THE JOURNAL OF BI~L~CICAL CHEMISTRY (~81990 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 265, No. 10, Issue of April 5, pp. 5460-5465, 1990 Printed in U.S. A. The Nuclear Matrix from Cells of Different Origin EVIDENCE FOR A COMMON SET OF MATRIX PROTEINS* (Received for publication, April 17, 1989) Nice Stuurman, Alexandra M. L. Meijne, Arnoud J. van der Pol, Luitzen de Jong, Roe1 van Driel, and Jos van RenswoudeS From the E. C. Slater Institute for Biochemical Research, University of Amsterdam, Past office Box 20151, Amsterdam 1000 HD, The Netherlands We compared the protein composition of the nuclear matrix isolated from several murine embryonal carci- noma cells and mature tissues by two-dimensional gel electrophoresis. Two nuclear matrix fractions were investigated: the “peripheral” nuclear matrix (matrix proteins that remain insoluble after reduction), and the “internal” nuclear matrix (matrix proteins released by reduction). The two subfractions have completely dif- ferent protein compositions. Although numerous dif- ferences in nuclear matrix protein composition among different cell types were observed, a limited set of polypeptides common to all mouse cell types was iden- tified. A majority of these common proteins was also present in cells from other mammalian species (i.e. rat and human). For this set of proteins, we coin the term “minimal matrix.” As expected, lamin B, known to be expressed throughout differentiation, is part of the common set of peripheral nuclear matrix proteins. Lamins A and C are not because these proteins were absent from undif- ferentiated embryonal carcinoma cells. Since these common nuclear matrix proteins occur in all mamma- lian nuclear matrices analyzed so far, it is likely that they have a basic role in nuclear organization and function. Recently, several authors have expanded on the idea that a proteinaceous network in the nucleus (the nuclear matrix) is involved in control and coordination of gene expression during differentiation (Blobel, 1985; Razin, 1987; Bodnar, 1988). It is thought that loops of chromatin are persistently as well as transiently attached to this network (Pienta and Coffey, 1984; Mirkovitch et al., 1984; Gassar and Laemmli (1987); Earn- shaw, 1988). It has been proposed that each chromatin loop constitutes an independent regulatory unit containing several genes. Processes such as DNA replication (Jackson et al., 1984; Dijkwel et al., 1986; Razin et al., 1986; Tubo and Mar- telli, 1987; Nakayasu and Berezney, 1989), transcription (Jackson and Cook, 1985; Razin et al., 1985; Buttyan and Olsson, 1986), RNA processing and transport (Schroder et al., 1987a, 198713; Zeitlin et al., 1987), and the regulation of DNA superhelicity (Berrios et al., 1985) have been shown to be associated with this nuclear matrix. These observations sup- port the hypothesis that the nuclear matrix is instrumental * This study was financed in part by a grant from The Netherlands Execut,ive Agency for the Stimulation of Computer Science (SPIN). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ To whom correspondence should be sent. Fax: 20-525 5124. in regulation and coordination of gene expression. Little is known about the nature of the proteins that constitute the nuclear matrix. It seems likely that the nuclear matrix has a similar basal structure in almost all mammalian cells. Therefore, we expect that the elements essential to this structure will tend to be conserved during evolution and remain present throughout differentiation. In addition, nu- merous differentiation state- and species-specific components will be present in each nuclear matrix. Recently, we and others (Fey and Penman, 1987, 1988; Stuurman et al., 1989) have shown that the protein composi- tion of the nuclear matrix is differentiation state dependent. Here, we present a detailed analysis of the matrix composition determined by two-dimensional gel electrophoresis (O’Farrell, 1975; O’Farrell et al., 1977). Nuclear matrices were prepared by extraction of nuclease-treated purified nuclei with buffers of high ionic strength. Two matrix subfractions were isolated, one enriched in nuclear envelope polypeptides (peripheral matrix), the other containing mainly internal matrix proteins (internal matrix) (Kaufmann and Shaper, 1984). We have looked for nuclear matrix polypeptides that are present in a variety of murine cell types and tissues and in cells that originate from other mammals (rat and human). The results show that a limited set of proteins is present in the nuclear matrix of all cell types studied, irrespective of species and differentiation state. We propose that these proteins are es- sential in constituting the nuclear matrix and coin the term minimal matrix for them. MATERIALS AND METHODS Cell Culture P19EC (MC Burney et al., 1982), PISMES-1 (Mummery et al., 1986), P19EPI-7, and P19END-2 (Mummery et al., 1985) cells were grown in monolavers in 0.1% (w/v) gelatin-coated tissue culture flasks ;n 45% (v/v) &lbecco’s minimum essential medium, 45% (v/v) Ham’s F-12. and 10% (v/v) fetal bovine serum (GIBCO) buffered with bicarbonate in a 5% COz atmosphere at 37 “C. F9 (a gift from Dr. B. Terrana, Sclavo Research Center, Siena, Italy), FSAC cl9 (Howe and Solter, 1981), Dif-5 (Nagarajan et al., 1983), PYS-2 (Lehman et al., 1974), and PC13 (Bernstine et al., 1973) cells were cultured in 90% (v/v) Dulbecco’s minimum essential medium, 10% (v/v) fetal calf serum in noncoated tissue culture flasks. All these cell lines originate from mice. Cells were subcultured using 0.05% (w/v) trypsin, 0.02% (w/v) EDTA (Flow Labs, Irvine, Ayrshire, United Kingdom). Human K562 erythroleukemia cells were grown in suspen- sion in 90% (v/v) RPM1 1640, 10% newborn calf serum (GIBCO). P19EC cells, growing in monolayer culture, were induced to differ- entiate by addition to the culture medium of all-trans-retinoic acid (Sigma) from a stock solution (lo-* M in dimethyl sulfoxide) to a final concentration of 10V6 M. Isolation of Nuclei Nuclei of tissue culture cells were isolated as described (Rao et al., 1986). Briefly, phosphate-buffered saline (150 mM NaCI, 6.7 mM NaPi 5460 by guest on March 15, 2018 http://www.jbc.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOURNAL OF BI~L~CICAL CHEMISTRY (~8 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 10, Issue of April 5, pp. 5460-5465, 1990 Printed in U.S. A.
