BIOLOGY OF REPRODUCTION (2014) 90(4):85, 1–11 Published online before print 5 March 2014. DOI 10.1095/biolreprod.113.117077 Minireview The Ovarian Reserve of Primordial Follicles and the Dynamic Reserve of Antral Growing Follicles: What Is the Link? 1 Danielle Monniaux, 2,3,4,5,6 Fre ´de ´rique Cle ´ment, 7 Rozenn Dalbie `s-Tran, 3,4,5,6 Anthony Estienne, 3,4,5,6 Ste ´phane Fabre, 3,4,5,6 Camille Mansanet, 3,4,5,6 and Philippe Monget 3,4,5,6 3 Institut National de la Recherche Agronomique (INRA), UMR85 Physiologie de la Reproduction et des Comportements, Nouzilly, France 4 Centre National de la Recherche Scientifique (CNRS), UMR7247, Nouzilly, France 5 Universite ´ Franc ¸ois Rabelais de Tours, Tours, France 6 Institut Franc ¸ais du Cheval et de l’Equitation (IFCE), Nouzilly, France 7 Institut National de Recherche en Informatique et en Automatique (INRIA), Paris-Rocquencourt Research Centre, Le Chesnay, France ABSTRACT The growing follicles develop from a reserve of primordial follicles constituted early in life. From this pre-established reserve, a second ovarian reserve is formed, which consists of gonadotropin-responsive small antral growing follicles and is a dynamic reserve for ovulation. Its size, evaluated by direct antral follicular count or endocrine markers, determines the success of assisted reproductive technologies in humans and embryo production biotechnologies in animals. Strong evidence indi- cates that these two reserves are functionally related. The size of both reserves appears to be highly variable between individuals of similar age, but the equilibrium size of the dynamic reserve in adults seems to be specific to each individual. The dynamics of both follicular reserves appears to result from the fine tuning of regulations involving two main pathways, the phosphatase and tensin homolog (PTEN)/phosphatidylinositol-3 kinase (PI3K)/3- phosphoinositide-dependent protein kinase-1 (PDPK1)/v-akt murine thymoma viral oncogene homolog 1 (AKT1) and the bone morphogenetic protein (BMP)/anti-Mu ¨ llerian hormone (AMH)/SMAD signaling pathways. Mutations in genes encoding the ligands, receptors, or signaling effectors of these pathways can accelerate or modulate the exhaustion rate of the ovarian reserves, causing premature ovarian insufficiency (POI) or increase in reproductive longevity, respectively. With female aging, the decline in primordial follicle numbers parallels the decrease in the size of the dynamic reserve of small antral follicles and the deterioration of oocyte quality. Recent progress in our knowledge of signaling pathways and their environmental and hormonal control during adult and fetal life opens new perspectives to improve the management of the ovarian reserves. aging, anti-Mu ¨llerian hormone, follicular development, ovary, premature ovarian failure INTRODUCTION Antral follicle count (AFC) has been the subject of increasing interest during the last decade in humans and large mammals. From the improvement of ovarian ultrasonography and the validation of new endocrine markers, it is now possible to evaluate the number of antral follicles larger than 1 mm in diameter present in the ovaries of women and large domestic or wild species [1, 2]. Presently, this evaluation is a routine practice of primary importance to estimating the ovarian activity of individual females for clinical or breeding applications. AFC helps the clinicians to establish a diagnosis for female infertility, and it participates in predicting the reproductive capacity of both human and animal species [3, 4]. AFC can also predict the ovarian response of an individual to gonadotropin-based stimulatory treatments, hence the success of assisted reproductive technologies in humans and embryo production biotechnologies in farm and wild animals [5, 6]. In all species, the antrum cavity is formed when the growing follicle reaches a diameter between 200 and 300 lm. The antrum grows by accumulating fluid derived from blood flowing through the thecal capillaries and secretion products of follicular cells; some of them, such as hyaluronan and proteoglycans, generate an osmotic gradient that participates in antrum growth [7]. In humans and large mammals, the follicles grow slowly after antrum formation up to the stage when they become gonadotropin dependent and enter a phase of rapid terminal development, occurring in a wave-like pattern [8–10]. Gonadotropin dependence is acquired at a given follicular diameter, characteristic of each species (e.g., 200 lm in rodents, 1 mm in pig, 2 mm in sheep, 3–5 mm in human and cow, and about 10 mm in horse [11]). Below this diameter, the small antral follicles constitute a pool of gonadotropin- responsive follicles, which is the reserve for ovulation and ovarian biotechnologies. This is a dynamic reserve, since it is 1 Supported by the INRA Prediction of Ovulation (PREDICTOV) project, by the Regulation of the Gonadotrope Axis (REGATE) INRIA/INRA Large Scale Initiative Action, by the Agence Nationale pour la Recherche (ANR-2010-BLAN-1608-01) and by French fellowships from the Re ´gion Centre and INRA to A.E. and C.M. 2 Correspondence: Danielle Monniaux, Physiologie de la Reproduction et des Comportements, Centre INRA Val-de-Loire, 37380 Nouzilly, France. E-mail: [email protected] Received: 20 December 2013. First decision: 30 January 2014. Accepted: 19 February 2014. Ó 2014 by the Society for the Study of Reproduction, Inc. eISSN: 1529-7268 http://www.biolreprod.org ISSN: 0006-3363 1 Article 85 Downloaded from www.biolreprod.org.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

BIOLOGY OF REPRODUCTION (2014) 90(4):85, 1–11Published online before print 5 March 2014.DOI 10.1095/biolreprod.113.117077
Minireview
The Ovarian Reserve of Primordial Follicles and the Dynamic Reserve of AntralGrowing Follicles: What Is the Link?1
Danielle Monniaux,2,3,4,5,6 Frederique Clement,7 Rozenn Dalbies-Tran,3,4,5,6 Anthony Estienne,3,4,5,6
Stephane Fabre,3,4,5,6 Camille Mansanet,3,4,5,6 and Philippe Monget3,4,5,6
3Institut National de la Recherche Agronomique (INRA), UMR85 Physiologie de la Reproduction et desComportements, Nouzilly, France4Centre National de la Recherche Scientifique (CNRS), UMR7247, Nouzilly, France5Universite Francois Rabelais de Tours, Tours, France6Institut Francais du Cheval et de l’Equitation (IFCE), Nouzilly, France7Institut National de Recherche en Informatique et en Automatique (INRIA), Paris-Rocquencourt Research Centre,Le Chesnay, France
ABSTRACT
The growing follicles develop from a reserve of primordialfollicles constituted early in life. From this pre-establishedreserve, a second ovarian reserve is formed, which consists ofgonadotropin-responsive small antral growing follicles and is adynamic reserve for ovulation. Its size, evaluated by direct antralfollicular count or endocrine markers, determines the success ofassisted reproductive technologies in humans and embryoproduction biotechnologies in animals. Strong evidence indi-cates that these two reserves are functionally related. The size ofboth reserves appears to be highly variable between individualsof similar age, but the equilibrium size of the dynamic reserve inadults seems to be specific to each individual. The dynamics ofboth follicular reserves appears to result from the fine tuning ofregulations involving two main pathways, the phosphatase andtensin homolog (PTEN)/phosphatidylinositol-3 kinase (PI3K)/3-phosphoinositide-dependent protein kinase-1 (PDPK1)/v-aktmurine thymoma viral oncogene homolog 1 (AKT1) and thebone morphogenetic protein (BMP)/anti-Mullerian hormone(AMH)/SMAD signaling pathways. Mutations in genes encodingthe ligands, receptors, or signaling effectors of these pathwayscan accelerate or modulate the exhaustion rate of the ovarianreserves, causing premature ovarian insufficiency (POI) orincrease in reproductive longevity, respectively. With femaleaging, the decline in primordial follicle numbers parallels thedecrease in the size of the dynamic reserve of small antralfollicles and the deterioration of oocyte quality. Recent progress
in our knowledge of signaling pathways and their environmentaland hormonal control during adult and fetal life opens newperspectives to improve the management of the ovarianreserves.
aging, anti-Mullerian hormone, follicular development, ovary,premature ovarian failure
INTRODUCTION
Antral follicle count (AFC) has been the subject ofincreasing interest during the last decade in humans and largemammals. From the improvement of ovarian ultrasonographyand the validation of new endocrine markers, it is now possibleto evaluate the number of antral follicles larger than 1 mm indiameter present in the ovaries of women and large domestic orwild species [1, 2]. Presently, this evaluation is a routinepractice of primary importance to estimating the ovarianactivity of individual females for clinical or breedingapplications. AFC helps the clinicians to establish a diagnosisfor female infertility, and it participates in predicting thereproductive capacity of both human and animal species [3, 4].AFC can also predict the ovarian response of an individual togonadotropin-based stimulatory treatments, hence the successof assisted reproductive technologies in humans and embryoproduction biotechnologies in farm and wild animals [5, 6].
In all species, the antrum cavity is formed when the growingfollicle reaches a diameter between 200 and 300 lm. Theantrum grows by accumulating fluid derived from bloodflowing through the thecal capillaries and secretion products offollicular cells; some of them, such as hyaluronan andproteoglycans, generate an osmotic gradient that participatesin antrum growth [7]. In humans and large mammals, thefollicles grow slowly after antrum formation up to the stagewhen they become gonadotropin dependent and enter a phaseof rapid terminal development, occurring in a wave-like pattern[8–10]. Gonadotropin dependence is acquired at a givenfollicular diameter, characteristic of each species (e.g., 200 lmin rodents, 1 mm in pig, 2 mm in sheep, 3–5 mm in human andcow, and about 10 mm in horse [11]). Below this diameter, thesmall antral follicles constitute a pool of gonadotropin-responsive follicles, which is the reserve for ovulation andovarian biotechnologies. This is a dynamic reserve, since it is
1Supported by the INRA Prediction of Ovulation (PREDICTOV) project,by the Regulation of the Gonadotrope Axis (REGATE) INRIA/INRALarge Scale Initiative Action, by the Agence Nationale pour laRecherche (ANR-2010-BLAN-1608-01) and by French fellowships fromthe Region Centre and INRA to A.E. and C.M.2Correspondence: Danielle Monniaux, Physiologie de la Reproductionet des Comportements, Centre INRA Val-de-Loire, 37380 Nouzilly,France. E-mail: [email protected]
Received: 20 December 2013.First decision: 30 January 2014.Accepted: 19 February 2014.� 2014 by the Society for the Study of Reproduction, Inc.eISSN: 1529-7268 http://www.biolreprod.orgISSN: 0006-3363
1 Article 85
Dow
nloaded from w
ww
.biolreprod.org.
emptied by the cyclic follicle-stimulating hormone (FSH)-orchestrated entry of follicles in the follicular waves of terminaldevelopment, and renewed by the continuous growth of smallerfollicles [12].
