The origin of species chapter 24. Evolutionary theory must explain how new species originate. Two basic patterns in which evolution of one species into one or more other species occurs.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The origin of species chapter 24.
Evolutionary theory must explain how new species originate.
Two basic patterns in which evolution of one species into one or more other species occurs.

The origin of species
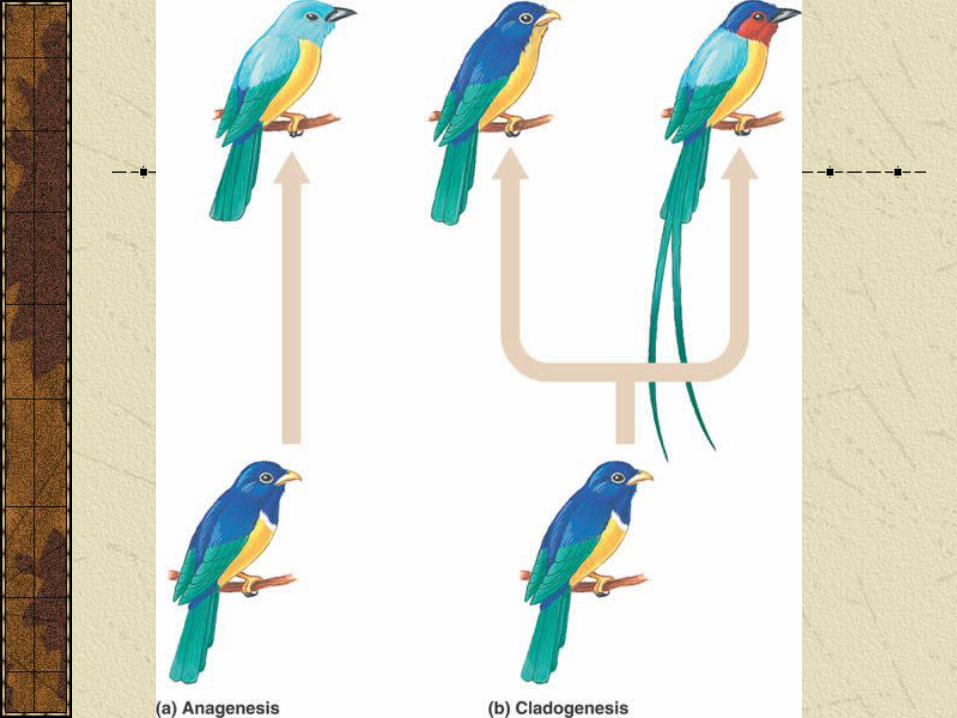
Anagenesis: accumulation of changes over time gradually transforms a species into a different species.
Cladogenesis: Gene pool splits into two or more pools which each give rise to new species.

The origin of species
Biological Species Concept
The species is a basic biological unit and humans seem to intuitively recognize species.

The origin of species
Why do species exist?
Why don’t we see a smooth continuous blending of one species into another?

The origin of species
Because intermediate forms are selected against.
If they were not selected against, then the two forms would merge into one as their gene pools mixed.

Species definitions
John Ray (1627-1705) gave first general definition of a species.
A species consists of all individuals that can breed together and produce fertile offspring.

A female donkey mated to a male horseproduces what?

A mule (which is sterile)Hence, donkeys and horses are separate species.

Biological Species Concept
Ray’s idea updated into the biological species concept.
“Species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups.” Ernst Mayr.

Reproductive Isolation
There are a large number of potential barriers that prevent different species producing viable, healthy adults.
These include both prezygotic and postzygotic isolating mechanisms (i.e., barriers that come respectively before and after mating).
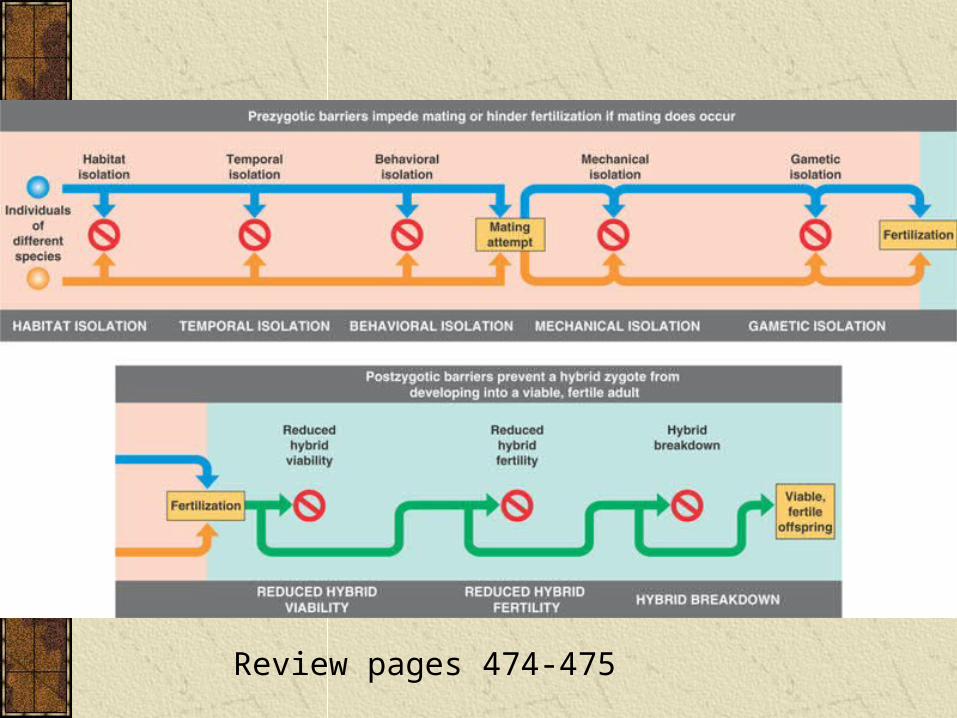
Review pages 474-475

But what about organisms that do not mate with another individual?
E.g. single-celled animals, bacteria, fungi and many plants reproduce asexually.
In practice, many organisms are assigned to species based on morphology or DNA.

