The lithium tolerance of the Arabidopsis cat2 mutant reveals a cross-talk between oxidative stress and ethylene Eduardo Bueso, Santiago Alejandro, Pablo Carbonell, Miguel A. Perez-Amador, Joaquı ´n Fayos, Jose M. Belle ´ s, Pedro L. Rodriguez and Ramo ´ n Serrano * Instituto de Biologı´a Molecular y Celular de Plantas, Universidad Polite ´ cnica de Valencia-C.S.I.C., Camino de Vera s/n, 46022 Valencia, Spain Received 4 May 2007; accepted 10 August 2007. * For correspondence (fax +34 963 87 7859; e-mail [email protected]). Summary In order to investigate the effects of a permanent increase in cellular H 2 O 2 on cation homeostasis we have studied a T-DNA insertion mutant of the Arabidopsis CATALASE 2 gene. This mutant (cat2-1) exhibits 20% of wild-type leaf catalase activity and accumulates more H 2 O 2 than the wild type under normal growth conditions. In addition to reduced size, a pale green color and great reduction in secondary roots, the cat2-1 mutant exhibited increased sensitivity to H 2 O 2 , NaCl, norspermidine, high light and cold stress. On the other hand, the germination of the cat2-1 mutant is more tolerant to lithium than the wild type. This novel phenotype cannot be explained by changes in lithium transport. Actually, the uptake of lithium (and of other toxic cations such as sodium and norspermidine) is increased in the cat2-1 mutant while K + levels were decreased. The lithium tolerance of this mutant seems to result both from insensitivity to the inhibitory ethylene induced by this cation and a reduced capability for ethylene production. Accordingly, induction by ethylene of responsive genes such as PR4 and EBP/ERF72 is decreased in cat2-1. Mutants insensitive to ethylene such as etr1-1 and ein3-3 are lithium tolerant, and inhibition of ethylene biosynthesis with 2-aminoisobutyrate protects against lithium toxicity. Microarray analysis of gene expression indicates that the expression of genes related to cation transport and ethylene synthesis and perception was not altered in the cat2-1 mutant, suggesting that H 2 O 2 modulates these processes at the protein level. These results uncover a cross-talk between oxidative stress, cation homeostasis and ethylene. Keywords: hydrogen peroxide, cation homeostasis, lithium tolerance, ethylene sensitivity, Arabidopsis. Introduction Hydrogen peroxide (H 2 O 2 ) is a toxic cellular metabolite that, together with other reactive oxygen species (ROS), is pro- duced in chloroplasts, mitochondria, peroxisomes and plasma membranes and is detoxified by enzymatic (cata- lases, ascorbate peroxidases, glutathione peroxidases) and non-enzymatic (ascorbate, glutathione, phenolic com- pounds, carotenoids) mechanisms (Allen, 1995; Apel and Hirt, 2004; Mittler et al., 2004). The generation of H 2 O 2 is increased in response to various biotic and abiotic stresses, and much of the injury caused by stress is associated with oxidative damage to cells (Allen, 1995) and with H 2 O 2 -in- duced programmed cell death (Laloi et al., 2004; Neill et al., 2002). However, it is now clear that H 2 O 2 generated by plasma membrane NADPH oxidases also functions as a signaling molecule in stress and growth responses (Apel and Hirt, 2004; Laloi et al., 2004; Mittler et al., 2004; Neill et al., 2002). It has been shown that H 2 O 2 mediates stomatal closure (Murata et al., 2001) and inhibition of seed germination and root elongation (Kwak et al., 2003) triggered by abscisic acid (ABA) and it is required for the elongation of root hairs (Foreman et al., 2003) and for auxin-regulated root gravitro- pism (Joo et al., 2001). Many effects of H 2 O 2 on plant cells are mediated by changes in cation transport. In particular H 2 O 2 activates Ca 2+ influx channels at the plasma membrane (Foreman et al., 2003; Kwak et al., 2003; Murata et al., 2001; Mustilli et al., 2002; Pei et al., 2000). Elevated cytosolic Ca 2+ blocks inward K + channels and activates anion and K + efflux channels (Blatt and Thiel, 1993; Hedrich et al., 1990; Schroeder and 1052 ª 2007 The Authors Journal compilation ª 2007 Blackwell Publishing Ltd The Plant Journal (2007) 52, 1052–1065 doi: 10.1111/j.1365-313X.2007.03305.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The lithium tolerance of the Arabidopsis cat2 mutant revealsa cross-talk between oxidative stress and ethylene
Eduardo Bueso, Santiago Alejandro, Pablo Carbonell, Miguel A. Perez-Amador, Joaquın Fayos, Jose M. Belles,
Pedro L. Rodriguez and Ramon Serrano*
Instituto de Biologıa Molecular y Celular de Plantas, Universidad Politecnica de Valencia-C.S.I.C., Camino de Vera s/n, 46022
Valencia, Spain
Received 4 May 2007; accepted 10 August 2007.*For correspondence (fax +34 963 87 7859; e-mail [email protected]).
Summary
In order to investigate the effects of a permanent increase in cellular H2O2 on cation homeostasis we have
studied a T-DNA insertion mutant of the Arabidopsis CATALASE 2 gene. This mutant (cat2-1) exhibits 20% of
wild-type leaf catalase activity and accumulates more H2O2 than the wild type under normal growth
conditions. In addition to reduced size, a pale green color and great reduction in secondary roots, the cat2-1
mutant exhibited increased sensitivity to H2O2, NaCl, norspermidine, high light and cold stress. On the other
hand, the germination of the cat2-1 mutant is more tolerant to lithium than the wild type. This novel phenotype
cannot be explained by changes in lithium transport. Actually, the uptake of lithium (and of other toxic cations
such as sodium and norspermidine) is increased in the cat2-1 mutant while K+ levels were decreased. The
lithium tolerance of this mutant seems to result both from insensitivity to the inhibitory ethylene induced by
this cation and a reduced capability for ethylene production. Accordingly, induction by ethylene of responsive
genes such as PR4 and EBP/ERF72 is decreased in cat2-1. Mutants insensitive to ethylene such as etr1-1 and
ein3-3 are lithium tolerant, and inhibition of ethylene biosynthesis with 2-aminoisobutyrate protects against
lithium toxicity. Microarray analysis of gene expression indicates that the expression of genes related to cation
transport and ethylene synthesis and perception was not altered in the cat2-1 mutant, suggesting that H2O2
modulates these processes at the protein level. These results uncover a cross-talk between oxidative stress,
cation homeostasis and ethylene.
Keywords: hydrogen peroxide, cation homeostasis, lithium tolerance, ethylene sensitivity, Arabidopsis.
Introduction
Hydrogen peroxide (H2O2) is a toxic cellular metabolite that,
together with other reactive oxygen species (ROS), is pro-
duced in chloroplasts, mitochondria, peroxisomes and
plasma membranes and is detoxified by enzymatic (cata-
lases, ascorbate peroxidases, glutathione peroxidases) and
non-enzymatic (ascorbate, glutathione, phenolic com-
pounds, carotenoids) mechanisms (Allen, 1995; Apel and
Hirt, 2004; Mittler et al., 2004). The generation of H2O2 is
increased in response to various biotic and abiotic stresses,
and much of the injury caused by stress is associated with
oxidative damage to cells (Allen, 1995) and with H2O2-in-
duced programmed cell death (Laloi et al., 2004; Neill et al.,
2002).
However, it is now clear that H2O2 generated by plasma
membrane NADPH oxidases also functions as a signaling
molecule in stress and growth responses (Apel and Hirt,
2004; Laloi et al., 2004; Mittler et al., 2004; Neill et al., 2002).
It has been shown that H2O2 mediates stomatal closure
(Murata et al., 2001) and inhibition of seed germination and
root elongation (Kwak et al., 2003) triggered by abscisic acid
(ABA) and it is required for the elongation of root hairs
(Foreman et al., 2003) and for auxin-regulated root gravitro-
pism (Joo et al., 2001).
Many effects of H2O2 on plant cells are mediated by
changes in cation transport. In particular H2O2 activates Ca2+
influx channels at the plasma membrane (Foreman et al.,
2003; Kwak et al., 2003; Murata et al., 2001; Mustilli et al.,
2002; Pei et al., 2000). Elevated cytosolic Ca2+ blocks inward
K+ channels and activates anion and K+ efflux channels
(Blatt and Thiel, 1993; Hedrich et al., 1990; Schroeder and
1052 ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd
The Plant Journal (2007) 52, 1052–1065 doi: 10.1111/j.1365-313X.2007.03305.x

Hagiwara, 1989) at the plasma membrane of guard cells. In
addition, H2O2 mediates the responses of plant roots to K+
starvation, which include the induction of high-affinity K+
transport activity and of genes related to ROS metabolism
and K+ transport (Shin and Schachtman, 2004).
These observations suggest that H2O2 could be a general
modulator of ion homeostasis. Our knowledge of this
regulatory system, however, is rather fragmentary and
although the basic mechanisms of transport have been
identified, the nature and regulation of many transporters
are only partially understood (Sanders and Bethke, 2000;
Serrano and Rodriguez-Navarro, 2001).
Ion homeostasis is particularly relevant for plant salt
tolerance, a limiting factor for agriculture in more than 40%
of irrigated land (Flowers and Yeo, 1988). ROS and H2O2
accumulate during salt stress, mostly because the associ-
ated desiccation perturbs electron transport chains in
membranes (Hasegawa et al., 2000; Hoekstra et al., 2001).
In addition to standard enzymatic and non-enzymatic anti-
oxidant defenses, the accumulation of compatible solutes
(osmolytes) protects against oxidative damage because
compounds such as mannitol, inositol and proline are
effective scavengers of hydroxyl radicals (Shen et al., 1997;
Smirnoff and Cumbes, 1989). However, the possibility exist
that the role of H2O2 during salt stress is not only that of a
toxic metabolite to be disposed but also that of a signaling
molecule for ion homeostasis.
Plant catalases form part of the major ROS scavenging
network. In Arabidopsis there are three genes, CAT1–3, all
encoding peroxisomal enzymes (McClung, 1997; Mittler
et al., 2004). Antisense plants with reduced catalase expres-
sion develop necrotic lesions under high light (Chamnong-
pol et al., 1996; Vandenabeele et al., 2004; Willekens et al.,
1997), are sensitive to oxidative and salt stress (Willekens
et al., 1997) and, under high light, express high levels of
pathogenesis-related, antioxidant, heat shock protein and
anthocyanin biosynthesis genes, among others (Chamnong-
pol et al., 1996; Vandenabeele et al., 2004; Vanderauwera
et al., 2005). The effect of catalase deficiency on ion
homeostasis, however, has not been investigated.
