Protist, Vol. 150, 283-295, October 1999 © Urban & Fischer Verlag http://www.urbanfischer.de/journals/protist ORIGINAL PAPER The Plastid in Plasmodium falciparum Asexual Blood Stages: a Three-Dimensional Ultrastructural Analysis Protist John Hopkinsa,b, Ruth Fowler"·b, Sanjeev Krishna c , lain Wilson d , Graham Mitchell b , and Lawrence Bannister"·1 a Department of Anatomy and Cell Biology, Guy's, King's College and St. Thomas's Hospitals' Medical and Dental Schools, Guy's Campus, Guy's Hospital London SE1 9RT, UK b Department of Immunobiology, Guy's, King's College and St. Thomas's Hospitals' Medical and Dental Schools, Guy's Campus, Guy's Hospital London SE1 9RT, UK c Department of Infectious Diseases, St.George's Hospital Medical School, Tooting, London SW17 OQT, UK d Parasitology Division, National Institute for Medical Research, Mill Hill, London NW7 1M, UK Submitted May 6, 1999; Accepted July 28, 1999 Monitoring Editor: Larry Simpson The plastid in Plasmodium fa/ciparum asexual stages is a tubular structure measuring about 0.5 IJm x 0.15 IJm in the merozoite, and 1.6 x 0.35 IJm in trophozoites. Each parasite contains a single plastid until this organelle replicates in late schizonts. The plastid always adheres to the (single) mitochon- drion, along its whole length in merozoites and early rings, but only at one end in later stages. Re- gions of the plastid are also closely related to the pigment vacuole, nuclear membrane and endoplas- mic reticulum. In merozoites the plastid is anchored to a band of 2-3 subpellicular microtubules. Re- constructions show the plastid wall is characteristically three membranes thick, with regions of addi- tional, complex membranes. These include inner and outer membrane complexes. The inner complex in the interior lumen is probably a rolled invagination of the plastid's inner membrane. The outer com- plex lies between the outer and middle wall membranes. The interior matrix contains ribosome-like granules and a network of fine branched filaments. Merozoites of P. berghei and P. knowlesi possess plastids similar in structure to those of P. fa/ciparum. A model is proposed for the transfer of mem- brane lipid from the plastid to other organelles in the parasite. Introduction Recent interest in the plastids (or apicoplasts) of apicomplexan parasites has centred on the prokary- ote-like nature of their DNA, RNA and ribosomes, and the implications of this for novel forms of 1Corresponding author; fax 44 171 9554915 e-mail [email protected] chemotherapy (for recent reviews see Feagin 1994; Fichera and Roos 1997; Jeffries and Johnson 1996; McFadden and Waller 1997; Williamson et al. 1994, 1996; Wilson and Williamson 1997; Wilson et al. 1994, 1996; see also Clough et al. 1997; Rogers et al. 1997). The morphological identity of the plastid was established finally by Kohler et al. (1997) who localized the characteristic plastid DNA in Toxo- plasma gondii to a membranous organelle bounded 1434-4610/99/150/03-283 $ 12.00/0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Protist, Vol. 150, 283-295, October 1999 © Urban & Fischer Verlaghttp://www.urbanfischer.de/journals/protist
ORIGINAL PAPER
The Plastid in Plasmodium falciparumAsexual Blood Stages:a Three-Dimensional Ultrastructural Analysis
Protist
John Hopkinsa,b, Ruth Fowler"·b, Sanjeev Krishnac, lain Wilsond, Graham Mitchellb, and
Lawrence Bannister"·1
a Department of Anatomy and Cell Biology, Guy's, King's College and St. Thomas's Hospitals' Medical and DentalSchools, Guy's Campus, Guy's Hospital London SE1 9RT, UK
b Department of Immunobiology, Guy's, King's College and St. Thomas's Hospitals' Medical and Dental Schools,Guy's Campus, Guy's Hospital London SE1 9RT, UK
c Department of Infectious Diseases, St.George's Hospital Medical School, Tooting, London SW17 OQT, UKd Parasitology Division, National Institute for Medical Research, Mill Hill, London NW7 1M, UK
Submitted May 6, 1999; Accepted July 28, 1999Monitoring Editor: Larry Simpson
The plastid in Plasmodium fa/ciparum asexual stages is a tubular structure measuring about 0.5 IJm x0.15 IJm in the merozoite, and 1.6 x 0.35 IJm in trophozoites. Each parasite contains a single plastiduntil this organelle replicates in late schizonts. The plastid always adheres to the (single) mitochondrion, along its whole length in merozoites and early rings, but only at one end in later stages. Regions of the plastid are also closely related to the pigment vacuole, nuclear membrane and endoplasmic reticulum. In merozoites the plastid is anchored to a band of 2-3 subpellicular microtubules. Reconstructions show the plastid wall is characteristically three membranes thick, with regions of additional, complex membranes. These include inner and outer membrane complexes. The inner complexin the interior lumen is probably a rolled invagination of the plastid's inner membrane. The outer complex lies between the outer and middle wall membranes. The interior matrix contains ribosome-likegranules and a network of fine branched filaments. Merozoites of P. berghei and P. knowlesi possessplastids similar in structure to those of P. fa/ciparum. A model is proposed for the transfer of membrane lipid from the plastid to other organelles in the parasite.
Introduction
Recent interest in the plastids (or apicoplasts) ofapicomplexan parasites has centred on the prokaryote-like nature of their DNA, RNA and ribosomes,and the implications of this for novel forms of
1Corresponding author;fax 44 171 9554915e-mail [email protected]
chemotherapy (for recent reviews see Feagin 1994;Fichera and Roos 1997; Jeffries and Johnson 1996;McFadden and Waller 1997; Williamson et al. 1994,1996; Wilson and Williamson 1997; Wilson et al.1994, 1996; see also Clough et al. 1997; Rogers etal. 1997). The morphological identity of the plastidwas established finally by Kohler et al. (1997) wholocalized the characteristic plastid DNA in Toxoplasma gondii to a membranous organelle bounded
1434-4610/99/150/03-283 $ 12.00/0

284 J. Hopkins et al.
by four membranes, long known to ultrastructuralinvestigators (e.g. the 'corps multilamellaire' ofVivier and Petitprez 1972). Ultrastructural profilesresembling the Toxoplasma organelle have alsobeen reported for Plasmodium (see Aikawa 1971;Aikawa et al. 1967; Hepler et al. 1966). Aikawa et al.(1967) described it in merozoites of Plasmodiume/ongatum as a rounded structure (hence the name'spherical body') bounded by membranes of 'undetermined number' and situated in an indentation ofthe mitochondrion, an association which they suggested implies metabolic interactions betweenthese two organelles.
The number of membranes surrounding apicomplexan plastids has generated some evolutionaryspeculation. The quadruple membranes in Toxoplasma and some other non-malarial apicomplexans resemble those of the plastids of chromistanalgae (McFadden and Gilson 1995), but phylogenetic analyses of nuclear 18S rDNAs provide strongevidence that apicomplexans, dinoflagellates andciliates form a separate monophyletic clade (Cavalier-Smith 1993). That plastids in dinoflagellates aresurrounded by only three membranes may be evidence that they were independently acquired(Palmer and Delwiche 1996), or were modified during the evolutionary divergence of dinoflagellatesand apicomplexans.