Speciation
Classically, speciation has been viewed as a three stage process:
1. Isolation of populations.
2. Divergence in traits of separated populations (e.g. mating system or habitat use).
3. Reproductive isolation of populations that maintains isolation when members of different populations come into contact.

Speciation
Two ways in which speciation can occur.
Allopatric speciation occurs when a gene pool is geographically divided into two
Sympatric speciation occurs without geographic separation of the populations..
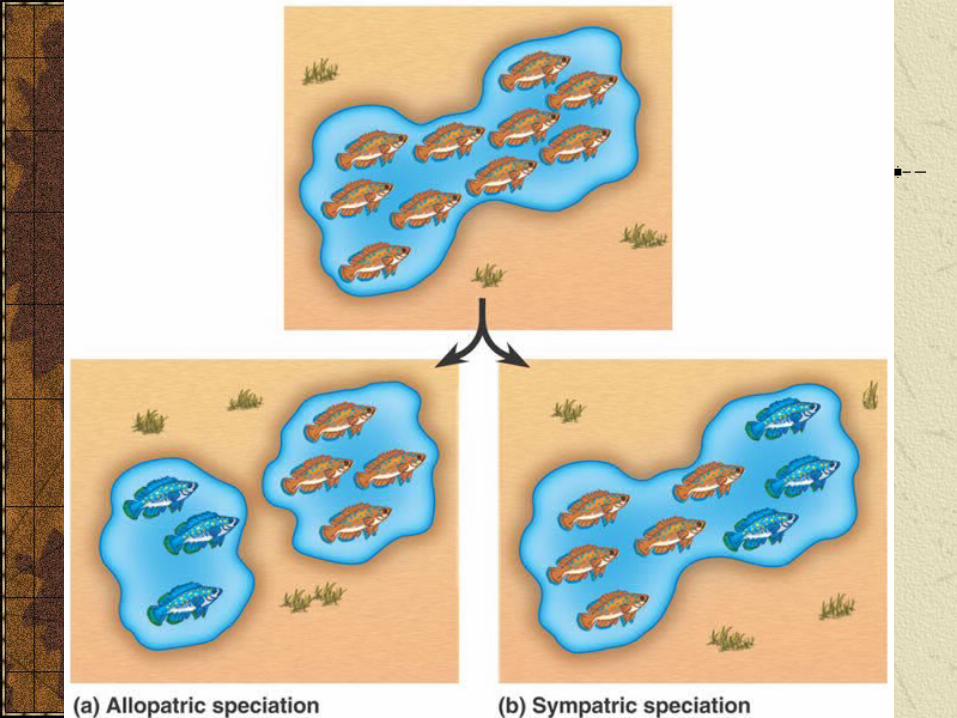

Allopatric speciationOccurs when a population is divided by a barrier.
Can occur because a barrier develops or because some members of population disperse to a new area.
Once separated, the gene pools diverge as each population adapts to its local environment. Over time isolating mechanisms are likely to develop.

Allopatric speciation
If after many generations members of the allopatric populations are brought back together they may or may not be able to produce fertile offspring.
Even if they can do so, those offspring may have intermediate characteristics which suit them to neither of the parental environments and thus they will be selected against.

Allopatric speciation
If intermediates are selected against, we would expect isolating mechanisms (barriers to reproduction) to be strongly favored by selection.
Ultimately the two populations would become different enough to be unable to interbreed successfully and so become new species.

Allopatric speciation
Examples.
Two species of closely related antelope squirrels live on opposite sides of the Grand Canyon. The canyon is a barrier to their dispersal.
In contrast, birds and other species that disperse well have not undergone speciation on opposite sides of the canyon

Allopatric speciation
Different Galapagos Islands contain different species of finches, which have evolved in the approximately 2 million years since the islands were first colonized from the South American mainland.

