Biochem. J. (2013) 450, 511–521 (Printed in Great Britain) doi:10.1042/BJ20121135 511 The membrane adaptor LAT is proteolytically cleaved following Fas engagement in a tyrosine phosphorylation-dependent fashion Antonio GARC ´ IA-BLESA*†, Mikolaj KLOSSOWICZ‡, Carmen L ´ OPEZ-OSUNA*†, Mario MART ´ INEZ-FLORENSA§, Bernard MALISSEN, Francisco J. GARC ´ IA-C ´ OZAR*†, Arkadiusz MIAZEK‡ and Enrique AGUADO*† 1 *Core Research Facility for Health Sciences, Department of Biomedicine, Biotechnology and Public Health (Immunology), University of C´ adiz, c/ Dr Mara˜ non, n ◦ 3, C´ adiz 1102, Spain, †Puerto Real University Hospital Research Unit, Ctra N-IV Km 665, 11510 Puerto Real (C´ adiz), Spain, ‡Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, R. Weigla Str. 12, 53–114 Wroclaw, Poland, §Institut d’Investigacions Biom` ediques, Centre Esther Koplowitz, c/ Rossell´ o, 149–153, 08036 Barcelona, Spain, and Centre d’Immunologie de Marseille-Luminy, Parc Scientifique et Technologique de Luminy, Case 906, 13288 Marseille cedex 09, France Engagement of the TCR (T-cell receptor) induces tyrosine phosphorylation of the LAT (linker for the activation of T-cells) adaptor, and thereby it recruits several cytosolic mediators for downstream signalling pathways. The Fas protein is essential for T-lymphocyte apoptosis, and following Fas engagement, many proteins are proteolytically cleaved, including several molecules that are important for the transduction of TCR intracellular signals. In the present study, we demonstrate that the adaptor LAT is also subject to a proteolytic cleavage in mature T-lymphocytes and thymocytes in response to Fas engagement, and also on TCR stimulation, and we identify three aspartic acid residues at which LAT is cleaved. Interestingly, these aspartic acid residues are located in proximity to several functionally important tyrosine residues of LAT, raising the possibility that their phosphorylation could modulate LAT cleavage. Consistent with that hypothesis, we show that induction of phosphorylation by pervanadate or H 2 O 2 in Jurkat cells and thymocytes inhibits Fas-mediated cleavage of LAT. Moreover, we show that LAT proteolysis is also enhanced during anergy induction of primary human T-cells, suggesting that LAT cleavage may act as a regulator of TCR-mediated activation of T-cells and not only as a transducer of cell death promoting stimuli. Key words: caspase, CD3, granzyme B, membrane adaptors, signalling, T-cell receptor (TCR). INTRODUCTION T-lymphocytes detect foreign peptides bound to MHC via the TCR (T-cell receptor) and convey this information to the inside of the cell through its associated CD (cluster of differentiation) 3 subunits. TCR engagement triggers phosphorylation of ITAM (immunoreceptor tyrosine-based activation motif) in the CD3 subunits, which on phosphorylation recruit ZAP-70 [ζ -chain (TCR)-associated protein kinase of 70 kDa] tyrosine kinase to the membrane, leading to the phosphorylation of several intracellular substrates, including the LAT (linker for the activation of T- cells) adaptor [1–3]. LAT is an integral transmembrane adaptor protein of 36–38 kDa expressed in peripheral T-lymphocytes, thymocytes, NK (natural killer) cells, mast cells, platelets and pre-B-cells [4]. Many of the functions of LAT are carried out by its four conserved C-terminal tyrosine residues, which on phosphorylation constitute docking sites for a number of cytosolic signalling effectors [4–7]. These tyrosine residues are essential for the transduction of intracellular signals generated by the TCR–CD3 complex in developing and mature T-cells [8–11]. Aside from tyrosine resides, other amino acids in LAT can be phosphorylated [12,13]. Indeed, we recently showed that several serine residues of LAT are essential for TCR- mediated activation of both PLC (phospholipase C)-γ 1 and MAPK (mitogen-activated protein kinase) [14]. Once recruited to the LAT signalosome, PLCγ 1 and other signalling effectors are then activated, leading to Ca 2 + mobilization and activation of multiple pathways, which in turn activate specific nuclear factors leading to anergy, or alternatively, to survival, activation, proliferation and differentiation of T-lymphocytes into effector cells [15–17]. In the course of immune responses, antigen-specific T-cells are activated and first undergo a phase of clonal expansion, followed by a contraction phase that restores normal T-cell numbers and requires apoptosis [18]. Apoptosis is a form of programmed cell death that is essential for many relevant biological processes [19]. There exist two major mechanisms of apoptosis that operate through the intrinsic and the extrinsic pathways. The intrinsic pathway is usually triggered in response to severe cell stress events such as DNA damage or withdrawal of survival factors and involves release of pro-apoptotic proteins from the mitochondria to the cytosol and activation of caspase enzymes [20]. In contrast, the extrinsic pathway is typically initiated upon engagement of members of the TNFR (tumour necrosis factor receptor) family. For instance, binding of Fas, also known as CD95 or APO-1 (apoptosis antigen 1), to its ligands leads to the recruitment and activation of caspase-8, which in turn activates effector caspases [21]. Although Fas engagement is essential for T-lymphocyte apoptosis and immune system homoeostasis [18], it can also transduce activation signals leading to T-cell proliferation on TCR engagement [22]. Paradoxically, it had been demonstrated previously that the simultaneous engagement of TCR, CD28 and Fas in naive human T-lymphocytes led to decreased activation Abbreviations used: CD, cluster of differentiation; ECL, enhanced chemiluminescence; ERK, extracellular-signal-regulated kinase; -fmk, fluoromethylketone; FBS, fetal bovine serum; GFP, green fluorescent protein; Grb2, growth-factor-receptor-bound protein 2; IL, interleukin; IRES, internal ribosome entry site; LAT, linker for the activation of T-cells; mAb, monoclonal antibody; PARP, poly(ADP-ribose) polymerase; PBMC, peripheral blood mononuclear cell; PHA, phytohaemagglutinin; PKC, protein kinase C; PLC, phospholipase C; SIN, self-inactivating; SLP-76, Src homology 2 domain- containing leucocyte protein of 76 kDa; TCR, T-cell receptor; ZAP-70, ζ -chain (T-cell receptor)-associated protein kinase of 70 kDa; z-, benzyloxycarbonyl-3. 1 To whom correspondence should be addressed (email [email protected]). c The Authors Journal compilation c 2013 Biochemical Society Biochemical Journal www.biochemj.org

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochem. J. (2013) 450, 511–521 (Printed in Great Britain) doi:10.1042/BJ20121135 511
The membrane adaptor LAT is proteolytically cleaved following Fasengagement in a tyrosine phosphorylation-dependent fashionAntonio GARCIA-BLESA*†, Mikolaj KLOSSOWICZ‡, Carmen LOPEZ-OSUNA*†, Mario MARTINEZ-FLORENSA§,Bernard MALISSEN‖, Francisco J. GARCIA-COZAR*†, Arkadiusz MIAZEK‡ and Enrique AGUADO*†1
*Core Research Facility for Health Sciences, Department of Biomedicine, Biotechnology and Public Health (Immunology), University of Cadiz, c/ Dr Maranon, n◦ 3, Cadiz 1102, Spain,†Puerto Real University Hospital Research Unit, Ctra N-IV Km 665, 11510 Puerto Real (Cadiz), Spain, ‡Institute of Immunology and Experimental Therapy, Polish Academy of Sciences,R. Weigla Str. 12, 53–114 Wrocław, Poland, §Institut d’Investigacions Biomediques, Centre Esther Koplowitz, c/ Rossello, 149–153, 08036 Barcelona, Spain, and‖Centre d’Immunologie de Marseille-Luminy, Parc Scientifique et Technologique de Luminy, Case 906, 13288 Marseille cedex 09, France
Engagement of the TCR (T-cell receptor) induces tyrosinephosphorylation of the LAT (linker for the activation of T-cells)adaptor, and thereby it recruits several cytosolic mediators fordownstream signalling pathways. The Fas protein is essential forT-lymphocyte apoptosis, and following Fas engagement, manyproteins are proteolytically cleaved, including several moleculesthat are important for the transduction of TCR intracellularsignals. In the present study, we demonstrate that the adaptor LATis also subject to a proteolytic cleavage in mature T-lymphocytesand thymocytes in response to Fas engagement, and also on TCRstimulation, and we identify three aspartic acid residues at whichLAT is cleaved. Interestingly, these aspartic acid residues arelocated in proximity to several functionally important tyrosine
residues of LAT, raising the possibility that their phosphorylationcould modulate LAT cleavage. Consistent with that hypothesis, weshow that induction of phosphorylation by pervanadate or H2O2
in Jurkat cells and thymocytes inhibits Fas-mediated cleavage ofLAT. Moreover, we show that LAT proteolysis is also enhancedduring anergy induction of primary human T-cells, suggesting thatLAT cleavage may act as a regulator of TCR-mediated activationof T-cells and not only as a transducer of cell death promotingstimuli.
Key words: caspase, CD3, granzyme B, membrane adaptors,signalling, T-cell receptor (TCR).
