DNA Repair 2 (2003) 581–591 The novel DNA glycosylase, NEIL1, protects mammalian cells from radiation-mediated cell death Thomas A. Rosenquist a,∗ , Elena Zaika a,b , Andrea S. Fernandes a,b , Dmitry O. Zharkov b,c , Holly Miller a,b , Arthur P. Grollman a,b a Department of Pharmacological Sciences, SUNY Stony Brook, Stony Brook, NY 11794, USA b Laboratory of Chemical Biology, Department of Pharmacological Sciences, SUNY Stony Brook, Stony Brook, NY 11794, USA c SB RAS Institute of Bioorganic Chemistry, Novosibirsk 630090, Russia Received 17 December 2002; accepted 21 January 2003 Abstract DNA damage mediated by reactive oxygen species generates miscoding and blocking lesions that may lead to mutations or cell death. Base excision repair (BER) constitutes a universal mechanism for removing oxidatively damaged bases and restoring the integrity of genomic DNA. In Escherichia coli, the DNA glycosylases Nei, Fpg, and Nth initiate BER of oxidative lesions; OGG1 and NTH1 proteins fulfill a similar function in mammalian cells. Three human genes, designated NEIL1, NEIL2 and NEIL3, encode proteins that contain sequence homologies to Nei and Fpg. We have cloned the corresponding mouse genes and have overexpressed and purified mNeil1, a DNA glycosylase that efficiently removes a wide spectrum of mutagenic and cytotoxic DNA lesions. These lesions include the two cis−thymineglycol (Tg) stereoisomers, guanine- and adenine-derived formamidopyrimidines, and 5,6-dihydrouracil. Two of these lesions, fapyA and 5S,6R thymine glycol, are not excised by mOgg1 or mNth1. We have also used RNA interference technology to establish embryonic stem cell lines deficient in Neil1 protein and showed them to be sensitive to low levels of -irradiation. The results of these studies suggest that Neil1 is an essential component of base excision repair in mammalian cells; its presence may contribute to the redundant repair capacity observed in Ogg1 −/− and Nth1 −/− mice. © 2003 Elsevier Science B.V. All rights reserved. Keywords: Base excision repair; DNA glycosylase; Oxidative DNA damage; Neil proteins; Fpg/Nei family; RNA interference 1. Introduction Metabolically produced reactive oxygen and nitro- gen species oxidize thousands of DNA bases per cell per day. Some of these lesions miscode, creating mu- tations; others block DNA replication, leading to cell death. Additional processes, including spontaneous ∗ Corresponding author. Tel.: +1-631-444-8054; fax: +1-631-444-3218. E-mail address: [email protected] (T.A. Rosenquist). deamination of cytosine and exposure to exogenous mutagens, also damage DNA. The base excision repair (BER) pathway is a univer- sal mechanism for replacing aberrant bases in DNA [1]. BER is initiated by DNA glycosylases that hy- drolyze the N-glycosidic bond, releasing the damaged base. Some DNA glycosylases also possess AP lyase activity, which cleaves the sugar–phosphate bond, in- troducing a nick in the DNA backbone 3 to the le- sion. The repair cycle is completed by the concerted action of AP-endonucleases, DNA polymerases, and DNA ligases (reviewed in [2]). 1568-7864/03/$ – see front matter © 2003 Elsevier Science B.V. All rights reserved. doi:10.1016/S1568-7864(03)00025-9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DNA Repair 2 (2003) 581–591
The novel DNA glycosylase, NEIL1, protects mammaliancells from radiation-mediated cell death
Thomas A. Rosenquista,∗, Elena Zaikaa,b, Andrea S. Fernandesa,b,Dmitry O. Zharkovb,c, Holly Miller a,b, Arthur P. Grollmana,b
a Department of Pharmacological Sciences, SUNY Stony Brook, Stony Brook, NY 11794, USAb Laboratory of Chemical Biology, Department of Pharmacological Sciences, SUNY Stony Brook, Stony Brook, NY 11794, USA
c SB RAS Institute of Bioorganic Chemistry, Novosibirsk 630090, Russia
Received 17 December 2002; accepted 21 January 2003
Abstract
DNA damage mediated by reactive oxygen species generates miscoding and blocking lesions that may lead to mutationsor cell death. Base excision repair (BER) constitutes a universal mechanism for removing oxidatively damaged bases andrestoring the integrity of genomic DNA. InEscherichia coli, the DNA glycosylases Nei, Fpg, and Nth initiate BER of oxidativelesions; OGG1 and NTH1 proteins fulfill a similar function in mammalian cells. Three human genes, designatedNEIL1,NEIL2andNEIL3, encode proteins that contain sequence homologies to Nei and Fpg. We have cloned the corresponding mouse genesand have overexpressed and purified mNeil1, a DNA glycosylase that efficiently removes a wide spectrum of mutagenic andcytotoxic DNA lesions. These lesions include the twocis−thymineglycol(Tg) stereoisomers, guanine- and adenine-derivedformamidopyrimidines, and 5,6-dihydrouracil. Two of these lesions, fapyA and 5S,6R thymine glycol, are not excised bymOgg1 or mNth1. We have also used RNA interference technology to establish embryonic stem cell lines deficient in Neil1protein and showed them to be sensitive to low levels of�-irradiation. The results of these studies suggest that Neil1 is anessential component of base excision repair in mammalian cells; its presence may contribute to the redundant repair capacityobserved inOgg1−/− andNth1−/− mice.© 2003 Elsevier Science B.V. All rights reserved.
Keywords:Base excision repair; DNA glycosylase; Oxidative DNA damage; Neil proteins; Fpg/Nei family; RNA interference
1. Introduction
Metabolically produced reactive oxygen and nitro-gen species oxidize thousands of DNA bases per cellper day. Some of these lesions miscode, creating mu-tations; others block DNA replication, leading to celldeath. Additional processes, including spontaneous
∗ Corresponding author. Tel.:+1-631-444-8054;fax: +1-631-444-3218.E-mail address:[email protected] (T.A. Rosenquist).
deamination of cytosine and exposure to exogenousmutagens, also damage DNA.
The base excision repair (BER) pathway is a univer-sal mechanism for replacing aberrant bases in DNA[1]. BER is initiated by DNA glycosylases that hy-drolyze the N-glycosidic bond, releasing the damagedbase. Some DNA glycosylases also possess AP lyaseactivity, which cleaves the sugar–phosphate bond, in-troducing a nick in the DNA backbone 3′ to the le-sion. The repair cycle is completed by the concertedaction of AP-endonucleases, DNA polymerases, andDNA ligases (reviewed in[2]).
