The Leishmania promastigote surface antigen-2 (PSA-2) is speci¢cally recognised by Th1 cells in humans with naturally acquired immunity to L. major Michael Kemp a; *, Emanuela Handman b , Ka îre Kemp a , Ahmed Ismail a;c , Mustafa D. Mustafa c , Anuar Y. Kordofani c , Klaus Bendtzen d , Arsalan Kharazmi a , Thor G. Theander a a Center for Medical Parasitology, Department of Clinical Microbiology, and Department of Infectious Diseases, RHIMA Centre, University Hospital (Rigshospitalet) and Institute for Medical Microbiology and Immunology, University of Copenhagen, Copenhagen, Denmark b The Walter and Eliza Hall Institute of Medical Research, Melbourne, Vic., Australia c Department of Pathology, University of Khartoum, Khartoum, Sudan d Institute for In£ammation Research, RHIMA Centre, University Hospital (Rigshospitalet), Copenhagen, Denmark Received 28 November 1997; revised 9 January 1998; accepted 9 January 1998 Abstract The promastigote surface antigen-2 (PSA-2) is a Leishmania parasite antigen, which can induce Th1-mediated protection against murine leishmaniasis when used as a vaccine. To evaluate PSA-2 as a human vaccine candidate the specific T-cell response to PSA-2 was characterised in individuals immune to cutaneous leishmaniasis. Peripheral blood mononuclear cells from Sudanese individuals with a past history of self-healing cutaneous leishmaniasis proliferated vigorously in response to PSA-2 isolated from Leishmania major, whereas the antigen did not activate cells from presumably unexposed Danes. Peripheral blood mononuclear cells from individuals with previous L. major infection had varying proliferative responses to PSA-2 derived from L. donovani promastigotes. Peripheral blood mononuclear cells activated by PSA-2 from L. major produced high amounts of interferon-Q and tumour necrosis factor-L, and little interleukin-4, thereby showing a Th1 cytokine pattern. Parallel cultures showed clear Th1 and Th2 response patterns to purified protein derivative of tuberculin or tetanus toxoid, respectively. Flow cytometric analysis revealed that PSA-2 induced blastogenesis in the CD3 positive population and that these cells were the major source of interferon-Q. The results show that Th1-like cells recognising PSA-2 are expanded during infection by L. major and that they maintain their Th1-like cytokine profile upon reactivation in vitro. Since immunity to cutaneous leishmaniasis is mediated by antigen-specific Th1-like cells, PSA-2 might be considered a vaccine candidate for human leishmaniasis. z 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V. Keywords : Leishmania major ; Interferon-Q ; Tumor necrosis factor-L ; Interleukin-4 ; Th1 1. Introduction Diseases caused by obligate intracellular parasites of the genus Leishmania are among the major health 0928-8244 / 98 / $19.00 ß 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V. PII S0928-8244(98)00015-7 * Corresponding author. Tel.: +45 32 68 36 42; Fax: +45 32 68 38 73; E-mail: [email protected] FEMS Immunology and Medical Microbiology 20 (1998) 209^218

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Leishmania promastigote surface antigen-2 (PSA-2) isspeci¢cally recognised by Th1 cells in humans with naturally
acquired immunity to L. major
Michael Kemp a;*, Emanuela Handman b, Kaîre Kemp a, Ahmed Ismail a;c,Mustafa D. Mustafa c, Anuar Y. Kordofani c, Klaus Bendtzen d,
Arsalan Kharazmi a, Thor G. Theander a
a Center for Medical Parasitology, Department of Clinical Microbiology, and Department of Infectious Diseases, RHIMA Centre, UniversityHospital (Rigshospitalet) and Institute for Medical Microbiology and Immunology, University of Copenhagen, Copenhagen, Denmark
b The Walter and Eliza Hall Institute of Medical Research, Melbourne, Vic., Australiac Department of Pathology, University of Khartoum, Khartoum, Sudan
d Institute for In£ammation Research, RHIMA Centre, University Hospital (Rigshospitalet), Copenhagen, Denmark
Received 28 November 1997; revised 9 January 1998; accepted 9 January 1998
Abstract
The promastigote surface antigen-2 (PSA-2) is a Leishmania parasite antigen, which can induce Th1-mediated protectionagainst murine leishmaniasis when used as a vaccine. To evaluate PSA-2 as a human vaccine candidate the specific T-cellresponse to PSA-2 was characterised in individuals immune to cutaneous leishmaniasis. Peripheral blood mononuclear cellsfrom Sudanese individuals with a past history of self-healing cutaneous leishmaniasis proliferated vigorously in response toPSA-2 isolated from Leishmania major, whereas the antigen did not activate cells from presumably unexposed Danes.Peripheral blood mononuclear cells from individuals with previous L. major infection had varying proliferative responses toPSA-2 derived from L. donovani promastigotes. Peripheral blood mononuclear cells activated by PSA-2 from L. majorproduced high amounts of interferon-Q and tumour necrosis factor-L, and little interleukin-4, thereby showing a Th1 cytokinepattern. Parallel cultures showed clear Th1 and Th2 response patterns to purified protein derivative of tuberculin or tetanustoxoid, respectively. Flow cytometric analysis revealed that PSA-2 induced blastogenesis in the CD3 positive population andthat these cells were the major source of interferon-Q. The results show that Th1-like cells recognising PSA-2 are expandedduring infection by L. major and that they maintain their Th1-like cytokine profile upon reactivation in vitro. Since immunityto cutaneous leishmaniasis is mediated by antigen-specific Th1-like cells, PSA-2 might be considered a vaccine candidate forhuman leishmaniasis. z 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V.