In the present work we have utilized a knock-out mutant of
the Arabidopsis CATALASE2 (CAT2, At4g35090) gene to
study the effects of a permanent increase in cellular H2O2 on
ion homeostasis. CAT2 is the most expressed gene in leaves;
it is induced by light and cold and regulated by the circadian
clock, with maximal expression in the early morning (McC-
lung, 1997). CAT2 expression is downregulated during leaf
senescence (Zimmermann et al., 2006). The cat2-1 mutant
exhibited altered monovalent cation homeostasis but the
most conspicuous phenotype was an unexpected lithium
tolerance despite increased lithium accumulation. Further
analysis demonstrated that lithium toxicity is caused by
increased ethylene production and that in the cat2-1 mutant
ethylene production and perception are both inhibited.
These results uncover a novel cross-talk between oxidative
stress, cation homeostasis and ethylene signaling.
Results and discussion
Molecular, biochemical and morphological
characterization of the cat2-1 mutant
The T-DNA of the cat2-1 mutant was inserted within the
fourth exon (total eight exons) after the first nucleotide of
codon 105 (total 492 codons) of the CAT2 (At4g35090) gene
and probably corresponds to a null mutation. Reverse
transcriptase-PCR analysis indicates that the level of
expression of CAT2 mRNA is undetectable (Figure 1).
CAT2 is a predominant catalase of Arabidopsis (McClung,
1997) and, accordingly, the cat2-1 mutant exhibited a great
reduction in catalase activity (Figure 2a). The enzymatic
activity of the mutant in leaves and roots was 21% and 47%,
respectively, of wild type activity, pointing to a greater
contribution of the CAT2 isoform in the aerial part. The
decrease in catalase activity of the cat2-1 mutant correlates
with a greater accumulation of H2O2 in leaves (Figure 2b).
Plants of the cat2-1 mutant grown under our normal
conditions of low light (60 lmol m)2 sec)1) have several
morphological alterations, such as reduced size (Figure 3a),
pale green color (Figure 3b) and reduced secondary roots
(Figure 3c). These results contrast with the absence of
visible phenotypes of tobacco (Chamnongpol et al., 1996)
and Arabidopsis (Vandenabeele et al., 2004) catalase anti-
sense plants grown under low light. Although the degree of
ACT8
CAT2
cat2-1 wt
cat2-1
1 972 2279
cat2(a)
(b)
f
cat2r
Figure 1. Molecular characterization of the cat2-1 mutant.
(a) Insertion of the T-DNA within the CAT2 gene. Blocks correspond to exons
and nucleotide positions are numbered. The position of the primers utilized
for PCR (cat2f and cat2r) is indicated.
(b) Reverse transcriptase-PCR analysis of the CAT2 and ACT8 mRNA in wild-
type Arabidopsis (wt) and in the cat2-1 mutant.
Lithium, ethylene and catalase mutant 1053
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065

reduction of catalase activity in some antisense lines was
similar to ours, the nature of the mutations (T-DNA insertion
versus antisense) may explain some differences in the
phenotypes. The T-DNA insertion of the cat2-1 mutant only
affects the CAT2 gene while antisense plants experience a
reduced expression of all three catalase genes (Mittler et al.,
2004).
Sensitivity of the cat2-1 mutant to abiotic stresses
The cat2-1 mutant was more sensitive than the wild type to
H2O2 (Figure 4a), NaCl (Figure 4b1,b2), relatively high light
(130 lmol m)2 sec)1) (Figure 4c) and cold (Figure 4d). The
first three phenotypes are similar to those of catalase anti-
sense plants previously described (Chamnongpol et al.,
1996; Vandenabeele et al., 2004) and corroborate the facts
that much of the damage caused by abiotic stresses is
mediated by H2O2 and that catalases play an important
protective role. The cold sensitivity of the cat2-1 mutant may
be related to the specific induction of the CAT2 gene by cold
stress, other catalase genes being repressed under these
conditions (Mittler et al., 2004).
All phenotypes of the cat2-1 mutant were complemented
by transformation with a CAT2 cDNA expressed from a
CaMV 35S promoter. Figure 3(c) shows the complementa-
tion of abundance of lateral roots and Figure 4(b1) demon-
strates the complementation of the sensitivity to NaCl.
0
150
300
450(a)
(b)Leaves Roots
Un
its/
g f
resh
wei
gh
t
Figure 2. Decreased catalase activity and increased H2O2 production in the
cat2-1 mutant.
(a) Catalase activity in extracts of wild-type (white bars) and cat2-1 (black bars)
leaves and roots. Values are the means of four independently performed
experiments and SE are indicated by vertical bars.
(b) Production of H2O2 in wild type (upper row) and cat2-1 (lower row) leaves
assayed with 3,3¢-diaminobenzidine. The experiment was repeated six times
with similar results.
wt
(a)
(b)
(c) wtcat2-1 35S:CAT2
cat2-1
cat2-1
wt
cat2-1
Figure 3. Morphological alterations of the cat2-1 mutant with respect to
control Arabidopsis (wt).
(a) Plants were grown on soil for 3 weeks and the reduced size of the mutant
can be observed.
(b) Seedlings were grown on plates for 2 weeks and the pale green color of the
mutant is apparent.
(c) Seedlings were grown on vertical plates for 2 weeks and the reduction of
secondary roots in the mutant is shown. The complementation of this root
phenotype in the mutant transformed with the CAT2 gene (cat2-1 35S:CAT2) is
also shown. All the experiments were repeated three times with similar
results.
1054 Eduardo Bueso et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065

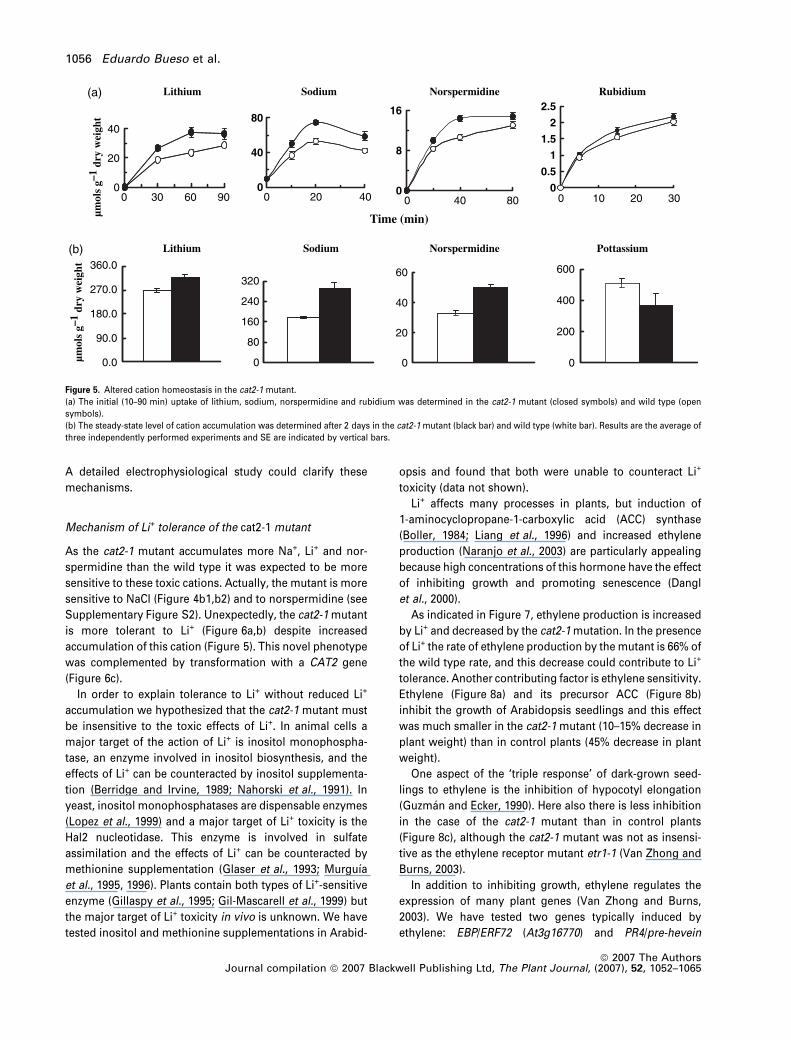
Altered cation homeostasis in the cat2-1 mutant
Seedlings of wild-type Arabidopsis and of the cat2-1 mutant
were incubated in liquid culture with different toxic cations
and the time course of uptake determined. The initial rates of
uptake (for the first 10–20 min) of Na+, Li+ and norspermi-
dine were 20–40% higher in the cat2-1 mutant than in the
wild type (Figure 5a) and the final levels accumulated after
2 days were also higher (15–60%) (Figure 5b).
The mechanism of uptake of toxic cations such as Na+ is
not well understood but it seems to involve non-selective
channels (Amtmann and Sanders, 1998; White, 1999),
including those of the cyclic nucleotide gated family
(Gobert et al., 2006). Initial rates of cation uptake by
seedlings will be mostly dependent on root uptake, while
cation accumulation at long times (days) must depend on
xylem loading and translocation to the shoot. The higher
levels of H2O2 in the cat2-1 mutant seem to stimulate both
processes.
The level of K+ in the cat2-1 mutant is 30% lower than in
wild-type plants (Figure 5b), pointing to an inhibition of K+
transport by H2O2. Similar results were obtained when
plants of wild-type Arabidopsis and of the cat2-1 mutant
were grown in soil and irrigated with 50 mM NaCl: the leaf
concentrations of Na+ were higher and those of K+ were
lower in the mutant than in the wild type (see Supplemen-
tary Figure S1).
The observed decrease in K+ accumulation by whole
plants of the cat2-1 mutant correlates with the situation of
guard cells during stomatal closure induced by H2O2 (Blatt
and Thiel, 1993; Foreman et al., 2003; Kwak et al., 2003;
Murata et al., 2001; Mustilli et al., 2002; Pei et al., 2000;
Schroeder and Hagiwara, 1989). Therefore, it is plausible
that a similar mechanism operates in the roots of cat2-1
plants: H2O2 activates Ca2+ uptake channels and elevated
cytosolic Ca2+ inhibits K+ uptake channels and activates
K+ efflux channels. This Ca2+signal may also activate
the non-specific channels utilized by toxic cations.
cat2-1 35S:CAT2 wt cat2-1
wt(a)
(b1)
(c) (d)
(b2)
cat2-1
cat2-1wt
cat2-1wt
cat2-1wt
Figure 4. Sensitivity of the cat2-1 mutant to several stresses. Wild-type Arabidopsis (wt) and the complemented mutant (cat2-1 35S:CAT2) are included as indicated.