1568-7864/03/$ – see front matter © 2003 Elsevier Science B.V. All rights reserved.doi:10.1016/S1568-7864(03)00025-9

582 T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591
While the genetics and biochemistry of oxidativedamage-specific DNA glycosylases in bacteria havelargely been elucidated, the mammalian counterpartsare relatively unexplored. InE.coli, Fpg, Nei, andNth act on overlapping sets of modified DNA bases,protecting cells from the deleterious effects of ox-idative DNA damage[3]. In mammalian cells, twoNth-type glycosylases that recognize oxidative lesionshave been described. NTH1 removes a wide varietyof oxidized pyrimidine derivatives, including 5,6-dihydroxy-5,6-dihydrothymine (thymine glycol (Tg))and 2,6-diamino-4-hydroxy-5-formamidopyrimidine(fapyG) but not 4,6-diamino-5-formamidopyrimidine(fapyA) [4–8]. Surprisingly, knockoutNth1 mice re-main healthy and, in extracts prepared from tissues ofNth1-null mice, at least two Tg DNA glycosylase ac-tivities are present[9,10]. Another NTH-family glyco-sylase, OGG1, with functional but not structural simi-larities to Fpg, is present in most eukaryotes[11–16].Humanogg1maps to a region of chromosome 3 as-sociated with lung cancer[13,15]andogg1mutationshave been observed in neoplastic tissues and cell lines[17,18]. Mice with null Ogg1function display a slightincrease in mutation rate in some tissues but show noincreased incidence of cancer[19–21]. In Ogg1-nullcells, 8-oxoG repair activity is detected in activelytranscribed sequences and it has been proposed thatan alternate pathway, transcription-coupled repair, isresponsible for limiting 8-oxoG mutagenesis in mam-malian cells[22,23]. It is also possible that additional8-oxoG glycosylases provide this function[24,25].
Three mammalian genes which contain the hallmarkhelix-two turn-helix motif and N-terminal homologywith DNA glycosylases of the Fpg/Nei family havebeen detected in recently released genomic and cDNAsequence databases[26–29]. These genes were desig-natedNeil1, Neil2 and Neil3(Nei-like), based on theirsequence similarity toNei[30]. In this paper we extendthe substrate-specificity analysis of mNeil1, showingthat prevalent oxidative lesions, including bothcis-Tgstereoisomers, are kinetically competent substrates forthis enzyme. mNeil1 also excises fapyG and fapyAfrom irradiated DNA. These formamidopyrimidinesare among the major cytotoxic products of oxidativeDNA damage. In addition, we used an RNA interfer-ence technique to establish murine embryonic stemcell lines with markedly reduced Neil1 activity. Thesecell lines show increased sensitivity to low-dose ioniz-
ing radiation. Taken together, these observations sug-gest that Neil1 activity may account for the mild phe-notype inOgg1−/− andNth1−/− mice.
2. Methods
2.1. Oligonucleotides
Adapter primers for amplifying, cloning, and se-quencing of murineNeil1, Neil2, Neil3, cDNAs werepurchased from Operon; oligonucleotides specific formouseNth1were kindly provided by McTigue and Delos Santos.
mNEIL1F1: 5′-TTTTTTCATATGCCAGAGGGCC-CAGAG-3′
mNEIL1R1: 5′-TTTTTTAAGCTTGGAGCTGTAG-GGGTTTCC-3′
mNEIL2F1: 5′-TTTTTTCATATGCCAGAAGGGC-CATCTG-3′
mNEIL2R1: 5′-TTTTTTAAGCTTGGAGGACGG-GAGATTCTG-3′
mNEIL2R2: 5′-TTTTTTAAGCTTTTACTTGGAG-GACGGGAGATTCTG-3′
mNEIL3F1: 5′-0TTTTTTCATATGGTGGAAGGG-CCAGGG-3′
mNEIL3R1: 5′-TTTTTTAAGCTTGCATCCTGGA-ACAATTTC-3′
mNTH1F1: 5′-AAAAAACATATGAACTCAGGG-GTGCGGATG-3′
mNTH1R1: 5′-AAAAAAAAGCTTGAGATCCTG-GGCAGCAGG-3′
Oligonucleotides were synthesized by standardphosphoramidite chemistry. 5R,6S- and 5S,6R-Tg ste-reoisomers were introduced into oligonucleotides asdescribed[31–33]. A fapyG analog, 2,6-diamino-4-hydroxy-5-N-methylformamidopyrimidine (meFapyG),was generated by postsynthetic modification in anoligonucleotide bearing a single guanine residue[34].By digesting withE. coli Fpg protein, this preparationwas shown to contain 30% meFapyG.
Unless otherwise indicated, enzyme substrates con-tained the following sequences: 5′-CTC TCC CTTCXC TCC TTT CCT CT-3′, where X: 5,6-dihydro-uracil (DHU), 8-oxoG, U, 5-hydroxycytosine (5OHC),5-hydroxyuracil (5OHU) or meFapyG; and 5′-GACAAG CGC AGX CAG CCG AAC AC-3′, where X:

T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591 583
Tg. The complementary strand contained G, C, T orA positioned opposite the modified base.
2.2. Enzymes
Uracil–DNA glycosylase and T4 polynucleotide ki-nase were purchased from New England Biolabs. Fpg[35] mOgg1[36] and Nei[34] proteins were purifiedand assayed as described. Nfo protein was the gen-erous gift of M. Saparbaev, Institut Gustave Roussy,France.
2.3. Cloning of mNeil1, mNeil2, mNeil3, mNth1
Total RNA was isolated from exponentially grow-ing mouse 129Sv/ev TC1 embryonic stem cells us-ing the Hi Pure RNA kit (Roche). TC1 cells were agift from Leder and co-workers[37]. OligodT-primedfirst strand cDNA was synthesized using a SuperscriptII kit (Life Technologies, #18089-011). PCR productswere sequenced in the SUNYSB DNA SequencingFacility. PCR products were first digested withNdeIand HindIII then ligated intoNdeI/HindIII-digestedpET24b (Novagen). These plasmids express insertedproteins with a C-terminal hexahistidine tag.
2.4. Expression and purification of mNeil1
Expression plasmids were transformed intoBL21(DE3)RIL cells (Stratagene). NEIL1-expressingcells were grown at 37◦C in 2xTY medium containing25�g/ml kanamycin and 25�g/ml chloramphenicol.After the OD600 reached 0.6–0.8, IPTG was addedto a final concentration of 0.2 mM and the incuba-tion was extended for 3 h. Cells were harvested bycentrifugation at 4oC and stored at−80◦C.
Cells were resuspended in 50 ml of a solution con-taining 10 mM Tris, 1 mM EDTA, pH 8.0, and 1 mMphenylmethylsulfonyl fluoride (PMSF). Lysozymeand DNase I were added to a final concentration of0.1 mg/ml and 1�g/ml, respectively. After incubationat room temperature for 20 min, NaCl was added (fi-nal concentration, 1 M) and the cell suspension stirredfor 30 min on ice. The cell lysate was centrifuged at25,000× g at 4◦C for 30 min. The supernatant wastreated with 100�l of 5% polyethylenimine solutionand the centrifugation step was repeated. Ammoniumsulfate was added to 50% saturation and allowed to
stand for 2 h. The precipitate was collected by cen-trifugation at 25,000× g at 4◦C for 20 min. Theresulting pellet was dissolved in buffer A (20 mMHEPES-NaOH, pH 7.5) containing 200 mM NaCl,1 mM EDTA and 1 mM DTT, then loaded onto a70 ml S-Sepharose column equilibrated with buffer Acontaining 200 mM NaCl. Proteins were eluted using400 ml of a 200–800 mM NaCl gradient in buffer A.Fractions containing NEIL1 were pooled. Imidazolewas added to a final concentration of 20 mM and thesolution loaded onto 5 ml HiTrap Chelating (Ni) col-umn (Pharmacia). Protein was eluted in 10 columnvolumes with 20–400 mM imidazole gradient in bufferB (20 mM HEPES-NaOH, pH 7.5, 0.5 M NaCl). Thepeak fractions were diluted two-fold with buffer A andadsorbed onto SP/F 10/100 (POROS) column. Neil1was eluted with 20 column volumes of 200–800 mMNaCl linear gradient in buffer A. Fractions were ana-lyzed by SDS-PAGE. Those containing at least 95%pure protein were pooled, dialyzed against the storagebuffer (50 mM HEPES-NaOH, pH 7.5, 400 mM NaCl,1 mM EDTA, 1 mM DTT, 50% glycerol), and storedat −20◦C. The molecular weight for the purifiedprotein was confirmed by mass spectroscopy.