INTRODUCTION
T-lymphocytes detect foreign peptides bound to MHC via theTCR (T-cell receptor) and convey this information to the insideof the cell through its associated CD (cluster of differentiation)3 subunits. TCR engagement triggers phosphorylation of ITAM(immunoreceptor tyrosine-based activation motif) in the CD3subunits, which on phosphorylation recruit ZAP-70 [ζ -chain(TCR)-associated protein kinase of 70 kDa] tyrosine kinase to themembrane, leading to the phosphorylation of several intracellularsubstrates, including the LAT (linker for the activation of T-cells) adaptor [1–3]. LAT is an integral transmembrane adaptorprotein of 36–38 kDa expressed in peripheral T-lymphocytes,thymocytes, NK (natural killer) cells, mast cells, platelets andpre-B-cells [4]. Many of the functions of LAT are carriedout by its four conserved C-terminal tyrosine residues, whichon phosphorylation constitute docking sites for a number ofcytosolic signalling effectors [4–7]. These tyrosine residues areessential for the transduction of intracellular signals generatedby the TCR–CD3 complex in developing and mature T-cells[8–11]. Aside from tyrosine resides, other amino acids in LATcan be phosphorylated [12,13]. Indeed, we recently showedthat several serine residues of LAT are essential for TCR-mediated activation of both PLC (phospholipase C)-γ 1 andMAPK (mitogen-activated protein kinase) [14]. Once recruitedto the LAT signalosome, PLCγ 1 and other signalling effectorsare then activated, leading to Ca2 + mobilization and activation
of multiple pathways, which in turn activate specific nuclearfactors leading to anergy, or alternatively, to survival, activation,proliferation and differentiation of T-lymphocytes into effectorcells [15–17].
In the course of immune responses, antigen-specific T-cells areactivated and first undergo a phase of clonal expansion, followedby a contraction phase that restores normal T-cell numbers andrequires apoptosis [18]. Apoptosis is a form of programmed celldeath that is essential for many relevant biological processes [19].There exist two major mechanisms of apoptosis that operatethrough the intrinsic and the extrinsic pathways. The intrinsicpathway is usually triggered in response to severe cell stressevents such as DNA damage or withdrawal of survival factors andinvolves release of pro-apoptotic proteins from the mitochondriato the cytosol and activation of caspase enzymes [20]. In contrast,the extrinsic pathway is typically initiated upon engagement ofmembers of the TNFR (tumour necrosis factor receptor) family.For instance, binding of Fas, also known as CD95 or APO-1(apoptosis antigen 1), to its ligands leads to the recruitment andactivation of caspase-8, which in turn activates effector caspases[21].
Although Fas engagement is essential for T-lymphocyteapoptosis and immune system homoeostasis [18], it can alsotransduce activation signals leading to T-cell proliferation onTCR engagement [22]. Paradoxically, it had been demonstratedpreviously that the simultaneous engagement of TCR, CD28 andFas in naive human T-lymphocytes led to decreased activation
Abbreviations used: CD, cluster of differentiation; ECL, enhanced chemiluminescence; ERK, extracellular-signal-regulated kinase; -fmk,fluoromethylketone; FBS, fetal bovine serum; GFP, green fluorescent protein; Grb2, growth-factor-receptor-bound protein 2; IL, interleukin; IRES, internalribosome entry site; LAT, linker for the activation of T-cells; mAb, monoclonal antibody; PARP, poly(ADP-ribose) polymerase; PBMC, peripheral bloodmononuclear cell; PHA, phytohaemagglutinin; PKC, protein kinase C; PLC, phospholipase C; SIN, self-inactivating; SLP-76, Src homology 2 domain-containing leucocyte protein of 76 kDa; TCR, T-cell receptor; ZAP-70, ζ-chain (T-cell receptor)-associated protein kinase of 70 kDa; z-, benzyloxycarbonyl-3.
1 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2013 Biochemical Society
Bio
chem
ical
Jo
urn
al
ww
w.b
ioch
emj.o
rg

512 A. Garcıa-Blesa and others
and proliferation [23]. In these cells, Fas triggering is able todisrupt the TCR signalling pathway by inhibiting recruitment ofZAP-70, PLC-γ 1 and PKC (protein kinase C)-θ . Consideringthat Fas engagement results in cleavage of Gads (GRB2-related adaptor protein 2), LAT and SLP-76 (Src homology2 domain-containing leucocyte protein of 76 kDa) adaptors,the present study analysed whether the negative impact ofFas engagement on TCR signalling resulted in Fas-mediatedcleavage of LAT. LAT seems to be cleaved by a caspase-me-diated process in response to Fas engagement, CD3 cross-linkingand by treatment of cells with the PKC inhibitor staurosporine.LAT cleavage takes place at three aspartic acid residues locatedin close proximity to several functionally important tyrosineresidues. Interestingly, phosphorylation of these tyrosine residuesblocks Fas-mediated cleavage, whereas Fas triggering preventsearly intracellular signals associated with the TCR–CD3 complex,suggesting a previously unrecognized negative regulatory loopthat involves cleavage of LAT which is dependent on the state ofLAT tyrosine phosphorylation.
EXPERIMENTAL
Antibodies, primers and plasmids
Stimulations were performed with the anti-human CD3 OKT3 oranti-mouse CD3ε clone 145–2C11 mAb (monoclonal antibody)(both from eBioscience). The anti-Fas (IgM), anti-phospho-LAT-Tyr226 and rabbit polyclonal anti-LAT 06-807 antibodieswere obtained from Millipore; anti-PLC-γ 1 mAb, anti-ERK(extracellular-signal-regulated kinase) and rabbit polyclonal anti-PARP [poly(ADP-ribose) polymerase]-1 H-250 were from SantaCruz Biotechnology; the antibodies binding phospho-ERK,phospho-PLC-γ 1, phospho-LAT-Tyr171 and phospho-LAT-Tyr171
were from Cell Signaling Technology; anti-phosphotyrosineRC20 was from Transduction Laboratories; and anti-Grb2(growth-factor-receptor-bound protein 2) and anti-phospho-LAT-Tyr132 were obtained from Abcam. Monoclonal mouse anti-IgG1 anti-β-actin antibody peroxidase conjugate was fromSigma–Aldrich; and monoclonal anti-mouse-CD69 antibody PE(phycoerythrin) was from BD Bioscience.
Mutation of LAT was performed with GeneTailorTM Site-Directed Mutagenesis System (Invitrogen) with specific pairsof primers. Mutagenesis was performed using a pENTR11plasmid (Invitrogen) containing the cDNA for wild-type LATas a template, followed by a linker containing a 6×His-tag 5′
and an IRES (internal ribosome entry site) sequence [14]. DNAsencoding wild-type or mutant LAT–6×His-IRES were subclonedin-frame with GFP (green fluorescent protein) in the SIN (self-inactivating) lentiviral transfer plasmid pHR’SIN-cPPT-Blast bymeans of site-specific recombination (Gateway LR Clonase,Invitrogen). The coding sequences of all LAT constructs wereverified by sequencing. The lentiviral transfer plasmid for theexpression of the wild-type LAT–GFP fusion protein has beendescribed recently [14].
Reagents
Ionomycin and PHA (phytohaemagglutinin) were purchasedfrom Sigma–Aldrich. The peptide caspase inhibitorsz-VAD-fmk (where z- is benzyloxycarbonyl- and -fmk is-fluoromethylketone), z-VDVAD-fmk (caspase 2), Ac(N-acetyl)-DEVD-cmk (chloromethylketone) (caspase 3-III),z-DQMD-fmk (caspase 3-V), z-IETD-fmk (caspase 8/granzymeB) and z-LEHD-fmk were purchased from Calbiochem. IL-2
(interleukin 2) was obtained through the AIDS Research andReference Reagent Program, Division of AIDS; NIAID; NIH.
Cell lines and PBMC (peripheral blood mononuclear cell) isolationand culture
Jurkat cells, the LAT-deficient JCaM2 cell line (generouslyprovided by Dr Arthur Weiss, UCSF, San Francisco, CA, U.S.A.),and subsequent stable transfectant lines were maintained asdescribed in [24]. VL3–3M2 cells were maintained as describedpreviously [25]. Human peripheral blood samples were obtainedfrom healthy donors with informed consent of the subjects andfollowing ethical approval. PBMCs were obtained from buffycoats of normal individuals by Ficoll-Hypaque density-gradientcentrifugation. PBMCs (24×106) were then cultured for 1 weekin the presence of 1 μg/ml of PHA and 40 international units/mlof recombinant IL-2 before anergy induction.
Thymocytes isolation and culture
For isolation of mouse thymocytes, thymi from C57Bl/6 micewere dissected and ground through a 70-μm cell strainer andwashed twice in PBS. All procedures using mice were approvedby the First Local Ethical Commission in Wrocław, Poland, underpermission number 06/2009.
In vitro induction of anergy in CD4 T-cells with ionomycin
CD4 T-cells were purified from cultured PBMCs with DynabeadsUntouchedTM Human CD4 T Cells kit (Invitrogen), and then withionomycin (Sigma–Aldrich) at a final concentration of 1 μM for16 h in DMEM (Dulbecco’s modified Eagle’s medium) with 10 %FBS (fetal bovine serum) at a concentration of 2×106 cells/ml ina volume of 2 ml. Anergy induction in these cells was evaluatedby the proliferative capacity in response to plate-bound anti-CD3and anti-CD28 antibodies, measured with the Click-iT EdU FlowCytometry Assay Kit (Invitrogen) following instructions from theproducer.
