The LIM domain protein Wtip interacts with the receptor tyrosine kinase Ror2 and inhibits canonical Wnt signalling Nicole Verhey van Wijk a,b,1 , Florian Witte a,b , Ann Carolin Feike c , Alexandra Schambony c , Walter Birchmeier d , Stefan Mundlos a,b , Sigmar Stricker a,b, * a Max Planck-Institute for Molecular Genetics, Development and Disease Group, Berlin, Germany b Institute for Medical Genetics, University Medicine Charité, Berlin, Germany c Developmental Biology Unit, Biology Department, University of Erlangen–Nuremberg, Erlangen, Germany d Max Delbrück Center for Molecular Medicine, Berlin, Germany article info Article history: Received 17 September 2009 Available online 26 September 2009 Keywords: Wtip Ror2 Brachydactyly Wnt abstract Wtip is a LIM domain protein of the Ajuba/Zyxin family involved in kidney and neural crest development; Ror2 is a receptor tyrosine kinase involved in the development of skeleton, heart, lung, genitalia and kid- neys. Here we describe Wtip as an intracellular interaction partner of Ror2. Full-length Ror2 recruits Wtip to the cell membrane, a mutant involved in human disease fails to do so. Both genes and proteins show overlapping expression in the mouse embryo. We show that Wtip is able to inhibit canonical Wnt signal- ling in mammalian cells and in Xenopus embryos linking Wtip to a crucial developmental pathway. Ó 2009 Elsevier Inc. All rights reserved. Introduction Ror2 is a receptor tyrosine kinase (RTK) that consists of extra- cellular immunoglobulin (IG)-like, cysteine-rich and kringle do- mains as well as an intracellular tyrosine kinase (TK) domain and, unique to the Ror family of RTKs, a C-terminal serine–pro- line–threonine-rich (PST) region [1]. Mutations in human ROR2 cause two distinct syndromes, brachydactyly type B (BDB) [2,3] and recessive Robinow syndrome (RRS) [4,5]. Ror2 null mice show skeletal defects including craniofacial abnormalities and a shorten- ing of long bones, especially in the limb zeugopode, as well as de- fects in heart, lung and external genitalia [6,7]. In cartilage, deficiency of Ror2 or overexpression of dominant-negative iso- forms severely impairs chondrocyte differentiation [8,9]. Ror2 was shown to be a receptor for Wnt5a, inducing a non- canonical cascade involving activation of cJun-N-terminal kinase (JNK) [10,11]. In addition, Wnt5a can negatively regulate canonical Wnt signalling via Ror2 [12]. Additionally, Ror2 mediates Wnt5a- induced filopodia formation via its interaction with filamin A (FLNa) [13] depending on JNK [14]. So far, only a small number of Ror2-interacting proteins have been identified. Apart from FLNa, BmpR1b, Dlxin-1, casein kinase 1e (CK1e), Glycogen-synthase ki- nase 3 (Gsk3), Src and the scaffolding protein 14-3-3b have been shown to interact with Ror2 [15–20]. The Wt1-interacting protein Wtip has originally been identified as an interaction partner of the Wilms tumour protein 1 (WT1) in a Y2H screen [21]. Wtip contains three LIM domains (LDs), which are generally thought to mediate protein–protein interactions [22–24] and exhibits high homology to the Ajuba/Zyxin family of LD pro- teins. Here, we describe the interaction of Wtip with the C-termi- nal part of Ror2 in yeast and in mammalian cells and provide comparative expression data of both genes and proteins. Function- ally, we show that Wtip is involved in the intracellular regulation of canonical Wnt signalling. Materials and methods Yeast two-hybrid screening. The cytoplasmic part of mouse Ror2 (Ror2-CP) or distally truncated cytoplasmic mRor2 (Ror2-BDB) were fused to the Tpr dimerisation domain and in frame to the LexA domain. All constructs used for screening procedures were checked for expression by Western blotting (not shown). Bait DNA was transformed into yeast strain L40 (Clontech) and screen- ing was performed against a mouse embryonic cDNA (E9.5–10.5) library [25,26] in VP16 yeast expression vector. Positive clones were picked, restreaked three times and assayed for LacZ activity using a filter b-galactosidase assay to reduce false positives. Antibodies, cells and transfections. Following antibodies were used: mouse anti-HA, rabbit anti-Flag (Sigma–Aldrich); mouse anti-LexA (Clontech); mouse anti-Gal4 (Santa Cruz), rabbit anti- 0006-291X/$ - see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2009.09.086 * Corresponding author. Address: Max Planck-Institute for Molecular Genetics, Development and Disease Group, Ihnestr. 73, 14195 Berlin, Germany. Fax: +49 30 84131385. E-mail address: [email protected] (S. Stricker). 1 Present address: Division of Molecular Neurobiology, MRC National Institute for Medical Research, London NW7 1AA, United Kingdom. Biochemical and Biophysical Research Communications 390 (2009) 211–216 Contents lists available at ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochemical and Biophysical Research Communications 390 (2009) 211–216
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
The LIM domain protein Wtip interacts with the receptor tyrosine kinaseRor2 and inhibits canonical Wnt signalling
Nicole Verhey van Wijk a,b,1, Florian Witte a,b, Ann Carolin Feike c, Alexandra Schambony c,Walter Birchmeier d, Stefan Mundlos a,b, Sigmar Stricker a,b,*
a Max Planck-Institute for Molecular Genetics, Development and Disease Group, Berlin, Germanyb Institute for Medical Genetics, University Medicine Charité, Berlin, Germanyc Developmental Biology Unit, Biology Department, University of Erlangen–Nuremberg, Erlangen, Germanyd Max Delbrück Center for Molecular Medicine, Berlin, Germany
a r t i c l e i n f o
Article history:Received 17 September 2009Available online 26 September 2009
Keywords:WtipRor2BrachydactylyWnt
0006-291X/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.bbrc.2009.09.086
* Corresponding author. Address: Max Planck-InstDevelopment and Disease Group, Ihnestr. 73, 1419584131385.