mNth1 protein was purified from 20 ml culturesgrown as described above and induced by addingIPTG to a final concentration of 1 mM. Cells wereresuspended in 300�l of 20 mM sodium phosphate,pH 8.0, 1 mM DTT, 1 mM PMSF, 1 mg/ml lysozymeand lysed by sonication. An equal volume of 1MNaCl, 20 mM sodium phosphate, 1 mM DTT, 10 mMimidazole, 1 mM PMSF, pH 8.0 was added and thelysate stored on ice for 30 min before centrifuging at16,000× g for 30 min. The supernatant was trans-ferred to a fresh tube and 100�l of Ni-CAM HC resin(Sigma), pre-equilibrated in 20 mM sodium phos-phate, pH 8.0, 10 mM imidazole, 1 mM DTT, wasadded. The resin was pelleted and washed three timesin equilibration buffer. mNTH1 protein was eluted andstored in 20 mM sodium phosphate, 200 mM imida-zole, 1 mM DTT, and 25% glycerol, pH 8.0 at−20◦.
2.5. Glycosylase/lyase assays
Oligonucleotides containing target adducts werelabeled at the 5′ end with [�-32P]-ATP and polynu-cleotide kinase.32P-labeled oligonucleotides werehybridized to a 1.3-fold excess of the unlabelled

584 T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591
complementary strand. To generate a substrate con-taining an abasic (AP) site, a uracil-containingoligonucleotide was treated for 5 min at 37◦C withuracil-DNA glycosylase in a solution containing25 mM sodium phosphate, pH 7.5, 100 mM NaCl,2 mM EDTA, and 1 mM DTT. Reaction volumes were10�l. Concentrations of substrate and enzymes usedin individual experiments are given in legends to thefigures. Reactions were terminated by the additionof 5�l formamide gel buffer and heated at 95◦C for1 min. Products were separated by electrophoresison 20% polyacrylamide/7 M urea gels and quantifiedusing a PhosphorImager (Molecular Dynamics).
2.6. Antibodies
Polyclonal anti-mNeil1 rabbit sera were pro-duced at Cocalico Biologicals (Reamstown, PA).Mouse monoclonal antisera specific for PCNA(PC10) and beta-actin (AC-74) were purchased fromSigma–Aldrich and used as per the manufacturer’srecommendations. Antibodies were detected with anECL kit (Amersham Biosciences).
2.7. RNAi vector construction
Adapter primers used for amplifying the mouseRNAseP H1 promoter were 5′-CCATGGAATTCCAT-GCAAATTACGCGCTGTG-3′ (mH1A) and 5′-GCA-AGCTTCGTGTCGACGTAGACCGGCCGCCACT-ATAAG-3′ (mH1B). The 110 base pair H1 promoterwas amplified from mouse 129Sv/ev genomic DNA,digested withEcoRI and HindIII, then ligated toEcoRI/HindIII-digested pUC18 to generate the vectorpmH1P. TheEcoRI fragment from pMJKO containingthe pgk-neogene was ligated into theEcoRI site ofpH1P to generate pH1P-neo. The following oligonu-cleotides were used to generate the hairpin-enco-ding insert: 5′-TTTTCTCGAGCCGTCCTCTGTCA-GCCGGAACTTCAAGAGA-3′ (NEIL1A), 5′-CCTT-AAGCTTTTCCAAAAAGTCCTCTGTCAGCCGG-AACTCTCTTGAA-3′(Neil1B). Neil1A and Neil1Boligonucleotides were annealed and extended withKlenow DNA polymerase. The resulting fragment wasdigested withXhoI and HindIII and ligated toSalI/HindIII-digested pH1P-neo to generate the plasmidpmNeil1HP. The plasmids, pNeil1HP or pH1P-neo,were electroporated into TC1 ES cells and colonies
selected with 300�g/ml Geneticin (Roche). Individ-ual colonies were cloned and frozen at passage 3.
2.8. Radiation survival curves
ES cells were trypsinized and resuspended in culturemedium at a concentration of 106 cells/ml. Cells wereexposed for 1, 2, or 3 min in a Gammacell-40 Cesiumirradiator at a dose rate of 0.75 Gy/min. Cells were pel-leted and resuspended in fresh media; 1000 cells wereseeded in gelatinized tissue culture dishes. Each timepoint was plated in triplicate. Colonies were grown forsix days, then fixed and stained in 2% ethanol, 0.2%crystal violet for 10 min at room temperature. Excesscrystal violet was removed by washing with water andcolonies counted under a microscope. Colony numberwas normalized to mock-irradiated cells to correct forthe plating efficiency of each cell line.
3. Results
3.1. Identification of sequences in mammaliancells homologous to nei
The E. coli Nei protein sequence was used in aTBLASTN homology search of the NCBI translatedhuman genome database. Three potential genes, des-ignated NEIL1, NEIL2 and NEIL3, were detectedand identified as potential Nei or Fpg homologs bythe NCBI sequence automatic annotation system.Representative hypothetical human protein sequencesare FLJ22402 (NEIL1), FLJ20378 (NEIL2) andFLJ10858 (NEIL3). Further BLAST searches identi-fied more than thirty expressed sequence tag (EST)clones for each gene, indicating that each homologis an expressed gene. We also identified homologouspartial cDNA sequences for each gene in mouse ESTclones.
We designed PCR primers to amplify the putativecoding region of each mouse gene. Each primer setamplified a major band from oligo-dT primed cDNAisolated from mouse embryonic stem cells (data notshown). ForNeil1 andNeil2, the same primers ampli-fied larger fragments when genomic DNA was usedas the template, indicating that the RT-PCR productswere spliced derivatives, as expected.mNeil3 primersfailed to amplify a product from genomic DNA,

T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591 585
Fig. 1. Sequence lineup ofE. coli Nei and Fpg with mouse Neil1, Neil2, and Neil3. Boxed sequences represent functional regions of theFpg/Nei proteins retained in the mammalian homologs. (A) N-terminal homology; (B) H2TH Fpg-type DNA binding domain; (C) putativezinc finger: note that mNeil1 does not retain the Nei/Fpg zinc finger domain; (D) second zinc finger in Neil3; (E) 32-amino acid stretchof Neil3 with 69% identity to a repeat near the C-terminus of mammalian topoisomerase III�.
possibly reflecting the presence of large introns inthis gene.
The mouse RT-PCR products were sequenced, re-vealing open reading frames in agreement with theconsensus sequence derived from the NCBI EST andGenomic Sequence databases.Fig. 1 depicts the se-quence homology between mouse Neil proteins andE.coli Nei and Fpg proteins. Each sequence has sectionshomologous to essential sections of the Nei/Fpg pro-tein family. Of special note, all three mammalian ho-mologs contain a putative helix-two turn-helix DNAbinding motif which, in mNeil2 and mNeil3, is fol-lowed by putative zinc finger domains. In bacterialhomologs, the N-terminal sequence is involved in en-zyme catalysis with the N-terminal Pro2 residue act-ing as nucleophile. Conserved glutamic acid residuesin the third and fifth position are believed to activatethis residue and stabilize the substrate in the transitionstate[38,39]. The Neil2 sequence contains proline inthe second position and glutamic acid in the third. TheNeil3 sequence has glutamic acid in the third posi-tion but lacks Pro 2. The mouse proteins share 84.7,79.9, and 80.2% sequence identity with human NEIL1,NEIL2, and NEIL3, respectively.
