THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 255. No. 15, Issue of August 10. pp. 7474-7479.1980 Prrnted in L’. S. A. Progesterone and Estrogen Control of Rates of Synthesis of Uterine Cathepsin D* (Received for publication, January 21, 1980) Sudha Elangovan and Bruce C. Moulton From the DeDartments of Obstetrics and Gvnecolom and of Biological Chemisty, University of Cincinnati, College of Medicine, Ckcinnati, Ohio 45267 ” Changes in cathepsin D activity (EC 3.4.23.5) in the mammalian uterus areassociated with changes in the balance of intracellular protein synthesis and degra- dation. To examine the hormonal control of uterine levels of cathepsin D, rates of synthesis of the enzyme protein were determined in ovariectomized rat uteri following estrogen or progesterone treatment. Rates of incorporation of [3HJIeucine into cathepsin D protein were determined using immunoprecipitation of the spe- cific enzyme protein from uterine homogenates. The antigen used to develop a specific goat antiserum was cathepsin D protein purified from rat spleen by acid dialysis, fractionation with ammonium sulfate and ace- tone, and affinity chromatography on pepstatin-Seph- arose 4B. The specificity of the goat immune serum was determined by double diffusion analysis and by sodium dodecyl sulfate-polyacrylamide gel electrophoresis of [3H]leucine-labeleduterine supernatant proteins pre- cipitated by the goat immune serum. The amount of radioactivity in the single uterine protein (Mr = 43,000) precipitated by the antiserum was used to calculate rates of uterine cathepsin D synthesis after estrogen or progestin treatment. Estradiol increased cathepsin D synthesis at the same rate as general protein synthesis after a lag period of 12 h. Progesterone and medroxy- progesterone acetate, however, specifically increased the rate of cathepsin D synthesis to a maximum at 6 h. To determine whether progestins increased cathepsin D content of specific tissues in the uterus, the antiserum to cathepsin D was used to localize the enzyme protein by immunohistochemical staining. Medroxyprogester- one acetate treatment greatly increased levels of ca- thepsin D protein detected in the endometrium. These data demonstrate that progestins specifically increase rates of endometrial cathepsin D synthesis indicating a physiological role in the maintenance of lysosomal function in this tissue. Considerable evidence has accumulated implicating the ly- sosome as the predominant site of intracellular protein deg- radation (1). Particularly under conditions of nutritional dep- rivation, increased ceUular autophagic activity is associated with increased rates of proteolysis which can be suppressed by compounds which inhibit lysosomal function (2,3). In liver, the control of rates of protein degradation by glucagon or insulin depends upon the control of rates of protein internal- ization within vacuolar components of the lysosomal system * This work was supported by Grants HD-07255 and HD-10721 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section I734 solely to indicate this fact. mediated by intracellular depletion of amino acids (4). Control of lysosomal protein degradation depends upon not only the control of rates of internalization of cytoplasmic protein but also the control of the lysosomal content of hydrolytic en- zymes. Other studies have shown that compounds such as leupeptin and pepstatin which inhibit the activity of cathepsin Bl and D also suppress protein degradation in several mam- malian c e b and tissues (5-10). Concurrent changes in cathep- sin BI and D activity with changes in rates of proteolysis have been observed (1 1). Increases in cathepsin D (EC 3.4.23.5), a major lysosomal proteinase, would increase the capacity of mammalian cells for protein degradation. Estrogen and progesterone increase levels of cathepsin D activity in the uterus (12, 13). Increased cellular lysosomal activity observed during pinocytosis by uterine epithelial cells (14), cellular autophagic activity (15), and the differentiation of stromal cells (16) may involve in- creased rates of intracellular protein degradation and require increases in cathepsin D. Significant changes are observed in cathepsin D activity during blastocyst implantation (12) and postpartum involution of the uterus (17). In this study, im- munochemical procedures have been developed so that the control of rates of cathepsin D synthesis by ovarian hormones could be examined. Estrogen treatment of ovariectomized rats increased rates of cathepsin D synthesis, but more specific increases in the synthesis of this enzyme wereobserved after progestin treatment, which resulted in greater levels of the enzyme protein in the uterine endometrium. EXPERIMENTAL PROCEDURES Materials-Aminohexyl (AH)-Sepharose was purchased from Pharmaeia, pepstatin from Protein Research Foundation, Japan, dicyclohexylcarbodimidefrom Eastman Kodak, acrylamide, bisacryl- amide, N,N,N’N’-tetramethylethylenediamine from Bio-Rad Labo- ratories, BSA,’ a-lactalbumin, ovalbumin, pepsin, trypsin, and SDS from Sigma, Freund‘s complete adjuvant from Difco, immunodiffu- sion plates from Meloy Laboratories, hemoglobin from Worthington. AU other chemicals were of analytical grade and obtained from various sources. Animals-Mature female rats (150 to 175 g) (Harlan/SD, Indian- apolis) maintained on Purina Lab Chow and water ad libitum were ovariectomized 3 to 4 weeks prior to use in experiments. Estradiol benzoate (5 pg/rat) or progesterone (2 mg/rat) was injected subcu- taneously in oil. Medroxyprogesterone acetate (2 mg/rat) was injected subcutaneously as an aqueous suspension. Purification of Enzyme-Cathepsin D was purified from rat spleen using acid pH precipitation, ammonium sulfate andacetone fraction- ation, and affinity chromatography on pepstatin-Sepharose (18-20). SDS-polyacrylamide gel electrophoresis (21) of the purified enzyme revealed a major protein with a M, of approximately 42,000 to 44 , OOO . ‘ The abbreviations used are: BSA, bovine serum albumin; SDS, sodium dodecyl sulfate; MPA, medroxyprogesterone acetate, pregn- 4-ene-3, 20-dione, 17-(acetyloxy)-6-methyl-, (64.. 1414

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 255. No. 15, Issue of August 10. pp. 7474-7479.1980 Prrnted in L’. S. A.
