THE JOGRNAL OF BIOLOGICAL CHEHISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology , Inc VOl. 269, No. 41, Issue of October ’ 14. PP. 25905-25910,1994 Printed in U.S.A. Purification and Characterization of the 26 S Proteasome from Spinach Leaves” (Received for publication, February 15, 1994, and in revised form, May 31, 1994) Koji Fujinami, Nobuyuki Tanahashi$, Keiji Tanaka$, Akira IchiharaS,Zdenka CejkaQ, Wolfgang Eaumeisterg, Miyuki Miyawaki, Takahide Sato, and Hiroki Nakagawan From the D e~a~men~ of ~~cul~ura~ Chemis~~, Faculty of ~ort~~ulture, Chiba ~niversity, Matsudo, Chiba 271, Japan, the #Institute for Enzyme Research, University of Ilbkushima, Tokushima 770, Japan, and the §Max Planck fnstitute for Biochemistry, 82152 Martinsried, Germany The 26 S proteasome complex catalyzingATP-depend- ent breakdown of ubiquit~-ligated proteins was puri- fied &om spinach leaves to nearhomogeneity bychro- matography on DEAE-cellulose, gel filtration on Biogel A-1.5, and glycerol density gradient centrifugation. The purified enzyme was shown to degrade multi-ubiquiti- nated, but not unmodified, lysozymes in an ATP-depend- ent fashion coupled with ATPase activity supplying energy €or proteolysis and isopeptidase activity to gen- erate free ubiquitin. By nondenaturing electrophoresis, the purified enzyme was separated into two distinct forms of the 26 S complex, named 26 Sa and 26 SP pro- teasomes, with different electrophoretic mobilities. The 26 S proteasome was found to consist of multiple polypeptides with molecular masses of 2335 and 39-115 kDa, which were thought to be those of a 20 S protea- some with multicatalytic proteinase activity and an as- sociated regulatory part with ATPase and deubiquiti- nating activities, respectively. The subunit multiplicity of the spinach 26 S proteasome closely resembled that of rat liver with minor differences in certain components. No sulfhydryl bond was involved in the assembly of this multicomponent polypeptide complex, Electron micros- copy showed that the 26 S proteasome complex had a ‘‘caterpillar‘‘-like shape, consisting of four central pro- tein layers, assumed to be the 20 S proteasome, with asymmetric V-shaped layers at each end. These struc- tural and functional characteristics of the spinach 26 S proteasome showed marked similarity to those of the mammalian 26 S proteasomes reported recently, sug- gesting that the 26 S proteasome is widely distributed in eukaryotic cells and is of general importance for cata- lyzing the soluble energy- and ubiquitin-dependent pro- teolytic pathway. Two large multisubunit proteases are abundant constituents of the cytosol and nuclear compartments of eukaryotic cells (for reviews, see Hershko and Ciechanover (19921, Goldberg (1992), Tanaka et al. (19921, and Rechsteiner et al. (1993)). The smaller of the two is most often referred to as the multicatalytic pro- teinase complex (Orlowski, 1990; Rivett, 1993) or the 20 S proteasome (Goldberg, 1992; Tanaka et al., 1992). This 20 S proteasome has been isolated from a variety of organisms in- cluding higher plants (Ozaki et al. 1992; Skoda and Malek, Research (0430~004 and 05454014) from the Ministry of Education, * This workwassupported in part by Grants-in-aid for Scientific Science, and Culture of Japan. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. ll To whom correspondence should be addressed. 1992). The larger of the two, discovered by its ability to degrade ubiquitin (Ub)l-lysoz~e conjugates in an ATP-dependent fash- ion, is called the 26 S protease complex (Hough et at., 1987; Waxman et al., 1987) or the 26 S proteasome (Goldberg, 1992). The 20 S proteasome is known to associate with multiple pro- tein components ATP-dependently to form the 26 S protease complex (Eytan et al., 1989; Driscoll and Goldberg, 1990; Orino et al., 1991; Kanayama et al., 1992). The abundance and wide distribution of proteasomes and their ability to degrade protein substrates ATP dependently suggest that they play an important role in nonlysosomal path- ways of protein turnover (here and Hershko and Ciechnover, (1992)). Proteasomes function in both ATP-dependent and ATP- independent pathways of protein breakdown, and their relative contributions to these two pathways seem to be determined by the relative proportions of the 20 and 26 S protease complexes (Hershko and Ciechanover, 1992; Goldberg, 1992;Tanaka et al., 1992; Rechsteiner et al., 1993). The 26 S proteasome is believed to be responsible for ATP-dependent degradation of Ub-protein conjugates (Hershko and Ciechanover, 1992; Rechsteiner et al., 19931, but it also appears to be involved in the Ub-independent degradation of certain proteins, such as ornithine decarboxyl- ase, ATP dependently (Murakami et aE., 1992). The existence of the 26 S proteasome in plants was suggested by the detection of ATP-dependent activities that degrade ubiquitinated proteins in viuo (.Vierstra and Sullivan, 1988; Hatfield and Vierstra, 19891, but no 26 S complex has yet been isolated from plants. In this work, we purified a 26 S proteasome complex that rapidly degradedUb-ligated proteins in an ATP-dependent manner from spinach leaves and examined some properties of an inte- gral ATPase and a Ub-specific isopeptidase of this complex. We also examined the subunit structure of the purified complex and obtained direct evidence by electrophoretic and immuno- chemical ~ c ~ i q u e s that the 26 S proteasome from spinach leaves is a heterogeneous multipolypeptide complex, consisting of the 20 S proteasome (23-35 kDa) and a characteristic set of other nonidentical protein components (39-115 m a ) . Finally, we analyze thegross structure of the 26 S proteasome by elec- tron microscopy and discuss the similarity of its molecular properties with those of other eukaryotic 26 S proteasomes. EXPERIMENTAL PROCEDURES Muterials-The materials used were obtained as follows: Ub (Sigma); six times crystallized egg white lysozyme (Seikagaku Kogyo Co., Tokyo, Japan); NalZ5I (3.7 GBq/ml)and[y-3zPlATP(925GBq/mmol)(Amer- sham Gorp.); succinyl-Leu-Leu-Vai-Tyr-l-methylcoumaryl-7-amide (Suc-LLVY-MCA, Peptide Institute, Minoh, Japan); DE~-cellulose (DE52) (Whatman); Biogel A-1.5 m (Bio-Rad); and ATP (OrientalYeast, Osaka, Japan). ’ The abbreviations used are: Ub, ubiquitin; Suc-LLVY-MCA, succi- nyl-Leu-Leu-Val-Tyr-4-methylcoumaryl-7-amide; PAGE, polyacrylam- ide gel electrophoresis. - 25905

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOGRNAL OF BIOLOGICAL CHEHISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology , Inc
VOl. 269, No. 41, Issue of October ’ 14. PP. 25905-25910,1994 Printed in U.S.A.