Allopatric speciation
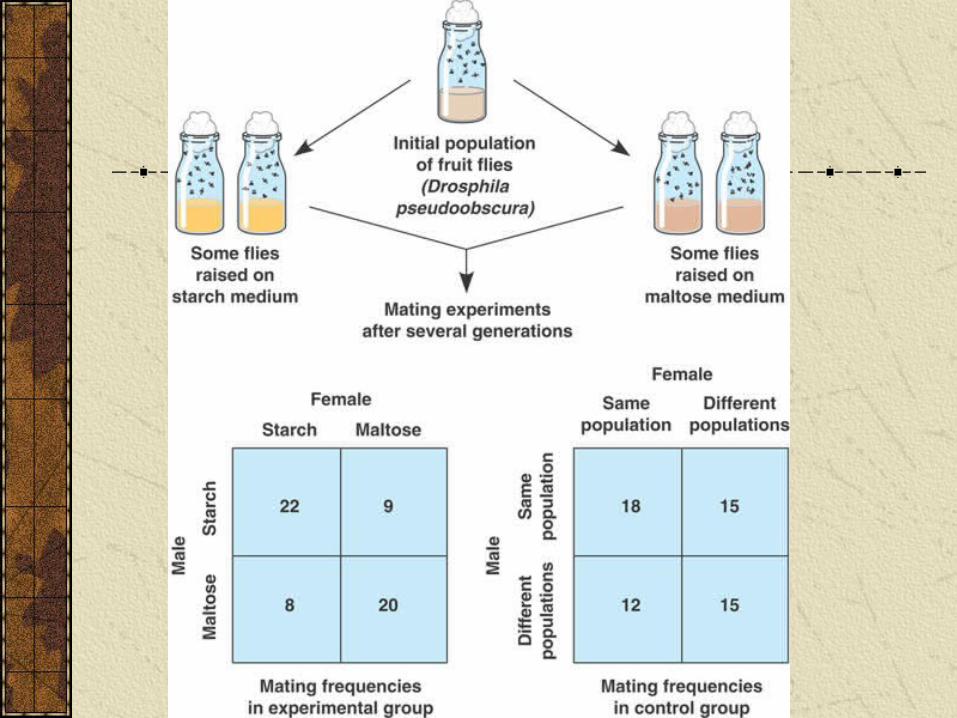
Diane Dodd investigated development of reproductive barriers in fruit flies.
Raised populations for several generations on either starch or maltose medium. Fly populations diverged each becoming better at digesting its food source.

Allopatric speciation
When flies from “starch populations” and from “maltose populations” brought together they were significantly more likely to mate with flies of their own population.
Indicates that reproductive barriers between species can begin to form quickly.

Sympatric Speciation
In sympatric speciation, speciation takes place in geographically overlapping populations.
Mechanisms of sympatric speciation include polyploidy and nonrandom mating that reduces gene flow.

Sympatric SpeciationPolyploidy is common in plants and many species have resulted from accidents in cell division that produce extra sets of chromosomes.
For example a diploid plant (2n chromosomes) may become a tetraploid (4n). The tetraploid cannot produce fertile young with diploid plants because young will be triploid (3n chromosomes), but can self-pollinate or mate with other tetraploids.
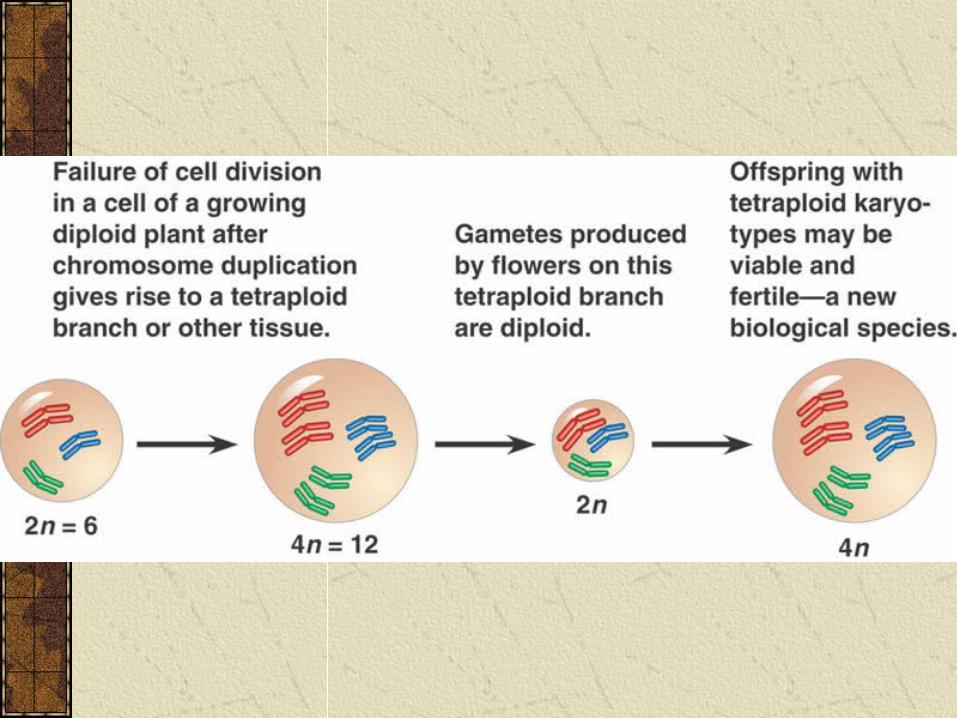

Sympatric Speciation
Polyploidy can thus result in speciation in just one generation.
Polyploidy can also occur when two different species produce a hybrid. The offspring are often sterile because chromosomes cannot pair up during meiosis. However, the plant can often reproduce asexually.