The Nuclear Matrix from Cells of Different Origin EVIDENCE FOR A COMMON SET OF MATRIX PROTEINS*
(Received for publication, April 17, 1989)
Nice Stuurman, Alexandra M. L. Meijne, Arnoud J. van der Pol, Luitzen de Jong, Roe1 van Driel, and Jos van RenswoudeS From the E. C. Slater Institute for Biochemical Research, University of Amsterdam, Past office Box 20151, Amsterdam 1000 HD, The Netherlands
We compared the protein composition of the nuclear matrix isolated from several murine embryonal carci- noma cells and mature tissues by two-dimensional gel electrophoresis. Two nuclear matrix fractions were investigated: the “peripheral” nuclear matrix (matrix proteins that remain insoluble after reduction), and the “internal” nuclear matrix (matrix proteins released by reduction). The two subfractions have completely dif- ferent protein compositions. Although numerous dif- ferences in nuclear matrix protein composition among different cell types were observed, a limited set of polypeptides common to all mouse cell types was iden- tified. A majority of these common proteins was also present in cells from other mammalian species (i.e. rat and human). For this set of proteins, we coin the term “minimal matrix.”
As expected, lamin B, known to be expressed throughout differentiation, is part of the common set of peripheral nuclear matrix proteins. Lamins A and C are not because these proteins were absent from undif- ferentiated embryonal carcinoma cells. Since these common nuclear matrix proteins occur in all mamma- lian nuclear matrices analyzed so far, it is likely that they have a basic role in nuclear organization and function.
Recently, several authors have expanded on the idea that a proteinaceous network in the nucleus (the nuclear matrix) is involved in control and coordination of gene expression during differentiation (Blobel, 1985; Razin, 1987; Bodnar, 1988). It is thought that loops of chromatin are persistently as well as transiently attached to this network (Pienta and Coffey, 1984; Mirkovitch et al., 1984; Gassar and Laemmli (1987); Earn- shaw, 1988). It has been proposed that each chromatin loop constitutes an independent regulatory unit containing several genes. Processes such as DNA replication (Jackson et al., 1984; Dijkwel et al., 1986; Razin et al., 1986; Tubo and Mar- telli, 1987; Nakayasu and Berezney, 1989), transcription (Jackson and Cook, 1985; Razin et al., 1985; Buttyan and Olsson, 1986), RNA processing and transport (Schroder et al., 1987a, 198713; Zeitlin et al., 1987), and the regulation of DNA superhelicity (Berrios et al., 1985) have been shown to be associated with this nuclear matrix. These observations sup- port the hypothesis that the nuclear matrix is instrumental
* This study was financed in part by a grant from The Netherlands Execut,ive Agency for the Stimulation of Computer Science (SPIN). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be sent. Fax: 20-525 5124.
in regulation and coordination of gene expression. Little is known about the nature of the proteins that
constitute the nuclear matrix. It seems likely that the nuclear matrix has a similar basal structure in almost all mammalian cells. Therefore, we expect that the elements essential to this structure will tend to be conserved during evolution and remain present throughout differentiation. In addition, nu- merous differentiation state- and species-specific components will be present in each nuclear matrix.
Recently, we and others (Fey and Penman, 1987, 1988; Stuurman et al., 1989) have shown that the protein composi- tion of the nuclear matrix is differentiation state dependent. Here, we present a detailed analysis of the matrix composition determined by two-dimensional gel electrophoresis (O’Farrell, 1975; O’Farrell et al., 1977). Nuclear matrices were prepared by extraction of nuclease-treated purified nuclei with buffers of high ionic strength. Two matrix subfractions were isolated, one enriched in nuclear envelope polypeptides (peripheral matrix), the other containing mainly internal matrix proteins (internal matrix) (Kaufmann and Shaper, 1984). We have looked for nuclear matrix polypeptides that are present in a variety of murine cell types and tissues and in cells that originate from other mammals (rat and human). The results show that a limited set of proteins is present in the nuclear matrix of all cell types studied, irrespective of species and differentiation state. We propose that these proteins are es- sential in constituting the nuclear matrix and coin the term minimal matrix for them.
