© CAB International 2013. Trichoderma: Biology and Applications (eds P.K. Mukherjee et al.) 43 3 The Influence of Light on the Biology of Trichoderma Sergio Casas-Flores 1 and Alfredo Herrera-Estrella 2 * 1 IPICYT, División de Biología Molecular, San Luis Potosí, México; 2 Laboratorio Nacional de Genómica para la Biodiversidad, CINVESTAV Irapuato, México 3.1 Introduction During evolution, almost all forms of life on earth have been exposed to different electro- magnetic radiation of internal and external origin, and are ultimately maintained by trans- formation of the energy contained in the elec- tromagnetic radiation emitted by the sun. Plants capture the sun’s energy and use it to synthesize energy-rich molecules (sugars) from molecules of carbon dioxide and water. These molecules then serve, directly or indirectly, as the source of energy for plants themselves and ultimately for all animals and decomposer organisms (such as bacteria and fungi). But many organisms use sunlight in other ways. Apart from its energetic content, sunlight has properties (such as intensity, duration, polarization or spectral composition) that pro- vide patterns, which in turn have important consequences for living organisms. In par- ticular, these patterns have the potential to be used as a source of information by a cognitive agent or can be used to produce thermody- namic work. In their natural habitat, all forms of life are continuously obtaining and decoding information from their environment (includ- ing that contained in light), which they use for their own benefit. In this sense, the human * E-mail: [email protected] visual system is the best example based on the quantity and complexity of meanings conferred to the spatio-temporal patterns of stimulation that light provokes on the retina. The use of light either as energy or as an information source therefore depends on the interaction of light with the molecular system of the organism under consideration. In general, the mechanism of photoper- ception starts when an electron in a photo- sensitive molecule passes from its ground state to an excited state by absorbing energy from the incoming photon. When the electron returns to its ground state (or intermediate states lying below the excited state), the energy difference is released in one (or a combina- tion) of several paths: it can be dissipated thermally, it can be returned as light (as in fluorescence or phosphorescence) or it can initiate a series of photochemical reactions. This last possibility is the one that leads to physiological responses in photosensitive organisms. The sun emits light in a wide wave- length range (Fig. 3.1). Infrared is the radia- tion of longer wavelength carrying less energy (Fig. 3.1); it fits with the energies required for vibrational movements of sev- eral molecular bonds, which absorb it and

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

© CAB International 2013. Trichoderma: Biology and Applications (eds P.K. Mukherjee et al.) 43
3 The Influence of Light on the Biology of Trichoderma
Sergio Casas-Flores1 and Alfredo Herrera-Estrella2*1IPICYT, División de Biología Molecular, San Luis Potosí, México;
2Laboratorio Nacional de Genómica para la Biodiversidad, CINVESTAV Irapuato, México
3.1 Introduction
During evolution, almost all forms of life on earth have been exposed to different electro-magnetic radiation of internal and external origin, and are ultimately maintained by trans-formation of the energy contained in the elec-tromagnetic radiation emitted by the sun. Plants capture the sun’s energy and use it to synthesize energy-rich molecules (sugars) from molecules of carbon dioxide and water. These molecules then serve, directly or indirectly, as the source of energy for plants themselves and ultimately for all animals and decomposer organisms (such as bacteria and fungi). But many organisms use sunlight in other ways.
Apart from its energetic content, sunlight has properties (such as intensity, duration, polarization or spectral composition) that pro-vide patterns, which in turn have important consequences for living organisms. In par-ticular, these patterns have the potential to be used as a source of information by a cognitive agent or can be used to produce thermody-namic work. In their natural habitat, all forms of life are continuously obtaining and decoding information from their environment (includ-ing that contained in light), which they use for their own benefit. In this sense, the human
* E-mail: [email protected]
visual system is the best example based on the quantity and complexity of meanings conferred to the spatio-temporal patterns of stimulation that light provokes on the retina. The use of light either as energy or as an information source therefore depends on the interaction of light with the molecular system of the organism under consideration.
In general, the mechanism of photoper-ception starts when an electron in a photo-sensitive molecule passes from its ground state to an excited state by absorbing energy from the incoming photon. When the electron returns to its ground state (or intermediate states lying below the excited state), the energy difference is released in one (or a combina-tion) of several paths: it can be dissipated thermally, it can be returned as light (as in fluorescence or phosphorescence) or it can initiate a series of photochemical reactions. This last possibility is the one that leads to physiological responses in photosensitive organisms.
The sun emits light in a wide wave-length range (Fig. 3.1). Infrared is the radia-tion of longer wavelength carrying less energy (Fig. 3.1); it fits with the energies required for vibrational movements of sev-eral molecular bonds, which absorb it and

44 S. Casas-Flores and A. Herrera-Estrella
transform it into molecular movement (heat). Radiation of shorter wavelength, con-taining more energy per photon, corresponds to ultraviolet (UV; Fig. 3.1), which can initiate photochemical reactions. Among the molecules that can be affected by UV, DNA is perhaps the most important because the result of one of such reactions can result in a change trans-mitted to the next generation as a mutation, if such a change is not repaired before DNA replication. UV radiation can also damage molecules through its capacity to initiate un -controlled free radical reactions, in most cases involving reactive oxygen species (ROS). Additionally, visible light can indirectly give rise to ROS as a product of photosensitive reactions through energy transfer from a mol-ecule that can be activated by light such as flavin or porphyrin. It is in this way that blue light is potentially harmful (Lledias and Hansberg, 2000; Aguirre et al., 2005).
Reactions of metabolism such as photo-synthesis and gluconeogenesis in plants, and glycolysis and respiration in plants and animals, imply redox reactions that involve
electron transfer and are tightly regulated. Electron transfer can occur only between molecules that are in close proximity; there-fore, metabolism would have never reached its complexity without the existence of elec-tron carrier molecules such as NAD(P), flavins and porphyrins. These molecules can be found freely in the cytoplasm or associated with proteins. In flavins such as riboflavin (vitamin B12), flavin mononucleotide (FMN), flavin adenine dinucleotide (FAD), and por-phyrins such as a haem group, the necessary energy for the transition of a valence electron to the next orbital is much lower than that required in most simple molecules, which is why such molecules absorb at longer wave-lengths. This absorption occurs in the range of visible light and makes solutions of these compounds appear coloured to our eyes. Such molecules are therefore known as pig-ments or chromophores. Consequently, all biological responses to visible light must be initiated by one of these pigments.
In the context described above it is easy to understand why sunlight is a significant
Increasing energy
Increasing wavelength
400 500 600 700 Wavelength (nm)
wavelength (μ)10–6 10–5 10–4 10–3 10–2 10–1 102 103 104 105 106 107 108 1091 10
MicrowaveUV
Visible light
Xrays
Gammarays
Cosmicrays
RadioTV
Thermal IR
Fig. 3.1. Solar radiation spectrum. Electromagnetic radiation of the sun comprises a broad spectrum from very long radio waves to very short gamma rays. This spectrum includes light visible to the human eye, which ranges from near ultraviolet (UV) to red light, passing by blue and green light. A decrease in the wavelength translates into an increase in energy and vice versa. The sun’s electromagnetic radiation ranging from UV to infrared (IR) regulates several biological processess including circadian rhythms, photomorphogenesis, phototropisms and synthesis of pigments, among others.

Light and the Biology of Trichoderma 45
element for life, and, besides the utilization of its energy and information, during evolution many mechanisms have been selected for to resist its negative effects. Many organisms synthesize pigments that act as a protective umbrella, such as melanin and carotenoids. Carotenoids possess, in addition, antioxidant power to neutralize the harmful effects of sin-glet oxygen. They also produce DNA repair enzymes that reverse damage provoked by UV light. Thus, light has an ambiguous role in life; on the one hand life itself depends on its energy and information, and on the other it is potentially harmful and even deadly. Life, including fungi, under different light conditions requires significant adjustments in numerous regulatory processes, which is reflected in the widespread effects on the reg-ulation of genes and enzymes (Tisch and Schmoll, 2009).
3.2 The Study of Light Responses in Fungi
Phycomyces was probably the first fungus in which the effect of light was analysed. Max Delbrück in his pioneering work concentrated precisely on studying the way this organism relates to light, in the capacity of this fungus to ‘see’. His work established the basis for future developments in fungal photobiology. He described in great detail the properties of the response to light of Phycomyces and initi-ated the genetic zapping of the signal trans-duction pathways but faced serious difficulties in the molecular characterization of the first step of these pathways, photoperception. Long before the work in Phycomyces, however, the study of the ‘informational’ use of light by plants had been initiated. Plant movements such as the orientation towards the sun or the escape from an excess of light are provoked or influenced by blue light and were consid-ered in ancient times by philosophers as a proof that plants have a soul. These phenom-ena caught the attention of Charles Darwin so strongly that he wrote a complete volume describing them, where he refers to the observation that a yellow–orange solution of potassium dichromate used as a light filter
eliminated phototropism (Darwin, 1880). This phenomenon was considered as ideal to solve the identity of the blue-light photoreceptor. By the late 1960s, the similarity of the action spectra for various biological responses to blue light in organisms as diverse as plants, bacteria, ferns and fungi had become evident. This suggested that all such responses should be controlled by the same type of photoreceptor of ancestral origin (Bergman et al., 1969), which was named the ‘near UV/blue’ receptor or ‘cryptochrome’, highlighting its hidden absorp-tion (cryptic) and its preponderance in lower plants (cryptogams) and fungi (Gressel, 1979).
3.3 The Influence of Light on Asexual Reproduction
A common mode of asexual reproduction in fungi is conidiation. Fungal conidia have diverse forms and functions, and all of them represent non-motile asexual propagules. Most Trichoderma species can be found living in soil as saprobes or colonizing plant roots. A variety of stress signals have been found to trigger this developmental process in Trichoderma includ-ing nutrient deprivation, desiccation, pH, the use of all available space to grow or when the colony reaches the end of the dish, among oth-ers (Horwitz et al., 1984a; Carreras-Villaseñor et al., 2012).
Conidiation is a developmental process that implies drastic physiological and struc-tural changes that can be induced by a brief pulse of light in Trichoderma. In contrast to what has been described in other organisms, this was the only obvious response of Trichodermato light and hence the reason that led to the use of this fungus as a simple photomorpho-genic model. Early studies on fungal photo-biology including physiological, biochemical and genetic aspects of light responses were carried out basically only in Trichoderma and Phycomyces as coordinated research efforts (Gressel and Rau, 1983).
The first description of the effect of light on conidiation of Trichoderma was made in 1951 (Lilly and Barnett, 1951). In their work, Lilly and Barnett described that in nutrient-rich medium in the dark, Trichoderma grows
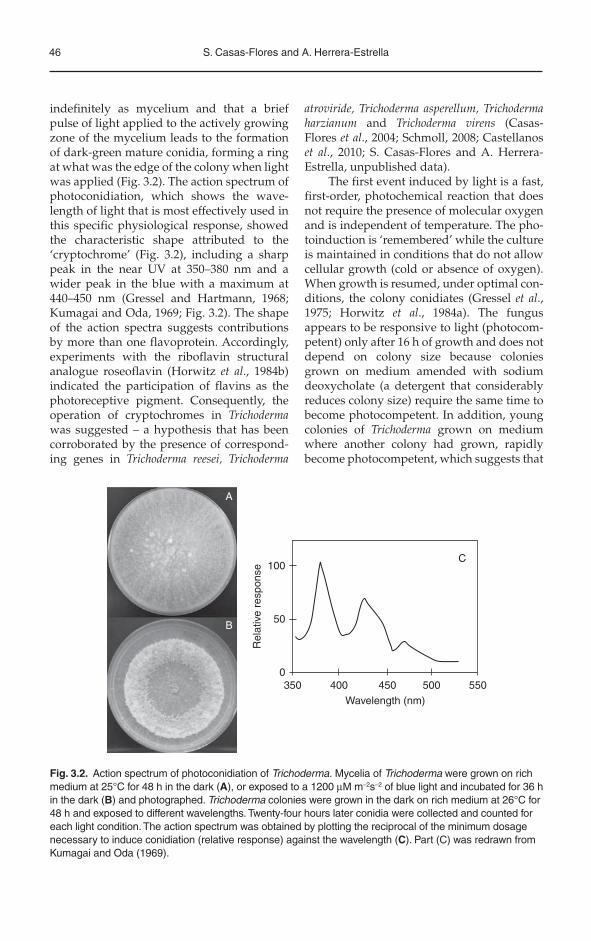
46 S. Casas-Flores and A. Herrera-Estrella
indefinitely as mycelium and that a brief pulse of light applied to the actively growing zone of the mycelium leads to the formation of dark-green mature conidia, forming a ring at what was the edge of the colony when light was applied (Fig. 3.2). The action spectrum of photoconidiation, which shows the wave-length of light that is most effectively used in this specific physiological response, showed the characteristic shape attributed to the ‘cryptochrome’ (Fig. 3.2), including a sharp peak in the near UV at 350–380 nm and a wider peak in the blue with a maximum at 440–450 nm (Gressel and Hartmann, 1968; Kumagai and Oda, 1969; Fig. 3.2). The shape of the action spectra suggests contributions by more than one flavoprotein. Accordingly, experiments with the riboflavin structural analogue roseoflavin (Horwitz et al., 1984b) indicated the participation of flavins as the photoreceptive pigment. Consequently, the operation of cryptochromes in Trichodermawas suggested – a hypothesis that has been corroborated by the presence of correspond-ing genes in Trichoderma reesei, Trichoderma
atroviride, Trichoderma asperellum, Trichoderma harzianum and Trichoderma virens (Casas-Flores et al., 2004; Schmoll, 2008; Castellanos et al., 2010; S. Casas-Flores and A. Herrera-Estrella, unpublished data).
The first event induced by light is a fast, first-order, photochemical reaction that does not require the presence of molecular oxygen and is independent of temperature. The pho-toinduction is ‘remembered’ while the culture is maintained in conditions that do not allow cellular growth (cold or absence of oxygen). When growth is resumed, under optimal con-ditions, the colony conidiates (Gressel et al.,1975; Horwitz et al., 1984a). The fungus appears to be responsive to light (photocom-petent) only after 16 h of growth and does not depend on colony size because colonies grown on medium amended with sodium deoxycholate (a detergent that considerably reduces colony size) require the same time to become photocompetent. In addition, young colonies of Trichoderma grown on medium where another colony had grown, rapidly become photocompetent, which suggests that
100C
A
B50
Rel
ativ
e re
spon
se
0350 400 450
Wavelength (nm)
500 550
Fig. 3.2. Action spectrum of photoconidiation of Trichoderma. Mycelia of Trichoderma were grown on rich medium at 25°C for 48 h in the dark (A), or exposed to a 1200 μM m–2s–2 of blue light and incubated for 36 h in the dark (B) and photographed. Trichoderma colonies were grown in the dark on rich medium at 26°C for 48 h and exposed to different wavelengths. Twenty-four hours later conidia were collected and counted for each light condition. The action spectrum was obtained by plotting the reciprocal of the minimum dosage necessary to induce conidiation (relative response) against the wavelength (C). Part (C) was redrawn from Kumagai and Oda (1969).

