The interaction between EAP30 and ELL is modulated by MCM2 Amit Mathews 1 , Linda Holland 1 , Krassimir Yankulov * Department of Molecular and Cellular Biology, University of Guelph, Guelph, Ontario, Canada N1G 2W1 article info Article history: Received 24 April 2009 Revised 17 September 2009 Accepted 2 October 2009 Available online 9 October 2009 Edited by Ivan Sadowski Keywords: Eleven–nineteen lysine-rich leukemia ELL-associated protein 30 Mini-chromosome maintenance 2 abstract ELL-associated protein 30 (EAP30) was initially characterized as a component of the Holo–ELL com- plex, which contains the elongation factor ELL. Both ELL and Holo–ELL stimulate RNA pol II elonga- tion in vitro. However, ELL and not Holo–ELL inhibits RNA pol II initiation. It is not clear how these two discrete functions of ELL are regulated. Here we report that mini-chromosome maintenance 2 (MCM2) binds to EAP30 and show that MCM2 competes with ELL for binding to EAP30 thus poten- tially modulating the stability of Holo–ELL. Structured summary: MINT-7277033: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with RPB1 (uniprotkb:P24928) by anti tag coimmunoprecipitation (MI:0007) MINT-7277085: EAP30 (uniprotkb:Q96H20) binds (MI:0407) to ELL (uniprotkb:P55199) by pull down (MI:0096) MINT-7277072: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with ELL (uniprotkb:P55199) by anti tag coimmunoprecipitation (MI:0007) MINT-7277100: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with ELL (uniprotkb:P55199) by competition binding (MI:0405) MINT-7277153: MCM2 (uniprotkb:P49736) binds (MI:0407) to ELL (uniprotkb:P55199) by pull down (MI:0096) MINT-7276989: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with MCM2 (uni- protkb:P49736) by pull down (MI:0096) MINT-7277005: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with RPB1 (uniprotkb:P24928) by pull down (MI:0096) MINT-7276960, MINT-7277168: MCM2 (uniprotkb:P49736) physically interacts (MI:0915) with EAP30 (uniprotkb:Q96H20) by two hybrid (MI:0018) MINT-7276971, MINT-7277121, MINT-7277137: MCM2 (uniprotkb:P49736) binds (MI:0407) to EAP30 (uniprotkb:Q96H20) by pull down (MI:0096) MINT-7277018, MINT-7277061: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with MCM2 (uniprotkb:P49736) by anti tag coimmunoprecipitation (MI:0007) Crown Copyright Ó 2009 Published by Elsevier B.V. on behalf of Federation of European Biochemical society. All rights reserved. 1. Introduction Eleven–nineteen lysine-rich leukemia (ELL) has been initially identified as a gene that undergoes translocations with MLL (mixed lineage leukemia) in acute myeloid leukemia [1]. Independently, ELL has been purified as a RNA pol II elongation factor [2], which also acts as a potent suppressor of the initiation of RNA pol II tran- scription [3]. Structure–function analyses have indicated that inhi- bition of initiation but not stimulation of elongation in vitro was suppressed by the removal of the first 50 N-terminal amino acids of ELL [3]. Interestingly, the MLL–ELL translocations, which are found in patients with acute myeloid leukemia, produce similar deletions of the N-terminus of ELL [1,4]. Studies in Drosophila have shown that dELL associates with ac- tively elongating RNA polymerase II and plays an essential role in development [5,6]; and that it regulates poised RNA pol II at the Hsp70 and other promoters [7]. Therefore, ELL has emerged as a general RNA pol II initiation/elongation factor. However, N-termi- nal dELL deletion mutants, which lack the domain essential for its targeting to sites of active transcription, still support viability 0014-5793/$36.00 Crown Copyright Ó 2009 Published by Elsevier B.V. on behalf of Federation of European Biochemical society. All rights reserved. doi:10.1016/j.febslet.2009.10.009 Abbreviations: ELL, Eleven–nineteen lysine-rich leukemia; EAP30, ELL-associated protein 30; MCM2, mini-chromosome maintenance 2; ChB, chromatography buffer; PAGE–SDS gels, polyacrylamide–sodium dodecyl sulfate gels * Corresponding author. Fax: +1 519 837 2075. E-mail address: [email protected] (K. Yankulov). 1 These authors contributed equally to this work. FEBS Letters 583 (2009) 3431–3436 journal homepage: www.FEBSLetters.org

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

FEBS Letters 583 (2009) 3431–3436
journal homepage: www.FEBSLetters .org
The interaction between EAP30 and ELL is modulated by MCM2
Amit Mathews 1, Linda Holland 1, Krassimir Yankulov *
Department of Molecular and Cellular Biology, University of Guelph, Guelph, Ontario, Canada N1G 2W1
a r t i c l e i n f o
Article history:Received 24 April 2009Revised 17 September 2009Accepted 2 October 2009Available online 9 October 2009
Edited by Ivan Sadowski
Keywords:Eleven–nineteen lysine-rich leukemiaELL-associated protein 30Mini-chromosome maintenance 2
0014-5793/$36.00 Crown Copyright � 2009 Publishedoi:10.1016/j.febslet.2009.10.009
Abbreviations: ELL,Eleven–nineteen lysine-rich leuprotein 30; MCM2, mini-chromosome maintenance 2;PAGE–SDS gels, polyacrylamide–sodium dodecyl sulfa
* Corresponding author. Fax: +1 519 837 2075.E-mail address: [email protected] (K. Yankul
