Epilepsia, 49(8):1348–1357, 2008 doi: 10.1111/j.1528-1167.2008.01581.x FULL-LENGTH ORIGINAL RESEARCH The renin-angiotensin system is upregulated in the cortex and hippocampus of patients with temporal lobe epilepsy related to mesial temporal sclerosis ∗ Gustavo Adolfo Arga˜ naraz, ∗ Ana Carla Konno, ∗ Sandra Regina Perosa, ∗ Joselita Ferreira Carvalho Santiago, †Mirian A. Boim, ‡Daniela Berguio Vidotti, ∗ Pedro Paulo Vasconcelos Varella, ‡Luciana Gilbert Costa, §Mauro Canzian, ¶Marim ´ elia Aparecida Porcionatto, #Elza M´ arcia Yacubian, #Am´ erico Ceiki Sakamoto, #Henrique Carrete Jr, #Ricardo Silva Centeno, ∗ D´ ebora Amado, ∗ Esper Abr ˜ ao Cavalheiro, ∗∗ Jos´ e Antonio Silva Junior, and ∗ ¶Maria da Grac ¸a Naffah Mazzacoratti Department of Neurology and Neurosurgery, Disciplines of ∗ Experimental Neurology and †Nephrology; and ‡Department of Pathology, Federal University of S˜ ao Paulo (UNIFESP), S˜ ao Paulo, SP, Brazil; §Department of Pathology, Medical School of S˜ ao Paulo University, S˜ ao Paulo, SP, Brazil; ¶Department of Biochemistry and #Department of Neurology and Neurosurgery, Discipline of Clinical Neurology, Federal University of S˜ ao Paulo (UNIFESP), S ˜ ao Paulo, SP, Brazil; and ∗∗ Department of Sciences of Rehabilitation, Nove de Julho University (UNINOVE), S˜ ao Paulo, SP, Brazil SUMMARY Purpose: As reported by several authors, an- giotensin II (AngII) is a proinflammatory molecule that stimulates the release of inflammatory cy- tokines and activates nuclear factor κ B (NFκ B), being also associated with the increase of cellular oxidative stress. Its production depends on the ac- tivity of the angiotensin converting enzyme (ACE) that hydrolyzes the inactive precursor angiotensin I (AngI) into AngII. It has been suggested that AngII underlies the physiopathological mechanisms of several brain disorders such as stroke, bipolar dis- order, schizophrenia, and disease. The aim of the present work was to localize and quantify AngII AT1 and AT2 receptors in the cortex and hip- pocampus of patients with temporal lobe epilepsy related to mesial temporal sclerosis (MTS) sub- mitted to corticoamygdalohippocampectomy for seizure control. Method: Immunohistochemistry, Western blot, and real-time PCR techniques were employed to analyze the expression of these receptors. Results: The results showed an upregulation of AngII AT1 receptor as well as its messenger ri- bonucleic acid (mRNA) expression in the cortex and hippocampus of patients with MTS. In addi- tion, an increased immunoexpression of AngII AT2 receptors was found only in the hippocampus of these patients with no changes in its mRNA levels. Discussion: These data show, for the first time, changes in components of renin-angiotensin sys- tem (RAS) that could be implicated in the phys- iopathology of MTS. KEY WORDS: Angiotensin II AT1 and AT2 recep- tors, Temporal lobe epilepsy, Cortex, Hippocam- pus. Accepted February 15, 2008; Online Early publication March 21, 2008. Address correspondence to Maria da Grac ¸a Naffah Mazzacoratti, Ph.D., Rua Botucatu 862, Edif´ ıcio Leal Prado, S˜ ao Paulo, SP, Brazil. CEP: 04023900. E-mail: [email protected] Wiley Periodicals, Inc. C 2008 International League Against Epilepsy The renin-angiotensin system (RAS) participates in the regulation of several physiological processes including body water balance, maintenance of blood pressure, fluc- tuation of reproductive hormones, sexual behavior, and regulation of pituitary gland hormones. In this system, angiotensinogen is the inactive precursor and after its 1348

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x
FULL-LENGTH ORIGINAL RESEARCH
The renin-angiotensin system is upregulated in thecortex and hippocampus of patients with temporallobe epilepsy related to mesial temporal sclerosis
∗Gustavo Adolfo Arganaraz, ∗Ana Carla Konno, ∗Sandra Regina Perosa,∗Joselita Ferreira Carvalho Santiago, †Mirian A. Boim, ‡Daniela Berguio Vidotti,
∗Pedro Paulo Vasconcelos Varella, ‡Luciana Gilbert Costa, §Mauro Canzian,¶Marimelia Aparecida Porcionatto, #Elza Marcia Yacubian, #Americo CeikiSakamoto, #Henrique Carrete Jr, #Ricardo Silva Centeno, ∗Debora Amado,
∗Esper Abrao Cavalheiro, ∗∗Jose Antonio Silva Junior, and∗¶Maria da Graca Naffah Mazzacoratti
Department of Neurology and Neurosurgery, Disciplines of ∗Experimental Neurology and †Nephrology; and‡Department of Pathology, Federal University of Sao Paulo (UNIFESP), Sao Paulo, SP, Brazil; §Department of
Pathology, Medical School of Sao Paulo University, Sao Paulo, SP, Brazil; ¶Department of Biochemistry and#Department of Neurology and Neurosurgery, Discipline of Clinical Neurology, Federal University of Sao Paulo
(UNIFESP), Sao Paulo, SP, Brazil; and ∗∗Department of Sciences of Rehabilitation, Nove de Julho University(UNINOVE), Sao Paulo, SP, Brazil
SUMMARYPurpose: As reported by several authors, an-giotensin II (AngII) is a proinflammatory moleculethat stimulates the release of inflammatory cy-tokines and activates nuclear factor κB (NFκB),being also associated with the increase of cellularoxidative stress. Its production depends on the ac-tivity of the angiotensin converting enzyme (ACE)that hydrolyzes the inactive precursor angiotensin I(AngI) into AngII. It has been suggested that AngIIunderlies the physiopathological mechanisms ofseveral brain disorders such as stroke, bipolar dis-order, schizophrenia, and disease. The aim of thepresent work was to localize and quantify AngIIAT1 and AT2 receptors in the cortex and hip-pocampus of patients with temporal lobe epilepsyrelated to mesial temporal sclerosis (MTS) sub-mitted to corticoamygdalohippocampectomy forseizure control.