Light and the Biology of Trichoderma 47
the accumulation of an extracellular factor is responsible for this phenomenon (Gressel and Galun, 1967). Using scanning electron microscopy, 3–7 h after exposure to light, abundant branching of aerial hyphae with an increased number of septa can be observed, as well as the formation of new aerial hyphae (Galun, 1971). Branches form conidiophores, and the new aerial hyphae elongate, branch and also form conidiophores. Both types of hyphae are photo-inducible but conidiophore development starts from aerial hyphae.
This developmental programme can be divided into a determination state, a morpho-genetic stage that includes re-programming of gene expression, and the appearance of the corresponding physiological and morpho-logical changes. Between these two states there is a ‘point of no return’, from which execution of the programme is irreversible. Considerable changes in contents of DNA, RNA and pro-teins in mycelia of colonies exposed to light were detected, whereas those maintained in the dark showed only minor changes (Gressel and Galun, 1967; Betina and Spisiaková, 1976). These changes were attributed to fructification structures and conidia. Pharmacological experi-ments showed that actinomycine D and rami-hyphin A blocked photoconidiation partially, whereas cycloheximide and 5-fluorouracil were completely inhibitory to conidiation induced by light (Betina and Spisiaková, 1976). Together these results suggested that de novo tran-scription and translation are required for pho-toconidiation. Accordingly, the use of the RNA synthesis inhibitor 5-fluorouracil per-mitted the establishment of a time-window of approximately 7 h after illumination in which this developmental programme can be sup-pressed once it was triggered by light, co -inciding with the time when conidiophores start to branch. After this time, application of 5-fluorouracil can diminish conidiation but cannot completely inhibit it (Galun and Gressel, 1966; Gressel and Galun, 1967; Betina and Spisiaková, 1976).
According to the Bunsen–Roscoe law of reciprocity, a given quantity of photons could be delivered in pulses of different duration but the final response should be the same. A deviation from this rule indicates the par-ticipation of more than one photoreceptor, as
in the case of phototropism in higher plants or the Phycomyces sporangiophore (Presti and Galland, 1987). Alternatively, it could point to complexities of a single photoreceptor system such as photoreceptor recycling. For T. atroviridephotoconidiation, reciprocity holds for pulses of blue light lasting from nanoseconds to minutes. On the basis of this data, it would appear that in Trichoderma photoconidiation is triggered by a single receptor system that is neither recycled to the photoreceptive form nor counted by enzymatic processes during or immediately following irradiation (Horwitz et al., 1990). This conclusion is supported by the observation that photoinduction is ‘remem-bered’ while the culture is maintained in conditions that do not allow cellular growth (cold or absence of oxygen) (Gressel et al.,1975; Horwitz et al., 1990). Recently, these data were reinterpreted pointing to a separa-tion between the initial photoreactions and the development of conidiation. The require-ment of an oxidative process for allowing this phenomenon was also suggested (Steyaert et al., 2010b).
3.4 Early Light Responses
Trichoderma develops aerial hyphae at the col-ony perimeter growing mycelium, but they soon fall over and fuse with the mycelial mat (Greensphan, 1970; Galun, 1971; revised in Horwitz et al., 1984a). When photoinduced, aerial hyphae remain upright at the growing edge of the colony when the light pulse was given. Application of a blue light pulse increases vacuolation, an observation that led to the proposal that this stimulus was a tran-sient ‘stress’ that could initiate changes allow-ing vacuolation, ending in conidiation (Gressel, 1980; Horwitz, 1984a).
Horwitz and co-workers registered elec-tric currents on the plasma membrane of Trichoderma, from 60 to 150 min after applica-tion of a blue light pulse (Horwitz et al.,1984c); Gresik et al. (1988) detected hyperpo-larization of the plasma membrane after the application of light, as well as an increase in the intracellular level of ATP and a rapid decrease in cyclic adenosine monophosphate

48 S. Casas-Flores and A. Herrera-Estrella
(cAMP), followed by a second pulse of cAMP that decreased slowly. These observations suggested the participation of a membrane protein, probably associated with G proteins (Gresik et al., 1988). Interestingly, with the advent of the genome era, opsin-like proteins, which belong to the family of G-protein coupled receptors (GPCRs), have been described for several fungi including three Trichodermaspecies, T. atroviride (210598), T. harzianum(536324) and T. asperellum (206217) (Fig. 3.3). In this sense, three different heterotrimeric G protein alpha subunits have been described in these fungi (Schmoll, 2008; S. Casas-Flores
and A. Herrera-Estrella, unpublished data). The participation of a signalling pathway involving a heterotrimeric G protein is fur-ther supported by the observation that addi-tion of dibutyril-cAMP (a membrane-permeable compound analogous to cAMP) to Trichodermacolonies growing in the dark allowed them to conidiate. Furthermore, the addition of atro-pine, a drug that reduces intracellular cAMP levels, inhibited conidiation after a pulse of blue light (Berrocal-Tito et al., 2000). Together these results indicated that a cAMP-dependent pathway participates in conidiation. However, the transcription of the rapidly
LOV
LOV
PAS PAS
PAS
GATA BLR-1
BLR-2
ENV1
CPD PhotolyasePhotolyase
Photolyase
Photolyase
Rhodopsin
RRDHisKA-like
ATPaseHisKAPHYGAFRed
Green
Blue
FADbinding
FADbinding
FADbinding
6–4 Photolyase
DASH-Cryptochrome
Opsin
Phytochrome
GATA
Fig. 3.3. Domain organization of potential Trichoderma spp. light receptors. The blue light regulator-1 photoreceptor (BLR-1) has a LOV domain that is hypothesized to use a flavin as chromophore to sense light; it also has two PAS domains involved in protein–protein interaction and dimerization, and a GATA zinc-finger DNA-binding domain. The blue light regulator-2 (BLR-2) contains one PAS domain, a histone deacetylase (HDAC) interaction domain, a coil–coil domain and a GATA zinc finger domain. ENV1 has a LOV domain, which presumably binds FAD as chromophore to sense blue light. The DNA 6-4 and CPD photolyases, and cryptochrome, are related flavoproteins that consist of a DNA photolyase domain, which binds a light-harvesting cofactor (folate) and a FAD-binding domain of DNA photolyase. Opsins are similar to microbial opsin-related proteins, which putatively bind rhodopsin to perceive green light. The phytochrome has a GAF small ligand-binding domain and a phytochrome region (PHY) that functions to seal the chromophore and stabilize the photoactivated far-red absorbing state. Phytochrome also has a histidine kinase A (phosphoacceptor and dimerization) domain (HisKA), a HisKA-like ATPase domain, and an RRD, a signal receiver domain that receives the signal from the sensor partner in a two-component system. It contains a phosphoacceptor site that is phophorylated by HisKA.

Light and the Biology of Trichoderma 49
induced gene, phr-1 (coding for a photolyase), was not induced by the cAMP analogue in the dark and atropine did not inhibit its transcrip-tion, despite the fact that fluence response curves suggested that both phenomena are conducted by the same photoreceptor (Berrocal-Tito et al., 2000).
3.5 The Search for Photoreceptors
Using a mutagenesis approach, Horwitz and co-workers (1985) first attempted to isolate the photoreceptor involved in photoconidia-tion, based on a model in which conidiation can be triggered by signals transduced through different receptors for different types of stress (including light), which converge in a common pathway for morphogenesis. They screened for T. atroviride mutants that did not conidiate in response to light but did in response to stress. Despite exhaustive screen-ings they could not obtain the desired type of mutants, which suggested that the photo-receptor could be essential. Interestingly, they isolated mutants that required more light than the parent strain to get an equivalent response and overproduced a yellow pig-ment (dimY, for dimsighted and yellow). It was then suggested that the mutants were affected in the photoreceptor (Horwitz et al.,1985). However, in contrast to Phycomyces and Neurospora, genetic mapping of the mutations was not possible owing to the lack of a sexual reproduction cycle for Trichoderma in the la -boratory. Horwitz and co-workers (1984a, 1985) estimated the number of genes involved in conidiation triggered by light and stress by complementation of heterokaryons (forced heterokaryons) or by the frequency with which they isolated non-conidiating mutants. Despite all the uncertainties involved in such estimations, all the approaches suggested that 12–30 genes were available for selection by their procedure for photoresponses. They also estimated that about 90 Trichoderma gene products are needed for conidiation (induced by either stress or light). They further pro-posed that strains that responded normally to stress should have normal dark reactions (Horwitz et al., 1984a, 1985). Twenty years
later, Casas-Flores and co-workers demon-strated that there is a cross talk between carbon deprivation and the light response, which is mediated by the BLR-1 and BLR-2 proteins (Casas-Flores et al., 2004). Finally, the so long sought ‘cryptochrome’ or flavin asso-ciated molecule responsible for almost all blue light responses observed in Trichodermahad been identified using a PCR-based strat-egy, by designing oligonucleotides directed against the zinc finger domain of GATA type transcription factors, also contained in the white collar (WC) proteins (Casas-Flores et al., 2004). The blr genes encode proteins homologous to the Neurospora crassa WC proteins (Liu and Bell-Pedersen, 2006), and are essential for photoconidiation, gene expression regulated by blue light and are important for mycelial growth (Casas-Flores et al., 2004; Rosales-Saavedra et al., 2006). In Trichoderma spp., BLR-1 and BLR-2 are PAS proteins with a GATA-type DNA-binding domain. BLR-1 has three PAS domains (Fig. 3.3) the first of which belongs to a spe-cialized sensory domain called LOV (Light–Oxygen–Voltage), similar to that initially described in plant phototropins. The LOV domain is a module sensitive to light broadly conserved in light-, oxygen- and voltage-sensing proteins from bacteria, fungi and plants. The BLR-1 LOV domain has all the amino acids necessary to interact with the chromophore FAD, including a cysteine that forms a covalent photoadduct with the fla-vin. BLR-2 has only one PAS domain pre-sumably used to interact with other proteins (Fig. 3.3). Thus, based on the structure of the BLR proteins, and the phenotype observed in blr-1 and blr-2 gene replacement mutants, it is conceivable that the corresponding proteins form a blue light photoreceptor complex in Trichoderma spp., and that this complex acts as a transcription factor (Casas-Flores et al., 2004), as described for the N. crassa WC-1/WC-2 proteins. Castellanos and co-workers (2010) reported similar results for mutants in the T. reesei homo-logues of the blr genes and showed that such mutations also influenced cellulase gene expression (Castellanos et al., 2010). In fact, overexpression of BLR-2, which has no LOV domain, resulted in an increment in