Purification and Characterization of the 26 S Proteasome from Spinach Leaves”
(Received for publication, February 15, 1994, and in revised form, May 31, 1994)
Koji Fujinami, Nobuyuki Tanahashi$, Keiji Tanaka$, Akira IchiharaS, Zdenka CejkaQ, Wolfgang Eaumeisterg, Miyuki Miyawaki, Takahide Sato, and Hiroki Nakagawan From the D e ~ a ~ m e n ~ of ~ ~ c u l ~ u r a ~ C h e m i s ~ ~ , Faculty of ~ o r t ~ ~ u l t u r e , Chiba ~niversity, Matsudo, Chiba 271, Japan, the #Institute for Enzyme Research, University of Ilbkushima, Tokushima 770, Japan, and the §Max Planck fnstitute for Biochemistry, 82152 Martinsried, Germany
The 26 S proteasome complex catalyzing ATP-depend- ent breakdown of ubiquit~-ligated proteins was puri- fied &om spinach leaves to near homogeneity by chro- matography on DEAE-cellulose, gel filtration on Biogel A-1.5, and glycerol density gradient centrifugation. The purified enzyme was shown to degrade multi-ubiquiti- nated, but not unmodified, lysozymes in an ATP-depend- ent fashion coupled with ATPase activity supplying energy €or proteolysis and isopeptidase activity to gen- erate free ubiquitin. By nondenaturing electrophoresis, the purified enzyme was separated into two distinct forms of the 26 S complex, named 26 Sa and 26 SP pro- teasomes, with different electrophoretic mobilities. The 26 S proteasome was found to consist of multiple polypeptides with molecular masses of 2335 and 39-115 kDa, which were thought to be those of a 20 S protea- some with multicatalytic proteinase activity and an as- sociated regulatory part with ATPase and deubiquiti- nating activities, respectively. The subunit multiplicity of the spinach 26 S proteasome closely resembled that of rat liver with minor differences in certain components. No sulfhydryl bond was involved in the assembly of this multicomponent polypeptide complex, Electron micros- copy showed that the 26 S proteasome complex had a ‘‘caterpillar‘‘-like shape, consisting of four central pro- tein layers, assumed to be the 20 S proteasome, with asymmetric V-shaped layers at each end. These struc- tural and functional characteristics of the spinach 26 S proteasome showed marked similarity to those of the mammalian 26 S proteasomes reported recently, sug- gesting that the 26 S proteasome is widely distributed in eukaryotic cells and is of general importance for cata- lyzing the soluble energy- and ubiquitin-dependent pro- teolytic pathway.
Two large multisubunit proteases are abundant constituents of the cytosol and nuclear compartments of eukaryotic cells (for reviews, see Hershko and Ciechanover (19921, Goldberg (1992), Tanaka et al. (19921, and Rechsteiner et al. (1993)). The smaller of the two is most often referred to as the multicatalytic pro- teinase complex (Orlowski, 1990; Rivett, 1993) or the 20 S proteasome (Goldberg, 1992; Tanaka et al., 1992). This 20 S proteasome has been isolated from a variety of organisms in- cluding higher plants (Ozaki et al. 1992; Skoda and Malek,
Research (0430~004 and 05454014) from the Ministry of Education, * This work was supported in part by Grants-in-aid for Scientific
Science, and Culture of Japan. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. ll To whom correspondence should be addressed.
1992). The larger of the two, discovered by its ability to degrade ubiquitin (Ub) l - ly soz~e conjugates in an ATP-dependent fash- ion, is called the 26 S protease complex (Hough et at., 1987; Waxman et al., 1987) or the 26 S proteasome (Goldberg, 1992). The 20 S proteasome is known to associate with multiple pro- tein components ATP-dependently to form the 26 S protease complex (Eytan et al., 1989; Driscoll and Goldberg, 1990; Orino et al., 1991; Kanayama et al., 1992).
The abundance and wide distribution of proteasomes and their ability to degrade protein substrates ATP dependently suggest that they play an important role in nonlysosomal path- ways of protein turnover (here and Hershko and Ciechnover, (1992)). Proteasomes function in both ATP-dependent and ATP- independent pathways of protein breakdown, and their relative contributions to these two pathways seem to be determined by the relative proportions of the 20 and 26 S protease complexes (Hershko and Ciechanover, 1992; Goldberg, 1992; Tanaka et al., 1992; Rechsteiner et al., 1993). The 26 S proteasome is believed to be responsible for ATP-dependent degradation of Ub-protein conjugates (Hershko and Ciechanover, 1992; Rechsteiner et al., 19931, but it also appears to be involved in the Ub-independent degradation of certain proteins, such as ornithine decarboxyl- ase, ATP dependently (Murakami et aE., 1992). The existence of the 26 S proteasome in plants was suggested by the detection of ATP-dependent activities that degrade ubiquitinated proteins in viuo (.Vierstra and Sullivan, 1988; Hatfield and Vierstra, 19891, but no 26 S complex has yet been isolated from plants. In this work, we purified a 26 S proteasome complex that rapidly degraded Ub-ligated proteins in an ATP-dependent manner from spinach leaves and examined some properties of an inte- gral ATPase and a Ub-specific isopeptidase of this complex. We also examined the subunit structure of the purified complex and obtained direct evidence by electrophoretic and immuno- chemical ~ c ~ i q u e s that the 26 S proteasome from spinach leaves is a heterogeneous multipolypeptide complex, consisting of the 20 S proteasome (23-35 kDa) and a characteristic set of other nonidentical protein components (39-115 m a ) . Finally, we analyze the gross structure of the 26 S proteasome by elec- tron microscopy and discuss the similarity of its molecular properties with those of other eukaryotic 26 S proteasomes.
EXPERIMENTAL PROCEDURES Muterials-The materials used were obtained as follows: Ub (Sigma);
six times crystallized egg white lysozyme (Seikagaku Kogyo Co., Tokyo, Japan); NalZ5I (3.7 GBq/ml) and [y-3zPlATP (925 GBq/mmol) (Amer- sham Gorp.); succinyl-Leu-Leu-Vai-Tyr-l-methylcoumaryl-7-amide (Suc-LLVY-MCA, Peptide Institute, Minoh, Japan); DE~-cellulose (DE52) (Whatman); Biogel A-1.5 m (Bio-Rad); and ATP (Oriental Yeast, Osaka, Japan).
’ The abbreviations used are: U b , ubiquitin; Suc-LLVY-MCA, succi- nyl-Leu-Leu-Val-Tyr-4-methylcoumaryl-7-amide; PAGE, polyacrylam- ide gel electrophoresis.
-
25905
25906 26 S Proteasome from Spinach Leaves Plant Materials-Spinach (Spinacia oleracea L.) was obtained from a
local market, and cells of the leaves were cultured under continuous white light on Murashige and Skoog's medium as described (Nakagawa et al., 1985).
Assay of Peptidase Actiuity-The fluorogenic substrate SUC-LLVY- MCA was incubated with a test preparation for 10 min at 30 "C in the presence or absence of 0.02% SDS in 100 mM Tris-HC1 (pH 8.0), as described (Tanaka et al., 1988). The reaction was stopped by the addi- tion of 100 pl of 10% SDS and 2 ml of 100 mM Tris-HCI (pH 9.0), and the fluorescence of the reaction products was measured.
