Plant Physiol. (1974) 54, 737-743 The Mode of Incorporation of 6-Benzylaminopurine into Tobacco Callus Transfer Rilbonucleic Acid ATDOUBLE LABELING DETERMINATION' Received for publication March 26. 1974 and in revised form Junie 13, 1974 GRAHAM C. WALKER AND NELSON J. LEONARD School of Chemical Sciences, University of Illinois, Urbana, Illinois 61801 DONALD J. ARMSTRONG, NORIMOTO MURAI, AND FOLKE SKOOG Institute of Plant Development, University of Wisconsin, Madison, Wisconsin 53706 ABSTRACT Cytokinin-dependent tobacco (Nicotiana tabacumn) tissue was grown in the presence of N'-benzyladenine (6-benzylamino- purine) labeled in the phenyl ring with 'H and in the 8-position of the purine ring with "4C. The 'H/'4C ratio of N'-benzyladeno- sine recovered from labeled tobacco callus transfer RNA prep- arations was compared with the corresponding ratio for N6- benzyladenine supplied in the medium. By this means, with suitable controls, the incorporation of the synthetic cytokinin, N6-benzyladenine, into the transfer RNA of tobacco callus tissue was shown to involve the intact moiety. The observed level of incorporation was up to one N6-benzyladenosine molecule per 104 transfer RNA molecules. Cytokinins have been found free in higher plants and plant pathogens (17, 29) as well as in the tRNAs of many organisms. Of the specific tRNAs that have been studied, only those which read a codon on the mRNA beginning with U (uridine) contain a cytokinin, and in those tRNAs which have been sequenced, the cytokinin is located adjacent to the anticodon on the 3'-side (24, 29, 30, 34). There has been considerable speculation as to whether the presence of cytokinins in tRNA is related to their mode of action in regulation of growth, but attempts to measure the incorporation of exogenous cytokinins into plant tissue tRNA have led to conflicting results. McCalla et al. (22) first reported radioactivity in RNA of plants treated with bzlPAde-8-_4C; Burrows et al. (4), Selivankina et al. (28), van Onckelen et al. (32), and Fox et al. (13, 14) reported the incorporation of cytokinins into sRNA although the purity of some of the sRNA was later questioned by Key (19). Dyson (8) and Srivastava (31) reported the incorporation of cytokinins 1 This research was supported at the University of Wisconsin by Research Grants GB-25812, GB-35260X, and GB-35260X1 from the National Science Foundation and by the Research Committee of the Graduate School with funds from the Wisconsin Alumni Research Foundation. It was supported at the University of Illinois by Research Grant GM-05829 from the National Institutes of Health. into both tRNA and rRNA, whereas Hall (16), Richmond et al. (26), and Bezemer-Sybrandy et al. (3) failed to detect any incorporation into RNA. Kende and Tavares (18), on the basis of measurements of incorporation of radioactive 6- benzylaminopurine and 6-benzylamino-9-methylpurine into soybean callus, claim that the methyl substituent in the 9 posi- tion inhibited incorporation of the latter cytokinin into RNA without inhibiting its biological activity. The validity of this report is subject to limitations in procedures employed. Fox et al.- (15) claim that the 9-methyl group is released from the 6-benzylamino-9-methylpurine in contact with tobacco and soybean callus and that in tests on tobacco tissue some 6- benzylaminopurine was recovered. Recently, Elliott and Murray (10) have reported that bzl6Ade2 is not incorporated into tRNA but that bzl6A, apparently arising from trans- benzylation of an adenosine in the tRNA, is found at a level of one molecule of bzl6A/750-2210 molecules of tRNA in soy- bean tissue grown on medium supplemented with bzl6Ade. Plant tissues requiring exogenous cytokinins in order to grow contain potentially active cytokinins as constituents of their tRNA. Chen and Hall (5) supplied the cytokinin require- ment of tobacco tissue with i6A yet found that the iA in the tRNA was not incorporated directly but arose from modifica- tion of the preformed tRNA. Recently, Burrows et al. (4) showed that from a tRNA preparation of tobacco callus tissue requiring exogenous cytokinin which had been grown on medium containing bzl6Ade, three endogenous cytokinins, 6-(4-hydroxy-3 -methyl-2-butenylamino)-9-,/-D-ribofuranosyl- purine, 6-(4-hydroxy-3-methyl-2-butenylamino)-2-methylthio- 9-/3-D-ribofuranosylpurine, and iA, as well as bzl6A, could be isolated and conclusively identified. The identification of bzl'A was of particular interest since the mechanism of incorporation was unclear. In organisms examined to date, the i'A moieties in the tRNA arise by transfer of the isopentenyl group of _2_ isopentenyl pyrophosphate to specific adenosine residues in the tRNA molecules (30). The enzyme or enzymes that catalyze this reaction, AY-isopentenyl pyrophosphate: transfer ribo- nucleic acid-A2-isopentenyltransferase, have been isolated from 'Abbreviations: bzl6Ade: N6-benzyladenine; bzl6Ado or bzl6A: N6-benzyladenosine; i6Ade: N6(A'-isopentenyl)adenine or 6-(3- methyl-2-butenylamino)purine, previously called 2iP; i6Ado or iPA: NM-(A2-isopentenyl)adenosine, previously called 2iPA; (G), as in benzoic acid-'H(G), signifies general label; CTAB: cetyltri- methylammonium bromide. 737

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Plant Physiol. (1974) 54, 737-743
The Mode of Incorporation of 6-Benzylaminopurine into TobaccoCallus Transfer Rilbonucleic AcidATDOUBLE LABELING DETERMINATION'
Received for publication March 26. 1974 and in revised form Junie 13, 1974
GRAHAM C. WALKER AND NELSON J. LEONARDSchool of Chemical Sciences, University of Illinois, Urbana, Illinois 61801
DONALD J. ARMSTRONG, NORIMOTO MURAI, AND FOLKE SKOOGInstitute of Plant Development, University of Wisconsin, Madison, Wisconsin 53706
ABSTRACT
Cytokinin-dependent tobacco (Nicotiana tabacumn) tissue wasgrown in the presence of N'-benzyladenine (6-benzylamino-purine) labeled in the phenyl ring with 'H and in the 8-positionof the purine ring with "4C. The 'H/'4C ratio of N'-benzyladeno-sine recovered from labeled tobacco callus transfer RNA prep-arations was compared with the corresponding ratio for N6-benzyladenine supplied in the medium. By this means, withsuitable controls, the incorporation of the synthetic cytokinin,N6-benzyladenine, into the transfer RNA of tobacco callus tissuewas shown to involve the intact moiety. The observed level ofincorporation was up to one N6-benzyladenosine molecule per104 transfer RNA molecules.