50 S. Casas-Flores and A. Herrera-Estrella
photosensitivity, a property directly associ-ated with photoreceptors (Esquivel-Naranjo and Herrera-Estrella, 2007).
Although BLR-1 and BLR-2 form a blue/UV-A receptor, these proteins also have functions independent of light. BLR proteins are required for conidiation induced by sud-den carbon deprivation and by cAMP addi-tion in darkness (Casas-Flores et al., 2004, 2006). We have also observed lower germina-tion of conidia independent of light in a Dblr-1strain, suggesting that BLR proteins also regulate functions specific to the conidia or to early stages of germination (A. Herrera-Estrella etal., unpublished data). Undoubtedly, BLR and WC proteins are the main photo-receptors in fungi that regulate practically all light responses studied so far but investigat-ing their functions in the dark will provide a better understanding about photoregulated circuits in the cells.
In addition to the putative TrichodermaBLR-1 photoreceptor, a secondary presump-tive blue light photoreceptor has been described (Schmoll et al., 2004). ENVOY (ENV1), the orthologue of VIVID from N. crassa, is a small protein that contains a PAS/LOV domain as its central functional characteristic (Fig. 3.3). VIVID is a blue light photoreceptor that functions downstream of the WC proteins to negatively regulate the responses initiated by the WC proteins. In con-stant light most of blue-light-regulated genes are down-regulated after 2 h in wild-type N. crassa, whereas a mutant in VIVID shows increased accumulation of carotenoids under constant illumination. This hyperpigmenta-tion is due to a sustained expression of carot-enoid synthesis genes in the light (Heintzen et al., 2001; Schwerdtfeger and Linden, 2001, 2003). ENV1 was discovered in T. reesei after a screening for novel signalling factors involved in regulation of the production of cellulases. Surprisingly, a presumed component of cel-lulase gene expression control was related to the light signalling machinery (Schmoll et al.,2004). It was found later that in T. reesei light induces the transcription of genes involved in cellulose degradation (Schmoll et al., 2005; Castellanos et al., 2010). In fact, several inves-tigations support the role of ENV1 in light signal transduction and cellulase production,
which extends the hypothesis that there is a cross talk between light and carbon source signalling, as proposed by Casas-Flores and co-workers in 2004 and 2006 (Schmoll et al.,2004, 2005; Castellanos et al., 2010). In dark-ness, env1 is transcribed at a very low basal level. Upon illumination, strong induction of transcription within minutes results in a 50- to 500-fold increase in transcript abundance in T. atroviride and T. reesei, respectively, which is mediated by the photoreceptor complex BLRC (Castellanos et al., 2010; A. Herrera-Estrella et al., unpublished results). In T. reesei, strains lacking the env1 gene show alterations in transcription of cellulase encod-ing genes in response to blue light and in the dark. The involvement of ENV1 in transcrip-tion of a number of genes in the dark was also demonstrated, thus suggesting a role of ENV1 in gene regulation in the dark (Schmoll et al.,2005). Interestingly, growth of a Denv1 strain exposed to continuous light was strongly reduced when compared with the wild-type strain, suggesting a role of the env1 product in tolerance to light (Schmoll et al., 2005). Similar effects to that observed on the growth pattern of the Denv1 strain had been observed earlier in T. atroviride blr mutants (Casas-Flores et al.,2004). Consequently, the growth inhibition by light observed in T. atroviride could be due to the absence of the env1 transcript.
The signalling function of ENV1 must involve additional, light-dependent auxiliary components because overexpression of ENV1 in darkness did not rescue typical light-dependent behaviour (Schuster et al., 2007). In fact, Denv1 mutants express photoregu-lated genes for a long time under constant illumination, indicating that ENV1 is a nega-tive regulator of the light input, switching off gene expression regulated through BLR-1 and BLR-2 (Castellanos et al., 2010). Although not yet proven to act as a photoreceptor (as its N. crassa orthologue does) in Trichoderma spp., the PAS/LOV domain protein ENV1 plays an important role in light responses. Thus, ENV1 regulates photoadaptation similar to the function of VIVID in N. crassa (Schmoll et al.,2005; Castellanos et al., 2010).
After the publication of the genome sequence of T. reesei (Martinez et al., 2008; http://genome.jgi-psf.org/Trire2/Trire2.

Light and the Biology of Trichoderma 51
home.html), four additional Trichodermagenomes have been sequenced and are pub-licly available: T. atroviride (http://genome.jgi-psf.org/Triat1/Triat1.home.html), T. virens(http://genome.jgi-psf.org/Trive1/Trive1.home.html), T. harzianum (http://genome.jgi-psf.org/Triha1/Triha1.home.html) and T. asperellum (http://genome.jgi-psf.org/Trias1/Trias1.home.html). In all five fungi, homo-logues of the photoreceptors blr-1 and blr-2,and env1 are present. The fact that light responses have been observed, even in Dblrmutants, indicates that Trichoderma spp. have additional genes encoding for photoreceptors (Fig. 3.3). In fact, there are a CPD photolyase (PHR-1), a cryptochrome DASH (Ta-12806, Tv-28006 and Tr-59726), a cryptochrome/6-4 photolyase (Ta-86846, Tv-37166 and Tr-77473) and a phytochrome (Fig. 3.3). The N. crassagenome has one CPD photolyase and a cryp-tochrome DASH, and A. nidulans has only one CPD photolyase but with cryptochrome-type roles, described for PHR-1 of T. atroviride(Berrocal-Tito et al., 2007). The cryp-tochromes/6-4 photolyases of Trichodermaform part of the subfamily of animal cryp-tochromes and 6-4 photolyases and have a COOH-terminal extension of 79 amino acids, similar to animal cryptochromes and the recently described 6-4 photolyase of Cercospora zeae-maydis, with roles in development and secondary metabolism in addition to their photolyase activity. Another notable differ-ence is that the Trichoderma DASH cryp-tochromes have a longer COOH-terminal region (Fig. 3.3) than those described in animals, plants, bacteria and other fungi. T. atroviride cryptochrome DASH has a shorter extension of 391 amino acids than T. virens and T. reesei, which have 710 and 661 amino acids, respectively. However, blast analyses of the COOH-terminal extension against the non-redundant database of the NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi) found no homology, suggesting that the Trichoderma cryptochromes might form a sub-division with novel features within the DASH cryptochromes.
Although responses to red and far-red light have been documented in some fungi, the function of a phytochrome (FphA) has been demonstrated only in Aspergillus nidulans.
It is involved in repression of sexual and stimulation of asexual reproduction under red light (Blumenstein et al., 2005). In Trichoderma, red light provokes a reduction in mycelia growth and also has an impact on the transcriptional regulation of some genes, indicating the participation of a phytochrome in these responses (Casas-Flores et al., 2004; Rosales-Saavedra et al., 2006). As do many fungal genomes, Trichoderma spp. has a phy-tochrome with all features contained in the phytochromes PHYT-1 and FphA described in N. crassa and A. nidulans, except that the putative second phytochrome (PHYT-2) of N. crassa does not contain a conserved PAS domain in its N terminus.
As mentioned above, light impacts on the cAMP-signalling pathway. It is well known that this second messenger is pro-duced by adenylyl cyclase, an enzyme the activity of which is modulated by heterotri-meric G-protein a-subunits, highlighting the possibility that a photoreceptor coupled to G-proteins (GPCR-like) could be involved in this signalling pathway. Three out of five Trichoderma genomes contain a gene encoding a putative opsin protein (T. atroviride-210598,T. asperellum-206217 and T. harzianum-536324)with homology to opsins NCU01735.3 (46% identity) and ops-1 (27% identity) from N. crassa, among others. The opsin has seven transmembrane spans typical of GPCRs asso-ciated with G-proteins. However, at least in T. atroviride and T. reesei, light responses are similar between them and the fact that two Trichoderma species do not have a gene encod-ing an opsin-type photoreceptor decreases the probability of its participation in regulat-ing signalling through the cAMP pathway, although it remains to be proven.
3.6 Circadian Rhythms
To adapt to their environment, organisms have evolved endogenous cellular timekeep-ers that control a number of daily physiologi-cal and molecular rhythms in most eukaryotic and some prokaryotic organisms. These time-keepers are usually named circadian clocks (Liu and Bell-Pedersen, 2006). In N. crassa