In the present paper we describe the detailedstructure of the plastid in Plasmodium falciparum. Apreliminary study showed us that it has a complexorganization, requiring three-dimensional information for an adequate analysis. We have therefore serially sectioned different asexual erythrocytic stagesof the parasite, and reconstructed the organizationof the plastid and its relations to other organelleswithin the parasite. This approach, which has onlyoccasionally been used for Plasmodium (e.g. Siomianny and Prensier 1986, to study the mitochondrionin P. falciparum trophozoites), has enabled us to address a series of questions, including the number ofmembranes forming its wall, its association with themitochondrion, and relation to other organelleswithin the parasite.
Results
The Plastid in the Ring, Trophozoite andEarly Schizont Stages
In all specimens examined up to the mid-schizontstage, a single plastid was present in each parasite(Figs. 1-4A-C), but thereafter the number increasedup t016 prior to the allocation of individual plastids
to budding merozoites (details of plastid replicationwill be given in another paper). In all stages the plastid is approximately cylindrical, with one or morebulges along its length which become more pronounced in trophozoites (Fig. 4C). As the intraerythrocytic cycle progresses the plastid lengthensconsiderably; in typical reconstructions the plastidin a merozoite is 0.5 mm long and in a young trophozoite 1.6 mm. In schizonts it increases considerablymore than this, although we have yet to completereconstructions of this stage. The merozoite plastidhas an approximately uniform diameter of about0.15 mm (Figs 4A, 5, 6), whereas in trophozoites andearly schizonts the width varies from 0.25 mm to0.35 mm depending on the region measured (Figs.4,7-21).
Detailed Structure of the Plastid
The internal morphology of the plastid (Figs. 9-17) isessentially similar throughout the different stages,although there are some changes in the details ofmembrane arrangements. We have studied theplastid most thoroughly in young trophozoiteswhere it is still quite short, and therefore relativelyeasy to reconstruct from serial sections (Figs. 4A,9-17), and where it is likely to be fully functional. Thedescription given here is based mainly on sequences taken from this stage.
Membrane number
Throughout the asexual cycle the plastid and mitochondrion are clearly distinct in structure because ofthe larger number of the plastid's membranes andits somewhat denser internal matrix. For its entirelength the plastid wall is composed of three membranes (Figs. 2, 3, 9, 16, 18, 20) designated here asouter, middle and inner, but in certain regions of theplastid there are also, variably, more numerousmembranes (see below). The basic three membranes have a characteristic intermembrane spacing, the two inner leaflets being very close to ortouching each other, while the outermost is separated from the middle membrane by a wide thoughvariable (20-40 nm) space. The outer membrane hasoccasional long leaf-like extensions into the surrounding cytoplasm (Fig.7, 8, 14, 15).
The profiles of all three wall membranes, but especiallythe inner two, are typically very irregular compared withthe generally smooth membranes of the parasite's mitochondrion (Fig. 2), a feature which initially made it difficultfor us to be sure of the number of membranes composingit. Although many micrographs clearly showed threemembranes, others suggested four or five, often appar-
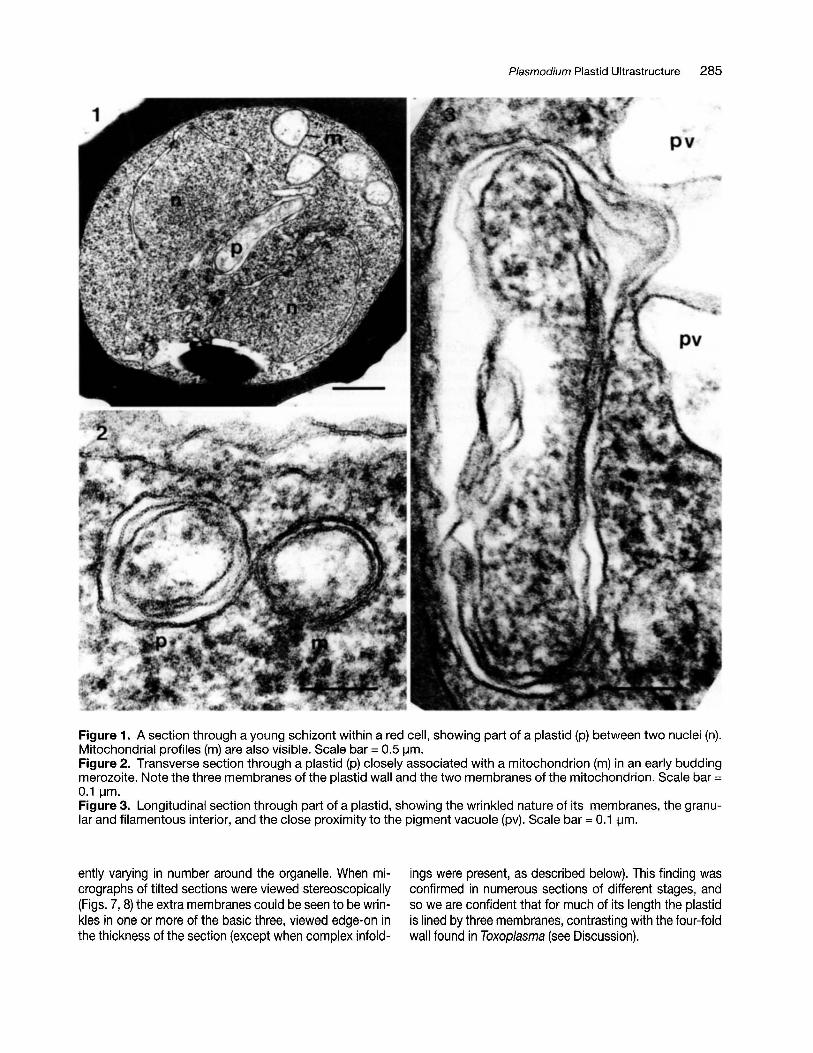
Plasmodium Plastid Ultrastructure 285
Figure 1. A section through a young schizont within a red cell, showing part of a plastid (p) between two nuclei (n).Mitochondrial profiles (m) are also visible. Scale bar = 0.5IJm.Figure 2. Transverse section through a plastid (p) closely associated with a mitochondrion (m) in an early buddingmerozoite. Note the three membranes of the plastid wall and the two membranes of the mitochondrion. Scale bar =0.1 IJm.Figure 3. Longitudinal section through part of a plastid, showing the wrinkled nature of its membranes, the granular and filamentous interior, and the close proximity to the pigment vacuole (pv). Scale bar = 0.1 IJm.
ently varying in number around the organelle. When micrographs of tilted sections were viewed stereoscopically(Figs. 7, 8) the extra membranes could be seen to be wrinkles in one or more of the basic three, viewed edge-on inthe thickness of the section (except when complex infold-
ings were present, as described below). This finding wasconfirmed in numerous sections of different stages, andso we are confident that for much of its length the plastidis lined by three membranes, contrasting with the four-foldwall found in Toxoplasma (see Discussion).