MATERIALS AND METHODS
Cell Culture
P19EC (MC Burney et al., 1982), PISMES-1 (Mummery et al., 1986), P19EPI-7, and P19END-2 (Mummery et al., 1985) cells were grown in monolavers in 0.1% (w/v) gelatin-coated tissue culture flasks ;n 45% (v/v) &lbecco’s minimum essential medium, 45% (v/v) Ham’s F-12. and 10% (v/v) fetal bovine serum (GIBCO) buffered with bicarbonate in a 5% COz atmosphere at 37 “C. F9 (a gift from Dr. B. Terrana, Sclavo Research Center, Siena, Italy), FSAC cl9 (Howe and Solter, 1981), Dif-5 (Nagarajan et al., 1983), PYS-2 (Lehman et al., 1974), and PC13 (Bernstine et al., 1973) cells were cultured in 90% (v/v) Dulbecco’s minimum essential medium, 10% (v/v) fetal calf serum in noncoated tissue culture flasks. All these cell lines originate from mice. Cells were subcultured using 0.05% (w/v) trypsin, 0.02% (w/v) EDTA (Flow Labs, Irvine, Ayrshire, United Kingdom). Human K562 erythroleukemia cells were grown in suspen- sion in 90% (v/v) RPM1 1640, 10% newborn calf serum (GIBCO). P19EC cells, growing in monolayer culture, were induced to differ- entiate by addition to the culture medium of all-trans-retinoic acid (Sigma) from a stock solution (lo-* M in dimethyl sulfoxide) to a final concentration of 10V6 M.
Isolation of Nuclei
Nuclei of tissue culture cells were isolated as described (Rao et al., 1986). Briefly, phosphate-buffered saline (150 mM NaCI, 6.7 mM NaPi
5460
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Evidence for a Common Set of Nuclear Matrix Proteins 5461
(pH 7.4))washed cells were incubated for 30 min in hypotonic lysis buffer (10 mM Tris/HCl (pH 7.5 at 4 “C), 2 mM MgCl,, 3 mM CaCl*, 3 mM dithiothreitol, 0.05% (v/v) Nonidet P-40, 1 mM PMSF,’ apro- tinin (Boehringer Mannheim) at 100 kallikrein-activating units/ml). After vigorous mixing (five times for 5 s each using a vortex at 2,500 rpm) and separation of nuclei from the postnuclear supernatant by centrifugation for 10 min at 1,000 X g, nuclei were centrifuged through a sucrose cushion (2 M sucrose, 3.5 mM MgCl,, 3 mM dithiothreitol, 1 mM PMSF) for 1 h at 72,000 X g.
Isolation of nuclei from tissues was essentially according to Blobel and Potter (1966). Freshly excised mouse or rat tissues (or, alterna- tively, tissues that had been stored at -70 “C) were minced in STM (250 mM sucrose, 50 mM Tris/HCl (pH 7.4 at 4 “C), 5 mM MgSO,) supplemented with 0.2 mM PMSF, washed three times with the same buffer by decantation, and subsequently homogenized at 4 “C using a motor-driven Potter-Elvehjem homogenizer equipped with a Teflon pestle (10 strokes at 500 rpm followed by 5 strokes at 1,000 rpm). After centrifugation at 1,000 x g, postnuclear supernatant was sepa- rated from the pellet. The pellet was washed three times with STM supplemented with 0.2 mM PMSF, and the nuclei were layered on sucrose cushions and centrifuged as described above.
Isolation of the Nuclear Matrix
It is important to avoid confusion about the nature and the no- menclature of the nuclear matrix fractions used in this study. Basi- cally two kinds of fractions were employed. One type of fraction resulted from applying a classical protocol for isolating nuclear mat- rices, including an extraction with a reducing agent (Kaufmann et al., 1983). Since the work of Kaufmann et al. (1983) has shown that the residues of nuclei in this type of fraction appear as rather empty ghosts, this fraction will be referred to as peripheral nuclear matrix throughout this paper. The other type of fraction used consists of the material released upon treatment of the matrices with reducing agents and is called internal nuclear matrix in agreement with the designa- tion given by Kaufmann and Shaper (1984).
Isolation of the Peripheral Matrix-Peripheral matrices were iso- lated from purified nuclei by the aforementioned method of Kauf- mann et al. (1983) with minor modifications. Sucrose cushion-purified nuclei were resuspended in 0.25 mg/ml DNase I (from bovine pan- creas, 2000 Kunitz units/mg; Sigma) and 0.25 mg/ml RNase A (type I-AS from bovine pancreas, 50-100 Kunitz units/mg; Sigma) in STM, supplemented with 1 mM PMSF and aprotinin at 100 kallikrein- inactivating units/ml, to a density of 3-5 X 10R nuclei/ml and incu- bated for 1 h on ice. The suspension was washed with STM supple- mented with 1 mM PMSF and resuspended at a density of 3 X 10R nuclei/ml in LS (low salt) buffer (10 mM Tris/HCl (pH 7.4 at 4 “C), 0.2 mM MgSO,, 1 mM PMSF) supplemented with 10 mM P-mercap- toethanol. HS (high salt) buffer (2 M NaCl buffer in LS) supple- mented with 10 mM @-mercaptoethanol was added dropwise to a final concentration of 1.6 M NaCl. After 15 min, the matrices were pelleted by centrifugation at 5000 x g. This low/high salt extraction was repeated once. Peripheral matrices from tissues were washed once with 1% (v/v) Nonidet P-40 in LS and once with LS. The peripheral matrix fraction was washed in LS buffer and stored frozen (-70 “C) in LS buffer supplemented with aprotinin at 100 kallikrein-inactivat- ing units/ml. In accordance with Kaufmann et al. (1983), these peripheral nuclear matrices appeared as structures containing few intranuclear material as judged by phase-contrast microscopy.