Method: Immunohistochemistry, Western blot,and real-time PCR techniques were employed toanalyze the expression of these receptors.Results: The results showed an upregulation ofAngII AT1 receptor as well as its messenger ri-bonucleic acid (mRNA) expression in the cortexand hippocampus of patients with MTS. In addi-tion, an increased immunoexpression of AngII AT2receptors was found only in the hippocampus ofthese patients with no changes in its mRNA levels.Discussion: These data show, for the first time,changes in components of renin-angiotensin sys-tem (RAS) that could be implicated in the phys-iopathology of MTS.KEY WORDS: Angiotensin II AT1 and AT2 recep-tors, Temporal lobe epilepsy, Cortex, Hippocam-pus.
Accepted February 15, 2008; Online Early publication March 21, 2008.Address correspondence to Maria da Graca Naffah Mazzacoratti,
Ph.D., Rua Botucatu 862, Edifıcio Leal Prado, Sao Paulo, SP, Brazil. CEP:04023900. E-mail: [email protected]
Wiley Periodicals, Inc.C© 2008 International League Against Epilepsy
The renin-angiotensin system (RAS) participates in theregulation of several physiological processes includingbody water balance, maintenance of blood pressure, fluc-tuation of reproductive hormones, sexual behavior, andregulation of pituitary gland hormones. In this system,angiotensinogen is the inactive precursor and after its
1348

1349
Angiotensin System is Related to Temporal Lobe Epilepsy
cleavage by the protease renin, the angiotensin I (AngI)is released. AngI, which has very low biological activity,is the main substrate for angiotensin converting enzyme(ACE), which generates angiotensin II (AngII), the mainactive peptide. After that, a cascade of events could be trig-gered, leading to the release of diverse angiotensin-relatedpeptides with a variety of biological effects (for review, seeWright & Harding, 2004).
Although widely distributed in peripheral systems, allRAS components have also been found in the brain wherethey actively modulate several functions, including ex-ploratory behavior, stress, anxiety, learning, and mem-ory acquisition (Chen et al., 1993; Meffert et al., 1996;Yamada et al., 1996; Wright & Harding, 2004; Saavedraet al., 2005). Interestingly, AngII has been considered tobe the major active peptide of the RAS, acting in theperiphery as a paracrine and endocrine hormone. Thispolypeptide could also be considered as a neurotransmitter/neuromodulator through its action on AT1 and AT2 re-ceptors present in several brain regions (Wright et al.,2002). In addition, growing evidence has emerged indicat-ing the RAS participation’s in some neurodegenerative dis-orders such as Alzheimer’s (Barnes et al., 1991; Tian et al.,2004), Parkinson’s (Grammatopoulos et al., 2005), andHuntington’s (Bird, 1980) diseases.
Recent works of Tchekalarova (Tchekalarova et al.,2005; Tchekalarova & Georgiev, 2005, 2006) indicatethat angiotensin-related peptides may have anticonvul-sant activity in pentylenetetrazol (PTZ), bicuculline- andpicrotoxin-induced seizures. Although the mechanisms bywhich RAS peptides could exert the anticonvulsant activ-ity are not completely understood, these authors suggestedthat the underlying process could involve the participationof other neurotransmitters (Wright et al., 2002). In thissense, AngII was shown to be able to increase the thresholdfor PTZ-induced seizures through a dopaminergic-relatedmechanism as dopamine (DA) receptor (D1/D2) agonistsor DA uptake inhibitors increase while DA antagonists at-tenuate the above-mentioned AngII action (Georgiev et al.,1985). Furthermore, an involvement of GABAergic in-terneurons mediating the inhibitory effect of AngII wasalso suggested in the basolateral amygdala, where GABAA
receptor antagonist effectively blocked the discharge ratedecrease normally induced by AngII (Tchekalarova et al.,2005). In addition, results in mice have shown that theincreased threshold of PTZ-induced seizures exerted byadenosine analogues was attenuated by AngII receptor an-tagonists (for review, see Tchekalarova & Georgiev, 2005).Interestingly, Das (2005) has shown that AngII stimulatesthe release of proinflammatory cytokines, activates NFκB,and increases oxidative stress—a group of activities thathas been considered to facilitate neuronal loss. These find-ings would justify the association of AngII with the phys-iopathology of stroke, bipolar disorder, schizophrenia, andAlzheimer’s disease. Thus, ACE inhibitors and AngII re-
ceptor antagonists should be considered important coadju-tants in the treatment of chronic inflammatory brain dis-eases.
Temporal lobe epilepsy related to mesial temporal lobesclerosis (MTS) is characterized by hippocampal sclero-sis, which includes atrophy, induration of shrunken tissue,granule cell loss in the dentate gyrus and extensive pyra-midal cell loss in Ammon’s horn, and astroglial prolifera-tion in the hippocampus (Babb et al., 1991). The predom-inant cell loss occurs in pyramidal neurons of the stratumpyramidale of Ammons horn. Neuronal population of thestratum oriens survives in most sclerotic hippocampi, alongwith interneurons scattered throughout the neuron-depletedpyramidal layer. In the dentate gyrus, many of the gran-ule cells are lost and neurons in the granule cell layer ap-pear to be more dispersed. Thus, several lines of evidencepoint to sclerotic hippocampus as the major structure in-volved in chronic seizures observed in MTS (for review,see Lanerolle & Lee, 2005).
Based on these data, the present work was designed tostudy the expression of AngII receptors (AT1 and AT2) inthe cortex and hippocampus obtained from patients withMTS, submitted to surgery for seizure control. With thispurpose, double staining procedure associated to Westernblot was employed to localize and quantify these alter-ations, respectively. In addition, AngII AT1 and AT2 re-ceptors mRNA levels were also analyzed using real-timePCR to study the synthesis of these two receptors in bothcortex and hippocampus tissues. All patients presented hip-pocampal sclerosis visualized by image analysis and bypathological studies. The expression of AngII receptorsand their respective mRNA were also studied in hippocam-pal and cortical areas removed during autopsy of subjectswithout neurological disease, which were used as controltissues.