Progesterone and Estrogen Control of Rates of Synthesis of Uterine Cathepsin D*
(Received for publication, January 21, 1980)
Sudha Elangovan and Bruce C. Moulton From the DeDartments of Obstetrics and Gvnecolom and of Biological Chemisty, University of Cincinnati, College of Medicine, Ckcinnati, Ohio 45267
”
Changes in cathepsin D activity (EC 3.4.23.5) in the mammalian uterus are associated with changes in the balance of intracellular protein synthesis and degra- dation. To examine the hormonal control of uterine levels of cathepsin D, rates of synthesis of the enzyme protein were determined in ovariectomized rat uteri following estrogen or progesterone treatment. Rates of incorporation of [3HJIeucine into cathepsin D protein were determined using immunoprecipitation of the spe- cific enzyme protein from uterine homogenates. The antigen used to develop a specific goat antiserum was cathepsin D protein purified from rat spleen by acid dialysis, fractionation with ammonium sulfate and ace- tone, and affinity chromatography on pepstatin-Seph- arose 4B. The specificity of the goat immune serum was determined by double diffusion analysis and by sodium dodecyl sulfate-polyacrylamide gel electrophoresis of [3H]leucine-labeled uterine supernatant proteins pre- cipitated by the goat immune serum. The amount of radioactivity in the single uterine protein (Mr = 43,000) precipitated by the antiserum was used to calculate rates of uterine cathepsin D synthesis after estrogen or progestin treatment. Estradiol increased cathepsin D synthesis at the same rate as general protein synthesis after a lag period of 12 h. Progesterone and medroxy- progesterone acetate, however, specifically increased the rate of cathepsin D synthesis to a maximum at 6 h. To determine whether progestins increased cathepsin D content of specific tissues in the uterus, the antiserum to cathepsin D was used to localize the enzyme protein by immunohistochemical staining. Medroxyprogester- one acetate treatment greatly increased levels of ca- thepsin D protein detected in the endometrium. These data demonstrate that progestins specifically increase rates of endometrial cathepsin D synthesis indicating a physiological role in the maintenance of lysosomal function in this tissue.
Considerable evidence has accumulated implicating the ly- sosome as the predominant site of intracellular protein deg- radation (1). Particularly under conditions of nutritional dep- rivation, increased ceUular autophagic activity is associated with increased rates of proteolysis which can be suppressed by compounds which inhibit lysosomal function (2,3). In liver, the control of rates of protein degradation by glucagon or insulin depends upon the control of rates of protein internal- ization within vacuolar components of the lysosomal system
* This work was supported by Grants HD-07255 and HD-10721 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section I734 solely to indicate this fact.
mediated by intracellular depletion of amino acids (4). Control of lysosomal protein degradation depends upon not only the control of rates of internalization of cytoplasmic protein but also the control of the lysosomal content of hydrolytic en- zymes. Other studies have shown that compounds such as leupeptin and pepstatin which inhibit the activity of cathepsin Bl and D also suppress protein degradation in several mam- malian c e b and tissues (5-10). Concurrent changes in cathep- sin BI and D activity with changes in rates of proteolysis have been observed (1 1).
Increases in cathepsin D (EC 3.4.23.5), a major lysosomal proteinase, would increase the capacity of mammalian cells for protein degradation. Estrogen and progesterone increase levels of cathepsin D activity in the uterus (12, 13). Increased cellular lysosomal activity observed during pinocytosis by uterine epithelial cells (14), cellular autophagic activity (15), and the differentiation of stromal cells (16) may involve in- creased rates of intracellular protein degradation and require increases in cathepsin D. Significant changes are observed in cathepsin D activity during blastocyst implantation (12) and postpartum involution of the uterus (17). In this study, im- munochemical procedures have been developed so that the control of rates of cathepsin D synthesis by ovarian hormones could be examined. Estrogen treatment of ovariectomized rats increased rates of cathepsin D synthesis, but more specific increases in the synthesis of this enzyme were observed after progestin treatment, which resulted in greater levels of the enzyme protein in the uterine endometrium.
EXPERIMENTAL PROCEDURES
Materials-Aminohexyl (AH)-Sepharose was purchased from Pharmaeia, pepstatin from Protein Research Foundation, Japan, dicyclohexylcarbodimide from Eastman Kodak, acrylamide, bisacryl- amide, N,N,N’N’-tetramethylethylenediamine from Bio-Rad Labo- ratories, BSA,’ a-lactalbumin, ovalbumin, pepsin, trypsin, and SDS from Sigma, Freund‘s complete adjuvant from Difco, immunodiffu- sion plates from Meloy Laboratories, hemoglobin from Worthington. AU other chemicals were of analytical grade and obtained from various sources.