3.2. Chromosomal positions of NEIL genes inmouse and human
Genomic sequences place the humanneil1, neil2,and neil3 genes on chromosomes 15q22, 8p22, and
4q33, respectively. The corresponding mouse geneticlocations are chromosomes 9 (31 cM), 14 (28 cM),and 8 (32 cM) for mNeil1, mNeil2,and mNeil3,respectively. The mouseNeil2 gene maps near theDisorganization (Ds) locus associated with dominantpleiotrophic developmental defects and is locatedon the cosmid believed to harbor theDs gene[40].However, theNeil2 coding exons are not altered inDNA obtained fromDs mice (J. Nadeau, personalcommunication).
4. Glycosylase/lyase activity and substratespecificity of mNEIL1
To establish the class of glycosylase to whichmNeil1 belongs, the enzyme was incubated with a5′ 32P-labelled duplex oligonucleotide containing asingle DHU residue. Products of this reaction werecompared with those produced by other enzymes(Fig. 2B). mNeil1 produced a single radioactive prod-uct co-migrating with the�-elimination fragmentproduced byE. coli Nei, indicating that mNeil1 uti-lizes a�–� elimination mechanism, like other mem-bers of the Fpg/Nei family. Other markers shownin Fig. 2B include an oligonucleotide containing a3′-OH terminus produced by AP endonuclease ac-tivity (Nfo protein) and a 3′ ring-opened unsaturatedsugar generated via a single�-elimination reac-tion (mNth). A �–� elimination mechanism is also
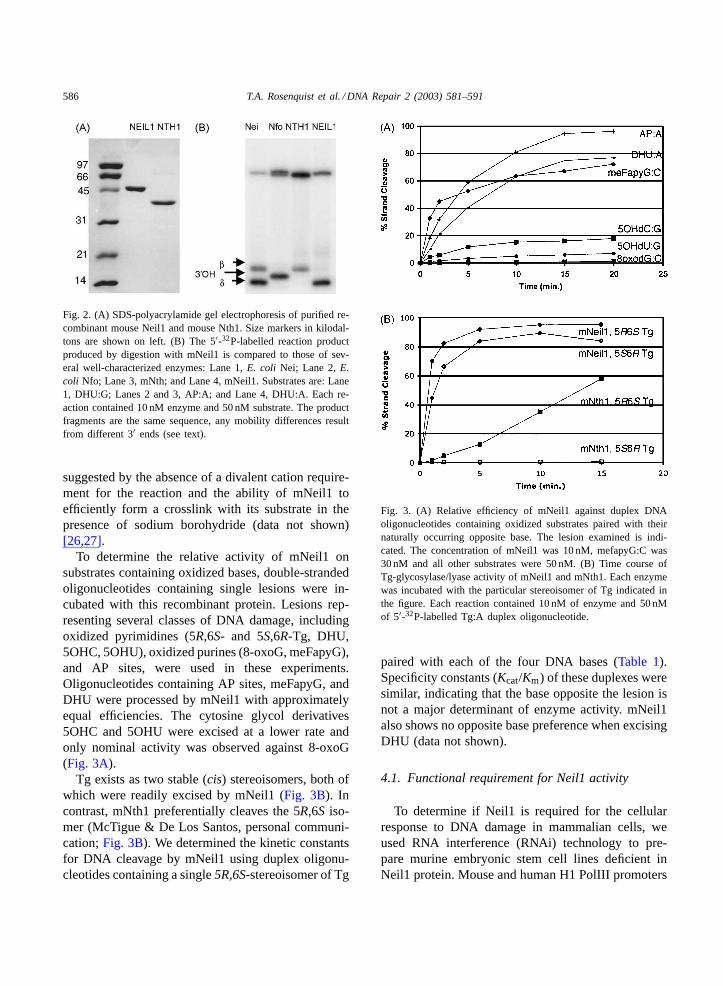
586 T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591
Fig. 2. (A) SDS-polyacrylamide gel electrophoresis of purified re-combinant mouse Neil1 and mouse Nth1. Size markers in kilodal-tons are shown on left. (B) The 5′-32P-labelled reaction productproduced by digestion with mNeil1 is compared to those of sev-eral well-characterized enzymes: Lane 1,E. coli Nei; Lane 2,E.coli Nfo; Lane 3, mNth; and Lane 4, mNeil1. Substrates are: Lane1, DHU:G; Lanes 2 and 3, AP:A; and Lane 4, DHU:A. Each re-action contained 10 nM enzyme and 50 nM substrate. The productfragments are the same sequence, any mobility differences resultfrom different 3′ ends (see text).
suggested by the absence of a divalent cation require-ment for the reaction and the ability of mNeil1 toefficiently form a crosslink with its substrate in thepresence of sodium borohydride (data not shown)[26,27].
To determine the relative activity of mNeil1 onsubstrates containing oxidized bases, double-strandedoligonucleotides containing single lesions were in-cubated with this recombinant protein. Lesions rep-resenting several classes of DNA damage, includingoxidized pyrimidines (5R,6S- and 5S,6R-Tg, DHU,5OHC, 5OHU), oxidized purines (8-oxoG, meFapyG),and AP sites, were used in these experiments.Oligonucleotides containing AP sites, meFapyG, andDHU were processed by mNeil1 with approximatelyequal efficiencies. The cytosine glycol derivatives5OHC and 5OHU were excised at a lower rate andonly nominal activity was observed against 8-oxoG(Fig. 3A).
Tg exists as two stable (cis) stereoisomers, both ofwhich were readily excised by mNeil1 (Fig. 3B). Incontrast, mNth1 preferentially cleaves the 5R,6S iso-mer (McTigue & De Los Santos, personal communi-cation;Fig. 3B). We determined the kinetic constantsfor DNA cleavage by mNeil1 using duplex oligonu-cleotides containing a single5R,6S-stereoisomer of Tg
Fig. 3. (A) Relative efficiency of mNeil1 against duplex DNAoligonucleotides containing oxidized substrates paired with theirnaturally occurring opposite base. The lesion examined is indi-cated. The concentration of mNeil1 was 10 nM, mefapyG:C was30 nM and all other substrates were 50 nM. (B) Time course ofTg-glycosylase/lyase activity of mNeil1 and mNth1. Each enzymewas incubated with the particular stereoisomer of Tg indicated inthe figure. Each reaction contained 10 nM of enzyme and 50 nMof 5′-32P-labelled Tg:A duplex oligonucleotide.
paired with each of the four DNA bases (Table 1).Specificity constants (Kcat/Km) of these duplexes weresimilar, indicating that the base opposite the lesion isnot a major determinant of enzyme activity. mNeil1also shows no opposite base preference when excisingDHU (data not shown).