METHODS
All experiments were performed under approval fromthe Institutional Ethics Committee of the UniversidadeFederal de Sao Paulo (UNIFESP). Surgical specimensfrom patients with intractable epilepsy were submitted tostandard corticoamygdalohippocampectomy at the Hos-pital Sao Paulo (UNIPETE, Brazil). All cases showingneoplasm, vascular malformations, post-traumatic and is-chemic lesions on preoperative MRI were excluded. Se-lected patients (n = 12) had detailed anamnesis, video-EEG recordings, and MRI studies. The age of patients withepilepsy varied from 19 to 64 years (38.8 ± 17 years). Con-trol tissues were obtained from autopsies (less than 5 hof postmortem) and the age of the subjects at death var-ied from 28 to 64 years (47.25 ± 18 years) (n = 12). Theantiepileptic drugs used by these patients for seizure con-trol included: carbamazepine, phenobarbital, diphenylhy-dantoin, clobazam, valproic acid, and topiramate. Surgery
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x
1350
G. A. Arganaraz et al.
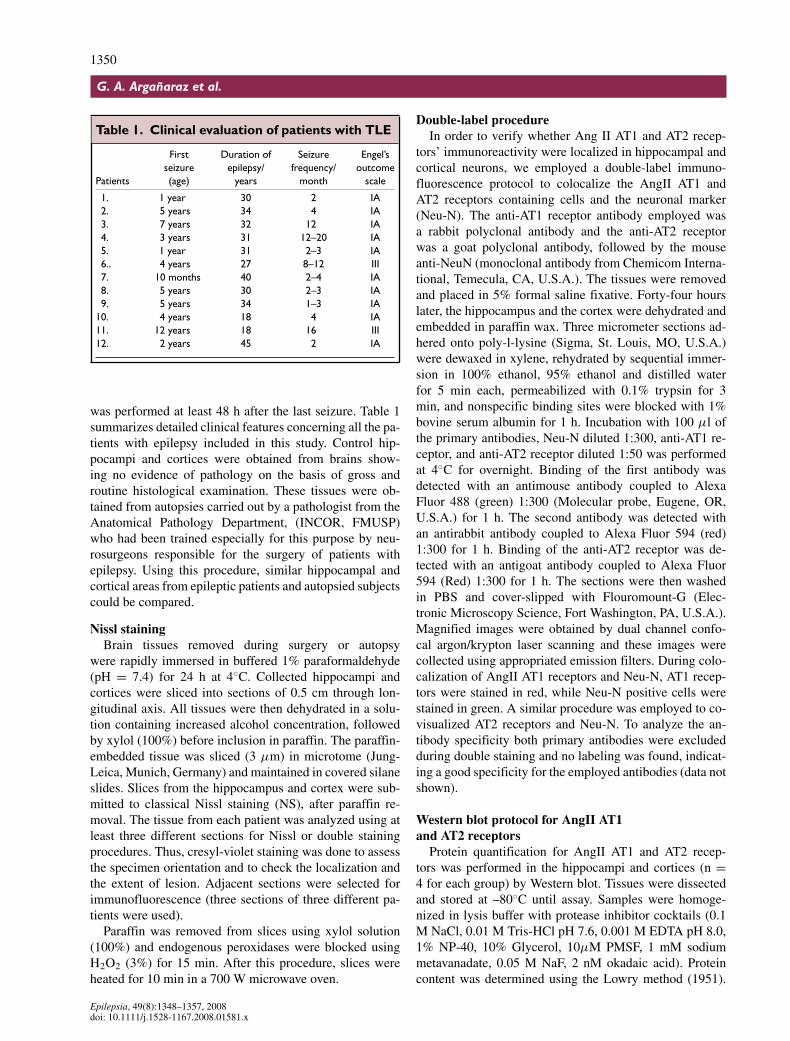
Table 1. Clinical evaluation of patients with TLE
First Duration of Seizure Engel’sseizure epilepsy/ frequency/ outcome
Patients (age) years month scale
1. 1 year 30 2 IA2. 5 years 34 4 IA3. 7 years 32 12 IA4. 3 years 31 12–20 IA5. 1 year 31 2–3 IA6.. 4 years 27 8–12 III7. 10 months 40 2–4 IA8. 5 years 30 2–3 IA9. 5 years 34 1–3 IA
10. 4 years 18 4 IA11. 12 years 18 16 III12. 2 years 45 2 IA
was performed at least 48 h after the last seizure. Table 1summarizes detailed clinical features concerning all the pa-tients with epilepsy included in this study. Control hip-pocampi and cortices were obtained from brains show-ing no evidence of pathology on the basis of gross androutine histological examination. These tissues were ob-tained from autopsies carried out by a pathologist from theAnatomical Pathology Department, (INCOR, FMUSP)who had been trained especially for this purpose by neu-rosurgeons responsible for the surgery of patients withepilepsy. Using this procedure, similar hippocampal andcortical areas from epileptic patients and autopsied subjectscould be compared.
Nissl staining
Brain tissues removed during surgery or autopsywere rapidly immersed in buffered 1% paraformaldehyde(pH = 7.4) for 24 h at 4◦C. Collected hippocampi andcortices were sliced into sections of 0.5 cm through lon-gitudinal axis. All tissues were then dehydrated in a solu-tion containing increased alcohol concentration, followedby xylol (100%) before inclusion in paraffin. The paraffin-embedded tissue was sliced (3 μm) in microtome (Jung-Leica, Munich, Germany) and maintained in covered silaneslides. Slices from the hippocampus and cortex were sub-mitted to classical Nissl staining (NS), after paraffin re-moval. The tissue from each patient was analyzed using atleast three different sections for Nissl or double stainingprocedures. Thus, cresyl-violet staining was done to assessthe specimen orientation and to check the localization andthe extent of lesion. Adjacent sections were selected forimmunofluorescence (three sections of three different pa-tients were used).
Paraffin was removed from slices using xylol solution(100%) and endogenous peroxidases were blocked usingH2O2 (3%) for 15 min. After this procedure, slices wereheated for 10 min in a 700 W microwave oven.
Double-label procedure
In order to verify whether Ang II AT1 and AT2 recep-tors’ immunoreactivity were localized in hippocampal andcortical neurons, we employed a double-label immuno-fluorescence protocol to colocalize the AngII AT1 andAT2 receptors containing cells and the neuronal marker(Neu-N). The anti-AT1 receptor antibody employed wasa rabbit polyclonal antibody and the anti-AT2 receptorwas a goat polyclonal antibody, followed by the mouseanti-NeuN (monoclonal antibody from Chemicom Interna-tional, Temecula, CA, U.S.A.). The tissues were removedand placed in 5% formal saline fixative. Forty-four hourslater, the hippocampus and the cortex were dehydrated andembedded in paraffin wax. Three micrometer sections ad-hered onto poly-l-lysine (Sigma, St. Louis, MO, U.S.A.)were dewaxed in xylene, rehydrated by sequential immer-sion in 100% ethanol, 95% ethanol and distilled waterfor 5 min each, permeabilized with 0.1% trypsin for 3min, and nonspecific binding sites were blocked with 1%bovine serum albumin for 1 h. Incubation with 100 μl ofthe primary antibodies, Neu-N diluted 1:300, anti-AT1 re-ceptor, and anti-AT2 receptor diluted 1:50 was performedat 4◦C for overnight. Binding of the first antibody wasdetected with an antimouse antibody coupled to AlexaFluor 488 (green) 1:300 (Molecular probe, Eugene, OR,U.S.A.) for 1 h. The second antibody was detected withan antirabbit antibody coupled to Alexa Fluor 594 (red)1:300 for 1 h. Binding of the anti-AT2 receptor was de-tected with an antigoat antibody coupled to Alexa Fluor594 (Red) 1:300 for 1 h. The sections were then washedin PBS and cover-slipped with Flouromount-G (Elec-tronic Microscopy Science, Fort Washington, PA, U.S.A.).Magnified images were obtained by dual channel confo-cal argon/krypton laser scanning and these images werecollected using appropriated emission filters. During colo-calization of AngII AT1 receptors and Neu-N, AT1 recep-tors were stained in red, while Neu-N positive cells werestained in green. A similar procedure was employed to co-visualized AT2 receptors and Neu-N. To analyze the an-tibody specificity both primary antibodies were excludedduring double staining and no labeling was found, indicat-ing a good specificity for the employed antibodies (data notshown).
