. (2013). Structural basis for molecular recognition at serotonin receptors. Science, 340(6132), 610-‐614.
Wang, S., Zhang, Z.-‐j., Guo, Y.-‐j., Teng, G.-‐j., & Chen, B.-‐a. (2009). Decreased expression of serotonin 1A receptor in the dentate gyrus in association with chronic mild stress: a rat model of post-‐stroke depression. Psychiatry research, 170(2), 245-‐251.
Wang, Z., & Woolverton, W. L. (2007). Estimating the relative reinforcing strength of (±)-‐3, 4-‐methylenedioxymethamphetamine (MDMA) and its isomers in rhesus monkeys: comparison to (+)-‐methamphetamine. Psychopharmacology, 189(4), 483-‐488.
Watanabe, K., Bois, F. Y., & Zeise, L. (1992). Interspecies extrapolation: a reexamination of acute toxicity data. Risk Analysis, 12(2), 301-‐310.
Watson, J. M., Burton, M. J., Price, G. W., Jones, B. J., & Middlemiss, D. N. (1996). GR127935 acts as a partial agonist at recombinant human 5-‐HT1Dα and 5-‐HT1Dβ receptors. European journal of pharmacology, 314(3), 365-‐372. doi:http://dx.doi.org/10.1016/S0014-‐2999(96)00579-‐1
Wee, S., Anderson, K. G., Baumann, M. H., Rothman, R. B., Blough, B. E., & Woolverton, W. L. (2005). Relationship between the serotonergic activity and reinforcing effects of a series of amphetamine analogs. Journal of Pharmacology and Experimental Therapeutics, 313(2), 848-‐854.
Weissmann, D., Mach, E., Oberlander, C., Demassey, Y., & Pujol, J.-‐F. (1986). Evidence for hyperdensity of 5HT1B binding sites in the substantia nigra of the rat after 5, 7-‐dihydroxytryptamine intraventricular injection. Neurochemistry international, 9(1), 191-‐200.
Welner, S., De Montigny, C., Desroches, J., Desjardins, P., & Suranyi-‐Cadotte, B. (1989). Autoradiographic quantification of serotonin1A receptors in rat brain following antidepressant drug treatment. Synapse, 4(4), 347-‐352.
Wilkins, C. (2011). Recent Trends in Illegal Drug Use in New Zealand, 2006-‐2010: Findings from the 2006, 2007, 2008, 2009 and 2010 Illicit Drug Monitoring System (IDMS): Social and Health Outcomes Research and Evaluation, School of Public Health, Massey University.
Wilkins, C., & Sweetsur, P. (2008). Trends in population drug use in New Zealand: findings from national household surveying of drug use in 1998, 2001, 2003, and 2006. NZ Med J, 121.
Winsauer, P., Rodriguez, F., Cha, A., & Moerschbaecher, J. (1999). Full and partial 5-‐HT1A receptor agonists disrupt learning and performance in rats. Journal of Pharmacology and Experimental Therapeutics, 288(1), 335-‐347.
Winstanley, C. A., Chudasama, Y., Dalley, J. W., Theobald, D. E., Glennon, J. C., & Robbins, T. W. (2003). Intra-‐prefrontal 8-‐OH-‐DPAT and M100907 improve visuospatial attention and decrease impulsivity on the five-‐choice serial reaction time task in rats. Psychopharmacology, 167(3), 304-‐314.
Winstanley, C. A., Eagle, D. M., & Robbins, T. W. (2006). Behavioral models of impulsivity in relation to ADHD: translation between clinical and preclinical studies. Clinical psychology review, 26(4), 379-‐395.

