ENVIRONMENTAL BIOTECHNOLOGY The quantity and quality of dissolved organic matter as supplementary carbon source impacts the pesticide-degrading activity of a triple-species bacterial biofilm Benjamin Horemans & Johanna Vandermaesen & Philip Breugelmans & Johan Hofkens & Erik Smolders & Dirk Springael Received: 5 February 2013 / Revised: 11 April 2013 / Accepted: 13 April 2013 / Published online: 8 May 2013 # Springer-Verlag Berlin Heidelberg 2013 Abstract Effects of environmental dissolved organic matter (eDOM) that consists of various low concentration carbonic compounds on pollutant biodegradation by bacteria are poorly understood, especially when it concerns synergistic xenobiotic-degrading consortia where degradation depends on interspecies metabolic interactions. This study examines the impact of the quality and quantity of eDOM, supplied as secondary C-source, on the structure, composition and pesticide-degrading activity of a triple-species bacterial con- sortium in which the members synergistically degrade the phenylurea herbicide linuron, when grown as biofilms. Biofilms developing on 10 mg L -1 linuron showed a steady-state linuron degradation efficiency of approximately 85 %. The three bacterial strains co-localized in the biofilms indicating syntrophic interactions. Subsequent feeding with eDOM or citrate in addition to linuron resulted into changes in linuron-degrading activity. A decrease in linuron- degrading activity was especially recorded in case of co-feeding with citrate and eDOM of high quality and was always associated with accumulation of the primary metabolite 3,4-dichloroaniline. Improvement of linuron degradation was especially observed with more recalci- trant eDOM. Addition of eDOM/citrate formulations altered biofilm architecture and species composition but without loss of any of the strains and of co- localization. Compositional shifts correlated with linuron degradation efficiencies. When the feed was restored to only linuron, the linuron-degrading activity rapidly changed to the level before the mixed-substrate feed. Meanwhile only minor changes in biofilm composition and structure were recorded, indicating that observed eDOM/citrate effects had been primarily due to repression/stimulation of linuron catabolic activity rather than to biofilm characteristics. Keywords Bacterial biofilms . Dissolved organic matter . Mixed substrate effects . Linuron biodegradation Introduction Biodegradation of pesticides often implicates the activity of bacterial consortia in which degradation occurs through metabolic synergistic interactions between the member or- ganisms. In such consortia, one member strain functions as the primary catalyst that degrades the parent compound into metabolites that are degraded by other members of the consortium. In turn, degradation of the metabolite(s) stimu- lates degradation of the parent compound (Murga et al. 1995; Dejonghe et al. 2003; Breugelmans et al. 2007). Synergistic metabolic interactions in consortia are often translated into the co-localization of the participating strains Electronic supplementary material The online version of this article (doi:10.1007/s00253-013-4928-4) contains supplementary material, which is available to authorized users. B. Horemans (*) : J. Vandermaesen : P. Breugelmans : E. Smolders : D. Springael Division of Soil and Water Management, Department of Earth and Environmental Sciences, Faculty of Bioscience Engineering, KU Leuven, Kasteelpark Arenberg 20–bus 2459, B-3001 Heverlee, Belgium e-mail: [email protected] J. Hofkens Molecular Imaging and Photonics, Faculty of Science, KU Leuven, Celestijnenlaan 200f, box 2404, B-3001 Heverlee, Belgium Appl Microbiol Biotechnol (2014) 98:931–943 DOI 10.1007/s00253-013-4928-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ENVIRONMENTAL BIOTECHNOLOGY
The quantity and quality of dissolved organic matteras supplementary carbon source impacts the pesticide-degradingactivity of a triple-species bacterial biofilm
Benjamin Horemans & Johanna Vandermaesen &
Philip Breugelmans & Johan Hofkens & Erik Smolders &
Dirk Springael
Received: 5 February 2013 /Revised: 11 April 2013 /Accepted: 13 April 2013 /Published online: 8 May 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Effects of environmental dissolved organic matter(eDOM) that consists of various low concentration carboniccompounds on pollutant biodegradation by bacteria arepoorly understood, especially when it concerns synergisticxenobiotic-degrading consortia where degradation dependson interspecies metabolic interactions. This study examinesthe impact of the quality and quantity of eDOM, supplied assecondary C-source, on the structure, composition andpesticide-degrading activity of a triple-species bacterial con-sortium in which the members synergistically degrade thephenylurea herbicide linuron, when grown as biofilms.Biofilms developing on 10 mg L−1 linuron showed asteady-state linuron degradation efficiency of approximately85 %. The three bacterial strains co-localized in the biofilmsindicating syntrophic interactions. Subsequent feeding witheDOM or citrate in addition to linuron resulted into changesin linuron-degrading activity. A decrease in linuron-degrading activity was especially recorded in case ofco-feeding with citrate and eDOM of high quality and
was always associated with accumulation of the primarymetabolite 3,4-dichloroaniline. Improvement of linurondegradation was especially observed with more recalci-trant eDOM. Addition of eDOM/citrate formulationsaltered biofilm architecture and species compositionbut without loss of any of the strains and of co-localization. Compositional shifts correlated with linurondegradation efficiencies. When the feed was restored toonly linuron, the linuron-degrading activity rapidlychanged to the level before the mixed-substrate feed.Meanwhile only minor changes in biofilm compositionand structure were recorded, indicating that observedeDOM/citrate effects had been primarily due torepression/stimulation of linuron catabolic activity ratherthan to biofilm characteristics.
Keywords Bacterial biofilms . Dissolved organic matter .
Mixed substrate effects . Linuron biodegradation
Introduction
Biodegradation of pesticides often implicates the activity ofbacterial consortia in which degradation occurs throughmetabolic synergistic interactions between the member or-ganisms. In such consortia, one member strain functions asthe primary catalyst that degrades the parent compound intometabolites that are degraded by other members of theconsortium. In turn, degradation of the metabolite(s) stimu-lates degradation of the parent compound (Murga et al.1995; Dejonghe et al. 2003; Breugelmans et al. 2007).Synergistic metabolic interactions in consortia are oftentranslated into the co-localization of the participating strains
Electronic supplementary material The online version of this article(doi:10.1007/s00253-013-4928-4) contains supplementary material,which is available to authorized users.
B. Horemans (*) : J. Vandermaesen : P. Breugelmans :E. Smolders :D. SpringaelDivision of Soil and Water Management, Department of Earth andEnvironmental Sciences, Faculty of Bioscience Engineering, KULeuven, Kasteelpark Arenberg 20–bus 2459,B-3001 Heverlee, Belgiume-mail: [email protected]
J. HofkensMolecular Imaging and Photonics, Faculty of Science, KULeuven, Celestijnenlaan 200f, box 2404,B-3001 Heverlee, Belgium
Appl Microbiol Biotechnol (2014) 98:931–943DOI 10.1007/s00253-013-4928-4

when grown in biofilms on solid surfaces. Biofilm growth isconsidered an important growth mode of microorganisms(Costerton et al. 1995; Hall-Stoodley et al. 2004) in naturalenvironments like soil and rivers but also in anthropogenicenvironments like wastewater treatment systems and hasbeen suggested to play an essential role in the in situ remov-al of organic contaminants (Singh et al. 2006). Co-localization and juxtapositioning of bacterial cells in biofilmsare suggested to optimize the synergistic metabolic interac-tions by minimizing metabolite diffusion distances betweenthe organisms (Nielsen et al. 2000; Breugelmans et al. 2008).
In the environment, organic xenobiotics and in particularpesticides do not form the only C-source for heterotrophicbacteria. In any environment, bacteria are exposed to othermore or less degradable C-sources present in dissolvedorganic matter (DOM). Environmental DOM (eDOM) con-sists of a broad range of organic compounds, all present atlow concentrations, and shows temporal and spatial varia-tions in quality and quantity, inherently linked to the eDOMsource (Volk et al. 1997). The quality of eDOM is related tothe extent that it can be biodegraded (Marschner and Kalbitz2003), and variability in eDOM biodegradation between 4and 93 % has been reported (Kalbitz et al. 2003a). eDOM ofhigh quality consists primarily of easily degradable organiccompounds and a low fraction of more slowly degradableand recalcitrant compounds (Kalbitz et al. 2003a). Thiseasily degradable fraction includes low molecular weightcompounds such as carbohydrates, carboxylic acids andamino acids (Thurman 1985) and is preferentially degraded(Volk et al. 1997; Amon et al. 2001). Low quality eDOMconsists primarily of more stable high molecular weightcompounds such as lignin-derived aromatic compounds in-cluding humic acids and fulvic acids (Kalbitz et al. 2003b).Dissolved organic carbon (DOC) concentrations, a measurefor eDOM quantity, in European surface water bodies rangefrom <0.5 to 72 mg L−1 DOC with a median value of5 mg L−1 DOC (Salminen 2005). In the USA, groundwatersamples contain <0.1 to 15 mg L−1 DOC with a medianvalue of 1.2 mg L−1 DOC (Leenheer et al. 1974). In agri-cultural soils, DOC concentrations range from 3 to32 mg L−1 (Brye et al. 2001; Siemens et al. 2003).
