ELSEVIER THE INFLUENCE OF CAMP BEFORE OR DURING BOVINE OOCYTE MATURATION ON EMBRYONIC DEVELOPMENTAL COMPETENCE Z. Guixue,’ A.M. Luciano,* K. Coenen,’ F. Gandolfi* and M.A. Sirardla ‘CRBR, Department of Animal Science, Laval University, Quebec, Canada GlK 7P4 *Department of Anatomy of Domestic Animals, University of Milan, 20134 Milan, Italy Received for publication: March 30, 2000 Accepted: November 13, 2000 ABSTRACT This study was designed to evaluate the effects of pretreatment with various forms of CAMP before or during bovine oocyte maturation on the acquisition of embryonic developmental competence. The objective of the 4 experiments was to induce differentiation of the early maturing oocyte in conditions of maintained meiotic arrest or normal maturation. To promote differentiation, different forms of cyclic AMP-dependent protein kinase (PKA) pathways were investigated. The factors studied included follicular fluid, invasive adenylate cyclase (iAC), dibutyryl-CAMP (dbcAMP) and 3-isobutyl-1-methyl-xanthine (IBMK) with or without cycloheximide (CHK). High concentrations of iAC pretreatment were beneficial to the oocyte competence in BSA-iAC maturation while harmful in normal maturation. Also, after 2 to 3 h IBMK-iAC pretreatment, another 6 h of CHX treatment with or without iAC was harmful to the embryonic developmental competence of fertilized oocytes even though it did not have any effect on cleavage rate. Experiment 4 was to assess the role of CAMP in acquisition of oocyte developmental competence before meiotic resumption. Results supported that the intracellular CAMP concentration during the interval between oocyte isolation from the follicle and the beginning of in vitro maturation is critical for requiring optimal developmental competence. Q 2001 by Elsev~er Science Inc. Key words: bovine oocyte, meiotic arrest, dbcAMP, iAC, developmental competence INTRODUCTION The status of growth and differentiation of a follicle have a clear impact on oocyte capacity to become an embryo. Generally, in vitro fertilization (IVF) protocols use cumulus- enclosed oocytes (CO&) aspirated from follicles 1 to 5 mm in diameter found in bovine ovaries collected at a slaughterhouse. However, 60 to 80% of those selected are not developmentally competent and do not reach the blastocyst stage. The oocytes collected from follicles smaller than 2 mm in diameter rarely develop beyond the g-cell stage, whereas oocytes obtained from follicles with diameters measuring 2 to 4 mm and >4 mm had comparable developmental competence (5, 10. 11. 26, 28). Some researchers believe that a growing follicle must reach a minimum of 3 to 4 mm diameter for the oocyte to acquire the capacity to form an embryo. This aCorrespondence: Dr Marc-Andre Sirard, CRBR, Department of Animal Science, Laval University, Quebec, Canada GlK 7P4. FAX 418-656-3766; E-Mail: Marc-Andre.Sirard@ crbr.ulaval.ca Theriogenology 55: 1733-l 743,200l 0093-691W01/$-see front matter 0 2001 Elsevier Science Inc. PII: 50093-691X(01)00516-7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER
THE INFLUENCE OF CAMP BEFORE OR DURING BOVINE OOCYTE MATURATION ON EMBRYONIC DEVELOPMENTAL COMPETENCE
Z. Guixue,’ A.M. Luciano,* K. Coenen,’ F. Gandolfi* and M.A. Sirardla
‘CRBR, Department of Animal Science, Laval University, Quebec, Canada GlK 7P4
*Department of Anatomy of Domestic Animals, University of Milan, 20134 Milan, Italy
Received for publication: March 30, 2000 Accepted: November 13, 2000
ABSTRACT
This study was designed to evaluate the effects of pretreatment with various forms of CAMP before or during bovine oocyte maturation on the acquisition of embryonic developmental competence. The objective of the 4 experiments was to induce differentiation of the early maturing oocyte in conditions of maintained meiotic arrest or normal maturation. To promote differentiation, different forms of cyclic AMP-dependent protein kinase (PKA) pathways were investigated. The factors studied included follicular fluid, invasive adenylate cyclase (iAC), dibutyryl-CAMP (dbcAMP) and 3-isobutyl-1-methyl-xanthine (IBMK) with or without cycloheximide (CHK). High concentrations of iAC pretreatment were beneficial to the oocyte competence in BSA-iAC maturation while harmful in normal maturation. Also, after 2 to 3 h IBMK-iAC pretreatment, another 6 h of CHX treatment with or without iAC was harmful to the embryonic developmental competence of fertilized oocytes even though it did not have any effect on cleavage rate. Experiment 4 was to assess the role of CAMP in acquisition of oocyte developmental competence before meiotic resumption. Results supported that the intracellular CAMP concentration during the interval between oocyte isolation from the follicle and the beginning of in vitro maturation is critical for requiring optimal developmental competence. Q 2001 by Elsev~er Science Inc.