Isolation of the Internal Matrix-Internal matrix proteins can be isolated after preparing matrices from nuclei stabilized by sodium tetrathionate oxidation (Kaufmann and Shaper, 1984; Kaufmann et al., 1986). In extraction of such matrices with 1 M NaCl, 20 mM dithiothreitol, internal matrix proteins were solubilized and can be separated from nuclear envelopes by centrifugation. Nuclei isolated in buffers lacking dithiothreitol were stabilized by incubation for 1 h with 0.5 mM sodium tetrathionate (Pierce Chemical Co.) in STM supplemented with 0.2 mM PMSF. Nuclei were washed three times with STM, 0.2 mM PMSF and digested with nucleases as described above. Nuclei were extracted with LS and HS as stated above; however, @-mercaptoethanol was omitted from the buffers. The mat- rices contained intranuclear material, including residual nucleoli, as judged by phase-contrast microscopy. Matrices were suspended in LS
’ The abbreviations used are: PMSF, phenylmethylsulfonyl fluo- ride; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electro- phoresis; NEPHGE, nonequilibrium pH gradient gel electrophoresis; LS, low salt; HS, high salt.
at a density of 2 x lOs/ml. An equal volume of HS supplemented with 40 mM dithiothreitol was added. At this point, the matrices became empty ghosts containing little if any intranuclear material visible by phase-contrast microscopy. After a 20-min incubation, the suspension was centrifuged at 10,000 x g for 10 min at 4 “C. The supernatant was dialyzed against 10 mM ammonium acetate (pH 7.4) with several changes of buffer and stored lyophilized at -70 “C.
Gel Electrophoresis Protein concentrations were determined using the microassay de-
scribed by Peterson (1977). Twenty or fifty micrograms of nuclear matrix protein was suspended in lysis buffer (as described by O’Far- rell, 1975). After 30 min, insoluble material was removed by centrif- ugation at 10,000 X g (pellet and supernatant had a similar protein composition as judged by SDS-PAGE, indicating that the composition of the supernatant is similar to the composition of the total matrix preparation) and layered on a NEPHGE tube gel as described by Bravo (1984). After electrophoresis (4.5 h at 400 V), the tube gels were placed on top of an 8% SDS-polyacrylamide gel, and gel electro- phoresis was carried out according to Laemmli (1970). The gels were stained with silver, employing the method of Morrisey (1981).
RESULTS
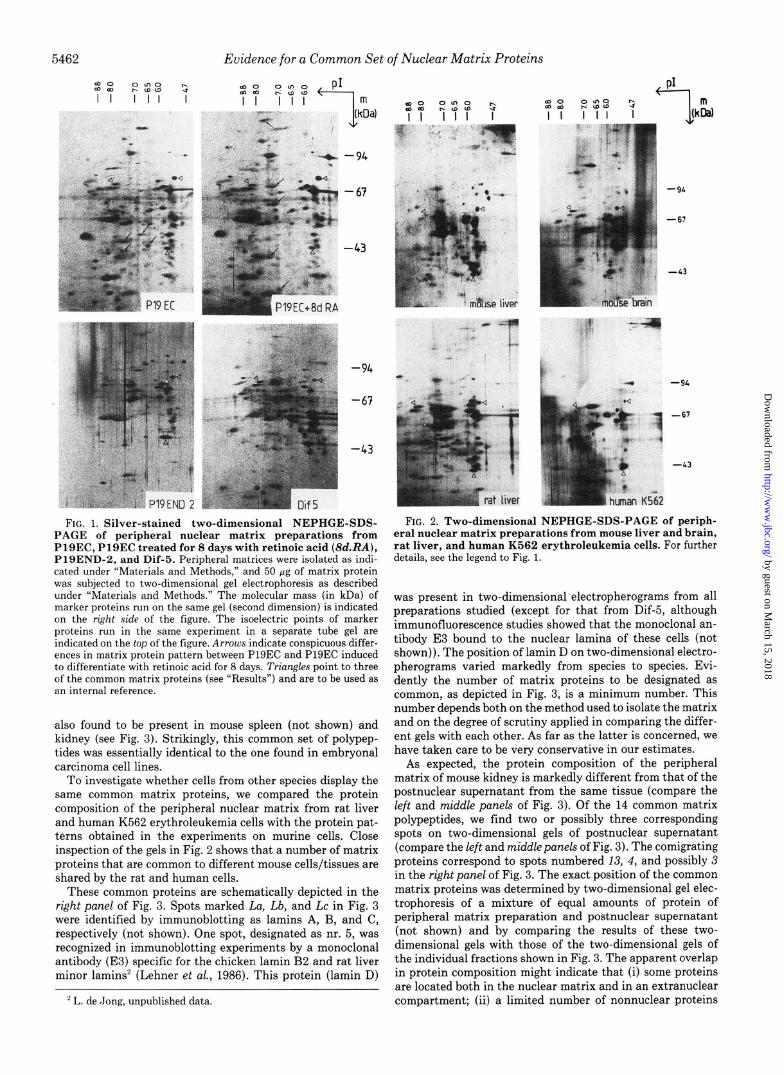
Protein Composition of the Peripheral Nuclear Matrix from Various Mouse Embryonal Carcinoma Cell Lines-Nuclei of tissue culture cells were isolated by hypotonic lysis and cen- trifugation through a sucrose cushion. They were devoid of cytoplasmic contaminants as judged by phase contrast mi- croscopy and immunofluorescent staining of isolated nuclei from P19END-2 cells with a monoclonal antibody (TROMA- 1) directed against the cytoplasmic endo-A cytokeratin (Kem- ler et al., 1981) (results not shown). The antibody did stain cytoplasmic filaments of fixed P19END-2 cells (results not shown). Previous work from our group showed that there are marked differences in protein composition of the nuclear matrix among cells in various differentiation states (Stuur- man et al., 1989). We examined the protein composition of the peripheral nuclear matrix from a variety of mouse em- bryonal carcinoma cell lines by two-dimensional NEPHGE- SDS-PAGE and studied the effect of in vitro differentiation of P19EC cells (induced by retinoic acid) on the protein composition of the peripheral nuclear matrix. The results are shown in Fig. 1. The protein composition of peripheral nuclear matrices from the undifferentiated cell line P19EC is, at first view, similar to that of peripheral matrix preparations of P19EC cells induced to differentiate by treatment with reti- noic acid for 8 days. On close inspection, however, a number of differences, as indicated by the arrows in Fig. 1, are found. The protein composition of peripheral matrix preparations of P19END-2 (a P19EC derivative) and Dif-5 (a F9 derivative) is even more different.