E-mail address: [email protected] (S. Strick1 Present address: Division of Molecular Neurobiolo
Medical Research, London NW7 1AA, United Kingdom.
a b s t r a c t
Wtip is a LIM domain protein of the Ajuba/Zyxin family involved in kidney and neural crest development;Ror2 is a receptor tyrosine kinase involved in the development of skeleton, heart, lung, genitalia and kid-neys. Here we describe Wtip as an intracellular interaction partner of Ror2. Full-length Ror2 recruits Wtipto the cell membrane, a mutant involved in human disease fails to do so. Both genes and proteins showoverlapping expression in the mouse embryo. We show that Wtip is able to inhibit canonical Wnt signal-ling in mammalian cells and in Xenopus embryos linking Wtip to a crucial developmental pathway.
� 2009 Elsevier Inc. All rights reserved.
Introduction nase 3 (Gsk3), Src and the scaffolding protein 14-3-3b have been
Ror2 is a receptor tyrosine kinase (RTK) that consists of extra-cellular immunoglobulin (IG)-like, cysteine-rich and kringle do-mains as well as an intracellular tyrosine kinase (TK) domainand, unique to the Ror family of RTKs, a C-terminal serine–pro-line–threonine-rich (PST) region [1]. Mutations in human ROR2cause two distinct syndromes, brachydactyly type B (BDB) [2,3]and recessive Robinow syndrome (RRS) [4,5]. Ror2 null mice showskeletal defects including craniofacial abnormalities and a shorten-ing of long bones, especially in the limb zeugopode, as well as de-fects in heart, lung and external genitalia [6,7]. In cartilage,deficiency of Ror2 or overexpression of dominant-negative iso-forms severely impairs chondrocyte differentiation [8,9].
Ror2 was shown to be a receptor for Wnt5a, inducing a non-canonical cascade involving activation of cJun-N-terminal kinase(JNK) [10,11]. In addition, Wnt5a can negatively regulate canonicalWnt signalling via Ror2 [12]. Additionally, Ror2 mediates Wnt5a-induced filopodia formation via its interaction with filamin A(FLNa) [13] depending on JNK [14]. So far, only a small numberof Ror2-interacting proteins have been identified. Apart from FLNa,BmpR1b, Dlxin-1, casein kinase 1e (CK1e), Glycogen-synthase ki-
ll rights reserved.
itute for Molecular Genetics,Berlin, Germany. Fax: +49 30
er).gy, MRC National Institute for
shown to interact with Ror2 [15–20].The Wt1-interacting protein Wtip has originally been identified
as an interaction partner of the Wilms tumour protein 1 (WT1) in aY2H screen [21]. Wtip contains three LIM domains (LDs), which aregenerally thought to mediate protein–protein interactions [22–24]and exhibits high homology to the Ajuba/Zyxin family of LD pro-teins. Here, we describe the interaction of Wtip with the C-termi-nal part of Ror2 in yeast and in mammalian cells and providecomparative expression data of both genes and proteins. Function-ally, we show that Wtip is involved in the intracellular regulationof canonical Wnt signalling.
Materials and methods
Yeast two-hybrid screening. The cytoplasmic part of mouse Ror2(Ror2-CP) or distally truncated cytoplasmic mRor2 (Ror2-BDB)were fused to the Tpr dimerisation domain and in frame to theLexA domain. All constructs used for screening procedures werechecked for expression by Western blotting (not shown). BaitDNA was transformed into yeast strain L40 (Clontech) and screen-ing was performed against a mouse embryonic cDNA (E9.5–10.5)library [25,26] in VP16 yeast expression vector. Positive cloneswere picked, restreaked three times and assayed for LacZ activityusing a filter b-galactosidase assay to reduce false positives.