When clinicians and zootechnicians talk about ovarianreserve, they usually refer to this dynamic reserve of smallantral follicles. However, the wording ‘‘ovarian reserve’’ canbe confusing, since the growing follicles themselves developfrom a first reserve of primordial follicles, which is constitutedearly in life. In this review, we consider these two differentovarian reserves: the former, the pre-established reserve ofprimordial follicles, and the latter, the dynamic reserve of smallantral follicles. The subject of this review concerns the generalfeatures of these reserves, the tools available for evaluating
them quantitatively and qualitatively, their interrelationships,and the regulation of the transitions between reservesthroughout life in physiological, pathological, and experimen-tal situations.
THE PRE-ESTABLISHED RESERVE OF PRIMORDIALFOLLICLES
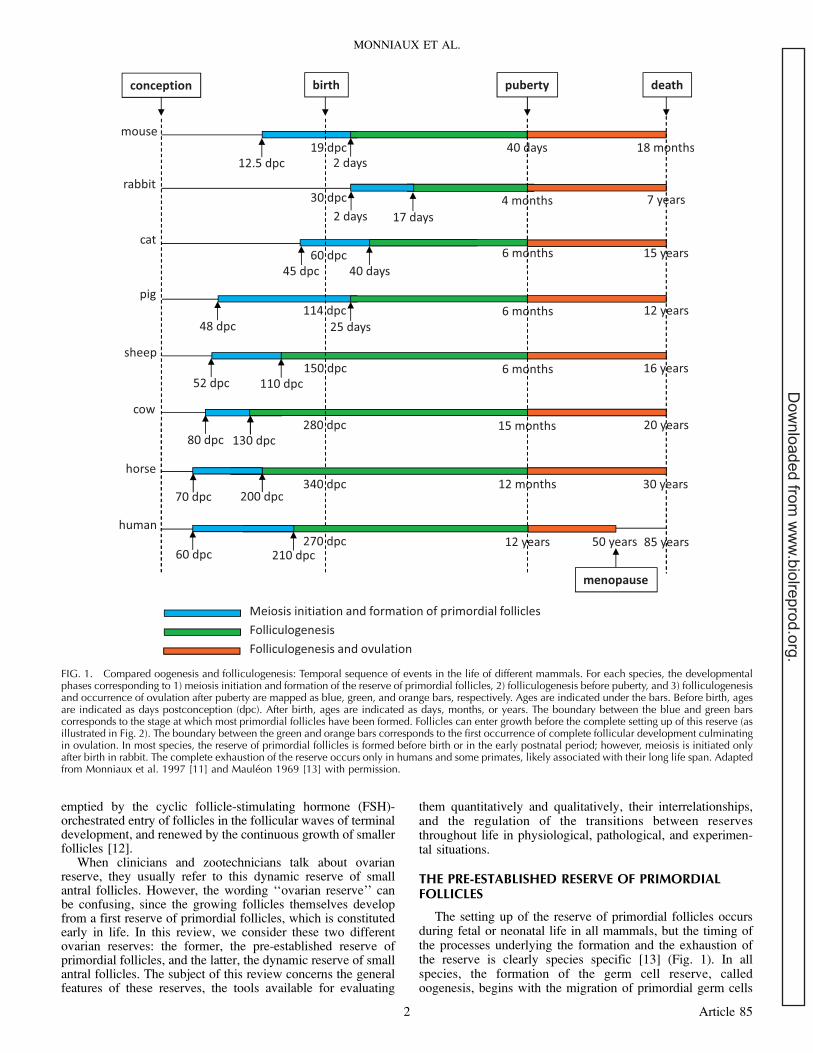
The setting up of the reserve of primordial follicles occursduring fetal or neonatal life in all mammals, but the timing ofthe processes underlying the formation and the exhaustion ofthe reserve is clearly species specific [13] (Fig. 1). In allspecies, the formation of the germ cell reserve, calledoogenesis, begins with the migration of primordial germ cells
FIG. 1. Compared oogenesis and folliculogenesis: Temporal sequence of events in the life of different mammals. For each species, the developmentalphases corresponding to 1) meiosis initiation and formation of the reserve of primordial follicles, 2) folliculogenesis before puberty, and 3) folliculogenesisand occurrence of ovulation after puberty are mapped as blue, green, and orange bars, respectively. Ages are indicated under the bars. Before birth, agesare indicated as days postconception (dpc). After birth, ages are indicated as days, months, or years. The boundary between the blue and green barscorresponds to the stage at which most primordial follicles have been formed. Follicles can enter growth before the complete setting up of this reserve (asillustrated in Fig. 2). The boundary between the green and orange bars corresponds to the first occurrence of complete follicular development culminatingin ovulation. In most species, the reserve of primordial follicles is formed before birth or in the early postnatal period; however, meiosis is initiated onlyafter birth in rabbit. The complete exhaustion of the reserve occurs only in humans and some primates, likely associated with their long life span. Adaptedfrom Monniaux et al. 1997 [11] and Mauleon 1969 [13] with permission.
MONNIAUX ET AL.
2 Article 85
Dow
nloaded from w
ww
.biolreprod.org.
into the gonadal ridges and their proliferation as oogonia withinovarian nests or cysts. Then oogonia enlarge and develop intoprimary oocytes, which initiate meiotic prophase. Thebreakdown of the cysts leads to the formation of 30-lm-diameter primordial follicles, each one consisting of a primaryoocyte arrested at the diplotene stage of prophase I of meiosis,surrounded by one layer of somatic cells, the pregranulosa, thecellular origin (ovarian or mesonephric surface epithelial cells)of which remains the subject of controversy [14, 15]. Abnormalfollicle formation is associated with a massive loss of oocytesat this stage. After forming, the primordial follicles may beginto grow either immediately or after a gap, depending on thespecies [16]. Alternatively, they become quiescent. In this lattercase, they will either degenerate or resume their growth severalmonths or years later. The molecular mechanisms underlying
oogenesis and follicle growth activation have been the subjectof recent reviews [17–20].
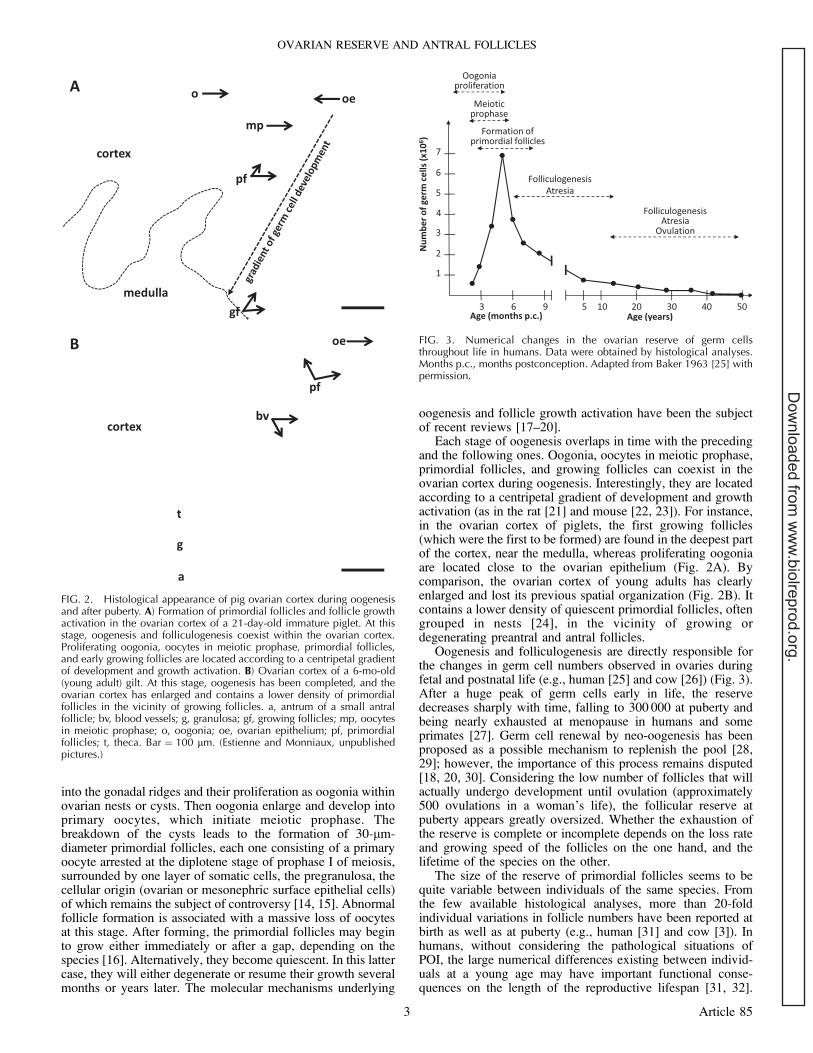
Each stage of oogenesis overlaps in time with the precedingand the following ones. Oogonia, oocytes in meiotic prophase,primordial follicles, and growing follicles can coexist in theovarian cortex during oogenesis. Interestingly, they are locatedaccording to a centripetal gradient of development and growthactivation (as in the rat [21] and mouse [22, 23]). For instance,in the ovarian cortex of piglets, the first growing follicles(which were the first to be formed) are found in the deepest partof the cortex, near the medulla, whereas proliferating oogoniaare located close to the ovarian epithelium (Fig. 2A). Bycomparison, the ovarian cortex of young adults has clearlyenlarged and lost its previous spatial organization (Fig. 2B). Itcontains a lower density of quiescent primordial follicles, oftengrouped in nests [24], in the vicinity of growing ordegenerating preantral and antral follicles.