Assay of Protease Actiuity-About 10,000 cpm of '2sII-lysozyme-Ub was incubated at 37 "C for 60 min in a total volume of 100 pl of reaction mixture consisting of 50 mM "is-HCI (pH 8.0). 5 mM MgCI,, 2 mM ATP and an ATP-regenerating system (10 pg/ml of creatine kinase and 10 mM phosphocreatine), 1 mM dithiothreitol, and a suitable amount oftest preparation. Then, the conversion of l""Ilysozyme to acid-soluble frag- ments was measured as described (Kanayama et al., 1992). ATP- dependent breakdown of '2sI-lysozyme-Ub conjugates was calculated as the difference between the activities with or without M e . For exact measurement of the activity without M e , which was usually less than 1.5%/hour, 5 mM EDTA was added to the assay mixture. The methods for preparation of radiolabeled and Ub-lysozyme conjugates were as de- scribed (Tamura et al., 1991).
Electrophoretic Analysis-Polyacrylamide gel electrophoresis (PAGE) was carried out in 2.58 polyacrylamide gel containing 0.5% agarose under nondenaturing conditions. SDS-PAGE was carried out by the method of Laemmli (Laemmli, 1970) in 12.5% slab gel. Protein was detected by staining with Coomassie Brilliant Blue. Low molecular weight marker proteins (Pharmacia Biotech Inc.) were used for SDS- PAGE.
Immunological Analysis-A polyclonal antibody against the spinach 20 S proteasome was prepared as described (Ozaki et al., 1992). Immu- noelectrophoretic blot analysis was carried out by the method of Towbin et al. (1979). Samples were separated by PAGE and transferred elec- trophoretically to Immobilon polyvinylidene difluoride membranes (Millipore Corp.). Anti-rabbit IgG conjugated with alkaline phosphatase was used as a second antibody with nitro blue tetrazolium and 5-bromo- 4-chloro-3-indolyl phosphate as a substrate of alkaline phosphatase.
Analysis of Cultured Spinach Cell Extract by Glycerol Density Gra- dient Centrifugation-Cultured spinach cells (1.0 g) were homogenized with buffer A (50 mM Tris-HC1 (pH 7.5) containing 2 mM ATP, 5 mM MgCI,, and 10 mM 2-mercaptoethanol) or buffer B (buffer A without ATP). The homogenates were centrifuged a t 15,000 x g for 30 min, and the resulting supernatants were used as crude cell extracts. These cell extracts (2 mg of protein) were loaded onto linear gradients of 2040% (v/v) glycerol in buffer A or buffer B and centrifuged at 25,000 rpm for 22 h in a Hitachi SRP28SA1 rotor. They were then collected in 30 fractions of 1 ml each a t 4 "C.
Electron Microscopy and Image Processing-Details of the methods used for electron microscopy were described elsewhere (Yoshimura et al., 1994). Briefly, the purified enzymes were negatively stained on grids with 2% uranyl acetate and examined in a Philips EM420 electron microscope a t a magnification of ~36 ,000 . Suitable areas of the micro- graphs were digitized with an EIKONIX microdensitometer. Approxi- mately 450 images of the double-ended and of the one-ended form of the 26 S proteasome complex were extracted interactively. The whole data set was subjected to an eigenvector-eigenvalue classification procedure (Van Heel and Frank, 1981; Frank and Van Heel, 1982) as implemented in the EM and SEMPER program systems (Saxton et al., 1979; Hegerl and Altbauer, 1982) to detect significant interimage variations.
Other Biochemical Analyses-ATPase activity was assayed as de- scribed by Armon et al. (1990) with ly-"2PIATP. Protein concentration was measured by the method of Bradford (1976) with bovine serum albumin as a standard.
RESULTS
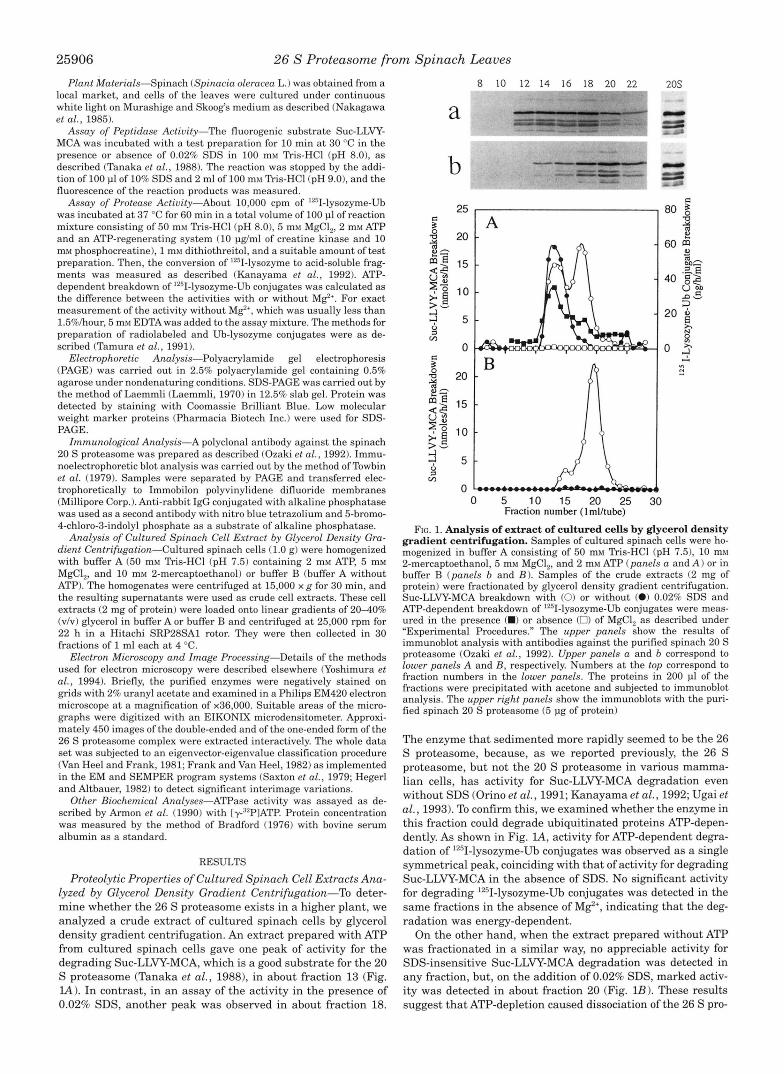
Proteolytic Properties of Cultured Spinach Cell Extracts Ana- lyzed by Glycerol Density Gradient Centrifugation-To deter- mine whether the 26 S proteasome exists in a higher plant, we analyzed a crude extract of cultured spinach cells by glycerol density gradient centrifugation. An extract prepared with ATP from cultured spinach cells gave one peak of activity for the degrading Suc-LLVY-MCA, which is a good substrate for the 20 S proteasome (Tanaka et al., 1988), in about fraction 13 (Fig. lA). In contrast, in an assay of the activity in the presence of 0.02% SDS, another peak was observed in about fraction 18.