Apart from proteins that seem to be specific for a certain cell line, a set of matrix proteins is present which is common to all cell lines (see below). Analysis of the peripheral matrix protein composition of PYS-2, PlSMES-1, P19EPI-7, F9, PC13, and FSAC cl9 (all mouse embryonal carcinoma cell lines) confirmed these observations (not shown).
Protein Composition of the Peripheral Nuclear Matrix from Mouse Tissues and Cells of Other Mammals---In a next step we compared the protein composition of the peripheral nu- clear matrix from established mouse cell lines with that of different mouse tissues. We isolated peripheral matrices from mouse liver and brain and analyzed them by two-dimensional NEPHGE-SDS-PAGE (Fig. 2). These experiments confirmed the observations made on peripheral nuclear matrices of tissue culture cell lines. Marked differences in protein composition were observed between peripheral matrix preparations from liver and brain. Only a subset of matrix proteins was common to peripheral matrices from both tissues. The same set was
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Evidence for a Common Set of Nuclear Matrix Proteins
(hOa) .,
-94
-63
-43
FIG. 1. Silver-stained two-dimensional NEPHGE-SDS- PAGE of peripheral nuclear matrix preparations from PlSEC, Pl9EC treated for 8 days with retinoic acid ($d.RA), P19END-2, and Dif-5. Peripheral matrices were isolated as indi- cated under “Materials and Methods,” and 50 Gg of matrix protein was subjected to two-dimensional gel electrophoresis as described under “Materials and Methods.” The molecular mass (in kDa) of marker proteins run on the same gel (second dimension) is indicated on the right side of the figure. The isoelectric points of marker proteins run in the same experiment in a separate tube gel are indicated on the top of the figure. Arrows indicate conspicuous differ- ences in matrix protein pattern between P19EC and P19EC induced to differentiate with retinoic acid for 8 days. Triangles point to three of the common matrix proteins (see “Results”) and are to be used as an internal reference.
also found to be present in mouse spleen (not shown) and kidney (see Fig. 3). Strikingly, this common set of polypep- tides was essentially identical to the one found in embryonal carcinoma cell lines.
To investigate whether cells from other species display the same common matrix proteins, we compared the protein composition of the peripheral nuclear matrix from rat liver and human K562 erythroleukemia cells with the protein pat- terns obtained in the experiments on murine cells. Close inspection of the gels in Fig. 2 shows that a number of matrix proteins that are common to different mouse cells/tissues are shared by the rat and human cells.
These common proteins are schematically depicted in the right panel of Fig. 3. Spots marked La, Lb, and Lc in Fig. 3 were identified by immunoblotting as lamins A, B, and C, respectively (not shown). One spot, designated as nr. 5, was recognized in immunoblotting experiments by a monoclonal antibody (E3) specific for the chicken lamin B2 and rat liver minor lamins’ (Lehner et al., 1986). This protein (lamin D)
’ L. de Jong, unpublished data.
-94
-67
-43
4 -94
1
-67
i
-43
r uman K562
FIG. 2. Two-dimensional N S-PAGE of periph- eral nuclear matrix preparations from mouse liver and brain, rat liver, and human K562 erythroleukemia cells. For further details, see the legend to Fig. 1.
was present in two-dimensional electropherograms from all preparations studied (except for that from Dif-5, although immunofluorescence studies showed that the monoclonal an- tibody E3 bound to the nuclear lamina of these cells (not shown)). The position of lamin D on two-dimensional electro- pherograms varied markedly from species to species. Evi- dently the number of matrix proteins to be designated as common, as depicted in Fig. 3, is a minimum number. This number depends both on the method used to isolate the matrix and on the degree of scrutiny applied in comparing the differ- ent gels with each other. As far as the latter is concerned, we have taken care to be very conservative in our estimates.