Western blot protocol for AngII AT1
and AT2 receptors
Protein quantification for AngII AT1 and AT2 recep-tors was performed in the hippocampi and cortices (n =4 for each group) by Western blot. Tissues were dissectedand stored at –80◦C until assay. Samples were homoge-nized in lysis buffer with protease inhibitor cocktails (0.1M NaCl, 0.01 M Tris-HCl pH 7.6, 0.001 M EDTA pH 8.0,1% NP-40, 10% Glycerol, 10μM PMSF, 1 mM sodiummetavanadate, 0.05 M NaF, 2 nM okadaic acid). Proteincontent was determined using the Lowry method (1951).
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x

1351
Angiotensin System is Related to Temporal Lobe Epilepsy
Samples were diluted in Laemmli buffer and boiled for5 min. Two standard curves were performed using sev-eral protein concentrations. A linear range was obtainedusing 40 μg of protein for AngII AT1 receptor and 80μg for AngII AT2 receptor. Thus, an equivalent amount ofprotein was electrophored on 10% polyacrylamide mini-gels and transferred to nitrocellulose membrane by elec-troblotting. Blocking was performed in 5% nonfat milkfor 2 h at room temperature. Thereafter, blots were thenprobed with an affinity-purified goat polyclonal antibodyraised against a peptide mapping near the carboxy termi-nus of rabbit AT1 receptor (1:150, Santa Cruz Biotechnol-ogy, Inc., Santa Cruz, CA, U.S.A.) or with a goat poly-clonal antibody raised against a peptide mapping the C-terminus of AT2 receptor (1:150, Santa Cruz Biotechnol-ogy, Inc) in TBS-T (50 mM Tris-HCl, 154 mM NaCl,pH7.5 + 0.1% Tween 20) plus 2% nonfat milk. Afterovernight incubation, blots were washed in TBS-T (3 ×10 min) and incubated for 2 h in biotinylated antigoator antirabbit immunoglobulin (Calbiochem, Darmstadt,Germany), diluted 1:150 in TBS-T plus 2% nonfat milk.Blots were washed in TBS-T (3 × 10 min) and incu-bated for 1 h in streptavidin-horseradish peroxidase (VectorLaboratories, Inc., Burlingame, CA, U.S.A.). After wash-ing, enhanced chemiluminescence reagents (ECL, Amer-shan Pharmacia Biotech, Cardiff, UK) were applied tothe blots. The membranes were then exposed to an x-rayfilm (Amersham) and the bands were quantified by den-sitometry. Equal protein loading was shown by strippingand reprobing the membrane with an anti-glyceraldehyde3-phosphate dehydrogenase (GAPDH) antibody (1:1000,Proteimax Biotecnologia Ltda., Sao Paulo, Brazil).
Western blot data analysis
The ratio between optical density of AngII AT1 and AT2receptors and the internal standard GAPDH bands are pre-sented as means ± standard deviation (SD). Significanceof differences was assessed by Student’s t-test and p-value<0.05 was considered significant.
Quantitative real-time TaqMan PCR for angiotensin
receptors
Dissected hippocampi and cortices were frozen in liquidnitrogen and stored at –80◦C. Thawed tissue was homoge-nized in 1 ml TRIzol reagent (Gibco BRL, Gaithersburg,MD, U.S.A.) and total RNA was isolated according to themanufacturer’s instructions. Samples were submitted to a20 μl reaction using TaqMan Amplification System witha Mastercycler Realplex (Eppendorf, Wesseling-Berzdorf,Germany). Real-time PCR was performed with 900 ng ofcDNA for AT1 and AT2 receptors and 100 ng of cDNA forGAPDH, used as an internal standard (GAPDH mRNAaccession number NM 002046). Beta2-microglobulin wasused as a second internal standard (β2 microglobulinmRNA accession number AF072097). Oligonucleotide
primer and fluorogenic probe sets for TaqMan real-timePCR were designed for AT1 and AT2 receptors andGAPDH or beta2-microglobulin using Assays-by-DesignService (Applied Biosystems, Foster City, CA, U.S.A.)to meet all TaqMan design guidelines. Probes weresynthesized with a reporter dye 6-carboxyfluorescein (6-FAM) covalently linked at the 5′ end and a quencher dye6-carboxy-tetramethyl-rhodamine (TAMRA) was linkedto the 3′ end of the probe. AT1 and AT2 probes used were:5′-FAM-CGACGCACAATGCTTGTAGCCAAAGTCA-TAMRA-3′ and 5′-FAM-TTGTTCTGGCCTTCATCATTTGCTGGC-TAMRA-3′, respectively. The utilized primerswere:
Primers Sense Anti-sense
AT1 5′AGA TGA CGG CTG 5′ AGG ACA AAA GCAreceptor CTC GAA GA 3′ TAG GGA G 3′
AT2 5′ CAA AGC ATT CTG 5′ TCC TTA TGC CTT TGGreceptor CAG CCT GA 3′ TTG TTG TTG AAG T 3′
Each reaction was carried out with 10 μl of RealMas-terMix Probe (Eppendorf) and 1 μl of a mix containingtwo primers (18 μM each) and a probe (5 μM), specificto mRNA of angiotensin AT1 receptor or angiotensin AT2receptor. The cycle conditions were: 50◦C for 2 min, then95◦C for 10 min, followed by 50 cycles of 95◦C for 15 s(melting step), 60◦C for 1 min (anneal/extend step). Eachgroup was composed of five subjects.