Oogenesis and folliculogenesis are directly responsible forthe changes in germ cell numbers observed in ovaries duringfetal and postnatal life (e.g., human [25] and cow [26]) (Fig. 3).After a huge peak of germ cells early in life, the reservedecreases sharply with time, falling to 300 000 at puberty andbeing nearly exhausted at menopause in humans and someprimates [27]. Germ cell renewal by neo-oogenesis has beenproposed as a possible mechanism to replenish the pool [28,29]; however, the importance of this process remains disputed[18, 20, 30]. Considering the low number of follicles that willactually undergo development until ovulation (approximately500 ovulations in a woman’s life), the follicular reserve atpuberty appears greatly oversized. Whether the exhaustion ofthe reserve is complete or incomplete depends on the loss rateand growing speed of the follicles on the one hand, and thelifetime of the species on the other.
The size of the reserve of primordial follicles seems to bequite variable between individuals of the same species. Fromthe few available histological analyses, more than 20-foldindividual variations in follicle numbers have been reported atbirth as well as at puberty (e.g., human [31] and cow [3]). Inhumans, without considering the pathological situations ofPOI, the large numerical differences existing between individ-uals at a young age may have important functional conse-quences on the length of the reproductive lifespan [31, 32].
FIG. 2. Histological appearance of pig ovarian cortex during oogenesisand after puberty. A) Formation of primordial follicles and follicle growthactivation in the ovarian cortex of a 21-day-old immature piglet. At thisstage, oogenesis and folliculogenesis coexist within the ovarian cortex.Proliferating oogonia, oocytes in meiotic prophase, primordial follicles,and early growing follicles are located according to a centripetal gradientof development and growth activation. B) Ovarian cortex of a 6-mo-old(young adult) gilt. At this stage, oogenesis has been completed, and theovarian cortex has enlarged and contains a lower density of primordialfollicles in the vicinity of growing follicles. a, antrum of a small antralfollicle; bv, blood vessels; g, granulosa; gf, growing follicles; mp, oocytesin meiotic prophase; o, oogonia; oe, ovarian epithelium; pf, primordialfollicles; t, theca. Bar ¼ 100 lm. (Estienne and Monniaux, unpublishedpictures.)
FIG. 3. Numerical changes in the ovarian reserve of germ cellsthroughout life in humans. Data were obtained by histological analyses.Months p.c., months postconception. Adapted from Baker 1963 [25] withpermission.
OVARIAN RESERVE AND ANTRAL FOLLICLES
3 Article 85
Dow
nloaded from w
ww
.biolreprod.org.
Despite its indisputable interest, an accurate estimation of thesize of the reserve of primordial follicles of an individualremains difficult, due to the lack of molecular markers. Fromthe results of a recent study combining laser capturemicrodissection with transcriptome analysis in sheep ovary,oocyte-specific factors, such as DEAD (Asp-Glu-Ala-Asp) boxpolypeptide 4 (DDX4) (previously known as VASA), which wasfound to be highly expressed in primordial follicles, might beproposed as possible molecular markers [33]; however, theirvalidation as accurate markers of the reserve in a fragment ofovarian cortex remains to be assessed.
THE DYNAMIC RESERVE OF SMALL ANTRAL FOLLICLES
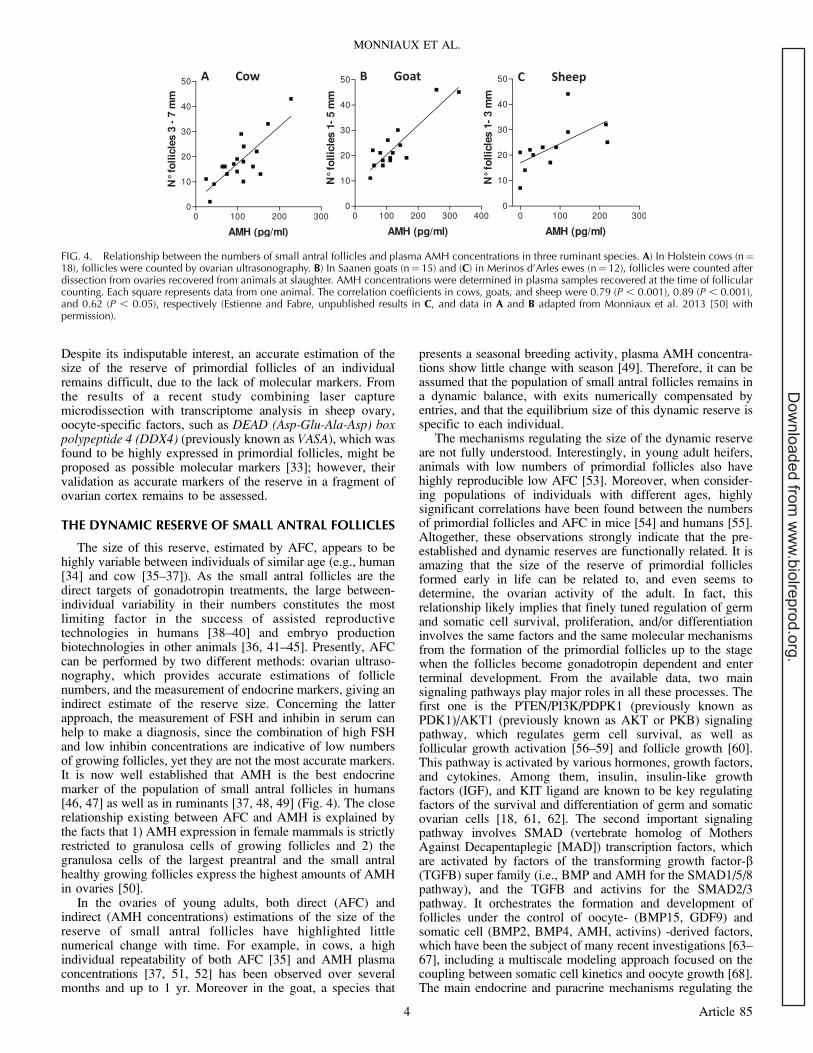
The size of this reserve, estimated by AFC, appears to behighly variable between individuals of similar age (e.g., human[34] and cow [35–37]). As the small antral follicles are thedirect targets of gonadotropin treatments, the large between-individual variability in their numbers constitutes the mostlimiting factor in the success of assisted reproductivetechnologies in humans [38–40] and embryo productionbiotechnologies in other animals [36, 41–45]. Presently, AFCcan be performed by two different methods: ovarian ultraso-nography, which provides accurate estimations of folliclenumbers, and the measurement of endocrine markers, giving anindirect estimate of the reserve size. Concerning the latterapproach, the measurement of FSH and inhibin in serum canhelp to make a diagnosis, since the combination of high FSHand low inhibin concentrations are indicative of low numbersof growing follicles, yet they are not the most accurate markers.It is now well established that AMH is the best endocrinemarker of the population of small antral follicles in humans[46, 47] as well as in ruminants [37, 48, 49] (Fig. 4). The closerelationship existing between AFC and AMH is explained bythe facts that 1) AMH expression in female mammals is strictlyrestricted to granulosa cells of growing follicles and 2) thegranulosa cells of the largest preantral and the small antralhealthy growing follicles express the highest amounts of AMHin ovaries [50].
In the ovaries of young adults, both direct (AFC) andindirect (AMH concentrations) estimations of the size of thereserve of small antral follicles have highlighted littlenumerical change with time. For example, in cows, a highindividual repeatability of both AFC [35] and AMH plasmaconcentrations [37, 51, 52] has been observed over severalmonths and up to 1 yr. Moreover in the goat, a species that
presents a seasonal breeding activity, plasma AMH concentra-tions show little change with season [49]. Therefore, it can beassumed that the population of small antral follicles remains ina dynamic balance, with exits numerically compensated byentries, and that the equilibrium size of this dynamic reserve isspecific to each individual.
The mechanisms regulating the size of the dynamic reserveare not fully understood. Interestingly, in young adult heifers,animals with low numbers of primordial follicles also havehighly reproducible low AFC [53]. Moreover, when consider-ing populations of individuals with different ages, highlysignificant correlations have been found between the numbersof primordial follicles and AFC in mice [54] and humans [55].Altogether, these observations strongly indicate that the pre-established and dynamic reserves are functionally related. It isamazing that the size of the reserve of primordial folliclesformed early in life can be related to, and even seems todetermine, the ovarian activity of the adult. In fact, thisrelationship likely implies that finely tuned regulation of germand somatic cell survival, proliferation, and/or differentiationinvolves the same factors and the same molecular mechanismsfrom the formation of the primordial follicles up to the stagewhen the follicles become gonadotropin dependent and enterterminal development. From the available data, two mainsignaling pathways play major roles in all these processes. Thefirst one is the PTEN/PI3K/PDPK1 (previously known asPDK1)/AKT1 (previously known as AKT or PKB) signalingpathway, which regulates germ cell survival, as well asfollicular growth activation [56–59] and follicle growth [60].This pathway is activated by various hormones, growth factors,and cytokines. Among them, insulin, insulin-like growthfactors (IGF), and KIT ligand are known to be key regulatingfactors of the survival and differentiation of germ and somaticovarian cells [18, 61, 62]. The second important signalingpathway involves SMAD (vertebrate homolog of MothersAgainst Decapentaplegic [MAD]) transcription factors, whichare activated by factors of the transforming growth factor-b(TGFB) super family (i.e., BMP and AMH for the SMAD1/5/8pathway), and the TGFB and activins for the SMAD2/3pathway. It orchestrates the formation and development offollicles under the control of oocyte- (BMP15, GDF9) andsomatic cell (BMP2, BMP4, AMH, activins) -derived factors,which have been the subject of many recent investigations [63–67], including a multiscale modeling approach focused on thecoupling between somatic cell kinetics and oocyte growth [68].The main endocrine and paracrine mechanisms regulating the
FIG. 4. Relationship between the numbers of small antral follicles and plasma AMH concentrations in three ruminant species. A) In Holstein cows (n¼18), follicles were counted by ovarian ultrasonography. B) In Saanen goats (n¼ 15) and (C) in Merinos d’Arles ewes (n¼ 12), follicles were counted afterdissection from ovaries recovered from animals at slaughter. AMH concentrations were determined in plasma samples recovered at the time of follicularcounting. Each square represents data from one animal. The correlation coefficients in cows, goats, and sheep were 0.79 (P , 0.001), 0.89 (P , 0.001),and 0.62 (P , 0.05), respectively (Estienne and Fabre, unpublished results in C, and data in A and B adapted from Monniaux et al. 2013 [50] withpermission).
