Systematic and Applied Microbiology 30 (2007) 297–308 The phylogeny of the genus Nitrobacter based on comparative rep-PCR, 16S rRNA and nitrite oxidoreductase gene sequence analysis Bram Vanparys a, , Eva Spieck b , Kim Heylen a , Lieven Wittebolle c , Joke Geets c , Nico Boon c , Paul De Vos a a Laboratory of Microbiology, Department of Biochemistry, Physiology and Microbiology, Ghent University, K.L. Ledeganckstraat 35, B-9000 Gent, Belgium b Biozentrum Klein Flottbek, Abteilung Mikrobiologie, University of Hamburg, Ohnhorststr. 18, D-22609 Hamburg, Germany c Laboratory of Microbial Ecology and Technology (LabMET), Ghent University, Coupure Links 653, B-9000 Gent, Belgium Received 11 October 2006 Abstract Strains of Nitrobacter mediate the second step in the nitrification process by oxidizing nitrite to nitrate. The phylogenetic diversity of the genus is currently not well investigated. In this study, a rep-PCR profile and the nearly complete 16S rRNA gene sequence of 30 strains, comprising a wide physiological as well as ecological diversity and encompassing representatives of the four species, were determined. The sequence diversity of the 16S rRNA gene between different species was low, indicating the need for additional phylogenetic markers. Therefore, primers were developed for amplifying the complete nxrX gene and a 380 bp fragment of the nxrB1 gene, which are both genes involved in the nitrite oxidation process. These genes confirmed the division into phylogenetic groups revealed by the 16S rRNA gene but showed a better discriminatory power. They can be a valuable additional tool for phylogenetic analysis within the genus Nitrobacter and can assist in the identification of new Nitrobacter isolates. r 2006 Elsevier GmbH. All rights reserved. Keywords: Nitrobacter; Nitrite oxidoreductase; nxrA; nxrB; nxrX; 16S rRNA; Rep-PCR; Phylogenetic diversity Introduction Nitrification, the microbiological conversion of am- monia to nitrate, is a key process in the nitrogen cycle. The process consists of two subsequent steps: (i) the oxidation of ammonia to nitrite and (ii) the oxidation of nitrite to nitrate. The latter reaction is performed by the nitrite oxidizing bacteria (NOB), which belong to four phylogenetically unrelated genera: Nitrobacter (a-Pro- teobacteria), Nitrococcus (g-Proteobacteria), Nitrospina (preliminarily assigned to the d-Proteobacteria) and Nitrospira (separate phylum; [11,40]). Although Nitrospira is currently regarded as more important than Nitrobacter in, for example, wastewater treatment plants and aquaria, Nitrobacter has been shown to coexist with Nitrospira in plants with temporally or spatially elevated nitrite concentrations [9]. Furthermore, Nitrobacter is still considered an important genus in fertilized as well as unfertilized soils [14], which are regarded as the privileged habitats of Nitrobacter [10,15]. Nitrobacter is also associated with landfill leachate treatment plants [20] and rivers [8], and is important in the erosion of rocks [23] and the deterioration of historic monuments [24]. ARTICLE IN PRESS www.elsevier.de/syapm 0723-2020/$ - see front matter r 2006 Elsevier GmbH. All rights reserved. doi:10.1016/j.syapm.2006.11.006 Corresponding author. Tel.: +32 9 264 51 01; fax: +32 9 264 53 46. E-mail address: [email protected] (B. Vanparys).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
0723-2020/$ - se
doi:10.1016/j.sy
�CorrespondE-mail addr
Systematic and Applied Microbiology 30 (2007) 297–308
www.elsevier.de/syapm
The phylogeny of the genus Nitrobacter based on comparative rep-PCR,
16S rRNA and nitrite oxidoreductase gene sequence analysis
Bram Vanparysa,�, Eva Spieckb, Kim Heylena, Lieven Wittebollec, Joke Geetsc,Nico Boonc, Paul De Vosa
aLaboratory of Microbiology, Department of Biochemistry, Physiology and Microbiology, Ghent University,
K.L. Ledeganckstraat 35, B-9000 Gent, BelgiumbBiozentrum Klein Flottbek, Abteilung Mikrobiologie, University of Hamburg, Ohnhorststr. 18, D-22609 Hamburg, GermanycLaboratory of Microbial Ecology and Technology (LabMET), Ghent University, Coupure Links 653, B-9000 Gent, Belgium
Received 11 October 2006
Abstract
Strains of Nitrobacter mediate the second step in the nitrification process by oxidizing nitrite to nitrate. Thephylogenetic diversity of the genus is currently not well investigated. In this study, a rep-PCR profile and the nearlycomplete 16S rRNA gene sequence of 30 strains, comprising a wide physiological as well as ecological diversity andencompassing representatives of the four species, were determined. The sequence diversity of the 16S rRNA genebetween different species was low, indicating the need for additional phylogenetic markers. Therefore, primers weredeveloped for amplifying the complete nxrX gene and a 380 bp fragment of the nxrB1 gene, which are both genesinvolved in the nitrite oxidation process. These genes confirmed the division into phylogenetic groups revealed by the16S rRNA gene but showed a better discriminatory power. They can be a valuable additional tool for phylogeneticanalysis within the genus Nitrobacter and can assist in the identification of new Nitrobacter isolates.r 2006 Elsevier GmbH. All rights reserved.
Keywords: Nitrobacter; Nitrite oxidoreductase; nxrA; nxrB; nxrX; 16S rRNA; Rep-PCR; Phylogenetic diversity
Introduction
Nitrification, the microbiological conversion of am-monia to nitrate, is a key process in the nitrogen cycle.The process consists of two subsequent steps: (i) theoxidation of ammonia to nitrite and (ii) the oxidation ofnitrite to nitrate. The latter reaction is performed by thenitrite oxidizing bacteria (NOB), which belong to fourphylogenetically unrelated genera: Nitrobacter (a-Pro-
teobacteria), Nitrococcus (g-Proteobacteria), Nitrospina
e front matter r 2006 Elsevier GmbH. All rights reserved.
apm.2006.11.006
ing author. Tel.: +329 264 51 01; fax: +32 9 264 53 46.
ess: [email protected] (B. Vanparys).
(preliminarily assigned to the d-Proteobacteria) andNitrospira (separate phylum; [11,40]).
Although Nitrospira is currently regarded as moreimportant than Nitrobacter in, for example, wastewatertreatment plants and aquaria, Nitrobacter has beenshown to coexist with Nitrospira in plants withtemporally or spatially elevated nitrite concentrations[9]. Furthermore, Nitrobacter is still considered animportant genus in fertilized as well as unfertilized soils[14], which are regarded as the privileged habitats ofNitrobacter [10,15]. Nitrobacter is also associated withlandfill leachate treatment plants [20] and rivers [8], andis important in the erosion of rocks [23] and thedeterioration of historic monuments [24].

ARTICLE IN PRESSB. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308298
Nitrobacter is a phylogenetically young group [30,35]and the genome is conserved within this genus [15]. Thegenus currently consists of four valid species, Nitrobac-
ter winogradskyi [44,48], Nitrobacter hamburgensis [6],Nitrobacter vulgaris [4] and Nitrobacter alkalicus [36].Previous studies indicated that the 16S rRNA gene haslimited discriminatory power within Nitrobacter [30,35]and might not be a suitable phylogenetic tool [2],although more data are necessary to confirm thisobservation. Sequence analysis of functional genes couldovercome this constraint, since protein coding genesshow more sequence diversity than the 16S rRNA gene.