Gene expression data analysis
Increased amount of reporter dye fluorescence duringthe 50 cycles of amplification were monitored using Ep-pendorf Cycle Manager. The PCR cycle when a given flu-orescence threshold is crossed by the amplification curveis considered the first parameter to analyze mRNA expres-sion and named Ct. The bigger the initial copies amount,the smaller the Ct number. A normalized value is obtainedby subtracting Ct of GAPDH or β2 microglobulin fromCt of AT1 and AT2 receptors, resulting in �Ct. As it is un-common to use �Ct as a relative expression data due to thislogarithmic characteristic, the 2−�Ct parameter was used toexpress the relative expression data, which were analyzedas mean ± SD. Hippocampi and cortices from patients withMTS and control tissue were compared using Student’s t-test. A value of p < 0.05 was considered to be statisticallysignificant.
RESULTS
Nissl-stained cortices from patients with epilepsy pre-sented no difference with regards to cellular distributionin all layers of temporal cortex when compared to con-trol tissues (data not shown). However, this staining re-vealed hippocampal sclerosis in all patients with epilepsy,
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x

1352
G. A. Arganaraz et al.
Figure 1.Nissl staining of control and sclerotic hippocampus:(A) control, (B) sclerotic hippocampus (scale bars 400um).Epilepsia C© ILAE
confirming data obtained during MRI analysis, which pre-viously demonstrated atrophy, signal changes, and abnor-mal shape of the hippocampus. The hippocampi of patientswith epilepsy presented intense cell loss mainly in the hilusas well as in the CA1 and CA3 regions, when comparedwith the control area. The hippocampi of the control groupshowed a normal pattern of cell distribution (Fig. 1).
The double staining showed intense AngII AT1 labelingof pyramidal neurons, colocalizing with Neu-N protein, incontrol and epileptic tissues. As shown (Fig. 2, top panel)in the cortex of patients with epilepsy the density of AngIIAT1 receptor is increased in each neuron, mainly in apicalregions as clusters, near the plasma membrane (Fig. 2, toppanel). This type of labeling was less visualized in controlcortex.
The double staining of AngII AT1 receptor and Neu-Nalso showed increased expression of AngII AT1 receptor inCA3 regions of sclerotic hippocampus and apical regionsof neurons were more stained (Fig. 2, top panel). In con-trast, granule cells from patients showed no immunoreac-tivity against AngII AT1 receptors (Fig. 3), demonstratingthat more vulnerable cells are preferentially stained. Onlyhilar neurons were double-labeled.
Western blot analysis indicated a 55 kDa band, identi-fied as Ang II AT1 receptor. Optical density analysis, af-ter normalization with GAPDH (35–40 kDa) confirmed anincreased expression of AT1 receptor in the hippocampus(0.425 ± 0.04, p = 0.038) and cortex (0.386 ± 0.03, p =0.012) of patients with epilepsy, when compared to controlhippocampus (0.331 ± 0.03) and cortex (0.277 ± 0.03),respectively (Fig. 4, top and lower panels). In addition,quantification of mRNA for AngII AT1 receptor by real-time PCR demonstrated an increased level of this mRNAin the hippocampus (0.55 ± 0.105, p = 0.00057) and cortex(2.5 ± 0.179, p = 5.49 × 10−9) of patients with epilepsy,when compared with the control hippocampus (0.33 ±0.034) and control cortex (0.86 ± 0.089) (Fig. 6, top panel).These data demonstrated an upregulation of AngII AT1
Figure 2.Double staining: colocalization of NeuN and AngII AT1or AT2 receptor in the temporal cortex and hippocam-pus (n = 3). NeuN is stained in green while AngIIAT1 and AT2 receptors are stained in red. The toppanel shows immunostaining of AngII AT1 receptorsof control tissues on right panel and epileptic tissue onleft panel. The lower panel shows immunostaining ofAngII AT2 receptors of control tissue on right paneland epileptic tissue on left panel.Epilepsia C© ILAE
receptors in the cortex and hippocampus of patients withMTS.
The double staining for AngII AT2 receptors and NeuNshowed that in the temporal cortex no visible differencewas found concerning AT2 receptor expression in the con-trol and epileptic tissue. This procedure allows us to visu-alize few neurons stained in control and epileptic cortex,showing no difference between them (Fig. 2, lowerpanel).Vessels and glial-processes were also found in bothtissues.
In contrast, an increased immunoreactivity againstAngII AT2 receptors was found in sclerotic hippocam-pus. Magnifications of this figure revealed few cells in
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x

1353
Angiotensin System is Related to Temporal Lobe Epilepsy
Figure 3.Double staining: colocalization of NeuN and AngII AT1receptor in the dentate gyrus of patient with hippocam-pal sclerosis. Note that the granular cells were notstained by anti-AT1 receptor while hilar cells are im-munolabeled.Epilepsia C© ILAE
Figure 4.Western blot analysis of AngII AT1 receptor in the cor-tex (top panel) and hippocampus (lower panel) ofcontrol and epileptic tissues. Representative Westernblot analysis: rows: 1, 2, 3, 4—control tissues; 5, 6, 7,8—epileptic tissues. ∗p = 0.012 (top panel); ∗p = 0.038(lower panel), control versus epileptic tissues (n = 4per group).Epilepsia C© ILAE
CA3 region immunostained against AngII AT2 receptors inthe hippocampus of these patients. In addition, the doublestaining of AngII AT2 receptor and Neu-N also revealedthat besides pyramidal cells, several fibers, glial processes,and vessels were visualized in the sclerotic hippocampus(Fig. 2, lower panel). However, granular cells were not la-beled in sclerotic hippocampus (data not shown).
Western blot analysis indicated a 50-kDa band, identi-fied as AngII AT2 receptor. Optical density determination,after normalization with GAPDH (35–40 Kda), showed anincreased expression of AngII AT2 receptor in the hip-pocampus (0.83 ± 0.07) of patients with epilepsy, whencompared with control tissue (0.307 ± 0.07, p = 0.0532 ×10−3). No difference was observed in the temporal cortex(2.17 ± 0.14, p = 0.288) of these patients, when comparedto controls (2.04 ± 0.13) (Fig. 5, lower panel).