Sympatric Speciation
Subsequently, various mechanisms can convert a sterile hybrid into a fertile polyploid called an allopolyploid.
The allopolyploids are fertile with each other, but not other species and so are a new species.
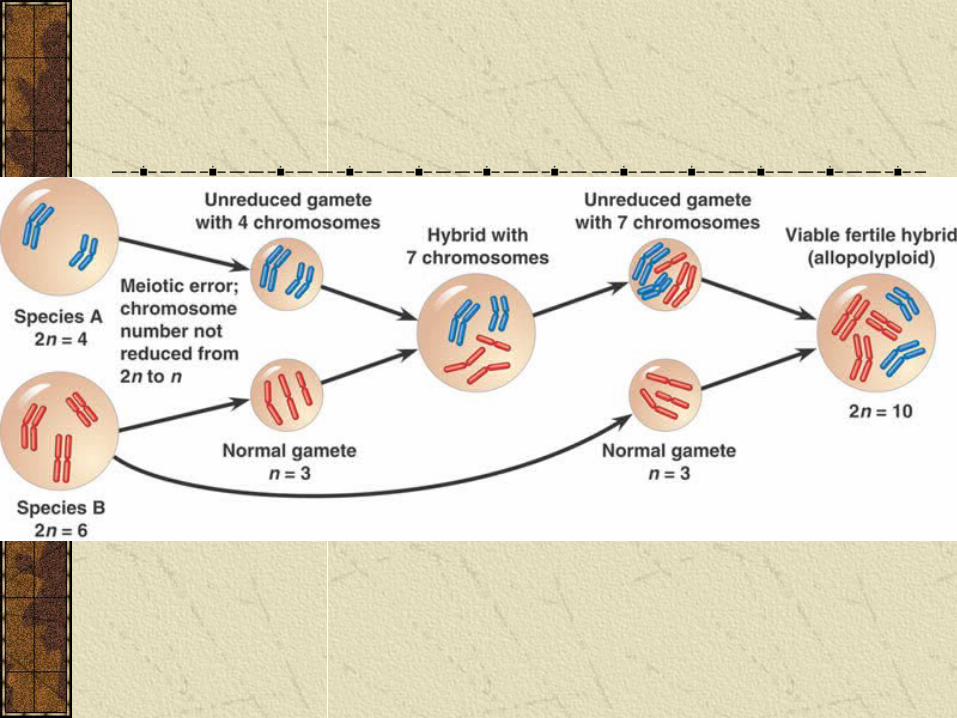

Sympatric Speciation
Many important crops are polyploids. For example, the wheat used for bread is an allohexaploid (six sets of chromosomes, with two sets from each of three different species).

Sympatric Speciation
Non-random mating. Reproductive isolation can occur when genetic factors enable a subpopulation to exploit a resource not used by the parental population.

Sympatric Speciation
Example: North American apple maggot fly. Original breeding habitat was hawthorn fruits on hawthorn trees, but about 200 years ago some populations colonized apple trees.
Apples mature faster than haws (hawthorn fruit) so apple-feeding flies have been selected for rapid development.

http://www.virginiafruit.ento.vt.edu/
AppleMaggot.html Larvae
http://bugguide.net/node/view/248877
Adult
http://www.eppo.int/
QUARANTINE/insects/
Rhagoletis_pomonella/RHAGPO_images.htm
Damaged apple

Sympatric speciation in apple-maggot flies
Natural selection favors divergence because hawthorn fruits ripen 3-4 weeks after apples. As a result, hawthorn fly larvae experience cool temperatures before pupating whereas apple fly larvae experience warmer temperatures.
Hawthorn flies and apple flies thus depend on different temperature signals to time their pupation and emergence the next spring and have different developmental timetables.

Sympatric Speciation
Apple-feeding flies now temporally isolated (isolated in time) from hawthorn-feeding flies. Speciation appears well underway.
A protein electrophoresis study by Feder et al. (1988,1990) showed that the two populations are genetically distinct.

Sympatric Speciation
Lake Victoria cichlids. Lake Victoria about 12,000 years old but home to more than 500 species of cichlids (fish).
There has been rapid speciation and some of it appears to have been caused by non-random mating in which females choose males based on their appearance.

Sympatric SpeciationResearchers studied two closely related species one which has a blue-tinged back and the other a red-tinged back.
In an aquarium with natural light females mated with males of their own species exclusively. However, in an aquarium under monochromatic orange light (where blue and red could not be distinguished), females mated indiscriminately and offspring were fertile.

Sympatric Speciation
Researchers concluded mate choice by females based on coloration is main barrier keeping gene pools separated.
Because fertile young are produced in interspecific crosses the speciation probably occurred recently.

Phylogeny and Systematics
Phylogeny is the evolutionary history of a species or group of species.
Systematics is science of understanding the diversity and relatedness of organisms.

Phylogeny and Systematics
Traditionally morphological similarities used to infer evolutionary relationships.
More recently, comparisons of DNA, RNA and other molecules used to infer relationships: molecular systematics.