Cytokinins have been found free in higher plants and plantpathogens (17, 29) as well as in the tRNAs of many organisms.Of the specific tRNAs that have been studied, only thosewhich read a codon on the mRNA beginning with U (uridine)contain a cytokinin, and in those tRNAs which have beensequenced, the cytokinin is located adjacent to the anticodonon the 3'-side (24, 29, 30, 34). There has been considerablespeculation as to whether the presence of cytokinins in tRNAis related to their mode of action in regulation of growth, butattempts to measure the incorporation of exogenous cytokininsinto plant tissue tRNA have led to conflicting results. McCallaet al. (22) first reported radioactivity in RNA of plants treatedwith bzlPAde-8-_4C; Burrows et al. (4), Selivankina et al. (28),van Onckelen et al. (32), and Fox et al. (13, 14) reported theincorporation of cytokinins into sRNA although the purity ofsome of the sRNA was later questioned by Key (19). Dyson(8) and Srivastava (31) reported the incorporation of cytokinins
1 This research was supported at the University of Wisconsin byResearch Grants GB-25812, GB-35260X, and GB-35260X1 fromthe National Science Foundation and by the Research Committeeof the Graduate School with funds from the Wisconsin AlumniResearch Foundation. It was supported at the University of Illinoisby Research Grant GM-05829 from the National Institutes ofHealth.
into both tRNA and rRNA, whereas Hall (16), Richmondet al. (26), and Bezemer-Sybrandy et al. (3) failed to detectany incorporation into RNA. Kende and Tavares (18), on thebasis of measurements of incorporation of radioactive 6-benzylaminopurine and 6-benzylamino-9-methylpurine intosoybean callus, claim that the methyl substituent in the 9 posi-tion inhibited incorporation of the latter cytokinin into RNAwithout inhibiting its biological activity. The validity of thisreport is subject to limitations in procedures employed. Foxet al.- (15) claim that the 9-methyl group is released from the6-benzylamino-9-methylpurine in contact with tobacco andsoybean callus and that in tests on tobacco tissue some 6-benzylaminopurine was recovered. Recently, Elliott andMurray (10) have reported that bzl6Ade2 is not incorporatedinto tRNA but that bzl6A, apparently arising from trans-benzylation of an adenosine in the tRNA, is found at a level ofone molecule of bzl6A/750-2210 molecules of tRNA in soy-bean tissue grown on medium supplemented with bzl6Ade.
Plant tissues requiring exogenous cytokinins in order togrow contain potentially active cytokinins as constituents oftheir tRNA. Chen and Hall (5) supplied the cytokinin require-ment of tobacco tissue with i6A yet found that the iA in thetRNA was not incorporated directly but arose from modifica-tion of the preformed tRNA. Recently, Burrows et al. (4)showed that from a tRNA preparation of tobacco callustissue requiring exogenous cytokinin which had been grownon medium containing bzl6Ade, three endogenous cytokinins,6-(4-hydroxy-3 -methyl-2-butenylamino)-9-,/-D-ribofuranosyl-purine, 6-(4-hydroxy-3-methyl-2-butenylamino)-2-methylthio-9-/3-D-ribofuranosylpurine, and iA, as well as bzl6A, could beisolated and conclusively identified. The identification of bzl'Awas of particular interest since the mechanism of incorporationwas unclear. In organisms examined to date, the i'A moietiesin the tRNA arise by transfer of the isopentenyl group of _2_isopentenyl pyrophosphate to specific adenosine residues inthe tRNA molecules (30). The enzyme or enzymes that catalyzethis reaction, AY-isopentenyl pyrophosphate: transfer ribo-nucleic acid-A2-isopentenyltransferase, have been isolated from
'Abbreviations: bzl6Ade: N6-benzyladenine; bzl6Ado or bzl6A:N6-benzyladenosine; i6Ade: N6(A'-isopentenyl)adenine or 6-(3-methyl-2-butenylamino)purine, previously called 2iP; i6Ado oriPA: NM-(A2-isopentenyl)adenosine, previously called 2iPA; (G), asin benzoic acid-'H(G), signifies general label; CTAB: cetyltri-methylammonium bromide.
737

Plant Physiol. Vol. 54. 19/4
yeast (20), rat liver (12), and E. coli (2, 27); Chen and Hall(5) have reported that this enzyme activity is present in cyto-kinin-dependent tobacco callus tissue. The 2-isopentenylpyrophosphate is derived from mevalonic acid (5, 25), butmevalonic acid itself does not satisfy the cytokinin require-ments of the plant tissues (23). Studies on the short termmetabolism of bzl6Ade have shown that it is in part degradedand in part rapidly transformed to its riboside and ribotideand more slowly to a stable 7-glucofuranosyl derivative (3, 7,9).A double labeling experiment has now been performed to
determine whether the incorporation of bzl6Ade into tobaccocallus tRNA involves the entire base or occurs by transfer ofthe benzyl sidechain to adenosine residues in the RNA. Cyto-kinin-dependent tobacco callus tissue was grown in thepresence of bzl6Ade labeled in the phenyl ring with 3H and inthe 8-position of the purine ring with "4C. The 3H/14C ratio ofbzl6A recovered from labeled tobacco callus tRNA prepara-tions was compared with the corresponding ratio for bzl6Adesupplied in the medium.