As expected, the protein composition of the peripheral matrix of mouse kidney is markedly different from that of the postnuclear supernatant from the same tissue (compare the left and middle panels of Fig. 3). Of the 14 common matrix polypeptides, we find two or possibly three corresponding spots on two-dimensional gels of postnuclear supernatant (compare the left and middlepanels of Fig. 3). The comigrating proteins correspond to spots numbered 13, 4, and possibly 3 in the right panel of Fig. 3. The exact position of the common matrix proteins was determined by two-dimensional gel elec- trophoresis of a mixture of equal amounts of protein of peripheral matrix preparation and postnuclear supernatant (not shown) and by comparing the results of these two- dimensional gels with those of the two-dimensional gels of the individual fractions shown in Fig. 3. The apparent overlap in protein composition might indicate that (i) some proteins are located both in the nuclear matrix and in an extranuclear compartment; (ii) a limited number of nonnuclear proteins
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FIG:. 3. Schematic representation of the protein spots common to all peripheral matrix preparations an- alyzed (right panel) and two-dimen- sional NEPHGE-SDS-PAGE of a peripheral matrix preparation from mouse kidney (middle panel) and postnuclear supernatant (PNS) from mouse kidney (left panel). The molecular mass scale (III kDa) 1s indl- cated on the rr:rht Hl:ht panel. La, Lb, I,(. lamin\ A, H, and C, respectively. M~ddl~pnnc~l C/K+.\ Indicate the protein spots ured to construct the r~ghrht panel; tor trrnnglcs, see the legend to Fig. 1. Left pnncl trrclcs and friar&+ indicate the positlonq at which the common matrix proteins migrate m two-dImensIona electropherograms of mixtures of equal amounts of protein of peripheral nuclear matrix preparation and postnuclear su- pernatant from mouse kidney (orlginal data not shown).
Evidence for a Common Set of Nuclear Matrix Proteins
contaminates the nuclear matrix fraction; or (iii) some matrix proteins fortuitously corn&rate with nonnuclear polypeptides. It is clear however that the vast majority of the nuclear matrix proteins, including the common set of peripheral matrix pro- teins, is different from those in the postnuclear supernatant.
Protein Composition of the Internal Nuclear Matrix--Mat- rices isolated under reducing conditions are considered to consist mainly of peripheral nuclear matrix proteins (cf. Kauf- mann et al., 1981, 1983). Therefore, it is important to deter- mine whether the internal nuclear matrix subfraction also contains a set of common proteins. Matrices derived from nuclei stabilized by oxidation with sodium tetrathionate con- tain a higher percentage of the original nuclear protein than matrices isolated in the presence of reducing agents (Kauf- mann et al., 1981). We isolated these disulfide bond-stabilized matrix proteins, which, according to Kaufmann and Shaper (1984), make up the internal nuclear matrix. The protein composition of this fraction (see Fig. 4) is quite different from that of the matrices isolated under reducing conditions (com- pare Figs. 2 and 4; see also Kaufmann and Shaper, 1984) and
5463
. a-0 (
M 0 - CT=.
FIG. 4. Two-dimensional NEP- HGE-SDS-PAGE of internal matrix proteins from mouse liver and brain. The molecular mass (in kDa) of marker proteins run on the same gel (second dimension) is indicated on the right side of the figure. The isoelectric points of marker proteins run in the same experiment in a separate tube gel are indicated on the top of the figure. Filled trranggles point to some of the pro- tein spots found in both tissues; arrows indicate some of the differences in pro- tein composition.
from that of the postnuclear supernatant (compare Figs. 3 and 4). However, comparison of the internal matrix proteins from mouse liver and mouse brain shows that this fraction too contains proteins common to both tissues (Fig. 4, indi- cated by triangles) as well as tissue-specific proteins (Fig. 4, indicated by arrows). The internal nuclear matrix proteins shared by mouse liver and brain are not identical to the common matrix proteins indicated in Fig. 3 (compare Figs. 3 and 4).
DISCUSSION
The composition of the nuclear matrix, as judged by two- dimensional gel electrophoresis, is of an astonishing complex- ity. In this report, we provide evidence that several nuclear matrix proteins, identified by their isoelectric point and mo- lecular weight, are shared by murine cells of various differ- entiation stages. Lamin B, known to be expressed throughout differentiation (Guilly et al., 1987; Worman et al., 1988; Paulin-Lavasseur et al., 1988), is part of this common set. Lamins A and C are not because they are absent from undif-
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5464 Evidence for a Common Set of Nuclear Matrix Proteins
ferentiated embryonal carcinoma cells (Lebel et al., 1987; Stewart and Burke, 1987; Stuurman et al., 1989). At least a high proportion of these murine common proteins is also present in rat and human cells, indicating evolutionary con- servation of these proteins. Since these polypeptides consti- tute a group of matrix proteins that occur in matrices of all cells analyzed thus far, we call them minimal matrix proteins.