Keywords: Leishmania major ; Interferon-Q ; Tumor necrosis factor-L ; Interleukin-4; Th1
1. Introduction
Diseases caused by obligate intracellular parasitesof the genus Leishmania are among the major health
0928-8244 / 98 / $19.00 ß 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V.PII S 0 9 2 8 - 8 2 4 4 ( 9 8 ) 0 0 0 1 5 - 7
FEMSIM 853 27-3-98
* Corresponding author. Tel. : +45 32 68 36 42;Fax: +45 32 68 38 73; E-mail: [email protected]
FEMS Immunology and Medical Microbiology 20 (1998) 209^218

problems in a number of countries. The visceral andcutaneous forms of leishmaniasis cause much humansu¡ering and enormous economic losses. Treatmentof the diseases is expensive and not always e¤cient.No e¡ective vaccine is currently available againstleishmaniasis. There is hope, however, that it maybe possible to develop e¡ective vaccines against theseinfections since immunity often develops after curefrom leishmaniasis, either spontaneously or aftermedical treatment. A thorough knowledge of the im-munoregulatory mechanisms leading to immunityand pathology is necessary for the development ofe¡ective vaccines.
In recent years much information has been ob-tained on the roles of parasite-speci¢c T cells inboth protection and pathology [1]. Results from invitro investigations of human T cells have suggestedthat conclusions from experimental infections ofmice with L. major may be extrapolated to the hu-man situation. In mice, there is a clear associationbetween resistance to disease and strong Th1 (inter-feron- Q (IFN-Q)-, interleukin (IL)-2-, and tumournecrosis factor-L (TNF-L)- producing) parasite-spe-ci¢c T-cell responses on the one hand, and suscept-ibility and Th2 (IL-4-, IL-5-, IL-6- and IL-10-pro-ducing) responses on the other. In humans who areimmune to L. major after natural exposure, either asa subclinical infection or as self-healing cutaneousleishmaniasis (CL), there is a strong Th1 response,which contributes to protection against parasite in-fection at the cellular level [2,3]. Even within theclinical spectrum of cutaneous leishmaniasis themilder presentations are associated with a predomi-nant Th1 response compared to more severe caseswhere Th2 cell activation is also evident [4]. Vaccinecandidates for the prophylaxis of cutaneous leishma-niasis would be expected to induce strong Th1 re-sponses in humans without expansion of cells witha Th2 cytokine secretion pattern. An important con-sideration for a vaccine against leishmaniasis is itsability to induce not only a Th1 initial response,but a T-cell memory of the Th1 type, so that uponinfection with L. major only this population will beexpanded.
Recovery from CL depends on the ability to mod-ulate an initial mixed Th1/Th2 response into a ¢xedTh1-type response [5,6]. In vitro studies have shownthat Th0 cells can be driven to di¡erentiate into ei-
ther Th1-like or Th2-like cells, but become ¢xed intheir phenotype at some point in time [7,8]. An un-answered question in T-cell immunology is whetherupon re-stimulation memory T cells maintain theoriginal £exibility of the Th0 phenotype, or are al-ready ¢xed in their respective Th1 or Th2 phenotype.
Several native and recombinant Leishmania anti-gens have been tested in murine vaccination experi-ments. Among these, the promastigote surface anti-gen 2 (PSA-2) has given good protection againstsubsequent challenge infection with L. major [9].The PSA-2 glycoprotein complex of L. major is aglycosylphosphatidylinositol membrane-anchoredpolypeptide [10]. In promastigotes there are threePSA-2 polypeptides, which migrate on SDS-PAGEwith apparent molecular masses of 96 kDa, 80 kDa,and 50 kDa [11]. In amastigotes a 50-kDa polypep-tide is expressed which shows extensive homology tothe promastigote proteins [9]. Moreover, homologuesof PSA-2 are expressed by all Leishmania speciesexcept L. braziliensis [12]. The amastigote PSA-2from L. major has been expressed as a recombinantproduct by both Escherichia coli and L. mexicana[11].
As PSA-2 from L. major can vaccinate miceagainst experimental infection with the parasite,studies on its potential use as a vaccine in humansare warranted. As a ¢rst step we have characterisedthe speci¢c T-cell response to PSA-2 in individualswith protective immunity to CL. Cytokine pro¢les ofperipheral blood mononuclear cells (PBMC) re-sponding to PSA-2 allowed these cells to be classi¢edaccording to the Th1/Th2 T-cell subset concept. ThePSA-2-reactive cells produced large amounts of in-terferon-Q (IFN-Q), which is a major activator ofmacrophages for resistance to Leishmania parasites.By intracellular staining with IFN-Q speci¢c antibod-ies and £ow cytometry, this cytokine was found to beproduced predominantly by T cells.