8 10 12 14 16 18 20 22
a
b 25
l4 0 5 10 15 20 25 30
Fraction number ( 1 mlltube)
FIG. 1. Analysis of extract of cultured cells by glycerol density gradient centrifugation. Samples of cultured spinach cells were ho- mogenized in buffer A consisting of 50 mM Tris-HC1 (pH 7.5), 10 mM 2-mercaptoethanol, 5 mM MgCI,, and 2 mM ATP (panels a and A) or in buffer B (panels b and B). Samples of the crude extracts (2 mg of protein) were fractionated by glycerol density gradient centrifugation. Suc-LLVY-MCA breakdown with (0) or without (0) 0.02% SDS and ATP-dependent breakdown of '2"I-lysozyme-Ub conjugates were meas- ured in the presence (W) or absence (0) of MgCI, as described under "Experimental Procedures." The upper panels show the results of immunoblot analysis with antibodies against the purified spinach 20 S proteasome (Ozaki et al., 1992). Upper panels a and b correspond to lower panels A and B , respectively. Numbers at the top correspond to fraction numbers in the lower panels. The proteins in 200 pl of the fractions were precipitated with acetone and subjected to immunoblot analysis. The upper right panels show the immunoblots with the puri- fied spinach 20 S proteasome (5 pg of protein)
The enzyme that sedimented more rapidly seemed to be the 26 S proteasome, because, as we reported previously, the 26 S proteasome, but not the 20 S proteasome in various mamma- lian cells, has activity for Suc-LLVY-MCA degradation even without SDS (Orino et al., 1991; Kanayama et al., 1992; Ugai et al., 1993). To confirm this, we examined whether the enzyme in this fraction could degrade ubiquitinated proteins ATP-depen- dently. As shown in Fig. l A , activity for ATP-dependent degra- dation of 1251-lysozyme-Ub conjugates was observed as a single symmetrical peak, coinciding with that of activity for degrading Suc-LLVY-MCA in the absence of SDS. No significant activity for degrading 1251-lysozyme-Ub conjugates was detected in the same fractions in the absence of M e , indicating that the deg- radation was energy-dependent.
On the other hand, when the extract prepared without ATP was fractionated in a similar way, no appreciable activity for SDS-insensitive Suc-LLVY-MCA degradation was detected in any fraction, but, on the addition of 0.02% SDS, marked activ- ity was detected in about fraction 20 (Fig. lB). These results suggest that ATP-depletion caused dissociation of the 26 S pro-
26 S Proteasome from Spinach Leaves
teasome complex liberating a latent 20 S proteasome. Interest- ingly, the SDS-activated putative 20 S enzyme in Fig. lA is larger than that in Fig. lB, which is apparently the same size as the purified 20 S proteasome, suggesting that the material with latent activity seen in Fig. IA consisted of the 20 S pro- teasome with additional associated components.
Next, by immunoblot analysis with antibodies against the 20 S proteasome (Ozaki et al., 1992) we examined whether the fractions of the 26 S proteasome and SDS-activated enzyme shown in Fig. IA actually contained the 20 S proteasome. Poly- clonal antibodies against the 20 S proteasome of spinach leaves reacted with several components in fractions 12-22, showing a similar pattern on immunostaining to that of the purified spin- ach 20 S proteasome (Fig. la), and, thus, confirming the pres- ence of the 20 S proteasome in fractions 12-22. Furthermore, in the ATP-depleted extract, anti-20 S proteasome antibodies mainly reacted with proteins recovered in about fraction 20 (Fig. lb). The patterns of these immunostainings resembled those obtained with the purified spinach 20 S proteasome (up- per right panels). These proteolytic properties of the spinach cell extract resembled those in human promyelocytic leukemia HL-60 cells reported previously (Orino et al., 1991). These re- sults indicate that the 26 S proteasome, which catalyzes ATP- dependent degradation of ubiquitinated proteins, is present in cultured spinach cells.
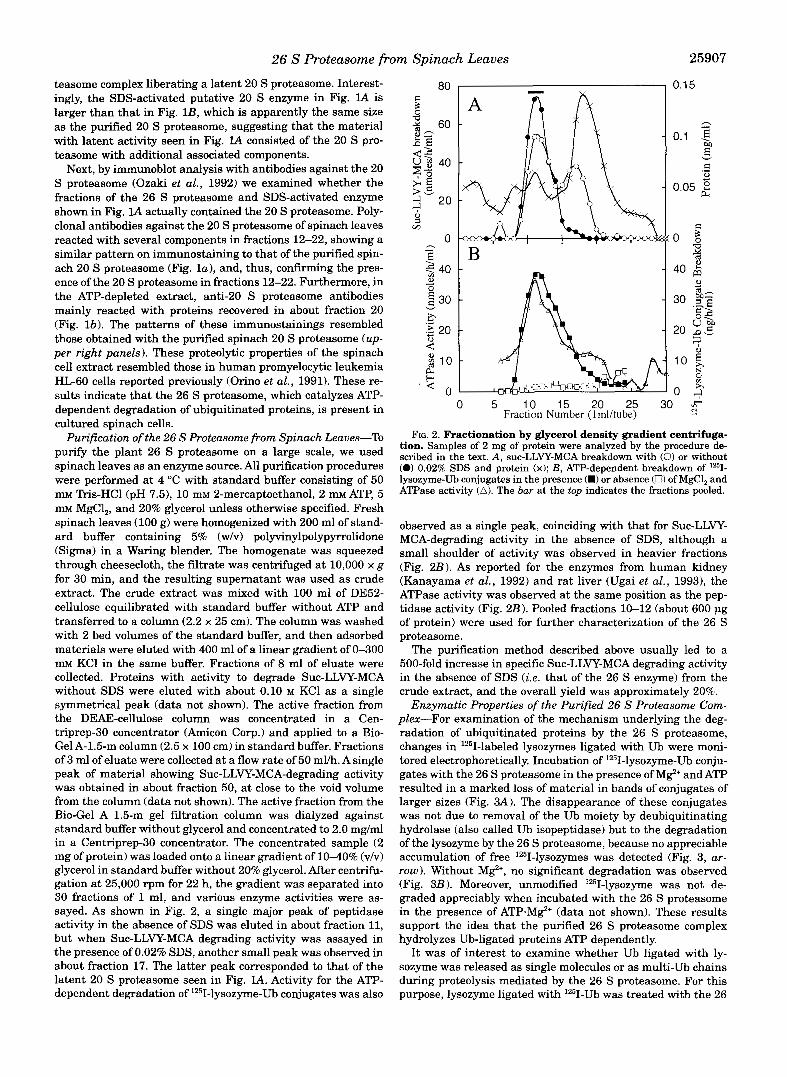