52 S. Casas-Flores and A. Herrera-Estrella
the two photoreceptors, WC-1, WC-2, and the clock protein FRQ have a crucial function in circadian rhythmicity (Brunner and Káldi, 2008). Research with Trichoderma spp., in con-trast, did not yet provide any hints as to the presence of circadian rhythms or their regula-tion by homologues of WC-1, WC-2 and FRQ in these fungi. Nevertheless, given the ubiquity of circadian regulation in countless organ-isms, it would be surprising if this mecha-nism was not operative in Trichodermaspp. Thus, it seems that robust circadian rhythms in conidiation cannot be observed under the culture conditions currently used in the laboratory.
Although multiple efforts to elucidate circadian clocks in Trichoderma spp. have been made, little is known about such phenomena in these organisms. Most Trichoderma species require light for conidiation. Colonies grow-ing in the dark were induced by light/day cycles at 8, 16, 24 and 48 h, determining that conidiation occurred, corresponding to the interval of illumination. When Trichoderma is returned to the dark after a pulse of light, conidiation is observed only at what was the colony perimeter at the time of the pulse. Such result allow the conclusion that conidia-tion in T. viride is not rhythmic, but can be synchronized by a light pulse (Betina and Zajacová, 1978). Similarly, under continuous light, conidiation is not rhythmic; however, banding patterns are formed under light/dark cycles. Interestingly, a Trichoderma mutant (B119) that conidiates rhythmically in the dark has been described and the composition of the growth medium influences the period length of conidiation. Medium containing sodium deoxycholate, an ionic detergent that delays the growth of Trichoderma, increased the interval between dark bands from 12 to 24 h. As in the wild-type strains, the applica-tion of light induced a ring of conidia but it also delayed the reappearance of the dark banding pattern (Deitzer et al., 1988). Recently, Steyaert and co-workers described rhythmic conidiation in Trichoderma pluroticola (Steyaert et al., 2010a). These authors described that dark-grown cultures of T. pluroticola form rings of green conidia at intervals of approxi-mately 24 h. Light did not, however, induce rhythmicity in conidiation (Steyaert et al.,
2010a). Nevertheless, similarly to what has been described for frq in N. crassa, the frq gene (Id 162169) of T. atroviride is regulated by light, and such regulation depends on functional blr-1 (Id 268453) and blr-2 (266278) genes – the orthologues of the N. crassa wc-1 and wc-2(M. García-Esquivel, E.U. Esquivel-Naranjo, A.S. Pérez-Martínez, M.A. Hernández-Oñate and A. Herrera-Estrella, unpublished data; Liu and Bell-Pedersen, 2006). Orthologous genes to those described as involved in the regula-tion of circadian rhythms (wc-1, wc-2 and frq)in several fungal species have been also found in other Trichoderma species [(BLR-1: T. virens (193551), T. reesei (121962), T. asperel-lum (260902), T. harzianum (551926); BLR-2 T. virens (126526), T. reesei (22699), T. asperel-lum (60682), T. harzianum (68771); FRQ T. virens (122964), T. reesei (121670), T. asperel-lum (349342) and T. harzianum (60964)], whose genomes have been sequenced. The role of their products in the regulation of circadian clocks has not been described, however. Further research on the molecular aspects of circadian rhythms in Trichoderma should lead us to under-standing this phenomenon in the genus.
3.7 The Influence of Light on Gene Expression
The first studies on the influence of light on gene expression were those carried out to determine the pattern of expression of phr-1,which is rapidly induced by blue light through the BLR proteins (Berrocal-Tito et al.,1999; Casas-Flores et al., 2004). A more thor-ough study included the use of cDNA micro-arrays representing 1438 T. atroviride genes. That study allowed the discovery of 40 genes reg-ulated by white light (2.8% of the genes printed in the array), 30 of which were up-regulated (2%) and 10 down-regulated (0.8%). Surprisingly, not all light-regulated genes appeared to be regulated by the BLR proteins but all those regulated through the BLR pro-teins strictly required both functional pro-teins. Those data provided further support to the suggestion that, like in Neurospora, the BLR-1 and BLR-2 proteins act as a complex (Casas-Flores et al., 2004; Rosales-Saavedra

Light and the Biology of Trichoderma 53
et al., 2006). The identification of blue-light down-regulated genes and their expression analysis in the wild-type strain as well as the Dblr-1 and Dblr-2 mutant strains, demon-strated that the T. atroviride BLR proteins can activate or repress gene transcription, a dual function also recently observed in A. nidulans(Rosales-Saavedra et al., 2006; Ruger-Herreros et al., 2011). The promoter region of the genes regulated by blue light through the BLR pro-teins contain GATA elements, similar to the consensus sequence of the light-responsive element (LRE) described in the genes regu-lated by light in N. crassa (He and Liu, 2005; Rosales-Saavedra et al., 2006). In fact env1expression was regulated by light in N. crassaunder the control of its own promoter, which contains LREs (Schmoll et al., 2005). The fact that the promoter region of the blue-light down-regulated genes of T. atroviride also contains LRE-like elements suggests a more complex mechanism for the control of tran-scriptional activity by the BLR proteins in this fungus, where combined arrangements of cis-acting elements and additional trans-factorscan be decisive for the regulatory output, as proposed for genes regulated by light in plants (Rosales-Saavedra et al., 2006; Jiao et al., 2007).
Analysis under different light conditions of a set of phr-1 promoter serial deletions and chimerical constructs fused to the Escherichia coli lacZ gene showed that LREs play an important role in blue-light signal transduc-tion. Contrasting with Neurospora, only one GATA box of the predicted LRE is sufficient to drive blue-light induction of phr-1. Binding of BLR-1 and BLR-2 to the LRE promoter of phr-1 was also demonstrated by mobility shift assays and chromatin immunoprecipitation, respectively. This supports the hypothesis that BLR-1 and BLR-2 work as a complex during light transcriptional responses in T. atroviride(M. Cervantes-Badillo, T. Muñoz-Centeno, E.E. Uresti-Rivera, G. Argüello-Astorga and S. Casas-Flores, unpublished data).
In a more recent analysis by high-throughput sequencing, we identified 331 light-regulated genes, 70 of which seem to be blr-independent, providing support for the existence of additional, functional, light receptors. Interestingly, 39 out of the
178 light-induced genes are related to differ-ent stress responses. Among these stress-induced genes 17 are related to oxidative stress. This set of genes includes key elements such as components of the mitogen-activated protein kinase (MAPK) (p38/Hog1) cascade related to this type of stress. In addition, key elements involved in the nucleotide excision repair and mismatch repair systems are induced, as well as phr-1 (A. Herrera-Estrella et al., unpublished results). Within the set of light-regulated genes at least ten transcription factors were found. Two of them are ortho-logues of the N. crassa short aerial hyphae tran-scription factors (sah1 and sah3), which as suggested by their names are involved in the correct development of aerial hyphae, a pre-requisite in Trichoderma for the formation of conidiophores. These latter data together with the previously described time-window in which photoconidiation may be halted using inhibitors of transcription, suggest that during 7 h after exposure to light there is a cascade of transcriptional events leading to conidiation.
Similarly to what has been observed for the BLR proteins, ENV1 has both positive and negative impact on gene expression, not only in light but also in darkness, and it is involved in regulation of genes that are not responsive to light (Schuster et al., 2007). Nevertheless, ENV1 is dependent on the presence of additional – presumably light responsive – factors to per-form its regulatory function. The variety of genes found to be regulated by light and partially by ENV1 in T. reesei ranges from transcription, translation and signal trans-duction to genes involved in metabolism and transport (Schuster et al., 2007). Interestingly, it was also found that env1 itself is sub-jected to light regulation (Schmoll et al., 2005; Castellanos et al., 2010; A. Herrera-Estrella et al., unpublished data).
Other genes found to be induced by light in T. reesei are those encoding hydrophobins (hfb1 and hfb2), hydrophobic proteins predom-inantly present in aerial structures of fungi (Nakari et al., 1997). It was found that the G-protein alpha subunit GNA1 is involved in this regulation (Seibel et al., 2009). Moreover, in T. atroviride the BLR-1 and BLR-2 proteins contribute to the adjustment of hydro-phobin levels to environmental conditions

54 S. Casas-Flores and A. Herrera-Estrella
(Mikus et al., 2009). The underlying regulatory mechanism triggering this light response of hydrophobin gene expression in Trichodermais likely to be highly sophisticated and involves light-dependent splicing of pre-mRNA (Vargovic et al., 2006).
3.8 The Effect of Light, cAMP and the Carbon Source on Growth
and Conidiation
It is well known that nitrogen and carbon sources, as well as their availability, may influence growth and development in fungi. Analysis of the effect of different cues on the utilization and assimilation of nutrients has been scarcely studied; it is therefore poorly understood.
The first molecular evidence of the effect of light on growth was that of mutations in the blue-light photoreceptor and its transcrip-tional partner, the BLR-2 protein. Deletions of any of the blr genes allowed a slightly faster growth under continuous light, which was very similar to the wild type growing in the dark. When growing in the dark, mutants also grew faster than the parental strain (Casas-Flores et al., 2004).
Later, a role for the T. reesei photorecep-tor ENV1 in growth stimulation by light was demonstrated. The growth rates of wild-type strains and a mutant strain lacking the PAS domain of the ENV1 protein were compared, when growing on media supplemented with different carbon sources, under dark or light conditions. In the presence of light the paren-tal strains indeed grew faster on many such media than the mutant (Schuster et al., 2007). Interestingly, inhibition by light was not observed on all carbon sources. These results together pointed to an inhibitory effect of light dependent on the carbon source (Schuster et al., 2007).
To study on a wide range, the possible effect of carbon source on growth and conidi-ation, and to investigate the roles of BLR-1 and BLR-2 proteins, oxidative stress and cAMP signalling on both processes in T. atroviride,the fungus was grown on media with differ-ent carbon sources, cAMP or menadione in
darkness or light conditions (Friedl et al.,2008a,b). Exposure of T. atroviride to constant light or 12-h dark/light periods showed sig-nificant stimulation of growth in the presence of light in 17 out of the 95 carbon sources tested (i.e. the hexoses d-fructose, d-manose, d-galacose, etc.). When the fungus was grown in the darkness, 48 out of 95 tested carbon sources induced conidiation in T. atroviride.These included polyols, sugar acids and N-acetyl-b-d-glucosamine as the unique sugar amine. In contrast, conidiation did not occur when growing on any amino acids, aliphatic acids or alcohols, excluding b-hydroxybutiric acid. Interestingly, the growth rate did not cor-relate with conidiation. Of the 48 carbon sources tested that allowed the fungus to conidiate, 9 sustained fast growth, 10 sustained moderate growth and 15 sustained slow growth. The remaining 14 carbon sources provided very poor mycelial growth of the fungus. These results pointed to a strong dependence of growth and conidiation on the carbon source. Similar results were observed when the 48 carbon sources that support conidiation in the dark were tested with light. The light stimulus increased the intensity and speed of conidial development.
The Dblr-1 mutant still showed photo-stimulation of growth on some carbon sources, whereas the Dblr-2 mutant showed similar pattern in the dark as in 12-h dark/light cycles. A stimulatory effect on the Dblr-2mutant was only observed in constant light. These results indicate that BLR proteins have different roles in light perception, pointing to a major role for BLR-1 in photostimulation of growth (Friedl et al., 2008). The same experi-ments were conducted for conidiation with the Dblr-1 and Dblr-2 mutants, and the result was that conidiation in the dark was strongly affected on most carbon sources. The Dblr-1and Dblr-2 strains conidiated moderately on 15 and 27 carbon sources, respectively. When growing on glucuronic acid or with sorbitol, conidiation was indeed enhanced in Dblr-2and Dblr-1, respectively. Addition of D-methyl-d-glucoside to the media did not induce conidiation in the wild-type strain; however, blr mutants clearly developed mature conidia. Only 11 of the carbon sources tested allowed conidiation of both the blr mutants, which

Light and the Biology of Trichoderma 55
indicates different roles for BLR proteins in the carbon-source-dependent regulation of conidia formation.
In this regard, recently it was demon-strated that BLR-2 is the limiting factor for photoperception and phototransduction in T. atroviride. The overexpression of the blr-2gene resulted in higher sensitivity to light by enhancing conidiation and expression of blue-light-responsive genes for which expres-sion is BLR dependent. In contrast, overex-pression of the blr-1 gene resulted in reduced photoconidiation and weaker expression of light-regulated genes (Esquivel-Naranjo and Herrera-Estrella, 2007). Interestingly, overex-pression of blr-2 resulted in reduced growth when T. atroviride was grown under 12-h dark/light conditions, including a period of strong sensitivity to light, followed by a period of insensitivity (Esquivel-Naranjo and Herrera-Estrella, 2007).
It is well known that light provokes pro-duction of reactive oxygen species and it con-tributes to oxidative stress (Aguirre et al., 2005). It was formerly proposed that light in Trichoderma could be a stress signal because a brief pulse of light evokes conidiation (Horwitz et al., 1984a). This asseveration was proposed under the premise that resistant structures such as conidia are produced when conditions are no longer suitable for apical extension. In T. atroviride addition of menadione (a super-oxide generator) to the media, with several carbon sources, mimics growth stimulated by light in the darkness, allowing the conclusion that the effect of light is, in part, due to the gen-eration of reactive oxygen species (Friedl et al., 2008). Furthermore, the finding that T. atrovir-ide was able to grow on 16 out of 95 carbon sources and growth was completely abolished on the others and with some non-carbohy-drates indicates that oxidative stress is also dependent on carbon signalling and/or metabolism (Friedl et al., 2008). These results are in agreement and extend the hypothesis that BLR-1 and BLR-2 proteins could act as intracellular redox and oxygen sensors, and that these proteins have a link with carbohy-drate metabolism (Casas-Flores et al., 2006).
Addition of cAMP to T. atroviride cultures growing in the darkness stimulated growth only on some of 95 carbon sources where
stimulation of growth was determined, which suggests that photostimulation of the assimi-lation of such carbon sources acts through the cAMP pathway (Friedl et al., 2008). Addition of cAMP to the wild-type strain growing in the dark influenced its growth in just four carbon sources (a-d-glucose, gentobiose, d-cellobiose and d-xylose), where an increase in growth in the presence of light had previ-ously been shown. This effect was not observed in the blr mutants, pointing to a link of their products and cAMP in light stimula-tion of growth.
Nemcovic and Farkas (1998) observed that addition of cAMP derivatives to the medium enhanced conidiation in T. viride, but the effect was more pronounced when colo-nies were exposed to sub-saturating doses of light. It was also determined that conidiation is dependent on the concentration of glucose in the media; conidia are more abundant on glucose-rich media. Casas-Flores and co-workers showed that blr mutants were not rescued by supplementation of the media with cAMP, which suggests that BLR pro-teins could act upstream of protein kinase A (Fig. 3.4) (Casas-Flores et al., 2006).
Addition of cAMP to the media essen-tially induced conidiation in the three strains on some but not all carbon sources. The wild type and blr mutants did not conidiate when growing on media with d-xylose, l-rhamnose, b-hydroxybutyric acid, and the stimulation of conidiation on d-saccharid acid, on which the wild-type strain did not conidiate without cAMP in the media. These data together sup-port the hypothesis stating that the BLR pro-teins could be acting as redox and oxygen sensors (Casas-Flores et al., 2006), by suggest-ing that the differences in conidiation observed on different carbon sources are due to differ-ent redox potentials in the cell during growth (Friedl et al., 2008a,b).
3.9 The Biosynthesis of Peptaibols and Secondary Metabolites
In several fungi a regulatory connection between sporulation and the production of secondary metabolism has been reported

