twenty-four defines a critical translational step in the Drosophila clock Chunghun Lim 1,* , Jongbin Lee 2,* , Changtaek Choi 2 , Valerie L. Kilman 1 , Juwon Kim 2 , Sung Mi Park 3 , Sung Key Jang 3 , Ravi Allada 1 , and Joonho Choe 2 1 Department of Neurobiology and Physiology, Northwestern University, Evanston, Illinois USA, 60208 2 Department of Biological Sciences, Korea Advanced Institute of Science and Technology, Daejeon, Korea, 305-701 3 Department of Life Sciences, Pohang University of Science and Technology, Pohang, Korea, 790-784 Abstract Daily oscillations of gene expression underlie circadian behaviours in multicellular organisms1. While attention has been focused on transcriptional and posttranslational mechanisms1 – 3, other posttranscriptional modes have been less clearly delineated. Here we report mutants of a novel Drosophila gene twenty-four (tyf) that display weak behavioural rhythms. Weak rhythms are accompanied by dramatic reductions in the levels of the clock protein PERIOD (PER) as well as more modest effects on TIMELESS (TIM). Nonetheless, PER induction in pacemaker neurons can rescue tyf mutant rhythms. TYF associates with a 5′-cap binding complex, poly(A)-binding protein (PABP) as well as per and tim transcripts. Furthermore, TYF activates reporter expression when tethered to reporter mRNA even in vitro. Taken together, these data suggest that TYF potently activates PER translation in pacemaker neurons to sustain robust rhythms, revealing a novel and important role for translational control in the Drosophila circadian clock. Transcriptional feedback loops are critical for setting time of eukaryotic circadian clocks. In Drosophila, the Clock (Clk)/cycle (cyc) dimer activates the transcription of period (per), timeless (tim), vrille (vri), Par domain protein 1 (Pdp1), and clockwork orange (cwo) genes, which in turn feed back to inhibit CLK-activated transcription or regulate Clk transcription2. These components are also modified posttranslationally to alter core clock timing2 – 3. Users may view, print, copy, download and text and data- mine the content in such documents, for the purposes of academic research, subject always to the full Conditions of use: http://www.nature.com/authors/editorial_policies/license.html#terms Correspondence and requests for materials should be addressed to R.A, [email protected] or J.C., [email protected]. * These authors contributed equally to this work Full Methods and any associated references are available in the online version of the paper at www.nature.com/nature. Supplementary Information accompanies the paper on www.nature.com/nature. Author Contributions R.A. and J.C. conceived the study; R.A., C.L., and J.C. designed the experiments; C.L. (under supervision of R.A.) and J.L (under supervision of J.C.) jointly completed Figs. 1 and 2, Supplementary Figs. 1, 4, 8, and 14, Supplementary Tables 2 and 3; J.L., S.M.P. and S.K.J. performed and analyzed the experiments in Supplementary Fig. 13; J.L., C.C., and J.K. performed the genome-wide behavioural screen; C.C. performed GST pull-down studies in Supplementary Fig. 12a; V.L.K. performed PDF quantification analysis in Supplementary Fig. 5b; C.L. performed and analyzed experiments in all remaining Figs, Supplementary Figs and Tables; C.L. and R.A. wrote the manuscript. Author Information Reprints and permissions information is available at www.nature.com/reprints. The authors declare no competing financial interests. NIH Public Access Author Manuscript Nature. Author manuscript; available in PMC 2011 August 17. Published in final edited form as: Nature. 2011 February 17; 470(7334): 399–403. doi:10.1038/nature09728. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

twenty-four defines a critical translational step in the Drosophilaclock
Chunghun Lim1,*, Jongbin Lee2,*, Changtaek Choi2, Valerie L. Kilman1, Juwon Kim2, SungMi Park3, Sung Key Jang3, Ravi Allada1, and Joonho Choe2
1 Department of Neurobiology and Physiology, Northwestern University, Evanston, Illinois USA,602082 Department of Biological Sciences, Korea Advanced Institute of Science and Technology,Daejeon, Korea, 305-7013 Department of Life Sciences, Pohang University of Science and Technology, Pohang, Korea,790-784
AbstractDaily oscillations of gene expression underlie circadian behaviours in multicellular organisms1.While attention has been focused on transcriptional and posttranslational mechanisms1–3, otherposttranscriptional modes have been less clearly delineated. Here we report mutants of a novelDrosophila gene twenty-four (tyf) that display weak behavioural rhythms. Weak rhythms areaccompanied by dramatic reductions in the levels of the clock protein PERIOD (PER) as well asmore modest effects on TIMELESS (TIM). Nonetheless, PER induction in pacemaker neurons canrescue tyf mutant rhythms. TYF associates with a 5′-cap binding complex, poly(A)-bindingprotein (PABP) as well as per and tim transcripts. Furthermore, TYF activates reporter expressionwhen tethered to reporter mRNA even in vitro. Taken together, these data suggest that TYFpotently activates PER translation in pacemaker neurons to sustain robust rhythms, revealing anovel and important role for translational control in the Drosophila circadian clock.
Transcriptional feedback loops are critical for setting time of eukaryotic circadian clocks. InDrosophila, the Clock (Clk)/cycle (cyc) dimer activates the transcription of period (per),timeless (tim), vrille (vri), Par domain protein 1 (Pdp1), and clockwork orange (cwo) genes,which in turn feed back to inhibit CLK-activated transcription or regulate Clk transcription2.These components are also modified posttranslationally to alter core clock timing2–3.
Users may view, print, copy, download and text and data- mine the content in such documents, for the purposes of academic research,subject always to the full Conditions of use: http://www.nature.com/authors/editorial_policies/license.html#terms
Correspondence and requests for materials should be addressed to R.A, [email protected] or J.C., [email protected].*These authors contributed equally to this work
Full Methods and any associated references are available in the online version of the paper at www.nature.com/nature.
Supplementary Information accompanies the paper on www.nature.com/nature.
Author Contributions R.A. and J.C. conceived the study; R.A., C.L., and J.C. designed the experiments; C.L. (under supervision ofR.A.) and J.L (under supervision of J.C.) jointly completed Figs. 1 and 2, Supplementary Figs. 1, 4, 8, and 14, Supplementary Tables 2and 3; J.L., S.M.P. and S.K.J. performed and analyzed the experiments in Supplementary Fig. 13; J.L., C.C., and J.K. performed thegenome-wide behavioural screen; C.C. performed GST pull-down studies in Supplementary Fig. 12a; V.L.K. performed PDFquantification analysis in Supplementary Fig. 5b; C.L. performed and analyzed experiments in all remaining Figs, Supplementary Figsand Tables; C.L. and R.A. wrote the manuscript.