125
Winstanley, C. A., Theobald, D. E., Dalley, J. W., Glennon, J. C., & Robbins, T. W. (2004). 5-‐HT2A and 5-‐HT2C receptor antagonists have opposing effects on a measure of impulsivity: interactions with global 5-‐HT depletion. Psychopharmacology, 176(3-‐4), 376-‐385.
Wise, R. A., & Bozarth, M. A. (1987). A psychomotor stimulant theory of addiction. Psychological review, 94(4), 469.
Wolf, W. A., & Kuhn, D. M. (1991). The 5-‐HT transporter is an additional site of action for the 5-‐HT agonists RU 24969 and TFMPP. Neurochemistry international, 19(1), 39-‐44.
Yamamoto, B. K., Nash, J. F., & Gudelsky, G. A. (1995). Modulation of methylenedioxymethamphetamine-‐induced striatal dopamine release by the interaction between serotonin and gamma-‐aminobutyric acid in the substantia nigra. Journal of Pharmacology and Experimental Therapeutics, 273(3), 1063-‐1070.
Yan, Q.-‐S., & Yan, S.-‐E. (2001a). Activation of 5-‐HT1B/1D receptors in the mesolimbic dopamine system increases dopamine release from the nucleus accumbens: a microdialysis study. European journal of pharmacology, 418(1), 55-‐64.
Yan, Q.-‐S., & Yan, S.-‐E. (2001b). Serotonin-‐1B receptor-‐mediated inhibition of [3H] GABA release from rat ventral tegmental area slices. Journal of Neurochemistry, 79(4), 914-‐922.
Yan, Q.-‐S., Zheng, S.-‐Z., Feng, M.-‐J., & Yan, S.-‐E. (2005). Involvement of 5-‐HT1B receptors within the ventral tegmental area in ethanol-‐induced increases in mesolimbic dopaminergic transmission. Brain research, 1060(1), 126-‐137.
Yan, Q.-‐S., Zheng, S.-‐Z., & Yan, S.-‐E. (2004). Involvement of 5-‐HT1B receptors within the ventral tegmental area in regulation of mesolimbic dopaminergic neuronal activity via GABA mechanisms: a study with dual-‐probe microdialysis. Brain research, 1021(1), 82-‐91.
Yen, C.-‐F., & Hsu, S.-‐Y. (2007). Symptoms of ecstasy dependence and correlation with psychopathology in Taiwanese adolescents. The Journal of nervous and mental disease, 195(10), 866-‐869.
Yokel, R. A., & Wise, R. A. (1975). Increased lever pressing for amphetamine after pimozide in rats: implications for a dopamine theory of reward. Science, 187(4176), 547-‐549.
Yokel, R. A., & Wise, R. A. (1978). Amphetamine-‐type reinforcement by dopaminergic agonists in the rat. Psychopharmacology, 58(3), 289-‐296.
Yoshimoto, K., & McBride, W. (1992). Regulation of nucleus accumbens dopamine release by the dorsal raphe nucleus in the rat. Neurochemical research, 17(5), 401-‐407.
Zhang, J., Engel, J. A., Jackson, D. M., Johansson, C., & Svensson, L. (1997). (−) Alprenolol potentiates the disrupting effects of dizocilpine on sensorimotor function in the rat. Psychopharmacology, 132(3), 281-‐288.
Zlebnik, N. E., & Carroll, M. E. (2015). Effects of the combination of wheel running and atomoxetine on cue-‐and cocaine-‐primed reinstatement in rats selected for high or low impulsivity. Psychopharmacology, 232(6), 1049-‐1059.
Zouk, H., McGirr, A., Lebel, V., Benkelfat, C., Rouleau, G., & Turecki, G. (2007). The effect of genetic variation of the serotonin 1B receptor gene on impulsive aggressive behavior and suicide. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 144(8), 996-‐1002.

126
Appendix A: Publication details and permissions
Aronsen, Webster, & Schenk (2014). RU 24969-produced adipsia and
hyperlocomotion: Differential role of 5HT1A and 5HT1B receptor mechanisms.
Pharmacology, Biochemistry and Behavior, 124, 1-4. DOI
10.1016/j.pbb.2014.05.008
Licensed content publisher: Elsevier
License Number: 3805061346001
License date: Feb 09, 2016
Schenk & Aronsen (2015). Contribution of Impulsivity and Serotonin Receptor
Neuroadaptations to the Development of an MDMA (“ecstasy”) Substance Use
Disorder, in Current Topics in Behavioral Neuroscience: The Misuse of Licit
and Illicit Drugs in Psychopharmacology Susan Schenk, Suzanne Nielson,
Raimondo Bruno (Eds), Springer. DOI 10.1007/7854_2015_421
Licensed content publisher: Springer
License Number: 3805070143872
License date: Feb 09, 2016
Aronsen, Bukholt, & Schenk (2016). Repeated administration of the 5-HT1B/1A agonist,
RU 24969, facilitates the acquisition of MDMA self-administration: Role of 5-
HT1A and 5-HT1B receptor mechanisms. Psychopharmacology, 233 (8), 1339-
1347. DOI 10.1007/s00213-016-4225-x
Licensed content publisher: Springer
License Number: 3805490889197
License date: Feb 10, 2016
Aronsen & Schenk (2016). MDMA self-administration fails to alter the behavioral
response to 5-HT1A and 5-HT1B agonists. Psychopharmacology, 233 (7), 1323-
1330. DOI 10.1007/s00213-016-4226-9

127
Licensed content publisher: Springer
License Number: 3805491012086
License date: Feb 10, 2016
Related Documents





![[MDMA]MDMA Neurochemistry](https://static.cupdf.com/doc/110x72/577dab601a28ab223f8c57f3/mdmamdma-neurochemistry.jpg)