2. Materials and methods
2.1. Donors and isolation of mononuclear cells
Leishmanin skin test-positive donors with a his-tory of self-healing CL were from El Izergab villagenorth of Khartoum, Sudan. The epidemiology of CL
FEMSIM 853 27-3-98
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218210

in this village and previous studies on CL in thepopulation have been described elsewhere [2,13].On regular follow-up visits, no new cases were regis-tered for several years after the epidemic had ceased.On a visit to the village by the end of 1995, new casesof CL had started to appear, but only in childrenborn after the end of the epidemic. Informed consentwas obtained from all individuals participating in thestudy. Clinical examination was performed beforeheparinised blood was collected and PBMC wereisolated by Lymphoprep (Nyegaard, Oslo, Norway)density centrifugation. The cells were frozen, stored,and transported in liquid nitrogen as previously de-scribed [2]. Blood from Danish volunteers withoutknown exposure to Leishmania parasites was col-lected and treated similarly. Before use, the cellswere rapidly thawed and washed. The viability ofthe cells was ascertained by trypan blue dye exclu-sion.
2.2. Antigens
PSA-2 was isolated from promastigotes of L. ma-jor (V121, a cloned line from an Israeli isolate LRC-L137) and L. donovani (Indian isolate LRC-L52)[11]. An E. coli-derived recombinant protein ofL. major amastigote PSA-2 from V121 was obtainedas a Schistosoma japonicum glutathione S-transferasefusion protein [9]. The term PSA-2 refers to the na-tive polypeptide puri¢ed from L. major promasti-gotes except where otherwise speci¢ed. In initial ex-periments, PSA-2 was found to induce optimalproliferative responses when used at a ¢nal concen-tration of 1 Wg ml31, and this concentration was usedthroughout the experiments. The amastigoterecombinant protein was tested at 1 Wg ml31 and5 Wg ml31 (¢nal concentrations).
Puri¢ed protein derivative of tuberculin (PPD)and tetanus toxoid were purchased from Statens Ser-uminstitut (Copenhagen, Denmark) and used at ¢nalconcentrations of 12 Wg ml31 and 3 Wg ml31, respec-tively.
2.3. Lymphoproliferation and cytokine production
PBMC were cultured in RPMI 1640 with 10 mMHEPES, 20 U ml31 penicillin and 20 mg ml31 strep-tomycin with 15% heat-inactivated pooled normal
human serum (NHS). The cells were incubatedwith antigens at 6.6U105 cells ml31 in volumes of170 ml in round-bottomed microculture plates(Nunc, Roskilde, Denmark). The cultures were incu-bated for 7 days at 37³C, 5% CO2 in a humidi¢edatmosphere and pulsed with [3H]thymidine (1.85MBq ml31, 20 ml per culture) (New England Nu-clear, Boston, MA, USA) for the last 24 h of incu-bation. The culture supernatants were recovered andstored at 320³C for later determination of IFN-Q.The cells were harvested onto glass-¢bre ¢lters andthe incorporation of [3H]thymidine into DNA wasdetermined by measurement of L-radiation, using aMatrix L-counter (Packard, Greve, Denmark). Alltests were done in triplicate. For each set of samplesthe median was recorded. The speci¢c response wasexpressed as the di¡erence between antigen-stimu-lated cultures and cultures incubated without anti-gen. This is referred to as `incremental counts permin' (cpm). A positive proliferative response was de-¢ned as stimulatory index (SI) s 2.5 and speci¢cresponse s 0.5 kcpm.
For the measurement of interleukin-4 (IL-4) re-lease by antigen-stimulated cultures of PBMC, par-allel cultures were incubated for 6 days and thenpulsed with 1 mM of ionomycin and 50 ng ml31 ofphorbol myristate acetate (PMA) (both from Sigma,St. Louis, MO, USA) for 24 h before the culturesupernatants from triplicate wells were harvested[14].
Tumour necrosis factor-L (TNF-L) and IL-5 weremeasured in culture supernatants after 7 days of in-cubation both with and without re-stimulation withPMA and ionomycin.
2.4. Cytokine measurements
IFN-Q and IL-4 in supernatants of cultures ofPBMC were measured by ELISA as described else-where [14]. IL-5 and TNF-L were measured by com-mercial ELISA kits (RpD systems, Abingdon, UK).The antigen-induced cytokine production was calcu-lated as the di¡erence between antigen-stimulatedand unstimulated cultures.
2.5. Flow cytometry
PBMC were incubated for 6 days with or without
FEMSIM 853 27-3-98
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218 211
PSA-2 as described with a cell density of 1U106 cellsml31. Monensin (Sigma) was added in a ¢nal con-centration of 1.5 mM 6 h before the end of the cul-ture period. The cells were then washed once withPBS and ¢xed by incubation in 2% formaldehyde(Sigma) in PBS for 20 min. at 4³C. The ¢xed cellswere washed twice and re-suspended in PBS contain-ing 0.5% BSA and 0.01% NaN3 (PBS/BSA/NaN3).1U106 cells were incubated with PE-conjugated anti-CD3 antibody (Dako, Glostrup, Denmark) at roomtemperature for 20 min. The cells were washed twicein PBS/BSA/NaN3 and then once in PBS/BSA/NaN3
with 0.1% w/v saponin (Sigma) (saponin bu¡er). Thepermeabilised cells were then stained for intracellularIFN-Q by incubation at room temperature for 20 minwith FITC-conjugated anti-IFN-Q antibody (Phar-mingen, San Diego, CA, USA) in saponin bu¡er,washed twice with saponin bu¡er and subsequentlytwice with staining bu¡er. The cells were analysed onan Epics XL-MCL £ow-cytometer (Coulter, Miami,FL, USA).