Lentiviral production and transfection of JCaM2 cells
HEK (human embryonic kidney)-293-FT packaging cells(Invitrogen) were plated in 12-well plates at a density of 3×105
cells per well the day before transfection. Cells were washed withOptiMEM (Invitrogen) before transfection and transfectedwith the corresponding expression vectors, together withgag–pol and VSV (vesicular stomatitis virus) capsid,using LipofectamineTM 2000 (Invitrogen), according to themanufacturer’s instructions. At 48 and 72 h, transfectionefficiency was evaluated by FACS analysis using a CyanADP-MLE flow cytometer (DakoCytomation, Denmark). Lentiviralsupernatants were collected 48 and 72 h after transfection.JCaM2 cells were plated at a density of 105 cells per well (5×105
cells/ml) in 6-well plates. Lentiviral supernatant was added,and cells were cultured for 48 h in a 37 ◦C and 5% CO2 incubator.After selection with 10 μg/ml blasticidin, expression of LAT andGFP was analysed by FACS.
Anti-CD3 and anti-Fas stimulation and preparation of cell lysates
JCaM2 transfected cells were starved overnight in RPMI 1640medium without FCS for five times, then stimulated with OKT3anti-CD3 mAb (eBioscience) in RPMI 1640 at 37 ◦C. Cells(2.5×107 cells/ml) were then lysed in 2× Laemmli buffer,followed by incubation at 95 ◦C for 5 min and sonication. For
c© The Authors Journal compilation c© 2013 Biochemical Society

Tyrosine phosphorylation of LAT 513
anti-Fas stimulation, cells were incubated with the indicatedconcentrations of anti-Fas mAb at 1×106 cells/ml in RPMI 1640supplemented with 10% FBS, and then pelleted and lysed asdescribed above. When needed, cells were treated with freshlyprepared 1% pervanadate (10 mM Na3VO4 containing 1% H2O2)for 5 min at 37 ◦C.
SDS/PAGE and Western blotting
For LAT detection by Western blot analysis, whole cell lysatesfrom 106 transfectant cells were obtained by resuspending cellsin Laemmli lysis buffer and separated in SDS/PAGE gels andtransferred on to PVDF membranes. The membranes wereincubated with the indicated primary antibodies followed bythe appropriate secondary antibody conjugated to horseradishperoxidase. Reactive proteins were visualized using theECL (enhanced chemiluminescence) system and exposition toHyperfilm ECL (GE Healthcare) or directly acquired withVersaDoc 5000 Imaging System (Bio-Rad Laboratories). For re-probing, PVDF membranes were incubated for 10 min at roomtemperature (25 ◦C) with 0.1 M NaOH, followed by thoroughwashes with blocking buffer.
Measurement of intracellular Ca2 + mobilization
Measurement of intracellular free Ca2 + was carried outusing Indo-1 AM (acetoxymethyl) (2 μM; Molecular Probes,Invitrogen) as described recently [14]. Calcium measurementswere performed using a Synergy MX Multi-Mode Reader(Biotek) at 37 ◦C. The cells were excited by light at a wavelengthof 340 nm, and the fluorescence emitted at 405 and 485 nmwas collected alternately per second. Calcium mobilization wasevaluated by the ratio of 405/485 nm fluorescence signal.
Statistical analysis
Data are expressed as means +− S.E.M. Statistical differences inthe results for the different parameters between groups wereevaluated by the two-tailed Student’s t test. A P value of <0.05was considered significant.
RESULTS
The adaptor LAT is proteolytically cleaved on Fas engagement
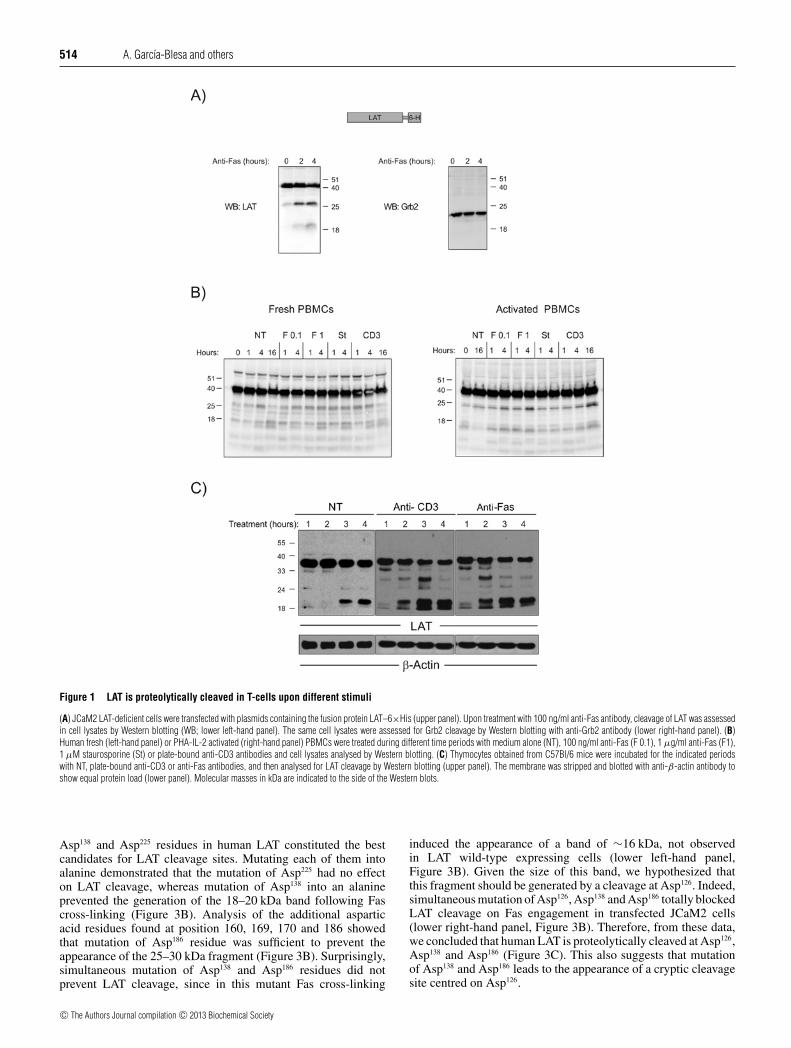
LAT is proteolytically cleaved in T-cells following Fas cross-linking [24,25]. In order to analyse the cleavage of LAT, wetransfected LAT-deficient JCaM2 cells with a lentiviral vectorcoding for wild-type LAT in frame with a 6×His tag andcontaining an IRES-GFP cassette [14] (upper panel, Figure 1A).Treatment of transfected cells with anti-Fas mAb for 2 h inducedthe appearance of a LAT cleavage product of approximately 25–30 kDa. A 4-h treatment produced the appearance of two bandswith electrophoretic mobilities of approximately 25–30 kDa and18–20 kDa respectively (lower left-hand panel, Figure 1A). Inorder to verify that LAT cleavage was not the result of a non-specific increase in proteolytic activity, we analysed the same celllysates for the presence of cleaved Grb2 molecules. Consistentwith previous data [26,27], Grb2 was not cleaved in response toFas engagement (lower right-hand panel, Figure 1A).
We then studied whether LAT was also cleaved in peripheralT-lymphocytes following Fas engagement. Freshly isolated PHAplus IL-2 activated PBMCs were left untreated or treated withanti-Fas mAb, staurosporine or plate-bound anti-CD3 antibodies.
As shown in Figure 1(B), resting and activated PBMCs also show amodest cleavage of LAT following anti-Fas mAb or staurosporinetreatments. Interestingly, CD3 cross-linking for 16 h also inducescleavage of LAT in activated T-cells.
As the 06-807 polyclonal antibody recognizes both human andmouse LAT, we also analysed LAT cleavage in thymocytes fromC57Bl/6 mice. LAT is cleaved in thymocytes upon CD3 and Fascross-linking (upper panel, Figure 1C). Interestingly, untreatedthymocytes also show some cleavage of LAT after 3 h of culture,although the intensity of this is weaker than the one observed inanti-CD3- or anti-CD95-treated cells (see the equal protein loadin the β-actin Western blot in lower panel of Figure 1C).