Phylogeny and Systematics
Phylogenetic trees are based on common ancestry and data from various sources used to construct them:
Fossil evidence
Molecular evidence
Morphological evidence

Phylogeny and SystematicsIn constructing phylogenies important to distinguish between homologous structures (similar due to common descent) and analagous structures (similar because of convergent evolution).Australian sugar glider a marsupial and North American flying squirrel a eutherian mammal are examples of convergent evolution. Both possess gliding membrane, but otherwise only distantly related.
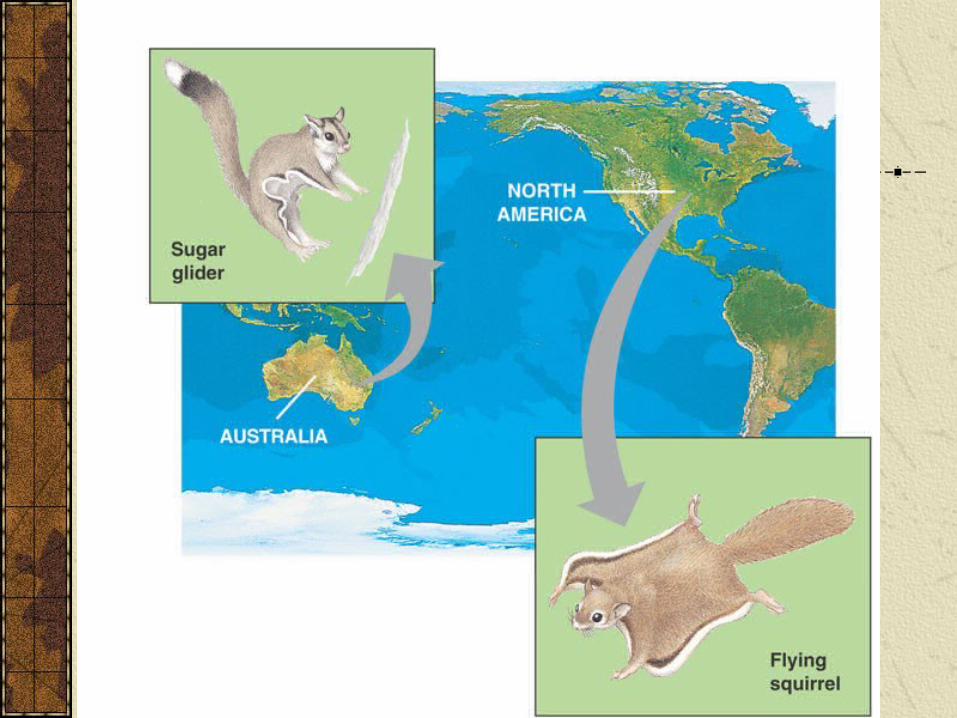

Phylogeny and Systematics
Analagous structures that have evolved independently are called homoplasies.
Deciding whether structures are homologous or analagous requires various types of evidence to be assessed.

Phylogeny and Systematics
Corroborating similarities in other structures
Fossil evidence
Complexity of characters being compared. The more points of resemblance there are between two structures the less likely it is they evolved independently.

Phylogeny and Systematics
Evaluating molecular homologies. Comparison of DNA sequences usually done using computer programs that match up sequences taking into account effects of insertions and deletions.
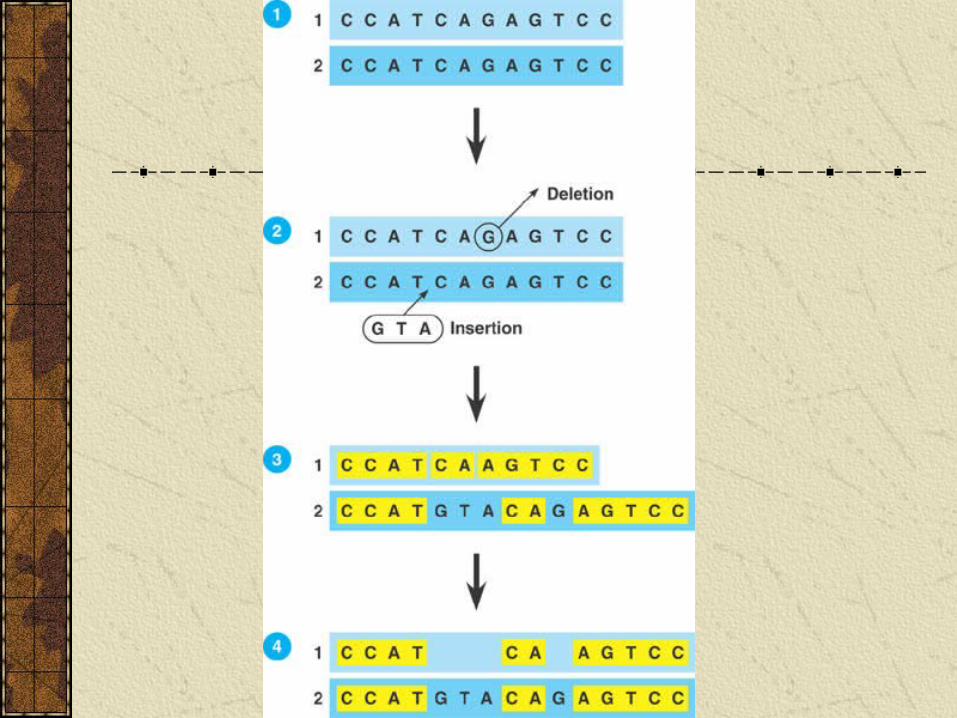

Phylogeny and Systematics
Science of systematics dates to Linnaeus in the 18th century who devised basic systems of binomial nomenclature and hierarchical classification in use today.
All organisms have a unique binomial name
E.g. Humans are Homo sapiens