Purification of the 26 S Proteasome from Spinach Leaves-To purify the plant 26 S proteasome on a large scale, we used spinach leaves as an enzyme source. All purification procedures were performed at 4 "C with standard buffer consisting of 50 mM "is-HC1 (pH 7.51, 10 mM 2-mercaptoethanol, 2 m ATP, 5 mM MgCl,, and 20% glycerol unless otherwise specified. Fresh spinach leaves (100 g) were homogenized with 200 ml of stand- ard buffer containing 5% (w/v) polyvinylpolypyrrolidone (Sigma) in a Waring blender. The homogenate was squeezed through cheesecloth, the filtrate was centrifuged at 10,000 x g for 30 min, and the resulting supernatant was used as crude extract. The crude extract was mixed with 100 ml of DE52- cellulose equilibrated with standard buffer without ATP and transferred to a column (2.2 x 25 cm). The column was washed with 2 bed volumes of the standard buffer, and then adsorbed materials were eluted with 400 ml of a linear gradient of 0-300 mM KC1 in the same buffer. Fractions of 8 ml of eluate were collected. Proteins with activity to degrade Suc-LLVY-MCA without SDS were eluted with about 0.10 M KC1 as a single symmetrical peak (data not shown). The active fraction from the DEAE-cellulose column was concentrated in a Cen- triprep-30 concentrator (Amicon Corp.) and applied to a Bio- Gel A-1.5-m column (2.5 x 100 cm) in standard buffer. Fractions of 3 ml of eluate were collected at a flow rate of 50 ml/h. A single peak of material showing Suc-LLVY-MCA-degrading activity was obtained in about fraction 50, at close to the void volume from the column (data not shown). The active fraction from the Bio-Gel A 1.5-m gel filtration column was dialyzed against standard buffer without glycerol and concentrated to 2.0 mg/ml in a Centriprep-30 concentrator. The concentrated sample (2 mg of protein) was loaded onto a linear gradient of 10-40% (v/v) glycerol in standard buffer without 20% glycerol. After centrifu- gation a t 25,000 rpm for 22 h, the gradient was separated into 30 fractions of 1 ml, and various enzyme activities were as- sayed. As shown in Fig. 2, a single major peak of peptidase activity in the absence of SDS was eluted in about fraction 11, but when Suc-LLVY-MCA degrading activity was assayed in the presence of 0.02% SDS, another small peak was observed in about fraction 17. The latter peak corresponded to that of the latent 20 S proteasome seen in Fig. lA. Activity for the ATP- dependent degradation of 1251-lysozyme-Ub conjugates was also
B
0 5 1.0 15 20 25 30 mm Fractlon N u m b e r ( l m l h b e ) N
FIG. 2. Fractionation by glycerol density gradient centrifuga- tion. Samples of 2 mg of protein were analyzed by the procedure de- scribed in the text. A, suc-LLVY-MCA breakdown with (0) or without (0) 0.02% SDS and protein (x); B, ATP-dependent breakdown of 1251- lysozyme-Ub conjugates in the presence (.) or absence (0) of MgC1, and ATPase activity (A). The bar at the top indicates the fractions pooled.
observed as a single peak, coinciding with that for Suc-LLVY- MCA-degrading activity in the absence of SDS, although a small shoulder of activity was observed in heavier fractions (Fig. 2 B ) . As reported for the enzymes from human kidney (Kanayama et al., 1992) and rat liver (Ugai et al., 19931, the ATPase activity was observed at the same position as the pep- tidase activity (Fig. 2B). Pooled fractions 10-12 (about 600 pg of protein) were used for further characterization of the 26 S proteasome.
The purification method described above usually led to a 500-fold increase in specific Suc-LLVY-MCA degrading activity in the absence of SDS (ie. that of the 26 S enzyme) from the crude extract, and the overall yield was approximately 20%.
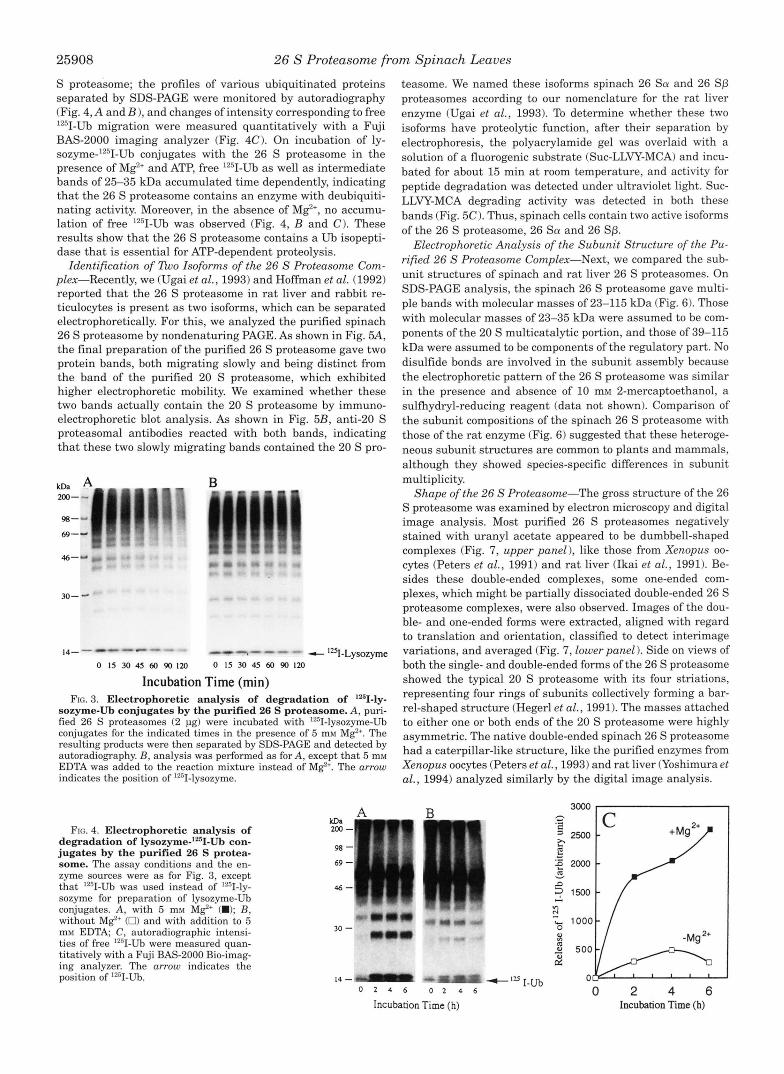
Enzymatic Properties of the Purified 26 S Proteasome Com- plex-For examination of the mechanism underlying the deg- radation of ubiquitinated proteins by the 26 S proteasome, changes in lz5I-1abeled lysozymes ligated with Ub were moni- tored electrophoretically. Incubation of '251-lysozyme-Ub conju- gates with the 26 S proteasome in the presence of Mg2' and ATP resulted in a marked loss of material in bands of conjugates of larger sizes (Fig. 3A). The disappearance of these conjugates was not due to removal of the Ub moiety by deubiquitinating hydrolase (also called Ub isopeptidase) but to the degradation of the lysozyme by the 26 S proteasome, because no appreciable accumulation of free 1251-lysozymes was detected (Fig. 3, ar- row). Without M e , no significant degradation was observed (Fig. 3B). Moreover, unmodified 1251-lysozyme was not de- graded appreciably when incubated with the 26 S proteasome in the presence of ATP.Mg2f (data not shown). These results support the idea that the purified 26 S proteasome complex hydrolyzes Ub-ligated proteins ATP dependently.