Animals-Mature female rats (150 to 175 g) (Harlan/SD, Indian- apolis) maintained on Purina Lab Chow and water ad libitum were ovariectomized 3 to 4 weeks prior to use in experiments. Estradiol benzoate (5 pg/rat) or progesterone (2 mg/rat) was injected subcu- taneously in oil. Medroxyprogesterone acetate (2 mg/rat) was injected subcutaneously as an aqueous suspension.
Purification of Enzyme-Cathepsin D was purified from rat spleen using acid pH precipitation, ammonium sulfate and acetone fraction- ation, and affinity chromatography on pepstatin-Sepharose (18-20). SDS-polyacrylamide gel electrophoresis (21) of the purified enzyme revealed a major protein with a M, of approximately 42,000 to 44,OOO.
‘ The abbreviations used are: BSA, bovine serum albumin; SDS, sodium dodecyl sulfate; MPA, medroxyprogesterone acetate, pregn- 4-ene-3, 20-dione, 17-(acetyloxy)-6-methyl-, (64..
1414

Hormonal Control of Rates of Uterine Cathepsin D Synthesis 7475
A faint additional band at a M, of 25,000 to 28,000 has also been observed by others using pepstatin-Sepharose purification of rat spleen cathepsin D (22). Cathepsin D activity was assayed by diges- tion of hemoglobin at pH 3.2 (12, 18). One unit of enzyme activity was defined as the amount of enzyme producing a final absorbance of 1.0 a t 660 nM after a IO-min digestion a t 37°C. Protein was determined by the method of Lowry et al. (23) using crystalline BSA as standard.
Preparation of Antiserum-Purified cathepsin D (920 pg, 153 enzyme activity units/mg of protein) was emulsified with equal vol- umes of Freund’s complete adjuvant and injected subcutaneously to initiate antibody production in a female goat. Booster doses (690 pg) were given in the same manner 1 week and 3 weeks later. Serum was collected before immunization to provide control serum and 4 weeks after the fmt injection of antigen. One unit of either uterine or spleen enzyme activity was precipitated by 63 pl of serum as determined by antibody titration. Double diffusion analysis was performed as de- scribed by Ouchterlony (24) using immunodiffusion plates.
Measurement of Uterine Cathepsin D Synthesis-Groups of 6 or 12 rats were given intrauterine applications of 32 pCi of [“Hlleucine (140 to 160 Ci/mmol, New England Nuclear) by transcervical injection (25) a t 0 and 1 h and killed a t 2 h. Uteri were removed, trimmed of extraneous connective tissue, and flushed with cold 0.15 M NaCl to remove unincorporated amino acid remaining in the lumen before freezing between two blocks of dry ice and stored a t -7OOC. Frozen uteri from each group were pooled and homogenized in 0.15 M NaCl using a Polytron with a PTIOST probe generator. After removal of samples for enzyme assay and for determination of acid-soluble and -insoluble radioactivity, cathepsin D enzyme protein of each homog- enate was partially purified through the ammonium sulfate step of the enzyme purification procedure. The ammonium sulfate precipi- tate was dissolved in 408 of the original homogenate volume in 50 mM phosphate buffer, pH 7.0, and assayed for enzyme activity to calculate recovery. After Centrifugation and filtration through a mem- brane filter (Metricel, 1.2-pm pore, Gelman), 50 pg of BSA was added to each sample. followed by 500pl of anti-BSA antibody and incubated overnight a t 4OC. Preprecipitation of the uterine supernatant with added BSA and anti-BSA antibody eliminated nonspecific precipita- tion by the anti-cathepsin D antibody. After centrifugation and filtra- tion to remove the immunoprecipitate, anti-cathepsin D serum was added equivalent to the enz.yme activity measured after the ammo- nium sulfate step. The immune reaction was incubated overnight a t 4°C and centrifuged; the supernatant was assayed for unprecipitated cathepsin D, and the pellet was washed three times with 0.15 M NaCl containing 0.1 M leucine. The immunoprecipitate was then solubilized and dissociated into subunits before electrophoresis on SDS-poly- acrylamide gels. Gels were sliced into 2-mm slices and incubated with tissue solubilizer overnight a t 37°C before addition of a toluene-based scintillation mixture and counting in a liquid scintillation counter.
An aliquot of the uterine homogenate (100 pl) was precipitated by the addition of 5 ml of cold 10% trichloroacetic acid for the determi- nation of acid-precipitable and acid-soluble counts. Acid-precipitable protein was washed five times with 5 8 trichloroacetic acid containing 0.1 M leucine on a thick glass filter (Gelman Instrument Co.). After the last wash, the filter with the precipitate was digested overnight a t 37°C with tissue solubilizer and counted after the addition of scintil- lation mixture. The acid-soluble fraction was assayed for total amino acid content by the ninhydrin reaction and for radioactivity after the addition of 6 ml of Instagel (Packard) to 4 ml of the supernatant. To determine whether progesterone or estradiol treatment altered the specific radioactivity of leucine in the intracellular amino acid pool, the specific radioactivity of the leucine was measured at various times after hormone treatment. Leucine in the acid-soluble fraction was separated by thin layer chromatography and assayed. using a sensitive isotope dilution procedure with a range of 100 to 1500 pmol (26) before the radioactivity was determined. The amount of radioactivity incorporated into cathepsin D protein was the sum of the radioactivity in the enzyme peak corrected for the recovery of the enzyme during partial purification (79.1 f 0.3%, S.E.) based on enzyme activity. To determine the rate of cathepsin D synthesis, the amount of radioac- tivity in the enzyme peak was expressed per cpm/pmol of amino acid in the acid-soluble fraction or per cpm/mg of protein in the acid- insoluble fraction.