The availability of supplementary C-sources affects bio-degradation of organic pollutants in different ways. Studieson the effect of additional C-sources on pollutant biodegra-dation have been mainly performed using single easilydegradable C-sources as auxiliary carbon source. Additionof such pure, easily degradable C-sources either stimulatesor inhibits biodegradation. Stimulation is explained by ei-ther additional production of the degraders’ biomass (Toppet al. 1988) or by an increased uptake (Ortega-Calvo andSaiz-Jimenez 1998; Wick et al. 2003), while inhibition isoften explained by carbon catabolite repression (CCR;McFall et al. 1997; Rentz et al. 2004). Apparently, the
concentration of the alternative C-source and pollutant playsan important role. While in carbon-limited environments, asimultaneous use of carbon compounds was observed(Schmidt and Alexander 1985; Keuth and Rehm 1991), athigh concentration levels, a diauxic use of organic carboncompounds occurred. Keuth and Rehm (1991) for instanceobserved the repression of phenanthrene metabolism byArthrobacter polychromogenes in the presence of 4 g L−1
glucose, but not at 0.45 g L−1. In case of xenobiotic-degrading consortia, such effects can be expected to beamplified since biodegradation activity relies on synergisticinteractions between the different members, and a negativeeffect on a single member could affect the functioning of theentire consortium. For instance, citrate was shown to affectthe composition and structure as well as the degradingactivity of a linuron-degrading consortium when grown inbiofilms (Breugelmans et al. 2010).
In contrast to the effects of single easily degradablecarbon sources, the effect of eDOM on pollutant degrada-tion has been far less studied. As reported above, eDOMconsists of a multitude of structurally different compoundsall present at low concentrations and can differ significantlyin quality and quantity in time and space. Recently, weshowed that the presence of eDOM of different quality andadded at different quantities did not considerably alter thelinuron-degrading activity of a triple-species bacterial con-sortium in suspended batch cultures (Horemans et al.2013c). Neither did the addition of eDOM affect the degra-dation of atrazine by Chelatobacter heintzii SalB in similarbatch systems (Cheyns et al. 2012). Suspended batch sys-tems are however quite different from continuous biofilmsystems. In batch systems, the culture is homogeneous, andadded supplementary C-sources like eDOM that act as apotential repressor or inducer of pollutant degradation willdeplete over time. In contrast, continuous biofilm systemsare heterogeneous in structure and are continuously fed withfresh material. Moreover, supplementary C-sources mightalso affect attachment of biofilm members. Therefore,pollutant-degrading biofilms might respond differently onthe presence of eDOM regarding pollutant degradation com-pared to batch systems. In this study, we examined theresponse of the linuron-degrading triple-species bacterialconsortium mentioned above to the co-occurrence of differ-ent eDOM formulations when grown as a biofilm in contin-uously fed flow chamber systems. It was examined howeDOMquality and quantity affect biofilm structure/compositionand functionality. In the consortium, three bacteria metabolicallycooperate in the mineralization of the phenyl urea herbicidelinuron (Dejonghe et al. 2003). Variovorax sp. WDL1 initiateslinuron degradation through hydrolysis producing themetabolites 3,4-dichloroaniline (3,4-DCA) and N,O-dimethylhydroxylamine (N,O-DMHA). WDL1 uses 3,4-DCA inefficiently as source of carbon and energy and
932 Appl Microbiol Biotechnol (2014) 98:931–943

partially excretes 3,4-DCA that serves as growth sub-strate for a second strain Comamonas testosteroniWDL7. N,O-DMHA serves as C-source for the thirdmember strain Hyphomicrobium sulfonivorans WDL6.The triple-species biofilms were first cultivated onlinuron as sole source of carbon and energy and subse-quently challenged with linuron in the presence of dif-ferent eDOM formulations of variable quality and twodifferent quantities. After the mixed-substrate feed, con-ditions were reverted again to linuron as sole C-sourceto examine whether the linuron-degrading activity wasrestored to this recorded prior to the mixed-substratefeed. eDOM was applied at DOC concentrations of 5and 20 mg L−1. In addition to eDOM formulations,citrate, previously shown to disrupt the linuron-degrading activity of the triple consortium biofilm(Breugelmans et al. 2010), was used as a supplementarysingle C-substrate. The effect of eDOM being a sourceof nutrients different from carbon was excluded byapplying macronutrients and trace elements at non-limiting concentrations and maintaining a constant pH 7.
Materials and methods
Chemicals Pestanal® analytical standards (99.9 %) oflinuron (3-(3,4-dichlorophenyl)-1-methoxy-1-methylurea)and 3,4-dichloroaniline (Sigma-Aldrich®, Belgium) werepurchased.
Bacterial cultures and cultivation The linuron-degradingconsortium (Dejonghe et al. 2003) consists of Variovorax sp.WDL1 (LMG27260), C. testosteroni WDL7 (LMG27261)and H. sulfonivorans WDL6 (LMG27262). A Rfp taggedderivative of C. testosteroni WDL7, designated WDL7-Rfp,and a Yfp tagged derivative H. sulfonivorans WDL6, desig-nated WDL6-Yfp, were used (Breugelmans et al. 2008).Bacterial cultivation and media preparation were describedpreviously (Horemans et al. 2013a). Bacterial cells wereharvested in the late exponential phase, washed repeatedlywith 10 mMMgSO4 solution and cell densities of each strainwere adjusted to an OD660nm of 0.25 corresponding to ap-proximately 2.5×108 cells mL−1.
Preparation of eDOM and citrate formulations UsedeDOM formulations were DOM extracts originating fromalgae exudates, maize leaf leachate, soil incubated maizeleaves and from the rivers Urftallsperre (Simmerath,Germany) and Rourbron (Hoge Venen, Belgium) as wellas fulvic acids and humic acids purified from eDOM ex-tracts. Preparation of these eDOM formulations was de-scribed previously (Horemans et al. 2013b), and theircharacteristics regarding composition and microbial
biodegradability by the linuron-degrading consortium asreported in Horemans et al. (2013b) are summarized inTable 1. Citrate was prepared as a trisodiumcitrate solutiondissolved in Milli-Q® water as described (Horemans et al.2013b). eDOM and citrate preparations were filter sterilized(0.22 μm) and adjusted to 5 and 20 mg L−1 DOC to which10 mg L−1 linuron was added. Macronutrients and trace ele-ments were added equal to those applied in the minimalmediumMMO (Dejonghe et al. 2003) and furthermore adjust-ed to 1.59 mM NaHCO3, 1.11 mM NaCl, 0.54 mM Na2SO4
and 0.32 mM NaNO3 to obtain an equal ionic compositionamong all eDOM/citrate formulations.
Continuous flow chamber experiments Biofilms were culti-vated at 25 °C in continuously fed flow chamber systems(©BioCentrum DTU, Denmark) as described previously(Breugelmans et al. 2008). A 350-μL suspension consistingof a mixture of WDL1, WDL6-Yfp and WDL7-Rfp (ap-proximately 2.5×108 cells mL−1 each) was used as inocu-lum. The suspension was injected into the flow chamberusing a syringe (Myjector® U−100 Insulin needle, Terumo),and the chambers were left without feeding for 1 h to letcells settle on the glass substratum. After inoculation, flowchambers were fed for 30 days with adjusted MMOcontaining 10 mg L−1 linuron at 3.5 mL h−1. At day 30,two flow chambers were disconnected for confocal laserscanning analysis (CLSM). The remaining flow chamberswere subsequently fed for 10 days with adjusted MMOcontaining one of the seven eDOM formulations or citrateat 5 or 20 mg L−1 DOC supplemented with 10 mg L−1
linuron. Four replicate biofilms were operated for each ofthe eDOM tested as well as for citrate. In addition, a setupfed with adjusted MMO containing 10 mg L−1 linuron assole C-source for the entire duration of the experimentserved as biotic control. For each treatment, two of the fourreplicates were stopped at day 40 for CLSM analysis. Feedconditions in the remaining flow chambers were thenreverted by irrigating again with adjusted MMO containing10 mg L−1 linuron as sole C-source for 8 days. At day 48, allremaining flow chambers were disconnected for CLSManalysis. Effluent samples were taken regularly and acidi-fied with 0.33 % (v/v) of a 5-M HCl solution to stopmicrobial activity. Concentrations of linuron and 3,4-DCAin effluent samples were analysed with HPLC.