286 J. Hopkins et al.
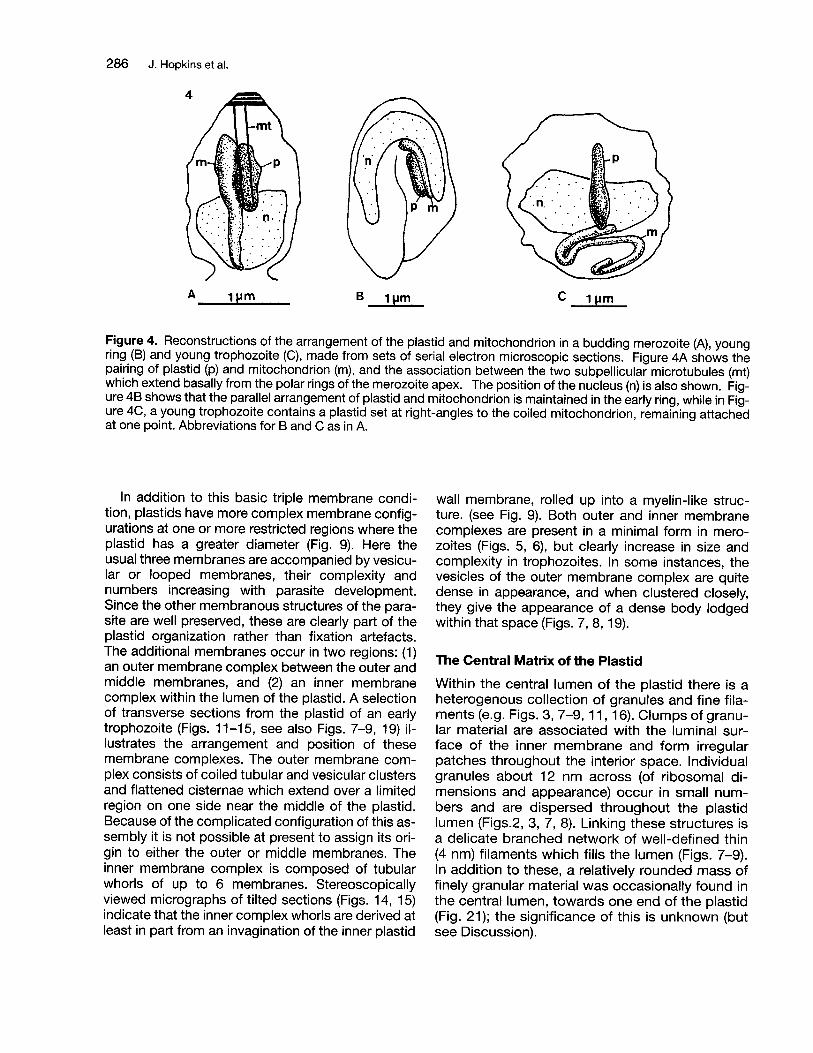
A 1IJm B c 1 pm
Figure 4. Reconstructions of the arrangement of the plastid and mitochondrion in a budding merozoite (A), youngring (8) and young trophozoite (C), made from sets of serial electron microscopic sections. Figure 4A shows thepairing of plastid (p) and mitochondrion (m), and the association between the two subpellicular microtubules (mt)which extend basally from the polar rings of the merozoite apex. The position of the nucleus (n) is also shown. Figure 48 shows that the parallel arrangement of plastid and mitochondrion is maintained in the early ring, while in Figure 4C, a young trophozoite contains a plastid set at right-angles to the coiled mitochondrion, remaining attachedat one point. Abbreviations for 8 and C as in A.
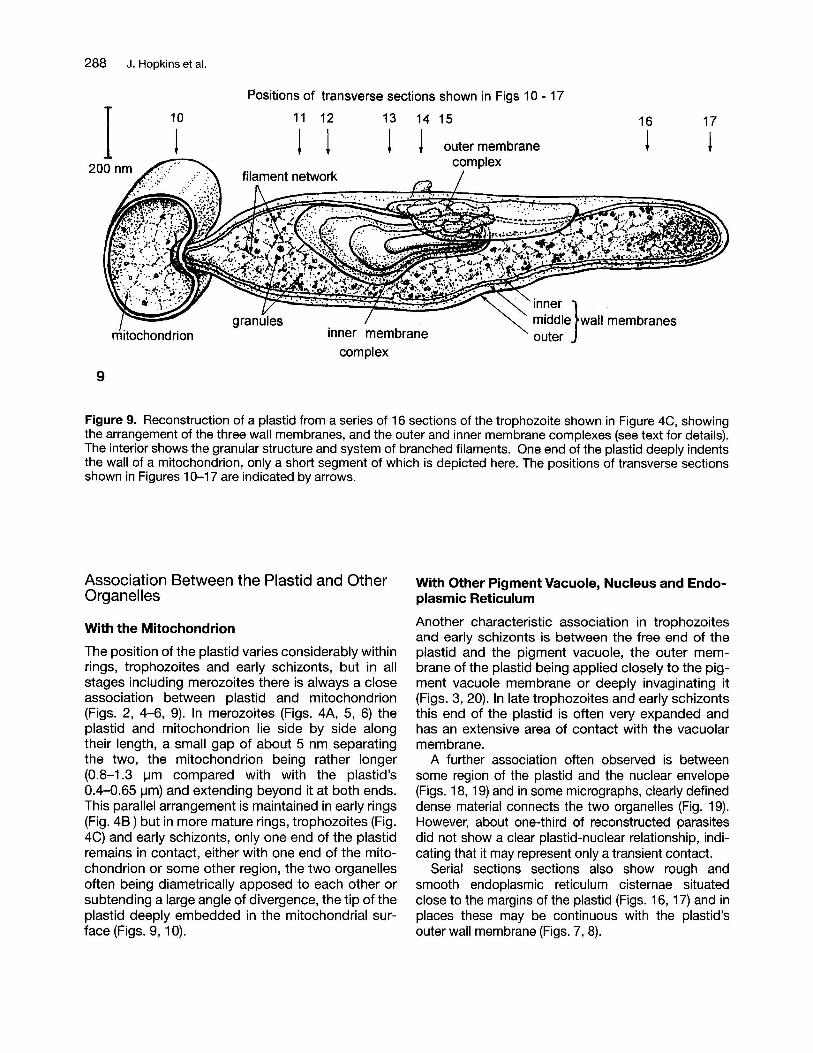
In addition to this basic triple membrane condition, plastids have more complex membrane configurations at one or more restricted regions where theplastid has a greater diameter (Fig. 9). Here theusual three membranes are accompanied by vesicular or looped membranes, their complexity andnumbers increasing with parasite development.Since the other membranous structures of the parasite are well preserved, these are clearly part of theplastid organization rather than fixation artefacts.The additional membranes occur in two regions: (1)an outer membrane complex between the outer andmiddle membranes, and (2) an inner membranecomplex within the lumen of the plastid. A selectionof transverse sections from the plastid of an earlytrophozoite (Figs. 11-15, see also Figs. 7-9,19) illustrates the arrangement and position of thesemembrane complexes. The outer membrane complex consists of coiled tubular and vesicular clustersand flattened cisternae which extend over a limitedregion on one side near the middle of the plastid.Because of the complicated configuration of this assembly it is not possible at present to assign its origin to either the outer or middle membranes. Theinner membrane complex is composed of tubularwhorls of up to 6 membranes. Stereoscopicallyviewed micrographs of tilted sections (Figs. 14, 15)indicate that the inner complex whorls are derived atleast in part from an invagination of the inner plastid
wall membrane, rolled up into a myelin-like structure. (see Fig. 9). Both outer and inner membranecomplexes are present in a minimal form in merozoites (Figs. 5, 6), but clearly increase in size andcomplexity in trophozoites. In some instances, thevesicles of the outer membrane complex are quitedense in appearance, and when clustered closely,they give the appearance of a dense body lodgedwithin that space (Figs. 7, 8, 19).