The £ow-cytometric data were analysed by PcLy-sisII software (Becton-Dickinson, Mountain View,CA, USA). The cell populations were de¢ned by
forward and side scatter gatings and limits for £uo-rescence-positive cells were set from adequate non-sense IgG controls of each donor.
3. Results
3.1. Antigen-induced proliferation of PBMC
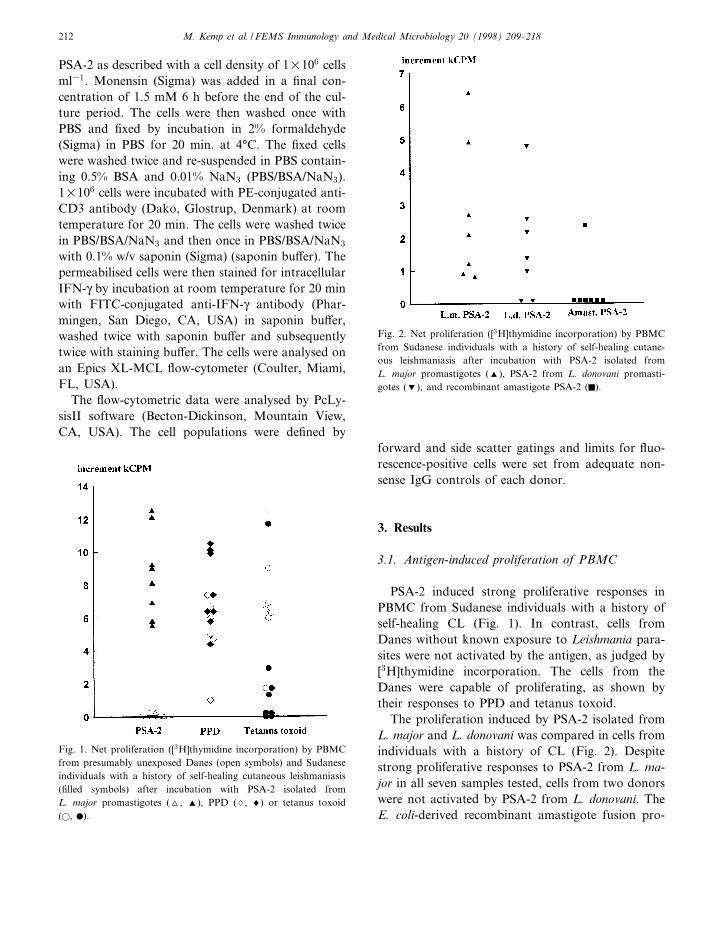
PSA-2 induced strong proliferative responses inPBMC from Sudanese individuals with a history ofself-healing CL (Fig. 1). In contrast, cells fromDanes without known exposure to Leishmania para-sites were not activated by the antigen, as judged by[3H]thymidine incorporation. The cells from theDanes were capable of proliferating, as shown bytheir responses to PPD and tetanus toxoid.
The proliferation induced by PSA-2 isolated fromL. major and L. donovani was compared in cells fromindividuals with a history of CL (Fig. 2). Despitestrong proliferative responses to PSA-2 from L. ma-jor in all seven samples tested, cells from two donorswere not activated by PSA-2 from L. donovani. TheE. coli-derived recombinant amastigote fusion pro-
FEMSIM 853 27-3-98
Fig. 2. Net proliferation ([3H]thymidine incorporation) by PBMCfrom Sudanese individuals with a history of self-healing cutane-ous leishmaniasis after incubation with PSA-2 isolated fromL. major promastigotes (R), PSA-2 from L. donovani promasti-gotes (S), and recombinant amastigote PSA-2 (F).
Fig. 1. Net proliferation ([3H]thymidine incorporation) by PBMCfrom presumably unexposed Danes (open symbols) and Sudaneseindividuals with a history of self-healing cutaneous leishmaniasis(¢lled symbols) after incubation with PSA-2 isolated fromL. major promastigotes (O, R), PPD (7, 8) or tetanus toxoid(a, b).
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218212
tein induced proliferation in cells from one of theseven donors only.