MONNIAUX ET AL.
4 Article 85
Dow
nloaded from w
ww
.biolreprod.org.
dynamic reserve of small antral follicles are summarized inFigure 5.
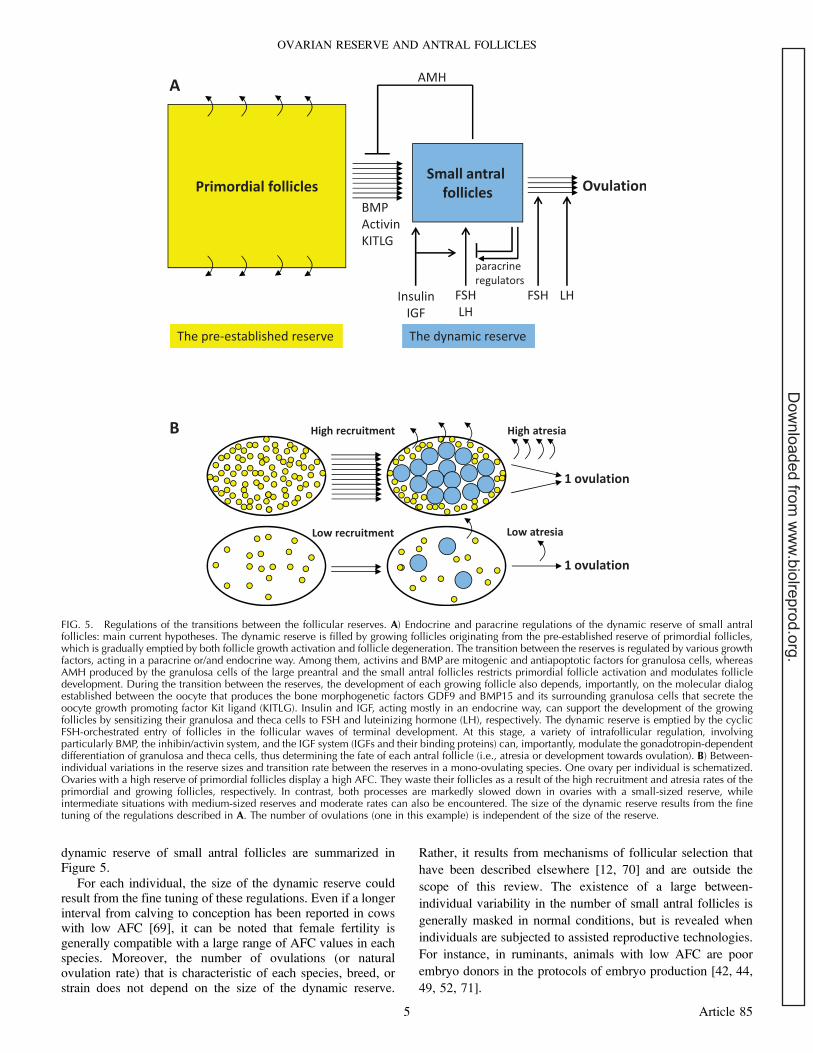
For each individual, the size of the dynamic reserve couldresult from the fine tuning of these regulations. Even if a longerinterval from calving to conception has been reported in cowswith low AFC [69], it can be noted that female fertility isgenerally compatible with a large range of AFC values in eachspecies. Moreover, the number of ovulations (or naturalovulation rate) that is characteristic of each species, breed, orstrain does not depend on the size of the dynamic reserve.
Rather, it results from mechanisms of follicular selection that
have been described elsewhere [12, 70] and are outside the
scope of this review. The existence of a large between-
individual variability in the number of small antral follicles is
generally masked in normal conditions, but is revealed when
individuals are subjected to assisted reproductive technologies.
For instance, in ruminants, animals with low AFC are poor
embryo donors in the protocols of embryo production [42, 44,
49, 52, 71].
FIG. 5. Regulations of the transitions between the follicular reserves. A) Endocrine and paracrine regulations of the dynamic reserve of small antralfollicles: main current hypotheses. The dynamic reserve is filled by growing follicles originating from the pre-established reserve of primordial follicles,which is gradually emptied by both follicle growth activation and follicle degeneration. The transition between the reserves is regulated by various growthfactors, acting in a paracrine or/and endocrine way. Among them, activins and BMP are mitogenic and antiapoptotic factors for granulosa cells, whereasAMH produced by the granulosa cells of the large preantral and the small antral follicles restricts primordial follicle activation and modulates follicledevelopment. During the transition between the reserves, the development of each growing follicle also depends, importantly, on the molecular dialogestablished between the oocyte that produces the bone morphogenetic factors GDF9 and BMP15 and its surrounding granulosa cells that secrete theoocyte growth promoting factor Kit ligand (KITLG). Insulin and IGF, acting mostly in an endocrine way, can support the development of the growingfollicles by sensitizing their granulosa and theca cells to FSH and luteinizing hormone (LH), respectively. The dynamic reserve is emptied by the cyclicFSH-orchestrated entry of follicles in the follicular waves of terminal development. At this stage, a variety of intrafollicular regulation, involvingparticularly BMP, the inhibin/activin system, and the IGF system (IGFs and their binding proteins) can, importantly, modulate the gonadotropin-dependentdifferentiation of granulosa and theca cells, thus determining the fate of each antral follicle (i.e., atresia or development towards ovulation). B) Between-individual variations in the reserve sizes and transition rate between the reserves in a mono-ovulating species. One ovary per individual is schematized.Ovaries with a high reserve of primordial follicles display a high AFC. They waste their follicles as a result of the high recruitment and atresia rates of theprimordial and growing follicles, respectively. In contrast, both processes are markedly slowed down in ovaries with a small-sized reserve, whileintermediate situations with medium-sized reserves and moderate rates can also be encountered. The size of the dynamic reserve results from the finetuning of the regulations described in A. The number of ovulations (one in this example) is independent of the size of the reserve.
OVARIAN RESERVE AND ANTRAL FOLLICLES
5 Article 85
Dow
nloaded from w
ww
.biolreprod.org.

OVARIAN RESERVE EXHAUSTION: GENETICDETERMINISM
Both ovarian reserves are exhausted at menopause inhumans and some primates, unlike in most mammals (Figs. 1and 3). In some pathological situations, a premature depletionof follicles is observed, which affects approximately 1%–2% ofwomen under the age of 40 yr and results in POI. Some geneticcomponent has been evidenced among the possible causes ofPOI [72].
Various natural or experimentally induced mutations ingenes encoding the ligands, receptors, or signaling effectors ofthe PTEN/PI3K/PDPK1/AKT1 or the SMAD signalingpathways can accelerate the exhaustion rate of the ovarianreserves and cause POI. The deletion of Pten, PDPK1,Ribosomal protein S6 (Rps6), or Forkhead box O3 (Foxo3,previously known as Foxo3A) in mouse oocytes induces adramatic activation and/or premature loss of the primordialfollicles, resulting in POI around the onset of sexual maturity[56–58]. In regard to the SMAD pathway, no data are availableon the exhaustion rate of the reserve of primordial follicles inSmad mutant mice [66]. However, mutations in some factors ofthe TGFB super family have been shown to affect the transitionof growing follicles between the two follicular reserves (Table1). In mice, the deletions of Amh [73, 74] or Inha [75–77], aswell as the overexpression of Bmp15 [78], accelerate thistransition, resulting in a premature depletion of the reserve ofprimordial follicles. In contrast, the deletion of Gdf9 blocks thegrowth of primary follicles, causing primary sterility [79], anda similar phenotype is observed in sheep carrying homozygousGDF9 mutations responsible for a loss of function of theprotein [80, 81]. Interestingly, the transition of folliclesbetween the reserves is not impaired in mice when Bmp15 isdeleted [82], whereas it is completely blocked in sheep carryinghomozygous loss-of-function mutations in BMP15 [83–86],indicating that the involvement of BMP15 in the control offollicular development is species specific. In humans, variousmutations in BMP15 and, to a lesser extent, in GDF9 andINHA, have been found to be associated with POI [72, 87, 88],and genetic variants of AMH and its receptor AMHR2 areassociated with different ages at menopause [89, 90],confirming the importance of these factors in the lifespan ofthe ovarian reserves.
As shown in the mouse genetic models described above, it isunusual to have beneficial effects of mutations on AFC withoutadverse side effects on the ovarian reserves in the long term(Table 1). However, in sheep carrying either heterozygous loss-of-function BMP15 mutations or the BMPR1BQ249R partial loss-of-function mutation, high AFC is not accompanied by thepremature depletion of the reserves [84, 91–93]. Rather, in thelatter genetic model, despite the presence of a low reserve ofprimordial follicles at birth, the ovarian reserves at 5 yr of ageare higher in ewes carrying the mutation, suggesting thatattenuation of the intraovarian signaling pathway of BMPs mayin fact be a successful means of limiting follicle consumption,hence preserving reproductive longevity and fertility [93].