Key words: bovine oocyte, meiotic arrest, dbcAMP, iAC, developmental competence
INTRODUCTION
The status of growth and differentiation of a follicle have a clear impact on oocyte capacity to become an embryo. Generally, in vitro fertilization (IVF) protocols use cumulus- enclosed oocytes (CO&) aspirated from follicles 1 to 5 mm in diameter found in bovine ovaries collected at a slaughterhouse. However, 60 to 80% of those selected are not developmentally competent and do not reach the blastocyst stage. The oocytes collected from follicles smaller than 2 mm in diameter rarely develop beyond the g-cell stage, whereas oocytes obtained from follicles with diameters measuring 2 to 4 mm and >4 mm had comparable developmental competence (5, 10. 11. 26, 28). Some researchers believe that a growing follicle must reach a minimum of 3 to 4 mm diameter for the oocyte to acquire the capacity to form an embryo. This
aCorrespondence: Dr Marc-Andre Sirard, CRBR, Department of Animal Science, Laval University, Quebec, Canada GlK 7P4. FAX 418-656-3766; E-Mail: Marc-Andre.Sirard@
crbr.ulaval.ca
Theriogenology 55: 1733-l 743,200l 0093-691W01/$-see front matter 0 2001 Elsevier Science Inc. PII: 50093-691X(01)00516-7

1734 Theriogenology
capacity increases as the follicle reaches a plateau or a reduction of its growth rate, either by dominance or early atresia (26). Alternatively, large follicles do not always yield oocytes capable of producing embryos, while some oocytes from medium size follicles already have this capacity (26). Competent oocytes could originate from early atretic follicles because oocytes are affected at the very last stage of follicular atresia. In addition, many healthy follicles do not always contain competent oocytes. Therefore, the degree of follicular differentiation appears to play a more important role in developmental competence than other factors such as follicular size or health status (4).
Because oocytes matured in vivo versus those matured in vitro do not differ significantly in the level of nuclear maturation, fertilization or cleavage rates, the most striking difference between these two types of oocytes is their developmental potential (26). This strongly suggests major differences at the level of cytoplasmic maturation between oocytcs matured in vivo and those matured in vitro (14-16). When oocytes are aspirated from follicles, their ability to synthesize protein is not affected, but they lose the capacity to make RNA in less than 2 h unless arrest of meiosis is maintained artificially (9, 26, 28). The acquisition of developmental potential involves the accumulation of important factors in the form of protein or stable mRNA. Standard maturation protocols usually involve the addition of serum, estradiol, FSH, LH or other factors to facilitate in vitro development. In recent years, research focused on methods to improve developmental competence by controlling the interval between the time the oocyte is aspirated and the completion of maturation. Some of these experiments yielded encouraging results (19, 21, 25, 27).
The second messenger, CAMP. is a differentiating factor found in follicular cells and oocytes. Cyclic AMP maintains meiotic arrest in GVBD-competent oocytes and also helps growing oocytes to acquire GVBD competence (6). Cyclic AMP is also involved in the signalling pathway of FSH (17, 18). Dibutyryl cyclic AMP (dbcAMP) is a CAMP analog (29). Invasive adenylate cyclase (iAC), an enzyme derived from Bordetella pertussis, increases the CAMP level by using the endogenous ATP pool (1,2, 12,31). Invasive adenylate cyclase has a biphasic effect on oocyte maturation since a high dose of iAC induces meiotic arrest, while a low dose sustains a high rate of maturation (21).
Isobutyl methyl xanthine (IBMX), a phosphodiesterase inhibitor, can maintain bovine CGCs in meiotic arrest for approximately 3 h while cycloheximide (CHX), a protein synthesis inhibitor, can maintain a reversible state of arrested meiosis for a longer period (20, 23, 24, 25, 29).
The aim of the experiments was to use CAMP to induce differentiation in the oocytes and their cumulus cells without inducing arrested meiosis. Additionally, cycloheximide was tested to maintain arrested meiosis while CAMP can have its differentiation effect directly on the oocyte or through the cumulus.
MATERIAL AND METHODS
Oocyte Recovery and Selection
Ovaries from non-pregnant or pregnant heifers or cows were removed within 30 min after slaughter and were transported to the laboratory within 4 to 6 h at 30 to 36°C in 0.9% NaCl

Theriogenology 1735
aqueous solution containing 100,000 III/L penicillin, 100 mg/L streptomycin and 250 @L arnphotericin B (Sigma-Aldrich, Oakville, Ontario, Canada). CGCs were aspirated from 2- to 5 mm follicles with an 18-g needle attached to a 10-mL syringe, pooled and selected according to cumulus morphology. After removal of oocytes, follicular fluid was obtained by centrifugation at 2,000 g for 3 min.
In Vitro Maturation
For in vitro maturation, groups of 10 CGCs were placed for 24 h in 50-pL droplets of maturation medium covered with filtered mineral oil (Sigma-Aldrich) and pre-incubated under the culture conditions for at least 2 h in an incubator at 385°C containing 5% CG2 and 95% air atmosphere with 100% humidity. Control maturation medium consisted of TCM-199 with Earle’s salts (Gibco BBL, Burlington, Ontario, Canada) and bicarbonate (Sigma-Aldrich) supplemented with 10% fetal calf serum (FCS; Medicorp, Montreal, Qdbec, Canada), 0.5 pg/mL FSH (National Institute of Diabetes and Digestive and Kidney Diseases; NIDDK, Bethesda, MD, USA), 5 pg/mL LH (NIDDK), 1 pg/mL estradiol-17P (Sigma-Aldrich), 0.2 mM pyruvic acid (Sigma-Aldrich) and 50 ug/mL gentamicin (Sigma-Aldrich). Invasive adenylate cyclase maturation medium consisted of TCM-199 with Earle’s salts and bicarbonate supplemented with 0.8% bovine serum albumin (BSA; Sigma-Aldrich), 0.01 pg/mL iAC (Alexis Biochemicals Corporation, San Diego, CA, USA), 0.2 mM pyruvic acid and 50 pg/mL gentamicin. The pH was adjusted to be similar to the control maturation medium with sodium hydroxide.