Author Information Reprints and permissions information is available at www.nature.com/reprints.
The authors declare no competing financial interests.
NIH Public AccessAuthor ManuscriptNature. Author manuscript; available in PMC 2011 August 17.
Published in final edited form as:Nature. 2011 February 17; 470(7334): 399–403. doi:10.1038/nature09728.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Regulation at multiple levels is thought to impose temporal delays in feedback allowingsustained oscillations on a circadian time scale.
To discover novel clock components, we performed a genome-wide behavioural screen.Using the KAIST-GenExel Drosophila library, we identified ~4000 EP lines containing Pelements bearing the Upstream Activating Sequence (UAS) for the yeast GAL4 transcriptionfactor inserted near transcription start sites. These flies were crossed with transgenic fliesexpressing GAL4 under the control of the tim promoter (tim-GAL4) to drive downstreamgene expression in clock cells4. One EP line identified by a long-period rhythm was theG10872 line that contains an insertion 893 bp upstream of the CG4857 transcription startsite (Supplementary Fig. 1a). Sequence analyses of the predicted amino acid sequence forCG4857 did not reveal any apparent functional domains or obvious vertebrate homologuesbut do reveal conservation with genes from different Drosophila species and other insects(Supplementary Fig. 2). We termed this novel gene twenty-four (tyf).
To characterize the phenotype in flies bearing tyf loss-of-function mutations, we generated a~2.5 kb deletion by imprecise P element excision (Supplementary Fig. 1a, tyfΔG14151; tyfΔ),deleting amino acids 79–449 of tyf and resulting in a frame-shift and premature termination.In addition, we identified a piggyBac insertion line that shows dramatically reduced levelsof tyf transcript (tyfe00614; tyfe) without affecting the transcript levels of adjacent genes(Supplementary Fig. 1b). Wild-type flies display morning and evening peaks under 12 hlight- 12 h dark (LD) cycles, anticipate the transitions between light-on and light-off bygradually increasing their activity, and maintain their locomotor rhythm in subsequentconstant dark (DD). In tyf mutants, morning anticipation of lights-on was reduced and theirrhythm was immediately less robust, resulting in weak but long periods in DD (Fig. 1 andSupplementary Table 1). Precise piggyBac excision in tyfe restored wild-type circadianbehaviour (Supplementary Table 1), indicating that the tyf gene disruption is responsible forits circadian phenotype. Analyses in trans-heterozygous females show that tyf alleles arerecessive and not complemented by deletions of the CG4857 locus (Supplementary Fig. 3).
While circadian clocks are evident in multiple tissues, brain clocks are largely responsiblefor circadian behaviours4. Neuroanatomical studies have established two oscillator modelsin which distinct groups of clock cells control morning and evening locomotor activity andbehavioural rhythms5,6. The neuropeptide gene, Pigment-dispersing factor (Pdf), expressedin ventral lateral neurons (LNvs) has been implicated in driving morning anticipation andresetting evening clocks in the dorsal LNs (LNds)/dorsal neurons (DNs)7–9. To map theneurons important for tyf effects, we generated tyf-GAL4 lines containing the tyf promoterregion (from −3.0 kb to +0.5 kb), and visualized its expression using a UAS-GFP reporter.tyf-GAL4 expression was relatively restricted to a subset of neurons in the adult brain(Supplementary Fig. 4a–f). Anti-PER antibody staining revealed that it is strongly expressedin PDF+ LNvs and weakly in LNds assessed in independent lines (data not shown). Incontrast, tyf-GAL4 expression was not detectable in the DNs. Consistent with the idea thattyf-GAL4 reflects endogenous tyf expression, tyf-GAL4 along with a UAS-tyf transgenefully rescues the behavioural phenotypes in tyf mutants (data not shown).
To map the loci of tyf function, we restricted TYF overexpression to the PDF+ LNvs usingPdf-GAL47. This results in a long period similar to the tim-GAL4 driver, while GAL4inhibition in PDF+ cells by a Pdf-GAL80 transgene5 suppressed the long period phenotype(Supplementary Table 2). Independent UAS-tyf insertions confirmed these results. TYFexpression restricted to PDF+ cells was also sufficient to rescue free-running locomotorrhythms in mutants (Supplementary Table 3). In addition, RNAi-mediated knockdown of tyfexpression in PDF+ cells phenocopied circadian behaviours in tyf hypomorphic mutants
Lim et al. Page 2
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(Supplementary Table 3). These data indicate that tyf expression in the PDF+ pacemakerneurons is necessary and sufficient for robust behavioural rhythms.
To determine tyf effects on the core clock, we analyzed molecular rhythms from headextracts, which largely reflect eye clocks10. We found that cycling expression of PER, TIM,and PDP1 proteins in tyf mutants is comparable to wild type (data not shown). tyf transcriptlevels were relatively constant in LD and not affected in clock mutants (Supplementary Fig.4g,h). We then focused on the behaviourally relevant pacemaker neurons. Anti-PDFimmunofluorescence revealed no overt defects in the neural projections from PDF+ LNvs oftyf mutants (Supplementary Fig. 5a). Adult-specific TYF expression using a drug-inducibleGAL4 was sufficient for behavioural rescue in tyf mutants (Supplementary Table 4 andSupplementary Fig. 5c), further reducing the likelihood that tyf phenotypes are due todevelopmental defects.
Strikingly, we found that PER protein was barely detectable in LN clock cells of tyf mutants(Fig. 2a). PER cycling was dampened but not absent (Fig. 2b and Supplementary Fig. 6). tyfmutant effects were less severe in the DNs with PER at ~50% of wild-type peak levels. TYFexpression in PDF+ neurons rescued PER cycling only in PDF+ clock cells of tyf mutants(Supplementary Fig. 7). Consistent with dramatic PER reductions, PDF levels increased indorsal projections from the small LNvs of tyf mutants (Supplementary Fig. 5b), as observedin per01 flies11. TIM levels were also reduced in tyf mutants, but to a lesser extent thanPER, with peak levels in tyf mutants reduced to ~50% of wild-type (Supplementary Fig. 8a).Such effects may be indirect through PER as we found that TIM reductions were alsoobserved in per01 flies and there was little effect of loss of tyf on TIM in per01 mutants.