LAT is cleaved by a caspase or granzyme-B-mediated process
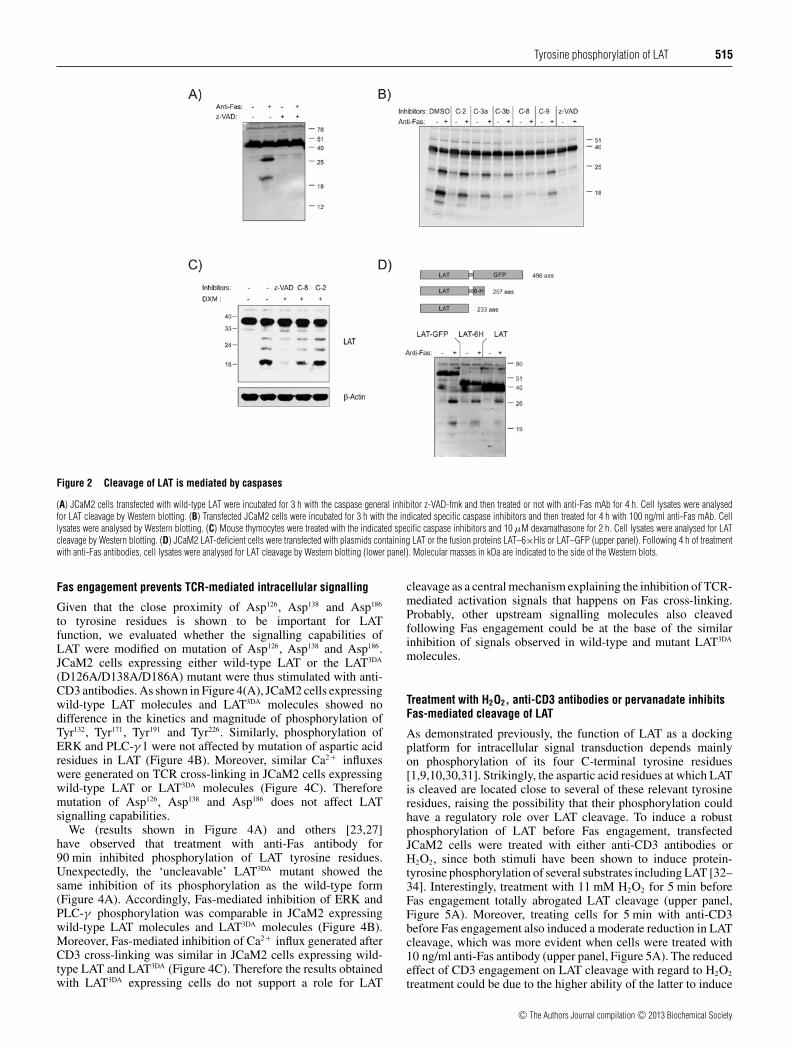
Given that the LAT adaptor is cleaved on Fas cross-linking,we wanted to analyse the involvement of caspases. Thereforewe analysed whether the general caspase inhibitor z-VAD-fmkaffected LAT cleavage. As shown in Figure 2(A), z-VAD-fmk treatment of LAT plus JCaM2 cells prevented Fas-mediatedLAT cleavage. To identify the protease(s) responsible for Fas-mediated LAT cleavage, LAT plus JCaM2 were incubated withinhibitors specific for caspases 2, 3, 9 and 8/granzyme-B. Asshown in Figure 2(B), inhibition of caspase 8/granzyme B with z-IETD-fmk almost totally abrogated LAT rupture. Also, inhibitionof caspases 3 and 9 modestly reduced Fas-mediated LAT cleavage,but this effect could be due to ‘cross-inhibition’ of caspase 8or granzyme B [28]. In contrast, inhibition of caspase 2 barelyaffected LAT cleavage. We also analysed whether LAT cleavagecould be inhibited by caspase inhibitors in mouse thymocytes. Asshown in Figure 2(C), treatment with the general caspase inhibitorz-VAD-fmk blocked LAT cleavage induced by dexamethasone inmouse thymocytes. Interestingly, although inhibition of caspase 2did not affect LAT cleavage, treatment with caspase 8/granzymeB inhibitor reduced LAT cleavage (Figure 2C). Together, thesedata suggest that caspase 8 or granzyme B are implicated in LATcleavage.
Proteolytic cleavage generates discrete N-terminal fragments ofLAT
To characterize LAT fragments resulting from its cleavage,we compared the size of fragments generated in JCaM2 cellsexpressing wild-type LAT or LAT molecules fused at their C-terminus with a GFP or a 6×His tag (upper panel, Figure 2D).Following a 4-h treatment with anti-Fas antibodies, cells werelysed and analysed for LAT cleavage by Western blotting. Asshown in Figure 2(D), proteolytic fragments generated in LAT–GFP, LAT–6×His and wild-type LAT-expressing cells had exactlythe same electrophorectic mobility. This result, together with thefact that the 06-807 anti-LAT antibody used in the Western blotwas generated against a peptide corresponding to a sequence nearthe N-terminus of the human LAT, demonstrates that the observedLAT fragments correspond to N-terminal portions of this adaptormolecule.
LAT is cleaved at three discrete aspartic acid residues
To predict the amino acid residues at which LAT is cleaved,we took into account the specificity of caspase 8/granzyme Binhibitors, but also the size of the fragments resulting fromLAT cleavage. By combining these prerequisites with the useof the MEROPS database [29], we determined candidate caspase8/granzyme B cleavage sites in the LAT sequence (Figure 3A).
c© The Authors Journal compilation c© 2013 Biochemical Society
514 A. Garcıa-Blesa and others
Figure 1 LAT is proteolytically cleaved in T-cells upon different stimuli
(A) JCaM2 LAT-deficient cells were transfected with plasmids containing the fusion protein LAT–6×His (upper panel). Upon treatment with 100 ng/ml anti-Fas antibody, cleavage of LAT was assessedin cell lysates by Western blotting (WB; lower left-hand panel). The same cell lysates were assessed for Grb2 cleavage by Western blotting with anti-Grb2 antibody (lower right-hand panel). (B)Human fresh (left-hand panel) or PHA-IL-2 activated (right-hand panel) PBMCs were treated during different time periods with medium alone (NT), 100 ng/ml anti-Fas (F 0.1), 1 μg/ml anti-Fas (F1),1 μM staurosporine (St) or plate-bound anti-CD3 antibodies and cell lysates analysed by Western blotting. (C) Thymocytes obtained from C57Bl/6 mice were incubated for the indicated periodswith NT, plate-bound anti-CD3 or anti-Fas antibodies, and then analysed for LAT cleavage by Western blotting (upper panel). The membrane was stripped and blotted with anti-β-actin antibody toshow equal protein load (lower panel). Molecular masses in kDa are indicated to the side of the Western blots.
Asp138 and Asp225 residues in human LAT constituted the bestcandidates for LAT cleavage sites. Mutating each of them intoalanine demonstrated that the mutation of Asp225 had no effecton LAT cleavage, whereas mutation of Asp138 into an alanineprevented the generation of the 18–20 kDa band following Fascross-linking (Figure 3B). Analysis of the additional asparticacid residues found at position 160, 169, 170 and 186 showedthat mutation of Asp186 residue was sufficient to prevent theappearance of the 25–30 kDa fragment (Figure 3B). Surprisingly,simultaneous mutation of Asp138 and Asp186 residues did notprevent LAT cleavage, since in this mutant Fas cross-linking
induced the appearance of a band of ∼16 kDa, not observedin LAT wild-type expressing cells (lower left-hand panel,Figure 3B). Given the size of this band, we hypothesized thatthis fragment should be generated by a cleavage at Asp126. Indeed,simultaneous mutation of Asp126, Asp138 and Asp186 totally blockedLAT cleavage on Fas engagement in transfected JCaM2 cells(lower right-hand panel, Figure 3B). Therefore, from these data,we concluded that human LAT is proteolytically cleaved at Asp126,Asp138 and Asp186 (Figure 3C). This also suggests that mutationof Asp138 and Asp186 leads to the appearance of a cryptic cleavagesite centred on Asp126.
c© The Authors Journal compilation c© 2013 Biochemical Society
Tyrosine phosphorylation of LAT 515
Figure 2 Cleavage of LAT is mediated by caspases
(A) JCaM2 cells transfected with wild-type LAT were incubated for 3 h with the caspase general inhibitor z-VAD-fmk and then treated or not with anti-Fas mAb for 4 h. Cell lysates were analysedfor LAT cleavage by Western blotting. (B) Transfected JCaM2 cells were incubated for 3 h with the indicated specific caspase inhibitors and then treated for 4 h with 100 ng/ml anti-Fas mAb. Celllysates were analysed by Western blotting. (C) Mouse thymocytes were treated with the indicated specific caspase inhibitors and 10 μM dexamathasone for 2 h. Cell lysates were analysed for LATcleavage by Western blotting. (D) JCaM2 LAT-deficient cells were transfected with plasmids containing LAT or the fusion proteins LAT–6×His or LAT–GFP (upper panel). Following 4 h of treatmentwith anti-Fas antibodies, cell lysates were analysed for LAT cleavage by Western blotting (lower panel). Molecular masses in kDa are indicated to the side of the Western blots.
Fas engagement prevents TCR-mediated intracellular signalling
Given that the close proximity of Asp126, Asp138 and Asp186
to tyrosine residues is shown to be important for LATfunction, we evaluated whether the signalling capabilities ofLAT were modified on mutation of Asp126, Asp138 and Asp186.JCaM2 cells expressing either wild-type LAT or the LAT3DA
(D126A/D138A/D186A) mutant were thus stimulated with anti-CD3 antibodies. As shown in Figure 4(A), JCaM2 cells expressingwild-type LAT molecules and LAT3DA molecules showed nodifference in the kinetics and magnitude of phosphorylation ofTyr132, Tyr171, Tyr191 and Tyr226. Similarly, phosphorylation ofERK and PLC-γ 1 were not affected by mutation of aspartic acidresidues in LAT (Figure 4B). Moreover, similar Ca2 + influxeswere generated on TCR cross-linking in JCaM2 cells expressingwild-type LAT or LAT3DA molecules (Figure 4C). Thereforemutation of Asp126, Asp138 and Asp186 does not affect LATsignalling capabilities.
We (results shown in Figure 4A) and others [23,27]have observed that treatment with anti-Fas antibody for90 min inhibited phosphorylation of LAT tyrosine residues.Unexpectedly, the ‘uncleavable’ LAT3DA mutant showed thesame inhibition of its phosphorylation as the wild-type form(Figure 4A). Accordingly, Fas-mediated inhibition of ERK andPLC-γ phosphorylation was comparable in JCaM2 expressingwild-type LAT molecules and LAT3DA molecules (Figure 4B).Moreover, Fas-mediated inhibition of Ca2 + influx generated afterCD3 cross-linking was similar in JCaM2 cells expressing wild-type LAT and LAT3DA (Figure 4C). Therefore the results obtainedwith LAT3DA expressing cells do not support a role for LAT
cleavage as a central mechanism explaining the inhibition of TCR-mediated activation signals that happens on Fas cross-linking.Probably, other upstream signalling molecules also cleavedfollowing Fas engagement could be at the base of the similarinhibition of signals observed in wild-type and mutant LAT3DA
molecules.