MATERIALS AND METHODS
6-Benzylaminopurine-8-'4C. Adenine-8-'4C (8.4 mg, 1 mCi,16.0 mCi/mmole), 8.4 mg (0.062 mmole) freshly preparedbenzylamine hydrochloride, and 0.5 ml (46 mmole) of freshlydistilled benzylamine were heated in a sealed ampoule at 175C overnight. The contents of the ampoule were then trans-ferred to a 15-ml centrifuge tube. 13 ml of hexane were added,and the solution was mixed with a Pasteur pipette. Aftercentrifugation the oily precipitate was washed with 3 ml ofhexane and recentrifuged. The semisolid brown oil was thendissolved in 0.75 ml of ethanol, and 8 ml of anhydrous etherwere added. The white precipitate which formed was removedby centrifugation, washed with 3 ml of ether, and the ethersupernatants were combined. After removal of the ether, theresidue was dissolved in 2 ml of chloroform and the solutionwas concentrated to a volume of about 0.1 ml before beingapplied to a silica column (2.5 g) equilibrated with chloroform-methanol (99:1). Following elution with 15 ml of the samesolvent and 21 ml of chloroform-methanol (98:2) the 6-benzylaminopurine-8-'4C was eluted from the column with 40ml of chloroform-methanol (95:5). The solvent was removedin vacuo to yield 6-benzyaminopurine-8-14C (6.9 mg, 49%,17.6 mCi/mmole) as a yellowish white powder; El'OH 270nm, mA H 245; chromatographically homogeneous as deter-mined by TLC on silica and on alumina in chloroform-methanol (80:20); radiochemically pure as established byTLC on silica gel followed by counting of the scraped-offsegments of the plate in dioxane scintillator; practice runs withunlabeled adenine gave material with mp 231 to 234 C mp231-232 C in ref. 13) and a nuclear magnetic resonancecorresponding to 6-benzylaminopurine.Benzylamine Hydrochloride-benzene-3H(G). Benzoic acid-
3H(G) (48 mg, 12.5 mCi, 29.2 mCi/mmole) and 0.5 ml ofthionyl chloride were heated under ruflux for 1 hr on thesteam bath. After partial removal of excess thionyl chloride bydistillation. 2 ml of ether were added followed by 2 ml of con-centrated aqueous ammonia (saturated with ammonia at -10C prior to use). The aqueous layer was extracted with etherand then several times with chloroform. Evaporation of thecombined organic layers yielded benzamide-3H(G) (21 mg,42%): chromatographically homogeneous on silica gel inchloroform-methanol (95: 5); in unlabeled practice runs thematerial produced by this method had an infrared spectrumidentical with authentic benzamide. The benzamide-3H(G)
(21 mg, 0.17 mmole) in 8 ml of ether was treated with 75 mgof lithium aluminum hydride (2.1 mmoles) in several portionsand refluxed gently for 12 hr. Then 75 dl of water, 75 ,ul of15% NaOH, and 225 ,ul of water were added successively,and the mixture was refluxed until the precipitate settled. Afterwashing the precipitate with more ether, the combined etherlayers were dried with MgSO4 and concentrated to 7.5 ml witha gentle stream of nitrogen. Passage of dry HCI over thesurface of the ether led to the precipitation of benzylaminehydrochloride-benzene-3H(G) (18 mg, 31 % from benzoic acid)as a white powder which was collected by centrifugation andwas chromatographically homogeneous on silica gel in chloro-form-methanol (95:5). In unlabeled practice runs this methodgave material with an infrared spectrum identical to that ofauthentic benzylamine hydrochloride.
6-Benzylaminopurine-benzene-3H(G). A mixture of thebenzylamine hydrochloride-benzene-3H(G) (18 mg, 0.125mmole, 29.2 mCi/mmole) described above, 20 mg of 6-chloro-purine (0.13 mmole), 0.15 ml of triethylamine (2.0 mmole),and 2 ml of 1-butanol was refluxed at 232 to 235 C for 3 hr.After removal of the 1-butanol in vaciuo, 5 ml of water wereadded. the solution was taken to dryness in vacuo, and theprocedure was repeated with 5 ml of absolute ethanol. Theresidue was mixed with a small amount of chloroform-methanol (99: 1), and the inhomogeneous mixture was appliedto an alumina column (8 g) equilibrated with the same solvent.Elution with chloroform-methanol (98:2) yielded 6-benzyl-aminopurine-benzene-3H(G) (22 mg, 78%, 28.3 mCi/mmole)as a white powder; XEtOH 270 nm, XEmt.H 240; chromatograph-ically homogeneous on silica gel thin layers in chloroform-methanol (95:5) and on alumina thin layers in chloroform-methanol (80:20); radiochemically pure as determined by TLSunder the preceding conditions followed by counting of thescraped-off segments of the plates; in practice runs materialprepared by this method had mp 231 to 233 C (mp 231-232 Cin ref. 13).Growth of Tobacco Tissue Cultures. Tobacco callus from
stock cultures (Nicotiana tabacuin var. Wisconsin No. 38)grown on RM- 1965 medium (21) containing 11.4 [uM IAA andunlabeled 0.044 uM bzl6Ade for at least one transfer was cul-tured on RM-1965 medium (21) containing 11.4 ,uM IAA and0.044 liM 3H:'4C-bzl6Ade (3H/'4C = 9.15/1). The labeledbzl"Ade was added to the medium prior to autoclaving. Fivepieces of callus (about 30 mg each) were planted per 125-mlErlenmeyer flask containing 50 ml of medium. The tissue washarvested after 24 to 28 days growth on labeled medium. Theaverage age of the cultures at harvest was 26 days, and theaverage yield of tissue (fresh weight) was 5.3 g per flask. Atthis time the tissue was growing and had achieved less thanhalf of the final yield possible on the medium.
Isolation of tRNA. The isolation of the tobacco callus tRNA(3500 A2- units from 14.2 kg of tissue) was carried out asfollows. The tissue was homogenized (Sorvall Omnimixer, 5min) in 0.33 tissue volumes (assuming 1 g of tissue = 1 ml) of0.4 M potassium acetate (pH 5.5, glacial acetic acid) and 0.67tissue volumes of buffer-saturated phenol. The homogenatewas stirred for 30 min at room temperature. The aqueous phasewas recovered by centrifugation (300g, 30 min) and stirredfor 30 min with 0.33 volume of fresh buffer-saturated phenol.After centrifugation, the RNA was precipitated from theaqueous phase by the addition of 2 volumes of cold ethanol.All subsequent steps were performed in the cold except asindicated. Supernatants and precipitates were separated bycentrifugation at 7500g for 30 min except as indicated. Theethanol precipitate from above was washed by resuspendingin 70%c ethanol containing 0.1 M sodium acetate (pH 6). The