4.1. Functional requirement for Neil1 activity
To determine if Neil1 is required for the cellularresponse to DNA damage in mammalian cells, weused RNA interference (RNAi) technology to pre-pare murine embryonic stem cell lines deficient inNeil1 protein. Mouse and human H1 PolIII promoters

T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591 587
Table 1Kinetic parameters of mNeil1 on thymine glycola,b
Km (nM) Kcat (min−1) Kcat/Km (min−1 nM−1)
Tg:A 0.62 +/− 0.14 0.022+/− 0.013 0.035+/− 0.023Tg:C 0.97+/− 0.16 0.033+/− 0.013 0.034+/− 0.006Tg:G 0.94+/− 0.24 0.034+/− 0.002 0.036+/− 0.009Tg:T 0.79+/− 0.18 0.035+/− 0.002 0.044+/− 0.01
a 5R,6S-Tg paired with each opposite base.b Values are the mean of three determinations+/− standard deviation.
are compact and have well-defined initiation and ter-mination sites[41]. Recently, Brummelkamp et al.[42] demonstrated that small hairpin RNAs expressedfrom the human H1 promoter may induce RNA in-terference of target genes. We used mouse embry-onic stem cells for our experiments as the compe-tency of these cell lines to carry out RNAi initiatedby long double-stranded RNAs has been established[43,44].
Using guidelines proposed by Brummelkamp andco-workers[42], we introduced a 19 bp sequence fromthe first coding exon ofmneil1as a hairpin behind themouse H1 promoter, as shown inFig. 4. The result-ing plasmid, pNEIL1HP, contains thepgk-neogenefor stable expression in ES-cells. The plasmid waselectroporated into ES-cells and G418r colonies wereisolated. ES-cell lines containing the same plasmidwithout a hairpin insert were derived and used as acontrol.
The expression level of mNEIL1 protein was de-termined in a panel of pNEIL1HP-transfected ES-cellclones. As shown inFig. 4C, most cell lines con-tained reduced levels of mNeil1 varying, at passage3, from 19 to 88% relative to wild-type. The level ofmNeil1 relative to other proteins was determined bycomparing its immunoblot signal to those generatedafter re-probing the blots with monoclonal antibodiesdirected against�-actin and PCNA. We also observedthat cell lines expressing low levels of mNeil1 grad-ually regained their ability to express the protein. Forthe�-irradiation studies reported here, cell lines wereused after 4–6 passages.
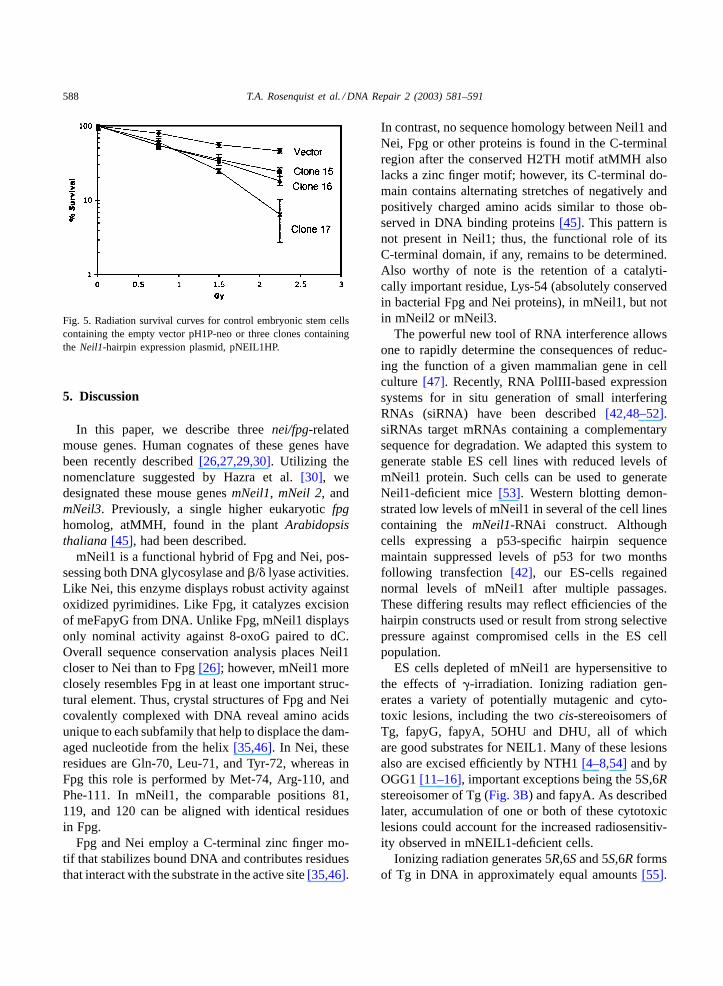
Several NEIL1HP lines showing a five-fold reduc-tion in mNeil1 relative to wild-type were selected forphenotypic characterization. Exponentially growingcells were exposed to low levels of�-irradiation andplated at low density. Colonies were counted and the
data normalized to non-irradiated cells. The survivalcurves shown inFig. 5 indicate that mNeil1-deficientlines are approximately twice as sensitive as controlcells to low doses of�-irradiation. Following 12 pas-sages, 5/5 cell lines regained their original abilityto express mNeil1 and showed normal sensitivity toradiation (data not shown).
Fig. 4. (A) Structure of theNeil1 hairpin expressing vectorpNEIL1HP. This vector contains a neomycin resistance gene(pgk-neo); the mouse H1 PolIII promoter (mH1P); and the hairpincorresponding to a portion ofNeil1 (NEIL1 hairpin). The vectorbackbone is pUC18. (B) The expressed hairpin insert fromNeil1.The first underlined sequence corresponds to nucleotides 87-105of the first Neil1 coding exon relative to the ATG start codon(amino acids 28–35 inFig. 1), the second underlined sequenceis the complement. (C) A Western blot of total protein from ei-ther control ES-cells (TC1) or 5 independent stable ES-cell linestransfected with pNEILHP (lanes 1–5). The blot was probed witha rabbit poly-clonal anti-mNeil1 antiserum (top panel). The blotwas re-probed with anti-�-actin (middle panel) and anti-PCNA(bottom panel) antibodies.
588 T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591
Fig. 5. Radiation survival curves for control embryonic stem cellscontaining the empty vector pH1P-neo or three clones containingthe Neil1-hairpin expression plasmid, pNEIL1HP.
5. Discussion
In this paper, we describe threenei/fpg-relatedmouse genes. Human cognates of these genes havebeen recently described[26,27,29,30]. Utilizing thenomenclature suggested by Hazra et al.[30], wedesignated these mouse genesmNeil1, mNeil 2, andmNeil3. Previously, a single higher eukaryoticfpghomolog, atMMH, found in the plantArabidopsisthaliana [45], had been described.
mNeil1 is a functional hybrid of Fpg and Nei, pos-sessing both DNA glycosylase and�/� lyase activities.Like Nei, this enzyme displays robust activity againstoxidized pyrimidines. Like Fpg, it catalyzes excisionof meFapyG from DNA. Unlike Fpg, mNeil1 displaysonly nominal activity against 8-oxoG paired to dC.Overall sequence conservation analysis places Neil1closer to Nei than to Fpg[26]; however, mNeil1 moreclosely resembles Fpg in at least one important struc-tural element. Thus, crystal structures of Fpg and Neicovalently complexed with DNA reveal amino acidsunique to each subfamily that help to displace the dam-aged nucleotide from the helix[35,46]. In Nei, theseresidues are Gln-70, Leu-71, and Tyr-72, whereas inFpg this role is performed by Met-74, Arg-110, andPhe-111. In mNeil1, the comparable positions 81,119, and 120 can be aligned with identical residuesin Fpg.