Treatment with H2O2, anti-CD3 antibodies or pervanadate inhibitsFas-mediated cleavage of LAT
As demonstrated previously, the function of LAT as a dockingplatform for intracellular signal transduction depends mainlyon phosphorylation of its four C-terminal tyrosine residues[1,9,10,30,31]. Strikingly, the aspartic acid residues at which LATis cleaved are located close to several of these relevant tyrosineresidues, raising the possibility that their phosphorylation couldhave a regulatory role over LAT cleavage. To induce a robustphosphorylation of LAT before Fas engagement, transfectedJCaM2 cells were treated with either anti-CD3 antibodies orH2O2, since both stimuli have been shown to induce protein-tyrosine phosphorylation of several substrates including LAT [32–34]. Interestingly, treatment with 11 mM H2O2 for 5 min beforeFas engagement totally abrogated LAT cleavage (upper panel,Figure 5A). Moreover, treating cells for 5 min with anti-CD3before Fas engagement also induced a moderate reduction in LATcleavage, which was more evident when cells were treated with10 ng/ml anti-Fas antibody (upper panel, Figure 5A). The reducedeffect of CD3 engagement on LAT cleavage with regard to H2O2
treatment could be due to the higher ability of the latter to induce
c© The Authors Journal compilation c© 2013 Biochemical Society

516 A. Garcıa-Blesa and others
Figure 3 LAT is cleaved at three aspartic acid residues
(A) LAT amino acid sequence. Aspartic acid residues where LAT could be cleaved are in bold and underlined. Functionally relevant tyrosine residues at positions 127, 132, 171, 191 and 226 arehighlighted. (B) JCaM2 cells transfected with wt (wild-type) LAT or the indicated mutants were treated for 4 h at 37◦C with 100 ng/ml anti-Fas antibody. LAT cleavage was assessed in total celllysates by Western blot analysis with anti-LAT antibody. Molecular masses in kDa are indicated to the side of the Western blots. (C) Schematic representation of full-length LAT and the differentproteolytic fragments.
tyrosine phosphorylation of intracellular substrates (middle lanes,Figure 5A). We also analysed the effect of pervanadate pre-treatment over LAT cleavage, since this constitutes a differentmethod for inducing its phosphorylation [35]. As shown in Fig-ure 5(B), pre-treatment of JCaM2 cells with freshly prepared 1 %pervanadate strongly inhibited Fas-mediated cleavage of LAT.
Then, we analysed the inhibitory effect of LAT phosphorylationover its cleavage in mouse thymocytes. Given that anti-CD3treatment induces LAT cleavage in thymocytes, we performedH2O2 pre-treatment before incubation with medium alone or anti-Fas antibody. Consistent with the results obtained in transfectedJCaM2 cells, H2O2 pre-treatment clearly reduced LAT cleavage:generation of proteolytic bands on 1-h incubation with mediumalone or anti-Fas antibody is almost totally abrogated in H2O2
pre-treated thymocytes (Figure 5C).It has been proposed that the Fas-triggered cleavage of other
adaptor molecules such as Gads could be a mechanism for
uncoupling signalling pathways downstream of the TCR–CD3complex during apoptosis, preventing the transduction of survivalsignals [26]. In this context, breakdown of LAT is of greatinterest since LAT acts upstream of Gads. Moreover, the specificinhibitory effect of H2O2 and anti-CD3 mAb pre-treatment overFas-triggered cleavage of LAT suggests that phosphorylationcould be a negative feedback mechanism acting over the cleavageof this molecule. In such a model, activation of T-cells through theTCR–CD3 complex leads to LAT phosphorylation, preventingthe cleavage that would disrupt TCR-mediated activation signals.In order to verify this model, we analysed the effect of pervanadatepre-treatment of cells again, but in order to achieve a more specificeffect, transfectant cells were pre-incubated with pervanadatefor 5 min and then pervanadate was diluted 20 times before in-cubation with anti-Fas antibody. As shown in Figure 6, pre-incubation of wild-type LAT expressing cells with pervanadateinhibited its Fas-triggered cleavage. The inhibitory effect of
c© The Authors Journal compilation c© 2013 Biochemical Society

Tyrosine phosphorylation of LAT 517
Figure 4 Mutation of aspartic acid residues of LAT does not modify itssignalling ability
JCaM2 cells expressing either LAT wt (wild-type) or the LAT3DA mutant were pre-treated for90 min with medium alone or 1 μg/ml anti-Fas antibody, and then stimulated for the indicatedperiods with anti-CD3 antibody. (A) Whole cell lysates obtained were probed by Western blottingfor the phosphorylation of specific LAT tyrosine residues with different antibodies. The uppermembrane was stripped and blotted with anti-LAT antibody to show equal protein expression inJCaM2-LAT wt and JCaM2-LAT3DA cells. (B) Whole cell lysates were used to analyse ERK andPLC-γ phosphorylation by using specific antibodies. Stripped membranes were also blotted withanti-ERK and anti-PLC-γ mAbs to show equal protein expression. (C) JCaM2 cells expressingLAT wt (thick continuous line) or the LAT3DA mutant (thin continuous line) were left untreatedor pre-treated with 100 ng/ml anti-Fas mAb for 90 min (dotted line and dashed line for LATwt and LAT3DA respectively); on loading with Indo-1 cells were stimulated with anti-CD3 mAb(1 μg/ml) at the indicated time (arrow). The intracellular Ca2 + concentration was determinedat 37◦C through the change in Indo-1 fluorescence. P-, phospho-.
pervanadate, however, was not as strong as that observed for H2O2
(Figure 5A), probably owing to the fact that concentration ofH2O2 was maintained during the incubation with anti-Fas anti-body. Indeed, by maintaining pervanadate concentration during
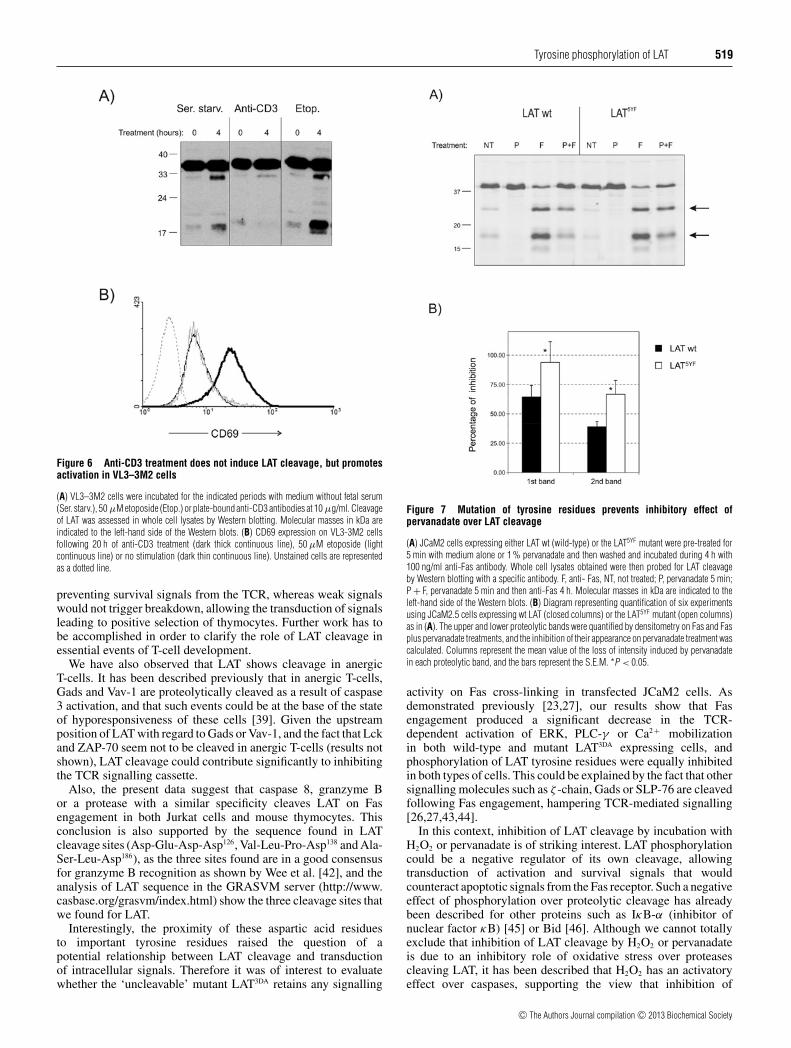
the 4-h incubation with anti-Fas, LAT cleavage was almost totallyinhibited (see Figure 5B). We further examined the pervanadateeffect on LAT cleavage in VL3–3M2 cells, an immortalizedCD4+ CD8+ cell line derived from a mouse thymic lymphoma[36]. As shown in Figure 5(D), VL3–3M2 cells pre-treated for10 min with 2% pervanadate results in a small reduction in theLAT cleavage induced by etoposide treatment. The weak effectobserved of pervanadate pre-treatment is probably due to thefact that upon pre-treatment with 2% pervanadate cells werewashed in order to remove it, and then incubated for 8 h in thepresence of etoposide. Indeed, sustained low-dose pervanadatetreatment of VL3-3M2 cells totally abrogated LAT cleavage, andLAT phosphorylation was evident. This effect is specific for LATsince PARP cleavage (a typical marker of apoptosis) is not affectedby pervanadate treatment (Figure 5D).