56 S. Casas-Flores and A. Herrera-Estrella
nutrientsignalling
ENV1
ENV1H2O2 O2⋅−
⋅HO
AAAAAAAAAAAAAAAAAAAAAAAA
conidiation sexualdevelopment
mycelialgrowth
gna1
gna3
bldblu
gna1
carbon sourceutilization
cellulaseexpression
AMP
LIGHT
GPCR GPCR
ENV1
cAMP
cAMP ATP
PKA
R R
R R
C C
C C
BLR-1
peptaibols
BLR-2
GγGNG1
GβGNB1
GγGNG1
GβGNB1
GαGNA3
AC
DNA repair
stresstolerance
GPCR
GβGNB1
PhLP1
GγGNG1
GαGNA3
GαGNA1
PDE
LIGHT
Fig. 3.4. Trichoderma spp. light signal perception and transduction. The BLR-1 and BLR-2 proteins putatively located in the nucleus sense blue light through a flavin (pentagon) attached to the LOV domain of the BLR-1 protein, which leads to the activation of blue light up-regulated (blu) or repression of blue light down-regulated (bld ) genes. Transcription of blu genes leads to stress tolerance, DNA repair, conidiation and synthesis of proteins such as GNA3 and the photoreceptor ENV1. The BLR proteins positively regulate carbon source utilization, conidiation and synthesis of peptaibols. Several lines of evidence also point to the activation of the BLR complex by reactive oxygen species (H2O2, O2•– and •HO) provoked by the interaction of light with biological molecules such as flavins and porphyrins, or by nutrient deprivation. Once ENV1 senses light, presumably through FAD, it positively regulates carbon source utilization, mycelial growth, cellulase gene expression and transcription of gna3, among other genes. ENV1 is also involved in the negative regulation of the activity of the BLR complex and represses the transcription of gna3 and the activity of the phosphodiesterase (PDE); the latter leads to the decrease of cAMP levels that had acumulated owing to the activation of GNA3, which in turn activates adenylyl cyclase (AC), allowing the synthesis of cAMP (cyclic adenosine monophosphate). Acumulation of cAMP leads to the release of the catalytic subunit (C) of protein kinase A (PKA) due to binding of cAMP to the regulatory subunit (R), to positively regulate conidiation, sexual development and mycelial growth. It has been proposed that PKA also positively regulates the BLR complex (dashed lines from PKA to BLR complex). The accumulation of cAMP has a positive effect on mycelial growth. The PKA catalytic subunit negatively regulates cellulase gene expression. ENV1 negatively regulates the putative interaction of the phosducin-like (PhPL chaperone) product with the GNB1/GNG1 heterodimer to dissociate the active complex that positively regulates conidiation, cellulase expression and sexual development. Nutrient signalling has been shown to have a positive effect on the PhPL/GNB1/GNG1 complex. The PhPL/GNB1/GNG1 complex exerts a negative regulation on blu genes. The GNA1 heterotrimeric G protein has a positive effect on its own transcription, cAMP accumulation, cellulase gene expression and presumably on the PKA catalytic subunit. In the inactive state, the three G-protein subunits (α, β, γ) are present in a complex, in association with G-protein-coupled receptors (GPCRs). Ligand binding to the GPCR leads to dissociation of the Gα subunit from the Gβγ dimer. Solid arrows represent positive regulation, whereas dotted arrows show a presumably positive regulation. Lines with a bar at the end indicate negative regulation. Grey arrows indicate light stimulation of photoreceptors.

Light and the Biology of Trichoderma 57
(Calvo et al., 2002). The fact that Dblr-1 and Dblr-2 strains do not conidiate in response to light allowed the hypothesis that they are altered in secondary metabolism and synthe-sis of peptaibols.
Peptaibols are a family of short peptides (£20 residues), synthesized by non-ribosomal peptide synthetases (NRPSs). Naturally occurring peptaibols are isolated from soil fungi and often exhibit antibacterial and anti-fungal activities. There are over 300 anno-tated sequences of non-ribosomal peptides (Whitmore and Wallace, 2004) and the list is continuously increasing because a single NRPS can produce up to three distinct pep-taibols. Neuhof and co-workers (2007) ana-lysed 28 phylogenetically related Trichodermastrains, identifying 48 different classes of peptaibols.
Recently, it was determined by matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry that T. atroviride did not produce peptaibols dur-ing vegetative growth; however, production of peptaibols was evident when the fungus initiated conidiation. Therefore, different stimuli that induce conidiation were tested to determine if in all cases conidiation and pro-duction of peptaibols were intimately corre-lated. Interestingly, conidiation induced by light promoted high production of peptai-bols, whereas in the dark peptaibols were observed in minimum quantities. Light-associated synthesis of peptaibols was depend-ent on blr-1 and blr-2 products. As expected, no conidiation was observed in the blr-1 and blr-2 mutants. On the other hand, carbon dep-rivation also induces the production of both peptaibols and conidia, independent of blr-1and blr-2, hence light independent. These data are apparently contradictory because it was reported that the carbon deprivation sig-nal is blr-1 and blr-2 dependent; however, such dependence is only observed when cul-tures are subject to sudden carbon depriva-tion (Casas-Flores et al., 2006). Mechanical injury of mycelia also triggered peptaibol production but it depended completely on light stimulation: no peptaibol production was observed in absence of light, in spite of sporulation. These discoveries were also observed in blr mutants. Together these results
indicate the existence of a BLR-independent pathway for peptaibol production stimu-lated by light (Fig. 3.4) (Komon-Zelazowska et al., 2007).
Recently, it was shown that mycelia of T. virens wild-type strain (Gv29.8) exposed to light exhibit a slight increase in the expres-sion of the veA transcript (Mukherjee and Kenerley, 2010). Mutants in T. virens veA are defective in induction of genes that encode for secondary metabolism enzymes and show a null phenotype in the synthesis of gliotoxin. The lack of gliotoxin correlates with low expression levels of gliP, the NRPS encod-ing gene responsible for gliotoxin production in Aspergillus nidulans. In A. nidulans, VeA interacts with the phytochrome (FphA)–white-collar light regulator complex (LreB/LreA) and is also part of the velvet complex VelB/VeA/LaeA, which are key components in secondary metabolism. This complex coor-dinates the light signal with fungal develop-ment and secondary metabolism (Bayram et al., 2008a,b). Furthermore, mutants in the wcoA gene, the blr-1 orthologue in Fusariumfujikuroi, sustained carotenoid synthesis in response to light. On the contrary, production of fusarin showed a drastic reduction in the light and less synthesis of gibberellins and more bikaverins when mycelia were growing under nitrogen-limiting conditions. These results indicate that blr-1 orthologous genes play pivotal roles in secondary metabolism in F. fujikuroi. It is noteworthy that F. fujikuroidoes not sporulate when exposed to light; on the contrary, this stimulus seems to have a repressing role on conidiation in this fungus. However, DwcoA showed reduced levels of conidiation on minimal agar when com-pared with the wild-type strain (Estrada and Avalos, 2008). Together these results indicate an important role for WcoA in nitrogen-regulated processes, as well as for gibberellin and bikaverin synthesis in a light-independent pathway.
The available data indicate notable differ-ences in the use of VeA and BLR-1, and their partners in secondary metabolism. Further studies on single, double and triple mutants as well as the demonstration of the interactions between the different components of the VelB/VeA/LaeA and phytochrome–white-collar

58 S. Casas-Flores and A. Herrera-Estrella
complexes in different fungi will allow us to understand the complexity in regulation by light in secondary metabolism.
3.10 Cyclic AMP/Protein Kinase A
A variety of environmental cues are transduced in many organisms by the heterotrimeric G protein (a, b and g) and heterotrimeric GPCRs play a pivotal role in this process. Basically, there are two main signal transduction pathways involving GPCR: (i) the cyclic adenosine monophosphate (cAMP) and (ii) phosphati-dylinositol (PI) pathways (Jae-Hyuk, 2006). In fungi, G proteins regulate mating, cell growth and division, morphogenesis, virulence and sec-ondary metabolite production (Li et al., 2007).
cAMP is an internal, and sometimes external, second messenger produced from ATP by adenylyl cyclase, which is activated by external and internal signals through the activation of adenylyl cyclase stimulatory Gs-coupled receptors and inhibited by ago-nists of adenylyl cyclase inhibitory Gi-coupledreceptors. Phosphodiesterase negatively reg-ulates cAMP intracellular concentrations. In eukaryotes cAMP activates protein kinase A (PKA), also known as cAMP-dependent protein kinase; in the absence of cAMP PKA remains in an inactivate state.
As mentioned earlier, it is well docu-mented that in Trichoderma a brief pulse of light provokes modifications of membrane properties (Horwitz et al., 1984c), an increase in cAMP levels (Farkas et al., 1985; Gresik et al., 1988), and a rapid and transient incre-ment in ATP intracellular concentrations and protein phosphorylation (Gresik et al., 1989). Interestingly, non-conidiating mutants showed no changes in intracellular levels of ATP (Farkas et al., 1985) and the addition of membrane-damaging agents inhibited photo-conidiation (Betina and Spisiaková, 1976). Furthermore, light induces adenylyl cyclase activity (Kolarova et al., 1992) and the addition of db-cAMP (dibutiryl cAMP), a membrane diffusible compound and analogue of cAMP, can mimic a light stimulus resulting in protein phosphorylation and conidiation in the dark, whereas that of an adenylate cyclase inhibitor
(atropin) blocks light induced conidiation (Gresik et al., 1989; Berrocal-Tito et al., 2000).
In this regard, an increase in cAMP dependent protein kinase (PKA) activity after a pulse of blue light was observed (Fig. 3.4). This activation occurred even in the Dblr-1 and Dblr-2mutant strains (Casas-Flores et al., 2006), sup-porting the existence of an alternative system for light perception linked to cAMP signalling and consistent with earlier biochemical data mentioned above. On the other hand, transfor-mants expressing an antisense version of pkr-1,a gene encoding the regulatory subunit of PKA, did not conidiate and overexpressed BLR-dependent blue-light-inducible genes after a pulse of blue light. In contrast, decreased levels of PKA activity, achieved by overexpression of the pkr-1 gene, result in conidiation, even in the dark, and result in no transcription of BLR-dependent blue-light-responsive genes. This evidence indicates the participation of the cAMP pathway in photoconidiation and transcription of BLR-dependent blue-light-responsive genes (Fig. 3.4). Together these results reinforce the participation of cAMP-dependent protein kinase in blue-light signal transduction and it has been proposed that the BLR proteins could be their targets (Fig. 3.4) (Casas-Flores et al., 2006).
In T. reesei, deletion of env1 leads to decreased intracellular levels of cAMP, whereas supplementation of culture medium with cAMP provoked an even more drastic growth phenotype when exposed to light, and the addition of caffeine (an inhibitor of phosphodiesterase) rescued the growth phe-notype of Denv1 strains (Tisch et al., 2011).
The respective molecular data suggest complex mechanisms involved in the cAMP-signalling pathway that regulates asexual reproduction and transcription of BLR-dependent blue-light-inducible genes in T. atroviride (Fig. 3.4). The light receptor responsible for the activation of the cAMP pathway has not been identified, however.
3.11 G proteins
In fungi, G proteins regulate morphogenesis, mating, cell growth/division, chemotaxis,