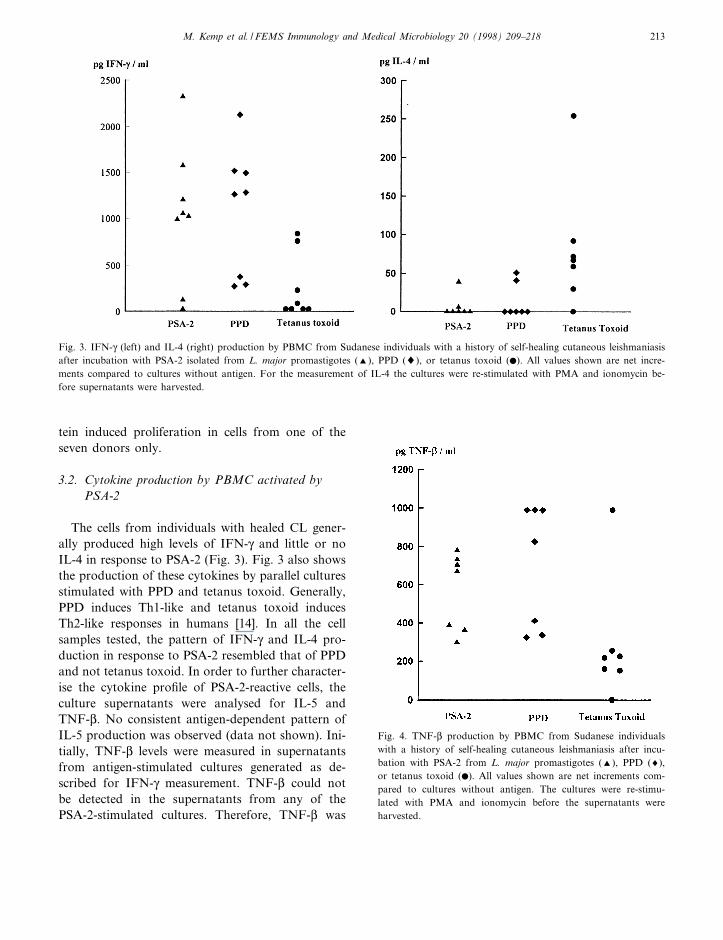
3.2. Cytokine production by PBMC activated byPSA-2
The cells from individuals with healed CL gener-ally produced high levels of IFN-Q and little or noIL-4 in response to PSA-2 (Fig. 3). Fig. 3 also showsthe production of these cytokines by parallel culturesstimulated with PPD and tetanus toxoid. Generally,PPD induces Th1-like and tetanus toxoid inducesTh2-like responses in humans [14]. In all the cellsamples tested, the pattern of IFN-Q and IL-4 pro-duction in response to PSA-2 resembled that of PPDand not tetanus toxoid. In order to further character-ise the cytokine pro¢le of PSA-2-reactive cells, theculture supernatants were analysed for IL-5 andTNF-L. No consistent antigen-dependent pattern ofIL-5 production was observed (data not shown). Ini-tially, TNF-L levels were measured in supernatantsfrom antigen-stimulated cultures generated as de-scribed for IFN-Q measurement. TNF-L could notbe detected in the supernatants from any of thePSA-2-stimulated cultures. Therefore, TNF-L was
FEMSIM 853 27-3-98
Fig. 4. TNF-L production by PBMC from Sudanese individualswith a history of self-healing cutaneous leishmaniasis after incu-bation with PSA-2 from L. major promastigotes (R), PPD (8),or tetanus toxoid (b). All values shown are net increments com-pared to cultures without antigen. The cultures were re-stimu-lated with PMA and ionomycin before the supernatants wereharvested.
Fig. 3. IFN-Q (left) and IL-4 (right) production by PBMC from Sudanese individuals with a history of self-healing cutaneous leishmaniasisafter incubation with PSA-2 isolated from L. major promastigotes (R), PPD (8), or tetanus toxoid (b). All values shown are net incre-ments compared to cultures without antigen. For the measurement of IL-4 the cultures were re-stimulated with PMA and ionomycin be-fore supernatants were harvested.
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218 213

FEMSIM 853 27-3-98
Fig. 5. Expression of intracellular IFN-Q (x-axis) and surface CD3 (y-axis) in PBMC cultured with and without PSA-2 for 7 days. Individ-ual limits for positive signals were de¢ned from isotype control antibodies. Numbers are the percentages of cells in each population.
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218214

subsequently measured in the supernatants from cul-tures that had been pulsed with PMA and ionomycinby the method previously described for measurementof IL-4. As shown in Fig. 4, in these cultures PPDand PSA-2 both induced higher values than tetanustoxoid did, except for cells from one of the sevendonors.
3.3. Flow cytometry
The phenotype of the cells activated by PSA-2 wasevaluated by £ow-cytometric analysis of forwardscatter signal and CD3 expression. In PBMC fromall of the ¢ve donors with a history of self-healingCL tested, incubation with PSA-2 for 7 days inducedincreased forward scatter signal predominantly in theCD3-positive cell population (data not shown). Theexpression of CD3 on cells containing intracellularIFN-Q was assessed by £ow cytometry (Fig. 5). In alldonors tested the majority of the cells expressingintracellular IFN-Q after incubation with PSA-2also expressed CD3. Low-grade staining for IFN-Qcould be detected in less than 6% of antigen-stimu-lated cell populations when stained without priorpermeabilisation, con¢rming the intracellular loca-tion of this cytokine.