(a) Seedlings growing on plates with 3 mM H2O2 for 10 days and the bleaching of mutant plants is apparent.
(b1) Seedlings growing on plates with 0.1 M NaCl for 2 weeks; the sensitivity of the mutant and its complementation are shown. (b2) Three-week-old plants grown in
soil irrigated with 0.1 M NaCl; the sensitivity of the mutant is apparent.
(c) Seedlings growing on plates under high light (130 lE m)2 sec)1) for 2 weeks, showing bleaching of the mutant.
(d) Three-week-old plants in soil incubated at )18�C for 30 min and further grown for 5 days, resulting in the death of mutant plants. All the experiments were
repeated three times with similar results.
Lithium, ethylene and catalase mutant 1055
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065
A detailed electrophysiological study could clarify these
mechanisms.
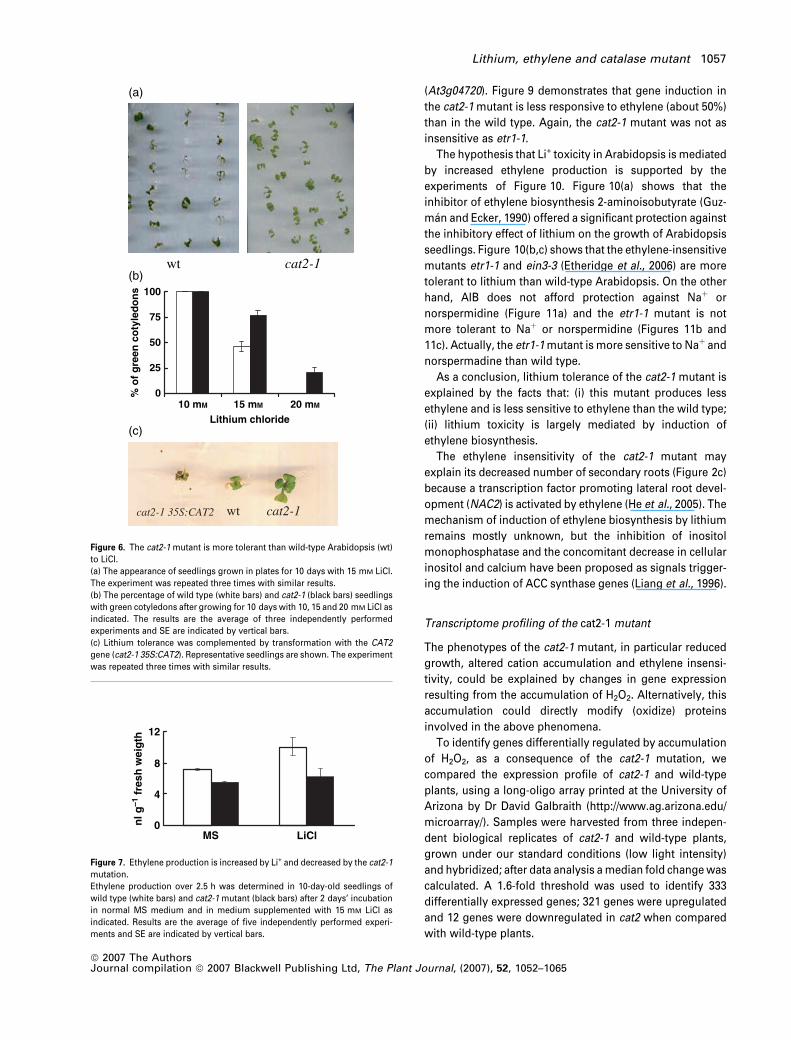
Mechanism of Li+ tolerance of the cat2-1 mutant
As the cat2-1 mutant accumulates more Na+, Li+ and nor-
spermidine than the wild type it was expected to be more
sensitive to these toxic cations. Actually, the mutant is more
sensitive to NaCl (Figure 4b1,b2) and to norspermidine (see
Supplementary Figure S2). Unexpectedly, the cat2-1 mutant
is more tolerant to Li+ (Figure 6a,b) despite increased
accumulation of this cation (Figure 5). This novel phenotype
was complemented by transformation with a CAT2 gene
(Figure 6c).
In order to explain tolerance to Li+ without reduced Li+
accumulation we hypothesized that the cat2-1 mutant must
be insensitive to the toxic effects of Li+. In animal cells a
major target of the action of Li+ is inositol monophospha-
tase, an enzyme involved in inositol biosynthesis, and the
effects of Li+ can be counteracted by inositol supplementa-
tion (Berridge and Irvine, 1989; Nahorski et al., 1991). In
yeast, inositol monophosphatases are dispensable enzymes
(Lopez et al., 1999) and a major target of Li+ toxicity is the
Hal2 nucleotidase. This enzyme is involved in sulfate
assimilation and the effects of Li+ can be counteracted by
methionine supplementation (Glaser et al., 1993; Murguıa
et al., 1995, 1996). Plants contain both types of Li+-sensitive
enzyme (Gillaspy et al., 1995; Gil-Mascarell et al., 1999) but
the major target of Li+ toxicity in vivo is unknown. We have
tested inositol and methionine supplementations in Arabid-
opsis and found that both were unable to counteract Li+
toxicity (data not shown).
Li+ affects many processes in plants, but induction of
1-aminocyclopropane-1-carboxylic acid (ACC) synthase
(Boller, 1984; Liang et al., 1996) and increased ethylene
production (Naranjo et al., 2003) are particularly appealing
because high concentrations of this hormone have the effect
of inhibiting growth and promoting senescence (Dangl
et al., 2000).
As indicated in Figure 7, ethylene production is increased
by Li+ and decreased by the cat2-1 mutation. In the presence
of Li+ the rate of ethylene production by the mutant is 66% of
the wild type rate, and this decrease could contribute to Li+
tolerance. Another contributing factor is ethylene sensitivity.
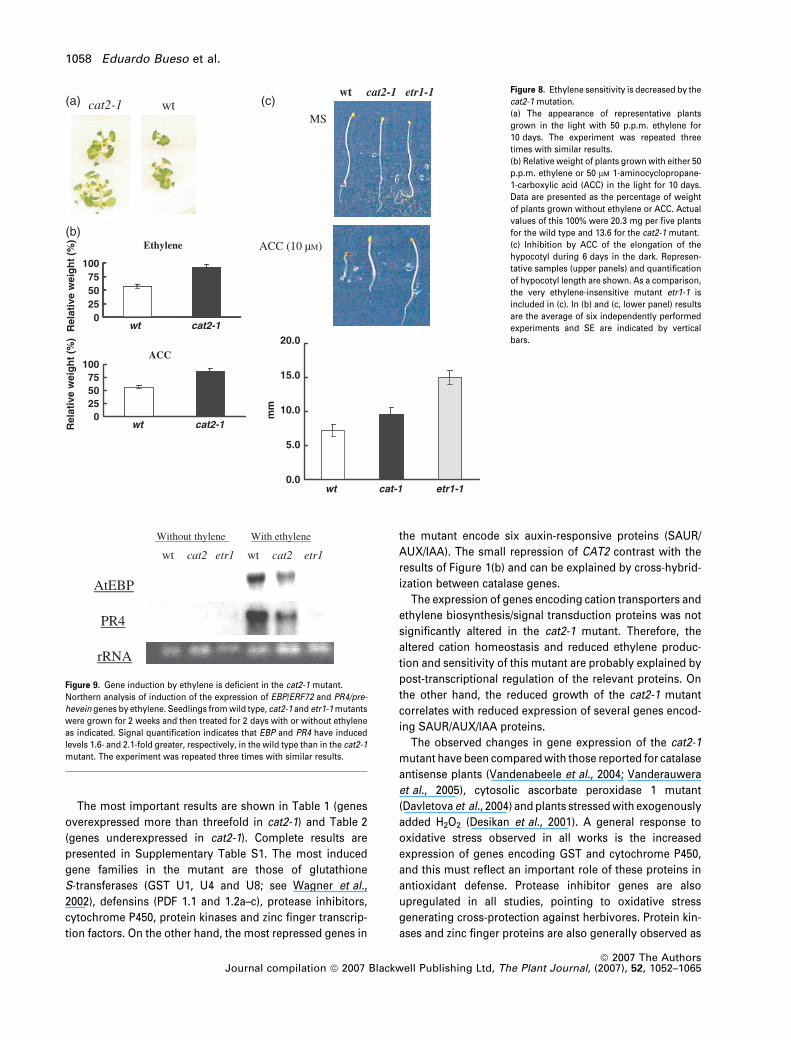
Ethylene (Figure 8a) and its precursor ACC (Figure 8b)
inhibit the growth of Arabidopsis seedlings and this effect
was much smaller in the cat2-1 mutant (10–15% decrease in
plant weight) than in control plants (45% decrease in plant
weight).
One aspect of the ‘triple response’ of dark-grown seed-
lings to ethylene is the inhibition of hypocotyl elongation
(Guzman and Ecker, 1990). Here also there is less inhibition
in the case of the cat2-1 mutant than in control plants
(Figure 8c), although the cat2-1 mutant was not as insensi-
tive as the ethylene receptor mutant etr1-1 (Van Zhong and
Burns, 2003).
In addition to inhibiting growth, ethylene regulates the
expression of many plant genes (Van Zhong and Burns,
2003). We have tested two genes typically induced by
ethylene: EBP/ERF72 (At3g16770) and PR4/pre-hevein
0
40
80
0 20 40
Sodium
0
8
16
0 40 80
NorspermidineLithium
00 30 60 90
20
40
(a)
(b)
µmol
s g–
1 dr
y w
eigh
t
0
0.5
1
2
2.5
0 10 20 30
1.5
Rubidium
Time (min)
Pottassium
0
200
400
600
Norspermidine
20
40
0
60
µmol
s g–
1 dr
y w
eigh
t
Lithium
360.0
270.0
180.0
90.0
0.0
Sodium
320
240
160
80
0
Figure 5. Altered cation homeostasis in the cat2-1 mutant.