738 WALKER ET AL.

Plant Physiol. Vol. 54, 1974 INCORPORATION OF 6-BENZYLAMINOPURINE INTO tRNA
washed pellet was drained as dry as possible and extracted twicewith 2 M potassium acetate (pH 6, glacial acetic acid). Foreach extraction, the pellet was suspended in 50 ml of solutionper kg of original tissue and stirred for 30 min. The super-natants from the two extractions were combined, and theRNA was precipitated with 2 volumes of ethanol. The ethanolprecipitate was washed as described above and extracted twicewith 0.5% cetyltrimethylammonium bromide in 0.45 M NaCl.For each extraction, the precipitate was suspended in 25 mlof CTAB solution per kg of original tissue and stirred for 30min. The cloudy supernatants from the two extractions werecombined and centrifuged for 1 hr at 10,000g. Transfer RNAwas precipitated from the supernatant of the last centrifuga-tion by dilution with 2 volumes of 0.5% CTAB in distilledwater (room temperature). The CTA-RNA precipitate wasconverted to the sodium form by repeated suspension in 0.4M sodium acetate (pH 6) followed by precipitation with 3volumes of ethanol. Washing was continued until the pelletdissolved in the acetate buffer and foam was absent. The RNAwas then dissolved in 0.1 M tris-HCl buffer (pH 7.3) contain-ing 0.1 M NaCl and applied to a DEAE-cellulose column(about 5 ml bed volume per kg tissue) equilibrated with thesame buffer. The column was washed with 0.1 M tris-HCl (pH7.3) containing 0.2 M NaCl (at least 30 bed volumes), and thetRNA eluted with 1 M NaCl in the same buffer. The tRNA wasrecovered from the column eluate by precipitation withethanol. The final preparation contained 4S RNA and asmall amount of SS RNA as judged by polyacrylamide gelelectrophoresis. Some carbohydrate material was present, butthe preparation was essentially free of high mol wt RNA andDNA. The final yield was about 250 A2, units per kg tissue.
Isolation of bzlrA from tRNA Preparations. Tobacco callustRNA was hydrolyzed to nucleosides with crude snake venom(Crotalus adamanteus, Sigma) and alkaline phosphatase (calfintestinal mucosa, type II, Sigma) (4). Hydrolysis was termi-nated after 24 hr by adjusting the pH to 6 and adding 2volumes of ethanol. The ethanol solution was chilled at 4 Cfor 1 hr and then centrifuged (15,OOOg, 20 min). The super-natant was taken to dryness in vacuo at 37 C. The nucleosidepreparation was fractionated according to the procedure ofArmstrong et al. (1). The dry solids from above were extracted6 times (10 min each) with water-saturated ethyl acetate (1 mlof solvent per 600 A., units tRNA for each extraction). Thecombined extracts were evaporated to dryness in vacuo atroom temperature. The ethyl acetate soluble nucleosides thusobtained were fractionated by chromatography on SephadexLH-20 columns in 35% aqueous ethanol (Fig. 1, Table II).Double distilled water and redistilled ethanol were used for allchromatography.
Counting Procedure. Aliquots of column fractions wereevaporated to dryness in scintillation vials and counted on aPackard Tri-Carb liquid scintillation spectrophotometer Model2003. Aquasol (New England Nuclear) was used as thescintillation fluid. The counting efficiency of "C was about51% in the "C channel and about 14% in the (CH + 14C)channel, whereas 'H was counted at an efficiency of about 32%.Count data were corrected for quenching using a method ofexternal standardization with an external source. UnhydrolyzedtRNA samples and the residue from the ethyl acetate ex-traction were combusted to "CO2 and 'H2O, which were thencounted separately and corrected for quenching by the use ofan internal standard.Rechromatography of the Isolated bzl6A. Eighty per cent of
the material from fraction VI, Figure 1. was taken to drynessand redissolved in 2 ml of double distilled water containingunlabeled bzl'A. A 1-ml aliquot of this sample was applied to
a Sephadex LH-20 column (30 g, 1.9 X 39 cm, 110 ml bedvolume) equilibrated with double distilled water. Six-ml frac-tions were collected. The fractions were pooled as indicated(Fig. 2) for determination of radioactivity (Table III).
Denaturation Experiment. A sample of the tRNA isolatedfrom tobacco callus grown on double labeled bzl'Ade, esti-mated to contain 500 A. units, was dissolved in 4.5 ml of 0.01M sodium acetate (pH 5, glacial acetic acid) containing 0.01 MNaCl and unlabeled bzl'A (0.75 mg) and bzl'AMP (1 mg).This solution heated slowly to 85 C and held at that tempera-ture for 10 min to melt the tRNA. The solution was cooled toroom temperature and the buffer concentration was adjusted bythe addition of 0.5 ml of 1 M sodium acetate (pH 5) contain-ing 1 M NaCl. This sample (bzl'A, bzl'AMP, and tRNA) waschromatographed on a Sephadex G-25 column (50 g, 2.6 x46 cm, 244 ml bed volume) in 0.1 M sodium acetate (pH 5)containing 0.1 M NaCl (Fig. 3). The bzl'A and bzl'AMP peaksfrom the gel filtration column were pooled separately andconcentrated to about 10 ml each by evaporation at 40 Cunder reduced pressure. The bzl'A sample was then desaltedon Sephadex G-25 in distilled water. The bzl'AMP wasdesalted on Sephadex LH-20 in distilled water. The tRNA fromthe column (475 A2., units) was recovered by ethanol precipita-tion. It was then hydrolyzed to nucleosides, and bzl'A wasisolated from the hydrolysate by ethyl acetate extraction andSephadex LH-20 chromatography in 35% ethanol (Fig. 4)as described above. Fractions from the LH-20 column andthe desalted compounds from the gel filtration column weretaken to dryness in scintillation vials and were counted in theusual manner. The results are shown in Table IV.