1 These authors contributed equally to this work.
a b s t r a c t
ELL-associated protein 30 (EAP30) was initially characterized as a component of the Holo–ELL com-plex, which contains the elongation factor ELL. Both ELL and Holo–ELL stimulate RNA pol II elonga-tion in vitro. However, ELL and not Holo–ELL inhibits RNA pol II initiation. It is not clear how thesetwo discrete functions of ELL are regulated. Here we report that mini-chromosome maintenance 2(MCM2) binds to EAP30 and show that MCM2 competes with ELL for binding to EAP30 thus poten-tially modulating the stability of Holo–ELL.
Structured summary:MINT-7277033: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with RPB1 (uniprotkb:P24928)by anti tag coimmunoprecipitation (MI:0007)MINT-7277085: EAP30 (uniprotkb:Q96H20) binds (MI:0407) to ELL (uniprotkb:P55199) by pull down(MI:0096)MINT-7277072: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with ELL (uniprotkb:P55199)by anti tag coimmunoprecipitation (MI:0007)MINT-7277100: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with ELL (uniprotkb:P55199)by competition binding (MI:0405)MINT-7277153: MCM2 (uniprotkb:P49736) binds (MI:0407) to ELL (uniprotkb:P55199) by pull down(MI:0096)MINT-7276989: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with MCM2 (uni-protkb:P49736) by pull down (MI:0096)MINT-7277005: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with RPB1 (uniprotkb:P24928)by pull down (MI:0096)MINT-7276960, MINT-7277168: MCM2 (uniprotkb:P49736) physically interacts (MI:0915) with EAP30(uniprotkb:Q96H20) by two hybrid (MI:0018)MINT-7276971, MINT-7277121, MINT-7277137: MCM2 (uniprotkb:P49736) binds (MI:0407) to EAP30(uniprotkb:Q96H20) by pull down (MI:0096)MINT-7277018, MINT-7277061: EAP30 (uniprotkb:Q96H20) physically interacts (MI:0915) with MCM2(uniprotkb:P49736) by anti tag coimmunoprecipitation (MI:0007)
Crown Copyright � 2009 Published by Elsevier B.V. on behalf of Federation of European Biochemicalsociety. All rights reserved.
1. Introduction scription [3]. Structure–function analyses have indicated that inhi-
Eleven–nineteen lysine-rich leukemia (ELL) has been initiallyidentified as a gene that undergoes translocations with MLL (mixedlineage leukemia) in acute myeloid leukemia [1]. Independently,ELL has been purified as a RNA pol II elongation factor [2], whichalso acts as a potent suppressor of the initiation of RNA pol II tran-
d by Elsevier B.V. on behalf of Fede
kemia; EAP30,ELL-associatedChB, chromatography buffer;te gels
ov).
bition of initiation but not stimulation of elongation in vitro wassuppressed by the removal of the first 50 N-terminal amino acidsof ELL [3]. Interestingly, the MLL–ELL translocations, which arefound in patients with acute myeloid leukemia, produce similardeletions of the N-terminus of ELL [1,4].
Studies in Drosophila have shown that dELL associates with ac-tively elongating RNA polymerase II and plays an essential role indevelopment [5,6]; and that it regulates poised RNA pol II at theHsp70 and other promoters [7]. Therefore, ELL has emerged as ageneral RNA pol II initiation/elongation factor. However, N-termi-nal dELL deletion mutants, which lack the domain essential forits targeting to sites of active transcription, still support viability
ration of European Biochemical society. All rights reserved.

3432 A. Mathews et al. / FEBS Letters 583 (2009) 3431–3436
thus suggesting that the essential role of ELL is distinct from itsfunction as a general pol II factor [8]. In agreement, over-expres-sion of ELL causes transformation of RAT1 cells [4] or growth inhi-bition/apoptosis in HEK293 cells [9] again suggesting a role inaddition to its function as a pol II transcription factor. A third lineof study has revealed that ELL acts as a positive co-regulator of themineralocorticoid receptor but negative regulator of the glucocor-ticoid receptor [10].
In rat liver extracts ELL has been found in a stable complex withthree proteins, EAP45, EAP30 and EAP22, to form the so-calledHolo–ELL [11]. Unlike ELL, Holo–ELL does not suppress transcrip-tional initiation in vitro suggesting that the associated proteinsregulate this activity [11]. Of these, EAP30 alone can interact withELL and reverse the inhibitory effect of ELL on initiation [12]. Sep-arately, EAP30 has been characterized as the VPS22 component ofthe endosomal trafficking complex ESCRT-II and has been impli-cated in telomere length regulation [13] or in the association withbicoid RNA [14]. It is not clear if this form of EAP30 regulates ELL orif it has a completely independent role. In summary, the signifi-cance of the ELL-EAP30 interaction remains elusive. Here we reportthat mini-chromosome maintenance 2 (MCM2) binds to EAP30 andinterferes with the ELL-EAP30 association.