(a) The initial (10–90 min) uptake of lithium, sodium, norspermidine and rubidium was determined in the cat2-1 mutant (closed symbols) and wild type (open
symbols).
(b) The steady-state level of cation accumulation was determined after 2 days in the cat2-1 mutant (black bar) and wild type (white bar). Results are the average of
three independently performed experiments and SE are indicated by vertical bars.
1056 Eduardo Bueso et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065
(At3g04720). Figure 9 demonstrates that gene induction in
the cat2-1 mutant is less responsive to ethylene (about 50%)
than in the wild type. Again, the cat2-1 mutant was not as
insensitive as etr1-1.
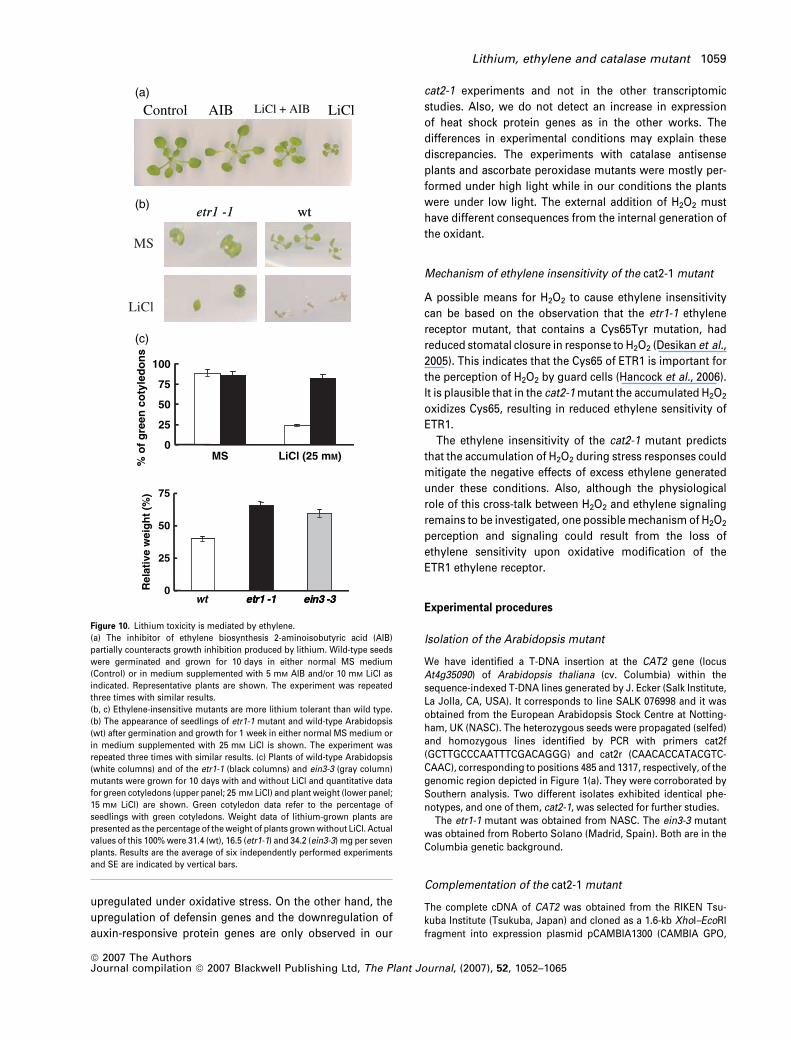
The hypothesis that Li+ toxicity in Arabidopsis is mediated
by increased ethylene production is supported by the
experiments of Figure 10. Figure 10(a) shows that the
inhibitor of ethylene biosynthesis 2-aminoisobutyrate (Guz-
man and Ecker, 1990) offered a significant protection against
the inhibitory effect of lithium on the growth of Arabidopsis
seedlings. Figure 10(b,c) shows that the ethylene-insensitive
mutants etr1-1 and ein3-3 (Etheridge et al., 2006) are more
tolerant to lithium than wild-type Arabidopsis. On the other
hand, AIB does not afford protection against Naþ or
norspermidine (Figure 11a) and the etr1-1 mutant is not
more tolerant to Naþ or norspermidine (Figures 11b and
11c). Actually, the etr1-1 mutant is more sensitive to Naþ and
norspermadine than wild type.
As a conclusion, lithium tolerance of the cat2-1 mutant is
explained by the facts that: (i) this mutant produces less
ethylene and is less sensitive to ethylene than the wild type;
(ii) lithium toxicity is largely mediated by induction of
ethylene biosynthesis.
The ethylene insensitivity of the cat2-1 mutant may
explain its decreased number of secondary roots (Figure 2c)
because a transcription factor promoting lateral root devel-
opment (NAC2) is activated by ethylene (He et al., 2005). The
mechanism of induction of ethylene biosynthesis by lithium
remains mostly unknown, but the inhibition of inositol
monophosphatase and the concomitant decrease in cellular
inositol and calcium have been proposed as signals trigger-
ing the induction of ACC synthase genes (Liang et al., 1996).
Transcriptome profiling of the cat2-1 mutant
The phenotypes of the cat2-1 mutant, in particular reduced
growth, altered cation accumulation and ethylene insensi-
tivity, could be explained by changes in gene expression
resulting from the accumulation of H2O2. Alternatively, this
accumulation could directly modify (oxidize) proteins
involved in the above phenomena.
To identify genes differentially regulated by accumulation
of H2O2, as a consequence of the cat2-1 mutation, we
compared the expression profile of cat2-1 and wild-type
plants, using a long-oligo array printed at the University of
Arizona by Dr David Galbraith (http://www.ag.arizona.edu/
microarray/). Samples were harvested from three indepen-
dent biological replicates of cat2-1 and wild-type plants,
grown under our standard conditions (low light intensity)
and hybridized; after data analysis a median fold change was
calculated. A 1.6-fold threshold was used to identify 333
differentially expressed genes; 321 genes were upregulated
and 12 genes were downregulated in cat2 when compared
with wild-type plants.
wt
(a)
(b)
(c)
cat2-1
0
25
50
75
100
10 mM 15 mM 20 mM Lithium chloride
% o
f g
reen
co
tyle
do
ns
cat2-1wtcat2-1 35S:CAT2
Figure 6. The cat2-1 mutant is more tolerant than wild-type Arabidopsis (wt)
to LiCl.
(a) The appearance of seedlings grown in plates for 10 days with 15 mM LiCl.
The experiment was repeated three times with similar results.
(b) The percentage of wild type (white bars) and cat2-1 (black bars) seedlings
with green cotyledons after growing for 10 days with 10, 15 and 20 mM LiCl as
indicated. The results are the average of three independently performed
experiments and SE are indicated by vertical bars.
(c) Lithium tolerance was complemented by transformation with the CAT2
gene (cat2-1 35S:CAT2). Representative seedlings are shown. The experiment
was repeated three times with similar results.
0
4
8
12
MS LiCl
nl g
–1 f
resh
wei
gth
Figure 7. Ethylene production is increased by Li+ and decreased by the cat2-1
mutation.
Ethylene production over 2.5 h was determined in 10-day-old seedlings of
wild type (white bars) and cat2-1 mutant (black bars) after 2 days’ incubation
in normal MS medium and in medium supplemented with 15 mM LiCl as
indicated. Results are the average of five independently performed experi-
ments and SE are indicated by vertical bars.
Lithium, ethylene and catalase mutant 1057
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065
The most important results are shown in Table 1 (genes
overexpressed more than threefold in cat2-1) and Table 2
(genes underexpressed in cat2-1). Complete results are
presented in Supplementary Table S1. The most induced
gene families in the mutant are those of glutathione
S-transferases (GST U1, U4 and U8; see Wagner et al.,
2002), defensins (PDF 1.1 and 1.2a–c), protease inhibitors,
cytochrome P450, protein kinases and zinc finger transcrip-
tion factors. On the other hand, the most repressed genes in
the mutant encode six auxin-responsive proteins (SAUR/
AUX/IAA). The small repression of CAT2 contrast with the
results of Figure 1(b) and can be explained by cross-hybrid-
ization between catalase genes.
The expression of genes encoding cation transporters and
ethylene biosynthesis/signal transduction proteins was not
significantly altered in the cat2-1 mutant. Therefore, the
altered cation homeostasis and reduced ethylene produc-
tion and sensitivity of this mutant are probably explained by
post-transcriptional regulation of the relevant proteins. On
the other hand, the reduced growth of the cat2-1 mutant
correlates with reduced expression of several genes encod-
ing SAUR/AUX/IAA proteins.
The observed changes in gene expression of the cat2-1
mutant have been compared with those reported for catalase
antisense plants (Vandenabeele et al., 2004; Vanderauwera
et al., 2005), cytosolic ascorbate peroxidase 1 mutant
(Davletova et al., 2004) and plants stressed with exogenously
added H2O2 (Desikan et al., 2001). A general response to
oxidative stress observed in all works is the increased
expression of genes encoding GST and cytochrome P450,
and this must reflect an important role of these proteins in
antioxidant defense. Protease inhibitor genes are also
upregulated in all studies, pointing to oxidative stress
generating cross-protection against herbivores. Protein kin-
ases and zinc finger proteins are also generally observed as
wtcat2-1
0255075
100
wt cat2-1Rel
ativ
e w
eig
ht
(%)
0255075
100
wt cat2-1Rel
ativ
e w
eig
ht
(%)
Ethylene
(a) (c)
(b)
ACC
MS
ACC (10 µM)
wt cat2-1 etr1-1
wt cat-1 etr1-1
mm
0.0
5.0
10.0
15.0
20.0
Figure 8. Ethylene sensitivity is decreased by the
cat2-1 mutation.
(a) The appearance of representative plants
grown in the light with 50 p.p.m. ethylene for
10 days. The experiment was repeated three
times with similar results.
(b) Relative weight of plants grown with either 50
p.p.m. ethylene or 50 lM 1-aminocyclopropane-
1-carboxylic acid (ACC) in the light for 10 days.
Data are presented as the percentage of weight
of plants grown without ethylene or ACC. Actual
values of this 100% were 20.3 mg per five plants
for the wild type and 13.6 for the cat2-1 mutant.
(c) Inhibition by ACC of the elongation of the
hypocotyl during 6 days in the dark. Represen-
tative samples (upper panels) and quantification
of hypocotyl length are shown. As a comparison,
the very ethylene-insensitive mutant etr1-1 is
included in (c). In (b) and (c, lower panel) results
are the average of six independently performed
experiments and SE are indicated by vertical
bars.