4. Discussion
There is a great need for identi¢cation of parasitecomponents that can be incorporated into futurevaccines against human leishmaniasis. Immunisationof mice with the parasite antigen PSA-2 has provideda high degree of protection against challenge infec-tion with L. major. Protection against L. amazonensishas been obtained in mice by immunisation withpuri¢ed gp46, a homologue of PSA-2 [15]. Recombi-nant vaccinia virus expressing gp46 was also e¡ectivein inducing T-cell responses and protection againstL. amazonensis in mice [16]. A major question for aLeishmania vaccine is its ability to induce and main-tain a Th1 type of response in humans. In this studywe attempted to correlate the T-cell memory in im-mune individuals with their ability to respond to aLeishmania antigen which is a vaccine candidatebased on animal studies.
We found that lymphocytes from immune donors
proliferated in response to incubation with PSA-2.By contrast, cells from presumably non-exposedDanish donors did not respond to the antigen, indi-cating that the PSA-2-speci¢c cells in the immuneindividuals had been expanded in response to theinfection with L. major.
Observations in humans and mice indicate thatvaccine candidates need to induce speci¢c activationof Th1-like cells. Moreover, they should not induceTh2 responses, since Th2 responses may contributeto the pathology of the disease in humans [1]. Inorder to characterise the cells recognising PSA-2 ac-cording to the Th1/Th2 concept, IFN-Q and IL-4were measured in supernatants of PSA-2-stimulatedcells. The cells produced high levels of IFN-Q and noor little IL-4 as did PPD-stimulated cultures. TheTh1-like pattern in these cultures was in markedcontrast to the high IL-4/low IFN-Q seen in tetanustoxoid-stimulated cultures from the same donors. Tofurther characterise the cytokine response by thecells, the levels of TNF-L and IL-5 were determinedin the supernatants. The Th1-associated cytokineTNF-L was not detectable in antigen-stimulated cul-tures. We therefore applied the PMA/ionomycin am-pli¢cation step previously used for determination ofIL-4 production [14]. With use of this technique de-tectable levels of TNF-L were obtained in the anti-gen-stimulated cultures. With one exception, therewas a clear antigen-dependent distribution of TNF-L with high levels in PPD-stimulated cultures andlow levels in cultures incubated with tetanus toxoid.Thus, the PMA/ionomycin ampli¢cation step issuited for determination of Th1- as well as Th2-as-sociated cytokines secreted in small quantities. As forIFN-Q, TNF-L was produced in relatively largeamounts by cells incubated with PSA-2 comparedto tetanus toxoid-stimulated cells. The associationbetween TNF-L and IFN-Q is in accordance withour previous observations of co-expression of thesetwo cytokines in Leishmania-reactive CD4� T-cellclones [17].
It was not possible to con¢rm the association ofIL-5 with Th2 responses as de¢ned by IFN-Q and IL-4. This lack of association was not restricted to PSA-2, but was also seen in cultures incubated with thecontrol antigens PPD and tetanus toxoid. Since bothIL-4 and TNF-L showed better antigen-dependentdiscrimination after ampli¢cation with PMA/iono-
FEMSIM 853 27-3-98
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218 215

mycin, IL-5 was also measured in supernatants ofcultures treated with PMA/ionomycin as describedfor IL-4 measurement. Higher levels of IL-5 wereobtained by this procedure, but still there was noclear antigen dependence (data not shown). Thus,in the present system, IL-5 did not seem to ¢t intothe Th1/Th2 `package' [18]. Studies at the single celllevel have suggested that IL-4 and IL-5 are producedby di¡erent cells in short-term cultures of humanPBMC [19].
The identity of the cells activated by PSA-2 wasestablished by £ow cytometry. Forward scatter sig-nal, re£ecting cell size, was increased in cells incu-bated with PSA-2. Increased size was predominantlyobserved within the CD3� T-cell population, indicat-ing PSA-2-induced blastogenesis predominantly orsolely in T cells.
IFN-Q is both a marker of activation of Th1 cellsand a powerful inducer of resistance against intra-cellular Leishmania parasites in macrophages. It istherefore essential to identify the cells that produceIFN-Q. Flow-cytometric analysis after staining forintracellular cytokine has become an important toolfor investigations of cytokine production at the sin-gle cell level [20]. We have applied the technique forantigen-stimulated cultures to identify the cellularsource of Leishmania-induced IFN-Q production[21]. The present results show that in PBMC incu-bated for 1 week with PSA-2, intracellular IFN-Q isfound predominantly in CD3-positive cells. It is rea-sonable to assume that IFN-Q at this time wasmainly produced by T cells in the cultures.
While PSA-2 varies quite considerably betweenspecies and developmental stages of Leishmania,there is a good degree of antigenic cross-reactivityas detected by monoclonal and polyclonal antibodies[11]. In mice good protection against infection withL. major has been found after immunisation withPSA-2 from L. major, as well as PSA-2 fromL. donovani (Handman, unpublished). To evaluatethe cross-recognition of PSA-2 from L. major andL. donovani by human T cells, the proliferative re-sponses were compared in PBMC incubated withPSA-2 from promastigotes of the two species.L. donovani is not endemic in the part of Sudanwhere the donors live, and there is no reason tobelieve that they have been exposed to L. donovani.Cells from some, but not all, of the immune donors
recognised PSA-2 from L. donovani in addition tothe L. major-derived antigen. In another set ofexperiments, PBMC were collected from Sudanesedonors 2 months after initiation of treatment ofpost-kala-azar dermal leishmaniasis caused byL. donovani. Cells from three out of eight of thesedonors proliferated in response to PSA-2 (data notshown). The cells responded to PSA-2 from bothL. major and L. donovani and the proliferative re-sponses to the two antigens were of the same mag-nitude.