The Central Matrix of the Plastid
Within the central lumen of the plastid there is aheterogenous collection of granules and fine filaments (e.g. Figs. 3, 7-9, 11, 16). Clumps of granular material are associated with the luminal surface of the inner membrane and form irregularpatches throughout the interior space. Individualgranules about 12 nm across (of ribosomal dimensions and appearance) occur in small numbers and are dispersed throughout the plastidlumen (Figs.2, 3, 7, 8). Linking these structures isa delicate branched network of well-defined thin(4 nm) filaments which fills the lumen (Figs. 7-9).In addition to these, a relatively rounded mass offinely granular material was occasionally found inthe central lumen, towards one end of the plastid(Fig. 21); the significance of this is unknown (butsee Discussion).
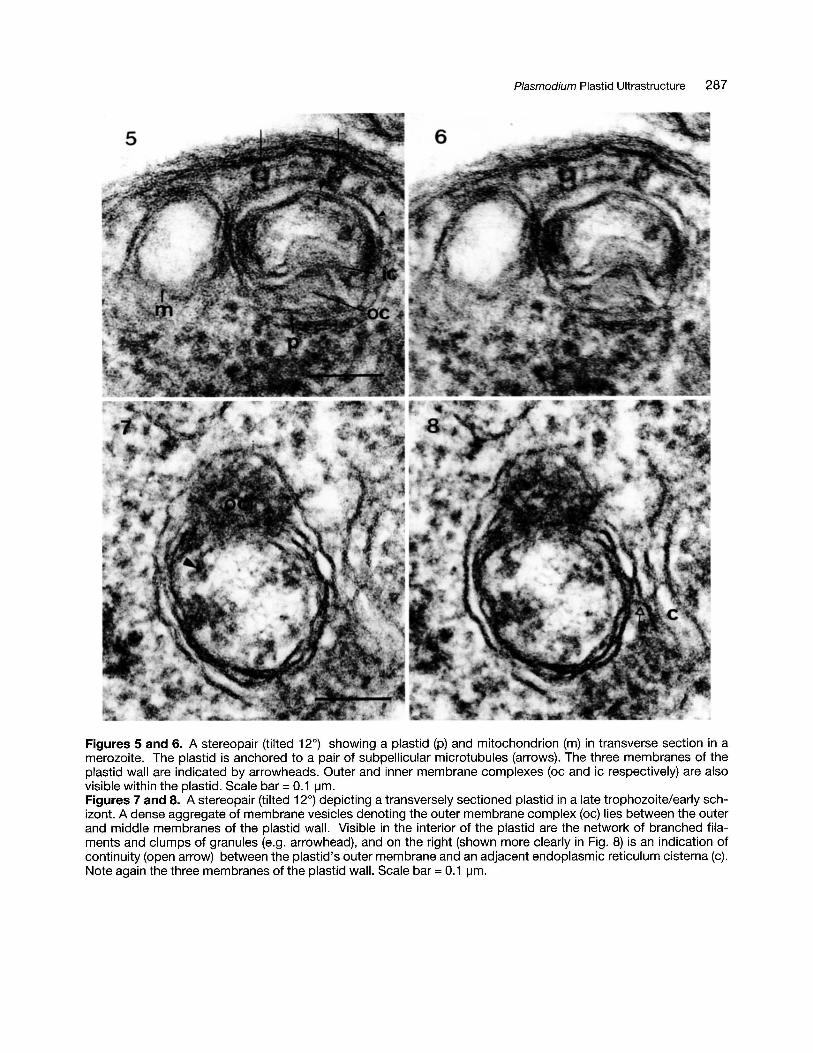
Plasmodium Plastid Ultrastructure 287
Figures 5 and 6. A stereopair (tilted 12°) showing a plastid (p) and mitochondrion (m) in transverse section in amerozoite. The plastid is anchored to a pair of subpellicular microtubules (arrows). The three membranes of theplastid wall are indicated by arrowheads. Outer and inner membrane complexes (oc and ic respectively) are alsovisible within the plastid. Scale bar = 0.1 ~m.
Figures 7 and 8. A stereopair (tilted 12°) depicting a transversely sectioned plastid in a late trophozoite/early schizont. A dense aggregate of membrane vesicles denoting the outer membrane complex (oc) lies between the outerand middle membranes of the plastid wall. Visible in the interior of the plastid are the network of branched filaments and clumps of granules (e.g. arrowhead), and on the right (shown more clearly in Fig. 8) is an indication ofcontinuity (open arrow) between the plastid's outer membrane and an adjacent endoplasmic reticulum cisterna (c).Note again the three membranes of the plastid wall. Scale bar =0.1 ~m.
288 J. Hopkins et al.
17
!16
113 14 15
t !11 12
! 1
Positions of transverse sections shown in Figs 10 - 17
10
!
mitochondrion
I
9
Figure 9. Reconstruction of a plastid from a series of 16 sections of the trophozoite shown in Figure 4C, showingthe arrangement of the three wall membranes, and the outer and inner membrane complexes (see text for details).The interior shows the granular structure and system of branched filaments. One end of the plastid deeply indentsthe wall of a mitochondrion, only a short segment of which is depicted here. The positions of transverse sectionsshown in Figures 10-17 are indicated by arrows.
Association Between the Plastid and OtherOrganelles
With the Mitochondrion
The position of the plastid varies considerably withinrings, trophozoites and early schizonts, but in allstages including merozoites there is always a closeassociation between plastid and mitochondrion(Figs. 2, 4-6, 9). In merozoites (Figs. 4A, 5, 6) theplastid and mitochondrion lie side by side alongtheir length, a small gap of about 5 nm separatingthe two, the mitochondrion being rather longer(0.8-1.3 I-Im compared with with the plastid's0.4-0.65 I-Im) and extending beyond it at both ends.This parallel arrangement is maintained in early rings(Fig. 48) but in more mature rings, trophozoites (Fig.4C) and early schizonts, only one end of the plastidremains in contact, either with one end of the mitochondrion or some other region, the two organellesoften being diametrically apposed to each other orsubtending a large angle of divergence, the tip of theplastid deeply embedded in the mitochondrial surface (Figs. 9, 10).
With Other Pigment Vacuole, Nucleus and Endoplasmic Reticulum
Another characteristic association in trophozoitesand early schizonts is between the free end of theplastid and the pigment vacuole, the outer membrane of the plastid being applied closely to the pigment vacuole membrane or deeply invaginating it(Figs. 3, 20). In late trophozoites and early schizontsthis end of the plastid is often very expanded andhas an extensive area of contact with the vacuolarmembrane.