Phylogeny and Systematics
Organisms are classified into hierarchical classifications that group closely related organisms and progressively include more and more organisms.
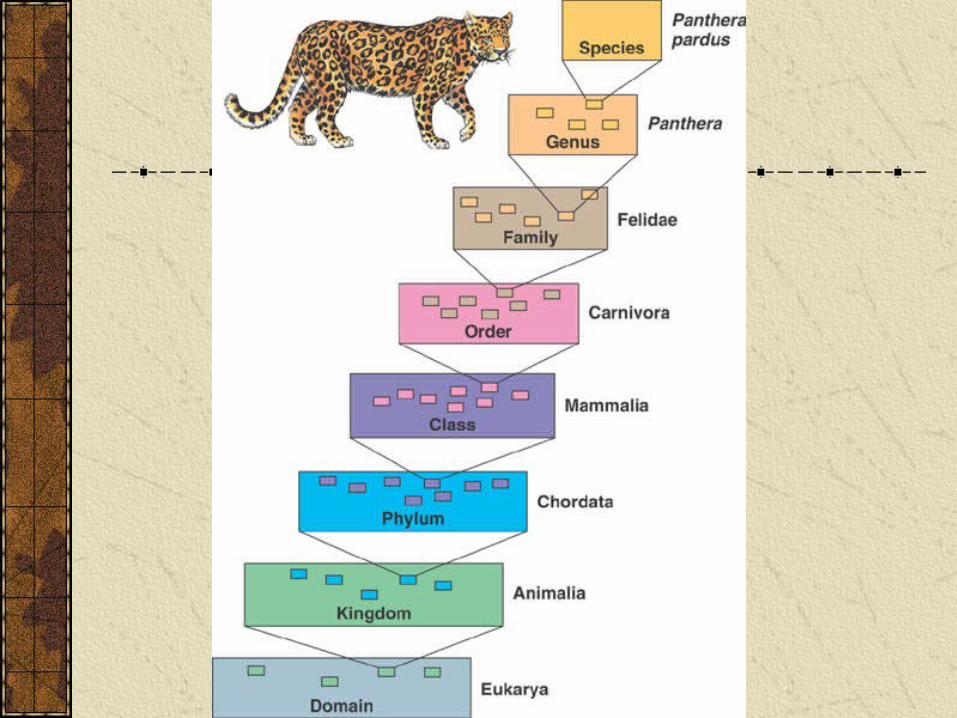

Phylogenetic trees
Branching diagrams called phylogenetic trees summarize evolutionary relationships and hierarchical classification is represented in finer branching of phylogenetic trees.
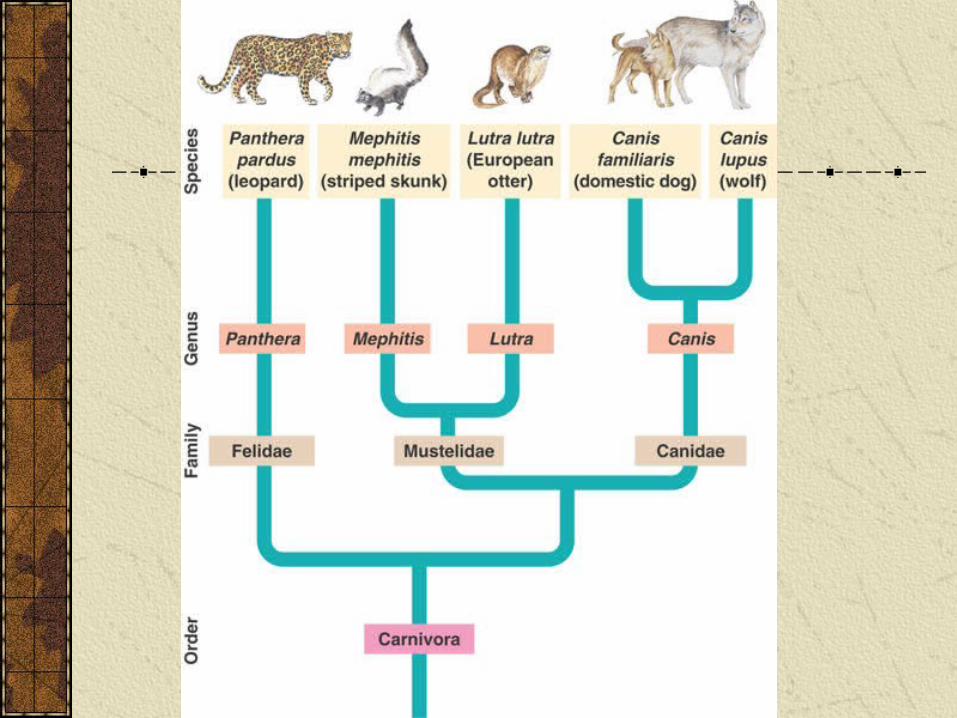