RESULTSIn order to investigate the mode of incorporation of bzl6Ade
into tRNA of tobacco callus tissue, the synthesis of the doublelabeled compound was undertaken. A tritium label waschosen for the benzyl group and an 8-"C label for the purinering because of a desire to avoid the use of the potentiallyexchangeable (11) 8-3H label which had been used in some pre-vious work (26). The 6-benzylaminopurine-8-"C was syn-thesized from adenine-8-"C and benzylamine in the presenceof an acid catalyst by a modification of the exchange amina-tion procedure of Whitehead and Traverso (33). The 6-benzyl-aminopurine-benzene-'H(G) was prepared from benzylamine-benzene-'H(G). Benzylamine hydrochloride-benzene-3H(G) wassynthesized from benzoic acid-3H(G) by conversion to the acidchloride, then to the amide, followed by lithium aluminumhydride reduction to yield benzylamine-benzene-3H(G), whichwas collected as the hydrochloride. Reaction of the amine with6-chloropurine using a modification of the method of Daly andChristensen (6) yielded 6-benzylaminopurine-benzene-3H(G).
The two compounds were mixed to give a 3H/"C ratio of9.15/1. The higher tritium content was designed to minimizeerror due to "C overlap into the 3H channel during liquidscintillation counting. The labeled compound was incor-porated into the growth medium for tobacco callus at a con-centration of 0.044 /M bzl6Ade. A total of 3500 A. units oftRNA was isolated from 14.2 kg of tobacco callus tissue. Theethyl acetate soluble nucleosides from 3000 A260 units of thispreparation were obtained by water-saturated ethyl acetateextraction. The radioactivity recovered in this fraction as wellas in the crude tRNA is shown in Table I. The samples oftRNA and the ethyl acetate residue were combusted to CO2and H,O before counting; the efficiency was determined by theuse of internal standards. Apparently there was considerablerandomization of the label into many compounds during thegrowth of the tobacco callus tissue.
739

Plant Physiol. Vol. 54, 1974
The ethyl acetate soluble nucleosides were then fractionatedby chromatography on a Sephadex LH-20 column in 35%ethanol (1). The UV absorption profile from the column isshown in Figure 1 along with a standard profile showing theelution volumes of bzl6A, bzl6Ade, and 2-methylthio-bzl6A. Theradioactivity in each fraction was determined by a methodemploying standardization with an external source and theresults are presented in Table II. Fraction III contains themajor unmodified nucleosides, and the enrichment of 14C re-flects the extensive scavenging of the purine ring observed byFox (13. 14) and McCalla (22). Most of the radioactivityeluted is in fraction VI (corresponding to bzl'A). This fractionwas well separated from the elution point of bzl6Ade andcontained very little UV-absorbing material. No significantcounts were observed in fractions IX and X correspondingto the elution point of the 2-methylthio derivative of bzl6A.As shown in Table II the 3H/'4C ratio in the fraction cor-responding to bzl6A was virtually the same as that of theoriginal bzl6Ade in the growth medium for the tobacco callustissue. The level of incorporation was about 1 mole bzl6A/ I0Omoles tRNA. As a further check on the identity and radio-chemical purity of the isolated bzl6A, a sample of unlabeled
Table 1. Inicorporationi ofDolible Labelec bzl6Ade in Eth-l!I Acetaite-so/Ifble Nlicleosidles from Tobacco Ca/llis tRNA
The 'H I C ratio of the original bzl6Ade was 9.15.
Total Radioactivitv(Normalized to 30mm
'Sample 14 206 units tRNA "1I 14C
"It 1:C
Unhydrolyzed tRNA' 24,800 59,100 0.42Ethyl acetate extract? 20,700 3,830 5.4Ethyl acetate residue2 8,050 61,900 0.13
Sum of extract + residLue 28,750 65,730 0.44
Determined on 30 A260 units tRNA.2 Determined on 120 A42o units of tRNA.
40
60
80
100
70
80
90
Table 1I. Fractioniationi of Ethyl Acetate-soluible Naicleosicdes oniSephadex LH-20
Distribution of radioactivity in fractions from a SephadexLH-20 (35'c ethanol) fractionation of the ethyl acetate-solublenucleosides from tobacco callus tRNA prepared from tissue grownon bzl6Ade (3H 14C 9.15). The radioactivity measured is of 201'aliquots (equivalent to 600 A260 units tRNA) of the pooled frac-tions from the elutioni profile shown in Figure 1.
Fraction 1I 14C nH 1iC
lpmil ra tio0I 4 0
II 3 0HII 29 52 0.56IV 14 19 0.72V 20 1VI (bzl6A) 3626 389 9.3VII (bzl6Ade) 20 0
VIlI 12 0IX (ms2bzl'A) 15 0X (ms2bzl0A) 18 0xI 0 1XII 0 1
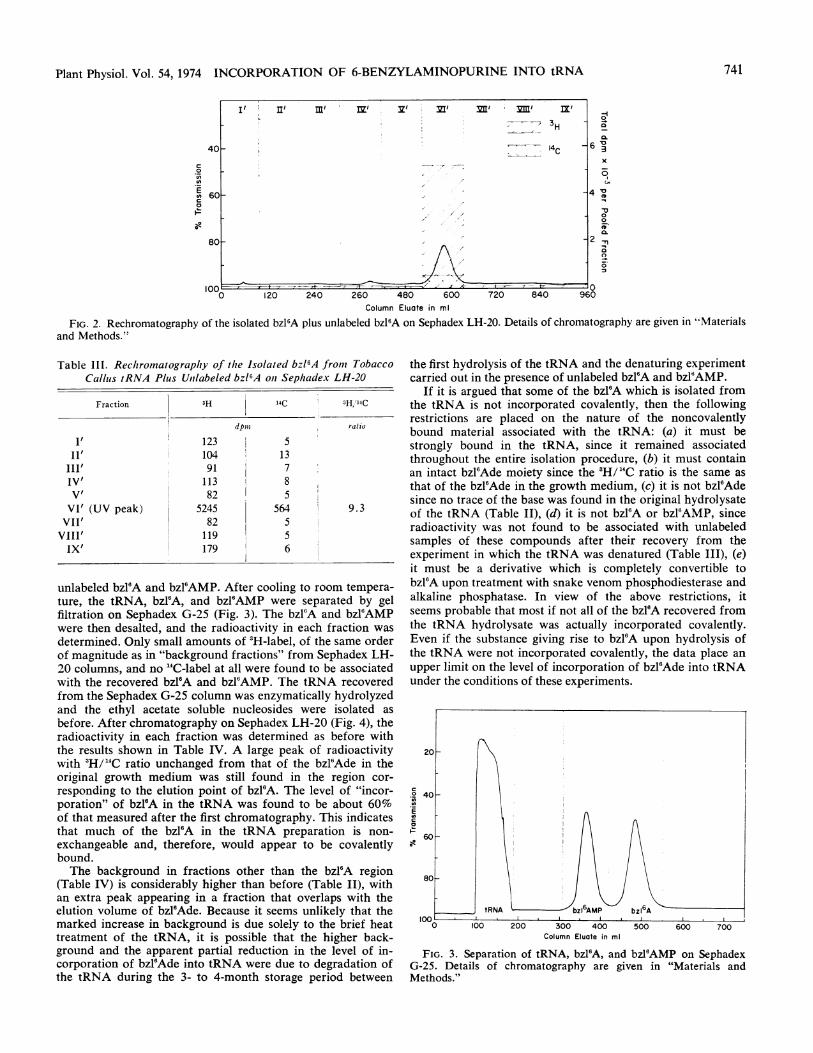
bzl'A was added to the remaining material from fraction VI,and the fraction was rechromatographed on Sephadex LH-20in distilled H20 (Fig. 2). Both the 3H and "4C peaks recoveredfrom the column coincided with the UV absorption peak fromthe unlabeled bzl'A marker and there was no change in the3H/'4C ratio (Table III).The noncovalent binding of labeled bzl6Ade to tRNA