2. Materials and methods
2.1. Two-hybrid screen
Two-hybrid screen was performed with pGBKT7-MCM2(1–230)as bait and Hela Matchmaker cDNA library (Clontech). Two-hybridinteraction and a-galactosidase assays were performed withpGBKT7-MCM2(1–230) and a set of pGAD-GH-EAP30 deletion mu-tants (shown in Fig. 2) according to the instructions of the manu-facturer (Clontech).
2.2. Vectors
Vectors for the expression of His6-MCM2 (pRSET-BM28) [15],GST-MCM2(1–169) and GST-MCM2(1–230) [16] were describedpreviously. EAP30 expression vectors were produced by PCR andsub-cloning of full-length EAP30 in pGEX2T (Pharmacia), pFLAG-CMV-2 (Sigma), pcDNA3.1 (Invitrogen), pET28b (Novagen) andpGAD-GH (Clontech), respectively. pGBKT7-MCM2(1-169) andpGBKT7-MCM2(1–230) were produced by PCR and sub-cloninginto pGBKT7. pET28-hisELL was produced by sub-cloning the NotIfragment from pFLAG-ELL [17].
2.3. Cell transformation and immunoprecipitation
HEK293 cells were grown in 10 cm plates to 50% confluency inDMEM/10% FBS. For the experiment in Fig. 1D, each plate wastransfected with 10 lg of pFLAG-EAP30 plus 20 lg of carrier plas-mid (pBluescript) by calcium phosphate-precipitation. For the co-expression of MYC-EAP30 and FLAG-ELL in Fig. 3a each plasmidwas used at 10 lg without carrier. Cells were harvested 36–48 hpost-transfection and extracts were prepared as in [16]. Lysateswere immunoprecipitated with 2 lg of anti-FLAG M2 (Sigma)and 20 ll of Protein-G-Agarose (Sigma) or 20 ll of anti-MYC-Aga-rose (Sigma). Beads were washed four times with 1 ml buffer andeluted in SDS-loading buffer. Western blot analysis was performedwith anti-RPB1(8WG16), anti-(BM28) [15] or anti-Enolase1 [18].Anti-MYC and anti-FLAG-M2 antibodies were from Sigma.
2.4. Expression of His6-tagged proteins
His6-ELL was expressed from pET28a-hisELL in BL21(LysS)DE3for 2 h at 30 �C. Cells were lysed in Chromatography buffer (ChB:
50 mM NaCl, 20 mM Na–HEPES 7.9, 0.2 mM EDTA, 0.2 mM EGTA,5 mM 2-glycerophosphate, 0.05% NP-40, 12% glycerol, 1 lg/mLpepstatin, 1 lg/mL leupeptin, 2 lg/mL aprotonin, 50 lg/mL PMSF)and extracts were spun for 15 min at 15 000 rpm in a SorvallSA300. His6-ELL was found in the insoluble pellet. About 100 llof the pellet was suspended in 1 ml of 7 M guanidine hydrochlo-ride and renatured by stepwise dialysis to 0.09 M guanidinehydrochloride. The renatured proteins were then loaded on Ni-NTA Agarose (Qiagen). His6-MCM2 was expressed from pRSET-BM28 in BL21(LysS)DE3 for 2 h at 30 �C as directed by the man-ufacturer (Invitrogen). His6-EAP30 was expressed from pET28b-EAP30 in BL21(LysS)DE3 for 2 h at 30 �C. In both cases, cell lysatesin ChB were spun for 15 min at 15 000 rpm, the supernatantswere supplemented with 0.1% NP-40 and loaded on Ni-NTA Aga-rose (Qiagen). Ni-NTA beads were then washed with ChB/2 mMImidazole and eluted stepwise with 50 mM and 300 mM Imidaz-ole in ChB. The peak fractions were pooled and snap-frozen. Forthe experiments in Fig. 3, the partially purified proteins werethawed, buffer-exchanged to ChB through Biospin P-6 columns(Bio-Rad) and immediately applied to beads containing GST-fu-sion proteins.
2.5. GST-affinity chromatography and GST pull-down assays
GST, GST-MCM2(169–230), GST-MCM2(1–230) and GST-EAP30were expressed from pGEX and cells were lysed in ChB. Lysateswere incubated with Glutathione-Sepharose-4B (Pharmacia) toyield about 1–2 mg of full-length protein per ml resin as estimatedby boiling an aliquot of the resin and running the sample on poly-acrylamide–sodium dodecyl sulfate gels (PAGE–SDS) gels. ControlGST beads contained about 5 mg protein per ml of resin. For theaffinity chromatography experiments in Fig. 1b and c 100 ll beadswere transferred to 1 ml columns (Bio-Rad) and loaded with 1 mlHeLa whole cell extract prepared as in [15]. Extracts were passedthree times through the beads, the beads were washed four timeswith 1 ml ChB and eluted with ChB/1 M NaCl. For the pull-downassays in Fig. 3b and c 15 ll beads were transferred to 1.5 ml tubesand were incubated with different combinations of His6-taggedproteins for 45 min at 4 �C. Beads were washed four times with1 ml ChB and then boiled in SDS-loading buffer before analysisby PAGE–SDS/Coomassie.