In Vitro Fertilization and In Vitro Development
For the fertilization procedure, 5 washed matured oocytes were placed for 18 to 19 h in 48- pL droplets of fertilization medium consisting of modified Tyrode lactate medium, 0.6% BSA fatty acid-free (Sigma-Aldrich), 0.2 mM pyruvic acid, 2 pg/mL heparin (Sigma-Aldrich), and 50 ug/mL gentamicin. After the transfer of oocytes, 2 pL of PHE (penicillamine 2 mM, hypotaurine 1 mM, epinephrine 250 mM; Sigma-Aldrich) were added to each droplet. Two straws of frozen semen from a pool of 5 bulls (Centre d’Indmination Artificielle du Quebec; CIAQ, St-Hyacinthe. Quebec, Canada) were thawed in a 35°C water bath for 1 min and were then separated by the swim-up procedure (22). To each droplet, 2 pL of sperm suspension (final
concentration = 1 x 106 cells/ml) were added. The fertilized oocytes were washed 3 times in Hepes buffered Tyrode’s medium (TLH) supplemented with 0.3% BSA, 0.2 mM pyruvic acid and 50 pg/mL gentamicin. The oocytes surrounded only with corona then were co-cultured with bovine oviductal vesicles (3) in 50-uL development droplets consisting of TCM-199, 10% FCS, 1% BSA, 0.2 mM pyruvic acid and 50 ug/mL gentamicin. After 2 d of culture, cleavage rates were recorded, and 50 pL of fresh medium was added. After 4 d of culture, embryos and oviductal vesicles were transferred into new drops. Finally, after 6.5 d of culture, embryos were fixed and the number of nuclei counted.

1736 Theriogenology
Experimental Design
Bovine oocytes commit to undergo meiosis soon after their removal from the follicles (8, 29). Therefore, to prevent priming of WCs, follicular fluid, IBMX and iAC were used during CGCs selection, rinsing and 15 min pretreatment in order to maintain the arrested meiosis.
In Experiment 1 we investigated the effect of pretreatment CGCs with different iAC concentrations before maturation on the embryonic development competence. CGCs were kept in follicular fluid supematant during the selection procedure and then transferred for 15 min at 25°C in follicular fluid or pretreatment mediums containing 0.1 pg/mL, 0.2 l.tg/mL or 0.3 l.tg/mL iAC in TCM-199 with Earle’s salts and bicarbonate supplemented with 25 mM Hepes buffer, 0.4% BSA (fraction V, Sigma-Aldrich), 0.5 mM IBMX (Sigma-Aldrich), 0.2 mM pyruvic acid and 50 @/mL gentamicin. Then, CGCs were distributed in control maturation medium after 2 quick washes in the same medium.
In Experiment 2 we investigated the influence of different concentrations of iAC during pretreatment before maturation with or without iAC. Control-WCs (Groups 1 and 2) were kept in follicular fluid supematant during the selection procedure and then transferred for 15 min at 25°C in follicular fluid before maturation in control or iAC maturation mediums. Invasive adenylate cyclase treated CGCs (Groups 3 to 8) were aspirated with a IO-mL syringe containing 0.5 to 1 mL of pretreatment medium containing 0.1 pg/mL iAC. Aspiration was continued until reaching approximate total volume of 3 mL (pretreatment medium + follicular fluid). Selection of oocytes was carried out in their corresponding supematant and then transferred for 15 min at 25°C in pretreatment medium containing 0.1 pg/mL, 0.2 pg/mL or 0.3 pg/mL iAC before maturation in control or iAC maturation medium.
The IBMX can maintain bovine CGCs in meiotic arrest for approximately 3 h while CHX can maintain a reversible state of meiotic arrest for a longer period (19, 25, 29). Therefore, CHX was used to replace the IBMX pretreatment for another 6 h.
In Experiment 3 we mainly evaluated the effect of long term pretreatment (6 h) with a combination of iAC and CHX before maturation. Control COCs (Groups 9 and 10) were kept in follicular fluid supematant during the selection procedure, transferred for 15 min at 25°C in to pretreatment medium containing 0.1 pg/mL iAC then matured for a period of 24 or 30 h in control maturation medium. All the other groups of CGCs (11 to 15) were aspirated and selected as were the iAC-treated CGCs in Experiment 2; and then transferred for 15 min at 25°C in pretreatment medium containing 0.1 l.tg/mL iAC. Oocytes of Groups 11 and 12 immediately were transferred on maturation to iAC maturation medium for a period of 24 or 30 h. For CHX- treated oocytes (Groups 13 to 15) a pretreatment of 6 h on meiotic arresting medium, with or without 0.01 l.tg/mL iAC, was added before control or iAC maturation of 24 h. The meiotic arresting medium consisted of TCM-199 with Earle’s salts and bicarbonate, 10% FCS, 0.2 mM pyruvic acid, 50 pg/mL gentamicin, and 2 l.tg/mL CHX (Sigma-Aldrich). To remove CHX after incubation, oocytes quickly were double rinsed in maturation medium.