What may be the interpretation of such a set of minimal matrix proteins? Evidently minimal matrix proteins are found in all mouse cell types and tissues and in cells from other mammalian species (rat, human). Most likely, their function is the same in all different cells. They probably fulfill a variety of indispensable nuclear functions. The minimal matrix pro- teins may represent structural, enzymatic, as well as regula- tory entities. Many nuclear processes such as RNA synthesis, DNA replication, and spatial organization of chromatin are found associated with the nuclear matrix (for reviews, see Nelson et al., 1984; Nigg, 1988). Virtually nothing is known about the matrix proteins that are involved in these processes. The minimal matrix proteins may play a role in differentia- tion state-independent nuclear processes. For example, the attachment of DNA to the nuclear matrix is likely to be mediated by minimal matrix proteins. Until now, the attach- ment of matrix-associated region sequences to the nuclear matrix has been found to be independent of cell type and species (Gasser and Laemmli, 1986; Cockerill and Garrard, 1986).
In analogy to what is known for the cytoplasmic pendant of the nuclear matrix, the cytoskeleton, it is likely that some of the minimal matrix proteins have an important structural role. Since cytoskeletal proteins are often evolutionary con- served and the same proteins are often found in many differ- entiation stages (Schliwa, 1986), we expect the same to hold for the minimal matrix proteins. Moreover, the procedure used in isolating the nuclear matrix tends to select for struc- tural proteins.
It must be stressed that the current set of minimal matrix proteins represents an underestimate of the true number of common proteins. In our approach, only proteins with an isoelectric point and molecular weight that do not change, for instance during differentiation, are classified as minimal ma- trix proteins. A difference in post-translational modification of a matrix protein among different cells, for example, would exclude such a protein from the current set of minimal matrix proteins.
In this study, two methods to isolate nuclear matrix frac- tions were employed, one resulting in an enrichment of pe- ripheral nuclear matrix proteins, the other in an enrichment of internal matrix proteins. In agreement with the data pro- vided by Kaufmann and Shaper (1984), the protein composi- tion of these two fractions, as analyzed by two-dimensional gel electrophoresis, is completely different. Importantly, in both fractions a set of common proteins is found. These two sets are not overlapping. We speculate that proteins from both sets play important roles in nuclear organization.
Apart from common proteins we also find numerous differ- ences in the protein composition of nuclear matrices isolated from different cells and tissues. This finding is in agreement with the data provided by Fey and Penman (1987, 1988). Since these differences are only based on differences in iso- electric point and molecular weight, it is not clear whether they are accounted for by entirely different proteins, by major differences in post-translational modification, or by different proteins with a limited degree of homology. An example of this last category is the putative lamin D, which has a different isoelectric point and molecular weight in different species but
a common antigenic epitope, since the protein is specifically recognized in several species by the same monoclonal anti- body.
Future research will focus on the function of nuclear matrix proteins. The identification of a minimal matrix as described in this report is helpful in selecting those nuclear matrix proteins that have an important function in nuclear organi- zation.
Acknowledgments-We wish to thank Dr. Benedetto Terrana for helpful advice on the culture of embryonal carcinoma cell lines and Dr. R. Kemmler for his gift of the monoclonal antibody TROMA-1.
REFERENCES Bernstine, E. G., Hooper, M. L., Grandchamp, S., and Ephrussi, B.
(1973) Proc. Natl. Acad. Sci. U. S. A. 70. 3899-3903 Berrios,‘M., Osheroff, N., and Fisher, P. A. (1985) Proc. Natl. Acad.
Sci. U. S. A. 82, 4142-4146 Blobel, G. (1985) Proc. Natl. Acad. Sci. U. S. A. 82, 8527-8529 Blobel, G., and Potter, V. R. (1966) Science 154, 1662-1666 Bodnar, J. W. (1988) J. Theor. Biol. 132,479-507 Bravo, R. (1984) in Two-dimensional Gel Electrophoresis of Proteins
(Celis, J. E., and Bravo, R., eds) pp. 3-36, Academic Press, London Buttyan, R., and Olsson, C. A. (1986) Biochem. Biophys. Res. Com-
mun. 138, 1334-1340 Cockerill, P. N., and Garrard, W. T. (1986) Cell 44, 273-282 Dijkwel, P. A., Wenink, P. W., and Poddighe, J. (1986) Nucleic Acids
Res. 14,3241-3249 Earnshaw, W. C. (1988) BioEssays 9, 147-150 Fey, E. G., and Penman, S. (1987) Prog. Clin. Biol. Res. 249, 263-
272 Fey, E. G., and Penman, S. (1988) Proc. Natl. Acad. Sci. U. 5’. A. 85,
121-125 Gasser, S. M., and Laemmli, U. K. (1986) Cell 46,521-530 Gasser, S. M., and Laemmli, U. K. (1987) Trends Genet. 3,16-22 Guilly, M. N., Bensussan, A., Bourge, J. F., Bornens, M., and Cour-
vahn, J. C. (1987) EMBO J. 6,3?95-3799 Howe. C. C.. and Solter. D. (1981) Deu. Biol. 84.239-243 Jackson, D. ‘A., and Cook, PI R. (i985) EMBO j 4,919-925 Jackson, D. A., McCready, S. J., and Cook, P. R. (1984) J. Cell Sci.