In contrast to strong effects on PER, tyf mutants normally expressed PDP1ε, CWO, andCLK proteins (Fig. 2a and Supplementary Fig. 8b). The oscillating expression of PDP1εprotein as well as tim and Pdp1ε transcripts in mutant flies was comparable to wild-type(Supplementary Fig. 8c,d). We reason that LD cycles, the clock neural network and/ormultiple feedback loops may buffer the molecular clock against loss of tyf function.
Given the robust reductions in PER may be responsible for the arrhythmic behaviour, wehypothesized that PER expression via the GAL4/UAS system could rescue tyf locomotorrhythms12. Indeed, we find that PER, but not TIM or CLK, overexpression specifically inPDF+ cells of tyf mutants restored wild-type levels of rhythmicity (Fig. 3, SupplementaryFig. 9, and Supplementary Table 5). These rescue data indicate that neither posttranslationalregulation of PER protein by TIM3 nor transcriptional activation of per gene expression byCLK2 would be limiting for normal circadian behaviour in tyf mutants. Moreover, thesebehavioural data suggest that PER is a major target of TYF in PDF neurons.
We next examined at what regulatory step PER expression in tyf mutants is compromised.We observed that PER protein, but not per RNA, levels were reduced in brain extracts (datanot shown). While consistent with posttranscriptional regulation, we cannot exclude thepossibility that this result could arise from the masking effect of low level non-cycling perRNA in non-circadian tissues. tyf effects were not evident on a CLK-activated per promoter-GAL4 transgene (Supplementary Fig. 10) but were evident on a per transgene lacking itsnatural promoter13 (Supplementary Fig. 11a), indicating that the per promoter is notnecessary nor sufficient for tyf effects. tyf effects were also still observed on a per(Δ)-HAHis transgene that lacks the DOUBLETIME (DBT)-binding domain, thus reducingDBT-mediated PER degradation14. We then examined PER in flies in which we rescued tyfby PER overexpression. Interestingly, neither PER levels driven by constitutive Pdf-GAL4nor oscillations were affected in tyf mutants (Supplementary Fig. 11b), indicating that tyf isnot required for posttranslational regulation of PER. The lack of most per 5′ and 3′ UTRs in
Lim et al. Page 3
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the UAS-per transgene15 and/or per overexpression itself may compensate for loss of tyf.Taken together, our observations suggest that tyf posttranscriptionally regulates perexpression.
We next asked whether TYF associates with posttranscriptional regulatory factors, circadianclock components and/or specific RNA targets using immunoprecipitation of an epitope-tagged TYF (TYF-V5). We confirmed that GAL4/UAS-driven TYF-V5 rescues behaviouralrhythms in tyf mutants (Fig. 3 and Supplementary Table 5) and that TYF-V5 expression iscytoplasmic at all times of day (Supplementary Fig. 4i). TYF-V5 driven by tim-GAL4 wasimmunoprecipitated from head extracts and then TYF-associating proteins were probed withdifferent antibodies. TYF did not associate with either PER or TIM (Fig. 4a). However,poly(A)-binding protein (PABP) was specifically co-immunoprecipitated with TYF at twotimes-of-day in a RNase-insensitive manner (Fig. 4a, data not shown). In vitro assaysdemonstrated that PABP interacts with the N-terminal region of TYF (Supplementary Fig.12a; TYF-N, amino acids 1–1167).
It has been proposed that PABP stimulates translation in part by binding to both 5′ cap-associating translation initiation factor eIF4G and the poly(A) tail in mRNAs, therebyfacilitating mRNA circularization16. Therefore, we also examined a possible association ofTYF with a cap-binding complex using 5′ cap (7-methylguanosine, m7-GTP) affinity beadsand S2 cell extracts expressing epitope-tagged TYF. We found that eIF4E, a translationinitiation factor directly recognizing the 5′ cap structure, PABP, and TYF were efficientlyand specifically pulled down by m7-GTP affinity beads in a RNase-insensitive manner (Fig.4b and Supplementary Fig. 12b). Addition of soluble m7-GTP but not GTP inhibited theircap association, validating the specificity. Interestingly, the N-terminal portion of TYF,capable of in vitro PABP binding (Supplementary Fig. 12a), exhibits strongly reducedaffinity for the 5′ cap, suggesting that PABP may not solely mediate TYF association withthe 5′ cap. Our results link TYF to RNA-binding proteins involved in translation.
We further checked whether TYF associates with clock gene transcripts. RNA in TYF-V5immunoprecipitation was analyzed by real-time RT-PCR (Fig. 4d). To quantify TYF-specific transcript enrichment, we normalized RNA levels in TYF immunoprecipitation toinput RNA (enrichment fold of TYF IP) and then subtracted the signal from a backgroundcontrol immunoprecipitation (PDFR-V5) also normalized to input. We find that TYF pullsdown significant amounts of per and tim RNAs relative to the amounts of Pdp1 at the timesof their peak expression levels (ZT15, p<0.029). Moreover, per RNA levels in TYFimmunoprecipitation were higher at their peak times than trough times (i.e., ZT15 v. ZT3)even after normalizing for input levels (p<0.026). As Pdp1 has comparable input levels toper and tim (data not shown), the low Pdp1 signal is unlikely to be explained by low inputlevels. However, differential anatomic distributions could contribute to these results17. Wealso did not detect significant TYF-specific signal for Clk and cyc RNAs inimmunoprecipitated RNA, although low input levels could explain the lack of detectableassociation (data not shown). We could not identify any RNA recognition motif in TYF,suggesting that RNA binding intermediaries, such as eIF4E and PABP, may mediate itsassociation with RNA.
To address whether TYF is incorporated into translating ribonucleocomplexes, wefractionated S2 cell extracts expressing TYF-3xHA by sucrose density gradient in theabsence or presence of EDTA to dissociate ribosomes (Supplementary Fig. 13).Sedimentation profiles demonstrated that ribosomal protein P0 and PABP were present inpolysomal fractions in an EDTA-sensitive manner. By contrast, a minor fraction of TYF co-sedimented with polysomes and exhibited a modest shift by EDTA. This pattern is similar tothat of eIF4E consistent with their association (Fig. 4b,c). Taken together, the association of
Lim et al. Page 4
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

TYF with eIF4E, PABP, and per RNA suggests a direct role in PER protein synthesis,possibly at a translation initiation step.