In Experiment 4 we investigated the role of CAMP in acquisition of the oocyte’s developmental competence before meiosis resumed. The CGCs were aspirated, selected and washed in oocyte washing medium alone (Groups 16 to 19) or supplemented with 2 j.tg/mL CHX
Theriogenology 1737
(Groups 20 to 23). The oocyte washing medium consisted of TLH supplemented with 10% FCS. 0.2 mM pyruvic acid and 50 pg/mL gentamicin. Selected oocytes then were incubated for 6 h in meiosis-arresting medium (Groups 20 to 23) or medium without CHX (Groups 16 to 19) containing 0, 0.01. 0.1 or 1 mM dbcAMP. The 6 h period was followed by 2 washes in oocyte washing medium and a 24-h control maturation.
Statistical Analysis
Each experiment consisted of 3 replicates. Individual comparisons were performed using the Chi-square analysis or an analysis of variance (ANOVA) followed by the New Multiple Range Test (30).
RESULTS
As shown in Table 1, maintaining CGCs in arrested meiosis with follicular fluid and exposing the CGCs to increasing doses of iAC (0.1, 0.2 and 0.3 pg/mL) for 15 min before maturation did not improve the oocyte’s cleavage rate nor did it improve embryonic developmental competence under normal maturation conditions.
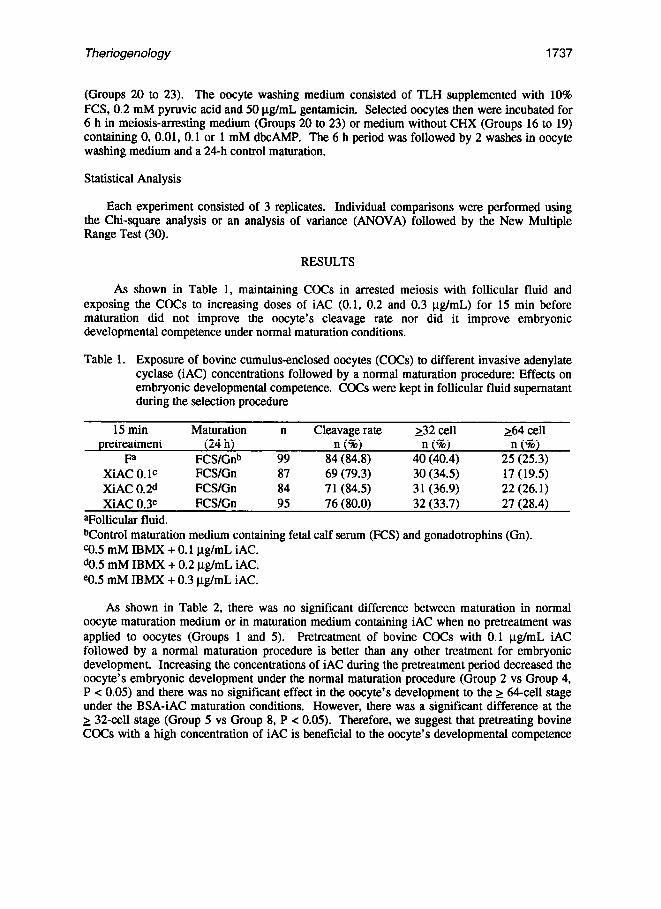
Table 1. Exposure of bovine cumulus-enclosed oocytes (CGCs) to different invasive adenylate cyclase (iAC) concentrations followed by a normal maturation procedure: Effects on embryonic developmental competence. CGCs were kept in follicular fluid supematant during the selection procedure
15 min Maturation n Cleavage rate 232 cell *4 cell pretreatment (24 h) n (%) n (%) n (%)
Fa FCS/Gnb 99 84 (84.8) 40 (40.4) 25 (25.3) XiAC 0. lc FCS/Gn 87 69 (79.3) 30 (34.5) 17 (19.5) XiAC 0.2d FCS/Gn 84 71 (84.5) 31 (36.9) 22 (26.1) XiAC 0.3e FCS/Gn 95 76 (80.0) 32 (33.7) 27 (28.4)
aFollicular fluid. bControl maturation medium containing fetal calf serum (FCS) and gonadotrophins (Gn). CO.5 mM IBMX + 0.1 pg/mL iAC. do.5 mM IBMX + 0.2 pg/mL iAC. e0.5 mM IBMX + 0.3 pg/mL iAC.