Interestingly, anti-CD3 mAb treatment of thymocytes, but notin T-lymphocytes or Jurkat cells, induces LAT cleavage. It iswidely accepted that TCR–CD3-mediated stimulation inducesphosphorylation of LAT, which should protect it from cleavage. Itis conceivable that the outcome of TCR engagement in thymocytesis different from mature T-cells, with negative-selecting strongsignals leading to LAT cleavage. Given that stimulation ofthymocytes with anti-CD3 mAb induce cell death even at lowconcentrations, we treated the VL3-3M2 thymocyte cell line(known to be resistant to CD3-induced apoptosis) with plate-bound anti-CD3 mAb or etoposide. As shown in Figure 6(A),treatment of VL3-3M2 cells with anti-CD3 antibodies does notinduce LAT cleavage following etoposide treatment, but it inducesthe expression of CD69, a typical marker of positive selection(Figure 6B). Therefore, this result, together with the effect of sus-tained low-dose pervanadate treatment over LAT phosphorylationand cleavage, supports the view that during thymic developmentthe interplay between LAT phosphorylation and cleavage couldhave an important role for positive and negative selection.
In addition, in order to confirm the inverse relationship betweenphosphorylation and cleavage, we mutated tyrosine residuesof LAT located near aspartic acid residues by which it iscleaved (Tyr127, Tyr132, Tyr171, Tyr191 and Tyr226, see Figure 3A),generating the LAT5YF (Y127F/Y132F/Y171F/Y191F/Y226F)mutant. Mutation of the five C-terminal tyrosine residues reducesthe inhibitory effect granted by pervanadate pre-treatment (seethe intensity of the proteolytic bands marked with arrows,Figure 7A). Figure 7(B) represents the quantification of upper andlower proteolytic bands of LAT in six independent experimentsusing JCaM2.5 cells expressing wild-type LAT or the LAT5YF
mutant. The upper and lower proteolytic bands were quantified bydensitometry following Fas and Fas plus pervanadate treatments,and the inhibition of their appearance after pervanadate treatmentwas calculated. As it can be seen, pervanadate diminished theintensity of both proteolytic bands in wild-type LAT expressingcells, but mutation of the five C-terminal tyrosine residuesdecreased the effect of pervanadate, with a P value lower than0.05.
Therefore these results support a model in which LAT phos-phorylation would negatively regulate its cleavage, counteractingthe inhibition of proximal TCR signalling elicited by Fasengagement.
Relationship between anergy induction and LAT cleavage
Clonal anergy is a process triggered by TCR engagement in theabsence of co-stimulatory signals and leading to the uncouplingof antigen receptors from their downstream signalling pathways
c© The Authors Journal compilation c© 2013 Biochemical Society

518 A. Garcıa-Blesa and others
Figure 5 LAT phosphorylation with H2O2, anti-CD3 or pervanadate antibodies lowers LAT cleavage
(A) JCaM2 cells (106) transfected with a lentiviral vector coding for wild-type LAT were treated during 5 min with 11 mM H2O2 or 1 μg of anti-CD3 antibody, and then incubated during 4 h with10 or 100 ng/ml of anti-Fas antibody. LAT cleavage (upper panel) and tyrosine phosphorylation (P-Tyr; lower panel) were analysed by Western blotting with specific antibodies. (B) JCaM2 cells(1×106) expressing wild-type LAT were treated during 5 min with pervanadate and then incubated during 4 h with 100 ng/ml anti-Fas antibody. F, anti-Fas; NT, not treated; P, pervanadate 5 min;P + F, pervanadate 5 min and then anti-Fas 4 h. (C) Mouse thymocytes were treated for 2 min with medium alone or H2O2 and then incubated for the indicated time periods with plate-boundanti-Fas antibody or NT. LAT cleavage was analysed in cell lysates by Western blotting with a specific antibody. (D) VL3-3M2 cells were pre-treated for 10 min with medium or 2 % pervanadate (PV)and then washed and treated with 50 μM etoposide for 8 h or 50 μM etoposide plus 0.5 % pervanadate. Whole cell lysates were obtained and assayed by Western blotting for LAT cleavage, LATphosphorylation and PARP cleavage. The membrane was revealed with anti-β-actin antibody to show equal protein load. Molecular masses in kDa are indicated to the left-hand side of the Westernblot.
[37,38]. It has been proposed that proteolytic mechanisms areimplicated in the uncoupling of the TCR–CD3 complex fromintracellular signalling machinery that takes place during anergy[17,39]. Therefore, given the central role of LAT for transducingsignals from the TCR, we sought to study its cleavage in anergicT-cells. Accordingly, PBMCs from healthy donors were culturedin the presence of PHA and IL-2, and then anergy was induced inpurified CD4 T-cells by 1 μM ionomycin for 20 h. Anergywas evaluated in these cells by their proliferative capacity inresponse to plate-bound anti-CD3 and anti-CD28 antibodies.As shown in Figure 7(A), ionomycin-treated PBMCs have aclearly diminished proliferative capacity when compared withuntreated cells. Interestingly, anergy induction induced cleavageof LAT in the absence of any other stimulus (Figure 7B),but treatment with anti-CD3 or anti-Fas antibodies did notenhance LAT cleavage. Therefore, these data further support amodel by which LAT cleavage could be one of the mechanismsinvolved in uncoupling TCR from intracellular activatory signals,contributing to maintain peripheral tolerance.
DISCUSSION
LAT is an essential membrane adaptor co-ordinating assemblyof signalling complexes on TCR engagement of T-lymphocytes[4], and is also crucial for proper thymic development [40]. Mostof the activatory capacity of LAT seems to relay in its four C-
terminal tyrosine residues [1,30]. However, LAT also constitutes anegative regulator of TCR signalling, since two strains of knockinmice harbouring mutant forms of LAT for several of its tyrosineresidues showed a Th2 lymphoproliferative disease [9–11]. Thisparadoxical finding revealed that LAT also constitutes a negativeregulator of TCR signalling whose molecular base is not known.
As demonstrated previously, LAT and Gads are cleavedfollowing Fas cross-linking in Jurkat cells [26], and cleavageof Gads has been proposed as a mechanism for the negativeregulation of TCR signalling. In the present study, wedemonstrated that LAT cleavage occurs not only in Jurkat cells,but also in activated T-lymphocytes, mouse thymocytes andanergic T-cells in response to apoptotic stimuli.
Interestingly, LAT is cleaved in thymocytes in the absenceof any treatment, and also on CD3 cross-linking. In agreementwith previous data reporting caspase 8 activity in double positivethymocytes [41], the degree of LAT cleavage is higher in thymo-cytes when compared with peripheral T-lymphocytes, withuntreated thymocytes showing a fast degradation that does notoccur in peripheral T-cells or Jurkat cells. It is widely acceptedthat T-cell development requires the appropriate transduction ofTCR-associated signals: high-strength binding of the TCR to itsMHC peptide leads to cell death, whereas intermediate affinityinteractions lead to survival and differentiation. In this context,cleavage of LAT could constitute a mechanism controlling theintensity of intracellular signals triggered by the TCR. It isconceivable that strong signals could lead to LAT cleavage
c© The Authors Journal compilation c© 2013 Biochemical Society
Tyrosine phosphorylation of LAT 519
Figure 6 Anti-CD3 treatment does not induce LAT cleavage, but promotesactivation in VL3–3M2 cells
(A) VL3–3M2 cells were incubated for the indicated periods with medium without fetal serum(Ser. starv.), 50 μM etoposide (Etop.) or plate-bound anti-CD3 antibodies at 10 μg/ml. Cleavageof LAT was assessed in whole cell lysates by Western blotting. Molecular masses in kDa areindicated to the left-hand side of the Western blots. (B) CD69 expression on VL3-3M2 cellsfollowing 20 h of anti-CD3 treatment (dark thick continuous line), 50 μM etoposide (lightcontinuous line) or no stimulation (dark thin continuous line). Unstained cells are representedas a dotted line.
preventing survival signals from the TCR, whereas weak signalswould not trigger breakdown, allowing the transduction of signalsleading to positive selection of thymocytes. Further work has tobe accomplished in order to clarify the role of LAT cleavage inessential events of T-cell development.
We have also observed that LAT shows cleavage in anergicT-cells. It has been described previously that in anergic T-cells,Gads and Vav-1 are proteolytically cleaved as a result of caspase3 activation, and that such events could be at the base of the stateof hyporesponsiveness of these cells [39]. Given the upstreamposition of LAT with regard to Gads or Vav-1, and the fact that Lckand ZAP-70 seem not to be cleaved in anergic T-cells (results notshown), LAT cleavage could contribute significantly to inhibitingthe TCR signalling cassette.
Also, the present data suggest that caspase 8, granzyme Bor a protease with a similar specificity cleaves LAT on Fasengagement in both Jurkat cells and mouse thymocytes. Thisconclusion is also supported by the sequence found in LATcleavage sites (Asp-Glu-Asp-Asp126, Val-Leu-Pro-Asp138 and Ala-Ser-Leu-Asp186), as the three sites found are in a good consensusfor granzyme B recognition as shown by Wee et al. [42], and theanalysis of LAT sequence in the GRASVM server (http://www.casbase.org/grasvm/index.html) show the three cleavage sites thatwe found for LAT.