Another promising way to increase the lifespan of theovarian reserves was proposed from the dramatic prolongationof the ovarian lifespan observed after Bax deletion in mice,suggesting that the mitochondrial factors of the Bcl2 family orthe death effector caspase cascade could modulate numericalchanges in the ovarian reserve through their pro- orantiapoptotic actions [94, 95]. However, up to now, nomutations in apoptosis regulator genes have been found to beassociated with the exhaustion rate of the ovarian reserves inhumans, so their importance in POI syndrome remainshypothetical.
DYNAMICS OF THE FOLLICULAR RESERVES DURINGLIFE
The reserve of primordial follicles is progressively depletedthroughout life, but the activation rate of primordial folliclesvaries as the ovary ages. In the young ovaries, the follicles thatwere formed first during oogenesis in the deepest part of thecortex massively activate first (Fig. 2), and are then relayed byfollicles located in outer parts of the cortex, and follicularactivation rate is progressively reduced [21]. Based on astatistical study of the human ovarian reserve size fromconception to menopause, the rate of follicular activation hasbeen estimated to increase from birth until approximately age14 yr, then to decrease toward menopause [31]. However, inanother study, an accelerated decay rate of the reserve wasfound before menopausal transition, resulting from enhancedfollicular activation rate in aging ovaries [96]. With aging, thedecline in primordial follicle numbers parallels the decrease in
TABLE 1. Experimental and natural mutations in genes encoding factors of the TGFB super family and their effects on the size of the two follicularreserves, the transition rate between them, and the occurrence of a premature depletion.
Animal species and genetic modelFirst
reserve* Transition rateSecondreserve$
Prematuredepletion� References
Mouse, Amh�/� Normal Accelerated rate andincreased atresia
High Yes [73, 74]
Mouse, Inha�/� Normal Accelerated High ND [76]Mouse, Bmp15�/� Normal Fairly normal Normal to low Reported in
some animals[82]
Mouse, Bmp15 overexpression Normal Accelerated rate andincreased atresia
Normal Yes [78]
Mouse, Gdf9�/� Normal No transition 0 / [79]Mouse, Gdf9þ/� Normal Normal Normal No [79]Mouse, Inha �/�Gdf9�/� Normal Accelerated Normal ND [75, 77]Mouse, Bmp15�/�Gdf9þ/� Normal Increased atresia Low Yes [82]Sheep, BMP15 homozygous loss-of-function mutations Normal No transition 0 / [83–85]Sheep, GDF9 homozygous loss-of-function mutations Normal No or few transition 0 to very low / [80, 81]Sheep, BMP15 heterozygous loss-of-function mutations Normal Unknown High No [91, 92]Sheep, BMPR1BQ249R partial loss-of-function mutation Low Slow High No [84, 92, 93]
* The pre-established reserve of primordial follicles.$ The dynamic reserve of small antral follicles.� ND, not determined, due to the premature death of animals by tumor development.
MONNIAUX ET AL.
6 Article 85
Dow
nloaded from w
ww
.biolreprod.org.

size of the dynamic reserve of small antral follicles [97–99] andAMH blood concentrations [100, 101].
These numerical changes are accompanied by changes inthe quality of the follicles of the dynamic reserve. In ruminants,oocytes recovered from small antral follicles before pubertydisplay a suboptimal developmental competence in in vitroembryo production systems compared to oocytes from adults,and this poor oocyte quality is associated with differences inRNA stores [102]. Similarly, in humans, during childhood andadolescence there is a high proportion of morphologicallyabnormal follicles that show reduced capacity for in vitrogrowth [103]. In mice, the loss of follicles during theprepubertal period has been proposed to be an active processnecessary to eliminate the follicles that are in excess,containing poor quality oocytes [104]. Compromised oocytequality also occurs in aged ovaries [98, 105, 106], due tounfavorable microenvironment (vascularization defects, oxida-tive stress, influence of toxic compounds), leading to a highprevalence of aneuploidy [107]. During the premenopausal
period, the high FSH endocrine levels, resulting from thedecline in the number of inhibin-secreting follicles, may alsohave deleterious effects upon follicle and oocyte quality [108,109], while increasing multiple ovulations [110]. Thesequalitative and quantitative follicular changes contribute tofemale infertility and constitute an important limit to thesuccess of assisted reproductive technologies in premenopausalwomen [111]. The dynamics of the follicular reserves and theassociated endocrine changes throughout life are summarizedin Figure 6.
THE MANAGEMENT OF THE OVARIAN RESERVES:FUTURE DIRECTIONS
As described above, the existence of a large, between-individual variability in AFC constitutes the most limitingfactor in the success of assisted reproductive technologies [36,38–45]. Moreover, between-individual variability in thetransition rates between ovarian reserves and in the gonado-tropin-regulated development of the antral follicles toward
FIG. 6. Schematic representation of the ovarian reserves, including their exhaustion dynamics and associated endocrine changes in AMH, FSH, andinhibin throughout life. The reserves are represented by two tanks, the first containing the primordial follicles, which can either degenerate after activationand entry into growth or develop and enter the second tank, containing the small antral, gonadotropin-responsive follicles. The endocrine changes arerepresented by a balance between FSH and inhibin. The fulcrum of the balance is AMH, the endocrine levels of which are proportional to the number ofhealthy small antral follicles present in the second tank. Left panel: in early life, the first tank contains a high number of primordial follicles; most of theprimordial follicles, which were formed first during oogenesis (red balls), are also the first to be activated and to enter the second tank. At that point, thereis a burst of growing follicles, mirrored by high AMH endocrine levels and associated with high and low inhibin (INH) and FSH endocrine levels,respectively. These growing follicles enclose oocytes with suboptimal developmental competence after in vitro fertilization. In vivo, all growing follicleswill degenerate by atresia before or after entering the second tank. Middle panel: in the adult age period, the number of primordial follicles has decreasedand the first tank contains follicles formed later during oogenesis (green balls), which are able to give rise to growing follicles containing oocytes withoptimal developmental competence. The second tank has reached an equilibrium size (with exits broadly compensated by entries), which is specific toeach individual, correlated with the size of the first tank and mirrored by AMH endocrine levels. The antral follicles present in the second tank can enterterminal development, and some of them will be selected for ovulation. Right panel: in the presenescence period, the first tank contains a low number ofaged primordial follicles (black balls), which are able to give rise to growing follicles containing low-quality oocytes. The second tank has a reduced size,associated with low to undetectable AMH endocrine levels that can, therefore, enhance follicular growth activation. The high FSH endocrine levelspresent at this stage could be responsible for the increased occurrence of multiple ovulations in mono-ovulating species during aging (period ofpremenopause in women).
OVARIAN RESERVE AND ANTRAL FOLLICLES
7 Article 85
Dow
nloaded from w
ww
.biolreprod.org.

ovulation can account for important differences betweenfemales in fertility and longevity of reproductive function.This variability likely has strong genetic components, but thecrucial question to answer now is: how to optimize thedynamics of the follicular reserves?
One strategy is to target the PTEN/PI3K/PDPK1/AKT1pathway, particularly by acting upon metabolism throughenvironmental and hormonal factors. This can apply to theimprovement of embryo production in animals, since increasesin insulin and IGF1 consecutive to a short-term feedingregimen with a high energy intake (flushing) have been shownto increase AFC and the follicular responses to gonadotropinsin ruminants [112, 113]. This could also be applied to humanclinics, particularly to restore the fertility of women withpolycystic ovary syndrome (PCOS), a common hyperandro-genic disorder often associated with metabolic alterations,chiefly insulin resistance and obesity. The ovaries of PCOSwomen are characterized by a slow transition betweenfollicular reserves, blocking of antral follicular development,and anovulation despite a high AFC [114, 115]; interestingly,improving insulin sensitivity by dietary therapy and exerciseregimens, or by administrating insulin-sensitizing drugs [116–118], can mobilize the growing follicles and restore ovulation.In a very interesting complementary perspective, new treat-ments for PCOS patients have been proposed recently toreinforce the activation of AKT1 signaling; they consist of thelocal disruption of the Hippo signaling pathway regulated byactin polymerization, as AKT1 and Hippo pathways can act inconcert to regulate follicular growth [119].
An alternative strategy is to target the BMP/AMH/SMADpathway. For example, in sheep, it has been recently shownthat the knockdown of AMH bioactivity by active immuniza-tion leads to a decline in the numbers of small growingfollicles, but an increase in AFC and ovulation numbers [120].In contrast, the administration of AMH would lower folliclegrowth activation and preserve more follicles for developmentlater in life, thereby causing a delay in the onset of menopause,as hypothesized in a lifelong model for the female reproductivecycle [121]. From these studies, AMH knockdown may haveshort-term therapeutic value in women who respond poorly toovarian stimulation, whereas AMH treatment might preservethe follicular reserves in the long term. From the sheep geneticmodels described above [84, 91–93], targeting the signalingpathway of BMPs by immunization or specific antagonists mayalso be a promising way to increase both AFC in the short termand reproductive longevity in the long term.
Moreover, the discovery of epigenetic regulation hashighlighted the importance of developmental programmingfor ovarian function. In sheep, testosterone exposure duringfetal life causes reproductive and metabolic disruptions,reproducing the human PCOS syndrome in adult animals[122]. This experimental situation might reflect, at least partly,some pathological cases, since endocrine disorders of PCOSoccur in female infants born to mothers with PCOS [115]. Asanother example, maternal nutrition and health during gestationin cows have been shown to influence AFC in prepubertal andadult offspring [123, 124]. All these observations indicate thatinteractions between genes and the maternal-fetal hormonalenvironment at the time of the formation of the reserve ofprimordial follicles may program postnatal ovarian function.
In conclusion, the pre-established reserve of primordialfollicles and the dynamic reserve of small antral follicles arefunctionally related and under both genetic and environmentalcontrol. From the recent improvements in the methodsavailable to estimate the reserve size, and our increasedknowledge of the cellular processes and signaling pathways
regulating their dynamics, new perspectives have been openedto improve the management of the ovarian reserves in the nearfuture.