The recombinant amastigote protein expressed byE. coli only activated cells from one of the donorstested. The lack of response to the recombinant pro-tein may be due to di¡erent processing and presen-tation of native and E. coli-derived protein by theantigen-presenting cells. Similar data have been ob-tained in the mouse model for leishmaniasis [9]where T cells from immune animals vaccinatedwith PSA-2 or recovered from infection did not rec-ognise E. coli-derived PSA-2. However, the yeast-de-rived recombinant PSA-2 or recombinant PSA-2 de-rived from transfected promastigotes was recognised.Recent studies have suggested that glycosylation maybe a very important factor in de¢ning T-cell epitopes[22,23]. Interestingly, vaccination of mice with re-combinant PSA-2 puri¢ed from E. coli did not con-fer protection against challenge infection, whereasrecombinant PSA-2 derived from L. mexicana did[9]. A similar discrepancy in the T-cell recognitionof proteins expressed by E. coli and Leishmania para-sites has been observed in another major glycopro-tein antigen, the surface protease gp63. In mice, pro-tection could be obtained by vaccination with nativebut not with E. coli-derived recombinant gp63 [24].Human T-cell lines recognising native gp63 and notresponding to E. coli-derived recombinant gp63 havebeen reported [25]. The recombinant vaccinia virusgp46, which protected mice against L. amazonensis,was glycosylated [16].
In this study we show that individuals naturallyimmune to leishmaniasis display a Th1 responseupon re-stimulation with the molecularly de¢nedPSA-2 antigen. This indicates that a Th1-type mem-ory has been ¢xed after natural infection and cureand is maintained in the immune individual. In ad-dition, the presence of Th1-type memory to PSA-2 inhumans, added to its demonstrated ability to protect
FEMSIM 853 27-3-98
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218216

in the mouse model, makes this antigen an attractivevaccine candidate for humans.
Acknowledgments
This investigation received ¢nancial support fromThe Danish Biotechnology Program, The Novo-Nordisk Foundation, and the Danish InternationalDevelopment Agency (DANIDA). E. Handman issupported by The Australian National Health andMedical Research Council and the Collaborative Re-search Centre for Vaccine Technology. The authorsare indebted to Anders Sjoë lander for critical reviewof the manuscript.
References
[1] Kemp, M., Theander, T.G. and Kharazmi, A. (1996) Thecontrasting roles of CD4+ T cells in intracellular infectionsin humans: Leishmaniasis as an example. Immunol. Today 17,13^16.
[2] Kemp, M., Hey, A.S., Kurtzhals, J.A., Christensen, C.B.,Gaafar, A., Mustafa, M.D., Kordofani, A.A., Ismail, A.,Kharazmi, A. and Theander, T.G. (1994) Dichotomy of thehuman T cell response to Leishmania antigens. I. Th1-likeresponse to Leishmania major promastigote antigens in indi-viduals recovered from cutaneous leishmaniasis. Clin. Exp.Immunol. 96, 410^415.
[3] Kemp, M., Hey, A.S., Bendtzen, K., Kharazmi, A. and The-ander, T.G. (1994) Th1-like human T-cell clones recognizingLeishmania gp63 inhibit Leishmania major in human macro-phages. Scand. J. Immunol. 40, 629^635.
[4] Gaafar, A., Kharazmi, A., Ismail, A., Kemp, M., Hey, A.,Christensen, C.B.V., Dafalla, M., El-Kadaro, A.Y., El-Has-san, A.M. and Theander, T.G. (1995) Dichotomy of the T-cellresponse to Leishmania antigens in patients su¡ering fromcutaneous leishmaniasis : absence or scarcity of Th1 activityis associated with severe infections. Clin. Exp. Immunol. 100,239^245.
[5] Morris, L., Aebisher, T., Handman, E. and Kelso, A. (1993)Resistance of BALB/c mice to Leishmania major infection isassociated with a decrease in the precursor frequency of anti-gen-speci¢c CD4+ cells secreting interleukin-4. Int. Immunol.5, 761^767.
[6] Morris, L., Troutt, A., Handman, E. and Kelso, A. (1996)Changes in the precursor frequencies of IL-4 and IFN-gammasecreting CD4+ cells correlate with resolution of lesions inmurine cutaneous leishmaniasis. J. Immunol. 149, 2715^2721.
[7] Murphy, E.E., Shibuya, K., Hosken, N.A., Openshaw, P.,Maino, V., Davis, K., Murphy, K. and O'Garra, A. (1996)
Reversibility of T helper 1 and 2 populations is lost after longterm stimulation. J. Exp. Med. 183, 901^913.
[8] O'Garra, A. and Murphy, K. (1994) Role of cytokines indetermining T-lymphocyte function. Curr. Opin. Immunol.6, 466.