Light and the Biology of Trichoderma 59
secondary metabolism and virulence (Li et al.,2007). As described in the previous section, the first evidence of the participation of G proteins in a development process in Trichoderma was the fact that the addition of cAMP to the growing medium induced conidiation in the dark, bypassing the require-ment of light or stress, whereas atropine, a compound that blocks cAMP production, inhibited conidiation even after a blue-light pulse (Betina et al., 1976; Berrocal-Tito et al.,2000). Later, Rocha-Ramírez and co-workers (2002) isolated from T. atroviride a Ga gene with highest similarity to Ga inhibitory pro-teins (Gai). T. atroviride strains bearing an overexpression construct of Gai were unable to conidiate under light conditions, in con-trast to wild type. Transgenic strains bearing the wild-type tga1 gene and at least one copy of the mutant allele (QL-1), the product of which is locked in the GTP-bound state and therefore in permanent signal transmission, showed the same behaviour as the overex-pressing transformants in light. Transfor-mants expressing an antisense version of tga1showed two phenotypes: one of them resem-bles the wild-type phenotype in response to light, whereas the other phenotype showed constitutive sporulation even in the dark, with no distinction of the ring of green conidia formed by the wild-type strain.
Recently, gna3, the orthologous gene of tga3 in T. reesei was characterized (Schmoll et al., 2009). Mycelia of T. reesei exposed to light showed that tga3 is expressed at low levels in the darkness, whereas a light pulse induced its transcription (Fig. 3.4). Analysis of gna3 mRNA in a Denv1 strain showed an increased expression level in the dark, whereas in light conditions the transcript was hardly detected. This is indicative of a negative role of ENV1 on the transcription of gna3 in the darkness (Fig. 3.4). A strain carrying a constitutively activated allele generated by replacing Q206 by L of gna3 showed similar behaviour in the transcription of the endoge-nous gna3 gene under light and dark condi-tions when compared with the wild-type strain. Nevertheless, the gna3QL strain showed fivefold higher cAMP levels than the wild-type strain in the light. Synthesis of cAMP by a gna3 antisense (gna3AS) strain
growing for 72 h in constant light presented reduced levels of cAMP, whereas overexpres-sion of gna3 did not increase the accumula-tion of the nucleotide above wild-type levels. This indicates that activation of GNA3 is important for this process. None of the strains showed an important alteration in conidia-tion. Growth of the gna3QL strain on carbon sources that do not normally induce cellulase expression in the wild type did not result in cellulase gene expression, regardless of the light condition. When both strains were grown on cellulose in the dark, expression of cbh1 was very similar to that found in the wild type. When the strains were cultured under light conditions, however, the cbh1gene showed a delay in mRNA transcription in the gna3AS strain, resembling its expres-sion in a Denv1 mutant, whereas the gna3QL strain presented a substantially increased level of cbh1 mRNA. Therefore, the role of GNA3 directly correlates with cellulase gene expression in the presence of light (Fig. 3.4). Similarly, a strain bearing a construct that constitutively expresses gna3 showed no dif-ferences in cbh1 gene transcription when grown in the dark. Upon light stimulation, however, cbh1 clearly accumulated to higher levels than in the wild-type strain. These data indicate that GNA3 modulates cellulase gene transcription in a light-dependent manner (Fig. 3.4). Transcription analysis of genes that encode the light-perception proteins BLR-1, BLR-2 and ENV1 in a gna3QL background showed no differences from the wild-type strains. Together these results and the up-regulation of gna3 in a Denv1 background indicate that GNA3 is a downstream compo-nent of the light-perception proteins BLR-1, BLR-2 and ENV1, which modulates the light response of the cellulase encoding genes cbh1and cbh2 (Fig. 3.4) (Schmoll et al., 2009).
In T. reesei, the gna1 gene encodes a class I Ga subunit. In contrast to what has been described for gna3, gna1 is not regu-lated by light (Fig. 3.4). Cultures of Dgna1and gna1QL growing on 1% microcrystalline cellulose as carbon source under light condi-tions showed no accumulation and increased transcription of cbh1 compared with the wild-type strain, respectively. No major changes were observed in the dark. These results

60 S. Casas-Flores and A. Herrera-Estrella
indicate that GNA1 plays a role in the regu-lation of cellulase gene expression on cellu-lose and that this role depends on light (Fig. 3.4) (Seibel et al., 2009).
To determine the hierarchical interac-tions of GNA1, GNA2 and ENV1, the light responses of their encoding genes were meas-ured in glycerol under different light condi-tions. T. reesei was grown on glycerol as carbon source because glucose inhibits inducer uptake, whereas the use of glycerol as carbon source allows cellulase gene expres-sion. The env1 transcript was very low in the darkness, whereas upon a pulse of light it increased 400-fold in the wild-type strain. In the Dgna1 background, the env1 messenger presented similar behaviour to that of the wild-type strain; however, after a light pulse the env1 transcript remained elevated for a longer period of time. It was therefore con-cluded that GNA1 has a negative regulation on the env1 transcript and its deletion enhances or prolongs the light response of env1 (Fig. 3.4) (Tisch et al., 2011). Furthermore, ENV1 showed a negative effect on the accu-mulation of the gna3 transcript (Schmoll et al.,2009). It was then concluded that there is a hierarchical signal transduction pathway of GNA1, ENV1 and GNA3, in that order.
When the env1 gene was deleted from the genomes of the gna1QL (GNA1QLE) and gna3QL (GNA3QLE) transformants, a severe growth phenotype was observed under constant light for both strains, resem-bling the phenotype of the Denv1 strain growing in the same conditions (Castellanos et al., 2010). Analysis of the hyphal exten-sion rates and conidiation of the double mutants (GNA1QLE and GNA3QLE), grow-ing under light and dark conditions, showed the same phenotypes as their corresponding parental strains GNA1QL and GNA3QL, whereas the Denv1 strain growing in the dark showed a wild-type phenotype. These results indicate that ENV1 is epistatic to GNA3 and GNA1, suggesting that GNA3 and GNA1 are downstream of ENV1 under light conditions.
As mentioned above, GNA3 strongly regulates cAMP levels (Fig. 3.4), whereas GNA1 has a minor effect. Mutants in the cAMP-dependent protein kinase (Schuster
et al., 2012) or in Denv1 grown in light indi-cated that neither mutant displayed hyphal avoidance or formed exploratory hyphae. These observations led to the hypothesis that ENV1 could be regulating cAMP levels (Fig. 3.4). Measurements of cAMP intracellu-lar levels in GNA1QLE and NA3QLE and Denv1 showed very low levels of the nucleo-tide in all strains. These results indicated that ENV1 is important in maintaining the intra-cellular cAMP levels and for hyphal growth in light (Fig. 3.4). The addition of cAMP to the media did not rescue the Denv1 phenotype, leading to the conclusion that decreased cAMP levels were not due to the activity of adenylate cyclase in the mutants. This sup-ported an alternative explanation, in which phosphodiesterases are the target of ENV1. The addition to the medium of phosphodies-terase inhibitors enhanced the growth of Denv1 in the presence of light. These results indicated that ENV1 negatively regulates phosphodiesterase activity in light, resulting in the accumulation of cAMP (Fig. 3.4) (Tisch et al., 2011).
Transcriptomic analysis of T. reesei grow-ing on media with microcrystalline cellulosefor 72 h in constant light and constant dark-ness showed that 248 genes were differen-tially regulated (2.7%). These results are in agreement with those reported for other asco-mycetes (Rosales-Saavedra et al., 2006; Chen et al., 2009a,b; Ruger-Herreros et al., 2011). Among these genes, a phosducin-like protein 1 (PhLPp1) was clearly induced by light after 60 min of exposure (Tisch et al., 2011).
Members of the phosducin gene family were initially proposed to act as down-regulators of G proteins by binding to G-protein bg dimers (Gbg) and inhibiting their ability to interact with the G-protein asubunit (Ga) and effectors. Recently, how-ever, it has been demonstrated that members of the phosducin family act as co-chaperones, to assist folding of proteins from their nascent polypeptides. On the other hand, phosducin-like proteins (PhLP1) have been shown to be essential for G-protein signalling by catalys-ing the folding and assembly of the Gbg dimer (Willardson and Howlett, 2007).
Deletion of env1 led to high expression of phlp1 in response to light, whereas no

Light and the Biology of Trichoderma 61
considerable differences were observed in blr-1 and blr-2 mutant background. These results point to a major role for ENV1 in the regulation of phlp1 by light, leading to the postulation of a possible role of PhLP1 in the light response through the G-protein b (GNB1) and g (GNG1) subunits. On the basis of these asseverations, the corresponding mutants were generated. Cultures of Dphlp1, Dgnb1and Dgng1 growing in constant light and con-stant darkness showed an increase in conidia-tion for the three strains, whereas only a minor negative effect on conidiation was observed in Dphlp1 in the darkness. The fact that the three mutants showed similar pheno-types points to the participation of their prod-ucts in the same pathway and that probably PhLP1 acts in the efficient folding of GNB1 (Fig. 3.4) (Tisch et al., 2011). Genome-wide transcriptional analysis by growing the three mutants and the wild-type strain on cellulose as a carbon source, under constant light or dark-ness, showed that the three genes are essen-tial for the regulation of all light-responsive genes. The analysis of the transcriptome also led to the identification of glycoside hydro-lase encoding genes (GH), which were down regulated in a Dgnb1 mutant growing under light conditions. From the 628 genes posi-tively regulated by PhLP1, GNB1 and GNG1 in light, the predominant functional group were the glycoside hydrolases with 21 target genes. Analysis of genes encoding for enzymes that degrade cellulosic substrates led to the observation of a positive effect by light on tran-script levels. Light did not affect negatively gene expression of any celulase-encoding gene. A total of 11 out of 16 hemicellulase encoding genes were differentially expressed at least twofold in light and darkness in the wild-type and/or the mutant strains. Thus, PhLP1, GNB1 and GNG1 are important for the regulation of cellulase gene expression through the heterotrimeric G-protein path-way (Fig. 3.4).
PhLP1, GNB1 and GNG1 were shown to regulate genes involved in sexual develop-ment, including the pheromone precursor gene hpp1 and the orthologue of the yeast pheromone transporter in ste6. The hpp1 and ste6 transcripts were considerably down-regulated in the Dphlp1 mutant compared
with the wild type strain and this gene was co-regulated with ste6 in Dphlp1 and Dgng1.The pheromone precursor showed basal lev-els in all three mutants in the dark, support-ing the importance of light for sexual reproduction. Expression levels of hpp1 were down-regulated in all three mutants when exposed to light and the lack of PhLP1 pro-voked a decrease even in the dark levels, whereas deletion of gnb1 led to up-regulation of ste6, reflecting a reaction to the decreased pheromone levels (Tisch et al., 2011).
3.12 MAPK
Mitogen-activated protein kinase (MAPK) pathways transduce a large variety of sig-nals regulating different cellular processes. A MAPK has been found to be implicated in light responses, oxidative stress occasioned by light and in the control of circadian rhythms in both fungi and animals (de Paula et al., 2008). Trichoderma spp. have three genes encoding MAPK named tmk1, tmk2 and tmk3. Many genes regulated by light identi-fied in Trichoderma spp., are implicated in different stresses such as oxidative stress and DNA damage. Among them, the tmk3gene encoding MAPK related to p38/Hog1 and an homologue to pbs2 MAPKK of Saccharomyces cerevisiae are induced by light through the BLR proteins (Schuster et al., 2007; A. Herrera-Estrella, unpublished data), suggesting that this stimulus also impacts upon a well-conserved stress response MAPK cascade. As it occurs in other model systems, Tmk3/Hog1 is phosphorylated in response to oxidative and osmotic stress among others, protecting the cell against harmful effects of various factors, including light (Delgado-Jarana et al., 2006; Esquivel-Naranjo, unpub-lished data). In N. crassa, OS-2 (p38/Hog1) activity shows circadian oscillations and it is regulated by the WC complex, presum-ably anticipating daily changes occasioned by sunlight (de Paula et al., 2008). Together these data suggest that light is seen as stress signal, triggering protection systems and perhaps anticipating harmful effects provoked by light.