REFERENCES
1. Jaiswal RS, Singh J, Adams GP. Developmental pattern of small antralfollicles in the bovine ovary. Biol Reprod 2004; 71:1244–1251.
2. Deb S, Batcha M, Campbell BK, Jayaprakasan K, Clewes JS, HopkissonJF, Sjoblom C, Raine-Fenning NJ. The predictive value of the automatedquantification of the number and size of small antral follicles in womenundergoing ART. Hum Reprod 2009; 24:2124–2132.
3. Ireland JJ, Smith GW, Scheetz D, Jimenez-Krassel F, Folger JK, IrelandJL, Mossa F, Lonergan P, Evans AC. Does size matter in females? Anoverview of the impact of the high variation in the ovarian reserve onovarian function and fertility, utility of anti-Mullerian hormone as adiagnostic marker for fertility and causes of variation in the ovarianreserve in cattle. Reprod Fertil Dev 2011; 23:1–14.
4. Broekmans FJ, Kwee J, Hendriks DJ, Mol BW, Lambalk CB. Asystematic review of tests predicting ovarian reserve and IVF outcome.Hum Reprod Update 2006; 12:685–718.
5. Broer SL, Dolleman M, Opmeer BC, Fauser BC, Mol BW, BroekmansFJ. AMH and AFC as predictors of excessive response in controlledovarian hyperstimulation: a meta-analysis. Hum Reprod Update 2011;17:46–54.
6. Baruselli PS, Sa Filho MF, Ferreira RM, Sales JN, Gimenes LU, VieiraLM, Mendanha MF, Bo GA. Manipulation of follicle development toensure optimal oocyte quality and conception rates in cattle. ReprodDomest Anim 2012; 47(Suppl 4):134–141.
7. Rodgers RJ, Irving-Rodgers HF. Formation of the ovarian follicularantrum and follicular fluid. Biol Reprod 2010; 82:1021–1029.
8. Fortune JE. Ovarian follicular growth and development in mammals.Biol Reprod 1994; 50:225–232.
9. Ireland JJ, Mihm M, Austin E, Diskin MG, Roche JF. Historicalperspective of turnover of dominant follicles during the bovine estrouscycle: key concepts, studies, advancements, and terms. J Dairy Sci 2000;83:1648–1658.
10. Gougeon A. Human ovarian follicular development: from activation ofresting follicles to preovulatory maturation. Ann Endocrinol (Paris) 2010;71:132–143.
11. Monniaux D, Huet C, Besnard N, Clement F, Bosc M, Pisselet C,Monget P, Mariana JC. Follicular growth and ovarian dynamics inmammals. J Reprod Fertil Suppl 1997; 51:3–23.
12. Scaramuzzi RJ, Baird DT, Campbell BK, Driancourt MA, Dupont J,Fortune JE, Gilchrist RB, Martin GB, McNatty KP, McNeilly AS,Monget P, Monniaux D, et al. Regulation of folliculogenesis and thedetermination of ovulation rate in ruminants. Reprod Fertil Dev 2011; 23:444–467.
13. Mauleon P. Oogenesis and folliculogenesis. In: Cole HH, Cupps PT(eds.), Reproduction in Domestic Animals. New York: Academic Press;1969:187–215.
14. Sawyer HR, Smith P, Heath DA, Juengel JL, Wakefield SJ, McNatty KP.Formation of ovarian follicles during fetal development in sheep. BiolReprod 2002; 66:1134–1150.
15. Hummitzsch K, Irving-Rodgers HF, Hatzirodos N, Bonner W, SabatierL, Reinhardt DP, Sado Y, Ninomiya Y, Wilhelm D, Rodgers RJ. A newmodel of development of the mammalian ovary and follicles. PLoS One2013; 8:e55578.
16. Fortune JE, Yang MY, Allen JJ, Herrick SL. Triennial ReproductionSymposium: the ovarian follicular reserve in cattle: what regulates itsformation and size? J Anim Sci 2013; 91:3041–3050.
17. Tingen C, Kim A, Woodruff TK. The primordial pool of follicles andnest breakdown in mammalian ovaries. Mol Hum Reprod 2009; 15:795–803.
18. Monget P, Bobe J, Gougeon A, Fabre S, Monniaux D, Dalbies-Tran R.The ovarian reserve in mammals: a functional and evolutionaryperspective. Mol Cell Endocrinol 2012; 356:2–12.
19. McLaughlin EA, McIver SC. Awakening the oocyte: controllingprimordial follicle development. Reproduction 2009; 137:1–11.
20. Kerr JB, Myers M, Anderson RA. The dynamics of the primordialfollicle reserve. Reproduction 2013; 146:R205–R215.
21. Hirshfield AN, DeSanti AM. Patterns of ovarian cell proliferation in ratsduring the embryonic period and the first three weeks postpartum. BiolReprod 1995; 53:1208–1221.
22. Byskov AG, Guoliang X, Andersen CY. The cortex-medulla oocyte
MONNIAUX ET AL.
8 Article 85
Dow
nloaded from w
ww
.biolreprod.org.

growth pattern is organized during fetal life: an in-vitro study of themouse ovary. Mol Hum Reprod 1997; 3:795–800.
23. Mork L, Maatouk DM, McMahon JA, Guo JJ, Zhang P, McMahon AP,Capel B. Temporal differences in granulosa cell specification in the ovaryreflect distinct follicle fates in mice. Biol Reprod 2012; 86:37.
24. Da Silva-Buttkus P, Marcelli G, Franks S, Stark J, Hardy K. Inferringbiological mechanisms from spatial analysis: prediction of a localinhibitor in the ovary. Proc Natl Acad Sci U S A 2009; 106:456–461.
25. Baker TG. A quantitative and cytological study of germ cells in humanovaries. Proc R Soc Lond B Biol Sci 1963; 158:417–433.
26. Erickson BH. Development and senescence of the postnatal bovineovary. J Anim Sci 1966; 25:800–805.
27. Walker ML, Herndon JG. Menopause in nonhuman primates? BiolReprod 2008; 79:398–406.
28. Johnson J, Canning J, Kaneko T, Pru JK, Tilly JL. Germline stem cellsand follicular renewal in the postnatal mammalian ovary. Nature 2004;428:145–150.
29. Tilly JL, Niikura Y, Rueda BR. The current status of evidence for andagainst postnatal oogenesis in mammals: a case of ovarian optimismversus pessimism? Biol Reprod 2009; 80:2–12.
30. Kerr JB, Brogan L, Myers M, Hutt KJ, Mladenovska T, Ricardo S,Hamza K, Scott CL, Strasser A, Findlay JK. The primordial folliclereserve is not renewed after chemical or c-irradiation mediated depletion.Reproduction 2012; 143:469–476.
31. Wallace WH, Kelsey TW. Human ovarian reserve from conception to themenopause. PLoS One 2010; 5:e8772.
32. Faddy MJ. Follicle dynamics during ovarian ageing. Mol Cell Endocrinol2000; 163:43–48.
33. Bonnet A, Bevilacqua C, Benne F, Bodin L, Cotinot C, Liaubet L,Sancristobal M, Sarry J, Terenina E, Martin P, Tosser-Klopp G, Mandon-Pepin B. Transcriptome profiling of sheep granulosa cells and oocytesduring early follicular development obtained by laser capture microdis-section. BMC Genomics 2011; 12:417.
34. Broekmans FJ, Faddy MJ, Scheffer G, te Velde ER. Antral follicle countsare related to age at natural fertility loss and age at menopause.Menopause 2004; 11:607–614.
35. Burns DS, Jimenez-Krassel F, Ireland JL, Knight PG, Ireland JJ.Numbers of antral follicles during follicular waves in cattle: evidence forhigh variation among animals, very high repeatability in individuals, andan inverse association with serum follicle-stimulating hormone concen-trations. Biol Reprod 2005; 73:54–62.
36. Monniaux D, Chupin D, Saumande J. Superovulatory responses of cattle.Theriogenology 1983; 19:55–81.
37. Rico C, Fabre S, Medigue C, di Clemente N, Clement F, Bontoux M,Touze JL, Dupont M, Briant E, Remy B, Beckers JF, Monniaux D. Anti-Mullerian hormone is an endocrine marker of ovarian gonadotropin-responsive follicles and can help to predict superovulatory responses inthe cow. Biol Reprod 2009; 80:50–59.
38. Fauser BC, Diedrich K, Devroey P. Predictors of ovarian response:progress towards individualized treatment in ovulation induction andovarian stimulation. Hum Reprod Update 2008; 14:1–14.
39. Hendriks DJ, Mol BW, Bancsi LF, Te Velde ER, Broekmans FJ. Antralfollicle count in the prediction of poor ovarian response and pregnancyafter in vitro fertilization: a meta-analysis and comparison with basalfollicle-stimulating hormone level. Fertil Steril 2005; 83:291–301.
40. Jayaprakasan K, Campbell B, Hopkisson J, Johnson I, Raine-Fenning N.A prospective, comparative analysis of anti-Mullerian hormone, inhibin-B, and three-dimensional ultrasound determinants of ovarian reserve inthe prediction of poor response to controlled ovarian stimulation. FertilSteril 2010; 93:855–864.
41. Cushman RA, DeSouza JC, Hedgpeth VS, Britt JH. Superovulatoryresponse of one ovary is related to the micro- and macroscopicpopulation of follicles in the contralateral ovary of the cow. Biol Reprod1999; 60:349–354.
42. Ireland JJ, Ward F, Jimenez-Krassel F, Ireland JL, Smith GW, LonerganP, Evans AC. Follicle numbers are highly repeatable within individualanimals but are inversely correlated with FSH concentrations and theproportion of good-quality embryos after ovarian stimulation in cattle.Hum Reprod 2007; 22:1687–1695.
43. Singh J, Dominguez M, Jaiswal R, Adams GP. A simple ultrasound testto predict the superstimulatory response in cattle. Theriogenology 2004;62:227–243.