Nitrite oxidation in Nitrobacter is mediated by thenitrite oxidoreductase enzyme (previously abbreviatedas NOR, but now called NXR [38]). The catalyticallyactive NXR enzyme of N. hamburgensis X14T consists oftwo subunits [25], encoded by nxrA and nxrB [21].Kirstein and Bock [21] showed that a nxrX gene,assumed to encode a peptidyl-prolyl cis–trans isomerasethat may assist in the folding of the nitrite oxidor-eductase enzyme [38], was located between nxrA andnxrB (further denominated as the nxrAXB gene cluster;Fig. 1). The NXR enzymes of the different Nitrobacter
species have serological homology [1,2] and recently thenxrAXB gene cluster was also shown to be present inrepresentatives of the other Nitrobacter species [38,42].A complete genome sequence analysis showed that N.
winogradskyi Nb-255T contains, besides the nxrA
(nxrA1) and nxrB (nxrB1) genes located in the nxrAXB
cluster, additional nxrA (nxrA2) and nxrB (nxrB2) genessegregated at distant points in the genome. The twocopies of nxrA (94%) and nxrB (97%) were onlymoderately related [38]. Currently, only a limitednumber of nxr gene sequences are publicly availableand no primers amplifying these genes have beendescribed in the literature. The value of the nxr genesin the phylogenetic analysis of Nitrobacter has hithertonot been evaluated.
In this study, a set of 30 Nitrobacter strains,comprising a wide physiological as well as ecologicaldiversity and encompassing representatives of the fourvalid species of the genus, was analysed. The nearlycomplete 16S rRNA gene sequence, a rep-PCR profileand, using newly developed primers, the complete nxrX
NxrA-1F
Nxr
NxrA1
bp 1
Fig. 1. Organization of the Nitrobacter nxrAXB gene cluster. Eac
indicated.
and a 380 bp fragment of the nxrB1 gene sequence of allstrains were determined.
Materials and methods
Strains and culture conditions
All strains used in this study are listed in Table 1.A frozen cell suspension of N. alkalicus AN2 was
obtained from D. Sorokin (Delft University of Technol-ogy, The Netherlands). An active liquid culture of N.
vulgaris DSM 10236T was obtained from DSMZ. Activecultures of N. winogradskyi ATCC 25391T and ATCC14123 and N. hamburgensis X14T were obtained from P.Bodelier (Netherlands Institute of Ecology, The Nether-lands). An active culture of Nitrospira moscoviensis
NCIMB 13793T was obtained from Zena Smith(NCIMB Ltd., UK). The other strains originated fromthe Institut fur Allgemeine Botanik, Abteilung Micro-biology, Universitat Hamburg (Germany), that pro-vided strains 339, 329, 219, 311 and 263 isolated by S.T.Watson. Strains Engel, 219, Nato I, F83KO, 339, GTOman, AB1, K48, K55 and 263 were cultivated in abasal mineral salt medium [11], supplemented with 2 g/lNaNO2. Strains 5F/3, LIP, 4111/1, BS5/19, AB3, R1.30,329, Termite 2, Yukatan, BB3, C2, D6/13 and Y werecultivated in a basal mineral salt medium [11] containing0.2 g/l NaNO2. Strain Io Acid was cultivated in an acidicmedium [34] at a pH of 5.5 and strain 311 in a marinemedium [45]. Cultures were grown at 28 1C andperiodically assayed for nitrite consumption. To obtaina sufficient amount of cells, cultures with 0.2 g/l NaNO2
were supplemented with nitrite up to eight times. Strainswere checked for contamination analogous to Steinmul-ler and Bock [39]. Cells were harvested by centrifuga-tion, suspended in 0.9% NaCl and stored at –20 1C uponDNA extraction.
DNA extraction
DNA was extracted by alkalic lyses. Cell suspensionswere centrifuged for 5min at 16,000g, whereupon
X-1F
NxrB-1F NxrB-1R
NxrX-1R
NxrX NxrB1
1 1 15423645 702
h arrow represents one gene. Primers used in this study are

ARTICLE IN PRESS
Table 1. Overview of the strains used in this study
Strain Origin Reference
Nitrobacter
N. alkalicus
AN2 Sediment soil, Siberia (Russia) [36]
N. hamburgensis
X14T Soil from botanical garden, Hamburg (Germany) [6]
Y Soil from Uxmal, Yucatan (Mexico) [6]
N. vulgaris
ZT¼ DSM 10236T Sand filter from waterworks, Hamburg (Germany) [4]
329 Soil, Yalta (Russia) [4]
339 Soil, Rhodos (Greece) [4]
AB1 Sewage system, Hamburg (Germany) [4]
BB3 Brackish water from river Elbe, Hamburg (Germany) [4]
C2 Salt water from river Elbe, Hamburg (Germany) [4]
K48 Sandstone from cathedral, Cologne (Germany) [4]
K55 Sandstone from cathedral, Cologne (Germany) [4]
N. winogradskyi
ATCC 25391T ¼ Nb-255T Soil [45]
ATCC 14123 ¼ AG ¼ 106 Soil [45]
219 Paramaribo river (Surinam) Watson, unpublished
311 Atlantic near coast of Africa Watson, unpublished
AB3 Sewage system, Hamburg (Germany) [7]
Engel Soil from botanical garden, Hamburg (Germany) [4]
F83KO Concrete from cooling tower, Cologne (Germany) [4]
Nato I Activated carbon filter, Hannover (Germany) [4]
R 1.30 Stone from cathedral, Regensburg (Germany) [4]
Nitrobacter sp.
263 ¼ ATCC 25393 Soil, Galapagos Islands (Ecuador) [22]
LIP Concrete, Lipetsk (Russia) [22]
4111/1 Concrete, Frankfurt (Germany) [22]
BS 5/19 Sulfidic ore mine, Baia Sprie (Romania) [22]
Gt Oman Oasis (Oman) [22]
Termite 2 Termite heap, Macheke (Zimbabwe) [22]
Yukatan Soil from Uxmal, Yucatan (Mexico) [22]
D6/13 Ore mine, Dubova (Romania) [22]
Io Acid Hoosier national forest (USA) [16]
5 F/3 Permafrost soil, Siberia (Russia) [3]
Nitrospira
N. moscoviensis
NCIMB 13793T Iron pipe, Moscow (Russia) [11]
B. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308 299
supernatant was removed, 20 ml of lysis buffer (0.5 ml10% SDS; 1 ml 1M NaOH; 18.5 ml MilliQ water) wasadded and the tube was placed at 95 1C for 15min.Subsequently, 180 ml MilliQ water was added, the tubewas centrifuged for 5min at 16,000g and the super-natant was transferred to a new tube. DNA extractswere stored at –20 1C until use.