Although Western blot has shown increased expressionof AngII AT2 receptor in the sclerotic hippocampus, thequantification of their mRNA expression by real-time PCRshowed similar results in the hippocampus of patients with
Figure 5.Western blot analysis of AngII AT2 receptor in the cor-tex and hippocampus of control and epileptic tissues.Representative Western blot analysis: rows 1, 2, 3, 4—control tissues; 5, 6, 7, 8—epileptic tissues. p = 0.2885(top panel), ∗∗p = 5.32.10–5 (lower panel), controlversus epileptic tissues (n = 4 per group).Epilepsia C© ILAE
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x

1354
G. A. Arganaraz et al.
Figure 6.Top panel: quantification of AngII receptors mRNAexpression in the cortex and hippocampus. Top: levelof AngII AT1 receptors mRNA in the human cortexmeasured by real-time PCR assay. All data have beennormalized for levels of β2 microtubulin expressionwithin the same sample and are expressed as relativelevels detected in each group sample as 2 – �Ct. Dataare expressed as mean + SD (n = 5 per group). Lower:level of AngII AT1 receptors mRNA in the human hip-pocampus measured by real-time PCR assay. All datahave been normalized for levels of β2 microtubulin ex-pression within the same sample and are expressedrelative to levels detected in each group sample as2–�Ct. Data are expressed as mean + SD (n = 5 perEpilepsia C© ILAE
epilepsy (0.13 ± 0.016) and control subjects (0.12 ± 0.024,p = 0.348) as well as in the cortex of patients (0.22 ±0.018) and control subjects (0.20 ± 0.028, p = 0.437)(Fig. 6, top and lower panel). All together, these datashowed that although AngII AT2 receptor is upregulated inthe hippocampus of epileptic patients, its synthesis is notinduced during an interictal period. Concerning the epilep-tic cortex, no difference was found related to the expres-sion of AngII AT2 receptor. The employment of GAPDHas housekeeping on RT–PCR did not modify the obtainedresults (data not shown).
DISCUSSION
In the present work we showed, for the first time, a rela-tionship between human temporal lobe epilepsy related tomesial temporal sclerosis (MTS) and the RAS. Increasedamount of AngII AT1 receptors was found in neurons fromcortex and hippocampus of patients with MTS. In addi-tion, mRNA for AngII AT1 receptor was increased in bothtissues from these patients showing that the expressionas well as the synthesis of this receptor is upregulated inpatients with MTS. This study also showed an upregu-lation of AngII AT2 receptor in the hippocampus of pa-tients with MTS, although its synthesis was not modifiedduring the interictal period of analysis. Altogether, theseresults strongly suggest the participation of angiotensinsystems in the pathophysiology of temporal lobe epilepsy(TLE) associated with mesial sclerosis.
AngII AT1 and AT2 receptors have similar but not iden-tical brain distribution in all mammalian species studied sofar. AngII AT1 receptor expression predominates in adultanimals while AngII AT2 receptors are highly expressed indeveloping brain (Tsutsumi & Saavedra, 1991; Saavedra,
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−group). Lower panel: quantification of AngII recep-tors mRNA expression in the cortex and hippocampus.Top: level of AngII AT2 receptors mRNA in the humancortex measured by real-time PCR assay. All data havebeen normalized for levels of β2 microtubulin expres-sion, within the same sample and are expressed asrelative levels detected in each group sample as 2 –�Ct. Data are expressed as mean + SD (n = 5 pergroup). Lower: level of AngII AT2 receptors mRNA inthe human hippocampus, measured by real-time PCRassay. All data have been normalized for the levels ofβ2 microtubulin, within the same sample and are ex-pressed relative to levels detected in each group sampleas 2 – �Ct. Data are expressed as mean + SD (n = 5per group).Epilepsia C© ILAE
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x

1355
Angiotensin System is Related to Temporal Lobe Epilepsy
2005). Despite the fact that AngII AT1 receptor is almostentirely responsible for the classical effects of AngII, ithas been found that AngII AT2 receptor is expressed ata low amount in many healthy adult tissues and is up-regulated in a variety of human diseases. AngII AT2 re-ceptors have been related to ionic fluxes, cell differenti-ation, neuronal tissue regeneration, and programmed celldeath in the brain. According to Wilms et al. (2005), AngIIAT2 receptor could also be involved in neuronal protectionfollowing brain injury, such as stroke or CNS trauma, whileAngII AT1 receptor might act as a receptor linked to neu-ronal injury. As reported by Das (2005), the binding ofpeptide AngII to the AT1 receptor is also involved inthe stimulation of leukotriens, prostaglandins, free radi-cals generation, activation of the NFκB and increase ofC-reactive protein levels, triggering the expression ofproinflammatory and pro-oxidant genes. In addition, theassociation between inflammatory processes and temporallobe epilepsy is already well known (Vezzani & Granata,2005). Several inflammatory targets are upregulated intissues involved in this syndrome (Vezzani et al., 1999;Vezzani et al. 2000) and proinflammatory cytokines havebeen linked to the pathogenesis of seizures. Jamali et al.(2006) showed a dysregulation of the neurotransmissionand complement systems in the entorhinal cortex of hu-mans with temporal lobe epilepsy, showing the partici-pation of immune system with infiltration of C3 positiveleucocytes and membrane attack complex deposition onneurons. In addition, Lerner-Natoli et al. (2000) showedthat during long-lasting seizures the neuronal firing acti-vates NF-κB and the major histocompatibility complex ex-pression by glial cells of hippocampal formation, whichare also related to inflammatory processes of TLE. Wilmset al. (2005) have shown that AngII AT1 receptor, althoughlinked to inflammatory cascade production, is also associ-ated to growth factors induction in neurons and glia. Thisreceptor is also related with the modulation and prolifera-tion in various cell types, showing a dual function for thisreceptor.
ACE, the main enzyme of RAS, is also related to thekallikrein-kinin system, which is linked to the inflamma-tory process. In this regard our group recently showed anoverexpression of kinin B1 and B2 receptors in the hip-pocampus of patients with temporal lobe epilepsy, associ-ated with mesial sclerosis (Perosa et al., 2007). Together,these data suggest that both systems could be involved inseizure modulation or in epileptogenic processes, via in-flammatory pathways.
Interestingly, ACE gene polymorphism has also beenassociated with Alzheimer’s disease being related to hip-pocampus and amygdala atrophy, showing that the RASsystem is involved with brain pathologies, linked to severecell loss and brain degeneration (Sleegers et al., 2005). Al-though the present work did not evaluate ACE expression,the differences reported here concerning the expression of
angiotensin-related receptors between control and sclerotichippocampus add new evidence to the participation of theRAS in brain diseases, associated with atrophy of impor-tant structures such as the hippocampus.