As shown in Table 2, there was no significant difference between maturation in normal oocyte maturation medium or in maturation medium containing iAC when no pretreatment was applied to oocytes (Groups 1 and 5). Pretreatment of bovine CGCs with 0.1 pg/mL iAC followed by a normal maturation procedure is better than any other treatment for embryonic development. Increasing the concentrations of iAC during the pretreatment period decreased the oocyte’s embryonic development under the normal maturation procedure (Group 2 vs Group 4, P < 0.05) and there was no significant effect in the oocyte’s development to the >64-cell stage under the BSA-iAC maturation conditions. However, there was a significant difference at the 1 32-cell stage (Group 5 vs Group 8, P < 0.05). Therefore, we suggest that pretreating bovine CGCs with a high concentration of iAC is beneficial to the oocyte’s developmental competence

1738
when maturation is carried out in media supplemented normal maturation procedure.
Theriogenology
with BSA-iAC and harmful under the
Table 2. Effects on the oocyte’s developmental capacity of pretreating bovine cumulus- enclosed oocytes for 15 min with increasing concentrations of invasive adenylate cyclase (iAC) before maturation with or without iAC
Group Aspiration/ 15 min pre- Maturation n Cleavage rate 132 cells ~64 cells selection treatment (24 h) n (8) n (%) n (%)
1 Fa F FCS/Gnb 111 83 (78.4)Cd 29 (26.l)Cd 18 (16.2)Cd 2 FXiACe XiAC 0. lr FCWGn 101 78 (77.2)Cd 27 (26.7)C“ 26 (25.7)C 3 FXiAC XiAC 0.2s FCS/Gn 102 70 (68.6)d 23 (22.5)cd 15 (14.7)Cd 4 FXiAC XiACO.3h FCSlGn 98 7 1 (72.4)Cd 20 (zo.q=’ 13 (13.3)“ 5 F F BSA-iAC1 124 92 (74.2)cd 24 (19.4)d 15 (12.1)d 6 FXiAC XiAC 0.1 BSA-iAC 105 78 (74.3)Cd 30 (28.6)cd 17 (16.2)Cd 7 FXiAC XiAC0.2 BSA-iAC 98 76 (77.4)Cd 25 (25.5)Cd 18 ( 18.4)Cd 8 FXiAC XiAC0.3 BSA-iAC 88 71 (80.7)C 29 (33.O)C 18 (20.5)C“
aFollicular fluid. bControl maturation medium containing fetal calf serum (FCS) and gonadotrophins (Gn). C*dDifferent letters within the same column indicate significant differences (P < 0.05). eFollicular fluid + pretreatment medium containing 0.5 mM IBMX + 0.1 pg/mL iAC. f0.5 mM IBMX + 0.1 pg/mL iAC. so.5 mM IBMX + 0.2 pg/mL iAC. ho.5 mM IBMX + 0.3 pg/mL iAC. iiAC maturation medium containing bovine serum albumin (BSA).
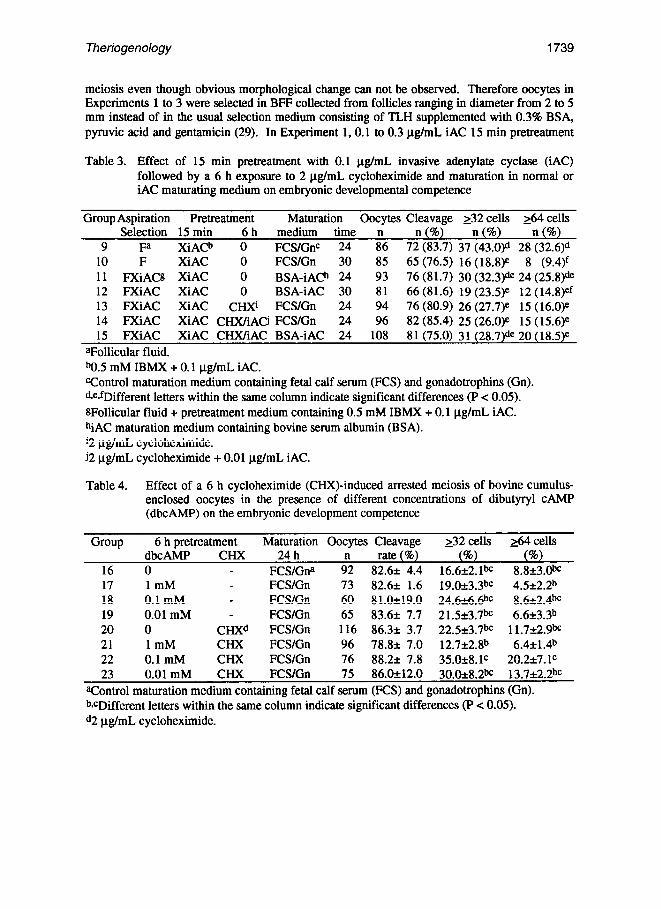
The results of Experiment 3 are shown in Table 3. Significant morphological differences were observed after the CGC pretreatment and maturation. The cumulus cells of CGCs maturing in normal maturation medium expanded well (Group 9). while those maturing in the iAC maturation medium (Group 11) showed little expansion. The 30 h maturation period had detrimental effects on IVM, with Groups 9 and 11 showing the best developmental competence (P < 0.05) at 2 32- and 2 64-cell stages. In addition, pretreatment of CGCs for 2 h (aspiration time + 15 min pretreatment) with IBMX-iAC followed by another 6 h in CHX with or without iAC was harmful to the later stages of embryonic development. Interestingly, this treatment did not seem to affect the oocyte’s cleavage rate.