Antibodies, cells and transfections. Following antibodies wereused: mouse anti-HA, rabbit anti-Flag (Sigma–Aldrich); mouseanti-LexA (Clontech); mouse anti-Gal4 (Santa Cruz), rabbit anti-

212 N. Verhey van Wijk et al. / Biochemical and Biophysical Research Communications 390 (2009) 211–216
Met (kind gift of Dr. U. Schaeper, Berlin); rabbit anti-Ror2 (kind giftof Prof. P. Knaus, Berlin), goat anti-Ror2 (R&D Systems), rabbit anti-WTIP (produced by Eurogentec). Secondary antibodies: anti-goatAlexa Fluor 488, anti-rabbit Alexa Fluor 488, anti-rabbit AlexaFluor 568 (Molecular Probes); anti-mouse and anti-rabbit IgG-per-oxidase conjugate (Oncogene). HEK293 cells were cultured in a-MEM (Cambrex), 10% FBS, 1 mM L-glutamine. Transient transfec-tion of HEK293 cells was done by standard Ca/PO4 precipitation.Cos-1 cells were cultured in DMEM (4.5 g/L glucose), 5% FBS,1 mM L-glutamine and transfected with Polyfectamine (Qiagen)according to manufacturer’s instructions.
Co-immunoprecipitation. Two days after transfection, HEK293cells were washed with cold PBS. Proteins were solubilised in lysisbuffer (50 mM Hepes, pH 7.5, 50 mM NaCl, 10 mM EDTA, 10% glyc-erol, 1% Triton X-100) supplemented with 100 mM NaF, 1 mMNa3VO4, 10 mM sodium pyrophosphate, 1 mM PMSF and 0.5%aprotinin. Proteins were precipitated with anti-Flag affinity aga-rose gel (Sigma–Aldrich), and further analysed by standard Wes-tern blotting procedure.
Immunofluorescence, in-situ hybridisation and immunohistochemistry.Cos-1 cells grown on cover slips were fixed in 4% PFA for 15 minand further processed for immunofluorescence as described inRef. [27]. In-situ hybridisation and immunohistochemistry wereperformed as described in Ref. [9]. A Wtip probe was generatedby cloning the Wtip-Y2H (nt498–957 of Wtip CDS) fragmentinto pCRII-TOPO (Invitrogen) and subsequent transcription withSP6 (Roche). Probes used to detect mRor2 are described inRef. [28].
TOPFLASH assay, injection and analysis of Xenopus laevis embryos.Cos-1 cells per well (105) were seeded in 24-well plates and transfec-ted using ExGen 500 (Fermentas) with either 150 ng of the Tcf/Lefreporter construct TOPFLASH or FOPFLASH (negative control),pCMRV to normalise for transfection efficiency, and constructs forDvl2, Ror2 and Wtip. Total amount of transfected DNA was kept con-stant by adding empty vector DNA. Cells were lysed 48 h after trans-fection with 1�passive lysis buffer (Promega) and luciferase activitywas measured according to the Dual-Glo Luciferase Reporter Assay(Promega). Transfections were performed as duplicates, all experi-ments were repeated at least three times, representative experi-ments are shown. Xenopus axis assays were performed asdescribed in Ref. [29].
Results
Identification of Wtip as Ror2-interacting protein
To identify interaction partners of Ror2 we performed a yeasttwo-hybrid screen with the cytoplasmic domain of Ror2 (Ror2-CP) as bait. We isolated a Wtip fragment encoding amino acids161–319 that contains the first two but not the third LIM domainof Wtip, suggesting that the third LIM domain is not crucial forbinding Ror2.
To confirm the interaction we analysed yeast colony growth onselection medium and LacZ expression in retransformation exper-iments in L40 yeast. Only yeast cells that have been transformedwith Ror2-CP and Wtip-Y2H are enabled to grow on selection med-ium (Fig. 1A, top row) or express a LacZ reporter (Fig. 1A, bottomrow). A truncated version of Ror2 lacking the distal PST domainfailed to interact with Wtip, indicating that the interaction occursvia the PST domain of Ror2.
Since several LIM domain proteins have been shown to interactwith RTKs and tyrosine-containing motifs [30–32] we investigatedthe specificity of the Ror2–Wtip interaction. For that we expressedWtip-Y2H in combination with several cytoplasmic parts of RTKsfused to a LexA domain in L40 yeast (Fig. 1B). We found that Wtip
did not interact with any of the kinases tested, indicating a highlyspecific interaction of Wtip with Ror2.
Confirmation of Wtip–Ror2 interaction in eukaryotic cells
First, we tested the interaction of the Wtip-Y2H-fragment withfull-length Ror2 by co-immunoprecipitation (Co-IP) in HEK293 cells(Fig. 1D). Specificity of the interaction was confirmed, since Wtip-Y2H did not interact with another tyrosine kinase, Trk-Met [26].