To investigate tyf effects on its associating RNAs, we performed a RNA-tethering assay incultured S2 cells18. TYF fused to the RNA-binding bacteriophage MS2 coat protein istethered to a luciferase reporter RNA containing MS2-binding sites (Fig. 5a). TYF activity ismonitored by assaying luciferase activity. TYF-MS2, compared to MS2 alone, enhancedluciferase expression in a MS2-binding sites-dependent manner (Fig. 5b). The C-terminalregion of TYF (TYF-C, amino acids 1161–1911) was necessary and sufficient for theactivation (Fig. 5b and Supplementary Fig. 14a–c). TYF-MS2 activation was augmented ifthe per or tim 3′ UTR is fused to the reporter gene but not the cyc 3′ UTR (Fig. 5b andSupplementary Fig. 14d). TYF without the MS2 domain could not activate reportercontaining both per 5′ and 3′ UTRs consistent with a requirement for other RNA bindingproteins (Supplementary Fig. 14h). Notably, we found that a transgenic fusion between theper coding region and luciferase lacking the per 3′ UTR (XLG-luc)19 was also reduced intyf mutants indicating that the per 3′ UTR is not necessary for tyf effects in vivo (data notshown). In contrast to reporter activity, reporter RNA levels and its nuclear/cytoplasmicdistribution were comparable between MS2 and TYF-MS2 transfected cells (SupplementaryFig. 14e,f). Moreover, analytical centrifugation through a sucrose cushion revealed thatmore reporter transcripts associate with high-density ribosomes in the presence of TYF-MS2(Supplementary Fig. 14g, p<0.027), further supporting a role in translational control.
To more directly test the translational activation function of TYF, we reconstituted thistethering system in in vitro translation assays. A C-terminal TYF region (TYFc3, aminoacids 1373–1911) fused to MS2, which robustly activated MS2 reporter expression intransfected cells (data not shown), was bacterially expressed, purified and incubated with invitro transcribed MS2 reporter RNAs and translation-competent S2 cell extracts. TYFc3-MS2 activated translation from a m7-G capped and poly(A)-tailed reporter RNA modestly(Fig. 5c, p<0.001) and a non-polyadenylated RNA even more strongly (2.9×, p<0.001).TYFc3-MS2 effects are evident without affecting reporter RNA levels (data not shown).Moreover, this TYF region is not sufficient to bind PABP in vitro (Supplementary Fig. 12a),suggesting that these effects are not mediated by PABP recruitment. These data clearlydemonstrate a translation activation function of TYF in vitro, supporting a possible role intranslation of poorly adenylated transcripts.
Relative to studies of transcriptional and posttranslational regulation, little is known aboutother posttranscriptional/translational mechanisms of core clock regulation in differentorganisms3,20. While a number of RNA-binding proteins are either rhythmically expressed1or important for behavioural rhythms21–24, direct links between specific transacting factors,specific clock gene transcripts, and in vivo core clock function have yet to be clearlyestablished, especially in metazoans. Indeed, a number of studies have indicated a role forposttranscriptional regulation in modulating per expression10,25–28. Here we demonstratewith multiple lines of evidence that TYF activates PER translation to sustain behaviouralrhythms, revealing a novel and important role for translational control in the Drosophilacircadian clock. We observe robust tyf effects on PER and lesser effects on TIM yet nodetectable reduction of other core clock components we assayed. This observation suggeststhat impairing tyf-dependent translation is not critical for the expression of most clockcomponents. Importantly, transgenic induction of PER, but not other clock componentsincluding TIM, can rescue tyf mutant phenotypes.
TYF function is especially important in pacemaker neurons. Both tyf and per expression inthe PDF+ LNv is sufficient to strongly rescue behavioural rhythms in tyf mutants. Moreover,tyf effects on PER are, by far, most evident in these pacemaker neurons. The brain
Lim et al. Page 5
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

pacemaker neurons are also among the few clock cells, which demonstrate robust free-running molecular rhythms in constant dark8,9. Thus, TYF-mediated translational controlmay be a specialization of networked pacemakers in the brain crucial for sustaining free-running rhythmicity.
Our data also strongly support the model that TYF acts at the level of translational control.TYF associates with per and tim RNAs, as well as translational regulatory components suchas the eIF4E-containing 5′ cap binding complex, and PABP, the latter of which areinsensitive to RNase treatment. In addition, TYF tethered to a reporter RNA via MS2 canactivate reporter expression without altering RNA levels in transfected cells andimportantly, in cell extracts providing exogenous RNA templates and purified TYF-MS2.
How might TYF control translation of its target RNAs? We observe specific effects on PERand TIM yet not on other clock components while we find that TYF interacts withtranslation components such as the eIF4E-containing cap binding complex and PABP. Wehypothesize that RNA-binding translational repressors associate with newly transcribed perRNA, temporarily postpone translation and thus, delay feedback PER repression on its owntranscription (Supplementary Fig. 15). Such a delay could contribute to the observed lagbetween protein and RNA particularly in pacemaker neurons, although posttranslationalmechanisms may also contribute at least in the eyes29. TYF, which does not have a knownRNA recognition motif, could then be recruited to target transcripts by these translationalrepressors, releasing them to stimulate initiation of per translation. We have not been able tobiochemically or genetically link TYF to RNA-binding proteins FMR, LARK, or thetranslation regulator Thor/4E-BP that have been shown to contribute to circadian clockfunction21,22,24. Nonetheless, TYF association with eIF4E and their similar polysomeprofiles implicates TYF as a novel translation initiation factor. In addition, TYF effects maybe more evident on poorly adenylated transcripts based on our in vitro data (Fig. 5c). Ofnote, the fly homolog of the clock-regulated deadenylase nocturnin30 has been shown to beimportant in DNs for circadian light responses but neither an LN function nor an RNA targethas been described24. Nevertheless, unique features of TYF-regulated transcripts maymediate the highly selective TYF effects on clock components in vivo.
Posttranscriptional regulation on per RNA has been considered to be modulatory to clockfunction. The identification of critical role for TYF highlights an important role for PERtranslation in the Drosophila neural clockwork. It will be of interest to determine if proteinsfunctionally analogous to TYF serve similarly important and specific functions in themammalian clock.
METHODS SUMMARYPlasmids
Total RNA from adult fly heads was isolated using Trizol reagent and reverse-transcribedusing Superscript III according to the manufacturer’s instructions (Invitrogen). tyf cDNAwas PCR-amplified by Platinum Pfx polymerase (Invitrogen) with the appropriate primersets and inserted into pUAST vector for regular germ-line transformation and into itsmodified version with attB site and C-terminal V5-tag for site-specific germ-linetransformation.