As shown in Table 4, CGC exposure to 0.1 mM dbcAMP during CID&induced arrested meiosis is better than 1 mM for embryonic development. In absence of CHX, the different concentrations of dbcAMP had no effect on developmental competence.
DISCUSSION
Follicular fluid from follicles of 2 to 5 mm in diameter shows a higher meiosis-inhibiting effect than does fluid from follicles of 5 to 10 mm (3, 7, 13). Bovine follicular fluid may act in association with CAMP to block spontaneous GVBD of mouse oocytes (32). The selection of 150 to 200 oocytes generally requires about 1 to 1.5 h during which oocytes may start resuming
Theriogenology 1739
meiosis even though obvious morphological change can not be observed. Therefore oocytes in Experiments 1 to 3 were selected in BFF collected from follicles ranging in diameter from 2 to 5 mm instead of in the usual selection medium consisting of TLH supplemented with 0.3% BSA, pyruvic acid and gentamicin (29). In Experiment LO.1 to 0.3 pg/mL IAC 15 min pretreatment
Table 3. Effect of 15 min pretreatment with 0.1 ug/mL invasive adenylate cyclase (iAC) followed by a 6 h exposure to 2 pg/mL cycloheximide and maturation in normal or iAC maturating medium on embryonic developmental competence
Group Aspiration Pretreatment Maturation Gocytes Cleavage 132 cells ~64 cells Selection 15 min 6 h medium time n n (%) n (%) n (%)
9 Fa XiACb 0 FCS/GnC 24 86 72 (83.7) 37 (43.W 28 (32.6Y’ . 10 F XiAC 0 FCS/Gn 30 85 65 (76.5) 16 (18.8je 8 (9.4)f
11 FXiACtt XiAC 0 BSA-iACh 24 93 76 (81.7) 30 (32.3)* 24 (25.8)de 12 FXiAC XiAC 0 BSA-iAC 30 81 66 (81.6) 19 (23.5)e 12 (14.8)ef 13 FXiAC XiAC CHXi FCS/Gn 24 94 76 (80.9) 26 (27.7)e 15 (16.0)e 14 FXiAC XiAC CI-IX/iAci FCS/Gn 24 96 82 (85.4) 25 (26.0)e 15 (15.6)e 15 FXiAC XiAC CHX/iAC BSA-iAC 24 108 81 (75.0) 31 (28.7)de 20 (18.5)e
aFollicular fluid. b0.5 mM IBMX + 0.1 pg/mL IAC. ‘Control maturation medium containing fetal calf serum (FCS) and gonadotrophins (Gn). Qe$rDifferent letters within the same column indicate significant differences (P < 0.05). sFollicular fluid + pretreatment medium containing 0.5 mM IBMX + 0.1 pg/mL iAC. hiAC maturation medium containing bovine serum albumin (BSA). ‘2 ug/mL cycloheximide. j2 ug/mL cycloheximide + 0.01 pg/mL iAC.
Table 4. Effect of a 6 h cycloheximide (CHX)-induced arrested meiosis of bovine cumulus- enclosed oocytes in the presence of different concentrations of dibutyryl CAMP (dbcAMP) on the embryonic development competence
Group 6 h pretreatment Maturation Oocytes Cleavage 132 cells ti4 cells dbcAMP CHX 24 h
16 0 FCSlGna 9: rate (8) (96) (a)
82.6+ 4.4 16.6+2.1& 8.8k3.w 17 1mM FCS/Gn 73 82.6* 1.6 19.0+3.3bc 4.5+2.2b 18 0.1 mM - FCS/Gn 60 81.Ok19.0 24.6&6.6bC 8.6+2.4bC 19 0.01 mM - FCS/Gn 65 83.6* 7.7 21.523.7h 6.6+3.3b 20 0 CHXd FCS/Gn 116 86.3* 3.7 22.5*3.7& 11.7+2.9& 21 1mM CHX FCWGn 96 78.8* 7.0 12.7k2.8” 6.4*1.4b 22 0.1 mM CHX FCSlGn 76 88.22 7.8 35.0*8.1C 20.2+7.1C 23 0.01 mM CHX FCS/Gn 75 86.Ok12.0 3O.Ozt8.2t’C 13.7&z!bc
aControl maturation medium containing fetal calf serum (FCS) and gonadotrophins (Gn). b*cDifferent letters within the same column indicate significant differences (P < 0.05). d2 ug/mL cycloheximide.

1740 Theriogenology
of oocytes did not affect embryonic developmental competence. Recently Luciano et al. (21) reported the importance of iAC in the washing medium before culture to improve the blastocyst rate. In our experiment although no detrimental effect were observed, the iAC priming was without effect. If we compare to in vivo conditions, a sharp increase in CAMP followed by a gradual decrease could mimic some of the effect of the LH surge.