Fpg and Nei employ a C-terminal zinc finger mo-tif that stabilizes bound DNA and contributes residuesthat interact with the substrate in the active site[35,46].
In contrast, no sequence homology between Neil1 andNei, Fpg or other proteins is found in the C-terminalregion after the conserved H2TH motif atMMH alsolacks a zinc finger motif; however, its C-terminal do-main contains alternating stretches of negatively andpositively charged amino acids similar to those ob-served in DNA binding proteins[45]. This pattern isnot present in Neil1; thus, the functional role of itsC-terminal domain, if any, remains to be determined.Also worthy of note is the retention of a catalyti-cally important residue, Lys-54 (absolutely conservedin bacterial Fpg and Nei proteins), in mNeil1, but notin mNeil2 or mNeil3.
The powerful new tool of RNA interference allowsone to rapidly determine the consequences of reduc-ing the function of a given mammalian gene in cellculture [47]. Recently, RNA PolIII-based expressionsystems for in situ generation of small interferingRNAs (siRNA) have been described[42,48–52].siRNAs target mRNAs containing a complementarysequence for degradation. We adapted this system togenerate stable ES cell lines with reduced levels ofmNeil1 protein. Such cells can be used to generateNeil1-deficient mice[53]. Western blotting demon-strated low levels of mNeil1 in several of the cell linescontaining the mNeil1-RNAi construct. Althoughcells expressing a p53-specific hairpin sequencemaintain suppressed levels of p53 for two monthsfollowing transfection [42], our ES-cells regainednormal levels of mNeil1 after multiple passages.These differing results may reflect efficiencies of thehairpin constructs used or result from strong selectivepressure against compromised cells in the ES cellpopulation.
ES cells depleted of mNeil1 are hypersensitive tothe effects of�-irradiation. Ionizing radiation gen-erates a variety of potentially mutagenic and cyto-toxic lesions, including the twocis-stereoisomers ofTg, fapyG, fapyA, 5OHU and DHU, all of whichare good substrates for NEIL1. Many of these lesionsalso are excised efficiently by NTH1[4–8,54]and byOGG1[11–16], important exceptions being the 5S,6Rstereoisomer of Tg (Fig. 3B) and fapyA. As describedlater, accumulation of one or both of these cytotoxiclesions could account for the increased radiosensitiv-ity observed in mNEIL1-deficient cells.
Ionizing radiation generates 5R,6Sand 5S,6R formsof Tg in DNA in approximately equal amounts[55].

T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591 589
Tg-oligonucleotides generated by oxidation withMnO4 or OsO4 also contain a mixture of isomers[56,57]. Recently, improved methods were developedfor the separation of oligonucleotides containing asingle Tg stereoisomer[7,31], facilitating compar-ison of the 5R,6S- and 5S,6R-Tg stereoisomers assubstrates for BER enzymes. mNth1 preferentiallyexcises5R,6S-Tg while mNeil1 is effective in remov-ing both stereoisomers. Similar observations havebeen made for mNth1 by McTigue et al., with mNth1(unpublished data). Thus, mNeil1 contributes to theoverall repair of this potentially lethal lesion in DNA.
The cytotoxicity of Tg, fapyG and fapyA has beenattributed to the ability of these lesions to block theprogression of replicative DNA polymerases[58–60].FapyG is a substrate for mOgg1 and NTH1, whilefapyA is excised only by mNEIL1 (Rosenquist andM. Dizdaraglu, unpublished data[27]). Therefore,irradiated and oxidatively damaged DNA containstwo potentially lethal lesions repaired primarilyby Neil1.
5,6-Dihydroxycytosine (cytosine glycol) is gener-ated when DNA is exposed to ionizing radiation[6] orwhen oligonucleotides containing cytosine are treatedwith OsO4 [56]. Cytosine glycol is relatively unstablein aqueous solution, breaking down to form 5OHC andthe miscoding lesions DHU and 5OHU[61,62]. Neil1shows activity against DHU and 5OHU and may par-ticipate in the repair of these pre-mutagenic lesions inDNA.
In conclusion, a novel DNA glycosylase, Neil1, hasbeen shown to protect murine ES cells from the dele-terious effects of ionizing radiation. The presence ofthis enzyme may account for the lack of an obviousphenotype inOgg1−/− andNth −/− mice.
Acknowledgements
This research was supported by NIH ES09113 (toTAR) and NIH CA17395 and CA47995 (to APG).D.O.Z. acknowledges support from the Russian Foun-dation for Basic Research (02-04-49605) and theRussian Ministry of Education (PD02-1.4-469). Wethank Cecilia Torres for synthesizing oligonucleotides,Robert Rieger for the mass spectrometric analysis,and Carlos De los Santos and Monica McTigue forhelpful discussions.
References
[1] S.S. David, S.D. Williams, Chemistry of glycosylases andendonucleases involved in base-excision repair, Chem. Rev.98 (1998) 1221–1261.
[2] E.C. Friedberg, G.C. Walker, W. Siede, DNA Repair andMutagenesis, ASM Press, 1995.
[3] S. Wallace, Biological consequences of free radical-damagedDNA bases, Free Radic. Biol. Med. 33 (2002) 1–14.
[4] T. Hilbert, R. Boorstein, H. Kung, P. Bolton, D. Xing,R. Cunningham, G. Teebor, Purification of a mamma-lian homologue of Escherichia coli endonuclease III:identification of a bovine pyrimidine hydrate-thymine glycolDNAse/AP lyase by irreversible cross linking to a thymineglycol-containing oligoxynucleotide, Biochemistry 35 (1996)2505–2511.
[5] R. Aspinwall, D. Rothwell, T. Roldan-Arjona, C. Anselmino,C. Ward, J. Cheadle, J. Sampson, T. Lindahl, P. Harris, I.Hickson, Cloning and characterization of a functional humanhomolog of Escherichia coliendonuclease III, Proc. Natl.Acad. Sci. U.S.A. 94 (1997) 109–114.
[6] M. Dizdaroglu, B. Karahalil, S. Senturker, T. Buckley, T.Roldan-Arjona, Excision of products of oxidative DNA basedamage by human NTH1 protein, Biochemistry 38 (1999)243–246.
[7] K. Asagoshi, H. Odawara, H. Nakano, T. Miyano, H.Terato, Y. Ohyama, S. Seki, H. Ide, Comparison of substratespecificities of Escherichia coli endonuclease III and itsmouse homologue (mNTH1) using defined oligonucleotidesubstrates, Biochemistry 39 (2000) 11389–11398.
[8] L. Luna, B.M.E. Hoff, T. Rognes, E. Seeberg, Cell-cycleregulation, Mutat. Res. 460 (2000) 95–104.
[9] M. Takao, S. Kanno, T. Shiromoto, R. Hasegawa, H. Ide, S.Ikeda, A.H. Sarker, S. Seki, J.Z. Xing, X.C. Le, M. Weinfeld,K. Kobayashi, J. Miyazaki, M. Muijtjens, J.H. Hoeijmakers,Y.A. van der Horst G, Novel nuclear and mitochondrialglycosylases revealed by disruption of the mouseNth1 geneencoding an endonuclease III homolog for repair of thymineglycols, EMBO J. 21 (2002) 3486–3493.
[10] M. Ocampo, W. Chaung, D. Marenstein, M. Chan, A.Altamirano, A. Basu, R. Boorstein, R. Cunningham, G.Teebor, Targeted deletion of mNth1 reveals a novel DNArepair enzyme activity, Mol. Cell Biol. 22 (2002) 6111–6121.