I t was of interest to examine whether Ub ligated with ly- sozyme was released as single molecules or as multi-Ub chains during proteolysis mediated by the 26 S proteasome. For this purpose, lysozyme ligated with lZ5I-LJb was treated with the 26
25908 26 S Proteasome from Spinach Leaves
S proteasome; the profiles of various ubiquitinated proteins separated by SDS-PAGE were monitored by autoradiography (Fig. 4, A and B ) , and changes of intensity corresponding to free "'I-Ub migration were measured quantitatively with a Fuji BAS-2000 imaging analyzer (Fig. 4C). On incubation of ly- sozyme-'2sI-Ub conjugates with the 26 S proteasome in the presence of Mg2' and ATP, free 1251-Ub as well as intermediate bands of 25-35 kDa accumulated time dependently, indicating that the 26 S proteasome contains an enzyme with deubiquiti- nating activity. Moreover, in the absence of M e , no accumu- lation of free '*'I-Ub was observed (Fig. 4, B and C). These results show that the 26 S proteasome contains a Ub isopepti- dase that is essential for ATP-dependent proteolysis.
Identification of llno Isoforms of the 26 S Proteasome Com- plex-Recently, we (Ugai et al., 1993) and Hoffman et al. (1992) reported that the 26 S proteasome in rat liver and rabbit re- ticulocytes is present as two isoforms, which can be separated electrophoretically. For this, we analyzed the purified spinach 26 S proteasome by nondenaturing PAGE. As shown in Fig. 5A, the final preparation of the purified 26 S proteasome gave two protein bands, both migrating slowly and being distinct from the band of the purified 20 S proteasome, which exhibited higher electrophoretic mobility. We examined whether these two bands actually contain the 20 S proteasome by immuno- electrophoretic blot analysis. As shown in Fig. 5B, anti-20 S proteasomal antibodies reacted with both bands, indicating that these two slowly migrating bands contained the 20 S pro-
30-
14"""" . "" - - - 0 15 30 45 60 90 I 2 0 0 15 30 45 60 90 120
- '251-Lysozyme
Incubation Time (min) FIG. 3. Electrophoretic analysis of degradation of '*51-ly-
sozyme-Ub conjugates by the purified 26 S proteasome. A, puri- fied 26 S proteasomes (2 pg) were incubated with '""I-lysozyme-Ub conjugates for the indicated times in the presence of 5 mM Mg2'. The resulting products were then separated by SDS-PAGE and detected by autoradiography. B, analysis was performed as for A, except that 5 m M EDTA was added to the reaction mixture instead of M e . The arrow indicates the position of lZsII-lysozyme.
FIG. 4. Electrophoretic analysis of degradation of lysozyme-'2sII-Ub con- jugates by the purified 26 S protea- some. The assay conditions and the en- zyme sources were as for Fig. 3, except that 12sI-Ub was used instead of '"I-ly- sozyme for preparation of lysozyme-Ub conjugates. A, with 5 mM Mg"' (W); B, without MgZ' (0) and with addition to 5 mM EDTA, C, autoradiographic intensi- ties of free 1z31-Ub were measured quan- titatively with a Fuji BAS-2000 Bio-imag- ing analyzer. The arrow indicates the position of 1251-Ub.
teasome. We named these isoforms spinach 26 Sa and 26 Sp proteasomes according to our nomenclature for the rat liver enzyme (Ugai et al., 1993). To determine whether these two isoforms have proteolytic function, after their separation by electrophoresis, the polyacrylamide gel was overlaid with a solution of a fluorogenic substrate (Suc-LLVY-MCA) and incu- bated for about 15 min at room temperature, and activity for peptide degradation was detected under ultraviolet light. Suc- LLVY-MCA degrading activity was detected in both these bands (Fig. 5 0 . Thus, spinach cells contain two active isoforms of the 26 S proteasome, 26 Sa and 26 Sp.
Electrophoretic Analysis of the Subunit Structure of the Pu- rified 26 S Proteasome Complex-Next, we compared the sub- unit structures of spinach and rat liver 26 S proteasomes. On SDS-PAGE analysis, the spinach 26 S proteasome gave multi- ple bands with molecular masses of 23-115 kDa (Fig. 6). Those with molecular masses of 23-35 kDa were assumed to be com- ponents of the 20 S multicatalytic portion, and those of 39-115 kDa were assumed to be components of the regulatory part. No disulfide bonds are involved in the subunit assembly because the electrophoretic pattern of the 26 S proteasome was similar in the presence and absence of 10 mM 2-mercaptoethanol, a sulfhydryl-reducing reagent (data not shown). Comparison of the subunit compositions of the spinach 26 S proteasome with those of the rat enzyme (Fig. 6) suggested that these heteroge- neous subunit structures are common to plants and mammals, although they showed species-specific differences in subunit multiplicity.
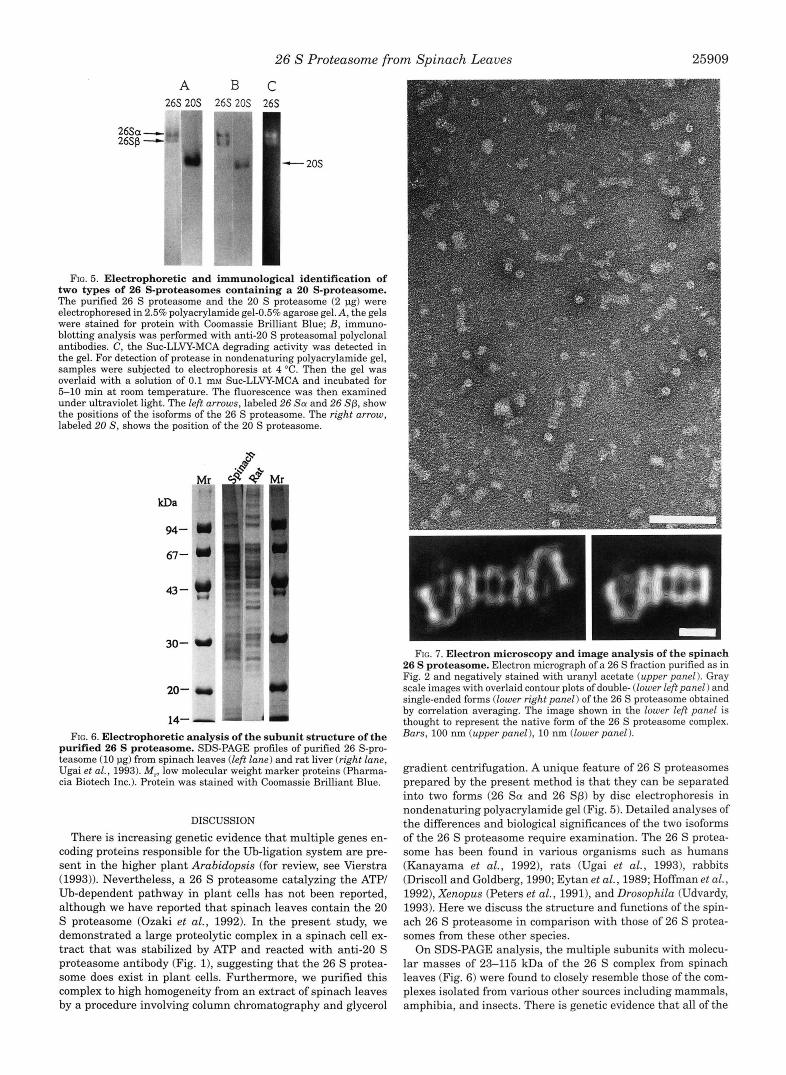
Shape of the 26 S Proteasome-The gross structure of the 26 S proteasome was examined by electron microscopy and digital image analysis. Most purified 26 S proteasomes negatively stained with uranyl acetate appeared to be dumbbell-shaped complexes (Fig. 7, upper panel 1, like those from Xenopus oo- cytes (Peters et al., 1991) and rat liver (Ikai et al., 1991). Be- sides these double-ended complexes, some one-ended com- plexes, which might be partially dissociated double-ended 26 S proteasome complexes, were also observed. Images of the dou- ble- and one-ended forms were extracted, aligned with regard to translation and orientation, classified to detect interimage variations, and averaged (Fig. 7, lower panel 1. Side on views of both the single- and double-ended forms of the 26 S proteasome showed the typical 20 S proteasome with its four striations, representing four rings of subunits collectively forming a bar- rel-shaped structure (Hegerl et al., 1991). The masses attached to either one or both ends of the 20 S proteasome were highly asymmetric. The native double-ended spinach 26 S proteasome had a caterpillar-like structure, like the purified enzymes from Xenopus ooc-ytes (Peters et al., 1993) and rat liver (Yoshimura et al., 1994) analyzed similarly by the digital image analysis.