CLSM and image analysis An IX81 inverted microscope(Olympus, Japan) coupled with a Fluoview FV1000 confo-cal scanning unit (Olympus, Japan) was used for CLSManalysis. Before analysis, cells in the flow chamber werestained with Syto62 (Invitrogen™, Belgium) by injection ofthe stain through the tubing in the flow chamber and subse-quently incubated for 30 min. Emission signals were sequen-tially acquired through differential excitation and emission.
Appl Microbiol Biotechnol (2014) 98:931–943 933
Previous studies showed that unspecific DNA stains such asSyto 62 do not penetrate fluorescent protein-labeled bacterialcells (Wuertz et al. 2001) and also did not stain the Rfp andYfp-labeled bacterial strains used in this study (Breugelmanset al. 2007), enabling easy discrimination between the threecommunity members. Syto62-stained WDL1 cells were visu-alized by excitation with a 635-nm laser in combination with a655–755-nm band pass emission filter. WDL6-Yfp cells werevisualized by excitation with a 488-nm laser in combinationwith a 505–530-nm band pass emission filter. WDL7-Rfpcells were visualized by excitation with a 561-nm laser incombination with a 565–630-nm band pass emission filter.CLSM images of the biofilms were taken at three differentlocations at as well the front, the middle and the back sectionof the chambers. Images (512×512 pixel frame; 0.414 μmpixel size) were captured at 1-μm increments in height with anUPL SAPO x60/1.35 objective (Olympus, Japan). Biofilmswere visualized with Imaris 7.2 software (Bitplane,USA) from which biofilm structural variables (relativebiovolume, average thickness, surface coverage, rough-ness and diffusion distance) and biofilm composition(relative biovolume of each strain in the biofilm) werecalculated using COMSTAT 1 software as describedpreviously (Heydorn et al. 2000).
Analytical methods Linuron and 3,4-DCA concentrationswere determined with a LaChrom Classic (Hitachi, Japan)HPLC system equipped with an Alltima C18 (Alltech®,Grace, USA) reversed phase column (100 mm length×4.2 mm inner diameter; particle size 3 μm). Sample volumesof 20 μL were eluted with acetonitrile/water (65:35) at0.8 mL min−1. 3,4-DCA and linuron were detected by UVabsorption at 210 nm, and concentrations were determinedusing the HSM manager (Hitachi, Japan). Limit of detection
and limit of quantification were respectively 2.4 and4.1 μg L−1 for linuron and 4 and 7 μg L−1 for 3,4-DCA.The absence of interference from eDOM with the detectionof linuron and 3,4-DCA during HPLC analysis was deter-mined prior to conducting the experiments by comparingsamples at a concentration range between 100 μg L−1 and10 mg L−1 with and without an eDOM formulation (data notshown). Also, influent linuron concentrations with and with-out DOM were regularly measured and did not show differ-ences. Citrate concentrations were measured by means ofion exclusion chromatography performed with a DionexICE AC-6 column and a AMMS ICE-II suppressor.
Data analysis Statistical multivariate analysis wasperformed using SOLO (Eigenvector Research Inc., USA).Partial least squares regression (PLSR) regression was ap-plied to correlate eDOM biodegradability with the structural(biomass, thickness, roughness and diffusion distance) andcompositional biofilm variables (relative WDL1, WDL6-Yfp and WDL7-Rfp abundance) and to correlate these lattervariables in turn with the residual linuron concentrations inthe effluent. Structural and compositional biofilm variableswere determined for biofilms analysed (1) after feeding withthe mixed linuron-eDOM/citrate feed (at day 40) and (2)after reverting feed conditions to linuron as sole C-source (atday 48). Parameters determined from images taken at thefront, middle and back section of the flow chambers were allconsidered as independent variables. The biodegradabledissolved organic carbon fraction (BDOC) was used as ameasure for eDOM biodegradability as previously deter-mined for the consortium consisting of WDL1, WDL6-Yfp and WDL7-Rfp (Table 1; Horemans et al. 2013b). Atcertain time points, the actual residual linuron concentrationin the effluent was calculated as a percentage of the applied
Table 1 Initial characteristics of DOM and BDOC values (percentage of initial DOC)
DOM sample SUVA254a αNH2
b Carbohydratesb Peptidesb BDOC (%)a
L g−1 TOC cm−1 μM compound mM C−1
Citrate 5.1±0.0 0.0±1.0 1.2±0.9 0±16 –
Algae exudates 9.0±0.9 23.3±5.2 2.6±1.4 40±14 44±2
Maize leaf leachate 15.4±0.3 24.3±2.9 43.8±2.5 68±10 54±4
Humic acids 25.5±0.3 2.6±1.3 9.1±1.3 82±22 39±2
Soil incubated leaf extracts 30.8±0.6 10.0±1.9 5.0±1.1 32±17 26±3
Fulvic acids 37.0±0.8 0.0±2.6 8.6±1.9 62±16 18±1
River DOM Urftallsperre 37.4±0.1 8.4±2.9 9.9±0.7 58±28 23±4
River DOM Rourbron 46.5±0.0 2.1±1.0 14.3±1.3 98±32 23±1
BDOC values were determined in a suspended batch culture in the presence of the consortium comprising WDL1, WDL6 and WDL7 after anincubation period of 14 days at 25 °C (Horemans et al. 2013b). The DOM samples are ordered according to increasing aromaticity (SUVA254).aMeasured values with 95 % confidence intervals (n=3)bMeasured values with 95 % confidence intervals (n=12)
934 Appl Microbiol Biotechnol (2014) 98:931–943

influent concentration, and 3,4-DCA accumulation was cal-culated as the percentage of 6.6 mg L−1 3,4-DCA, theequivalent of 10 mg L−1 linuron. In addition, linuron deg-radation curves for each biofilm setup were generated show-ing linuron effluent concentrations in function of time. Theintegrated area [(linuron)×time] under the linuron degrada-tion curves was calculated for each setup between day 30and day 40. The ratio of the integrated areas (area percent-age) of a DOM/citrate fed setup over that of the controlsetup without DOM/citrate was used as a measure of thetotal residual linuron in the effluent between day 30 andday 40 during the mixed-substrate feed of DOM andlinuron, a value that we designated as total residual linuron(TRL). PLSR was performed on the biofilm structural andcompositional variables determined at day 40 and day 48 asthe independent variables and BDOC (percentage) and theTRL (area percentage) as dependent variables. Prior toPLSR, all variables were normalized over their mean valueand the standard deviation to avoid scaling effects. Sampleswere designated as an outlier when the Hotelling T2 valueand Q residual value exceeded the 95 % confidence bound-aries and were eliminated from analysis. Unpaired Student’st test (0.05 confidence level; n=3) was used to determinesignificant differences in biofilm properties and linurondegradation amongst different biofilm setups.