44. Mossa F, Duffy P, Naitana S, Lonergan P, Evans AC. Associationbetween numbers of ovarian follicles in the first follicle wave andsuperovulatory response in ewes. Anim Reprod Sci 2007; 100:391–396.
45. Taneja M, Bols PE, Van de Velde A, Ju JC, Schreiber D, Tripp MW,Levine H, Echelard Y, Riesen J, Yang X. Developmental competence of
juvenile calf oocytes in vitro and in vivo: influence of donor animalvariation and repeated gonadotropin stimulation. Biol Reprod 2000; 62:206–213.
46. La Marca A, Sighinolfi G, Radi D, Argento C, Baraldi E, Artenisio AC,Stabile G, Volpe A. Anti-Mullerian hormone (AMH) as a predictivemarker in assisted reproductive technology (ART). Hum Reprod Update2010; 16:113–130.
47. Loh JS, Maheshwari A. Anti-Mullerian hormone—is it a crystal ball forpredicting ovarian ageing? Hum Reprod 2011; 26:2925–2932.
48. Lahoz B, Alabart JL, Cocero MJ, Monniaux D, Echegoyen E, Sanchez P,Folch J. Anti-Mullerian hormone concentration in sheep and itsdependence of age and independence of BMP15 genotype: an endocrinepredictor to select the best donors for embryo biotechnologies.Theriogenology 2014; 81:347–357.
49. Monniaux D, Baril G, Laine AL, Jarrier P, Poulin N, Cognie J, Fabre S.Anti-Mullerian hormone as a predictive endocrine marker for embryoproduction in the goat. Reproduction 2011; 142:845–854.
50. Monniaux D, Drouilhet L, Rico C, Estienne A, Jarrier P, Touze JL, SapaJ, Phocas F, Dupont J, Dalbies-Tran R, Fabre S. Regulation of anti-Mullerian hormone production in domestic animals. Reprod Fertil Dev2013; 25:1–16.
51. Rico C, Medigue C, Fabre S, Jarrier P, Bontoux M, Clement F,Monniaux D. Regulation of anti-Mullerian hormone production in thecow: a multiscale study at endocrine, ovarian, follicular, and granulosacell levels. Biol Reprod 2011; 84:560–571.
52. Rico C, Drouilhet L, Salvetti P, Dalbies-Tran R, Jarrier P, Touze JL,Pillet E, Ponsart C, Fabre S, Monniaux D. Determination of anti-Mullerian hormone concentrations in blood as a tool to select Holsteindonor cows for embryo production: from the laboratory to the farm.Reprod Fertil Dev 2012; 24:932–944.
53. Ireland JL, Scheetz D, Jimenez-Krassel F, Themmen AP, Ward F,Lonergan P, Smith GW, Perez GI, Evans AC, Ireland JJ. Antral folliclecount reliably predicts number of morphologically healthy oocytes andfollicles in ovaries of young adult cattle. Biol Reprod 2008; 79:1219–1225.
54. Kevenaar ME, Meerasahib MF, Kramer P, van de Lang-Born BM, deJong FH, Groome NP, Themmen AP, Visser JA. Serum anti-Mullerianhormone levels reflect the size of the primordial follicle pool in mice.Endocrinology 2006; 147:3228–3234.
55. Hansen KR, Hodnett GM, Knowlton N, Craig LB. Correlation of ovarianreserve tests with histologically determined primordial follicle number.Fertil Steril 2011; 95:170–175.
56. Reddy P, Liu L, Adhikari D, Jagarlamudi K, Rajareddy S, Shen Y, Du C,Tang W, Hamalainen T, Peng SL, Lan ZJ, Cooney AJ, et al. Oocyte-specific deletion of Pten causes premature activation of the primordialfollicle pool. Science 2008; 319:611–613.
57. John GB, Gallardo TD, Shirley LJ, Castrillon DH. Foxo3 is a PI3K-dependent molecular switch controlling the initiation of oocyte growth.Dev Biol 2008; 321:197–204.
58. Reddy P, Adhikari D, Zheng W, Liang S, Hamalainen T, Tohonen V,Ogawa W, Noda T, Volarevic S, Huhtaniemi I, Liu K. PDK1 signaling inoocytes controls reproductive aging and lifespan by manipulating thesurvival of primordial follicles. Hum Mol Genet 2009; 18:2813–2824.
59. Li J, Kawamura K, Cheng Y, Liu S, Klein C, Duan EK, Hsueh AJ.Activation of dormant ovarian follicles to generate mature eggs. ProcNatl Acad Sci U S A 2010; 107:10280–10284.
60. Li Q, He H, Zhang YL, Li XM, Guo X, Huo R, Bi Y, Li J, Fan HY, ShaJ. Phosphoinositide 3-kinase p110delta mediates estrogen- and FSH-stimulated ovarian follicle growth. Mol Endocrinol 2013; 27:1468–1482.
61. Hutt KJ, McLaughlin EA, Holland MK. Kit ligand and c-Kit have diverseroles during mammalian oogenesis and folliculogenesis. Mol HumReprod 2006; 12:61–69.
62. Monget P, Bondy C. Importance of the IGF system in early folliculo-genesis. Mol Cell Endocrinol 2000; 163:89–93.
63. Shimasaki S, Moore RK, Otsuka F, Erickson GF. The bonemorphogenetic protein system in mammalian reproduction. EndocrRev 2004; 25:72–101.
64. Knight PG, Satchell L, Glister C. Intra-ovarian roles of activins andinhibins. Mol Cell Endocrinol 2012; 359:53–65.
65. Pangas SA. Bone morphogenetic protein signaling transcription factor(SMAD) function in granulosa cells. Mol Cell Endocrinol 2012; 356:40–47.
66. Matzuk MM, Burns KH. Genetics of mammalian reproduction: modelingthe end of the germline. Annu Rev Physiol 2012; 74:503–528.
67. Visser JA, Themmen AP. Role of anti-Mullerian hormone and bonemorphogenetic proteins in the regulation of FSH sensitivity. Mol CellEndocrinol 2014; 382:460–465.
OVARIAN RESERVE AND ANTRAL FOLLICLES
9 Article 85
Dow
nloaded from w
ww
.biolreprod.org.

68. Clement F, Michel P, Monniaux D, Stiehl T. Coupled somatic cellkinetics and germ cell growth: multiscale model-based insight on ovarianfollicular development. Multiscale Model Simul 2013; 11:719–746.
69. Mossa F, Walsh SW, Butler ST, Berry DP, Carter F, Lonergan P, SmithGW, Ireland JJ, Evans AC. Low numbers of ovarian follicles �3 mm indiameter are associated with low fertility in dairy cows. J Dairy Sci 2012;95:2355–2361.
70. Clement F, Monniaux D. Multiscale modelling of ovarian follicularselection. Prog Biophys Mol Biol 2013; 113:398–408.
71. Durocher J, Morin N, Blondin P. Effect of hormonal stimulation onbovine follicular response and oocyte developmental competence in acommercial operation. Theriogenology 2006; 65:102–115.
72. Persani L, Rossetti R, Cacciatore C, Fabre S. Genetic defects of ovarianTGF-beta-like factors and premature ovarian failure. J Endocrinol Invest2011; 34:244–251.
73. Durlinger AL, Kramer P, Karels B, de Jong FH, Uilenbroek JT,Grootegoed JA, Themmen AP. Control of primordial follicle recruitmentby anti-Mullerian hormone in the mouse ovary. Endocrinology 1999;140:5789–5796.
74. Visser JA, Durlinger AL, Peters IJ, van den Heuvel ER, Rose UM,Kramer P, de Jong FH, Themmen AP. Increased oocyte degeneration andfollicular atresia during the estrous cycle in anti-Mullerian hormone nullmice. Endocrinology 2007; 148:2301–2308.
75. Wu X, Chen L, Brown CA, Yan C, Matzuk MM. Interrelationship ofgrowth differentiation factor 9 and inhibin in early folliculogenesis andovarian tumorigenesis in mice. Mol Endocrinol 2004; 18:1509–1519.
76. Myers M, Middlebrook BS, Matzuk MM, Pangas SA. Loss of inhibinalpha uncouples oocyte-granulosa cell dynamics and disrupts postnatalfolliculogenesis. Dev Biol 2009; 334:458–467.
77. Myers M, Mansouri-Attia N, James R, Peng J, Pangas SA. GDF9modulates the reproductive and tumor phenotype of female inha-nullmice. Biol Reprod 2009; 88:86.
78. McMahon HE, Hashimoto O, Mellon PL, Shimasaki S. Oocyte-specificoverexpression of mouse bone morphogenetic protein-15 leads toaccelerated folliculogenesis and an early onset of acyclicity in transgenicmice. Endocrinology 2008; 149:2807–2815.
79. Dong J, Albertini DF, Nishimori K, Kumar TR, Lu N, Matzuk MM.Growth differentiation factor-9 is required during early ovarian folliculo-genesis. Nature 1996; 383:531–535.
80. McNatty KP, Galloway SM, Wilson T, Smith P, Hudson NL, O’ConnellA, Bibby AH, Heath DA, Davis GH, Hanrahan JP, Juengel JL.Physiological effects of major genes affecting ovulation rate in sheep.Genet Sel Evol 2005; 37(Suppl 1):S25–S38.
81. Nicol L, Bishop SC, Pong-Wong R, Bendixen C, Holm LE, Rhind SM,McNeilly AS. Homozygosity for a single base-pair mutation in theoocyte-specific GDF9 gene results in sterility in Thoka sheep.Reproduction 2009; 138:921–933.
82. Yan C, Wang P, DeMayo J, DeMayo FJ, Elvin JA, Carino C, Prasad SV,Skinner SS, Dunbar BS, Dube JL, Celeste AJ, Matzuk MM. Synergisticroles of bone morphogenetic protein 15 and growth differentiation factor9 in ovarian function. Mol Endocrinol 2001; 15:854–866.
83. Braw-Tal R, McNatty KP, Smith P, Heath DA, Hudson NL, Phillips DJ,McLeod BJ, Davis GH. Ovaries of ewes homozygous for the X-linkedInverdale gene (FecXI) are devoid of secondary and tertiary follicles butcontain many abnormal structures. Biol Reprod 1993; 49:895–907.