In the present work, the expression of AngII AT1 recep-tors was increased in the cortex and in the hippocampusof patients with temporal lobe epilepsy, while the AT2 re-ceptor was elevated only in the hippocampus of these pa-tients. These results suggest that changes in the release ofAngII may occur via not yet known mechanisms and thatthis polypeptide would have pro- or antiepileptogenic func-tion depending on its receptor activation. In this sense, ouridea follows those of Tchekalarova & Georgiev (2005) whodemonstrated the involvement of this system in seizuregenesis and/or control. According to them, the activationof AngII AT1 receptor elicits neuronal depolarization, viathe inhibition of potassium channels or by opening a nons-elective sodium–calcium channel, facilitating the dischargefiring in several brain regions. Increased c-Fos expressionwith consequent expression of immediate-early genes wasalso found, showing neuronal activation. In this context,our findings suggest that the upregulation of AngII AT1 re-ceptor is related to increased tissue excitability, in the hip-pocampus and cortical areas.
In addition, dopaminergic system mobilization hasbeen also reported. In vivo and in vitro studies re-vealed that AngII AT1 receptor activation potentates thedepolarization-induced release of DA as well as the sus-ceptibility to seizures. In this sense, a previous paper fromour group (Naffah-Mazzacoratti et al., 1996) has shown in-creased utilization rate of DA in the cortical tissue of pa-tients with temporal lobe epilepsy, a finding that now couldbe attributed to the present results on the upregulation ofangiotensin system. Tchekalarova & Georgiev (2006) re-ported the modulation of PTZ-induced seizures by AngIIAT1 receptor associated with a noradrenergic mechanism,mediated by the peptides AngII and AngIII.
Recently, Stragier et al. (2006) have also shown thatAngIV is able to inhibit pilocarpine-induced seizures viathe activation of somatostatin receptor 2. In this work,the authors observed that the administration of AngIV in-creased the hippocampal release of DA and serotonin, andthat this effect is blocked by somatostatin receptor 2 an-tagonist, a finding that confirms the participation of thesemonoamines in the modulation of seizures induced byangiotensin-related peptides.
The anticonvulsant effect of AngII has also been at-tributed to the observation that this compound may havea direct action on the GABA-benzodiazepine receptor–ionophore complex, probably through allosteric mech-anisms or by an interaction of AngII receptor withthe GABAA receptor (Tchekalarova & Georgiev, 2005).In addition, Schelman et al. (2004) and Jing et. al.(2004) found that AngII acting on AT2 receptors attenu-ates the N-methyl-D-aspartate (NMDA) receptor-mediated
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x

1356
G. A. Arganaraz et al.
neurotoxicity, via antiapoptotic mechanisms, altering theexpression level of Bcl-2 protein and rescuing cells fromdeath. In the present work, we found an increased expres-sion of AngII AT2 receptor in neurons from the hippocam-pus of patients with MTS, a region with severe cell loss.These data could be related with an AngII AT2 receptormediated effect of neuroprotection or regenerative process.In agreement with this idea, Grammatopoulos et al. (2004)reported that AngII attenuates chemical hypoxia-inducedcaspase-3 activation in primary cortical neuronal culturesvia the AT2 receptor and stimulates the p21 ras/mitogenactivated protein kinase pathway, which is known to in-duce neuronal survival signals. In addition, oxidative stressinduced apoptosis via upregulation of Jun-N-terminal ki-nases (JNK) could also be inhibited by AT2 receptor stim-ulation. Furthermore, Lucius et al. (1998) showed using theoptic nerve of adult rats that AngII, acting through the AT2receptor induces neurite outgrowth. Thus, the increased ex-pression of these receptors in the hippocampus may be re-lated to an attempt to control tissue degeneration.
According to Wilms et al. (2005), a cross-talking be-tween AngII AT1 and AT2 receptors may occur. In thissense, the understanding of how distinct signal transduc-tion elements are targeted by AT1 and AT2 receptors indifferent types of neurons will pave the way for a deeperinsight into the role of these receptors during health anddiseases of the nervous system.
In conclusion, the present work shows for the firsttime changes in the RAS in temporocortical areas of pa-tients with MTS and suggests its participation in the phys-iopathologic mechanisms underlying epileptogenesis.
ACKNOWLEDGMENTS
The research has been supported by FAPESP (CInAPCe), CAPES,CNPq, and FADA. The authors are grateful to all professionals fromUNIPETE, responsible for the clinical care and surgery of all patients.
Conflict of interest: We confirm that we have read the Journal’s positionon issues involved in ethical publication and affirm that this report is con-sistent with those guidelines. In addition, none of the authors has anyconflict of interest to disclose.
REFERENCES
Babb TL, Kupfer WR, Pretorius JK, Crandalle PH, Levesque MF. (1991)Synaptic recurrent excitation of granule cells mossy fiber in humanepileptic fascia dentata. Neuroscience 42:351–363.
Barnes NM, Cheng CHK, Costall B, Naylor RJ, Williams TJ, WischikCM. (1991) Angiotensin converting enzyme is increased in tempo-ral cortex from patients with Alzheimer disease. Eur J of Pharmacol200:289–292.
Bird ED. (1980) Chemical pathology of Huntington disease. Annu RevPharmacol Toxicol 20:533–551.
Chen L, Prakash O, Re RN. (1993) The interaction of insulin and an-giotensin II on the regulation of human neuroblastoma cell growth.Mol Chem Neuropathol 18:189–196.
Das UN. (2005) Is angiotensin II and endogenous pro-inflammatorymolecule? Med Sci Monit 11:RA155–162.
Georgiev V, Gyorgy L, Getova D, Markovska V. (1985) Some centraleffects of angiotensin II. Interaction with DAergic Transmission. ActaPhysiol et Pharmacol Bulg 11:19–26.
Grammatopoulos TN, Morris K, Bachar C, Moore S, Andre R, Weyhen-meyer JA. (2004) Angiotensin II attenuates chemical hypoxia-inducedcaspase-3 activation in primary cortical neuronal cultures. Brain ResBull 62:297–303.
Grammatopoulos TN, Ahmadi F, Jones SM, Fariss MW, WeyhenmeyerJA, Zawada WM. (2005) Angiotensin II protects cultured midbraindopaminergic neurons against rotenone-induced cell death. Brain Res1045:64–71.
Jamali S, Bartolomei F, Robaglia-Schlupp A, Massacrier A, Peragut JC,Regis J, Dufour H, Ravid, Roll P, Pereira S, Royer B, Roeckel-Trevisiol N, Fontaine M, Guye M, Boucraut J, Chauvel P, Cau P,Szepetowski P. (2006) Large-scale expression of human mesial tem-poral lobe epilepsy: evidence for dysregulation of the neurotransmis-sion and complement systems in entorhinal cortex. Brain 129:625–641.