Immunohistochemical Localization of Cathepsin D-Uterine tis- sue samples for the immunohistochemical localization of cathepsin D were excised, frozen in a hexane-dry ice mixture, and stored a t -70°C before 10-pm sections were cut with a cryostat a t -20°C. Sections on slides were incubated in acetone a t -10°C for 6 to 8 min and then air- dried. Goat antiserum to cathepsin D and preimmune serum were
diluted 1:W with FA buffer (Difco), and 100 pl of either diluted serum was incubated with tissue sections a t room temperature for 1 h. Slides were then washed three times with FA buffer before incu- bation with loop1 of fluorescein-conjugated rabbit anti-goat 7 S globin (IgG, Meloy Laboratories) a t room temperature for 1 h. After three washes with FA buffer to remove unreacted rabbit IgG, sections were mounted in FA mounting fluid (Difco). Slides were stored overnight a t -70°C and then examined with a Zeiss Photomicroscope I1 equipped with phase contrast and epifluorescence optics. Phase con- trast microscopy was used to scan histological sections before exam- ination for fluorescence. Photographs were taken on Kodak Tri-X film (ASA 400) with constant exposure times and printed on Polycon- trast Rapid I1 RCF Kodak paper with constant exposure times.
RESULTS
Immunological Similarity between Rat Spleen and Rat Uterine Cathepsin D-Cathepsin D protein was purified from rat spleen tissue for use as an antigen to develop an antiserum to uterine cathepsin D. Although cathepsin D had been puri- fied from bovine uterus (21), the amount and specific activity of the enzyme in rat spleen were greater than in uterine tissue. Previous studies of cross-reactivity of an antiserum to chicken liver cathepsin D (27) offered some assurance that an antise- rum to rat spleen cathepsin D would also precipitate the uterine enzyme. Determinations of rates of synthesis of uterine cathepsin D required that the antiserum specifically precipi- tate the uterine enzyme from a mixture of radiolabeled pro- teins. Ouchterlony double diffusion analysis was used to de- termine the immunological similarity of the spleen and uterine cathepsin D proteins. As shown in Fig. 1, various concentra- tions of antiserum and y-globulin to rat spleen cathepsin D showed a single precipitin band without “spurs” after reaction with partially purified uterine cathepsin D. These observa- tions indicated that the antiserum was specific to a single uterine protein, and that the spleen and uterine enzymes were immunologically similar. Immunotitration of the antiserum with partially purified rat uterine or spleen cathepsin D gave the same immunological equivalence point providing further evidence of the immunological similarity of the proteins.
Specificity of Immunoprecipitation of Uterine Cathepsin D-To determine the specificity of the immunoprecipitate,
FIG. 1. Ouchterlony diffusion analysis of uterine cathepsin D interaction with antiserum to spleen cathepsin D. The center well contained partially purified uterine cathepsin D (0.023 enzyme unit). Surrounding wells contained the following: cathepsin D anti- serum, 0 dilution (well I ) , cathepsin D antiserum, ‘,$X (well 2). or cathepsin D antiserum y-globulin. 0 dilution (well 3). Ihx (well 4) . and %X (well 5) . Gels were incubated a t 25°C at pH 7.0 using PO,- buffered saline, washed with 0.15 M NaCl for 4 h to remove unprecip- itated protein, and stained with 0.1% Coomassie brilliant blue R for 1 h before destaining.
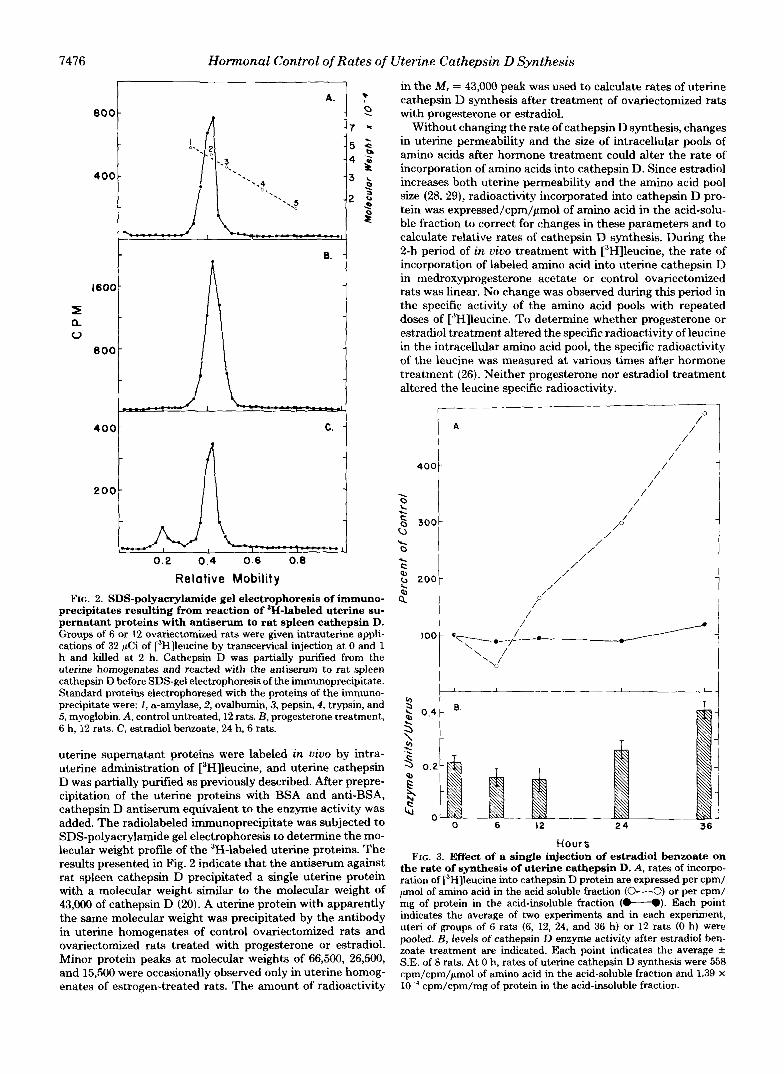
7476 Hormonal Control of Rates of Uterine Cathepsin D Synthesis
A.