Phylogenetic trees
In a phylogenetic tree the tips of the branches specify particular species and the branch points represent common ancestors.
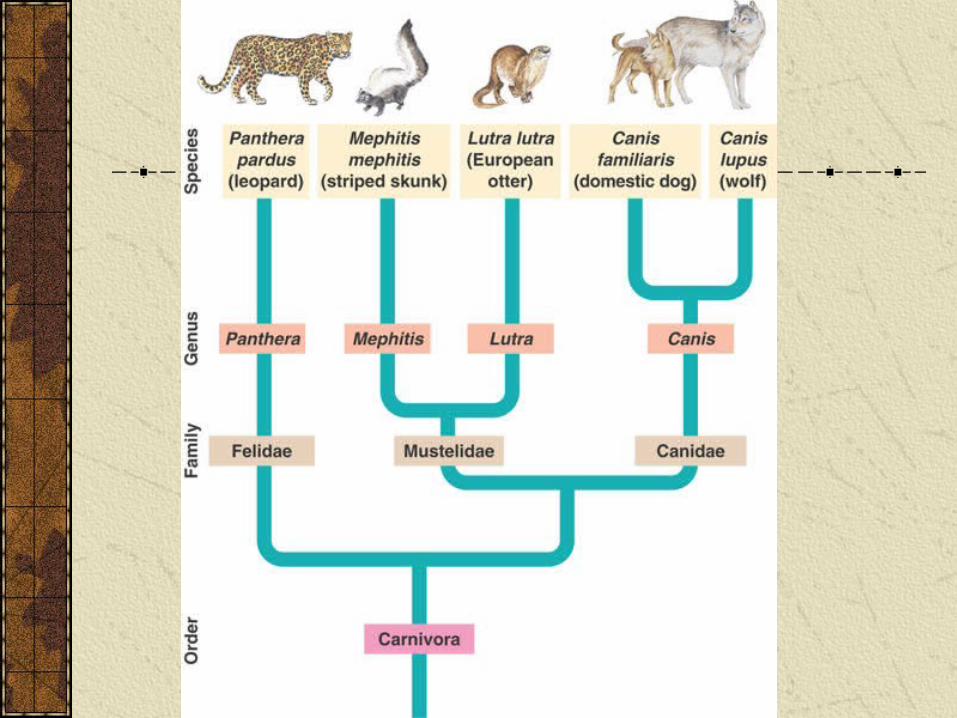

Cladistics and construction of phylogenetic trees
Cladograms are diagrams that display patterns of shared characteristics.
If shared characteristics are due to common ancestry the cladogram forms basis of a phylogenetic tree.

Cladograms
Within a tree a clade is defined as a group that includes an ancestral species and all of its descendants.
Cladistics is analysis of how species may be grouped into clades.

Shared derived characters
Cladograms are largely constructed using shared derived characters.
These are characteristics that are evolutionary novelties, new developments that are unique to a particular clade.

Shared derived characters
Shared derived characters are unique to the clade. For example, for mammals hair is a shared derived character

Shared primitive characters
Shared primitive characters are characters that are shared beyond the taxon we are interested in. Among vertebrates the backbone is an example because it evolved in ancestor of all vertebrates.
If you go back far enough in time shared primitive characters will be shared derived characters. Thus, the backbone is a shared derived character that distinguishes vertebrates from all other animals.

Constructing a cladogram
Outgroup comparison is used to begin building a cladogram.
An outgroup is a close relative of the members of the ingroup (the various species being studied) that provides a basis for comparison with the others.

Constructing a cladogram
The outgroup in a cladogram is determined using data different from that being used to construct the cladistic tree.
The outgroup “roots” the tree. The outgroup is based on the assumption that homologies in the outgroup and ingroup are primitive characters.

Constructing a cladogram
Having the outgroup for comparison enables researchers to focus on those characters derived after the separation from the outgroup to figure out relationships among species in the ingroup.

Constructing a cladogram
Cladogram of various vertebrates: leopard, tuna, salamander, turtle and lamprey.
Use lancelet as outgroup (is a chordate, but has no backbone).
Table summarizes data about character traits and which organisms possess them.
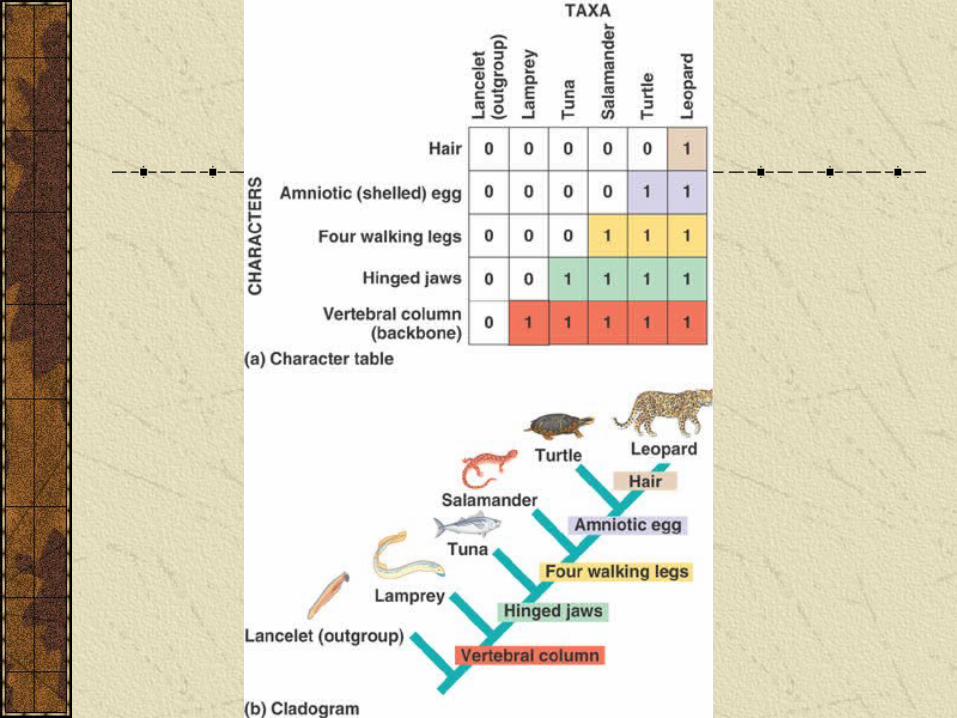

Constructing a cladogram
In the cladogram new characters are marked on the tree where they originate and these characters are possessed by all subsequent groups.
The cladogram of vertebrates is a step towards constructing a phylogenetic tree, but such a tree would need to be based on much more data. Unfortunately, additional data and additional species make it hard to decide on a best tree.