AtEBP
wt cat2 etr1
PR4
wt cat2 etr1
Without thylene With ethylene
rRNA
Figure 9. Gene induction by ethylene is deficient in the cat2-1 mutant.
Northern analysis of induction of the expression of EBP/ERF72 and PR4/pre-
hevein genes by ethylene. Seedlings from wild type, cat2-1 and etr1-1 mutants
were grown for 2 weeks and then treated for 2 days with or without ethylene
as indicated. Signal quantification indicates that EBP and PR4 have induced
levels 1.6- and 2.1-fold greater, respectively, in the wild type than in the cat2-1
mutant. The experiment was repeated three times with similar results.
1058 Eduardo Bueso et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065
upregulated under oxidative stress. On the other hand, the
upregulation of defensin genes and the downregulation of
auxin-responsive protein genes are only observed in our
cat2-1 experiments and not in the other transcriptomic
studies. Also, we do not detect an increase in expression
of heat shock protein genes as in the other works. The
differences in experimental conditions may explain these
discrepancies. The experiments with catalase antisense
plants and ascorbate peroxidase mutants were mostly per-
formed under high light while in our conditions the plants
were under low light. The external addition of H2O2 must
have different consequences from the internal generation of
the oxidant.
Mechanism of ethylene insensitivity of the cat2-1 mutant
A possible means for H2O2 to cause ethylene insensitivity
can be based on the observation that the etr1-1 ethylene
receptor mutant, that contains a Cys65Tyr mutation, had
reduced stomatal closure in response to H2O2 (Desikan et al.,
2005). This indicates that the Cys65 of ETR1 is important for
the perception of H2O2 by guard cells (Hancock et al., 2006).
It is plausible that in the cat2-1 mutant the accumulated H2O2
oxidizes Cys65, resulting in reduced ethylene sensitivity of
ETR1.
The ethylene insensitivity of the cat2-1 mutant predicts
that the accumulation of H2O2 during stress responses could
mitigate the negative effects of excess ethylene generated
under these conditions. Also, although the physiological
role of this cross-talk between H2O2 and ethylene signaling
remains to be investigated, one possible mechanism of H2O2
perception and signaling could result from the loss of
ethylene sensitivity upon oxidative modification of the
ETR1 ethylene receptor.
Experimental procedures
Isolation of the Arabidopsis mutant
We have identified a T-DNA insertion at the CAT2 gene (locusAt4g35090) of Arabidopsis thaliana (cv. Columbia) within thesequence-indexed T-DNA lines generated by J. Ecker (Salk Institute,La Jolla, CA, USA). It corresponds to line SALK 076998 and it wasobtained from the European Arabidopsis Stock Centre at Notting-ham, UK (NASC). The heterozygous seeds were propagated (selfed)and homozygous lines identified by PCR with primers cat2f(GCTTGCCCAATTTCGACAGGG) and cat2r (CAACACCATACGTC-CAAC), corresponding to positions 485 and 1317, respectively, of thegenomic region depicted in Figure 1(a). They were corroborated bySouthern analysis. Two different isolates exhibited identical phe-notypes, and one of them, cat2-1, was selected for further studies.
The etr1-1 mutant was obtained from NASC. The ein3-3 mutantwas obtained from Roberto Solano (Madrid, Spain). Both are in theColumbia genetic background.
Complementation of the cat2-1 mutant
The complete cDNA of CAT2 was obtained from the RIKEN Tsu-kuba Institute (Tsukuba, Japan) and cloned as a 1.6-kb XhoI–EcoRIfragment into expression plasmid pCAMBIA1300 (CAMBIA GPO,
Control (a)
(b)
(c)
AIB LiCl + AIB LiCl
etr1 -1 wt
MS
LiCl
etr1 -1 wt
0
25
50
75
100
MS LiCl (25 mM)
% o
f g
reen
co
tyle
do
ns
etr1 -1 ein3 -30
25
50
75
wt etr1 -1 ein3 -3
Rel
ativ
e w
eig
ht
(%)
Figure 10. Lithium toxicity is mediated by ethylene.
(a) The inhibitor of ethylene biosynthesis 2-aminoisobutyric acid (AIB)
partially counteracts growth inhibition produced by lithium. Wild-type seeds
were germinated and grown for 10 days in either normal MS medium
(Control) or in medium supplemented with 5 mM AIB and/or 10 mM LiCl as
indicated. Representative plants are shown. The experiment was repeated
three times with similar results.
(b, c) Ethylene-insensitive mutants are more lithium tolerant than wild type.
(b) The appearance of seedlings of etr1-1 mutant and wild-type Arabidopsis
(wt) after germination and growth for 1 week in either normal MS medium or
in medium supplemented with 25 mM LiCl is shown. The experiment was
repeated three times with similar results. (c) Plants of wild-type Arabidopsis
(white columns) and of the etr1-1 (black columns) and ein3-3 (gray column)
mutants were grown for 10 days with and without LiCl and quantitative data
for green cotyledons (upper panel; 25 mM LiCl) and plant weight (lower panel;
15 mM LiCl) are shown. Green cotyledon data refer to the percentage of
seedlings with green cotyledons. Weight data of lithium-grown plants are
presented as the percentage of the weight of plants grown without LiCl. Actual
values of this 100% were 31.4 (wt), 16.5 (etr1-1) and 34.2 (ein3-3) mg per seven
plants. Results are the average of six independently performed experiments
and SE are indicated by vertical bars.
Lithium, ethylene and catalase mutant 1059
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065

Table 1 Genes induced in the cat2-1mutant with respect to the wild type undernormal (low-light) conditions. Only geneswith ratios cat2/wt greater than or equal to3 are presented. Complete results aregiven in Supplementary Table S1
TAIR code Gene annotation Ratio (cat2/wt)
DefenseAt3g22600 Protease inhibitor 8.58At5g44420 Plant defensin (PDF1.2a) 8.03At2g29490 Glutathione S-transferase (GST U1) 7.83At1g75830 Plant defensin (PDF1.1) 6.51At2g43510 Trypsin inhibitor 5.81At1g45145 Thioredoxin H-type 5 5.55At4g12490 Protease inhibitor 5.23At2g29460 Glutathione S-transferase (GST U4) 5.17At5g44430 Plant defensin (PDF1.2c) 4.63At5g43580 Protease inhibitor 4.46At3g09940 Monodehydroascorbate reductase 4.3At2g26020 Plant defensin (PDF1.2b) 4.16At3g09270 Glutathione S-transferase (GST U8) 3.77At1g72900 Disease resistance protein (TIR-NBS class) 3.44At4g12500 Protease inhibitor 3.41At5g38900 DSBA oxidoreductase family protein 3.35At1g66090 Disease resistance protein (TIR-NBS class) 3.35At4g12480 Protease inhibitor 3.33At3g53980 Protease inhibitor 3.24At3g53980 Protease inhibitor 3.11At1g74590 Glutathione S-transferase (GST U10) 3.04At1g57630 Disease resistance protein (TIR class) 3.04
MetabolismAt2g29350 Tropinone reductase 8.53At4g34135 UDP-glucosyl transferase 5.45At5g20960 Aldehyde oxidase 1 (AAO1) 5.22At3g26830 Cytochrome P450 71B15 5.12At3g60140 Glycosyl hydrolase 4.63At3g54150 Embryo-abundant protein-related 4.58At1g61800 Glucose-6-phosphate/phosphate translocator 4.42At1g67980 Caffeoyl-CoA 3-O-methyltransferase 4.2At1g32350 Alternative oxidase 4.06At3g44300 Nitrilase 2 (NIT2) 4.05At2g15480 UDP-glucoronosyl/UDP-glucosyl transferase 3.48At1g60730 Aldo/keto reductase 3.45At4g37370 Cytochrome P450 3.34At4g13310 Cytochrome P450 71A20 3.31At1g33030 O-methyltransferase 3.22At2g24850 Aminotransferase 3.12At2g15390 Xyloglucan fucosyltransferase 3.1At2g30750 Cytochrome P450 71A12 3.08At3g22370 Alternative oxidase 1a, mitochondrial (AOX1A) 3.08At5g22860 Serine carboxypeptidase S28 3.02
CalciumAt3g01830 Calmodulin-related protein 4.31At5g48410 Glutamate receptor 3.62At1g21550 Calcium-binding protein 3.15At3g63380 Calcium-transporting ATPase 3.05
KinasesAt2g19190 Light-responsive receptor protein kinase 5.21At5g67080 Protein kinase 4.65At4g18250 Receptor serine/threonine kinase 4.17At4g23190 Protein kinase 3.59At1g79680 Wall-associated kinase 3.54At5g01540 Lectin protein kinase 3.22At3g59700 Lectin protein kinase 3.21At5g25930 Leucine-rich repeat family protein/protein kinase 3.16At4g04490 Protein kinase 3.1
Transcription factorsAt3g28210 Zinc finger (AN1-like) 4.07
1060 Eduardo Bueso et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065

Canberra, Australia) and transformed into Agrobacterium tum-efaciens strain C58C1. Wild type Arabidopsis (cv. Columbia) andthe cat2-1 mutant were transformed by A. tumefaciens infiltration(Bechtold et al., 1993) and hygromycin B-resistant plants wereselected. All the cat2-1 transformed plants recovered the wild-typemorphology and one line was selected as cat2-1 35S:CAT2. Mostwild-type transformed plants exhibited no phenotype, but a smallfraction (5–10%) resembled the original cat2-1 mutant. This can beexplained by co-suppression of the CAT2 gene.
Plant growth and stress treatments
Arabidopsis seedlings were grown in vitro on Murashige andSkoog (MS) medium (1% sucrose) under 16-h light (60 lE m)2
sec)1)/8-h dark conditions. Plates (90 mm) with solid agar mediumor Erlenmeyer flasks (100 ml) with 20-ml liquid medium wereemployed. For in vitro stress experiments, 3 mM H2O2, 100 mM
NaCl, 10–20 mM LiCl or 10–50 lM ACC were added as indicated.The relatively high-light stress involved 130 lE m)2 sec)1 with thesame photoperiod.
Arabidopsis plants were grown in pots containing a 1:3 vermic-ulite-soil mixture, also under 16-h light (60 lE m)2 sec)1)/8-h darkconditions and irrigated with water. For salt stress, 100 mM NaClwas included.