L
4001 C.
"
0.2 0.4 0.6 0.0
1
- ! d
- 2
L
L
L
Relative Mobility FIG. 2. SDS-polyacrylamide gel electrophoresis of immuno-
precipitates resulting from reaction of 3H-labeled uterine su- pernatant proteins with antiserum to rat spleen cathepsin D. Groups of 6 or 12 ovariectomized rats were given intrauterine appli- cations of 32 gCi of [3H]leucine by transcervical injection at 0 and 1 h and killed at 2 h. Cathepsin D was partially purified from the uterine homogenates and reacted with the antiserum to rat spleen cathepsin D before SDS-gel electrophoresis of the immunoprecipitate. Standard proteins electrophoresed with the proteins of the immuno- precipitate were: 1, a-amylase, 2, ovalbumin, 3, pepsin, 4, trypsin, and 5, myoglobin. A, control untreated, 12 rats. €3, progesterone treatment, 6 h, 12 rats. C, estradiol benzoate, 24 h, 6 rats.
uterine supernatant proteins were labeled in vivo by intra- uterine administration of [3H]leucine, and uterine cathepsin D was partially purified as previously described. After prepre- cipitation of the uterine proteins with BSA and anti-BSA, cathepsin D antiserum equivalent to the enzyme activity was added. The radiolabeled immunoprecipitate was subjected to SDS-polyacrylamide gel electrophoresis to determine the mo- lecular weight profile of the 3H-labeled uterine proteins. The results presented in Fig. 2 indicate that the antiserum against rat spleen cathepsin D precipitated a single uterine protein with a molecular weight similar to the molecular weight of 43,000 of cathepsin D (20). A uterine protein with apparently the same molecular weight was precipitated by the antibody in uterine homogenates of control ovariectomized rats and ovariectomized rats treated with progesterone or estradiol. Minor protein peaks at molecular weights of 66,500, 26,500, and 15,500 were occasionally observed only in uterine homog-
in the M , = 43,000 peak was used to calculate rates of uterine cathepsin D synthesis after treatment of ovariectomized rats with progesterone or estradiol.
Without changing the rate of cathepsin D synthesis, changes in uterine permeability and the size of intracellular pools of amino acids after hormone treatment could alter the rate of incorporation of amino acids into cathepsin D. Since estradiol increases both uterine permeability and the amino acid pool size (28,29), radioactivity incorporated into cathepsin D pro- tein was expressed/cpm/pmol of amino acid in the acid-solu- ble fraction to correct for changes in these parameters and to calculate relative rates of cathepsin D synthesis. During the 2-h period of in vivo treatment with [3H]leucine, the rate of incorporation of labeled amino acid into uterine cathepsin D in medroxyprogesterone acetate or control ovariectomized rats was linear. No change was observed during this period in the specific activity of the amino acid pools with repeated doses of ["Hlleucine. To determine whether progesterone or estradiol treatment altered the specific radioactivity of leucine in the intracellular amino acid pool, the specific radioactivity of the leucine was measured at various times after hormone treatment (26). Neither progesterone nor estradiol treatment altered the leucine specific radioactivity.