To determine if the full-length Wtip is also capable of associatingwith Ror2, the full-length Wtip-FLAG was expressed in HEK293cells together with HA-tagged full-length and truncated Ror2 con-structs [19] (see Fig. 1E). Full-length Wtip was able to specificallybind to full-length Ror2 but not to the truncated Ror2-BDB con-struct (Fig. 1E), confirming the data obtained in yeast. In addition,we found that Wtip under the same conditions is not able to asso-ciate with the closely related tyrosine kinase Ror1 (data not shown).
In order to spot the essential domain of Ror2 responsible forinteracting with Wtip, we used constructs of either the whole cyto-plasmic part of Ror2 (Ror2-IC) or deleted the second serine/threo-nine-rich domain (Ror2-IC–DST2) or the proline- and the secondST-rich (Ror2-IC–DP/ST2) domains, respectively. As shown inFig. 1F, full-length Wtip was able to precipitate all of these Ror2-variants indicating that the interaction between Ror2 and Wtiptakes place at the first ST-rich domain of Ror2.
Recently, it has been described that LIM domain proteins canform homodimers [33]. Co-IP analysis of full-length Wtip-HA andfull-length Wtip-FLAG in HEK293 cells revealed that Wtip is ableto homodimerise. Wtip was not able to interact with a Wtip pro-tein lacking all three LD (WtipDLD1,2,3), thus the LDs in generalare indispensable for homodimerisation of Wtip (Fig. 1G). In con-trast, Wtip lacking the two C-terminally located LD (WtipDLD2,3)was weakly precipitated by Wtip suggesting that LD1 is essentialfor homodimerisation of Wtip (Fig. 1G).
To further confirm a functional interaction of Wtip and Ror2 ineukaryotic cells, we analysed the distribution of Wtip alone or incombination with full-length or truncated Ror2 in Cos-1 cells(Fig. 1H). As reported by Srichai et al. [21], Wtip was predomi-nantly found located in cytoplasmic spots, which was also ob-served in other cell lines (HEK293, HeLa, not shown). Ror2 andRor2-BDB were localised to the cell membrane. Importantly, co-expression of Ror2-HA together with Wtip-FLAG altered the sub-cellular localisation of Wtip (Fig. 1H, lower panel), so that it wasnow predominantly co-localised with Ror2 at the plasma mem-brane. In agreement with our Co-IP data, Ror2-BDB-HA was notable to recruit Wtip to the plasma membrane.
Analysis of Wtip and Ror2 gene and protein expression in mouseembryos
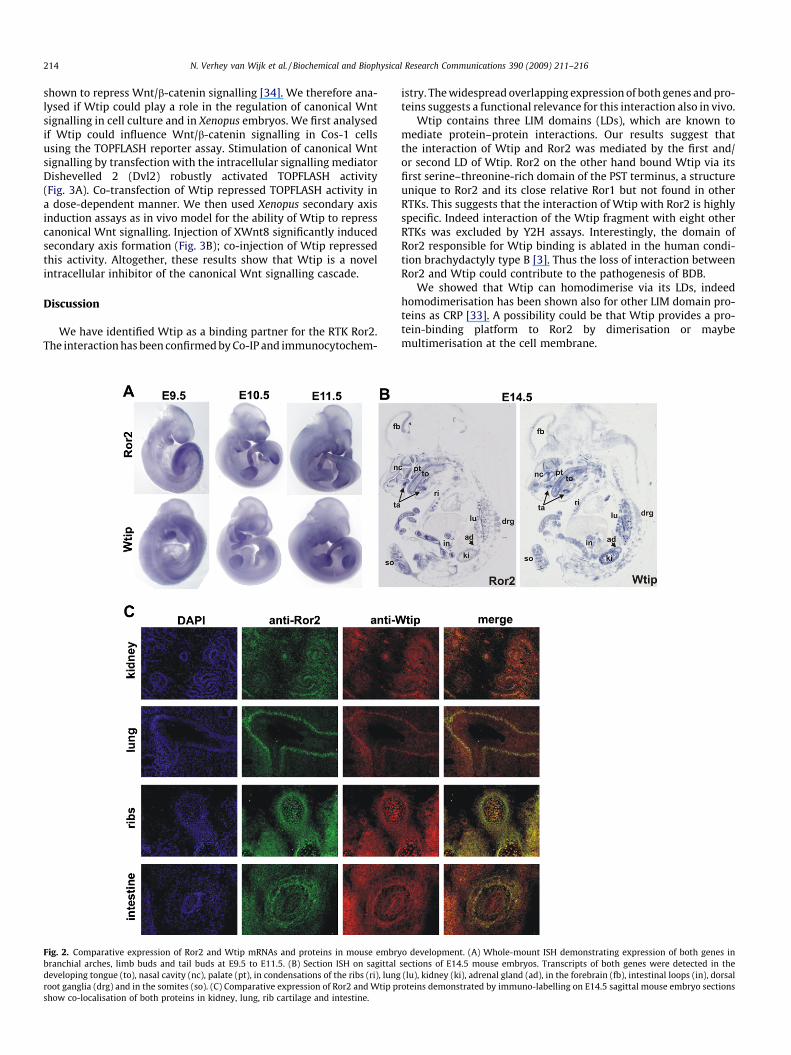
To gain an overall insight into the expression of Wtip and Ror2,we first performed whole-mount in-situ hybridisation on mouseembryos. At embryonic (E) stages E9.5 to E11.5, Wtip and Ror2 bothare expressed in the branchial arches, the otic vesicle, the outgrow-ing limb buds, in somites, craniofacial mesenchyme and in the tail(Fig. 2A).