[11] P.A. van der Kemp, D. Thomas, R. Barbey, R. de Oliveira,S. Boiteux, Cloning and expression in Escherichia coli of theOGG1gene ofSaccharomyces cerevisiae, which codes for aDNA glycosylase that excises 7,8-dihydro-8-oxoguanine and2,6-diamino-4-hydroxy-5-N-methylformamidopyrimidine,Proc. Natl. Acad. Sci. U.S.A. 93 (1996) 5197–5202.
[12] T.A. Rosenquist, D.O. Zharkov, A.P. Grollman, Cloningand characterization of a mammalian 8-oxoguanine DNAglycosylase, Proc. Natl. Acad. Sci. U.S.A. 94 (1997) 7429–7434.
[13] R. Lu, H.M. Nash, G.L. Verdine, A mammalian DNA repairenzyme that excises oxidatively damaged guanine maps toa locus frequently lost in lung cancer, Curr. Biol. 7 (1997)397–407.

590 T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591
[14] H. Aburatani, Y. Hippo, T. Ishida, R. Takashima, C.Matsuba, T. Kodama, M. Takao, A. Yasui, K. Yamamoto,M. Asano, K. Fukasawa, T. Yoshinari, H. Inoue, E. Ohtsuka,S. Nishimura, Cloning and characterization of mammalian8-hydroxyguanine-specific DNA glycosylase/AP-lyase, afunctional mutM homologue, Cancer Res. 57 (1997) 2151–2156.
[15] K. Arai, K. Morishita, K. Shinmura, T. Kohno, S. Kim, T.Nohmi, M. Taniwaki, S. Ohwada, J. Yokota, Cloning of ahuman homolog of the yeastOGG1 gene that is involved inthe repair of oxidative DNA damage, Oncogene 14 (1997)2857–2861.
[16] J.P. Radicella, C. Dherin, C. Desmaze, M.S. Fox, S. Boiteux,Cloning and characterization ofhOGG1, a human homologof the OGG1 gene ofSaccharomyces cerevisiae, Proc. Natl.Acad. Sci. U.S.A. 94 (1997) 8010–8015.
[17] K. Shinmura, S. Yamaguchi, T. Saitoh, T. Kohno, J. Yokota,Somatic mutations and single nucleotide polymorphisms ofbase excision repair genes involved in the repair of 8-hydroxy-guanine in damaged DNA, Cancer Lett. 166 (2001) 65–69.
[18] Y.J. Park, E.Y. Choi, J.Y. Choi, J.-G. Park, H.J. You, M.-H. Chung, Genetic changes ofhOGG1 and the activity ofoh8Gua glycosylase in colon cancer, Eur. J. Cancer 37 (2001)340–346.
[19] A. Klungland, I. Rosewell, S. Hollenbach, E. Larsen, G. Daly,B. Epe, E. Seeberg, T. Lindahl, D.E. Barnes, Accumulationof premutagenic DNA lesions in mice defective in removalof oxidative base damage, Proc. Natl. Acad. Sci. U.S.A. 96(1999) 13300–13305.
[20] O. Minowa, T. Arai, M. Hirano, Y. Monden, S. Nakai, M.Fukuda, M. Itoh, H. Takano, Y. Hippou, H. Aburatani, K.Masumura, T. Nohmi, S. Nishimura, T. Noda,Mmh/Ogg1gene inactivation results in accumulation of 8-hydroxyguaninein mice, Proc. Natl. Acad. Sci. U.S.A. 97 (2000) 4156–4161.
[21] M. Osterod, S. Hollenbach, J.G. Hengstler, D.E. Barnes, T.Lindahl, B. Epe, Age-related and tissue-specific accumulationof oxidative DNA base damage in 7,8-dihydro-8-oxoguanine-DNA glycosylase (Ogg1) deficient mice, Carcinogenesis 22(2001) 1459–1463.
[22] F. Le Page, E.E. Kwoh, A. Avrutskaya, A. Gentil, S.A.Leadon, A. Sarasin, P.K. Cooper, Transcription-coupled repairof 8-oxoguanine: requirement for XPG TFIIH, Cell 101(2000) 159–171.
[23] F. Le Page, A. Klungland, D.E. Barnes, A. Sarasin, S. Boiteux,Transcription coupled repair of 8-oxoguanine in murine cells:the ogg1 protein is required for repair in nontranscribedsequences but not in transcribed sequences, Proc. Natl. Acad.Sci. U.S.A. 97 (2000) 8397–8402.
[24] T.K. Hazra, T. Izumi, L. Maidt, R.A. Floyd, S. Mitra, Thepresence of two distinct 8-oxoguanine repair enzymes inhuman cells: their potential complementary roles in preventingmutation, Nucleic Acids Res. 26 (1998) 5116–5122.
[25] T. Bessho, K. Tano, H. Kasai, E. Ohtsuka, S. Nishimura,Evidence for two DNA repair enzymes for 8-hydroxyguanine(7,8-dihydro-8-oxoguanine) in human cells, J. Biol. Chem.268 (1993) 19416–19421.
[26] V. Bandaru, S. Sunkara, S.S. Wallace, J.P. Bond, A novelhuman glycosylase that removes oxidative DNA damage and
is homologous toEscherichia coliendonuclease VIII, DNARepair 1 (2002) 517–529.
[27] T.K. Hazra, T. Izumi, I. Boldogh, B. Imhoff, Y.W. Kow, P.Jaruga, M. Dizdaroglu, S. Mitra, Identification and charac-terization of a human DNA glycosylase for repair of modifiedbases in oxidatively damaged DNA, Proc. Natl. Acad. Sci.U.S.A. 99 (2002) 3523–3528.
[28] I. Morland, V. Rolseth, L. Luna, T. Rognes, M. Bjoras,E. Seeberg, Human DNA glycosylases of the bacterialFpg/MutM superfamily: an alternative pathway for the repairof 8-oxoguanine and other oxidation products in DNA,Nucleic Acids Res. 30 (2002) 4926–4936.
[29] M. Takao, S. Kanno, K. Kobayashi, Q. Zhang, S. Yonei,G. van der Horst, A. Yasui, A back-up glycosylase in Nth1knock-out mice is a functional Nei (endonuclease VIII)homologues, J. Biol. Chem. 277 (2002) 42205–42213.
[30] T.K. Hazra, Y.W. Kow, Z. Hatahet, B. Imhoff, I. Boldogh,S.K. Mokkapati, S. Mitra, T. Izumi, Identification andcharacterization of a novel human DNA glysosylase for repairof cytosine derived lesions, J. Biol. Chem. (2002).
[31] R. Rieger, C. Iden, 49th ASMS Conference on Mass Spectro-scopy and Allied Topics, Chicago, 2001, pp. ThPK 273.
[32] A.K. Basu, E.L. Loechler, S.A. Leadon, J.M. Essigmann,Genetic effects of thymine glycol: site-specific mutagenesisand molecular modeling studies, Proc. Natl. Acad. Sci. U.S.A.86 (1989) 7677–7681.
[33] J.Y. Kao, I. Goljer, T.A. Phan, P.H. Bolton, Characterizationof the effects of a thymine glycol residue on the structure,dynamics, and stability of duplex DNA by NMR, J. Biol.Chem. 268 (1993) 17787–17793.