Results
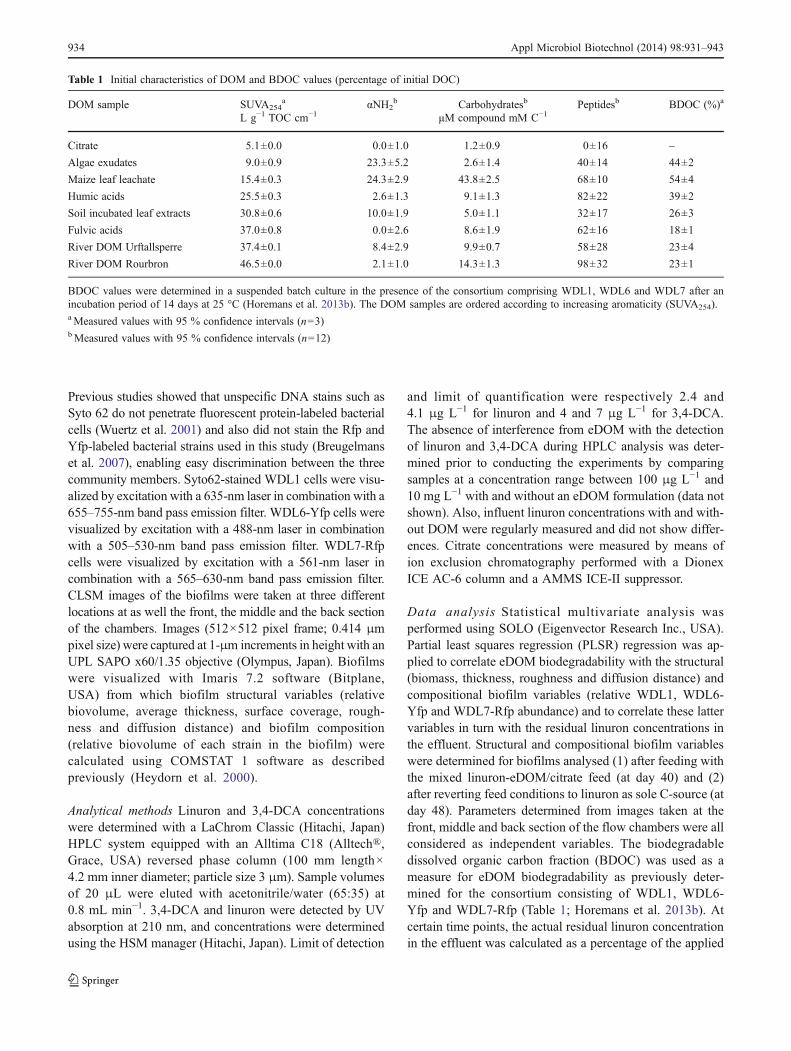
Linuron degradation in mixed-substrate conditions Flowchambers were first fed with 10 mg L−1 linuron as solecarbon and energy source. Linuron degradation was signif-icant after a lag phase of 10 days after which the residuallinuron concentration in the effluent progressively decreasedwith approximately 1 mg L−1 linuron per day till a steady-state effluent concentration of 1.1±0.3 mg L−1 linuron cor-responding to a linuron degradation efficiency (linuron con-centration in the effluent compared to this in the influent) of89 % was reached from day 20 onwards (Fig. 1a). 3,4-DCAdid not accumulate in the effluent. At day 30, the linuronfeed was replaced by a mixed-substrate feed containing10 mg L−1 linuron and eDOM/citrate at a concentration ofeither 5 or 20 mg L−1 DOC. Meanwhile, as a control, someflow chambers remained fed with 10 mg L−1 linuron withoutsupplementary C-source. At day 40, after 10 days of sup-plying the mixed-substrate, the linuron effluent concentra-tion in the control setup remained around 0.95 ±0.05 mg L−1. In contrast, clear changes in linuron degrada-tion were observed in the mixed-substrate feed setups. Theresponse of biofilms regarding their linuron degrading ac-tivity to eDOM addition is illustrated by means of the TRLin the effluent between day 30 and day 40 for all setups inFig. 1b and by means of the linuron degradation efficiency
(effluent concentration compared to influent concentration)for selected setups in Fig. 1a. The addition of citrate resultedin a decrease in linuron degradation efficiency (Fig. 1a) andan increase in TRL (Fig. 1b) concomitant with a significantaccumulation of 3,4-DCA (Fig. 1a). At 5 and 20 mg L−1
DOC, maximal linuron degrading efficiency was respective-ly 83±2 and 51±1 % and 3,4-DCA accumulated maximallyat 11±2 and 49±1 %. TRL values were 376±28 and 620±40 %, respectively. Citrate was degraded for maximally 100and 31±9 % as compared to the added influent concentra-tion of 5 and 20 mg L−1 DOC, respectively. Inhibition oflinuron degradation was also observed in the setup fed withmaize leaf leachate, but only when a feed concentration of20 mg L−1 DOC was applied (Fig. 1a, b). In this setup, amaximal linuron-degrading efficiency of 80±9 % was reg-istered and a TRL of 186±53 %. Concomitant with thedecrease in linuron degradation, 3,4-DCA accumulated tilla maximum of 23±9 % (Fig. 1a). However, at 5 mg L−1
DOC, maize leaf leachate stimulated linuron degradationsince more linuron was removed compared to the controlsetup and 3,4-DCA was not accumulated (Fig. 1a, b).Stimulation of linuron degradation was also observed withthe other remaining eDOM formulations when supplied at5 mg L−1 DOC and for some eDOM at 20 mg L−1 DOC andfollowed a similar evolution in linuron degradation efficien-cy as shown in Fig. 1a for maize leaf leachate at 5 mg L−1
DOC. The degree of stimulation depended on the eDOMformulation, and no detectable accumulation of 3,4-DCAoccurred. No significant effects were observed on linurondegradation for biofilms fed with 20 mg L−1 DOC of theriver eDOM from Urftallsperre and algae exudates (data notshown).
After the mixed feed of linuron and eDOM (day 40),conditions were reverted to the feed with linuron as sole C-source. In all setups, effluent linuron concentrations rapidlyrestored to 1.0±0.1 mg L−1 linuron, i.e. identical to theeffluent concentrations prior to the mixed substrate feedand to those in the control setup (Fig. 1a). Especially setupsin which eDOM inhibited linuron degradation, such as thesetups fed with citrate and maize leaf leachate, showedinstant linuron degradation. In all other setups, this restora-tion was rather slow.
Biofilm development before and after mixed-substrateconditions CLSM images of biofilms grown in the controlsetup fed with linuron as sole C-source are shown in Fig. 2.Replicates of each setup showed equal biofilm morphol-ogies and compositions at all examined locations in the flowchamber. As previously observed by Breugelmans et al.(2008) triple-species biofilms grown on linuron as sole C-source with inorganic N were dominated by WDL6-Yfp thatformed large mounds surrounded by structures ofintermingled WDL1 and WDL7-Rfp cells as a reflection
Appl Microbiol Biotechnol (2014) 98:931–943 935
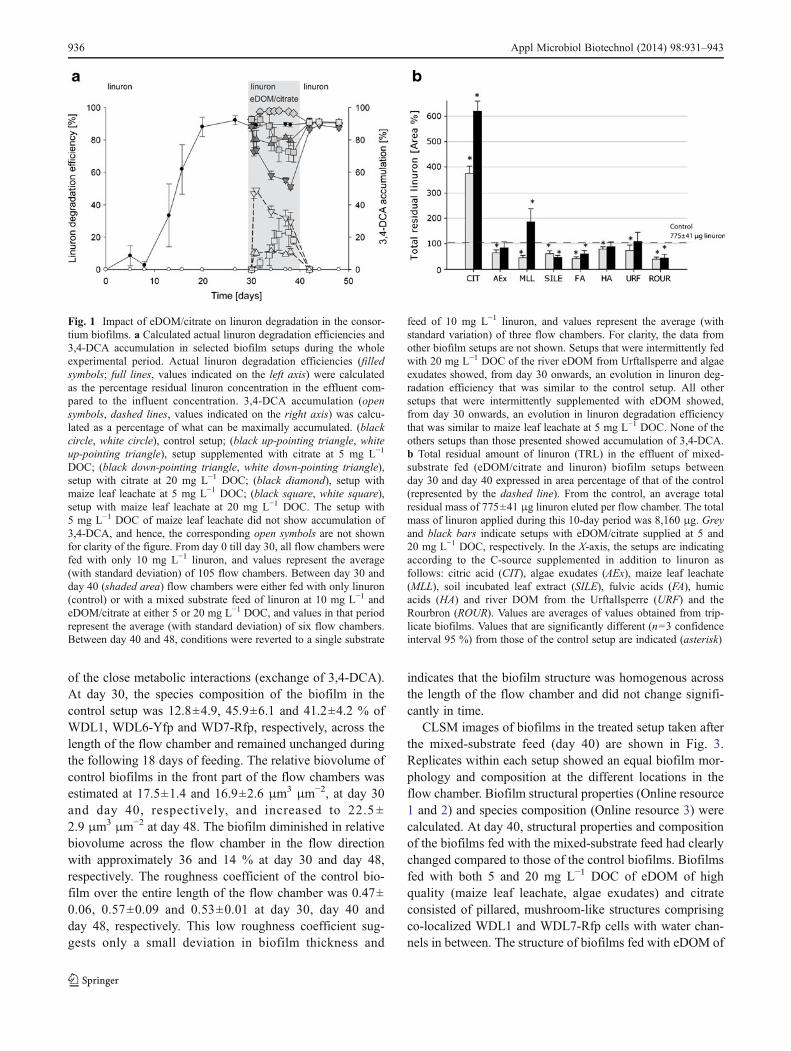
of the close metabolic interactions (exchange of 3,4-DCA).At day 30, the species composition of the biofilm in thecontrol setup was 12.8±4.9, 45.9±6.1 and 41.2±4.2 % ofWDL1, WDL6-Yfp and WD7-Rfp, respectively, across thelength of the flow chamber and remained unchanged duringthe following 18 days of feeding. The relative biovolume ofcontrol biofilms in the front part of the flow chambers wasestimated at 17.5±1.4 and 16.9±2.6 μm3 μm−2, at day 30and day 40, respectively, and increased to 22.5 ±2.9 μm3 μm−2 at day 48. The biofilm diminished in relativebiovolume across the flow chamber in the flow directionwith approximately 36 and 14 % at day 30 and day 48,respectively. The roughness coefficient of the control bio-film over the entire length of the flow chamber was 0.47±0.06, 0.57±0.09 and 0.53±0.01 at day 30, day 40 andday 48, respectively. This low roughness coefficient sug-gests only a small deviation in biofilm thickness and
indicates that the biofilm structure was homogenous acrossthe length of the flow chamber and did not change signifi-cantly in time.