Jing G, Grammatopoulos T, Ferguson P, Schelman W, Weyhernmeter J.(2004) Inhibitory effects of angiotensin on NMDS-induced cytotoxi-city in primary neuronal cultures. Brain Res Bull 62:397–403.
de Lanerolle NC, Lee TS. (2005) New facets of neuropathology andmolecular profile of human temporal lobe epilepsy. Epilepsy Behav7:190–203.
Lerner-Natoli M, Montpied P, Rousset MC, Bockaert J, Rondouin G.(2000) Sequencial expression of surface antigens and transcriptionfactor NFκB by hippocampal cells in excitotoxicity and experimen-tal epilepsy. Epilepsy Res 41:141–154.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. (1951) Protein mea-surement with the Folin phenol reagent. J Biol Chem 193:265–275.
Lucius R, Gallinat S, Rosentiel P, Herdegen T, Sievers J, Unger T. (1998)The angiotensin II type 2 (AT2) receptor promotes axonal regenera-tion in the optic nerve of adults rats. J Exp Med 188:661–670.
Meffert S, Stoll M, Stecklings UM, Bottari SP, Unger T. (1996) The an-giotensin II AT2 receptors inhibits proliferation and promotes differ-entiation in PC12W cells. Mol Cell Endocrinol 122:59–67.
Naffah-Mazzacoratti MG, Amado D, Cukiert A, Gronich G, Marino R,Calderazzo L, Cavalheiro EA. (1996) Monomines and their metabo-lites in cerebrospinal fluid and temporal cortex of epileptic patients.Epilepsy Res 25:133–137.
Perosa SR, Argnaraz GA, Goto EM, Costa LGP, Konno AC, VarellaPPV, Santiago JFC, Pesquero JB, Canzian M, Amado D, YacubianEM, Carrete H Jr, Centeno RS, Cavalheiro EA, Silva JA Jr, Naffah-Mazzacoratti MD. (2007) Kinin B1 and B2 receptors are overex-pressed in the hippocampus of humans with temporal lobe epilepsy.Hippocampus 17:26–33.
Saavedra JM. (2005) Brain angiotensin II: new development, unansweredquestions and therapeutic opportunities. Cell Mol Neurobiol 25:485–512.
Saavedra JM, Ando H, Armando I, Baiardi G, Bregonzio C, Juorio A,Macova M. (2005) Anti-stress and anti-anxiety effects of centrallyacting angiotensin II AT1 receptor antagonists. Regul Pept 128:227–238.
Schelman WR, Andres R, Ferguson P, Orr B, Kang E, Weyhenmeyer JA.(2004) Angiotensin II attenuates NMDA receptor-mediated neuronalcell death and prevents the associated reduction in Bcl-2 expression.Brain Res Mol Brain Res 128:20–29.
Sleegers K, den Heijer T, van Dijk EJ, Hofman A, Bertoli-Avella AM,Koudstaal PJ, Breteler MM, van Duijn CM. (2005) ACE gene is as-sociated with Alzheimer’s disease and atrophy of hippocampus andamygdala. Neurobiol Aging 26:1153–1159.
Stragier B, Clinckers R, Meurs A, De Bundel D, Sarre S, Ebinger G,Michotte Y, Smolders I. (2006) Involvement of the somatostatin-2 re-ceptor in anti-convulsant effect of angiotensin IV against pilocarpine-induced limbic seizures in rat. J Neurochem 98:1100–1113.
Tchekalarova J, Georgiev V. (2005) Angiotensin peptides modulatorysystem: how is it implicated in the control of seizure susceptibility ?Life Sci 76:955–970.
Tchekalarova J, Sotiriou E, Georgiev V, Kostopoulos G, AngelatouF. (2005) Up-regulation of adenosine A1 receptor binding inpentylenetetrazol kindling in mice: effects of angiotensin IV. BrainRes 1032:94–103.
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x

1357
Angiotensin System is Related to Temporal Lobe Epilepsy
Tchekalarova J, Georgiev V. (2006) AngII and AngII modulate PTZseizure threshold in non-stressed and stressed mice: possible involve-ment of noradrenergic mechanism. Neuropeptides 40:339–348.
Tian J, Shi J, Bailey K, Harris JM. Pritchard A, Lambert JC, Chartier-Harlin MC, Pickering-Brown SM, Lendon CL, Mann DM. (2004) Apolymorphism in the angiotensin 1-converting enzyme gene is associ-ated with damage to cerebral cortical white matter in Alzheimer dis-ease. Neurosci Lett 354:103–106.
Tsutsumi K, Saavedra JM. (1991) Characterization and development ofangiotensin II receptor subtypes (AT1 e AT2) in rat brain. Am J Phys-iol 261:R209–R2016.
Vezzani A, Conti M, De Luigi A, Ravizza T, Moneta D, Marchesi F,De Simoni MG. (1999) Interleukin 1 beta immunoreactivity and mi-croglia are enhanced in the rat hippocampus by focal kainate applica-tion: functional evidence for enhancement of electrographic seizures.J Neurosci 19:5054–5065.
Vezzani A, Moneta D, Conti M, Richichi C, Ravizza T, De Luigi A, DeSimoni MG, Sperk G, Andell-Jonsson S, Lundkvist J. Iverfeldt K,
Bartfai T. (2000). Powerful anticonvulsant action of IL-1 receptorantagonist on intracerebral injection and astrocytic overexpression inmice. Proc Natl Acad Sci USA 97:11534–11539.
Vezzani A, Granata T. (2005) Brain inflammation in epilepsy: experimen-tal and clinical evidence. Epilepsia 46:1724–1743.
Wilms H, Rosenstiel P, Unger T, Deuschl G, Lucius R. (2005) Neuro-protection with angiotensin receptor antagonists: a review of the ev-idence and potential mechanisms. Am J of Cardiovasc Drugs 5:245–253.
Wright JW, Reichert JR, Davis CJ, Harding JW. (2002) Neuronal plas-ticity and the brain rennin-angiotensin system. Neurosci Biochem Rev26:529–552.
Wright JW, Harding J. (2004) The brain angiotensin system and extracel-lular matrix molecules in neural plasticity, learning and memory. ProgNeurobiol 72:263–293.
Yamada T, Horiuchi M, Dzau VJ. (1996) Angiotensin II type 2 receptormediates programmed cell death. Proc Natl Acad Sci USA 93:156–160.
Epilepsia, 49(8):1348–1357, 2008doi: 10.1111/j.1528-1167.2008.01581.x
Related Documents