Fly stocksAll flies were reared with standard cornmeal-yeast-agar medium at 25°C under LD (12-hlight/12-h dark) cycles. EP lines G10872 and G14151 were obtained from KAIST-GenExelDrosophila library. To generate a tyf deletion line (tyfΔ), P-element excision lines wereestablished from the G14151 line and molecularly characterized by genomic DNA-PCR
Lim et al. Page 6
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

with appropriate primer sets. Df(1)HC244, Df(1)rb23, and UAS-mCD8-GFP lines wereobtained from Bloomington Drosophila stock center. UAS-tyfRNAi line was obtained fromNational Institute of Genetics (Japan). Several independent germ-line transformants wereestablished from w1118 embryos injected with UAS-tyf transgenic construct (BestGene Inc.).
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWe thank Issac Edery, Jeffrey Hall, Haig Keshishian, Michael Rosbash, Francois Rouyer, Amita Sehgal,Bloomington Drosophila stock center, Harvard Exelixis Drosophila stock collection, KAIST-GenExel Drosophilalibrary, and National Institute of Genetics for fly strains; Paul Hardin, Elisa Izaurralde, Akira Nakamura, MichaelRosbash, and Nahum Sonenberg for antibodies; Jens Lykke-Andersen for plasmids; Kent E. Duncan for helpfulsuggestions on in vitro translation assays. This work was supported by grants from the Brain Research Center of the21st Century Frontier Research Program through the National Research Foundation of Korea funded by theMinistry of Education, Science and Technology, the Republic of Korea (J.C.) and from the National Institutes ofHealth (R01NS059042, R01NS052903, R01MH067870; R.A.)
References1. Doherty CJ, Kay SA. Circadian control of global gene expression patterns. Annu Rev Genet. 2010;
44:419–444. [PubMed: 20809800]
2. Zheng X, Sehgal A. Probing the relative importance of molecular oscillations in the circadian clock.Genetics. 2008; 178:1147–1155. [PubMed: 18385110]
3. Harms E, Kivimae S, Young MW, Saez L. Posttranscriptional and posttranslational regulation ofclock genes. J Biol Rhythms. 2004; 19:361–373. [PubMed: 15534317]
4. Dubruille R, Emery P. A plastic clock: how circadian rhythms respond to environmental cues inDrosophila. Mol Neurobiol. 2008; 38:129–145. [PubMed: 18751931]
5. Stoleru D, Peng Y, Agosto J, Rosbash M. Coupled oscillators control morning and eveninglocomotor behaviour of Drosophila. Nature. 2004; 431:862–868. [PubMed: 15483615]
6. Grima B, Chelot E, Xia R, Rouyer F. Morning and evening peaks of activity rely on different clockneurons of the Drosophila brain. Nature. 2004; 431:869–873. [PubMed: 15483616]
7. Renn SC, Park JH, Rosbash M, Hall JC, Taghert PH. A pdf neuropeptide gene mutation andablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms inDrosophila. Cell. 1999; 99:791–802. [PubMed: 10619432]
8. Peng Y, Stoleru D, Levine JD, Hall JC, Rosbash M. Drosophila free-running rhythms requireintercellular communication. PLoS Biol. 2003; 1:E13. [PubMed: 12975658]
9. Lin Y, Stormo GD, Taghert PH. The neuropeptide pigment-dispersing factor coordinates pacemakerinteractions in the Drosophila circadian system. J Neurosci. 2004; 24:7951–7957. [PubMed:15356209]
10. Zeng H, Hardin PE, Rosbash M. Constitutive overexpression of the Drosophila period proteininhibits period mRNA cycling. EMBO J. 1994; 13:3590–3598. [PubMed: 8062834]
11. Park JH, et al. Differential regulation of circadian pacemaker output by separate clock genes inDrosophila. Proc Natl Acad Sci U S A. 2000; 97:3608–3613. [PubMed: 10725392]
12. Yang Z, Sehgal A. Role of molecular oscillations in generating behavioral rhythms in Drosophila.Neuron. 2001; 29:453–467. [PubMed: 11239435]
13. Frisch B, Hardin PE, Hamblen-Coyle MJ, Rosbash M, Hall JC. A promoterless period genemediates behavioral rhythmicity and cyclical per expression in a restricted subset of theDrosophila nervous system. Neuron. 1994; 12:555–570. [PubMed: 8155319]
14. Kim EY, Ko HW, Yu W, Hardin PE, Edery I. A DOUBLETIME kinase binding domain on theDrosophila PERIOD protein is essential for its hyperphosphorylation, transcriptional repression,and circadian clock function. Mol Cell Biol. 2007; 27:5014–5028. [PubMed: 17452449]