16S rRNA gene sequencing
Amplification of both strands of the nearly complete(1495bp; positions 27–1522 in the Escherichia coli number-ing system) 16S rRNA gene sequence was performed asdescribed previously [18]. The PCR-amplified 16S rRNA
gene products were purified using the Nucleofasts 96 PCRclean up membrane system (Machery-Nagel, Germany).For each sequence reaction, a mixture was made using 1mlpurified product, 1ml of BigDyeTM Termination RR mixversion 3.1 (Perkin Elmer), 1.5ml of BigDyeTM buffer (5� ),1.5ml MilliQ and 3ml (20ngml�1) of one of the eightsequencing primers used (forward primer, position 339–358,50-tcctacgggaggcagcact-30; forward, 519–536, 50-cag-cagccgcggtaatac-30; forward, 908–926, 50-aactcaaaggaatt-gacgg-30; forward, 1093–1112, 50-agtcccgcaacgagcgcaac-30;reverse, 358–339, 50-actgctgcctcccgtaggag-30; reverse,536–519, 50-gtattaccgcggctgctg-30; reverse, 1112–1093, 50-gttgcgctcgttgcgggact-30 and reverse, 1241–1222, 50-gctaca-cacgtgctacaatg-30). The thermal programme consisted of 30cycles (15 s at 96 1C, 1 sec at 351C and 4min at 60 1C).

ARTICLE IN PRESSB. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308300
Sequencing products were purified and sequenced asdescribed previously [26]. Sequence assembly was performedwith BioNumerics version 4.5 (Applied Maths, Belgium). Aphylogenetic tree was constructed based on the neighbor-joining method.
Nxr primer design and gene sequencing
The organization of the nxrAXB cluster with indicationof the primers used in this study is represented in Fig. 1.Primers were designed using the four nxrA (AF344872-AF344875) and thirteen nxrB gene sequences (AY508477-AY508483; L76185-L76190; X66067) available in theEMBL database at the time using the software Kodonversion 2.0 (Applied-Maths, Belgium). PCR amplificationof the complete nxrX gene and the 50-end of the nxrB1 genewas performed using the primers nxrA-1F and nxrB-1R.NxrA-1F (50-gcatggatccggtgtggatca-30) is situated at posi-tion 3242–3262 of the nxrA sequence of N. hamburgensis
X14T (AF344872). NxrB-1R (50-ccgtgctgttgayctcgttga-30
[y ¼ c or t]) is situated at position 470–490 of the nxrB
sequence of N. hamburgensis X14T (L76186). Amplificationwas performed using a 25ml PCR mixture containing 2.5mlPCR buffer (10� ), 2.5ml dNTP’s (2mM each), 0.5mlforward primer (5mM), 0.5ml reversed primer (5mM), 0.5mlAmpliTaq DNA polymerase (1Uml�1), 17.5ml sterileMilliQ water and 1.0ml DNA extract (50mgml�1). Thethermal profile consisted of 10min at 95 1C, 30 cycles of1min at 95 1C, 1min at 55 1C and 2min at 72 1C, and a finalelongation step of 12min at 72 1C. The PCR products werepurified using the Nucleofasts 96 PCR clean up membranesystem. Sequencing reactions were performed analogous tothe 16S rRNA gene sequencing reactions, using the samemixture composition and thermal profile. The completenxrX gene was sequenced using the primers nxrX-1F (50-cgtcgtgcgcaagatggaga-30; position 3573–3592 of the nxrA
sequence of N. hamburgensis X14T AF344872; [42]) andnxrX-1R (50-cccggcttggtctccacgt-30; position 80–98 of thenxrB sequence of N. hamburgensis X14T L76186; [42]). In afew cases, the nxrX-1F primer generated sequences withambiguous positions at the 50-end. For these stains, theamplification primer nxrA-1F was additionally used as asequence primer. A 380bp fragment of the nxrB1 gene wassequenced using the primers nxrB-1F (50-acgtggagac-caagccggg-30; reverse complement of nxrX-1R) and nxrB-1R. Sequence assembly was performed with BioNumericsversion 4.5. Phylogenetic trees were constructed based onthe neighbor-joining method.
Rep-PCR
Rep-PCR genomic fingerprinting was performed withthe REP-, (GTG)5- and BOX-primers [43] using thePCR conditions described previously [33]. Electrophor-esis was performed as described by Heyrman et al. [17].
The patterns were digitalized, UPGMA clustering of thePearson correlation similarity values of the resultingband patterns was performed and cophenetic correlationvalues were calculated using BioNumerics v. 4.5.Cophenetic correlation is a parameter to express theconsistency of a cluster, which is similar to the bootstrapmethod for sequence clusters, with values above 80representing reliable clusters. The method calculates thecorrelation between the dendrogram-derived similaritiesand the matrix similarities.
Nucleotide sequence accession numbers
The 16S rRNA gene sequences have been deposited inthe GenBank/EMBL/DDBJ under accession numbersAM114522 and AM286374–AM286398. The nxrX genesequences have been deposited in the GenBank/EMBL/DDBJ under accession numbers AM114516–114520 andAM286349–AM286373. The nxrB1 gene sequences havebeen deposited in the GenBank/EMBL/DDBJ underaccession numbers AM286319–AM286348.
Results
16S rRNA gene sequence analysis
In total, 30 Nitrobacter strains were analysed in thisstudy. The strain list (Table 1) comprised members ofthe four valid species and strains unidentified at thespecies level or with a preliminary taxonomic position.They were obtained from a wide range of habitats andgeographical locations. A nearly complete 16S rRNAgene sequence from all strains was determined, and aphylogenetic clustering of these sequences together withreference sequences from the GenBank database isrepresented in Fig. 2. A 100% similarity was shownbetween the 16S rRNA gene sequence of N. winograds-
kyi ATCC 25391T ¼ Nb-255T and N. hamburgensis
X14T determined in this study and those from thecomplete genome sequences of the respective strains([38]; Copeland et al., unpublished). All sequences werehighly (498.8%) related to each other. Based onpairwise sequence similarities and by visual analysis ofthe dendrogram, five groups were delineated.
Group 1 contained six strains with 100% sequencesimilarity and included the type strain of N. winograds-
kyi (ATCC 25391T) and other strains of this species.Group 2 showed 99.9% sequence similarity to group 1and contained three sequences with 100% sequencesimilarity, including N. winogradskyi ATCC 14123(former type strain of ‘‘N. agilis’’). Group 3 showed99.9% sequence similarity to groups 1 and 2, andcontained the three sequenced members of N. alkalicus
and strain Termite 2, which shared 99.9–100% sequence

ARTICLE IN PRESS
1%
N. alkalicus AN2 (AF069957)*
N. alkalicus AN4 (AF069958)*
N. alkalicus AN1T (AF069956)*
Nitrobacter sp. Termite2 (AM286391)
N. winogradskyi ATCC 25391T (CP000115)*
N. winogradskyi Engel (AM286374)
N. winogradskyi F83KO (AM286377)
Nitrobacter sp. LIP (AM286385)
N. winogradskyi R1.30 (AM286389)
N. winogradskyi 311 (AM286397)
N. winogradskyi ATCC 14123 (L35506)*
N. winogradskyi Nato I (AM286376)
N. winogradskyi AB3 (AM286388)
N. vulgaris K48 (AM286381)
N. vulgaris K55 (AM286382)
N. vulgaris 329 (AM286390)
N. vulgaris 339 (AM286378)
N. vulgaris DSM10236T (AM114522)
Nitrobacter sp. GT Oman (AM286379)
N. vulgaris AB1 (AM286380)
Nitrobacter sp. 5F/3 (AM286384)
Nitrobacter sp. 4111/1 (AM286386)
Nitrobacter sp. Yukatan (AM286392)
N. vulgaris C2 (AM286393)
Nitrobacter sp. D6/13 (AM286394)
Nitrobacter sp. Io Acid (AM286395)
N. vulgaris BB3 (AM286398)
N. hamburgensis Y (AM286396)
Nitrobacter sp. 263 (AM286383)
N. winogradskyi 219 (AM286375)
Nitrobacter sp. BS5/19 (AM286387)
N. hamburgensis X14T (NC_007964)*
Rhodopseudomonas palustris DSM123T(AB175650)*
62
63
84
71
66
60
59
32
37
31
76
64
87
100
1
2
3
4
5
Fig. 2. Phylogenetic dendrogram based on neighbour-joining clustering after multiple alignment (1372 bp) of the 16S rRNA gene
sequences of the isolates analysed in this study and reference strains from GenBank (indicated with *). Bootstrap values (expressed
as percentages of 1000 replications) are shown at the branch points. Accession numbers are shown in parentheses.