To obtain information about the expression of Wtip and Ror2during organogenesis, we analysed sagittal sections from mouseembryos stage E14.5 by section in-situ hybridisation. Wtip andRor2 showed a broadly overlapping expression in the developingmouse embryo (Fig. 2B).
To compare Wtip and Ror2 protein expression, we performedimmunohistochemistry for Wtip and Ror2 on sagittal sections ofE14.5 mouse embryos (Fig. 2C). Co-expression of Ror2 and Wtipproteins was verified in kidneys, in lung epithelium, in the cartilag-inous condensations of the developing ribs and in intestinal loops.

Fig. 1. Interaction of Wtip and Ror2. (A) Identification of Wtip as a binding partner of Ror2 by Y2H: confirmation of Ror2/Wtip interaction on selective medium (-THULL) andinduction of LacZ. (B) Wtip interacts with Ror2, but not with 7 other RTKs. (C) Depiction of Ror2 and Wtip constructs used. (D) Wtip-Y2H interacts with full-length Ror2, butnot with Met in HEK293 cells. (E) Wtip interacts with full-length Ror2 but not with a BDB truncation. (F) Wtip interacts with a form of Ror2 that carries only the first serine–threonine-rich domain. (G) Homodimerisation of Wtip via its LIM domains. (H) Ror2 but not Ror2-BDB recruits Wtip to the cell membrane. Cos-1 cells were transfected withconstructs indicated, nuclear staining was performed with DAPI (blue). Top row: single transfections, bottom row: co-transfections. (For interpretation of the references tocolour in this figure legend, the reader is referred to the web version of this paper.)
N. Verhey van Wijk et al. / Biochemical and Biophysical Research Communications 390 (2009) 211–216 213
Taken together the analysis of Wtip/Ror2 association by co-immunoprecipitation, together with the subcellular localisationof Wtip and Ror2 and their co-expression (mRNA and protein) indi-cates a functional interaction of these proteins in vivo.
Wtip inhibits canonical Wnt signalling in vitro and in vivo
Ror2 has been implicated in the modulation of canonical Wntsignalling [12] and the closely related LIM protein Ajuba was
214 N. Verhey van Wijk et al. / Biochemical and Biophysical Research Communications 390 (2009) 211–216
shown to repress Wnt/b-catenin signalling [34]. We therefore ana-lysed if Wtip could play a role in the regulation of canonical Wntsignalling in cell culture and in Xenopus embryos. We first analysedif Wtip could influence Wnt/b-catenin signalling in Cos-1 cellsusing the TOPFLASH reporter assay. Stimulation of canonical Wntsignalling by transfection with the intracellular signalling mediatorDishevelled 2 (Dvl2) robustly activated TOPFLASH activity(Fig. 3A). Co-transfection of Wtip repressed TOPFLASH activity ina dose-dependent manner. We then used Xenopus secondary axisinduction assays as in vivo model for the ability of Wtip to represscanonical Wnt signalling. Injection of XWnt8 significantly inducedsecondary axis formation (Fig. 3B); co-injection of Wtip repressedthis activity. Altogether, these results show that Wtip is a novelintracellular inhibitor of the canonical Wnt signalling cascade.
Discussion
We have identified Wtip as a binding partner for the RTK Ror2.The interaction has been confirmed by Co-IP and immunocytochem-
Fig. 2. Comparative expression of Ror2 and Wtip mRNAs and proteins in mouse embrybranchial arches, limb buds and tail buds at E9.5 to E11.5. (B) Section ISH on sagittaldeveloping tongue (to), nasal cavity (nc), palate (pt), in condensations of the ribs (ri), lungroot ganglia (drg) and in the somites (so). (C) Comparative expression of Ror2 and Wtip pshow co-localisation of both proteins in kidney, lung, rib cartilage and intestine.
istry. The widespread overlapping expression of both genes and pro-teins suggests a functional relevance for this interaction also in vivo.
Wtip contains three LIM domains (LDs), which are known tomediate protein–protein interactions. Our results suggest thatthe interaction of Wtip and Ror2 was mediated by the first and/or second LD of Wtip. Ror2 on the other hand bound Wtip via itsfirst serine–threonine-rich domain of the PST terminus, a structureunique to Ror2 and its close relative Ror1 but not found in otherRTKs. This suggests that the interaction of Wtip with Ror2 is highlyspecific. Indeed interaction of the Wtip fragment with eight otherRTKs was excluded by Y2H assays. Interestingly, the domain ofRor2 responsible for Wtip binding is ablated in the human condi-tion brachydactyly type B [3]. Thus the loss of interaction betweenRor2 and Wtip could contribute to the pathogenesis of BDB.