Measurement of catalase activity
Plants were grown for 3 weeks on liquid medium and leaves androots were separated for extraction as described by Moran et al.(1994). Catalase activity was measured in the extracts at 25�C with8 mM H2O2 by the initial rate of decrease of absorbance at 240 nm(Aebi, 1984). One unit is defined as 1 lmol min)1 referred to the FWof extracted tissue. Controls with boiled extracts had no significantactivity.
Table 1 (Continuted)TAIR code Gene annotation Ratio (cat2/wt)
At2g41835 Zinc finger (C2H2 type, AN1-like) 3.96At5g27420 Zinc finger (C3HC4-type RING finger) 3.84At4g11370 Zinc finger (C3HC4-type RING finger) 3.64At5g18270 No apical meristem (NAM) 3.29At3g04070 No apical meristem (NAM) 3.27At5g13080 WRKY family transcription factor 3.06
MiscellaneousAt1g26400 FAD-binding domain-containing protein 6.21At1g26380 FAD-binding domain-containing protein 6.04At5g62150 Peptidoglycan-binding LysM domain 5.88At4g18430 Ras-related GTP-binding protein 4.95At2g26400 Acireductone dioxygenase (ARD/ARD¢) 4.3At2g26560 Patatin 4.11At4g15610 Integral membrane family protein 3.8At1g26420 FAD-binding domain-containing protein 3.73At4g20860 FAD-binding domain 3.7At3g26500 Leucine-rich repeat family protein 3.56At1g66700 S-adenosyl-L-methionine:carboxyl methyltransferase 3.52At5g66640 LIM domain-containing protein-related 3.43At1g26390 FAD-binding domain-containing protein 3.33At4g23700 Cation/hydrogen exchanger, putative (CHX17) 3.24At1g03850 Glutaredoxin family protein 3.22At2g04100 MATE efflux family protein 3.14At5g54710 Ankyrin repeat family protein 3.08At2g02990 Ribonuclease 1 (RNS1) 3.06At4g01870 TolB protein-related 3.06At3g04720 Hevein-like protein (HEL) 3.06
UnknownAt1g36640 Expressed protein 6.72At4g08555 Expressed protein 6.32At4g37290 Expressed protein 5At2g18680 Expressed protein 4.55At2g14777 Hypothetical protein 4.53At5g22545 Expressed protein 4.24At2g40095 Expressed protein 4.01At1g10585 Expressed protein 3.98At3g10320 Expressed protein 3.93At3g13950 Expressed protein 3.92At2g32190 Expressed protein 3.85At4g39670 Expressed protein 3.82At2g23270 Expressed protein 3.23
Lithium, ethylene and catalase mutant 1061
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065
Detection of H2O2
Plants were grown on soil for 3 weeks and detection of H2O2 wasperformed by infiltrating excised leaves over 8 h with a solution of1 mg ml)1 3,3¢-diaminobenzidine and processed as described(Orozco-Cardenas and Ryan, 1999).
Determination of cation transport and accumulation
Arabidopsis plants were grown in liquid culture for 3 weeks andtransferred to fresh media containing 2.4 mM norspermidine,20 mM LiCl or 120 mM NaCl as indicated. Plants were sampledimmediately after transfer (zero time control) and at different
incubation times, washed with a cold solution of 20 mM MgCl2 andthen with cold water.
In the case of inorganic cations, the washed tissue was dried at65�C for 2 days, weighed and extracted with 0.1 M nitric acid (1 ml/10 mg DW) for 30 min. Cell debris was removed by filtration andLi+, Na+ and K+ were determined by atomic absorption spectropho-tometry in a Varian SpectrAA-10 Plus spectrometer (http://www.varianinc.com/).
For norspermidine determination the washed tissue was weighed,frozen in liquid nitrogen, ground in a mortar and extracted with cold5% perchloric acid (1 ml/100 mg FW). Analysis by HPLC of benzoy-lated polyamines was as described by Flores and Galston (1982).
Measurement of ethylene production
Seedlings were grown for 2 weeks on plates with normal MSmedium and then transferred to fresh normal medium or mediumcontaining 15 mM LiCl for 24 h. Plants were incubated at room
AIB AIB + Na NaControl
IB + Ne NeIB + Ne NeAIB + Ne Ne
MS
NaCl
Ne
wt etr1-1
0
25
50
75(c)
(b)
(a)
MS Ne 2 mM Na 100 mM
% o
f g
erm
inat
ion
AIBControl
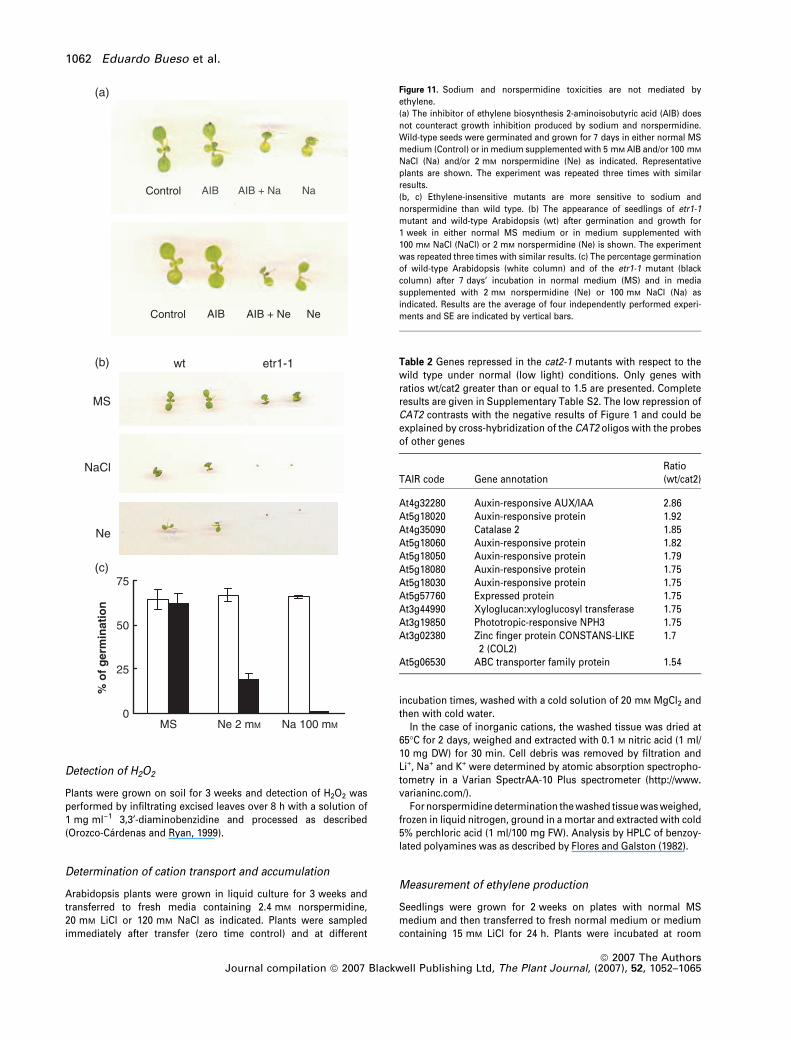
Figure 11. Sodium and norspermidine toxicities are not mediated by
ethylene.
(a) The inhibitor of ethylene biosynthesis 2-aminoisobutyric acid (AIB) does
not counteract growth inhibition produced by sodium and norspermidine.
Wild-type seeds were germinated and grown for 7 days in either normal MS
medium (Control) or in medium supplemented with 5 mM AIB and/or 100 mM
NaCl (Na) and/or 2 mM norspermidine (Ne) as indicated. Representative
plants are shown. The experiment was repeated three times with similar
results.
(b, c) Ethylene-insensitive mutants are more sensitive to sodium and
norspermidine than wild type. (b) The appearance of seedlings of etr1-1
mutant and wild-type Arabidopsis (wt) after germination and growth for
1 week in either normal MS medium or in medium supplemented with
100 mM NaCl (NaCl) or 2 mM norspermidine (Ne) is shown. The experiment
was repeated three times with similar results. (c) The percentage germination
of wild-type Arabidopsis (white column) and of the etr1-1 mutant (black
column) after 7 days’ incubation in normal medium (MS) and in media
supplemented with 2 mM norspermidine (Ne) or 100 mM NaCl (Na) as
indicated. Results are the average of four independently performed experi-
ments and SE are indicated by vertical bars.
Table 2 Genes repressed in the cat2-1 mutants with respect to thewild type under normal (low light) conditions. Only genes withratios wt/cat2 greater than or equal to 1.5 are presented. Completeresults are given in Supplementary Table S2. The low repression ofCAT2 contrasts with the negative results of Figure 1 and could beexplained by cross-hybridization of the CAT2 oligos with the probesof other genes
TAIR code Gene annotationRatio(wt/cat2)
At4g32280 Auxin-responsive AUX/IAA 2.86At5g18020 Auxin-responsive protein 1.92At4g35090 Catalase 2 1.85At5g18060 Auxin-responsive protein 1.82At5g18050 Auxin-responsive protein 1.79At5g18080 Auxin-responsive protein 1.75At5g18030 Auxin-responsive protein 1.75At5g57760 Expressed protein 1.75At3g44990 Xyloglucan:xyloglucosyl transferase 1.75At3g19850 Phototropic-responsive NPH3 1.75At3g02380 Zinc finger protein CONSTANS-LIKE
2 (COL2)1.7
At5g06530 ABC transporter family protein 1.54
1062 Eduardo Bueso et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065

temperature in closed vials for 2.5 h and ethylene determined bygas chromatography as described (Naranjo et al., 2003).
RNA analysis
Total RNA was extracted and Northern blot analysis performed asdescribed (Naranjo et al., 2003). Probes for the PR4 and EBP geneswere synthesized by random-priming from templates prepared byPCR amplification of Arabidopsis DNA with gene-specific primers.The sequences of the primers were the following: for PR4(At3g04720) forward CGAACTTGTCCCGGTAACAT and reverseGCCGCAAGAAGCTTGACCAC; for EBP (At3g16770) forwardGGAGGCAGTGAAGAAGC and reverse CCACCTCGGAAACGG-ACTC. Signal quantification was carried out using a Fujifilm BAS-1500 phosphorimager (http://www.fujifilm.com/).
Reverse transcriptase-PCR was as described (Naranjo et al., 2003)with 30 cycles of amplification. Primers for CAT2 were as describedabove and those for ACT8 (At1g49240) were the following: forwardAGTGGTCGTACAACCGGTATTGT and reverse GAGGATAGCAT-GTGGAAGTGAGAA.