3. Results and discussion
3.1. EAP30 interacts with the N-terminus of MCM2
We have performed an yeast two-hybrid screen using theMCM2 N-terminal amino acids 1–230 ((MCM2(1–230)) as bait.Among other meaningful preys we pulled out EAP30 (Fig. 1a) andconfirmed the EAP30-MCM2 interaction by affinity chromatogra-phy with bacterially over-expressed proteins (Fig. 1b). Removalof the N-terminal 169 amino acids ((MCM2(169–230)) dramati-cally reduced this interaction in both assays (Fig. 1a and b). Onthe other hand, the previously reported association with RNA polII ‘‘holoenzyme” [16] was critically dependent on the 169–230and not on the 1–169 amino acids of MCM2. The MCM2(1–169)protein proved to be unstable in both systems thus precluding di-rect test on its interaction with EAP30. Together, these analyseshave shown that EAP30 associates with MCM2 via a domain dis-tinct from the one mediating the MCM2-RNApol II ‘‘holoenzyme”interaction.
We extended the initial observations by analysis of the associa-tion of EAP30 with native proteins in mammalian cell extracts. Inthe first set of experiments, GST and GST-EAP30 were expressedin E. coli and immobilized on Glutathione-Sepharose. Subse-quently, HeLa whole cell extract prepared as in [15] was passed
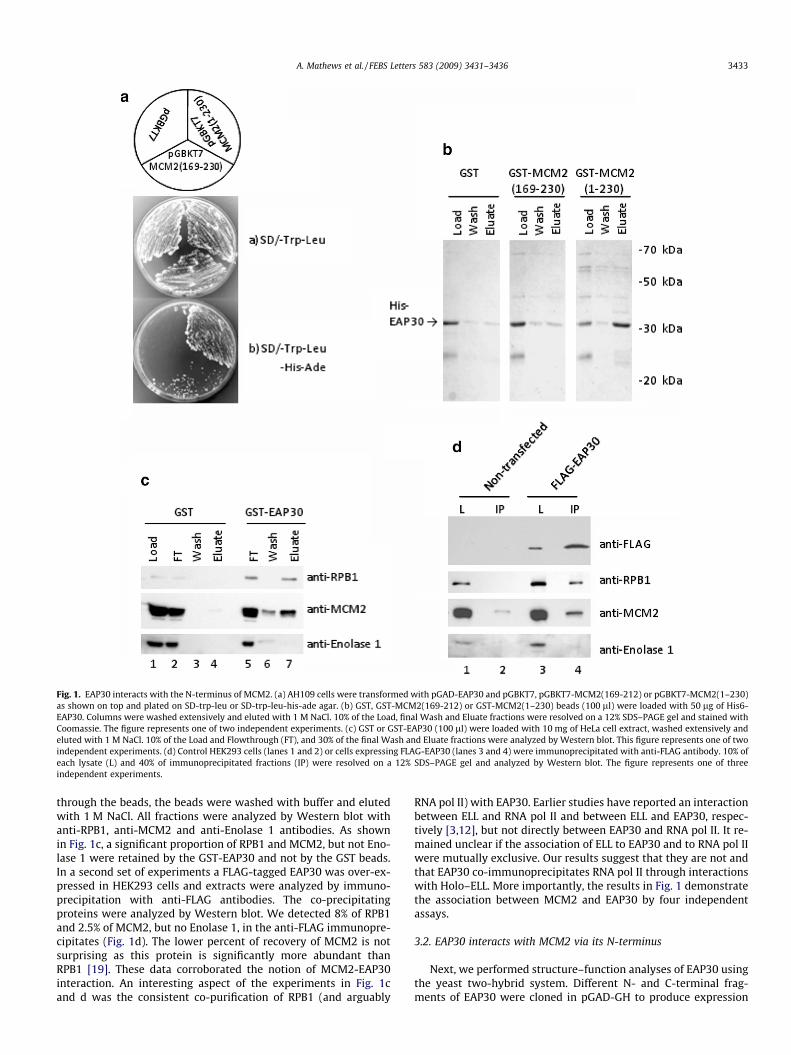
Fig. 1. EAP30 interacts with the N-terminus of MCM2. (a) AH109 cells were transformed with pGAD-EAP30 and pGBKT7, pGBKT7-MCM2(169-212) or pGBKT7-MCM2(1–230)as shown on top and plated on SD-trp-leu or SD-trp-leu-his-ade agar. (b) GST, GST-MCM2(169-212) or GST-MCM2(1–230) beads (100 ll) were loaded with 50 lg of His6-EAP30. Columns were washed extensively and eluted with 1 M NaCl. 10% of the Load, final Wash and Eluate fractions were resolved on a 12% SDS–PAGE gel and stained withCoomassie. The figure represents one of two independent experiments. (c) GST or GST-EAP30 (100 ll) were loaded with 10 mg of HeLa cell extract, washed extensively andeluted with 1 M NaCl. 10% of the Load and Flowthrough (FT), and 30% of the final Wash and Eluate fractions were analyzed by Western blot. This figure represents one of twoindependent experiments. (d) Control HEK293 cells (lanes 1 and 2) or cells expressing FLAG-EAP30 (lanes 3 and 4) were immunoprecipitated with anti-FLAG antibody. 10% ofeach lysate (L) and 40% of immunoprecipitated fractions (IP) were resolved on a 12% SDS–PAGE gel and analyzed by Western blot. The figure represents one of threeindependent experiments.