Interestingly, the proximity of these aspartic acid residuesto important tyrosine residues raised the question of apotential relationship between LAT cleavage and transductionof intracellular signals. Therefore it was of interest to evaluatewhether the ‘uncleavable’ mutant LAT3DA retains any signalling
Figure 7 Mutation of tyrosine residues prevents inhibitory effect ofpervanadate over LAT cleavage
(A) JCaM2 cells expressing either LAT wt (wild-type) or the LAT5YF mutant were pre-treated for5 min with medium alone or 1 % pervanadate and then washed and incubated during 4 h with100 ng/ml anti-Fas antibody. Whole cell lysates obtained were then probed for LAT cleavageby Western blotting with a specific antibody. F, anti- Fas, NT, not treated; P, pervanadate 5 min;P + F, pervanadate 5 min and then anti-Fas 4 h. Molecular masses in kDa are indicated to theleft-hand side of the Western blots. (B) Diagram representing quantification of six experimentsusing JCaM2.5 cells expressing wt LAT (closed columns) or the LAT5YF mutant (open columns)as in (A). The upper and lower proteolytic bands were quantified by densitometry on Fas and Fasplus pervanadate treatments, and the inhibition of their appearance on pervanadate treatment wascalculated. Columns represent the mean value of the loss of intensity induced by pervanadatein each proteolytic band, and the bars represent the S.E.M. *P < 0.05.
activity on Fas cross-linking in transfected JCaM2 cells. Asdemonstrated previously [23,27], our results show that Fasengagement produced a significant decrease in the TCR-dependent activation of ERK, PLC-γ or Ca2 + mobilizationin both wild-type and mutant LAT3DA expressing cells, andphosphorylation of LAT tyrosine residues were equally inhibitedin both types of cells. This could be explained by the fact that othersignalling molecules such as ζ -chain, Gads or SLP-76 are cleavedfollowing Fas engagement, hampering TCR-mediated signalling[26,27,43,44].
In this context, inhibition of LAT cleavage by incubation withH2O2 or pervanadate is of striking interest. LAT phosphorylationcould be a negative regulator of its own cleavage, allowingtransduction of activation and survival signals that wouldcounteract apoptotic signals from the Fas receptor. Such a negativeeffect of phosphorylation over proteolytic cleavage has alreadybeen described for other proteins such as IκB-α (inhibitor ofnuclear factor κB) [45] or Bid [46]. Although we cannot totallyexclude that inhibition of LAT cleavage by H2O2 or pervanadateis due to an inhibitory role of oxidative stress over proteasescleaving LAT, it has been described that H2O2 has an activatoryeffect over caspases, supporting the view that inhibition of
c© The Authors Journal compilation c© 2013 Biochemical Society

520 A. Garcıa-Blesa and others
Figure 8 LAT is cleaved in anergic T-lymphocytes
PBMCs from healthy donors were cultured in the presence of PHA and IL-2, and then anergywas induced over purified CD4 T-cells with 1 μM ionomycin for 20 h. (A) Evaluation of anergyinduction by the proliferative capacity in response to plate-bound anti-CD3 and anti-CD28antibodies, measured with the Click-iT EdU Flow Cytometry Assay Kit (Invitrogen). (B) Cellsincubated for 20 h with medium alone (untreated) or 1 μM ionomycin were incubated for 4 hwith plate-bound anti-CD3, soluble anti-Fas antibody or medium alone, and LAT cleavage wasanalysed by Western blotting. Molecular masses in kDa are indicated to the right-hand side ofthe Western blots.
LAT cleavage could be due to its phosphorylation [47,48]. Insupport to this model, we show a reduced inhibitory effect ofpervanadate treatment in the LAT5YF mutant, in which its fiveC-terminal tyrosine residues have been mutated, preventing theirphosphorylation and, consequently, its putative inhibitory effectover cleavage. However, by maintaining the pervanadate or H2O2
during anti-Fas treatment, cleavage of LAT5YF mutant was alsototally inhibited (results not shown). This could be due to theremaining tyrosine residues in the LAT5YF molecule; alternatively,pervanadate might induce a double effect directly inhibitingtyrosine phosphatases, but indirectly activating serine/threoninekinases that could in turn phosphorylate LAT. However, mutationof functionally relevant Ser38, Ser40, Ser106, Ser164 and Ser180 had noeffect on the inhibition of LAT cleavage mediated by pervanadate(results not shown), but some of the remaining 28 serine residuesor nine threonine residues of LAT could be implicated.
In summary, our results suggest an inverse correlation betweenLAT phosphorylation and its cleavage. It has been previouslyshown that LAT is able to bind to the open/active isoformof Lck, and this interaction seems to negatively regulate Lckkinase [49]. Interestingly, Kabouridis et al. [50] have recentlydemonstrated that interaction between LAT and the open form ofLck is mediated by a negatively charged stretch between aminoacids 112 and 126 of LAT. Therefore LAT cleavage at Asp126
would leave a proteolytic fragment still able to bind Lck, andthis could constitute a new mechanism for negative regulationof the TCR–CD3 signalling cassette. Moreover, this group hasalso shown that a truncated form of LAT ending at residue136 co-immunoprecipitated more efficiently with Lck than full-length LAT, possibly because in this truncated form the acidicdomain is better exposed for association with Lck. The strikingsimilarity between this 136-truncated LAT and the putative LAT-138 truncated form of LAT that is generated following Fasengagement suggests a new mechanism by which the Fas pathwaycan regulate TCR-mediated activation.
AUTHOR CONTRIBUTION
All authors contributed to discussions of experimental design anddata analysis; Antonio Garcıa-Blesa did all of the experimentalstudies unless otherwise indicated; Mikolai Klossowicz andArkadius Miazek performed experiments involving mousethymocytes and VL3-3M2 cell line. Carmen Lopez-Osuna andMario Martınez-Florensa performed mutagenesis and Westernblots and provided technical assistance; Francisco Garcıa-Cozar,Bernard Malissen and Arkadius Miazek provided suggestions;Enrique Aguado directed the study and wrote the manuscript.
ACKNOWLEDGEMENT
We thank Dr Arthur Weiss (University of California, SanFrancisco, San Francisco, CA, U.S.A.) for the JCaM2 cell line.
FUNDING
This work was supported by the Polish National Science Centre[grant number NN401057237 (to A.M.)], Consejerıa de Salud deAndalucıa, Junta de Andalucıa [grant numbers SAS 111206 (toE.A.) and PI-0007/2007 (to F.G.-C.)], Spanish Ministry of Scienceand Innovation (MICINN) [grant number SAF2009-09449 (toF.G.-C.)] and Consejerıa de Economıa, Innovacion y Ciencia,Junta de Andalucia [grant number P08-CTS-04348 (to F.G.-C.)].
REFERENCES
1 Lin, J. and Weiss, A. (2001) Identification of the minimal tyrosine residues required forlinker for activation of T cell function. J. Biol. Chem. 276, 29588–29595
2 Aguado, E., Martinez-Florensa, M. and Aparicio, P. (2006) Activation of T lymphocytesand the role of the adapter LAT. Transplant Immunol. 17, 23–26
3 Guy, C. S. and Vignali, D. A. A. (2009) Organization of proximal signal initiation at theTCR:CD3 complex. Immunol. Rev. 232, 7–21
4 Zhang, W., Sloan-Lancaster, J., Kitchen, J., Trible, R. P. and Samelson, L. E. (1998) LAT:the ZAP-70 tyrosine kinase substrate that links T cell receptor to cellular activation. Cell92, 83–92
5 Asada, H., Ishii, N., Sasaki, Y., Endo, K., Kasai, H., Tanaka, N., Takeshita, T., Tsuchiya, S.,Konno, T. and Sugamura, K. (1999) Grf40, a novel Grb2 family member, is involved in Tcell signaling through interaction with SLP-76 and LAT. J. Exp. Med. 189, 1383–1390
6 Liu, S. K., Fang, N., Koretzky, G. A. and McGlade, C. J. (1999) The hematopoietic-specificadaptor protein gads functions in T-cell signaling via interactions with the SLP-76 andLAT adaptors. Curr. Biol. 9, 67–75
c© The Authors Journal compilation c© 2013 Biochemical Society

Tyrosine phosphorylation of LAT 521
7 Paz, P. E., Wang, S., Clarke, H., Lu, X., Stokoe, D. and Abo, A. (2001) Mapping the Zap-70phosphorylation sites on LAT (linker for activation of T cells) required for recruitment andactivation of signalling proteins in T cells. Biochem. J. 356, 461–471
8 Zhang, W., Sommers, C. L., Burshtyn, D. N., Stebbins, C. C., DeJarnette, J. B., Trible,R. P., Grinberg, A., Tsay, H. C., Jacobs, H. M., Kessler, C. M. et al. (1999) Essential role ofLAT in T cell development. Immunity 10, 323–332