A further association often observed is betweensome region of the plastid and the nuclear envelope(Figs. 18, 19) and in some micrographs, clearly defineddense material connects the two organelles (Fig. 19).However, about one-third of reconstructed parasitesdid not show a clear plastid-nuclear relationship, indicating that it may represent only a transient contact.
Serial sections sections also show rough andsmooth endoplasmic reticulum cisternae situatedclose to the margins of the plastid (Figs. 16, 17) and inplaces these may be continuous with the plastid'souter wall membrane (Figs. 7, 8).
Plasmodium Plastid Ultrastructure 289
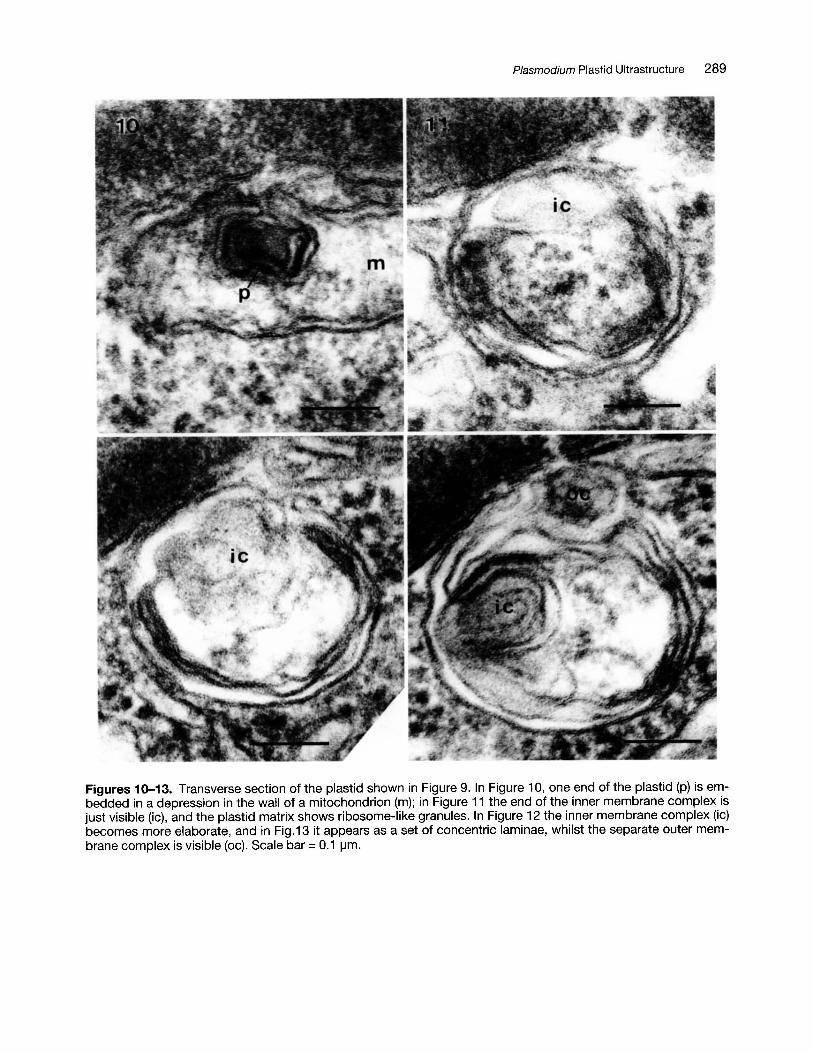
Figures 10-13. Transverse section of the plastid shown in Figure 9. In Figure 10, one end of the plastid (p) is embedded in a depression in the wall of a mitochondrion (m); in Figure 11 the end of the inner membrane complex isjust visible (ic), and the plastid matrix shows ribosome-like granules. In Figure 12 the inner membrane complex (ic)becomes more elaborate, and in Fig.13 it appears as a set of concentric laminae, whilst the separate outer membrane complex is visible (oc). Scale bar =0.1 \-1m.
290 J. Hopkins et al.
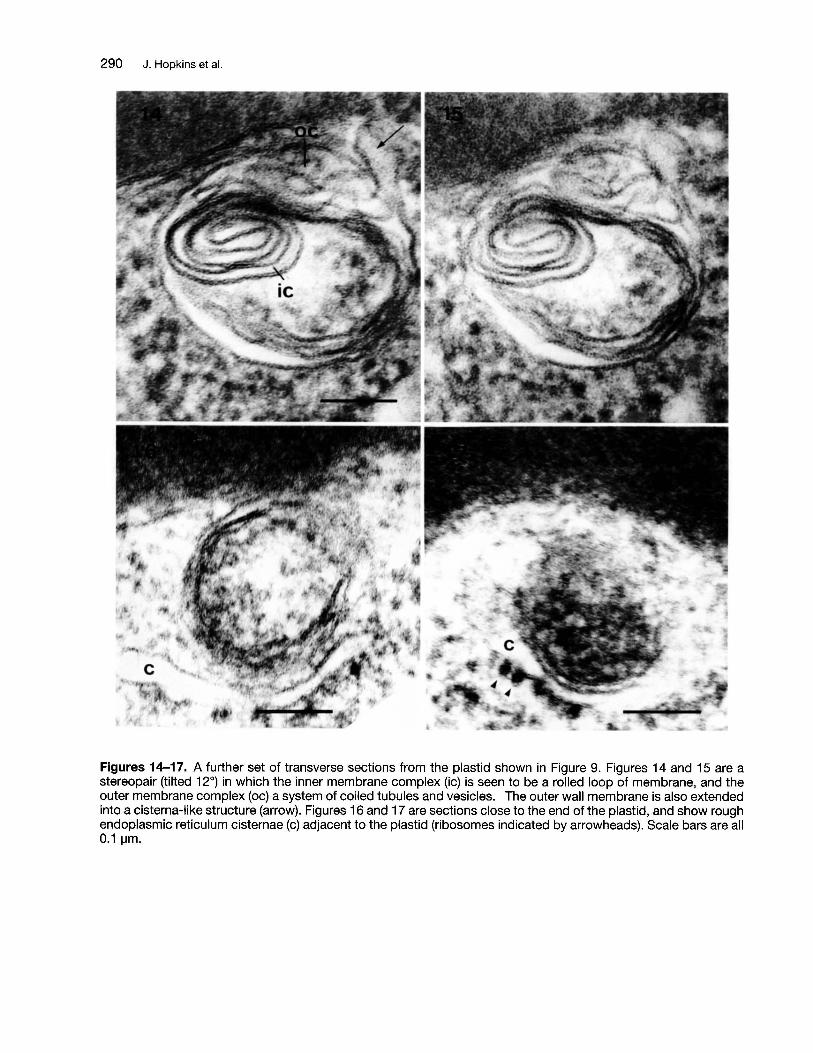
Figures 14-17. A further set of transverse sections from the plastid shown in Figure 9. Figures 14 and 15 are astereopair (tilted 12°) in which the inner membrane complex (ic) is seen to be a rolled loop of membrane, and theouter membrane complex (oc) a system of coiled tubules and vesicles. The outer wall membrane is also extendedinto a cisterna-like structure (arrow). Figures 16 and 17 are sections close to the end of the plastid, and show roughendoplasmic reticulum cisternae (c) adjacent to the plastid (ribosomes indicated by arrowheads). Scale bars are all0.1 IJm.