1, (suppl.) 59-79 Kaufmann, S. H., and Shaper, J. H. (1984) Exp. Cell Res. 155,477-
495 Kaufmann, S. H., Coffey, D. S., and Shaper, J. H. (1981) Exp. Cell
Res. 132,105-123 Kaufmann, S. H., Gibson, W., and Shaper, J. H. (1983) J. Biol. Chem.
258,2710-2719 Kaufmann, S. H., Fields, A. P., and Shaper, J. H. (1986) Methods
Achieu. Exp. Pathol. 12, 141-171 Kemler, R., Brulet, B., Schnebelen, M., Gaillard, J., and Jacob, F.
(1981) J. Embryo. Exp. Morphol. 64,45-60 Laemmli, U. K. (1970) Nature 227,680-685 Lebel, S., Lampron, C., Royal, A., and Raymond, Y. (1987) J. Cell
Biol. 105, 1099-1104 Lehman, J. M., Speers, W. C., Schwartzendruber, D. E., and Pierce,
G. B. (1974) J. Cell Physiol. 79, 429-440 Lehner, C. F., Kurer, V., Eppenberger, H. M., and Nigg, E. A. (1986)
J. Biol. Chem. 261,13293-13301 MC Burney, M. M., and Rogers, B. J. (1982) Dev. Biol. 89, 503-508 Mirkovitch, J., Mirault, M.-E., and Laemmli, U. K. (1984) Cell 39,
223-232 Morrisey, J. H. (1981) Anal. Biochem. 117, 307-310 Mummery, C. L., Feijen, A., van der Saag, P. T., van den Brink, C.
E., and de Laat, S. W. (1985) Deu. Biol. 109, 402-410 Mummery, C. L., Feijen, A., Moolenaar, W. H., van den Brink, C. E.,
and de Laat, S. W. (1986) Exp. Cell Res. 165, 229-242 Nagarajan, L., Jetten, A. M., and Anderson, W. B. (1983) Exp. Cell
Res. 147,315-327 Nakayasu, H., and Berezney, R. (1989) J. Cell Biol. 108, l-11 Nelson. W. G.. Pienta. K. J., Barrach, E. R., and Coffey, D. S. (1986)
Ann;. Rev. Biophys. Biophys. Chem. 15, 457-475 Nice. E. A. (1988) Znt. Reu. Cvtol. 110. 27-92 O’Farrell, PI H. (i975) J. Bioi. Chem. 250, 4007-4021 O’Farrell, P. Z., Goodman, H. M., and O’Farrell, P. H. (1977) Cell
12, 1133-1142
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Evidence for a Common Set of Nuclear Matrix Proteins 5465
Paulin-Lavasseur, M., Scherbarth, A., and Traub, P. (1988) Eur. J. Cell Biol. 47, 121-131
Peterson, G. L. (1987) Anal. Biochem. 83, 346-356 Pienta, K. J., and Coffey, D. S. (1984) J. Cell Sci. 1, (suppl.) 123-135 Rao, K., Harford, J. B., Rouault, T., MC Clelland, A., Ruddle, F. H.,
and Klausner, R. D. (1986) Mol. Cell. Biof. 6,236-240 Razin. S. V. (1987) BioEssavs 6. 19-22 Razin; S. V.; Yarovaya, 0.” V., ‘and Georgiev, G. P. (1985) Nucleic
Acids Res. 13,7427-7444 Razin, S. V., Kekelidze, M. G., Lukanidin, E. M., and Scherrer, K.
(1986) Nucleic Acids Res. 14, 8189-8207 Razin, S. V., Chernokhvostov, V. V., Vassetzky, E. S., Razina, M. V.,
and Georgiev, G. P. (1988) Nucleic Acids Res. 16, 3617-3633
Schliwa, M. (1986) Cell Biol. Monogr. 13, 5-131 Schroder, H. C., Trolltsch, D., Friese, U., Bachmann, M., and Muller,
W. E. G. (1987a) J. Biol. Chem. 262,8917-8925 Schroder, H. C., Trolltsch, D., Wenger, R., Bachmann, M., Diehl-
Seifest, B., and Muller, W. E. G. (1987b) Eur. J. Biochem. 167, 239-245
Stewart, C., and Burke, B. (1987) Cell 51,383-392 Stuurman, N., Van Driel, R., de Jong, L., Meijne, A. M. L., and Van
Renswoude, J. (1989) Exp. Cell Res. 180, 460-466 Tubo, R. A., and Martelli, A. M. (1987) Biochemistry 26,5710-5718 Worman, H. J., Lazaridis, I., and Georgatos, S. D. (1988) J. Biol.
Chem. 263, 12135-12141 Zeitlin, S., Parent, A., Silverstein, S., and Efstratiadis, A. (1987) Mol.
Cell. Biol. 7, 111-120
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

RenswoudeN Stuurman, A M Meijne, A J van der Pol, L de Jong, R van Driel and J van
matrix proteins.The nuclear matrix from cells of different origin. Evidence for a common set of
1990, 265:5460-5465.J. Biol. Chem.
http://www.jbc.org/content/265/10/5460Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/10/5460.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Related Documents