CLSM images of biofilms in the treated setup taken afterthe mixed-substrate feed (day 40) are shown in Fig. 3.Replicates within each setup showed an equal biofilm mor-phology and composition at the different locations in theflow chamber. Biofilm structural properties (Online resource1 and 2) and species composition (Online resource 3) werecalculated. At day 40, structural properties and compositionof the biofilms fed with the mixed-substrate feed had clearlychanged compared to those of the control biofilms. Biofilmsfed with both 5 and 20 mg L−1 DOC of eDOM of highquality (maize leaf leachate, algae exudates) and citrateconsisted of pillared, mushroom-like structures comprisingco-localized WDL1 and WDL7-Rfp cells with water chan-nels in between. The structure of biofilms fed with eDOM of
Fig. 1 Impact of eDOM/citrate on linuron degradation in the consor-tium biofilms. a Calculated actual linuron degradation efficiencies and3,4-DCA accumulation in selected biofilm setups during the wholeexperimental period. Actual linuron degradation efficiencies (filledsymbols; full lines, values indicated on the left axis) were calculatedas the percentage residual linuron concentration in the effluent com-pared to the influent concentration. 3,4-DCA accumulation (opensymbols, dashed lines, values indicated on the right axis) was calcu-lated as a percentage of what can be maximally accumulated. (blackcircle, white circle), control setup; (black up-pointing triangle, whiteup-pointing triangle), setup supplemented with citrate at 5 mg L−1
DOC; (black down-pointing triangle, white down-pointing triangle),setup with citrate at 20 mg L−1 DOC; (black diamond), setup withmaize leaf leachate at 5 mg L−1 DOC; (black square, white square),setup with maize leaf leachate at 20 mg L−1 DOC. The setup with5 mg L−1 DOC of maize leaf leachate did not show accumulation of3,4-DCA, and hence, the corresponding open symbols are not shownfor clarity of the figure. From day 0 till day 30, all flow chambers werefed with only 10 mg L−1 linuron, and values represent the average(with standard deviation) of 105 flow chambers. Between day 30 andday 40 (shaded area) flow chambers were either fed with only linuron(control) or with a mixed substrate feed of linuron at 10 mg L−1 andeDOM/citrate at either 5 or 20 mg L−1 DOC, and values in that periodrepresent the average (with standard deviation) of six flow chambers.Between day 40 and 48, conditions were reverted to a single substrate
feed of 10 mg L−1 linuron, and values represent the average (withstandard variation) of three flow chambers. For clarity, the data fromother biofilm setups are not shown. Setups that were intermittently fedwith 20 mg L−1 DOC of the river eDOM from Urftallsperre and algaeexudates showed, from day 30 onwards, an evolution in linuron deg-radation efficiency that was similar to the control setup. All othersetups that were intermittently supplemented with eDOM showed,from day 30 onwards, an evolution in linuron degradation efficiencythat was similar to maize leaf leachate at 5 mg L−1 DOC. None of theothers setups than those presented showed accumulation of 3,4-DCA.b Total residual amount of linuron (TRL) in the effluent of mixed-substrate fed (eDOM/citrate and linuron) biofilm setups betweenday 30 and day 40 expressed in area percentage of that of the control(represented by the dashed line). From the control, an average totalresidual mass of 775±41 μg linuron eluted per flow chamber. The totalmass of linuron applied during this 10-day period was 8,160 μg. Greyand black bars indicate setups with eDOM/citrate supplied at 5 and20 mg L−1 DOC, respectively. In the X-axis, the setups are indicatingaccording to the C-source supplemented in addition to linuron asfollows: citric acid (CIT), algae exudates (AEx), maize leaf leachate(MLL), soil incubated leaf extract (SILE), fulvic acids (FA), humicacids (HA) and river DOM from the Urftallsperre (URF) and theRourbron (ROUR). Values are averages of values obtained from trip-licate biofilms. Values that are significantly different (n=3 confidenceinterval 95 %) from those of the control setup are indicated (asterisk)
936 Appl Microbiol Biotechnol (2014) 98:931–943

lower quality [fulvic acids, soil incubated leaf extract, rivereDOM (Rourbron)] resembled that of the biofilms of thecontrol setup. However, the biofilm composition differedsignificantly from this in the control setup for all treatedsetups and depended on the applied DOM and its quantity.Whereas WDL6-Yfp comprised about 45.9±6.1 % of thebiofilm in the control setup, its abundance decreased signif-icantly in setups fed with a mixed-substrate feed. Especiallyfor the feed containing either algae exudates, humic acids,river eDOM (Urftallsperre and Rourbron) at 5 mg L−1 DOC,WDL6-Yfp cells could not be detected anymore. In case ofsetups fed with maize leaf leachate, soil incubated leafextract and fulvic acids at 5 mg L−1 DOC, WDL6-Yfp wasstill present, but had decreased in relative abundance (20–35 %) compared to the control biofilm (40–45 %). As in thecontrol setup, WDL6-Yfp formed large mounds in the soilincubated leaf extract and maize leaf leachate fed biofilms.A decrease in WDL6-Yfp abundance to approximately 10 %or less was also observed when DOM was applied at20 mg L−1 DOC (Online resource 3). In contrast toWDL6-Yfp, the abundance of WDL1 increased significant-ly after the mixed-substrate feed for almost all treatments atboth 5 and 20 mg L−1 DOC from 25 % to approximately40 % for the slowly degradable eDOM up to 70 and 80 %for the easily degradable eDOM, respectively. This increasein WDL1 was at the expense of both WDL6-Yfp andWDL7-Rfp. The exception was in setups fed with humicacids, where WDL7-Rfp became the dominant strain (approx-imately 90 %; Online resource 3). No significant differencesbetween mixed-substrate fed biofilms and control biofilmswere recorded regarding biovolumes (Online resource 2).
CLSM images of biofilms in the treated setup taken afterrenewal of the single substrate linuron feed (day 48) areshown in Online resource 4. Replicates of each setup
showed an equal biofilm morphology and composition atthe different locations in the flow chamber. Biofilm struc-tural properties (Online resource 1 and 2) and species com-position (Online resource 3) were calculated. Structuralproperties and composition of the biofilms (day 48)remained significantly different among the biofilms. Totalbiovolumes did not change and differences in structuralproperties in biofilms previously fed on a mixed feed oflinuron and eDOM/citrate were difficult to distinguish be-fore (day 40) and after (day 48) the last linuron feed (Onlineresource 1 and 2). Between day 40 and day 48, only subtlechanges occurred in both species composition and structureof the biofilms previously fed with a mixed feed of citrate,algae exudates and maize leaf leachate. On the other hand,biofilms previously fed with a mixture of linuron and moreslowly degradable eDOM, i.e. soil incubated leaf extracts,fulvic acids, river DOM from both Urftallsperre andRourbron, showed a shift in species composition toward ahigher abundance of WDL1 from 40 % to approximately 70–80 % at the expense of both WDL6-Yfp and WDL7-Rfp.