Plasmodium Plastid Ultrastructure 291
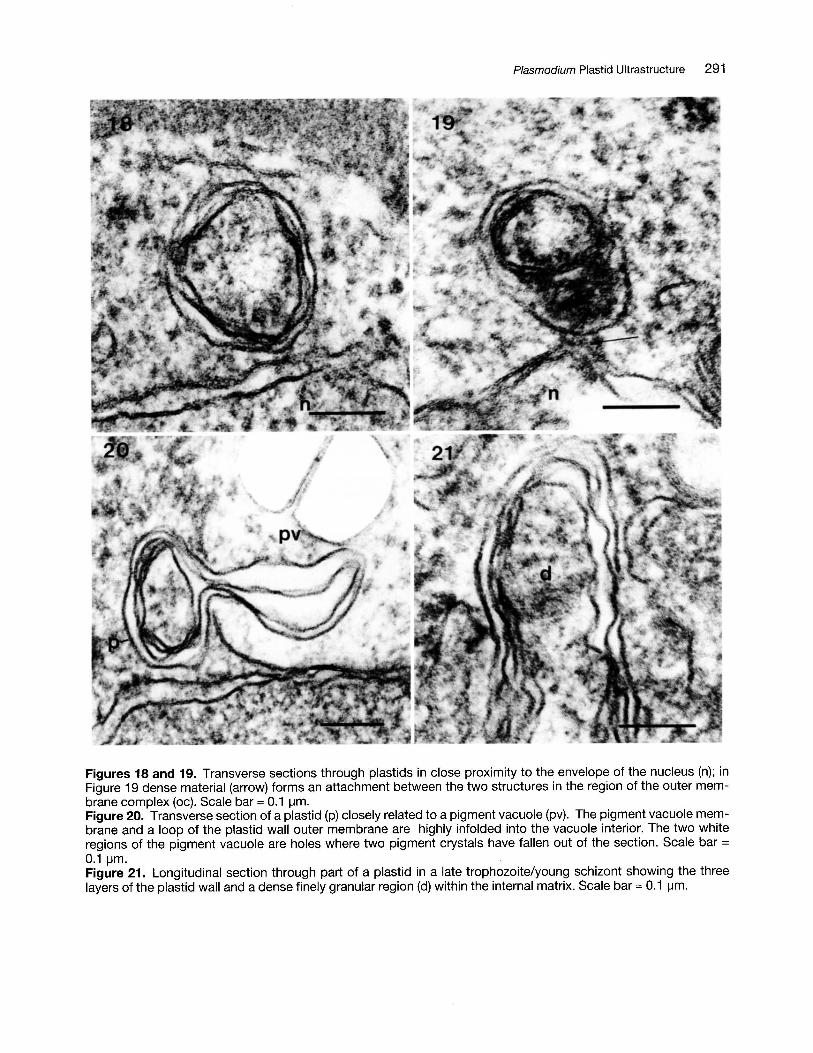
Figures 18 and 19. Transverse sections through plastids in close proximity to the envelope of the nucleus (n); inFigure 19 dense material (arrow) forms an attachment between the two structures in the region of the outer membrane complex (oc). Scale bar = 0.1 ~m.
Figure 20. Transverse section of a plastid (p) closely related to a pigment vacuole (pv). The pigment vacuole membrane and a loop of the plastid wall outer membrane are highly infolded into the vacuole interior. The two whiteregions of the pigment vacuole are holes where two pigment crystals have fallen out of the section. Scale bar =0.1 ~m.
Figure 21. Longitudinal section through part of a plastid in a late trophozoite/young schizont showing the threelayers of the plastid wall and a dense finely granular region (d) within the internal matrix. Scale bar = 0.1 ~m.
292 J. Hopkins et al.
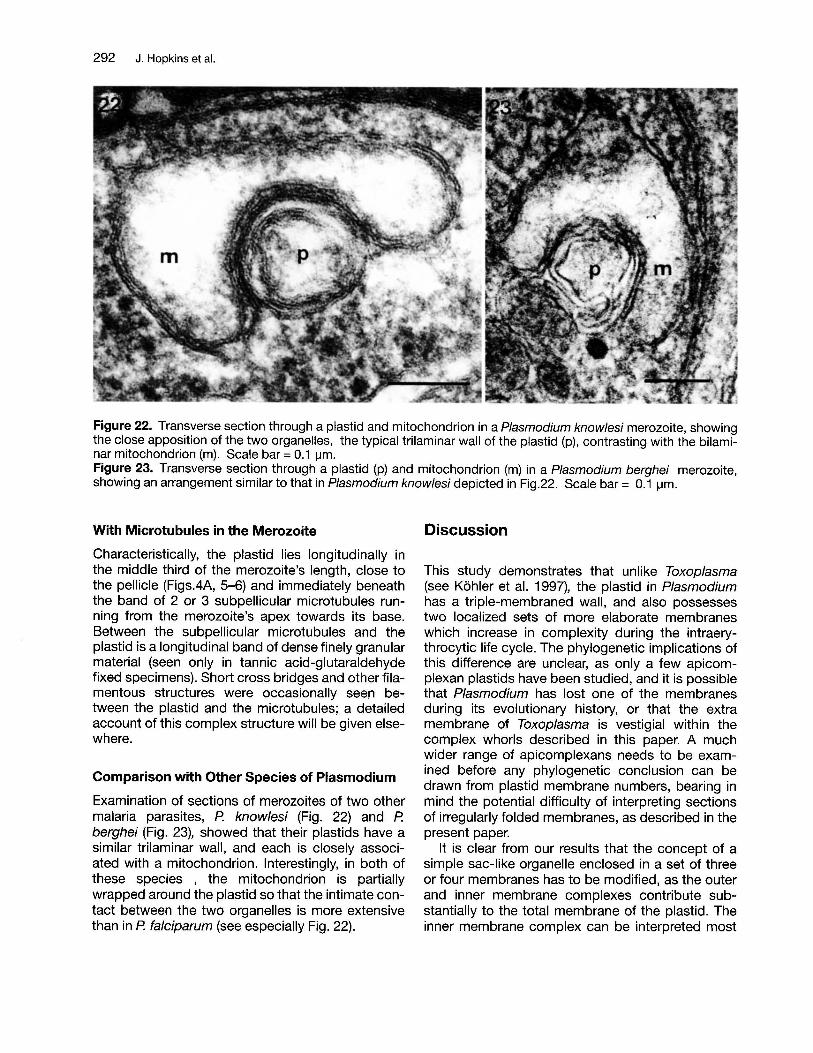
Figure 22. Transverse section through a plastid and mitochondrion in a Plasmodium knowlesi merozoite, showingthe close apposition of the two organelles, the typical trilaminar wall of the plastid (p), contrasting with the bilaminar mitochondrion (m). Scale bar = 0.1 ~m.
Figure 23. Transverse section through a plastid (p) and mitochondrion (m) in a Plasmodium berghei merozoite,showing an arrangement similar to that in Plasmodium knowlesi depicted in Fig.22. Scale bar = 0.1 ~m.