Rhodopseudomonas palustris DSM123T was used as an outgroup. Bar, % nucleotide substitution.
B. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308 301
similarity to each other. Group 4 contained elevenstrains sharing 99.9–100% sequence similarity andincluded the type strain of N. vulgaris (DSM 10236T)
and several other strains identified as belonging to thisspecies by Bock et al. [4]. Group 4 showed 99.4–99.6%sequence similarity to groups 1–3. K48, K55 and 329,

ARTICLE IN PRESSB. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308302
also belonging to N. vulgaris according to Bock et al. [4],fell outside this group. Group 5 contained three strainswith intragroup sequence similarities lower than in theother groups (99.5–99.7%). Sequence similarities withstrains of other groups were between 98.9% and 99.6%.The two strains of N. hamburgensis shared only 98.8%sequence similarity and were interspersed with themembers of group 5.
nxrX and nxrB1 gene sequence analysis
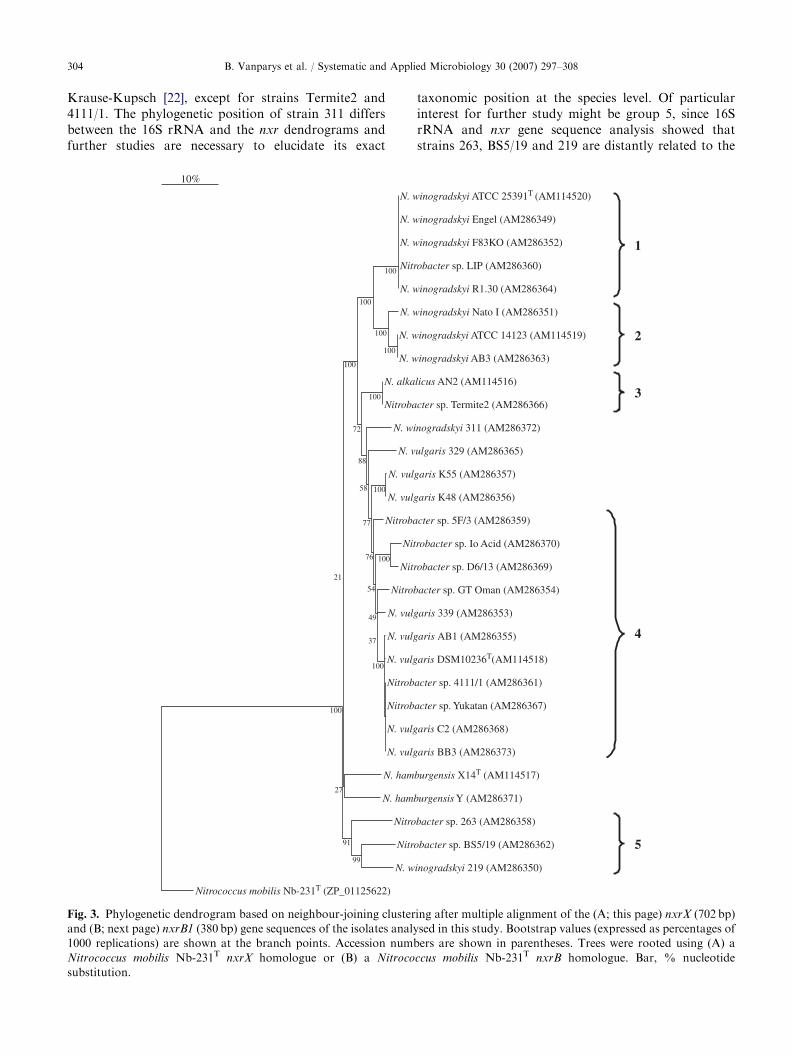
Unambiguous sequences for the complete nxrX gene(684–705 bp, depending on the strain) and a 380 bpfragment of the nxrB1 gene were obtained for all 30Nitrobacter strains, while no amplicon was obtained forthe representative of Nitrospira. Since 100% similaritywas found between the nxrX and nxrB1 gene sequencesdetermined in this study for N. winogradskyi ATCC25391T and those obtained by Starkenburg et al. [38],our amplification and sequence analysis protocol wasconsidered valid. As is common for protein codinggenes, the nucleotide sequence similarity between strainswas lower for the nxrX (ranging from 79.9% to 100%)and nxrB1 (88.5–100%) genes than for the 16S rRNAgene (95.8–100%).
The phylogenetic tree for the nxrX gene (Fig. 3A) was inagreement with the tree based on the corresponding aminoacid sequences (data not shown), although the relationshipbetween the representatives of N. hamburgensis and thoseof group 5 showed some difference. All groups delineatedin the 16S rRNA gene tree were retrieved in the nxrX
dendrograms. However, due to the lower sequencesimilarities and higher bootstrap values, the grouping wasconsidered to be more reliable. Alignment of the nxrX
amino acid sequences showed multiple strain- or group-dependent indels in the region from amino acid position 7to 19 (with N. winogradskyi ATCC 14123 used as areference). For example, a sequence of six amino acids waspresent in N. hamburgensis Y, while this sequence wasabsent in N. hamburgensis X14T. At amino acid position16, a histidine was present in all members of group 2 butwas absent in the members of group 1. Furthermore, atposition 216, an insert of one amino acid was present in therepresentatives of N. hamburgensis (threonine) and group 5(lysine for strain 263 and valine for strains BS5/19 and219), but it was absent in all other strains.
Although some minor variability was observed in therelatedness between the different groups, the phyloge-netic tree for the nxrB1 gene (Fig. 3B) was generally inagreement with the nxrX trees and the five groups couldbe separated from each other. On the contrary, aminoacid sequences of all strains were nearly identical to eachother, and therefore did not allow delineation of thedifferent groups. The alignment of the nxrB1 amino acidsequences showed no indels.