30 - 2 I /
14 - I-Ub 0 1 4 6 0 1 4 6 0 2 4 6
IncubationTime (h) Incubation Time (h)
26 S Proteasome from Spinach Leaves 25909
A B C
m q 26s 20s 26s 20s 26s
2fiSa-,. 26sp - - 20s
Ffc. 5. Electrophoretic and immunological identification of two types of 26 S-proteasomes containing a 20 S-proteasome. The purified 26 S proteasome and the 20 S proteasome (2 pg) were electrophoresed in 2.5% polyacrylamide gel-0.5% agarose gel. A, the gels were stained for protein with Coomassie Brilliant Blue; B, immuno- blotting analysis was performed with anti-20 S proteasomal polyclonal antibodies. C, the Suc-LLVY-MCA degrading activity was detected in the gel. For detection of protease in nondenaturing polyacrylamide gel, samples were subjected to electrophoresis at 4 "C. Then the gel was overlaid with a solution of 0.1 mM Suc-LLVY-MCA and incubated for 5-10 min a t room temperature. The fluorescence was then examined under ultraviolet light. The left arrows, labeled 26 Sa and 26 Sp, show the positions of the isoforms of the 26 S proteasome. The right arrow, labeled 20 S, shows the position of the 20 S proteasome.
20- r
FIG. 6. Electrophoretic analysis of the subunit structure of the purified 26 S proteasome. SDS-PAGE profiles of purified 26 S-pro- teasome (10 1.18) from spinach leaves (left lane) and rat liver (right lane, Ugai et al., 1993). M,, low molecular weight marker proteins (Pharma- cia Biotech Inc.). Protein was stained with Coomassie Brilliant Blue.
DISCUSSION There is increasing genetic evidence that multiple genes en-
coding proteins responsible for the Ub-ligation system are pre- sent in the higher plant Arabidopsis (for review, see Vierstra (1993)). Nevertheless, a 26 S proteasome catalyzing the ATP/ Ub-dependent pathway in plant cells has not been reported, although we have reported that spinach leaves contain the 20 S proteasome (Ozaki et al., 1992). In the present study, we demonstrated a large proteolytic complex in a spinach cell ex- tract that was stabilized by ATP and reacted with anti-20 S proteasome antibody (Fig. l ) , suggesting that the 26 S protea- some does exist in plant cells. Furthermore, we purified this complex to high homogeneity from an extract of spinach leaves by a procedure involving column chromatography and glycerol
FIG. 7. Electron microscopy and image analysis of the spinach 26 S proteasome. Electron micrograph of a 26 S fraction purified as in Fig. 2 and negatively stained with uranyl acetate (upper panel 1. Gray scale images with overlaid contour plots of double- (lower left panel) and single-ended forms (lower right panel) of the 26 S proteasome obtained by correlation averaging. The image shown in the lower left panel is thought to represent the native form of the 26 S proteasome complex. Bars, 100 nm (upper panel ), 10 nm (lower panel).
gradient centrifugation. A unique feature of 26 S proteasomes prepared by the present method is that they can be separated into two forms (26 Sa and 26 Sp) by disc electrophoresis in nondenaturing polyacrylamide gel (Fig. 5). Detailed analyses of the differences and biological significances of the two isoforms of the 26 S proteasome require examination. The 26 S protea- some has been found in various organisms such as humans (Kanayama et al., 1992), rats (Ugai et al., 19931, rabbits (Driscoll and Goldberg, 1990; Eytan et al., 1989; Hoffman et al., 1992), Xenopus (Peters et al., 19911, and Drosophila (Udvardy, 1993). Here we discuss the structure and functions of the spin- ach 26 S proteasome in comparison with those of 26 S protea- somes from these other species.
On SDS-PAGE analysis, the multiple subunits with molecu- lar masses of 23-115 kDa of the 26 S complex from spinach leaves (Fig. 6) were found to closely resemble those of the com- plexes isolated from various other sources including mammals, amphibia, and insects. There is genetic evidence that all of the

25910 26 S Proteasome from Spinach Leaves
genes involved in encoding 20 S proteasomes have been highly conserved during evolution (Tanaka et al., 1992). Moreover, most of the multiple distinct polypeptides ranging from 39 to over 115 kDa that are associated with the 20 S proteasome to form the 26 S proteasome (Ugai et al., 1993) are also similar in size in 26 S proteasomes from various sources (Fig. 6), indicat- ing that the 26 S proteasome is a ubiquitous complex of mul- tiple, nonidentical components. However the mechanism of as- sembly of the multiple proteasome subunits is unknown.
We found by electron microscopy in conjunction with digital image analysis that the “26 S” proteasome complex purified from spinach leaves is caterpillar-shaped with two large V- or U-like terminal complexes in opposite orientation attached to a smaller, four-layered central structure (Fig. 7). Similar ultra- structures of 26 S proteasome complexes from Xenopus and rat liver were reported by Peters et al. (1993) and Yoshimura et al. (19941, respectively. In previous studies, we showed that the 20 S proteasome from a higher plant (Ozaki et al., 1992) was similar in shape to various mammalian 20 S proteasomes (Tanaka et al., 1988). Thus, the gross structures of 20 and 26 S proteasomes have been conserved in eukaryotes during evolu- tion, and the conserved configuration may indicate its critical role in the functions of this complex protease.