It is clear why not all oocytes are capable of embryonic development when recovered from small follicles obtained at slaughter. To continue the oocyte endogenous developmental program, the influence from a larger follicle could be beneficial if the oocyte were maintain in a responsive mode. Culture of intact antral follicles for 24 h is an alternative method for keeping maintenance of bovine oocytes in meiotic arrest but these oocytes do not acquire a greater developmental competence in vitro (9). Bovine oocytes start condensing their DNA soon after removal from the follicular environment (8. 29). Perhaps oocytes acquire some developmental capacity in a very short period before resumption of meiosis. Therefore, oocytes were kept in meiotic arrest by aspirating and selecting them in BFF supplemented with a high dose of IAC. These oocytes then were exposed to different concentrations of iAC to try to enhance their developmental competence. The addition of iAC had either positive or negative effects on the oocyte’s developmental competence depending on the type of media used for the maturation procedure (Table 2). In presence of iAC alone during maturation (without serum or hormones), oocytes do not perform as well compared to the regular culture medium. But when the oocytes are primed with iAC at levels 10 to 30 times higher during the pretreatment, more developmental capacity is observed with increasing iAC (19.4% vs 338, P c 0.05). The ability of iAC to penetrate the oocyte membrane allows it to remain active even after repeated washing of the COCs following exposure to concentrations as low as 0.01 ug/mL iAC. This toxin can potentially mimic the roles of LH and FSH on CCC’s maturation. It could also act in synergy with those hormones to promote maturation. In vitro maturation of bovine oocytes in the presence of FSH retards nuclear maturation initially via a CAMP-mediated pathway, while it enhances fertilizing ability and developmental ability of the oocytes (17). The time required for completion of nuclear maturation was influenced by gonadotropins used to supplement the maturation medium. The LH enhanced the speed of nuclear maturation when compared to FSH (18).
Arresting meiosis could improve the response to differentiation factors such as CAMP. In Experiment 3, adding 6 h of maturation (24 to 30) did not improve competence. However, the addition of a 6 h oocyte pretreatment with CHX or CHX plus iAC partially overcome this aging effect (16 vs 9%). On the other hand, there were no significant differences when maturation occured in media containing BSA and iAC.
The IBMX can maintain bovine COCs in meiotic arrest for approximately 3 h while CHX can maintain a reversible state of meiotic arrest for a longer period (20, 25, 29). In this condition, a narrow range of dbcAMP concentration is required for optimal developmental capacity of bovine oocytes (Table 4). This finding suggests that a low concentration of intracellular CAMP during arrested meiosis before in vitro maturation was effective in promoting cytoplasm maturation of bovine oocytes. The level of intracellular CAMP concentration during the interval between oocyte isolation from the follicle and the beginning of in vitro maturation seems critical for requiring optimal development competence.

Theriogenology 1741
Initiation of nuclear maturation is marked by the breakdown of the nuclear envelope, or germinal vesicle, and is triggered by LH. Although a decrease in intraoocyte levels of CAMP precedes germinal vesicle breakdown (GVBD), the gonadotropin induction of ovulation and oocyte maturation is associated with major increases of CAMP in ovarian follicles (31). Based on these results, our experiment first involves exposure of CGCs to a high concentration of dbcAMP and then maturation in media deprived of dbcAMP. Experiment 4 shows that among the concentrations of dbcAMP used, 0.1 mM is capable of increasing the oocyte’s developmental competence during cycloheximideinduced arrested meiosis (Table 4). This is the second study in which the presence of a very well defined amount (0.1 vs 1 mM) of CAMP results in a significant improvement of developmental competence (21). In our case the beneficial effect is observed during a period of arrested meiosis and not in the absence of CHX. supporting the hypothesis that the oocyte can be influence during that period. The difference between the dbcAMP treatment and the iAC is one of precision where the amount present inside the oocyte is better controlled in quantity and in time with the agonist than with the enzyme (whose activity cannot be removed precisely).
In conclusion, this study confirms the importance of the priming period with the invasive adenylate cyclase and the possibility of replacing the impact of FSH-LH and estradiol by CAMP only.
1.
2.
3.
4.
5.
6.
7.
8.
9.
REFERENCES
Aktas H, Wheeler MB, First NL, Leibfried-Rutledge ML. Maintenance of meiotic arrest by increasing [cAMP]i may have physiological relevance in bovine oocytes. J Reprod Fertil 1995b;105:237-245. Aktas H, Wheeler MB, Rosenkrans CF Jr, First NL, Leibfried-Rutledge ML. Maintenance of bovine oocytes in prophase of meiosis I by high [cAMP]i. J Reprod Fertil 1995a;105:227-235. Bilodeau S, Fortier MA, Sirard MA. Effect of adenylate cyclase stimulation on meiotic resumption and cyclic AMP content of zona-free and cumulus-enclosed bovine oocytes in vitro. J Reprod Fertil 1993;97:5-11. Blondin P. Sirard MA. Gocyte and follicular morphology as determining characteristics for developmental competence in bovine oocytes. Mol Reprod Dev 1995;41:54-62. Brackett BG, Bousquet D, Boice ML, Donawick WJ, Evans JF, Dressel MA. Normal development following in vitro fertilization in the cow. Biol Reprod 1982; 27:147-158. Chesnel F, Wigglesworth K, Eppig JJ. Acquisition of meiotic competence by denuded mouse oocytes: participation of somatic-cell product(s) and CAMP. Dev Biol 1994;161:285-295. Dostal J, Pavlok A. Isolation and characterization of maturation inhibiting compound in bovine follicular fluid. Reprod Nutr Dev 1996;36:681-690. First NL, Leibfried-Rutledge ML, Sirard MA. Cytoplasmic control of oocyte maturation and species differences in the development of maturational competence. In: First NL, Haseltine (eds.), Meiotic inhibition: Molecular control of meiosis. New York, Alan R Liss inc., 1988;1-46. Fouladi Nashta AA, Waddington D, Campbell KH. Maintenance of bovine oocytes in meiotic arrest and subsequent development in vitro: A comparative evaluation of antral follicle culture with other methods. Biol Reprod 1998;59:255-262.