Rep-PCR
In this study, a banding profile of rep-PCR productsamplified with REP-, BOX- and (GTG)5-primers wasdetermined for all 30 strains. Banding profiles usingREP-primers showed too few bands to allow reliablegrouping (data not shown). An UPGMA clustering ofPearson’s correlation similarity coefficients for thecombined normalized BOX- and (GTG)5 bandingpatterns is represented in Fig. 4. The reproducibility ofthe technique was analysed by replicate analysis ofNitrobacter sp. 263 (i.e. two independent PCR amplifi-cation reactions and electrophoresis on different gels). A96% Pearson correlation similarity was shown betweenthe replicates. All strains of group 1 were located in acluster that could be reliably (cophenetic correlationvalue of 88%) separated from all other strains. Thestrains of group 2 grouped together, although N.
winogradskyi ATCC 14123 was more distantly located.However, no good banding profile could be obtained forthis strain, even after repeated attempts. Both strains ofgroup 3 clustered together, since they showed nearlyidentical banding patterns. All strains of group 4 werelocated in a cluster that could be reliably (copheneticcorrelation of 95%) separated from all other strains. Asin the 16S rRNA and nxr gene analyses, strains K48,K55 and 329 fell outside group 4. The members of N.
hamburgensis did not cluster separately from otherspecies. Furthermore, the three strains of group 5 werelocated at distantly related positions in the dendrogrambut showed low similarities compared with other strains.
Discussion
Although generally a good target for studyingphylogenetic relationships [41], the 16S rRNA genelacks discriminatory power in recently evolved taxa [13].Previous studies [30,35] concluded that the 16S rRNAgene showed a low diversity within the genus Nitrobac-
ter. These studies were, however, based on only threesequences and even to date the public databases containvery few 16S rRNA sequences of identified Nitrobacter
strains. Here, the nearly complete 16S rRNA genesequence of a set of 30 strains comprising a widephysiological as well as ecological diversity and encom-passing representatives of the four species was deter-mined. The results showed that the 16S rRNA gene wasindeed conserved within Nitrobacter. One misidentifiedbasepair during sequencing could hence result in acompletely different position of the strain in the 16SrRNA gene dendrogram. Although the species could beseparated from each other, there is a need to comple-ment the 16S rRNA gene sequence data with that ofother genes to obtain more reliable phylogeneticinformation. Therefore, primers were developed that

ARTICLE IN PRESSB. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308 303
targeted the nitrite oxidoreductase genes of Nitrobacter.Unambiguous sequences were obtained for the completenxrX gene and a 380 bp fragment of the nxrB1 genefrom all strains. The nxrX and nxrB1 dendrograms werein agreement with the 16S rRNA gene dendrogram butshowed a higher discriminatory power. Both nxrX andnxrB1 dendrograms were able to differentiate clearly theNitrobacter species from each other. Thus, sequencing ofthe nxrX and/or nxrB1 gene can be a valuable additionaltool for phylogenetic analysis within Nitrobacter andcan assist in the identification of new Nitrobacter
isolates.Previously, amplified ribosomal DNA restriction
analysis [15] and restriction fragment length polymorph-ism analysis [27,28] has been applied to Nitrobacter, butwithout representatives of N. alkalicus [15] or N.
alkalicus and N. vulgaris [27,28]. In this study, a rep-PCR profile was obtained for all 30 strains. The resultsconfirmed the observation of Navarro et al. [27] thatstrains from different geographical areas and/or ecolo-gical niches can be genotypically very similar. Forexample, N. vulgaris C2, isolated from German saltwater, and Nitrobacter sp. Yukatan, isolated fromMexican soil, showed 99.4% Pearson correlationsimilarity while the 16S rRNA and nxr genes of bothstrains were identical. Strains with very similar bandingpatterns showed identical 16S rRNA and nxr genesequences and the rep-PCR technique can hence beapplied as a fast initial method for dereplication of setsof Nitrobacter isolates.
A combination of the data from the rep-PCR bandingpatterns and the 16S rRNA, nxrX and nxrB1 genesequence analysis, and data from the literature providedinsights into the phylogenetic relationships betweendifferent Nitrobacter strains and species, which led tothe following conclusions.
(1) N. winogradskyi ATCC 14123 was originallydescribed as the only strain of a separate species, ‘‘N.
agilis’’ [29]. Later, it was stated that both species couldnot be phenotypically separated from each other[19,31,46] and that ‘‘N. agilis’’ should hence be asubjective synonym of N. winogradskyi [44]. Althoughfurther studies showed that ‘‘N. agilis’’ could bedistinguished from N. winogradskyi by immunofluores-cence [12], different growth characteristics in thepresence of organic matter [39] and different rRNAgene restriction patterns [27], ‘‘N. agilis’’ is currentlyconsidered as an invalid name. Based on DNA-DNAhybridization data, Navarro et al. [27] suggested that‘‘N. agilis’’ might be a subspecies of N. winogradskyi.This current study showed that N. winogradskyi ATCC25391T and N. winogradskyi ATCC 14123 have a 99.9%16S rRNA gene sequence similarity and belong todifferent groups. In the nxrX and nxrB1 dendrogram, N.
winogradskyi ATCC 14123 clustered into group 2 andcould be reliably separated (bootstrap values of 100%
and 68%, respectively) from group 1 containing N.
winogradskyi ATCC 25391T. Although phenotypicanalysis is necessary to determine whether both groupscan be considered as different species, 16S rRNA geneand nxr sequence analyses indicated additional genomicdifferences between N. winogradskyi ATCC 25391T andN. winogradskyi ATCC 14123 suggesting a separatetaxonomic position at the subspecies level.
(2) N. vulgaris was originally described as a newspecies containing 17 strains. DNA–DNA hybridizationstudies, however, showed DNA relatedness valuesbetween 55% and 97% [4], while 70% is consideredthe cut-off value for species delineation [47]. Eightstrains belonging to N. vulgaris were included in thisstudy. K48 and K55 had very similar rep-PCR patterns,and 16S rRNA and nxr gene sequences. Both strainsformed a cluster reliably separated from the clustercontaining the N. vulgaris type strain and from otherNitrobacter species. These data were in agreement withthe DNA–DNA hybridization values of Bock et al. [4]and indicated that K48 and K55 might belong to ahitherto undescribed Nitrobacter species or N. vulgaris
subspecies, although phenotypic data distinguishingthem from the other N. vulgaris strains are lacking atthe moment. Both strains were isolated from naturalstones of historical buildings and it was previouslyshown that this habitat is a good niche for the isolationof new lineages of nitrifiers [5,32]. N. vulgaris 329showed DNA relatedness values at the threshold valuefor species delineation compared with other strains of N.
vulgaris. Also, in rep-PCR, 16S rRNA and nxr genesequence analyses, 329 clustered outside the N. vulgaris
strains and could hence belong to a separate species or aN. vulgaris subspecies.
(3) N. hamburgensis was described based on twostrains [6], both of which were included in this study.While DNA-DNA hybridizations showed 100% DNArelatedness [6], some dissimilarity between both strainswas found with the rep-PCR profiles (87.7% similarity)as well as with nxrX (94.1%) and nxrB1 (87.0%) genesequence analysis. Further study is necessary to eluci-date the phylogenetic relationship between both strains.