[9] Handman, E., Symons, F.M., Baldwin, T.M., Curtis, J.M.and Scheerlinck, J.P. (1995) Protective vaccination with pro-mastigote surface antigen 2 from Leishmania major is medi-ated by a TH1 type of immune response. Infect. Immun. 63,4261^4267.
[10] Murray, P.J., Spithill, T.W. and Handman, E. (1989) ThePSA-2 glycoprotein complex of Leishmania major is a glyco-sylphosphatidylinositol-linked promastigote surface antigen.J. Immunol. 143, 4221^4226.
[11] Symons, F.M., Murray, P.J., Ji, H., Simpson, R.J., Osborn,A.H., Cappai, R. and Handman, E. (1994) Characterizationof a polymorphic family of integral membrane proteins inpromastigotes of di¡erent Leishmania species. Mol. Biochem.Parasitol. 67, 103^113.
[12] McMahon Pratt, D., Traub-Cseko, Y., Lohman, K.L., Rog-ers, D.D. and Beverley, S.M. (1992) Loss of the GP46/M-2surface glycoprotein gene family in the Leishmania braziliensiscomplex. Mol. Biochem. Parasitol. 50, 151^160.
[13] Kadaro, A.Y., Ghalib, H.W., Ali, M.S., El-Toum, I., Ismail,A., Gaafar, A., Kemp, M., Kordofani, A.A.Y., Reed, S., El-Hassan, A.M., Kharazmi, A., Hag-Ali, M. and Mustafa,M.D. (1993) Prevalence of cutaneous leishmaniasis along theNile river North of Khartoum (Sudan) in the aftermath of anepidemic in 1985. Am. J. Trop. Med. Hyg. 48, 44^49.
[14] Kurtzhals, J.A.L., Hansen, M.B., Hey, A. and Poulsen, L.K.(1992) Measurement of antigen-dependent interleukin-4 pro-duction by human peripheral blood mononuclear cells ^ In-troduction of an ampli¢cation step using ionomycin and phor-bol myristate acetate. J. Immunol. Methods 156, 239^245.
[15] Champsi, J. and McMahon Pratt, D. (1988) Membrane gly-coprotein M-2 protects against Leishmania amazonensis infec-tion. Infect. Immun. 52, 3272^3279.
[16] McMahon Pratt, D., Rodriguez, D., Rodriguez, J.-R., Zhang,Y., Manson, K., Bergman, C., Rivas, L., Rodriguez, J.F.,Lohman, K.L., Ruddle, N.H. and Esteban, M. (1993) Re-combinant vaccinia viruses expressing gp46/M-2 protectagainst Leishmania infection. Infect. Immun. 61, 3351^3359.
[17] Kemp, M., Kurtzhals, J.A., Bendtzen, K., Poulsen, L.K.,Hansen, M.B., Koech, D.K., Kharazmi, A. and Theander,T.G. (1993) Leishmania donovani-reactive Th1- and Th2-like T-cell clones from individuals who have recovered fromvisceral leishmaniasis. Infect. Immun. 61, 1069^1073.
[18] Kelso, A. (1995) Th1 and Th2 subsets: paradigm lost? Immu-nol. Today 16, 374^379.
[19] Jung, T., Schauer, U., Rieger, C., Wagner, K., Einsle, K.,Neumann, C. and Heusser, C. (1995) Interleukin-4 and inter-leukin-5 are rarely co-expressed by human T cells. Eur.J. Immunol. 25, 2413^2416.
[20] Schauer, U., Jung, T., Krug, N. and Frew, A. (1996) Measure-ment of intracellular cytokines. Immunol. Today 17, 305^306.
[21] Kemp, K., Hviid, L., Kharazmi, A. and Kemp, M. (1997)Interferon-gamma production by T cells and NK cells in vitro
FEMSIM 853 27-3-98
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218 217

in response to antigens from the two intracellular pathogensMycobacterium tuberculosis and Leishmania major. Scand.J. Immunol. 46, 495^499.
[22] Haurum, J.S., Arsequell, G., Lellouch, A.C., Wong, S.Y.C.,Dwek, R.A., McMichael, A.J. and Elliott, T. (1994) Recog-nition of carbohydrate by major histocompatibility complexclass I-restricted, glycopeptide-speci¢c cytotoxic T lympho-cytes. J. Exp. Med. 180, 739^744.
[23] Jensen, T., Galli-Stampino, L., Mouritsen, S., Frische, K.,Peters, S., Meldal, M. and Werdelin, O. (1996) T cell recog-
nition of Tn-glycosylated peptide antigens. Eur. J. Immunol.26, 1342^1349.
[24] Handman, E., Button, L.L. and McMaster, R.W. (1996)Leishmania major : production of recombinant gp63, its anti-genicity and immunogenicity in mice. Exp. Parasitol. 70, 427^435.
[25] Button, L.L., Reiner, N.E. and McMaster, R.W. (1991) Mod-i¢cation of gp63 genes from diverse species of Leishmania forexpression of recombinant protein at high levels in Escherichiacoli. Mol. Biochem. Parasitol. 44, 213^224.
FEMSIM 853 27-3-98
M. Kemp et al. / FEMS Immunology and Medical Microbiology 20 (1998) 209^218218
Related Documents