[34] J. Tchou, H. Kasai, S. Shibutani, M.H. Chung, J. Laval, A.P.Grollman, S. Nishimura, 8-oxoguanine (8-hydroxyguanine)DNA glycosylase and its substrate specificity, Proc. Natl.Acad. Sci. U.S.A. 88 (1991) 4690–4694.
[35] R. Gilboa, D. Zharkov, G. Golan, A. Fernandes, S.Gerchman, E. Matz, J. Kycia, A. Grollman, G. Shoham,Structure of formamidopyrimidine-DNA glycosylase cova-lently complexed to DNA, J. Biol. Chem. 277 (2002) 19811–19816.
[36] D. Zharkov, T. Rosenquist, S. Gerchman, A. Grollman,Substrate specificity and reaction mechanism of murine8-oxoguanine-DNA glycosylase, J. Biol. Chem. 275 (2000)28607–28617.
[37] C. Deng, A. Wynshaw-Boris, M. Shen, C. Daugherty, D.Ornitz, P. Leder, Murine FGFR-1 is required for earlypostimplantation growth and axial organization, Genes Dev.8 (1994) 3045–3057.
[38] O. Lavrukhin, R. Lloyd, Involvement of phylogeneticallyconserved acidic amino acid residues in catalysis by an oxi-dative DNA damage enzyme formamidopyrimidine glyco-sylase, Biochemistry 39 (2000) 15266–15271.
[39] M. Sugahara, T. Mikawa, T. Kumasaka, M. Yamamoto, R.Kato, K. Fukuyama, Y. Inoue, S. Kuramitsu, Crystal Structureof a repair enzyme of oxidatively damaged DNA, MutM(Fpg), from an exteme thermophile, Thermus thermophilusHB8, EMBO J. 19 (2000) 3857–3869.
[40] N.H. Robin, J.H. Nadeau, Disorganization in Mice andHumans, Am. J. Med. Genet. 101 (2001) 334–338.

T.A. Rosenquist et al. / DNA Repair 2 (2003) 581–591 591
[41] E. Myslinski, J.-C. Ame, A. Krol, P. Carpon, An unusuallycompact external promoter for RNA polymerase III trans-cription of the human H1 RNA gene, Nucleic Acids Res. 29(2001) 2502–2509.
[42] T.R. Brummelkamp, R. Bernards, R.A. Agami, System forstable expression of short interfering RNAs in mammaliancells, Science 296 (2002) 550–553.
[43] S. Yang, S. Tutton, E. Pierce, K. Yoon, Specific double-stranded RNA interference in undifferentiated mouseembryonic stem cells, Mol. Cell Biol. 21 (2001) 7807–7816.
[44] E. Billy, V. Brondani, H. Zhang, U. Muller, W. Filipowicz,Specific interference with gene expression induced bylong, Proc. Natl. Acad. Sci. U.S.A. 98 (2001) 14428–14433.
[45] T. Ohtsubo, O. Matsuda, K. Iba, I. Terashima, M. Sekiguchi,Y. Nakabeppu, Molecular cloning of AtMMH, an Arabidopsisthaliana ortholog of theEscherichia colimutM gene, andanalysis of functional domains of its product, Mol. Gen.Genet. 259 (1998) 577–590.
[46] D. Zharkov, G. Golan, R. Gilboa, A. Fernandes, S.E.Gerchman, J.H. Kycia, R.A. Rieger, A.P. Grollman,G. Shoham, Structural analysis of anEscherichia coliendonuclease VIII covalent reaction intermediate, EMBO J.21 (2002) 789–800.
[47] M. McManus, P. Sharp, Gene silencing in mammals by smallinterfering RNAs, Nature Rev. Genet. 3 (2002) 737–747.
[48] N. Lee, T. Dohjima, G. Bauer, H. Li, M. Li, A. Ehsani, P.Salvaterra, J. Rossi, Expression of small interfering RNAstargeted against HIV-1 rev transcripts in human cells, NatureBiotechnol. 20 (2002) 500–505.
[49] P. Paddison, A. Caudy, E. Bernstein, G. Hannon, D. Conklin,Short Hairpin (shRNAs) unduce sequence-specific silencingin mammalian cells, Genes Dev. 16 (2002) 948–958.
[50] C. Paul, P. Good, I. Winer, D. Engelke, Effective expressionof small interfering RNA in human cells, Nat. Biotechnol.20 (2002) 505–508.
[51] G. Sui, C. Soohoo, E. Affar, F. Gay, Y. Shi, W. Forrester,A DNA vector-based RNAi technology to suppress geneexpression in mammalian cells, Proc. Natl. Acad. Sci. U.S.A.99 (2002) 5515–5520.
[52] J.-Y. Yu, S. DeRuiter, D. Turner, RNA interference byexpression of short-interfering RNAs and hairpin RNAs in
mammalian cells, Proc. Natl. Acad. Sci. U.S.A. 99 (2002)6047–6052.
[53] M. Carmell, L. Zhang, D. Conklin, G. Hannon, T. Rosenquist,Germline transmission of RNAi in mice, Nature StructuralBiol. 10 (2003) 91–92.
[54] L. Eide, L. Luna, E. Gustad, P.T. Henderson, J.M. Essigmann,B. Demple, E. Seeberg, Human endonuclease III actsprefernetially on DNA damage opposite guanine residues inDNA, Biochemistry 40 (2001) 6653–6659.
[55] G. Teebor, A. Cummings, K. Frenkel, A. Shaw, L. Voituriez,J. Cadet, Quantitative measurement of the diastereoisomersof cis thymidine glycol in�-irradiated DNA, Free. Radic.Res. Commun. 2 (1987) 303–309.
[56] P. Fischhaber, V. Gerlach, W. Feaver, Z. Hatahet, S.S. Wallace,E. Friedberg, Human DNA polymerase k bypasses andextends beyond thymine glycols during translesion synthesisin vitro, preferentially incorporating corect nucleotides, J.Biol. Chem. 277 (2002) 37604–37611.
[57] M. Lustig, J. Cadet, R. Boorstein, T. GW, Synthesis ofthe diastereomers of thymidine glycol, determination ofconcentrations and rates of interconversion of theircis–transepimers at equilibrium and demonstration of differential alkalilability within DNA, Nucleic Acids Res. 20 (1992) 4839–4845.
[58] J. Clark, G. Beardsley, Template length, sequence context,and 3′–5′ exonuclease activity modulate replicative bypassof thymine glycol lesions in vitro, Biochemistry 28 (1989)775–779.
[59] D. Wang, D. Kreutzer, J. Essigmann, Mutagenicity and repairof oxidative DNA damage: insights from studies using definedlesions, Mutat. Res. 400 (1998) 99–115.
[60] K. Asagoshi, H. Terato, Y. Ohyama, H. Ide, Effects ofa guanine-derived formamidopyrimidine lesion on DNAreplication: translesion DNA synthesis, nucleotide insertion,and extension kinetics, J. Biol. Chem. 277 (2002) 14589–14597.
[61] S. Tremblay, D. Thierry, J. Cadet, J.R. Wagner,2′-Deoxycytidine glycols, a missing link in the freeradical-mediated oxidation of DNA, J. Biol. Chem. 274 (1999)20833–20838.
[62] D.A. Kreutzer, J.M. Essigman, Oxidized, deaminatedcytosines are a source of C→ T transitions in vivo, Proc.Natl. Acad. Sci. U.S.A. 95 (1998) 3578–3582.
Related Documents