(4) Besides a selection of strains previously assigned tothe four currently known species of Nitrobacter, thisstudy also included strains unidentified at the specieslevel or with a preliminary taxonomic position deducedfrom protein pattern analysis [22]. The combination ofrep-PCR, 16S rRNA and nxr gene sequences allowedpresumptive allocation at the species level. Strain LIPwas highly related to members of N. winogradskyi, whilestrain Termite 2 most probably belongs to N. alkalicus.Stains 5F/3, Gt Oman, Io Acid, D6/13, 4111/1 andYukatan were provisionally assigned to N. vulgaris. Ourresults counter the conclusion of Seewaldt et al. [35] thatstrain Yukatan can be treated as belonging to N.
winogradskyi and were generally in agreement with
ARTICLE IN PRESSB. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308304
Krause-Kupsch [22], except for strains Termite2 and4111/1. The phylogenetic position of strain 311 differsbetween the 16S rRNA and the nxr dendrograms andfurther studies are necessary to elucidate its exact
10%
N. w
N. w
N. w
Nitr
N. w
N. w
N. w
N. w
N. alka
Nitroba
N. wi
N. v
N. vul
N. vulg
Nitroba
Nit
Nitr
Nitrob
N. vulg
N. vulg
N. vulg
Nitrob
Nitrob
N. vulg
N. vulg
N. ham
N. hamb
Nitro
Nitr
N. w
Nitrococcus mobilis Nb-231T (ZP_01125622)
100
100
100
100
100
100
100
100
37
49
54
76
77
58
88
72
100
27
21
99
91
100
Fig. 3. Phylogenetic dendrogram based on neighbour-joining cluster
and (B; next page) nxrB1 (380 bp) gene sequences of the isolates analy
1000 replications) are shown at the branch points. Accession numb
Nitrococcus mobilis Nb-231T nxrX homologue or (B) a Nitroco
substitution.
taxonomic position at the species level. Of particularinterest for further study might be group 5, since 16SrRNA and nxr gene sequence analysis showed thatstrains 263, BS5/19 and 219 are distantly related to the
inogradskyi ATCC 25391T (AM114520)
inogradskyi Engel (AM286349)
inogradskyi F83KO (AM286352)
obacter sp. LIP (AM286360)
inogradskyi R1.30 (AM286364)
inogradskyi Nato I (AM286351)
inogradskyi ATCC 14123 (AM114519)
inogradskyi AB3 (AM286363)
licus AN2 (AM114516)
cter sp. Termite2 (AM286366)
nogradskyi 311 (AM286372)
ulgaris 329 (AM286365)
garis K55 (AM286357)
aris K48 (AM286356)
cter sp. 5F/3 (AM286359)
robacter sp. Io Acid (AM286370)
obacter sp. D6/13 (AM286369)
acter sp. GT Oman (AM286354)
aris 339 (AM286353)
aris AB1 (AM286355)
aris DSM10236T(AM114518)
acter sp. 4111/1 (AM286361)
acter sp. Yukatan (AM286367)
aris C2 (AM286368)
aris BB3 (AM286373)
burgensis X14T (AM114517)
urgensis Y (AM286371)
bacter sp. 263 (AM286358)
obacter sp. BS5/19 (AM286362)
inogradskyi 219 (AM286350)
1
2
3
4
5
ing after multiple alignment of the (A; this page) nxrX (702 bp)
sed in this study. Bootstrap values (expressed as percentages of
ers are shown in parentheses. Trees were rooted using (A) a
ccus mobilis Nb-231T nxrB homologue. Bar, % nucleotide

ARTICLE IN PRESS
10%
N. winogradskyi ATCC 25391T (AM286323)
N. winogradskyi Engel (AM286324)
N. winogradskyi F83KO (AM286327)
N. winogradskyi R1.30 (AM286339)
Nitrobacter sp. LIP (AM286335)
N. winogradskyi Nato I (AM286326)
N. winogradskyi ATCC 14123 (AM286322)
N. winogradskyi AB3 (AM286338)
N. winogradskyi 311 (AM286347)
Nitrobacter sp. Termite2 (AM286341)
N. alkalicus AN2 (AM286319)
N. vulgaris 329 (AM286340)
N. vulgaris K48 (AM286331)
N. vulgaris K55 (AM286332)
Nitrobacter sp. 5F/3 (AM286334)
Nitrobacter sp. GT Oman (AM286329)
Nitrobacter sp. Io Acid (AM286345)
N. vulgaris 339 (AM286328)
Nitrobacter sp. D6/13 (AM286344)
N. vulgaris DSM10236T(AM286321)
N. vulgaris AB1 (AM286330)
Nitrobacter sp. 4111/1 (AM286336)
Nitrobacter sp. Yukatan (AM286342)
N. vulgaris C2 (AM286343)
N. vulgaris BB3 (AM286348)
N. hamburgensis X14T (AM286320)
N. hamburgensis Y (AM286346)
Nitrobacter sp. 263 (AM286333)
N. winogradskyi 219 (AM286325)
Nitrobacter sp. BS5/19 (AM286337)
Nitrococcus mobilis Nb-231T(ZP_01125873)
100
97
100
68
48
100
100
86
51
39
84
73
85
100
100
88
43
88
81
72
78
61
48
100
1
2
3
4
5
Fig. 3. (Continued)
B. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308 305
strains of all currently known Nitrobacter species, andthey might hence belong to another species. In contrastto Watson (unpublished), our results indicate that 219does not belong to N. winogradskyi.
Immunological analysis showed that the nitriteoxidizing system (NOS) from Nitrospira and Nitrospina
differs from the NXR enzyme of Nitrobacter [2,37]. Nosequence information for the NOS of Nitrospira and
Nitrospina is currently available. The nxr primers werehence solely based on Nitrobacter sequences and noamplicon was obtained for Nitrospira moscoviensis
NCIMB 13793T. During the course of this research, acomplete genome sequence of Nitrococcus mobilis Nb-231T was deposited in GenBank. The genome did notshow the nxrAXB gene cluster found in Nitrobacter andthe nxrA and nxrB genes of N. mobilis Nb-231 and N.

ARTICLE IN PRESS
100
97
100
89
89
87
92
97
99
98
100
99
96
95
95
83
83
100
100
85
90
100
69
92
82
88
76
77
(GTG)5 Box
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
N. vulgaris K48
N. vulgaris K55
N. hamburgensis X14T
N. hamburgensis Y
Nitrobacter sp. 311
N. vulgaris 329
Nitrobacter sp. 219
Nitrobacter sp. Yukatan
N. vulgaris BB3
N. vulgaris C2
Nitrobacter sp. 4111/1
N. vulgaris AB1
N. vulgaris DSM 10236T
N. vulgaris 339
Nitrobacter sp. D6/13
Nitrobacter sp. Io Acid
Nitrobacter sp. GT Oman
Nitrobacter sp. 5F/3
Nitrobacter sp. 263
N. winogradskyi Nato I
Nitrobacter sp. AB3
Nitrobacter sp. Termite2
N. alkalicus AN2
N. winogradskyi ATCC 14123
N. winogradskyi Engel
N. winogradskyi F83KO
N. winogradskyi R1.30
Nitrobacter sp. LIP
N. winogradskyi ATCC25391T
Nitrobacter sp. BS5/19
% Pearson correlation
1
2
3
4
5
2
5
5
100
80 85 90 95 100
Fig. 4. Grouping of combined normalized BOX- and (GTG)5-patterns of the isolates in a dendrogram based on UPGMA clustering
of Pearson’s correlation similarity coefficients. Cophenetic correlation (i.e. a parameter to express the consistency of a cluster) values
are shown at the branch points.
B. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308306
winogradsky Nb-255T showed, respectively, a related-ness of 69.3% and 69.0%. When more complete genomesequences of NOB become available, it will be clearerwhether primers amplifying the NOS of all currentlyknown genera of NOB can be developed.
Conclusions
In conclusion, the data in this study gave insights intothe phylogenetic diversity and relationships in Nitro-
bacter and indicated that taxonomic re-evaluations
might be necessary. At the moment, the phenotypicdifferences between different Nitrobacter species andstrains are not well investigated. A thorough knowledgeof the phylogenetic relatedness between strains may leadto a well-founded choice of representative isolates forcomparative phenotypic studies of this environmentallyimportant group. Furthermore, the newly developed nxr
primers were able to amplify successfully a large varietyof Nitrobacter strains. Nxr gene-based dendrogramswere largely in agreement with the 16S rRNA gene tree,but showed a better discriminatory power. Theseprimers could potentially be suitable tools for the

ARTICLE IN PRESSB. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308 307
culture independent monitoring of Nitrobacter inenvironmental samples.
Acknowledgements
This work was supported by Project Grant G.O.A.1205073 (2003-2008) from the ‘Ministerie van deVlaamse Gemeenschap, Bestuur Wetenschappelijk On-derzoek’ (Belgium), the FWO project G20156.02 and aPh.D. Grant (no. 41428) from the Institute forthe Promotion of Innovation through Science andTechnology in Flanders (IWT-Vlaanderen). We thankDr. D. Sorokin (Delft University of Technology, TheNetherlands), Dr. P. Bodelier (Netherlands Institute ofEcology, The Netherlands) and Zena Smith (NCIMBLtd., UK) for supply of biological material.
References
[1] J. Aamand, T. Ahl, E. Spieck, Monoclonal antibodies
recognizing nitrite oxidoreductase of Nitrobacter hambur-
gensis, N. winogradskyi, and N. vulgaris, Appl. Environ.
Microbiol. 62 (2000) 2352–2355.
[2] S. Bartosch, I. Wolgast, E. Spieck, E. Bock, Identification
of nitrite-oxidizing bacteria with monoclonal antibodies
recognizing the nitrite oxidoreductase, Appl. Environ.
Microbiol. 65 (1999) 4126–4133.
[3] S. Bartosch, C. Hartwig, E. Spieck, E. Bock, Immunolo-
gical detection of Nitrospira-like bacteria in various soils,
Microbiol. Ecol. 43 (2002) 26–33.
[4] E. Bock, H.P. Koops, U.C. Moller, M. Rudert, A new
facultative nitrite oxidizing bacterium Nitrobacter vulgaris
sp. nov, Arch. Microbiol 153 (1990) 105–110.
[5] E. Bock, W. Sand, The microbiology of masonry
biodeterioration, J. Appl. Bacteriol. 74 (1993) 503–514.
[6] E. Bock, H. Sundermeyer-Klinger, E. Stackebrandt, New
facultative lithoautotrophic nitrite-oxidizing bacteria,
Arch. Microbiol. 136 (1983) 281–284.
[7] E. Bock, P.A. Wilderer, A. Freitag, Growth of Nitrobac-
ter in the absence of dissolved oxygen, Water Res. 22
(1988) 245–250.
[8] A. Cebron, J. Garnier, Nitrobacter and Nitrospira genera
as representatives of nitrite-oxidizing bacteria: detection,
quantification and growth along the lower Seine River
(France), Water Res. 39 (2005) 4979–4992.
[9] H. Daims, J.L. Nielsen, P.H. Nielsen, K.H. Schleifer, M.
Wagner, In situ characterization of Nitrospira-like nitrite-
oxidizing bacteria active in wastewater treatment plants,
Appl. Environ. Microbiol. 67 (2001) 5273–5284.
[10] V. Degrange, R. Lensi, R. Bardin, Activity, size and
structure of a Nitrobacter community as affected by
organic carbon and nitrite in sterile soil, FEMS Micro-
biol. Ecol. 24 (1998) 173–180.
[11] S. Ehrich, D. Behrens, E. Lebedeva, W. Ludwig, E. Bock,
A new obligately chemolithoautotrophic, nitrite-oxidizing
bacterium, Nitrospira moscoviensis sp. nov. and its
phylogenetic relationship, Arch. Microbiol. 164 (1995)
16–23.
[12] C.B. Fliermans, B.B. Bohlool, E.L. Schmidt, Autecologi-
cal study of the chemoautotroph Nitrobacter by immuno-
fluorescence, Appl. Microbiol. 27 (1974) 124–129.
[13] G.E. Fox, J.D. Wisotzkey, P. Jurtshuk, How close is
close: 16S rRNA sequence identity may not be sufficient
to guarantee species identity, Int. J. Syst. Bacteriol. 42
(1992) 166–170.
[14] T.E. Freitag, L. Chang, C.D. Clegg, J.I. Prosser, Influence
of inorganic nitrogen management regime on the diversity
of nitrite-oxidizing bacteria in agricultural grassland soils,
Appl. Environ. Microbiol. 71 (2005) 8323–8334.
[15] G.L. Grundman, P. Normand, Microscale diversity of the
genus Nitrobacter in soil on the basis of analysis of genes
encoding rRNA, Appl. Environ. Microbiol. 66 (2000)
4543–4546.
[16] T.R. Hankinson, E.L. Schmidt, An acidophilic and a
neutrophilic Nitrobacter strain isolated from the numeri-
cally predominant nitrite-oxidizing population of an
acid forest soil, Appl. Environ. Microbiol. 54 (1988)
1536–1540.
[17] J. Heyrman, N.A. Logan, H.-J. Busse, A. Balcaen, L.
Lebbe, M. Rodriguez-Diaz, J. Swings, P. De Vos,
Virgibacillus carmonensis sp. nov., Virgibacillus necropolis
sp. nov. and Virgibacillus picturae sp. nov., three novel
species isolated from deteriorated mural paintings,
transfer of the species of the genus Salibacillus to
Virgibacillus, as Virgibacillus marismortui comb. nov.
and Virgibacillus salexigens comb. nov., and emended
description of the genus Virgibacillus, Int. J. Syst. Evol.
Microbiol. 53 (2003) 501–511.
[18] J. Heyrman, J. Swings, 16S rDNA sequence analysis of
bacterial isolates from biodeteriorated mural paintings in
the Servilia Tomb (necropolis of Carmona, Seville,
Spain), Syst. Appl. Microbiol. 24 (2001) 417–422.
[19] H. Kalthoff, S. Fehr, H. Sundemeyer, L. Renwrantz, E.
Bock, A comparison by means of antisera and lectins of
surface structures of Nitrobacter winogradskyi and N.
agilis, Curr. Microbiol. 2 (1979) 375–380.
[20] D.J. Kim, S.H. Kim, Effect of nitrite concentration on the
distribution and competition of nitrite-oxidizing bacteria
in nitratation reactor systems and their kinetic character-
istics, Water Res. 40 (2006) 887–894.