With Microtubules in the Merozoite
Characteristically, the plastid lies longitudinally inthe middle third of the merozoite's length, close tothe pellicle (FigsAA, 5-6) and immediately beneaththe band of 2 or 3 subpellicular microtubules running from the merozoite's apex towards its base.Between the subpellicular microtubules and theplastid is a longitudinal band of dense finely granularmaterial (seen only in tannic acid-glutaraldehydefixed specimens). Short cross bridges and other filamentous structures were occasionally seen between the plastid and the microtubules; a detailedaccount of this complex structure will be given elsewhere.
Comparison with Other Species of Plasmodium
Examination of sections of merozoites of two othermalaria parasites, P. know/esi (Fig. 22) and P.berghei (Fig. 23), showed that their plastids have asimilar trilaminar wall, and each is closely associated with a mitochondrion. Interestingly, in both ofthese species , the mitochondrion is partiallywrapped around the plastid so that the intimate contact between the two organelles is more extensivethan in P. falciparum (see especially Fig. 22).
Discussion
This study demonstrates that unlike Toxoplasma(see Kohler et al. 1997), the plastid in Plasmodiumhas a triple-membraned wall, and also possessestwo localized sets of more elaborate membraneswhich increase in complexity during the intraerythrocytic life cycle. The phylogenetic implications ofthis difference are unclear, as only a few apicomplexan plastids have been studied, and it is possiblethat Plasmodium has lost one of the membranesduring its evolutionary history, or that the extramembrane of Toxoplasma is vestigial within thecomplex whorls described in this paper. A muchwider range of apicomplexans needs to be examined before any phylogenetic conclusion can bedrawn from plastid membrane numbers, bearing inmind the potential difficulty of interpreting sectionsof irregularly folded membranes, as described in thepresent paper.
It is clear from our results that the concept of asimple sac-like organelle enclosed in a set of threeor four membranes has to be modified, as the outerand inner membrane complexes contribute substantially to the total membrane of the plastid. Theinner membrane complex can be interpreted most

simply as a rolled-up myelin-like invagination of theinner wall membrane. The outer membrane complexis not so obviously continuous with either the outeror middle membranes, and its vesicles and tubulescould be derived by budding from either the middleor outer membrane.
Recently, Waller et al. (1998) have demonstratedthat the plastids of P. fa/ciparum and Toxoplasmagondii are active in membrane lipid synthesis (as arethe chloroplasts of higher plants which can exportlipids to other cell structures: see the review by Joyard et al. 1998). Combining this information withthe structural data we describe here, an interestingpossibility emerges, i.e. that the apicomplexan plastid is essentially a membrane lipid factory for theparasite, budding off lipid in vesicular form betweenthe inner and outer membranes and transporting itto adjacent organelles via extensions of its surface.The enlargement of the plastid and the increasingcomplexity of its internal membranes in trophozoitesand early schizont stages may therefore denote anincreasing demand for membrane lipids as othermembranous organelles of the parasite increase insize and activity, although of course some membrane generation would also be needed for plastidreplication in the schizont stage. If this view of plastid function is correct, plastids in other apicomplexan genera are likely to have similar membraneconfigurations, and a detailed analysis of plastidstructure in representative genera of apicomplexanswould be of much interest.
Turning to the association between the plastidand mitochondrion, first described in Plasmodiumelongatum merozoites by Aikawa et al. (1967), wehave shown that in Plasmodium falciparum thetwo organelles invariably adhere to each other,with varying degrees of contact throughout theasexual cycle. As suggested by Aikawa et al.(1967), this duality may infer a metabolic interaction between the two structures. The attachmentmay also have a mechanical role, ensuring pairedallocations of the two organelles to buddingmerozoites at the end of schizogony.
The association between plastid and mitochondrion is particularly striking in merozoites,where in P. falciparum the position of both arespecified by the plastid's connection to the bandof subpellicular microtubules running along oneside of this stage (see Bannister and Mitchell1995; Read et al. 1993). Microtubules have beenshown in other organisms to be responsible formitochondrial movements (see e.g. Baumann andMurphy 1995; Knabe and Kuhn 1996), and couldbe the means of propelling plastids, mitochondriaand other organelles into merozoites during
Plasmodium Plastid Ultrastructure 293
merozoite assembly (see Bannister and Mitchell1995; Tilney and Tilney 1996). However, experiments in our laboratory with anti-microtubuledrugs have as yet failed to show inhibition of mitochondrial placement during the formation ofmerozoites (Fowler et al. 1998), and the deeplyplaced location of the plastid in P. berghei and P.knowlesi suggest that they may not be anchoredto microtubules in the same way in these species.Further work is in progress to clarify this issue.
There are three other plastid associations whichmay be significant, i.e. with the pigment vacuole,the nuclear membrane and endoplasmic reticulum.Although proximity does not prove a functional relationship, there are clear signs of structural connection or interaction between the plastid andthese structures. As proposed above, the plastidmay deliver membrane lipid to such organelles(and also perhaps to the mitochondrion), but in theinstances of the nucleus and endoplasmic reticulum, the traffic may consist of nuclear-encodedproteins moving in the opposite direction. i.e. intothe plastid from their sites of synthesis in the endoplasmic reticulum and cisterna of the nuclear envelope, as shown biochemically by Waller et al.(1998). Intermittent fusion between the plastidouter membrane, including its leaf-like extensions,with these cisternae could afford a route by whichsuch proteins are conveyed to the plastid.
The inner matrix of the plastid also deservescomment. The 12 nm granules are clearly goodcandidates for a ribosomal identity on the basis oftheir size and appearance. The fine filaments ofthe matrix are approximately the width of DNAstrands, but the results of Waller et al. (1998) fromfluorescence labeling indicate that the DNA isconcentrated in a restricted region of the plastid.The dense material we show in Figure 21 may berelevant to this finding. Further work is in progressto localise DNA ultrastructurally within the plastid.
Methods
Cultures of Plasmodium falciparum (IT04 strain)were grown in continuous culture and loosely synchronised, maturing schizonts were concentratedon Percoll cushions by standard techniques (seee.g. Fowler et al. 1998). After collection, the cellswere incubated for a further 2h at 37°C. Thereafterthe cells were processed for electron microscopyaccording to a modification of the protocol described by Bannister and Mitchell (1989). Briefly, theparasites were concentrated again by brief centrifugation at 6000xg and fixed in suspension for 2 hrs in

294 J. Hopkins et al.
2.5% glutaraldehyde in 0.075M sodium cacodylatebuffer (pH 7.2) at room temperature, in some caseswith the addition of 1% tannic acid (final pH 7.2).Cells were washed x3 in cacodylate buffer, thenconcentrated as above into pellets which were postfixed in 1% osmium tetroxide (1 h), briefly washed inbuffer, then block stained for 1 h in 1% aqueousuranyl acetate. Pellets were then dehydrated in anacetone series and embedded in Medium TAABresin. Serial sections were cut at a thickness of 85nm on a Reichert Ultracut E ultramicrotome,mounted on slotted grids covered with a thin film ofpioloform, and contrasted with uranyl acetate andlead citrate. Depending on orientation and externalshape, rings, trophozoites and schizonts each required up to 75 serial sections for complete reconstruction, while merozoites needed up to 32 sections. Sectioned cells were photographed atx12,000 and printed at x53,000 for survey reconstructions, and at x40,000 (printed at x120,000) formore detailed reconstructions. Micrographs weretraced on to acetate sheets, and reconstructedgraphically from these. For detailed morphology,thinner (50 nm) sections were also prepared and examined. In some cases serial sections and thinnersingle sections were tilted using a goniometer stageand photographed at 0° and 12° at x100,000, thenviewed with a stereoscope, to analyse complexmembrane configurations and other internal details.