62 S. Casas-Flores and A. Herrera-Estrella
3.13 Light and Sexual Development in Trichoderma/Hypocrea
Recently, sexual development has been established in the laboratory for T. reesei(Hypocrea jecorina) but so far in no other Trichoderma spp., although numerous, pre-sumably sexually competent, strains have been tested (Kuhls et al., 1996). Asexual spor-ulation in T. reesei – albeit induced by light (Eveleigh, 1985) – is much less sensitive to the light status than in many other fungi including T. atroviride and T. virens, in that the original strain QM6a sporulates well also in darkness, albeit to a somewhat decreased extent than in light. Sexual development, however, requires light to occur, which has not only been shown with the strains described (Seidl et al., 2009; Chen et al., 2012) but is also apparent with other natural iso-lates of H. jecorina (for a more complete description on the effect of light on sexual development see Chapter 4, this volume).
Mating of Denv1, Dblr-1 and Dblr-2(derived form CBS999.97) (Seidl et al., 2009) with the wild-type strain was fruitful and discharge of ascospores was not abated; these data indicated that deletion of photo-receptors does not affect male fertility (Seibel et al., 2012). Sexual reproduction in fully male and female wild-type back-ground, bearing mutation in blr-1 and blr-2,showed altered fruiting body formation compared with the wild type with fewer, but larger, fruiting bodies. The blr-1 and blr-2 products repress the formation of stro-mata under constant light and are necessary to speed up the formation of stromata under a 12-h light/12-h dark regime (Chen et al., 2012). In contrast, crosses of MAT1-1 and MAT1-2 bearing a deletion in env1 provoked a defect in sexual development character-ized by the lack of fruiting bodies, and ENV1 was shown to be essential for female fertility in T. reesei CBS999.97.
Recently, it was demonstrated that genes involved in sexual development such as the pheromone precursor gene hpp1 and the orthologous gene to the pheromone transporter in yeast ste6 are regulated by
PhLP1, and the G-protein b (GNB1) and g (GNG1) subunits. It has been proposed that phosducin-like proteins mediate light transduction through the G-protein signal transduction pathway (Fig. 3.4) (Tisch et al., 2011) (see Chapter 4, this volume).
3.14 Conclusions and Perspectives
During the last decade, significant advances have been made towards understanding light perception and the molecular events triggered by this environmental cue in Trichoderma. The consequences of exposure to light on growth and development are also better understood. We are, however, still far from elucidating the extremely com-plex network involved in all observed responses to light. So far, all efforts have been focused on understanding blue-light responses through what is perhaps the main blue-light perception system, composed of two basic elements the BLR complex and ENV1. Nevertheless, it should still be proven whether there are physical interactions among them, as shown for their N. crassaorthologues. It should also be investigated how these photoreceptors are interacting with the signalling cascades affected upon exposure to blue light. It is fundamental to understand the role, if any, of all other poten-tial photoreceptors found in the Trichodermagenome. Is it for instance possible that, like in Aspergillus, the switch between sexual and asexual reproduction is not only influenced by blue light but also by red light? Is it pos-sible that a higher order light-perception complex, involving several receptors, is formed?
On the other hand, it is clear that light influences growth, reproduction and carbon source utilization. Accordingly, it is only logi-cal to expect that such a strong influence as light is reflected in the ecological behaviour of these fungi. In this sense, it would be plaus-ible that mutants affected in their light per-ception machinery would be disadvantaged, even if released in the soil.

Light and the Biology of Trichoderma 63
References
Aguirre, J., Rios-Momberg, M., Hewitt, D. and Hansberg, W. (2005) Reactive oxygen species and development in microbial eukaryotes. Trends in Microbiology 13, 111–118.
Bayram, O., Krappmann, S., Ni, M., Bok, J.W., Helmstaedt, K., Valerius, O., Braus-Stromeyer, S., Kwon, N.J., Keller, N.P., Jae-Hyuk, Y. and Braus, G. (2008a) VelB/VeA/LaeA complex coordinates light signalwith fungal development and secondary metabolism. Science 320, 1504–1506.
Bayram, O., Krappmann, S., Seiler, S., Vogt, N. and Braus, G.H. (2008b) Neurospora crassa ve-1 affects asexual conidiation. Fungal Genetics and Biology 45, 127–138.
Bergman, K., Burke, P.V., Cerda-Olmedo, E., David, C.N., Delbruck, M., Foster, K.W., Goodell, E.W., Heisenberg, M., Meissner, G., Zalokar, M., Dennison, D.S. and Shropshire Jr, W. (1969) Phycomyces. Bacteriology Review 33, 99–157.
Berrocal-Tito, G., Sametz-Baron, L., Eichenberg, K., Horwitz, B.A. and Herrera-Estrella, A. (1999) Rapid blue light regulation of a Trichoderma harzianum photolyase gene. Journal of Biological Chemistry 274, 14288–14294.
Berrocal-Tito, G.M., Rosales-Saavedra, T., Herrera-Estrella, A. and Horwitz, B.A. (2000) Characterization of blue-light and developmental regulation of the photolyase gene phr1 in Trichoderma harzianum. Photochemistry and Photobiology 71, 662–668.
Berrocal-Tito, G., Esquivel-Naranjo, E.U., Horwitz, B.A. and Herrera-Estrella, A. (2007) Trichoderma atroviridePHR1, a fungal photolyase responsible for DNA repair, autoregulates its own photoinduction. Eukaryotic Cell 6, 1682–1692.
Betina, V. and Spisiaková, J. (1976) Suppression of photo-induced sporulation in Trichoderma viride by inhibitors. Folia Microbiologica 21, 362–370.
Betina, V. and Zajacová, J. (1978) Inhibition of photo-induced Trichoderma viride conidiation by inhibitors of RNA synthesis. Folia Microbiologica 23, 460–464.
Blumenstein, A., Vienken, K., Tasler, R., Purschwitz, J., Veith, D., Frankenberg-Dinkel, N. and Fischer, R. (2005) The Aspergillus nidulans phytochrome FphA represses sexual development in red light. Current Biology 15, 1833–1838.
Brunner, M. and Káldi, K. (2008) Interlocked feedback loops of the circadian clock of Neurospora crassa. Molecular Microbiology 68, 255–262.
Calvo, A.M., Wilson, R.A., Bok, J.W. and Keller, N.P. (2002) Relationship between secondary metabolism and fungal development. Microbiology and Molecular Biology Reviews 66, 447–459.
Carreras-Villaseñor, N., Sánchez-Arreguin, J.A. and Herrera-Estrella, A. (2012) Trichoderma: sensing the environment for survival and dispersal. Microbiology 158, 3–16.
Casas-Flores, S., Rios-Momberg, M., Bibbins, M., Ponce-Noyola, P. and Herrera-Estrella, A. (2004) BLR-1 and BLR-2, key regulatory elements of photoconidiation and mycelial growth in Trichoderma atroviride. Microbiology 150, 3561–3569.
Casas-Flores, S., Rios-Momberg, M., Rosales-Saavedra, T., Martinez-Hernandez, P., Olmedo-Monfil, V. and Herrera-Estrella, A. (2006) Cross talk between a fungal blue-light perception system and the cyclic AMP signaling pathway. Eukaryotic Cell 5, 499–506.
Castellanos, F., Schmoll, M., Martinez, P., Tisch, D., Kubicek, C.P., Herrera-Estrella, A. and Esquivel-Naranjo, E.U. (2010) Crucial factors of the light perception machinery and their impact on growth and cellulase gene transcription in Trichoderma reesei. Fungal Genetics and Biology 47, 468–476.
Chen, C.H. and Loros, J.J. (2009a) Neurospora sees the light: Light signaling components in a model system. Communicative and Integrative Biology 2, 448–451.
Chen, C.H., Ringelberg, C.S., Gross, R.H., Dunlap, J.C. and Loros, J.J. (2009b) Genomewide analysis of light-inducible responses reveals hierarchical light signaling in Neurospora. EMBO Journal 28, 1029–1042.
Chen, C.L., Kuo, H.C., Tung, S.Y., Hsu, P.W.C., Wang, C.L., Seibel, C., Schmoll, M., Chen, R.S. and Wang, T.F. (2012) Blue light acts as a double-edged sword in regulating sexual development of Hypocrea jecorina(Trichoderma reesei). PLoS ONE 7, e44969.
Darwin, C. (1880) The Power of Movement in Plants. Available at: http://darwin-online.org.uk/EditorialIntroductions/Freeman_ThePowerofMovementinPlants.html (accessed 25 September 2012).
de Paula, R.M., Lamb, T.M., Bennett, L. and Bell-Pedersen, D. (2008) A connection between MAPK path-ways and circadian clocks. Cell Cycle 7, 2630–2634.
Deitzer, G.F., Horwitz, B.A. and Gressel, J. (1988) Rhythms in blue-light-induced conidiation of wild type and a mutant strain of Trichoderma harzianum. Photochemistry and Photobiology 47, 424–431.