Lim et al. Page 7
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

15. Blanchardon E, et al. Defining the role of Drosophila lateral neurons in the control of circadianrhythms in motor activity and eclosion by targeted genetic ablation and PERIOD proteinoverexpression. Eur J Neurosci. 2001; 13:871–888. [PubMed: 11264660]
16. Sonenberg N, Hinnebusch AG. Regulation of translation initiation in eukaryotes: mechanisms andbiological targets. Cell. 2009; 136:731–745. [PubMed: 19239892]
17. Kula-Eversole E, et al. Surprising gene expression patterns within and between PDF-containingcircadian neurons in Drosophila. Proc Natl Acad Sci U S A. 2010; 107:13497–13502. [PubMed:20624977]
18. Keryer-Bibens C, Barreau C, Osborne HB. Tethering of proteins to RNAs by bacteriophageproteins. Biol Cell. 2008; 100:125–138. [PubMed: 18199049]
19. Veleri S, Brandes C, Helfrich-Forster C, Hall JC, Stanewsky R. A self-sustaining, light-entrainablecircadian oscillator in the Drosophila brain. Curr Biol. 2003; 13:1758–1767. [PubMed: 14561400]
20. Staiger D, Koster T. Spotlight on post-transcriptional control in the circadian system. Cell Mol LifeSci. (in press). 10.1007/s00018-010-0513-5
21. Dockendorff TC, et al. Drosophila lacking dfmr1 activity show defects in circadian output and failto maintain courtship interest. Neuron. 2002; 34:973–984. [PubMed: 12086644]
22. Sofola O, et al. The Drosophila FMRP and LARK RNA-binding proteins function together toregulate eye development and circadian behavior. J Neurosci. 2008; 28:10200–10205. [PubMed:18842880]
23. Kadener S, et al. A role for microRNAs in the Drosophila circadian clock. Genes Dev. 2009;23:2179–2191. [PubMed: 19696147]
24. Nagoshi E, et al. Dissecting differential gene expression within the circadian neuronal circuit ofDrosophila. Nat Neurosci. 2010; 13:60–68. [PubMed: 19966839]
25. So WV, Rosbash M. Post-transcriptional regulation contributes to Drosophila clock gene mRNAcycling. EMBO J. 1997; 16:7146–7155. [PubMed: 9384591]
26. Stanewsky R, Jamison CF, Plautz JD, Kay SA, Hall JC. Multiple circadian-regulated elementscontribute to cycling period gene expression in Drosophila. EMBO J. 1997; 16:5006–5018.[PubMed: 9305642]
27. Majercak J, Sidote D, Hardin PE, Edery I. How a circadian clock adapts to seasonal decreases intemperature and day length. Neuron. 1999; 24:219–230. [PubMed: 10677039]
28. Suri V, Lanjuin A, Rosbash M. TIMELESS-dependent positive and negative autoregulation in theDrosophila circadian clock. EMBO J. 1999; 18:675–686. [PubMed: 9927427]
29. Suri V, Hall JC, Rosbash M. Two novel doubletime mutants alter circadian properties andeliminate the delay between RNA and protein in Drosophila. J Neurosci. 2000; 20:7547–7555.[PubMed: 11027213]
30. Green CB, et al. Loss of Nocturnin, a circadian deadenylase, confers resistance to hepatic steatosisand diet-induced obesity. Proc Natl Acad Sci U S A. 2007; 104:9888–9893. [PubMed: 17517647]
Lim et al. Page 8
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
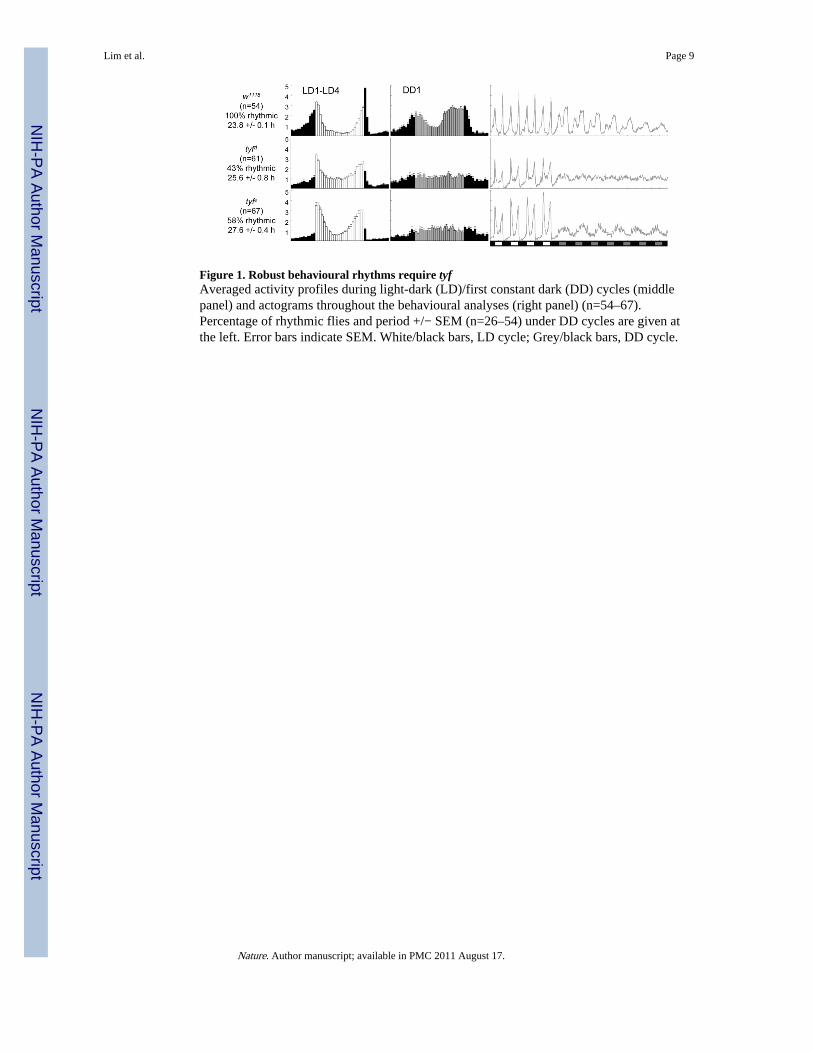
Figure 1. Robust behavioural rhythms require tyfAveraged activity profiles during light-dark (LD)/first constant dark (DD) cycles (middlepanel) and actograms throughout the behavioural analyses (right panel) (n=54–67).Percentage of rhythmic flies and period +/− SEM (n=26–54) under DD cycles are given atthe left. Error bars indicate SEM. White/black bars, LD cycle; Grey/black bars, DD cycle.
Lim et al. Page 9
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
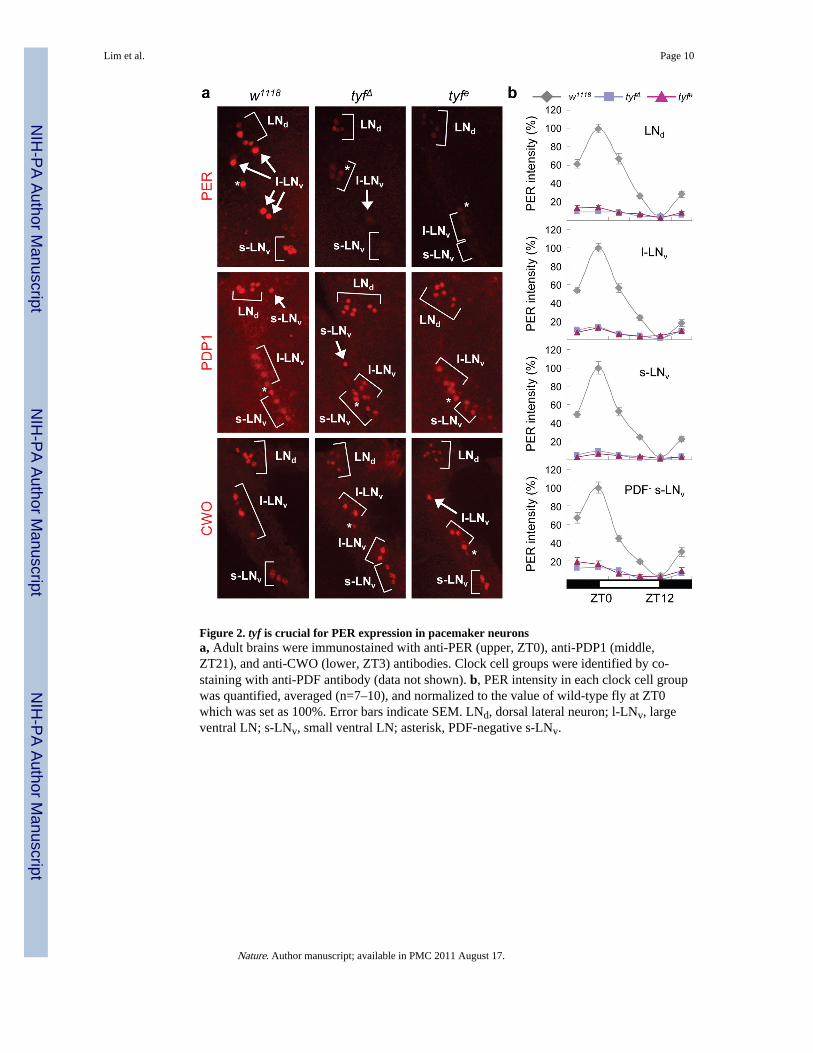
Figure 2. tyf is crucial for PER expression in pacemaker neuronsa, Adult brains were immunostained with anti-PER (upper, ZT0), anti-PDP1 (middle,ZT21), and anti-CWO (lower, ZT3) antibodies. Clock cell groups were identified by co-staining with anti-PDF antibody (data not shown). b, PER intensity in each clock cell groupwas quantified, averaged (n=7–10), and normalized to the value of wild-type fly at ZT0which was set as 100%. Error bars indicate SEM. LNd, dorsal lateral neuron; l-LNv, largeventral LN; s-LNv, small ventral LN; asterisk, PDF-negative s-LNv.