throughout an isolation procedure has been reported recentlyfor soybean callus (10). However, in our experiments withtobacco callus, done with different extraction procedures, notrace of bzl'Ade binding was detected. Nevertheless thepossibility existed that the bzl6A which was isolated from thehydrolyzed tRNA had been bound in a noncovalent fashion. Inan attempt to clarify the situation, a sample of the tRNA wasslowly heated to a denaturing temperature in the presence of
Fraction No. II U E EZ at m MZI ixa x xi xiII
01
Standards
bz16A\ zI6Ade s2bzl6A60 120 180 240 300 360 420 480 540 600
Column Eluate in ml
FIG. I. Chromatography of ethyl acetate-soluble nucleosides on Sephadex LH-20. The ethyl acetate-soluble nucleosides from 2880 A260, unitsof tRNA were dissolved in 2.4 ml of 35%c ethanol and were applied to a Sephadex LH-20 column (30 g, 1.9 X 42 cm, 118 ml bed volume) equil-ibrated with the same solvent. Six-ml fractions were collected. The fractions were pooled as indicated and 20%c aliquots were taken for determi-nation of radioactivity (Table 11). The cytokinin standards (bzl'Ade, bzl6A, and ms2bzl6A) were dissolved in 2.4 ml of 35%- ethanol and wereapplied to the same LH-20 column used for fractionation of the ethyl acetate-soluble nucleosides.
740 WALKER ET AL.
c
.,0
.Ecp
Plant Physiol. Vol. 54, 1974 INCORPORATION OF 6-BENZYLAMINOPURINE INTO tRNA
in.Ecn
741
0
0.cx3x
0
10-o
_-n
Column Eluate in ml
FIG. 2. Rechromatography of the isolated bzl'A plus unlabeled bzl6A on Sephadex LH-20. Details of chromatography are given in 'Materialsand Methods."
Table 11I. Rechromatography of the Isolated bzl'A from TobaccoCalluts tRNA Plus Unllabeled bzl'A oni Sephadex LH-20
Fraction 3H 14C 3H/14C
dpwl ratio
I' 123 5II' 104 13
III' 91 7IV' 113 8V' 82 5VI' (UV peak) 5245 564 9.3VII' 82 5VIII' 119 5IX' 179 6
unlabeled bzl'A and bzl'AMP. After cooling to room tempera-ture, the tRNA, bzl'A, and bzl'AMP were separated by gelfiltration on Sephadex G-25 (Fig. 3). The bzl6A and bzl6AMPwere then desalted, and the radioactivity in each fraction was
determined. Only small amounts of 3H-label, of the same orderof magnitude as in "background fractions" from Sephadex LH-20 columns, and no "4C-label at all were found to be associatedwith the recovered bzl6A and bzl'AMP. The tRNA recoveredfrom the Sephadex G-25 column was enzymatically hydrolyzedand the ethyl acetate soluble nucleosides were isolated as
before. After chromatography on Sephadex LH-20 (Fig. 4), theradioactivity in each fraction was determined as before withthe results shown in Table IV. A large peak of radioactivitywith 3H/"C ratio unchanged from that of the bzl'Ade in theoriginal growth medium was still found in the region cor-
responding to the elution point of bzl6A. The level of "incor-poration" of bzl6A in the tRNA was found to be about 60%of that measured after the first chromatography. This indicatesthat much of the bzl6A in the tRNA preparation is non-
exchangeable and, therefore, would appear to be covalentlybound.The background in fractions other than the bzl'A region
(Table IV) is considerably higher than before (Table II), withan extra peak appearing in a fraction that overlaps with theelution volume of bzl6Ade. Because it seems unlikely that themarked increase in background is due solely to the brief heattreatment of the tRNA, it is possible that the higher back-ground and the apparent partial reduction in the level of in-corporation of bzl6Ade into tRNA were due to degradation ofthe tRNA during the 3- to 4-month storage period between
the first hydrolysis of the tRNA and the denaturing experimentcarried out in the presence of unlabeled bzl'A and bzl'AMP.
If it is argued that some of the bzl'A which is isolated fromthe tRNA is not incorporated covalently, then the followingrestrictions are placed on the nature of the noncovalentlybound material associated with the tRNA: (a) it must bestrongly bound in the tRNA, since it remained associatedthroughout the entire isolation procedure, (b) it must containan intact bzl'Ade moiety since the 3H/ 4C ratio is the same asthat of the bzl'Ade in the growth medium, (c) it is not bzl6Adesince no trace of the base was found in the original hydrolysateof the tRNA (Table II), (d) it is not bzl'A or bzl'AMP, sinceradioactivity was not found to be associated with unlabeledsamples of these compounds after their recovery from theexperiment in which the tRNA was denatured (Table III), (e)it must be a derivative which is completely convertible tobzl'A upon treatment with snake venom phosphodiesterase andalkaline phosphatase. In view of the above restrictions, itseems probable that most if not all of the bzl6A recovered fromthe tRNA hydrolysate was actually incorporated covalently.Even if the substance giving rise to bzl'A upon hydrolysis ofthe tRNA were not incorporated covalently, the data place anupper limit on the level of incorporation of bzl'Ade into tRNAunder the conditions of these experiments.