For comparison with Plasmodium fa/ciparum,electron microscopy was carried out on specimensof Plasmodium knowlesi prepared for a previousstudy (Bannister and Mitchell 1989), and on Plasmodium berghei kindly provided by Dr. A.w. Thomas(Primate Biomedical Research Centre, Rijkswijk,Netherlands), prepared as in Kocken et al. (1998).
Acknowledgements
The authors thank the Wellcome Trust (Grants No.037082 and 048244) and the Special Trustees forSt.Thomas' Hospital for supporting this study. Sanjeev Krishna is a Wellcome Trust Senior ResearchFellow in Clinical Science. The authors are also indebted to the Electron Microscope Unit at UMDS(Guy's site) for photographic assistance, and to theMedical Research Council of the UK and ProfessorB. Boycott for provision of serial sectioning facilities.
References
Aikawa M (1971) Plasmodium: The fine structure ofmalarial parasites. Exp Parasitol 30: 284-320
Aikawa M, Huff CG, Sprinz H (1967) Fine structure ofthe asexual stages of Plasmodium e/ongatum. J CellBioi 34: 229-249
Bannister LH, Mitchell GH (1989) The fine structure ofsecretion by Plasmodium knowlesi merozoites duringred cell invasion. J Protozool 36: 362-367
Bannister LH, Mitchell GH (1995) The role of the cytoskeleton in Plasmodium falciparum merozoite biology: an electron microscopic view. Ann Trop Med Parasitol 89: 105-111
Baumann 0, Murphy DB (1995) Microtubule-associated movement of mitochondria and small particles inAcanthamoeba castel/anii. Cell Motil Cytoskeleton 32:305-317
Cavalier-Smith T (1993) Kingdom protozoa and its 18phyla. Microbiol Rev 57: 953-994
Clough B, Strath M, Preiser P, Denny P, Wilson I R(1997) Thiostrepton binds to malarial plastid rRNA.FEBS Letters 406: 123-125
Feagin JE (1994) The extrachromosomal DNAs of apicomplexan parasites. Annu Rev Microbiol48: 81-104
Fichera ME, Roos DS (1997) A plastid organelle as adrug target in apicomplexan parasites. Nature 390:407-409
Fowler RE, Fookes RE, Lavin FM, Bannister LH,Mitchell GH (1998) Microtubules in Plasmodium falciparum merozoites and their importance for invasion oferythrocytes. Parasitology 117: 425-433
Hepler PK, Huff CG, Sprinz H (1966) The fine structureof the exo-erythrocytic stages of Plasmodium fal/ax. JCell Bioi 30: 333-358
Jeffries AC, Johnson AM (1996) The growing importance of the plastid-like DNAs of the Apicomplexa. Int JParasitol 26: 1139-1150
Joyard J, Teyssiere E, Miege C, Berny-Seigneurin D,Marf~chal E, Block MA, Dorne AJ, Rollande N, AjlaniE, Douce R (1998) The biochemical machinery of plastid envelope membranes. Plant Physiol118: 715-723
Knabe W, Kuhn HJ (1996) The role of microtubules andmicrotubule-organising centres during the migration ofmitochondria. J Anat 189: 383-391
Kocken CHM, van derWel AM, Dubbeld MA, NarumDL, van de Rijke FM, van Gemert GJ, van der LindeX, Bannister LH, Janse C, Waters AP, Thomas AW(1998) Precise timing of expression of a Plasmodiumfalciparum-derived transgene in Plasmodium berghei isa critical determinant of subsequent cellular localization. J Bioi Chem 273: 15119-15124
Kohler S, Delwiche CF, Denny PD, Tilney LG, Webster P, Wilson RJM, Palmer JD, Roos DS (1997) Aplastid of probable green algal origin in apicomplexanparasites. Science 275: 1485-1487

McFadden G, Gilson P (1995) Something borrowed,something green: lateral transfer of chloroplasts by secondary endosymbiosis. Trends Ecol Evo110: 12-17
McFadden GI, Waller RF (1997) Plastids in parasites ofhumans. BioEssays 19: 1033-1040
Palmer JD, Delwiche CF (1996) Second-hand chloroplasts and the case of the disappearing nucleus. ProcNatl Acad Sci USA 93: 7432-35
Read M, Sherwin T, Holloway SP, Gull K, Hyde JE(1993) Microtubular organisation visualised by immunofluorescence microscopy during erythrocytic schizogony in Plasmodium falciparum and investigation ofpost-translational modifications of parasite tubulin. Parasitology 106: 223-232
Rogers MJ, BukhmanYV, McCutchan TF, Draper DE(1997) Interaction of thiostrepton with an RNA fragmentderived from the plastid-encoded ribosomal RNA of themalaria parasite. RNA 3: 815-820
Siomianny C, Prensier G (1986) Application of the serial sectioning and tridimensional reconstruction techniques to the morphological study of the Plasmodiumfalciparum mitochondrion. J Parasitol 72: 595-598
Tilney LG, Tilney MS (1996) The cytoskeleton of protozoan parasites. Curr Opin Cell Bioi 8: 43-48
Vivier E, Petitprez A (1972) Donees ultrastructuralescomplementaires, morphologiques et cytochimiques,sur Toxoplasma gondii. Protistologica 8: 199-221
Plasmodium Plastid Ultrastructure 295
Waller RF, Keeling PJ, Donald RGK, Striepen B,Handman E., Lang-Unnasch N, Cowman AF, BesraGS, Roos OS, McFadden GI (1998) Nuclear-encodedproteins target to the plastid in Toxoplasma gondii andPlasmodium fa/ciparum. Proc Natl Acad Sci USA 95:12352-12357
Williamson DH, Preiser PR, Wilson RJM (1996) Organelle DNAs: the bit players in malaria parasite DNAreplication. Parasitol Today 12: 357-362
Williamson DH, Gardner MJ, Preiser P, Moore OJ,Rangachari K, Wilson RJM (1994) The evolutionaryorigin of the malaria parasite's 35 kb circular DNA; newevidence supports a possible rhodophyte ancestry. MolGen Genet 243: 249-252
Wilson RJM, Williamson DH (1997) Extrachromosomal DNA in the Apicomplexa. Microbiol Mol Bioi Rev 6:1-16
Wilson RJM, Williamson DH, Preiser PR (1994)Malaria and other apicomplexans: the "plant" connection. Infect Agents Dis 3: 29-37
Wilson, RJM, Denny PO, Preiser PR, Rangachari K,Roberts K, Roy A, Whyte A., Strath M, Moore OJ,Moore PW, Williamson DH (1996) Complete gene mapof the plastid-like DNA of the malaria parasite Plasmodium fa/ciparum. J Mol Bioi 261: 155-172
Related Documents