1742 Theriogenology
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
Greve T, Callesen H, Hyttel P. Follicular correlates with in-vitro fertilization in cattle. J Reprod Fertil 1989Suppl38: 117- 126. Hashimoto S, Saeki K, Nagao Y, Minami N, Yamada M, Utsumi K. Effects of cumulus cell density during in vitro maturation of the developmental competence of bovine oocytes. Theriogenology 1998;49:1451-1463. Heikinheimo 0, Gibbons WE. The molecular mechanisms of oocyte maturation and early embryonic development are unveiling new insights into reproductive medicine. Mol Hum Reprod 1998;4:745-756. Hinrichs K. Martin MG, Schmidt AL, Friedman PP. Effect of follicular components on meiotic arrest and resumption in horse oocytes. J Reprod Fertil 1995; 104: 149- 156. Hyttel P, Callesen H, Greve T. Ultrastructural features of preovulatory oocyte maturation in superovulated cattle. J Reprod Fertil 1986a;76:645-656. Hyttel P, Greve T, Callesen H. Ultrastructural aspects of oocytes maturation and fertilization in cattle. J Reprod Fertil 1989;Suppl38:35-47. Hyttel P, Xu KP, Smith S, Greve T. Ultrastructure of in vitro oocyte maturation in cattle. J Reprod Fertil 1986b;78:615-625. Izadyar F, Zeinstra E, Bevers MM. Follicle-stimulating hormone and growth hormone act differently on nuclear maturation while both enhance developmental competence of in vitro matured bovine oocytes. Mol Reprod Dev 1998;51:339-345. Kuran M, Hutchinson JS, Broadbent PJ. The response of bovine granulosa cells to different gonadotrophins in culture. Anim Reprod Sci 1996;45: l- 12. Lonergan P, Dinnyes A, Fair T, Yang X, Boland M. Bovine oocyte and embryo development following meiotic inhibition with butyrolactone I. Mol Reprod Dev 2000;57: 204-209. Lonergan P, Khatir H, Carolan C, Mermillod P. Bovine blastocyst production in vitro after inhibition of oocyte meiotic resumption for 24 h. J Reprod Fertil 1997;109:355-365. Luciano AM, Pocar P, Milanesi E, Modina S, Rieger D, Lauria A, Gandolfi F. Effect of different levels of intracellular CAMP on the in vitro maturation of cattle oocytes and their subsequent development following in vitro fertilization. Mol Reprod Dev 1999;54:86-91. Parrish JJ, Susko-Parrish JL, Leibfried-Rutledge ML, Critser ES, Eyestone WH, First NL. Bovine in vitro fertilization with frozen-thawed semen. Theriogenology 1986;25:591-600. Rose-Hellekant TA, Bavister BD. Roles of protein kinase A and C in spontaneous maturation and in forskolin or 3-isobutyl-1-methylxanthine maintained meiotic arrest of bovine oocytes. Mol Reprod Dev 1996;44:241-249. Saeki K, Nagao Y, Kishi M, Nagai M. Development capacity of bovine oocytes following inhibition of meiotic resumption by cycloheximide or 6-dimethylaminopurine. Theriogenology 1997;48:1161-1172. Simon M, Jilek F, Fulka J Jr. Effect of cycloheximide upon maturation of bovine oocytes. Reprod Nutr Dev 1989;29:533-540. Sirard MA, Blondin P. Oocyte maturation and IVF in cattle. Proc 13th Int Congr Anim Reprod; Research and Practice 1996;42:417-426. Sirard MA, Coenen K. Effects of inhibition of meiotic resumption upon the subsequent development of bovine oocytes in vitro. J Reprod Dev 1995;41:255-262. Sirard MA, Coenen K. The effect of hormones during in vitro meiotic inhibition with cycloheximide on subsequent development of bovine oocytes. Biol Reprod 1994;50 (Suppl 1):361 abst. Sirard MA, First NL. In vitro inhibition of oocyte nuclear maturation in the bovine. Biol Reprod 1988;39:229-234.

Theriogenology 1743
30. Snedecor GW, Cochran WG. Statistical methods. Ames: Iowa State University Press, 1980;507.
31. Tsafriri A, Chun SY, Zhang R, Hsueh AI, Conti M. Oocyte maturation involves compartmentalization and opposing changes of CAMP levels in follicular somatic and germ cells: studies using selective phosphodiesterase inhibitors. Dev Biol 1996;178:393- 402.
32. Tsafriri A, Dekel N, Bar-Ami S. The role of oocyte maturation inhibition in follicular regulation of oocyte maturation. J Reprod Fertil 1982$X541-551.
Related Documents