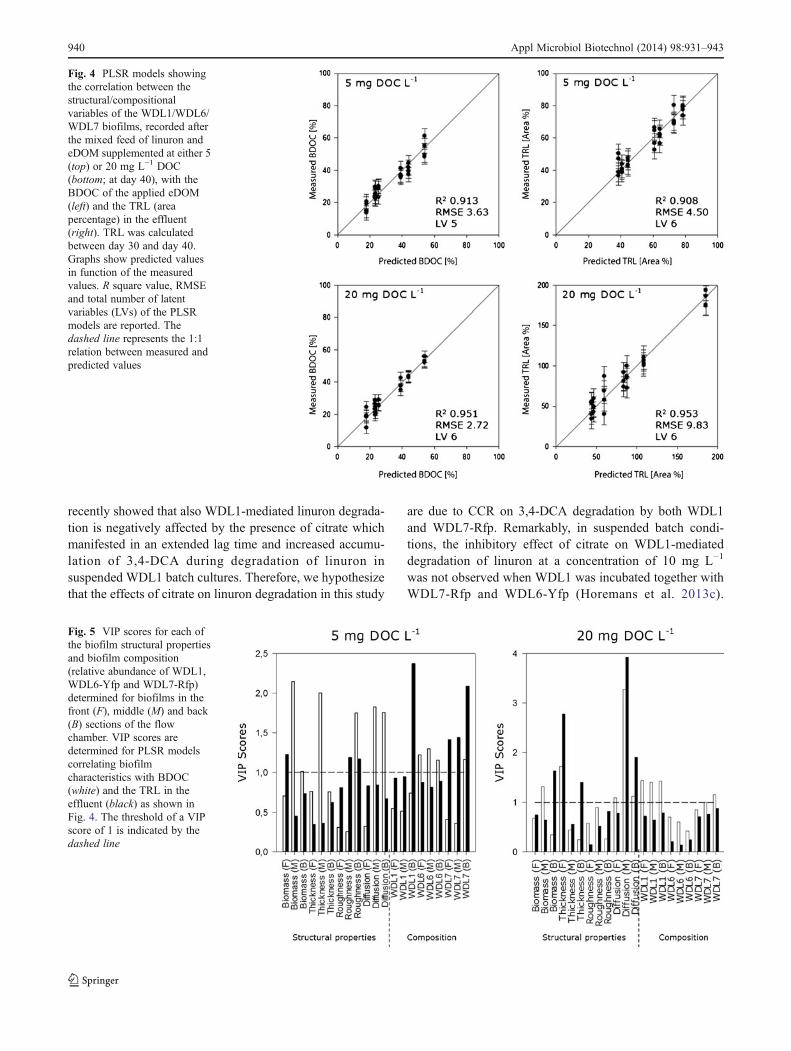
Correlation of biofilm properties with eDOM biodegrad-ability and linuron degradation PLSR describes the vari-ance in biofilm properties determined after the mixed-substrate feed in relation to the co-variance with eDOMbiodegradability (BDOC) and the TRL as recorded duringthe mixed-substrate feed of 10 days. Preceding multivariateanalysis designated the citrate fed biofilms as outliers com-pared to the eDOM fed biofilms. Therefore, the dataobtained with citrate were omitted. PLSR models showingthe predicted BDOC values and the TRL in function of themeasured values are depicted in Fig. 4. A PLSR model thatperfectly predicts the dependent variable is represented by acurve showing a 1:1 relation in the graph. Biofilm properties
Fig. 2 Representative CLSMimages of WDL1/WDL6-Yfp/WDL7-Rfp biofilms at the frontsection of the flow chamber incontrol setups fed with10 mg L−1 linuron as recordedon day 30 (a), day 40 (b) andday 48 (c). Images are topviews of 3D representations ofCLSM micrographs withshadow projection (a1, b1, c1)and cross-sectional views in thex, y and z-plane (a2, b2, c2).WDL1 (blue), WDL7-Rfp (red)and WDL6-Yfp (green)
Appl Microbiol Biotechnol (2014) 98:931–943 937

after the mixed-substrate feed are highly correlated bothwith the BDOC and the TRL. This accounts for the feedcontaining eDOM at as well 5 as 20 mg L−1 DOC.Apparently, the biofilm morphology and compositionchanged significantly depending on the biodegradability ofthe applied eDOM. Biofilm morphology and compositioncorrelated in turn with the linuron-degrading activity asrepresented by TRL. By using the Variable Importance inProjection (VIP) scores (Fig. 5) for the biofilm propertiesthat were used in the PLSR models, we could determinewhether these properties weighed significantly on the PLSR
model. A VIP score above threshold 1 indicates that thevariable weighs significantly on the model. For setups ex-posed to a mixed-substrate feed with eDOM at 5 mg L−1
DOC, structural properties weighed significantly on themodel that links biofilm properties with the BDOC ofeDOM. In contrast, for the PLSR model that links biofilmproperties with the TRL, the species composition in theentire flow chamber weighed significantly. In case of amixed feed with 20 mg L−1 DOC, several structural biofilmproperties in the middle section of the flow chamber andWDL1 abundance in the entire flow chamber weighed more
Fig. 3 Representative CLSM images of WDL1/WDL6-Yfp/WDL7-Rfpbiofilms at the front section of the flow chamber at day 40 after feeding with10 mg L−1 linuron for 30 days followed by feeding for 10 days with themixed-substrate feed containing 10mgL−1 linuron and the indicated eDOM/citrate either at 5 mg L−1 DOC or 20 mg L−1 DOC. The setups are indicatedaccording to the used supplementary C-source in addition to linuron as
follows: citric acid (CIT), algae exudates (AEx), maize leaf leachate (MLL),soil incubated leaf extract (SILE), fulvic acids (FA), humic acids (HA) andriver DOM from the Urftallsperre (URF) and the Rourbron (ROUR). Imagesare top views of 3D representations of CLSM micrographs with shadowprojection (top) and cross-sectional views in the x, y and z-plane (bottom).WDL1 (blue), WDL7-Rfp (red) and WDL6-Yfp (green)
938 Appl Microbiol Biotechnol (2014) 98:931–943

strongly on the model that describes BDOC. VIP scoresrevealed that only a few structural properties of the biofilmsweighed significantly for the PLSRmodel that describes TRL.Those properties were the thickness of the biofilm in the frontof the flow chamber and the diffusion distance of the biofilmin the middle and back section of the flow chamber.
Discussion
The application of eDOM in addition to linuron to a maturelinuron-degrading triple-species biofilm clearly changed itslinuron-degrading activity. Negative effects of the supple-mentary C-source added were especially observed witheDOM of high quality, i.e. high biodegradability and withcitrate. Negative effects of citrate and maize leaf leachate
were accompanied by 3,4-DCA accumulation. The negativeeffect of citrate on linuron degradation and accumulation of3,4-DCA by the consortium grown as biofilms has beenpreviously reported (Breugelmans et al. 2010) under slightlydifferent conditions, i.e. using a linuron concentration of25 mg L−1 and a citrate concentration of 7.2 mg L−1 DOC.The effect of citrate on linuron degradation though washighly similar showing approximately 50 % of linuron deg-radation and 10 % of 3,4-DCA accumulation. The authorshypothesized that citrate exerted CCR on the degradation ofthe metabolite 3,4-DCA by WDL7-Rfp and disturbed thecooperation between WDL1 and WDL7-Rfp which is need-ed for efficient linuron degradation. The removal of 3,4-DCA has indeed been found to be important for efficientlinuron and 3,4-DCA degradation by WDL1 (Dejonghe etal. 2003; Breugelmans et al. 2007). On the other hand, we
Fig. 3 (continued)
Appl Microbiol Biotechnol (2014) 98:931–943 939
recently showed that also WDL1-mediated linuron degrada-tion is negatively affected by the presence of citrate whichmanifested in an extended lag time and increased accumu-lation of 3,4-DCA during degradation of linuron insuspended WDL1 batch cultures. Therefore, we hypothesizethat the effects of citrate on linuron degradation in this study
are due to CCR on 3,4-DCA degradation by both WDL1and WDL7-Rfp. Remarkably, in suspended batch condi-tions, the inhibitory effect of citrate on WDL1-mediateddegradation of linuron at a concentration of 10 mg L−1
was not observed when WDL1 was incubated together withWDL7-Rfp and WDL6-Yfp (Horemans et al. 2013c).
Fig. 4 PLSR models showingthe correlation between thestructural/compositionalvariables of the WDL1/WDL6/WDL7 biofilms, recorded afterthe mixed feed of linuron andeDOM supplemented at either 5(top) or 20 mg L−1 DOC(bottom; at day 40), with theBDOC of the applied eDOM(left) and the TRL (areapercentage) in the effluent(right). TRL was calculatedbetween day 30 and day 40.Graphs show predicted valuesin function of the measuredvalues. R square value, RMSEand total number of latentvariables (LVs) of the PLSRmodels are reported. Thedashed line represents the 1:1relation between measured andpredicted values
Fig. 5 VIP scores for each ofthe biofilm structural propertiesand biofilm composition(relative abundance of WDL1,WDL6-Yfp and WDL7-Rfp)determined for biofilms in thefront (F), middle (M) and back(B) sections of the flowchamber. VIP scores aredetermined for PLSR modelscorrelating biofilmcharacteristics with BDOC(white) and the TRL in theeffluent (black) as shown inFig. 4. The threshold of a VIPscore of 1 is indicated by thedashed line
940 Appl Microbiol Biotechnol (2014) 98:931–943

Apparently, the system behaves differently between batchand biofilm systems. This can be explained by the inherentdifferences between a continuously fed biofilm setup and abatch setup. In batch conditions, WDL7 removes citrate as arepressor, and inhibition on linuron degradation might beminimal. In a continuously fed biofilm setup, however, therepressor citrate is continuously resupplied, and the repres-sion is maintained. Indeed, in the setup fed with a citrateconcentration at 20 mg L−1 DOC, where the greatest effectswere observed, approximately 40 % of the citrate was notremoved during passage through the flow chamber andhence could exert CCR. CCR of xenobiotic degradationpathways has been reported with various single compoundsubstrates including TCA cycle intermediates, carbohy-drates and amino acids (Müller et al. 1996; McFall et al.1997; Petruschka et al. 2001; Rentz et al. 2004).