Gene expression using long oligonucleotide microarrays
Long oligonucleotide microarrays were provided by Dr DavidGalbraith (the University of Arizona, http://www.ag.arizona.edu/microarray/). Oligonucleotides were the Operon ArabidopsisGenome Oligo Set Version 3.0 (Operon, https://www.operon.com/),which contains 29 110 70-mers probes, represents 26 173 protein-coding genes, 28 964 protein-coding gene transcripts and 87miRNAs. The design is based on the ATH1 release 5.0 of the TIGRArabidopsis genome annotation database (http://www.tigr.org/tdb/e2k1/ath1/) and release 4.0 of the miRNA Registry at theSanger Institute (http://www.sanger.ac.uk/Software/Rfam/mirna/index.shtml).
Oligos were printed onto aminosilane-coated slides (Telechem,http://www.arrayit.com/). After printing, the slides were baked for1 h at 80�C and stored dry in the dark at room temperature. Justbefore use, DNA was rehydrated by placing the slide upside downover a water bath at 60�C for 10 sec, and snap drying onto a heatblock at 65�C. Rehydration was repeated three times. DNA wasimmobilized by UV light by exposing the slides DNA-side upto 65 mJ in a UV cross-linker (Stratagene, http://www.stratagene.com/). Slides were then washed twice in 0.1% SDS and four times inwater, 2 min each wash at room temperature. Slides were thendipped in 96% ethanol for 1 min and dried by centrifugation at1500 g for 1 min at room temperature. Slides were pre-hybridizedfor 30 min at 42�C with 100 ll of pre-hybridization solution [6 · SSC(Sigma, http://www.sigmaaldrich.com/), 1% BSA (Sigma) and 0.5%SDS (Sigma)], under a 60 · 22 mm coverslip LifterSlip (Erie Scien-tific, http://www.eriesci.com/) in a microarray hybridization cham-ber (ArrayIt Hybridization Cassette, TeleChem). Slides were thenrinsed five times in H2O in a slide rack at room temperature anddried by centrifugation at 1500 g for 1 min. Slides were hybridizedimmediately.
Total RNA was extracted from wild-type parental Col-0 andcat2-1 mutant plants, and used in an amplification reaction usingthe MessageAmp II aRNA Amplification kit (Ambion, http://www.ambion.com/), basically as described by the manufacturer.5-(3-aminoallyl)-UTP (aa-UTP) (Ambion), was incorporated intothe amplified RNA (aRNA) in the amplification reaction andat ratio 1:1 with UTP. After purification 7.5 lg of aminoallyl-aRNA(aa-aRNA) was used in a coupling eaction with either fluores-cent Cy3 or Cy5 as reactive N-hydroxyl succinimidal dyes
(NHS-dyes) (Amersham Pharmacia Biotech, http://www5.amershambiosciences.com/). For this, each aliquot of Cy3 or Cy5was dissolved in 20 ll of DMSO and used in four couplingreactions. aa-aRNA was dissolved in 15 ll of freshly made 0.1 M
NaCO3 pH 9.0, and mixed with 5 ll of either Cy3 or Cy5 dye.Coupling was allowed for 1 h at 25�C in the dark. The reaction wasstopped with 35 ll of 100 mM NaOAc pH 5.2 and incubated for5 min in the dark. Fluorescent aa-aRNA was purified using aMegaClear purification column (Ambion) as described by themanufacturer. Coupled aa-aRNA (250–300 pmol Cy dye in 5–7 lgaa-aRNA) was dried in a SpeedVac and dissolved in 2.5 ll offiltered water. Both coupled aa-aRNAs (cy3 and Cy5) were mixedand fragmented in the presence of 20 lg of tRNA (Ambion) and20 lg of poly(A) (Roche, http://www.roche.com/) using the RNAfragmentation Reagent (Ambion) as described by the manufac-turer, in a final volume of 10 ll. Fragmented aa-aRNA was useddirectly in the hybridization mix, which contains 50 ll deionizedformamide (Sigma), 30 ll 20 · SSC, 5 ll 100 · Denhardt’s solu-tion (Sigma) and 5 ll 10% SDS in a final volume of 100 ll. Thehybridization mixture was denatured at 95�C for 5 min, spunbriefly and applied by capillary between a pre-treated slide (seeabove) and a 60 · 42 mm coverslip LifterSlip (Erie Scientific).Slides were incubated overnight at 37�C in a microarray hybrid-ization chamber (ArrayIt Hybridization Cassette, TeleChem). Thenext morning, slides were washed sequentially once in 1 · SSC0.1% SDS 5 min at 30�C; once in 0.2 · SSC 0.1% SDS 5 min at30�C; twice in 0.1 · SSC 2 min each at 30�C; and finally 0.01 ·SSC 10 sec at 25�C. Slides were dried by centrifugation at 1500 g
1 min at room temperature.Hybridized microarray slides were scanned at 532 nm for Cy3 and
635 nm for Cy5, with a GenePix 4000B scanner (Axon MolecularDevices, http://www.moleculardevices.com), at 10 nm resolutionand 100% laser power. Photomultiplier tube voltages were adjustedmanually to equal the overall signal intensity for each channel, toincrease the signal- to-noise ratio, and to reduce the number ofspots with saturated pixels. Spot intensities were quantified usingGENEPIX PRO 6.0 microarray-analysis software (Axon MolecularDevices). Data were normalized by mean global intensity and withlowess (locally weighted scatter plot) correction (Yang et al., 2001)with GENEPIX PRO 6.0 and ACUITY 4.0 software (Axon MolecularDevices), respectively.
Three biological replicates were obtained for each control paren-tal Columbia and cat2-1 mutant plants. These were sets of mutantand parental plants grown in parallel as described above (16-h lightand 8-h night; 22�C in growth chambers), but consecutively in time.Samples were harvested at the same time in the photoperiod toavoid differences in diurnal or circadian regulation of gene expres-sion. Two pairs of samples (cat2-1 and Col) were labeled Cy5 for Coland Cy3 for cat2-1 plants, while the third pair was reversed (dye-swap). After image analysis, spots with a net intensity in bothchannels lower than twice the median signal background wereremoved as low-signal spots, and only probes for which weobtained valid data in the three slides were considered for furtheranalysis (12 871 spots).
To identify genes based on fold-change, we select those geneswhose spots showed a log2 ratio >0.8 or <)0.8 in at least two slides,and only genes with a mean log2 ratio in the three replicates greateror less that >0.68 or <)0.68 (1.6-fold difference) were selected (361spots). Finally, spots with inconsistent data (i.e. genes whose spotsshowed positive and negative values of the log2 ratio) wereremoved (12 spots removed). With these criteria, a final data setof 349 probes, corresponding to 321 upregulated (337 probes) and12 downregulated (12 probes) genes, was selected as differentiallyexpressed between cat2 and Columbia plants.
Lithium, ethylene and catalase mutant 1063
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065

Acknowledgements
This work was supported by grant BFU2005-06388-C04-01 of theSpanish ‘Ministerio de Educacion y Ciencia’, Madrid.
Supplementary Material
The following supplementary material is available for this articleonline:Figure S1. Na+ and K+ levels in plants grown in soil for 3 weeksirrigated with water.Figure S2. The cat2-1 mutant is more sensitive to norspermidinethan the wild type.Table S1. Gene expression data from DNA microarray analysis of catand wt plants.This material is available as part of the online article from http://www.blackwell-synergy.comPlease note: Blackwell Publishing are not responsible for the contentor functionality of any supplementary materials supplied by theauthors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
References
Aebi, H. (1984) Catalase in vitro. Methods Enzymol. 105, 121–126.Allen, R.D. (1995) Dissection of oxidative stress tolerance using
transgenic plants. Plant Physiol. 107, 1049–1054.Amtmann, A. and Sanders, D. (1998) Mechanism of Na+ uptake by
plant cells. Adv. Bot. Res. 29, 75–112.Apel, K. and Hirt, H. (2004) Reactive oxygen species: metabolism,
oxidative stress, and signal transduction. Annu. Rev. Plant Biol.55, 373–399.
Bechtold, N., Ellis, F. and Pelletier, G. (1993) In planta Agrobacte-rium mediated gene transfer by infiltration of adult Arabidopsisthaliana plants. C. R. Acad. Sci. Paris, Life Sci. 316, 1194–1199.
Berridge, M.J. and Irvine, R.F. (1989) Inositol phosphates and cellsignaling. Nature, 341, 197–205.
Blatt, M.R. and Thiel, G. (1993) Hormonal control of ion channelgating. Annu. Rev. Plant Physiol. Plant Mol. Biol. 44, 543–567.
Boller, T. (1984) Superinduction of ACC synthase in tomato pericarpby lithium ions. In Ethylene: Biochemical, Physiological and Ap-plied Aspects (Fuchs, Y. and Chalutz, E., eds). The Hague: Nijhoff/Junk, pp. 87–88.
Chamnongpol, S., Willekens, H., Langebartels, C., Van Montagu,
M., Inze, D. and Van Camp, W. (1996) Transgenic tobacco with areduced catalase activity develops necrotic lesions and inducespathogenesis-related expression under high light. Plant J. 10,491–503.
Dangl, J.L., Dietrich, R.A. and Thomas, H. (2000) Senescence andprogrammed cell death. In Biochemistry and Molecular Biologyof Plants (Buchanan, B., Gruissem, W. and Jones, R., eds).Maryland: American Society of Plant Physiologists, Rockville, pp.1044–1100.
Davletova, S., Rizhsky, L., Liang, H., Shengqiang, Z., Oliver, D.J.,
Coutu, J., Shulaev, V., Schlauch, K. and Mittler, R. (2004) Cyto-solic ascorbate peroxidase 1 is a central component of the reac-tive oxygen gene network of Arabidopsis. Plant Cell, 17, 268–281.
Desikan, R., A.-H.-Mackerness, S., Hancock, J.T. and Neill, S.J.
(2001) Regulation of the Arabidopsis transcriptome by oxidativestress. Plant Physiol. 127, 159–172.
Desikan, R., Hancock, J.T., Bright, J., Harrison, J., Weir, I., Hooley, T.
and Neill, S.J. (2005) A role for ETR1 in hydrogen peroxidesignaling in stomatal guard cells. Plant Physiol. 137, 831–834.
Etheridge, N., Hall, B.P. and Schaller, G.E. (2006) Progress report:ethylene signaling and responses. Planta, 223, 387–391.