Identifying the “best” trees
When constructing a phylogenetic tree that involves many species there are billions of possible ways to arrange a tree.
We try to build tress that are the most likely. Generally these are trees that are the most parsimonious (require the fewest evolutionary changes) to construct.

Homology and analogy
The most parsimonious tree may not always correct.
If analogy versus homology mistakes are made the tree will be incorrect.
Which of the next two trees is the best tree?
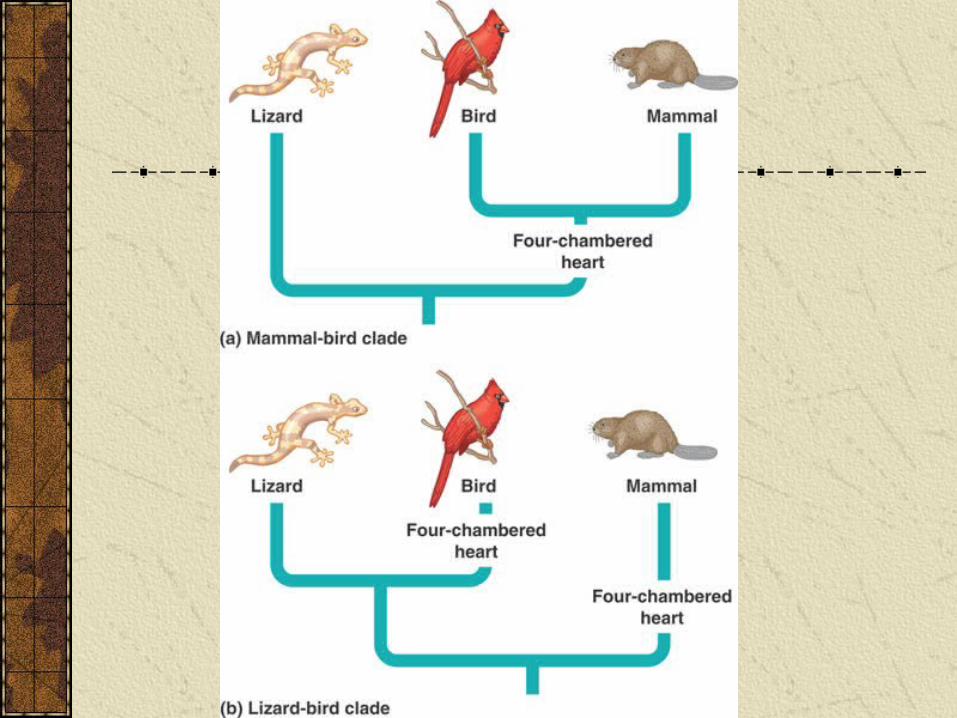

Homology and analogy
If mammal and bird four-chambered hearts are homologous, then tree A is most parsimonious.
However, lots of data suggest birds and reptiles more closely related, so tree B is better tree. Four-chambered heart evolved more than once.

Phylogenetic trees are hypotheses
Important to remember that phylogenetic trees are hypotheses for the evolutionary pathways.
Trees that we will have most confidence in will be supported by multiple lines of evidence (e.g. molecular, morphological and fossil evidence).

Molecular clocks
Trees of relatedness can be dated by using fossil evidence, but also by using molecular clocks.
Based on observation that some genes appear to evolve at fairly constant rates.

Molecular clocks
Assumption is that the number of changes in genes is proportional to the amount of time since two species branched from their common ancestor.
Molecular clocks are calibrated against the fossil record.

Molecular clocks
Molecular clocks are not perfect as genes may evolve in fits and starts (because of effects of selection) and not be very clocklike.

Molecular clocks
Some good markers to use for molecular clocks are silent mutations (changes in genes that do not change the amino acid coded for) because these will have no effect on selection. However, these are most useful over only relatively short time periods.

Applying a molecular clock: HIV
HIV is descended from viruses found in chimps and monkeys (SIV simian immunodeficiency virus).
To date the time the virus jumped to humans scientists have compared current HIV-1 M samples to some from tissue samples preserved in 1959.

Applying a molecular clock: HIV
Samples showed virus has evolved at steady rate and by extrapolating back using the molecular clock have estimated that HIV-1 M first infected humans in the 1930’s.
Related Documents






![Evolutionary Phylogenetic Networks: Models and Issues...1 Introduction In Charles Darwin’s Origin of Species [17], the depiction of an evolutionary history of species took the shape](https://static.cupdf.com/doc/110x72/6003f8dcf6d8c3703e43d90f/evolutionary-phylogenetic-networks-models-and-issues-1-introduction-in-charles.jpg)