In addition to citrate, the only eDOM that affectedlinuron degradation negatively was maize leaf leachate butonly at a concentration of 20 mg L−1 DOC. Maize leafleachate contains a large fraction of carbohydrates(Table 1) suggesting that high concentrations of especiallycarbohydrates and organic acids affect linuron degradation.This effect of maize leaf leachate was not observed insuspended batch systems (Horemans et al. 2013c) empha-sizing again the difference between batch and biofilm sys-tems. Also with maize leaf leachate, 3,4-DCA wasaccumulating in the effluent suggesting that the mechanismof inhibition of linuron degradation is similar to this ob-served with citrate, i.e. directly through CCR of 3,4-DCAdegradation. A similar effect of maize leaf leachate onlinuron degradation by WDL1 was previously observed insuspended batch cultures (Horemans et al. 2013c).Previously, phenanthrene degradation by Pseudomonasputida ATCC17484 was inhibited in the presence of plantroot extracts and exudates containing organic acids, aminoacids and carbohydrates (Rentz et al. 2004). Easily degrad-able eDOM did however not always affect linuron degrada-tion negatively. Maize leaf leachate applied at 5 mg L−1 DOCaffected linuron degradation positively, and no 3,4-DCA accu-mulation was observed. This result shows the concentration-dependent character of the effect the presence of eDOM has(Horemans et al. 2013c; Keuth and Rehm 1991). Similarly,algae exudates that, as maize leaf leachate, mainly consist ofrelatively easily degradable C-compounds, stimulated linurondegradation at 5 mg L−1 DOC but did not affect linurondegradation when supplied at 20 mg L−1 DOC. Algae exudatescontain mainly free amino acids instead of carbohydrates(Table 1). Amino acids were previously shown to stimulatebiodegradation of the phenyl urea herbicide isoproturon as itprovided the essential co-factors for an auxotrophic strain(Sørensen et al. 2002) but was also reported to exert CCR onphenanthrene degradation (Rentz et al. 2004). We hypothesizethat due to the subtle balance of simultaneously occurring
negative and positive effects, no effect was observed in caseof algae exudates at 20 mg L−1 DOC.
Low-quality eDOM rather stimulated linuron degradationduring the mixed-substrate feeds, except in case of humicacids and river eDOM (Urftallsperre) at 20 mg L−1 DOCwhere no difference was observed compared to the controlbiofilm. For eDOM exhibiting positive effects, no signifi-cant differences in stimulation were observed betweeneDOM applied at either 5 or 20 mg L−1 DOC, indicatingthat the effects on linuron degradation of more recalcitranteDOM were irrespective of the applied eDOM quantity.Supplementary C-sources can stimulate concomitant organ-ic xenobiotic degradation in different ways. The availabilityof supplementary organic compounds can enhance pollutantdegradation by providing additional carbon and energy forgrowth (Schmidt and Alexander 1985; Topp et al. 1988;Hess et al. 1990; Wick et al. 2003) resulting into an in-creased biomass of degraders and hence improved degrada-tion. The members of the consortium and especially WDL1and WDL7 can use a substantial part of the eDOM formu-lations applied in this study including eDOM of low qualityof which they probably use the more degradable fraction(Horemans et al. 2013b). However, in biofilms, stimulationwas recorded without substantial changes in total biomass.Alternatively, the eDOM provided organic co-factors thatimprove cell metabolism (Sørensen et al. 2002). Humicacids have also been reported to enhance pollutant degrada-tion by improving the bioavailability of the pollutants, butthis process probably only plays a role in case of organicpollutants with a higher octanol-water partitioning coeffi-cient than linuron (Ortega-Calvo and Saiz-Jimenez 1998;Smith et al. 2011). Improved removal by sorption to humicacids that accumulate in the biofilm is unlikely since thiseffect would also have been observed in the setup fed with20 mg L−1 DOC. A possible explanation is that an increasedexpression of the linuron catabolic pathway occurred due toinduction by specific (aromatic) compounds present ineDOM. Similar observations were made for the mineraliza-tion of chlorinated phenols in humic acid containing lakewater (Larsson et al. 1988). Also in batch conditions, theaddition of humic acids improved linuron degradation byVariovorax sp. WDL1 (Horemans et al. 2013c).
Changes in linuron degradation activity can also be dueto changes in biofilm structure and composition. Therefore,we looked for correlations between biofilm properties andTRL values. From PLSR, it was deduced that the biode-gradability of the intermittently applied supplementaryeDOM as well as the linuron-degrading activity (based onTRL) during the mixed feed correlated with biofilm archi-tecture and species composition. Further analysis of thePLSR models revealed which biofilm properties weighedsignificantly on the models. At 5 mg L−1 DOC, TRL largelycorrelated with species composition. When biofilms were
Appl Microbiol Biotechnol (2014) 98:931–943 941

fed with a mixed feed of linuron and eDOM at 20 mg L−1
DOC, TRL correlated most with both the species composi-tion and some structural properties. As such, eDOM quan-tity appears to add up to the effect that a certain eDOMformulation has on biofilm development and its activity.Despite the observed correlation between biofilm propertiesand TRL, it remains unclear if eDOM formulations alteredbiofilm properties and whether this in turn directly alteredthe linuron degradation activity during the mixed feed. Thefact that linuron degradation activity restored immediatelyafter reverting the feed to only linuron, while biofilm mor-phology and species composition remained largelyunchanged, indicates that other factors were rather dominantin determining the effect of eDOM on linuron degradationactivity during the mixed feed. As discussed above, thisfactor is likely the effect eDOM/citrate has on the singlecell activity of each consortium member by means of CCRon 3,4-DCA degradation in case of negative effects orstimulation in case of positive effects and the additional cellproliferation of certain consortium members on additionalC-sources. We therefore suggest that the changes in biofilmproperties were as such rather a result of the alterations incell activity then that they directly affected linuron degrada-tion. Interestingly, in all setups, both WDL1 and WDL7-Rfpremained an important fraction of the consortium in thebiofilm, and in many cases, this was also the case forWDL6-Yfp. Moreover, the co-localization of WDL1 andWDL7-Rfp, considered to be required for efficient linurondegradation, was always conserved. We attribute this toearlier observed synergism between consortium membersin the degradation of eDOM and of other single carbonsubstrates different from linuron (Horemans et al. 2013a,b). Apparently, this multiple substrate synergism ensures acertain integrity of the consortium in biofilms in caselinuron is not the only C-source. The disappearance ofWDL6-Yfp from the biofilms had no detrimental effect onthe linuron-degrading activity, suggesting that WDL6-Yfphas in the first place a self-beneficiary commensal relationwith the other strains in the biofilm.
We conclude that effect of supplementary C-sources inaddition to linuron on the linuron-degrading activity of theconsortium is different in continuously fed biofilms com-pared to suspended batch cultures. Moreover, the linuron-degrading activity of the studied linuron-degrading consor-tium appears to remain rather stable and robust under con-ditions of co-feeding with eDOM in addition to linuron.eDOM nevertheless can affect linuron degradation activityby the consortium. eDOM of high quality seems to affectlinuron degradation negatively especially at higher DOCconcentrations probably due to the presence of CCR in-voked by carbon sources that are present in the eDOM.eDOM of low quality however seems to stimulate degrada-tion by hitherto unknown mechanisms. The robustness of
the consortiums activity seems to be at least partially due tosynergism in the utilization of C compounds present in theeDOM that ensures integrity in composition and co-localization. Currently, we are examining how the presenceof eDOM in addition to linuron affects linuron degradationin the case linuron is present at micropollutant concentra-tions (micrograms per liter range) as often observed ingroundwater and surface water.
Acknowledgments This research was funded by IWT (Innovation byScience and Technology) project SB/73381, OT10/03 project of KULeuven, the Inter-University Attraction Pole (IUAP) “μ-manager” of theBelgian Science Policy (BELSPO, P7/25) and EU project BIOTREAT(EU grant no. 266039).