A. Mathews et al. / FEBS Letters 583 (2009) 3431–3436 3433
through the beads, the beads were washed with buffer and elutedwith 1 M NaCl. All fractions were analyzed by Western blot withanti-RPB1, anti-MCM2 and anti-Enolase 1 antibodies. As shownin Fig. 1c, a significant proportion of RPB1 and MCM2, but not Eno-lase 1 were retained by the GST-EAP30 and not by the GST beads.In a second set of experiments a FLAG-tagged EAP30 was over-ex-pressed in HEK293 cells and extracts were analyzed by immuno-precipitation with anti-FLAG antibodies. The co-precipitatingproteins were analyzed by Western blot. We detected 8% of RPB1and 2.5% of MCM2, but no Enolase 1, in the anti-FLAG immunopre-cipitates (Fig. 1d). The lower percent of recovery of MCM2 is notsurprising as this protein is significantly more abundant thanRPB1 [19]. These data corroborated the notion of MCM2-EAP30interaction. An interesting aspect of the experiments in Fig. 1cand d was the consistent co-purification of RPB1 (and arguably
RNA pol II) with EAP30. Earlier studies have reported an interactionbetween ELL and RNA pol II and between ELL and EAP30, respec-tively [3,12], but not directly between EAP30 and RNA pol II. It re-mained unclear if the association of ELL to EAP30 and to RNA pol IIwere mutually exclusive. Our results suggest that they are not andthat EAP30 co-immunoprecipitates RNA pol II through interactionswith Holo–ELL. More importantly, the results in Fig. 1 demonstratethe association between MCM2 and EAP30 by four independentassays.
3.2. EAP30 interacts with MCM2 via its N-terminus
Next, we performed structure–function analyses of EAP30 usingthe yeast two-hybrid system. Different N- and C-terminal frag-ments of EAP30 were cloned in pGAD-GH to produce expression

Fig. 2. The N-terminus of EAP30 interacts with MCM2(1–230). EAP30 deletion mutants fused to the GAL4 Activation Domain in pGAD-GH were co-transformed with pGBKT7-MCM2(1–230) into AH109 cells and assayed for a-galactosidase activity. For each construct three colonies were assayed and the mean activities are plotted as a percentagerelative to full-length GAL4AD-EAP30(1–258) in the graph on the left. Bracketed numbers on the left correspond to the EAP30 amino acid residues encoded in the construct.Transformants were also grown for on SC-trp-leu-his-ade plates and scored for growth (+), weak growth (+/�) and no growth (�) as shown on the right. The figure representsone of two independent experiments. Error bars represent S.D. of triplicate a-galactosidase assays.
3434 A. Mathews et al. / FEBS Letters 583 (2009) 3431–3436
cassettes for GAL4-AD-EAP30 fusion proteins (Fig. 2). These weretransformed in AH109 cells (Clontech) harbouring pGAL4-DBD-MCM2(1–230). Western blot indicated that all EAP30-fusion pro-teins were expressed at similar levels (not shown). Subsequently,we tested the ability of these fusion proteins to support growthon quadruple drop-out medium or by measuring a-galactosidaseactivity of cells grown in double drop-out medium. The results ofboth assays showed that removal of the N-terminal 48 amino acidsof EAP30 reduced binding to MCM2 by 75% while removal of the Nterminal 1–98 amino acids precluded the association of the twoproteins. C-terminal deletion reduced binding to MCM2 as mea-sured by the a-galactosidase assay but still supported growth(Fig. 2). Together, the data show that the N-terminus of EAP30plays a critical role in its interaction with MCM2.
We attempted similar analyses with ELL cloned in pGBKT7. Sur-prisingly, neither the cell growth nor the a-galactosidase assays re-vealed any interaction between EAP30 and ELL (not shown).Because ELL expression was detected by Western blot, we attributethis lack of interaction to misfolding of ELL or to blocking of its N-terminus in the fusion protein. We concluded that the two-hybridsystem is not adequate for analysis of the EAP30-ELL interaction.
3.3. MCM2 and ELL compete for binding to EAP30
Despite the negative result in the two-hybrid assay, there is awell-documented interaction between EAP30 with ELL [12]. Wetherefore attempted analyses by transient transfection in HEK293cells using FLAG-ELL and MYC-EAP30. Indeed, we noticed thatabout 20% of FLAG-ELL in the load was immunoprecipitated bythe anti-MYC antibodies (Fig. 3a, lane 5). Interestingly, no endoge-nous MCM2 was seen in these samples (Fig. 3a, lane 5). In contrast,when MYC-EAP30 was expressed alone, about 3% of endogenousMCM2 was immunoprecipitated (Fig. 3a, lane 6). It is noteworthythat HEK293 do not express significant amounts of endogenousELL [9]. So, it is conceivable that over-expression of ELL competeswith MCM2 for EAP30. Still, the complexity of cell extracts com-bined with the over-expression of EAP30 and ELL offer limited con-fidence in this conclusion.