The purified 26 S proteasome complex showed similar enzy- matic properties to the enzyme complexes from other sources. For instance, it rapidly degraded 1261-lysozyme-Ub conjugates in an ATP-dependent fashion (Figs. 2 and 3), suggested that covalent modification of substrate proteins through ubiqutina- tion is essential for their proteolysis by the 26 S proteasome. Thus, the plant 26 S proteasome may recognize multiubiquitin chains bound to proteins for their specific breakdown. However, details of the mechanisms underlying these processes are un- known. In addition, it had intrinsic ATPase activity. This newly identified ATPase seems to be an essential component of the 26 S complex for catalyzing the ATP-dependent breakdown of Ub- conjugated proteins. Presumably the role of the ATPase is to supply energy continuously for the selective degradation of ubiquitinated proteins by the active 26 S complex. Moreover, recently multiple ATPase components were found to be associ- ated with the 26 S proteasome, and these putative ATPases are suggested to operate as regulatory subunits by specifically in- teracting with different substrates (Rechsteiner et al., 1993). However, further characterization of the 26 S proteasome com- plex as an ATP-dependent protease is necessary to clarify the role of energy requirement at the molecular level. In the pre- sent work, we also found that the purified 26 S proteasome complex contains an isopeptidase whose activity is closely as- sociated with the proteolytic reaction, as judged by the Mg2‘ requirement for its action (Fig. 4). Recently Ub-isopeptidase was reported to be associated with 26 S proteasomes from rab- bit reticulocytes (Eytan et al., 1993) and rat liver (Ugai et al., 1993). Thus, this activity may also be common to all 26 S proteasomes.
A remarkable finding in this work was that the structure and properties of 26 S proteasomes are very similar in a plant to those in other organisms ranging from Drosophila to man. These facts indicate that the 26 S proteasome must have been conserved evolutionally and play important roles in living or- ganisms, such as in cell cycle progression. Studies on the amino acid sequences of the subunits of plant proteasomes would pro- vide information on the evolutional tree of this complex.
REFERENCES
Armon, T., Ganoth, D., and Hershko, A. (1990) J. B i d . Chem. 265,20723-20726 Bradford, M. M. (1976) Anal. Biochem. 72, 248-254 Driscoll, J., and Goldberg, A. L. (1990) J. Biol. Chem. 265, 47894792 Eytan, E., Ganoth, D., Arnon, T., and Hershko, A. (1989) Proc. Natl. Acad. Sci.
U. S. A. 86. 7751-7755 Eytan, E., &on, T., Heller, H., Beck, S., and Hershko, A. (1993) J. Biol. Chem.
268.46684674 Frank,’J., and Van Heel, M. (1982) J. Mol. B i d . 161, 1134-1137 Goldberg, A. L. (1992) Eur J. Biochem. 205, 9-23 Hatfield, P. M., and Vierstra, R. D. (1989) Biochemistry 28, 735-742 Hegerl, R., and Altbauer, A. (1982) Ultramicroscopy 9, 109-116 Hegerl, R., Pfeifer, G., Piihler, G., Dahlmann, B., and Baumeister, W. (1991) FEBS
Hershko, A,, and Ciechanover, A. (1992) Annu. Rev. Biochem. 61, 761-807 Hoffman, L., Pratt, G., and Rechsteiner, M. (1992) J. Biol. Chem. 267, 22362-
Hough, R., Pratt, G., and Rechsteiner, M. (1987) J. Bid . Chem. 262, 8303-8313 Ikai, A., Nishigai, M., Tanaka, K., and Ichihara, A. (1991) FEBS Lett. 292, 21-24 Kanayama, H., Tamura, T., Ugai, S., Kagawa, S., Tanahashi, N., Yoshimura, T.,
Laemmli, U. K. (1970) Nature 227, 680485 Murakami, Y., Matsufuji, S., Kameji, T., Hayashi, S., Igarashi, K., Tamura, T.,
Nakagawa, H., Tanaka, H., Oba, T., Ogura, N., and Iizuka, M. (1985) Plant Cell
Orino, E., Tanaka, K., Tamura, T., Sone, S., Ogura, T., and Ichihara, A. (1991)
Orlowski, M. (1990) Biochemistry 29, 10289-10297 Ozaki, M., Fujinami, K., Tanaka, K., Amemiya, Y., Sato, T., Ogura, N., and Naka-
Peters, J.-M., Hams, J. R., and Kleinschmidt, J. A. (1991) Eur J. Cell Biol. 56,
Peters, J.-M., Cejka, Z., Hams, J. R., Kleinschmidt, J. A., and Baumeister, W.
Rivett, A. J. (1993) Biochem. J. 291, 1-10 Rechsteiner, M., Hoffman, L., and Dubiel, W. (1993) J. B i d . Chem. 268,6065-6068
Saxton, W. O., Pitt, T. J., and Horner, M. (1979) Ultramicroscopy 4, 343-354
Tamura, T., Tanaka, K., Tanahashi, N., and Ichihara, A. (1991) FEBS Lett. 292, Skoda, B., and Malek, L. (1992) Plant Physiol. 99,1515-1519
Tanaka, K., Yoshimura, T., Kumatori, A,, Ichihara, A,, Ikai, A,, Nishigai, M.,
Tanaka, K., Tamura, T., Yoshimura, T., and Ichihara, A. (1992) New Biol. 4,173-
Towbin, H. S., Staehelin, J., and Gordon, J. (1979) Proc. Natl. Acad. Sci. U. S. A.
Udvardy, A. (1993) J. Biol. Chem. 268,9055-9062 Ugai, S., Tamura, T., Tanahashi, N., Takai, S., Komi, N., Chung, c.-H., Tanaka, K.,
and Ichihara, A. (1993) J. Biochem. (Tokyo) 113,754-768
Vlerstra, R. D. (1993) Annu. Rev. Plant Physiol. Plant Mol. B i d . 44, 385-410 Van Heel, M., and Frank, J. (1981) Ultramicroscopy 6, 187-194
Vierstra, R. D., and Sullivan, M. L. (1988) Biochemistry 27, 3290-3295 Waxman, L., Fagan, J. M., and Goldberg, A. L. (1987) J. Biol. Chem. 262, 2451-
2457 Yoshimura, T., Kameyama, K., Takagi, T., Ikai, A,, Tokunaga, F., Koide, T., Tana-
hashi, N., Tamura, T., Cejka, Z., Baumeister, W., Tanaka, K., and Ichihara, A. (1993) J. Struct. B i d . 111, 200-211
Lett. 283, 117-121
22368
Tanaka, K., and Ichihara, A. (1992) Eur J . Biochem. 206, 567-578
Tanaka, K., and Ichihara, A. (1992) Nature 360,597-599
Rep. 4, 148-150
FEBS Lett. 284,206-210
gawa, H. (1992) J. Biol. Chem. 267,21678-21684
422-432
(1993) J. Mol. B i d . 234, 932-937
154-158
Kameyama, K., and Takagi, T. (1988) J. Biol. Chem. 263, 16209-16217
187
76,435M354
Related Documents