r"" 0
400 i A / /
/ /
/ /
/ P
/ / 1
i
I
J Hours
FIG. 3. Effect of a single injection of estradiol benzoate on the rate of synthesis of uterine cathepsin D. A, rates of incorpo- ration of i3H]leucine into cathepsin D protein are expressed per cpm/ p o l of amino acid in the acid soluble fraction (O---O) or per cpm/ mg of protein in the acid-insoluble fraction (t".). Each point indicates the average of two experiments and in each experiment, uteri of groups of 6 rats (6, 12, 24, and 36 h) or 12 rats (0 h) were pooled. B, levels of cathepsin D enzyme activity after estradiol ben- zoate treatment are indicated. Each point indicates the average f S.E. of 8 rats. At 0 h, rates of uterine cathepsin D synthesis were 558 com/cDm/umol of amino acid in the acid-soluble fraction and 1.39 X
enates of estrogen-treated rats. The amount of radioactivity cpm/cpm/mg of protein in the acid-insoluble fraction. . . _ . ,

Hormonal Control of Rates of Uterine Cathepsin D Synthesis 7477
Rates of cathepsin D synthesis were also determined by expressing radioactivity incorporated into cathepsin D pro- tein/cpm/mg of protein in the acid-insoluble fraction. Calcu- lation of cathepsin D synthetic rates by this means not only corrected for changes in uterine amino acid pool sizes, but also compared the rate of cathepsin D synthesis with the synthetic rates of other uterine proteins. Estradiol increases the general synthetic rate of uterine proteins (30, 31), so expression of rates of cathepsin D synthesis on this basis determined the specificity of the hormonal effects on rates of cathepsin D synthesis.
Effects of Estradiol on the Rate of Uterine Cathepsin D Synthesis-The effects of estradiol on the rate of uterine cathepsin D synthesis after a single injection of the hormone are shown in Fig. 3. After a lag period of at least 6 h, estradiol increased the rate of synthesis of cathepsin D at 12 h and further increases in the rate of synthesis were oberved up to 36 h. The result of the increased rate of cathepsin D synthesis was an accumulation of enzyme activity in the uterus begin- ning 24 h after the injection of estradiol. Rates of cathepsin D synthesis were also compared with the synthetic rates of other uterine proteins by expressing radioactivity in cathepsin D per radioactivity in acid-insoluble protein. Data presented in Fig. 3 indicate that, although estradiol increased the rate of cathepsin D synthesis, the rates of cathepsin D synthesis
I t E.
h 3 0.61
1
1
\ $ 1 T 3 0.4 3 ; 9 0.2 Lu
0 0 6 ' 12 24
Hours FIG. 4. Effect of a single injection of progesterone on the
rate of synthesis of uterine cathepsin D. A , rates of incorporation of [3H]leucine into cathepsin D protein are expressed per cpm/pmol of amino acid in the acid soluble fraction (O---O) or per cpm/mg of protein in the acid-insoluble fraction (U). Each point indicates the average of two experiments and in each experiment, uteri of groups of 12 rats at each time interval were pooled. B, levels of cathepsin D enzyme activity after progesterone treatment are indi- cated. Each point indicates the average & S.E. of 8 rats. At 0 h, rates of uterine cathepsin D synthesis were 2760 cpm/cpm/pmol of amino acid in the acid-soluble fraction and 5.20 X cpm/cpm/mg of protein in the acid-insoluble fraction.
d 1 I 0 6 12 24
Hours
FIG. 5. Effect of medroxyprogesterone acetate on the rate of synthesis of uterine cathepsin D. Rates of incorporation of ["Hlleucine into cathepsin D protein are expressed per cpm/pmol of amino acid in the acid-soluble fraction (O---O) or per cpm/mg of protein in the acid-insoluble fraction (M). Each point indicates the average of two experiments and in each experiment, uteri of groups of 6 (6, 12, and 24 h) or 12 rats (0 h) were pooled. At 0 h, rates of uterine cathepsin D synthesis were 1166 cpm/cpm/pmol of amino acid in the acid-soluble fraction and 1.72 x lo" cpm/cpm/mg of protein in the acid-insoluble fraction.
relative to the synthesis of other uterine proteins were not increased by estradiol treatment. Increases in uterine cathep- sin D synthesis after estradiol treatment appeared to be a part of the general increase in protein synthesis after stimulation of uterine growth by the hormone.
Effect of Progesterone or MPA on the Rate of Uterine Cathepsin D Synthesis-The effects of progesterone on the rate of uterine cathepsin D synthesis after a single injection of progesterone are shown in Fig. 4. Rates of cathepsin D syn- thesis expressed per radioactivity in either the acid-insoluble or -soluble fractions increased to a maximum level at 6 h after the hormone injection and then returned to control levels by 12 h. The result of the increased rate of synthesis of the enzyme protein was an increase in the uterine content of enzyme activity beginning some 12 h after progesterone treat- ment, and further increase in enzyme activity was observed at 24 h.
A potential explanation for the transitory increase in ca- thepsin D synthesis after progesterone treatment could be the relatively short half-life of serum levels of progesterone. When progesterone was injected subcutaneously in oil, the half-life of serum levels of the hormone was approximately 10 h (32). To examine this possibility, medroxyprogesterone acetate (MPA), a progestin with prolonged action, was injected, and rates of uterine cathepsin D synthesis determined, after var- ious periods of time. As shown in Fig. 5, MPA treatment resulted in a large and sustained increase in the rate of cathepsin D synthesis. The rate of cathepsin D synthesis expressed per cpm in the acid-soluble fraction reached a maximum at 6 h, and then decreased. The rate expressed per cpm in the acid-insoluble fraction reached a maximum at 12 h, and then decreased slightly by 24 h. Increases in rates of uterine cathepsin D synthesis after progesterone or MPA treatment were not accompanied by increases in uterine weight.