We showed that Wtip can homodimerise via its LDs, indeedhomodimerisation has been shown also for other LIM domain pro-teins as CRP [33]. A possibility could be that Wtip provides a pro-tein-binding platform to Ror2 by dimerisation or maybemultimerisation at the cell membrane.
o development. (A) Whole-mount ISH demonstrating expression of both genes insections of E14.5 mouse embryos. Transcripts of both genes were detected in the(lu), kidney (ki), adrenal gland (ad), in the forebrain (fb), intestinal loops (in), dorsal
roteins demonstrated by immuno-labelling on E14.5 sagittal mouse embryo sections

Fig. 3. Inhibition of canonical Wnt signalling by Wtip. (A) TOPFLASH assay stimulated with Dishevelled-2 (Dvl2). (B) Axis induction assay in Xenopus embryos showssecondary axis induction induced by XWnt8 is inhibited significantly by Wtip. Representative embryos are shown for each injection experiment, secondary axes are markedwith arrows.
N. Verhey van Wijk et al. / Biochemical and Biophysical Research Communications 390 (2009) 211–216 215
Zyxin LIM proteins are known to localise to the cell membraneespecially at sites of cell–cell contacts or matrix attachment sites.In addition, Zyxin family members have been shown to shuttle be-tween membrane and nucleus and are thought to coordinate cellattachment events with nuclear responses [21 and referencestherein]. We found that Ror2 was able to recruit Wtip to the cellmembrane in an overexpression situation. It remains to be deter-mined, if both proteins endogenously localise to cell–cell contactsites or focal adhesions and could contribute to their function.
We show that Wtip can antagonise canonical Wnt signalling.Ror2 has also been implicated in several Wnt pathways and wasshown to inhibit canonical signalling as well [11,12,35,36]. It isworthwhile noting that two key players of the canonical Wnt cas-cade, CK1e and Gsk3, have been shown to interact with Ror2[15,18]. CK1e was shown to phosphorylate Ror2 on its serine–thre-onine-rich 2 domain resulting in Ror2 autophosphorylation [15].Gsk3a and Gsk3b were shown to phosphorylate Ror2 after stimu-lation with Wnt5a [18]. Moreover it was shown that Gsk3 is neces-sary for Wnt5a induced cell migration [18]. Furthermore,Haraguchi et al. [34] have recently shown that Ajuba, a LD contain-ing protein of the Zyxin family closely related to Wtip, is phosphor-ylated by Gsk3b. Ajuba was shown to negatively regulate canonicalWnt signalling by enforcing the interaction of Gsk3b with b-cate-nin. Ajuba thereby increased the Gsk3b-mediated phosphorylationand thus destabilisation of b-catenin. At this point we cannot stateon which level the inhibition of canonical Wnt signalling takesplace, however it is tempting to speculate that Wtip could functionalong a similar mechanism. In conclusion, we have demonstratedthat the receptor tyrosine kinase Ror2, via its C-terminal domain,interacts with the LIM domain protein Wtip, recruiting it to the cellsurface. Furthermore, Wtip is a novel negative regulator of thecanonical Wnt signalling pathway.
Acknowledgments
This project was funded by grants form the Deutsche Fors-chungsgemeinschaft to S.S. and S.M. (SFB 577) and to A.S. (965 2-
3). We acknowledge the expert technical assistance from KathrinSeidel and Norbert Brieske. We thank Ute Schaeper for help withY2H experiments.
References
[1] P. Masiakowski, R.D. Carroll, A novel family of cell surface receptors withtyrosine kinase-like domain, J. Biol. Chem. 267 (1992) 26181–26190.
[2] M. Oldridge, A.M. Fortuna, M. Maringa, P. Propping, S. Mansour, C. Pollitt, T.M.DeChiara, R.B. Kimble, D.M. Valenzuela, G.D. Yancopoulos, A.O. Wilkie,Dominant mutations in ROR2, encoding an orphan receptor tyrosine kinase,cause brachydactyly type B, Nat. Genet. 24 (2000) 275–278.
[3] G.C. Schwabe, S. Tinschert, C. Buschow, P. Meinecke, G. Wolff, G. Gillessen-Kaesbach, M. Oldridge, A.O. Wilkie, R. Komec, S. Mundlos, Distinct mutations inthe receptor tyrosine kinase gene ROR2 cause brachydactyly type B, Am. J.Hum. Genet. 67 (2000) 822–831.
[4] A.R. Afzal, A. Rajab, C.D. Fenske, M. Oldridge, N. Elanko, E. Ternes-Pereira, B.Tuysuz, V.A. Murday, M.A. Patton, A.O. Wilkie, S. Jeffery, Recessive Robinowsyndrome, allelic to dominant brachydactyly type B, is caused by mutation ofROR2, Nat. Genet. 25 (2000) 419–422.