[21] K. Kirstein, E. Bock, Close genetic relationship between
Nitrobacter hamburgensis nitrite oxidoreductase and E.
coli nitrate reductases, Arch. Microbiol. 160 (1993)
447–453.
[22] T. Krause-Kupsch, Entwicklung einer Schnellmethode
zur Identifizierung und Klassifizierung nitritoxidierender
Bakterien. Dissertation Universitat Hamburg, 1993.
[23] E.V. Lebedeva, N.N. Lialikova, IuIu Bugel’skii, Partici-
pation of nitrifying bacteria in the disintegration of
serpentinous ultrabasic rock, Mikrobiologiia 47 (1978)
1101–1107.
[24] R. Mansch, E. Bock, Biodeterioration of natural stone
with special reference to nitrifying bacteria, Biodegrada-
tion 9 (1998) 47–64.
[25] M. Meincke, E. Bock, D. Kastrau, P.M.H. Kroneck,
Nitrite oxidoreductase from Nitrobacter hamburgensis:
redox centrers and their catalytic role, Arch. Microbiol.
158 (1992) 127–131.

ARTICLE IN PRESSB. Vanparys et al. / Systematic and Applied Microbiology 30 (2007) 297–308308
[26] S. Naser, F.L. Thompson, B. Hoste, D. Gevers, K.
Vandemeulebroecke, I. Cleenwerck, C.C. Thompson, M.
Vancanneyt, J. Swings, Phylogeny and identification of
enterococci by atpA gene sequence analysis, J. Clin.
Microbiol. 43 (2005) 2224–2230.
[27] E. Navarro, M.P. Fernandez, F. Grimont, A. Clays-
Josserand, R. Bardin, Genomic heterogeneity of the genus
Nitrobacter, Int. J. Syst. Bacteriol. 42 (1992) 554–560.
[28] E. Navarro, P. Simonet, P. Normand, R. Bardin,
Characterization of natural populations of Nitrobacter
spp. using PCR/RFLP analysis of the ribosomal inter-
genic spacer, Arch. Microbiol. 157 (1992) 107–115.
[29] D.H. Nelson, Isolation and characterization of Nitrosomo-
nas and Nitrobacter, Zbl. Bakt. II. Abt. 83 (1931) 280–311.
[30] S. Orso, M. Gouy, E. Navarro, P. Normand, Molecular
phylogenetic analysis of Nitrobacter spp, Int. J. Syst.
Bacteriol. 44 (1994) 83–86.
[31] P.H.C. Pan, Lack of distinction between Nitrobacter agilis
and Nitrobacter winogradskyi, J. Bacteriol. 108 (1971)
1416–1418.
[32] U. Purkhold, M. Wagner, G. Timmermann, A. Pommer-
ening-Roser, H.-P. Koops, 16S rRNA and amoA-based
phylogeny of 12 novel betaproteobacterial ammonia-
oxidizing isolates: extension of the dataset and proposal
of a new lineage within the nitrosomonads, Int. J. Syst.
Evol. Microbiol. 53 (2003) 1485–1494.
[33] J.L.W. Rademaker, F.J. de Bruijn, Characterization and
classification of microbes by rep-PCR genomic finger-
printing and computer assisted pattern analysis, in: G.
Gaetano-Anolles, P.M. Gresshoff (Eds.), DNA Markers:
Protocols, Applications and Overviews, Wiley, New
York, 1997, pp. 151–171.
[34] E.L. Schmidt, L.W. Belser, Nitrifying bacteria, in: R.H.
Miller, D.R. Keeney (Eds.), Methods of Soil Analysis
Properties, American Society of Agronomy, Madison, 1982.
[35] E. Seewaldt, K.H. Schleifer, E. Bock, E. Stackebrandt,
The close phylogenetic relationship of Nitrobacter and
Rhodopseudomonas palustris, Acta Microbiol. 131 (1982)
287–290.
[36] D.Y. Sorokin, G. Muyzer, T. Brinkhoff, J.G. Kuenen,
M.S.M. Jetten, Isolation and characterization of a novel
facultatively alkaliphilic Nitrobacter species, N. alkalicus
sp. nov, Arch. Microbiol. 170 (1998) 345–352.
[37] E. Spieck, S. Ehrich, J. Aamand, E. Bock, Isolation and
immunocytochemical location of the nitrite-oxidizing
system in Nitrospira moscoviensis, Arch. Microbiol. 169
(1998) 225–230.
[38] S.R. Starkenburg, P.S.G. Chain, L.A. Sayavedra-Soto, L.
Hauser, M.L. Land, F.W. Larimer, S.A. Malfatti, M.G.
Klotz, et al., Genome sequence of the chemolithoauto-
trophic nitrite-oxidizing bacterium Nitrobacter winograds-
kyi Nb-255,, Appl. Environ. Microbiol. 72 (2006)
2050–2063.
[39] W. Steinmuller, E. Bock, Growth of Nitrobacter in the
presence of organic matter. I. Mixotrophic growth, Arch.
Microbiol. 108 (1976) 299–304.
[40] A. Teske, E. Alm, J.M. Regan, S. Toze, B.E. Rittmann,
D.A. Stahl, Evolutionary relationships among ammonia-
and nitrite-oxidizing bacteria, J. Bacteriol. 176 (1994)
6623–6630.
[41] P. Vandamme, B. Pot, M. Gillis, P. De Vos, K. Kersters,
J. Swings, Polyphasic taxonomy, a consensus approach to
bacterial systematics, Microbiol. Rev. 60 (1996) 407–438.
[42] B. Vanparys, P. Bodelier, P. De Vos, Validation of the
correct start codon of nxrX/nxrX and universality of the
nxrAXB/nxrAXB gene cluster in Nitrobacter species,
Curr. Microbiol. 53 (2006) 255–257.
[43] J. Versalovic, M. Schneider, F.J. de Bruijn, J.R. Lupksi,
Genomic fingerprinting of bacteria using repetitive
sequence-based polymerase chain reaction, Methods
Mol. Cell. Biol. 5 (1994) 25–40.
[44] S.W. Watson, Taxonomic considerations of the family
Nitrobacteraceae Buchanan, Int. J. Syst. Bacteriol. 21
(1971) 254–270.
[45] S.W. Watson, E. Bock, F.W. Valois, J.B. Waterbury, U.
Schlosser, Nitrospira marina gen. nov. sp. nov.: a
chemolithotrophic nitrite-oxidizing bacterium, Arch. Mi-
crobiol. 144 (1986) 1–7.
[46] S.W. Watson, M. Mandel, Comparison of the morphol-
ogy and deoxyribonucleic acid composition of 27 strains
of nitrifying bacteria, J. Bacteriol. 107 (1971) 563–569.
[47] L.G. Wayne, D.J. Brenner, R.R. Colwell, P.A.D.
Grimont, O. Kandler, M.I. Krichevsky, L.H. Moore,
W.E.C. Moore, R.G.E. Murray, E. Stackebrandt, M.P.
Starr, H.G. Truper, Report of the ad hoc committee on
reconciliation of approaches to bacterial systematics, Int.
J. Syst. Bacteriol. 37 (1987) 463–464.
[48] S. Winogradsky, Contributions a la morphologie des
organismes de la nitrifications, Arch. Sci. Biol. 1 (1982)
86–137.
Related Documents