9 Aguado, E., Richelme, S., Nunez-Cruz, S., Miazek, A., Mura, A. M., Richelme, M., Guo,X. J., Sainty, D., He, H. T., Malissen, B. and Malissen, M. (2002) Induction of T helpertype 2 immunity by a point mutation in the LAT adaptor. Science 296, 2036–2040
10 Sommers, C. L., Park, C. S., Lee, J., Feng, C., Fuller, C. L., Grinberg, A., Hildebrand,J. A., Lacana, E., Menon, R. K., Shores, E. W. et al. (2002) A LAT mutation that inhibits Tcell development yet induces lymphoproliferation. Science 296, 2040–2043
11 Nunez-Cruz, S., Aguado, E., Richelme, S., Chetaille, B., Mura, A. M., Richelme, M.,Pouyet, L., Jouvin-Marche, E., Xerri, L., Malissen, B. and Malissen, M. (2003) LATregulates γ δ T cell homeostasis and differentiation. Nat. Immunol. 4, 999–1008
12 Matsuda, S., Miwa, Y., Hirata, Y., Minowa, A., Tanaka, J., Nishida, E. and Koyasu, S.(2004) Negative feedback loop in T-cell activation through MAPK-catalyzed threoninephosphorylation of LAT. EMBO J. 23, 2577–2585
13 Zahedi, R. P., Lewandrowski, U., Wiesner, J., Wortelkamp, S., Moebius, J., Schutz, C.,Walter, U., Gambaryan, S. and Sickmann, A. (2007) Phosphoproteome of resting humanplatelets. J. Proteome Res. 7, 526–534
14 Martinez-Florensa, M., Garcia-Blesa, A., Yelamos, J., Munoz-Suano, A.,Dominguez-Villar, M., Valdor, R., Alonso, A., Garcia-Cozar, F., Aparicio, P., Malissen, B.and Aguado, E. (2011) Serine residues in the LAT adaptor are essential forTCR-dependent signal transduction. J. Leukocyte Biol. 89, 63–73
15 Winslow, M. M., Neilson, J. R. and Crabtree, G. R. (2003) Calcium signalling inlymphocytes. Curr. Opin. Immunol. 15, 299–307
16 Malissen, B., Aguado, E. and Malissen, M. (2005) Role of the LAT adaptor in T-celldevelopment and Th2. Adv. Immunol. 87, 1–25
17 Macian, F., Garcia-Cozar, F., Im, S. H., Horton, H. F., Byrne, M. C. and Rao, A. (2002)Transcriptional mechanisms underlying lymphocyte tolerance. Cell 109, 719–731
18 Snow, A. L., Pandiyan, P., Zheng, L., Krummey, S. M. and Lenardo, M. J. (2010) Thepower and the promise of restimulation-induced cell death in human immune diseases.Immunol. Rev. 236, 68–82
19 Danial, N. N. and Korsmeyer, S. J. (2004) Cell death: critical control points. Cell 116,205–219
20 Coultas, L. and Strasser, A. (2003) The role of the Bcl-2 protein family in cancer. Semin.Cancer Biol. 13, 115–123
21 Strasser, A., Jost, P. J. and Nagata, S. (2009) The many roles of FAS receptor signaling inthe immune system. Immunity 30, 180–192
22 Alderson, M. R., Armitage, R. J., Maraskovsky, E., Tough, T. W., Roux, E., Schooley, K.,Ramsdell, F. and Lynch, D. H. (1993) Fas transduces activation signals in normal humanT-lymphocytes. J. Exp. Med. 178, 2231–2235
23 Strauss, G., Lindquist, J. A., Arhel, N., Felder, E., Karl, S., Haas, T. L., Fulda, S., Walczak,H., Kirchhoff, F. and Debatin, K. M. (2009) CD95 co-stimulation blocks activation of naiveT cells by inhibiting T cell receptor signaling. J. Exp. Med. 206, 1379–1393
24 Finco, T. S., Kadlecek, T., Zhang, W., Samelson, L. E. and Weiss, A. (1998) LAT is requiredfor TCR-mediated activation of PLCγ 1 and the Ras pathway. Immunity 9, 617–626
25 Groves, T., Katis, P., Madden, Z., Manickam, K., Ramsden, D., Wu, G. and Guidos, C. J.(1995) In vitro maturation of clonal CD4+ CD8 + cell lines in response to TCRengagement. J. Immunol. 154, 5011–5022
26 Berry, D. M., Benn, S. J., Cheng, A. M. and McGlade, C. J. (2001) Caspase-dependentcleavage of the hematopoietic specific adaptor protein Gads alters signalling from the Tcell receptor. Oncogene 20, 1203–1211
27 Yankee, T. M., Draves, K. E., Ewings, M. K., Clark, E. A. and Graves, J. D. (2001)CD95/Fas induces cleavage of the GrpL/Gads adaptor and desensitization of antigenreceptor signaling. Proc. Natl. Acad. Sci. U.S.A 98, 6789–6793
28 Berger, A. B., Sexton, K. B. and Bogyo, M. (2006) Commonly used caspase inhibitorsdesigned based on substrate specificity profiles lack selectivity. Cell Res. 16, 961–963
29 Rawlings, N. D., Barrett, A. J. and Bateman, A. (2012) MEROPS: the database ofproteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 40, D343–D350
30 Zhang, W., Trible, R. P., Zhu, M., Liu, S. K., McGlade, C. J. and Samelson, L. E. (2000)Association of Grb2, Gads, and phospholipase C-γ 1 with phosphorylated LAT tyrosineresidues. Effect of LAT tyrosine mutations on T cell antigen receptor-mediated signaling.J. Biol. Chem. 275, 23355–23361
31 Zhu, M., Janssen, E. and Zhang, W. (2003) Minimal requirement of tyrosine residues oflinker for activation of T cells in TCR signaling and thymocyte development. J. Immunol.170, 325–333
32 Schieven, G. L., Mittler, R. S., Nadler, S. G., Kirihara, J. M., Bolen, J. B., Kanner, S. B. andLedbetter, J. A. (1994) ZAP-70 tyrosine kinase, CD45, and T cell receptor involvement inUV- and H2O2-induced T cell signal transduction. J. Biol. Chem. 269, 20718–20726
33 Griffith, C. E., Zhang, W. and Wange, R. L. (1998) ZAP-70-dependent and -independentactivation of Erk in Jurkat T cells. J. Biol. Chem. 273, 10771–10776
34 Haas, A., Weckbecker, G. and Welzenbach, K. (2008) Intracellular phospho-flowcytometry reveals novel insights into TCR proximal signaling events. A comparison withWestern blot. Cytometry 73A, 799–807
35 Jiang, Y. and Cheng, H. (2007) Evidence of LAT as a dual substrate for Lck and Syk in Tlymphocytes. Leuk. Res. 31, 541–545
36 Lieberman, M., Decleve, A., Ricciardi-Castagnoli, P., Boniver, J., Finn, O. J. and Kaplan,H. S. (1979) Establishment, characterization and virus expression of cell lines derivedfrom radiation- and virus-induced lymphomas of C57BL/Ka mice. Int. J. Cancer 24,168–177
37 Fields, P. E., Gajewski, T. F. and Fitch, F. W. (1996) Blocked ras activation in anergicCD4 + T cells. Science 271, 1276–1278
38 Li, W., Whaley, C. D., Mondino, A. and Mueller, D. L. (1996) Blocked signal transductionto the ERK and JNK protein kinases in anergic CD4 + T cells. Science 271, 1272–1276
39 Puga, I., Rao, A. and Macian, F. (2008) Targeted cleavage of signaling proteins by caspase3 inhibits T cell receptor signaling in anergic T cells. Immunity 29, 193–204
40 Zhang, W., Irvin, B. J., Trible, R. P., Abraham, R. T. and Samelson, L. E. (1999) Functionalanalysis of LAT in TCR-mediated signaling pathways using a LAT-deficient Jurkat cellline. Int. Immunol. 11, 943–950
41 Jiang, D., Zheng, L. and Lenardo, M. J. (1999) Caspases in T-cell receptor-inducedthymocyte apoptosis. Cell Death. Differ. 6, 402–411
42 Wee, L., Er, E., Ng, L. and Tong, J. C. (2011) In silico prediction of the granzyme Bdegradome. BMC Genomics 12, S11
43 Gastman, B. R., Johnson, D. E., Whiteside, T. L. and Rabinowich, H. (1999)Caspase-mediated degradation of T-cell receptor ζ -chain. Cancer Res. 59, 1422–1427
44 Wieckowski, E., Wang, G. Q., Gastman, B. R., Goldstein, L. A. and Rabinowich, H. (2002)Granzyme B-mediated degradation of T-cell receptor ζ chain. Cancer Res. 62, 4884–4889
45 Barkett, M., Xue, D., Horvitz, H. R. and Gilmore, T. D. (1997) Phosphorylation of IκB-αinhibits its cleavage by caspase CPP32 in vitro. J. Biol. Chem. 272, 29419–29422
46 Desagher, S., Osen-Sand, A., Montessuit, S., Magnenat, E., Vilbois, F., Hochmann, A.,Journot, L., Antonsson, B. and Martinou, J. C. (2001) Phosphorylation of bid by caseinkinases I and II regulates its cleavage by caspase 8. Mol. Cell 8, 601–611
47 Matsura, T., Kai, M., Fujii, Y., Ito, H. and Yamada, K. (1999) Hydrogen peroxide-inducedapoptosis in HL-60 cells requires caspase-3 activation. Free Radical Res. 30, 73–83
48 Tamura, H., Ohtsuru, A., Kamohara, Y., Fujioka, H., Yanaga, K., Kanematsu, T. andYamashita, S. (2003) Bax cleavage implicates caspase-dependent H2O2-inducedapoptosis of hepatocytes. Int. J. Mol. Med. 11, 369–374
49 Kabouridis, P. S. (2003) Selective interaction of LAT (linker of activated T cells) with theopen-active form of Lck in lipid rafts reveals a new mechanism for the regulation of Lck inT cells. Biochem. J. 371, 907–915
50 Kabouridis, P. S., Isenberg, D. A. and Jury, E. C. (2011) A negatively charged domain ofLAT mediates its interaction with the active form of Lck. Mol. Membr. Biol. 28, 487–494
Received 20 July 2012/6 December 2012; accepted 14 December 2012Published as BJ Immediate Publication 14 December 2012, doi:10.1042/BJ20121135
c© The Authors Journal compilation c© 2013 Biochemical Society
Related Documents