[5] H. van Bokhoven, J. Celli, H. Kayserili, E. van Beusekom, S. Balci, W. Brussel, F.Skovby, B. Kerr, E.F. Percin, N. Akarsu, H.G. Brunner, Mutation of the geneencoding the ROR2 tyrosine kinase causes autosomal recessive Robinowsyndrome, Nat. Genet. 25 (2000) 423–426.
[6] T.M. DeChiara, R.B. Kimble, W.T. Poueymirou, J. Rojas, P. Masiakowski, D.M.Valenzuela, G.D. Yancopoulos, Ror2, encoding a receptor-like tyrosine kinase, isrequired for cartilage and growth plate development, Nat. Genet. 24 (2000) 271–274.
[7] S. Takeuchi, K. Takeda, I. Oishi, M. Nomi, M. Ikeya, K. Itoh, S. Tamura, T. Ueda, T.Hatta, H. Otani, T. Terashima, S. Takada, H. Yamamura, S. Akira, Y. Minami,Mouse Ror2 receptor tyrosine kinase is required for the heart development andlimb formation, Genes Cells 5 (2000) 71–78.
[8] G.C. Schwabe, S. Turkmen, G. Leschik, S. Palanduz, B. Stover, T.O. Goecke, S.Mundlos, Brachydactyly type C caused by a homozygous missense mutation inthe prodomain of CDMP1, Am. J. Med. Genet. A 124 (2004) 356–363.
[9] S. Stricker, N. Verhey van Wijk, F. Witte, N. Brieske, K. Seidel, S. Mundlos,Cloning and expression pattern of chicken Ror2 and functionalcharacterization of truncating mutations in Brachydactyly type B andRobinow syndrome, Dev. Dyn. 235 (2006) 3456–3465.
[10] I. Oishi, H. Suzuki, N. Onishi, R. Takada, S. Kani, B. Ohkawara, I. Koshida, K.Suzuki, G. Yamada, G.C. Schwabe, S. Mundlos, H. Shibuya, S. Takada, Y. Minami,The receptor tyrosine kinase Ror2 is involved in non-canonical Wnt5a/JNKsignalling pathway, Genes Cells 8 (2003) 645–654.
[11] A. Schambony, D. Wedlich, Wnt-5A/Ror2 regulate expression of XPAPC throughan alternative noncanonical signaling pathway, Dev. Cell 12 (2007) 779–792.
[12] A.J. Mikels, R. Nusse, Purified Wnt5a protein activates or inhibits beta-catenin-TCF signaling depending on receptor context, PLoS Biol. 4 (2006) e115.

216 N. Verhey van Wijk et al. / Biochemical and Biophysical Research Communications 390 (2009) 211–216
[13] M. Nishita, S.K. Yoo, A. Nomachi, S. Kani, N. Sougawa, Y. Ohta, S. Takada, A.Kikuchi, Y. Minami, Filopodia formation mediated by receptor tyrosine kinaseRor2 is required for Wnt5a-induced cell migration, J. Cell Biol. 175 (2006) 555–562.
[14] A. Nomachi, M. Nishita, D. Inaba, M. Enomoto, M. Hamasaki, Y. Minami,Receptor tyrosine kinase Ror2 mediates Wnt5a-induced polarized cellmigration by activating c-Jun N-terminal kinase via actin-binding proteinfilamin A, J. Biol. Chem. 283 (2008) 27973–27981.
[15] S. Kani, I. Oishi, H. Yamamoto, A. Yoda, H. Suzuki, A. Nomachi, K. Iozumi, M.Nishita, A. Kikuchi, T. Takumi, Y. Minami, The receptor tyrosine kinase Ror2associates with and is activated by casein kinase I epsilon, J. Biol. Chem. 279(2004) 50102–50109.
[16] Y. Liu, J.F. Ross, P.V. Bodine, J. Billiard, Homodimerization of Ror2 tyrosinekinase receptor induces 14-3-3(beta) phosphorylation and promotesosteoblast differentiation and bone formation, Mol. Endocrinol. 21 (2007)3050–3061.
[17] T. Matsuda, H. Suzuki, I. Oishi, S. Kani, Y. Kuroda, T. Komori, A. Sasaki, K.Watanabe, Y. Minami, The receptor tyrosine kinase Ror2 associates with themelanoma-associated antigen (MAGE) family protein Dlxin-1 and regulates itsintracellular distribution, J. Biol. Chem. 278 (2003) 29057–29064.
[18] H. Yamamoto, S.K. Yoo, M. Nishita, A. Kikuchi, Y. Minami, Wnt5a modulatesglycogen synthase kinase 3 to induce phosphorylation of receptor tyrosinekinase Ror2, Genes Cells 12 (2007) 1215–1223.