References
Amon RMW, Fitzner H-P, Benner R (2001) Linkages betweenbioreactivity, chemical composition and diagenetic state of marinedissolved organic matter. Limnol Oceanogr 46:287–297
Breugelmans P, D’Huys P-J, De Mot R, Springael D (2007)Characterization of novel linuron-mineralizing bacterial consortiaenriched from long-term linuron-treated agricultural soils. FEMSMicrobiol Ecol 62:374–385. doi:10.1111/j.1574-6941.2007.00391.x
Breugelmans P, Horemans B, Hofkens J, Springael D (2010) Responseto mixed substrate feeds of the structure and activity of a linuron-degrading triple-species biofilm. Res Microbiol 161:660–666.doi:10.1016/j.resmic.2010.06.006
Breugelmans P, Barken KB, Tolker-Nielsen T, Hofkens J, Dejonghe W,Springael D (2008) Architecture and spatial organization in atriple-species bacterial biofilm synergistically degrading thephenylurea herbicide linuron. FEMS Microbiol Ecol 64:271–282. doi:10.1111/j.1574-6941.2008.00470.x
Brye K, Norman J, Bundy L, Gower S (2001) Nitrogen and carbonleaching in agroecosystems and their role in denitrification poten-tial. J Environ Qual 30:58–70. doi:10.2134/jeq2001.30158x
Cheyns K, Calcoen J, Martin-Laurent F, Bru D, Smolders E, SpringaelD (2012) Effects of dissolved organic matter (DOM) at environ-mentally relevant carbon concentrations on atrazine degradationby Chelatobacter heintzii SalB. Appl Microbiol Biotechnol95:1333–1341. doi:10.1007/s00253-011-3741-1
Costerton JW, Lewandowski DE, Caldwell DE, Korber DR, Lappin-ScottHM (1995) Microbial biofilms. Ann Rev Microbiol 49:711–745
Dejonghe W, Berteloot E, Goris J, Boon N, Crul K, Maertens S, HöfteM, Vos PD, Verstraete W, Top EM (2003) Synergistic degradationof linuron by a bacterial consortium and isolation of a singlelinuron-degrading Variovorax strain. Appl Environ Microbiol69:1532–1541. doi:10.1128/AEM.69.3.1532-1541.2003
Hall-Stoodley L, Costerton JW, Stoodley P (2004) Bacterial biofilms:from the natural environment to infectious diseases. Nat RevMicrobiol 2:95–108
Hess TF, Schmidt SK, Silverstein J, Howe B (1990) Supplementalsubstrate enhancement of 2,4-dinitrophenol mineralization by abacterial consortium. Appl Environ Microbiol 56:1551–1558
Heydorn A, Nielsen AT, Hentzer M, Sternberg C, GivskovM, Ersbäll BK,Molin S (2000) Quantification of biofilm structures by the novelcomputer program COMSTAT. Microbiology 146:2395–2407
Horemans B, Smolders E, Springael D (2013a) Carbon source utiliza-tion profiles suggest additional metabolic interactions in a syner-gistic linuron degrading bacterial consortium. FEMS MicrobiolEcol 84:24–34. doi:10.1111/1574-6941.12033
942 Appl Microbiol Biotechnol (2014) 98:931–943

Horemans B, Vandermaesen J, Smolders E, Springael D (2013b)Cooperative dissolved organic carbon assimilation by a linurondegrading bacterial consortium. FEMS Microbiol Ecol 84:35–46.doi:10.1111/1574-6941.12036
Horemans B, Vandermaesen J, Vanhaecke L, Smolders E, Springael D(2013c) Variovorax sp. mediated biodegradation of the phenyl ureaherbicide linuron at micropollutant concentrations and effects of nat-ural dissolved organic matter as supplementary carbon source. ApplMicrobiol Biotechnol (in press). doi: 10.1007/s00253-013-4690-7
Kalbitz K, Schmerwitz J, Schwesig D, Matzne E (2003a) Biodegradationof soil-derived dissolved organic matter as related to its properties.Geoderma 113:273–291. doi:10.1016/S0016-7061(02)00365-8
Kalbitz K, Schwesig D, Schmerwitz J, Kaiser K, Haumaier L, Glaser B,Ellerbrock R, Leinweber P (2003b) Changes in properties of soil-derived dissolved organic matter induced by biodegradation. SoilBiol Biochem 35:1129–1142. doi:10.1016/S0038-0717(03)00165-2
Keuth S, Rehm H-J (1991) Biodegradation of phenanthrene byArthrobacter polychromogenes isolated from a contaminated soil.Appl Microbiol Biotechnol 34:804–808. doi:10.1007/bf00169354
Larsson P, Okla L, Tranvik L (1988) Microbial degradation of xeno-biotic, aromatic pollutants in humic water. Appl EnvironMicrobiol 54:1864–1867
Leenheer JA, Malcom RL, McKinley PW (1974) Occurrence ofdissolved organic carbon in selected groundwater samples in theUnited States. J Res US Geol Surv 2:361–369
Marschner B, Kalbitz K (2003) Controls of bioavailability and biode-gradability of dissolved organic matter in soils. Geoderma113:211–235. doi:10.1016/S0016-7061(02)00362-2
McFall SM, Abraham B, Narsolis CG, Chakrabarty AM (1997) Atricarboxylic acid cycle intermediate regulating transcription of achloroaromatic biodegradative pathway: fumarate-mediated re-pression of the clcABD operon. J Bacteriol 179:6729–6735
Müller C, Petruschka L, Cuypers H, Burchhardt G, Herrmann H (1996)Carbon catabolite repression of phenol degradation in Pseudomonasputida is mediated by the inhibition of the activator protein PhlR. JBacteriol 178:2030–2036
Murga R, Stewart PS, Daly D (1995) Quantitative analysis of biofilmthickness variability. Biotechnol Bioeng 45:503–510. doi:10.1002/bit.260450607
Nielsen AT, Tolker-Nielsen T, Barken KB, Molin S (2000) Role ofcommensal relationships on the spatial structure of a surface-attached microbial consortium. Environ Microbiol 2:59–68.doi:10.1046/j.1462-2920.2000.00084.x
Ortega-Calvo JJ, Saiz-Jimenez C (1998) Effect of humic fractions andclay on biodegradation of phenanthrene by a Pseudomonasfluorescens strain isolated from soil. Appl Environ Microbiol64:3123–3126
Petruschka LP, Burchhardt GB, Müller CM, Weihe CW, Herrmann HH(2001) The cyo operon of Pseudomonas putida is involved incarbon catabolite repression of phenol degradation. Mol GenetGenomics 266:199–206. doi:10.1007/s004380100539
Rentz JA, Alvarez PJJ, Schnoor JL (2004) Repression of Pseudomonasputida phenanthrene degrading activity by plant root extracts andexudates. Environ Microbiol 6:574–583. doi:10.1111/j.1462-2920.2004.00589.x
Salminen R (2005) Geochemical atlas of Europe. Part 1: backgroundinformation, methodology and maps. Geological Survey ofFinland, Espoo
Schmidt SK, Alexander M (1985) Effects of dissolved organiccarbon and second substrates on the biodegradation of organ-ic compounds at low concentrations. Appl Environ Microbiol49:822–827
Siemens J, Haas M, Kaupenjohann M (2003) Dissolved organic matterinduced denitrification in subsoils and aquifers. Geoderma113:253–271. doi:10.1016/S0016-7061(02)00364-6
Singh R, Paul D, Jain RK (2006) Biofilms: implications in bioremedia-tion. Trends Microbiol 14:389–397. doi:10.1016/j.tim.2006.07.001
Smith KEC, Thullner M, Wick LY, Harms H (2011) Dissolvedorganic carbon enhances the mass transfer of hydrophobicorganic compounds from nonaqueous phase liquids (NAPLs)into the aqueous phase. Environ Sci Technol 45:8741–8747.doi:10.1021/es202983k
Sørensen SR, Ronen Z, Aamand J (2002) Growth in coculture stimu-lates metabolism of the phenylurea herbicide isoproturon bySphingomonas sp. strain SRS2. Appl Environ Microbiol68:3478–3485. doi:10.1128/aem.68.7.3478-3485.2002
Thurman EM (1985) Organic geochemistry of natural waters.Nijhoff/Junk
Topp E, Crawford RL, Hanson RS (1988) Influence of readily metab-olizable carbon on pentachlorophenol metabolism by apentachlorophenol-degrading Flavobacterium sp. Appl EnvironMicrobiol 54:2452–2459
Volk CJ, Volk CB, Kaplan LA (1997) Chemical composition of bio-degradable dissolved organic matter in streamwater. LimnolOceanogr 42:39–44
Wick LY, Pasche N, Bernasconi SM, Pelz O, Harms H (2003)Characterization of multiple-substrate utilization by anthracene-degrading Mycobacterium frederiksbergense LB501T. ApplEnviron Microbiol 69:6133–6142. doi:10.1128/AEM.69.10.6133-6142.2003
Wuertz S, Hendrickx L, Kuehn M, Rodenacker K, Hausner M, Ron JD(2001) In situ quantification of gene transfer in biofilms. Methodsin enzymology, Volume 336 ed.), pp. 139–143. Academic Press.doi: 10.1016/S0076-6879(01)36585-0
Appl Microbiol Biotechnol (2014) 98:931–943 943
Related Documents