84. McNatty KP, Juengel JL, Wilson T, Galloway SM, Davis GH. Geneticmutations influencing ovulation rate in sheep. Reprod Fertil Dev 2001;13:549–555.
85. Bodin L, Di Pasquale E, Fabre S, Bontoux M, Monget P, Persani L,Mulsant P. A novel mutation in the bone morphogenetic protein 15 genecausing defective protein secretion is associated with both increasedovulation rate and sterility in Lacaune sheep. Endocrinology 2007; 148:393–400.
86. Galloway SM, McNatty KP, Cambridge LM, Laitinen MP, Juengel JL,Jokiranta TS, McLaren RJ, Luiro K, Dodds KG, Montgomery GW,Beattie AE, Davis GH, et al. Mutations in an oocyte-derived growthfactor gene (BMP15) cause increased ovulation rate and infertility in adosage-sensitive manner. Nat Genet 2000; 25:279–283.
87. Otsuka F, McTavish KJ, Shimasaki S. Integral role of GDF-9 and BMP-15 in ovarian function. Mol Reprod Dev 2011; 78:9–21.
88. Chand AL, Harrison CA, Shelling AN. Inhibin and premature ovarianfailure. Hum Reprod Update 2010; 16:39–50.
89. Kevenaar ME, Themmen AP, Rivadeneira F, Uitterlinden AG, Laven JS,van Schoor NM, Lips P, Pols HA, Visser JA. A polymorphism in theAMH type II receptor gene is associated with age at menopause ininteraction with parity. Hum Reprod 2007; 22:2382–2388.
90. Braem MG, Voorhuis M, van der Schouw YT, Peeters PH, Schouten LJ,
Eijkemans MJ, Broekmans FJ, Onland-Moret NC. Interactions betweengenetic variants in AMH and AMHR2 may modify age at naturalmenopause. PLoS One 2013; 8:e59819.
91. Shackell GH, Hudson NL, Heath DA, Lun S, Shaw L, Condell L, BlayLR, McNatty KP. Plasma gonadotropin concentrations and ovariancharacteristics in Inverdale ewes that are heterozygous for a major gene(FecX1) on the X chromosome that influences ovulation rate. BiolReprod 1993; 48:1150–1156.
92. McNatty KP, Smith P, Hudson NL, Heath DA, Tisdall DJ, O WS, Braw-Tal R. Development of the sheep ovary during fetal and early neonatallife and the effect of fecundity genes. J Reprod Fertil Suppl 1995; 49:123–135.
93. Ruoss C, Tadros A, O’Shea T, McFarlane J, Almahbobi G. Ovarianfollicle development in Booroola sheep exhibiting impaired bonemorphogenetic protein signalling pathway. Reproduction 2009; 138:689–696.
94. Perez GI, Robles R, Knudson CM, Flaws JA, Korsmeyer SJ, Tilly JL.Prolongation of ovarian lifespan into advanced chronological age byBax-deficiency. Nat Genet 1999; 21:200–203.
95. Morita Y, Tilly JL. Oocyte apoptosis: like sand through an hourglass.Dev Biol 1999; 213:1–17.
96. Gougeon A, Ecochard R, Thalabard JC. Age-related changes of thepopulation of human ovarian follicles: increase in the disappearance rateof non-growing and early-growing follicles in aging women. Biol Reprod1994; 50:653–663.
97. Scheffer GJ, Broekmans FJ, Dorland M, Habbema JD, Looman CW, teVelde ER. Antral follicle counts by transvaginal ultrasonography arerelated to age in women with proven natural fertility. Fertil Steril 1999;72:845–851.
98. Adams GP, Jaiswal R, Singh J, Malhi P. Progress in understandingovarian follicular dynamics in cattle. Theriogenology 2008; 69:72–80.
99. Ginther OJ, Gastal MO, Gastal EL, Jacob JC, Siddiqui MA, Beg MA.Effects of age on follicle and hormone dynamics during the oestrouscycle in mares. Reprod Fertil Dev 2008; 20:955–963.
100. de Vet A, Laven JS, de Jong FH, Themmen AP, Fauser BC.Antimullerian hormone serum levels: a putative marker for ovarianaging. Fertil Steril 2002; 77:357–362.
101. van Rooij IA, Broekmans FJ, Scheffer GJ, Looman CW, Habbema JD,de Jong FH, Fauser BJ, Themmen AP, te Velde ER. Serum antimullerianhormone levels best reflect the reproductive decline with age in normalwomen with proven fertility: a longitudinal study. Fertil Steril 2005; 83:979–987.
102. Romar R, De Santis T, Papillier P, Perreau C, Thelie A, Dell’Aquila ME,Mermillod P, Dalbies-Tran R. Expression of maternal transcripts duringbovine oocyte in vitro maturation is affected by donor age. ReprodDomest Anim 2011; 46:e23–30.
103. Anderson RA, McLaughlin M, Wallace WH, Albertini DF, Telfer EE.The immature human ovary shows loss of abnormal follicles andincreasing follicle developmental competence through childhood andadolescence. Hum Reprod 2014; 29:97–106.
104. Bristol-Gould SK, Kreeger PK, Selkirk CG, Kilen SM, Cook RW, KippJL, Shea LD, Mayo KE, Woodruff TK. Postnatal regulation of germ cellsby activin: the establishment of the initial follicle pool. Dev Biol 2006;298:132–148.
105. Hewitt DA, England GC. The effect of oocyte size and bitch age uponoocyte nuclear maturation in vitro. Theriogenology 1998; 49:957–966.
106. Carnevale EM. The mare model for follicular maturation andreproductive aging in the woman. Theriogenology 2008; 69:23–30.
107. Battaglia DE, Goodwin P, Klein NA, Soules MR. Influence of maternalage on meiotic spindle assembly in oocytes from naturally cyclingwomen. Hum Reprod 1996; 11:2217–2222.
108. Dursun P, Gultekin M, Yuce K, Ayhan A. What is the underlying causeof aneuploidy associated with increasing maternal age? Is it associatedwith elevated levels of gonadotropins? Med Hypotheses 2006; 66:143–147.
109. McTavish KJ, Jimenez M, Walters KA, Spaliviero J, Groome NP,Themmen AP, Visser JA, Handelsman DJ, Allan CM. Rising follicle-stimulating hormone levels with age accelerate female reproductivefailure. Endocrinology 2007; 148:4432–4439.
110. Beemsterboer SN, Homburg R, Gorter NA, Schats R, Hompes PG,Lambalk CB. The paradox of declining fertility but increasing twinningrates with advancing maternal age. Hum Reprod 2006; 21:1531–1532.
111. Broekmans FJ, Soules MR, Fauser BC. Ovarian aging: mechanisms andclinical consequences. Endocr Rev 2009; 30:465–493.
112. Gong JG, Armstrong DG, Baxter G, Hogg CO, Garnsworthy PC, WebbR. The effect of increased dietary intake on superovulatory response toFSH in heifers. Theriogenology 2002; 57:1591–1602.
MONNIAUX ET AL.
10 Article 85
Dow
nloaded from w
ww
.biolreprod.org.

113. Armstrong DG, Gong JG, Webb R. Interactions between nutrition and
ovarian activity in cattle: physiological, cellular and molecular
mechanisms. Reprod Suppl 2003; 61:403–414.
114. Franks S, Stark J, Hardy K. Follicle dynamics and anovulation in
polycystic ovary syndrome. Hum Reprod Update 2008; 14:367–378.
115. Dumesic DA, Richards JS. Ontogeny of the ovary in polycystic ovary
syndrome. Fertil Steril 2013; 100:23–38.
116. Moran LJ, Brinkworth GD, Norman RJ. Dietary therapy in polycystic
ovary syndrome. Semin Reprod Med 2008; 26:85–92.
117. Harrison CL, Lombard CB, Moran LJ, Teede HJ. Exercise therapy in
polycystic ovary syndrome: a systematic review. Hum Reprod Update
2011; 17:171–183.
118. Diamanti-Kandarakis E, Dunaif A. Insulin resistance and the polycystic
ovary syndrome revisited: an update on mechanisms and implications.
Endocr Rev 2012; 33:981–1030.
119. Kawamura K, Cheng Y, Suzuki N, Deguchi M, Sato Y, Takae S, Ho CH,
Kawamura N, Tamura M, Hashimoto S, Sugishita Y, Morimoto Y, et al.
Hippo signaling disruption and Akt stimulation of ovarian follicles forinfertility treatment. Proc Natl Acad Sci U S A 2013; 110:17474–17479.
120. Campbell BK, Clinton M, Webb R. The role of anti-Mullerian hormone(AMH) during follicle development in a monovulatory species (sheep).Endocrinology 2012; 153:4533–4543.
121. Margolskee A, Selgrade JF. A lifelong model for the female reproductivecycle with an antimullerian hormone treatment to delay menopause. JTheor Biol 2013; 326:21–35.
122. Padmanabhan V, Veiga-Lopez A. Sheep models of polycystic ovarysyndrome phenotype. Mol Cell Endocrinol 2013; 373:8–20.
123. Evans AC, Mossa F, Walsh SW, Scheetz D, Jimenez-Krassel F, IrelandJL, Smith GW, Ireland JJ. Effects of maternal environment duringgestation on ovarian folliculogenesis and consequences for fertility inbovine offspring. Reprod Domest Anim 2012; 47(Suppl 4):31–37.
124. Mossa F, Carter F, Walsh SW, Kenny DA, Smith GW, Ireland JL,Hildebrandt TB, Lonergan P, Ireland JJ, Evans AC. Maternalundernutrition in cows impairs ovarian and cardiovascular systems intheir offspring. Biol Reprod 2013; 88:92.
OVARIAN RESERVE AND ANTRAL FOLLICLES
11 Article 85
Dow
nloaded from w
ww
.biolreprod.org.
Related Documents