[19] M. Sammar, S. Stricker, G.C. Schwabe, C. Sieber, A. Hartung, M. Hanke, I. Oishi, J.Pohl, Y. Minami, W. Sebald, S. Mundlos, P. Knaus, Modulation of GDF5/BRI-bsignalling through interaction with the tyrosine kinase receptor Ror2, GenesCells 9 (2004) 1227–1238.
[20] S. Akbarzadeh, L. Wheldon, S. Sweet, S. Talma, F. Mardakheh, J. Heath, Thedeleted in brachydactyly B domain of ROR2 is required for receptor activationby recruitment of Src, PLoS One 3 (3) (2008) e1873.
[21] M.B. Srichai, M. Konieczkowski, A. Padiyar, D.J. Konieczkowski, A. Mukherjee,P.S. Hayden, S. Kamat, M.A. El-Meanawy, S. Khan, P. Mundel, S.B. Lee, L.A.Bruggeman, J.R. Schelling, J.R. Sedor, A WT1 co-regulator controls podocytephenotype by shuttling between adhesion structures and nucleus, J. Biol.Chem. 279 (2004) 14398–14408.
[22] I.B. Dawid, J.J. Breen, R. Toyama, LIM domains: multiple roles as adapters andfunctional modifiers in protein interactions, Trends Genet. 14 (1998)156–162.
[23] J.L. Kadrmas, M.C. Beckerle, The LIM domain: from the cytoskeleton to thenucleus, Nat. Rev. Mol. Cell Biol. 5 (2004) 920–931.
[24] K.L. Schmeichel, M.C. Beckerle, The LIM domain is a modular protein-bindinginterface, Cell 79 (1994) 211–219.
[25] J. Behrens, J.P. von Kries, M. Kuhl, L. Bruhn, D. Wedlich, R. Grosschedl, W.Birchmeier, Functional interaction of beta-catenin with the transcriptionfactor LEF-1, Nature 382 (1996) 638–642.
[26] K.M. Weidner, S. Di Cesare, M. Sachs, V. Brinkmann, J. Behrens, W. Birchmeier,Interaction between Gab1 and the c-Met receptor tyrosine kinase isresponsible for epithelial morphogenesis, Nature 384 (1996) 173–176.
[27] A.N. Albrecht, U. Kornak, A. Boddrich, K. Suring, P.N. Robinson, A.C. Stiege, R.Lurz, S. Stricker, E.E. Wanker, S. Mundlos, A molecular pathogenesis fortranscription factor associated poly-alanine tract expansions, Hum. Mol.Genet. 13 (2004) 2351–2359.
[28] G.C. Schwabe, B. Trepczik, K. Suring, N. Brieske, A.S. Tucker, P.T. Sharpe, Y.Minami, S. Mundlos, Ror2 knockout mouse as a model for the developmentalpathology of autosomal recessive Robinow syndrome, Dev. Dyn. 229 (2004)400–410.
[29] V. Bryja, D. Gradl, A. Schambony, E. Arenas, G. Schulte, Beta-arrestin is anecessary component of Wnt/beta-catenin signaling in vitro and in vivo, Proc.Natl. Acad. Sci. USA 104 (2007) 6690–6695.
[30] K. Durick, R.Y. Wu, G.N. Gill, S.S. Taylor, Mitogenic signaling by Ret/ptc2requires association with enigma via a LIM domain, J. Biol. Chem. 271 (1996)12691–12694.
[31] R. Wu, K. Durick, Z. Songyang, L.C. Cantley, S.S. Taylor, G.N. Gill, Specificity ofLIM domain interactions with receptor tyrosine kinases, J. Biol. Chem. 271(1996) 15934–15941.
[32] R.Y. Wu, G.N. Gill, LIM domain recognition of a tyrosine-containing tight turn,J. Biol. Chem. 269 (1994) 25085–25090.
[33] R. Feuerstein, X. Wang, D. Song, N.E. Cooke, S.A. Liebhaber, The LIM/doublezinc-finger motif functions as a protein dimerization domain, Proc. Natl. Acad.Sci. USA 91 (1994) 10655–10659.
[34] K. Haraguchi, M. Ohsugi, Y. Abe, K. Semba, T. Akiyama, T. Yamamoto, Ajubanegatively regulates the Wnt signaling pathway by promoting GSK-3beta-mediated phosphorylation of beta-catenin, Oncogene 27 (2008) 274–284.
[35] J. Billiard, D.S. Way, L.M. Seestaller-Wehr, R.A. Moran, A. Mangine, P.V. Bodine,The orphan receptor tyrosine kinase Ror2 modulates canonical Wnt signalingin osteoblastic cells, Mol. Endocrinol. 19 (2005) 90–101.
[36] A. Winkel, S. Stricker, P. Tylzanowski, V. Seiffart, S. Mundlos, G. Gross, A.Hoffmann, Wnt-ligand-dependent interaction of TAK1 (TGF-beta-activatedkinase-1) with the receptor tyrosine kinase Ror2 modulates canonical Wnt-signalling, Cell. Signal. 20 (2008) 2134–2144.
Related Documents