7478 Hormonal Control of Rates of Uterine Cathepsin D Synthesis
FIG. 6 . Immunofluorescent localization of cathepsin D in uterine cross-sections. Ovariectomized rats were injected with con- trol vehicle ( A and B ) or with medroxyprogesterone acetate (C and D). Cross-sections A and C were treated with cathepsin 1) antiserum; cross sections R and D were treated with preimmune serum. M indicates the uterine myometrium and E indicates the endometrium. Bar in Fig. 6A indicates 50 pm.
Effect of MPA on the Immunohistochemical Localization of Cathepsin D-Control of uterine function by progesterone and estrogen depends upon the effects of these hormones on specific tissues comprising the uterus. To determine whether progestins increased the cathepsin D content of specific tissues in the uterus, the antiserum to cathepsin D was used to localize the enzyme protein in the uterus by immunohisto- chemical staining of frozen sections. In the section of an ovariectomized rat uterus shown in Fig. 6 A , concentrations of cathepsin D indicated by fluorescence were localized in the thin layer of cells of the uterine serosal membrane. Some fluorescence was also observed in the smooth muscle layers of the uterus, but very little fluorescence was observed in the endometrium. Fig. 6C shows the distribution of uterine ca- thepsin D a t 24 h after MPA treatment of an ovariectomized rat. MPA treatment greatly increased the fluorescence ob- served in the uterine endometrium without altering levels of fluorescence in the smooth muscle or serosal cell layers. Fig. 6, B and D show the fluorescence of ovariectomized rat uterine sections after staining with goat preimmune serum. The min- imal fluorescence in these figures indicated the specificity of the fluorescence for cathepsin D in Figs, 6.4 and 6C.
DISCUSSION
Extreme changes in size and function of the uterus occurring during the various phases of normal physiological function require extensive capacity for intracellular degradation. The massive resorption of various uterine tissues during postpar- tum involution, as well as the cyclic changes in the uterine endometrium during the reproductive cycle, would require precise hormonal control of both lysosomal function and
enzyme content (33). More subtle lysosomal mechanisms would appear to be involved in the pinocytosis of the uterine luminal epithelium during the preimplantation period of preg- nancy (14), the penetration of the luminal epithelium with the arrival of the blastocyst (15). and the extensive remodeling of the endometrial stroma during decidualization (16). Changes in rates of protein synthesis and degradation during postpar- tum involution and blastocyst implantation are accompanied by significant changes in cathepsin D activity (12, 17). While the contribution of cathepsin D to the intracellular degrada- tion of protein remains unclear, the control of cellular lyso- somal activity in the uterus must depend in part upon the hormonal control of levels of lysosomal enzymes.
The results obtained in our experiments indicate that in- creased levels of cathepsin D activity in the ovariectomized rat uterus after estradiol or progesterone treatment result from increases in the rates of synthesis of the enzyme protein. Both estradiol and progesterone increased the rates of cathep- sin D synthesis, but significant differences between their ef- fects were observed. Increases in uterine cathepsin D synthesis after estradiol treatment appeared to be a part of a general increase in protein synthesis rather than the induction of a specific enzyme protein. Rates of incorporation of labeled amino acid into cathepsin D protein relative to rates of protein synthesis in general remained constant after estradiol treat- ment. Cathepsin D activity increased only some 12 h after estradiol treatment during the period of uterine growth.
In contrast to the effects of estradiol on cathepsin D syn- thesis, progesterone and medroxyprogesterone acetate in- creased the rate of synthesis of cathepsin D without initiating increases in general protein synthesis or uterine growth. Pro- gesterone treatment had an immediate effect on the synthesis of cathepsin D protein which led to more rapid increases in cathepsin D activity. Few studies have demonstrated “positive effects” of progestins on uterine biochemistry, despite the expectation that these hormones would prepare the uterine endometrial cell for blastocyst implantation by genetic acti- vation and increased synthesis of specific proteins (34). Pro- gesterone maintains endometrial growth and differentiation during decidualization, but suppression or tissue redirection of increases in cellular synthetic activity resulting from estra- diol treatment have been more frequently observed. The suppressive effects of progesterone may depend upon in- creases in levels of progesterone receptor after estradiol treat- ment (35).
Our studies have demonstrated an increased rate of synthe- sis of a specific uterine protein in response to progestin treat- ment without pretreatment of the ovariectomized rats with estradiol. Maintenance of the increased rates of cathepsin D synthesis evidently depends upon stable levels of serum pro- gestins, since rates of synthesis rapidly returned to control levels after treatment with progesterone, which has a short half-life in the serum. Levels of cathepsin D activity in the uterine myometrium increase after progesterone or estradiol treatment (13), but immunohistochemical staining of uterine tissues with cathepsin D antiserum after progestin treatment, indicated that the predominant effect of the hormone was on endometrial levels of enzyme protein. Disappearance of the endometrial epithelium in response to the implanting blasto- cyst is a significant feature of blastocyst implantation in rodents (36). Deterioration and eventual removal of the uter- ine epithelial cells might reasonably depend upon the auto- phagic activity of epithelial cell lysosomes and their content of hydrolytic enzymes. The accumulation of cathepsin D within the endometrium during early pregnancy in response to progesterone secretion would provide the uterine epithelial cells with the potential for increased autophagic activity, and

Hormonal Control of Rates of Uterine Cathepsin D Synthesis 7479
increased intracellular protein degradation facilitating the dis- 15. El-Shershaby, A. M., and HincNiffe, J. R. (1975) J. Embryol. appearance of the cells during blastocyst implantation. EX^. MoFhol. 33, 1067-1080
16. Jollie, W. P., and Bencosme, S. A. (1965) Am. J. Anat. 116, 217- Acknowledgments-We thank Dr. Roger Ganschow for assistance
with the methods of measuring amino acid incorporation into immu- noprecipitable protein, Dr. James Lessard for assistance with the immunofluorescence microscopy, and Dr. Anne Martin for allowing us to use her laboratory facilities for measurements of the specific radioactivity of leucine. Drs. Kenneth Barker and Paul Russell pro- vided many helpful suggestions, and Carol Bates and Beth Koenig provided excellent technical assistance. We thank Vickie Stidham for the careful preparation of the manuscript.