Lim et al. Page 10
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
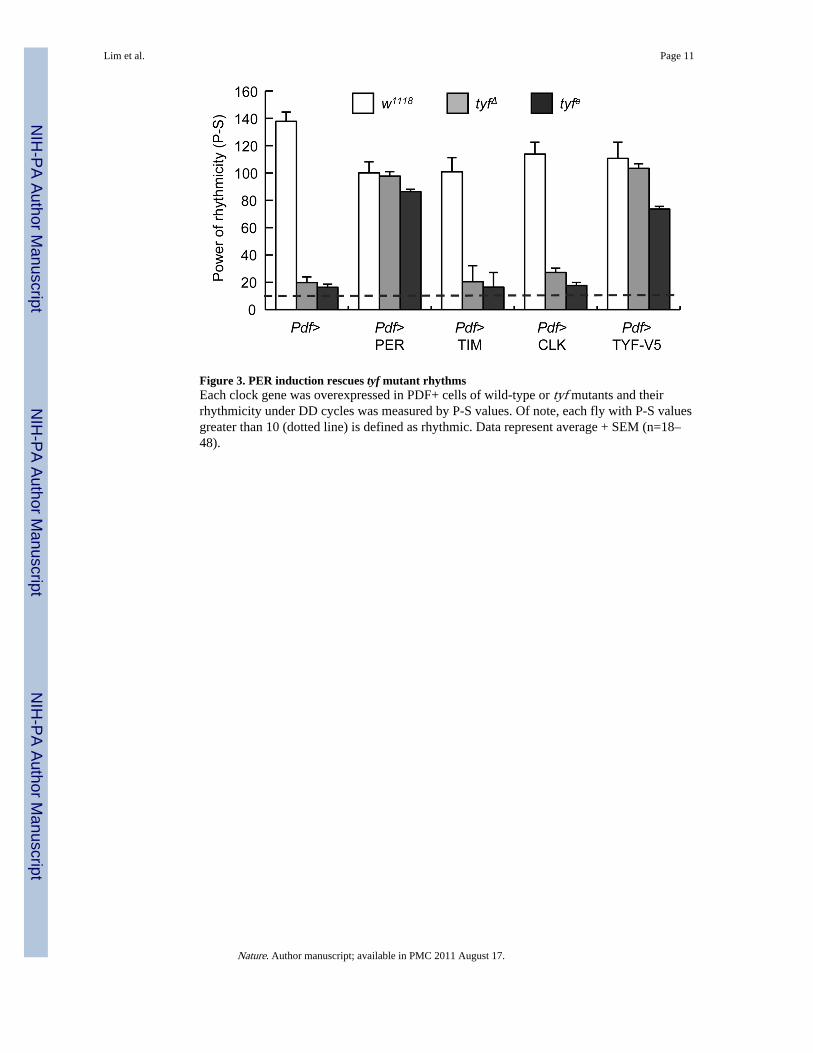
Figure 3. PER induction rescues tyf mutant rhythmsEach clock gene was overexpressed in PDF+ cells of wild-type or tyf mutants and theirrhythmicity under DD cycles was measured by P-S values. Of note, each fly with P-S valuesgreater than 10 (dotted line) is defined as rhythmic. Data represent average + SEM (n=18–48).