20-
.2 40-
tRNA 1bz6AMP bzl6A1000 100 200 300 400 500 600 700
Column Eluate in ml
FIG. 3. Separation of tRNA, bzl'A, and bzlAMP on SephadexG-25. Details of chromatography are given in "Materials andMethods."

WALKER ET AL.
Fraction No.iE~~~~~~~~3l JI/ X ' XI ' Is
40- bzl16AIo
.60I
60-
80 F
10060 120 1 80 240 300 360 420 480 540 600
Column Eluote in mlFIG. 4. Isolation of bzl'A from tRNA purified by heat denaturation and gel filtration. The ethyl acetate-soluble nucleosides from 475 A260units of tRNA were dissolved in 2 ml of 35'c ethanol and applied to a Sephadex LH-20 column (30 g, 1.9 X 42 cm, 118 ml bed volume) equili-brated with the same solvent. Six-ml fractions were collected and pooled as indicated for determination of radioactivity (Table IV). The elution
point of bzl6A was determined by applying unlabeled bzlIA to the same column.
Table IV. Sephadex LH-20 Fractioioaitioln of Etl/ lv ANiucleosides from tRNA after the Deniatitriiug EL
t11
dpn,t
I"
III"IVl"IV"
Vi"
VII" (bzl'A)
Ix"
Xl"
XII"Recovered bzleA
Recovered bzl'AMP
130
125
293
289
69
55
1601
134
401
52
39
49
43
118
1240
211
7
170
12
46
1
0
2
0
0
DISCUSSION AND CONCLUSION"
The equivalence of the -H/"C ratio of the b,from hydrolysates of tobacco tissue tRNA prep;that of the labeled bzl¢Ade fed to the tissue demoany direct incorporation of cytokinins into tRNA i
the intact base. This result is consistent with the ezof Fox (13, 14) and conclusively rules out any mevolving transfer of the N5-sidechains of free c
adenosine residues in RNA. The extent of incorrfound to be extremely low, about 1 bzl'A moletRNA molecules. These results are inconsistent wings of Elliott and Murray (10), who reported thawas incorporated as an intact molecule, placing arof about 1 bzleA molecule/16,400 tRNA molecula transbenzylation reaction involving transfer ofsidechain from a free cytokinin to an adenosinitRNA may take place. The 3H/"C ratios obtaineperiments rule out the transbenzylation reaction.ment of the upper limit for cytokinin incorporapaper is felt to be more reliable than that ofMurray (10), since the experiment was performed1000 times larger scale, and the labeled bzl6Adestudy did not contain the potentially exchangeablein the 8-position of the purine ring. Although n4
were added during the isolation procedure as in thby Elliott and Murray (10), no bzl6Ade was found t
cetate-soliuble in the tRNA hydrolysate by means of Sephadex LH-20 column;periment chromatography, which clearly resolves bzl'A and bzl¢Ade.
Neither bzl'A nor bzl'AMP coud be shown to be associatedFitII4C with the tRNA, and the possibility of the bzl'A arising from
the noncovalent association of a bzl'Ade derivative with atRNA seems quite remote as discussed above.
Possibilities for the incorporation of a bzl'Ade derivativeinto a tRNA include transcriptional errors, additions to the
.24 ends of completed RNA, or incorporation by some type of1~ excision and repair process. Increasing the concentration of
bzl'Ade in the medium 10 fold, from 0.04 ~M (which is nearlyoptimal for growth) to 0.44 /m, did not significantly increase9*4 the amount of incorporation into tRNA, although it did in-crease the randomization of the label. It is concluded that,
8.8 of the bzl6Ade administered exogenously to cytokinin depend-ent tobacco callus cultures for about a 3-week growth period,a small amount is incorporated intact into bzl6A recoverablefrom tRNA hydrolysates, but that the level of bzl5A recoveredfrom the tRNA preparations is far lower than that of endoge-nous cytokinin-active ribonucleosides (4). Hence, any rolethat the incorporated bzl6A in tRNA might play in growth
s regulation would appear to be other than as a replacement forendogenous cytokinins in their known functions as constituents
zl6A isolated of tRNA (24, 29, 30). Such a role may not be readily detect-arations and able by the present methodology. Research on the function of)nstrates that cytokinin molecules as allosteric effectors or as derepressorsmust involve as well as a continuing detailed study of their metabolic fatearlier studies might be a more effective approach to the mode of actionechanism in- of cytokinins in the regulation of growth.ytokinins tooration waso Acknouwledgmesnts-We thank Prof. R. F. Nystrom, for his valuable advicec /10 000 and assistance aind Dr. John C. Coll, University of Illinois, for assisting lis inthe synthesis of benzvlamine-benzene-3H(G). We thank Mrs. Anna Hilden,
'ith the find- University of 'Wisconsin, for assistance in the cultivation of callus and in cy-to-t no bzleAde kinin bioassavs.upper limit[es,pand that LITERATURE CITEDles, and thatthe benzyl 1. ARMSTRONG, D. J., W. J. BURROWS, P. K. EVANS, AND F. SKOOG. 1969.
e residue in Isolation of Cytokinins from tRNA. Biochem. Biophys. Res. Commun. 37:451-456.d in our ex- 2. BARTZ, J. K. AND D. SOLL. 1972. NC- (A2-Isopentenyl) adenosine: biosynthesisThe assign- in vitro in transfer RN'A by an enzyme purified from Escherichia colt.ition in this Biochemie 54: 31-39.
Elliott and 3. BEZENIER-SYBRAN-DY, J. M. AN-D N. VELDSTRA. 1971. The metabolism of 6-benzylaminopurine in Lemna minor. Physiol. Plant. 25: 1-7.I on about a 4. BURROWS, W. J., F. SKOOG. AND N. J. LEONARD. 1971. Isolation and identifi-used in this cation of cytokinins located in the transfer ribonucleic acid of tobacco
(11) tritium callus grown in the presence of 6-benzylaminopurine. Biochemistry 10:ocytokinins ~ 2189-2194.N6cytokinmns 5. CHEN, C. AND R. H. HLL... 1969. Biosynthesis of N6-(A2-isopentenfl adleno-
iat described sine in the transfer rihonucleic acid of cultured tobacco pith tissule. Phyrto-to be present cheemistry 8: 1687-169.5.