We revisited the idea of competition between ELL and MCM2for EAP30 by performing pull-down assays in a defined system
with bacterially expressed His6-ELL, His6-MCM2 and GST-EAP30.His6-ELL was expressed and renatured as in [12]. This proceduregenerates ELL, which interacts with EAP30 and RNA pol II [12].His6-MCM2 was expressed as in [15]. Both His6-ELL and His6-MCM2 were partially purified on Ni-NTA-Agarose and snap-frozen.Purity and abundance of the purified proteins were estimated byPAGE–SDS gels (Fig. 3b, lanes 3 and 4). To avoid aggregation duringdialysis, His6-ELL and His6-MCM2 were buffer-exchanged throughspin columns immediately before adding to GST-EAP30 beads.His6-ELL or His6-MCM2 did not bind to GST (Fig. 3b, lanes 16 and17). The GST-EAP30 beads were incubated for 45 min with differ-ent concentrations of His6-ELL and His6-MCM2 (Fig. 3b, lanes 5–14), washed and analyzed by PAGE–SDS/Coomassie stain. InFig. 3b we show that His6-MCM2 alone (lane 5) or His6-ELL alone(lane 10) were retained by GST-EAP30 at levels permitting visual-ization by Coomassie stain. When His6-MCM2 was premixed withHis6-ELL, even low amounts of His6-ELL were efficient in the dis-placing of EAP30-bound His6-MCM2 (Fig. 3b, lanes 6–9). In con-trast, when EAP30-bound His6-ELL was challenged by His6-MCM2, little His6-ELL was displaced (Fig. 3b, lanes 11–14). Thesedata indicate that His6-ELL and His6-MCM2 independently interactwith EAP30, however the EAP30-ELL interaction is favoured.
We performed a similar experiment with GST-MCM2(1–230) asan affinity ligand (Fig. 3c). These beads were incubated with His6-EAP30 (lane 5) or His6-ELL (lane 10) alone and with increasingamounts of His6-ELL (lanes 6–9) or His6-EAP30 (lanes 11–14),respectively. We observed that His6-ELL and His6-EAP30 bind indi-vidually to GST-MCM2(1–230) (Fig. 3d, lanes 5 and 10), but mixingthem precludes the binding of both. We conclude that MCM2, ELLand EAP30 can compete for interacting with each other withEAP30-ELL being the preferred interaction.
MCM2 is measured at more than 106 copies in proliferating cells[19], however its abundance is dramatically reduced in quiescentcells including hepatocytes [20]. The abundance of ELL in quiescentand proliferating cells has not been established [21,22], howeverthe highest levels of expression of ELL mRNA are observed in theliver [21]. Interestingly, Holo–ELL had been purified from rat liverswhere this complex is likely to be unchallenged by MCM2. Con-versely, Holo–ELL, which is maintained mostly through the ELL–EAP30 interaction [12], may be unstable in proliferating cells due
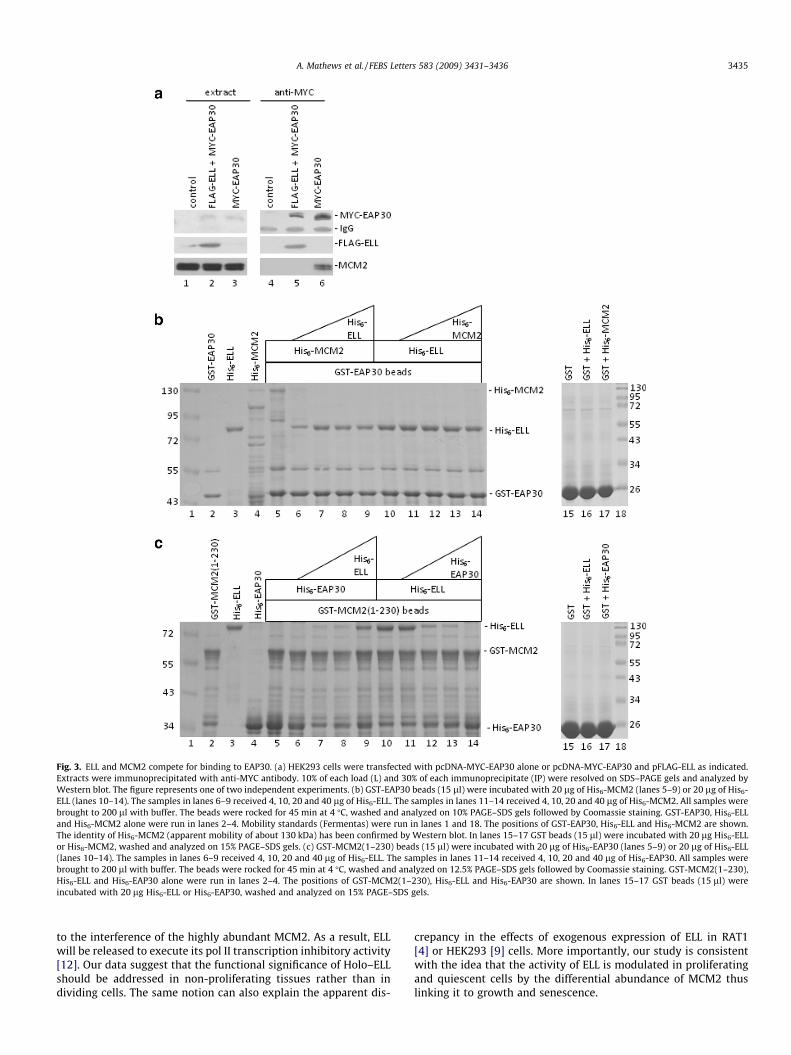
Fig. 3. ELL and MCM2 compete for binding to EAP30. (a) HEK293 cells were transfected with pcDNA-MYC-EAP30 alone or pcDNA-MYC-EAP30 and pFLAG-ELL as indicated.Extracts were immunoprecipitated with anti-MYC antibody. 