Lim et al. Page 11
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
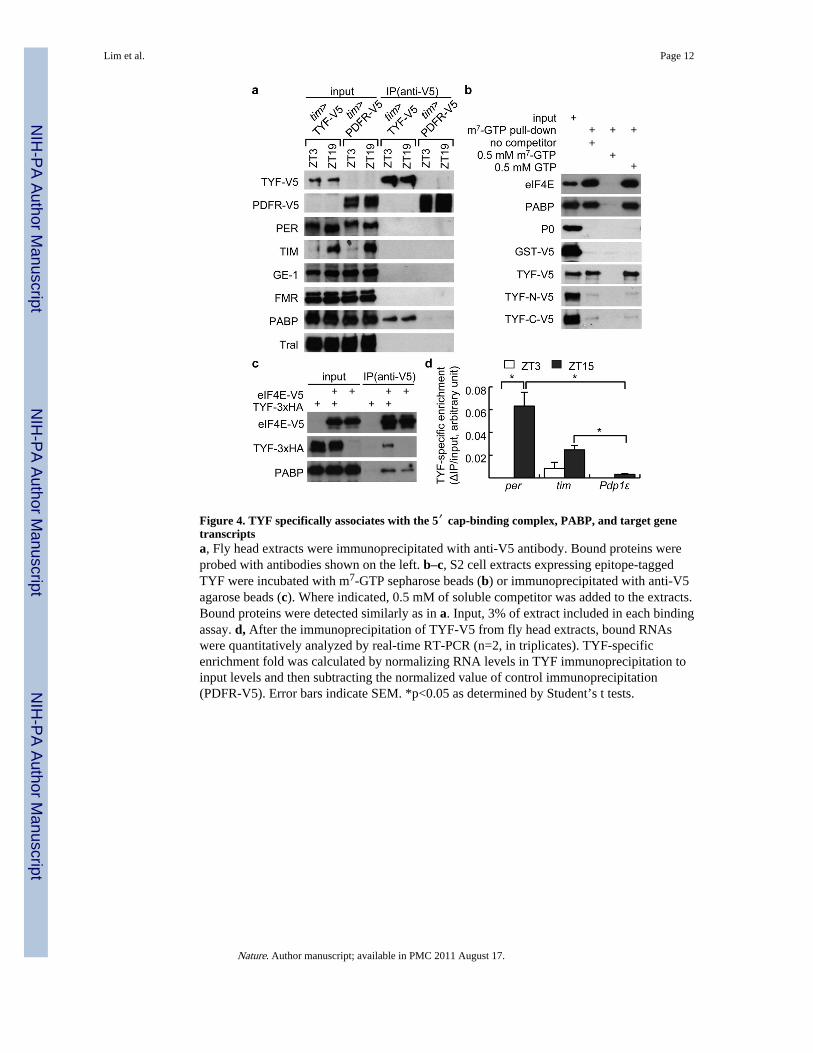
Figure 4. TYF specifically associates with the 5′ cap-binding complex, PABP, and target genetranscriptsa, Fly head extracts were immunoprecipitated with anti-V5 antibody. Bound proteins wereprobed with antibodies shown on the left. b–c, S2 cell extracts expressing epitope-taggedTYF were incubated with m7-GTP sepharose beads (b) or immunoprecipitated with anti-V5agarose beads (c). Where indicated, 0.5 mM of soluble competitor was added to the extracts.Bound proteins were detected similarly as in a. Input, 3% of extract included in each bindingassay. d, After the immunoprecipitation of TYF-V5 from fly head extracts, bound RNAswere quantitatively analyzed by real-time RT-PCR (n=2, in triplicates). TYF-specificenrichment fold was calculated by normalizing RNA levels in TYF immunoprecipitation toinput levels and then subtracting the normalized value of control immunoprecipitation(PDFR-V5). Error bars indicate SEM. *p<0.05 as determined by Student’s t tests.
Lim et al. Page 12
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
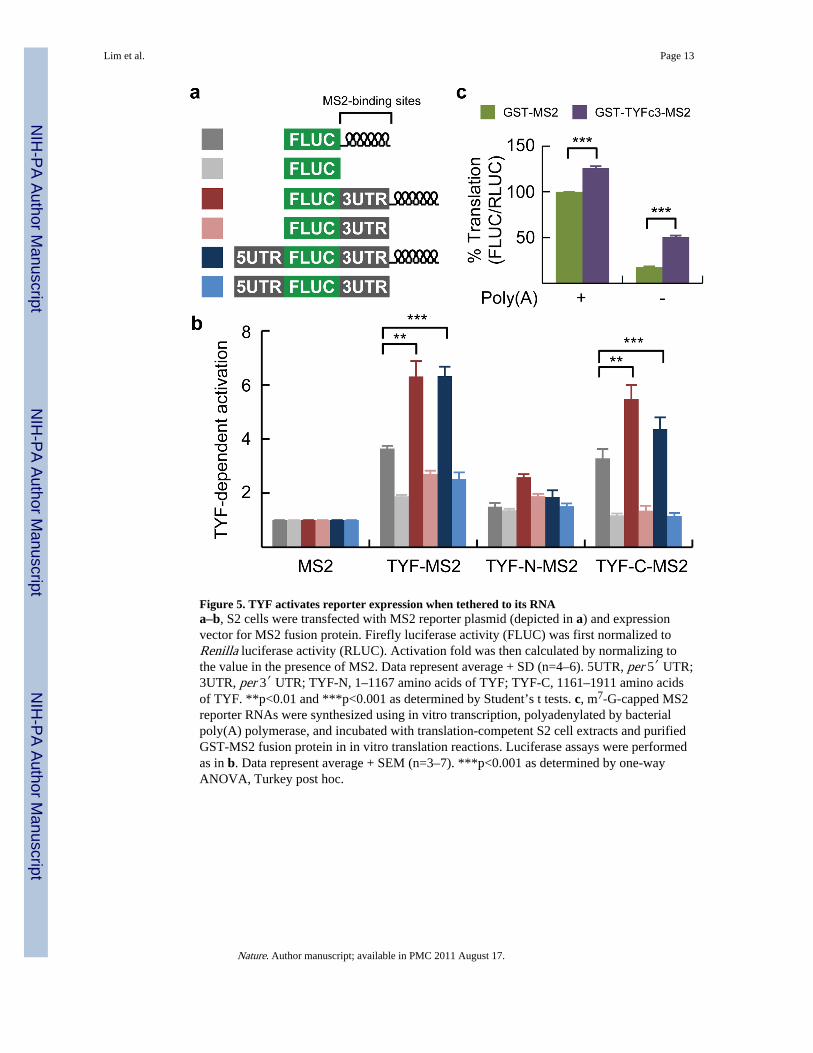
Figure 5. TYF activates reporter expression when tethered to its RNAa–b, S2 cells were transfected with MS2 reporter plasmid (depicted in a) and expressionvector for MS2 fusion protein. Firefly luciferase activity (FLUC) was first normalized toRenilla luciferase activity (RLUC). Activation fold was then calculated by normalizing tothe value in the presence of MS2. Data represent average + SD (n=4–6). 5UTR, per 5′ UTR;3UTR, per 3′ UTR; TYF-N, 1–1167 amino acids of TYF; TYF-C, 1161–1911 amino acidsof TYF. **p<0.01 and ***p<0.001 as determined by Student’s t tests. c, m7-G-capped MS2reporter RNAs were synthesized using in vitro transcription, polyadenylated by bacterialpoly(A) polymerase, and incubated with translation-competent S2 cell extracts and purifiedGST-MS2 fusion protein in in vitro translation reactions. Luciferase assays were performedas in b. Data represent average + SEM (n=3–7). ***p<0.001 as determined by one-wayANOVA, Turkey post hoc.
Lim et al. Page 13
Nature. Author manuscript; available in PMC 2011 August 17.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Related Documents







![MSc in Translational (Neuroscience) · PDF fileMSc in Translational Pathology [Neuroscience] Why Translational Pathology? The MSc Translational Pathology (Neuroscience) course combines](https://static.cupdf.com/doc/110x72/5a7454947f8b9a0d558bb440/msc-in-translational-neuroscience-a-msc-in-translational-pathology-neuroscience.jpg)