64 S. Casas-Flores and A. Herrera-Estrella
Delgado-Jarana, J., Sousa, S., González, F. and Llobell, A. (2006) ThHog1 controls the hyperosmotic stress response in Trichoderma harzianum. Microbiology 152, 1687–1700.
Esquivel-Naranjo, E.U. and Herrera-Estrella, A. (2007) Enhanced responsiveness and sensitivity to blue light by blr-2 overexpression in Trichoderma atroviride. Microbiology 153, 3909–3922.
Estrada, A.F. and Avalos, J. (2008) The White Collar protein WcoA of Fusarium fujikuroi is not essential for photocarotenogenesis, but is involved in the regulation of secondary metabolism and conidiation. Fungal Genetics and Biology 45, 705–718.
Eveleigh, D.E. (1985) Trichoderma. In: Demainand, A.L. and Solomon N.A. (eds) Biology of Industrial Microorganisms. Benjamin Cummings Publishing Co., California, USA, pp. 487–509.
Farkas, V., Sulová, Z. and Lehotsky, J. (1985) Effect of light on the concentration of adenine nucleotides in Trichoderma viride. Journal of General Microbiology 131, 317–320.
Friedl, M.A., Kubicek, C.P. and Druzhinina, I.S. (2008a) Carbon source dependence and photostimulation of conidiation in Hypocrea atroviridis. Applied and Environmental Microbiology 74, 245–250.
Friedl, M.A., Schmoll, M., Kubicek, C.P. and Druzhinina, I.S. (2008b) Photostimulation of Hypocrea atroviridisgrowth occurs due to a cross-talk of carbon metabolism, blue light receptors and response to oxida-tive stress. Microbiology 154, 1229–1241.
Galun, E. (1971) Scanning electron microscopy of intact Trichoderma colonies. Journal of Bacteriology 108, 938–940.
Galun, E. and Gressel, J. (1966) Morphogenesis in Trichoderma: suppression of photoinduction by 5-fluorouracil. Science 151, 696–698.
Greenshpan, D. (1970) Morphogenetic studies on the fungus Trichoderma viride. PhD thesis, Weizmann Institute of Science, Rehovot, Israel.
Gresik, M., Kolarova, N. and Farkas, V. (1988) Membrane potential, ATP, and cyclic AMP changes induced by light in Trichoderma viride. Experimental Mycology 12, 295–301.
Gresik, M., Kolarova, N. and Farkas, V. (1989) Light-stimulated phosphorylation of proteins in cell-free extracts from Trichoderma viride. FEBS Letters 248, 185–187.
Gressel, J. (1979) Yearly review: Blue light photoreception. Photochemistry and Photobiology 30, 749–754.Gressel, J. (1980) Photocontrol of transcription. In: Senger, H. (ed.) The Blue Light Syndrome. Springer, Berlin,
Heidelberg, New York, pp. 133–153.Gressel, J. and Galun, E. (1967) Morphogenesis in Trichoderma: photoinduction and RNA. Developmental
Biology 15, 575–598.Gressel, J. and Hartmann, K.M. (1968) Morphogenesis in Trichoderma: Action spectrum of photoinduced
sporulation. Planta 79, 271–274.Gressel, J. and Rau, W. (1983) Photocontrol of fungal development. In: Shropshire Jr, W. and Mohr, H. (eds)
Encyclopedia of Plant Physiology. Vol. 16B. Springer, Berlin, pp. 603–639.Gressel, J., Bar-Lev, S. and Galun, E. (1975) Blue light induced response in the absence of free oxygen. Plant
and Cell Physiology 16, 367–370.He, Q. and Liu, Y. (2005) Molecular mechanism of light responses in Neurospora: from light-induced tran-
scription to photoadaptation. Genes and Development 19, 2888–2899.Heintzen, C., Loros, J.J. and Dunlap, J.C. (2001) The PAS protein VIVID defines a clock-associated feedback
loop that represses light input, modulates gating, and regulates clock resetting. Cell 104, 453–464.Horwitz, B.A., Gressel, J. and Malkin, S. (1984a) The quest for Trichoderma cryptochrome. In: Senger, H.
(ed.) Blue Light Effects in Biological Systems. Springer-Verlag, Berlin, pp. 237–249.Horwitz, B.A., Malkin, S. and Gressel, J. (1984b) Roseoflavin inhibition of photoconidiation in a Trichoderma
riboflavin auxotroph: indirect evidence for flavin requirement for photoreactions. Photochemistry and Photobiology 40, 763–769.
Horwitz, B.A., Weisenseel, M.H., Dorn, A. and Gressel, J. (1984c) Electric currents around growing Trichoderma hyphae, before and after photoinduction of conidiation. Plant Physiology 74, 912–916.
Horwitz, B.A., Gressel, J. and Malkin, S. (1985) Photoperception mutants in Trichoderma: mutants that sporulate in response to stress but not light. Current Genetics 9, 605–613.
Horwitz, B.A., Perlman, A. and Gressel, J. (1990) Induction of Trichoderma sporulation by nanosecond laser pulses: evidence against cryptochrome cycling. Photochemistry and Photobiology 51, 99–104.
Jae-Hyuk, Y. (2006) Heterotrimeric G protein signaling and RGSs in Aspergillus nidulans. Journal of Microbiology 44, 145–154.
Jiao, Y., Lau, O.S. and Deng, X.W. (2007) Light regulated transcriptional networks in higher plants. Nature Reviews Genetics 8, 217–230.
Kolarova, N., Haplová, J. and Gresík, M. (1992) Light-activated adenyl cyclase from Trichoderma viride. FEMS Microbiology Letters 72, 275–278.

Light and the Biology of Trichoderma 65
Komon-Zelazowska, M., Neuhof, T., Dieckmann, R., von Döhren, H., Herrera Estrella, A., Kubicek, C.P. and Druzhinina, I.S. (2007) Formation of atroviridin by Hypocrea atroviridis is conidiation associated and positively regulated by blue light and the G proteína GNA3. Eukaryotic Cell 6, 2332–2342.
Kuhls, K., Lieckfeldt, E., Samuels, G.J., Kovacs, W., Meyer, W., Petrini, O., Gams, W., Börner, T. and Kubicek, C.P. (1996) Molecular evidence that the asexual industrial fungus Trichoderma reesei is a clonal derivative of the ascomycete Hypocrea jecorina. Proceedings of the National Academy of Sciences of the USA 93, 7755–7760.
Kumagai, T. and Oda, Y. (1969) An action spectrum for photoinduced sporulation in the fungus Trichoderma viride. Plant and Cell Physiology 10, 387–392.
Li, L., Wright, S.J., Krystofova, S., Park, G. and Borkovich, K.A. (2007) Heterotrimeric G proteins signaling in filementous fungi. Annual Review of Microbiology 61, 423–452.
Lilly, V.G. and Barnett, H.L. (1951) Physiology of the Fungi. McGraw-Hill Book Company Inc., USA.Liu, Y. and Bell-Pedersen, D. (2006) Circadian rhythms in Neurospora crassa and other filementous fungi.
Eukaryotic Cell 5, 1184–1193.Lledias, F. and Hansberg, W. (2000) Catalase modification as a marker of singlet oxygen. Methods in
Enzymology 319, 110–119.Mikus, M., Hatvani, L., Neuhof, T., Komon-Zelazowska, M., Dieckman, R., Schwecke, I.S., Druzhinina, I.S.,
von Döhren, H. and Kubicek, C.P. (2009) Differential regulation and posttranslational processing of the class II hydrophobin genes from the biocontrol fungus Hypocrea atroviridis. Applied and Environmental Microbiology 75, 3222–3229.
Mukherjee, P.K. and Kenerley, C.M. (2010) Regulation of morphogenesis and biocontrol properties in Trichoderma virens by a VELVET protein, Vel1. Applied and Environmental Microbiology 76, 2345–2352.
Nakari-Setälä, T., Aro, N., IImén, M., Muñoz, G., Kalkkinen, N. and Penttilä, M. (1997) Differential expression of the vegetative and spore-bound hydrophobins of Trichoderma reesei–cloning and characterization of the hfb2 gene. European Journal of Biochemistry 248, 415–423.
Nemkovic, M. and Farkas, V. (1998) Stimulation of conidiation by derivatives of cAMP in Trichoderma viride. Folia Microbiologica 43, 399–402.
Nuhof, T., Dieckmenn, R., Druzhinina, I.S., Kubicek, C.P. and von Döhren, H. (2007) Intact-cell MALDI-TOF mass spectrometry analysis of peptaibol formation by the genus Trichoderma/Hypocrea: can molecular phylogeny of species predict peptaibol structures? Microbiology 153, 3417–3437.
Presti, D.E. and Galland, P. (1987) Photoreceptor biology of Phycomyces. In: Cerda-Olmedo, E. and Lipson, E.D. (eds) Phycomyces. Cold Spring Harbor Laboratory Press, New York.
Rocha-Ramirez, V., Omero, C., Chet, I., Horwitz, B.A. and Herrera- Estrella, A. (2002) Trichoderma atrovirideG-protein a-subunit gene tga1 is involved in mycoparasitic coiling and conidiation. Eukaryotic Cell 1, 594–605.
Rosales-Saavedra, T., Esquivel-Naranjo, E.U., Casas-Flores, S., Martínez Hernández, P., Ibarra-Laclette, E., Cortes-Penagos, C. and Herrera-Estrella, A. (2006) Novel light-regulated genes in Trichoderma atroviride:a dissection by cDNA microarrays Microbiology 152, 3305–3317.
Ruger-Herreros, C., Rodríguez-Romero, J., Fernández-Barranco, R., Olmedo, M., Fischer, R., Corrochano, L.M. and Canovas, D. (2011) Regulation of conidiation by light in Aspergillus nidulans. Genetics 188, 809–822.
Schmoll, M. (2008) The information highways of a biotechnological workhorse – signal transduction in Hypocrea jecorin. BMC Genomics 9, 430–454.
Schmoll, M., Zeilinger, S., Mach, R.L. and Kubicek, C.P. (2004) Cloning of genes expressed early during cellulase induction in Hypocrea jecorina by a rapid subtraction hybridization approach. Fungal Genetics and Biology 41, 877–887.
Schmoll, M., Franchi, L. and Kubicek, C.P. (2005) Envoy, a PAS/LOV domain protein of Hypocrea jecorina(anamorph Trichoderma reesei), modulates cellulase gene transcription in response to light. EukaryoticCell 4, 1998–2007.
Schmoll, M., Schuster, A., Silva, R.N. and Kubicek, C.P. (2009) The G-alpha protein GNA3 of Hypocrea jecorina(anamorph Trichoderma reesei) regulates cellulase gene expression in the presence of light. EukaryoticCell 8, 410–420.
Schuster, A., Kubicek, C.P., Friedl, M.A., Druhinina, I.S. and Schmoll, M. (2007) Impact of light on Hypocrajecorina and the multiple cellular roles of ENVOY in this process. BMC Genomics 8, 449–465.
Schuster, A., Tisch, D., Seidl-Seiboth, V., Kubicek, C.P. and Schmoll, M. (2011) Roles of proteína kinase A and adenilate cyclase in light-modulated cellulase regulation in Trichoderma reesei. Applied and Environmental Microbiology 78, 2168–2178.
Schwerdtfeger, C. and Linden, H. (2001) Blue light adaptation and desensitization of light signal transduction in Neurospora crassa. Molecular Microbiology 39, 1080–1087.

66 S. Casas-Flores and A. Herrera-Estrella
Schwerdtfeger, C. and Linden, H. (2003) VIVID is a flavoprotein and serves as a fungal blue light photoreceptor for photoadaptation. EMBO Journal 22, 4846–4855.
Seibel, C., Gremel, G., do Nascimento Silva, R., Schuster, A., Kubicek, C.P. and Schmoll, M. (2009) Light-dependent roles of the G protein a subunit GNA1 of Hypocrea jecorina (anamorph Trichoderma reesei).BMC Biology 7, 58.
Seibel, C., Tisch, D., Kubicek, C.P. and Schmoll, M. (2012) ENVOY is a mejor determinant in regulation of sexual development in Hypocrea jecorina (Trichoderma reesei). Eukaryotic Cell 11, 885–896.
Steyaert, J.M., Weld, R.J., Loguercio, L.L. and Stewart, A (2010a) Rhythmic conidiation in the blue-light fungus Trichoderma pleuroticola. Fungal Biology 114, 219–223.
Steyaert, J.M., Weld, R.J., Mendoza-Mendoza, A. and Stewart, A. (2010b) Reproduction without sex: conidiation in the filamentous fungus Trichoderma. Microbiology 156, 2887–2900.
Tisch, D. and Schmoll, M. (2009) Light regulation of metabolic pathways in fungi. Applied Microbiology and Biotechnology 85, 1259–1277.
Tisch, D., Kubicek, C.P. and Schmoll, M. (2011) New insights into the mechanism of light modulated signal-ing by heterotrimeric G-proteins: ENVOY acts on gna1 and gna3 and adjusts cAMP levels in Trichoderma reesei (Hypocrea jecorina). Fungal Genetics and Biology 48, 631–640.
Vargovic, P., Pokorny, R., Holker, U., Hofer, M. and Varecka, L. (2006) Light accelerates the splicing of srh1homologue gene transcripts in aerial mycelia of Trichoderma viride. FEMS Microbiology Letters 254, 240–244.
Whitmore, L. and Wallace, B.A. (2004) The peptaibol database: a database for sequences and structures of naturally occurring peptaibols. Nucleic Acids Research 32, 593–594.
Willardson, B.M. and Howlett, A.C. (2007) Function of phosducin-like proteins in G protein signaling and chaperone-assisted protein folding. Cellular Signaling 19, 2417–2427.
Related Documents