Flores, H.E. and Galston, A.W. (1982) Analisis of polyamines inhigher plants by high performance liquid chromatography. PlantPhysiol. 69, 701–706.
Flowers, T.J. and Yeo, A.R. (1988) Ion relations of salt tolerance. InSolute Transport in Plant Cells and Tissues (Baker, D.A. and Hall,J.L., eds). New York: Longman, pp. 392–416.
Foreman, J., Demidchik, V., Bothwell, J.H.F. et al. (2003) Reactiveoxygen species produced by NADPH oxidase regulate plant cellgrowth. Nature, 422, 442–446.
Gillaspy, G.E., Keddie, J.S., Oda, K. and Grissem, W. (1995) Plantinositol monophosphatase is a lithium-sensitive enzyme encodedby a multigene family. Plant Cell, 7, 2175–2185.
Gil-Mascarell, R., Lopez-Coronado, J.M., Belles, J.M., Serrano, R.
and Rodriguez, P.L. (1999) The Arabidopsis HAL2-like gene familyincludes a novel sodium-sensitive phosphatase. Plant J. 17,373–383.
Glaser, H.-U., Thomas, D., Gaxiola, R., Montrichard, F., Surdin-
Kerjan, Y. and Serrano, R. (1993) Salt tolerance and methioninebiosynthesis in Saccharomyces cerevisiae involve a putativephosphatase gene. EMBO J. 12, 3105–3110.
Gobert, A., Park, G., Amtmann, A., Sanders, D. and Maathuis, F.J.
(2006) Arabidopsis thaliana cyclic nucleotide gated channel 3forms a non-selective ion transporter involved in germination andcation transport. J. Exp. Bot. 57, 791–800.
Guzman, P. and Ecker, J.R. (1990) Exploiting the triple response ofArabidopsis to identify ethylene-related mutants. Plant Cell 2,513–523.
Hancock, J., Desikan, R., Harrison, J., Bright, J., Hooley, R and Neill,
S. (2006) Doing the unexpected: proteins involved in hydrogenperoxide perception. J. Exp. Bot. 57, 1711–1718.
Hasegawa, P.M., Bressan, R.A., Zhu, J.-K. and Bohnert, H.J. (2000)Plant cellular and molecular responses to high salinity. Annu.Rev. Plant Physiol. Plant Mol. Biol. 51, 463–499.
He, X.J., Mu, R.L., Cao, W.H., Zhang, Z.G., Zhang, J.S. and Chen,
S.Y. (2005) AtNAC2, a transcription factor downstream ofethylene and auxin signaling pathways, is involved in saltstress response and lateral root development. Plant J. 44, 903–9916.
Hedrich, R., Busch, H. and Raschke, K. (1990) Ca2 + and nucleotidedependent regulation of voltage dependent anion channels in theplasma membrane of guard cells. EMBO J. 9, 3889–3892.
Hoekstra, F.A., Golovina, E.A. and Buitink, J. (2001) Mechanisms ofplant desiccation tolerance. Trends Plant Sci. 6, 431–438.
Joo, J.H., Bae, Y.S. and Lee, J.S. (2001) Role of auxin-inducedreactive oxygen species in root gravitropism. Plant Physiol. 126,1055–1060.
Kwak, J.M., Mori, I.C., Pei, Z.M., Leonhardt, N., Torres, M.A., Dangl,
J.L., Bloom, R.E., Bodde, S., Jones, J.D. and Schroeder, J.I. (2003)NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 22, 2623–2633.
Laloi, C., Apel, K. and Danon, A. (2004) Reactive oxygen signalling:the latest news. Curr. Opin. Plant Biol. 7, 323–328.
Liang, X.., Shen, N.F. and Theologis, A. (1996) Li+-regulated1-aminocyclopropane- 1-carboxylate synthase gene expressionin Arabidopsis thaliana. Plant J. 10, 1027–1036.
Lopez, F., Leube, M., Gil-Mascarell, R., Navarro-Avino, J.P. and
Serrano, R. (1999) The yeast inositol monophosphatase is a lith-ium- and sodium-sensitive enzyme encoded by a non-essentialgene pair. Mol. Microbiol. 31, 1255–1264.
McClung, C.R. (1997) Regulation of catalases in Arabidopsis. FreeRedical Biol. Med. 23, 489–496.
1064 Eduardo Bueso et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065

Mittler, R., Vanderauwera, S., Gollery, M. and Van Breusegem, F.
(2004) Reactive oxygen gene networks of plants. Trends Plant Sci.10, 490–498.
Moran, J.F., Becana, M., Iturbe-Ormaetxe, I., Frechilla, S., Klucas,
R.V. and Pedro Aparicio-Tejo, P. (1994) Drought induces oxidativestress in pea plants. Planta, 194, 346–352.
Murata, Y., Pei, Z.M., Mori, I.C. and Schroeder, J.I. (2001) Abscisicacid activation of plasma membrane Ca2+ channels in guard cellsrequires cytosolic NAD(P)H and is differentially disruptedupstream and downstream of reactive oxygen species productionin abi1-1 and abi2-1 protein phosphatase 2C mutants. Plant Cell,13, 2513–2523.
Murguıa, J.R., Belles, J.M. and Serrano, R. (1995) ) A salt-sensitive3¢(2¢),5¢-bisphosphate nucleotidase involved in sulfate activation.Science, 267, 232–234.
Murguıa, J.R., Belles, J.M. and Serrano, R. (1996) The yeast HAL2nucleotidase is an in vivo target of salt toxicity. J. Biol. Chem. 271,29029–29033.
Mustilli, A.C., Merlot, S., Vavasseur, A., Fenzi, F. and Giraudat, J.
(2002) Arabidopsis OST1 protein kinase mediates the regulationof stomatal aperture by abscisic acid and acts upstream of reac-tive oxygen species production. Plant Cell, 14, 3089–3099.
Nahorski, S.R., Ragan, C.I. and Challiss, R.A.J. (1991) Lithium andthe phosphooinositide cycle: an example of uncompetitive inhi-bition and its pharmacological consequences. Trends Pharmacol.Sci. 12, 297–303.
Naranjo, M.A., Romero, C., Belles, J.M., Montesinos, C., Vicente, O.
and Serrano, R. (2003) Lithium treatment induces a hypersensi-tive-like response in tobacco. Planta, 217, 417–424.
Neill, S., Desikan, R. and Hancock, J. (2002) Hydrogen peroxidesignallling. Curr. Opin. Plant Biol. 5, 388–395.
Orozco-Cardenas, M. and Ryan, C.A. (1999) Hydrogen peroxide isgenerated systemically in plant leaves by wounding and systeminvia the octadecanoid pathway. Proc. Natl. Acad. Sci. U.S.A. 96,6553–6557.
Pei, Z.M., Murata, Y., Benning, G., Thomine, S., Klusener, B., Allen,
G.J., Grill, E. and Schroeder, J.I. (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signaling inguard cells. Nature, 406, 731–734.
Sanders, D. and Bethke, P. (2000) Membrane transport. In Bio-chemistry and Molecular Biology of Plants (Buchanan, B.B.,Gruissem, W. and Jones, R.L. eds). Maryland: American Societyof Plant Physiology, Rockville, Chapter 3, pp.110–158.
Schroeder, J.I. and Hagiwara, S. (1989) Cytosolic calcium regulatesion channels in the plasma membrane of Vicia faba guard cells.Nature, 338, 427–430.
Serrano, R. and Rodriguez-Navarro, A. (2001) Ion homeostasis dur-ing salt stress in plants. Curr. Opin. Cell Biol. 13, 399–404.
Shen, B., Jensen, R.G. and Bohnert, H.J. (1997) Mannitol protectsagainst oxidation by hydroxyl radicals. Plant Physiol. 115,527–532.
Shin, R. and Schachtman, D.P. (2004) Hydrogen peroxide mediatesplant root cell response to nutrient deprivation. Proc. Natl. Acad.Sci. U.S.A. 101, 8827–88332.
Smirnoff, N. and Cumbes, Q.J. (1989) Hydroxyl radical scaveng-ing activity of compatible solutes. Phytochemistry, 28, 1057–1060.
Van Zhong, G. and Burns, J.K. (2003) Profiling ethylene-regulatedgene expression in Arabidopsis thaliana by microarray analysis.Plant Mol. Biol. 53, 117–131.
Vandenabeele, S., Vanderauwera, S., Vuylsteke, M., Rombauts, S.,
Langebartels, C., Seidlitz, H.K., Zabeau, M., Van Montagu, M.,
Inze, D. and Van Breusegem, F. (2004) Catalase deficiency dras-tically affects gene expression induced by high light in Arabid-opsis thaliana. Plant J. 39, 45–58.
Vanderauwera, S., Zimmermann, P., Rombauts, S., Vandenabeele,
S., Langebartels, C., Gruissem, W., Inze, D. and van Breusegem, F.
(2005) Genome- wide analysis of hydrogen peroxide-regulatedgene expression in Arabidopsis reveals a high light-inducedtranscriptional cluster involved in anthocyanin biosynthesis.Plant Physiol. 139, 806–821.
Wagner, U., Edwards, R., Dixon, D.P. and Mauch, F. (2002) Probingthe diversity of the Arabidopsis glutathione S-transferase genefamily. Plant Mol. Biol. 49, 515–5532.
White, P.J. (1999) The molecular mechanism of sodium influx toroot cells. Trends Plant Sci. 4, 245–246.
Willekens, H., Chamnongpol, S., Davey, M., Schraudner, M.,
Langebartels, C., Van Montagu, M., Inze, D. and Van Camp, W.
(1997) Catalase is a sink for H2O2 and is indispensable for stressdefence ion C3 plants. EMBO J. 16, 4806–4816.
Yang, Y.H., Dudoit, S., Luu, P. and Speed, T.P. (2001) Normalizationfor cDNA microarray. In Microarrays: Optical Technologies andInformatics (Bittner, M.L., Chen, Y., Dorsel, A.N. and Dougherty,E.R., eds). CA: SPIE, Society for Optical Engineering, San Jose,pp. 141–152.
Zimmermann, P., Heinlein, C., Orendi, G. and Zentgraf, U. (2006)Senescence-specific regulation of catalases in Arabidopsis thali-ana (L.) Heynh. Plant Cell Environ. 29, 1049–1060.
Lithium, ethylene and catalase mutant 1065
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 1052–1065
Related Documents