10% of each load (L) and 30% of each immunoprecipitate (IP) were resolved on SDS–PAGE gels and analyzed byWestern blot. The figure represents one of two independent experiments. (b) GST-EAP30 beads (15 ll) were incubated with 20 lg of His6-MCM2 (lanes 5–9) or 20 lg of His6-ELL (lanes 10–14). The samples in lanes 6–9 received 4, 10, 20 and 40 lg of His6-ELL. The samples in lanes 11–14 received 4, 10, 20 and 40 lg of His6-MCM2. All samples werebrought to 200 ll with buffer. The beads were rocked for 45 min at 4 �C, washed and analyzed on 10% PAGE–SDS gels followed by Coomassie staining. GST-EAP30, His6-ELLand His6-MCM2 alone were run in lanes 2–4. Mobility standards (Fermentas) were run in lanes 1 and 18. The positions of GST-EAP30, His6-ELL and His6-MCM2 are shown.The identity of His6-MCM2 (apparent mobility of about 130 kDa) has been confirmed by Western blot. In lanes 15–17 GST beads (15 ll) were incubated with 20 lg His6-ELLor His6-MCM2, washed and analyzed on 15% PAGE–SDS gels. (c) GST-MCM2(1–230) beads (15 ll) were incubated with 20 lg of His6-EAP30 (lanes 5–9) or 20 lg of His6-ELL(lanes 10–14). The samples in lanes 6–9 received 4, 10, 20 and 40 lg of His6-ELL. The samples in lanes 11–14 received 4, 10, 20 and 40 lg of His6-EAP30. All samples werebrought to 200 ll with buffer. The beads were rocked for 45 min at 4 �C, washed and analyzed on 12.5% PAGE–SDS gels followed by Coomassie staining. GST-MCM2(1–230),His6-ELL and His6-EAP30 alone were run in lanes 2–4. The positions of GST-MCM2(1–230), His6-ELL and His6-EAP30 are shown. In lanes 15–17 GST beads (15 ll) wereincubated with 20 lg His6-ELL or His6-EAP30, washed and analyzed on 15% PAGE–SDS gels.
A. Mathews et al. / FEBS Letters 583 (2009) 3431–3436 3435
to the interference of the highly abundant MCM2. As a result, ELLwill be released to execute its pol II transcription inhibitory activity[12]. Our data suggest that the functional significance of Holo–ELLshould be addressed in non-proliferating tissues rather than individing cells. The same notion can also explain the apparent dis-
crepancy in the effects of exogenous expression of ELL in RAT1[4] or HEK293 [9] cells. More importantly, our study is consistentwith the idea that the activity of ELL is modulated in proliferatingand quiescent cells by the differential abundance of MCM2 thuslinking it to growth and senescence.

3436 A. Mathews et al. / FEBS Letters 583 (2009) 3431–3436
Acknowledgements
We thank Dr. A. Shilatifard and Dr. M. Thirman for generousdonation of EAP30 and ELL plasmids, and Dr. D. Miller for anti-Eno-lase1 antibodies. L.H. was supported by a bursary from CIHR. Thisstudy was supported by a grant from NSERC to K.Y.
References
[1] Thirman, M.J., Levitan, D.A., Kobayashi, H., Simon, M.C. and Rowley, J.D. (1994)Cloning of ELL, a gene that fuses to MLL in a t(11;19)(q23;p13.1) in acutemyeloid leukemia. Proc. Natl. Acad. Sci. USA 91, 12110–12114.
[2] Shilatifard, A., Lane, W.S., Jackson, K.W., Conaway, R.C. and Conaway, J.W.(1996) An RNA polymerase II elongation factor encoded by the human ELLgene. Science 271, 1873–1876.
[3] Shilatifard, A., Haque, D., Conaway, R.C. and Conaway, J.W. (1997) Structureand function of RNA polymerase II elongation factor ELL. Identification of twooverlapping ELL functional domains that govern its interaction withpolymerase and the ternary elongation complex. J. Biol. Chem. 272, 22355–22363.