REFERENCES 1. Segal, H. L., and Doyle, D. J., eds (1978) Protein Turnover and
Lysosome Function, pp. 9-291, Academic Press, New York 2. Goldberg, A. L., and St. John, A. C. (1976) Annu. Rev. Biochem.
3. Seglen, P. O., Grinde, B., and Solheim, A. E. (1979) Eur. J.
4. Schworer, C. M., and Mortimore, G. E. (1979) Proc. Natl. Acad.
5. Dean, R. T. (1975) Nature (Lond.) 257,414-416 6. Hopgood, M. F., Clark, M. G., and Bdard, F. J . (1977) Biochem.
J. 164,399-407 7. Amenta, J . S., Hlivko, T. J., McBee, A. G., Shinozuka, H., and
Brocher, S. (1978) Exp. Cell Res. 115, 357-366 8. Poole, B., Ohkurna, S., and Warburton, M. (1978) in Protein
Turnover and Lysosome Function (Segal, H. L., and Doyle, D. J., eds) pp. 43-58, Academic Press, New York
9. Ward, W. F., Chua, B. L., Li, J. B., Morgan, H. E., and Mortimore, G. E. (1979) Bioehem. Biophys. Res. Commun. 87,92-98
10. Libby, P., and Goldberg, A. L. (1978) Science 199, 534-536 11. Amenta, J. S., Sargus, M. J., and Baccino, F. M. (1977) Biochim.
12. Moulton, B. C. (1974) Biol. Reprod. 10, 543-548 13. Sloane, B. F., and Bird, J. W. C. (1977) Am. J. Physiol. 232,
14. Parr, M. B., and Parr, E. L. (1978) J. Reprod. Fertil. 52, 183-188
45, 747-803
Biochem. 95.215-225
Sei. U. S. A. 76,3169-3173
Biophys. Acta 476,253-261
E423-E431
17. 18.
19. 20.
21. 22.
224 Woessner, J. F., Jr. (1965) Biochem. J. 97,855-866 Woessner, J. F., Jr., and Shamberger, R. J., Jr. (1971) J. Biol.
Barrett, A. J. (1970) Biochem. J. 117, 601-607 Kazakova, 0. V., and Orekhovich, V. N. (1976) Biochem. Biophys.
Weber, K., and Osborn, M. (1969) J. Biol. Chem. 244, 4406-4412 Yamamoto, K., Katsuda, N., Himeno, M., and Keitaro, D. (1979)
Chem. 246, 1951-1960
Res. Commun. 72, 747-752
Eur. J. Biochem. 95,459-467 23. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. and Randall, R. J.
24. Ouchterlony, 0. (1966) in Immunological Methods (Ackroyd, J .
25. Barker, K. L. (1976) Endocrinology 81,791-797 26. Martin, A. F., Rabinowitz, M., Blough, R., Prior, G., and Zak, R.
(1977) J. Biol. Chem. 252,3422-3429 27. Barrett, A. J. (1977) in Proteinases in Mammalian Cells and
Tissues (Barrett, A. J., ed) Vol. 2, pp. 181-208, Elsevier/North Holland, Amsterdam.
(1951) J. Biol. Chem. 193,265-275
F., ed) pp. 55-78, F. A. Davis Co., Philadelphia
28. Szego, C. M. (1965) Fed. Proc. 24, 1343-1352 29. Kalman, S. M., and Lombrozo, M. E. (1961) J. Pharmacol. Exp.
30. Mueller, G. C. (1953) J. Biol. Chem. 204, 77-90 31. Noteboom, W. D. and Gorski, J. (1963) Proc. Natl. Acad. Sci. U.
32. Pepe, G. J. and Rothchild, I. (1973) Endocrinology 93,1193-1199 33. Woessner, J. F., Jr. (1969) in Lysosomes in Biology and Pathol-
ogy, (Dingle, J. T., and Fell, H. B., eds) Vol. 1, pp. 299-329, North Holland Publishing Co., Amsterdam
34. OMalley, B. W., and Strott, C. A. (1973) in Handbook of Physi- ology Section 7: Endocrinology, (Greep, R. O., ed) Vol. 2, Part
C. 1, pp. 591-602, American Physiological Society, Washington D.
35. Leavitt, W. W., Toft, D. O., Strott, C. A., and O'Mdey, B. W. (1974) Endocrinology 94, 1041-1053
36. Schlafke, S., and Enders, A. C. (1975) Biol. Reprod. 12.41-65
Ther. 31, 265-269
S. A. 50,250-255
Related Documents