742 Plant Physiol. Vol. 54, 1974
14C

Plant Physiol. Vol. 54, 1974 INCORPORATION OF 6-BENZYLAMINOPURINE INTO tRNA
6. DALY, J. W. AND B. E. CHRISTENSEN. 1956. The preparation of certain 6-substituted and 6,9-disubstituted purines. J. Org. Chem. 21: 177-179.
7. DELEUZE, G. G., J. D. MCCHESNEY, AND J. E. Fox. 1972. Identification ofa stable cytokinin metabolite. Biochem. Biophys. Res. Commun. 48: 1426-1432.
8. DysoN, W. H. 1969. Metabolism of urea and purine cytokinins. Ph.D. thesis.University of Kansas, Lawrence.
9. DYSON, W. H., J. E. Fox, AND J. D. MCCHESNEY. 1972. Short term metabo-lism of urea and purine cytokinins. Plant Physiol. 49: 506-513.
10. ELLIOTT, D. C. AN-D A. W. MURRAY. 1972. A quantitative limit for cytokininincorporation into transfer ribonucleic acid by soya-bean callus tissue. Bio-chem. J. 130: 1157-1160.
11. EVANS, E. A., H. C. SHEPPARD, AND J. C. TuRNER. 1970. Validity of tritiumtracers. Stability of tritium atoms in purines, pyrimidines, nucleosides, andnucleotides. J. Label. Compounds 6: 76-87.
12. FITTLER, F., L. K. KLINE, AN-D R. H. HALL. 1968. N6-(A2-Isopentenyl)adeno-sine: biosynthesis in vitro by an enzyme extract from yeast and rat liver.Biochem. Biophys. Res. Commun. 31: 571-576.
13. Fox, J. E. 1966. Incorporation of a kinin, N,6-benzyladenine into solubleRNA. Plant Physiol. 41: 75-82.
14. Fox, J. E. AND C. CHEN. 1967. Characterization of labeled ribonucleic acidfrom tissue grown on "4C-containing cytokinins. J. Biol. Chem. 242: 4490-4494.
15. Fox, J. E., C. K. SOOD, B. BUCKWALTER, AND J. D. MCCHESNEY. 1971. Themetabolism and biological activity of a 9-substituted cytokinin. PlantPhysiol. 47: 275-281.
16. HALL, R. H. 1968. Cytokinins in the transfer-RNA: their significance to thestructure of t-RNA. In: F. Wightman and G. Setterfield, eds., Biochem-istry and Physiology of Plant Growth Substances, Runge Press, Ottawa.pp. 47-56.
17. KENDE, H. 1971. The cytokinins. Int. Rev. Cytol. 31: 301-338.18. KENDE, H. AND J. E. TAVARES. 1968. On the significance of cytokinin in-
corporation into RNA. Plant Physiol. 43: 1244-1248.19. KEY, J. L. 1969. Hormones and nucleic acid metabolism. Annu. Rev. Plant
Physiol. 20: 449-474.20. KLINE, L., K. F. FITTLER, AN-D R. H. HALL. 1969. N6- (A2-Isopentenyl) adeno-
sine. Biosynthesis in transfer ribonucleic acid in vitro. Biochemistrv 8:4361-4375.
21. LIN\SMIAIER, E. M. AND F. SKOOG. 1965. Organic growth factor requirementsof tobacco tissue cultures. Physiol. Plant. 18: 100-127.
22. 'MCCALLA, D. R., D. J. MORRE, AND D. OSBORNE. 1962. The metabolism of a
kinin, benzyladenine. Biochim. Biophys. Acta 55: 5222-5228.23. MCCHESN-EY, J. D. 1970. Stimulation of in vitro growth of plant tissues by
DL-mev-alonic acid. Can. J. Bot. 48: 2357-2359.24. NISHINIURA, S. 1973. 'Minor components in transfer RNA: Their charac-
terization, location, and function. Prog. Nucleic Acid Res. Mol. Biol. 12:49-85.
25. PETERKOFSKY, A. 1968. The incorporation of mevalonic acid into the N6-(A2-isopentenyl) adenosine of transfer ribonucleic acid in LactobacillUusacidophilus. Biochemistry 7: 472-482.
26. RICHMOND, A., A. BACK, A-ND B. SACHS. 1970. A study of the hypotheticalrole of cytokinins in completion of tRNA. Planta 90: 57-65.
27. ROSENBAUMI, N. AND WM. L. GEFTER. 1972. A2-Isopentenylpyrophosplhate:transfer ribonucleic acid-A2-isopentenyltransferase from Escherichia colt.J. Biol. Chem. 247: 5675-5680.
28. SELIVANKINA, S. V., V. A. KUROEDOV, AND 0. N. KULAEVA. 1972. Dynamicsof cytokinin action on label incorporation into tRNA of cut barley leaves.Fiziol. Rast. 19: 508-516.
29. SKOOG, F. AN-D D. J. ARMSTRONG. 1970. Cytokinins. Annu. Rev. Plant Physiol.21: 359-384.
30. SULL, D. 1971. Enzymatic modification of transfer RNA. Science 173: 293-299.
31. SRIVASTAVA, B. I. S. 1967. Effect of kinetin on biochemical changes in excisedbarley leaves and in tobacco pith tissue culture. Ann. N.Y. Acad. Sci. 144:260-278.
32. VAN ONCKELEN, H. A. AN'D R. VERBEEE. 1971. Incorporation of a cytokininN'- (furfurylamino)purine-8-14C in the nucleic acid fraction of barley.Bull. Soc. Roy. Bot. Belg. 104: 161-168.
33. WHITEHEAD, C. W. A-ND J. J. TRAVERSO. 1960. Exchange amination. Alkyland arylamino-pyrimidines and purines. J. Amer. Chem. Soc. 82: 3971-3974.
34. ZACHAU, H. G. 1971. Transfer ribonucleic acids. Angew. Chem. Intl. Ed.Engl. 8: 711-727.
743
Related Documents