[4] Kanda, Y., Mitani, K., Kurokawa, M., Yamagata, T., Yazaki, Y. and Hirai, H.(1998) Overexpression of the MEN/ELL protein, an RNA polymerase IIelongation factor, results in transformation of Rat1 cells with dependence onthe lysine-rich region. J. Biol. Chem. 273, 5248–5252.
[5] Gerber, M., Ma, J., Dean, K., Eissenberg, J.C. and Shilatifard, A. (2001) DrosophilaELL is associated with actively elongating RNA polymerase II ontranscriptionally active sites in vivo. EMBO J. 20, 6104–6114.
[6] Eissenberg, J.C., Ma, J., Gerber, M.A., Christensen, A., Kennison, J.A. andShilatifard, A. (2002) DELL is an essential RNA polymerase II elongationfactor with a general role in development. Proc. Natl. Acad. Sci. USA 99, 9894–9899.
[7] Smith, E.R., Winter, B., Eissenberg, J.C. and Shilatifard, A. (2008) Regulation ofthe transcriptional activity of poised RNA polymerase II by the elongationfactor ELL. Proc. Natl. Acad. Sci. USA 105, 8575–8579.
[8] Gerber, M.A., Shilatifard, A. and Eissenberg, J.C. (2005) Mutational analysis ofan RNA polymerase II elongation factor in Drosophila melanogaster. Mol. CellBiol. 25, 7803–7811.
[9] Johnstone, R.W., Gerber, M., Landewe, T., Tollefson, A., Wold, W.S. andShilatifard, A. (2001) Functional analysis of the leukemia protein ELL:evidence for a role in the regulation of cell growth and survival. Mol. CellBiol. 21, 1672–1681.
[10] Pascual-Le Tallec, L., Simone, F., Viengchareun, S., Meduri, G., Thirman, M.J. andLombes, M. (2005) The elongation factor ELL (eleven–nineteen lysine-richleukemia) is a selective coregulator for steroid receptor functions. Mol.Endocrinol. 19, 1158–1169.
[11] Shilatifard, A. (1998) Identification and purification of the Holo–ELL complex.Evidence for the presence of ELL-associated proteins that suppress thetranscriptional inhibitory activity of ELL. J. Biol. Chem. 273, 11212–11217.
[12] Schmidt, A.E., Miller, T., Schmidt, S.L., Shiekhattar, R. and Shilatifard, A. (1999)Cloning and characterization of the EAP30 subunit of the ELL complex thatconfers derepression of transcription by RNA polymerase II. J. Biol. Chem. 274,21981–21985.
[13] Rog, O., Smolikov, S., Krauskopf, A. and Kupiec, M. (2005) The yeast VPS genesaffect telomere length regulation. Curr. Genet. 47, 18–28.
[14] Irion, U. and St Johnston, D. (2007) Bicoid RNA localization requires specificbinding of an endosomal sorting complex. Nature 445, 554–558.
[15] Yankulov, K., Todorov, I., Romanowski, P., Licatalosi, D., Cilli, K., McCracken, S.,Laskey, R. and Bentley, D.L. (1999) MCM proteins are associated with RNApolymerase II holoenzyme. Mol. Cell Biol. 19, 6154–6163.
[16] Holland, L., Gauthier, L., Bell-Rogers, P. and Yankulov, K. (2002) Distinct partsof minichromosome maintenance protein 2 associate with histone H3/H4 andRNA polymerase II holoenzyme. Eur. J. Biochem. 269, 5192–5202.
[17] Luo, R.T., Lavau, C., Du, C., Simone, F., Polak, P.E., Kawamata, S. and Thirman,M.J. (2001) The elongation domain of ELL is dispensable but its ELL-associatedfactor 1 interaction domain is essential for MLL–ELL-induced leukemogenesis.Mol. Cell Biol. 21, 5678–5687.
[18] Subramanian, A. and Miller, D.M. (2000) Structural analysis of alpha-enolase.Mapping the functional domains involved in down-regulation of the c-mycprotooncogene. J. Biol. Chem. 275, 5958–5965.
[19] Burkhart, R., Schulte, D., Hu, D., Musahl, C., Gohring, F. and Knippers, R. (1995)Interactions of human nuclear proteins P1Mcm3 and P1Cdc46. Eur. J. Biochem.228, 431–438.
[20] Freeman, A., Hamid, S., Morris, L., Vowler, S., Rushbrook, S., Wight, D.G.,Coleman, N. and Alexander, G.J. (2003) Improved detection of hepatocyteproliferation using antibody to the pre-replication complex: an associationwith hepatic fibrosis and viral replication in chronic hepatitis C virus infection.J. Viral Hepat. 10, 345–350.
[21] Thirman, M.J., Diskin, E.B., Bin, S.S., Ip, H.S., Miller, J.M. and Simon, M.C. (1997)Developmental analysis and subcellular localization of the murine homologueof ELL. Proc. Natl. Acad. Sci. USA 94, 1408–1413.
[22] Polak, P.E., Simone, F., Kaberlein, J.J., Luo, R.T. and Thirman, M.J. (2003) ELL andEAF1 are Cajal body components that are disrupted in MLL–ELL leukemia. Mol.Biol. Cell 14, 1517–1528.
Related Documents











